Embed Size (px)
Citation preview

Biologia 69/9: 1190—1201, 2014Section ZoologyDOI: 10.2478/s11756-014-0420-6
Littoral macroinvertebrates of acidified lakesin the Bohemian Forest
Lenka Ungermanová1, Kateřina Kolaříková1*, Evžen Stuchlík1, Takaaki Senoo1,Jakub Horecký1, Jiří Kopáček2, Pavel Chvojka3, Jolana Tátosová1, Peter Bitušík4
& Arne Fjellheim5
1Institute for Environmental Studies, Faculty of Science, Charles University, Benátská 2, 128 01 Prague, Czech Republic;e-mail: [email protected] Centre ASCR, Institute of Hydrobiology, Na Sádkách 7, 37005 České Budějovice, Czech Republic3Department of Entomology, National Museum, Kunratice 1, 148 00 Prague, Czech Republic4Faculty of Science, Matthias Belius University, Tajovského 40, 974 01 Banská Bystrica, Slovakia5Uni Research, Uni Environment, Thormohlensgt. 49 B, 5006 Bergen, Norway
Abstract: Mountain lakes in the Bohemian Forest, on both the Czech and German sides, were atmospherically acidifiedmainly in the 1960s–1980s and have since been recovering from acidification. In 2007, we performed the first completestudy on littoral macroinvertebrates in all eight lakes. The goals of the study were to 1) compare macroinvertebrates inthe lakes during the process of recovery and 2) investigate relations between the occurrence of taxa and water chemistry.Lake water pH varied from 4.6 to 5.7, concentrations of dissolved reactive Al and labile Al ranged from 118–601 and 11–470 µg L−1, respectively, and DOC concentrations were < 6 mg L−1. Altogether 73 taxa were identified from all lakes;a positive relationship was found between pH and the number of macroinvertebrate taxa. The highest number of taxawas found in the least acidic lakes Laka and Grosser Arbersee, including the mollusk Pisidium casertanum. In contrast,the lowest diversity was found in the most acidified Čertovo jezero. Cluster analyses of macroinvertebrates and waterchemistry suggested pH as the key factor influencing the occurrence of macroinvertebrate taxa. An interesting finding wasthe occurrence of the boreo-montane water beetle Nebrioporus assimilis in Prášilské jezero, which is the first documentedrecord of this species in the Czech Republic since 1960.
Key words: lakes; macroinvertebrates; acidification; recovery; forest catchment
Abbreviations: ANC acid neutralizing capacity; DOC dissolved organic carbon; L-Al labile aluminium;N-Al non-labilealuminium; R-Al reactive aluminium; TP total phosphorus
Introduction
Anthropogenic acidification has strongly affected sur-face freshwaters in the second half of the 20th century asa result of S and N emissions (Schindler 1988), causingmajor chemical and biological changes in ecosystems.Despite a significant decrease of acidification over thelast decades, especially in North America and Europe,the negative effects on water ecosystems still remain animportant environmental problem. Current global cli-mate change is a confounding factor which can magnifythe negative consequences of acidification or can dis-rupt recovery (Laudon & Westling 2005). Because of adelayed response time, the biological recovery of acidi-fied sites occurs later than chemical restoration (Jeffrieset al. 2003; Posch et al. 2003; Wright et al. 2005).In Central Europe, scientists have devoted much
attention to the acidification of mountain lakes in the
Czech Republic, Slovakia, Poland, and Germany, par-ticularly in the Bohemian Forest and the Tatra Moun-tains (e.g., Vranovský et al. 1994; Kopáček et al. 1998;Kownacki et al. 2000; Kubovčík & Bitušík 2006; Hořickáet al. 2006; Krno et al. 2006; Stuchlík et al. 2006; Fjell-heim et al. 2009). Over the past half-century, thesemountain areas witnessed the most pronounced changesin acidic deposition and lake water composition amongall European lake districts (Evans et al. 2001), due tosteep increases in emission and deposition fluxes of sul-fur and nitrogen compounds in the 1950s–1980s andtheir abrupt reduction after 1989 (Kopáček et al. 2001).The Bohemian Forest lake district (at the border be-tween the Czech Republic and Germany) has currentlyexhibited an apparent chemical recovery from acidi-fication, with increasing water pH and declining Alconcentrations (Majer et al. 2003; Veselý et al. 2003;Oulehle et al. 2012). This process is relatively slow,
* Corresponding author
c©2014 Institute of Zoology, Slovak Academy of Sciences
Littoral macroinvertebrates of acidified lakes 1191
Fig. 1. The location of the Bohemian Forest lakes on the Czech side (Šumava) and the German side (Bohmerwald). Map source:www.sumava.xf.cz (modified)
Table 1. Basic characteristics of the lakes and their catchments (modified after Nedbalová et al. 2006).
CN CT PL PR RA KA LA GA
Latitude N 49◦11′ 49◦10′ 48◦47′ 49◦05′ 48◦58′ 49◦08′ 49◦07′ 49◦06′Longitude E 13◦11′ 13◦12′ 13◦52′ 13◦24′ 13◦24′ 13◦09′ 13◦20′ 13◦07′Elevation m a.s.l. 1008 1027 1087 1079 1071 918 1085 935Lake area ha 18.8 10.7 7.6 4.2 5.7 9.4 2.6 7.7Catchment area km2 1.24 0.89 0.67 0.65 0.58 2.79 1.02 2.58Max. depth m 40 35 19 17 13 9 3 16Lake volume 106m3 2.92 1.86 0.61 0.35 0.18 0.25 0.05 0.45Orientation E E NE E SE N NE E
however, compared to the rapid reductions in CentralEuropean emissions of SO2 (86%), NOx (53%), andNH3 (35%), and related acidic deposition in the Bo-hemian Forest over the last two decades (Kopáček etal. 2011). As a consequence, the carbonate bufferingsystem of the most acidified lakes will probably notbe re-established before ∼2050 (Majer et al. 2003).The reason for this delay is continuing leaching ofsulfate and nitrate from the lake catchments, and isalso dependent on soil characteristics, bedrock geology,and vegetation vigor (Kopáček et al. 2002). Changesin lake biota have been even more delayed than thechemical recovery of the Bohemian Forest lakes; nev-ertheless the first signs of zooplankton and macro-zoobenthos recovery have already been documented(Vrba et al. 2003; Nedbalová et al. 2006; Soldán et al.2012).The main aim of this study was to evaluate the
present distribution of macroinvertebrates in the lit-toral of the Bohemian Forest lakes in relation to lakewater chemistry. Furthermore, these are the first datafrom the all eight Bohemian Forest lakes obtained us-ing a homogenous sampling method, and should serveas a basis for future comparisons and assessments of theongoing process of recovery.
Material and methods
Study sitesThe study was performed in eight lakes of glacial originsituated in the Bohemian Forest, both in its Czech part(Šumava): Černé jezero (CN), Čertovo jezero (CT), Plešnéjezero (PL), Prášilské jezero (PR), and Laka (LA), and Ger-man part (Bohmerwald): Rachelsee (RA), Grosser Arbersee(GA), and Kleiner Arbersee (KA) (Fig. 1). Nearly all lakeshave rocky bottoms in the littoral, except for LA and KAwhere the bottom is formed by a layer of peat. The ba-sic characteristics of the lakes and their catchments are de-scribed in Table 1. The bedrock (schist, granite, and gneiss)in the lake catchments is sensitive to acid atmospheric de-position and magnified the effects of acidification on theseecosystems in the 20th century (Veselý 1994). Lake catch-ments are mainly formed of steep slopes predominantly cov-ered by shallow acidic soils, mostly podzols and cambisols.The forests around the lakes are dominated mainly by ma-ture Norway spruce (generally 80 to 150-year-old) mixedwith fir and beech (Veselý 1994; Vrba et al. 2003). All theBohemian Forest lakes are fishless at present, naturally oras a consequence of acidification (Vrba et al. 2003).
Sampling and analytical methodsSampling of water and benthic macroinvertebrates was car-ried out in spring (May) 2007 in the Czech lakes and inautumn (September – October) 2007 in all lakes.
1192 L. Ungermanová et al.
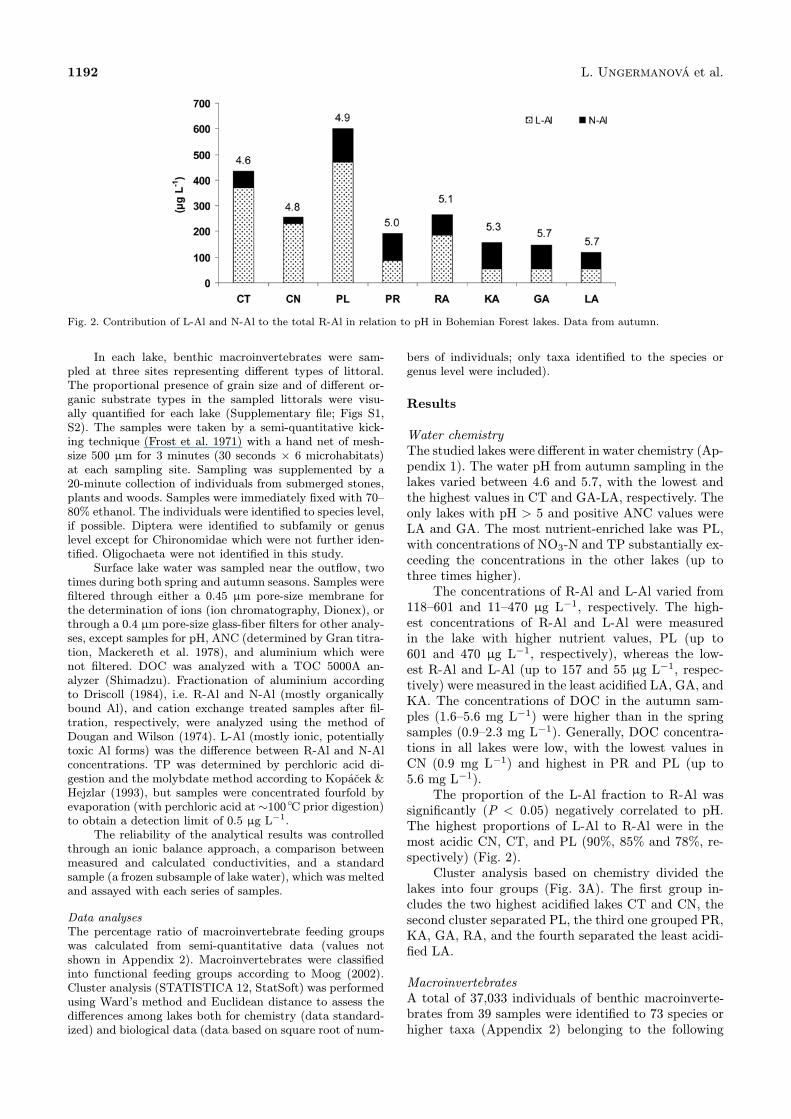
Fig. 2. Contribution of L-Al and N-Al to the total R-Al in relation to pH in Bohemian Forest lakes. Data from autumn.
In each lake, benthic macroinvertebrates were sam-pled at three sites representing different types of littoral.The proportional presence of grain size and of different or-ganic substrate types in the sampled littorals were visu-ally quantified for each lake (Supplementary file; Figs S1,S2). The samples were taken by a semi-quantitative kick-ing technique (Frost et al. 1971) with a hand net of mesh-size 500 µm for 3 minutes (30 seconds × 6 microhabitats)at each sampling site. Sampling was supplemented by a20-minute collection of individuals from submerged stones,plants and woods. Samples were immediately fixed with 70–80% ethanol. The individuals were identified to species level,if possible. Diptera were identified to subfamily or genuslevel except for Chironomidae which were not further iden-tified. Oligochaeta were not identified in this study.
Surface lake water was sampled near the outflow, twotimes during both spring and autumn seasons. Samples werefiltered through either a 0.45 µm pore-size membrane forthe determination of ions (ion chromatography, Dionex), orthrough a 0.4 µm pore-size glass-fiber filters for other analy-ses, except samples for pH, ANC (determined by Gran titra-tion, Mackereth et al. 1978), and aluminium which werenot filtered. DOC was analyzed with a TOC 5000A an-alyzer (Shimadzu). Fractionation of aluminium accordingto Driscoll (1984), i.e. R-Al and N-Al (mostly organicallybound Al), and cation exchange treated samples after fil-tration, respectively, were analyzed using the method ofDougan and Wilson (1974). L-Al (mostly ionic, potentiallytoxic Al forms) was the difference between R-Al and N-Alconcentrations. TP was determined by perchloric acid di-gestion and the molybdate method according to Kopáček &Hejzlar (1993), but samples were concentrated fourfold byevaporation (with perchloric acid at∼100◦C prior digestion)to obtain a detection limit of 0.5 µg L−1.
The reliability of the analytical results was controlledthrough an ionic balance approach, a comparison betweenmeasured and calculated conductivities, and a standardsample (a frozen subsample of lake water), which was meltedand assayed with each series of samples.
Data analysesThe percentage ratio of macroinvertebrate feeding groupswas calculated from semi-quantitative data (values notshown in Appendix 2). Macroinvertebrates were classifiedinto functional feeding groups according to Moog (2002).Cluster analysis (STATISTICA 12, StatSoft) was performedusing Ward’s method and Euclidean distance to assess thedifferences among lakes both for chemistry (data standard-ized) and biological data (data based on square root of num-
bers of individuals; only taxa identified to the species orgenus level were included).
Results
Water chemistryThe studied lakes were different in water chemistry (Ap-pendix 1). The water pH from autumn sampling in thelakes varied between 4.6 and 5.7, with the lowest andthe highest values in CT and GA-LA, respectively. Theonly lakes with pH > 5 and positive ANC values wereLA and GA. The most nutrient-enriched lake was PL,with concentrations of NO3-N and TP substantially ex-ceeding the concentrations in the other lakes (up tothree times higher).The concentrations of R-Al and L-Al varied from
118–601 and 11–470 µg L−1, respectively. The high-est concentrations of R-Al and L-Al were measuredin the lake with higher nutrient values, PL (up to601 and 470 µg L−1, respectively), whereas the low-est R-Al and L-Al (up to 157 and 55 µg L−1, respec-tively) were measured in the least acidified LA, GA, andKA. The concentrations of DOC in the autumn sam-ples (1.6–5.6 mg L−1) were higher than in the springsamples (0.9–2.3 mg L−1). Generally, DOC concentra-tions in all lakes were low, with the lowest values inCN (0.9 mg L−1) and highest in PR and PL (up to5.6 mg L−1).The proportion of the L-Al fraction to R-Al was
significantly (P < 0.05) negatively correlated to pH.The highest proportions of L-Al to R-Al were in themost acidic CN, CT, and PL (90%, 85% and 78%, re-spectively) (Fig. 2).Cluster analysis based on chemistry divided the
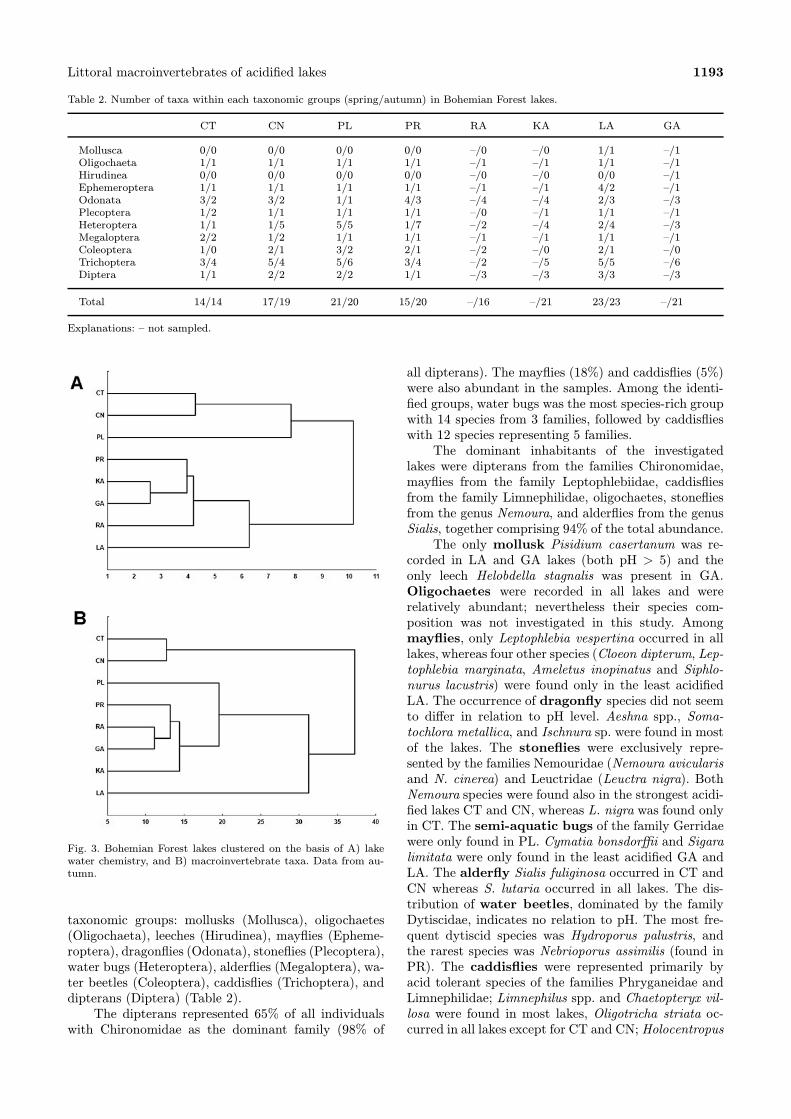
lakes into four groups (Fig. 3A). The first group in-cludes the two highest acidified lakes CT and CN, thesecond cluster separated PL, the third one grouped PR,KA, GA, RA, and the fourth separated the least acidi-fied LA.
MacroinvertebratesA total of 37,033 individuals of benthic macroinverte-brates from 39 samples were identified to 73 species orhigher taxa (Appendix 2) belonging to the following
Littoral macroinvertebrates of acidified lakes 1193
Table 2. Number of taxa within each taxonomic groups (spring/autumn) in Bohemian Forest lakes.
CT CN PL PR RA KA LA GA
Mollusca 0/0 0/0 0/0 0/0 –/0 –/0 1/1 –/1Oligochaeta 1/1 1/1 1/1 1/1 –/1 –/1 1/1 –/1Hirudinea 0/0 0/0 0/0 0/0 –/0 –/0 0/0 –/1Ephemeroptera 1/1 1/1 1/1 1/1 –/1 –/1 4/2 –/1Odonata 3/2 3/2 1/1 4/3 –/4 –/4 2/3 –/3Plecoptera 1/2 1/1 1/1 1/1 –/0 –/1 1/1 –/1Heteroptera 1/1 1/5 5/5 1/7 –/2 –/4 2/4 –/3Megaloptera 2/2 1/2 1/1 1/1 –/1 –/1 1/1 –/1Coleoptera 1/0 2/1 3/2 2/1 –/2 –/0 2/1 –/0Trichoptera 3/4 5/4 5/6 3/4 –/2 –/5 5/5 –/6Diptera 1/1 2/2 2/2 1/1 –/3 –/3 3/3 –/3
Total 14/14 17/19 21/20 15/20 –/16 –/21 23/23 –/21
Explanations: – not sampled.
Fig. 3. Bohemian Forest lakes clustered on the basis of A) lakewater chemistry, and B) macroinvertebrate taxa. Data from au-tumn.
taxonomic groups: mollusks (Mollusca), oligochaetes(Oligochaeta), leeches (Hirudinea), mayflies (Epheme-roptera), dragonflies (Odonata), stoneflies (Plecoptera),water bugs (Heteroptera), alderflies (Megaloptera), wa-ter beetles (Coleoptera), caddisflies (Trichoptera), anddipterans (Diptera) (Table 2).The dipterans represented 65% of all individuals
with Chironomidae as the dominant family (98% of
all dipterans). The mayflies (18%) and caddisflies (5%)were also abundant in the samples. Among the identi-fied groups, water bugs was the most species-rich groupwith 14 species from 3 families, followed by caddisflieswith 12 species representing 5 families.The dominant inhabitants of the investigated
lakes were dipterans from the families Chironomidae,mayflies from the family Leptophlebiidae, caddisfliesfrom the family Limnephilidae, oligochaetes, stonefliesfrom the genus Nemoura, and alderflies from the genusSialis, together comprising 94% of the total abundance.The only mollusk Pisidium casertanum was re-
corded in LA and GA lakes (both pH > 5) and theonly leech Helobdella stagnalis was present in GA.Oligochaetes were recorded in all lakes and wererelatively abundant; nevertheless their species com-position was not investigated in this study. Amongmayflies, only Leptophlebia vespertina occurred in alllakes, whereas four other species (Cloeon dipterum, Lep-tophlebia marginata, Ameletus inopinatus and Siphlo-nurus lacustris) were found only in the least acidifiedLA. The occurrence of dragonfly species did not seemto differ in relation to pH level. Aeshna spp., Soma-tochlora metallica, and Ischnura sp. were found in mostof the lakes. The stoneflies were exclusively repre-sented by the families Nemouridae (Nemoura avicularisand N. cinerea) and Leuctridae (Leuctra nigra). BothNemoura species were found also in the strongest acidi-fied lakes CT and CN, whereas L. nigra was found onlyin CT. The semi-aquatic bugs of the family Gerridaewere only found in PL. Cymatia bonsdorffii and Sigaralimitata were only found in the least acidified GA andLA. The alderfly Sialis fuliginosa occurred in CT andCN whereas S. lutaria occurred in all lakes. The dis-tribution of water beetles, dominated by the familyDytiscidae, indicates no relation to pH. The most fre-quent dytiscid species was Hydroporus palustris, andthe rarest species was Nebrioporus assimilis (found inPR). The caddisflies were represented primarily byacid tolerant species of the families Phryganeidae andLimnephilidae; Limnephilus spp. and Chaetopteryx vil-losa were found in most lakes, Oligotricha striata oc-curred in all lakes except for CT and CN; Holocentropus
1194 L. Ungermanová et al.
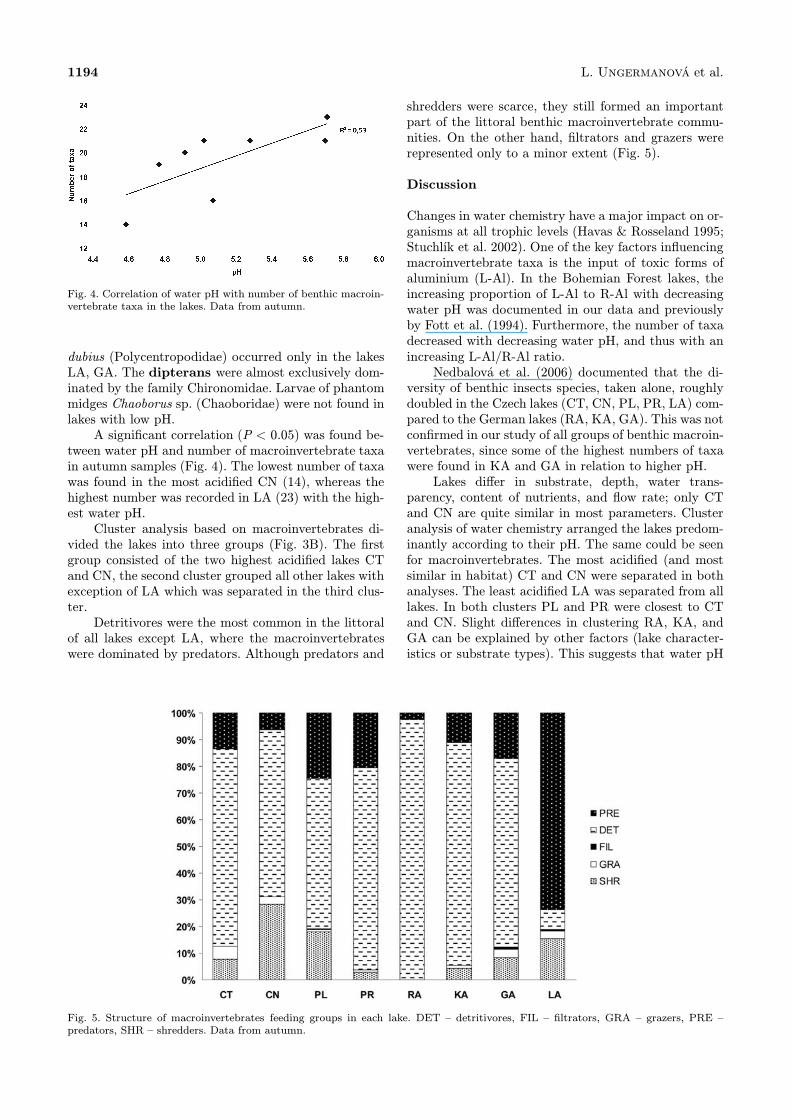
Fig. 4. Correlation of water pH with number of benthic macroin-vertebrate taxa in the lakes. Data from autumn.
dubius (Polycentropodidae) occurred only in the lakesLA, GA. The dipterans were almost exclusively dom-inated by the family Chironomidae. Larvae of phantommidges Chaoborus sp. (Chaoboridae) were not found inlakes with low pH.A significant correlation (P < 0.05) was found be-
tween water pH and number of macroinvertebrate taxain autumn samples (Fig. 4). The lowest number of taxawas found in the most acidified CN (14), whereas thehighest number was recorded in LA (23) with the high-est water pH.Cluster analysis based on macroinvertebrates di-
vided the lakes into three groups (Fig. 3B). The firstgroup consisted of the two highest acidified lakes CTand CN, the second cluster grouped all other lakes withexception of LA which was separated in the third clus-ter.Detritivores were the most common in the littoral
of all lakes except LA, where the macroinvertebrateswere dominated by predators. Although predators and
shredders were scarce, they still formed an importantpart of the littoral benthic macroinvertebrate commu-nities. On the other hand, filtrators and grazers wererepresented only to a minor extent (Fig. 5).
Discussion
Changes in water chemistry have a major impact on or-ganisms at all trophic levels (Havas & Rosseland 1995;Stuchlík et al. 2002). One of the key factors influencingmacroinvertebrate taxa is the input of toxic forms ofaluminium (L-Al). In the Bohemian Forest lakes, theincreasing proportion of L-Al to R-Al with decreasingwater pH was documented in our data and previouslyby Fott et al. (1994). Furthermore, the number of taxadecreased with decreasing water pH, and thus with anincreasing L-Al/R-Al ratio.Nedbalová et al. (2006) documented that the di-
versity of benthic insects species, taken alone, roughlydoubled in the Czech lakes (CT, CN, PL, PR, LA) com-pared to the German lakes (RA, KA, GA). This was notconfirmed in our study of all groups of benthic macroin-vertebrates, since some of the highest numbers of taxawere found in KA and GA in relation to higher pH.Lakes differ in substrate, depth, water trans-
parency, content of nutrients, and flow rate; only CTand CN are quite similar in most parameters. Clusteranalysis of water chemistry arranged the lakes predom-inantly according to their pH. The same could be seenfor macroinvertebrates. The most acidified (and mostsimilar in habitat) CT and CN were separated in bothanalyses. The least acidified LA was separated from alllakes. In both clusters PL and PR were closest to CTand CN. Slight differences in clustering RA, KA, andGA can be explained by other factors (lake character-istics or substrate types). This suggests that water pH
Fig. 5. Structure of macroinvertebrates feeding groups in each lake. DET – detritivores, FIL – filtrators, GRA – grazers, PRE –predators, SHR – shredders. Data from autumn.

Littoral macroinvertebrates of acidified lakes 1195
is the driving factor for both the water chemistry andfor macroinvertebrates.Moiseenko (2003) considered Ephemeroptera,
Crustacea and Mollusca to be the most acid sensitivegroups of macroinvertebrates, usually absent in waterswith pH < 5. No crustaceans were found in the Bo-hemian Forest lakes, and only one mollusk, Pisidiumcasertanum, found by us in two lakes with the high-est pH (LA and GA), was previously found also inKA (Soldán et al. 2012). Økland & Kuiper (1982) sug-gested that P. casertanum is the only species of thegenus that may be found in strongly acidified waters.According to Horsák (2006), P. casertanum is the re-gionally most acid tolerant aquatic mollusk, which cansurvive at pH 4.7 and appears to be calcifuge to someextent. Norwegian studies showed a tolerance limit ofpH ∼ 4.7 (Fjellheim & Raddum 1990). Šporka (1992)explained this high tolerance by its living in a sedimentenvironment where pH is higher than on the sedimentsurface. However, our results and conclusions of Soldánet al. (2012) indicate that this species occurs only inthe least acidified Bohemian Forest lakes (pH > 5.3).One of the possible explanations for this could be lowcontent of Ca (< 1 mg L−1) (Raddum & Skjelkvĺle1995). The most frequentmayfly, Leptophlebia vesper-tina, considered one of the most acid-tolerant mayflieswith a tolerance limit of pH ∼ 4 (Vrba et al. 2003;Schartau et al. 2008), was found to be present in allthe Bohemian Forest lakes, both currently and eventhroughout the whole acidification period (Vrba et al.2003; Soldán et al. 2012). The mayflies Cloeon dipterumand Leptophlebia marginata have been recorded in theBohemian Forest lakes only recently (summarized inSoldán et al. 2012), whereas Ameletus inopinatus andSiphlonurus lacustris were also found before the acid-ification period (Landa & Soldán 1989). According toBraukmann & Biss (2004), A. inopinatus and S. lacus-tris are acid-tolerant taxa occurring in periodically crit-ically acidic waters (pH < 6.5 with periods < 5.0). C.dipterum, A. inopinatus and S. lacustris were identifiedonly in samples from the least acidified LA; neverthe-less, they all were also recently found in other lakes,including the strongest acidified CN (reviewed by Sol-dán et al. 2012).The dragonflies, water bugs, and alderflies are
in general considered to be acid-tolerant groups. Wesuppose that their occurrence is mainly limited by theavailability of suitable food sources and by the presenceof other predators. Nevertheless, Soldán et al. (2012)concluded that some species of water bugs are probablyless acid-tolerant, as they have been observed in sub-stantially higher numbers during the past two decadesof recovery.The stoneflies are considered an acid-tolerant
group in general, but acidification can significantly re-duce their diversity, as documented in the families Leuc-tridae and Nemouridae. Some species, though, havedemonstrated a high resistance to acid conditions (Tix-ier & Guérold 2005). In the Bohemian Forest lakes,all stoneflies found (Nemoura cinerea, N. avicularis,
and Leuctra nigra) are considered to be acid-resistant(Braukmann & Biss 2004; Schartau et al. 2008). Thesespecies occurred only in small numbers in the littoral ofthe Bohemian Forest lakes, while they were abundantin the inflows and outflows (unpublished data).The water beetles are generally tolerant to acid-
ification, especially the adults, which breathe atmo-spheric oxygen and have exoskeletons effectively pro-tecting their bodies (Havas & Rosseland 1995). The dis-tribution of water beetles in the studied lakes is morelikely dependent on food supply or habitat suitability.The majority of beetles found in the Bohemian Forestlakes belonged to the family Dytiscidae. The dominat-ing dytiscid Hydroporus palustris is abundant in theCzech Republic and lives in all types of standing andflowing waters (Boukal et al. 2007). The second mostfrequent beetle, Deronectes latus, also occurs through-out the Czech Republic, but only locally and rarely(Boukal et al. 2007). The presence of Nebrioporus as-similis was unexpected, as this species had been foundin the Czech Republic only once, near the town ofPříbram, probably in the Brdy hills in the 1950s, andsince then it was considered regionally extinct (Boukalet al. 2007). Its occurrence in PR was again confirmedin 2010 and since then the species has been reclassifiedas critically endangered (Boukal et al. 2012). The otherspecies of beetles found in the Bohemian Forest lakesare common.The caddisflies contributed substantially to the
species diversity of benthic macroinvertebrates due topresence of acid-tolerant and tyrphophilous species.Mystacides azurea and species of the family Phryganei-dae and Limnephilidae were the only taxa found in thestrongest acidified lakes CN and CT, already in theearly 1990s (reviewed in Soldán et al. 2012). Recently,M. azurea still dominates in these two lakes, but cad-disfly species diversity in both lakes has substantiallyincreased since that time. The absence of Oligotrichastriata in CT and CN is noteworthy, since this specieswas also not found in these lakes during the preacidifi-cation period (Soldán et al. 2012). It is an acid-tolerantspecies (Braukmann & Biss 2004), so its absence in thestony littoral lacking moss and macrophytes in CT andCN (Supplementary file; Figs S1, S2) can be explainedrather by different habitat preferences. In other lakes,species of the families Limnephilidae, Polycentropodi-dae and Phryganeidae dominated, and species of thefamily Molannidae (Molanna nigra orMolannodes tinc-tus) also occurred in most of the lakes. The caddisflyHolocentropus dubius was found only in the least acid-ified LA and GA; however, its occurrence is probablydetermined more by habitat preferences (phytal) (Grafet al. 2008) than pH value.The dipterans were dominated by the family Chi-
ronomidae, and this group had the highest species rich-ness: 48 species/taxa of chironomids have been collectedso far in the Bohemian Forest lakes (Soldán et al. 2012).A recent investigation based on pupal exuviae analysis(Bitušík & Svitok 2006) showed that differences in tax-onomic composition among the Bohemian Forest lakes

1196 L. Ungermanová et al.
could be explained significantly by lake alkalinity (asa proxy of pH). However, chironomids have so far notreflected the effects of acidification in the investigatedlakes in such a dramatic way as mayflies and stoneflies(Vrba et al. 2003).Although our data on macroinvertebrates from all
Bohemian Forest lakes (Appendix 2) do not enable ananalysis of recovery processes, they have been used tosummarize the historical changes in macroinvertebratetaxa composition in the Bohemian Forest lakes (referredas Ungermanová 2009 in Soldán et al. 2012).Acidification can cause changes in occurrence of
macroinvertebrate taxa not only directly through thetoxicity of H+ , L-Al and heavy metals or by the dis-turbance of Ca2+ metabolism, but also indirectly byinfluencing the food supply (Økland & Økland 1986;Ormerod et al. 1987; Havas & Rosseland 1995). Al-though the total biomass of organisms often remainsunchanged, the disappearance of acid-sensitive taxamay cause the disruption and simplification of foodchains (Hendrey & Wright 1976). Detritivores entirelydominated in all investigated lakes except LA, whereasgrazers were always present in minor amounts. Grazers,living on the surface of the bottom substrate or aquaticplants, have been documented as being reduced in acid-ified waters (e.g. Dangles & Guérold 2000; Horecký etal. 2006), while shredders and detritivores are less af-fected because they often burrow in the sediment, wherethey find more suitable chemical conditions. However,particular instars can differ in their environmental aswell as in their food strategy during ontogenesis (Moog2002). A decline in the diversity or abundance of grazerscan also be a result of biomass reduction and changes inthe bacterial composition (Ledger & Hildrew 2001). Inthe studied lakes, predators were abundant among themacroinvertebrates, predominantly represented by wa-ter bugs, water beetles, dragonflies and water midges.In strongly acidified waters, these groups often becometop predators due to the extinction of fish and occurin increased abundances (Layer et al. 2010). Neverthe-less, this trend was not observed in our data as all lakesare fishless and the highest proportion of predators wasobserved in LA, one of the least acidified lakes. We didnot find any increasing proportion of predators with de-creasing pH, although this conclusion must be carefullyconsidered as the chironomids were not included. Thehigh proportion of predators in LA may rather be ex-plained by the highest occurrence of macrophytes andmoss as suitable microhabitats (Fig. S2).
Acknowledgements
This research was supported by the Czech Science Founda-tion (GACR grants No. 526/09/0567 and 206/07/1200), theEnvironment Project of the European Commission EURO-LIMPACS (GOCE-CT-2003-505540), the Ministry of theEnvironment of the Czech Republic in the framework ofthe Convention of Long-Range Transboundary Air Pollution(LRTAP), and by the Grant Agency of Charles University(GAUK No. 264711). This work was also partly supported
by the Ministry of Culture of the Czech Republic (DKRVO2013/11, National Museum, 00023272). We wish to expressour gratitude Jindřiška Bojková, Martin Fikáček, Jiří Há-jek, Lucie Juřičková, Miroslav Papáček, Jan Rucki, TomášSoldán, and Jan Špaček for their participation on speciesidentification. The authors wish to thank David Hardekopfand Jaroslav Vrba for their contribution to this research.
References
Bitušík P. & Svitok M. 2006. Structure of chironomid as-semblages along environmental and geographical gradientsin the Bohemian Forest lakes (Central Europe): An ex-ploratory analysis. Biologia 61 (Suppl. 20): S467–S476.DOI: 10.2478/s11756-007-0063-y
Boukal D.S., Boukal M., Fikáček M., Hájek J., Klečka J., SkalickýS., Šťastný J. & Trávníček D. 2007. Katalog vodních broukůČeské republiky [Catalogue of water beetles of the Czech Re-public]. Klapalekiana 43 (Suppl.): 1–289.
Boukal D.S., Fikáček M., Hájek J., Konvička O., Křivan V., Se-jkora R., Skalický S., Straka M., Sychra J. & Trávníček D.2012. Nové a zajímavé nálezy vodních brouků z území Českérepubliky (Coleoptera: Sphaeriusidae, Dytiscidae, Helophori-dae, Hydrophilidae, Georissidae, Hydraenidae, Scirtidae,Elmidae, Dryopidae, Limnichidae, Heteroceridae) [New andinteresting records of water beetles from the Czech Repub-lic (Coleoptera: Sphaeriusidae, Dytiscidae, Helophoridae, Hy-drophilidae, Georissidae, Hydraenidae, Scirtidae, Elmidae,Dryopidae, Limnichidae, Heteroceridae]. Klapalekiana 48 (1-2): 1–21.
Braukmann U. & Biss R. 2004. Conceptual study – An im-proved method to assess acidification in German streamsby using benthic macroinvertebrates. Limnologica – Ecologyand Management of Inland Waters 34 (4): 433–450. DOI:10.1016/S0075-9511(04)80011-2
Dangles O. & Guérold J. 2000. Structural and functional re-sponses of benthic macroinvertebrates to acid precipita-tion in two forested headwater streams (Vosges Mountains,northeastern France). Hydrobiologia 418 (1): 25–31. DOI:10.1023/A:1003805902634
Dougan W.K. & Wilson A.L. 1974. The absorptiometric de-termination of aluminium in water. A comparison of somechromogenic reagents and the development of an improvedmethod. Analyst 99 (1180): 413–430. DOI: 10.1039/AN9749900413
Driscoll C.T. 1984. A procedure for the fractionation of aqueousaluminium in dilute waters. Int. J. Environ. Anal. Chem. 16(4): 93–104. DOI: 10.1080/03067318408076957
Evans C.D., Cullen J.M., Alewell C., Marchetto A., Moldan F.,Kopáček J., Prechtel A., Rogora M., Vesely J. & Wright R.F.2001. Recovery from acidification in European surface waters.Hydrol. Earth Syst. Sci. 5 (3): 283–297. DOI: 10.5194/hess-5-283-2001
Fjellheim A. & Raddum G.G. 1990. Acid precipitation: Biologicalmonitoring of streams and lakes. Sci. Total Envir. 96 (1-2):57–66. DOI: 10.1016/0048-9697(90)90006-G
Fjellheim A., Raddum G.G., Vandvik V., Cogalniceanu D., Bog-gero A., Brancelj A., Galas J., Sporka F., Vidinova Y., BitusikP., Dumnicka E., Galdean N., Kownacki A., Krno I., PredaE., Rısnoveanu G. & Stuchlik E. 2009. Diversity and distribu-tion patterns of benthic invertebrates along alpine gradients.A study of remote European freshwater lakes. Adv. Limnol.62: 167–190. DOI: 10.1127/advlim/62/2009/167
Fott J., Pražáková M., Stuchlík E. & Stuchlíková Z. 1994. Acidi-fication of lakes in Šumava (Bohemia) and in the High TatraMountains (Slovakia). Hydrobiologia 274 (1-3): 37–47. DOI:10.1007/BF00014625
Frost S., Huni A. & Kershaw W.E. 1971. Evaluation of a kickingtechnique for sampling stream bottom fauna. Can. J. Zool.49 (2): 167–173. DOI: 10.1139/z71-026

Littoral macroinvertebrates of acidified lakes 1197
Graf W., Murphy J., Dahl J., Zamora-Munoz C. & Lopez-Rodriguez M.J. 2008. Trichoptera. Distribution and Eco-logical Preferences of European Freshwater Organisms. Vol-ume 1. Pensoft Publishers, Sofia-Moscow, 388 pp. ISBN:9789546424419
Havas M. & Rosseland B.O. 1995. Response of zooplankton, ben-thos, and fish to acidification: an overview. Water Air SoilPollut. 85 (1): 51–62. DOI: 10.1007/BF00483688
Hendrey G.R. & Wright R.F. 1976. Acid precipitation in Norway:Effects on aquatic fauna J. Great Lakes Res. 2 (Suppl. 1):192–207. DOI: 10.1016/S0380-1330(76)72327-X
Horecký J., Stuchlík E., Chvojka P., Hardekopf D.W., Mihalje-vič M. & Špaček J. 2006. Macroinvertebrate community andchemistry of the most atmospherically acidified streams in theCzech Republic. Water Air Soil Pollut. 173 (1-4): 261–272.DOI: 10.1007/s11270-005-9071-0
Horsák M. 2006. Mollusc community patterns and species re-sponse curves along a mineral richness gradient: a case studyin fens. J. Biogeograph. 33 (1): 98–107. DOI: 10.1111/j.1365-2699.2005.01359.x
Hořická Z., Stuchlík E., Hudec I., Černý M. & Fott J. 2006. Acidi-fication and the structure of crustacean zooplankton in moun-tain lakes: The Tatra Mountains (Slovakia, Poland). Biologia61 (18 Suppl.): S121–S134. DOI: 10.2478/s11756-006-0125-6
Jeffries D.S., Clair T.A., Couture S., Dillon P.J., Dupont J.,Keller W.B., McNicol D.K., Turner M.A., Vet R. & Wee-ber R. 2003. Assessing the recovery of lakes in southeasternCanada from effects of acidic deposition. Ambio 32 (3): 176–182. DOI: 10.1579/0044-7447-32.3.176
Kopáček J. & Hejzlar J. 1993. Semi-micro determination of to-tal phosphorus in fresh waters with perchloric acid diges-tion. Int. J. Environ. Anal. Chem. 53 (3): 173–183. DOI:10.1080/03067319308045987
Kopáček J., Hejzlar J., Stuchlík E., Fott J. & Veselý J. 1998.Reversibility of acidification of mountain lakes after reductionin nitrogen and sulphur emissions in Central Europe. Limnol.Oceanogr. 43 (2): 357–36. DOI: 10.4319/lo.1998.43.2.0357
Kopáček J., Stuchlík E., Veselý J., Schaumburg J., Anderson I.C.,Fott J., Hejzlar J. & Vrba J. 2002. Hysteresis in reversal ofCentral European mountain lakes from atmospheric acidi-fication. Water Air Soil Pollut. Focus 2 (2): 91–114. DOI:10.1023/A:1020190205652
Kopáček J., Turek J., Hejzlar J. & Porcal P. 2011. Bulk depositionand throughfall fluxes of elements in the Bohemian Forest(Central Europe) from 1998 to 2009. Boreal Envir. Res. 16(6): 495–508.
Kopáček J., Veselý J. & Stuchlík E. 2001. Sulphur and nitrogenfluxes and budgets in the Bohemian Forest and Tatra Moun-tains during the industrial revolution (1850-2000). Hydrol.Earth Syst. Sci. 5: 391–405. DOI: 10.5194/hess-5-391-2001
Kownacki A., Galas J., Dumnicka E. & Mielewczyk S. 2000. In-vertebrate communities in permanent and temporary highmountain lakes (Tatra Mts.). Ann. Limnol. – Int. J. Lim-nol.36 (3): 181–188. DOI: 10.1051/limn/2000016
Krno I., Šporka F., Galas J., Hamerlík L., Zaťovičová Z. & BitušíkP. 2006. Littoral benthic macroinvertebrates of mountainlakes in the Tatra Mountains (Slovakia, Poland). Biologia 61(Suppl. 18): S147–S166. DOI: 10.2478/s11756-006-0127-4
Kubovčík V. & Bitušík P. 2006. Subfossil chironomids (Diptera,Chironomidae) in three Tatra Mountain lakes (Slovakia) onan acidification gradient. Biologia 61 (Suppl. 18): S213–S220. DOI: 10.2478/s11756-006-0133-6
Landa V. & Soldán T. 1989. Rozšíření jepic v ČSSR a jeho změnyv souvislosti se změnami kvality vody v povodí Labe [Distri-bution of mayflies (Ephemeroptera) in Czechoslovakia and itschanges in connection with water quality changes in the Elbebasin]. Studie ČSAV, 17, Academia, Praha, 172 pp.
Laudon H. & Westling O. 2005. Drought induced episodes: Canthey counteract the acidification recovery in southern Swe-den? p. 384. In: Brimblecombe P., Hiroshi H., Houle D. &Novak M. (eds), Acid Rain 2005, 7th International Confer-ence on Acid Deposition, Prague, Czech Republic, June 12– 17, 2005 . Conference Abstracts, ČHMÚ Praha, 384 pp.ISBN: 978-1-4020-5884-4
Layer K., Hildrew A., Monteith D. & Woodward G. 2010. Long-term variation in the littoral food web of an acidified moun-tain lake. Global Change Biol. 16 (11): 3133–3143. DOI:10.1111/j.1365-2486.2010.02195.x
Ledger M.E. & Hildrew A.G. 2001. Growth of an acid-tolerantstonefly on epilithic biofilms from streams of contrasting pH.Freshwater Biol. 46 (11): 1457–1470. DOI: 10.1046/j.1365-2427.2001.00764.x
Mackereth F.J.H., Heron J. & Talling J.F. 1978. Water Analysis:Some Revised Methods for Limnologists. Freshwater Biologi-cal Association Scientific Publication no. 36. Titus Wilson &Sons Ltda, Kendal, 120 pp. ISBN: 0900386312
Majer V., Cosby B.J., Kopáček J. & Veselý J. 2003. Modellingreversibility of central European mountain lakes from acidi-fication: Part I – The Bohemian Forest. Hydrol. Earth Syst.Sci. 7: 494–509. DOI: 10.5194/hess-7-494-2003
Moiseenko T.I. 2003. Effects of acidification on aquatic ecosys-tems. Russ. J. Ecol. 36 (2): 93–102. DOI: 10.1007/s11184-005-0017-y
Moog O. (ed.) 2002. Fauna Aquatica Austriaca. A ComprehensiveSpecies Inventory of Austrian Aquatic Organisms with Eco-logical Notes. 2nd Edition 2002. Wasserwirtschaftskataster,Bundesministerium fur Land- und Forstwirtschaft, Umweltund Wasserwirtschaft, Vienna. xx pp. ISBN: 3-85 174-044-0
Nedbalová L., Vrba J., Fott J., Kohout L., Kopáček J., Macek M.& Soldán T. 2006. Biological recovery of the Bohemian Forestlakes from acidification. Biologia 61 (20 Suppl.): S453–S466.DOI: 10.2478/s11756-007-0071-y
Økland K.A. & Kuiper J.G.J. 1982. Distribution of small mussels(Sphaeriidae) in Norway, with notes on their ecology. Mala-cologia 22 (1-2): 469–477.
Økland J. & Økland K.A. 1986. The effects of acid depositionon benthic animals in lakes and streams. Experientia 42 (5):471–486. DOI: 10.1007/BF01946685
Ormerod S.J., Boole P., McCahon C.P., Weatherley N.S., PascoeD. & Edwards R.W. 1987. Short-term experimental acidifica-tion of a Welsh stream: comparing the biological effects ofhydrogen ions and aluminium. Freshwater Biol. 17 (2): 341–356. DOI: 10.1111/j.1365-2427.1987.tb01054.x
Oulehle F., Cosby B.J., Wright R.F., Hruška J., Kopáček J., KrámP., Evans C.D. & Moldan F. 2012. Modelling soil nitrogen:The MAGIC model with nitrogen retention linked to car-bon turnover using decomposer dynamics. Envir. Pollut. 165:158–166. DOI: 10.1016/j.envpol.2012.02.021
Posch M., Hettelingh J.-P. & Slotweg J. (eds) 2003. Manual forDynamic Modelling of Soil Response to atmospheric Depo-sition, RIVM Report 259101012. Bilthoven, Netherlands, 71pp.
Raddum G.G. & Skjelkvĺle B.L. 1995. Critical limits of acidifi-cation to invertebrates in different regions of Europe. WaterAir Soil Pollut. 85 (2): 475–480. DOI: 10.1007/BF00476874
Schartau A.K., Moe S.J., Sandin L., McFarland B. & RaddumG.G. 2008. Macroinvertebrate indicators of lake acidifica-tion: analysis of monitoring data from UK, Norway and Swe-den. Aquat. Ecol. 42 (2): 293–305. DOI: 10.1007/s10452-008-9186-7
Schindler D.W. 1988. Effects of acid rain on freshwater ecosys-tems. Science 239 (4836): 149–157. DOI: 10.1126/sci-ence.239.4836.149
Soldán T., Bojková J., Vrba J., Bitušík P., Chvojka P., PapáčekM., Peltanová J., Sychra J. & Tátosová J. 2012. Aquatic in-sects of the Bohemian Forest glacial lakes: Diversity, long-term changes and influence of acidification. Silva Gabreta 18(3): 123–283.
Stuchlík E., Appleby P., Bitušík P., Curtis C., Fott J., KopáčekJ., Pražáková M., Rose N., Strunecký O. & Wright R.F. 2002.Reconstruction of long-term changes in lake water chem-istry, zooplankton and benthos of a small, acidified high-mountain lake: Magic modelling and paleolimnogical anal-ysis. Water Air Soil Pollut. Focus 2 (2): 127–138. DOI:10.1023/A:1020198424308
Stuchlík E., Kopáček J., Fott J. & Hořická Z. 2006. Chem-ical composition of the Tatra Mountain lakes: Responseto acidification. Biologia 61 (18 Suppl.): S11–S20. DOI:10.2478/s11756-006-0116-7

1198 L. Ungermanová et al.
Šporka F. 1992. Máloštetinavce (Oligochaeta) jazier v ZápadnýchTatrách. Zborník prác o Tatranskom národnom parku 32:139–148.
Tixier G. & Guérold F. 2005. Plecoptera response of acidifica-tion in several head water streams in the Vosges Mountains(northeastern France). Biodivers. Conserv. 14 (6): 1525–1539. DOI:.10.1007/s10531-004-9790-3
Ungermanová L. 2009. Makrozoobentos litorální zóny acidifiko-vaných šumavských jezer [Macrozoobenthos of the littoralzone of acidified lakes in the Bohemian Forest Mountains].Diplomová práca. Univerzita Karlova v Praze, Přírodovědec-ká fakulta, Ústav pro životní prostředí [Ms., Diploma thesis,Charles University in Prague], 53 pp.
Veselý J. 1994. Investigation of the nature of the Šumava lakes: areview. Čas. Nár. Muz. Praha, Řada Přírodověd. 163 (1-4):103–120.
Veselý J., Kopáček J. & Norton S.A. 2003. Increasing temper-ature decreases aluminium concentrations in Central Euro-pean lakes recovering from acidification. Limnol. Oceanogr.,48 (6): 2346–2354. DOI: 10.4319/lo.2003.48.6.2346
Vranovský M., Krno K., Šporka F. & Tomajka J. 1994. The effectof anthropogenic acidification on the hydrofauna of the lakesof the West Tatra Mountains (Slovakia). Hydrobiologia 274(1-3): 163–170. DOI: 10.1007/BF00014639
Vrba J., Kopáček J. & Fott J. 2002. Šumavská ledovcová jezera napřelomu tisíciletí [Glacial Lakes in Šumava/Bohemian Forestlake on the turn of centuries]. Živa 6: 265–269.
Vrba J., Kopáček J., Fott J., Kohout L., Nedbalová L., PražákováM., Soldán T. & Schaumburg J. 2003. Long-term studies(1871–2000) on acidification and recovery of lakes in the Bo-hemian Forest (central Europe). Sci. Tot. Envir. 310 (1-3):73–85. DOI: 10.1016/S0048-9697(02)00624-1
Wright R.F., Larssen T., Camarero L., Cosby B.J., Ferrier R.C.,Helliwell R., Forsius M., Jenkins A., Kopáček J., Majer V.,Moldan F., Posch M., Rogora M. & Schopp W. 2005. Recoveryof acidified European surface water. Environ. Sci. Technol.38/ 39 (3): 64A–72A. PMID: 15757325
Received November 18, 2013Accepted July 7, 2014
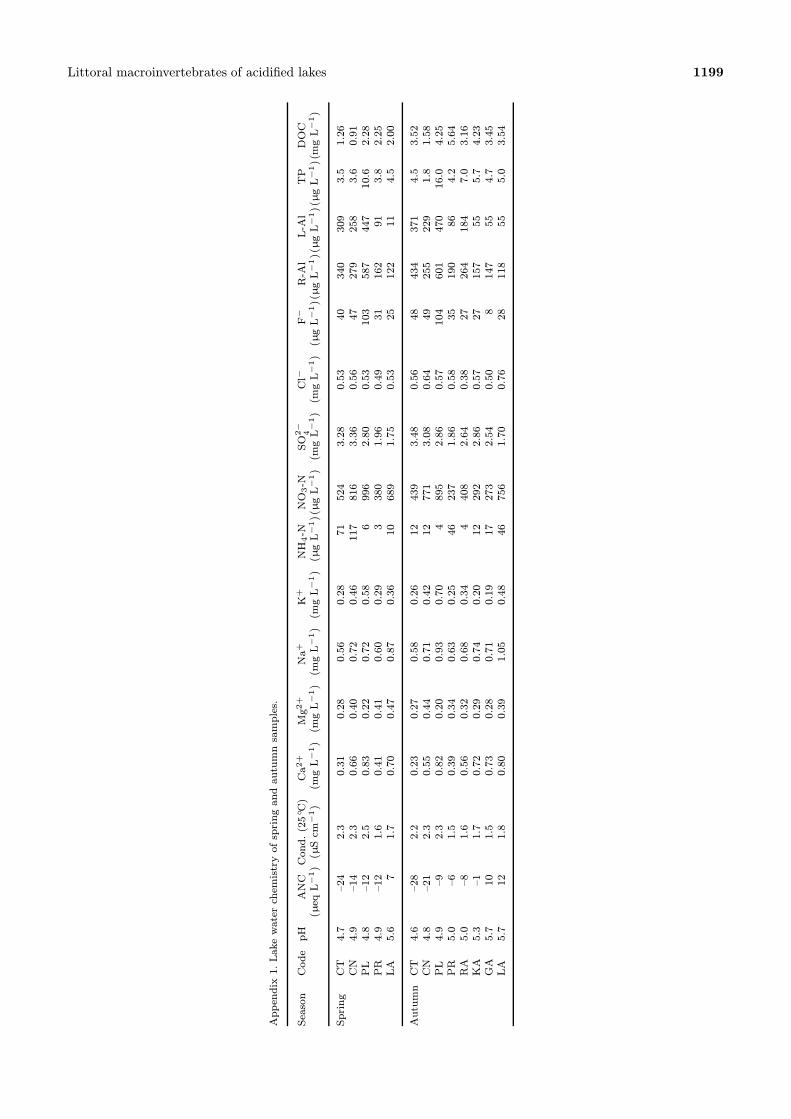
Littoral macroinvertebrates of acidified lakes 1199
Appendix1.Lakewaterchemistryofspringandautumnsamples.
Season
CodepH
ANCCond.(25◦ C)
Ca2+
Mg2+
Na+
K+
NH4-N
NO3-N
SO2−4
Cl−
F−
R-Al
L-Al
TP
DOC
(µeqL−1)(µScm
−1)(mgL−1)(mgL−1)(mgL−1)(mgL−1)(µgL−1)(
µgL−1)(mgL−1)(mgL−1)(µgL−1)(
µgL−1)(
µgL−1)(
µgL−1)(mgL−1)
Spring
CT
4.7
–24
2.3
0.31
0.28
0.56
0.28
71524
3.28
0.53
40340
309
3.5
1.26
CN
4.9
–14
2.3
0.66
0.40
0.72
0.46
117
816
3.36
0.56
47279
258
3.6
0.91
PL
4.8
–12
2.5
0.83
0.22
0.72
0.58
6996
2.80
0.53
103
587
447
10.6
2.28
PR
4.9
–12
1.6
0.41
0.41
0.60
0.29
3380
1.96
0.49
31162
913.8
2.25
LA
5.6
71.7
0.70
0.47
0.87
0.36
10689
1.75
0.53
25122
114.5
2.00
AutumnCT
4.6
–28
2.2
0.23
0.27
0.58
0.26
12439
3.48
0.56
48434
371
4.5
3.52
CN
4.8
–21
2.3
0.55
0.44
0.71
0.42
12771
3.08
0.64
49255
229
1.8
1.58
PL
4.9
–92.3
0.82
0.20
0.93
0.70
4895
2.86
0.57
104
601
470
16.0
4.25
PR
5.0
–61.5
0.39
0.34
0.63
0.25
46237
1.86
0.58
35190
864.2
5.64
RA
5.0
–81.6
0.56
0.32
0.68
0.34
4408
2.64
0.38
27264
184
7.0
3.16
KA
5.3
–11.7
0.72
0.29
0.74
0.20
12292
2.86
0.57
27157
555.7
4.23
GA
5.7
101.5
0.73
0.28
0.71
0.19
17273
2.54
0.50
8147
554.7
3.45
LA
5.7
121.8
0.80
0.39
1.05
0.48
46756
1.70
0.76
28118
555.0
3.54
1200 L. Ungermanová et al.
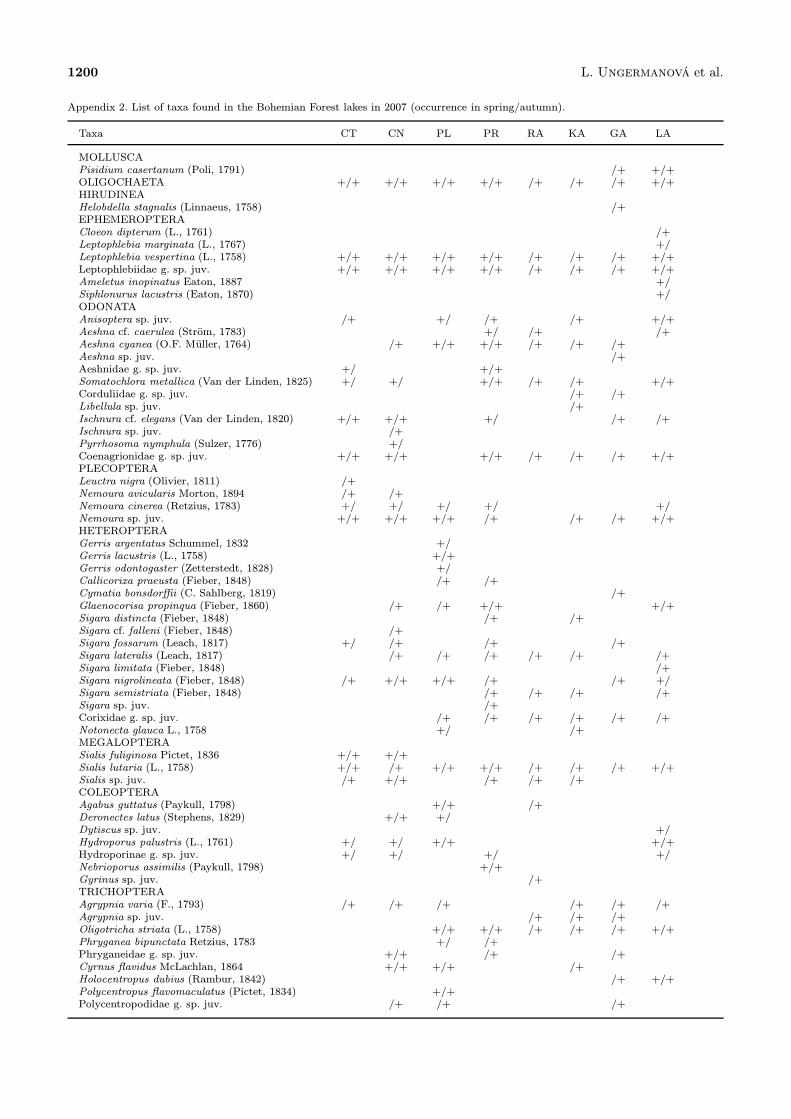
Appendix 2. List of taxa found in the Bohemian Forest lakes in 2007 (occurrence in spring/autumn).
Taxa CT CN PL PR RA KA GA LA
MOLLUSCAPisidium casertanum (Poli, 1791) /+ +/+OLIGOCHAETA +/+ +/+ +/+ +/+ /+ /+ /+ +/+HIRUDINEAHelobdella stagnalis (Linnaeus, 1758) /+EPHEMEROPTERACloeon dipterum (L., 1761) /+Leptophlebia marginata (L., 1767) +/Leptophlebia vespertina (L., 1758) +/+ +/+ +/+ +/+ /+ /+ /+ +/+Leptophlebiidae g. sp. juv. +/+ +/+ +/+ +/+ /+ /+ /+ +/+Ameletus inopinatus Eaton, 1887 +/Siphlonurus lacustris (Eaton, 1870) +/ODONATAAnisoptera sp. juv. /+ +/ /+ /+ +/+Aeshna cf. caerulea (Strom, 1783) +/ /+ /+Aeshna cyanea (O.F. Muller, 1764) /+ +/+ +/+ /+ /+ /+Aeshna sp. juv. /+Aeshnidae g. sp. juv. +/ +/+Somatochlora metallica (Van der Linden, 1825) +/ +/ +/+ /+ /+ +/+Corduliidae g. sp. juv. /+ /+Libellula sp. juv. /+Ischnura cf. elegans (Van der Linden, 1820) +/+ +/+ +/ /+ /+Ischnura sp. juv. /+Pyrrhosoma nymphula (Sulzer, 1776) +/Coenagrionidae g. sp. juv. +/+ +/+ +/+ /+ /+ /+ +/+PLECOPTERALeuctra nigra (Olivier, 1811) /+Nemoura avicularis Morton, 1894 /+ /+Nemoura cinerea (Retzius, 1783) +/ +/ +/ +/ +/Nemoura sp. juv. +/+ +/+ +/+ /+ /+ /+ +/+HETEROPTERAGerris argentatus Schummel, 1832 +/Gerris lacustris (L., 1758) +/+Gerris odontogaster (Zetterstedt, 1828) +/Callicorixa praeusta (Fieber, 1848) /+ /+Cymatia bonsdorffii (C. Sahlberg, 1819) /+Glaenocorisa propinqua (Fieber, 1860) /+ /+ +/+ +/+Sigara distincta (Fieber, 1848) /+ /+Sigara cf. falleni (Fieber, 1848) /+Sigara fossarum (Leach, 1817) +/ /+ /+ /+Sigara lateralis (Leach, 1817) /+ /+ /+ /+ /+ /+Sigara limitata (Fieber, 1848) /+Sigara nigrolineata (Fieber, 1848) /+ +/+ +/+ /+ /+ +/Sigara semistriata (Fieber, 1848) /+ /+ /+ /+Sigara sp. juv. /+Corixidae g. sp. juv. /+ /+ /+ /+ /+ /+Notonecta glauca L., 1758 +/ /+MEGALOPTERASialis fuliginosa Pictet, 1836 +/+ +/+Sialis lutaria (L., 1758) +/+ /+ +/+ +/+ /+ /+ /+ +/+Sialis sp. juv. /+ +/+ /+ /+ /+COLEOPTERAAgabus guttatus (Paykull, 1798) +/+ /+Deronectes latus (Stephens, 1829) +/+ +/Dytiscus sp. juv. +/Hydroporus palustris (L., 1761) +/ +/ +/+ +/+Hydroporinae g. sp. juv. +/ +/ +/ +/Nebrioporus assimilis (Paykull, 1798) +/+Gyrinus sp. juv. /+TRICHOPTERAAgrypnia varia (F., 1793) /+ /+ /+ /+ /+ /+Agrypnia sp. juv. /+ /+ /+Oligotricha striata (L., 1758) +/+ +/+ /+ /+ /+ +/+Phryganea bipunctata Retzius, 1783 +/ /+Phryganeidae g. sp. juv. +/+ /+ /+Cyrnus flavidus McLachlan, 1864 +/+ +/+ /+Holocentropus dubius (Rambur, 1842) /+ +/+Polycentropus flavomaculatus (Pictet, 1834) +/+Polycentropodidae g. sp. juv. /+ /+ /+
Littoral macroinvertebrates of acidified lakes 1201
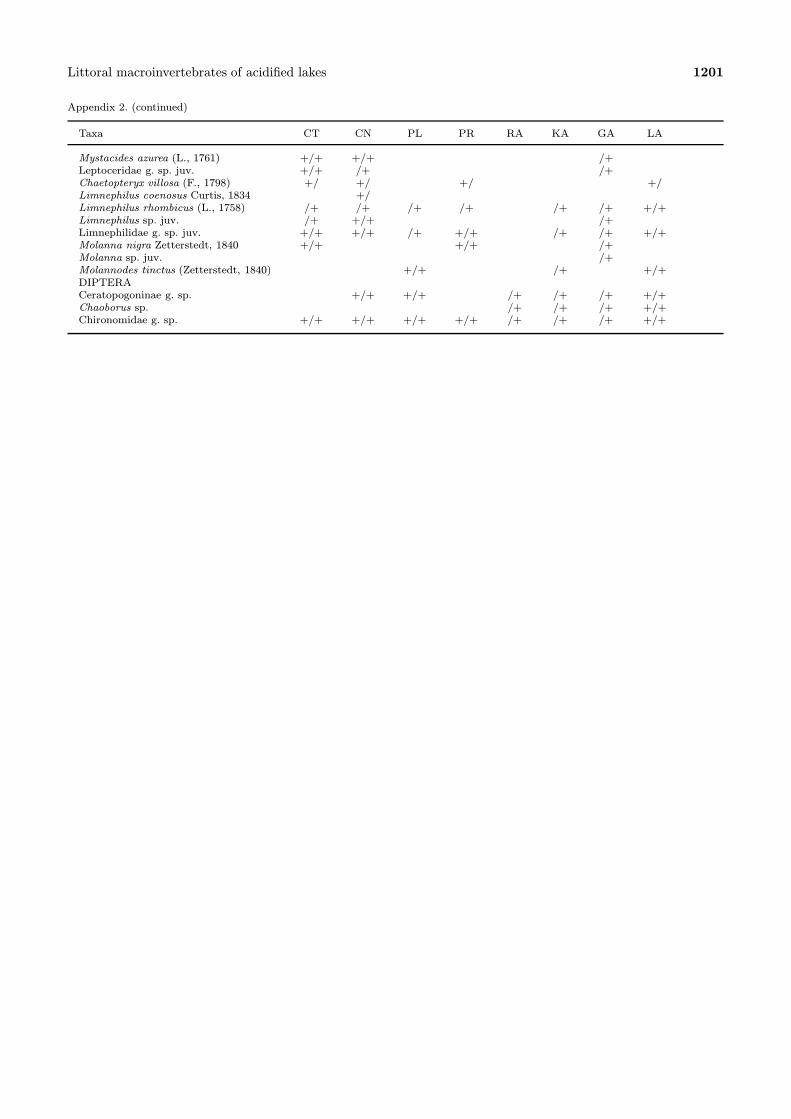
Appendix 2. (continued)
Taxa CT CN PL PR RA KA GA LA
Mystacides azurea (L., 1761) +/+ +/+ /+Leptoceridae g. sp. juv. +/+ /+ /+Chaetopteryx villosa (F., 1798) +/ +/ +/ +/Limnephilus coenosus Curtis, 1834 +/Limnephilus rhombicus (L., 1758) /+ /+ /+ /+ /+ /+ +/+Limnephilus sp. juv. /+ +/+ /+Limnephilidae g. sp. juv. +/+ +/+ /+ +/+ /+ /+ +/+Molanna nigra Zetterstedt, 1840 +/+ +/+ /+Molanna sp. juv. /+Molannodes tinctus (Zetterstedt, 1840) +/+ /+ +/+DIPTERACeratopogoninae g. sp. +/+ +/+ /+ /+ /+ +/+Chaoborus sp. /+ /+ /+ +/+Chironomidae g. sp. +/+ +/+ +/+ +/+ /+ /+ /+ +/+
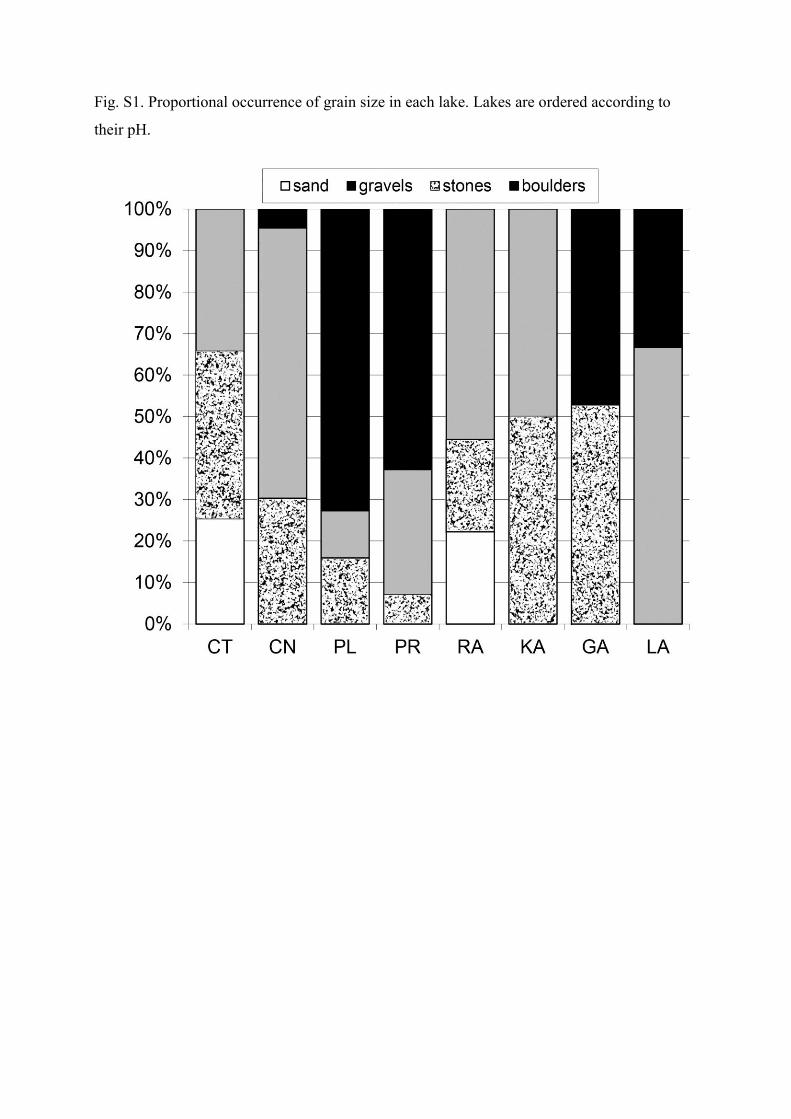
Fig. S1. Proportional occurrence of grain size in each lake. Lakes are ordered according to
their pH.
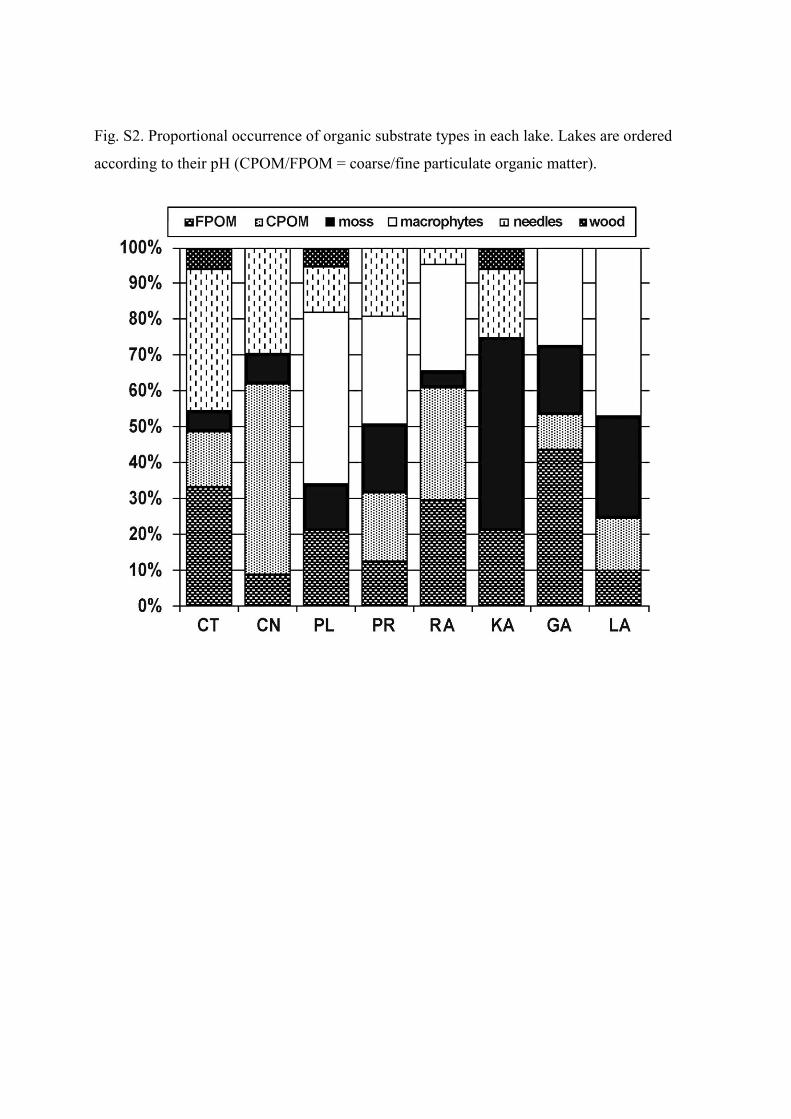
Fig. S2. Proportional occurrence of organic substrate types in each lake. Lakes are ordered
according to their pH (CPOM/FPOM = coarse/fine particulate organic matter).





























