Embed Size (px)
Citation preview

106
Biochimica et Biophysica Acta, 437 (1976) 106--115 © Elsevier Scientific Publishing Company, Amsterdam -- Printed in The Netherlands
BBA 27936
INTERACTION BETWEEN DERMATAN SULPHATE CHAINS
I. AFFINITY CHROMATOGRAPHY OF COPOLYMERIC GALACTOS- AMINOGLYCANS ON DERMATAN SULPHATE-SUBSTITUTED AGAROSE
L.-)k. FRANSSON
Department o f Physiological Chemistry 2, University o f Lund, P.O. Box 750, S-220 07 Lund (Sweden)
(Received December 24th, 1975)
Summary
(1) Binding of copolymeric as well as homopolymeric galactosaminoglycans to dermatan sulphate-substituted gels has been demonstrated. Material bound in the presence of 0.15 M NaC1 was eluted with either 1 M urea, 0.5 M guani- dine • HC1 or 0.5 M NaC1. Homopolymeric galactosaminoglycans were also dis- placed by 0.5 M sodium acetate. The interaction was not dependent on di- valent cations.
(2) Dermatan sulphate has been fractionated into aggregating and non- aggregating species by gel chromatography in the presence of 0.5 M sodium acetate. In the presence of 0.1 M sodium acetate or 0.5 M guanidine • HC1 no aggregation was observed.
(3) Crosslinks formed during periodate oxidation at physiological ionic strength have been ascribed to chain-chain interaction.
(4) Chondroitin 4-sulphate, heparan sulphate and heparin also showed in- teraction with gels substituted with copolymeric galactosaminoglycans, while chondroitin 6-st, lphate, hyaluronate and keratan sulphate did not.
(5) Binding of copolymeric galactosaminoglycan chains to dermatan sul- phate- or chondroitin sulphate-substituted gels was most pronounced when the copolymeric chains contained similar proportions of L-iduronic and D-glucu- ronic acid.
I n t r o d u c t i o n
A number of natural glycans form gels by co-operative interaction between individual chains. Such glycans constitute extraneous coats in bacteria or extra-

1 0 7
cellular matrices in algae and higher plants; for references see ref. 1. The pri- mary structure of gel-forming glycans is characterized by a 'masked repeating structure', i.e. the regular repeating unit (monosaccharide or disaccharide) is modified or replaced by other types of building blocks to a variable extent.
In connective tissues of higher animals the extracellular space is composed of fibrous elements imbedded in an amorphous gel matrix. This matrix contains polyanionic polysaccharide-protein conjugates (proteoglycans). These macro- molecules have certain functions like water immobilization, ion binding and exclusion phenomena. Further, by interaction with the fibrous elements, they are considered to be of great importance for the general appearance of a partic- ular connective tissue.
In cartilage, chondroitin sulphate proteoglycans interact with hyaluronate [2] to form large aggregates [3,4], which are held together by noncovalent bonds between the protein portion of proteochondroitin sulphate and t h e hyaluronate chain [5]. In fibrous connective tissue dermatan sulphate proteo- glycans are characteristic components of the intercellular matrix [6]. Proteo- dermatan sulphate has been isolated from heart valves [7], skin [8] and sclera [9] but little information concerning its structure is available. Proteodermatan sulphate shows unusually strong interaction with collagen [10], which appears to be mediated by the dermatan sulphate side-chains [11]. Dermatan sulphate* has a 'masked repeating structure', i.e. the principal repeating unit L-iduronic acid-GalNAc-SO4 is to a variable degree replaced by the chondroitin sulphate repeating unit glucuronic acid-GalNAc-SO4 [12,13].
During periodate oxidation of dermatan sulphate-chondroitin sulphate copolymers in the presence of 0.2 M NaC104, hemiacetal-type crosslinkages were formed between oxidized and unoxidized uronic acid residues in separate chains [14]. This observation suggested the presence of contact between co- polymeric chains. The main purpose of the present report is to demonstrate binding of free dermatan sulphate chains to dermatan sulphate-substituted agarose.
Experimental
Materials Chondroitin 4-sulphate from beef nasal septum was the same preparation as
described previously [15]. Chondroitin 6-sulphate from human nucleus pulp- osus was a gift from Dr. C.A. Antonopoulos, University of Patras, Patras, Greece. Beef lung dermatan sulphate (~-heparin) and a preparation of heparan sulphate from pig intestinal mucosa were gifts from Dr. J.A. Cifonelli, Univer- sity of Chicago, Chigaco, U.S.A. Pig skin dermatan sulphate was obtained after solubilization of the tissue by papain [16] or trypsin [17]. The galactosamino- glycans were further fractionated by ethanol (25 and 50% ethanol fractions
* D e r m a t a n su lpha te d e n o t e s a p o l y m e r where L-iduronic ac id -GaiNAc-S04 units comprise the m a j o r i t y o f the repeat ing periods. C h o n d r o i t i n su lpha te con ta ins only g lucuronic ac id-GaiNAc- (4 or 6)-SO4 periods. D e r m a t a n su lpha t e - chondro i t i n su lpha te c o p o l y m e r s wi th L- iduronie acid/ glucuron ic acid ra t ios approaching one m a y be referred to s imply as c o p o l y m e r i c ga iac tosamino- glycans. In this c o n t e x t , a h o m o p o l y m e r denote s a g lycan c o m p o s e d solely of e i the r L- iduronic ac id-conta in ing or g lucuron ic ac id-conta in ing repeat periods.

108
were obtained) [16]. Dermatan sulphate preparations from beef, horse or hu- man aorta were prepared in the same manner [18]. Galactosaminoglycans pre- cipitated at 25, 35 and 50% ethanol were isolated. A crude heparin preparation was obtained from AB Vitrum, Stockholm, Sweden, and purified by repeated precipitation with cetylpyridinium chloride [19]. Hyaluronic acid and keratan sulphate were prepared from pig skin [20] and human nucleus pulposus [21], respectively. Cartilage proteoglycan (monomer) was a gift from Dr. Sven Inerot of this laboratory. Chondroitinase-ABC was the same preparation as described previously [15].
Sepharose 6B, regular as well as epoxy-activated or crosslinked (CL), AH- Sepharose 4B and CH-Sepharose 4B were purchased from Pharmacia Fine Chemicals, Uppsala, Sweden. Other chemicals were of reagent grade.
Preparation of glycan-substituted agarose Three different procedures were utilized for the at tachment of various poly-
saccharides to agarose. Galactosaminoglycans were coupled to epoxy-activated Sepharose 6B (Method A), and dermatan sulphate prepared from pig skin after trypsin digestion was attached to the six-carbon, spacer group of CH-Sepharose 4B by a cabodiimide procedure (Method B), both according to the description of the manufacturer. Glucosamine was used as the blocking group with the latter gel. After partial oxidation with periodate, dermatan sulphate was coupled to the amino group of AH-Sepharose 4B via an aldimine which was subsequently stabilized by reduct ion (Method C). Samples of the polymer (50--100 mg) were oxidized for 10 or 20 min, according to procedures de- scribed elsewhere (in 0.05 M citrate buffer, pH 3.0 [14] }. After dialysis against distilled water the retentate was mixed with I g of swollen, washed gel (final volume 10 ml) and pH was kept between 12 and 13 by addition of 2 M NaOH for 1 h. Reduct ion was performed by treatment with NaBH4 (250=500 mg) for 1 h at room temperature. Subsequently, the gel was washed 3 times with water. In order to substitute free amino groups the gel was treated with a 5-fold molar excess of glyoxylic acid at pH 12--13 for 30 min followed by reduction with a 10-fold molar excess of borohydride. This procedure was repeated twice and the gel was finally washed with 2 M NaC1 followed by water.
Methods
Hexuronic acids were determined by the manual carbazole [22] and orcinol [23] methods. The former method was also employed in an automated proce- dure [24]. Hexose was estimated by the anthrone method as modified by Goa [25]. The galactosaminoglycan content of substi tuted agarose was determined by digestion with chondroitinase-ABC according to the procedure of Yamagata et al. [26]. Lyophilized samples of the gel were incubated according to Assay 3 [26], centrifuged and the absorbance at 232 nm was recorded against a sub- strate blank. The amount of glycan bound was calculated from a standard curve of dermatan sulphate assayed by the same procedure. The nonsulphated L- iduronic acid residues of dermatan sulphate-chondroitin sulphate copolymers were selectively oxidized by periodate as described elsewhere [14].

109
Affinity chromatography The gel was suspended in starting buffer, deaerated and columns (6 mm × 50
mm) were packed. Samples were applied dissolved in the starting buffer and elution was performed either stepwise or with a linear gradient. In the latter case the total elution volume was 20 ml. Fractions (1 ml) were analyzed for hexuronic acid or hexose as described above. Further details are described in the legends to the figures. All glycosaminoglycan solutions were freshly made if not stated otherwise.
Results
Correlation between interaction and gel chromatographic behaviour Homopolymeric and copolymeric galactosaminoglycans were subjected to
affinity chromatography on agarose substi tuted with pig skin dermatan sul- phate. As shown in Fig. 1, binding of pig skin dermatan sulphate to this matrix could be demonstrated (A). Bound material was displaced by 1 M urea, 0.5 M guanidine • HC1 or 0.5 M NaC1. In the presence of 0.15 M NaC1 (A and B) the capacity of the gel was approximately 0.5 mg (approx. 15% of the bound dermatan sulphate)*. A port ion of the bound material (approx. 25%) could be eluted with 0.5 M sodium acetate (B). The two homopolymers (beef lung dermatan sulphate and chondroitin 4-sulphate) showed little or no binding to dermatan sulphate-substituted agarose in the presence of 0.5 M sodium acetate (C and D). It should be added that, in the presence of 0.15 M NaCI, beef lung dermatan sulphate (0.2 mg) was bound to the gel. However, the bound material was quantitatively eluted with 0.5 M sodium acetate. No binding of any glycan was observed to a blank column (substi tuted only with glyoxalate).
In order to investigate possible aggregation of copolymeric chains galactos- aminoglycans were chromatographed on crosslinked Sepharose (CL6B) in either 0.5 M sodium acetate or 0.5 M guanidine. HC1 (Fig. 2). Whereas the homopolymers (beef lung dermatan sulphate and chondroitin 4-sulphate) were eluted in the same position in both solvents (A and D; C and F), copolymeric pig skin dermatan sulphate tended to be less retarded in the presence of 0.5 M sodium acetate (B and E). However, in the presence of 0.1 M sodium acetate the elution positions were the same as in 0.5 M guanidine • HCI. Pig skin derma- tan sulphate was subfractionated by gel chromatography into Fractions I and II (Fig. 3). These subfractions were also subjected to affinity chromatography on dermatan sulphate-substituted agarose (Fig. 4). Fraction I was firmly bound to gel in the presence of 0.5 M sodium acetate (A), while Fraction II was not bound bu t slightly retarded (B).
As shown previously [14], periodate oxidation of dermatan sulphate per- formed in the presence of 0.2 M NaC104 yields hemiacetal-type crosslinkages between oxidized and unoxidized uronic acid residues in separate chains. When the oxidation is performed at lower ionic strength, however, these crosslinkages
* The m a j o r p o r t i o n (85%) o f the d e r m a t a n su lpha t e l igand w a s apparent ly n o t access ib le for b inding . This can be pa r t ly e x p l a i n e d b y s ter ic e f f e c t s ; the l igands m a y be present w i t h i n the gel m a t r i x to a cons iderab le e x t e n t . Moreover , l igands w i t h c o m p l e m e n t a r y b inding z o n e s m i g h t bind to o ne another . Thus , on ly a f r a c t i o n o f the l igand chains , i .e. tho se that are present on t he e x t e r i o r o f the gel part ic les and have n o t m a d e c o n t a c t w i t h o th e r l igands , are avai lable for binding.
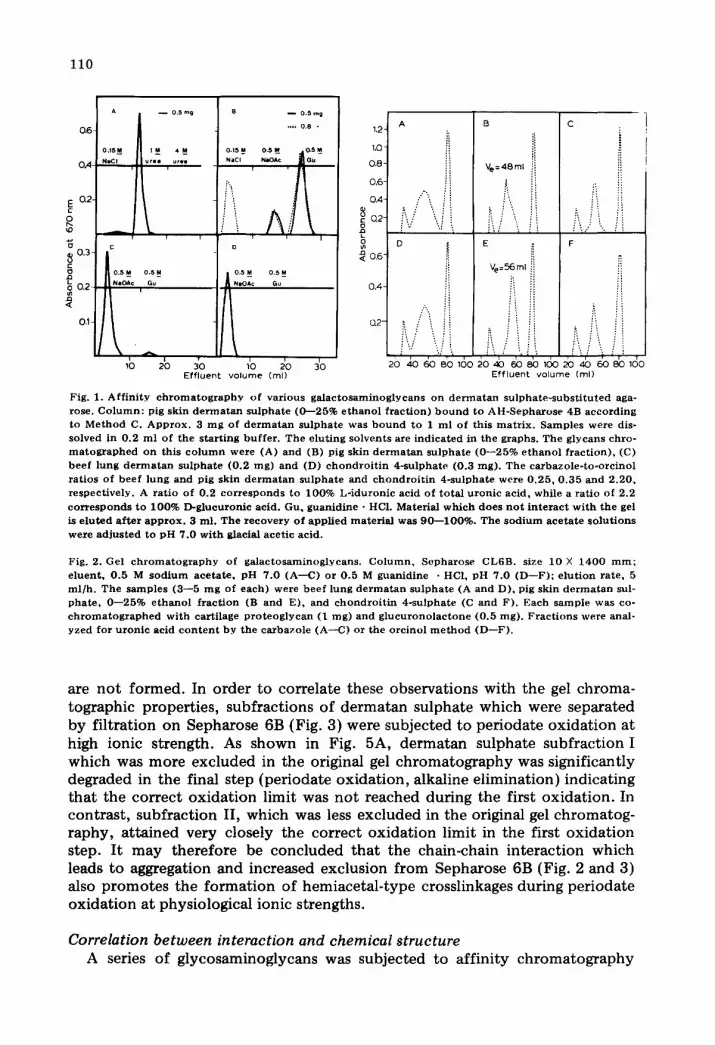
110
O&
0A-
0,2-
~o
~ o.~ ,<
03
o.m~ N a C l
B 0 . 5 mg
.. . . 0 .8 .
~_M 4M_ 0.ram O.SM_ l ~ s . l u r e a urea N I C I FMOA¢ GU -- ||
' 'It F I t I I
O
0.5 M 0 , 5 _M
N i O A c Gu
t I , I i I
10 20 30 10 20 30 Effluent volume (ml)
o.s~_ O.S_M N I O A ¢ Gu
t
1,2
1.O
).8"
),6"
).4'
D.2"
),6
).4-
).2-
A
i!
/'"...,..,i!
B C
Ve=48ml i
i i i i i i...i L i i D ~ E ~- F
"
2o ~o ~ Bb ~6o ~o ~ @o 8b ~6o 2b 4b go 8b ~do Effluent volume (ml)
Fig. 1. Aff in i ty c h r o m a t o g r a p h y of var ious ga l ac tosaminog lycans on d e r m a t a n su lpha te - subs t i tu ted aga- rose. C o l u m n : pig skin d e r m a t a n su lpha te ((}---25% e thano l f rac t ion) b o u n d to AH-Sepha rose 4B accord ing to Me thod C. A p p r o x . 3 m g of d e r m a t a n su lpha te was b o u n d to 1 ml of this ma t r i x . Samples were dis- solved in 0.2 m l of the s tar t ing buf fe r . The e lut ing solvents are ind ica ted in the graphs . The glycans chro- m a t o g r a p h e d on this c o l u m n were (A) and (B) pig skin d e r m a t a n su lphate (0 - -25% e th an o l f rac t ion) , (C) beef lung d e r m a t a n su lpha te (0 .2 mg) and (D) c h o n d r o i t i n 4-sulphate (0.3 rag). Th e carbazole - to-orc ino l ra t ios of beef lung and pig skin d e r m a t a n su lpha te and c hon d ro i t i n 4-sulphate were 0 .25 , 0 .35 and 2 .20 , respect ive ly . A ra t io of 0.2 co r r e sponds to 100% L- iduronic acid of to ta l u ron ic acid , while a ra t io of 2.2 co r r e s ponds to 100% D-glucuronic acid. Gu, guanid ine • HCI. Material wh ich does n o t i n t e r ac t w i th the gel is e lu ted a f t e r a p p r o x . 3 ml . The r e c o v e r y of appl ied ma te r i a l was 90 - -100%. The s o d i u m ace ta t e so lu t ions were ad jus t ed to p H 7.0 wi th glacial acet ic acid.
Fig. 2. Gel c h r o m a t o g r a p h y of ga lac tosaminoglycans . C o l u m n , Sepharose CL6B. size 10 X 1400 m m ; e luent , 0 .5 M s o d i u m ace ta te , p H 7.0 (A--C) or 0.5 M guanidine • HCI, pH 7.0 (D--F) ; e lu t ion ra te , 5 ml /h . The samples (3 - -5 m g of each) were bee f lung d e r m a t a n su lpha te (A and D), pig skin d e r m a t a n sul- pha te , 0 - -25% e thano l f rac t ion (B and E), and c hond ro i t i n 4-sulphate (C and F). Each sample was co- c h r o m a t o g z a p h e d wi th cart i lage p r o t e o g l y c a n (1 mg) and g l u c u r o n o l a c t o n e (0 .5 rag). F rac t ions were anal- yzed for u ron ic acid c o n t e n t by the carbazole (A--C) or the orc inol m e t h o d (D--F) .
are not formed. In order to correlate these observations with the gel chroma- tographic properties, subfractions of dermatan sulphate which were separated by filtration on Sepharose 6B (Fig. 3) were subjected to periodate oxidation at high ionic strength. As shown in Fig. 5A, dermatan sulphate subfraction I which was more excluded in the original gel chromatography was significantly degraded in the final step (periodate oxidation, alkaline elimination) indicating that the correct oxidation limit was not reached during the first oxidation. In contrast, subfraction II, which was less excluded in the original gel chromatog- raphy, attained very closely the correct oxidation limit in the first oxidation step. It may therefore be concluded that the chain-chain interaction which leads to aggregation and increased exclusion from Sepharose 6B (Fig. 2 and 3) also promotes the formation of hemiacetal-type crosslinkages during periodate oxidation at physiological ionic strengths.
Correlation between interaction and chemical structure A series of glycosaminoglycans was subjected to affinity chromatography
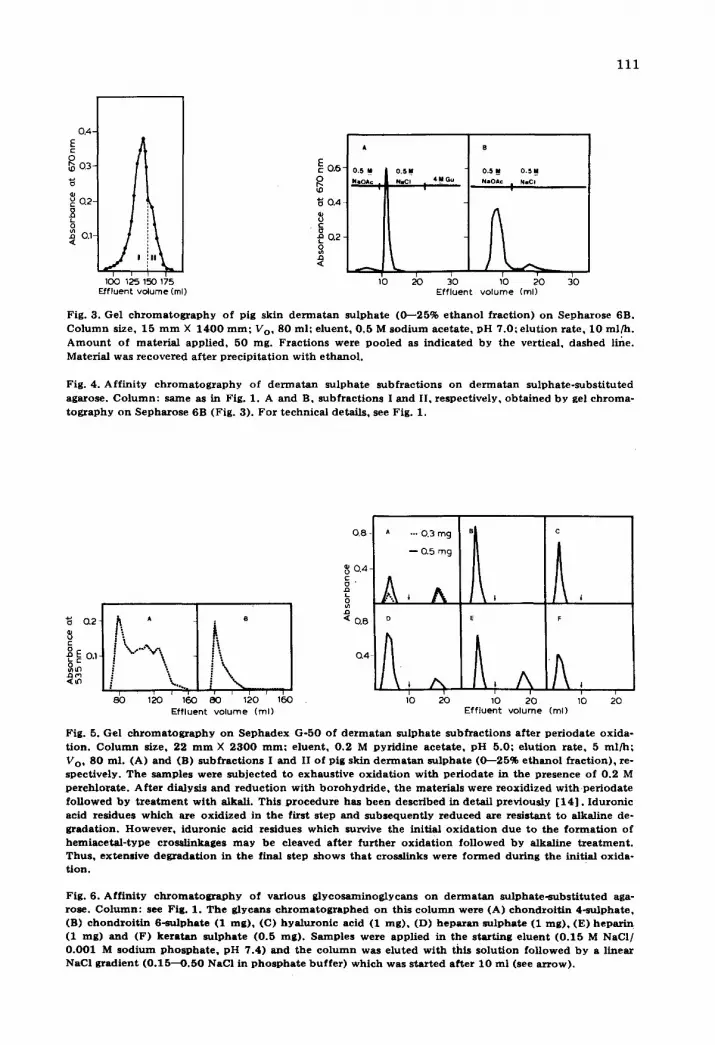
111
0 . 4 -
E
8 a2-
~a 0.1- <
1 1 ~ i i 125 150 t75
Eff luent Volume (ml)
Eco,6- o ~p
0.4
o : 0.2
o
k
o,5 ~ | o .sN N~mOA¢ . S NalCI 4 M G u
I -
o.5_m o .su NIOA¢ | NmCl
| I 10 20 30 10 20 30
Ef f luent vo lume (ml)
Fig. 3. Gel c h r o m a t o g r a p h y o f p ig sk in d e r m a t a n s u l p h a t e ( 0 - - 2 5 % e t h a n o l f r a c t i o n ) o n S e p h a r o s e 6B. C o l u m n size, 1 5 m m X 1 4 0 0 r a m ; Vo, 8 0 ml ; e l u e n t , 0 . 5 M s o d i u m a c e t a t e , p H 7 .0 ; e l u t i o n r a t e , 1 0 m l / h . A m o u n t o f m a t e r i a l a p p l i e d , 5 0 rag . F r a c t i o n s we re p o o l e d as i n d i c a t e d b y the ver t i ca l , d a s h e d l ine. Mate r ia l was r e c o v e r e d a f t e r p r e c i p i t a t i o n w i t h e t h a n o l .
Fig. 4. A f f i n i t y c h r o m a t o g r a p h y o f d e r m a t a n s u l p h a t e s u b f r a c t i o n s o n d e r m a t a n s u l p h a t e - s u b s t i t u t e d aga rose . C o l u m n : s ame as in Fig. 1. A a n d B, s u b f r a c t i o n s I a n d II, r e spec t i ve ly , o b t a i n e d b y gel c h r o m a - t o g r a p h y o n S e p h a r o s e 6B (Fig . 3) . F o r t e c h n i c a l de ta i l s , see Fig . 1.
t . B
\
0.8 - A ... 0.3 m 9
- - 0.5 m 9
0 4 -
~ 0.8 D
0.4 ~ . /N
10 20 8o 120 leo 80 120' 1~0 - L 2o Eff luent vo lume (m l )
/ c i t E F
i/A 10 20 10
E f f luen t vo lume (ml )
Fig. 5. Gel c h r o m a t o g r a p h y o n S e p h a d e x G - 5 0 o f d e r m a t a n s u l p h a t e s u b f r a c t i o n s a f t e r p e r i o d a t e o x i d a - t i on . C o l u m n size, 22 m m X 2 3 0 0 r a m ; e l u e n t , 0 . 2 M p y r i d i n e a c e t a t e , p H 5 .0 ; e l u t i o n r a t e , 5 m l / h ; Vo , 8 0 ml . (A) a n d (B) s u b f r a c t i o n s I a n d II o f p ig sk in d e r m a t a n s u l p h a t e (0---25% e t h a n o l f r a c t i o n ) , re- spec t ive ly . T h e s a m p l e s we re s u b j e c t e d t o e x h a u s t i v e o x i d a t i o n w i t h p e r i o d a t e in t h e p r e s e n c e o f 0 .2 M p e r c h l o r a t e . A f t e r d ia lys i s a n d r e d u c t i o n w i t h b o r o h y d r i d e , t h e m a t e r i a l s we re r e o x i d i z e d w i t h p e r i o d a t e f o l l o w e d b y t r e a t m e n t w i t h a lkal i . Th i s p r o c e d u r e h a s b e e n d e s c r i b e d in de t a i l p r e v i o u s l y [ 1 4 ] . I d u r o n i c ac id r e s i d u e s w h i c h a re o x i d i z e d in t h e f i rs t s tep a n d s u b s e q u e n t i y r e d u c e d are r e s i s t an t t o a lka l ine de- g r a d a t i o n . H o w e v e r , i d u r o n i c ac id r e s idues w h i c h survive the in i t ia l o x i d a t i o n d u e t o t he f o r m a t i o n o f h e m i a c e t a l - t y p e c ro s s i i nkages m a y b e c l eaved a f t e r f u r t h e r o x i d a t i o n f o l l o w e d b y a lka l ine t r e a t m e n t . T h u s , ex t ens ive d e g r a d a t i o n in t h e f ina l s t ep s h o w s t h a t c ross l inks we re f o r m e d d u r i n g the in i t i a l o x i d a - t i o n .
Fig . 6 . A f f i n i t y c h r o m a t o g r a p h y o f v a r i o u s g l y c o s a m i n o g l y c a n s o n d e r m a t a n s u l p h a t e - s u b s t i t u t e d aga- rose . C o l u m n : see Fig. 1. T h e g l y c a n s c h r o m a t o g r a p h e d o n th i s c o l u m n w e r e (A) c h o n d r o i t i n 4 - s u l p h a t e , (B) c h o n d r o i t i n 6 - s u l p h a t e (1 m g ) , (C) h y a l u r o n i c ac id (1 m g ) , (D) h e p a r a n s u l p h a t e (1 r a g ) , (E) h e p a r i n (1 rag) a n d (F) k e r a t a n s u l p h a t e (0 .5 m g ) . S a m p l e s we re a p p l i e d in t he s t a r t i n g e l u e n t ( 0 . 1 5 M NaCI / 0 . 0 0 1 M s o d i u m p h o s p h a t e , p H 7 .4 ) a n d t h e c o l u m n was e l u t e d w i t h th i s s o l u t i o n f o l l o w e d b y a l i nea r NaCI g r a d i e n t ( 0 . 1 5 - - O . 5 0 NaCI in p h o s p h a t e b u f f e r ) w h i c h w a s s t a r t e d a f t e r 1 0 m l (see a r r o w ) .

112
0 4 A •
®o.~ i i
10 20 10 20 10 20 10 20 10 20 E f f l uen t v o l u m e (ml)
Fig. 7. Aff in i ty c h r o m a t o g r a p h y of var ious ga l ac tosaminog lycan f rac t ions on de rvaa tan sulphate-subst i - t u t ed agarose. C o l u m n : pig skin d e r m a t a n su lpha te (0 - -25% e thano l f rac t ion) b o u n d to CH~Sepharose 4B accord ing to Me thod B. A p p r o x . 2 m g of d e r m a t a n su lpha te were b o u n d to 1 ml of this ma t r ix . The g lycans (0 .5- -1 m g dissolved in 0.2 ml of s ta r t ing bu f fe r ) c h r o m a t o g r a p h e d on this c o l u m n were (A) pig skin d e r m a t a n su lpha te (0 - -25% e thano l f rac t ion) , (B) bee f lung d e r m a t a n su lpha te , (C--E) subfrac- t ions 0- -25%, 25 - -35% and 35- -50% e thano l of horse ao r t a d e r m a t a n su lpha te , respect ive ly . The car- bazole - to-orc ino l ra t ios of the l a t t e r subf rae t ions were 0 .47 , 1.21 and 1.80, respect ively . Elu t ion was p e r f o r m e d wi th a linea~ C1- g rad ien t as descr ibed in the legend to Fig. 6. Solid l ine, samples appl ied di rect - ly: d o t t e d line, samples appl ied a f t e r s t and ing a t r o o m t e m p e r a t u r e for 48 h.
on dermatan sulphate-substituted agarose in the presence of 0.15 M NaCl. As shown in Fig. 6 chondroitin 4-sulphate, heparan sulphate and heparin showed interaction under these conditions, while chondroitin 6-sulphate, hyaluronate and keratan sulphate did not. The capacity for binding dermatan sulphate was more than twice that of chondroitin 4-sulphate, heparin and heparan sulphate (cf. Figs. 1A and 6A). No binding was observed when nonsubsti tuted gel was used.
In the present experimental design binding of free chains to one another should compete with binding of free chains to the matrix-bound material. Indeed, when a solution of copolymeric dermatan sulphate was kept at room temperature the capacity for binding to dermatan sulphate-substituted gel gradually declined (see Fig. 7A). In contrast, solutions of the homopolymers beef lung dermatan sulphate (Fig. 7B) or chondroitin 4-sulphate were not af- fected in the same manner. Interactions between dermatan sulphate chains (either between free chains or free and bound chains) apparently depend on the presence of both L-iduronic and glucuronic acids in one of the chains. (Heterologous interactions between dermatan sulphate and heparan sulphate might similarly depend on the presence of the two epimeric hexuronic acids.) This was further illustrated by the finding that copolymers with L-iduronic/ glucuronic acid ratios approaching unity were bound to a larger extent than were the homopolymers (Fig. 7 B--E, cf. also Fig. 6A). Similar results were obtained when various copolymers were chromatographed on chondroitin 4- sulphate-substituted agarose (Fig. 8). Whereas neither beef lung dermatan sul- phate (A) nor chondroitin 4-sulphate (F) showed binding to this matrix, co- polymeric galactosamlnoglycans (B--E) were bound to some extent. Binding of horse aorta dermatan sulphate (subfraction 25--35% ethanol) to chondroitin 4-sulphate-substituted agarose was markedly lower as compared with binding to dermatan sulphate gels (cf. Figs. 7D and 8D). The same results were ob- tained when these samples were chromatographed on a column of agarose sub- sti tuted with beef lung dermatan sulphate (homopolymer with L-iduronic acid as the uronic acid component) .
Moreover, binding between chondroitin 4-sulphate and dermatan sulphate
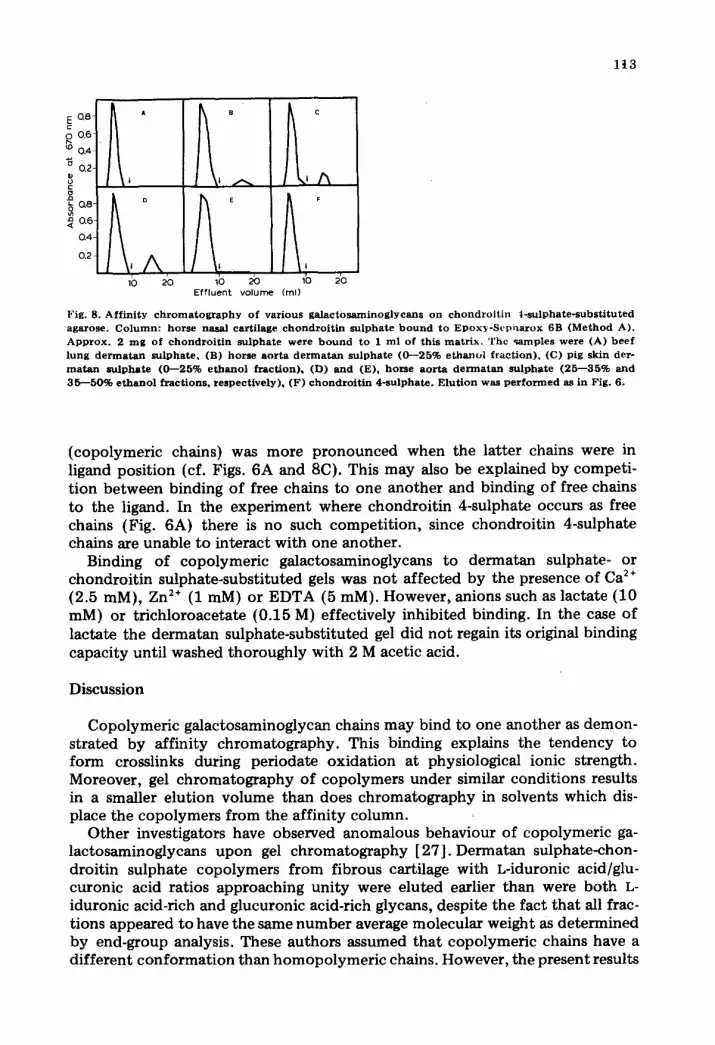
l i3
LL 0.8 a s c
o 0.6-
o.4 0.2
c
'~ 0~8 D E F
0.4
0,2
10 20 10 20 10 2O Effluent. volume (ml)
Fig. 8. Aff in i ty c h r o m a t o g r a p h y o f var ious ga l ac tosaminog lyeans o n c h o n d r o i t i n t - su lpha te - subs t i tu ted agarose. C o l um n : horse nasal cart i lage c h o n d r o i t i n s u l p h a t e b o u n d to Epox?c-Scpharox 6B (Method A). A p p r o x . 2 m g o f c h o n d r o i t i n su lphate w e r e b o u n d to I ml of this ma t r ix . The ~amples were (A) b e e f lung d e r m a t a n su lpha te , (B) h o r s e aor ta d e n n a t a n su lpha te (0 - -25% e thano l f rac t ion) , (C) pig skin dero m a t a n su lpha te (0 - -25% e tha no l f rac t ion) , (D) a nd (E), horse a o r t a de r rna t an su lpha te (25 - -35% and 35 - -50% e t h a n o l f rac t ions , respec t ive ly) , (F) c h o n d r o i t i n 4-sulphate . E lu t ion was p e r f o r m e d as in Fig. 6.
(copolymeric chains) was more pronounced when the latter chains were in ligand position (cf. Figs. 6A and 8C). This may also be explained by competi- t ion between binding of free chains to one another and binding of free chains to the ligand. In the experiment where chondroitin 4-sulphate occurs as free chains (Fig. 6A) there is no such competi t ion, since chondroitin 4-sulphate chains are unable to interact with one another.
Binding of copolymeric galactosaminoglycans to dermatan sulphate- or chondroitin sulphate-substituted gels was not affected by the presence of Ca 2 ÷ (2.5 mM), Zn 2÷ (1 raM) or EDTA (5 mM). However, anions such as lactate (10 mM) or trichloroacetate (0.15 M) effectively inhibited binding. In the case of lactate the dermatan sulphate-substituted gel did not regain its original binding capacity until washed thoroughly with 2 M acetic acid.
Discussion
Copolymeric galactosaminoglycan chains may bind to one another as demon- strated by affinity chromatography. This binding explains the tendency to form crosslinks during periodate oxidation at physiological ionic strength. Moreover, gel chromatography of copolymers under similar conditions results in a smaller elution volume than does chromatography in solvents which dis- place the copolymers from the affinity column.
Other investigators have observed anomalous behaviour of copolymeric ga- lactosaminoglycans upon gel chromatography [27]. Dermatan sulphate-chon- droitin sulphate copolymers from fibrous cartilage with L-iduronic acid/glu- curonic acid ratios approaching unity were eluted earlier than were both L- iduronic acid-rich and glucuronic acid-rich glycans, despite the fact that all frac- tions appeared to have the same number average molecular weight as determined by end-group analysis. These authors assumed that copolymeric chains have a different conformat ion than homopolymeric chains. However, the present results

1 1 4
show that chain-chain interaction leading to aggregation is a more plausible explanation.
Since guanidine, urea and trichloroacetate prevented binding between co- polymeric chains it may be suggested that both hydrogen bonding and hydro- phobic interactions play a role in this type of binding.
The presence of both glucuronic acid-GalNAc-SO4 and L-iduronic acid- GalNAc-SO4 repeat periods in the same chain are essential for interaction to occur. However, copolymeric chains interact with homopolymeric chains in the presence of 0.15 M NaC1, but not in 0.5 M sodium acetate. In the latter solvent only copolymeric chains interact. The highest degree of association is ob- served in a solution of copolymeric chains containing nearly equimolar propor- tions of the epimeric uronic acids. It is of interest to note that chondroitin 6-sulphate showed no binding to dermatan sulphate. Thus, only the 4-sulphated regions of copolymeric chains may be expected to contain the binding zones. In these regions glucuronic acid- and L-iduronic acid-containing units may be arranged in clusters or in an alternating fashion [28] . In a succeeding report the copolymeric sequences of aggregating and non-aggregating chains are com- pared.
In a recent s tudy on the formation and excretion of copolymeric galactos- aminoglycans by fibroblasts in culture [29] it was observed that alternating se- quences were particularly prominent in copolymers isolated from the cell layer after trypsin digestion. The presence of copolymers with these features in the pericellular pool may be partly explained by selective aggregation of proteo- dermatan sulphate monomers. The present s tudy also demonstrated binding of heparan sulphate and heparin to copolymeric galactosaminoglycans. Since heparan sulphate is associated with the cell surface [30] , th is phenomenon may be an additional factor in concentrating certain proteodermatan sulphate species to the immediate vicinity of the cell.
The chain-chain interactions described in this report may be one reason for the pronounced heterogeneity observed when proteodermatan sulphates are subjected to gel chromatography [8,9,31]. Investigations on this matter are currently being performed in this laboratory.
Acknowledgements
The author is indebted to Drs. I. Carlstedt, L. CSster, A. MalmstrSm, M. Paulsson and I. Sj5berg for many stimulating discussions and to Mrs. Birgitta Havsmark for expert technical assistance. Grants from: Swedish Medical Re- search Council (B72-13X-139-08B), Gustaf V's 80-~rsfond, and the Medical faculty, University of Lund.
References
1 Rees , D .A . ( 1 9 7 2 ) B i o c h e m . J. 1 2 6 , 2 5 7 - - 2 7 3 2 H a r d i n g h a m , T .E . a n d Mui r , H. ( 1 9 7 2 ) B i o c h i m . B i o p h y s . A c t a 2 7 9 , 4 0 1 - - 4 0 5 3 H a r d i n g h a m , T .E . a n d Mui r , H. ( 1 9 7 4 ) B i o c h e m . J . 139 , 5 6 5 - - 5 8 1 4 Hasca l l , V .C . a n d He ineg~rd , D. ( 1 9 7 4 ) J . Biol . C h e m . 2 4 9 , 4 2 3 2 - - 4 2 4 1 5 He ineg~rd , D. a n d Hasca l l , V.C. ( 1 9 7 4 ) J . Biol . C h e m . 2 4 9 , 4 2 5 0 - - 4 2 5 6 6 F r a n s s o n , L. -A. ( 1 9 7 0 ) In Chemis try and Molecular Bio logy o f the I n t e r c e l l u l a r M a t r i x (Balazs , E .A . ,
ed . ) , pp . 8 2 3 - - 8 4 2 , A c a d e m i c Press , L o n d o n

1 1 5
7 Toole, B.P. and Lowther, D.A. (1965) Biochim. Biophys. Acta 101 ,364- -366 8 0 b r i n k , B. (1972) Biochim. Biophys. Acta 264, 354--361 9 C~ster, L. and Fransson, L.--~. (1972) Scand. J. Clin. Lab. Invest. 29, suppl. 123, 9--
10 Toole, B.P. and Lowther, D.A. (1968) Arch. Biochem. Biophys. 128, 567--578 11 Obrink, B. (1973) Eur. J. Biochem. 33, 387--400 12 Fransson, L.-A. and Rod in , L. (1967) J. Biol. Chem. 242, 4161---4169 13 Fransson, L.-A. and Havsmark, B. (1970) J. Biol. Chem. 245, 4 7 7 0 - 4 7 8 3 14 Fransson, L.-A. (1974) Carbohydr. Res. 36, 339--348 15 Fransson, L.-A. and MalmstrSm, A. (1971) Eur. J. Biochem. 18, 422--439 16 Fransson, L.-A. (1968) Bioehim. Biophys. Acta 156 ,311 - -316 17 Stern, E.L., Cifonelli, J.A., Fransson, L.-A., Lindahl, B., Rod in , L., Schiller, S. and Spach, M.L.
(1968) Arkiv Kemi 39, 583--591 18 Fransson, L.-A., Anseth, A., Antonopoulos , C.A. and Gardell, S. (1970) Carbohydr. Res. 15, 73--89 19 Lindahl, U., Cifonelli, J.A., Lindahl, B. and Rod in , L. (1965) J. Biol. Chem. 240, 2817--2820 20 Fransson, L.-A. and Rod in , L. (1967) J. Biol. Chem. 242, 4161--4169 21 Antonopoulos , C.A., Fransson, L.-A., Gardell, S. and Heineg~rd, D. (1969) Aeta Chem. Scand. 23,
2616--2620 22 Dische, Z. (1949) J. Biol. Chem. 167 ,189 - -198 23 Brown, A.H. (1946) Arch. Biochem. Biophys. 11 ,269 - -275 24 Heineg~rd, D. (1973) Chem. Scripta 4, 199--201 25 Goa, J. (1955) Scand. J. Clin. Lab. Invest. suppl. 22, 7 26 Yamagata, T., Saito, H., Habuchi, O. and Suzuki, S. (1968) J. Biol. Chem. 243, 1523--1535 27 Habuchi, H., Yamagata, T., Iwata, H. and Suzuki, S. (1973) J. Biol. Chem. 248 ,601 . ~ -6028 28 Fransson, L.-A., C~ster. L., Malmstr~m, A. and Sj~berg, I. (1974) Biochem. J. ! 4 3 , 3 6 9 - - 3 7 8 29 Malmstr~m, A.. Carlstedt, I., Aberg, L. and Fransson, L.-A. (1975) Biochem. J. 15 i , 477---489 30 Kraemer, P.M. (1971) Biochemistry 10, 1437--1451 31 Antonopoulos , C.A., Axelsson, I., H e i n e g ~ l , D. and Garden, S. (1974) Biochim. Biophys. Acta 338,
108--119





























