Embed Size (px)
Citation preview

Lactisole Interacts with the Transmembrane Domains of HumanT1R3 to Inhibit Sweet Taste*□S
Received for publication, December 20, 2004, and in revised form, January 20, 2005Published, JBC Papers in Press, January 24, 2005, DOI 10.1074/jbc.M414287200
Peihua Jiang‡, Meng Cui§, Baohua Zhao‡, Zhan Liu‡¶, Lenore A. Snyder‡, Lumie M. J. Benard‡¶,Roman Osman§, Robert F. Margolskee‡�¶, and Marianna Max‡**
From the Department of ‡Neuroscience and §Physiology and Biophysics, Mount Sinai School of Medicine, New York, NewYork 10029 and the ¶Howard Hughes Medical Institute, Mount Sinai School of Medicine, New York, New York 10029
The detection of sweet-tasting compounds is mediatedin large part by a heterodimeric receptor comprised ofT1R2�T1R3. Lactisole, a broad-acting sweet antagonist,suppresses the sweet taste of sugars, protein sweeteners,and artificial sweeteners. Lactisole’s inhibitory effect isspecific to humans and other primates; lactisole does notaffect responses to sweet compounds in rodents. By het-erologously expressing interspecies combinations ofT1R2�T1R3, we have determined that the target for lac-tisole’s action is human T1R3. From studies with mouse/human chimeras of T1R3, we determined that the molec-ular basis for sensitivity to lactisole depends on only a fewresidues within the transmembrane region of humanT1R3. Alanine substitution of residues in the transmem-brane region of human T1R3 revealed 4 key residues re-quired for sensitivity to lactisole. In our model of T1R3’sseven transmembrane helices, lactisole is predicted todock to a binding pocket within the transmembrane re-gion that includes these 4 key residues.
Taste is a primal sense that enables diverse organisms toidentify and ingest sweet-tasting nutritious foods and to rejectbitter-tasting environmental poisons (1). Taste perception canbe categorized into five distinct qualities: salty, sour, bitter,umami (amino acid taste), and sweet (1). Salty and sour dependon the actions of ion channels. Bitter, umami, and sweet de-pend on G-protein-coupled receptors (GPCRs)1 and coupledsignaling pathways. Sweet taste in large part depends on aheterodimeric receptor comprised of T1R2�T1R3 (2–5).
The T1R taste receptors (T1R1, T1R2, and T1R3) are mostclosely related to metabotropic glutamate receptors (mGluRs),Ca2�-sensing receptors (CaSRs), and some pheromone recep-tors (6–10). All of these receptors are class-C GPCRs, with the
large clam shell-shaped extracellular amino-terminal domain(ATD) characteristic of this family. Following the ATD is acysteine-rich region that connects the ATD to the heptahelicaltransmembrane domain (TMD); following the TMD is a shortintracellular carboxyl-terminal tail. The solved crystal struc-ture of the ATD of mGluR1 identifies a “Venus flytrap module”(VFTM) involved in ligand binding (11). The canonical agonistglutamate binds within the VFTM in a cleft formed by the twolobes of this module to stabilize a closed active conformation ofthe mGluR1 ATD. In contrast, several positive and negativeallosteric modulators of class-C GPCRs have been identifiedand shown to act via binding not within the VFTM but insteadwithin the TMD (12–16).
Over the past few decades, multiple models of the sweetreceptor’s hypothetical ligand binding site have been generatedbased on the structures of existing sweeteners but withoutdirect knowledge of the nature of the sweet receptor itself. Aconsensus feature of these models is the presence of A-H-Bgroups, in which the AH group is a hydrogen donor and the Bgroup is an electronegative center. These models have explan-atory and predictive value for some, but not all sweeteners,suggesting that not all sweet ligands bind in the same mannerto the receptor. Based on homology to mGluR1, the canonicalligand binding site of the sweet receptor is likely to reside inthe cleft between lobes 1 and 2 of the ATD of T1R2. However,additional binding sites may exist elsewhere in T1R2 or in theT1R3 component of the heterodimeric sweet taste receptor. Thecysteine-rich region of human T1R3 (hT1R3), which has beenshown to be critical for determining responses to intenselysweet proteins, brazzein and monellin (17), is one such poten-tial binding site. Xu et al. (18) and we (19) have recently shownthat mutations in VFTM of human T1R2 (hT1R2) affect thesweet receptor’s responses to aspartame, whereas mutations inthe TMD affect receptor responses to cyclamate.
Lactisole, an arakyl carboxylic acid composed of a carboxylgroup and a hydrophobic phenoxyl group, inhibits sweet andumami taste perception by humans but not by rats (20). Here,we report that lactisole inhibits the hT1R2�hT1R3 humansweet taste receptor by binding to the TMD of hT1R3. Thecarboxyl group and aromatic ring of lactisole are necessary forthe suppression of sweet taste. In addition, we have identified7 critical residues within the TMD of T1R3 that form a poten-tial binding pocket for lactisole and are responsible for thesweet receptor’s sensitivity to lactisole. We present a model inwhich lactisole is docked to a potential binding site within theTMD of T1R3.
EXPERIMENTAL PROCEDURES
Materials—Acesulfame-K, cyclamate, D-tryptophan, neohesperidindihydrochalcone (NHDC), saccharin, sucrose, thaumatin, and lactisolewere obtained from Sigma. Sucralose was obtained from McNeil Spe-
* This work was supported in part by National Institute of HealthGrants 1 F32 DC007021-01 (to P. J.), DC003055 and DC003155 (toR. F. M), and MH58811 (to M. M.). The costs of publication of thisarticle were defrayed in part by the payment of page charges. Thisarticle must therefore be hereby marked “advertisement” in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
□S The on-line version of this article (available at http://www.jbc.org)contains four supplementary figures.
�An Associate Investigator of Howard Hughes Medical Institute.** To whom correspondence should be addressed: Dept. of Physiology
and Biophysics, Mount Sinai School of Medicine, Box 1677, 1425 Mad-ison Ave., New York, NY 10029. Tel.: 212-659-8612; Fax: 212-849-2599;E-mail: [email protected].
1 The abbreviations used are: GPCR, G-protein-coupled receptors;CaSR, Ca2�-sensing receptor; mGluR, metabotropic glutamate recep-tor; VFTM, Venus flytrap module; ATD, amino-terminal domain; TM,transmembrane; TMD, TM domain; NHDC, neohesperidin dihydro-chalcone; HEK, human embryonic kidney; HEK293E, HEK293 EBNA;EBNA, Epstein-Barr virus nuclear antigen; DPBS, Dulbecco’s phos-phate-buffered saline; h, human; m, mouse.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 280, No. 15, Issue of April 15, pp. 15238–15246, 2005© 2005 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org15238
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from

cialty (New Brunswick, NJ). Brazzein was a gift from Dr. Goran Helle-kant (see Supplemental Fig. 1 for structures of sweet ligands). Unlessnoted otherwise, acesulfame-K, cyclamate, and D-tryptophan were usedat 10 mM final concentration, NHDC at 0.25 mM, saccharin at 1 mM,sucralose at 1 mM, sucrose at 100 mM, brazzein at 0.25%, thaumatin at0.1%, and lactisole at 1.25 mM.
Residue Numbering—The general numbering of residues in the TMDof T1R3 follows T1R3’s primary sequence. Superscripted residue numbersfollow the generic numbering system of Ballesteros and Weinstein (21).
Preparation of Chimeras and Point Mutations—Human and mouseT1R2 and T1R3 clones were as described (17). The G�16-gust44 con-struct was as described (22). In preliminary studies, the G�16-gust44construct gave larger and/or more reliable responses than did G�16-i3;therefore, G�16-gust44 was used in all experiments. Human/mousechimeras of T1R3 were constructed using an overlapping PCR strategyas described (17). The integrity of all DNA constructs was confirmed bysequencing. Point mutations in T1R clones were made by the sameoverlapping PCR strategy or by site-directed mutagenesis (Stratagene,La Jolla, CA). Combinations of hT1R2 with the following chimeras werenot functional (data not shown): mT1R3.h.712–852, mT1R3.h.729–852,mT1R3.h.751–852, and mT1R3.h.797–852.
Heterologous Expression—HEK293 EBNA (HEK293E) cells werecultured at 37 °C in Dulbecco’s modified Eagle’s medium supplementedwith 10% fetal bovine serum and transfected as described (17). Cells forcalcium imaging were seeded onto 6-well plates (Corning); cells in eachwell were co-transfected using Mirus TransIT-293 (Panvera, Madison,WI) with plasmid DNAs encoding T1Rs (0.6 �g of each) and G�16-gust44(0.5 �g). After 24 h, the transfected cells were trypsinized and seededonto 96-well assay plates (Corning, Corning, NY) at about 40,000 cells/well in low glucose Dulbecco’s modified Eagle’s medium (Invitrogen)supplemented with 10% dialyzed fetal bovine serum (Invitrogen) and1� GlutaMax-1 (Invitrogen). After an additional 24 h, the cells werewashed once with Dulbecco’s phosphate-buffered saline (DPBS), loadedwith 75 �l of 3 �M Fluo-4 (Molecular Probe) in DPBS, incubated for 2 h,and then washed twice with DPBS and maintained at 37 °C in 50 �l ofDPBS. The dye-loaded transfected cells in plates were placed into aFlexStation II system (Molecular Devices) to monitor fluorescence (ex-citation, 488 nm; emission, 525 nm; cutoff, 515 nm) change after theaddition of 50 �l of DPBS supplemented with 2� tastants. For eachtrace, tastant or tastant � lactisole was added 30 s after the start of thescan, scanning continued for an additional 150 s, and data were col-lected every 2 s.
Data Analysis—After obtaining a calcium mobilization trace for eachsample, calcium mobilization in response to tastants or tastant � lactisolewas quantified as the percentage of change (peak fluorescence � baselinefluorescence level, denoted as �F) from its own baseline fluorescence level(denoted as F). Peak fluorescence intensity occurred about 20–30 s afterthe addition of tastants. As controls, buffer alone or compounds that donot taste sweet evoked no change of fluorescence (�F/F � 0, S.E. is about1%). The data were expressed as the mean � S.E. of the �F/F value ofthree independent samples, similarly presented in Jiang et al. (17). Theanalysis was done automatically by an in-house written SAS program.The bar graph and curving-fitting routines were carried out using Graph-Pad Prism 3.0 (GraphPad Software, Inc.).
Homology Modeling—The multiple sequence alignment among T1Rs,hT1R2, hT1R3, mT1R2, mT1R3, mGluR1, and bovine rhodopsin wasgenerated by the ClustalW program (23) with some manual adjust-ments to the non-homologous regions (see Supplemental Fig. 2, multi-ple sequence alignment). The sequence identities (similarities) for TMhelices 1–7 between hT1R3 and bovine rhodopsin are 22.6% (61.3%),13.3% (40%), 22.6% (48.4), 25.0% (33.3%), 11.1% (40.7%), 11.8%(61.8%), and 16% (60%), respectively. Based on the sequence alignment,the hT1R3 TM homology model was constructed by residue replacementusing the InsightII (Accelrys, San Diego, CA) biopolymer module. Thenon-conserved proline kink within TM helix 6 of the model was built bymanually shifting 3 residues according to the template. Extracellularloop 3 between TM helix 6 and TM helix 7 was generated by ab initioloop prediction (24, 25). The other intracellular and extracellular loopregions were generated by the modeler program (26, 27) based on thebovine rhodopsin template. The side chains of the residues in the 7 TMregion of hT1R3 and all the loops were refined by the following proce-dure: 500 steps of steepest descent followed by 2 ps of heating from 0 to300 K, and 20 ps equilibration and 100 ps of production run at 300 K torelax possible bad contacts of the model followed by energy minimiza-tion 1000 steps of steepest descent and 10,000 steps of adopted basisNewton-Raphson or until the root mean square energy gradient of 0.05kcal/mol/Å was achieved. During the simulation, the �-carbon atoms ofthe helixes were fixed to avoid possible distortion, and the distance-de-
pendent dielectric constant (� � 4Rij) was used to mimic solvationeffects and the membrane environment. All model refinements werecarried out by the CHARMM program (28) using the CHARMM27all-hydrogen force field.
Molecular Docking—The geometry of lactisole (S(-)2-(4-methoxyphe-noxy)-propanoic acid) was fully optimized by the ab initio quantumchemistry method at the HF/6–31G* level and followed by a singlepoint calculation with the polarized continuum model solvation methodto obtain the electrostatic potentials using the Gaussian98 package(29). Then, the CHELPG charge-fitting scheme (30) was used to calcu-late partial charges for lactisole. The missing force field parameters forlactisole were obtained from similar parameters taken fromCHARMM27 or Quanta (Accelrys, San Diego, CA).
Lactisole was docked into the pocket of the TM region of hT1R3 usingthe automatic docking program autodock V3.0.5 (31), which uses apowerful Lamarckian genetic algorithm for conformational samplingand docking. The docked conformations of lactisole were analyzed bythe cluster analysis of the autodock. The final docked conformation wasselected by comparing the available mutagenesis results followed bysome manual adjustments of the positions of lactisole and the sidechains of hT1R3 before employing model refinement by MD simula-tions. A similar MD protocol was used for the docked lactisole-hT1R3complex structure refinement that except the �-carbon atoms of thehelixes were restricted by 1.0 kcal/mol/Å2 harmonic restraint forceinstead of fixing the �-carbon atoms during the simulations.
RESULTS
Inhibition of Sweet Taste by Lactisole Requires hT1R3—Inhuman psychophysical studies, lactisole inhibits the sweettaste of sucrose, saccharin, and several other compounds (20,32, 33). In vitro lactisole inhibits sucrose-induced responses ofcells heterologously expressing hT1R2�hT1R3 (2). We exam-ined lactisole’s effect on in vitro responses of hT1R2�hT1R3 toa group of chemically diverse sweeteners (Fig. 1A). Lactisoleinhibited the [Ca2�]i responses of hT1R2�hT1R3-expressingHEK293E cells to sucrose (disaccharide), D-tryptophan (D-a-mino acid), cyclamate (sulfamate), saccharin, acesulfame-K (N-sulfonylamides), NHDC (a dihydrochalcone), sucralose (chlori-nated disaccharide), and brazzein and thaumatin (proteins).
Lactisole suppresses sweetness in humans but not in mice.To determine whether one or both components of the het-erodimeric human sweet receptor is required for sensitivity tolactisole, we tested the responses of human, mouse and hu-man � mouse mismatched heterodimers to D-tryptophan withand without lactisole (Fig. 1B). As expected, the fully humanpair (hT1R2�hT1R3) was sensitive to lactisole, whereas thefully mouse pair (mT1R2�mT1R3) was not. One mismatchedpair (hT1R2�mT1R3) behaved like the fully mouse het-erodimer, showing no sensitivity to lactisole. As reported pre-viously (17), the other mismatched pair (mT1R2�hT1R3) doesnot produce a functional receptor, precluding us from analyzingits sensitivity to lactisole. These results demonstrate thathT1R3 is required for lactisole sensitivity. Below, by usinghuman/mouse chimeras of T1R3, we show that hT1R2 is notrequired for sensitivity to lactisole (Fig. 2).
Lactisole Acts on the TMD of hT1R3—To identify the portionof hT1R3 required for the sweet receptor’s sensitivity to lacti-sole, we examined the responses of heterodimeric receptorscomprised of hT1R2 plus human/mouse chimeras in which wecombined varying portions of hT1R3 with the complementaryportion of mT1R3 (Fig. 2A). Heterodimers of hT1R2 plus T1R3chimeras containing most or all of the extracellular region ofhT1R3 coupled to the TMD and carboxyl-terminal tail ofmT1R3 (i.e. h.1–547.mT1R3 and h.1–567.mT1R3) responded toD-tryptophan but were not inhibited by lactisole (Fig. 2B). Incontrast, heterodimers of hT1R2 plus T1R3 chimeras contain-ing most or all of the extracellular region of mT1R3 coupled tothe TMD and carboxyl-terminal tail of hT1R3 (i.e.mT1R3.h.548–852 and mT1R3.h.568–852) responded toD-tryptophan and were inhibited by lactisole. These results
Lactisole Inhibits the Sweet Taste Receptor 15239
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from
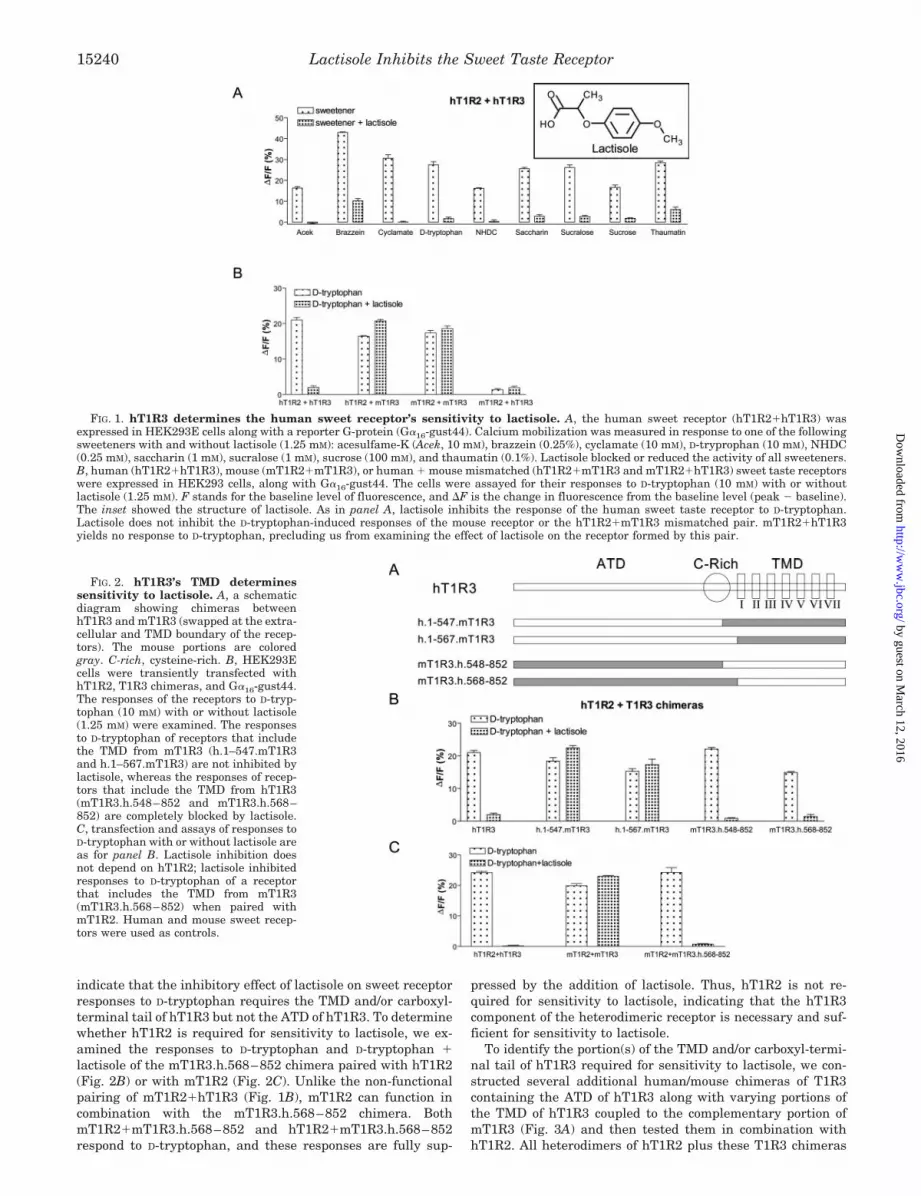
indicate that the inhibitory effect of lactisole on sweet receptorresponses to D-tryptophan requires the TMD and/or carboxyl-terminal tail of hT1R3 but not the ATD of hT1R3. To determinewhether hT1R2 is required for sensitivity to lactisole, we ex-amined the responses to D-tryptophan and D-tryptophan �lactisole of the mT1R3.h.568–852 chimera paired with hT1R2(Fig. 2B) or with mT1R2 (Fig. 2C). Unlike the non-functionalpairing of mT1R2�hT1R3 (Fig. 1B), mT1R2 can function incombination with the mT1R3.h.568–852 chimera. BothmT1R2�mT1R3.h.568–852 and hT1R2�mT1R3.h.568–852respond to D-tryptophan, and these responses are fully sup-
pressed by the addition of lactisole. Thus, hT1R2 is not re-quired for sensitivity to lactisole, indicating that the hT1R3component of the heterodimeric receptor is necessary and suf-ficient for sensitivity to lactisole.
To identify the portion(s) of the TMD and/or carboxyl-termi-nal tail of hT1R3 required for sensitivity to lactisole, we con-structed several additional human/mouse chimeras of T1R3containing the ATD of hT1R3 along with varying portions ofthe TMD of hT1R3 coupled to the complementary portion ofmT1R3 (Fig. 3A) and then tested them in combination withhT1R2. All heterodimers of hT1R2 plus these T1R3 chimeras
FIG. 1. hT1R3 determines the human sweet receptor’s sensitivity to lactisole. A, the human sweet receptor (hT1R2�hT1R3) wasexpressed in HEK293E cells along with a reporter G-protein (G�16-gust44). Calcium mobilization was measured in response to one of the followingsweeteners with and without lactisole (1.25 mM): acesulfame-K (Acek, 10 mM), brazzein (0.25%), cyclamate (10 mM), D-tryprophan (10 mM), NHDC(0.25 mM), saccharin (1 mM), sucralose (1 mM), sucrose (100 mM), and thaumatin (0.1%). Lactisole blocked or reduced the activity of all sweeteners.B, human (hT1R2�hT1R3), mouse (mT1R2�mT1R3), or human � mouse mismatched (hT1R2�mT1R3 and mT1R2�hT1R3) sweet taste receptorswere expressed in HEK293 cells, along with G�16-gust44. The cells were assayed for their responses to D-tryptophan (10 mM) with or withoutlactisole (1.25 mM). F stands for the baseline level of fluorescence, and �F is the change in fluorescence from the baseline level (peak � baseline).The inset showed the structure of lactisole. As in panel A, lactisole inhibits the response of the human sweet taste receptor to D-tryptophan.Lactisole does not inhibit the D-tryptophan-induced responses of the mouse receptor or the hT1R2�mT1R3 mismatched pair. mT1R2�hT1R3yields no response to D-tryptophan, precluding us from examining the effect of lactisole on the receptor formed by this pair.
FIG. 2. hT1R3’s TMD determinessensitivity to lactisole. A, a schematicdiagram showing chimeras betweenhT1R3 and mT1R3 (swapped at the extra-cellular and TMD boundary of the recep-tors). The mouse portions are coloredgray. C-rich, cysteine-rich. B, HEK293Ecells were transiently transfected withhT1R2, T1R3 chimeras, and G�16-gust44.The responses of the receptors to D-tryp-tophan (10 mM) with or without lactisole(1.25 mM) were examined. The responsesto D-tryptophan of receptors that includethe TMD from mT1R3 (h.1–547.mT1R3and h.1–567.mT1R3) are not inhibited bylactisole, whereas the responses of recep-tors that include the TMD from hT1R3(mT1R3.h.548–852 and mT1R3.h.568–852) are completely blocked by lactisole.C, transfection and assays of responses toD-tryptophan with or without lactisole areas for panel B. Lactisole inhibition doesnot depend on hT1R2; lactisole inhibitedresponses to D-tryptophan of a receptorthat includes the TMD from mT1R3(mT1R3.h.568–852) when paired withmT1R2. Human and mouse sweet recep-tors were used as controls.
Lactisole Inhibits the Sweet Taste Receptor15240
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from

responded to D-tryptophan (Fig. 3B). The heterodimer of hT1R2plus the T1R3 chimera containing the extracellular domainand entire TMD from hT1R3 (i.e. hT1R2�h.1–812.mT1R3)showed inhibition by lactisole comparable with that obtainedwith hT1R2�hT1R3. Heterodimers of hT1R2 with T1R3chimeras containing the hT1R3 ATD and hT1R3 TM helices1–6 (i.e. hT1R2�h.1–787.mT1R3) or TM helices 1–5 (i.e.hT1R2�h.1–751.mT1R3) showed moderate inhibition by lacti-sole. Heterodimers of hT1R2 with T1R3 chimeras containingthe hT1R3 ATD and hT1R3 TM helices 1–2 (i.e. hT1R2�h.1–638.mT1R3), TM helices 1–3 (i.e. hT1R2�h.1–669.mT1R3), orTM helices 1–4 (i.e. hT1R2�h.1–711.mT1R3 or hT1R2�h.1–729.mT1R3) showed no inhibition by lactisole. These resultsindicate that the inhibitory effect of lactisole on sweet receptorresponses to D-tryptophan requires human-specific residueswithin or adjacent to human TM helices 5, 6, and 7. Thedifference in lactisole sensitivity displayed by chimeras h.1–787.mT1R3 and h.1–812.mT1R3 indicates that residues withinor adjacent to human TM helix 7 in particular are required forsensitivity to lactisole. Thus, there are multiple human-specificresidues required for lactisole sensitivity within the regioncontaining TM helices 5–7.
T1R3 chimeras in which we replaced TM helices 5 and 6 orTM helix 5 alone of hT1R3 with the corresponding TM helicesof mT1R3 (i.e. hT1R3(m734–792) and hT1R3(m734–756), re-spectively) functioned in combination with hT1R2 to respond toD-tryptophan but displayed no sensitivity to lactisole (Fig. 3C).Conversely, T1R3 chimeras in which we replaced TM helices 5and 6 or TM5 alone of mT1R3 with the corresponding TM
helices of hT1R3 (i.e. mT1R3(h729–787) and mT1R3(h729–751), respectively) functioned in combination with hT1R2 torespond to D-tryptophan and were sensitive to lactisole (Fig.3C). Thus, at least some residues required for the human-specific sensitivity to lactisole lie within or adjacent to TM helix5 of hT1R3.
Alanine 7335.46 in hT1R3 TM Helix 5 Is Required for Sensi-tivity to Lactisole—In the TM helix 5 region of hT1R3 (aminoacids 729–751), only 4 residues (Phe-7305.43, Ala-7335.46, Ala-7355.48, and Thr-7395.52) differ between the human and mouseforms of T1R3. To identify which residue(s) within this regionare required for sensitivity to lactisole, we individually substi-tuted each of these 4 residues with the corresponding mouseT1R3 residues (hT1R3(F7305.43L), hT1R3(A7335.46V),hT1R3(A7355.48I), and hT1R3(T7395.52M)). We then examinedthe responses of heterodimers of hT1R2 with these substitutedforms of hT1R3. All mutants responded to D-tryptophan; how-ever, only the A7335.46V mutant showed diminished sensitivityto lactisole (Fig. 4A). In comparison with wild type hT1R3, theA7335.46V mutant showed a right shift in its dose-responsecurve to lactisole (A7335.46V mutant IC50 of 1.9 � 10�3 M
versus wild type IC50 of 4.1 � 10�5 M) (Fig. 4D) but wasunchanged in its dose-response curve to D-tryptophan(A7335.46V mutant EC50 of 3.5 mM versus wild type EC50 of 4.3mM) (Fig. 4C). Thus, valine substitution at position 7335.46
selectively affects the receptor’s sensitivity to lactisole.As noted above, the results with chimeras h.1–787.mT1R3
and h.1–812.mT1R3 (Fig. 3B) indicate that residues within oradjacent to human TM helix 7 are required for maximal sensi-
FIG. 3. TMD 5–7 from hT1R3 speci-fies lactisole sensitivity. A, a schematicdiagram showing chimeras made betweenhT1R3 and mT1R3 within the TDM. Themouse portions are colored gray. C-rich,cysteine-rich. B, hT1R2 and T1R3 chime-ras were expressed in HEK293E cells,along with G�16-gust44. Receptors con-taining T1R3 chimeras in which humansequences make up TM helices 1–2(h.1–638.mT1R3), TM helices 1–3 (h.1–669.mT1R3), or TM helices 1–4 (h.1–711.mT1R3, and h.1–729.mT1R3) are in-sensitive to lactisole. Receptorscontaining T1R3 chimeras in which hu-man sequences make up TM helices 1–5(h.1–751.mT1R3) or TM helices 1–6 (h.1–787.mT1R3) are partially sensitive to lac-tisole. Receptors containing T1R3 chime-ras in which human sequences make upTM helices 1–7, but lack the human car-boxyl terminus (h.1–812.mT1R3), arefully sensitive to lactisole. C, transfectionand assays of responses to D-tryptophanwith or without lactisole are as for panelB. Receptors containing human/mouseT1R3 chimeras in which human se-quences make up all but TM helix 5(hT1R3(m734–756) or TM helices 5–6(hT1R3(m734–792) are insensitive to lac-tisole. Receptors containing human/mouse T1R3 chimeras in which humansequences make up only TM helix 5(mT1R3(h729–751) or TM helices 5–6(mT1R3(h729–787) are sensitive tolactisole.
Lactisole Inhibits the Sweet Taste Receptor 15241
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from
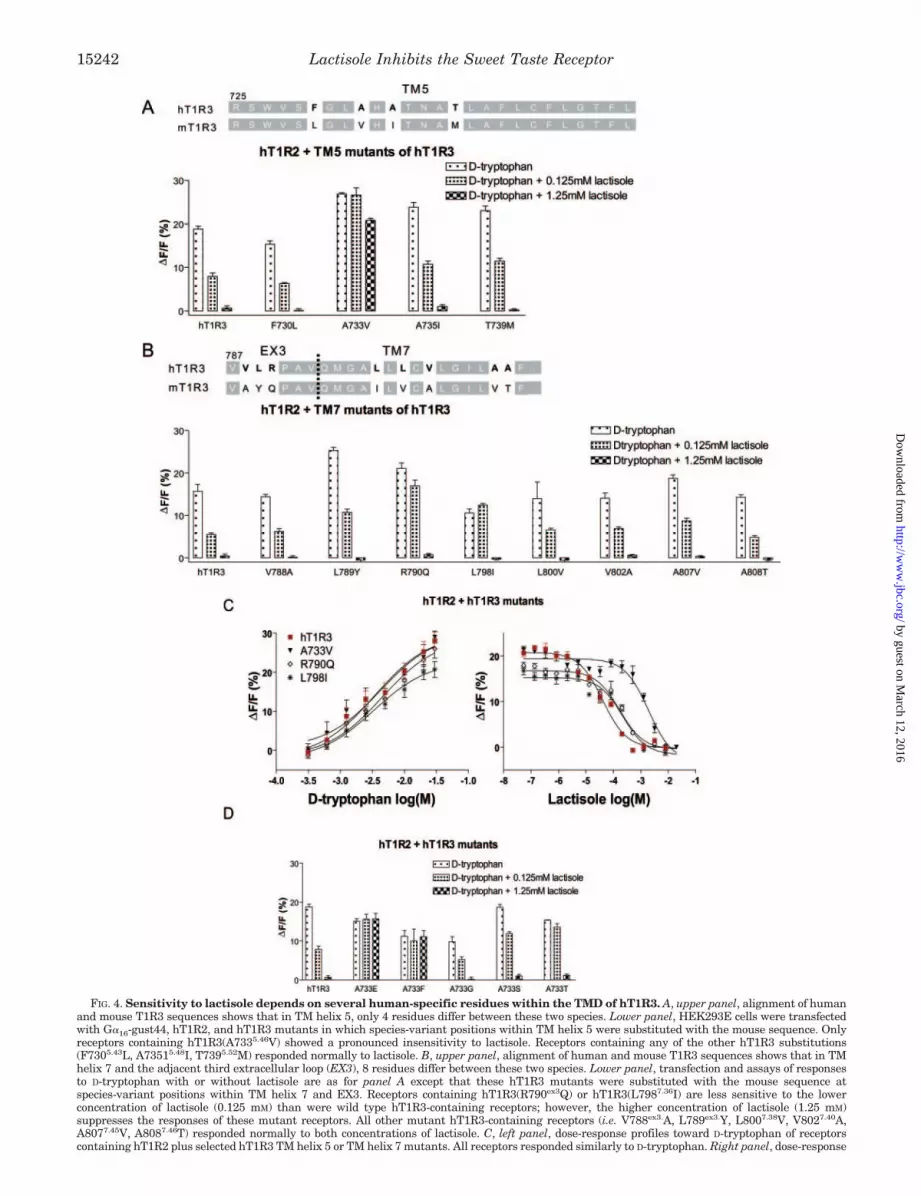
FIG. 4. Sensitivity to lactisole depends on several human-specific residues within the TMD of hT1R3. A, upper panel, alignment of humanand mouse T1R3 sequences shows that in TM helix 5, only 4 residues differ between these two species. Lower panel, HEK293E cells were transfectedwith G�16-gust44, hT1R2, and hT1R3 mutants in which species-variant positions within TM helix 5 were substituted with the mouse sequence. Onlyreceptors containing hT1R3(A7335.46V) showed a pronounced insensitivity to lactisole. Receptors containing any of the other hT1R3 substitutions(F7305.43L, A73515.48I, T7395.52M) responded normally to lactisole. B, upper panel, alignment of human and mouse T1R3 sequences shows that in TMhelix 7 and the adjacent third extracellular loop (EX3), 8 residues differ between these two species. Lower panel, transfection and assays of responsesto D-tryptophan with or without lactisole are as for panel A except that these hT1R3 mutants were substituted with the mouse sequence atspecies-variant positions within TM helix 7 and EX3. Receptors containing hT1R3(R790ex3Q) or hT1R3(L7987.36I) are less sensitive to the lowerconcentration of lactisole (0.125 mM) than were wild type hT1R3-containing receptors; however, the higher concentration of lactisole (1.25 mM)suppresses the responses of these mutant receptors. All other mutant hT1R3-containing receptors (i.e. V788ex3.A, L789ex3.Y, L8007.38V, V8027.40A,A8077.45V, A8087.46T) responded normally to both concentrations of lactisole. C, left panel, dose-response profiles toward D-tryptophan of receptorscontaining hT1R2 plus selected hT1R3 TM helix 5 or TM helix 7 mutants. All receptors responded similarly to D-tryptophan. Right panel, dose-response
Lactisole Inhibits the Sweet Taste Receptor15242
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from

tivity to lactisole. In the extracellular loop 3 and TM helix 7region of hT1R3 (amino acids 787–812), 8 residues (Val-788ex3,Leu-789ex3, Arg-790ex3, Leu-7987.36, Leu-8007.38, Val-8027.40,Ala-8077.45, and Ala-8087.46) differ between the human andmouse forms of T1R3. To identify which residue(s) within thisregion are required for sensitivity to lactisole, we individuallysubstituted each of these 8 residues with the correspondingmouse T1R3 residues (hT1R3(V788ex3A), hT1R3(L789ex3Y),hT1R3(R790ex3Q), hT1R3(L7987.36I), hT1R3(L8007.38V),hT1R3(V8027.40A), hT1R3(A8077.45V), and hT1R3(A8087.46T))and then examined the responses of these substituted forms ofhT1R3 heterodimerized with hT1R2. All of these mutants re-sponded to D-tryptophan; only the R790ex3Q and L7987.36I mu-tants showed diminished sensitivity to lactisole (Fig. 4B). Bothof these mutants were unchanged in their dose-response curvesto D-tryptophan (Fig. 4C) and right-shifted in their dose-re-sponse curves to lactisole (Fig. 4C). The effect on lactisolesensitivity of mutating either of these residues was much lesspronounced than that seen with A7335.46V (R790ex3Q mutantIC50 of 1.7 � 10�4 M, L7987.36I mutant IC50 of 2.2 � 10�4 M).Thus, substitutions at positions 733 (TM helix 5), in particular,790 (third extracellular domain) and 798 (TM helix 7), all affectthe receptor’s sensitivity to lactisole.
Ala-7335.46 Substitutions Affect hT1R3 Responses to Lacti-sole—To investigate the physicochemical effects of residue Ala-7335.46 within TM helix 5 of hT1R3 on receptor activity towardlactisole, we made various substitutions at this position (Fig.4D). Substitution of Ala-7335.46 by glutamate or phenylalanineabolished the receptor’s sensitivity to lactisole. Substitution ofAla-7335.46 by glycine or serine did not affect the receptor’ssensitivity toward lactisole. The threonine substitution(A7335.46 T) led to slightly reduced receptor sensitivityto lactisole.
Furthermore, we substituted mT1R3 Val-7385.46, Gln-795EX3, and Ile-8037.36 with human corresponding residuesAla, Arg, and Leu either individually or in combination to testwhether this would make the mutants mT1R3 sensitive tolactisole. Indeed, mT1R3(V7385.46A) showed sensitivity to lac-tisole (Supplemental Fig. 3).
hT1R3 Sensitivity to Lactisole Also Involves Residues in TMHelices 3 and 6—The solved structure of rhodopsin has beenused as a template, after sequence alignment, to model thestructures of the TMDs of certain family C receptors (mGluR1,mGluR5, and CaSR (13–15, 34)). Such models can be validatedby mutating residues predicted to be within binding pocketsfollowed by characterization of the mutant receptor’s proper-ties. Rhodopsin’s retinol binding pocket contains 17 residues,all of which are within 6.0 Å of retinol in the rhodopsin crystalstructure. Based on the alignment of T1R3 with rhodopsin, wepicked 17 residues lining a potential binding pocket of T1R3.Each of these residues was substituted with alanine, and themutants were tested for responsiveness to D-tryptophan andlactisole (Fig. 5A).
Receptors with alanine substitutions at any of five positions(Leu-6443.36, Thr-6453.37, Tyr-7716.44, Gln-7947.32, andIle-8057.43) showed no response to D-tryptophan (Fig. 5A) or toother sweeteners such as aspartame (data not shown). Thesepositions might be required for generating the active state ofthe receptor or for the structural integrity of the receptor.
Because these mutants did not respond to D-tryptophan, theycould not be tested for sensitivity to lactisole. Of the remaining12 mutants, Gln-6363.28, Ser-6403.32 His-6413.33, His-721ex2–49,Arg-723ex2–51, Ser-7295.42, Val-7766.49, Phe-7786.51,Val-7796.52, and Leu-7826.55 had normal to somewhat dimin-ished responses to D-tryptophan; the other two mutants (His-7345.47, Trp-7756.48) had severely diminished responses to D-tryptophan (Fig. 5A). Responses to lactisole of these mutantsvaried from near wild type (Gln-6363.28, His-721ex2–49, Arg-723ex2–51, Ser-7295.42, Val-7766.49) to diminished (Val-7796.52)or absent sensitivity (His-6413.33, Phe-7786.51) to enhanced sen-sitivity (Ser-6403.32, Leu-7826.55) (Fig. 5A). H7345.47A andW7756.48A showed such severely diminished responses to D-tryptophan that it was impossible to unequivocally evaluatelactisole’s effect on these mutants, although the responses ofboth appeared to be diminished.
Dose-response curves to D-tryptophan and lactisole (in thepresence of 10 mM D-tryptophan) were obtained for those mu-tants with marked changes in sensitivity to lactisole (i.e. His-6413.33, Phe-7786.51, Ser-6403.32, Leu-7826.55) (Fig. 5B). Re-sponses to D-tryptophan of these four mutants were comparablewith that of wild type (Fig. 5B). Responses to lactisole of thesemutants were severely diminished (F7786.51A, IC50 1.8 � 10�3
M), completely absent (H6413.33A), or enhanced (S6403.32A andL7826.55A, IC50 1.4 � 10�5 M and 1.2 � 10�5 M, respectively)(Fig. 5B). Thus, these four positions are likely components ofthe lactisole binding pocket.
Molecular Modeling of hT1R3’s TMD—We molecularly mod-eled the TM region of wild type hT1R3 and automaticallydocked lactisole to the predicted binding pocket (see “Experi-mental Procedures”). The optimal structure of hT1R3 was ob-tained after building the structure using rhodopsin as a tem-plate (35), manual adjustment of the potential kink introducedby Pro-7806.53 in TM helix 6, and energy minimization. Lacti-sole is a racemic mixture, with only the S-form active as asweetness suppressor.2 Therefore, we used the S-form of lacti-sole in our docking experiment. After running the AUTODOCKprogram, we found that lactisole can take on a few conforma-tional states when automatically docked into the TMD bindingpocket. By comparing the possible models with our mutagene-sis data, we found that only one of the docked structures couldsatisfy the constraints placed on it by the mutagenesis data(Fig. 6). In this model, His-6413.33 is 3 Å from the carboxylicgroup of S-lactisole, Phe-7786.51 is close to the phenoxyl ring ofS-lactisole, and the methoxyl group of S-lactisole is pointingtoward Ala-7335.46. From this model, we also see that Gln-637is close to the carboxylic group of S-lactisole. To further test ourmodel, we mutated Gln-6373.29 to glutamate to test whetherthe negatively charged group has a detrimental effect on lacti-sole’s activity. Indeed, this mutant was devoid of lactisole-induced suppression of D-tryptophan activity (SupplementalFig. 4).
DISCUSSION
Lactisole Broadly Inhibits Human Sweet Taste—Lactisole, anarakyl carboxylic acid, inhibits the perception of sweet andumami taste in humans but not in rodents (20). In humans,
2 Gordon Birch, personal communication.
profiles toward D-tryptophan (10 mM) plus varying amounts of lactisole of receptors containing hT1R2 plus selected hT1R3 TM helix 5 or TM helix7 mutants. All receptors containing hT1R3 mutants were right-shifted in their sensitivity to lactisole, and the A7335.46V mutant was mostprofoundly affected (IC50s to lactisole are: 1.3 � 10�3 M (A7335.46V), 1.7 � 10�4 M (R790ex3Q), and 2.2 � 10�4 M (L7987.36I). D, additional hT1R3mutants were generated in which Ala-7335.46 within TM helix 5 was substituted to Gly, Glu, Phe, Ser, and Thr. Transfection and assays ofresponses to D-tryptophan with or without lactisole are as for panel A. Receptors containing hT1R3 mutants showed a range of sensitivity tolactisole from normal (hT1R3(A7335.46G)) to slightly diminished (hT1R3(A7335.46S) and hT1R3(A7335.46T) show reduced responsiveness at 0.125mM lactisole) to completely insensitive (hT1R3(A7335.46E) and hT1R3(A7335.46F)).
Lactisole Inhibits the Sweet Taste Receptor 15243
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from

lactisole inhibits the perception of sweetness of many, but not all,sweeteners (32). Schiffman et al. (32) observed that lactisolereduced the sweet intensity of 12 of 15 sweeteners tested, includ-ing three sugars (sucrose, fructose, and glucose), two terpenoidglycosides (rebaudioside-A and stevioside), two dipeptide deriva-tives (alitame and aspartame), two N-sulfonylamides (acesul-fame-K and saccharin), two polyhydric alcohols (mannitol andsorbitol), and one sulfamate (sodium cyclamate). Lactisole hadlittle or no effect on the perceived sweetness of monoammoniumglycyrrhizinate (a terpenoid glycoside), neohesperidin dihydroch-alcone (a dihydrochalcone), or thaumatin (a protein).
Lactisole’s inhibition of both sweet and umami taste suggeststhat it might target a shared component of these two signalingpathways, e.g. T1R3. Consistent with this inference, prelimi-nary in vitro studies have shown that lactisole inhibits thesucrose-induced responses of cells heterologously expressinghT1R2�hT1R3 (2). More recently, and independently of thepresent study, Xu et al. (18) have determined that lactisole’seffects require the human form of T1R3’s TMD and/or cytoplas-mic domain but do not require human-specific elements withinT1R3’s ATD. These authors did not narrow down the specificregion within the TMD/cytoplasmic domain that serves as lac-tisole’s binding pocket.
Lactisole Acts on hT1R3’s TMD—In the present study, we setout to molecularly identify the target of lactisole’s broadly
acting inhibitory effects on human sweet taste, to determinewhy humans but not mice are susceptible to lactisole’s effects,and to understand mechanistically how lactisole inhibits sweettaste. First, we used a cell-based assay to determine that lac-tisole acts to inhibit heterologously expressed human sweettaste receptor (hT1R2�hT1R3) responses to all members of apanel of chemically diverse sweeteners. Using this same assay,we determined that lactisole does not inhibit the responses ofthe mouse sweet taste receptor (mT1R2�mT1R3), consistentwith its in vivo ineffectiveness in mice. Next, by comparingdifferences in sensitivity toward lactisole among interspecies(mouse � human) combinations of T1R2�T1R3, we demon-strated that lactisole’s target is hT1R3; either hT1R2 or mT1R2in combination with the appropriate T1R3 partner (hT1R3 orits chimeras) equivalently supports sensitivity to lactisole.Then, using human/mouse chimeras of T1R3 in combinationwith T1R2, we determined that it is the TM domain of hT1R3that specifies sensitivity to lactisole; the species of origin of theextracellular ATD and the C-rich region of T1R3 were irrele-vant to sensitivity to lactisole. Using additional interspecieschimeras, we identified which human TM helices were requiredfor sensitivity to lactisole, and then, using substitution mu-tants, we determined which specific human versus mouse var-iant residues within these TMs mattered. Specifically, Ala-7335.34 in TM helix 5, Leu-7987.36 in TM helix 7, and Arg-790ex3
FIG. 5. Alanine scan of candidate lactisole binding pocket residues in hT1R3’s TMD. A, residues of hT1R3 identified by homology torhodopsin as potentially contributing to the lactisole binding pocket were replaced by alanine. hT1R2 and hT1R3 mutants along with G�16-gust44were co-transfected into HEK293E cells. The responses of the transfected cells to D-tryptophan (10 mM) were examined in the presence or absenceof lactisole (1.25 mM). Mutant-containing receptors that were unresponsive to D-tryptophan (L6443.36A, T6453.37A, Y7716.44A, Q7947.32A, andI8057.43A) could not be tested for sensitivity to lactisole. H6413.33A, H7345.47A, W7756.48A, F7786.51A, and V7796.52A mutant-containing receptorshad reduced or absent sensitivity to lactisole, although the responses of H7345.47A andW7756.51A to D-tryptophan were sufficiently impaired thatquantifying the effect of lactisole on the D-tryptophan response of each of these mutants was not possible. S6403.32A and L7826.55A mutant-containing receptors were more sensitive to lactisole. Q6363.28A, H721Ex2–49A, R723 Ex2–51A, S729 5.42A, and V7766.52A mutant-containing receptorshad lactisole sensitivity comparable with wild type (WT). B, left panel, dose-response curves toward D-tryptophan of receptors containing hT1R2plus selected alanine scan mutants. All receptors responded similarly to D-tryptophan. Right panel, dose-response curves toward D-tryptophan (10mM) plus varying concentrations of lactisole of receptors containing hT1R2 plus selected alanine scan mutants. Receptors containing H641 3.33Alost all sensitivity to lactisole. Receptors containing F778 6.51A showed sensitivity toward lactisole that is greatly right-shifted when compared withthat of wild type (IC50 1.8 � 10�3 M). Receptors containing S6403.32A or L782 6.55A showed enhanced sensitivity toward lactisole (IC50 1.4 � 10�5
M and 1.2 � 10�5 M, respectively).
Lactisole Inhibits the Sweet Taste Receptor15244
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from

in extracellular loop 3 were found to be critically important forthe human-specific sensitivity to lactisole.
Our results with lactisole sensitivity of expressedhT1R2�hT1R3 are generally consistent with the human psy-chophysical studies. The only notable difference was that invitro responses of hT1R2�hT1R3 to neohesperidin dihydro-chalcone and thaumatin were suppressed by lactisole,whereas in vivo, human subjects found the sweetness of thesetwo compounds to be insensitive to lactisole. The reasonfor the discordance is unknown. One intriguing possibilityis that sweet receptors other than, or in addition to,T1R2�T1R3 respond to these two compounds and underlie
the lactisole-insensitive responses seen in psychophysicaltests.
hT1R3’s Lactisole Binding Pocket—To obtain a better mech-anistic understanding of how lactisole interacts with and in-hibits the human sweet taste receptor, we carried out twoadditional sets of studies, both of which depended on modelingthe TMD of hT1R3 based on homology to the known structureof rhodopsin. First, alignment with the retinol binding pocketof rhodopsin was used to identify a potential ligand bindingpocket within hT1R3’s TMD. Then, alanine-scanning mutagen-esis of these potential ligand binding residues in hT1R3 iden-tified 4 residues that had large effects on the sensitivity tolactisole: Ser-6403.32 and His-6413.33 in TM helix 3 and Phe-7786.51 and Leu-7826.55 in TM helix 6.
In the second study, we constructed a homology model ofhT1R3’s entire TMD based on the solved crystal structure ofrhodopsin and used this model to identify a plausible binding sitefor lactisole. Our model predicts that lactisole binds to hT1R3within a pocket formed by TM helices 3, 5, and 6. This modelprovides a framework to explain the results from our mutagen-esis experiments, identify the contributions of residues in each ofthese three TM helices, and make a number of testable predic-tions. For this model to be useful, it needs to explain mechanis-tically the two types of altered responses to lactisole we observedin our mutants: (a) reduced sensitivity to lactisole (seen withmutation of residues His-6413.33, Ala-7335.46, and Phe-7786.51)and (b) increased sensitivity to lactisole (seen with mutation ofresidues Ser-6403.32 and Leu-7826.55).
How might mutations of His-6413.33, Ala-7335.46, and Phe-7786.51 reduce lactisole’s ability to inhibit receptor activity? Atphysiological pH, His-6413.33 within TM3 is likely protonated,and its positive charge is predicted to form a salt bridge withthe negatively charged carboxyl group of lactisole. Alaninesubstitution of His-6413.33 would eliminate this potential saltbridge, explaining this mutant’s total loss of sensitivityto lactisole.
Within TM helix 5, replacement of Ala-7335.46 by valine (themouse equivalent), glutamate, or phenylalanine greatly re-duced the receptor’s sensitivity to lactisole, whereas glycine,serine, or threonine substitutions here had modest or no effecton sensitivity to lactisole. These effects are consistent with ourmodel, which places lactisole’s methoxy group in close proxim-ity with the side chain of Ala-7335.46. Larger aliphatic or aro-matic side chains at this position are likely to reduce sensitivitytoward lactisole by their steric effects. Tolerance of serine andthreonine substitutions suggests that these residues’ hydroxylgroups may have favorable hydrophobic interactions with lac-tisole’s methoxy group. Within TM helix 6, replacement ofPhe-7786.51 (conserved among class-C GPCRs) with alaninegreatly reduced the receptor’s sensitivity to lactisole. Ourmodel shows sufficient proximity of Phe-7786.51’s side chain tothe phenoxyl ring of lactisole for a �-� interaction to occur.
Alanine substitution of Ser-6403.32 or Leu-7826.55A markedlyincreased the sensitivity of the receptor to inhibition by lacti-sole. In our model, Leu-6443.36, Phe-7786.51, and Leu-7826.55
contribute to the lactisole binding pocket via hydrophobic in-teractions with lactisole. The Leu-7826.55A substitution wouldmaintain hydrophobicity and diminish any steric constraints.Replacement of Leu-7826.55 with asparagine or threonineslightly reduced the sensitivity of the receptor to lactisole (datanot shown), consistent with our proposal that decreased hydro-phobicity of this pocket would diminish the strength of theinteraction between the receptor and lactisole.
The Ser-6403.32A substitution may reduce steric constraintsby replacing a polar side group with a smaller hydrophobic one.Smaller side chains at either 6403.32 or 7826.55 may allow
FIG. 6. Molecular model of hT1R3 TMD binding pocket withdocked lactisole. An overall view (upper panel) and a detailed view ofthe pocket (lower panel) of a model of the T1R3 TMD binding pocketwith lactisole docked within the pocket are shown. Note the followingsalient points of the model: lactisole’s negatively charged propionic acidmoiety is in close enough proximity to positively charged His-641 3.33 ofTM helix 3 to form a salt bridge; lactisole’s phenyl ring is close enoughand properly oriented to Phe-778 6.51 of TM helix 6 to form �-� inter-actions; lactisole’s methoxyl group extends into the hydrophobic pocketand faces Ala-733 5.46 of TM helix 5.
Lactisole Inhibits the Sweet Taste Receptor 15245
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from

lactisole to better bridge the pocket and lock it into the groundstate. Consistent with this proposal is our recent observationthat substitution of Ser-6403.32 with valine (similar in size buthydrophobic) decreased sensitivity to lactisole (data not shown)
Out-of-Pocket Expanses—Our model implicates TM helices 3,5, and 6 of hT1R3 in lactisole binding; however, our chimericand mutational studies also identified Leu-7987.36 in TM helix7 and Arg-790ex3 in extracellular loop 3 (which connects TMhelices 6 and 7) as human-specific residues that affect respon-siveness to lactisole. These residues appear to be too far awayfrom the predicted binding pocket to be exerting direct effectson lactisole binding. TM helix 7 and extracellular loop 3 mayaffect agonist/antagonist activity by acting on TM helix 6. Ac-cording to our model, Arg-790ex3 is close enough to Glu-717 ex2
of extracellular loop 2 to form a salt bridge, and this may helpto stabilize the top of the TM bundle and “cap” the lactisolebinding site.
Common Motifs with Other GPCRs—T1R3’s binding pocketfor lactisole shares a great deal of similarity with ligand bindingpockets of many other GPCRs. For instance, the Asp in position3.32 is the principal anchoring residue of all monamine receptors(Asp3.32) and CaSR (15, 36). Phe-6843.32 in CaSR is involved inthe recognition of allosteric modulators (16). Ala-1173.32 and Thr-1183.33 as well as other TM helix 3 residues in rhodopsin makeimportant contacts mainly with the retinylidene group of 11-cisretinal (35). Position 3.33 (Tyr-1063.33) in the thyrotropin-releas-ing hormone receptor forms a hydrogen-bonding interaction withthyrotropin-releasing hormone (37). In rhodopsin, Thr-1183.33,Tyr-2686.51, and Ile-189ex2 determine the position of the C9-methyl group of the retinylidene group (35). Position 6.51 ispivotal for recognition of agonist or antagonist of many GPCRs,including CaSR, mGluR1, rhodopsin, and thyrotropin-releasinghormone receptor (13, 15, 38). Position 6.55 participates in ligandrecognition in serotonin 5-HT2B receptor and M3 muscarinicacetylcholine receptor (ACM3) (39, 40).
How does the heterodimeric sweet receptor transit into theactive form, and how does lactisole block this? In the case ofrhodopsin, the activation mechanism occurs as TM helix 6 tiltsaway from TM helix 3; this results in the cytoplasmic end of TMhelix 6 coming closer to the cytoplasmic end of TM helix 5(41–43). We speculate that a similar activation mechanismholds for the sweet taste receptor. Lactisole’s sweet antagonismcould be mediated by its binding to TM helices 3, 5, and 6 ofhT1R3 so as to restrict movement required for conversion to theactive state. In effect, lactisole locks the receptor into theground state. This would explain how lactisole binding tothe TMD of hT1R3 can “act at a distance” to antagonize sweet-eners, which are thought to bind to the ATD of T1R2.
Acknowledgments—We thank all the members of the Margolskeelaboratory for help and discussion during the course of this study.
REFERENCES
1. Lindemann, B. (2001) Nature 413, 219–2252. Li, X., Staszewski, L., Xu, H., Durick, K., Zoller, M., and Adler, E. (2002) Proc.
Natl. Acad. Sci. U. S. A. 99, 4692–46963. Nelson, G., Hoon, M. A., Chandrashekar, J., Zhang, Y., Ryba, N. J., and Zuker,
C. S. (2001) Cell 106, 381–3904. Zhao, G. Q., Zhang, Y., Hoon, M. A., Chandrashekar, J., Erlenbach, I., Ryba,
N. J., and Zuker, C. S. (2003) Cell 115, 255–2665. Damak, S., Rong, M., Yasumatsu, K., Kokrashvili, Z., Varadarajan, V., Zou, S.,
Jiang, P., Ninomiya, Y., and Margolskee, R. F. (2003) Science 301, 850–8536. Hoon, M. A., Adler, E., Lindemeier, J., Battey, J. F., Ryba, N. J., and Zuker,
C. S. (1999) Cell 96, 541–551
7. Bachmanov, A. A., Li, X., Reed, D. R., Ohmen, J. D., Li, S., Chen, Z., Tordoff,M. G., de Jong, P. J., Wu, C., West, D. B., Chatterjee, A., Ross, D. A., andBeauchamp, G. K. (2001) Chem. Senses 26, 925–933
8. Montmayeur, J. P., Liberles, S. D., Matsunami, H., and Buck, L. B. (2001) Nat.Neurosci. 4, 492–498
9. Max, M., Shanker, Y. G., Huang, L., Rong, M., Liu, Z., Campagne, F., Wein-stein, H., Damak, S., and Margolskee, R. F. (2001) Nat. Genet. 28, 58–63
10. Kitagawa, M., Kusakabe, Y., Miura, H., Ninomiya, Y., and Hino, A. (2001)Biochem. Biophys. Res. Commun. 283, 236–242
11. Kunishima, N., Shimada, Y., Tsuji, Y., Sato, T., Yamamoto, M., Kumasaka, T.,Nakanishi, S., Jingami, H., and Morikawa, K. (2000) Nature 407, 971–977
12. Binet, V., Brajon, C., Le Corre, L., Acher, F., Pin, J. P., and Prezeau, L. (2004)J. Biol. Chem. 279, 29085–29091
13. Malherbe, P., Kratochwil, N., Knoflach, F., Zenner, M. T., Kew, J. N.,Kratzeisen, C., Maerki, H. P., Adam, G., and Mutel, V. (2003) J. Biol. Chem.278, 8340–8347
14. Pagano, A., Ruegg, D., Litschig, S., Stoehr, N., Stierlin, C., Heinrich, M.,Floersheim, P., Prezeau, L., Carroll, F., Pin, J. P., Cambria, A., Vranesic, I.,Flor, P. J., Gasparini, F., and Kuhn, R. (2000) J. Biol. Chem. 275,33750–33758
15. Petrel, C., Kessler, A., Maslah, F., Dauban, P., Dodd, R. H., Rognan, D., andRuat, M. (2003) J. Biol. Chem. 278, 49487–49494
16. Petrel, C., Kessler, A., Dauban, P., Dodd, R. H., Rognan, D., and Ruat, M.(2004) J. Biol. Chem. 279, 18990–18997
17. Jiang, P., Ji, Q., Liu, Z., Snyder, L. A., Benard, L. M., Margolskee, R. F., andMax, M. (2004) J. Biol. Chem. 279, 45068–45075
18. Xu, H., Staszewski, L., Tang, H., Adler, E., Zoller, M., and Li, X. (2004) Proc.Natl. Acad. Sci. U. S. A. 101, 14258–14263
19. Jiang, P., Cui, M., Ji, Q., Snyder, L., Lui, Z., Benard, L., Margolskee, R.,Osman, R., and Max, M. (2005) Chem. Senses 30, Suppl. 1, 117–118
20. Sclafani, A., and Perez, C. (1997) Physiol. Behav. 61, 25–2921. Ballesteros, J. A., and Weinstein, H. (1995) Methods Neurosci. 25, 366–42822. Ueda, T., Ugawa, S., Yamamura, H., Imaizumi, Y., and Shimada, S. (2003)
J. Neurosci. 23, 7376–738023. Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994) Nucleic Acids Res. 22,
4673–468024. Fiser, A., Do, R. K., and Sali, A. (2000) Protein Sci. 9, 1753–177325. Fiser, A., and Sali, A. (2003) Bioinformatics 19, 2500–250126. Marti-Renom, M. A., Stuart, A. C., Fiser, A., Sanchez, R., Melo, F., and Sali, A.
(2000) Annu. Rev. Biophys. Biomol. Struct. 29, 291–32527. Sali, A., and Blundell, T. L. (1993) J. Mol. Biol. 234, 779–81528. Brooks, B. R., Bruccoleri, R. E., Olafson, B. D., States, D. J., Swaminathan, S.,
and Karplus, M. (1983) J. Comp. Chem. 4, 187–21729. Frisch, M. J., Trucks, G. W., Schlegel, H. B., Scuseria, G. E., Robb, M. A.,
Cheeseman, J. R., Zakrzewski, Montgomery, V. G., Jr., Stratmann, R. E.,Burant, J. C., Dapprich, S., Millam, J. M., Daniels, A. D., Kudin, K. N.,Strain, M. C., Farkas, O., Tomasi, J., Barone, V., Cossi, M., Cammi, R.,Mennucci, B., Pomelli, C., Adamo, C., Clifford, S., Ochterski, J., Petersson,G. A., Ayala, P. Y., Cui, Q., Morokuma, K., Malick, D. K., Rabuck, A. D.,Raghavachari, K., Foresman, J. B., Cioslowski, J., Ortiz, J. V., Baboul,A. G., Stefanov, B. B., Liu, G., Liashenko, A., Piskorz, P., Komaromi, I.,Gomperts, R., Martin, R. L., Fox, D. J., Keith, T., Al-Laham, M. A., Peng,C. Y., Nanayakkara, A., Gonzalez, C., Challacombe, M., Gill, P. M. W.,Johnson, B. G., Chen, M., Wong, W., Andres, J. L., Head-Gordon, M.,Replogle, E. S., and Pople, J. A. (1998) Gaussian 98 (Revision A.x), GaussianInc., Pittsburgh, PA
30. Breneman, C. M., and Wiberg, K. B. (1990) J. Comp. Chem. 11, 361–37331. Morris, G. M., Goodsell, D. S., Halliday, R. S., Huey, R., Hart, W. E., Belew,
R. K., and Olson, A. J. (1998) J. Comp. Chem. 19, 1639–166232. Schiffman, S. S., Booth, B. J., Sattely-Miller, E. A., Graham, B. G., and Gibes,
K. M. (1999) Chem. Senses 24, 439–44733. Johnson, C., Birch, G. G., and MacDougall, D. B. (1994) Chem. Senses 19,
349–35834. Malherbe, P., Knoflach, F., Broger, C., Ohresser, S., Kratzeisen, C., Adam, G.,
Stadler, H., Kemp, J. A., and Mutel, V. (2001) Mol. Pharmacol. 60, 944–95435. Palczewski, K., Kumasaka, T., Hori, T., Behnke, C. A., Motoshima, H., Fox,
B. A., Trong, I. L., Teller, D. C., Okada, T., Stenkamp, R. E., Yamamoto, M.,and Miyano, M. (2000) Science 289, 739–745
36. Strader, C. D., and Dixon, R. A. (1991) Adv. Exp. Med. Biol. 287, 209–22037. Perlman, J. H., Thaw, C. N., Laakkonen, L., Bowers, C. Y., Osman, R., and
Gershengorn, M. C. (1994) J. Biol. Chem. 269, 1610–161338. Perlman, J. H., Laakkonen, L. J., Guarnieri, F., Osman, R., and Gershengorn,
M. C. (1996) Biochemistry 35, 7643–765039. Manivet, P., Schneider, B., Smith, J. C., Choi, D. S., Maroteaux, L., Keller-
mann, O., and Launay, J. M. (2002) J. Biol. Chem. 277, 17170–1717840. Schmidt, C., Li, B., Bloodworth, L., Erlenbach, I., Zeng, F. Y., and Wess, J.
(2003) J. Biol. Chem. 278, 30248–3026041. Farrens, D. L., Altenbach, C., Yang, K., Hubbell, W. L., and Khorana, H. G.
(1996) Science 274, 768–77042. Ward, S. D., Hamdan, F. F., Bloodworth, L. M., and Wess, J. (2002) J. Biol.
Chem. 277, 2247–225743. Ghanouni, P., Steenhuis, J. J., Farrens, D. L., and Kobilka, B. K. (2001) Proc.
Natl. Acad. Sci. U. S. A. 98, 5997–6002
Lactisole Inhibits the Sweet Taste Receptor15246
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Supplemental Figure 1:
Chemical structures of sweet ligands.
Supplemental Figure 2:
Alignment of sequences of hT1R3, hT1R2, mT1R2, mT1R3, mGluR1 and rhodopsin.
Supplemental Figure 3: Humanized mT1R3 is sensitive to lactisole.
HEK293E cells were transfected with Gα16-gust44, hT1R2 and mT1R3 mutants in which species-
variant positions within TM helices 5 and 7 of T1R3 had been shown to affect the receptor’s sensitivity
to lactisole. Responses of these mutants to D-tryptophan (30mM) were examined in the presence or
absence of lactisole (1.25mM). Receptors containing wildtype mT1R3, mT1R3(Q795R), mT1R3(I803L)
and mT1R3(Q795R I803L) are insensitive to lactisole. Receptors containing mT1R3(V738A) or
mT1R3(V738A Q795R) are sensitive to lactisole. Receptors containing mT1R3(V738A I803L) or
mT1R3(V738AQ795RI803L) have diminished responses to D-tryptophan making it hard to evaluate the
effect of lactisole.
Supplemental Figure 4: hT1R3(Q637E) is not sensitive to lactisole.
HEK293E cells were transfected with hT1R2, hT1R3(Q6373.29E) and Gα16-gust44. Responses of the
mutant to D-tryptophan (10mM) were examined in the presence or absence of lactisole (1.25mM).
Receptor with hT1R3(Q6373.29E) showed no sensitivity to lactisole.
Acesulfame K Cyclamate D-tryptophan
Neohesperidin dihydrochalcone Saccharin
Sucralose Sucrose
Supplemental Fig. 1

ht1r2 557 VFLEWHEAPTIAVALLAALGFLSTLAILVIFWRHFQTPIVRSAGGPMCFLMLTLLLVAYM mt1r2 561 AFLEWHEVPTIVVTILAALGFISTLAILLIFWRHFQTPMVRSAGGPMCFLMLVPLLLAFG ht1r3 560 RFLAWGEPAVLLLLLLLSLALGLVLAALGLFVHHRDSPLVQASGGPLACFGLVCLGLVCL mt1r3 565 KFLAWGEPVVLSLLLLLCLVLGLALAALGLSVHHWDSPLVQASGGSQFCFGLICLGLFCL mGluR1 584 RYLEWSDIESIIAIAFSCLGILVTLFVTLIFVLYRDTPVVKSSSRELCYIILAGIFLGYV Rhodo 32 AEPWQFSMLAAYMFLLIMLGFPINFLTLYVTVQHKKLRTPLNYILLNLAVADLFMVFGGF HHHHHHHHHHHHHHHHHHHHHHHHHHHHHHH HHHHHHHHHHHHHHHHHHHHH ht1r2 617 VVPVYVGPP-----KVSTCLCRQALFPLCFTICISCIAVRSFQIVCAFKMASR-FPRAYS mt1r2 621 MVPVYVGPP-----TVFSCFCRQAFFTVCFSVCLSCITVRSFQIVCVFKMARR-LPSAYG ht1r3 620 SVLLFPGQP-----SPARCLAQQPLSHLPLTGCLSTLFLQAAEIFVESELPLS-WADRLS mt1r3 625 SVLLFPGRP-----SSASCLAQQPMAHLPLTGCLSTLFLQAAETFVESELPLS-WANWLC mGluR1 644 CPFTLIAKP-----TTTSCYLQRLLVGLSSAMCYSALVTKTNRIARILAGSKKKICTRKP Rhodo 92 TTTLYTSLHGYFVFGPTGCNLEGFFATLGGEIALWSLVVLAIERYVVVCKPMSNFRFG-- HHHHHHHHH HHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHH ht1r2 671 YWVRYQGPYVSMAFITVLKMVIVVIGMLATGLSPTTRTDPDDPKITIVSCNP-------- mt1r2 675 FWMRYHGPYVFVAFITAVKVALVAGNMLATTINPIGRTDPDDPNIIILSCHP-------- ht1r3 674 GCLRGPWAWLVVLLAMLVEVALCTWYLVAFPPEVVTDWH-MLPTEALVHCRT-------- mt1r3 679 SYLRGLWAWLVVLLATFVEAALCAWYLIAFPPEVVTDWS-VLPTEVLEHCHV-------- mGluR1 699 RFMSAWAQVIIASILISVQLTLVVTLIIMEPPMPILSYP--SIKEVYLICN--------- Rhodo 150 ----------ENHAIMGVAFTWVMALACAAPPLVGWSRY-IPEGMQ-CSCGIDYYTPHEE HHHHHHHHHHHHHHHHHHHHHHHH ht1r2 723 --NYRNSLLFNTSLDLLLSVVGFSFAYMGKELP------------TNYNEAKFITLSMTF mt1r2 727 --NYRNGLLFNTSMDLLLSVLGFSFAYVGKELP------------TNYNEAKFITLSMTF ht1r3 725 --RSWVSFGLAHATNATLAFLCFLGTFLVRSQP------------GRYNRARGLTFAMLA mt1r3 730 --RSWVSLGLVHITNAMLAFLCFLGTFLVQSQP------------GRYNRARGLTFAMLA mGluR1 748 --TSNLGVVAPVGYNGLLIMSCTYYAFKTRNVP------------ANFNEAKYIAFTMYT Rhodo 198 TNFVIYMFVVHFIIPLIVIFFCYGQLVFTVKEAAAQQQESATTQKAEKEVTRMVIIMVIA HHHHHHHHHHHHHHHHHHHHHHHHHH HHHHHHHHHHHHHH ht1r2 769 YFTSSVSLCTFMSAYSGVLV-TIVDLLVTVLNLLAISLGYFGPKCYMILFYPERNTPAYF mt1r2 773 SFTSSISLCTFMSVHDGVLV-TIMDLLVTVLNFLAIGLGYFGPKCYMILFYPERNTSAYF ht1r3 771 YFITWVSFVPLLANVQVVLR-PAVQMGALLLCVLGILAAFHLPRCYLLMRQPGLNTPEFF mt1r3 776 YFITWVSFVPLLANVQVAYQ-PAVQMGAILVCALGILVTFHLPKCYVLLWLPKLNTQEFF mGluR1 794 TCIIWLAFVPIYFGSNY--K-IITTCFAVSLSVTVALGCMFTPKMYIIIAKPERNVRSAF Rhodo 261 FLICWLPYAGVAFYIFTHQGSDFGPIFMTIPAFFAKTSAVYNPVIYIMMNKQFRNCMVTT HHHHHHHHHHHHHHHHH HHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHH ht1r2 828 NSMIQGYTM mt1r2 832 NSMIQGYTM ht1r3 830 LGGGPGDAQ mt1r3 835 LGRNAKKAA mGluR1 851 TTSDVVRMH Rhodo 321 LCCGKNPLG Q7RTX0 new TAS1R3 (human) SWISS-PROT; Q7RTX0 Q8TE23 TAS1R2 (human) SWISS-PROT; Q8TE23 Q91VA4 T1R3 OR TAS1R3 (mouse) SWISS-PROT; Q91VA4 Q923J8 TAS1R2 (mouse) SWISS-PROT; Q923J8 MGR1_MOUSE SWISS-PROT; P97772 Rhodo P02699
Supplemental Fig. 2

hT1R2 + mT1R3 mutants
hT1R
3
mT1R3
mT1R3(V
738A
)
mT1R3(Q
795R
)
mT1R3(I
803L
)
mT1R3(V
738A
Q795R
)
mT1R3(V
738A
I803L
)
mT1R3(Q
795R
I803L
)
mT1R3(V
738A
Q795R
I803L
)
0
10
20
30
40D-tryptophanD-tryptophan + lactisole
∆F
/F (
%)
Supplemental Fig. 3

hT1R2 + mutant
hT1R3 hT1R3(Q637E)0
10
20 D-tryptophanD-tryptophan + lactisole
∆F
/F (
%)
Supplemental Fig. 4

Roman Osman, Robert F. Margolskee and Marianna MaxPeihua Jiang, Meng Cui, Baohua Zhao, Zhan Liu, Lenore A. Snyder, Lumie M. J. Benard,
Sweet TasteLactisole Interacts with the Transmembrane Domains of Human T1R3 to Inhibit
doi: 10.1074/jbc.M414287200 originally published online January 24, 20052005, 280:15238-15246.J. Biol. Chem.
10.1074/jbc.M414287200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2005/01/25/M414287200.DC1.html
http://www.jbc.org/content/280/15/15238.full.html#ref-list-1
This article cites 42 references, 23 of which can be accessed free at
by guest on March 12, 2016
http://ww
w.jbc.org/
Dow
nloaded from





























