Embed Size (px)
Citation preview

Cell Tissue Res (1988) 252:307-315 Cell and Tissue Reseat �9 Springer-Verlag 1988
Localization of vasopressin-, vasoactive intestinal polypeptide-, peptide histidine isoleucine- and somatostatin-mRNA in rat suprachiasmatic nucleus J. Patrick Card z, Sandra Fitzpatrick-McElligott 1, Illana Gozes 2, and Frank Baldino, Jr. 1 t Neurobiology Group, Medical Products Department, E.I. du Pont de Nemours and Co., Wilmington, Delaware, USA; 2 Department of Hormone Research, Wcizmann Institute of Science, Rehovot, Israel
Summary. Messenger RNAs (mRNA) coding for vasoactive intestinal polypeptide (VIP), peptide histidine isoleucine (PHI), somatostatin and vasopressin were localized in the suprachiasmatic nucleus (SCN) of the rat hypothalamus using in situ hybridization histochemistry. Specific mRNA coding for each of these peptides was distributed in areas coextensive with the immunohistochemical localization of the appropriate peptide. The autoradiographic signal pro- duced with probes to VIP and PHI created dense concentra- tions of silver grains over neuronal perikarya in the ventro- lateral SCN, and the coextensive distribution of both VIP- and PHI-mRNAs suggests that both peptides are synthe- sized within the same neurons. The distribution of somato- statin-mRNA was distinct from that of VIP and PHI. La- beled neurons are observed at the interface of the two SCN subdivisions and the distribution of these neurons is identi- cal to those shown to contain somatostatin immunoreactivi- ty. Vasopressin-mRNA is also differentially concentrated within neurons in the dorsomedial subdivision of the SCN in an area that is coextensive with vasopressin-immunoreac- tive perikarya. The discrete pattern of hybridization for each of these mRNAs indicates that each of these peptides are synthesized in SCN neurons and reaffirms the differen- tial distribution of each of these chemically defined cell populations within cytoarchitecturally distinct subdivisions of the nucleus.
Key words: Suprachiasmatic nucleus - In situ hybridization histochemistry - Vasoactive intestinal polypeptide - Peptide histidine isoleucine - Somatostatin - Vasopressin - Rat (Sprague-Dawley)
The critical role of the suprachiasmatic nucleus (SCN) in the regulation of circadian rhythms has stimulated an in- tense effort to delineate the functional organization of SCN neurons and their connections (cf. Rusak and Zucker 1979; Moore 1983; Rosenwasser and Adler 1986). An important outgrowth of these efforts has been the demonstration that the SCN is composed of two major subdivisions, which are distinguished by the differential concentration of chemi- cally distinct neurons as well as the terminal arborization
Send offpr&t requests to: Dr. J. Patrick Card, Medical Products Department, E.I. du Pont de Nemours and Company, Experimen- tal Station, E400/3444, Wilmington, Delaware 19898, USA
of visual afferents (cf. Moore and Card 1985; van den Pol and Tsujimoto 1985, for reviews). The dorsomedial aspect of the SCN contains vasopressin- and somatostatin-immu- noreactive neurons, but lacks a direct visual projection. In contrast, the ventrolateral aspect of the SCN receives dense and overlapping visual projections from the retina (Moore and Lenn 1972; Hendrickson et al. 1972) and intergenicu- late leaflet of the thalamus (Card and Moore 1982; Moore et al. 1984) and is also characterized by the differential con- centration of neurons containing vasoactive intestinal poly- peptide (VIP), peptide histidine isoleucine (PHI), and bom- besin (Card et al. 1981; van den Pol and Tsujimoto 1985). Since visual input is the primary entraining influence upon SCN neuronal activity, the selective arborization of visual afferents within a subdivision of the nucleus which is also distinguished by the chemical content of the neurons sug- gests that the two major SCN subdivisions may be differen- tially involved in the process of entrainment. This is appar- ent in recent studies which have presented evidence of dis- tinct differences in the functional activity of neurons in the ventrolateral and dorsomedial SCN subdivisions. Fol- lowing selective lesions of either of the SCN subfields, van den Pol and Powley (1979) reported loss of behavioral rhythms only in animals with lesions that selectively im- pinged upon the dorsomedial aspect of the nucleus. Similar indications that the dorsomedial SCN may be critical to the regulation of circadian rhythms are evident in the elec- trophysiological investigations of Shibata and collaborators (1984). These investigators have examined the influence of photoperiod upon the activity of individual SCN neurons in hypothalamic slice preparations. Their studies demon- strate that bilateral orbital enucleation or constant darkness reduces neuronal activity in the ventrolateral subdivision, but does not alter the activity of neurons in the dorsomedial subdivision. Thus, the SCN is subdivided not only by the differential concentration of visual afferents and chemically distinct neurons, but also by the functional activity of the neurons in these subdivisions. However, the extent to which the initiation, maintenance and modulation of circadian rhythmicity is dependent on alterations in gene expression of peptides in the SCN has been addressed only recently. Uhl and Reppert (1986) demonstrated that vasopressin- mRNA is present in SCN neurons and that levels of the message fluctuate significantly during the light and dark phases of the circadian cycle. The purpose of the present investigation was to utilize high-resolution in situ hybridiza- tion histochemistry to localize mRNA encoding VIP/PHI- 27-, vasopressin- and somatostatin-precursors in discrete

308
populations of SCN neurons. The cellular distribution of these specific mRNA transcripts is reported in terms of the distribution of immunoreactivity for each of these pep- tides during the light phase of the photoperiod.
Materials and methods
Animals and tissue preparation. Adult male Sprague Dawley rats (n = 15) weighing 150-300 g were used in the investiga- tion. All animals were maintained in a standardized photo- period (12 h light; light on at 0600) with food and water freely available. Each animal was anesthetized with an over- dose of sodium pentobarbitol and sacrificed by transcardiac perfusion fixation during the light phase of the photoperi- od. Perfusion solutions were sterilized and consisted of 25ml of 0.1 M phosphate-buffered saline followed by 200-300 ml of 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) containing 2% sodium periodate and 1.4% lysine-HCl (McLean and Nakane 1974). The brain was re- moved, postfixed in sterile fixative for 1 h at 4 ~ C, stored in sterile 0.1 M phosphate buffer overnight at 4 ~ C, cryo- protected and sectioned serially at 30 ~tm with a cryostat. Sections were cut in the coronal plane throughout the ros- trocaudal extent of the SCN, mounted on gelatin-coated slides and stored at - 2 0 ~ C prior to processing for in situ hybridization histochemistry. Tissue processed for immuno- histochemical analysis was sectioned with a freezing micro- tome.
Immunohistochemistry. Hsu's (Hsu et al. 1981) avidin-biotin modification of the immunoperoxidase method (Stern- berger 1979) was used for immunohistochemical localiza- tion of each of the peptides analyzed. The VIP antiserum was generously provided by Dr. J. Walsh (Center for Ulcer Research and Education, UCLA). The PHI antiserum was purchased from Peninsula Laboratories (Belmont, Califor- nia) and antisera to somatostatin and vasopressin were pur- chased from Immuno Nuclear Corporation (Minneapolis, Minnesota). Specificity of the antisera was established by elimination of immunoreactivity following preabsorbtion of each antiserum with a 10 pM concentration of the antigen it was generated against. All antisera were also subjected to extensive cross-blocking studies with antigens to all of the other antisera used in the investigation. Persistence of immunoreactivity in these studies served to further establish the specificity of each antiserum. Particular attention was paid to the cross-blocking studies which involved the VIP and PHI antisera. These two peptides are derived from the same precursor and contain extensive sequence homology (Itoh et al. 1983). Immunoreactivity identical to that ob- served with unblocked antisera was observed when VIP antiserum (1/1000 dilution) was preabsorbed with PHI anti- gen or when PHI antiserum (1/2000 dilution) was preincu- bated with VIP antigen. Consequently, it is clear that these antisera are specifically identifying the neuronal localization of the VIP and PHI antigens rather than demonstrating cross reactivity of the antisera.
Synthesis of oligonucleotides. Oligodeoxyribonucleotides were synthesized on an automated DNA synthesizer (Model 380A, Applied Biosystems Inc.) using the phosphite/phos- photriester method of Matteucci and Caruthers (1981). Each oligonucleotide was purified by reverse-phase HPLC on a ~tBondapak C18 column (Waters Associates; Fritz
et al. 1978). Probes were synthesized complementary to the glycoprotein-coding region of the rat mRNA coding for arginine vasopressin (24 mer) 5 'GGCAGAATCCACG- GACTCTTGTGT3' ) , the 3' coding region for rat somatos- tatin mRNA (39 mer), (5 'GGATGTGAATGTCTTCC- AGAAGAAGTTCTTGCAGCCAGC3 ' ) and 5' portions of the exons coding for PHI (33 mer) (5 'GTCGCT- GGTGAAAACTCCATCAGCATGCCTGGC3 ' ) and VIP (29 mer) ( 5 ' G T G T C G T T T G A C C G G C A C G G G G T C T T - CCG3') rat mRNA.
Probe labeling. The probes were labeled with the 3' tailing method (Lewis et al. 1986). The reaction is carried out in a mixture containing 15pmol of probe, 70pmol of [12SI]dCTP or [35S]dATP, and 20 units of terminal deoxy- nucleotidyl transferase (Boehringer Mannheim) in 100 mM potassium cacodylate (pH 7.2), 0.2 mM dithiothreitol and 2 mM CaC12 in a final volume of 25 Ixl. The reaction mix- ture was incubated at 37 ~ C for 1.5 h and then terminated by a 5-min incubation at 65 ~ C. Reaction products were separated from both the enzyme and unincorporated radio- labeled nucleotides by using NENSORB 20 (Dupont New England Nuclear) and were used without further purifica- tion for in situ hybridization histochemistry.
In situ hybridization h&tochem&try. The method used in our laboratory for in situ hybridization histochemistry has been previously described (Baldino and Davis 1986; Lewis et al. 1986; Davis et al. 1986). Briefly, tissue sections were delipidated in a graded series of alcohols (50%, 70%, 100%), rehydrated and rinsed in sterile water treated with diethylpyrocarbonate (DEP). This was followed by a 15-min incubation with proteinase K (1 ~tg/ml in 10 mM Tris-HC1, 2 mM CaClz, pH 7.4) at 37~ and a 30-min rinse in 2 x SSC (0.3 M sodium chloride/0.3 M sodium ci- trate at pH 7.2). After incubation in hybridization buffer for 1 h, the sections were then hybridized overnight at 37~ with radiolabeled oligonucleotide probes (1.5- 3.0 • 10 6 c.p.m, diluted in 300 ~tl of hybridization buffer per slide). The hybridization buffer contained 0.6 M sodium chloride, 10 mM tris-HC1 (pH 7.5), 10 • Denhardts Solu- tion (0.02% Ficoll, 0.02% polyvinylpyrrolidone, 0.02% bo- vine serum albumin), 1 mM EDTA, 0,05% yeast tRNA, 0.05% salmon sperm DNA, 10% (wt/vol) dextran sulfate and 50% formamide. Washing stringency varied for each probe, but in general the sections were rinsed in decreasing concentrations of sodium citrate to a final rinse of 0.5 x SSC at 37 ~ C. For preliminary observations, the slides were ex- posed to X-Ray film (Kodak XAR-5) for 3 5 days at -- 70 ~ C. X-Ray films were developed in a Kodak X-OMAT processor. Subsequently, the slides were dipped in Kodak NTB2 emulsion and kept in the dark at 4~ for 7 to 21 days. The emulsion-coated slides were developed with Ko- dak D-19 for 2 min at 16~ and fixed for 5 min, dehy- drated, cleared and examined by both dark-field and bright- field light microscopy.
Probe specificity. The specificity of each probe was deter- mined by the sensitivity of the hybridization to RNase pre- treatment, the thermal stability of the hybrid and the ability of the probe to recognize a single species of RNA on North- ern blots. Tissue sections were pretreated with ribonuclease 1 (10 pg/ml) in 50 mM Tris-HC1, 10 mM NaC1, 10 mM EDTA (pH 8.0) for 30 rain prior to hybridization. The ther-
mal stability of the D N A - R N A hybrids were determined as follows. The sections were hybridized overnight, rinsed at 25~ in 0.5 x SSC for 1 h, dried and then exposed to X-Ray film to obtain a 100% premelt value. Using one slide for each of the indicated temperatures the slides were re-rinsed in 0.5 x SSC and reexposed to X-ray film for the same length of time as for the premelt film. The loss of specific signal was determined by comparing the premelt and postmelt optical density measurements. Spot densito- metric values were obtained with a D U M A S image analysis system. Extrapolation of the melting curve at 50% melt gave an experimental Tm value for each synthetic probe which was then compared with the calculated theoretical Tm value. The theoretical Tm value was calculated as fol- lows:
Tm = 81.5 + 16.6 (log[Na + ])
+ 0.41 (% G + C) - 675/(probe length) - 1.0 (% mismatch) - 0.65% (Formamide)
For Northern-blot analysis, total R N A was isolated from hypothalamic homogenates by urea-lithium chloride extraction (Auffray et al. 1980). Poly A + R N A was iso- lated by affinity chromatography on oligo(dT)-cellulose (Aviv and Leder 1972). Hypothalamic R N A (2 lag/lane) was electrophoresed in a 1% agarose gel and transferred to ni- trocellulose. After the filter was baked for 1.5 h at 80 ~ C, it was placed in a hybridization solution [4 x SSC, 10 x Denhardt ' s solution, 3 m M glycine, denatured salmon sperm D N A (146 p.g/ml), yeast t R N A (33 lag/ml) for 1 h at 35 ~ C. Hybridization was carried out by adding a 3zp_ labeled oligonucleotide probe (3 x 10 6 c.p.m./ml) and incu- bated overnight at 40 ~ C. The probes were phosphorylated on their 5' end with [gamma-a2p]ATP (100 gCI/34 pmol) and T4 polynucleotide kinase according to previously pub- lished procedures (Wolfson et al. 1985). The filter was then washed (twice at room temperature and once at 40 ~ C) in 2 x SSC for 30 min and air-dried. Autoradiography on X- ray film (Kodak XAR-5) was performed overnight at - 70 ~ C with intensifying screens (Du Pont Cronex Lighten- ing Plus).
Results
Our data demonstrate a close correlation of peptide and m R N A localization in SCN neurons containing vasopres- sin, somatostatin, VIP and PHI. Both immunohistochemis- try and in situ hybridization demonstrate groups of neurons which are differentially concentrated within either the dor- somedial or ventrolateral subfields of the SCN and the loca- tion of these neurons is consistent with previous immunocy- tochemical studies (Krisch 1978; van Leeuwen et al. 1978; Dierickx and Vandesande 1979; Card et al. 1981; van den Pol and Tsujimoto 1985). In addition, controls for specifici- ty of the immunohistochemical and in situ hybridization procedures have provided further assurance of the specifici- ty of each technique. In all cases, immunoreactivity was blocked by preabsorption of each antiserum with the anti- gen that it was generated against. In addition, crossblocking preabsorption of each antiserum with all other antigens involved in the analysis did not compromise the immunore- activity that was observed with unblocked antisera in paired controls. Specificity o f the oligonucleotide probes and the in situ hybridization procedure were established as follows.
309
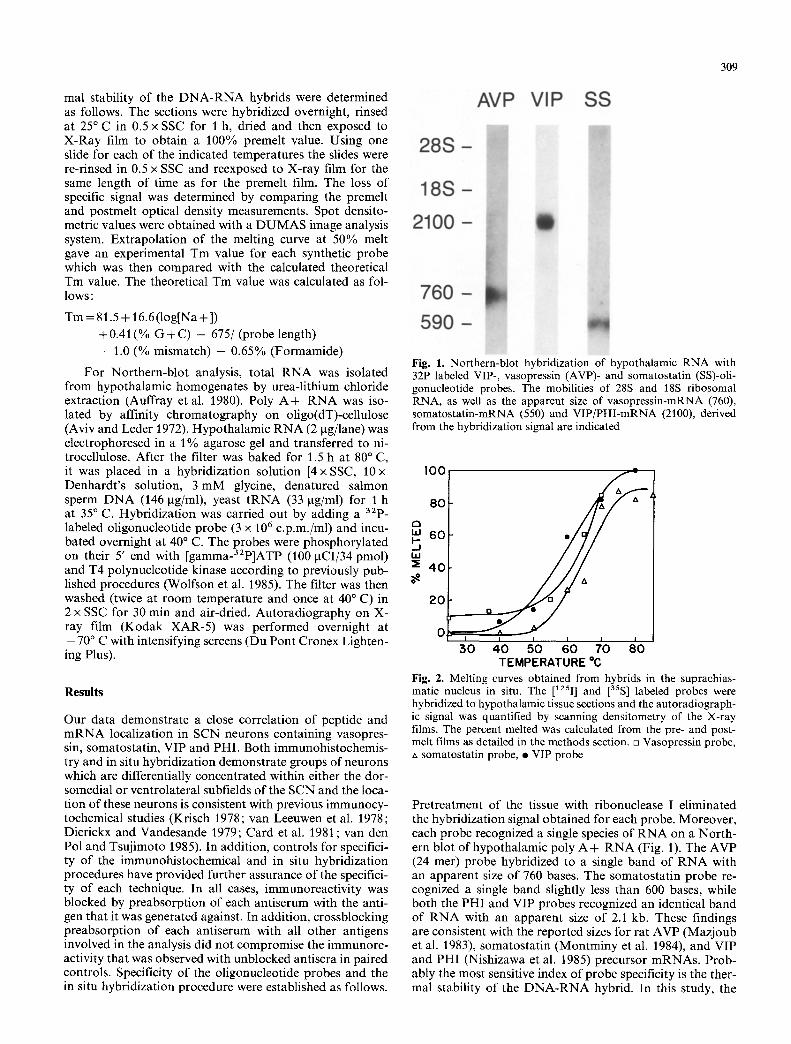
Fig. 1. Northern-blot hybridization of hypothalamic RNA with 32P labeled VIP-, vasopressin (AVP)- and somatostatin (SS)-oli- gonucleotide probes. The mobilities of 28S and 18S ribosomal RNA, as well as the apparent size of vasopressin-mRNA (760), somatostatin-mRNA (550) and VIP/PHI-mRNA (2100), derived from the hybridization signal are indicated
I 0 0
80
' " 60 I - ..J w
4 0
20 I
O, I
3O I I I I I
4 0 50 60 70 80 TEMPERATURE *C
Fig. 2. Melting curves obtained from hybrids in the suprachias- matic nucleus in situ. The [x2~I] and [35S] labeled probes were hybridized to hypothalamic tissue sections and the autoradiograph- ic signal was quantified by scanning densitometry of the X-ray films. The percent melted was calculated from the pre- and post- melt films as detailed in the methods section. [] Vasopressin probe, zx somatostatin probe, �9 VIP probe
Pretreatment of the tissue with ribonuclease I eliminated the hybridization signal obtained for each probe. Moreover, each probe recognized a single species o f R N A on a Nor th- ern blot of hypothalamic poly A + R N A (Fig. 1). The AVP (24 mer) probe hybridized to a single band of R N A with an apparent size o f 760 bases. The somatostatin probe re- cognized a single band slightly less than 600 bases, while both the PHI and VIP probes recognized an identical band of R N A with an apparent size of 2.1 kb. These findings are consistent with the reported sizes for rat AVP (Mazjoub et al. 1983), somatostatin (Montminy et al. 1984), and VIP and PHI (Nishizawa et al. 1985) precursor mRNAs. Prob- ably the most sensitive index of probe specificity is the ther- mal stability o f the D N A - R N A hybrid. In this study, the
310
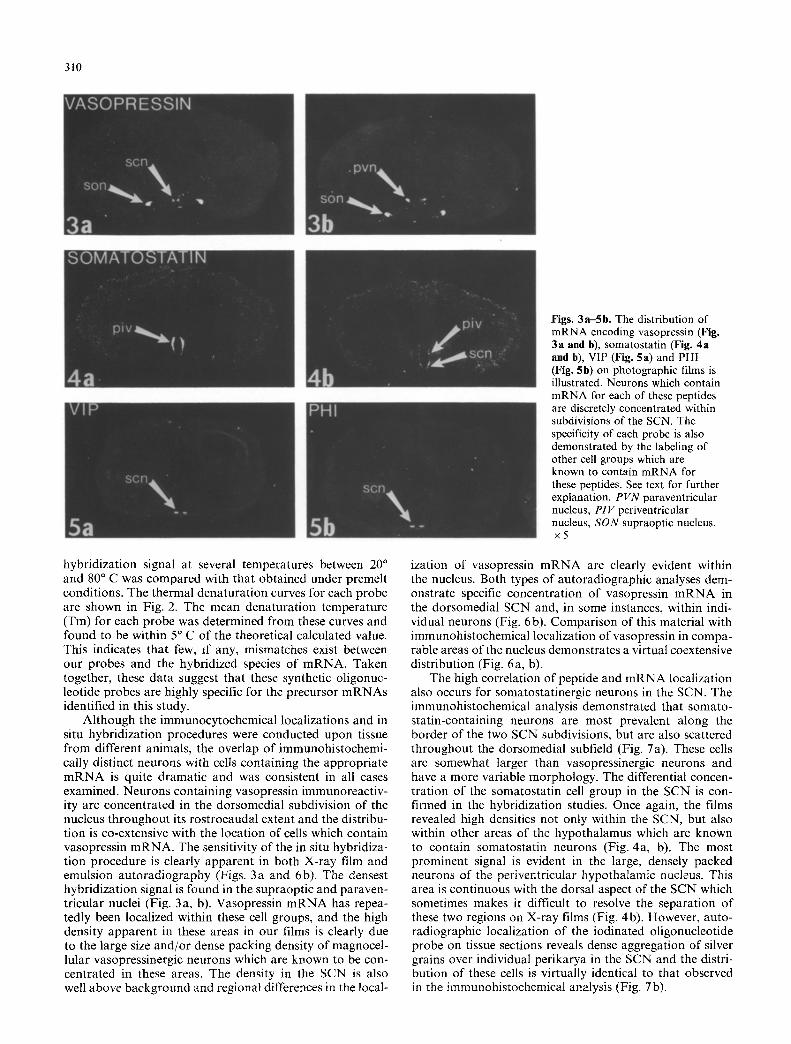
Figs. 3a-Sb. The distribution of mRNA encoding vasopressin (Fig. 3a and b), somatostatin (Fig. 4a and b), V1P (Fig. 5a) and PHI (Fig. 5b) on photographic films is illustrated. Neurons which contain mRNA for each of these peptides are discretely concentrated within subdivisions of the SCN. The specificity of each probe is also demonstrated by the labeling of other cell groups which are known to contain mRNA for these peptides. See text for further explanation. P V N paraventricular nucleus, PIV periventricular nucleus, S O N supraoptic nucleus. x5
hybridization signal at several temperatures between 20 ~ and 80 ~ C was compared with that obtained under premelt conditions. The thermal denaturation curves for each probe are shown in Fig. 2. The mean denaturation temperature (Tin) for each probe was determined from these curves and found to be within 5 ~ C of the theoretical calculated value. This indicates that few, if any, mismatches exist between our probes and the hybridized species of mRNA. Taken together, these data suggest that these synthetic oligonuc- leotide probes are highly specific for the precursor m R N A s identified in this study.
Although the immunocytochemical localizations and in situ hybridization procedures were conducted upon tissue from different animals, the overlap of immunohistochemi- cally distinct neurons with cells containing the appropriate m R N A is quite dramatic and was consistent in all cases examined. Neurons containing vasopressin immunoreactiv- ity are concentrated in the dorsomedial subdivision of the nucleus throughout its rostrocaudal extent and the distribu- tion is co-extensive with the location of cells which contain vasopressin m R N A . The sensitivity of the in situ hybridiza- tion procedure is clearly apparent in both X-ray film and emulsion autoradiography (Figs. 3a and 6b). The densest hybridization signal is found in the supraoptic and paraven- tricular nuclei (Fig. 3a, b). Vasopressin m R N A has repea- tedly been localized within these cell groups, and the high density apparent in these areas in our films is clearly due to the large size and/or dense packing density of magnocel- lular vasopressinergic neurons which are known to be con- centrated in these areas. The density in the SCN is also well above background and regional differences in the local-
ization of vasopressin m R N A are clearly evident within the nucleus. Both types of autoradiographic analyses dem- onstrate specific concentration of vasopressin m R N A in the dorsomedial SCN and, in some instances, within indi- vidual neurons (Fig. 6b). Comparison of this material with immunohistochemical localization of vasopressin in compa- rable areas o f the nucleus demonstrates a virtual coextensive distribution (Fig. 6a, b).
The high correlation of peptide and m R N A localization also occurs for somatostatinergic neurons in the SCN. The immunohistochemical analysis demonstrated that somato- statin-containing neurons are most prevalent along the border of the two SCN subdivisions, but are also scattered throughout the dorsomedial subfield (Fig. 7a). These cells are somewhat larger than vasopressinergic neurons and have a more variable morphology. The differential concen- tration of the somatostatin cell group in the SCN is con- firmed in the hybridization studies. Once again, the films revealed high densities not only within the SCN, but also within other areas of the hypothalamus which are known to contain somatostatin neurons (Fig. 4a, b). The most prominent signal is evident in the large, densely packed neurons o f the periventricular hypothalamic nucleus. This area is continuous with the dorsal aspect of the SCN which sometimes makes it difficult to resolve the separation of these two regions on X-ray films (Fig. 4b). However, auto- radiographic localization of the iodinated oligonucleotide probe on tissue sections reveals dense aggregation of silver grains over individual perikarya in the SCN and the distri- bution of these cells is virtually identical to that observed in the immunohistochemical analysis (Fig. 7b).
311
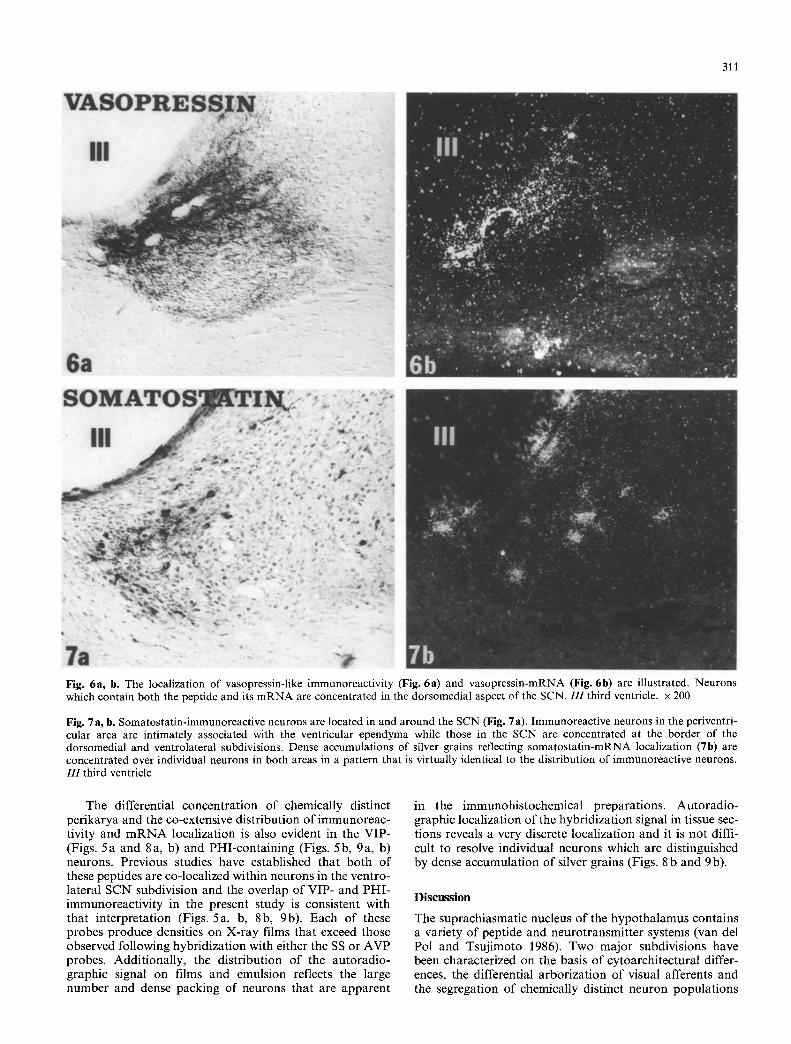
Fig. 6a, b. The localization of vasopressin-like immunoreactivity (Fig. 6a) and vasopressin-mRNA (Fig. 6b) are illustrated. Neurons which contain both the peptide and its mRNA are concentrated in the dorsomedial aspect of the SCN. 111 third ventricle, x 200
Fig. 7a, b. Somatostatin-immunoreactive neurons are located in and around the SCN (Fig. 7a). Immunoreactive neurons in the periventri- cular area are intimately associated with the ventricular ependyma while those in the SCN are concentrated at the border of the dorsomedial and ventrolateral subdivisions. Dense accumulations of silver grains reflecting somatostatin-mRNA localization (7b) are concentrated over individual neurons in both areas in a pattern that is virtually identical to the distribution of immunoreactive neurons. I I I third ventricle
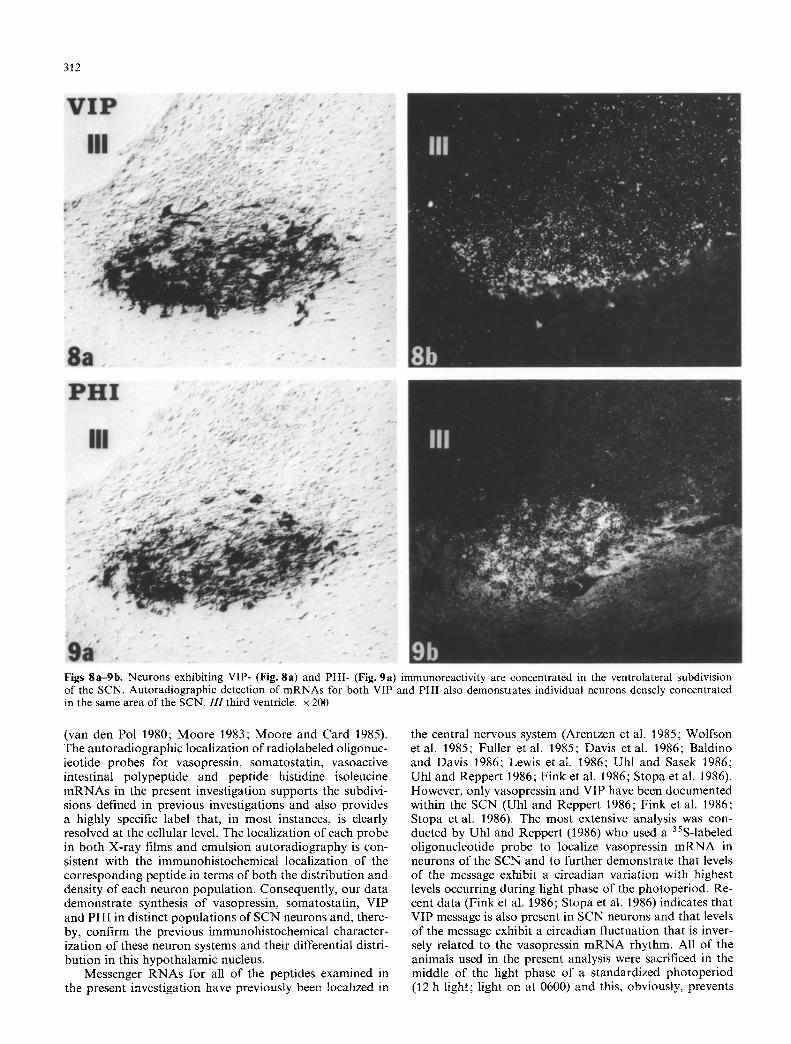
The differential concentrat ion of chemically distinct per ikarya and the co-extensive dis tr ibut ion of immunoreac- tivity and m R N A localizat ion is also evident in the VIP- (Figs. 5a and 8a, b) and PHI-conta in ing (Figs. 5b, 9a, b) neurons. Previous studies have established that both of these peptides are co-localized within neurons in the ventro- lateral SCN subdivision and the overlap of VIP- and PHI- immunoreact ivi ty in the present s tudy is consistent with that interpretat ion (Figs. 5a, b, 8b, 9b). Each o f these probes produce densities on X-ray films that exceed those observed following hybr idizat ion with either the SS or AVP probes. Addi t ional ly , the dis tr ibut ion of the au torad io- graphic signal on films and emulsion reflects the large number and dense packing o f neurons that are apparent
in the immunohis tochemical preparat ions. Au to rad io - graphic local izat ion of the hybr idizat ion signal in tissue sec- tions reveals a very discrete local izat ion and it is not diffi- cult to resolve individual neurons which are dist inguished by dense accumulat ion of silver grains (Figs. 8 b and 9 b).
Discussion
The suprachiasmat ic nucleus of the hypotha lamus contains a variety of pept ide and neurot ransmit ter systems (van del Pol and Tsuj imoto 1986). Two major subdivisions have been characterized on the basis o f cytoarchi tectural differ- ences, the differential a rbor iza t ion of visual afferents and the segregation of chemically distinct neuron popula t ions
312
Figs 8a-gb. Neurons exhibiting VIP- (Fig. 8a) and PHI- (Fig. 9a) immunoreactivity are concentrated in the ventrolateral subdivision of the SCN. Autoradiographic detection of mRNAs for both VIP and PHI also demonstrates individual neurons densely concentrated in the same area of the SCN. 111 third ventricle, x 200
(van den Pol 1980; Moore 1983; Moore and Card 1985). The autoradiographic localization of radiolabeled oligonuc- leotide probes for vasopressin, somatostatin, vasoactive intestinal polypeptide and peptide histidine isoleucine mRNAs in the present investigation supports the subdivi- sions defined in previous investigations and also provides a highly specific label that, in most instances, is clearly resolved at the cellular level. The localization of each probe in both X-ray films and emulsion autoradiography is con- sistent with the immunohistochemical localization of the corresponding peptide in terms of both the distribution and density of each neuron population. Consequently, our data demonstrate synthesis of vasopressin, somatostatin, VIP and PHI in distinct populations of SCN neurons and, there- by, confirm the previous immunohistochemical character- ization of these neuron systems and their differential distri- bution in this hypothalamic nucleus.
Messenger RNAs for all of the peptides examined in the present investigation have previously been localized in
the central nervous system (Arentzen et al. 1985; Wolfson et al. 1985; Fuller et al. 1985; Davis et al. 1986; Baldino and Davis 1986; Lewis et al. 1986; Uhl and Sasek 1986; Uhl and Reppert 1986; Fink et al. 1986; Stopa et al. 1986). However, only vasopressin and VIP have been documented within the SCN (Uhl and Reppert 1986; Fink et al. 1986; Stopa et al. 1986). The most extensive analysis was con- ducted by Uhl and Reppert (1986) who used a 35S-labeled oligonucleotide probe to localize vasopressin mRNA in neurons of the SCN and to further demonstrate that levels of the message exhibit a circadian variation with highest levels occurring during light phase of the photoperiod. Re- cent data (Fink et al. 1986; Stopa et al. 1986) indicates that VIP message is also present in SCN neurons and that levels of the message exhibit a circadian fluctuation that is inver- sely related to the vasopressin mRNA rhythm. All of the animals used in the present analysis were sacrificed in the middle of the light phase of a standardized photoperiod (12 h light; light on at 0600) and this, obviously, prevents

313
us from identifying any circadian fluctuations of message. However, our findings are consistent with those of Uhl and Reppert (1986) in that vasopressin message was clearly distinguishable during the light phase of the photoperiod. Interestingly, our data also demonstrate that the VIP and PHI message are present at high levels and this would ap- pear to conflict with the marginally detectable low levels of VIP message that Stopa and colleagues (1986) reported during the light phase. We are presently conducting quanti- tative analyses of the levels of VIP-, PHI-, somatostatin- and vasopressin-mRNA within the SCN during different phases of the photoperiod in an effort to verify previous findings concerning vasopressin (Uhl and Reppert 1986) and VIP (Stopa et al. 1986) and also to determine if levels of PHI- and somatostatin-mRNA exhibit circadian varia- tions.
One of the primary attributes of the present investiga- tion is the sensitivity of the technique and the resolution of the label with respect to individual SCN neurons. Both the X-ray films and the emulsion autoradiography demon- strate message distribution that is essentially coextensive with the location of the neurons exhibiting immunoreactivi- ty to antisera generated against the appropriate peptide. Further evidence that the oligonucleotide probes used in the present study are highly specific for each species of mRNA detected in the SCN are demonstrated by the melt- ing profiles of the DNA-RNA hybrids, the failure of each probe to hybridize to tissue sections pretreated with ribonu- clease I, and the ability of each probe to recognize a discrete band of poly A § RNA on Northern blots. In addition, all of the probes identified other neurons in the central nervous system which have previously been shown to con- tain the appropriate antigen. This was most apparent in examining the hypothalamic localization of the vasopressin and somatostatin oligonucleotide probes. In addition to the SCN, the vasopressin probe also identified magnocellular neurons in the paraventricular and supraoptic nuclei as well as individual neurons in and around the nucleus circularis. The abundance of vasopressin-mRNA in these large neu- rons was clearly reflected as a dense, cellular accumulation of silver grains which was easily distinguished from back- ground. Similarly, the somatostatin probe characteristically labeled the group of large, densely packed somatostatin- ergic neurons in the periventricular nucleus which have pre- viously been identified in both immunohistochemical (cf. Johansson et al. 1984) and in situ hybridization (Arentzen et al. 1985; Uhl and Reppert 1986) analyses. Taken together with the fact that the VIP and PHI probes also identified other CNS neurons previously shown to contain these pep- tides, the above observations provide clear evidence of the specificity of our oligonucleotide labeling in the SCN.
Delineation of the nucleotide sequence of the neuropep- tides in the SCN has raised interesting questions regarding their posttranslational processing. The rat VIP precursor contains the sequences of both VIP and PHI-27, and has post-translational processing sites for the generation of these two peptides (Nishizawa et al. 1985). In comparison with the human VIP and PHM-27 nucleotide sequence (Bodner et al. 1985), the rat sequence is approximately 80% homologous with the human sequence in both the VIP and PHI-27 domain. The rat PHI-27 differs from human PHM- 27 by only 4 amino acids. These findings suggest that the coding regions for rat and human VIP precursor are highly conserved. Although PHI-27 and VIP immunoreactive ma-
terial has been largely localized within the same neurons in the ventrolateral subdivision of the SCN (Moore et al. 1985; Daikoku et al. 1985), it has been suggested that the complexity of RNA processing, especially in RNA's with two discontinuous exon segments, could favor alternate processing events similar to those reported for the calcitonin gene (Rosenfeld et al. 1983). However, to our knowledge, there has been only one report of differential localization of VIP and PHI-27 in the rat CNS (Hrkfelt et al. 1983). Our data does not allow us to conclude that mRNAs for VIP and PHI occur in the same SCN neurons. However, the co-extensive distribution of both mRNAs combined with the demonstrated co-localization of both peptides (Moore et al. 1985; Daikoku et al. 1985) in SCN neurons would argue for this conclusion, and thereby indicate that the exon coding regions for both VIP and PHI are tran- scribed in these neurons. This would also suggest that alter- nate processing of the two products of the precursor mRNA does not occur during the light phase of the photoperiod.
The somatostatin peptides are produced in a variety of tissues and exhibit an extremely wide distribution within the CNS. Three forms of somatostatin (somatostatin-28, somatostatin-14 and somatostatin-28 (1-12) have been shown to possess biological activity (Brazeau et al. 1981). Probable cleavage sites have also been identified within the pre-prosomatostatin molecule which would suggest that each form of somatostatin is processed post-translationally from a single exon encoding the somatostatin precursor (Benoit et al. 1982). The distribution of specific transcripts encoding the somatostatin precursor as well as immunore- active material for this peptide within the rat hypothalamus, neocortex, pyriform cortex and amygdala has previously been reported (Arentzen et al. 1985; Uhl and Sasek 1986). However, somatostatin m R N A has not previously been lo- calized in neurons of the SCN. In the present investigation, primary transcripts coding for pre-prosomatostatin were identified within neurons concentrated at the border of the two SCN subfields. Immunohistochemical studies (van den Pol and Tsujimoto 1985; the present analysis) have utilized antisera directed against the C-terminus of the somatosta- tin-28 molecule to identify neurons in the same portion of the nucleus. Nevertheless, the extent to which prosoma- tostatin is post-translationally processed in these neurons remains unknown.
The clear role that the suprachiasmatic nuclei play in the regulation of circadian rhythms of behavior and hor- monal secretion has been the subject of extensive analysis (cf. Rusak and Zucker 1979; Rosenwaser and Adler 1986, for reviews). The existence of two distinct subdivisions of the nucleus has been established by a number of different criteria and is supported by this study which demonstrates the synthesis of four different peptides within the SCN. Furthermore, we demonstrate for the first time that PHI- m R N A is present within the ventrolateral subdivision of the SCN and is codistributed with VIP-mRNA. These data indicate that both PHI-27 and VIP are synthesized from the same precursor mRNA in these neurons. The initial demonstration that SCN neurons exhibit a circadian rhythm in metabolic activity as measured by uptake of 2- deoxyglucose provided important insights into the role of the SCN in the regulation of circadian rhythms (Schwartz and Gainer 1977; Schwartz et al. 1980). Consequently, the ability to assess the activity of subpopulations of SCN neu- rons through the measurement of m R N A synthesis, as re-

314
cent ly r epor t ed for vasopress in (Uh l and Reppe r t 1986) and VIP (S topa et al. 1986) should p rov ide even grea ter insight in to the means in which i n f o r m a t i o n is processed by S C N neurons in the regula t ion and en t r a inmen t o f these rhythms.
Acknowledgments. We gratefully acknowledge the expert technical assistance of Terry O'Kane and the secretarial work of Sherry Vari.
References
Arentzen R, Baldino F Jr, Davis LG, Higgins GA, Lin Y, Manning RW, Wolfson B (1985) In situ hybridization of putative soma- tostatin m R N A within hypothalamus of the rat using synthetic oligonucleotide probes. J Cell Biochem 27:415-422
Auffray C, Nugeotte R, Chambraud B, Rougeow F (1980) Muse immunoglobulin genes: a bacterial plasmid containing the en- tire coding sequence for a pre-2A heavy chain. Nucleic Acids Res8:1231 1235
Aviv H, Leder P (1972) Purification of biologically active globin messenger R N A by chromatography on oligo thymidylic acid- cellulose. PNAS USA 69:1408-1412
Baldino F Jr, Davis LG (1986) Glucocorticoid regulation of vaso- pressin messenger RNA. In: Uhl G (ed) In situ Hybridization in Brain. Plenum Press, New York, pp 97 116
Benoit R, Ling N, Alford B, Guillemin R (1982) Seven peptides derived from pro-somatostatin in rat brain. Biochem Biophys Res Comm 107 : 944-950
Bodner M, Fridkin M, Gozes I (1985) Coding sequences for va- soactive intestinal polypeptide and PHM-27 peptide are located on two adjacent exons in the human genome. PNAS USA 82:3548-3551
Brazeau P, Ling M, Bech F, Bohlen P, Benert R, Guillemin R (1981) High biological activity of the synthetic replicates soma- tostatin-28 and somatostatin-25. Regul Pept 1 : 255
Card JP, Moore RY (1982) Ventral lateral geniculate nucleus effer- ents to the rat suprachiasmatic nucleus exhibit avian pancreatic polypeptide-like immunoreactivity. J Comp Neurol 206:390-396
Card JP, Brecha N, Karten HJ, Moore RY (1981) Immunocyto- chemical localization of vasoactive intestinal polypeptide con- taining cells and processes in the suprachiasmatic nucleus of the rat: Light and electron microscopic analysis. J Neurosci 1:1289-1303
Daikoku S, Okamura Y, Kawano H, Tsuruo Y, Maegawa M, Shibasaki Y (1985) CRF-containing neurons of the rat hypo- thamus. Cell Tissue Res 240:575-584
Davis LG, Arentzen R, Reid JM, Manning RW, Wolfson B, Law- rence KL, Baldino F Jr (1986) Glucocorticoid sensitivity of vasopressin m R N A levels in the paraventricular nucleus of the rat. PNAS USA 83 : 1145-1149
Dierickx K, Vandesande F (1979) Immunocytochemical localiza- tion of somatostatin-containing neurons in the rat hypothala- mus. Cell Tissue Res 201 : 349-359
Fink JS, Montminy MR, Tsukada T, Hoefler H, Specht LA, Le- chan RM, Wolfe H, Mandel G, Goodman RH (1986) In situ hybridization of somatostatin and vasoactive intestinal peptide m R N A in the rat nervous system. Contrasting patterns of onto- geny. In: Uhl GR (ed) In Situ Hybridization in Brain. Plenum, New York, pp 181-191
Fritz H-J, Belagage R, Brown EL, Fritz RH, Jones RA, Lees RG, Khorana HG (1978) High-pressure liquid chromatography in polynucleotide synthesis. Biochemistry 17 : 1257-1267
Fuller P J, Clements JA, Funder JW (1985) Localization of arginine vasopressin-neurophysin II messenger ribonucleic acid in the hypothalamus of control and Brattleboro rats by hybridization histochemistry with a synthetic pentadecamer oligonucleotide probe. Endocrinology 116 �9 2366-2368
Hendrickson AE, Wagoner N, Cowan WM (1972) An autoradio-
graphic and electron microscopic study of retinohypothalamic connections. Z Zellforsch Mikrosk Anat 135:1-26
H6kfelt T, Fahrenkrug J, Tatemoto K, Mutt V, Werner N, Hulting AL, Tereniu L, Cheng KJ (1983) The PHI (PHI-27)/corticotro- pin releasing factor/enkephalin immunoreactive neuron: possi- ble morphological basis for integrated control of prolactin, cor- ticotropin and growth hormone secretion. PNAS USA 80: 895-898
Hsu S, Raine L, Fanger H (1981) Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: A compari- son between ABC and unlabeled antibody (PAP) procedures. J Histochem Cytochem 29:577-580
Itoh N, Obata K-I, Yanadihara N, Okamoto H (1983) Human preprovasoactive intestinal polypeptide contains a novel PHI- 27-like peptide, PHM-27. Nature 304: 547-549
Johansson O, H6kfelt T, Elde RP (1984) Immunohistochemical distribution of somatostatin-like immunoreactivity in the cen- tral nervous system of the adult rat. Neuroscience 13:265-339
Krisch B (1978) Light- and electron microscopic localization of vasopressin or a vasopressin-like substance in the neurons of the rat suprachiasmatic nucleus. Cell Tissue Res 194:361-365
Leeuwen FW van, Swaab DF, de Raay C (1978) Immunoelectron microscopic localization of vasopressin in the rat suprachias- matic nucleus. Cell Tissue Res 193 : 1 10
Lewis ME, Arentzen R, Baldino F Jr (1986) Rapid, high-resolution in situ hybridization histochemistry with radioiodinated syn- thetic oligonucleotides. J Neurosci Res 16:11%124
Matteucci MD, Caruthers MH (1981) Synthesis of deoxynucleo- tides on a polymer support. J Am Chem Soc 103:3185 3191
Mazjoub JA, Rich A, van Bloom J, Hadener JF (1983) Vasopressin and oxytocin mRNA regulation in the rat assessed by hybrid- ization with synthetic oligonucleotides. J Biol Chem 258:14061 14064
McLean IW, Nakane PK (1974) Periodate-lysine-paraformalde- hyde fixative. A new fixative for immunoelectron microscopy. J Histochem Cytochem 22:1077 1083
Montiminy MR, Goodman RH, Horovitch S J, Habener JF (1984) Primary structure of the gene encoding rat preprosomatostatin. PNAS USA 81:3337-3340
Moore RY (1983) Organization and function of a central nervous system circadian oscillator: the suprachiasmatic nucleus. Fed Proc 42 : 2783-2789
Moore RY, Card JP (1985) Visual pathways and the entrainment of circadian rhythms. In: Wurtman R J, Baum M J, Potts JT (eds) The Medical and Biological Effects of Light. Part II. Indi- rect Effects of Light : Biological Rhythms. Annals of the New York Academy of Sciences 453:123-133
Moore RY, Lenn NJ (1972) A retinohypothalamic projection in the rat. J Comp Neurol 146:1-14
Moore RY, Gustafson EL, Card JP (1984) Identical immunoreac- tivity of afferents to the rat suprachiasmatic nucleus with anti- sera against avian pancreatic polypeptide, molluscan cardioex- citatory peptide and neuropeptide Y. Cell Tissue Res 236:41-46
Moore RY, Speh J, Card JP (1985) Vasoactive intestinal polypep- tide (VIP) and peptide histidine isoleucine (PHI) co-localize in neurons of the rat suprachiasmatic nucleus (SCN). [Abstr] vol 332.4, Society Neurosci Meeting, p 1139
Morel A, Nicolas P, Cohen P (1983) Evidence for a predominant form of M r - 15,000 prosomatostatin in the mouse hypothala- mus. Relationship with somatostatin-14 and -28. J Biol Chem 258 : 8273-8276
Nishizawa M, Hayakawa Y, Yanaihara N, Okamoto H (1985) Nucleotide sequence divergence and functional constraint in VIP precursor mRNA evolution between human and rat. FEBS Lett 183:55-59
Pol AN van den (1980) The hypothalamic suprachiasmatic nucleus of the rat: intrinsic anatomy. J Comp Neurol 191:661-702
Pol AN van den, Powley TL (1979) A fine-grained anatomical analysis of the role of the rat suprachiasmatic nucleus in circa- dian rhythms of feeding and drinking. Brain Res 160: 307 326
Pol AN van den, Tsujimoto KL (1985) Neurotransmitters of the

315
hypothalamic suprachiasmatic nucleus: immunocytochemical analysis of 25 neuronal antigens. Neuroscience 15 : 1049-1086
Rosenfeld MG, Mermod J-J, Amara SG, Swanson LW, Saw- chenko PE, Rivier J, Vale WW, Evans RM (1983) Production of a novel neuropeptide encoded by the calcitonin gene via tissue-specific R N A processing. Nature 304:129-135
Rosenwasser AM, Adler NT (1986) Structure and function in circa- dian timing systems: evidence for multiple coupled circadian oscillators. Neurosci Behav Rev 10:431448
Rusak B, Zucker I (1979) Neural regulation of circadian rhythms. Physiol Rev 59:449-526
Schwartz WJ, Gainer H (1977) Suprachiasmatic nucleus: use of 14C-labeled deoxyglucose uptake as a functional marker. Science 197:1089-1091
Schwartz WJ, Davidsen LC, Smith CB (1980) In vivo metabolic activity of a putative circadian oscillator: the rat suprachias- matic nucleus. J Comp Neurol 189:157-167
Shibata S, Liou SY, Ueki S, Oomura Y (1984) Influence of environ- mental light-dark cycle and enucleation on activity of supra- chiasmatic neurons in slice preparations. Brain Res 302:75- 81
Sternberger LA (1979) The unlabeled antibody peroxidase-antiper- oxidase (PAP) method. In: Cohen S, McCluskey RT (eds) Im- munohistochemistry. Prentice-Hall, New Jersey, pp 104-169
Stopa EG, Jonassen JA, Minamitani N, King JC, Wolfe H, Mob- taker H, Albers HE (1986) m R N A of vasoactive intestinal poly- peptide and peptide histidine-isoleucine within the suprachias- matic nucleus. [Abstr] vol 166.5, Society for Neurosciences Meeting
Uhl GR, Reppert SM (1986) Suprachiasmatic nucleus vasopressin messenger RNA: circadian variation in normal and Brattleboro rats. Science 232 : 309-393
Uhl GR, Sasek CA (1986) Somatostatin mRNA: Regional varia- tion in hybridization densities in individual neurons. J Neurosci 6:3258-3264
Wolfson B, Manning RW, Davis LG, Arentzen R, Baldino F Jr (1985) Co-localization of corticotropin releasing factor and va- sopressin m R N A in neurones after adrenalectomy. Nature 315:59-61
Accepted November 9, 1987





























