Embed Size (px)
Citation preview

Influenza-induced, helper-independentCD8� T cell responses use CD40
costimulation at the late phase of theprimary response
Shirley G. K. Seah,*,† Jamie L. Brady,* Emma M. Carrington,* Wy Ching Ng,‡
Robyn M. Sutherland,* Manuela S. Hancock,* Nicole L. La Gruta,‡ Lorena E. Brown,‡
Stephen J. Turner,‡ Andrew M. Lew,*,†,2 and Yifan Zhan*,1,2
*Walter and Eliza Hall Institute of Medical Research, Parkville, Australia; and Departments of †Medical Biology and‡Microbiology and Immunology, University of Melbourne, Parkville, Australia
RECEIVED JUNE 1, 2012; REVISED SEPTEMBER 12, 2012; ACCEPTED OCTOBER 11, 2012. DOI: 10.1189/jlb.0612266
ABSTRACTThe helper-dependent pathway of priming CD8� T cellsinvolves “licensing” of DCs by CD40L on CD4� T cells.The helper-independent (“helpless”) pathways elicitedby many viruses, including influenza, are less widely un-derstood. We have postulated that CD40L can be up-regulated on DCs by such viruses, and this promotespriming of CD8� T cells via CD40. Most studies on co-stimulation have been performed in the presence ofCD4� T cells, and so the role of CD40L costimulationunder helpless circumstances has not been fully eluci-dated. Here, we investigated such a role for CD40L us-ing CD40L KO mice. Although the number of influenza-specific CD8� T cells was unaffected by the absence ofCD4� T cells, it was markedly decreased in the ab-sence of CD40L. Proliferation (the number ofCD44�BrdU� influenza-specific CD8� T cells) in the pri-mary response was diminished in CD40L KO mice atDay 8 but not at Day 5 after infection. MLR studies indi-cated that CD40L expression on DCs was critical forCD8� T cell activation. Adoptive transfer of CD40 KOCD8� T cells compared with WT cells confirmed thatCD40 on such cells was critical for the generation of pri-mary anti-influenza CD8� T cell responses. The late effectalso corresponded with the late expression of CD40 byinfluenza-specific CD8� T cells. We suggest that costimu-lation via CD40L on DCs and CD40 on CD8� T cells is im-portant in optimizing primary CD8� T cell responses dur-ing influenza infection. J. Leukoc. Biol. 93: 145–154;2013.
IntroductionCD40–CD40L interactions have been shown to be importantfor CD4� T cell help in the primary CD8� CTL response [1,2]. The engagement of CD40L on CD4� T cells with CD40 onDCs results in the conditioning of DCs (termed DC licensing),which renders them capable of priming and activating CD8� Tcells [3]. DCs, after being licensed by CD4� T cells, producethe proinflammatory cytokine IL-12 [4], which promotes theexpansion and the development of effector function of CD8�
T cells [5].Many of these early studies used noninfectious agents [1, 2],
and indeed, primary CD8� T cell responses to certain virusinfections, such as HSV infection [6, 7], do require CD4� Tcell help. Early CD4� T cell help reduces CD8� T cell exhaus-tion and helps CD8� T cells to maintain HSV latency [8]. Inaddition, CD4� T cell help promotes the expression of CD25and production of IFN-� and TNF by HSV-specific CD8� Tcells [6] and facilitates the migration of CD8� T cells to in-fected sites and draining LNs [9, 10].
However, during other infections, such as those with influ-enza, ectromelia, vesicular stomatitis, vaccinia, and lymphocyticchoriomeningitis viruses, the primary CD8� T cell responsesare independent of CD4� T cell help [11–15]. There may beseveral mechanisms that render CD8� T cells independent ofCD4� T cell help. The up-regulation of CD40L on DCs duringinfluenza infection would seem one such mechanism [7].Other studies have indicated that CD40 on DCs can directlycostimulate CD8� T cells [16]. Induction of type I IFNs andIFN-� by viral infection may bypass CD4� T cell help in theinduction and proliferation of CD8� T cell responses [17, 18].
Although there have been many studies about the role ofcostimulation in CD8� T cell responses [19–26], few have fo-
1. Correspondence: WEHI, 1G Royal Parade, Parkville 3052, Australia.E-mail: [email protected]
2. These authors contributed equally to this work.
Abbreviations: APC�allophycocyanin, CD40L�CD40 ligand, CTV�celltrace violet, HSV�herpes simplex virus, KO�knockout,LCMV�lymphochoriomeningitis virus, MLR�mixed leukocyte reaction,NHMRC�National Health and Medical Research Council,NP�nucleoprotein, PA�influenza virus polymerase acidic protein,VAD-FMK�Val-Ala-Asp-fluoromethyl ketone
The online version of this paper, found at www.jleukbio.org, includessupplemental information.
Article
0741-5400/13/0093-145 © Society for Leukocyte Biology Volume 93, January 2013 Journal of Leukocyte Biology 145

cused on CD40L/CD40 [7, 16, 27]. During acute influenzainfection, the absence of CD40 signaling resulted in a decreasein influenza-specific CD8� T cells and IFN-� production inmice [7, 28]. Likewise, during vesicular stomatitis virus infec-tion, CD40L-deficient mice have reduced activated, proliferat-ing CD8� T cells and IFN-� production [12]. These studiesinvolving CD40L costimulation only investigated the outcomeof the immune response. However, studies with LCMV infec-tions revealed that the initial CD8� T cell response was notaffected by anti-CD40 treatment, but an increased cell loss wasobserved over time [29]. This finding suggested that CD40Lcostimulation might play a role in the development and/ormaintenance of antigen-specific CD8� T cells during thecourse of an infection. A caveat to many of these studies wasthat they were performed in the presence of CD4� T cells, sothe effect of costimulation could not be distinguished at thelevel of CD4� versus CD8� T cells.
Therefore, we sought to investigate the role of CD40L co-stimulation on the development of primary CD8� T cell re-sponse as a result of acute influenza virus infection in micedepleted of CD4� T cells to avoid any contribution of CD40Lon those cells. We found that unlike HSV infection or OVA-challenged mice, the direct CD40L costimulation of CD8� Tcells was more important than CD4� T cell help in elicitingprimary CD8� T cell responses for influenza virus infection. Inaddition, CD40L costimulation was crucial in the late but notearly phase of the primary response. As well as for viral infec-tions that induce CTL without the need for help, these find-ings may also provide useful insights into attempts at inducingCTLs in HIV patients with reduced CD4� T cell counts.
MATERIALS AND METHODS
MiceC57Bl/6 mice were used as WT control mice. CD40L KO mice [30] andCD40 KO mice [31] were back-crossed 10 times to C57Bl/6. CD80/86 KOmice [32] and BALB/c mice were used as allogeneic responders or stimula-tors in MLR experiments. All mice were 6–12 weeks old and maintained inspecific, pathogen-free conditions at the Walter and Eliza Hall Institute.Experiments were performed according to the guidelines of the institute’sAnimal Ethics Committee.
Influenza virus and intranasal infectionInfluenza Mem71 virus (a H3N1 genetic reassortant of A/Memphis/1/71X A/Bellamy/42) was used for most experiments. In cases where the pre-cursosr frequency was very low, we resorted to using TCR transgenic mice(adoptively transferred OT-I cells) [33] and recombinant influenza virusA/HKx31-OVA expressing the cognate OVA257–263 peptide (X31-OVA) forOT-I [34]. Mice were depleted of CD4� T cells by i.p. injections of 0.5 mgGK1.5, 1 day before and after infection. Penthrane-anesthetized mice wereinfected intranasally with 104.5 of Mem71 virus in a volume of 30 �l.
Determination of viral titersLungs were collected in 5 ml RPMI medium containing penicillin andstreptomycin. Lungs were sieved, and after clarification, the supernatantwas stored at �70°C. The titer of virus was determined by plaqueassay [35].
ELISAELISA was performed using serum from infected mice [36]. The antibodytiter was expressed as the reciprocal of the dilution of serum giving an OD2� above background.
Tetramer enrichment for influenza-specific CD8� TcellsInfluenza-specific CD8� T cells were enriched by positive selection [37].Briefly, single-cell suspensions were prepared from a pool of all major LNsand spleen. Cells were incubated in FcR block (1% mouse serum, 1% ratserum, and 40 �g/ml 2.4G2 Fc�III/II mAb) with PE-conjugated tetramericcomplexes of the H-2Db influenza PA224–233 (SSLENFRAYV) peptide atroom temperature for 1 h in the dark. After washing, anti-PE magneticbeads (Miltenyi Biotec, Auburn, CA, USA) were added and incubated onice for 30 min. After washing, the cells were passed over a LS magnetic col-umn (Miltenyi Biotec). Initial effluent was passed over the column again,followed by 3 � 3-ml washes of the column with cold buffer. The columnwas removed from the magnet and again washed with 5 ml cold buffer.The cells were incubated on ice for 30 min with an antibody cocktail ofanti-CD3e-PerCP-Cy5.5 (clone 145-2C11), anti-CD8-APC-Cy7 (clone 53-6.7),anti-CD44-PE-Cy7 (clone IM7), anti-CD62L-PE-Cy7 (clone MEL14), anti-CD40-biotin/strepavidin-FITC (clone 3/23), anti-CD4 (clone RM4-5)/-CD45R (clone RA3-6B2)/F4/80 (clone BM8)-APC, and additional anti-CD11c (clone HL3)/-CD11b (clone M1/70)-APC when looking at naïveprecursors. Cells were washed and kept on ice for further staining forBrdU incorporation.
Intracellular staining for IFN-� productionIntracellular staining for the production of IFN-� was carried out as de-scribed previously [38]. One million cells were cultured for 6 h with 50 Uhuman rIL-2 (Hoffmann-LaRoche, Basel, Switzerland) and 1 �l Golgi Plug,with or without 1 �M PA224–233 peptide. Surface staining was carried outwith FITC-conjugated rat anti-mouse CD8 (clone 53-6.7; BD Biosciences,San Jose, CA, USA) before intracellular IFN-� staining using the cytofix/cytoperm kit (BD Biosciences) and PE-conjugated anti-mouse IFN-� mAb(clone XMG1.2; BD Biosciences), as instructed by the manufacturer.
ELISPOT assay for IFN-�-producing CD8� T cellsPeptide-specific IFN-�-producing CD8� T cells were enumerated by theELISPOT assay, as described previously [39]. Ninety-six-well flat-bottompolyvinyl chloride microtitre plates (Dynatech, Alexandria, VA, USA) werecoated overnight with rat anti-mouse IFN-� antibodies (clone RA-6A2;PharMingen, San Diego, CA, USA) at 5 �g/ml in PBS. Unoccupied siteswere blocked with 10 mg/ml of BSA (CSL, Victoria, Australia) in PBS for 1h. Plates were washed with PBS containing 0.05% Tween 20, 5 � 105 irradi-ated syngeneic spleen cells, and 10 U human rIL-2 (Hoffmann-LaRoche),and cells from mediastinal LNs were added into the wells and incubatedwith or without NP366–374 peptide (ASNENMETM) for 18 h at 37°C, 5%CO2. Plates were washed and incubated with biotinylated anti-mouse IFN-�antibodies (clone XMG1.2; PharMingen) for 2 h at room temperature, fol-lowed by streptavidin-alkaline phosphatase for another 2 h. ELISPOT sub-strate (100 �l) [40], containing 1 mg 5-bromo-4-chloro-3-indolyphos-phate/ml 2-amino-2-methyl-1-propynol buffer (Sigma-Aldrich, St. Louis,MO, USA), was added, and plates were incubated for 30 min at 37°C, 5%CO2. Plates were washed with distilled water and air-dried before countingIFN-�-producing cells under an inverted microscope.
In vivo CTL assaysMice were immunized with 2 � 107-irradiated OVA-coated spleen cellsfrom a B6 or bm1 donor with 1 �g LPS i.v. Seven days later, mice weregiven i.v. 2 � 107 CFSEhigh-labeled SIINFEKL (Mimotopes, Clayton, Austra-lia) peptide-pulsed cells and CFSElow-labeled control, unpulsed cells inequal ratios. Spleen cells were analyzed by flow cytometry 18–24 h post-transfer.
146 Journal of Leukocyte Biology Volume 93, January 2013 www.jleukbio.org

BrdU incorporationMice were given 100 mg/kg BrdU (Sigma-Aldrich) i.p. once and 0.5mg/ml BrdU with 10 mM glucose in the drinking water for 2 days beforetotal LN and spleens were harvested. Single-cell suspensions were preparedand subjected to tetramer enrichment, and staining for surface markerswas carried out as described above. Intracellular staining for BrdU was thenperformed according to the BrdU Flow Kit (BD Biosciences).
Adoptive transfer of OT-I cells and analysis of OT-Icell proliferationTotal LNs were harvested from 6- to 12-week-old OT-I/Ly5.1 transgenicmice and single-cell suspension prepared. OT-I cells were then enriched bymagnetic-bead depletion with rat anti-mouse mAb M1/70 (anti-MAC-1),Ter119 (anti-erythrocytes), F4/80 (anti-Mac-3), RB6-8C5 (anti-Gr-1), M5/114 (anti-MHC class II), and GK1.5 (anti-CD4), followed by goat anti-ratIgG-coupled magnetic beads (BioMag, Qiagen, Hilden, Germany). Thenonbead-bound fraction-enriched CD8� T cells were washed once and re-suspended in PBS. Cells were then stained using CellTrace Violet, accord-ing to the manufacturer’s instructions (Invitrogen, Carlsbad, CA, USA).Cells were washed and resuspended in PBS. OT-I cells were adoptivelytransferred by i.v. injection of 0.5 � 106 V�2�CD8� T cells into X31-OVA-infected, CD4� T cell-depleted mice.
Spleen and mediastinal LNs from X31-OVA-infected mice were har-vested at Day 5 post-transfer of OT-I cells. OT-I CD8� T cells were en-riched from organs of experimental mice by magnetic depletion as de-scribed above. The enriched cells were incubated on ice for 30 min with anantibody cocktail of anti-V�2-biotin (clone B20.1)/streptavidin-FITC, anti-CD8-APC-Cy7 (clone 53-6.7), and anti-CD45-1-APC (clone A20). Cells werewashed and resuspended in FACS buffer containing 1 �g/ml PI to excludedead cells during flow cytometry analysis. A total of 104 CaliBRITE APCbeads (BD Biosciences) was added to each sample prior to flow cytometryto allow quantification of OT-I cells.
DC enrichmentDCs from spleens of WT or CD40L KO mice were enriched [41]. Briefly,spleens were digested with collagenase-DNAase. Light-density cells were se-lected by centrifugation in a 1.077-g/cm3 Nycodenz medium (Rodeløkka,Oslo, Norway). CD11c� DCs were isolated using FITC anti-CD11c antibody(BD Biosciences) and anti-FITC magnetic beads (Miltenyi Biotec).
Analysis of OT-I cell proliferation in vitroPurified CD11c� DCs (1.25�104) from WT or CD40L KO mice were in-fected with X31-OVA, at fivefold MOI, at 37°C for 45 min in serum-freeRPMI. The inoculum was removed, and 5 � 104 CTV-labeled OT-I cellswere added. The cells were incubated at 37°C at 5% CO2 for 60 h or 72 hin RPMI, supplemented with 10% FCS and 0.05 mM 2-ME (Sigma-Aldrich).Cells were pelleted, and supernatant was harvested and kept frozen at�70°C for cytokine assays. The cells were washed with cold buffer beforestaining with anti-TCR-� chain-APC (clone H57-597; BD Biosciences), anti-CD8-APC.Cy7 (clone 53-6.7; BD Biosciences), anti-CD40-PE (clone 3/23;BD Biosciences), and anti-CD44-PE.Cy7 (clone IM7; eBioscience, San Di-ego, CA, USA). Cells were washed and kept on ice before analysis by flowcytometry.
MLRCD8� T cells were enriched using anti-CD8-PE-conjugated antibody (BDBiosciences) and anti-PE magnetic MACS beads (Miltenyi Biotec). All en-riched cells were washed and resuspended in RPMI, supplemented with10% heat-inactivated FCS and 0.05 mM 2-ME (Sigma-Aldrich). B6 CD40LKO or BALB/c WT 1 � 105 CD8� T cells were cocultured with BALB/cWT or B6 CD40L KO DCs, respectively, at a 1:1 ratio for 4 or 6 days. Triti-ated thymidine was added 16 h before harvest.
Pan-caspase stainingTo enumerate cells undergoing apoptosis, staining of activated caspase wasperformed using CaspACE FITC-VAD-FMK (Promega, Madison, WI, USA).This broad caspase inhibitor VAD-FMK compound, which was FITC-conju-gated, is cell-permeable and binds to activated caspase. The inhibitor (10mM) was incubated with the cells at 37°C at 5% CO2 for 20 min before thecells were harvested. The cells were washed with cold buffer before stainingwith anti-TCR-� chain-APC (clone H57-597; BD Biosciences), anti-CD8-AP-C.Cy7 (clone 53-6.7; BD Biosciences), anti-CD40-PE (clone 3/23; BD Biosci-ences), and anti-CD44-PE.Cy7 (clone IM7; eBioscience). Cells were washedand kept on ice before analysis by flow cytometry.
Transfer of CD40 KO CD8� T cells to RAG hostsSpleen and LNs of either CD40 KO or WT mice were enriched for CD8�
cells [42]. Three million cells (84-88% CD8�) were transferred i.v. intoRAG1 KO mice. To eliminate residual CD4� T cells, 0.5 mg GK1.5 was in-jected i.p. Mice were rested for 4 weeks after transfer (when the level ofhomeostatic proliferation has subsided) and infected with influenza as de-scribed above.
Data analysisFlow cytometric data were analyzed using FlowJo software (version 3.6.1;Tree Star, Ashland, OR, USA). The nonparametric Mann-Whitney U-testwas used to determine statistical significance (Prism 4; GraphPad Software,San Diego, CA, USA).
RESULTS
Antigen (viral) load is similar between CD40L KOand WT miceTo investigate the role of costimulation in CTL induction un-der helpless circumstances, WT and CD40L KO mice were an-tibody-depleted of CD4� T cells; this resulted in �99% deple-tion (Supplemental Fig. 1). CD4� T cell-depleted mice wereinfected intranasally with the same dose of influenza virus. Toensure that the virus replicated to a similar extent in the dif-ferent strains of mice, i.e., the antigen load would be equiva-lent among the different strains, we chose a time-point whenthe virus had grown enough to be easily and accurately mea-sured but before any clearing effect had been educed by theadaptive immune system [43]. We therefore harvested thelungs at Day 5 postinfection and found that the two groups ofinfected mice had similar viral loads (Fig. 1). Thus, this estab-lished that any differences in CTL response magnitude wasnot a result of differences in antigen load. This does not pre-clude that there may be differences in inflammatory nichesbetween the two strains of mice during influenza infection.
Generation of helpless antigen-specific CD8� Tresponses is impaired in the absence of CD40LcostimulationWe wanted to determine the requirement for costimulation ingenerating influenza-specific CTL responses in WT and CD40LKO mice in the absence of CD4� T cells. The abundance ofinfluenza-specific CD8 T cells in B6 mice is highest against theepitope, residues 224–233 of acid PA bound to the MHC classI molecule H-2Db during primary infection [37]. Therefore,we decided to focus on PA-specific CD8� T cells in our study.To ensure that the different groups of mice had equivalent,
Seah et al. CD40L costimulation of primary anti-influenza CTL
www.jleukbio.org Volume 93, January 2013 Journal of Leukocyte Biology 147

naïve precursor frequencies, the number of naïve, PA-specificCD8� T cells in uninfected mice was determined after te-tramer enrichment. We found that the number of PA-specific,naïve precursors was similar between the different groups ofmice (Fig. 2A).
At Day 8 postinfection, we found that CD4� T cell defi-ciency did not lead to a reduction in PA-specific CD8� T cellnumbers in WT mice (13.0�1.4�103 cells in untreated WTmice vs. 17.5�3.1�103 cells in GK1.5-treated WT mice;P�0.52; Fig. 2B and Supplemental Fig. 2A). In contrast, theloss of CD40L costimulation resulted in a dramatic decrease inPA-specific CD8� T cell numbers (2.8�0.7�103 cells in CD40LKO mice). However, the additional loss of CD4� T cells inCD40L KO mice did not reduce the number of PA-specificCD8� T cells any further than that found in the absence ofCD40L costimulation alone (3.7�0.4�103 cells in GK1.5-
treated CD40L KO mice). Hence, this demonstrated that theoptimal generation of PA-specific CD8� T cells was dependenton the direct CD40L costimulation of CD8� T cells.
CD40L affects the numbers of proliferatedantigen-specific CD8� T cellsWe next looked at the activation of PA-specific CD8� T cellsby staining for CD44 and found that the proportion of unacti-vated, PA-specific CD8� T cells (viz. CD44�) was similar be-tween CD40L KO mice and WT mice, 8 days after infection(Fig. 3A). Furthermore, proliferation of activated, PA-specificCD8� T cells assessed by BrdU incorporation showed only amodest reduction in the proportion of CD44�BrdU� PA-spe-cific CD8� T cells in CD40L KO mice compared with WTmice (52.1%�5.3 and 70.3%�3.1, respectively; P�0.05; Fig. 3Band Supplemental Fig. 2B). In contrast, the reduction in abso-lute numbers of CD44�BrdU� PA-specific CD8� T cells wasdramatic (sixfold less) in CD40L KO mice compared with WTmice (1.9�0.3 and 12.4�2.2�103 cells, respectively; P�0.005;Fig. 3C). Therefore, the strongest effect of CD40L deficiencywas the reduction of the absolute numbers of proliferated cellsrather than its proportion.
To determine whether the loss of CD40L costimulation af-fected the production of IFN-� by helpless CD8� T cells, agroup of CD4� T cell-depleted WT mice was treated with theblocking anti-CD40L antibody (MR1) prior to infection. Whenstimulated by PA-peptide ex vivo, the reduction in the IFN-�production by helpless CD8� T cells in mice under CD40Lblockade was slight and not statistically significant (P�0.06;Fig. 3D). To investigate whether the loss of CD4� T cells hadan effect on the activation of CD8� T cells, an ELISPOT assaywas carried out. Cells from CD4-intact or CD4-depleted CD40LKO mice were stimulated with NP366–374 peptide, and thenumber of IFN-�-producing, NP-specific CD8� T cells was enu-merated. Results indicated that the loss of CD4� T cells hadno effect on the activation of NP-specific CD8� T cells (Sup-plemental Fig. 2C).
To investigate whether the blockade of CD40L costimulationat various time-points would affect the generation of CD8� Tcells, mice were treated with mAb MR1 at various time-points(at Day 1 or Day 5). Spleen cells were analyzed at 18–24 h to
Figure 1. Antigen load in lungs of infected mice at Day 5. To depleteCD4� T cells, all mice were treated with 0.5 mg GK1.5 antibody 1 daybefore and 1 day after intranasal infection with 104.5 PFU Mem71 vi-rus. Lungs were harvested at Day 5 postinfection, and viral titers weredetermined by plaque assay using Madin-Darby canine kidney cells.Data are shown as mean � sem of n � 12, 6, and 9 for WT, CD40LKO, and naïve mice, respectively, for Day 5. Data were pooled fromtwo independent experiments. The detection limit of the virus assay is1.8 log.
Figure 2. Number of PA-specific CD8� T cells before andafter infection. (A) The total number of naïve PA precur-sors was determined in uninfected littermates treated with0.5 mg GK1.5 1 day before harvest. Data are shown asmean � sem of n � 15 (WT) and n � 6 (CD40L KO),pooled from two independent experiments. (B) To depleteCD4� T cells, mice were treated with 0.5 mg GK1.5 anti-body, 1 day before and 1 day after intranasal infection, with104.5 PFU Mem71 virus. Total LNs and spleen were har-vested from infected mice at Day 8 postinfection to deter-mine the total number of PA-specific CD8� T cells. Dataare shown as mean � sem of n � 8, 5, 14, 9, and 12 for WT,CD40L KO, GK1.5-treated WT, GK1.5-treated CD40L KO, andnaïve mice, respectively. Data were pooled from two indepen-dent experiments. Mann-Whitney U-test. ***P � 0.0005;**P � 0.005.
148 Journal of Leukocyte Biology Volume 93, January 2013 www.jleukbio.org

determine the killing of target cells by antigen-specific CD8�
T cells. Results demonstrated that there was no difference be-tween blockade of CD40L costimulation before infection (Day1) or at Day 5 postinfection (Supplemental Fig. 3). These re-sults suggested that the requirements of CD40L costimulationmight occur at a later time-point.
CD40L is not critical during the early phase ofprimary responseTo explore further how CD40L costimulation affects the earlydevelopment of antigen-specific CD8� T cells responses, welooked at an earlier time-point (viz. 5 days postinfection), al-though the numbers were expected to be low [37]. Surpris-ingly, we found that the total number of PA-specific CD8�
T cells in CD40L KO mice was similar to that in WT mice(150�15 and 134�15 cells, respectively; P�0.45; Fig. 4A).The percentage of PA-specific CD8� T cells that wereCD44�BrdU� in CD40L KO mice was also similar to WT mice(16.6�3.5% and 18.1�3.1%, respectively; P�0.89; Fig. 4B) andlikewise, for the absolute numbers of CD44�BrdU� PA-specificCD8� T (27.7�5.9 and 25.3�3.8 cells, respectively; P�0.74;Fig. 4C). Hence, CD40L was not critical in the early phase ofthe primary response.
Figure 4. Activation and proliferation of PA-specific CD8� T cells atDay 5 postinfection. To deplete CD4� T cells, all mice were treatedwith 0.5 mg GK1.5 antibody, 1 day before and 1 day after intranasalinfection with 104.5 PFU Mem71 virus. Mice were given BrdU by i.p.on Day 3 postinfection, together with BrdU in drinking water on Days3 and 4 postinfection. Total LNs and spleen were harvested at Day 5postinfection. (A) The absolute number of PA-specific CD8� T cellswas determined by counting the entire tetramer-specific population.(B) The percentage and (C) total number of activated and proliferat-ing PA-specific CD8� T cells were determined by staining for CD44and BrdU, respectively. Data are shown as mean � sem of n � 12, 6,and 9 for WT, CD40L KO, and naïve mice, respectively. Statisticalanalysis was performed using the Mann-Whitney U-test. Data werepooled from two repeated experiments.
Figure 3. Activation and proliferation of PA-specific CD8� T cells atDay 8 postinfection. To deplete CD4� T cells, all mice were treatedwith 0.5 mg GK1.5 antibody, 1 day before and 1 day after intranasalinfection with 104.5 PFU Mem71 virus. Mice were given BrdU i.p. onDay 6 postinfection, together with BrdU in drinking water on Days 6and 7 postinfection. Total LNs and spleen were harvested at Day 8postinfection. (A) Activation of PA-specific CD8� T cells was deter-mined by staining for CD44. (B) The percentage and (C) absolutenumber of CD44�BrdU� PA-specific CD8� T cells were determined bycounting the entire tetramer-specific population. Data are shown asmean � sem of n � 14, 9, and 9 for WT, CD40L KO, and naïve mice,respectively, and were pooled from two independent experiments.*P � 0.05; ***P � 0.0005. (D) A group of WT mice was given addi-tional treatment of MR1 antibody to block CD40L costimulation.Spleen cells from infected mice were stimulated with PA peptide andanalyzed for IFN-� production by intracellular staining. Data areshown as mean � sem of n � 5, 4, and 2 for GK1.5-treated, GK1.5 �MR1-treated, and naïve mice, respectively, and are representative oftwo separate experiments. Statistical analysis was performed using theMann-Whitney -test.
Seah et al. CD40L costimulation of primary anti-influenza CTL
www.jleukbio.org Volume 93, January 2013 Journal of Leukocyte Biology 149
As the above numbers of endogenous, PA-specific CD8� Tcells at this early time-point were so low, we wanted to confirmthe observation in some other way. Therefore, we took advan-tage of the X31-OVA influenza virus that expresses the OVAepitope in the neuraminidase stalk and adoptively transferredCellTrace Violet-labeled OT-I cells to accurately map the con-tribution of CD40L costimulation to amplification of antigen-specific T cells. We infected WT or CD40L KO mice with X31-OVA before adoptively transferring dye-labeled OT-I into in-fected mice. The OT-I cells were harvested 5 days post-transfer, and proliferation profiles were analyzed. We foundthat the proliferation profiles of OT-I cells in WT and CD40LKO mice were similar in spleen and mediastinal LNs (Supple-mental Fig. 4A and B). In addition, there was no impairmentin the activation of OT-I cells with regards to CD44 expression(Supplemental Fig. 4C and D). Therefore, the results obtainedwith PA-specific CD8� T cells and OT-I cells indicated thatCD40L costimulation does not affect the early activation andexpansion of antigen-specific CD8� T cell responses in vivo.
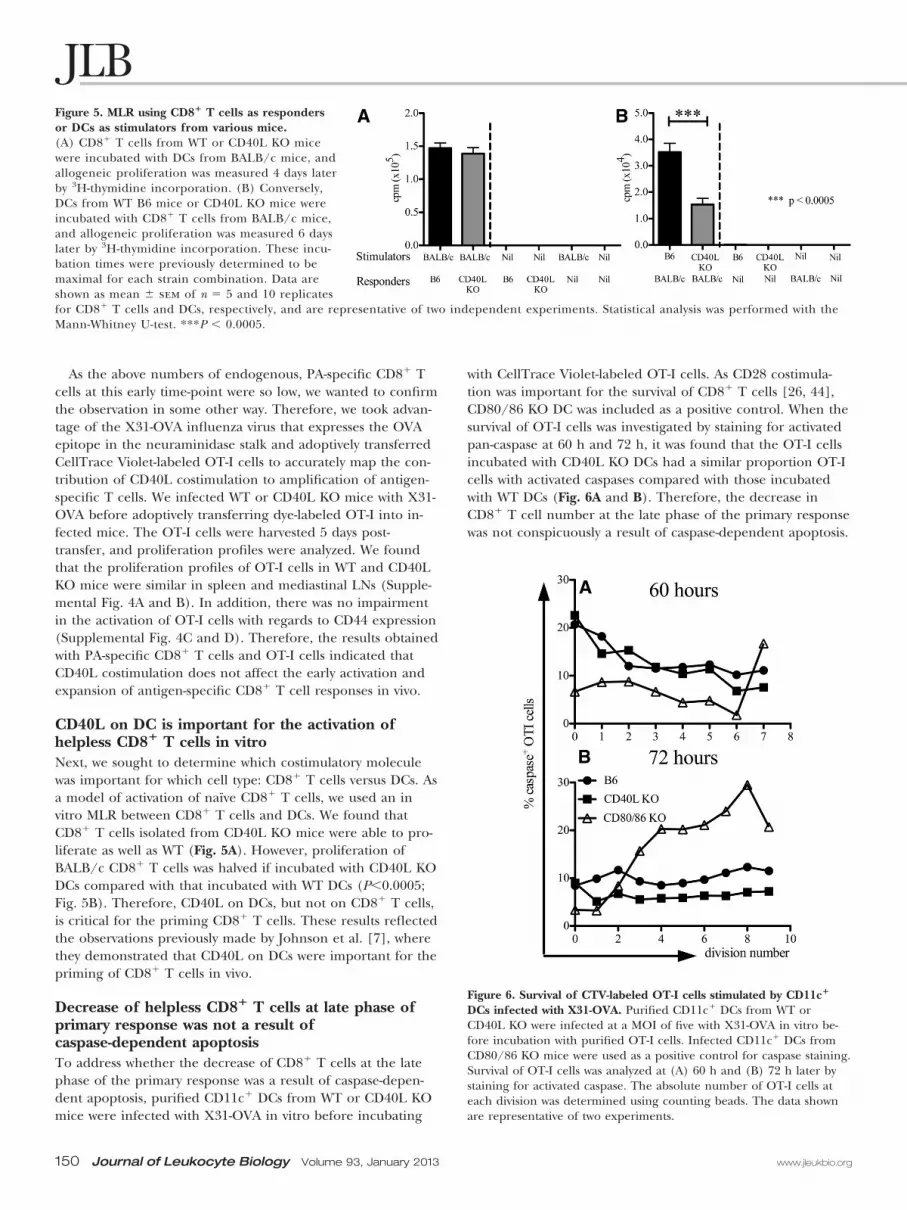
CD40L on DC is important for the activation ofhelpless CD8� T cells in vitroNext, we sought to determine which costimulatory moleculewas important for which cell type: CD8� T cells versus DCs. Asa model of activation of naïve CD8� T cells, we used an invitro MLR between CD8� T cells and DCs. We found thatCD8� T cells isolated from CD40L KO mice were able to pro-liferate as well as WT (Fig. 5A). However, proliferation ofBALB/c CD8� T cells was halved if incubated with CD40L KODCs compared with that incubated with WT DCs (P�0.0005;Fig. 5B). Therefore, CD40L on DCs, but not on CD8� T cells,is critical for the priming CD8� T cells. These results reflectedthe observations previously made by Johnson et al. [7], wherethey demonstrated that CD40L on DCs were important for thepriming of CD8� T cells in vivo.
Decrease of helpless CD8� T cells at late phase ofprimary response was not a result ofcaspase-dependent apoptosisTo address whether the decrease of CD8� T cells at the latephase of the primary response was a result of caspase-depen-dent apoptosis, purified CD11c� DCs from WT or CD40L KOmice were infected with X31-OVA in vitro before incubating
with CellTrace Violet-labeled OT-I cells. As CD28 costimula-tion was important for the survival of CD8� T cells [26, 44],CD80/86 KO DC was included as a positive control. When thesurvival of OT-I cells was investigated by staining for activatedpan-caspase at 60 h and 72 h, it was found that the OT-I cellsincubated with CD40L KO DCs had a similar proportion OT-Icells with activated caspases compared with those incubatedwith WT DCs (Fig. 6A and B). Therefore, the decrease inCD8� T cell number at the late phase of the primary responsewas not conspicuously a result of caspase-dependent apoptosis.
Figure 5. MLR using CD8� T cells as respondersor DCs as stimulators from various mice.(A) CD8� T cells from WT or CD40L KO micewere incubated with DCs from BALB/c mice, andallogeneic proliferation was measured 4 days laterby 3H-thymidine incorporation. (B) Conversely,DCs from WT B6 mice or CD40L KO mice wereincubated with CD8� T cells from BALB/c mice,and allogeneic proliferation was measured 6 dayslater by 3H-thymidine incorporation. These incu-bation times were previously determined to bemaximal for each strain combination. Data areshown as mean � sem of n � 5 and 10 replicatesfor CD8� T cells and DCs, respectively, and are representative of two independent experiments. Statistical analysis was performed with theMann-Whitney U-test. ***P � 0.0005.
Figure 6. Survival of CTV-labeled OT-I cells stimulated by CD11c�
DCs infected with X31-OVA. Purified CD11c� DCs from WT orCD40L KO were infected at a MOI of five with X31-OVA in vitro be-fore incubation with purified OT-I cells. Infected CD11c� DCs fromCD80/86 KO mice were used as a positive control for caspase staining.Survival of OT-I cells was analyzed at (A) 60 h and (B) 72 h later bystaining for activated caspase. The absolute number of OT-I cells ateach division was determined using counting beads. The data shownare representative of two experiments.
150 Journal of Leukocyte Biology Volume 93, January 2013 www.jleukbio.org

CD40 on CD8 T cells is critical for helplessinfluenza-induced expansionAlthough the in vitro DC:CD8� T cell MLR assays indicatedthat CD40L on DCs (and thus, CD40 on CD8� T cells) wascritical for priming, the in vivo influenza experiments usingCD40L KO mice do not ascribe the causative cell type (DCs,NK cells, or CD8� T cells). We therefore transferred purifiedCD8� T cells from CD40 KO or WT mice (and therefore, allother cell types, including NK cells, are of WT genotype) and
determined the number of PA-specific T cells. Mice weretreated with GK1.5 to eliminate any residual CD4� T cells; thiswas shown to be effective (Fig. 7A). The “take” of the trans-ferred CD8� T cells was comparable (Fig. 7A). Eight days afterinfection, the numbers of PA-specific CD40 KO CD8� T cellswere reduced dramatically compared with that of WT(160�103 vs. 10�103; P�0.05 Mann Whitney U-test; Fig. 7Band C). This finding cogently shows the role of CD40 onCD8� T cells for helpless anti-influenza CTL induction.
11.11.2
Total CD8+ T cells
0.5 3.3 0.05 0.06 0.05
CD4
Infected Uninfected
CD
8
PA-specific CD8+ T cells
% o
f PA
tetra
mer
+ cel
ls/to
tal l
euko
cyte
Percentage and number of PA tetramer+ cells of individual mice
**
Infected
PA tetramer
CD
8
donor CD8+ WT CD40-/- WT CD40-/-host WT Rag-/- Rag-/- Rag-/- Rag-/-
Infected Uninfected
host WT Rag-/- Rag-/- Rag-/- Rag-/-
0
1
2
0
200
400
600
*
DonorCD8+ T cell
Num
ber o
f PA
tetra
mer
+ cel
ls/ m
ouse
(x10
00)
WT CD40-/- WT CD40-/- WT CD40-/- WT CD40-/-
Uninfected Infected Uninfected
donor CD8+ WT CD40-/- WT CD40-/-
Days
%CD
40+ /
PA+ C
D8+ T
cel
ls%
CD40
+/P
A+ C
D8+ T
cel
ls
0 5 6 7 80.0
0.5
1.0
1.5
2.0
2.5
Days
abso
lute
num
bero
fCD
40+
PA+
CD8+
Tce
lls(x
103 )
0 5 6 7 80
10
20
30 ** p < 0.05
A
B
C
D
Figure 7. The role of CD40 on PA-specific CD8� Tcells. (A–C) RAG1 KO mice were injected i.v. with 3million CD8 T cells isolated from the spleens andLNs of CD40 KO or WT mice. Four weeks later, themice were infected intranasally with influenza virus,and spleen was harvested at Day 8 after infection. (A)FACS plot showing gross numbers of CD8 and CD4 Tcells. (B) FACS plot showing percentages of PA-spe-cific CD8 T cells. (C) Percentages and numbers ofPA-specific CD8 T cells shown in column graphs.Each dot respresents an indivual mouse; mean andsem are also indicated. (D and E) Expression ofCD40 on PA-specific cD8 T cells. To deplete CD4� Tcells, all mice were treated with 0.5 mg GK1.5 anti-body 1 day before and 1 day after intranasal infectionwith 104.5 PFU Mem71 virus. All major LNs andspleen were harvested from infected mice to deter-mine (D) the total number of PA-specific CD8� Tcells expressing CD40 and (E) percentage of PA-spe-cific CD8� T cells expressing CD40. Mean � sem isindicated; n � 4–6 for infected mice; n � 4 for naïvemice. Four sets of pooled, naïve mice (n�4) wereused in E to obtain a sufficient number for the per-centage of CD40� PA-specific CD8� T cells. Statisticalanalysis was done using the Mann-Whitney U-test.**P � 0.01; *P � 0.05.
Seah et al. CD40L costimulation of primary anti-influenza CTL
www.jleukbio.org Volume 93, January 2013 Journal of Leukocyte Biology 151

Previous studies have shown that the up-regulation of CD40expression was not immediate and the timing depended onthe cell type and infection/stimulus involved [45, 46]. There-fore, we postulated that the requirements of CD40L costimula-tion at the late phase could be a result of the delayed up-regu-lation of CD40 on PA-specific CD8� T cells. Indeed, the num-ber of CD40-expressing, PA-specific CD8� T cells increaseddramatically only by Day 7 (from undetectable on Day 6 to1.2�0.7�103 cells) and Day 8 (2.0�0.5�103 cells) postinfec-tion (Fig. 7D). The percentage of PA-specific CD8� T cellsexpressing CD40 in infected mice steadily increased after Day5 (Fig. 7E). Therefore, the delayed up-regulation of CD40 in-dicated that CD40–CD40L costimulation was only required atthe late phase. These findings are in accord with our blockingstudies above, whereby blockade at Day 5 after infection withanti-CD40L was effective at reducing CTL responsess (Supple-mental Fig. 3).
DISCUSSION
Early studies that demonstrated the role of CD40–CD40L inCD4� T cell help were carried out using OVA antigens [1, 2].Like the nonmicrobial OVA antigens, certain virus infections,such as HSV, require CD4� T cell help in the generation of aprimary CD8� T cell response, whereas others, such as influ-enza and LCMV that induce inflammation, do not [17, 18].There may be several determining factors that render CD8� Tcells independent of CD4� T cell help, and such factors maydiffer according to the infectious models used and the magni-tude of inflammation elicited. We have shown that up-regula-tion of CD40L on DCs can bypass the need for help by MLRherein and by mixed bone marrow chimaeras previously [7].Whether selective induction of inflammatory cytokines (e.g.,IL-12, IFNs) by certain viruses influence the up-regulation ofCD40L on DCs, which allows CD8� T cells to be help-indepen-dent, remains to be determined.
The role of CD4� T cell help in influenza has been studiedby many, including ourselves, using MHC class II-deficient ani-mals [14, 47, 48] or treatment with depleting anti-CD4 anti-body [13, 49, 50]. Such studies have found that in the absenceof CD4� T cell help but intact costimulation pathways, strongCD8� T cell responses were elicited in response to influenza.
In this investigation, we showed that the presence or ab-sence of CD4� T cell help did not influence the number ofantigen-specific CD8� T cells in the CD40L-deficient mouse.Hence, unlike demonstrated previously for noninfectious anti-gens [1, 2], the direct CD40L costimulation of CD8� T cellsand not costimulation via CD4� T cells was important in thegeneration of primary anti-influenza CD8� T cell responses. Inother words, whereas lack of help had little effect, the lack ofdirect CD40L/CD40 costimulation for CD8� T cells resultedin low numbers of PA-specific CD8� T cells. These results con-cur with those demonstrated previously by Hernandez et al.[16], where the direct CD40–CD40L interaction between DCsand CD8� T cells is required for the generation of anti-LCMVT cell responses in the absence of CD4� T cell help, althoughtheir conclusion was that CD40L on CD8� T cells was impor-tant. Previous reports have shown that CD40 can be expressed
by T cells, especially activated ones [7, 51, 52]. One reportfound that direct signaling to CD40 on CD8� T cells was notrequired during influenza infection [28]. However, such stud-ies were not performed in the absence of CD4� T cells. Weopine that there are at least two pathways whereby CD40L isimportant: the classical axis of CD40L� CD4 T cells licensingCD40� DCs that then activate CD8� T cells and virus-inducedCD40L up-regulation on DCs that activate CD40� CD8 T cells,which is independent of CD4� T cell help. Thus, in the pres-ence of CD4� T cells, the former pathway would mask the ef-fect of the latter pathway.
There are few, if any, studies on the effects of CD40L co-stimulation at the initiation of activation and expansion ofCD8� T cells. We found that the loss of CD40L costimulationin helpless mice had little effect on the early priming and acti-vation of antigen-specific CD8� T cells (i.e., Day 5). However,the effect of CD40L costimulation was observed only at thelate phase of the primary response, as demonstrated by thereduction of activated, PA-specific CD8� T cells, Day 8 postin-fection. We have also demonstrated that CD40L costimulationwas not required for the activation of PA-specific CD8� T cells,although the efficient production of IFN-� production wasslightly affected during the CD40L costimulation blockade.Our investigation shared similar results with a very recent re-port [53], demonstrating that CD40 costimulation was impor-tant at the late phase of the primary response; CD40 on CD8�
T cells is crucial and nonredundant for their progressive dif-ferentiation into memory cells.
The effect of CD40L deficiency was greatest for the reduc-tion of the absolute number of proliferated cells rather thanits proportion at Day 8 of infection. There are at least threepossible explanations (not mutually exclusive) for this result.First, the stimulated, antigen-specific T cells die more rapidlyin the absence of CD40L/CD40 costimulation. Our pan-caspase staining does not support this. However, it should benoted that apoptotic assays have the caveat that they do notmeasure cells that have died and not recovered (e.g., cells thathave been phagocytosed). At best, these assays provide a snap-shot of what is occurring. Second, the proportion of cells acti-vated is unaffected, but there is a delay in time to the first divi-sion of cells in the absence of costimulation [54], and so, thefinal output of proliferated cells would be reduced. Third, ifCD40 signaling were important for proliferation/survival, thenthe late onset of CD40 expression by antigen-specific CD8� Tcells would explain why the differential is more conspicuous inthe late phase of the primary response.
Previous studies had shown that CD4� T cells were criticalfor the development of memory CD8� T cells during influ-enza infection [14, 55–57]. Hence, investigations on the effectsof CD40–CD40L costimulation on the development of mem-ory CD8� T cells in the absence of CD4� T cell help duringinfluenza infection were not carried out. It is nevertheless ofnote that CD40/CD40L interactions are involved in the CD4�
T cell help of CTL memory induction [53, 58, 59].Our findings would suggest that unlike what we have dem-
onstrated previously for CD28 costimulation [50], CD40L/CD40 might be more important in the downstream expansionof antigen-specific CD8� T cells in the late phase of the pri-
152 Journal of Leukocyte Biology Volume 93, January 2013 www.jleukbio.org

mary response. This notion accords with earlier studies, whereCD28 was critical for initiating T cell responses, whereasCD40L was required for sustained Th1 responses [60]. Re-cently, CD40 agonists, such as dacetuzumab, are in clinical tri-als as anticancer therapies [61–64]. Our results suggest thatCD40 agonists, especially for those defective in CD4� T cells(e.g., people with HIV/AIDS), may promote antiviral CTL re-sponses, and the timing of such agonists may be critical.
AUTHORSHIP
S.G.K.S. planned and performed the experiments and wrotethe draft manuscript. J.L.B. and E.M.C. contributed to experi-ments. W.C.N. performed the intracellular staining assay.M.S.H. performed genotyping for all of the mice used. R.M.S.,N.L.L.G., L.E.B., Y.Z., and S.J.T. contributed to brainstormingand discussions. A.M.L., Y.Z., and S.J.T. contributed to editingthe manuscript and supervising the project.
ACKNOWLEDGMENTS
This work was supported by the NHMRC of Australia programand project grants (516700, 575543, 637324, and 10073), Juve-nile Diabetes Research Foundation grants (112613), VictorianState Government Operational Infrastructure Support andAustralian Government NHMRC IRIIS, Australian ResearchCouncil Future Fellowship (S.J.T.), and DSO National Labora-tories Scholarship, Singapore (S.G.K.S.). We thank Dr. IanBarr and Chris Durrant (World Health Organization Collabo-rating Centre for Reference and Research on Influenza) forhelp in virus culture and Dr. Gabrielle Belz for providing re-agents.
REFERENCES
1. Bennett, S. R., Carbone, F. R., Karamalis, F., Flavell, R. A., Miller, J. F.,Heath, W. R. (1998) Help for cytotoxic-T-cell responses is mediated byCD40 signalling. Nature 393, 478–480.
2. Schoenberger, S. P., Toes, R. E., van der Voort, E. I., Offringa, R., Me-lief, C. J. (1998) T-cell help for cytotoxic T lymphocytes is mediated byCD40-CD40L interactions. Nature 393, 480–483.
3. Ridge, J. P., Di Rosa, F., Matzinger, P. (1998) A conditioned dendriticcell can be a temporal bridge between a CD4� T-helper and a T-killercell. Nature 393, 474–478.
4. Filatenkov, A. A., Jacovetty, E. L., Fischer, U. B., Curtsinger, J. M., Me-scher, M. F., Ingulli, E. (2005) CD4 T cell-dependent conditioning ofdendritic cells to produce IL-12 results in CD8-mediated graft rejectionand avoidance of tolerance. J. Immunol. 174, 6909–6917.
5. Curtsinger, J. M., Johnson, C. M., Mescher, M. F. (2003) CD8 T cellclonal expansion and development of effector function require pro-longed exposure to antigen, costimulation, and signal 3 cytokine. J. Im-munol. 171, 5165–5171.
6. Rajasagi, N. K., Kassim, S. H., Kollias, C. M., Zhao, X., Chervenak, R.,Jennings, S. R. (2009) CD4� T cells are required for the priming ofCD8� T cells following infection with herpes simplex virus type 1. J.Virol. 83, 5256–5268.
7. Johnson, S., Zhan, Y., Sutherland, R. M., Mount, A. M., Bedoui, S.,Brady, J. L., Carrington, E. M., Brown, L. E., Belz, G. T., Heath, W. R.,Lew, A. M. (2009) Selected Toll-like receptor ligands and viruses pro-mote helper-independent cytotoxic T cell priming by upregulatingCD40L on dendritic cells. Immunity 30, 218–227.
8. Frank, G. M., Lepisto, A. J., Freeman, M. L., Sheridan, B. S., Cherpes,T. L., Hendricks, R. L. (2010) Early CD4(�) T cell help prevents partialCD8(�) T cell exhaustion and promotes maintenance of herpes sim-plex virus 1 latency. J. Immunol. 184, 277–286.
9. Kumamoto, Y., Mattei, L. M., Sellers, S., Payne, G. W., Iwasaki, A. (2011)CD4� T cells support cytotoxic T lymphocyte priming by controllinglymph node input. Proc. Natl. Acad. Sci. USA 108, 8749–8754.
10. Nakanishi, Y., Lu, B., Gerard, C., Iwasaki, A. (2009) CD8(�) T lympho-cyte mobilization to virus-infected tissue requires CD4(�) T-cell help.Nature 462, 510–513.
11. Rahemtulla, A., Fung-Leung, W. P., Schilham, M. W., Kundig, T. M.,Sambhara, S. R., Narendran, A., Arabian, A., Wakeham, A., Paige, C. J.,Zinkernagel, R. M.. (1991) Normal development and function of CD8�cells but markedly decreased helper cell activity in mice lacking CD4.Nature 353, 180–184.
12. Andreasen, S. O., Christensen, J. E., Marker, O., Thomsen, A. R. (2000)Role of CD40 ligand and CD28 in induction and maintenance of antivi-ral CD8� effector T cell responses. J. Immunol. 164, 3689–3697.
13. Allan, W., Tabi, Z., Cleary, A., Doherty, P. C. (1990) Cellular events inthe lymph node and lung of mice with influenza. Consequences of de-pleting CD4� T cells. J. Immunol. 144, 3980–3986.
14. Belz, G. T., Wodarz, D., Diaz, G., Nowak, M. A., Doherty, P. C. (2002)Compromised influenza virus-specific CD8(�)-T-cell memory inCD4(�)-T-cell-deficient mice. J. Virol. 76, 12388–12393.
15. Buller, R. M., Holmes, K. L., Hugin, A., Frederickson, T. N., Morse III,H. C. (1987) Induction of cytotoxic T-cell responses in vivo in the ab-sence of CD4 helper cells. Nature 328, 77–79.
16. Hernandez, M. G., Shen, L., Rock, K. L. (2007) CD40-CD40 ligand in-teraction between dendritic cells and CD8� T cells is needed to stimu-late maximal T cell responses in the absence of CD4� T cell help. J.Immunol. 178, 2844–2852.
17. Wiesel, M., Kratky, W., Oxenius, A. (2011) Type I IFN substitutes for Tcell help during viral infections. J. Immunol. 186, 754–763.
18. Zimmerli, S. C., Harari, A., Cellerai, C., Vallelian, F., Bart, P. A., Panta-leo, G. (2005) HIV-1-specific IFN-�/IL-2-secreting CD8 T cells supportCD4-independent proliferation of HIV-1-specific CD8 T cells. Proc. Natl.Acad. Sci. USA 102, 7239–7244.
19. Bertram, E. M., Lau, P., Watts, T. H. (2002) Temporal segregation of4-1BB versus CD28-mediated costimulation: 4-1BB ligand influences Tcell numbers late in the primary response and regulates the size of theT cell memory response following influenza infection. J. Immunol. 168,3777–3785.
20. Halstead, E. S., Mueller, Y. M., Altman, J. D., Katsikis, P. D. (2002) Invivo stimulation of CD137 broadens primary antiviral CD8� T cell re-sponses. Nat. Immunol. 3, 536–541.
21. Liu, Y., Wenger, R. H., Zhao, M., Nielsen, P. J. (1997) Distinct costimu-latory molecules are required for the induction of effector and memorycytotoxic T lymphocytes. J. Exp. Med. 185, 251–262.
22. Lumsden, J. M., Roberts, J. M., Harris, N. L., Peach, R. J., Ronchese, F.(2000) Differential requirement for CD80 and CD80/CD86-dependentcostimulation in the lung immune response to an influenza virus infec-tion. J. Immunol. 164, 79–85.
23. Tan, J. T., Whitmire, J. K., Ahmed, R., Pearson, T. C., Larsen, C. P.(1999) 4-1BB ligand, a member of the TNF family, is important for thegeneration of antiviral CD8 T cell responses. J. Immunol. 163, 4859–4868.
24. Shahinian, A., Pfeffer, K., Lee, K. P., Kundig, T. M., Kishihara, K., Wake-ham, A., Kawai, K., Ohashi, P. S., Thompson, C. B., Mak, T. W. (1993)Differential T cell costimulatory requirements in CD28-deficient mice.Science 261, 609–612.
25. Zimmermann, C., Seiler, P., Lane, P., Zinkernagel, R. M. (1997) Antivi-ral immune responses in CTLA4 transgenic mice. J. Virol. 71, 1802–1807.
26. Dolfi, D. V., Duttagupta, P. A., Boesteanu, A. C., Mueller, Y. M., Oliai,C. H., Borowski, A. B., Katsikis, P. D. (2011) Dendritic cells and CD28costimulation are required to sustain virus-specific CD8� T cell re-sponses during the effector phase in vivo. J. Immunol. 186, 4599–4608.
27. Zhan, Y., Corbett, A. J., Brady, J. L., Sutherland, R. M., Lew, A. M.(2000) CD4 help-independent induction of cytotoxic CD8 cells to allo-geneic P815 tumor cells is absolutely dependent on costimulation. J. Im-munol. 165, 3612–3619.
28. Lee, B. O., Hartson, L., Randall, T. D. (2003) CD40-deficient, influenza-specific CD8 memory T cells develop and function normally in a CD40-sufficient environment. J. Exp. Med. 198, 1759–1764.
29. Bartholdy, C., Kauffmann, S. O., Christensen, J. P., Thomsen, A. R.(2007) Agonistic anti-CD40 antibody profoundly suppresses the immuneresponse to infection with lymphocytic choriomeningitis virus. J. Immu-nol. 178, 1662–1670.
30. Xu, J., Foy, T. M., Laman, J. D., Elliott, E. A., Dunn, J. J., Waldschmidt,T. J., Elsemore, J., Noelle, R. J., Flavell, R. A. (1994) Mice deficient forthe CD40 ligand. Immunity 1, 423–431.
31. Kawabe, T., Naka, T., Yoshida, K., Tanaka, T., Fujiwara, H., Suematsu,S., Yoshida, N., Kishimoto, T., Kikutani, H. (1994) The immune re-sponses in CD40-deficient mice: impaired immunoglobulin class switch-ing and germinal center formation. Immunity 1, 167–178.
32. Borriello, F., Sethna, M. P., Boyd, S. D., Schweitzer, A. N., Tivol, E. A.,Jacoby, D., Strom, T. B., Simpson, E. M., Freeman, G. J., Sharpe, A. H.(1997) B7-1 and B7-2 have overlapping, critical roles in immunoglobu-lin class switching and germinal center formation. Immunity 6, 303–313.
33. Hogquist, K. A., Jameson, S. C., Heath, W. R., Howard, J. L., Bevan,M. J., Carbone, F. R. (1994) T cell receptor antagonist peptides inducepositive selection. Cell 76, 17–27.
Seah et al. CD40L costimulation of primary anti-influenza CTL
www.jleukbio.org Volume 93, January 2013 Journal of Leukocyte Biology 153

34. Jenkins, M. R., Webby, R., Doherty, P. C., Turner, S. J. (2006) Additionof a prominent epitope affects influenza A virus-specific CD8� T cellimmunodominance hierarchies when antigen is limiting. J. Immunol.177, 2917–2925.
35. Tannock, G. A., Paul, J. A., Barry, R. D. (1984) Relative immunogenicityof the cold-adapted influenza virus A/Ann Arbor/6/60 (A/AA/6/60-ca), recombinants of A/AA/6/60-ca, and parental strains with similarsurface antigens. Infect. Immun. 43, 457–462.
36. Deliyannis, G., Boyle, J. S., Brady, J. L., Brown, L. E., Lew, A. M. (2000)A fusion DNA vaccine that targets antigen-presenting cells increases pro-tection from viral challenge. Proc. Natl. Acad. Sci. USA 97, 6676–6680.
37. La Gruta, N. L., Rothwell, W. T., Cukalac, T., Swan, N. G., Valkenburg,S. A., Kedzierska, K., Thomas, P. G., Doherty, P. C., Turner, S. J. (2010)Primary CTL response magnitude in mice is determined by the extentof naive T cell recruitment and subsequent clonal expansion. J. Clin.Invest. 120, 1885–1894.
38. Lau, Y. F., Deliyannis, G., Zeng, W., Mansell, A., Jackson, D. C., Brown,L. E. (2006) Lipid-containing mimetics of natural triggers of innate im-munity as CTL-inducing influenza vaccines. Int. Immunol. 18, 1801–1813.
39. Deliyannis, G., Jackson, D. C., Ede, N. J., Zeng, W., Hourdakis, I., Saka-betis, E., Brown, L. E. (2002) Induction of long-term memory CD8(�)T cells for recall of viral clearing responses against influenza virus. J.Virol. 76, 4212–4221.
40. Sedgwick, J. D., Holt, P. G. (1983) A solid-phase immunoenzymatic tech-nique for the enumeration of specific antibody-secreting cells. J. Immu-nol. Methods 57, 301–309.
41. Vremec, D., Shortman, K. (2008) The isolation and identification ofmurine dendritic cell populations from lymphoid tissues and their pro-duction in culture. Methods Mol. Biol. 415, 163–178.
42. Seah, S. G., Sutherland, R. M., La Gruta, N. L., Brown, L. E., Car-rington, E. M., Belz, G. T., Brady, J. L., Turner, S. J., Zhan, Y., Lew,A. M. (2011) The linear range for accurately quantifying antigen-specificT-cell frequencies by tetramer staining during natural immune re-sponses. Eur. J. Immunol. 41, 1499–1500.
43. Wiley, J. A., Hogan, R. J., Woodland, D. L., Harmsen, A. G. (2001) Anti-gen-specific CD8(�) T cells persist in the upper respiratory tract follow-ing influenza virus infection. J. Immunol. 167, 3293–3299.
44. Boise, L. H., Minn, A. J., Noel, P. J., June, C. H., Accavitti, M. A., Lind-sten, T., Thompson, C. B. (1995) CD28 costimulation can promote Tcell survival by enhancing the expression of Bcl-XL. Immunity 3, 87–98.
45. Maisch, T., Kropff, B., Sinzger, C., Mach, M. (2002) Upregulation ofCD40 expression on endothelial cells infected with human cytomegalovi-rus. J. Virol. 76, 12803–12812.
46. Vowinkel, T., Wood, K. C., Stokes, K. Y., Russell, J., Krieglstein, C. F.,Granger, D. N. (2006) Differential expression and regulation of murineCD40 in regional vascular beds. Am. J. Physiol. Heart Circ. Physiol. 290,H631–H639.
47. Bodmer, H., Obert, G., Chan, S., Benoist, C., Mathis, D. (1993) Environ-mental modulation of the autonomy of cytotoxic T lymphocytes. Eur. J.Immunol. 23, 1649–1654.
48. Tripp, R. A., Sarawar, S. R., Doherty, P. C. (1995) Characteristics of theinfluenza virus-specific CD8� T cell response in mice homozygous fordisruption of the H-2lAb gene. J. Immunol. 155, 2955–2959.
49. Mozdzanowska, K., Maiese, K., Gerhard, W. (2000) Th cell-deficientmice control influenza virus infection more effectively than Th- and Bcell-deficient mice: evidence for a Th-independent contribution by Bcells to virus clearance. J. Immunol. 164, 2635–2643.
50. Seah, S. G., Carrington, E. M., Ng, W. C., Belz, G. T., Brady, J. L.,Sutherland, R. M., Hancock, M. S., La Gruta, N. L., Brown, L. E.,Turner, S. J., Zhan, Y., Lew, A. M. (2012) Unlike CD4� T-cell help,CD28 costimulation is necessary for effective primary CD8� T-cell influ-enza-specific immunity. Eur. J. Immunol. 42, 1744–1754.
51. Bourgeois, C., Rocha, B., Tanchot, C. (2002) A role for CD40 expres-sion on CD8� T cells in the generation of CD8� T cell memory. Science297, 2060–2063.
52. Munroe, M. E., Bishop, G. A. (2007) A costimulatory function for T cellCD40. J. Immunol. 178, 671–682.
53. Meunier, S., Rapetti, L., Beziaud, L., Pontoux, C., Legrand, A., Tanchot,C. (2012) Synergistic CD40 signaling on APCs and CD8 T cells drivesefficient CD8 response and memory differentiation. J. Leukoc. Biol. 91,859–869.
54. Gett, A. V., Hodgkin, P. D. (2000) A cellular calculus for signal integra-tion by T cells. Nat. Immunol. 1, 239–244.
55. Janssen, E. M., Lemmens, E. E., Wolfe, T., Christen, U., von Herrath,M. G., Schoenberger, S. P. (2003) CD4� T cells are required for sec-ondary expansion and memory in CD8� T lymphocytes. Nature 421,852–856.
56. Shedlock, D. J., Shen, H. (2003) Requirement for CD4 T cell help ingenerating functional CD8 T cell memory. Science 300, 337–339.
57. Sun, J. C., Bevan, M. J. (2003) Defective CD8 T cell memory followingacute infection without CD4 T cell help. Science 300, 339–342.
58. Ahmed, K. A., Wang, L., Munegowda, M. A., Mulligan, S., Gordon, J. R.,Griebel, P., Xiang, J. (2012) Direct in vivo evidence of CD4� T cell re-quirement for CTL response and memory via pMHC-I targeting andCD40L signaling. J. Leukoc. Biol. 92, 289–300.
59. Feau, S., Arens, R., Togher, S., Schoenberger, S. P. (2011) AutocrineIL-2 is required for secondary population expansion of CD8(�) mem-ory T cells. Nat. Immunol. 12, 908–913.
60. Howland, K. C., Ausubel, L. J., London, C. A., Abbas, A. K. (2000) Theroles of CD28 and CD40 ligand in T cell activation and tolerance. J. Im-munol. 164, 4465–4470.
61. Ruter, J., Antonia, S. J., Burris, H. A., Huhn, R. D., Vonderheide, R. H.(2010) Immune modulation with weekly dosing of an agonist CD40 anti-body in a phase I study of patients with advanced solid tumors. CancerBiol. Ther. 10, 983–993.
62. Hussein, M., Berenson, J. R., Niesvizky, R., Munshi, N., Matous, J., So-becks, R., Harrop, K., Drachman, J. G., Whiting, N. (2010) A phase Imultidose study of dacetuzumab (SGN-40; humanized anti-CD40 mono-clonal antibody) in patients with multiple myeloma. Haematologica 95,845–848.
63. Furman, R. R., Forero-Torres, A., Shustov, A., Drachman, J. G. (2010) Aphase I study of dacetuzumab (SGN-40, a humanized anti-CD40 mono-clonal antibody) in patients with chronic lymphocytic leukemia. Leuk.Lymphoma 51, 228–235.
64. Advani, R., Forero-Torres, A., Furman, R. R., Rosenblatt, J. D., Younes,A., Ren, H., Harrop, K., Whiting, N., Drachman, J. G. (2009) Phase Istudy of the humanized anti-CD40 monoclonal antibody dacetuzumabin refractory or recurrent non-Hodgkin’s lymphoma. J. Clin. Oncol. 27,4371–4377.
KEY WORDS:cytotoxic T lymphocyte � dendritic cells � mixed leukocyte reaction
154 Journal of Leukocyte Biology Volume 93, January 2013 www.jleukbio.org




























