Embed Size (px)
Citation preview

Neurobiology of Learning and Memory 93 (2010) 117–126
Contents lists available at ScienceDirect
Neurobiology of Learning and Memory
journal homepage: www.elsevier .com/ locate/ynlme
Intracranial self-stimulation recovers learning and memory capacityin basolateral amygdala-damaged rats
Pilar Segura-Torres *, Laura Aldavert-Vera, Anna Gatell-Segura, Diego Redolar-Ripoll 1,Ignacio Morgado-BernalDepartament de Psicobiologia i de Metodologia de les Ciències de la Salut, Institut de Neurociències, Universitat Autònoma de Barcelona, 08193 Bellaterra, Barcelona, Spain
a r t i c l e i n f o a b s t r a c t
Article history:Received 15 April 2009Revised 3 September 2009Accepted 9 September 2009Available online 15 September 2009
Keywords:Rewarding brain stimulationIntracranial self-stimulationBasolateral amygdala lesionActive avoidanceMemory modulationMemory recovery
1074-7427/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.nlm.2009.09.001
* Corresponding author. Fax: +34 93 581 20 01.E-mail address: [email protected] (P. Segura-To
1 Universitat Oberta de Catalunya, Rambla de PobleSpain.
We studied the capacity of post-training intracranial self-stimulation (SS) to reverse or ameliorate learn-ing and memory impairments caused by amygdala damage in rats. A first experiment showed that lesionsof the basolateral amygdala (BLA) slow down acquisition of two-way active avoidance conditioning(2wAA). In a second experiment we observed that a post-training SS treatment administered immedi-ately after each 2wAA conditioning session is able to completely reverse the disruptive effects of theBLA lesions, and the facilitative effect lasts for 10 days. A third experiment allowed us to differentiatethe strong recuperative effects of the SS treatment from the slight effect caused by overtraining the sameconditioning response. We concluded that SS is able to counteract the behavioral deficit induced by BLAdamage, probably by activating alternative undamaged brain structures related to learning and memory,such as the hippocampus.
� 2009 Elsevier Inc. All rights reserved.
1. Introduction
Post-training intracranial self-stimulation (SS) facilitates theacquisition and retention of several tasks related to both implicitand explicit memory systems in rats (Coulombe & White, 1980,1982; Huston & Mueller, 1978; Huston, Mueller, & Mondadori,1977; Redolar-Ripoll, Aldavert-Vera, Soriano-Mas, Segura-Torres,& Morgado-Bernal, 2002; Ruiz-Medina, Morgado-Bernal, Redolar-Ripoll, Aldavert-Vera, & Segura-Torres, 2008a; Segura-Torres,Capdevila-Ortis, Marti-Nicolovius, & Morgado-Bernal, 1988;Soriano-Mas, Redolar-Ripoll, Aldavert-Vera, Morgado-Bernal, &Segura-Torres, 2005). Beside being especially effective in subjectswith a naturally low learning capacity (Aldavert-Vera, Segura-Tor-res, Costa-Miserachs, & Morgado-Bernal, 1996) or a cognitive deficitcaused by aging (Aldavert-Vera et al., 1997; Redolar-Ripoll et al.,2003), experiments from our laboratory have shown that SS is alsoable to ameliorate two-way active avoidance (2wAA) conditioningdeficits caused by extensive bilateral lesions of the parafascicularnucleus of the thalamus (PF) (Redolar-Ripoll et al., 2003). In thepresent study, we aim to investigate if SS can be also able to func-tionally compensate the impairment caused by the damage of other
ll rights reserved.
rres).Nou, 156, Barcelona 08018,
brain structures belonging to neuroanatomic systems morecritically involved in 2wAA conditioning, such as the amygdala.
The amygdala has been identified as a central structure in theemotional memory system. In general terms, it is well establishedthat amygdala damage in humans and animals may impair emo-tional learning and memory (Bechara, 2005; Cahill, 2000; LeDoux,2000, 2003; Rodrigues, Schafe, & LeDoux, 2004). There is also evi-dence that post-training treatments stimulating amygdala func-tioning can positively influence learning and retention inappetitively or aversively motivated tasks in rats (Lalumiere &McGaugh, 2005). The basolateral area of the amygdala (lateral, ba-sal, and accessory basal nuclei) (Pitkanen, Savander, & LeDoux,1997), is especially important for associative learning related toemotional stimuli (Dwyer & Killcross, 2006; Sah & Westbrook,2008), and for 2wAA conditioning in particular (Savonenko, Werka,Nikolaev, Zielinski, & Kaczmarek, 2003). With reference to theacquisition of 2wAA, it has been proposed that the reactive re-sponse that responds to stimuli that predict danger by elicitinghard-wired defense responses (the classical component of 2wAA)would be mediated by direct pathways from the lateral nucleus(LA) to the central (Ce) nucleus of the amygdala, and that the abil-ity of a conditioned stimulus to reinforce the acquisition of new or‘‘active” responses (instrumental component of 2wAA) would bemediated by projections from the LA to the BLA (basolateral nu-cleus composed by the basal and basal accessory nuclei) (Amora-panth, LeDoux, & Nader, 2000; Savonenko, Filipkowski, Werka,Zielinski, & Kaczmarek, 1999).

118 P. Segura-Torres et al. / Neurobiology of Learning and Memory 93 (2010) 117–126
Another major function of the amygdala is the enhancement ofmemory consolidation for emotionally arousing experiences(McGaugh, 2004). Considerable evidence suggests that the basolat-eral complex of the amygdala (LA and BLA nuclei) is a key structurein a memory-modulatory system that regulates, in concert withother brain regions, stress and glucocorticoid effects on differentmemory functions (Nathan, Griffith, McReynolds, Hahn, & Roo-zendaal, 2004; Roozendaal, Okuda, Van der Zee, & McGaugh, 2006).
In this context, we have carried out three experiments. The firstevaluates the degree of impairment of 2wAA conditioning that canbe caused by BLA lesions. A second experiment determineswhether the same kind of post-training SS treatment that in ourprevious research has been able to reverse 2wAA impairments inrats with PF lesions, is also able to reverse the observed impair-ments in rats with BLA damage. To find out more about the natureof the observed effects, a third experiment compared the facilita-tive effect of the SS treatment with the improved performanceproduced by supplementary 2wAA training in rats with BLAlesions.
2. Experiment 1: effects of basolateral amygdala (BLA) lesionson 2wAA conditioning
This experiment was intended to test the effects of bilateralelectrolytic lesion of BLA on subsequent 2wAA distributed condi-tioning. Rats were trained daily until they reached an establishedlearning criterion.
2.1. Material and methods
2.1.1. SubjectsWe used 32 naive male Wistar rats, obtained from our labora-
tory breeding stock, with a mean age of 92.19 days (SD = 4.20) atthe beginning of the experiment, and mean weight of 446.47 g(SD = 37.34) at the time of surgery. All rats were housed singly, al-ways kept under conditions of controlled temperature (20–24 �C)and humidity (40–70%), and subjected to an artificial light/dark-ness cycle of 12/12 h (lights on at 08:00 h). Food and water wereavailable ad libitum. The rats were tested during the first half ofthe light cycle. The experiments were carried out in compliancewith the European Community Council Directive for care and useof laboratory animals (CEE 86/609) and the Generalitat de Catalu-nya Decret (DOGC 2073 10/7/ 1995, DARP protocol number 2181).
2.1.2. Stereotaxic surgeryBefore surgery rats were randomly distributed into three exper-
imental groups: BLA-lesion (rats that would receive a bilateral le-sion in the BLA nucleus, n = 16), BLA-sham (rats that wouldreceive a bilateral sham implantation without lesion in the BLA nu-cleus, n = 8), and Control (rats that would receive a sham stereo-taxic surgery without lesion or sham implantation, n = 8).Stereotaxic surgery (Model 1504, David Kopf Instruments) wasperformed under general anesthesia (150 mg/kg Ketolar� Ketaminechlorhydrate and 0.08 mg/kg Rompun� Xylazin; i.p.). Rats in theBLA-lesion group were submitted to bilateral electrolytic lesionsusing a direct current of 2 mA (Cibertec GL-2 electrical stimulator)for 9–15 s depending on the antero-posterior coordinate, with abipolar insulated stainless steel electrode (250 lm in diameter).Although electrolytic lesions are considered as non-specific, theycan be restricted to simple regions with minimal infringementonto neighboring structures (Nader, Majidishad, Amorapanth, &LeDoux, 2001). Moreover, it has been shown that this kind of le-sions is more effective than other kind of lesions, such as ibotenicacid, to affect conditioning when applied to the amygdala (Lanuza,Nader, & Ledoux, 2004). The incisor bar was set at �2.7 mm below
the interaural line, and the following stereotaxic coordinates wereused: AP = �2.12, �2.8, �3.3, �4.16 mm from bregma; L = ±4.9,±4.9, ±5.1, ±5.3; P = �9.0, �9.2, �9.0, and 9.0 with the cranium sur-face as dorsal reference (Paxinos & Watson, 1998).
2.1.3. ProcedureOnce the rats had recovered from surgery (15 days), they were
trained in 2wAA conditioning conducted in an automated shuttle-box (50 cm long � 24 cm wide � 25 cm high, Letica Li-916, PAN-LAB, Barcelona, Spain) enclosed in a sound-attenuating box venti-lated by an extractor fan. The conditioning box was illuminatedby a fluorescent bulb located on the sound-attenuating box. Thetwo compartment floors (without any physical separation betweenthem) were independently electrifiable and constructed of stain-less steel bars (3.9 mm in diameter, 8.8 mm apart) that formed ashock grid. The conditioned stimulus was a 60-dB, 1-kHz tone of3 s duration. The grid delivered a scrambled footshock uncondi-tioned stimulus (0.5 mA intensity, maximum duration of 30 s) pro-vided by a shock generator. The current supplied by the shockerwas a positive semiwave of 100 Hz. The shuttle-box was connectedto a computer that controlled the training schedule of 1 min(±10 s). Rats received one daily 10-trial 2wAA session until theyobtained 80% or more correct responses in two consecutive ses-sions. Rats that did not achieve the established learning criterionwere allowed to perform a maximum of 15 sessions (distributedin 3 blocks of 5 sessions separated by 2 days of rest). Just beforethe first acquisition session, the rats were allowed to ambulatefreely in the shuttle-box for 10 min to become familiarised withthe learning environment. During the learning training, animalsavoided the shock by crossing to the adjacent compartment whenthe conditioned stimulus was on. Apart from the number of avoid-ance responses made in each training session (considered as the le-vel of performance), inter-trial crossings and crossings during thefree ambulation period were also scored and considered measuresof locomotor activity.
2.1.4. HistologyAt the end of the experiment, histological analyses were per-
formed to verify the extent and location of the lesions. The animalswere given an overdose of sodium pentobarbital (150 mg/kg, i.p.)and transcardially perfused with 0.9% physiological saline followedby 10% formalin (water and 37–40% formaldehyde). The animals’brains were removed and placed in a 30% sucrose solution beforebeing cut into 40 lm sections on a freezing stage microtome (Cryo-cut 1800 with microtome 2020, JUNG). The tissue sections werestained following the acetylcholinesterase method and examinedunder a microscope (Olympus BX41, Olympus Optical CO., LTD Ja-pan) connected to a photographic camera (Olympus DP70). Theywere examined separately by three people who were not awareof either the group or the performance of the subjects. Lesionswere assessed neuroanatomically by examining sections for areasof neuronal loss. The damaged areas were represented by drawingthem onto standardised sections of the brain from the atlas of Pax-inos and Watson (1998).
2.1.5. Data analysesThe statistical computer package program SPSS 14.0 was used
to process the data. The main analyses were carried out consider-ing the independent variable as qualitative (three experimentalgroups), and the dependent variables as quantitative (performancein 2wAA conditioning sessions and locomotor activity). Multivari-ate analyses of variance (MANOVA) were performed with their cor-responding contrast analyses. Survival analyses were also carriedout to analyse and compare the mean number of sessions requiredby each experimental group to reach the established learningcriterion (80% avoidance responses during two consecutive 2wAA
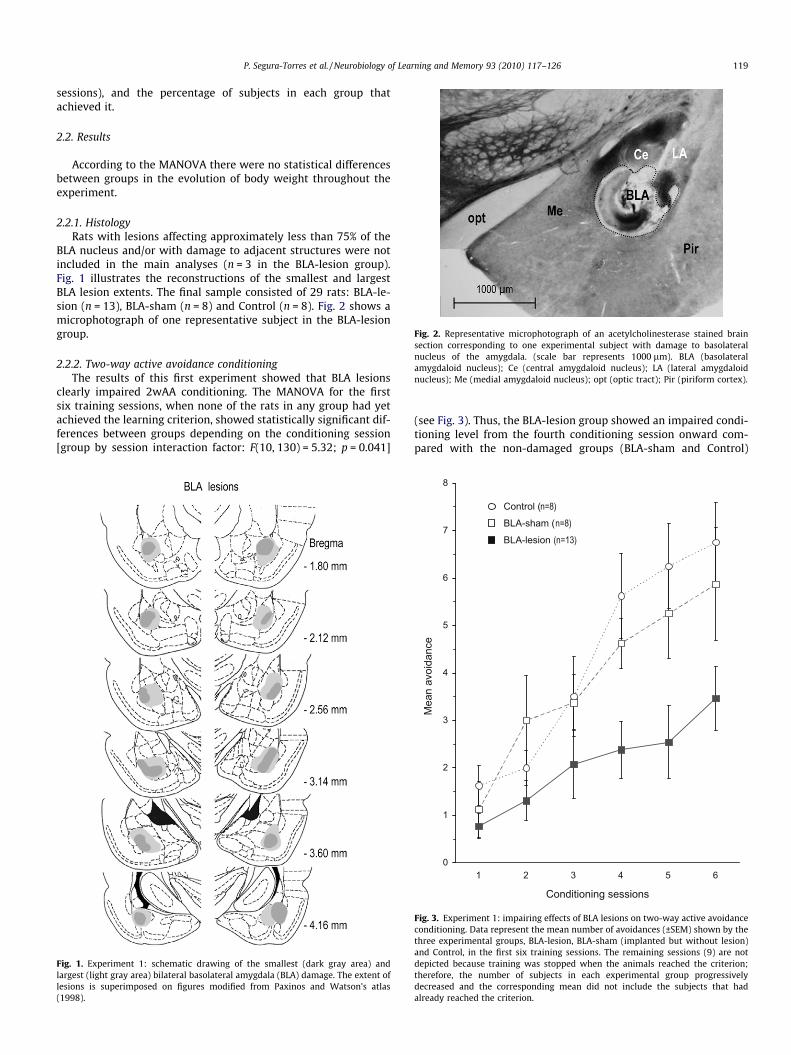
Fig. 2. Representative microphotograph of an acetylcholinesterase stained brainsection corresponding to one experimental subject with damage to basolateralnucleus of the amygdala. (scale bar represents 1000 lm). BLA (basolateralamygdaloid nucleus); Ce (central amygdaloid nucleus); LA (lateral amygdaloidnucleus); Me (medial amygdaloid nucleus); opt (optic tract); Pir (piriform cortex).
P. Segura-Torres et al. / Neurobiology of Learning and Memory 93 (2010) 117–126 119
sessions), and the percentage of subjects in each group thatachieved it.
2.2. Results
According to the MANOVA there were no statistical differencesbetween groups in the evolution of body weight throughout theexperiment.
2.2.1. HistologyRats with lesions affecting approximately less than 75% of the
BLA nucleus and/or with damage to adjacent structures were notincluded in the main analyses (n = 3 in the BLA-lesion group).Fig. 1 illustrates the reconstructions of the smallest and largestBLA lesion extents. The final sample consisted of 29 rats: BLA-le-sion (n = 13), BLA-sham (n = 8) and Control (n = 8). Fig. 2 shows amicrophotograph of one representative subject in the BLA-lesiongroup.
2.2.2. Two-way active avoidance conditioningThe results of this first experiment showed that BLA lesions
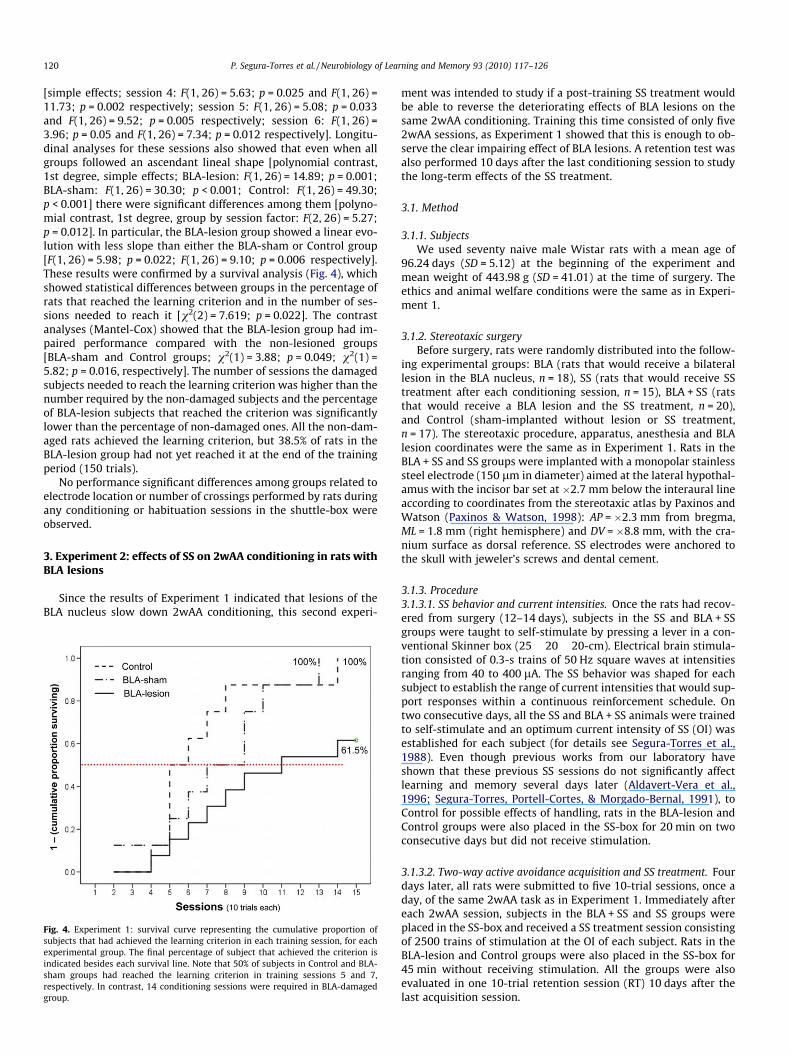
clearly impaired 2wAA conditioning. The MANOVA for the firstsix training sessions, when none of the rats in any group had yetachieved the learning criterion, showed statistically significant dif-ferences between groups depending on the conditioning session[group by session interaction factor: F(10, 130) = 5.32; p = 0.041]
Fig. 1. Experiment 1: schematic drawing of the smallest (dark gray area) andlargest (light gray area) bilateral basolateral amygdala (BLA) damage. The extent oflesions is superimposed on figures modified from Paxinos and Watson’s atlas(1998).
(see Fig. 3). Thus, the BLA-lesion group showed an impaired condi-tioning level from the fourth conditioning session onward com-pared with the non-damaged groups (BLA-sham and Control)
0
1
2
3
4
5
6
7
8
1 2 3 4 5 6
Conditioning sessions
Mea
nav
oida
nce
Control (n=8)
BLA-lesion (n=13)BLA-sham (n=8)
Fig. 3. Experiment 1: impairing effects of BLA lesions on two-way active avoidanceconditioning. Data represent the mean number of avoidances (±SEM) shown by thethree experimental groups, BLA-lesion, BLA-sham (implanted but without lesion)and Control, in the first six training sessions. The remaining sessions (9) are notdepicted because training was stopped when the animals reached the criterion;therefore, the number of subjects in each experimental group progressivelydecreased and the corresponding mean did not include the subjects that hadalready reached the criterion.
120 P. Segura-Torres et al. / Neurobiology of Learning and Memory 93 (2010) 117–126
[simple effects; session 4: F(1, 26) = 5.63; p = 0.025 and F(1, 26) =11.73; p = 0.002 respectively; session 5: F(1, 26) = 5.08; p = 0.033and F(1, 26) = 9.52; p = 0.005 respectively; session 6: F(1, 26) =3.96; p = 0.05 and F(1, 26) = 7.34; p = 0.012 respectively]. Longitu-dinal analyses for these sessions also showed that even when allgroups followed an ascendant lineal shape [polynomial contrast,1st degree, simple effects; BLA-lesion: F(1, 26) = 14.89; p = 0.001;BLA-sham: F(1, 26) = 30.30; p < 0.001; Control: F(1, 26) = 49.30;p < 0.001] there were significant differences among them [polyno-mial contrast, 1st degree, group by session factor: F(2, 26) = 5.27;p = 0.012]. In particular, the BLA-lesion group showed a linear evo-lution with less slope than either the BLA-sham or Control group[F(1, 26) = 5.98; p = 0.022; F(1, 26) = 9.10; p = 0.006 respectively].These results were confirmed by a survival analysis (Fig. 4), whichshowed statistical differences between groups in the percentage ofrats that reached the learning criterion and in the number of ses-sions needed to reach it [v2(2) = 7.619; p = 0.022]. The contrastanalyses (Mantel-Cox) showed that the BLA-lesion group had im-paired performance compared with the non-lesioned groups[BLA-sham and Control groups; v2(1) = 3.88; p = 0.049; v2(1) =5.82; p = 0.016, respectively]. The number of sessions the damagedsubjects needed to reach the learning criterion was higher than thenumber required by the non-damaged subjects and the percentageof BLA-lesion subjects that reached the criterion was significantlylower than the percentage of non-damaged ones. All the non-dam-aged rats achieved the learning criterion, but 38.5% of rats in theBLA-lesion group had not yet reached it at the end of the trainingperiod (150 trials).
No performance significant differences among groups related toelectrode location or number of crossings performed by rats duringany conditioning or habituation sessions in the shuttle-box wereobserved.
3. Experiment 2: effects of SS on 2wAA conditioning in rats withBLA lesions
Since the results of Experiment 1 indicated that lesions of theBLA nucleus slow down 2wAA conditioning, this second experi-
Fig. 4. Experiment 1: survival curve representing the cumulative proportion ofsubjects that had achieved the learning criterion in each training session, for eachexperimental group. The final percentage of subject that achieved the criterion isindicated besides each survival line. Note that 50% of subjects in Control and BLA-sham groups had reached the learning criterion in training sessions 5 and 7,respectively. In contrast, 14 conditioning sessions were required in BLA-damagedgroup.
ment was intended to study if a post-training SS treatment wouldbe able to reverse the deteriorating effects of BLA lesions on thesame 2wAA conditioning. Training this time consisted of only five2wAA sessions, as Experiment 1 showed that this is enough to ob-serve the clear impairing effect of BLA lesions. A retention test wasalso performed 10 days after the last conditioning session to studythe long-term effects of the SS treatment.
3.1. Method
3.1.1. SubjectsWe used seventy naive male Wistar rats with a mean age of
96.24 days (SD = 5.12) at the beginning of the experiment andmean weight of 443.98 g (SD = 41.01) at the time of surgery. Theethics and animal welfare conditions were the same as in Experi-ment 1.
3.1.2. Stereotaxic surgeryBefore surgery, rats were randomly distributed into the follow-
ing experimental groups: BLA (rats that would receive a bilaterallesion in the BLA nucleus, n = 18), SS (rats that would receive SStreatment after each conditioning session, n = 15), BLA + SS (ratsthat would receive a BLA lesion and the SS treatment, n = 20),and Control (sham-implanted without lesion or SS treatment,n = 17). The stereotaxic procedure, apparatus, anesthesia and BLAlesion coordinates were the same as in Experiment 1. Rats in theBLA + SS and SS groups were implanted with a monopolar stainlesssteel electrode (150 lm in diameter) aimed at the lateral hypothal-amus with the incisor bar set at �2.7 mm below the interaural lineaccording to coordinates from the stereotaxic atlas by Paxinos andWatson (Paxinos & Watson, 1998): AP = �2.3 mm from bregma,ML = 1.8 mm (right hemisphere) and DV = �8.8 mm, with the cra-nium surface as dorsal reference. SS electrodes were anchored tothe skull with jeweler’s screws and dental cement.
3.1.3. Procedure3.1.3.1. SS behavior and current intensities. Once the rats had recov-ered from surgery (12–14 days), subjects in the SS and BLA + SSgroups were taught to self-stimulate by pressing a lever in a con-ventional Skinner box (25 � 20 � 20-cm). Electrical brain stimula-tion consisted of 0.3-s trains of 50 Hz square waves at intensitiesranging from 40 to 400 lA. The SS behavior was shaped for eachsubject to establish the range of current intensities that would sup-port responses within a continuous reinforcement schedule. Ontwo consecutive days, all the SS and BLA + SS animals were trainedto self-stimulate and an optimum current intensity of SS (OI) wasestablished for each subject (for details see Segura-Torres et al.,1988). Even though previous works from our laboratory haveshown that these previous SS sessions do not significantly affectlearning and memory several days later (Aldavert-Vera et al.,1996; Segura-Torres, Portell-Cortes, & Morgado-Bernal, 1991), toControl for possible effects of handling, rats in the BLA-lesion andControl groups were also placed in the SS-box for 20 min on twoconsecutive days but did not receive stimulation.
3.1.3.2. Two-way active avoidance acquisition and SS treatment. Fourdays later, all rats were submitted to five 10-trial sessions, once aday, of the same 2wAA task as in Experiment 1. Immediately aftereach 2wAA session, subjects in the BLA + SS and SS groups wereplaced in the SS-box and received a SS treatment session consistingof 2500 trains of stimulation at the OI of each subject. Rats in theBLA-lesion and Control groups were also placed in the SS-box for45 min without receiving stimulation. All the groups were alsoevaluated in one 10-trial retention session (RT) 10 days after thelast acquisition session.
P. Segura-Torres et al. / Neurobiology of Learning and Memory 93 (2010) 117–126 121
3.1.4. Histology and data analysesAs in Experiment 1, histological analyses were performed at the
end of the experiment to verify the extent and location of the BLAlesions. To determine the location of the SS electrode tip, tissuesections from rats in the SS and BLA + SS groups were stained withcresyl violet. Data were analysed as in Experiment 1, with a learn-ing criterion of 50% avoidance responses for the survival analysis.
3.2. Results
The MANOVA did not show any statistical differences betweengroups in the evolution of body weight during the experiment.
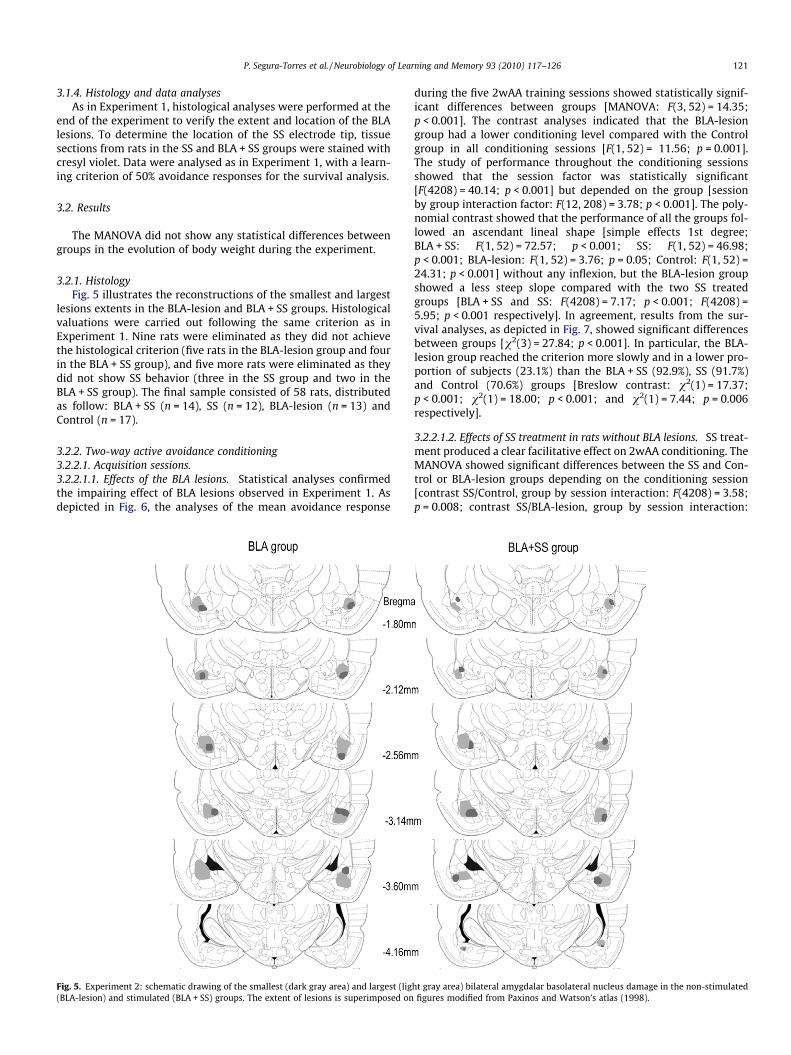
3.2.1. HistologyFig. 5 illustrates the reconstructions of the smallest and largest
lesions extents in the BLA-lesion and BLA + SS groups. Histologicalvaluations were carried out following the same criterion as inExperiment 1. Nine rats were eliminated as they did not achievethe histological criterion (five rats in the BLA-lesion group and fourin the BLA + SS group), and five more rats were eliminated as theydid not show SS behavior (three in the SS group and two in theBLA + SS group). The final sample consisted of 58 rats, distributedas follow: BLA + SS (n = 14), SS (n = 12), BLA-lesion (n = 13) andControl (n = 17).
3.2.2. Two-way active avoidance conditioning3.2.2.1. Acquisition sessions.3.2.2.1.1. Effects of the BLA lesions. Statistical analyses confirmedthe impairing effect of BLA lesions observed in Experiment 1. Asdepicted in Fig. 6, the analyses of the mean avoidance response
Fig. 5. Experiment 2: schematic drawing of the smallest (dark gray area) and largest (lig(BLA-lesion) and stimulated (BLA + SS) groups. The extent of lesions is superimposed on
during the five 2wAA training sessions showed statistically signif-icant differences between groups [MANOVA: F(3, 52) = 14.35;p < 0.001]. The contrast analyses indicated that the BLA-lesiongroup had a lower conditioning level compared with the Controlgroup in all conditioning sessions [F(1, 52) = 11.56; p = 0.001].The study of performance throughout the conditioning sessionsshowed that the session factor was statistically significant[F(4208) = 40.14; p < 0.001] but depended on the group [sessionby group interaction factor: F(12, 208) = 3.78; p < 0.001]. The poly-nomial contrast showed that the performance of all the groups fol-lowed an ascendant lineal shape [simple effects 1st degree;BLA + SS: F(1, 52) = 72.57; p < 0.001; SS: F(1, 52) = 46.98;p < 0.001; BLA-lesion: F(1, 52) = 3.76; p = 0.05; Control: F(1, 52) =24.31; p < 0.001] without any inflexion, but the BLA-lesion groupshowed a less steep slope compared with the two SS treatedgroups [BLA + SS and SS: F(4208) = 7.17; p < 0.001; F(4208) =5.95; p < 0.001 respectively]. In agreement, results from the sur-vival analyses, as depicted in Fig. 7, showed significant differencesbetween groups [v2(3) = 27.84; p < 0.001]. In particular, the BLA-lesion group reached the criterion more slowly and in a lower pro-portion of subjects (23.1%) than the BLA + SS (92.9%), SS (91.7%)and Control (70.6%) groups [Breslow contrast: v2(1) = 17.37;p < 0.001; v2(1) = 18.00; p < 0.001; and v2(1) = 7.44; p = 0.006respectively].
3.2.2.1.2. Effects of SS treatment in rats without BLA lesions. SS treat-ment produced a clear facilitative effect on 2wAA conditioning. TheMANOVA showed significant differences between the SS and Con-trol or BLA-lesion groups depending on the conditioning session[contrast SS/Control, group by session interaction: F(4208) = 3.58;p = 0.008; contrast SS/BLA-lesion, group by session interaction:
ht gray area) bilateral amygdalar basolateral nucleus damage in the non-stimulatedfigures modified from Paxinos and Watson’s atlas (1998).
0
1
2
3
4
5
6
7
8
9
1 2 3 4 5
Conditioning sessions
Mea
nav
oida
nce
Retention(10 days)
Control (n=17)BLA-lesion (n=13)
SS (n=12)
BLA+SS (n=14)
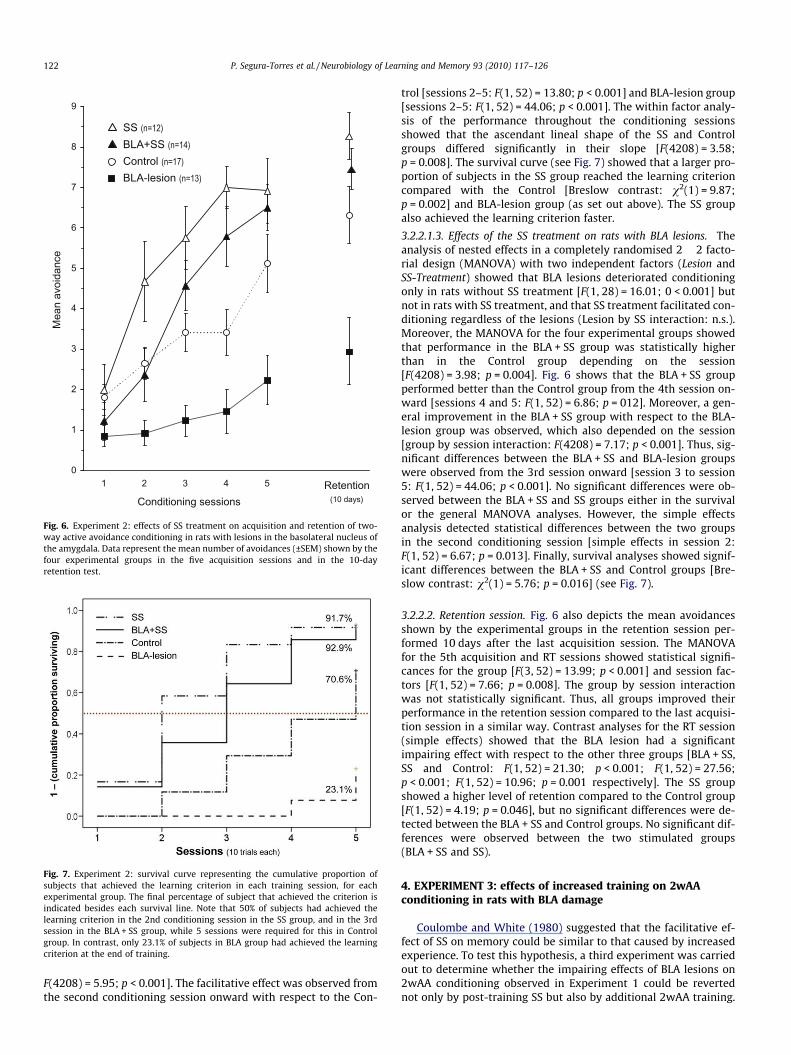
Fig. 6. Experiment 2: effects of SS treatment on acquisition and retention of two-way active avoidance conditioning in rats with lesions in the basolateral nucleus ofthe amygdala. Data represent the mean number of avoidances (±SEM) shown by thefour experimental groups in the five acquisition sessions and in the 10-dayretention test.
Fig. 7. Experiment 2: survival curve representing the cumulative proportion ofsubjects that achieved the learning criterion in each training session, for eachexperimental group. The final percentage of subject that achieved the criterion isindicated besides each survival line. Note that 50% of subjects had achieved thelearning criterion in the 2nd conditioning session in the SS group, and in the 3rdsession in the BLA + SS group, while 5 sessions were required for this in Controlgroup. In contrast, only 23.1% of subjects in BLA group had achieved the learningcriterion at the end of training.
122 P. Segura-Torres et al. / Neurobiology of Learning and Memory 93 (2010) 117–126
F(4208) = 5.95; p < 0.001]. The facilitative effect was observed fromthe second conditioning session onward with respect to the Con-
trol [sessions 2–5: F(1, 52) = 13.80; p < 0.001] and BLA-lesion group[sessions 2–5: F(1, 52) = 44.06; p < 0.001]. The within factor analy-sis of the performance throughout the conditioning sessionsshowed that the ascendant lineal shape of the SS and Controlgroups differed significantly in their slope [F(4208) = 3.58;p = 0.008]. The survival curve (see Fig. 7) showed that a larger pro-portion of subjects in the SS group reached the learning criterioncompared with the Control [Breslow contrast: v2(1) = 9.87;p = 0.002] and BLA-lesion group (as set out above). The SS groupalso achieved the learning criterion faster.
3.2.2.1.3. Effects of the SS treatment on rats with BLA lesions. Theanalysis of nested effects in a completely randomised 2 � 2 facto-rial design (MANOVA) with two independent factors (Lesion andSS-Treatment) showed that BLA lesions deteriorated conditioningonly in rats without SS treatment [F(1, 28) = 16.01; 0 < 0.001] butnot in rats with SS treatment, and that SS treatment facilitated con-ditioning regardless of the lesions (Lesion by SS interaction: n.s.).Moreover, the MANOVA for the four experimental groups showedthat performance in the BLA + SS group was statistically higherthan in the Control group depending on the session[F(4208) = 3.98; p = 0.004]. Fig. 6 shows that the BLA + SS groupperformed better than the Control group from the 4th session on-ward [sessions 4 and 5: F(1, 52) = 6.86; p = 012]. Moreover, a gen-eral improvement in the BLA + SS group with respect to the BLA-lesion group was observed, which also depended on the session[group by session interaction: F(4208) = 7.17; p < 0.001]. Thus, sig-nificant differences between the BLA + SS and BLA-lesion groupswere observed from the 3rd session onward [session 3 to session5: F(1, 52) = 44.06; p < 0.001]. No significant differences were ob-served between the BLA + SS and SS groups either in the survivalor the general MANOVA analyses. However, the simple effectsanalysis detected statistical differences between the two groupsin the second conditioning session [simple effects in session 2:F(1, 52) = 6.67; p = 0.013]. Finally, survival analyses showed signif-icant differences between the BLA + SS and Control groups [Bre-slow contrast: v2(1) = 5.76; p = 0.016] (see Fig. 7).
3.2.2.2. Retention session. Fig. 6 also depicts the mean avoidancesshown by the experimental groups in the retention session per-formed 10 days after the last acquisition session. The MANOVAfor the 5th acquisition and RT sessions showed statistical signifi-cances for the group [F(3, 52) = 13.99; p < 0.001] and session fac-tors [F(1, 52) = 7.66; p = 0.008]. The group by session interactionwas not statistically significant. Thus, all groups improved theirperformance in the retention session compared to the last acquisi-tion session in a similar way. Contrast analyses for the RT session(simple effects) showed that the BLA lesion had a significantimpairing effect with respect to the other three groups [BLA + SS,SS and Control: F(1, 52) = 21.30; p < 0.001; F(1, 52) = 27.56;p < 0.001; F(1, 52) = 10.96; p = 0.001 respectively]. The SS groupshowed a higher level of retention compared to the Control group[F(1, 52) = 4.19; p = 0.046], but no significant differences were de-tected between the BLA + SS and Control groups. No significant dif-ferences were observed between the two stimulated groups(BLA + SS and SS).
4. EXPERIMENT 3: effects of increased training on 2wAAconditioning in rats with BLA damage
Coulombe and White (1980) suggested that the facilitative ef-fect of SS on memory could be similar to that caused by increasedexperience. To test this hypothesis, a third experiment was carriedout to determine whether the impairing effects of BLA lesions on2wAA conditioning observed in Experiment 1 could be revertednot only by post-training SS but also by additional 2wAA training.
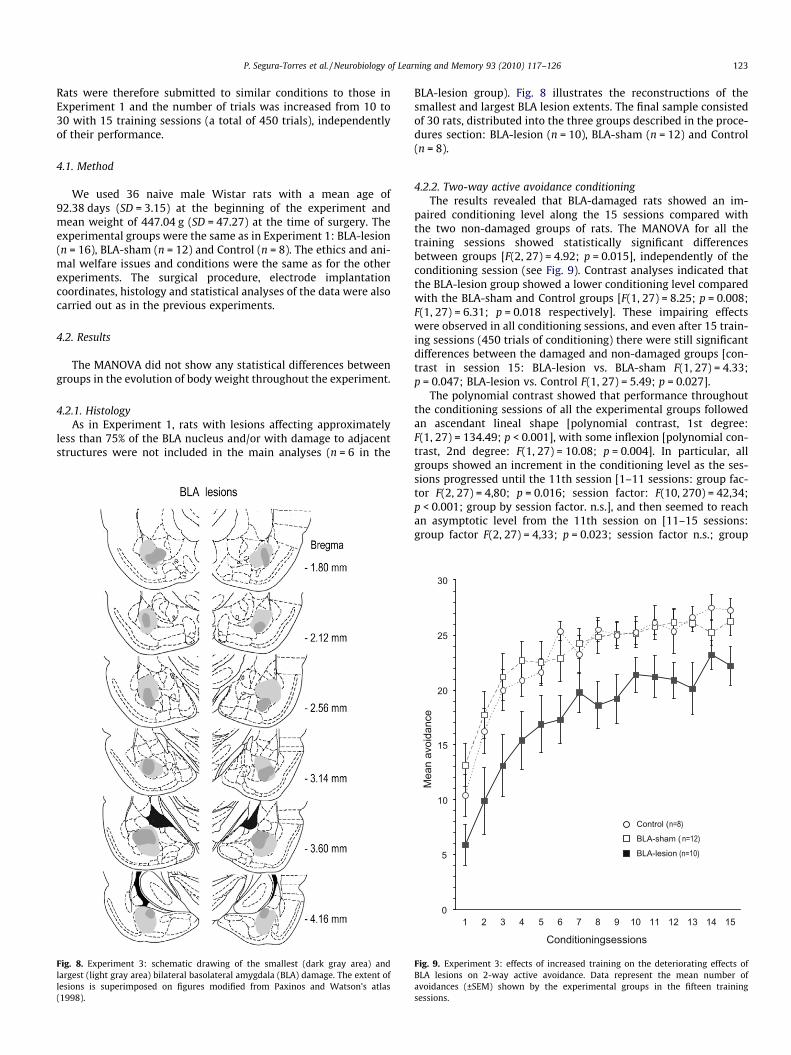
P. Segura-Torres et al. / Neurobiology of Learning and Memory 93 (2010) 117–126 123
Rats were therefore submitted to similar conditions to those inExperiment 1 and the number of trials was increased from 10 to30 with 15 training sessions (a total of 450 trials), independentlyof their performance.
4.1. Method
We used 36 naive male Wistar rats with a mean age of92.38 days (SD = 3.15) at the beginning of the experiment andmean weight of 447.04 g (SD = 47.27) at the time of surgery. Theexperimental groups were the same as in Experiment 1: BLA-lesion(n = 16), BLA-sham (n = 12) and Control (n = 8). The ethics and ani-mal welfare issues and conditions were the same as for the otherexperiments. The surgical procedure, electrode implantationcoordinates, histology and statistical analyses of the data were alsocarried out as in the previous experiments.
4.2. Results
The MANOVA did not show any statistical differences betweengroups in the evolution of body weight throughout the experiment.
4.2.1. HistologyAs in Experiment 1, rats with lesions affecting approximately
less than 75% of the BLA nucleus and/or with damage to adjacentstructures were not included in the main analyses (n = 6 in the
Fig. 8. Experiment 3: schematic drawing of the smallest (dark gray area) andlargest (light gray area) bilateral basolateral amygdala (BLA) damage. The extent oflesions is superimposed on figures modified from Paxinos and Watson’s atlas(1998).
BLA-lesion group). Fig. 8 illustrates the reconstructions of thesmallest and largest BLA lesion extents. The final sample consistedof 30 rats, distributed into the three groups described in the proce-dures section: BLA-lesion (n = 10), BLA-sham (n = 12) and Control(n = 8).
4.2.2. Two-way active avoidance conditioningThe results revealed that BLA-damaged rats showed an im-
paired conditioning level along the 15 sessions compared withthe two non-damaged groups of rats. The MANOVA for all thetraining sessions showed statistically significant differencesbetween groups [F(2, 27) = 4.92; p = 0.015], independently of theconditioning session (see Fig. 9). Contrast analyses indicated thatthe BLA-lesion group showed a lower conditioning level comparedwith the BLA-sham and Control groups [F(1, 27) = 8.25; p = 0.008;F(1, 27) = 6.31; p = 0.018 respectively]. These impairing effectswere observed in all conditioning sessions, and even after 15 train-ing sessions (450 trials of conditioning) there were still significantdifferences between the damaged and non-damaged groups [con-trast in session 15: BLA-lesion vs. BLA-sham F(1, 27) = 4.33;p = 0.047; BLA-lesion vs. Control F(1, 27) = 5.49; p = 0.027].
The polynomial contrast showed that performance throughoutthe conditioning sessions of all the experimental groups followedan ascendant lineal shape [polynomial contrast, 1st degree:F(1, 27) = 134.49; p < 0.001], with some inflexion [polynomial con-trast, 2nd degree: F(1, 27) = 10.08; p = 0.004]. In particular, allgroups showed an increment in the conditioning level as the ses-sions progressed until the 11th session [1–11 sessions: group fac-tor F(2, 27) = 4,80; p = 0.016; session factor: F(10, 270) = 42,34;p < 0.001; group by session factor. n.s.], and then seemed to reachan asymptotic level from the 11th session on [11–15 sessions:group factor F(2, 27) = 4,33; p = 0.023; session factor n.s.; group
Conditioningsessions
Mea
n av
oida
nce
0
5
10
15
20
25
30
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Control (n=8)
BLA-lesion (n=10)BLA-sham ( n=12)
Fig. 9. Experiment 3: effects of increased training on the deteriorating effects ofBLA lesions on 2-way active avoidance. Data represent the mean number ofavoidances (±SEM) shown by the experimental groups in the fifteen trainingsessions.
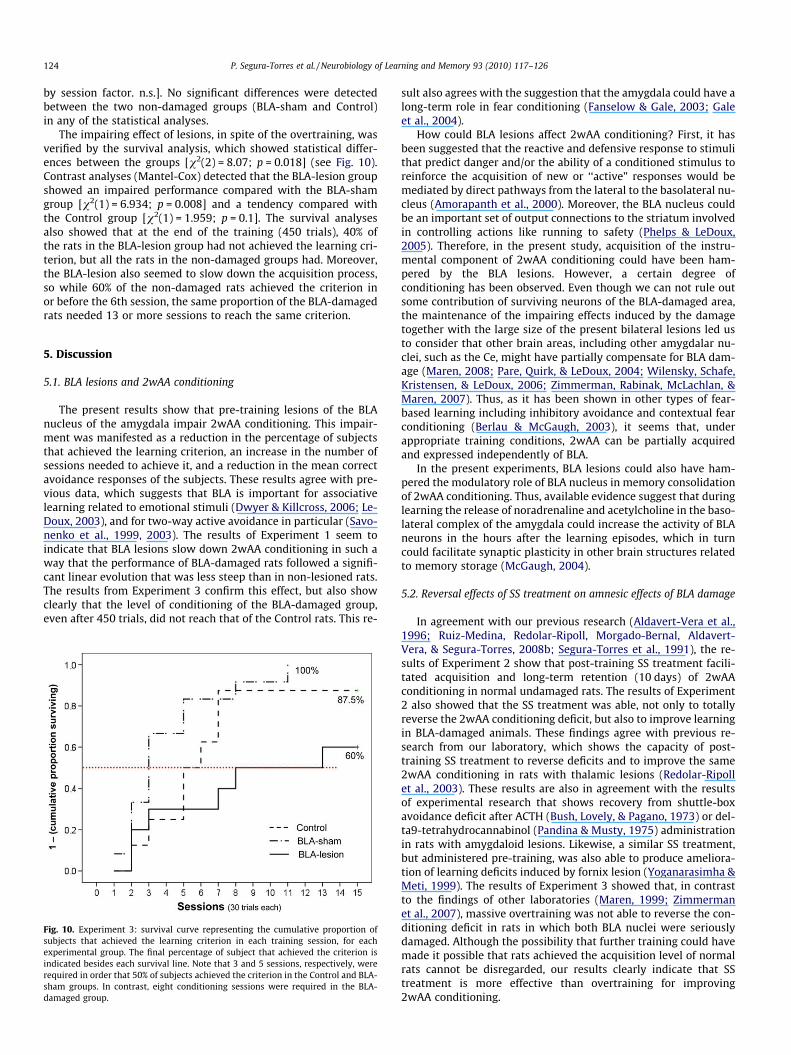
124 P. Segura-Torres et al. / Neurobiology of Learning and Memory 93 (2010) 117–126
by session factor. n.s.]. No significant differences were detectedbetween the two non-damaged groups (BLA-sham and Control)in any of the statistical analyses.
The impairing effect of lesions, in spite of the overtraining, wasverified by the survival analysis, which showed statistical differ-ences between the groups [v2(2) = 8.07; p = 0.018] (see Fig. 10).Contrast analyses (Mantel-Cox) detected that the BLA-lesion groupshowed an impaired performance compared with the BLA-shamgroup [v2(1) = 6.934; p = 0.008] and a tendency compared withthe Control group [v2(1) = 1.959; p = 0.1]. The survival analysesalso showed that at the end of the training (450 trials), 40% ofthe rats in the BLA-lesion group had not achieved the learning cri-terion, but all the rats in the non-damaged groups had. Moreover,the BLA-lesion also seemed to slow down the acquisition process,so while 60% of the non-damaged rats achieved the criterion inor before the 6th session, the same proportion of the BLA-damagedrats needed 13 or more sessions to reach the same criterion.
5. Discussion
5.1. BLA lesions and 2wAA conditioning
The present results show that pre-training lesions of the BLAnucleus of the amygdala impair 2wAA conditioning. This impair-ment was manifested as a reduction in the percentage of subjectsthat achieved the learning criterion, an increase in the number ofsessions needed to achieve it, and a reduction in the mean correctavoidance responses of the subjects. These results agree with pre-vious data, which suggests that BLA is important for associativelearning related to emotional stimuli (Dwyer & Killcross, 2006; Le-Doux, 2003), and for two-way active avoidance in particular (Savo-nenko et al., 1999, 2003). The results of Experiment 1 seem toindicate that BLA lesions slow down 2wAA conditioning in such away that the performance of BLA-damaged rats followed a signifi-cant linear evolution that was less steep than in non-lesioned rats.The results from Experiment 3 confirm this effect, but also showclearly that the level of conditioning of the BLA-damaged group,even after 450 trials, did not reach that of the Control rats. This re-
Fig. 10. Experiment 3: survival curve representing the cumulative proportion ofsubjects that achieved the learning criterion in each training session, for eachexperimental group. The final percentage of subject that achieved the criterion isindicated besides each survival line. Note that 3 and 5 sessions, respectively, wererequired in order that 50% of subjects achieved the criterion in the Control and BLA-sham groups. In contrast, eight conditioning sessions were required in the BLA-damaged group.
sult also agrees with the suggestion that the amygdala could have along-term role in fear conditioning (Fanselow & Gale, 2003; Galeet al., 2004).
How could BLA lesions affect 2wAA conditioning? First, it hasbeen suggested that the reactive and defensive response to stimulithat predict danger and/or the ability of a conditioned stimulus toreinforce the acquisition of new or ‘‘active” responses would bemediated by direct pathways from the lateral to the basolateral nu-cleus (Amorapanth et al., 2000). Moreover, the BLA nucleus couldbe an important set of output connections to the striatum involvedin controlling actions like running to safety (Phelps & LeDoux,2005). Therefore, in the present study, acquisition of the instru-mental component of 2wAA conditioning could have been ham-pered by the BLA lesions. However, a certain degree ofconditioning has been observed. Even though we can not rule outsome contribution of surviving neurons of the BLA-damaged area,the maintenance of the impairing effects induced by the damagetogether with the large size of the present bilateral lesions led usto consider that other brain areas, including other amygdalar nu-clei, such as the Ce, might have partially compensate for BLA dam-age (Maren, 2008; Pare, Quirk, & LeDoux, 2004; Wilensky, Schafe,Kristensen, & LeDoux, 2006; Zimmerman, Rabinak, McLachlan, &Maren, 2007). Thus, as it has been shown in other types of fear-based learning including inhibitory avoidance and contextual fearconditioning (Berlau & McGaugh, 2003), it seems that, underappropriate training conditions, 2wAA can be partially acquiredand expressed independently of BLA.
In the present experiments, BLA lesions could also have ham-pered the modulatory role of BLA nucleus in memory consolidationof 2wAA conditioning. Thus, available evidence suggest that duringlearning the release of noradrenaline and acetylcholine in the baso-lateral complex of the amygdala could increase the activity of BLAneurons in the hours after the learning episodes, which in turncould facilitate synaptic plasticity in other brain structures relatedto memory storage (McGaugh, 2004).
5.2. Reversal effects of SS treatment on amnesic effects of BLA damage
In agreement with our previous research (Aldavert-Vera et al.,1996; Ruiz-Medina, Redolar-Ripoll, Morgado-Bernal, Aldavert-Vera, & Segura-Torres, 2008b; Segura-Torres et al., 1991), the re-sults of Experiment 2 show that post-training SS treatment facili-tated acquisition and long-term retention (10 days) of 2wAAconditioning in normal undamaged rats. The results of Experiment2 also showed that the SS treatment was able, not only to totallyreverse the 2wAA conditioning deficit, but also to improve learningin BLA-damaged animals. These findings agree with previous re-search from our laboratory, which shows the capacity of post-training SS treatment to reverse deficits and to improve the same2wAA conditioning in rats with thalamic lesions (Redolar-Ripollet al., 2003). These results are also in agreement with the resultsof experimental research that shows recovery from shuttle-boxavoidance deficit after ACTH (Bush, Lovely, & Pagano, 1973) or del-ta9-tetrahydrocannabinol (Pandina & Musty, 1975) administrationin rats with amygdaloid lesions. Likewise, a similar SS treatment,but administered pre-training, was also able to produce ameliora-tion of learning deficits induced by fornix lesion (Yoganarasimha &Meti, 1999). The results of Experiment 3 showed that, in contrastto the findings of other laboratories (Maren, 1999; Zimmermanet al., 2007), massive overtraining was not able to reverse the con-ditioning deficit in rats in which both BLA nuclei were seriouslydamaged. Although the possibility that further training could havemade it possible that rats achieved the acquisition level of normalrats cannot be disregarded, our results clearly indicate that SStreatment is more effective than overtraining for improving2wAA conditioning.

P. Segura-Torres et al. / Neurobiology of Learning and Memory 93 (2010) 117–126 125
How could post-training SS facilitate learning and memory inrats with damaged bilateral BLA or other brain structures? Firstof all, the fact that the damaged animals treated with SS did notshow alterations either in SS behavior, escape responses to theunconditioned stimulus or crossings into shuttle-box comparedto the Controls let us to suggest that the SS compensating effectwould not be mainly mediated by changes in reward, emotionalor motor systems. On the other hand, SS in the medial forebrainbundle produces extensive brain activation and neural changesthat can be related to its capacity to improve learning and memoryconsolidation. Thus, SS induces widespread increased expression ofc-Fos and other genes, most of which are related to neural plastic-ity, in memory related brain areas such as the amygdala, prefrontalcortex, hippocampus and dorsal striatum (Arvanitogiannis, Flores,& Shizgal, 1997; Arvanitogiannis, Tzschentke, Riscaldino, Wise, &Shizgal, 2000; Huguet et al., 2009). SS also increases the levels ofexcitatory neurotransmitters (Shankaranarayana Rao, Raju, & Meti,1998c) and induces long-lasting structural changes, such as in-creases in the dendritic intersection and length, in the numericaldensity of synapses and in the spine and in the number of excres-cences in the hippocampus (Shankaranarayana Rao, Raju, & Meti,1998a, 1998b). All of these SS-induced changes could overcomethe learning deficit produced by BLA lesions by activating otherundamaged regions or memory systems, such as the hippocampus.Recently it has been shown that these neuroanatomical and phys-iological SS effects in the hippocampus are related to its functionalproperties capable of ameliorating memory and restoring memorydeficits caused by stress (Ramkumar, Srikumar, ShankaranarayanaRao, & Raju, 2008).
Furthermore, although 2wAA conditioning in the present stan-dard-delay paradigm does not seem to depend on relational-hippo-campal memory system, the hippocampus seems to contribute tothis conditioning. Accordingly, learning-specific changes (Datta,Saha, Prutzman, Mullins, & Mavanji, 2005; Saha & Datta, 2005)and c-fos and zif/268 mRNA accumulation in the rat hippocampus(Nikolaev, Kaminska, Tischmeyer, Matthies, & Kaczmarek, 1992)after two-way active avoidance response training have been ob-served. Thus, it is possible that, after a certain amount of trainingexperience, SS treatment might turn on alternative hippocampal-dependent abilities, such as using contextual cues, to guide learn-ing that could replace some amygdala-dependent abilities lost(such as using discrete stimuli to guide behavior). Furthermore, re-cent work blurs the anatomical segregation of cue and context con-ditioning and challenges the necessity for the hippocampus andamygdala in fear learning (Maren, 2008). Specific experimentsaddressing particular SS action modes must be designed to confirmthis suggestion and improve our knowledge about the psychobio-logical nature of SS.
Acknowledgments
This research was supported by a Ministerio de Educación yCiencia (MEC) Grant (SEJ2005-02547/PSIC) and a Generalitat ofCatalunya Grant (2005SGR).
References
Aldavert-Vera, L., Costa-Miserachs, D., Massanes-Rotger, E., Soriano-Mas, C., Segura-Torres, P., & Morgado-Bernal, I. (1997). Facilitation of a distributed shuttle-boxconditioning with posttraining intracranial self-stimulation in old rats.Neurobiology of Learning and Memory, 67(3), 254–258.
Aldavert-Vera, L., Segura-Torres, P., Costa-Miserachs, D., & Morgado-Bernal, I.(1996). Shuttle-box memory facilitation by posttraining intracranial self-stimulation: Differential effects in rats with high and low basic conditioninglevels. Behavioral Neuroscience, 110(2), 346–352.
Amorapanth, P., LeDoux, J. E., & Nader, K. (2000). Different lateral amygdala outputsmediate reactions and actions elicited by a fear-arousing stimulus. NatureNeuroscience, 3(1), 74–79.
Arvanitogiannis, A., Flores, C., & Shizgal, P. (1997). Fos-like immunoreactivity in thecaudal diencephalon and brainstem following lateral hypothalamic self-stimulation. Behavioural Brain Research, 88(2), 275–279.
Arvanitogiannis, A., Tzschentke, T. M., Riscaldino, L., Wise, R. A., & Shizgal, P. (2000).Fos expression following self-stimulation of the medial prefrontal cortex.Behavioural Brain Research, 107(1–2), 123–132.
Bechara, A. (2005). Decision making, impulse control and loss of willpower to resistdrugs: A neurocognitive perspective. Nature Neuroscience, 8(11), 1458–1463.
Berlau, D. J., & McGaugh, J. L. (2003). Basolateral amygdala lesions do not preventmemory of context-footshock training. Learning and Memory, 10(6), 495–502.
Bush, D. F., Lovely, R. H., & Pagano, R. R. (1973). Injection of ACTH induces recoveryfrom shuttle-box avoidance deficits in rats with amygdaloid lesions. Journal ofComparative and Physiological Psychology, 83(1), 168–172.
Cahill, L. (2000). Neurobiological mechanisms of emotionally influenced, long-termmemory. Progress in Brain Research, 126, 29–37.
Coulombe, D., & White, N. (1980). The effect of post-training lateral hypothalamicself-stimulation on aversive and appetitive classical conditioning. Physiologyand Behavior, 25(2), 267–272.
Coulombe, D., & White, N. (1982). The effect of post-training hypothalamic self-stimulation on sensory preconditioning in rats. Canadian Journal of Psychology,36(1), 57–66.
Datta, S., Saha, S., Prutzman, S. L., Mullins, O. J., & Mavanji, V. (2005). Pontine-wavegenerator activation-dependent memory processing of avoidance learninginvolves the dorsal hippocampus in the rat. Journal of Neuroscience Research,80(5), 727–737.
Dwyer, D. M., & Killcross, S. (2006). Lesions of the basolateral amygdala disruptconditioning based on the retrieved representations of motivationallysignificant events. The Journal of Neuroscience, 26(32), 8305–8309.
Fanselow, M. S., & Gale, G. D. (2003). The amygdala, fear, and memory. Annals theNew York Academy of Sciences, 985, 125–134.
Gale, G. D., Anagnostaras, S. G., Godsil, B. P., Mitchell, S., Nozawa, T., Sage, J. R., et al.(2004). Role of the basolateral amygdala in the storage of fear memories acrossthe adult lifetime of rats. The Journal of Neuroscience, 24(15), 3810–3815.
Huguet, G., Aldavert-Vera, L., Kádár, E., Peña de Ortiz, S., Morgado-Bernal, I., &Segura-Torres, P. (2009). Intracranial self-stimulation to the lateralhypothalamus, a memory improving treatment, results in hippocampalchanges in gene expression. Neuroscience, 162, 359–374.
Huston, J. P., Mueller, C. C., & Mondadori, C. (1977). Memory facilitation by posttrialhypothalamic stimulation and other reinforcers: A central theory ofreinforcement. Biobehavioral Reviews, 1, 143–150.
Huston, J. P., & Mueller, C. C. (1978). Enhanced passive avoidance learning andappetitive T-maze learning with post-trial rewarding hypothalamicstimulation. Brain Research Bulletin, 3(3), 265–270.
Lalumiere, R. T., & McGaugh, J. L. (2005). Memory enhancement induced by post-training intrabasolateral amygdala infusions of beta-adrenergic or muscarinicagonists requires activation of dopamine receptors: Involvement of right, butnot left, basolateral amygdala. Learning and Memory, 12(5), 527–532 (ColdSpring Harbor, NY).
Lanuza, E., Nader, K., & Ledoux, J. E. (2004). Unconditioned stimulus pathways to theamygdala: Effects of posterior thalamic and cortical lesions on fearconditioning. Neuroscience, 125(2), 305–315.
LeDoux, J. E. (2000). Emotion circuits in the brain. Annual Review of Neuroscience, 23,155–184.
LeDoux, J. E. (2003). The emotional brain, fear, and the amygdala. Cellular andMolecular Neurobiology, 23(4–5), 727–738.
Maren, S. (1999). Neurotoxic basolateral amygdala lesions impair learning andmemory but not the performance of conditional fear in rats. The Journal ofNeuroscience, 19(19), 8696–8703.
Maren, S. (2008). Pavlovian fear conditioning as a behavioral assay for hippocampusand amygdala function: Cautions and caveats. The European Journal ofNeuroscience, 28(8), 1661–1666.
McGaugh, J. L. (2004). The amygdala modulates the consolidation of memories ofemotionally arousing experiences. Annual Review of Neuroscience, 27, 1–28.
Nader, K., Majidishad, P., Amorapanth, P., & LeDoux, J. E. (2001). Damage to thelateral and central, but not other, amygdaloid nuclei prevents the acquisition ofauditory fear conditioning. Learning and Memory, 8(3), 156–163.
Nathan, S. V., Griffith, Q. K., McReynolds, J. R., Hahn, E. L., & Roozendaal, B. (2004).Basolateral amygdala interacts with other brain regions in regulatingglucocorticoid effects on different memory functions. Annals of the New YorkAcademy of Sciences, 1032, 179–182.
Nikolaev, E., Kaminska, B., Tischmeyer, W., Matthies, H., & Kaczmarek, L. (1992).Induction of expression of genes encoding transcription factors in the rat brainelicited by behavioral training. Brain Research Bulletin, 28(3), 479–484.
Pandina, R. J., & Musty, R. E. (1975). Effects of delta9-tetrahydrocannabinol on activeavoidance acquisition and passive avoidance retention in rats with amygdaloidlesions. Pharmacology, 13(4), 297–308.
Pare, D., Quirk, G. J., & LeDoux, J. E. (2004). New vistas on amygdala networks inconditioned fear. Journal of Neurophysiology, 92(1), 1–9.
Paxinos, G., & Watson, C. (1998). The rat brain in stereotaxic coordinates. San Diego:Academic Press.
Phelps, E. A., & LeDoux, J. E. (2005). Contributions of the amygdala to emotionprocessing: From animal models to human behavior. Neuron, 48(2), 175–187.
Pitkanen, A., Savander, V., & LeDoux, J. E. (1997). Organization of intra-amygdaloidcircuitries in the rat: An emerging framework for understanding functions ofthe amygdala. Trends in Neurosciences, 20(11), 517–523.

126 P. Segura-Torres et al. / Neurobiology of Learning and Memory 93 (2010) 117–126
Ramkumar, K., Srikumar, B. N., Shankaranarayana Rao, B. S., & Raju, T. R. (2008). Self-stimulation rewarding experience restores stress-induced CA3 dendritic atrophy,spatial memory deficits and alterations in the levels of neurotransmitters in thehippocampus. Neurochemical Research, 33(9), 1651–1662.
Redolar-Ripoll, D., Aldavert-Vera, L., Soriano-Mas, C., Segura-Torres, P., & Morgado-Bernal, I. (2002). Intracranial self-stimulation facilitates memory consolidation,but not retrieval: Its effects are more effective than increased training.Behavioural Brain Research, 129(1–2), 65–75.
Redolar-Ripoll, D., Soriano-Mas, C., Guillazo-Blanch, G., Aldavert-Vera, L., Segura-Torres, P., & Morgado-Bernal, I. (2003). Posttraining intracranial self-stimulationameliorates the detrimental effects of parafascicular thalamic lesions on activeavoidance in young and aged rats. Behavioral Neuroscience, 117(2), 246–256.
Rodrigues, S. M., Schafe, G. E., & LeDoux, J. E. (2004). Molecular mechanismsunderlying emotional learning and memory in the lateral amygdala. Neuron,44(1), 75–91.
Roozendaal, B., Okuda, S., Van der Zee, E. A., & McGaugh, J. L. (2006). Glucocorticoidenhancement of memory requires arousal-induced noradrenergic activation inthe basolateral amygdala. Proceedings of the National Academy of Sciences of theUnited States of America, 103(17), 6741–6746.
Ruiz-Medina, J., Morgado-Bernal, I., Redolar-Ripoll, D., Aldavert-Vera, L., & Segura-Torres, P. (2008a). Intracranial self-stimulation facilitates a spatial learning andmemory task in the Morris water maze. Neuroscience, 154(2), 424–430.
Ruiz-Medina, J., Redolar-Ripoll, D., Morgado-Bernal, I., Aldavert-Vera, L., & Segura-Torres, P. (2008b). Intracranial self-stimulation improves memory consolidationin rats with little training. Neurobiology of Learning and Memory, 89, 574–581.
Sah, P., & Westbrook, R. F. (2008). Behavioural neuroscience. The circuit of fear.Nature, 454(7204), 589–590.
Saha, S., & Datta, S. (2005). Two-way active avoidance training-specific increases inphosphorylated cAMP response element-binding protein in the dorsalhippocampus, amygdala, and hypothalamus. The European Journal ofNeuroscience, 21(12), 3403–3414.
Savonenko, A., Filipkowski, R. K., Werka, T., Zielinski, K., & Kaczmarek, L. (1999).Defensive conditioning-related functional heterogeneity among nuclei of therat amygdala revealed by c-fos mapping. Neuroscience, 94(3), 723–733.
Savonenko, A., Werka, T., Nikolaev, E., Zielinski, K., & Kaczmarek, L. (2003). Complexeffects of NMDA receptor antagonist APV in the basolateral amygdala on
acquisition of two-way avoidance reaction and long-term fear memory.Learning and Memory, 10(4), 293–303.
Segura-Torres, P., Capdevila-Ortis, L., Marti-Nicolovius, M., & Morgado-Bernal, I.(1988). Improvement of shuttle-box learning with pre- and post-trialintracranial self-stimulation in rats. Behavioural Brain Research, 29(1–2),111–117.
Segura-Torres, P., Portell-Cortes, I., & Morgado-Bernal, I. (1991). Improvement ofshuttle-box avoidance with post-training intracranial self-stimulation, in rats:A parametric study. Behavioural Brain Research, 42(2), 161–167.
Shankaranarayana Rao, B. S., Raju, T. R., & Meti, B. L. (1998a). Alterations in thedensity of excrescences in CA3 neurons of hippocampus in rats subjected toself-stimulation experience. Brain Research, 804(2), 320–324.
Shankaranarayana Rao, B. S., Raju, T. R., & Meti, B. L. (1998b). Long-lasting structuralchanges in CA3 hippocampal and layer V motor cortical pyramidal neuronsassociated with self-stimulation rewarding experience. A quantitative Golgistudy. Brain Research Bulletin, 47(1), 95–101.
Shankaranarayana Rao, B. S., Raju, T. R., & Meti, B. L. (1998c). Self-stimulation oflateral hypothalamus and ventral tegmentum increases the levels ofnoradrenaline, dopamine, glutamate, and AChE activity, but not 5-hydroxytryptamine and GABA levels in hippocampus and motor cortex.Neurochemical Research, 23(8), 1053–1059.
Soriano-Mas, C., Redolar-Ripoll, D., Aldavert-Vera, L., Morgado-Bernal, I., & Segura-Torres, P. (2005). Post-training intracranial self-stimulation facilitates ahippocampus-dependent task. Behavioural Brain Research, 160(1), 141–147.
Wilensky, A. E., Schafe, G. E., Kristensen, M. P., & LeDoux, J. E. (2006). Rethinking thefear circuit: The central nucleus of the amygdala is required for the acquisition,consolidation, and expression of pavlovian fear conditioning. The Journal ofNeuroscience, 26(48), 12387–12396.
Yoganarasimha, D., & Meti, B. L. (1999). Amelioration of fornix lesion inducedlearning deficits by self-stimulation rewarding experience. Brain Research,845(2), 246–251.
Zimmerman, J. M., Rabinak, C. A., McLachlan, I. G., & Maren, S. (2007). The centralnucleus of the amygdala is essential for acquiring and expressing conditionalfear after overtraining. Learning and Memory, 14(9), 634–644.





























