Embed Size (px)
Citation preview

Biochemical and Biophysical Research Communications 397 (2010) 220–225
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
Amiloride reduces the sweet taste intensity by inhibiting the human sweettaste receptor
Takamasa Imada a, Takumi Misaka a,*, Satoshi Fujiwara b, Shinji Okada a, Yusuke Fukuda a, Keiko Abe a
a Department of Applied Biological Chemistry, Graduate School of Agricultural and Life Sciences, The University of Tokyo, Tokyo 113-8657, Japanb T. Hasegawa Co., Ltd., Kawasaki-shi, Kanagawa 211-0022, Japan
a r t i c l e i n f o
Article history:Received 8 May 2010Available online 21 May 2010
Keywords:TasteSweetTaste receptorAmilorideInhibitor
0006-291X/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.bbrc.2010.05.088
* Corresponding author. Address: Department of AGraduate School of Agricultural and Life Sciences, ThYayoi, Bunkyo-ku, Tokyo 113-8657, Japan. Fax: +81 3
E-mail address: [email protected] (T
a b s t r a c t
In mammals, sweet taste perception is mediated by the heterodimeric G-protein-coupled receptor,T1R2/T1R3. An interesting characteristic of this sweet taste receptor is that it has multiple ligand bind-ing sites. Although there have been several studies on agonists of sweet taste receptors, little is knownabout antagonists of these receptors. In this study, we constructed a cell line stably expressing thehuman sweet taste receptor (hT1R2/hT1R3) and a functional chimeric G-protein (hGa16gust44) usingthe Flp-In system for measuring the antagonistic activity against the receptor. This constructed cell lineresponded quite intensely and frequently to the compounds applied for activation of hT1R2/hT1R3. Inthe presence of 3 mM amiloride, the responses to sweet tastants such as sugar, artificial sweetener, andsweet protein were significantly reduced. The inhibitory activity of amiloride toward 1 mM aspartamewas observed in a dose-dependent manner with an IC50 value of 0.87 mM. Our analysis of a cell lineexpressing hT1R3 mutants (hT1R3-A733V or hT1R3-F778A) made us to conclude that the target siteof amiloride is distinct from that of lactisole, a known sweet taste inhibitor. Our results strongly indi-cate that amiloride reduces the sweet taste intensity by inhibiting the human sweet taste receptor andalso that this receptor has multiple inhibitor binding sites.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
Taste is an important sensation for vertebrates, contributing toingestion of nutrients and avoidance of poisonous substances.Sweet taste is very important because it is an indicator of thechemical energy in foods [1]. In mammals, the perception of sweettaste is mediated by the heterodimeric G-protein-coupled receptor(GPCR), T1R2/T1R3 [2–6]. Interestingly, sweet compounds have awide variety of structural features as evidenced by the vastly dif-ferent chemical structures of sugars, artificial sweetener, sweetproteins, and so on [7]. Previous studies have indicated that thesweet taste receptor has multiple ligand binding sites [7]. Forexample, aspartame is recognized at the extracellular domain ofhuman T1R2 (hT1R2) [8], while cyclamate is received at the trans-membrane domain of human T1R3 (hT1R3) [8,9]. Sweet proteinssuch as monellin, brazzein [10], and neoculin [11] have beenshown to interact with the extracellular domain of hT1R3.
Although several studies have focused on agonists for sweettaste receptors, little is known about antagonists. For example,
ll rights reserved.
pplied Biological Chemistry,e University of Tokyo, 1-1-15841 8118.
. Misaka).
several compounds such as lactisole [12,13] and gymnemic acid[14,15] and ions such as Zn2+ [16,17] are known to repress theintensity of sweet taste according to human sensory evaluation.However, the interactions of these agonists with hT1R2/hT1R3are not well defined. Lactisole is the only antagonist whoseinhibitory mechanism has been clarified by molecular biologicalmethods. It has been demonstrated that this compound inhibitsthe activation of hT1R2/hT1R3 through interactions with the trans-membrane domain of hT1R3 [8,18,19]. These reports have identi-fied the specific residues in the transmembrane domain of T1R3that differ between human T1R3 and mouse T1R3, and they criti-cally determined the human-specific sensitivity to lactisole.
On the other hand, it is well known that amiloride partially re-duces the intensity of salty taste in humans and other mammals[20–23]. Amiloride blocks epithelial sodium ion channels (ENaC)[24] expressed in taste bud cells and therefore reduces the saltytaste sensation. Schiffman et al. [23] carried out human sensorytests and found that amiloride represses not only the salty tastebut also the sweet taste. In the presence of amiloride, the tasteintensities of various sweeteners significantly reduced [23], butthe mechanism remains to be clarified.
At the beginning of this study, we constructed a cell line stablyexpressing the human sweet taste receptor (hT1R2/hT1R3) and achimeric G-protein (hGa16gust44) as the basis for evaluating the

T. Imada et al. / Biochemical and Biophysical Research Communications 397 (2010) 220–225 221
sweet taste sensation in humans. We also investigated whetheramiloride inhibits the activation of human sweet taste receptor.Our results strongly indicated that the effect of amiloride on thesweet taste reduction was just the inhibition of hT1R2/hT1R3.
2. Materials and methods
2.1. Sample solutions
The Ca2+-containing buffer (assay buffer) included 10 mM4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 130mM NaCl, 10 mM glucose, 5 mM KCl, 2 mM CaCl2, and 1.2 mMMgCl2 (pH adjusted to 7.4 with NaOH). Ligands were diluted withthe assay buffer at the desired concentrations.
2.2. Construction of stable cell lines and cell culture
The entire coding region of hT1R2, hT1R3, and the chimericG-protein a-subunit, hGa16gust44 [25], were subcloned intopcDNA5/FRT vector (Invitrogen, Carlsbad, CA, USA) modified by6-nucleotide mutation according to the procedure described inthe text of Supplemental information and Fig. 1. The constructwas confirmed by sequencing using an ABI 3130 or 310 DNA genet-ic analyzer (Applied Biosystems, Foster City, CA, USA).
Flp-In 293 cells (Invitrogen) were cultured at 37 �C inDulbecco’s modified Eagle’s medium (Sigma–Aldrich Japan, Tokyo,Japan) supplemented with 10% fetal bovine serum (Invitrogen).The cell lines were constructed according to the manufacturer’sprotocol for the Flp-In pcDNA5/FRT Complete System (Invitrogen).Flp-In 293 cells were transfected with both the constructedplasmid and pOG44 (Invitrogen) using Lipofectamine 2000 (Invit-rogen). Then, 48 h after transfection, the cells were selected bytreatment with 100 lg/ml of hygromycin B (Invitrogen) for 2–3 weeks. Antibiotic resistant cells were collected, cultured, andused for measuring the cellular response to sweet tastants. Thecells for these measurements were cultured in low glucose(1.0 g/l) Dulbecco’s modified Eagle’s medium with 10% fetal bovineserum.
2.3. Measurement of cellular responses by calcium imaging analysis
On a day before the analysis, cells were trypsinized and seededinto 96-well Lumox multiwell black-wall plates (Greiner Bio-One,Frickenhausen, Germany). After 20–26 h, the cells were washedwith assay buffer, and then loaded with 5 lM of fura-2/AM (Invit-rogen) in assay buffer for 30 min at 27 �C. The cells were washedagain with assay buffer and incubated in 100 ll of assay bufferfor up to 15 min at room temperature. The cells were stimulatedwith sweet tastants by adding 100 ll of 2� ligands. The intensitiesof fura-2 fluorescence emissions resulting from excitation at 340and 380 nm were measured at 510 nm using a computer-con-trolled filter exchanger (Lambda 10-3; Sutter, San Rafael, CA,USA), a CoolSNAP HQ2 charge-coupled device camera (Photomet-rics, Tucson, AZ, USA), and an inverted fluorescence microscope(IX-71; Olympus, Tokyo, Japan). The images were recorded at 4 sintervals and analyzed using MetaFluor software (Molecular De-vices, Sunnyvale, CA, USA). Changes in intracellular calcium ionconcentration were estimated from changes in the ratio of fluores-cence intensity at the two excitation wavelengths (F340/F380).
2.4. Measurement of cellular responses by cell-based assay
The trypsinized cells were seeded at a density of 70,000 cellsper well into 96-well black-wall CellBIND surface plates (Corning,Corning, NY, USA) and 23 h after, washed with assay buffer prior
to load with a calcium indicator dye from the FLIPR Calcium 4 As-say Kit (Molecular Devices) by dilution with assay buffer. The cellswere incubated for 45 min at 27 �C, and measurements were madeusing FlexStation 3 (Molecular Devices). Fluorescence changes(excitation at 485 nm, emission at 525 nm, and cutoff at 515 nm)were monitored at 2 s intervals, an aliquot of 100 ll of assay buffersupplemented with 2� ligands were added at 20 s, and scanningwas continued for an additional 100 s. The response of each wellwas represented as DRFU (delta relative fluorescence unit), whichwas defined as (maximum fluorescence value) – (minimum fluo-rescence value). The responses were averaged from at least threewells receiving the same stimulus. Plots of the amplitudes versusconcentrations were fitted by Hill’s equations.
3. Results
3.1. Construction of cell lines stably expressing the human sweet tastereceptor and its mutants
To construct the cell lines used in this study, the Flp-In systemand Flp-In 293 cells were treated as described in Section 2, Supple-mental text and Fig. 1. Flp-In system allows the generation of iso-genic stable cell lines because an expression vector is integratedinto the genome via Flp recombinase-mediated DNA recombina-tion at the FRT site at a specific location of the genome of Flp-In293 cells. When the cell line expressing the functional humansweet taste receptor was constructed, the expression plasmidswere designed to contain the entire coding region for hT1R2,hT1R3, and hGa16gust44 as indicated in Fig. 1A–C. When the celllines expressing hT1R3 mutant were constructed, the hT1R3 cDNAfragment with the point mutation was used instead of using thewild type hT1R3 cDNA (Fig. 1A) to create the entire construct(Fig. 1C). Each plasmid was integrated into the Flp-In 293 cellsvia Flp recombinase-mediated DNA recombination at the specificFRT site. Cells were selected by hygromycin resistance and usedfor the measurement of cell responses to sweet tastants. In thisstudy, we generated four types of cell lines as summarized inFig. 1D.
3.2. Response of the cell line stably expressing the human sweet tastereceptor to sweet tastants
To verify the response of the generated cell line to various sweettastants, calcium imaging analysis was conducted to examinecellular responses to sweet tastants. In the case of the cells stablyexpressing hT1R2/hT1R3, intense and frequent responseswere observed upon treatment with compounds that activatehT1R2/hT1R3—aspartame, saccharin, acesulfame-K, cyclamate,and sucrose (Fig. 2A).
When the cell line expressing hT1R3, but not hT1R2 (Fig. S1A),was used for measuring cell responses, the cells did not respond toany of the sweeteners tested (Fig. S1B). These results strongly indi-cated that the responses of the cells stably expressing the humansweet taste receptor were due to expression of hT1R2/hT1R3 andhGa16gust44, and also that these proteins were functionally ex-pressed in the cell line (Fig. 2A).
To examine the relationship between the concentration ofsweet tastant and cellular responses, the cell-based assay was usedto investigate stimulatory responses of the hT1R2/hT1R3-express-ing cells. When aspartame was used as a sweet tastant, its responsewas clearly observed in a dose-dependent manner with an EC50
value of 1.2 ± 0.07 mM (Fig. 2B). We also confirmed the dose-dependent response of the hT1R2/hT1R3-expressing cells to othersweet tastants with EC50 values at 0.37 ± 0.05 mM (to saccharin),0.94 ± 0.05 mM (to acesulfame-K), and 4.6 ± 0.25 mM (to

Fig. 1. Expression construct of hT1R2/hT1R3 and hGa16gust44. (A–C) Schematic illustration of the plasmid construction. The expression plasmids were designed to containthe entire coding region for each of hT1R2, hT1R3, and hGa16gust44. I, internal ribosome entry site (IRES) sequence; G, hGa16gust44; pA, polyadenylation signal; F, FRT site;and Hygr, hygromycin resistance gene. (D) Summary of cell lines expressing hT1Rs generated in this study.
222 T. Imada et al. / Biochemical and Biophysical Research Communications 397 (2010) 220–225
cyclamate). This response, quite similar to the data reported byother independent groups [8,9,26,27], strongly indicates that thehuman sweet receptor was fully functional in our cell line stablyexpressing hT1R2/hT1R3.
3.3. Amiloride inhibits the human sweet taste receptor
In human sensory tests, amiloride inhibits the sweet taste sen-sation of sucrose, aspartame, and other sweet compounds [23], andso we examined the effect of amiloride on the response of hT1R2/hT1R3-expressing cells to a variety of sweet tastants (Fig. 3A). As aresult, in the presence of 3 mM amiloride, all responses of the cellsto sweet tastants such as sucrose (disaccharide), aspartame (pep-tide), saccharin, acesulfame-K (an N-sulfonylamide), cyclamate(sulfamate), neohesperidin dihydrochalcone (NHDC, a dihydroch-alcone), thaumatin and neoculin (proteins), significantly reduced.Moreover, inhibitory activity of amiloride toward 1 mM aspartamewas observed in a dose-dependent manner with an IC50 value of0.87 ± 0.20 mM (Fig. 3B).
To examine whether the inhibitory effect of amiloride was spe-cific to the response mediated by the human sweet taste receptor,the response of hT1R2/hT1R3-expressing cells to ATP was exam-ined. ATP activates endogenous GPCR of the cultured cells [28–31] and changes in the intracellular calcium ion concentrationwere clearly observed after the addition of ATP (Fig. 3C). Thedose-response relationship to ATP in the presence of 3 mM of amil-oride was found comparable to that of ATP alone (Fig. 3C). This re-sult strongly indicates that amiloride does not always reduce thecellular response as measured by the cell-based assay and thatthe suppression of receptor activity by amiloride is specific tohT1R2/hT1R3.
3.4. Amiloride and lactisole target different site on the human sweettaste receptor
Lactisole, a known hT1R2/hT1R3 inhibitor, has been shown tointeract with the transmembrane domain of hT1R3 [8,18,19]. Togain knowledge about the site targeted by amiloride, we generated
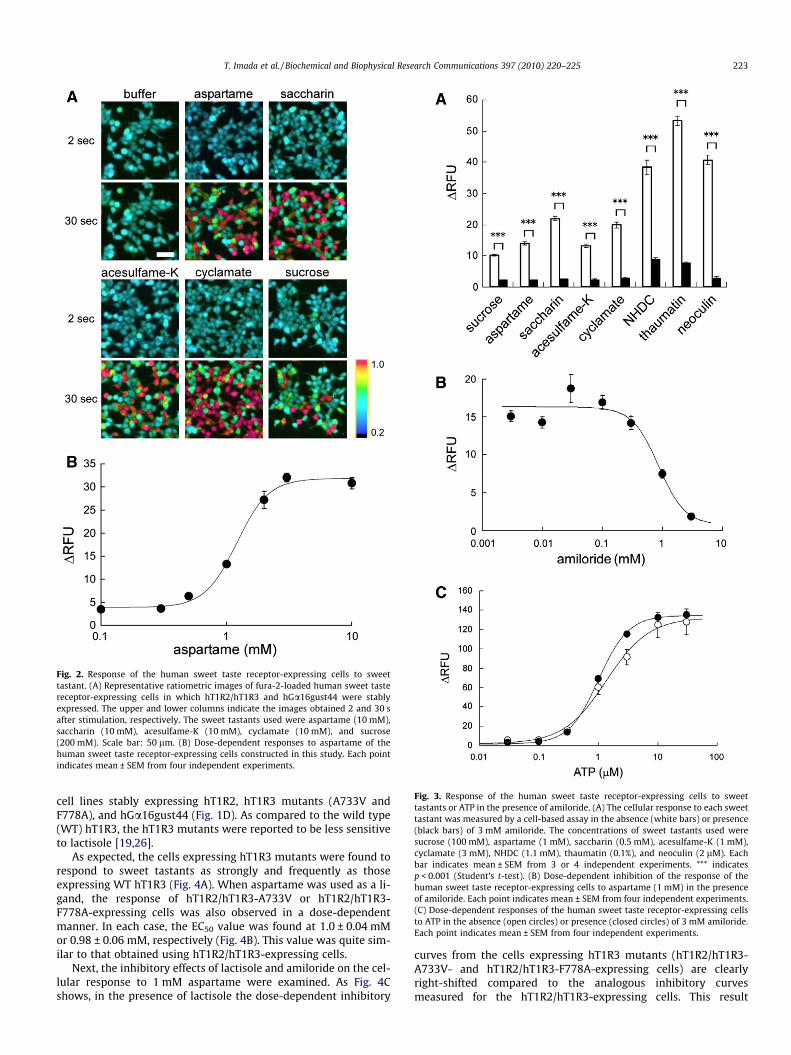
Fig. 3. Response of the human sweet taste receptor-expressing cells to sweettastants or ATP in the presence of amiloride. (A) The cellular response to each sweettastant was measured by a cell-based assay in the absence (white bars) or presence(black bars) of 3 mM amiloride. The concentrations of sweet tastants used weresucrose (100 mM), aspartame (1 mM), saccharin (0.5 mM), acesulfame-K (1 mM),cyclamate (3 mM), NHDC (1.1 mM), thaumatin (0.1%), and neoculin (2 lM). Eachbar indicates mean ± SEM from 3 or 4 independent experiments. *** indicatesp < 0.001 (Student’s t-test). (B) Dose-dependent inhibition of the response of thehuman sweet taste receptor-expressing cells to aspartame (1 mM) in the presenceof amiloride. Each point indicates mean ± SEM from four independent experiments.(C) Dose-dependent responses of the human sweet taste receptor-expressing cellsto ATP in the absence (open circles) or presence (closed circles) of 3 mM amiloride.Each point indicates mean ± SEM from four independent experiments.
Fig. 2. Response of the human sweet taste receptor-expressing cells to sweettastant. (A) Representative ratiometric images of fura-2-loaded human sweet tastereceptor-expressing cells in which hT1R2/hT1R3 and hGa16gust44 were stablyexpressed. The upper and lower columns indicate the images obtained 2 and 30 safter stimulation, respectively. The sweet tastants used were aspartame (10 mM),saccharin (10 mM), acesulfame-K (10 mM), cyclamate (10 mM), and sucrose(200 mM). Scale bar: 50 lm. (B) Dose-dependent responses to aspartame of thehuman sweet taste receptor-expressing cells constructed in this study. Each pointindicates mean ± SEM from four independent experiments.
T. Imada et al. / Biochemical and Biophysical Research Communications 397 (2010) 220–225 223
cell lines stably expressing hT1R2, hT1R3 mutants (A733V andF778A), and hGa16gust44 (Fig. 1D). As compared to the wild type(WT) hT1R3, the hT1R3 mutants were reported to be less sensitiveto lactisole [19,26].
As expected, the cells expressing hT1R3 mutants were found torespond to sweet tastants as strongly and frequently as thoseexpressing WT hT1R3 (Fig. 4A). When aspartame was used as a li-gand, the response of hT1R2/hT1R3-A733V or hT1R2/hT1R3-F778A-expressing cells was also observed in a dose-dependentmanner. In each case, the EC50 value was found at 1.0 ± 0.04 mMor 0.98 ± 0.06 mM, respectively (Fig. 4B). This value was quite sim-ilar to that obtained using hT1R2/hT1R3-expressing cells.
Next, the inhibitory effects of lactisole and amiloride on the cel-lular response to 1 mM aspartame were examined. As Fig. 4Cshows, in the presence of lactisole the dose-dependent inhibitory
curves from the cells expressing hT1R3 mutants (hT1R2/hT1R3-A733V- and hT1R2/hT1R3-F778A-expressing cells) are clearlyright-shifted compared to the analogous inhibitory curvesmeasured for the hT1R2/hT1R3-expressing cells. This result
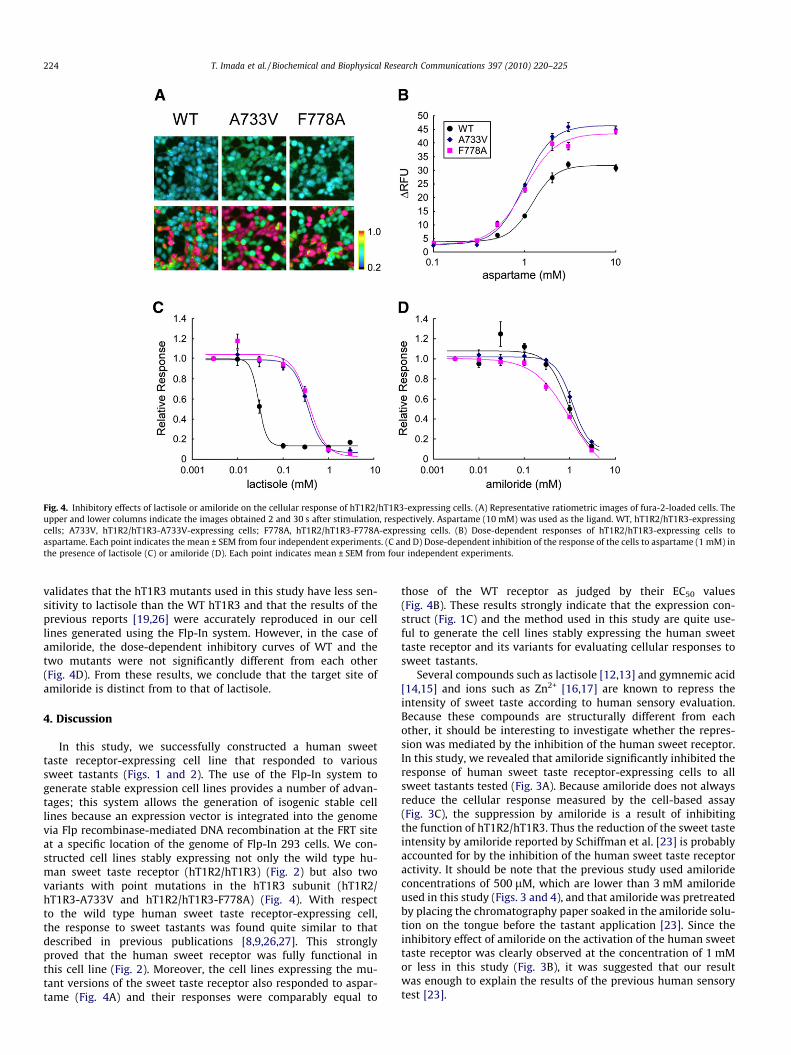
Fig. 4. Inhibitory effects of lactisole or amiloride on the cellular response of hT1R2/hT1R3-expressing cells. (A) Representative ratiometric images of fura-2-loaded cells. Theupper and lower columns indicate the images obtained 2 and 30 s after stimulation, respectively. Aspartame (10 mM) was used as the ligand. WT, hT1R2/hT1R3-expressingcells; A733V, hT1R2/hT1R3-A733V-expressing cells; F778A, hT1R2/hT1R3-F778A-expressing cells. (B) Dose-dependent responses of hT1R2/hT1R3-expressing cells toaspartame. Each point indicates the mean ± SEM from four independent experiments. (C and D) Dose-dependent inhibition of the response of the cells to aspartame (1 mM) inthe presence of lactisole (C) or amiloride (D). Each point indicates mean ± SEM from four independent experiments.
224 T. Imada et al. / Biochemical and Biophysical Research Communications 397 (2010) 220–225
validates that the hT1R3 mutants used in this study have less sen-sitivity to lactisole than the WT hT1R3 and that the results of theprevious reports [19,26] were accurately reproduced in our celllines generated using the Flp-In system. However, in the case ofamiloride, the dose-dependent inhibitory curves of WT and thetwo mutants were not significantly different from each other(Fig. 4D). From these results, we conclude that the target site ofamiloride is distinct from to that of lactisole.
4. Discussion
In this study, we successfully constructed a human sweettaste receptor-expressing cell line that responded to varioussweet tastants (Figs. 1 and 2). The use of the Flp-In system togenerate stable expression cell lines provides a number of advan-tages; this system allows the generation of isogenic stable celllines because an expression vector is integrated into the genomevia Flp recombinase-mediated DNA recombination at the FRT siteat a specific location of the genome of Flp-In 293 cells. We con-structed cell lines stably expressing not only the wild type hu-man sweet taste receptor (hT1R2/hT1R3) (Fig. 2) but also twovariants with point mutations in the hT1R3 subunit (hT1R2/hT1R3-A733V and hT1R2/hT1R3-F778A) (Fig. 4). With respectto the wild type human sweet taste receptor-expressing cell,the response to sweet tastants was found quite similar to thatdescribed in previous publications [8,9,26,27]. This stronglyproved that the human sweet receptor was fully functional inthis cell line (Fig. 2). Moreover, the cell lines expressing the mu-tant versions of the sweet taste receptor also responded to aspar-tame (Fig. 4A) and their responses were comparably equal to
those of the WT receptor as judged by their EC50 values(Fig. 4B). These results strongly indicate that the expression con-struct (Fig. 1C) and the method used in this study are quite use-ful to generate the cell lines stably expressing the human sweettaste receptor and its variants for evaluating cellular responses tosweet tastants.
Several compounds such as lactisole [12,13] and gymnemic acid[14,15] and ions such as Zn2+ [16,17] are known to repress theintensity of sweet taste according to human sensory evaluation.Because these compounds are structurally different from eachother, it should be interesting to investigate whether the repres-sion was mediated by the inhibition of the human sweet receptor.In this study, we revealed that amiloride significantly inhibited theresponse of human sweet taste receptor-expressing cells to allsweet tastants tested (Fig. 3A). Because amiloride does not alwaysreduce the cellular response measured by the cell-based assay(Fig. 3C), the suppression by amiloride is a result of inhibitingthe function of hT1R2/hT1R3. Thus the reduction of the sweet tasteintensity by amiloride reported by Schiffman et al. [23] is probablyaccounted for by the inhibition of the human sweet taste receptoractivity. It should be note that the previous study used amilorideconcentrations of 500 lM, which are lower than 3 mM amilorideused in this study (Figs. 3 and 4), and that amiloride was pretreatedby placing the chromatography paper soaked in the amiloride solu-tion on the tongue before the tastant application [23]. Since theinhibitory effect of amiloride on the activation of the human sweettaste receptor was clearly observed at the concentration of 1 mMor less in this study (Fig. 3B), it was suggested that our resultwas enough to explain the results of the previous human sensorytest [23].

T. Imada et al. / Biochemical and Biophysical Research Communications 397 (2010) 220–225 225
However, the inhibitory mechanism of amiloride was not fullyclarified. As in the case of lactisole, amiloride was found to inhibitthe activation of the human sweet taste receptor to many types ofsweet tastants (Fig. 3A). Because the dose-dependent inhibitorycurves of amiloride for WT hT1R2/hT1R3-, hT1R2/hT1R3-A733V-,and hT1R2/hT1R3-F778A-expressing cells were not significantlydifferent (Fig. 4D), we concluded that the target site of amilorideis distinct from that of lactisole. Our result strongly indicates thatthe human sweet taste receptor has multiple inhibitor bindingsites interacting with amiloride as well as with lactisole (trans-membrane region in hT1R3 [8,9,18]). Further experiments are inprogress to elucidate the target site of amiloride by using a pointmutant of the receptor.
Recently, it was reported that the attractive salty taste is med-iated by ENaC in mouse fungiform papillae [32]. It is known thatamiloride acts as an inhibitor of ENaC with an IC50 value of 0.1–0.5 lM [24]. In this study, the IC50 of hT1R2/hT1R3 inhibition byamiloride was found to be 0.87 ± 0.20 mM (Fig. 4D). This indicatesthat amiloride acts as a sweet as well as a salty taste blocker,although the activity of sweet taste-blocking is lower than thatof salty taste-blocking.
5. Conclusion
We succeeded in constructing a cell line stably expressing thehuman sweet taste receptor (hT1R2/hT1R3) and hGa16gust44which is usable for measuring the antagonistic activity againstthe receptor. We also found that amiloride has an inhibitory effecton the human sweet taste receptor, indicating that amiloride re-duces the sweet taste intensity by inhibiting the human sweettaste receptor.
Acknowledgments
This study was partly supported by a grant from the Researchand Development Program for New Bio-industry Initiatives of theBio-oriented Technology Research Advancement Institution(BRAIN); a Grant-in-Aid for Scientific Research from the Ministryof Education, Culture, Sports, Science and Technology of Japan(18688005, 20688015, and 21658046 to T.M., and 20380183 toK.A.); and the Iijima Memorial Foundation for the Promotion ofFood Science and Technology (to T.M.).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.bbrc.2010.05.088.
References
[1] J. Chandrashekar, M.A. Hoon, N.J. Ryba, C.S. Zuker, The receptors and cells formammalian taste, Nature 444 (2006) 288–294.
[2] M.A. Hoon, E. Adler, J. Lindemeier, J.F. Battey, N.J. Ryba, C.S. Zuker, Putativemammalian taste receptors: a class of taste-specific GPCRs with distincttopographic selectivity, Cell 96 (1999) 541–551.
[3] J.P. Montmayeur, S.D. Liberles, H. Matsunami, L.B. Buck, A candidate tastereceptor gene near a sweet taste locus, Nat. Neurosci. 4 (2001) 492–498.
[4] G. Nelson, M.A. Hoon, J. Chandrashekar, Y. Zhang, N.J. Ryba, C.S. Zuker,Mammalian sweet taste receptors, Cell 106 (2001) 381–390.
[5] M. Max, Y.G. Shanker, L. Huang, M. Rong, Z. Liu, F. Campagne, H. Weinstein, S.Damak, R.F. Margolskee, Tas1r3, encoding a new candidate taste receptor, isallelic to the sweet responsiveness locus Sac, Nat. Genet. 28 (2001) 58–63.
[6] M. Kitagawa, Y. Kusakabe, H. Miura, Y. Ninomiya, A. Hino, Molecular geneticidentification of a candidate receptor gene for sweet taste, Biochem. Biophys.Res. Commun. 283 (2001) 236–242.
[7] P. Temussi, The sweet taste receptor: a single receptor with multiple sites andmodes of interaction, Adv. Food Nutr. Res. 53 (2007) 199–239.
[8] H. Xu, L. Staszewski, H. Tang, E. Adler, M. Zoller, X. Li, Different functional rolesof T1R subunits in the heteromeric taste receptors, Proc. Natl. Acad. Sci. USA101 (2004) 14258–14263.
[9] P. Jiang, M. Cui, B. Zhao, L.A. Snyder, L.M. Benard, R. Osman, M. Max, R.F.Margolskee, Identification of the cyclamate interaction site within thetransmembrane domain of the human sweet taste receptor subunit T1R3, J.Biol. Chem. 280 (2005) 34296–34305.
[10] P. Jiang, Q. Ji, Z. Liu, L.A. Snyder, L.M. Benard, R.F. Margolskee, M. Max, Thecysteine-rich region of T1R3 determines responses to intensely sweet proteins,J. Biol. Chem. 279 (2004) 45068–45075.
[11] A. Koizumi, K. Nakajima, T. Asakura, Y. Morita, K. Ito, A. Shmizu-Ibuka, T.Misaka, K. Abe, Taste-modifying sweet protein, neoculin, is received at humanT1R3 amino terminal domain, Biochem. Biophys. Res. Commun. 358 (2007)585–589.
[12] S.S. Schiffman, B.J. Booth, E.A. Sattely-Miller, B.G. Graham, K.M. Gibes, Selectiveinhibition of sweetness by the sodium salt of +/-2-(4-methoxyphenoxy)propanoic acid, Chem. Senses 24 (1999) 439–447.
[13] A. Sclafani, C. Perez, Cypha [propionic acid, 2-(4-methoxyphenol) salt] inhibitssweet taste in humans, but not in rats, Physiol. Behav. 61 (1997) 25–29.
[14] Y. Kurihara, Antisweet activity of gymnemic acid A1 and its derivatives, LifeSci. 8 (1969) 537–543.
[15] R.M. Warren, C. Pfaffmann, Suppression of sweet sensitivity by potassiumgymnemate, J. Appl. Physiol. 14 (1959) 40–42.
[16] R.S. Keast, T.M. Canty, P.A. Breslin, Oral zinc sulfate solutions inhibit sweettaste perception, Chem. Senses 29 (2004) 513–521.
[17] R.S.J. Keast, The effect of zinc on human taste perception, J. Food Sci. 68 (2003)1871–1877.
[18] M. Winnig, B. Bufe, W. Meyerhof, Valine 738 and lysine 735 in the fifthtransmembrane domain of rTas1r3 mediate insensitivity towards lactisole ofthe rat sweet taste receptor, BMC Neurosci. 6 (2005) 22.
[19] P. Jiang, M. Cui, B. Zhao, Z. Liu, L.A. Snyder, L.M. Benard, R. Osman, R.F.Margolskee, M. Max, Lactisole interacts with the transmembrane domainsof human T1R3 to inhibit sweet taste, J. Biol. Chem. 280 (2005) 15238–15246.
[20] D.V. Smith, C.A. Ossebaard, Amiloride suppression of the taste intensity ofsodium chloride: evidence from direct magnitude scaling, Physiol. Behav. 57(1995) 773–777.
[21] A.M. Tennissen, Amiloride reduces intensity responses of human fungiformpapillae, Physiol. Behav. 51 (1992) 1061–1068.
[22] N.B. McCutcheon, Human psychophysical studies of saltiness suppression byamiloride, Physiol. Behav. 51 (1992) 1069–1074.
[23] S.S. Schiffman, E. Lockhead, F.W. Maes, Amiloride reduces the taste intensity ofNa+ and Li+ salts and sweeteners, Proc. Natl. Acad. Sci. USA 80 (1983) 6136–6140.
[24] J. Teiwes, R.D. Toto, Epithelial sodium channel inhibition in cardiovasculardisease, a potential role for amiloride, Am. J. Hypertens. 20 (2007) 109–117.
[25] T. Ueda, S. Ugawa, H. Yamamura, Y. Imaizumi, S. Shimada, Functionalinteraction between T2R taste receptors and G-protein alpha subunitsexpressed in taste receptor cells, J. Neurosci. 23 (2003) 7376–7380.
[26] M. Winnig, B. Bufe, N.A. Kratochwil, J.P. Slack, W. Meyerhof, The binding sitefor neohesperidin dihydrochalcone at the human sweet taste receptor, BMCStruct. Biol. 7 (2007) 66.
[27] V. Galindo-Cuspinera, M. Winnig, B. Bufe, W. Meyerhof, P.A. Breslin, A TAS1Rreceptor-based explanation of sweet ‘water-taste’, Nature 441 (2006) 354–357.
[28] P. Thomas, T.G. Smart, HEK293 cell line: a vehicle for the expression ofrecombinant proteins, J. Pharmacol. Toxicol. Methods 51 (2005) 187–200.
[29] S. Bruzzone, S. Kunerth, E. Zocchi, A. De Flora, A.H. Guse, Spatio-temporalpropagation of Ca2+ signals by cyclic ADP-ribose in 3T3 cells stimulated viapurinergic P2Y receptors, J. Cell Biol. 163 (2003) 837–845.
[30] K. Paemeleire, P.E. Martin, S.L. Coleman, K.E. Fogarty, W.A. Carrington, L.Leybaert, R.A. Tuft, W.H. Evans, M.J. Sanderson, Intercellular calcium waves inHeLa cells expressing GFP-labeled connexin 43, 32, or 26, Mol. Biol. Cell 11(2000) 1815–1827.
[31] F. De Smedt, L. Missiaen, J.B. Parys, V. Vanweyenberg, H. De Smedt, C. Erneux,Isoprenylated human brain type I inositol 1, 4, 5-trisphosphate 5-phosphatasecontrols Ca2+ oscillations induced by ATP in Chinese hamster ovary cells, J.Biol. Chem. 272 (1997) 17367–17375.
[32] J. Chandrashekar, C. Kuhn, Y. Oka, D.A. Yarmolinsky, E. Hummler, N.J. Ryba, C.S.Zuker, The cells and peripheral representation of sodium taste in mice, Nature464 (2010) 297–301.





























