Embed Size (px)
Citation preview

Neuropharmacology 53 (2007) 318e329www.elsevier.com/locate/neuropharm
Intracellular cholesterol changes induced by translocator protein(18 kDa) TSPO/PBR ligands
Angela Maria Falchi a, Barbara Battetta b, Francesca Sanna b, Marco Piludu a, Valeria Sogos a,Mariangela Serra c, Marta Melis a, Martina Putzolu a, Giacomo Diaz a,*
a Department of Cytomorphology, Cittadella Universitaria, University of Cagliari, Monserrato, 09100 Cagliari, Italyb Department of Biomedical Science & Biotechnology, Cittadella Universitaria, University of Cagliari, 09100 Cagliari, Italy
c Department of Experimental Biology & Center of Excellence for Neurobiology of Drug Dependence, Cittadella Universitaria,University of Cagliari, 09100 Cagliari, Italy
Received 10 April 2006; received in revised form 4 May 2007; accepted 20 May 2007
Abstract
One of the main functions of the translocator protein (18 kDa) or TSPO, previously known as peripheral-type benzodiazepine receptor, is theregulation of cholesterol import into mitochondria for steroid biosynthesis. In this paper we show that TSPO ligands induce changes in the dis-tribution of intracellular cholesterol in astrocytes and fibroblasts. NBD-cholesterol, a fluorescent analog of cholesterol, was rapidly removedfrom membranes and accumulated into lipid droplets. This change was followed by a block of cholesterol esterification, but not by modificationof intracellular cholesterol synthesis. NBD-cholesterol droplets were in part released in the medium, and increased cholesterol efflux was ob-served in [3H]cholesterol-prelabeled cells. TSPO ligands also induced a prominent shrinkage and depolarization of mitochondria and depletionof acidic vesicles with cytoplasmic acidification. Consistent with NBD-cholesterol changes, MTT assay showed enhanced accumulation offormazan into lipid droplets and inhibition of formazan exocytosis after treatment with TSPO ligands. The effects of specific TSPO ligandsPK 11195 and Ro5-4864 were reproduced by diazepam, which binds with high affinity both TSPO and central benzodiazepine receptors,but not by clonazepam, which binds exclusively to GABA receptor, and other amphiphilic substances such as DIDS and propranolol. All theseeffects and the parallel immunocytochemical detection of TSPO in potentially steroidogenic cells (astrocytes) and non-steroidogenic cells(fibroblasts) suggest that TSPO is involved in the regulation and trafficking of intracellular cholesterol by means of mechanisms not necessarilyrelated to steroid biosynthesis.� 2007 Elsevier Ltd. All rights reserved.
Keywords: TSPO; Translocator protein (18 kDa); PBR; Cholesterol; Mitochondria; Lipid droplets; MTT; Acidic vesicles; Immunocytochemistry
1. Introduction
One of the most extensively characterized functions of thetranslocator protein (18 kDa) or TSPO (Papadopoulos et al.,2006), previously known as peripheral-type benzodiazepinereceptor (PBR), is the mitochondrial import of cholesterol(Hauet et al., 2005; Miller, 1988; Simpson and Waterman,1983) delivered to mitochondria by cholesterol-transfer
* Corresponding author. Tel.: þ39 070 6754081; fax: þ39 070 675 4003.
E-mail address: [email protected] (G. Diaz).
0028-3908/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.neuropharm.2007.05.016
proteins (StAR, MLN64 and, less specifically, SCP-2) (Chan-derbhan et al., 1982; Clark et al., 1994; Watari et al., 1997).This represents the first and rate-determining step in steroidhormone biosynthesis. Accordingly, the localization, structureand pharmacology of TSPO and the effects induced by TSPOligands have been widely investigated in steroidogenic cells(for review see Casellas et al., 2002; Gavish et al., 1999;Papadopoulos, 2004). On the other hand, effects of TSPOligands have also been found in non-steroidogenic cells, affect-ing different physiological mechanisms such as respiration inheart, kidney and liver (Moreno-Sanchez et al., 1991; Veenmanand Gavish, 2006), generation of reactive oxygen species in
319A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
neurons (Jayakumar et al., 2002) and HL60 human leukemiacells (Fennell et al., 2001), anion transport in kidney (Basileet al., 1988), mitochondrial permeability transition in cardio-myocytes (Chelli et al., 2001), inhibition of cell proliferationin human fibroblasts and fibrosarcoma cells (Kletsas et al.,2004) and apoptosis in various cell lines (Chelli et al., 2004;Decaudin et al., 2002). Increased tumorigenicity has beenfound in TSPO antisense knockdown cells (Levin et al.,2005; Weisinger et al., 2004). Many of these effects havebeen attributed to the intimate association of TSPO witha complex of proteins present in mitochondrial membranes,including the voltage-dependent anion channel (VDAC), theadenine nucleotide transporter (McEnery et al., 1992), andthe mitochondrial transition pore complex (Kroemer et al.,1997). However, in some cases, effects of TSPO ligandshave been observed also in TSPO-deficient (Hans et al.,2005) and TSPO-knockdown cells (Gonzalez-Polo et al.,2005; Kletsas et al., 2004), raising the issue of the possiblepresence of TSPO-independent mechanisms of actions. Oneof these mechanisms may rely on the inhibition of the mito-chondrial inner membrane anionic channel (IMAC) exertedby several TSPO ligands, namely Ro5-4864 (a 40-chloro deriv-ative of diazepam) and PK 11195 (an isoquinoline carboxa-mide derivative) (Beavis, 1989, 1992). This hypothesis hasrecently been supported by data obtained by Aon et al.(2003) in cardiomyocytes where mitochondrial potential
oscillations were prevented by PK 11195, Ro5-4864 but alsoby DIDS, a stilbene-2,20-disulfonate which inhibits IMAC.
In the present study we investigated the acute effect ofTSPO ligands on the intracellular distribution of cholesterol,using the fluorescent analog NBD-cholesterol, and the con-comitant effects on mitochondria and acidic vesicles, usingpotential-sensitive (TMRM) and pH-sensitive (acridine or-ange) probes. We also evaluated cholesterol synthesis, esterifi-cation and efflux. Our findings indicate consistent changes inthe distribution, esterification and efflux of intracellular cho-lesterol, accompanied by mitochondrial depolarization anddepletion of acidic vesicles. These changes were observed indifferent cell types such as rat astrocytes, fibroblasts and3T3 mouse fibroblasts, which represent potentially steroido-genic and non-steroidogenic cells. This suggests that choles-terol changes induced by TSPO ligands are not necessarilyrelated to steroid biosynthesis.
2. Materials and methods
2.1. Cell cultures
Mouse Swiss 3T3 fibroblasts (ATCC collection) and primary cultures of rat
newborn astrocytes and rat newborn fibroblasts were used in this study. Exper-
imental procedures meet the UK and EC ethical policy guidelines and regula-
tions for the use of laboratory animals. Astrocytes were characterized by
positivity to glial fibrillary acidic protein antibody. Rat fibroblasts were
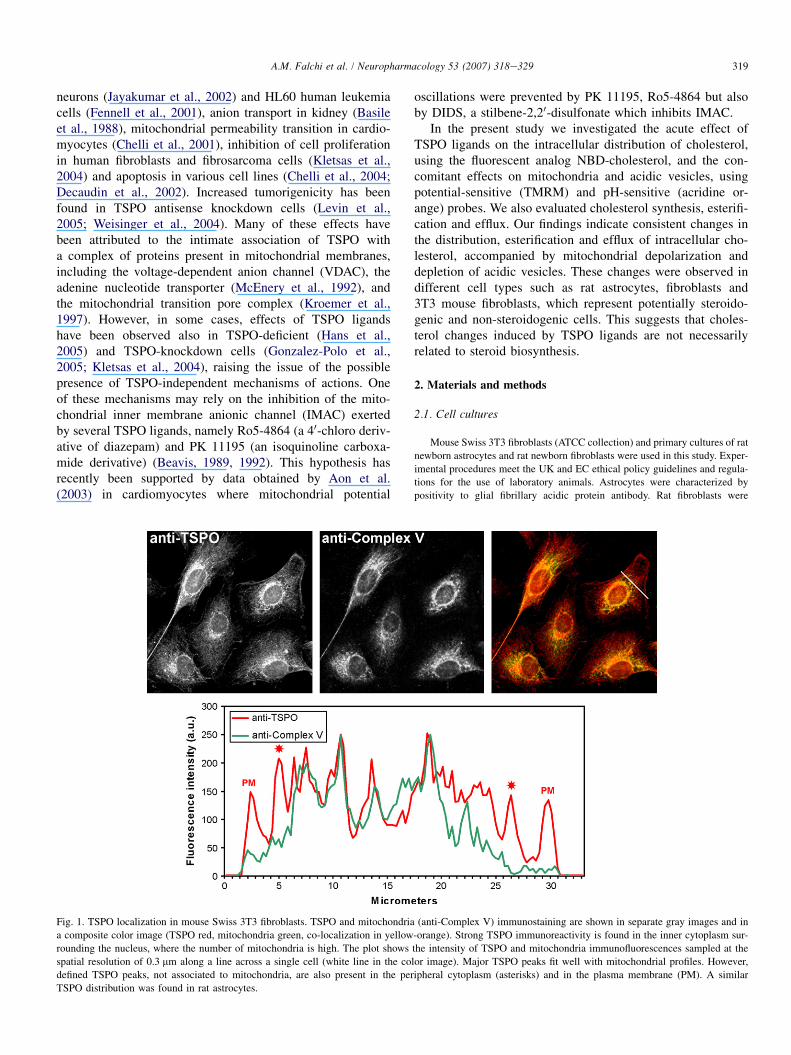
Fig. 1. TSPO localization in mouse Swiss 3T3 fibroblasts. TSPO and mitochondria (anti-Complex V) immunostaining are shown in separate gray images and in
a composite color image (TSPO red, mitochondria green, co-localization in yellow-orange). Strong TSPO immunoreactivity is found in the inner cytoplasm sur-
rounding the nucleus, where the number of mitochondria is high. The plot shows the intensity of TSPO and mitochondria immunofluorescences sampled at the
spatial resolution of 0.3 mm along a line across a single cell (white line in the color image). Major TSPO peaks fit well with mitochondrial profiles. However,
defined TSPO peaks, not associated to mitochondria, are also present in the peripheral cytoplasm (asterisks) and in the plasma membrane (PM). A similar
TSPO distribution was found in rat astrocytes.
320 A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
characterized by positivity to vimentin and negativity to desmin and factor
VIII antibodies. All cells were grown in Dulbecco’s modified Eagle’s medium
with high glucose, supplemented with 10% fetal bovine serum, in a 5% CO2
incubator at 37 �C. Experiments were performed when cells were subconflu-
ent. Cells were plated in glass-bottomed dishes (MatTek, Ashland, MA) to al-
low the use of immersion objectives in fluorescence and contrast microscopy.
Before treatments, serum-supplemented medium was substituted with serum-
free medium. Experiments were replicated in different cell types using the
same probe and drug concentrations, incubation times and data acquisition
parameters.
2.2. Drugs
Cells were treated in vivo with: 30e60 mM 7-chloro-5-(4-chlorophenyl)-
1,3-dihydro-1-methyl-2H-1,4-benzodiazepin-2-one (Ro5-4864), acutely added
(a.a.); 30e60 mM 1-(2-chlorophenyl-N-methyl-1-methylpropyl)-3-isoquino-
line-carboxamide (PK 11195), a.a.; 100e400 mM 2H-1,4-benzodiazepin-
2-one (diazepam), a.a.; 100e400 mM 5-(2-chlorophenyl)-7-nitro-3H-1,4-ben-
zodiazepin-2(1H)-one (clonazepam), a.a.; 100e400 mM 4,40-diisothiocya-
nato-stilbene-2,20-disulfonate (DIDS), a.a.; 60e400 mM propranolol, a.a.;
20e40 mM carbonyl cyanide 3-chloro-phenylhydrazone (CCCP), a.a.; 2 mM
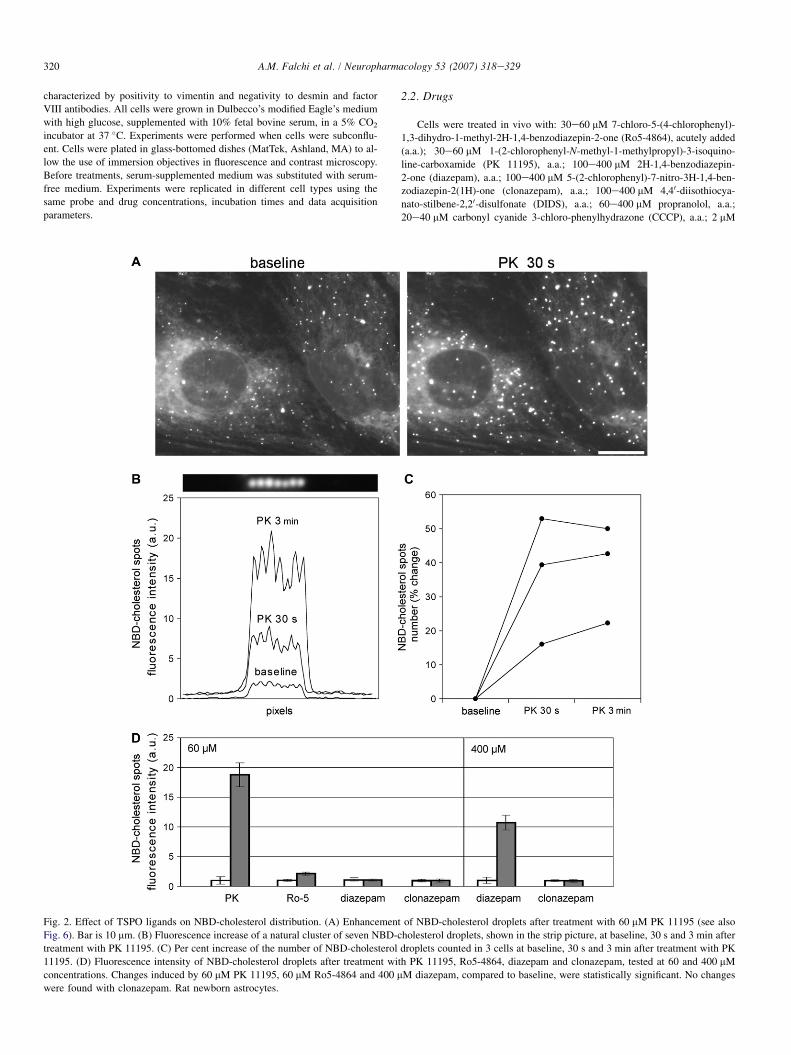
Fig. 2. Effect of TSPO ligands on NBD-cholesterol distribution. (A) Enhancement of NBD-cholesterol droplets after treatment with 60 mM PK 11195 (see also
Fig. 6). Bar is 10 mm. (B) Fluorescence increase of a natural cluster of seven NBD-cholesterol droplets, shown in the strip picture, at baseline, 30 s and 3 min after
treatment with PK 11195. (C) Per cent increase of the number of NBD-cholesterol droplets counted in 3 cells at baseline, 30 s and 3 min after treatment with PK
11195. (D) Fluorescence intensity of NBD-cholesterol droplets after treatment with PK 11195, Ro5-4864, diazepam and clonazepam, tested at 60 and 400 mM
concentrations. Changes induced by 60 mM PK 11195, 60 mM Ro5-4864 and 400 mM diazepam, compared to baseline, were statistically significant. No changes
were found with clonazepam. Rat newborn astrocytes.
321A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
cyclosporin A (CsA) for 30 min; 4 mM Sandoz-compound 58-035, an inhibitor
of acylCoA/cholesterol acyltransferase 1 (ACAT1) for 30 min or 24 h; 1 mg/ml
phalloidin for 30 min; 1 mM paclitaxel for 45 min; 100 nM colchicine for
30 min or 2 h. Vehicles were: Me2SO for PK 11195, Ro 5-4864, diazepam,
CCCP, propranolol, DIDS, paclitaxel and Sandoz 58-035; ethanol for CsA;
water for phalloidin and colchicine. Stock solutions were 1000-fold concen-
trated, to not exceed the 0.1% concentration of vehicle in the medium. The
same concentration of vehicle was added to control cells. Ro5-4864, PK
11195, diazepam, propranolol, DIDS, CsA, CCCP, colchicine, phalloidin
and paclitaxel were from Sigma (St. Louis, MO, USA). Sandoz 58-035 was
kindly donated by Novartis (Dayton, NJ, USA).
2.3. Probes
Cells were supravitally stained with the following probes (ex, em ¼ fluor-
escence excitation and emission): 5 mM 22-[N-nitrobenz-2-oxa-1,3-diazol-4-yl)
amino]-23,24-bisnor-5-cholen-3-ol (NBD-cholesterol) for 1 h (ex 460 � 25,
em 535 � 20); 300 nM Nile Red for 15 min (ex 460 � 25, em 535 � 20 for
nonpolar lipids; ex 540 � 12, em 590 LP for all lipids); 100 nM acridine
orange for 10 min (ex 540 � 12.5, em 590 LP); 100 nM tetramethylrhodamine
methyl ester perchlorate (TMRM) for 30 min (ex 540 � 12.5, em 590 LP);
100 nM Mito Tracker Green FM (MitoTracker) for 20 min (ex 460 � 25,
em 535 � 20); 20 mM dihydrofluorescein diacetate (DHF) for 30 min (ex
460 � 25, em 535 � 20); 1.25 mM MitoSOX for 10 min (ex 540 � 12.5, em
590 LP); 45 mM thiazolyl blue tetrazolium bromide (MTT) for 5e20 min;
10 mM calcein for 15 min (ex 480 � 20, em 520 � 20). NBD-cholesterol is
a fluorescent analog of cholesterol where the terminal segment of the alkyl
tail is replaced by the NBD fluorophore. NBD-cholesterol fluorescence is
environment-sensitive, being weak in aqueous media and very bright in non-
polar media (Rukmini et al., 2001). NBD-cholesterol has been successfully
used to study the mechanism of sterol transfer to mitochondrial membranes
by the steroidogenic acute regulatory protein (StAR) and sterol carrier pro-
tein-2 (SCP-2) (Petrescu et al., 2001). Owing to the overlap of NBD-cholesterol
and Nile Red emissions in the 535 � 20 band, for co-localization cells were
first incubated with NBD-cholesterol, then fixed in 4% paraformaldehyde to
block cytoplasm movements, photographed mapping the field position, over-
stained with Nile Red and re-photographed. MTT is a colorless compound
readily taken up by living cells and reduced to formazan (FMZ), an intensely
purple substance which can be observed in brightfield microscopy. FMZ is
extruded by exocytosis forming characteristic needle-like crystals on the cell
surface. Acridine orange is a pH-sensitive probe, which selectively stains
acidic vesicles. TMRM is a potential-sensitive probe, which selectively accu-
mulates in the mitochondrial matrix. MitoTracker is a potential-insensitive
mitochondrial probe, which binds covalently the mitochondrial matrix, thus
enabling structural changes to be detected even after mitochondrial depola-
rization. DHF is a non-fluorescent derivative of fluorescein, which reverts to
fluorescent fluorescein upon oxidation by reactive oxygen species (ROS).
MitoSOX is a fluorigenic dye, which selectively detects superoxide in mito-
chondria. However, MitoSOX fluorescence is not permanent as it leaves mito-
chondria after depolarization. Calcein is a membrane-impermeant fluorescent
dye used to test plasma membrane integrity. Vehicles were: Me2SO for
TMRM, DHF, MitoSOX, Nile Red, MitoTracker and calcein; chloroform for
NBD-cholesterol; water for MTT and acridine orange. Stock solutions were
1000-fold concentrated, to not exceed the 0.1% concentration of vehicle in
the medium. MTT was from Sigma (St. Louis, MO, USA); Nile Red and cal-
cein from Fluka (Buchs, SG, Switzerland); acridine orange from Merck
(Darmstadt, Germany). Other fluorescent probes were from Molecular Probes
(Eugene, OR, USA).
2.4. TSPO immunocytochemistry
Cells were fixed in 4% paraformaldehyde in PBS for 30 min, washed three
times in PBS for 5 min and incubated overnight with a polyclonal anti-TSPO
antibody (6361-PC-100, R&D Systems, Minneapolis, MN, USA). In some ex-
periments, the anti-TSPO antibody was used in combination with a mitochon-
drial antibody (anti-Complex V, Molecular Probes) to verify the association of
TSPO with mitochondria. Co-localization was semiquantitatively assessed by
linear profiles of BPR and Complex V immunofluorescences sampled at the
spatial resolution of 0.3 mm (Fig. 1).
2.5. Imaging
Observations were made using an Olympus IX 71 inverted microscope
(Olympus, Tokyo, Japan) equipped with 20� and 60� planapochromatic ob-
jectives (Olympus UPlanSApo series) with efficient chromatic correction,
which minimized the focus and planar drift between different fluorescence fil-
ters. This was a critical requisite for the co-localization of probes, as the size
of small lipid droplets was near to the microscope resolution limit. The nom-
inal resolutions of images taken with 20� and 60� objectives were 0.3 and
0.1 mm/pixel, respectively. The use of the inverted microscope excluded any
risk of dye contamination, as there was no contact between the immersion ob-
jective and the medium. Images were taken with a 12-bit cooled CCD camera
(Sensicam PCO, Kelheim, Germany), electronically coupled to a mechanical
shutter interposed between the 100 W Hg lamp and the microscope, to limit
illumination of cells for the time strictly required for the acquisition of images.
Excitation light was attenuated with a 6% transmittance neutral density filter.
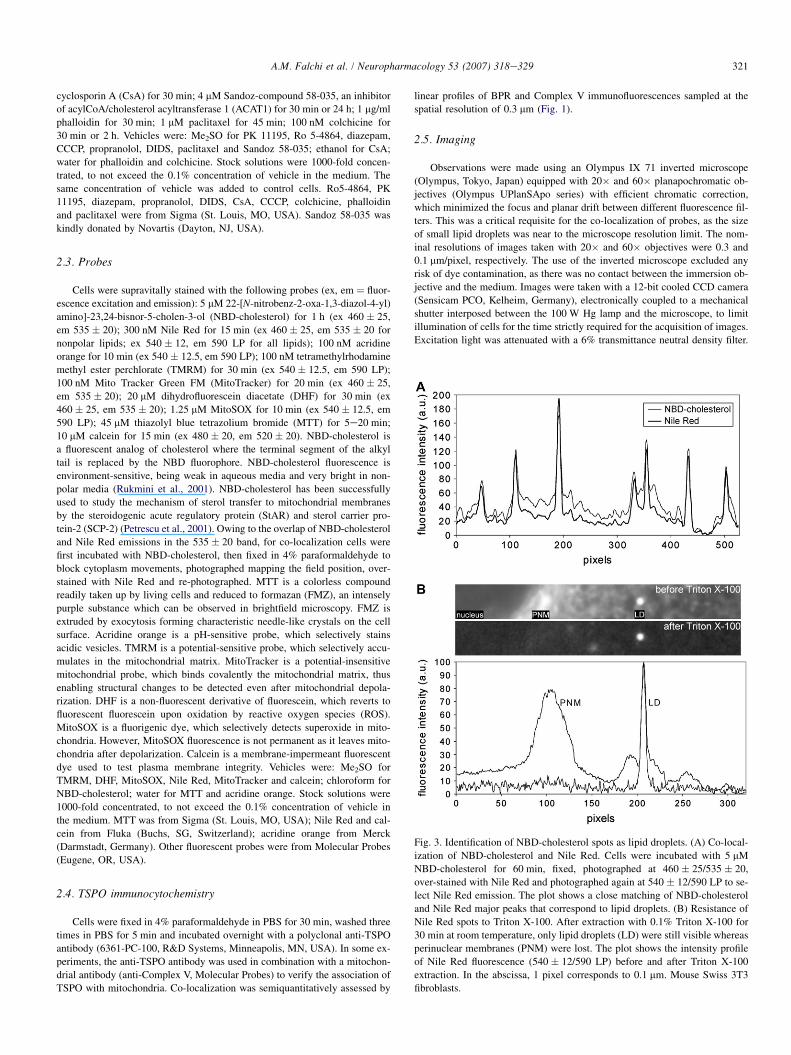
Fig. 3. Identification of NBD-cholesterol spots as lipid droplets. (A) Co-local-
ization of NBD-cholesterol and Nile Red. Cells were incubated with 5 mM
NBD-cholesterol for 60 min, fixed, photographed at 460 � 25/535 � 20,
over-stained with Nile Red and photographed again at 540 � 12/590 LP to se-
lect Nile Red emission. The plot shows a close matching of NBD-cholesterol
and Nile Red major peaks that correspond to lipid droplets. (B) Resistance of
Nile Red spots to Triton X-100. After extraction with 0.1% Triton X-100 for
30 min at room temperature, only lipid droplets (LD) were still visible whereas
perinuclear membranes (PNM) were lost. The plot shows the intensity profile
of Nile Red fluorescence (540 � 12/590 LP) before and after Triton X-100
extraction. In the abscissa, 1 pixel corresponds to 0.1 mm. Mouse Swiss 3T3
fibroblasts.
322 A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
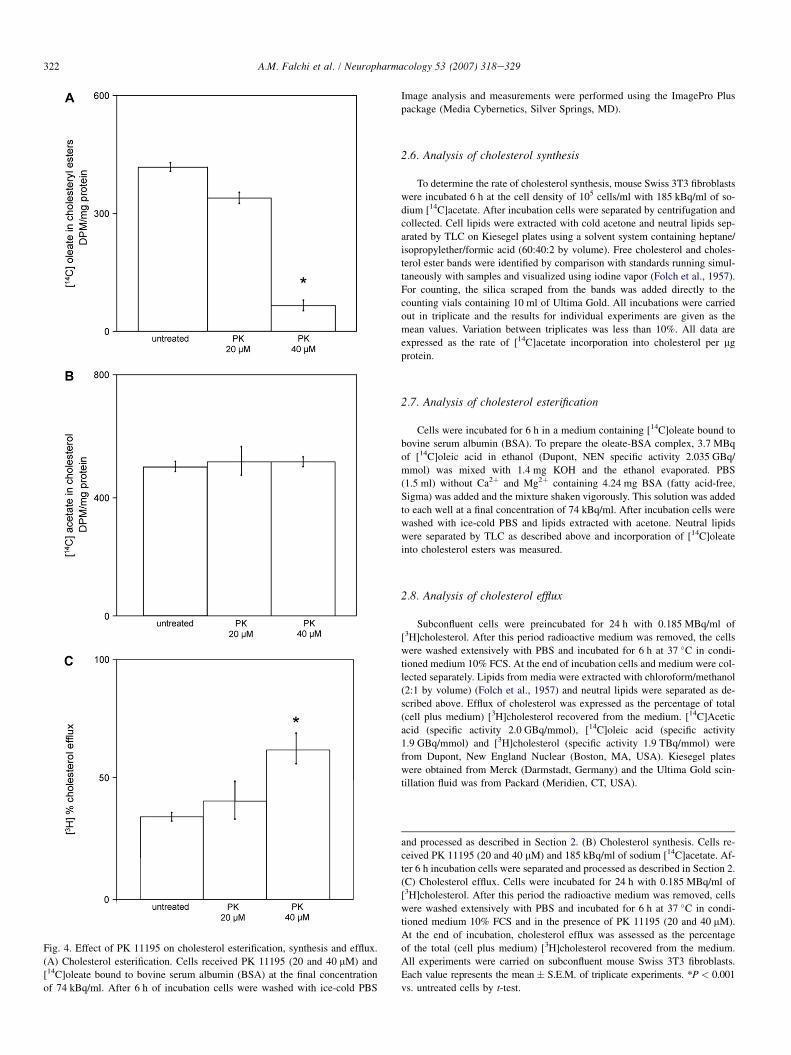
Fig. 4. Effect of PK 11195 on cholesterol esterification, synthesis and efflux.
(A) Cholesterol esterification. Cells received PK 11195 (20 and 40 mM) and
[14C]oleate bound to bovine serum albumin (BSA) at the final concentration
of 74 kBq/ml. After 6 h of incubation cells were washed with ice-cold PBS
Image analysis and measurements were performed using the ImagePro Plus
package (Media Cybernetics, Silver Springs, MD).
2.6. Analysis of cholesterol synthesis
To determine the rate of cholesterol synthesis, mouse Swiss 3T3 fibroblasts
were incubated 6 h at the cell density of 105 cells/ml with 185 kBq/ml of so-
dium [14C]acetate. After incubation cells were separated by centrifugation and
collected. Cell lipids were extracted with cold acetone and neutral lipids sep-
arated by TLC on Kiesegel plates using a solvent system containing heptane/
isopropylether/formic acid (60:40:2 by volume). Free cholesterol and choles-
terol ester bands were identified by comparison with standards running simul-
taneously with samples and visualized using iodine vapor (Folch et al., 1957).
For counting, the silica scraped from the bands was added directly to the
counting vials containing 10 ml of Ultima Gold. All incubations were carried
out in triplicate and the results for individual experiments are given as the
mean values. Variation between triplicates was less than 10%. All data are
expressed as the rate of [14C]acetate incorporation into cholesterol per mg
protein.
2.7. Analysis of cholesterol esterification
Cells were incubated for 6 h in a medium containing [14C]oleate bound to
bovine serum albumin (BSA). To prepare the oleate-BSA complex, 3.7 MBq
of [14C]oleic acid in ethanol (Dupont, NEN specific activity 2.035 GBq/
mmol) was mixed with 1.4 mg KOH and the ethanol evaporated. PBS
(1.5 ml) without Ca2þ and Mg2þ containing 4.24 mg BSA (fatty acid-free,
Sigma) was added and the mixture shaken vigorously. This solution was added
to each well at a final concentration of 74 kBq/ml. After incubation cells were
washed with ice-cold PBS and lipids extracted with acetone. Neutral lipids
were separated by TLC as described above and incorporation of [14C]oleate
into cholesterol esters was measured.
2.8. Analysis of cholesterol efflux
Subconfluent cells were preincubated for 24 h with 0.185 MBq/ml of
[3H]cholesterol. After this period radioactive medium was removed, the cells
were washed extensively with PBS and incubated for 6 h at 37 �C in condi-
tioned medium 10% FCS. At the end of incubation cells and medium were col-
lected separately. Lipids from media were extracted with chloroform/methanol
(2:1 by volume) (Folch et al., 1957) and neutral lipids were separated as de-
scribed above. Efflux of cholesterol was expressed as the percentage of total
(cell plus medium) [3H]cholesterol recovered from the medium. [14C]Acetic
acid (specific activity 2.0 GBq/mmol), [14C]oleic acid (specific activity
1.9 GBq/mmol) and [3H]cholesterol (specific activity 1.9 TBq/mmol) were
from Dupont, New England Nuclear (Boston, MA, USA). Kiesegel plates
were obtained from Merck (Darmstadt, Germany) and the Ultima Gold scin-
tillation fluid was from Packard (Meridien, CT, USA).
and processed as described in Section 2. (B) Cholesterol synthesis. Cells re-
ceived PK 11195 (20 and 40 mM) and 185 kBq/ml of sodium [14C]acetate. Af-
ter 6 h incubation cells were separated and processed as described in Section 2.
(C) Cholesterol efflux. Cells were incubated for 24 h with 0.185 MBq/ml of
[3H]cholesterol. After this period the radioactive medium was removed, cells
were washed extensively with PBS and incubated for 6 h at 37 �C in condi-
tioned medium 10% FCS and in the presence of PK 11195 (20 and 40 mM).
At the end of incubation, cholesterol efflux was assessed as the percentage
of the total (cell plus medium) [3H]cholesterol recovered from the medium.
All experiments were carried on subconfluent mouse Swiss 3T3 fibroblasts.
Each value represents the mean � S.E.M. of triplicate experiments. *P < 0.001
vs. untreated cells by t-test.
323A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
3. Results
3.1. TSPO immunocytochemistry
TSPO immunoreactivity was found in both astrocytes and3T3 fibroblasts. The intensity of immunoreaction was muchhigher in astrocytes than in 3T3 fibroblasts. However, theintracellular distribution of TSPO immunostaining was similarin the two types of cells. Dual immunostaining of TSPO andmitochondria (Complex V) showed a strongly preferentiallocalization of TSPO in mitochondria, mostly concentratedin the perinuclear cytoplasm. However, TSPO immunostainingwas also detected in peripheral regions of cytoplasm, devoid ofmitochondria, and in the plasma membrane (Fig. 1).
3.2. Enhancement of NBD-cholesterol droplets
In untreated cells, the fluorescent cholesterol analog NBD-cholesterol produced a moderate fluorescence of organellemembranes (endoplasmic reticulum, mitochondria, etc.) andfew intensely fluorescent spots apparently similar to lipid drop-lets. Surprisingly, treatment with TSPO ligands resulted in anincrease of fluorescence of NBD-cholesterol spots, both inastrocytes (Fig. 2A) and fibroblasts (Fig. 5). The change wasvery rapid, being apparent as soon as 30 s after treatment. The
strongest effect was observed with 60 mM PK 11195, resultingin an increase of both the fluorescence intensity and number ofNBD-cholesterol spots (Fig. 2B, C). Diazepam (400 mM) andRo5-4864 (60 mM) produced only an increase of fluorescenceintensity (Fig. 2D). The increase of fluorescence could not beattributed to an extra NBD-cholesterol uptake or endocytosis,as cells were washed several times after incubation with thefluorescent probe. Moreover, the fluorescence increase of NBD-cholesterol spots was accompanied by a decrease of the perinu-clear fluorescence, suggesting a process of NBD-cholesteroltransfer from cytoplasmic membranes to isolated spots.
To verify the nature of NBD-cholesterol spots, cells wereoverstained with Nile Red, a specific stain of lipid droplets.Though Nile Red can be used in vivo, cells were fixed to blockorganelle movements and co-localize probes at micron resolu-tion. All NBD-cholesterol spots were stained by Nile Red, thusrevealing the nature of lipid droplets (Fig. 3A). In addition,NBD-cholesterol spots resisted extraction with 0.1% TritonX-100 for 30 min at room temperature, whereas fluorescenttraces of cytoplasmic membranes were lost (Fig. 3B). Thesefindings support the hypothesis that NBD-cholesterol is ini-tially partitioned between cytoplasmic membranes and lipiddroplets. Treatment with TSPO ligands induces a redistributionof NBD-cholesterol, which preferentially accumulates in lipiddroplets.
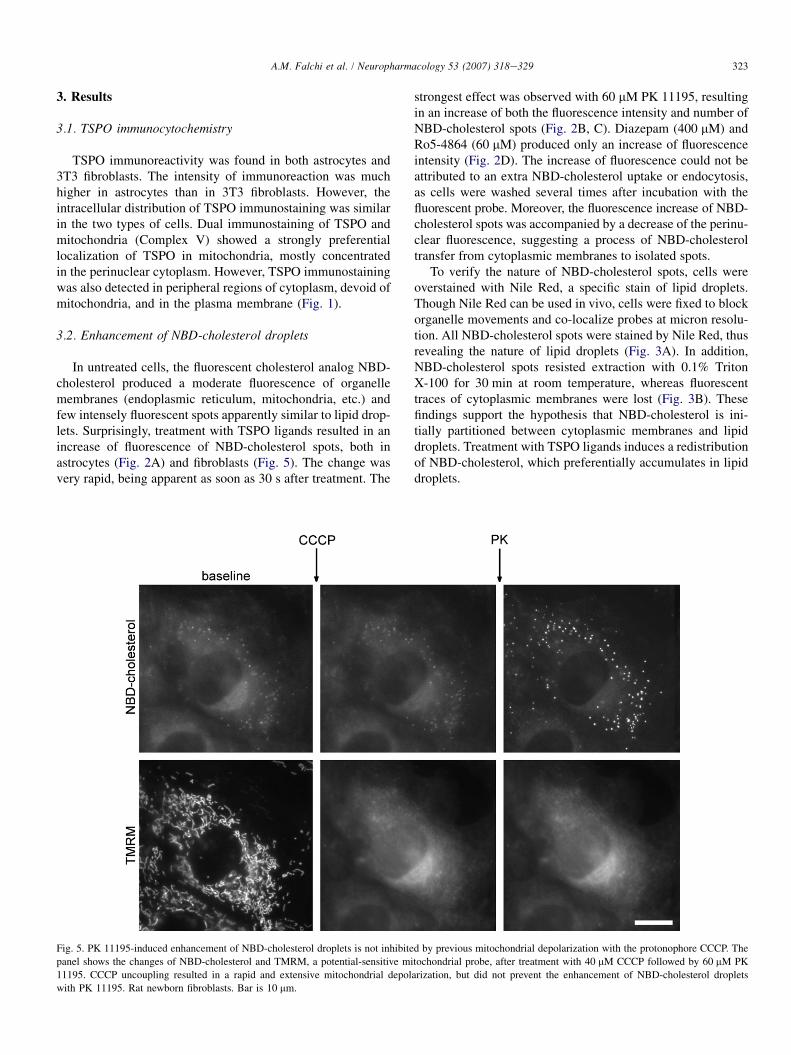
Fig. 5. PK 11195-induced enhancement of NBD-cholesterol droplets is not inhibited by previous mitochondrial depolarization with the protonophore CCCP. The
panel shows the changes of NBD-cholesterol and TMRM, a potential-sensitive mitochondrial probe, after treatment with 40 mM CCCP followed by 60 mM PK
11195. CCCP uncoupling resulted in a rapid and extensive mitochondrial depolarization, but did not prevent the enhancement of NBD-cholesterol droplets
with PK 11195. Rat newborn fibroblasts. Bar is 10 mm.
324 A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
Although NBD-cholesterol has been proven to be a suitablesubstrate for esterification by acylCoA/cholesterol acyltrans-ferase (ACAT) (Lada et al., 2004), overnight treatment withthe Sandoz-compound 58-035, a potent ACAT inhibitor, didnot prevent the enhancement of NBD-cholesterol droplets in-duced by TSPO ligands. Experiments with radioactive markers,in the absence of NBD-cholesterol, showed that PK 11195itself (Ro5-4864 not tested) is a strong inhibitor of cholesterolesterification (Fig. 4A). Owing to the relatively long (6 h)duration of PK 11195 treatments required to assess cholesterolesterification and cholesterol synthesis, PK 11195 concentra-tions were slightly lower than those used in acute administration
(20e40 mM rather than 30e60 mM). PK 11195 (40 mM) in-duced a complete block of cholesterol esterification, whereasthe effect of 20 mM PK 11195 was remarkably lower. However,PK 11195 did not modify endogenous cholesterol synthesis(Fig. 4B) nor did it affect cell viability, as evaluated by trypanblue exclusion test. The relative agreement between the rangeof effective concentrations required to inhibit cholesterol ester-ification and to enhance NBD-cholesterol droplets suggests theexistence of a common mechanism of action.
The accumulation of unesterified NBD-cholesterol in lipiddroplets does not contrast with the composition of lipid drop-lets which are known to contain mostly esterified cholesterol.
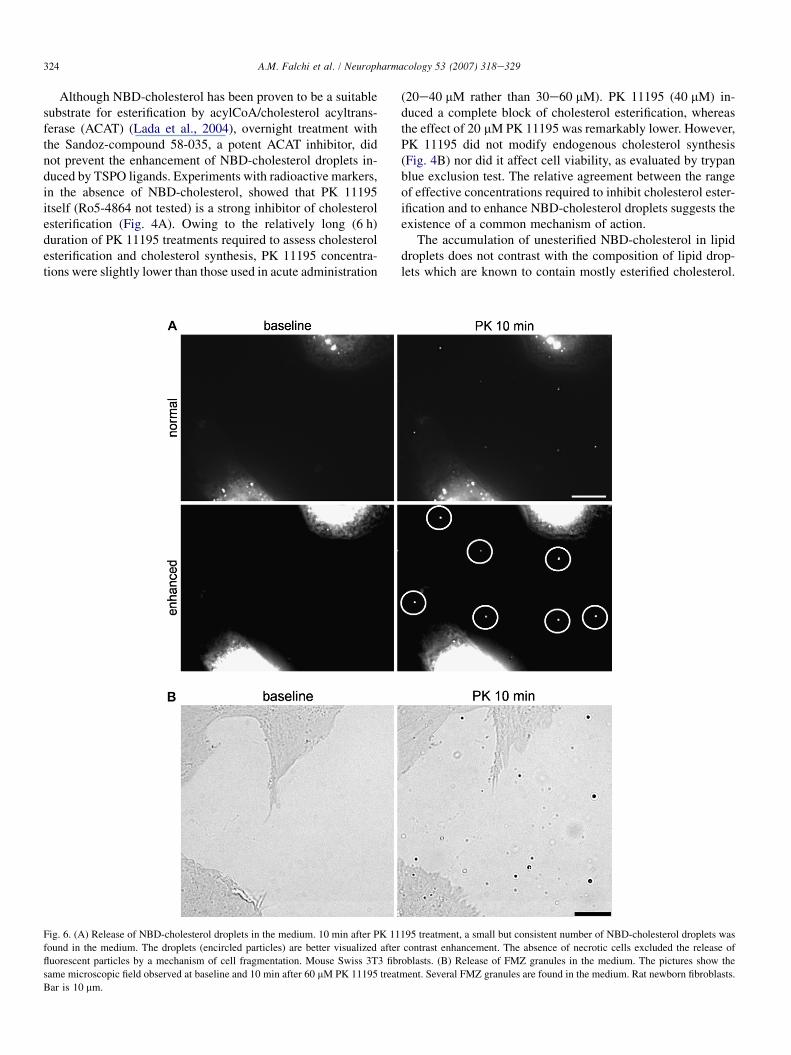
Fig. 6. (A) Release of NBD-cholesterol droplets in the medium. 10 min after PK 11195 treatment, a small but consistent number of NBD-cholesterol droplets was
found in the medium. The droplets (encircled particles) are better visualized after contrast enhancement. The absence of necrotic cells excluded the release of
fluorescent particles by a mechanism of cell fragmentation. Mouse Swiss 3T3 fibroblasts. (B) Release of FMZ granules in the medium. The pictures show the
same microscopic field observed at baseline and 10 min after 60 mM PK 11195 treatment. Several FMZ granules are found in the medium. Rat newborn fibroblasts.
Bar is 10 mm.
325A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
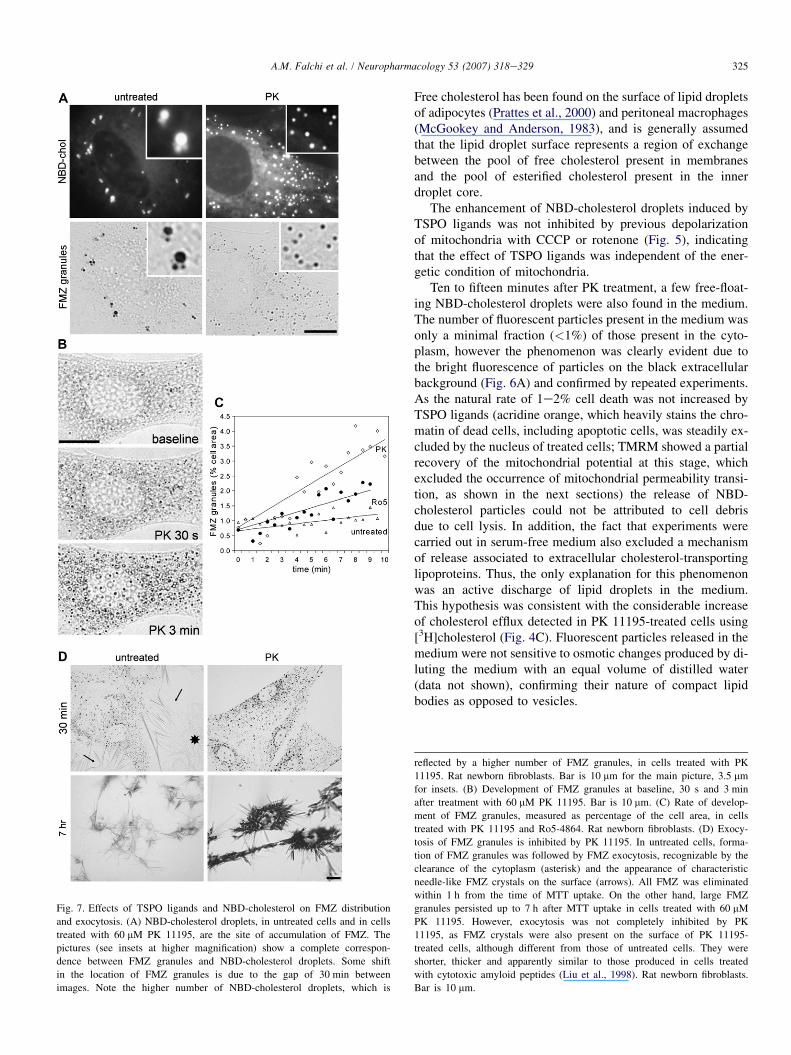
Fig. 7. Effects of TSPO ligands and NBD-cholesterol on FMZ distribution
and exocytosis. (A) NBD-cholesterol droplets, in untreated cells and in cells
treated with 60 mM PK 11195, are the site of accumulation of FMZ. The
pictures (see insets at higher magnification) show a complete correspon-
dence between FMZ granules and NBD-cholesterol droplets. Some shift
in the location of FMZ granules is due to the gap of 30 min between
images. Note the higher number of NBD-cholesterol droplets, which is
Free cholesterol has been found on the surface of lipid dropletsof adipocytes (Prattes et al., 2000) and peritoneal macrophages(McGookey and Anderson, 1983), and is generally assumedthat the lipid droplet surface represents a region of exchangebetween the pool of free cholesterol present in membranesand the pool of esterified cholesterol present in the innerdroplet core.
The enhancement of NBD-cholesterol droplets induced byTSPO ligands was not inhibited by previous depolarizationof mitochondria with CCCP or rotenone (Fig. 5), indicatingthat the effect of TSPO ligands was independent of the ener-getic condition of mitochondria.
Ten to fifteen minutes after PK treatment, a few free-float-ing NBD-cholesterol droplets were also found in the medium.The number of fluorescent particles present in the medium wasonly a minimal fraction (<1%) of those present in the cyto-plasm, however the phenomenon was clearly evident due tothe bright fluorescence of particles on the black extracellularbackground (Fig. 6A) and confirmed by repeated experiments.As the natural rate of 1e2% cell death was not increased byTSPO ligands (acridine orange, which heavily stains the chro-matin of dead cells, including apoptotic cells, was steadily ex-cluded by the nucleus of treated cells; TMRM showed a partialrecovery of the mitochondrial potential at this stage, whichexcluded the occurrence of mitochondrial permeability transi-tion, as shown in the next sections) the release of NBD-cholesterol particles could not be attributed to cell debrisdue to cell lysis. In addition, the fact that experiments werecarried out in serum-free medium also excluded a mechanismof release associated to extracellular cholesterol-transportinglipoproteins. Thus, the only explanation for this phenomenonwas an active discharge of lipid droplets in the medium.This hypothesis was consistent with the considerable increaseof cholesterol efflux detected in PK 11195-treated cells using[3H]cholesterol (Fig. 4C). Fluorescent particles released in themedium were not sensitive to osmotic changes produced by di-luting the medium with an equal volume of distilled water(data not shown), confirming their nature of compact lipidbodies as opposed to vesicles.
reflected by a higher number of FMZ granules, in cells treated with PK
11195. Rat newborn fibroblasts. Bar is 10 mm for the main picture, 3.5 mm
for insets. (B) Development of FMZ granules at baseline, 30 s and 3 min
after treatment with 60 mM PK 11195. Bar is 10 mm. (C) Rate of develop-
ment of FMZ granules, measured as percentage of the cell area, in cells
treated with PK 11195 and Ro5-4864. Rat newborn fibroblasts. (D) Exocy-
tosis of FMZ granules is inhibited by PK 11195. In untreated cells, forma-
tion of FMZ granules was followed by FMZ exocytosis, recognizable by the
clearance of the cytoplasm (asterisk) and the appearance of characteristic
needle-like FMZ crystals on the surface (arrows). All FMZ was eliminated
within 1 h from the time of MTT uptake. On the other hand, large FMZ
granules persisted up to 7 h after MTT uptake in cells treated with 60 mM
PK 11195. However, exocytosis was not completely inhibited by PK
11195, as FMZ crystals were also present on the surface of PK 11195-
treated cells, although different from those of untreated cells. They were
shorter, thicker and apparently similar to those produced in cells treated
with cytotoxic amyloid peptides (Liu et al., 1998). Rat newborn fibroblasts.
Bar is 10 mm.
326 A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
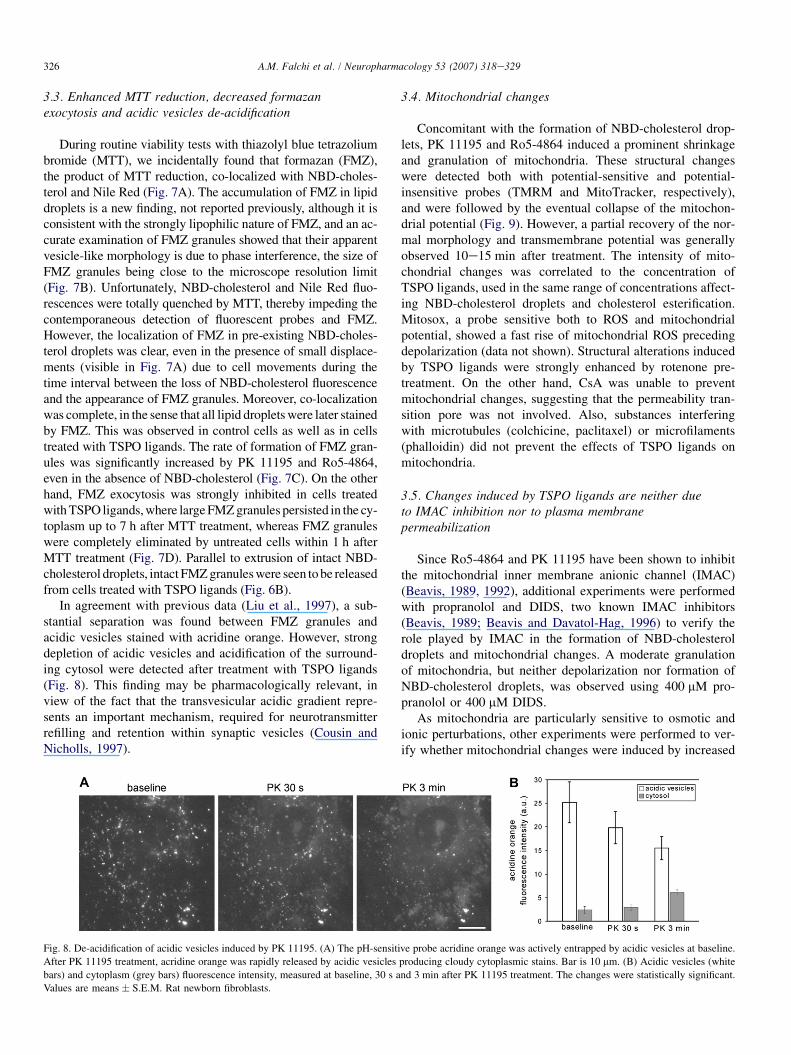
3.3. Enhanced MTT reduction, decreased formazanexocytosis and acidic vesicles de-acidification
During routine viability tests with thiazolyl blue tetrazoliumbromide (MTT), we incidentally found that formazan (FMZ),the product of MTT reduction, co-localized with NBD-choles-terol and Nile Red (Fig. 7A). The accumulation of FMZ in lipiddroplets is a new finding, not reported previously, although it isconsistent with the strongly lipophilic nature of FMZ, and an ac-curate examination of FMZ granules showed that their apparentvesicle-like morphology is due to phase interference, the size ofFMZ granules being close to the microscope resolution limit(Fig. 7B). Unfortunately, NBD-cholesterol and Nile Red fluo-rescences were totally quenched by MTT, thereby impeding thecontemporaneous detection of fluorescent probes and FMZ.However, the localization of FMZ in pre-existing NBD-choles-terol droplets was clear, even in the presence of small displace-ments (visible in Fig. 7A) due to cell movements during thetime interval between the loss of NBD-cholesterol fluorescenceand the appearance of FMZ granules. Moreover, co-localizationwas complete, in the sense that all lipid droplets were later stainedby FMZ. This was observed in control cells as well as in cellstreated with TSPO ligands. The rate of formation of FMZ gran-ules was significantly increased by PK 11195 and Ro5-4864,even in the absence of NBD-cholesterol (Fig. 7C). On the otherhand, FMZ exocytosis was strongly inhibited in cells treatedwith TSPO ligands, where large FMZ granules persisted in the cy-toplasm up to 7 h after MTT treatment, whereas FMZ granuleswere completely eliminated by untreated cells within 1 h afterMTT treatment (Fig. 7D). Parallel to extrusion of intact NBD-cholesterol droplets, intact FMZ granules were seen to be releasedfrom cells treated with TSPO ligands (Fig. 6B).
In agreement with previous data (Liu et al., 1997), a sub-stantial separation was found between FMZ granules andacidic vesicles stained with acridine orange. However, strongdepletion of acidic vesicles and acidification of the surround-ing cytosol were detected after treatment with TSPO ligands(Fig. 8). This finding may be pharmacologically relevant, inview of the fact that the transvesicular acidic gradient repre-sents an important mechanism, required for neurotransmitterrefilling and retention within synaptic vesicles (Cousin andNicholls, 1997).
3.4. Mitochondrial changes
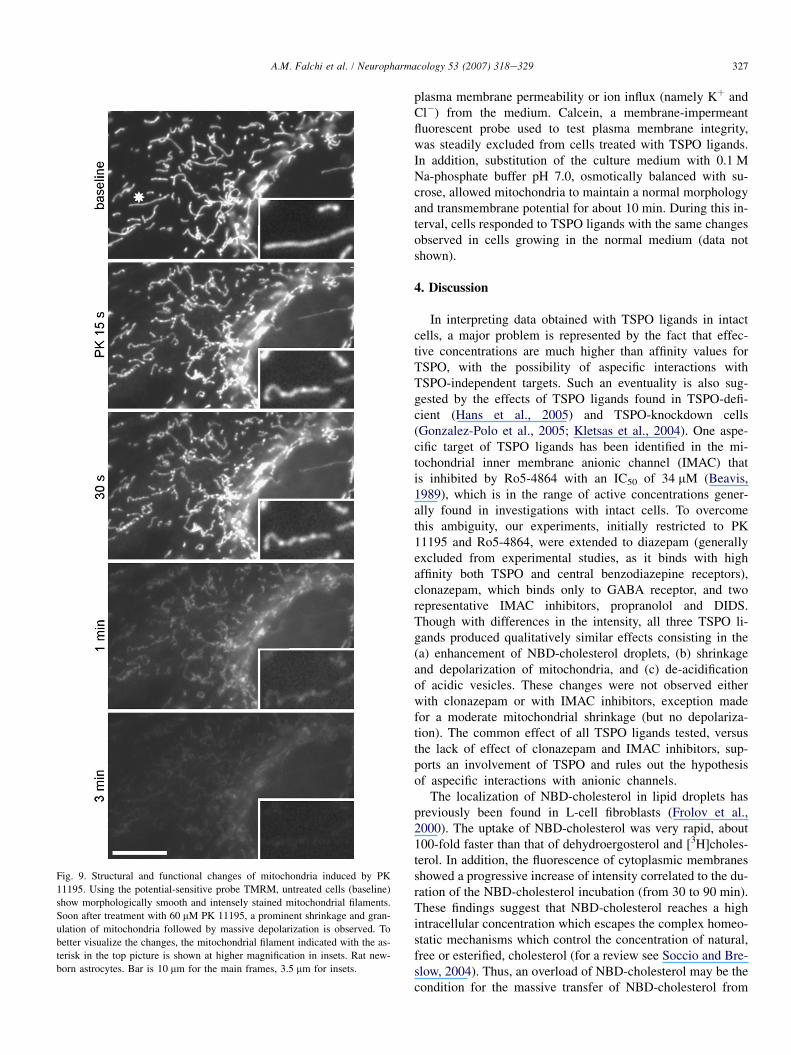
Concomitant with the formation of NBD-cholesterol drop-lets, PK 11195 and Ro5-4864 induced a prominent shrinkageand granulation of mitochondria. These structural changeswere detected both with potential-sensitive and potential-insensitive probes (TMRM and MitoTracker, respectively),and were followed by the eventual collapse of the mitochon-drial potential (Fig. 9). However, a partial recovery of the nor-mal morphology and transmembrane potential was generallyobserved 10e15 min after treatment. The intensity of mito-chondrial changes was correlated to the concentration ofTSPO ligands, used in the same range of concentrations affect-ing NBD-cholesterol droplets and cholesterol esterification.Mitosox, a probe sensitive both to ROS and mitochondrialpotential, showed a fast rise of mitochondrial ROS precedingdepolarization (data not shown). Structural alterations inducedby TSPO ligands were strongly enhanced by rotenone pre-treatment. On the other hand, CsA was unable to preventmitochondrial changes, suggesting that the permeability tran-sition pore was not involved. Also, substances interferingwith microtubules (colchicine, paclitaxel) or microfilaments(phalloidin) did not prevent the effects of TSPO ligands onmitochondria.
3.5. Changes induced by TSPO ligands are neither dueto IMAC inhibition nor to plasma membranepermeabilization
Since Ro5-4864 and PK 11195 have been shown to inhibitthe mitochondrial inner membrane anionic channel (IMAC)(Beavis, 1989, 1992), additional experiments were performedwith propranolol and DIDS, two known IMAC inhibitors(Beavis, 1989; Beavis and Davatol-Hag, 1996) to verify therole played by IMAC in the formation of NBD-cholesteroldroplets and mitochondrial changes. A moderate granulationof mitochondria, but neither depolarization nor formation ofNBD-cholesterol droplets, was observed using 400 mM pro-pranolol or 400 mM DIDS.
As mitochondria are particularly sensitive to osmotic andionic perturbations, other experiments were performed to ver-ify whether mitochondrial changes were induced by increased
Fig. 8. De-acidification of acidic vesicles induced by PK 11195. (A) The pH-sensitive probe acridine orange was actively entrapped by acidic vesicles at baseline.
After PK 11195 treatment, acridine orange was rapidly released by acidic vesicles producing cloudy cytoplasmic stains. Bar is 10 mm. (B) Acidic vesicles (white
bars) and cytoplasm (grey bars) fluorescence intensity, measured at baseline, 30 s and 3 min after PK 11195 treatment. The changes were statistically significant.
Values are means � S.E.M. Rat newborn fibroblasts.
327A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
Fig. 9. Structural and functional changes of mitochondria induced by PK
11195. Using the potential-sensitive probe TMRM, untreated cells (baseline)
show morphologically smooth and intensely stained mitochondrial filaments.
Soon after treatment with 60 mM PK 11195, a prominent shrinkage and gran-
ulation of mitochondria followed by massive depolarization is observed. To
better visualize the changes, the mitochondrial filament indicated with the as-
terisk in the top picture is shown at higher magnification in insets. Rat new-
born astrocytes. Bar is 10 mm for the main frames, 3.5 mm for insets.
plasma membrane permeability or ion influx (namely Kþ andCl�) from the medium. Calcein, a membrane-impermeantfluorescent probe used to test plasma membrane integrity,was steadily excluded from cells treated with TSPO ligands.In addition, substitution of the culture medium with 0.1 MNa-phosphate buffer pH 7.0, osmotically balanced with su-crose, allowed mitochondria to maintain a normal morphologyand transmembrane potential for about 10 min. During this in-terval, cells responded to TSPO ligands with the same changesobserved in cells growing in the normal medium (data notshown).
4. Discussion
In interpreting data obtained with TSPO ligands in intactcells, a major problem is represented by the fact that effec-tive concentrations are much higher than affinity values forTSPO, with the possibility of aspecific interactions withTSPO-independent targets. Such an eventuality is also sug-gested by the effects of TSPO ligands found in TSPO-defi-cient (Hans et al., 2005) and TSPO-knockdown cells(Gonzalez-Polo et al., 2005; Kletsas et al., 2004). One aspe-cific target of TSPO ligands has been identified in the mi-tochondrial inner membrane anionic channel (IMAC) thatis inhibited by Ro5-4864 with an IC50 of 34 mM (Beavis,1989), which is in the range of active concentrations gener-ally found in investigations with intact cells. To overcomethis ambiguity, our experiments, initially restricted to PK11195 and Ro5-4864, were extended to diazepam (generallyexcluded from experimental studies, as it binds with highaffinity both TSPO and central benzodiazepine receptors),clonazepam, which binds only to GABA receptor, and tworepresentative IMAC inhibitors, propranolol and DIDS.Though with differences in the intensity, all three TSPO li-gands produced qualitatively similar effects consisting in the(a) enhancement of NBD-cholesterol droplets, (b) shrinkageand depolarization of mitochondria, and (c) de-acidificationof acidic vesicles. These changes were not observed eitherwith clonazepam or with IMAC inhibitors, exception madefor a moderate mitochondrial shrinkage (but no depolariza-tion). The common effect of all TSPO ligands tested, versusthe lack of effect of clonazepam and IMAC inhibitors, sup-ports an involvement of TSPO and rules out the hypothesisof aspecific interactions with anionic channels.
The localization of NBD-cholesterol in lipid droplets haspreviously been found in L-cell fibroblasts (Frolov et al.,2000). The uptake of NBD-cholesterol was very rapid, about100-fold faster than that of dehydroergosterol and [3H]choles-terol. In addition, the fluorescence of cytoplasmic membranesshowed a progressive increase of intensity correlated to the du-ration of the NBD-cholesterol incubation (from 30 to 90 min).These findings suggest that NBD-cholesterol reaches a highintracellular concentration which escapes the complex homeo-static mechanisms which control the concentration of natural,free or esterified, cholesterol (for a review see Soccio and Bre-slow, 2004). Thus, an overload of NBD-cholesterol may be thecondition for the massive transfer of NBD-cholesterol from

328 A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
cytoplasmic membranes to lipid droplets, induced by TSPO li-gands. On the other hand, the fact that TSPO ligands alonewere capable of this change suggests a specific interaction be-tween TSPO and cholesterol. The almost complete block ofcholesterol esterification and the nearly two-fold increase ofcholesterol efflux, detected with methods that did not involvethe use of NBD-cholesterol, support this hypothesis. It shouldbe verified whether the release of NBD-cholesterol droplets,in the absence of cholesterol-transporting lipoproteins, alsooccurs in vivo, as this would represent an as yet unknownform of sterol release.
The increased rate of formation of FMZ granules inducedby TSPO ligands is consistent with the increased mass oflipid droplets, which provide a higher number of availablesites for FMZ accumulation. The transfer of cholesterol tolipid bodies may also explain the inhibition of FMZ exocy-tosis, in agreement with the fact that FMZ exocytosis dependson the availability of free (membrane) cholesterol (Liu et al.,1998).
The detection of mitochondrial alterations by MitoTracker,a probe scarcely influenced by the mitochondrial potential,proved the structural nature of changes induced by TSPO li-gands. The observed granulation and fragmentation of mito-chondria is also consistent with data recently obtained by Aonet al. (2003) in cardiomyocytes with PK 11195 and Ro5-4864at micromolar concentrations. In these cells, TSPO ligands ef-fectively blocked cell-wide oscillations of mitochondrial poten-tial and propagation of mitochondrial ROS locally induced byphotoactivation. As cardiomyocyte mitochondria are linked innetwork by specific inter-organelle junctions (Amchenkovaet al., 1988), it is likely that mitochondrial fragmentation wouldalso interrupt propagation of mitochondrial potential fluctua-tions and ROS. The structural and functional changes of mito-chondria correlate with the prevalent localization of TSPO inthis organelle, as confirmed by immunocytochemical data ob-tained in astrocytes and 3T3 fibroblasts. The different intensityof the immunoreaction, which was lower in 3T3 fibroblasts, isconsistent with the fact that 3T3 fibroblasts have been foundto express TSPO mRNA levels comparable to those of steroido-genic cells, but lower levels of protein transcripts (Giatzakis andPapadopoulos, 2004).
At present, no hypotheses can be put forward as to the mech-anisms by which TSPO ligands induce the various changesfound in this study. However, despite their apparent heteroge-neity, several common features, which constitute a basis forcomprehension, can be delineated. In summary, TSPO ligandsinduced rapid and simultaneous effects in distinct subcellularcompartments, including membrane-delimited organelles (mito-chondria and acidic vesicles), lipid droplets and lipid-basedinclusions (formazan). These effects were detected both in po-tentially steroidogenic (astrocytes) and non-steroidogenic (3T3fibroblasts) cells, thus ruling out an exclusive relationshipwith steroid biosynthesis. This is also confirmed by the unre-sponsiveness of NBD-cholesterol changes to disruption of themitochondrial electrochemical gradient, as the mitochondrialimport of StAR has been shown not to occur in de-energizedmitochondria (King and Stocco, 1996; King et al., 1999).
Acknowledgment
This work was supported in part by grant RBAU01C-CAJ_003 from MIUR-FIRB to G.D.
References
Amchenkova, A.A., Bakeeva, L.E., Chentsov, Y.S., Skulachev, V.P.,
Zorov, D.B., 1988. Coupling membranes as energy-transmitting cables.
I. Filamentous mitochondria in fibroblasts and mitochondrial clusters in
cardiomyocytes. J. Cell Biol. 107, 481e495.
Aon, M.A., Cortassa, S., Marban, E., O’Rourke, B., 2003. Synchronized whole
cell oscillations in mitochondrial metabolism triggered by a local release
of reactive oxygen species in cardiac myocytes. J. Biol. Chem. 278,
44735e44744.
Basile, A.S., Lueddens, H.W., Skolnick, P., 1988. Regulation of renal periph-
eral benzodiazepine receptors by anion transport inhibitors. Life Sci. 42,
715e726.
Beavis, A.D., 1989. On the inhibition of the mitochondrial inner membrane
anion uniporter by cationic amphiphiles and other drugs. J. Biol. Chem.
264, 1508e1515.
Beavis, A.D., 1992. Properties of the inner membrane anion channel in intact
mitochondria. J. Bioenerg. Biomembr. 24, 77e90.
Beavis, A.D., Davatol-Hag, H., 1996. The mitochondrial inner membrane an-
ion channel is inhibited by DIDS. J. Bioenerg. Biomembr. 28, 207e214.
Casellas, P., Galiegue, S., Basile, A.S., 2002. Peripheral benzodiazepine recep-
tors and mitochondrial function. Neurochem. Int. 40, 475e486.
Chanderbhan, R., Noland, B.J., Scallen, T.J., Vahouny, G.V., 1982. Sterol car-
rier protein2. Delivery of cholesterol from adrenal lipid droplets to mito-
chondria for pregnenolone synthesis. J. Biol. Chem. 257, 8928e8934.
Chelli, B., Falleni, A., Salvetti, F., Gremigni, V., Lucacchini, A., Martini, C.,
2001. Peripheral-type benzodiazepine receptor ligands: mitochondrial per-
meability transition induction in rat cardiac tissue. Biochem. Pharmacol.
61, 695e705.
Chelli, B., Lena, A., Vanacore, R., Pozzo, E.D., Costa, B., Rossi, L.,
Salvetti, A., Scatena, F., Ceruti, S., Abbracchio, M.P., Gremigni, V.,
Martini, C., 2004. Peripheral benzodiazepine receptor ligands: mitochon-
drial transmembrane potential depolarization and apoptosis induction in
rat C6 glioma cells. Biochem. Pharmacol. 68, 125e134.
Clark, B.J., Wells, J., King, S.R., Stocco, D.M., 1994. The purification, clon-
ing, and expression of a novel luteinizing hormone-induced mitochondrial
protein in MA-10 mouse Leydig tumor cells. Characterization of the
steroidogenic acute regulatory protein (StAR). J. Biol. Chem. 269, 28314e28322.
Cousin, M.A., Nicholls, D.G., 1997. Synaptic vesicle recycling in cultured cer-
ebellar granule cells: role of vesicular acidification and refilling. J. Neuro-
chem. 69, 1927e1935.
Decaudin, D., Castedo, M., Nemati, F., Beurdeley-Thomas, A., De Pinieux, G.,
Caron, A., Pouillart, P., Wijdenes, J., Rouillard, D., Kroemer, G.,
Poupon, M.F., 2002. Peripheral benzodiazepine receptor ligands reverse
apoptosis resistance of cancer cells in vitro and in vivo. Cancer Res. 62,
1388e1393.
Fennell, D.A., Corbo, M., Pallaska, A., Cotter, F.E., 2001. Bcl-2 resistant mi-
tochondrial toxicity mediated by the isoquinoline carboxamide PK11195
involves de novo generation of reactive oxygen species. Br. J. Cancer
84, 1397e1404.
Folch, J., Lees, M., Sloane Stanley, G.H., 1957. A simple method for the iso-
lation and purification of total lipids from animal tissues. J. Biol. Chem.
226, 497e509.
Frolov, A., Petrescu, A., Atshaves, B.P., So, P.T., Gratton, E., Serrero, G.,
Schroeder, F., 2000. High density lipoprotein-mediated cholesterol uptake
and targeting to lipid droplets in intact L-cell fibroblasts. A single- and
multiphoton fluorescence approach. J. Biol. Chem. 275, 12769e12780.
Gavish, M., Bachman, I., Shoukrun, R., Katz, Y., Veenman, L., Weisinger, G.,
Weizman, A., 1999. Enigma of the peripheral benzodiazepine receptor.
Pharmacol. Rev. 51, 629e650.

329A.M. Falchi et al. / Neuropharmacology 53 (2007) 318e329
Giatzakis, C., Papadopoulos, V., 2004. Differential utilization of the promoter
of peripheral-type benzodiazepine receptor by steroidogenic versus non-
steroidogenic cell lines and the role of Sp1 and Sp3 in the regulation of
basal activity. Endocrinology 145, 1113e1123.
Gonzalez-Polo, R.A., Carvalho, G., Braun, T., Decaudin, D., Fabre, C.,
Larochette, N., Perfettini, J.L., Djavaheri-Mergny, M., Youlyouz-
Marfak, I., Codogno, P., Raphael, M., Feuillard, J., Kroemer, G., 2005.
PK11195 potently sensitizes to apoptosis induction independently from
the peripheral benzodiazepine receptor. Oncogene 24, 7503e7513.
Hans, G., Wislet-Gendebien, S., Lallemend, F., Robe, P., Rogister, B.,
Belachew, S., Nguyen, L., Malgrange, B., Moonen, G., Rigo, J.M., 2005.
Peripheral benzodiazepine receptor (PBR) ligand cytotoxicity unrelated
to PBR expression. Biochem. Pharmacol. 69, 819e830.
Hauet, T., Yao, Z.X., Bose, H.S., Wall, C.T., Han, Z., Li, W., Hales, D.B.,
Miller, W.L., Culty, M., Papadopoulos, V., 2005. Peripheral-type benzodi-
azepine receptor-mediated action of steroidogenic acute regulatory protein
on cholesterol entry into leydig cell mitochondria. Mol. Endocrinol. 19,
540e554.
Jayakumar, A.R., Panickar, K.S., Norenberg, M.D., 2002. Effects on free
radical generation by ligands of the peripheral benzodiazepine receptor
in cultured neural cells. J. Neurochem. 83, 1226e1234.
King, S.R., Stocco, D.M., 1996. ATP and a mitochondrial electrochemical gra-
dient are required for functional activity of the steroidogenic acute regula-
tory (StAR) protein in isolated mitochondria. Endocr. Res. 22, 505e514.
King, S.R., Liu, Z., Soh, J., Eimerl, S., Orly, J., Stocco, D.M., 1999. Effects of
disruption of the mitochondrial electrochemical gradient on steroidogene-
sis and the Steroidogenic Acute Regulatory (StAR) protein. J. Steroid Bio-
chem. Mol. Biol. 69, 143e154.
Kletsas, D., Li, W., Han, Z., Papadopoulos, V., 2004. Peripheral-type benzo-
diazepine receptor (PBR) and PBR drug ligands in fibroblast and
fibrosarcoma cell proliferation: role of ERK, c-Jun and ligand-activated
PBR-independent pathways. Biochem. Pharmacol. 67, 1927e1932.
Kroemer, G., Zamzami, N., Susin, S.A., 1997. Mitochondrial control of apo-
ptosis. Immunol. Today 18, 44e51.
Lada, A.T., Davis, M., Kent, C., Chapman, J., Tomoda, H., Omura, S.,
Rudel, L.L., 2004. Identification of ACAT1- and ACAT2-specific inhibi-
tors using a novel, cell-based fluorescence assay: individual ACAT unique-
ness. J. Lipid Res. 45, 378e386.
Levin, E., Premkumar, A., Veenman, L., Kugler, W., Leschiner, S., Spanier, I.,
Weisinger, G., Lakomek, M., Weizman, A., Snyder, S.H., Pasternak, G.W.,
Gavish, M., 2005. The peripheral-type benzodiazepine receptor and tumor-
igenicity: isoquinoline binding protein (IBP) antisense knockdown in the
C6 glioma cell line. Biochemistry 44, 9924e9935.
Liu, Y., Peterson, D.A., Kimura, H., Schubert, D., 1997. Mechanism of cellular
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) re-
duction. J. Neurochem. 69, 581e593.
Liu, Y., Peterson, D.A., Schubert, D., 1998. Amyloid beta peptide alters intra-
cellular vesicle trafficking and cholesterol homeostasis. Proc. Natl. Acad.
Sci. U.S.A. 95, 13266e13271.
McEnery, M.W., Snowman, A.M., Trifiletti, R.R., Snyder, S.H., 1992. Isolation
of the mitochondrial benzodiazepine receptor: association with the volt-
age-dependent anion channel and the adenine nucleotide carrier. Proc.
Natl. Acad. Sci. U.S.A. 89, 3170e3174.
McGookey, D.J., Anderson, R.G., 1983. Morphological characterization of the
cholesteryl ester cycle in cultured mouse macrophage foam cells. J. Cell
Biol. 97, 1156e1168.
Miller, W.L., 1988. Molecular biology of steroid hormone synthesis. Endocr.
Rev. 9, 295e318.
Moreno-Sanchez, R., Hogue, B.A., Bravo, C., Newman, A.H., Basile, A.S.,
Chiang, P.K., 1991. Inhibition of substrate oxidation in mitochondria by
the peripheral-type benzodiazepine receptor ligand AHN 086. Biochem.
Pharmacol. 41, 1479e1484.
Papadopoulos, V., 2004. In search of the function of the peripheral-type ben-
zodiazepine receptor. Endocr. Res. 30, 677e684.
Papadopoulos, V., Baraldi, M., Guilarte, T.R., Knudsen, T.B., Lacapere, J.-J.,
Lindemann, P., Norenberg, M.D., Nutt, D., Weizman, A., Zhang, M.-R.,
Gavish, M., 2006. Translocator protein (18 kDa): new nomenclature for
the peripheral-type benzodiazepine receptor based on its structure and
molecular function. Trends Pharmacol. Sci. 27, 402e409.
Petrescu, A.D., Gallegos, A.M., Okamura, Y., Strauss 3rd, J.F., Schroeder, F.,
2001. Steroidogenic acute regulatory protein binds cholesterol and modu-
lates mitochondrial membrane sterol domain dynamics. J. Biol. Chem.
276, 36970e36982.
Prattes, S., Horl, G., Hammer, A., Blaschitz, A., Graier, W.F., Sattler, W.,
Zechner, R., Steyrer, E., 2000. Intracellular distribution and mobilization
of unesterified cholesterol in adipocytes: triglyceride droplets are sur-
rounded by cholesterol-rich ER-like surface layer structures. J. Cell Sci.
113 (Pt 17), 2977e2989.
Rukmini, R., Rawat, S.S., Biswas, S.C., Chattopadhyay, A., 2001. Cholesterol
organization in membranes at low concentrations: effects of curvature
stress and membrane thickness. Biophys. J. 81, 2122e2134.
Simpson, E.R., Waterman, M.R., 1983. Regulation by ACTH of steroid
hormone biosynthesis in the adrenal cortex. Can. J. Biochem. Cell Biol.
61, 692e707.
Soccio, R.E., Breslow, J.L., 2004. Intracellular cholesterol transport. Arterios-
cler. Thromb. Vasc. Biol. 24, 1150e1160.
Veenman, L., Gavish, M., 2006. The peripheral-type benzodiazepine receptor
and the cardiovascular system. Implications for drug development. Phar-
macol. Ther. 110, 503e524.
Watari, H., Arakane, F., Moog-Lutz, C., Kallen, C.B., Tomasetto, C.,
Gerton, G.L., Rio, M.C., Baker, M.E., Strauss 3rd, J.F., 1997. MLN64 con-
tains a domain with homology to the steroidogenic acute regulatory protein
(StAR) that stimulates steroidogenesis. Proc. Natl. Acad. Sci. U.S.A. 94,
8462e8467.
Weisinger, G., Kelly-Hershkovitz, E., Veenman, L., Spanier, I., Leschiner, S.,
Gavish, M., 2004. Peripheral benzodiazepine receptor antisense knockout
increases tumorigenicity of MA-10 Leydig cells in vivo and in vitro.
Biochemistry 43, 12315e12321.









![The translocator protein ligand [18F]DPA-714 images glioma and activated microglia in vivo](https://img.dokumen.tips/doc/110x75/6346d69f391b5ca53f0d32de/the-translocator-protein-ligand-18fdpa-714-images-glioma-and-activated-microglia.jpg)



















