Embed Size (px)
Citation preview

logy 553 (2006) 73–81www.elsevier.com/locate/ejphar
European Journal of Pharmaco
Involvement of I2-imidazoline binding sites in positive and negativemorphine analgesia modulatory effects
Francesco Gentili a, Claudia Cardinaletti a, Antonio Carrieri b, Francesca Ghelfi a, Laura Mattioli c,Marina Perfumi c, Cristian Vesprini a, Maria Pigini a,⁎
a Dipartimento di Scienze Chimiche, Università degli Studi di Camerino, via S. Agostino 1, 62032 Camerino, Italyb Dipartimento Farmaco-Chimico, Università degli Studi di Bari, via E. Orabona 4, 70125 Bari, Italy
c Dipartimento di Medicina Sperimentale e Sanità Pubblica, Università degli Studi di Camerino, via Scalzino 3, 62032 Camerino, Italy
Received 8 February 2006; received in revised form 5 September 2006; accepted 7 September 2006Available online 26 September 2006
Abstract
Some studies, suggesting the involvement of I2-imidazoline binding sites (I2-IBS) in morphine analgesia modulation, prompted us to examine onmice antinociceptive assays the effect produced by 1 (phenyzoline), that in view of its high I2-IBS affinity and high I2-IBS selectivity with regard to I1-IBS, α2-adrenoreceptors and μ-opioid receptors might be considered the first interesting I2-IBS ligand. The study was also applied to its ortho phenylderivative 2 (diphenyzoline), designed and prepared in order to produce a possible modification of the biological profile of 1. Diphenyzoline (2)retains a significant I2-IBS selectivity with regard to I1-IBS, α2-adrenoreceptors and μ-opioid receptors. Moreover, by the functional assays 1 and 2proved inactive at all α2-adrenoreceptors subtypes up to 10−3 M. As expected, phenyzoline and diphenyzoline, which are structurally related,highlighted an interesting “positive” or “negative”, respectively, morphine analgesia modulatory effect. In fact, 1 (s.c. 10 mg/kg) enhanced morphineanalgesia (60% and 40% in mouse tail-flick andmouse hot-plate, respectively), while 2 (s.c. 10 mg/kg) decreased it (−41% and −20%, respectively).The ability to decrease morphine analgesia had never been observed before in I2-IBS ligands. These effects were not affected by i.p. treatment ofanimals with yohimbine (a selective α2-adrenoreceptor antagonist, 0.625mg/kg) or efaroxan (an I1-IBS/α2-adrenoreceptor antagonist, 1.0 mg/kg). Incontrast, they were completely reversed by i.p. treatment of animals with idazoxan (an I2-IBS/α2-adrenoreceptor antagonist, 2 mg/kg). Moreover,compound 2, in mouse tail-flick test, was able to potentiate by 23% the naloxone-induced decrease of morphine analgesia. Therefore, the results ofthis study indicate the crucial involvement of I2-IBS in the morphine analgesia modulatory effects of 1 and 2.© 2006 Elsevier B.V. All rights reserved.
Keywords: Morphine analgesia modulation; I2-IBS (I2-imidazoline binding sites); I2-IBS ligands; Phenyzoline; Diphenyzoline
1. Introduction
The existence of imidazoline binding sites (IBS) washypothesized about 20 years ago (Bousquet et al., 1984).Findings from different laboratories have shown that they arewidely distributed throughout the tissues of various species,including man, in both central and peripheral nervous systems.The numerous studies with radioligands and those of functionaltype (Molderings, 1997), apart from confirming the existence ofIBS, have also revealed their heterogeneity. At present, theyappear to be divided into I1-IBS, recognized preferentially by
⁎ Corresponding author. Tel.: +39 0737 402257; fax: +39 0737 637345.E-mail address: [email protected] (M. Pigini).
0014-2999/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.ejphar.2006.09.031
[3H]-clonidine and related compounds and I2-IBS by [3H]-idazoxan; still uncertain is the existence of I3-IBS subtypes.Even though as a result of immunological studies it has beenpossible to ascribe the IBS nature to distinct proteins in humanand rat brain, the structures of these binding proteins have notbeen identified yet (Dardonville and Rozas, 2004).
The I1-IBS, located on the plasma membrane of neurons, arepresent at a fairly high density in the region of the medullaoblungata, that contains the sites of the hypotensive action forimidazoline-like and related drugs (Heemskerk et al., 1998).Pharmacological studies suggested that I1-IBS are involved inthe regulation of cardiovascular function (Bousquet, 2001;Molderings and Gothert, 1999), in the modulation of ocularpressure (Ogidigben and Potter, 2001), and in the secretion ofrenal sodium (Smyth and Penner, 1999).

74 F. Gentili et al. / European Journal of Pharmacology 553 (2006) 73–81
The I2-IBS, located principally in the outer membrane ofmitochondria of peripheral and central tissues (Tesson and Parini,1991; Tesson et al., 1992) are divided into I2A- and I2B-IBSaccording to their affinity for amiloride, high or low, respectively(Parini et al., 1996). Biochemical and pharmacological studiessuggested a possible structural and functional correlation betweenI2-IBS and monoamine oxidases (MAOs), two mitochondrialenzymes involved in the oxidate deamination of neurotransmitters(Raddatz et al., 2000; Eglen et al., 1998). I2-IBS are involved inCNS pathologies such as Parkinson's disease (Reynolds et al.,1996), depression (Escriba et al., 1999), tolerance and addictionto opiods (Ruiz-Durantez et al., 2003). It has been described thatI2-IBS are present in brain areas involved in perception andresponse to painful stimuli (Ruggiero et al., 1998).
Since a potentiation of the analgesic effect of morphine byagmatine (proposed as endogenous ligand of IBS) (Kolesnikov etal., 1996) has been observed and a significant decrease of the IBSdensity in different brain regions after chronic morphine treatment(Su et al., 2001), it is reasonable to hypothesize IBS involvement inthe modulation of pain and the pharmacological effects of opioids.However, since agmatine binds to I2-IBS, I1-IBS and α2-adrenoreceptors in many regions of the brain (Li et al., 1994), itcould therefore be difficult to ascribe its modulatory activity onmorphine antinociception to its binding to one of these receptortypes.
Recently a number of studies suggested that some substancesdisplaying high affinity toward the I2-IBS with regard to α2-adrenoceptors such as BFI (Hudson et al., 1995; Alemany et al.,1997) and tracizoline (Pigini et al., 1997), named alsovalldemossine or LSL 61122 (Ozaita et al., 1997) were able,analogously to agmatine, to regulate opioid-induced analgesia(Sanchez-Blazquez et al., 2000), and to attenuate the develop-ment of tolerance and dependence (Boronat et al., 1998). More-over, the I2-IBS ligands exhibiting no antinociceptive effects bythemselves but able to potentiate the analgesic effect ofmorphine, have been indicated as agonists, while those, suchas idazoxan, that by co-treatment completely reverse this poten-tiation, have been considered antagonists (Sanchez-Blazquezet al., 2000).
Despite the intense efforts in the field, there is still a great dealof uncertainty about the involvement of I2-IBS in the modulationof pain, also because the most commonly used ligands are gen-erally scantly selective with regard to the two main subtypes I1-and I2-IBS, and a possible role of the I1-IBS in analgesia cannotbe ruled out (Millan, 2002).
In the present study, our lead is compound 1 (Phenyzoline)(Fig. 1), selected for its very high I2-IBS affinity and high I2-IBS
Fig. 1
selectivitywith regard to I1-IBS andα2-adrenoceptors (pKi I2=8.60;pKi I1=5.43; pKi α2=5.70; I2/I1=1479; I2/α2=794) (Gentili et al.,2003). Analogously to what was demonstrated by us for ligandsinteracting with α2-adrenoceptors (Gentili et al., 2002) and I1-IBS(Gentili et al., 2005), in order to produce a possible modulation ofbiological profile of 1, we designed and prepared its ortho phenylderivative, compound 2 (Diphenyzoline), already known (Jones andDimopoulos, 2000) but never considered from this point of view(Fig. 1). The affinity values at I2-IBS, I1-IBS and α2-adrenoceptorsof 2 and the values atμ-opioid receptors of 1 and 2were determined.Compounds 1 and 2 were tested on two algesiometric paradigms:mouse hot-plate and mouse tail-flick tests; the previously studiedtracizoline (Pigini et al., 1997) or valldemossine (Sanchez-Blazquezet al., 2000) has been included in this studywith the aim to verify thevalidity of our experimental conditions. To confirm the role playedby I2-IBS in themodulation ofmorphine analgesia, the effects of thepre-treatment with idazoxan (a mixed I2-IBS/α2-adrenoceptorantagonist), efaroxan (a mixed I1-IBS/α2-adrenoceptor antagonist),and yohimbine (a selective α2-adrenoceptor antagonist) wereevaluated; a pre-treatment with naloxone (an opioid receptorantagonist) was also performed. In addition, 1 and 2were subjectedto functional assays at α2-adrenoceptor subtypes.
Finally, the molecular structures of 1, 2, tracizoline andidazoxan were superposed.
2. Materials and methods
2.1. Animals
Male CD-1 mice (Charles River Breeding Laboratory,Bloomington, MA, USA), weighing 25–35 g were employed.Animals were kept in a room with a reverse 12:12 h light/darkcycle (lights off at 10:00 a.m.), temperature of 20–22 °C andhumidity of 45–55%. They were offered free access to tap waterand food pellets (4RF18, Mucedola, Settimo Milanese, Italy).The animals were used only in one experimental session. Animaltesting was carried out according to the European CommunityCouncil Directive of 24 November 1986 (86/609/EEC).
2.2. Drugs
Phenyzoline (1) and tracizoline were synthesized as reportedby us elsewhere (Pigini et al., 1997). Diphenyzoline (2) wasprepared from 2-biphenyl-carboxaldehyde according to the pro-cedures used for the synthesis of the corresponding α-methylderivative (Gentili et al., 2005).
.

75F. Gentili et al. / European Journal of Pharmacology 553 (2006) 73–81
Morphine-hydrochloride was purchased from Salars (S.p.a,Como, Italy), naloxone-hydrochloride and efaroxan-hydrochlo-ride from Tocris (Cookson Ltd, UK), while idazoxan-hydro-chloride and yohimbine-hydrochloride were both purchasedfrom Sigma (St. Louis, MO).
Phenyzoline (1), diphenyzoline (2), and tracizoline, weretested as oxalate salts. In the in vivo studies, drugs were dis-solved in distilled water immediately before use, with the ex-ception ofmorphine-hydrochloride, efaroxan-hydrochloride andyohimbine-hydrochloride that were dissolved in saline. 1, 2,tracizoline and morphine-hydrochloride were subcutaneously(s.c.) injected, while naloxone-hydrochloride, idazoxan-hydro-chloride, efaroxan-hydrochloride and yohimbine-hydrochloridewere intraperitoneally (i.p.) administered.
2.3. Radioligand binding assays at I2-IBS, I1-IBS, α2-adrenoceptors and μ-opioid receptors
Affinity values at I2-IBS, I1-IBS and α2-adrenoceptors ofdiphenyzoline (2) were evaluated following already describedprocedures (Gentili et al., 2003) using membranes of rabbitkidney, rat pheochromocytoma cells (PC 12) and rat cortex,respectively. The radioligands used were [3H]-idazoxan (5 nM,I2), [
125I]-p-iodoclonidine (0.5 nM, I1), [3H]-clonidine (2 nM,
α2), and non-specific binding was defined by inclusion of 10 μMcirazoline (I2, 10%), 10 μM BDF 6143 (I1, 35%) and 10 μMphentolamine (α2, 25%).
Affinity values at μ-opioid receptors of phenyzoline (1) anddiphenyzoline (2) and naloxone, for useful comparison, wereevaluated in membranes of rat brain (Yoburn et al., 1991; Diazet al., 2000). Male Sprague Dawley rats (200–300 g, CharlesRiver, Italy) were killed by cervical dislocation, and the brains(without cerebellum) were homogenized (2×20 s) in 50 vol ofcold 50 mM Tris buffer pH 7.4, using a Politron homogenizer(Kinematica). Homogenates were centrifuged at 48,000 ×g for15 min, resuspended in 50 vol of the same buffer, incubated at37 °C for 15 min and centrifuged and resuspended twice more.The final pellets were frozen and stored at −80 °C until use. Onexperimental section, pellets were resuspended in 100 vol of50 mM Tris buffer pH 7.4, membranes were incubated in a finalvolume of 1 ml for 30 min at 25 °C with 1 nM [3H]DAMGO inabsence or presence of competing drugs. Non-specific bindingwas determined in the presence of 1 μM naloxone. The in-cubation was stopped by addition of cold Tris buffer pH 7.4 and
Table 1Binding affinities (pKi
a)
Compd pKi (I2) pKi (I1) pKi (α2)
Tracizoline 8.72±0.13 7.72±0.14 4.85±0.1 Phenyzoline 8.60±0.04 5.43±0.13 5.70±0.2 Diphenyzoline 6.80±0.05 5.20±0.01 5.15±0.Naloxone n.d.c n.d.c n.d.c
apKi affinity values for I2-IBS, I1-IBS, α2-adrenoceptors and μ-opiod receptor were m(rabbit kidney membranes), [125I]-p-iodoclonidine (rat pheochromocytoma cells,membranes), respectively. Values are the mean±S.E.M. of, in each case, a minimum opKi (I2) and pKi (α2) (I2/α2), pKi (I2) and pKi (μ) (I2/μ) values.
cNot determined.
rapid filtration through 0.2% polyethyleneimine pre-treatedSchleicher and Schuell GF52 filters. The filters were thenwashed with cold buffer and the radioactivity retained on thefilters was counted by liquid scintillation spectrometry.
2.4. Data analysis
The inhibition of specific binding of the radioligands bythe tested drugs was analyzed to estimate the IC50 value usingthe non-linear curve-fitting program Allfit (DeLean et al.,1978). The IC50 value was converted to an affinity constant(Ki) by the equation of Cheng and Prusoff (1973). Values arethe mean±S.E.M. of, in each case, a minimum of six ex-periments and are expressed as pKi (− log Ki) ±S.E.M.
2.5. Antinociceptive assays
To study the modulatory effects of phenyzoline (1), dipheny-zoline (2) and tracizoline in morphine antinociception, the tail-flick and the hot-plate tests were used; it is known that eachmethod reflects nociception at different levels of the central ner-vous system: the tail-flick test reflects predominantly a spinalresponse, whereas the hot-plate test a sovraspinal response (Lipp,1991; Langerman et al., 1995; Gardmark et al., 1998; Malmbergand Bannon, 1999).
2.5.1. Tail-flick testThe latency time was assessed by a tail-flick analgesymeter
(U. Basile, Italy). Radiant heat was focused on a blackened spot1–2 cm from the tip of the mouse-tail and the latency to tail-flickwas recorded. Beam intensity was adjusted to give a tail-flicklatency of 2–3 s in controls. In order to avoid tissue damage, acut-off latency of 12–15 s was used. Each mouse was tested 1and 0.5 h before vehicle or compounds administration todetermine baseline latency. Then mice received s.c. tracizoline(Sanchez-Blazquez et al., 2000), phenyzoline (1) or diphe-nyzoline (2) (10 mg/kg) and related vehicle, 30 min before s.c.morphine (5.0 mg/kg) administration or its vehicle. The anti-nociceptive activity was evaluated 30 min after morphineinjection.
In order to demonstrate the selective involvement of I2-IBSin the effects of phenyzoline (1) and diphenyzoline (2) on anti-nociception induced by morphine, animals were pre-treatedwith different drugs at doses effectively blocking the responses
pKi (μ) I2/I1b I2/α2
b I2/μb
15 n.d.c 10 7431 –06 b5 1479 794 398117 5.7±0.01 40 45 13
8.85±0.04 – – –
easured by assessing the ability of the test compounds to displace [3H]-idazoxanPC 12), [3H]-clonidine (rat cortex membranes), and [3H]DAMGO (rat brainf six experiments. bAntilog of the difference between pKi (I2) and pKi (I1) (I2/I1),

Fig. 2. Effect of tracizoline (10 mg/kg SC) or phenyzoline (1) (10 mg/kg s.c.) onmorphine (5.0 mg/kg, s.c.) analgesia in the tail-flick test (Panel A) and in the hot-plate test (Panel B). The reaction latencies were expressed as a percent of theMaximum Possible Effect (%MPE). Each column represents the mean±S.E.M.of 12–14 animals. Significant differences: ++pb0.01 compared to control group;⁎⁎pb0.01 compared to morphine alone-treated group, using Student's t-test;where not indicated, the difference was not statistically significant.
76 F. Gentili et al. / European Journal of Pharmacology 553 (2006) 73–81
mediated by I2-IBS, I1-IBS and α2-adrenoceptors systems. Inparticular, mice were pre-treated with idazoxan (2.0 mg/kg, i.p.,a mixed I2-IBS/α2-adrenoceptor antagonist) (Kolesnikov et al.,1996), efaroxan (1 mg/kg, i.p., a mixed I1-IBS/α2-adrenoceptorantagonist) (Santos et al., 2005), or vehicle (10 ml/kg, i.p.)respectively 5 and 15 min before being subjected to the testedcompounds injections; with yohimbine (0.625 mg/kg, i.p., aselective α2-adrenoceptor antagonist) (Yesilyurt and Uzbay,2001) 15 min before tested compounds injections; the tail-flicklatencies were assessed 30 min after the morphine injection.Finally, in the case of diphenyzoline (2), a pre-treatment withthe potent opioid receptor antagonist naloxone was also per-formed. First of all, to check the dose of naloxone which wasable to reduce by 50% the effect of morphine, mice received ani.p. dose of naloxone (0.5, 1.0 and 3.0 mg/kg) or vehicle, 40 minbefore morphine injection (5.0 mg/kg) 30 min before the tail-flick test. Then, they received i.p. naloxone (0.5 mg/kg) or itsvehicle 10 min before s.c. compound 2 (10 mg/kg) or relatedvehicle. 30 min after the second injection, all groups receiveds.c. morphine (5.0 mg/kg), 30 min before the tail-flick test.
2.5.2. Hot-Plate TestPain reflexes in response to a thermal stimulus were measured
using the hot-plate test. The surface of the hot-plate (Basile,Milano) was heated to a constant temperature of 55 °C to give alatency time of 15–18 s in controls. Mice were placed on the hot-plate, and the latency to respond with either a hind paw lick orjump was measured. In order to avoid tissue damage, a cut-offlatency of 60 s was used. Each mouse was tested 1 and 0.5 hbefore vehicle or compounds administration to determine base-line latency. Then animals received s.c. tracizoline, phenyzoline(1) and diphenyzoline (2) (10 mg/kg) or related vehicle, 30 minbefore s.c. morphine (5.0 mg/kg) or related vehicle administra-tion. The antinociceptive activity was evaluated 30 min aftermorphine injection.
2.5.3. Functional assays on α2-adrenoceptor subtypes. Cyto-sensor microphysiometry
With the purpose of further investigating the possible roleplayed by α2-adrenergic system in the morphine analgesia mod-ulatory effects of phenyzoline (1) and diphenyzoline (2), thesecompounds were also tested on human α2-adrenoceptor subtypesexpressed in Chinese hamster ovary (CHO) cells by use of aCytosensor microphysiometer instrument (Molecular Devices,CA, U.S.A.). With this instrument it is possible to determinethe functional properties of the tested compounds, measuringthe rate of extracellular acidification after treatment of the cellswithincreased doses of the ligands. Our experiments were performedaccording to the previously described procedures and (−)-noradrenaline was used as agonist reference (Gentili et al., 2004).
2.5.4. Statistical analysisAntinociceptive effect was expressed as a percent of the
Maximum Possible Effect (MPE) according to the followingformula (Sanchez-Blazquez et al., 2000): %MPE=(measuredlatency−basal latency) / (cut-off time−basal latency)×100%.Data are reported as means±S.E.M. The results were analyzed
by multifactorial split-plot analysis of variance (ANOVA).Post-hoc comparisons were made by means of the Student–Newman–Keuls test. Statistical significance was set at pb0.05.
2.6. Molecular modeling
Molecular superposition of phenyzoline (1), diphenyzoline(2), tracizoline and idazoxan, was performed by means of theTFIT module implemented in the QXP software package(McMartin and Bohacek, 1997). The internal geometry of allthe ligands was randomly perturbed through 5000 cycles ofMonte Carlo conformational search. All the compounds werebuilt in their protonated state using the fragment library of thesame software; for idazoxan (S)-equatorial configuration and forthe other compounds planar conformations were selectedaccording to our previously published I2-IBS CoMFA model(Carrieri et al., 1997). The conformation achieved by the ligandsin the molecular superposition was for all of them less than1 kcal/mol with respect to the relative local energy minimum.

Fig. 3. Effect of phenyzoline (1) (10 mg/kg s.c.) on morphine (5.0 mg/kg, s.c.)analgesia after idazoxan (2.0 mg/kg, i.p.) (Panel A), efaroxan (1.0 mg/kg, i.p.)(Panel B) and yohimbine (0.625 mg/kg, i.p.) (Panel C) pre-treatment, in the tail-flick test. The reaction latencies were expressed as a percent of the MaximumPossible Effect (%MPE). Each column represents the mean±S.E.M. of 8–12animals. Significant differences: ⁎pb0.05, ⁎⁎pb0.01 compared to morphinealone-treated group; °°pb0.01 compared to morphine/1 combination treatedgroup, using Student's t test; where not indicated, the difference was notstatistically significant.
77F. Gentili et al. / European Journal of Pharmacology 553 (2006) 73–81
3. Results
3.1. Radioligand binding assays at I2-IBS, I1-IBS, α2-adrenoceptors and μ-opioid receptors
The affinity values, expressed as pKi, at I2-IBS, I1-IBS, α2-adrenoceptors and μ-opioid receptors of compounds 1 and 2 arereported in Table 1, together with those of tracizoline andnaloxone for useful comparison. They show that 2, similarly to1, displays good affinity for I2-IBS (pKi 6.80), and low affinityfor the I1-IBS and α2-adrenoceptors (pKi 5.20 and 5.15,respectively). Both the compounds 1 and 2 display low affinityfor μ-opioid receptors (pKib5 and 5.7, respectively). Highaffinity is instead shown by naloxone (pKi 8.85).
3.2. Antinociceptive tests
In the algesiometric tests, tracizoline used alone had noanalgesic effect, but produced a 50% increase of the analgesicefficacy of the morphine test dose, as evidenced by increased
Fig. 4. Effect of diphenyzoline (2) (10 mg/kg, s.c.) on morphine (5.0 mg/kg, s.c.)analgesia in the tail-flick test (Panel A) and in the hot-plate test (Panel B). Thereaction latencies were expressed as a percent of the Maximum Possible Effect(%MPE). Each column represents the mean±S.E.M. of 11–12 animals.Significant differences: ++pb0.01 compared to control group; ⁎pb0.05,⁎⁎pb0.01 compared to morphine alone-treated group, using Student's t-test;where not indicated, the difference was not statistically significant.
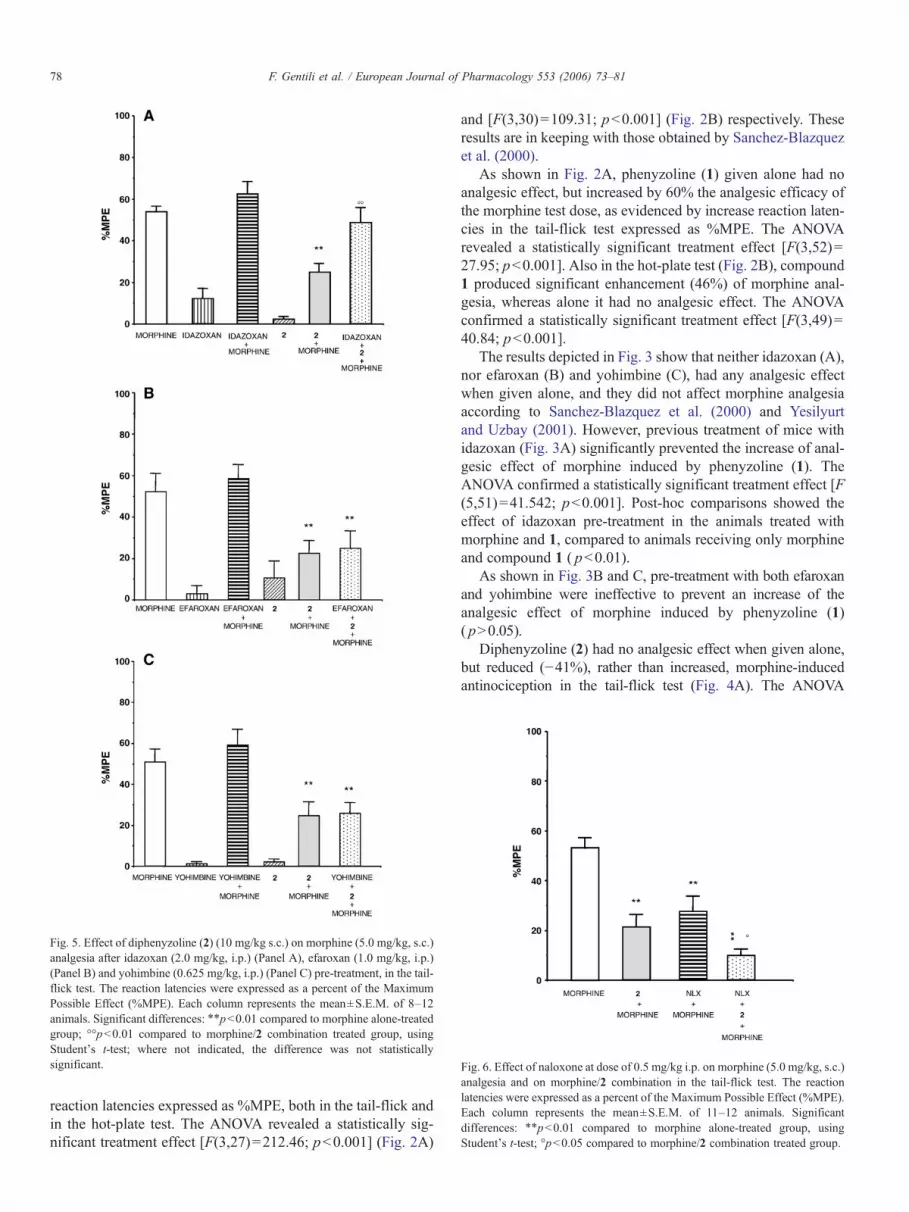
Fig. 6. Effect of naloxone at dose of 0.5 mg/kg i.p. on morphine (5.0 mg/kg, s.c.)analgesia and on morphine/2 combination in the tail-flick test. The reactionlatencies were expressed as a percent of the Maximum Possible Effect (%MPE).Each column represents the mean±S.E.M. of 11–12 animals. Significantdifferences: ⁎⁎pb0.01 compared to morphine alone-treated group, usingStudent's t-test; °pb0.05 compared to morphine/2 combination treated group.
Fig. 5. Effect of diphenyzoline (2) (10 mg/kg s.c.) on morphine (5.0 mg/kg, s.c.)analgesia after idazoxan (2.0 mg/kg, i.p.) (Panel A), efaroxan (1.0 mg/kg, i.p.)(Panel B) and yohimbine (0.625 mg/kg, i.p.) (Panel C) pre-treatment, in the tail-flick test. The reaction latencies were expressed as a percent of the MaximumPossible Effect (%MPE). Each column represents the mean±S.E.M. of 8–12animals. Significant differences: ⁎⁎pb0.01 compared to morphine alone-treatedgroup; °°pb0.01 compared to morphine/2 combination treated group, usingStudent's t-test; where not indicated, the difference was not statisticallysignificant.
78 F. Gentili et al. / European Journal of Pharmacology 553 (2006) 73–81
reaction latencies expressed as %MPE, both in the tail-flick andin the hot-plate test. The ANOVA revealed a statistically sig-nificant treatment effect [F(3,27)=212.46; pb0.001] (Fig. 2A)
and [F(3,30)=109.31; pb0.001] (Fig. 2B) respectively. Theseresults are in keeping with those obtained by Sanchez-Blazquezet al. (2000).
As shown in Fig. 2A, phenyzoline (1) given alone had noanalgesic effect, but increased by 60% the analgesic efficacy ofthe morphine test dose, as evidenced by increase reaction laten-cies in the tail-flick test expressed as %MPE. The ANOVArevealed a statistically significant treatment effect [F(3,52)=27.95; pb0.001]. Also in the hot-plate test (Fig. 2B), compound1 produced significant enhancement (46%) of morphine anal-gesia, whereas alone it had no analgesic effect. The ANOVAconfirmed a statistically significant treatment effect [F(3,49)=40.84; pb0.001].
The results depicted in Fig. 3 show that neither idazoxan (A),nor efaroxan (B) and yohimbine (C), had any analgesic effectwhen given alone, and they did not affect morphine analgesiaaccording to Sanchez-Blazquez et al. (2000) and Yesilyurtand Uzbay (2001). However, previous treatment of mice withidazoxan (Fig. 3A) significantly prevented the increase of anal-gesic effect of morphine induced by phenyzoline (1). TheANOVA confirmed a statistically significant treatment effect [F(5,51)=41.542; pb0.001]. Post-hoc comparisons showed theeffect of idazoxan pre-treatment in the animals treated withmorphine and 1, compared to animals receiving only morphineand compound 1 ( pb0.01).
As shown in Fig. 3B and C, pre-treatment with both efaroxanand yohimbine were ineffective to prevent an increase of theanalgesic effect of morphine induced by phenyzoline (1)( pN0.05).
Diphenyzoline (2) had no analgesic effect when given alone,but reduced (−41%), rather than increased, morphine-inducedantinociception in the tail-flick test (Fig. 4A). The ANOVA

Fig. 7. Dose–response curves of (−)-Noradrenaline (empty square), phenyzoline (1) (full circle) and diphenyzoline (2) (full triangle) in CHO cells stably expressing thethree human α2-AR subtypes, α2A, α2B, α2C. Agonist potency was determined by use of a Cytosesor microphysiometer instrument, data points represent the mean ofthree to six separate experiments.
79F. Gentili et al. / European Journal of Pharmacology 553 (2006) 73–81
confirmed a statistically significant treatment effect [F(3,42)=27.02; pb0.001]. Also in the hot-plate test (Fig. 4B), compound 2reduced morphine analgesia (−20%), whereas alone it had noanalgesic effect. The ANOVA confirmed a statistically significanttreatment effect [F(3,51)=48.12; pb0.01].
The results depicted in Fig. 5 confirm that neither idazoxan(A), nor efaroxan (B) and yohimbine (C), had any analgesic effectwhen given alone, and they did not affect morphine analgesia.However, previous treatment with idazoxan (Fig. 5A) completelyreversed the decrease of morphine-analgesia induced by 2. TheANOVA confirmed a statistically significant treatment effect [F(5,52)=20.370; pb0.001]. Post-hoc comparisons showed theeffect of idazoxan pre-treatment in the animals treated withmorphine/compound 2 combination, compared to animals re-ceiving only morphine/compound 2 combination ( pb0.01).
As shown in Fig. 5B and C, pre-treatment with both efaroxanand yohimbine were ineffective in reversing the decrease ofanalgesic effect of morphine induced by 2, as confirmed by theStudent–Newman–Keuls test (pN0.05).
In the preliminary experiment, treatment with opioidantagonist naloxone at doses of 0.5, 1.0 or 3.0 mg/kg produceddose-dependent reductions in morphine-antinociception, 58%,70% or 90%, respectively, in the tail-flick test. The ANOVAconfirmed a statistically significant treatment effect [F(3,36)=15.47; pb0.001] (data not shown).
As shown in Fig. 6, diphenyzoline (2) in the tail-flick testsynergically and significantly enhanced (23%) the decrease ofmorphine analgesia produced by the opioid antagonist naloxone
Fig. 8. Molecular overlay of (S)-idazoxan (white), tracizoline (green),phenyzoline (1) (cyan) and diphenyzoline (2) (red) derived by QXP.
(0.5 mg/kg). The ANOVA confirmed a statistically significanttreatment effect [F(3,36)=10.55; pb0.001]. Post-hoc compar-isons showed the effect of pre-treatment with naloxone bothin the animals receiving morphine alone ( pb0.01) and themorphine/compound 2 combination ( pb0.05).
3.3. Functional assays on α2-adrenoceptor subtypes. Cytosensormicrophysiometry
As shown in Fig. 7, neither phenyzoline (1) nor diphenyzo-line (2) had any agonist activity (up to 1.10−3 M.) at all α2-adrenoreceptor subtypes.
3.4. Molecular modeling
Themain evidence deriving from themolecular superpositionof the examined compounds is that they seem to share a similarconformation, as can be perceived from Fig. 8. In fact, thecommon structural features, namely the charged imidazolinering, the phenyl moiety and the two atoms bridge, are closelysuperposed and therefore should occupy the same receptorspace. As a consequence of this reliable alignment, a super-position energy of −133.00 kcal/mol is calculated (the lower thevalue, the better the fit). A second interesting result concerns thearomatic substituent characterizing compound 2 which pointstowards a region not occupied by the other three ligands. In thecase of idazoxan, the molecule shares a more puckered and lessflexible conformation due to the benzodioxane system.
4. Discussion
Pain represents a notable medical problem: it could be tackledby the rational design of compounds acting both at the opioidreceptors (morphine-like compounds) and non-opioid receptors.Among them, the α2-adrenergic system might also be included,whose agonist ligands, endowed with specific analgesic pro-perties, are also able to synergically interact with opioids.Nevertheless, their side effects such as sedation (Hunter et al.,1997; Lakhlani et al., 1997;Mizobe et al., 1996), epileptogenesis(Janumpalli et al., 1998), hypothermia (Hunter et al., 1997) andhypotension (MacMillan et al., 1996) limit their therapeuticutility.

80 F. Gentili et al. / European Journal of Pharmacology 553 (2006) 73–81
Therefore, the discovery of I2-IBS ligands able to enhance theantinociception induced by morphine (Sanchez-Blazquez et al.,2000) proved particularly interesting.
The important role attributed to I2-IBS in the morphineanalgesia modulation and the fact that the compounds mainlystudied, such as tracizoline and BFI, also possess significant I1-IBS affinity (Sanchez-Blazquez et al., 2000; Gentili et al., 2003;Eglen et al., 1998), prompted us to examine the effect producedby phenyzoline (1) that, in view of its high I2-IBS selectivity,might be considered the first interesting I2-IBS ligand (Table 1).
The study was also applied to its ortho phenyl derivative,diphenyzoline (2) designed and prepared in order to produce apossible modification of the biological profile of 1. The resultsreported in Table 1 show that 2 has good affinity for I2-IBS, andretains a significant I2-IBS selectivity vs I1-IBS and α2-adrenoceptors (I2/I1 40; I2/α2 45).
In algesiometric tests, phenyzoline (1) and diphenyzoline (2)exhibited no antinociceptive effects by themselves, but wereable to produce significant morphine analgesia modulation. Infact, while 1 enhanced morphine analgesia by 60% and 46% inthe tail-flick and the hot-plate, respectively, 2 decreased it by−41% and −20%, respectively. Moreover, compound 2 in thetail-flick test, was able to potentiate by 23% the naloxone-induced decrease of morphine antinociceptive effect. The abilityto reduce morphine analgesia had never been observed before inI2-IBS ligands. These effects proved to be mediated by I2-IBS,since they were completely reversed by the I2-IBS/α2-adreno-ceptor antagonist idazoxan, but not by the I1-IBS/α2-adreno-ceptor antagonist efaroxan, and the selective α2-adrenoceptorantagonist yohimbine. The involvement of α2-adrenoceptors inthe effects produced by 1 and 2 on morphine analgesia modu-lation could also be ruled out by the functional assays wherethese compounds proved inactive at all α2-adrenoceptor sub-types up to 10−3 M. Taken as a whole, these data indicate that I2-IBS, but not I1-IBS and α2-adrenoceptors, are involved in themorphine analgesia modulatory effects of 1 and 2.
In addition we superposed the structures of the I2-IBS ligands1, 2, tracizoline and (S)-idazoxan in an attempt to find themolecular determinants responsible for their observed differentbiological profile. All the compounds share similar highly fittedstructural features, such as the charged imidazoline, the phenylring and the two bridge spacing atoms, which proved to beconstrained in the benzodioxane scaffold of idazoxan. Interest-ingly, the aromatic substituent characterizing the diphenyzoline(2) points toward a region not occupied by the other three li-gands. It is probable that this phenyl substituent, introduced inthe ortho position of the aromatic ring of phenyzoline (1), pro-duced the determinant biological profile modulation of the latter.We previously observed that the same chemical modification instructurally related compounds preferentially interacting withα2-adrenoceptors (Gentili et al., 2002) and I1-IBS (Gentili et al.,2005), produced a drastic biological profile modulation (fromantagonist to agonist activity). This observation prompts us tosuppose the existence of some structural analogies between theα2-adrenoceptors, I1-IBS and I2-IBS, and to hypothesize that thephenyl substituent of 2, through π–π stacking interactions withan aromatic cluster, triggered the I2-binding proteins activation.
Therefore, diphenyzoline (2) could be defined as a “putative I2-IBS agonist”. Moreover, idazoxan, probably due to its veryreduced conformational freedom and enhanced steric hindrance,retained the character of “neutral antagonist” according toSanchez-Blazquez et al. (2000). Consequently, phenyzoline (1)such as tracizoline might be considered “putative I2-IBS inverseagonists”.
Finally, from the binding data, it emerged that phenyzoline(1) and diphenyzoline (2) had low affinity for μ-opioid receptors(pKib5 and 5.7, respectively; the pKi of naloxone, inserted foruseful comparison, is 8.85) (Table 1); in view of the afore-mentioned results, this slight affinity suggests that the significantmodulatory effects observed were not produced by the inter-action of 1 and 2 with μ-opioid receptor orthosteric sites. There-fore, we can speculate that the “putative I2-IBS inverse agonists”1 and tracizoline (positive regulators), and “putative I2-IBSagonist” 2 (negative regulator), interacting with I2-IBS couldinduce a perturbation of μ-opioid receptors to higher or lower,respectively, affinity state conformation for opioid ligands.
Phenyzoline (1) and diphenyzoline (2) constitute an impor-tant basis for future work, but in order to gain more insight intovery interesting “positive” and “negative” modulatory effectsmediated by I2-IBS, the design and the synthesis of new I2-IBSsubtype selective ligands are already being planned.
Acknowledgement
The present workwas supported by a grant from theUniversityof Camerino and Cofinanziamento MIUR 2005, Italy.
References
Alemany, R., Olmos, G., Garcìa-Sevilla, J.A., 1997. Labelling of I2B-imidazoline receptors by [3H]2-(2-benzofuranyl)-2-imidazoline (2-BFI) inrat brain and liver: characterization, regulation and relation to monoamineoxidase enzymes. Naunyn-Schmiedeberg's Arch. Pharmacol. 356, 39–47.
Boronat, M.A., Olmos, G., García-Sevilla, J.A., 1998. Attenuation of toleranceto opioid-induced antinociception and protection against morphine-induceddecrease of neurofilament proteins by idazoxan and other I2-imidazolineligands. Br. J. Pharmacol. 125, 175–185.
Bousquet, P., 2001. I1 receptors, cardiovascular function, and metabolism. Am.J. Hypertens. 14, 317S–321S.
Bousquet, P., Feldman, J., Schwartz, J., 1984. Central cardiovascular effects ofalpha adrenergic drugs: differences between catecholamines and imidazo-lines. J. Pharmacol. Exp. Ther. 230, 232–236.
Carrieri, A., Brasili, L., Leonetti, F., Pigini, M., Giannella, M., Bousquet, P.,Carotti, A., 1997. 2-D and 3-D modeling of imidazoline receptor ligands:insights into pharmacophore. Bioorg. Med. Chem. 5, 843–856.
Cheng, Y.C., Prusoff, W.H., 1973. Relationship between the inhibition constant(Ki) and the concentration of inhibitor which causes 50 percent inhibition(IC50) of an enzymatic reaction. Biochem. Pharmacol. 22, 3099–3108.
Dardonville, C., Rozas, I., 2004. Imidazoline binding sites and their ligands: anoverview of the different chemical structures. Med. Res. Rev. 24, 639–661.
DeLean, A., Munson, P.J., Rodbard, D., 1978. Simultaneous analysis of familiesof sigmoidal curves: application to bioassay, radioligand assay, andphysiological dose–response curves. Am. J. Physiol. 235, E97–E102.
Diaz, A., Pazos, A., Florez, J., Hurle, M.A., 2000. Autoradiographic mapping ofμ-opioid receptors during opiate tolerance and supersensitivity in the ratcentral nervous system. Naunyn-Schmiedeberg's Arch. Pharmacol. 362,101–109.

81F. Gentili et al. / European Journal of Pharmacology 553 (2006) 73–81
Eglen, R.M.,Hudson,A.L., Kendall, D.A.,Nutt, D.J.,Morgan,N.G.,Wilson,V.G.,Dillon, M.P., 1998. ‘Seeing through a glass darkly’: casting light onimidazoline ‘I’ sites. Trends Pharmacol. Sci. 19, 381–390.
Escriba, P.V., Ozaita, A., Garcia-Sevilla, J.A., 1999. Pharmacologicalcharacterization of imidazoline receptor proteins identified by immunologictechniques and other methods. Ann. N. Y. Acad. Sci. 881, 8–25.
Gardmark, M., Hoglund, A.U., Hammarlund-Udenaes, M., 1998. Aspects ontail-flick, hot-plate and electrical stimulation tests for morphine antinocicep-tion. Pharmacol. Toxicol. 83, 252–258.
Gentili, F., Bousquet, P., Brasili, L., Caretto, M., Carrieri, A., Dontenwill, M.,Giannella, M., Marucci, G., Perfumi, M., Piergentili, A., Quaglia, W.,Rascente, C., Pigini, M., 2002. α2-Adrenoreceptors profile modulation andhigh antinociceptive activity of (S)-(−)-2-[1-(biphenyl-2-yloxy)ethyl]4,5-dihydro-1H-imidazole. J. Med. Chem. 45, 32–40.
Gentili, F., Bousquet, P., Brasili, L., Dontenwill, M., Feldman, J., Ghelfi, F.,Giannella, M., Piergentili, A., Quaglia, W., Pigini, M., 2003. Imidazolinebinding sites (IBS) profile modulation: key role of the bridge in determiningI1-IBS or I2-IBS selectivity within a series of 2-phenoxymethylimidazolineanalogues. J. Med. Chem. 46, 2169–2176.
Gentili, F., Ghelfi, F., Giannella, M., Piergentili, A., Pigini, M., Quaglia, W.,Vesprini, C., Crassous, P.A., Paris, H., Carrieri, A., 2004. α2-Adrenorecep-tors profile modulation. 2.1 Biphenyline analogues as tools for selectiveactivation of the α2C-subtype. J. Med. Chem. 47, 6160–6173.
Gentili, F., Bousquet, P., Carrieri, A., Feldman, J., Ghelfi, F., Giannella, M.,Piergentili, A., Quaglia, W., Vesprini, C., Pigini, M., 2005. Rational designof the new antihypertensive I1-receptor ligand 2-(2-biphenyl-2-yl-1-methyl-ethyl)-4,5-dihydro-1H-imidazole. Lett. Drug Des. Discov. 2, 571–578.
Heemskerk, F.M., Dontenwill, M., Greney, H., Vonthron, C., Bousquet, P.,1998. Evidence for the existence of imidazoline-specific binding sites insynaptosomal plasma membranes of the bovine brainstem. J. Neurochem.71, 2193–2202.
Hudson, A.L., Mallard, N.J., Nutt, D.J., Chapleo, C.B., 1995. Affinity andselectivity of 2-(2-benzofuranyl)-2-imidazoline for mammalian brain non-adrenoceptor idazoxan binding sites (I2-sites). Br. J. Pharmacol. 114(Suppl.), 411P.
Hunter, J.C., Fontana, D.J., Hedley, L.R., Jasper, J.R., Lewis, R., Link, R.E.,Secchi, R., Sutton, J., Eglen, R.M., 1997. Assessment of the role of α2-adrenoceptor subtypes in the antinociceptive, sedative and hypothermicaction of dexmedetomidine in transgenic mice. Br. J. Pharmacol. 122,1339–1344.
Janumpalli, S., Butler, L.S., MacMillan, L.B., Limbird, L.E., McNamara, J.O.,1998. A point mutation (D79N) of the α2A adrenergic receptor abolishes theantiepileptogenic action of endogenous norepinephrine. J. Neurosci. 18,2004–2008.
Jones, R.C.F., Dimopoulos, P., 2000. A new protocol for the synthesis of N(1)-unsubstituted 2-substituted 2-imidazolines. Tetrahedron 56, 2061–2074.
Kolesnikov, Y., Jain, S., Pasternak, G.W., 1996. Modulation of opioid analgesiaby agmatine. Eur. J. Pharmacol. 296, 17–22.
Lakhlani, P.P., MacMillan, L.B., Guo, T.Z., McCool, B.A., Lovinger, D.M.,Maze, M., Limbird, L.E., 1997. Substitution of a mutant alpha2a-adrenergicreceptor via “hit and run” gene targeting reveals the role of this subtype insedative, analgesic, and anesthetic-sparing responses in vivo. Proc. Natl.Acad. Sci. U. S. A. 94, 9950–9955.
Langerman, L., Zakowski, M.I., Piskoun, B., Grant, G.J., 1995. Hot plate versustail flick: evaluation of acute tolerance to continuous morphine infusion inthe rat model. J. Pharmacol. Toxicol. Methods 34, 23–27.
Li, G., Regunathan, S., Barrow, C.J., Eshraghi, J., Cooper, R., Reis, D.J., 1994.Agmatine an endogenous clonidine-displacing substance in the brain.Science 263, 966–969.
Lipp, J., 1991. Possible mechanism of morphine analgesia. Clin. Neurophar-macol. 14, 131–147.
MacMillan, L.B., Hein, L., Smith, M.S., Piascik, M.T., Limbird, L.E., 1996.Central hypotensive effects of the alpha2a-adrenergic receptor subtype.Science 273, 801–803.
Malmberg, A.B., Bannon, A.W., 1999. Models of nociception: hot-plate, tail-flick,and formalin tests in rodents. In: Crawley, J.N., Gerfen, C.R., Rogawski,M.A.,
Sibley, D.R., Skolnick, P., Wray, S. (Eds.), Current Protocols in Neuroscience.John Wiley & Sons, New York, NY, pp. 8.9.1–8.9.15.
McMartin, C., Bohacek, R.S., 1997. QXP: powerful, rapid computer algorithmsfor structure-based drug design. J. Comput.-Aided Mol. Des. 11, 333–344.
Millan, M.J., 2002. Descending control of pain. Prog. Neurobiol. 66, 355–474.Mizobe, T., Maghsoudi, K., Sitwala, K., Tianzhi, G., Ou, J., Maze, M., 1996.
Antisense technology reveals the alpha2A adrenoceptor to be the subtypemediating the hypnotic response to the highly selective agonist, dexmede-tomidine, in the locus coeruleus of the rat. J. Clin. Invest. 98, 1076–1080.
Molderings, G.J., 1997. Imidazoline receptors: basic knowledge, recentadvances and future prospects for therapy and diagnosis. Drugs Future 22,757–772.
Molderings, G.J., Gothert, M., 1999. Imidazoline binding sites and receptors incardiovascular tissue. Gen. Pharmacol. 32, 17–22.
Ogidigben, M.J., Potter, D.E., 2001. Central imidazoline I1 receptors modulateaqueous hydrodynamics. Curr. Eye Res. 22, 358–366.
Ozaita, A., Olmos, G., Boronat, M.A., Lizcano, J.M., Unzeta, M., Garcìa-Sevilla, J.A, 1997. Inhibition of monoamine oxidase A and B activities byimidazol(ine)/guanidine drugs, nature of the interaction and distinction fromI2-imidazoline receptors in rat liver. Br. J. Pharmacol. 121, 901–912.
Parini, A., Moudanos, C.G., Pizzinat, N., Lanier, S.M., 1996. The elusive familyof imidazoline binding sites. Trends Pharmacol. Sci. 17, 13–16.
Pigini, M., Bousquet, P., Carotti, A., Dontenwill, M., Giannella, M., Moriconi, R.,Piergentili, A., Quaglia, W., Tayebati, S.K., Brasili, L., 1997. Imidazolinereceptors: qualitative structure-activity relationships and discovery of tracizo-line and benazoline. Two ligands with high affinity and unprecedentedselectivity. Bioorg. Med. Chem. 5, 833–841.
Raddatz, R., Savic, S.L., Bakthavachalam, V., Lesnick, J., Jasper, J.R., McGrath,C.R., Parini, A., Lanier, S.M., 2000. Imidazoline-binding domains onmonoamine oxidase B and subpopulations of enzyme. J. Pharmacol. Exp.Ther. 292, 1135–1145.
Reynolds, G.P., Boulton, R.M., Pearson, S.J., Hudson, A.L., Nutt, D.J., 1996.Imidazoline binding sites in Huntington's and Parkinson’s disease putamen.Eur. J. Pharmacol. 301, R19–R21.
Ruggiero, D.A., Regunathan, S., Wang, H., Milner, T.A., Reis, D.J., 1998.Immunocytochemical localization of an imidazoline receptor protein in thecentral nervous system. Brain Res. 780, 270–293.
Ruiz-Durantez, E., Torrecilla,M., Pineda, J., Ugedo, L., 2003. Attenuation of acuteand chronic effects of morphine by the imidazoline receptor ligand 2-(2-benzofuranyl)-2-imidazoline in rat locus coeruleus neurons. Br. J. Pharmacol.138, 494–500.
Sanchez-Blazquez, P., Boronat, M.A., Olmos, G., Garcìa-Sevilla, J.A., Garzon,J., 2000. Activation of I2-imidazoline receptors enhances supraspinalmorphine analgesia in mice: a model to detect agonist and antagonistactivities at these receptors. Br. J. Pharmacol. 130, 146–152.
Santos, A.R.S., Gadotti, V.M., Oliveira, G.L., Tibola, D., Paszcuk, A.F., Neto,A., Spindola, H.M., Souza, M.M., Rodrigues, A.L.S., Calixto, J.B., 2005.Mechanisms involved in the antinociception caused by agmatine in mice.Neuropharmacology 48, 1021–1034.
Smyth, D.D., Penner, S.B., 1999. Peripheral and central imidazoline receptor-mediated natriuresis in the rat. Ann. N. Y. Acad. Sci. 881, 344–357.
Su, R.B., Li, J., Li, X., Qin, B.Y., 2001. Down-regulation of MAO-B activityand imidazoline receptors in rat brain following chronic treatment ofmorphine. Acta Pharmacol. Sin. 22, 639–644.
Tesson, F., Parini, A., 1991. Identification of an imidazoline-guanidiniumreceptive site in mitochondria from rabbit cerebral cortex. Eur. J. Pharmacol.208, 81–83.
Tesson, F., Limon, I., Parini, A., 1992. Tissue-specific localization ofmitochondrialimidazoline-guanidinium receptive site. Eur. J. Pharmacol. 219, 335–338.
Yesilyurt, O., Uzbay, I.T., 2001. Agmatine potentiates the analgesic effect ofmorphine by an α2-adrenoceptor-mediated mechanism in mice. Neuropsy-chopharmacology 25, 98–103.
Yoburn, B.C., Lutfy, K., Candido, J., 1991. Species differences in μ- and δ-opioid receptors. Eur. J. Pharmacol. 193, 105–108.





























