Embed Size (px)
Citation preview

JOURNAL OF ULTRASTRUCTURE AND MOLECULAR STRUCTURE RESEARCH 102, 249-254 (1989)
Influence of Exogenous Sucrose on the Greening of Oat’
GABRIELLA PASQUA,* FRANCESCA D&LA VECCHU,P NICOLETTA RASCIO,~ AND GIORGIO CASADORO~
*Dipartimento di Biologia Vegetnle, lJniversitc2 “La Sapienza”, Roma, and fDipartimento di Biologia, Universiti di Paduva, Padova, Italy
Received June 21, 1989, and in revised form August 2, 1989
The greening of roots and leaves has been studied in whole oat seedlings grown on White’s medium either with or without 2% sucrose. The added nutrient promotes chlo- rophyll synthesis and chloroplast differentiation in the roots. Yet it manifests a negative effect in the folk tis- sues where it accelerates the decline in chlorophyll as well BS the chloroplast ultrastructural alterations usually associated with senescence. The negative effect of the nutrient in the leaves is probably a consequence of the addition of exogenous sucrose to the endogenous sugars produced by photosynthesis. The foliar tissues would therefore be in the presence of high sucrose concentra- tions, which are known to be harmful for the photosyn- thetic apparatus. SDS-PAGE analysis of thylakoid poiy peptides from root and leaf chloroplasts has revealed or- gan-specific differences in the electrophoretic patterns. 0 1989 Academic Press, Inc.
INTRODUCTION
It has been shown by many researchers that roots can differentiate photosynthetically active chloro- plasts when grown in the light. However, it seems that the greening must be imposed by long light exposures (Kirk and Tilney-Bassett, 1978).
Observed in cross section, the chloroplast differ- entiation does not take place uniformly throughout the root in different species. For instance, in wheat and barley roots the chloroplasts are usually re- stricted to the innermost cortical layers, while in flax they develop all over the cortical parenchyma (Fadeel, 1962). Whatever the case, both the number of chloroplasts and the amount of chlorophyll are much lower than in the leaves (Kirk and Tilney- Bassett, 1978).
The fine structure of the root chloroplasts ob- served in some species (Heltne and Bonnett, 1970; Wroblewski, 1973; Oliveira, 1982; Hldyer-Hansen et al., 1989) is quite similar to that of the correspond-
l N. Raacio and G. Casadoro dedicate this paper to Professor Mario Oreenigo on the occasion of his 65th birthday.
ing leaf chloroplasts. However, despite this similar- ity, in lemon the root thylakoid polypeptide pattern exhibits some peculiarities (H$yer-Hansen et aE., 1989), thus suggesting an expression organ-specific of some polypeptides.
Many researches on the root greening were car- ried out with cultured excised roots. In 1934 White demonstrated the importance of exogenous sucrose for growing excised roots of tomato (review in Weston, 1975). This importance was also demon- strated by R&her et al. (1976) while studying the differentiation of roots from cultured leaf explants of Digitalis purpurea L.
More recently, Kumar etal. (1983,1984) found out that exogenous sucrose aids the development of a photosynthetic apparatus in cultured explants of carrot roots. However, the amount is critical since excess or high concentrations of sucrose have an in- hibitory effect, as already seen with Dig&&s pur- purea cultured explants (Riicher et aZ., 1976). The optimal amount appears to be 2% sucrose (Kumar et al., 1984; Dodds and Roberts, 1985). In some cases exogenous sucrose was also shown to have a protec- tive effect on plastid senescence (review in De Vec- chi, 1971; Rascio et al., 1988).
Research dealing with the effects of exogenous su- crose on the plastid differentiation have usually been performed separately on excised leaves or roots. These experimental models bring in a number of unknown variables which can add to the specific effects of the supplied nutrient.
Since roots and leaves exhibit a different tendency to become green, we thought it of some interest to study the effects of exogenous sucrose on the green- ing of both roots and leaves of intact seedlings which represent an experimental system where all the in- teractions sustaining the developmental processes are always maintained.
MATERIALS AND METHODS
Plunt material. Kernels of Avenu sativa L. cv. Perona were surface-sterilized using sodium hypochlorite, throug%ly rinsed in sterile distilled water, and then germinated on filter paper moist-
249 0889-1606/99 $3.00
Copyright 6 1989 by Academic Prem, Inc. All rights of reproduction in any form reserved.
250 PASQUA ET AL.
ened with sterile distilled water in petri dishes. Germination was carried out for 3 days in the dark at 26°C. Three roots, 6-8 cm long, had usually developed from each kernel.
Cultures. The 3-day-old seedlings were divided into two groups and cultured in flasks containing modified White’s me- dium (Thomas and Davey, 1975) either with or without 2% su- cmse. The two different types of cultures were kept in a growth chamber at 26°C under continuous light. The light source con- sisted of 6 x F58W Sylvania Growlux and 15 x TLD 58W/33 Phil- ips Cool White. The photon fluence rate was 100 ~.LE . mm2 . set-’ and it was measured by a LI-COR L1185B Quantum Radiometer Photometer at the sample level.
Electron microscopy. Tissue samples collected from the root and from the first leaf were fixed and prepared for transmission electron microscopy observations as in Rascio et al. (1979).
Chlorophyll determination. Chlorophylls were determined spectmphotometrically according to Moran and Porath (1980) us- ing the extinction coefficients proposed by Inskeep and Bloom (1985).
Thylakoid membrane extraction and SDS-PAGE analy- sis. Thylakoid membranes were extracted and purified by flotation according to Chua and Bennoun (1975). Analysis by SDS-PAGE was carried out in slab gels with a 7.5-1546 linear gradient. Apparent molecular weights of protein bands were de- termined by comparison with a set of low-molecular-weight markers (phosphorylase b: 92,500; bovine serum albumin: 66,200; ovoalbumin: 45,000; carbonic anhydrase: 31,000; soybean trypsin inhibitor: 21,500; and lysozyme: 14,400; all from Bio-Rad Rich- mond, CA). The terminology of the chlorophyll proteins is accord- ing to Machold et aZ. (1979).
RESULTS
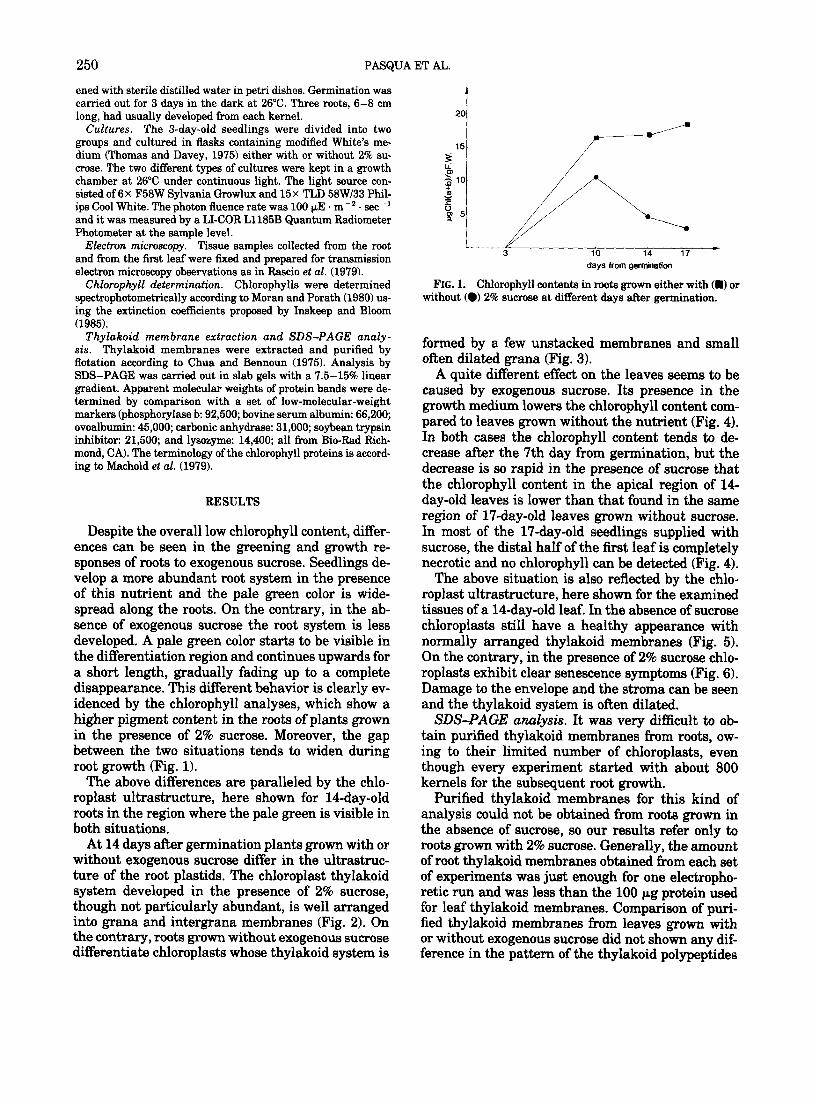
Despite the overall low chlorophyll content, differ- ences can be seen in the greening and growth re- sponses of roots to exogenous sucrose. Seedlings de- velop a more abundant root system in the presence of this nutrient and the pale green color is wide- spread along the roots. On the contrary, in the ab- sence of exogenous sucrose the root system is less developed. A pale green color starts to be visible in the differentiation region and continues upwards for a short length, gradually fading up to a complete disappearance. This different behavior is clearly ev- idenced by the chlorophyll analyses, which show a higher pigment content in the roots of plants grown in the presence of 2% sucrose. Moreover, the gap between the two situations tends to widen during root growth (Fig. 1).
The above differences are paralleled by the chlo- roplast ultrastructure, here shown for lCday-old roots in the region where the pale green is visible in both situations.
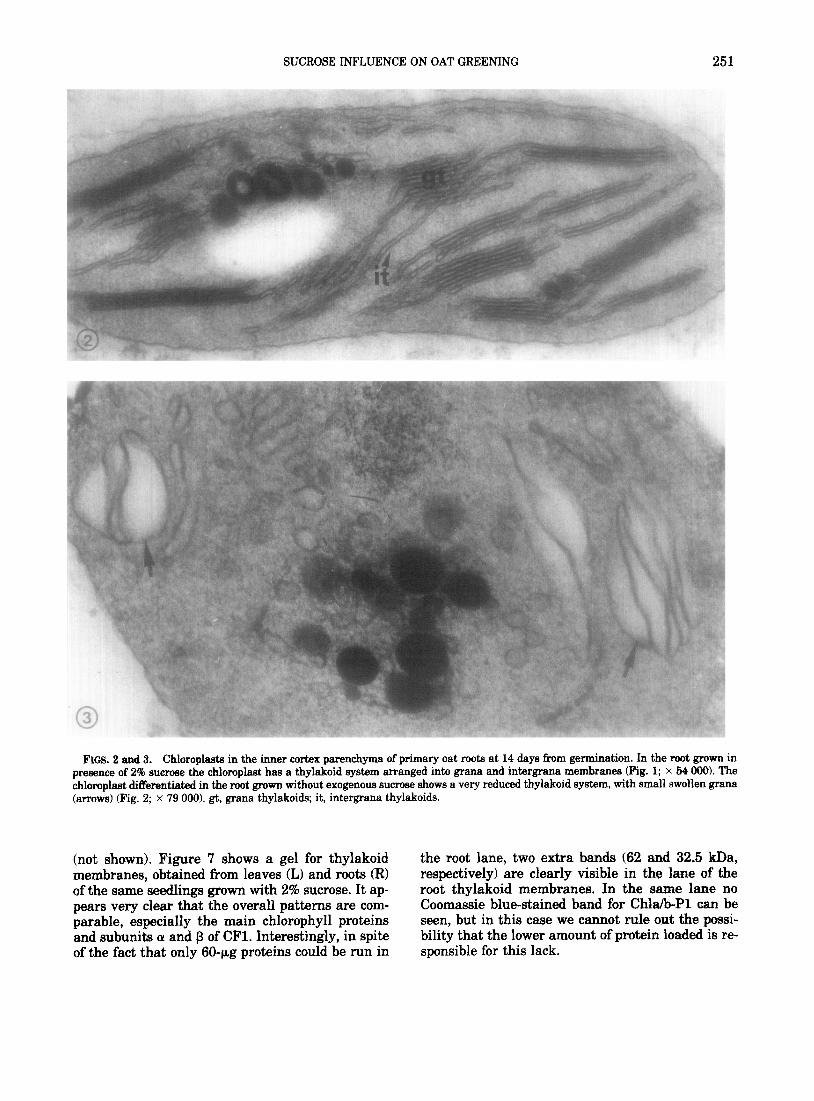
At 14 days after germination plants grown with or without exogenous sucrose differ in the ultrastruc- ture of the root plastids. The chloroplast thylakoid system developed in the presence of 2% sucrose, though not particularly abundant, is well arranged into grana and intergrana membranes (Fig. 2). On the contrary, roots grown without exogenous sucrose differentiate chloroplasts whose thylakoid system is
10 14 17 - days from gemWation
FIG. 1. Chlorophyll contents in roots grown either with (ml or without (0) 2% sucmse at different days after germination.
formed by a few unstacked membranes and small ofien dilated grana (Fig. 3).
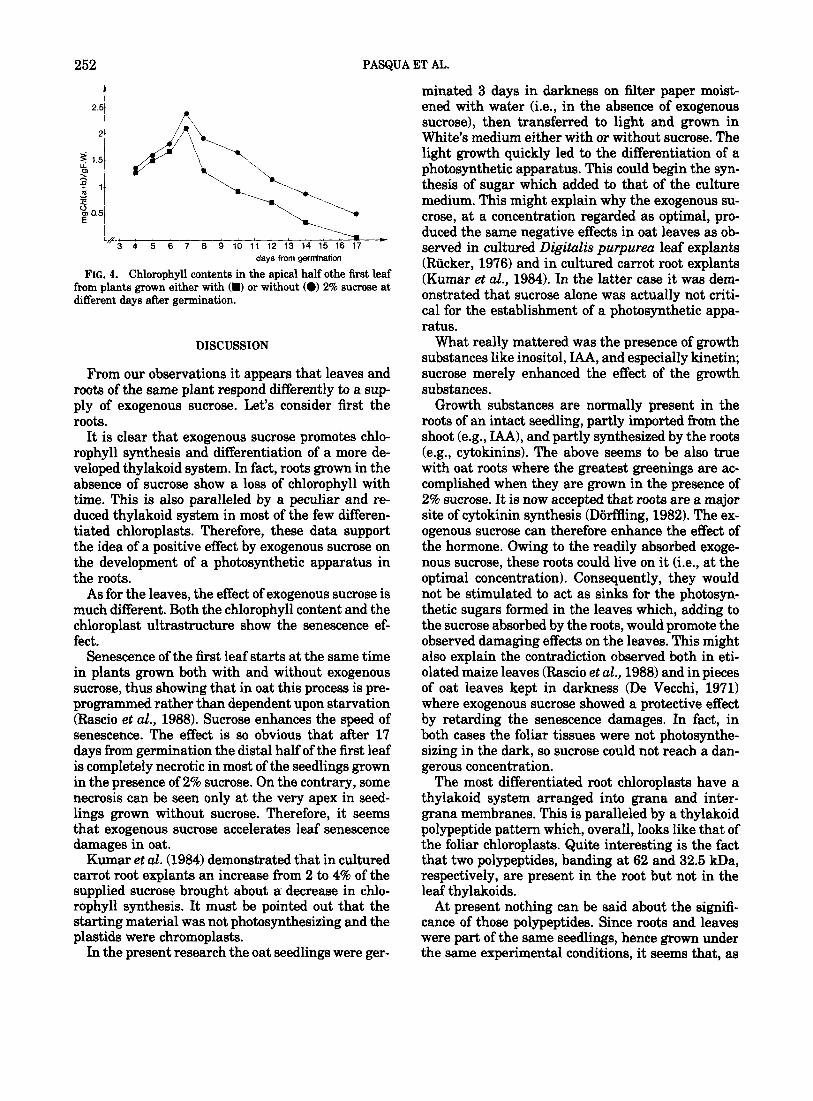
A quite different effect on the leaves seems to be caused by exogenous sucrose. Its presence in the growth medium lowers the chlorophyll content com- pared to leaves grown without the nutrient (Fig. 4). In both cases the chlorophyll content tends to de- crease after the 7th day from germination, but the decrease is so rapid in the presence of sucrose that the chlorophyll content in the apical region of 14- day-old leaves is lower than that found in the same region of 17-day-old leaves grown without sucrose. In most of the 17-day-old seedlings supplied with sucrose, the distal half of the first leaf is completely necrotic and no chlorophyll can be detected (Fig. 4).
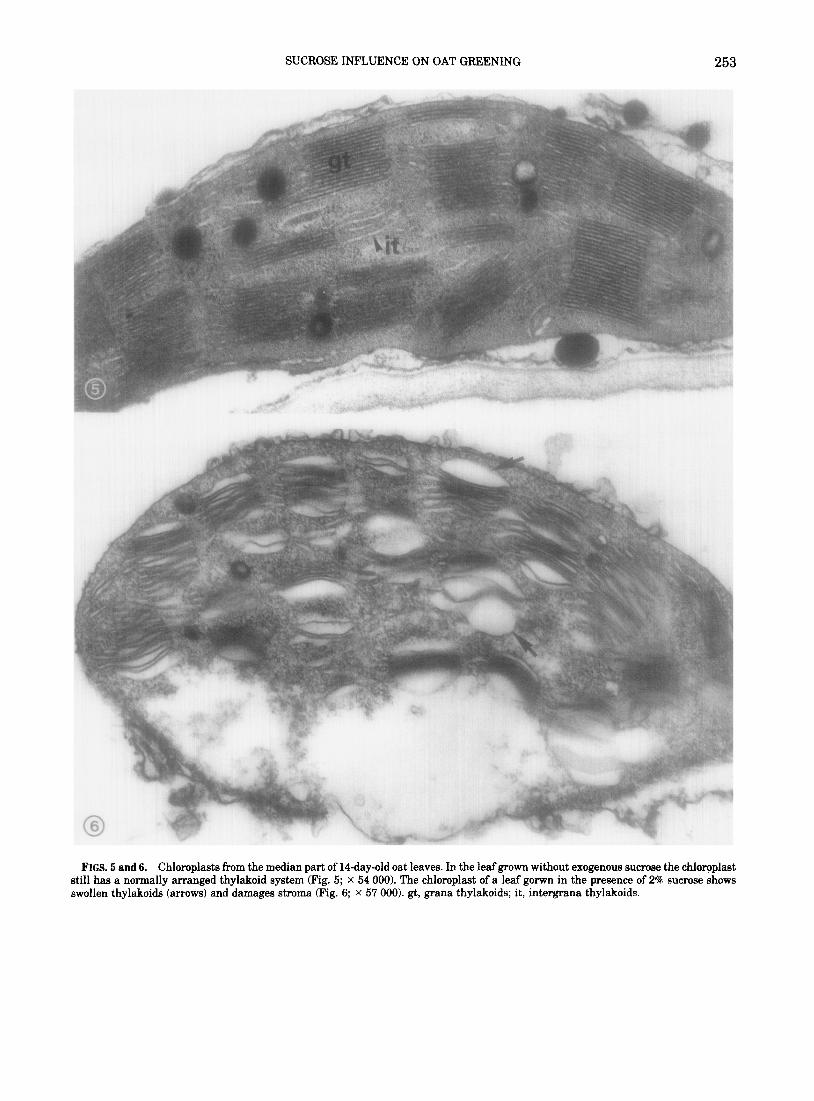
The above situation is also reflected by the chlo- roplast ultrastructure, here shown for the examined tissues of a lCday-old leaf. In the absence of sucrose chloroplasts still have a healthy appearance with normally arranged thylakoid membranes (Fig. 5). On the contrary, in the presence of 2% sucrose chlo- roplasts exhibit clear senescence symptoms (Fig. 6). Damage to the envelope and the stroma can be seen and the thylakoid system is often dilated.
SDS-PAGE analysis. It was very difficult to ob- tain purified thylakoid membranes from roots, ow- ing to their limited number of chloroplasts, even though every experiment started with about 800 kernels for the subsequent root growth.
Purified thylakoid membranes for this kind of analysis could not be obtained from roots grown in the absence of sucrose, so our results refer only to roots grown with 2% sucrose. Generally, the amount of root thylakoid membranes obtained from each set of experiments was just enough for one electropho- retie run and was less than the 100 pg protein used for leaf thylakoid membranes. Comparison of puri- fied thylakoid membranes from leaves grown with or without exogenous sucrose did not shown any dif- ference in the pattern of the thylakoid polypeptides
SUCROSE INFLUENCE ON OAT GREEMNG 251
FIGS. 2 and 3. Chloroplasts in the inner cortex parenchyma of primary oat roots at 14 days from germination. In the root grown in presence of 2% sucrose the chloroplast has a thylakoid system arranged into grana and intergrana membranes (Fig. 1; x 54 990). The chloroplast differentiated in the root grown without exogenous sucrose shows a very reduced thylakoid system, with small swollen grana (arrows) (Fig. 2; x 79 000). gt, grana thylakoids; it, intergrana thylakoids.
(not shown). Figure 7 shows a gel for thylakoid the root lane, two extra bands (62 and 32.5 kDa, membranes, obtained from leaves (L) and roots (R) respectively) are clearly visible in the lane of the of the same seedlings grown with 2% sucrose. It ap- root thylakoid membranes. In the same lane no pears very clear that the overall patterns are com- Coomassie blue-stained band for Chla/b-Pl can be parable, especially the main chlorophyll proteins seen, but in this case we cannot rule out the possi- and subunits (Y and j3 of CFl. Interestingly, in spite bility that the lower amount of protein loaded is re- of the fact that only 60-pg proteins could be run in sponsible for this lack.
252 PASQUA ET AL.
L I 77-b - 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
days from germination
FIG. 4. Chlorophyll contents in the apical half othe first leaf from plants grown either with (m) or without (0) 2% sucrose at different days after germination.
DISCUSSION
From our observations it appears that leaves and roots of the same plant respond differently to a sup- ply of exogenous sucrose. Let’s consider first the roots.
It is clear that exogenous sucrose promotes chlo- rophyll synthesis and differentiation of a more de- veloped thylakoid system. In fact, roots grown in the absence of sucrose show a loss of chlorophyll with time. This is also paralleled by a peculiar and re- duced thylakoid system in most of the few differen- tiated chloroplasts. Therefore, these data support the idea of a positive effect by exogenous sucrose on the development of a photosynthetic apparatus in the roots.
As for the leaves, the effect of exogenous sucrose is much different. Both the chlorophyll content and the chloroplast ultrastructure show the senescence ef- fect.
Senescence of the first leaf starts at the same time in plants grown both with and without exogenous sucrose, thus showing that in oat this process is pre- programmed rather than dependent upon starvation (Rascio et al., 1988). Sucrose enhances the speed of senescence. The effect is so obvious that after 17 days from germination the distal half of the first leaf is completely necrotic in most of the seedlings grown in the presence of 2% sucrose. On the contrary, some necrosis can be seen only at the very apex in seed- lings grown without sucrose. Therefore, it seems that exogenous sucrose accelerates leaf senescence damages in oat.
Kumar et al. (1984) demonstrated that in cultured carrot root explants an increase from 2 to 4% of the supplied sucrose brought about a decrease in chlo- rophyll synthesis. It must be pointed out that the starting material was not photosynthesizing and the plastids were chromoplasts.
In the present research the oat seedlings were ger-
minated 3 days in darkness on filter paper moist- ened with water (i.e., in the absence of exogenous sucrose), then transferred to light and grown in White’s medium either with or without sucrose. The light growth quickly led to the differentiation of a photosynthetic apparatus. This could begin the syn- thesis of sugar which added to that of the culture medium. This might explain why the exogenous su- crose, at a concentration regarded as optimal, pro- duced the same negative effects in oat leaves as ob- served in cultured Digitalis purpurea leaf explants (Riicker, 1976) and in cultured carrot root explants (Kumar et al., 1984). In the latter case it was dem- onstrated that sucrose alone was actually not criti- cal for the establishment of a photosynthetic appa- ratus.
What really mattered was the presence of growth substances like inositol, IAA, and especially kinetin; sucrose merely enhanced the effect of the growth substances.
Growth substances are normally present in the roots of an intact seedling, partly imported from the shoot (e.g., IAA), and partly synthesized by the roots (e.g., cytokinins). The above seems to be also true with oat roots where the greatest greenings are ac- complished when they are grown in the presence of 2% sucrose. It is now accepted that roots are a major site of cytokinin synthesis (DiirfAing, 1982). The ex- ogenous sucrose can therefore enhance the effect of the hormone. Owing to the readily absorbed exoge- nous sucrose, these roots could live on it (i.e., at the optimal concentration). Consequently, they would not be stimulated to act as sinks for the photosyn- thetic sugars formed in the leaves which, adding to the sucrose absorbed by the roots, would promote the observed damaging effects on the leaves. This might also explain the contradiction observed both in eti- olated maize leaves (Rascio et al., 1988) and in pieces of oat leaves kept in darkness (De Vecchi, 1971) where exogenous sucrose showed a protective effect by retarding the senescence damages. In fact, in both cases the foliar tissues were not photosynthe- sizing in the dark, so sucrose could not reach a dan- gerous concentration.
The most differentiated root chloroplasts have a thylakoid system arranged into grana and inter- grana membranes. This is paralleled by a thylakoid polypeptide pattern which, overall, looks like that of the foliar chloroplasts. Quite interesting is the fact that two polypeptides, banding at 62 and 32.5 kDa, respectively, are present in the root but not in the leaf thylakoids.
At present nothing can be said about the signifi- cance of those polypeptides. Since roots and leaves were part of the same seedlings, hence grown under the same experimental conditions, it seems that, as
SUCROSE INFLUENCE ON OAT GREENING 253
FIGS. 5 and 6. Chloroplasts from the median part of 14-day-old oat leaves. In the leafgrown without exogenous sucrose the chloroplast still has a normally arranged thylakoid system (Fig. 5; x 54 000). The chloroplast of a leaf gorwn in the presence of 2% sucrose shows swollen thylakoids (arrows) and damages stroma (Fig. 6; x 57 000). gt, grana thylakoids; it, intergrana thylakoids.

254 PASQUA ET AL.
R L
97,400- w -c+ -Chla -PI
66,200-
42,700-
21,500-
14,400-
, (tCF1 -62 kDa ’ pCF1
,32.5 kDa -Chla/b-PI \ C hla/b-P2
FIG. 7. Electrophoregram of chloroplast thylakoid polypep- tides from roots (R) and leaves (L) of plants grown with 2% su- crose. Note in the root lane the two extra bands of 62 and 32.5 kDa. The Chla/b-Pl present in the leaf lane is not detectable in the root one.
already observed with other thylakoid polypeptides in lemon root chloroplasts (Hflyer-Hansen et al., 1989), in oat root the different membrane com-
position can be regarded as the result of an or- gan-specific expression of genes for chloroplast pro- teins.
Dr. Pessali of the Institute Sperimentale per la Cerealiooltura (Badia Polesine, Rovigo) is thanked for the gift of the oat kernels. This work was supported by a Grant from M.P.I.
REFERENCES CHUA, N. H., AND BENNOUN, P. (1975) Proc. Natl. Acad. Sci. USA
72, 2175-2179. DE VECCHI, L. (1971) Zsmel. J. Rot. 169, 169-183. DODD& J. H., AND ROBERTS, L. W. (1985) Experiments in Plant
Tissue Culture, 2nd ed., Cambridge Univ. Press, London/New York.
D(IRFFLING, K. (1982) Das Hormonsystem der Pflanzen, George Thieme Verlag, Stuttgart New York.
FADEEL, A. A. (1962) Physoil. Plant. 15, 130-147. HELTNE, J., AND BONNETT, H. T. (1970) Pkzntu 92, 1-12. HBYER-HANSEN, G., RASCIO, N., AND CASADORO, G. (1989)
Pluntu (submitted). INSKEEP, W. P., AND BLOOM, P. R. (1985) Plant Physid. 47,483-
485. KIRK, J. T. O., AND TILNEY-BASSETT, R. A. E. (1978) The Plas-
tids, their Chemistry, Structure, Growth and Inheritance, Elsevier, Amsterdam.
KUMAR, A., BENDER, L., AND NEUMANN, K. H. (1984) Plant Cell, Tissue Organ Cult. 3, 11-28.
KUMAR, A., BENDER, L., PAULER, B., NEUMANN, K. H., SENGER, H., AND JESKE, C. (1983) Plant Cell, Tissue Organ Cult. 2, 161-177.
MACHOLD, O., SIMPSON, D. J., AND MPILLER, B. L. (1979) Carls- berg Res. Commun. 44,235-254.
MORAN, R., AND PORATH, D. (1980) Plant Physid. 65, 47M79. OLNElR.4, L. (1982) New Phytol. 91, 263-275. FtAsCIo, N., CASADORO, G., AND DI CHIo, L. (1979) Protoplasmu
109,45-52. hWX0, N., MARIANI, P., DALLA VECCHlA, F., AND CHITANO, P.
(1988) J. Ultra&u&. Mokc. Struct. Res. 99, 226-233. ROCKER, W., JENTZSCH, K., AND WICHTL, M. (1976) 2. Pfbzen-
physiol. 80, 323-335. THOMAS, E., AND DAVEY, M. R. (1975) From Single Cells to
Plants, Wyheham Publ., London. WESTON, D. G. (1975) Can&. J. Rot. 53, 1769-1773. WHITE, P. R. (1934) PIant Physiol. 9, 585-600. WROBLEWSKI, R. (1973) J. Submicrosc. Cytd. 5,97-105.





























