Embed Size (px)
Citation preview

This article was downloaded by: [UNIVERSITY OF ADELAIDELIBRARIES]On: 04 April 2013, At: 18:35Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number:1072954 Registered office: Mortimer House, 37-41 Mortimer Street,London W1T 3JH, UK
Bioacoustics: TheInternational Journalof Animal Sound and itsRecordingPublication details, including instructionsfor authors and subscription information:http://www.tandfonline.com/loi/tbio20
INDIVIDUAL VOCAL TRAITSOF MOTHER AND PUP FURSEALSBRAD PAGE a , SIMON D. GOLDSWORTHY a &MARK A. HINDELL aa Antarctic Wildlife Research Unit, Schoolof Zoology, University of Tasmania, Hobart,Tasmania, AustraliaVersion of record first published: 13 Apr2012.
To cite this article: BRAD PAGE , SIMON D. GOLDSWORTHY & MARK A.HINDELL (2002): INDIVIDUAL VOCAL TRAITS OF MOTHER AND PUP FUR SEALS,Bioacoustics: The International Journal of Animal Sound and its Recording,13:2, 121-143
To link to this article: http://dx.doi.org/10.1080/09524622.2002.9753491
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private studypurposes. Any substantial or systematic reproduction, redistribution,

reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or makeany representation that the contents will be complete or accurateor up to date. The accuracy of any instructions, formulae, and drugdoses should be independently verified with primary sources. Thepublisher shall not be liable for any loss, actions, claims, proceedings,demand, or costs or damages whatsoever or howsoever causedarising directly or indirectly in connection with or arising out of theuse of this material.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

Bioacoustics The International Journal of Animal Sound and its Recording, 2002, Vol. 13, pp. 121~ 143 0952-4622/02 $10 © 2002 AB Academic Publishers
INDIVIDUAL VOCAL TRAITS OF MOTHER AND PUP FUR SEALS
BRAD PAGE*t, SIMON D. GOLDSWORTHYt AND MARK A. HINDELL
Antarctic Wildlife Research Unit, School of Zoology, University of Tasmania, Hobart, Tasmania, Australia.
ABSTRACT
This study investigates individual vocal traits in three species of fur seal: Antarctic fur seal Arctocephalus gazella, Subantarctic fur seal A. tropicalis, New Zealand fur seal A. forsteri and their hybrids. The attraction calls of mothers and pups were highly stereotyped within each individual and markedly different between individuals. Pup attraction calls (emitted by females) were more variable between individual A. tropicalis and A. forsteri than between individual hybrids and A. gazella. However, such a pattern was not evident for female attraction calls (emitted by pups). Pup attraction calls and female attraction calls were both generally tonal calls overlain with regions of pulses. Female attraction calls had relatively high fundamental frequencies. The importance of individual vocalisations for mothers and pups is best appreciated in relation to maternal fitness and offspring survival: individual vocalisations are favoured by natural selection as they facilitate rapid reunion when mothers return from foraging trips, ensuring maternal resources are not invested in unrelated pups.
Keywords: Vocalisation, Arctocephalus, fur seal, individual recognition
INTRODUCTION
The ability to recognise offspring is critical if mobile young intermingle when parents leave them to search for food. Parents need to be able to discriminate between their offspring and unrelated young and locate them when they return from foraging. Therefore in colonial animals there should be strong selection for offspring recognition, if recognition
*Corresponding author; email: [email protected] tCurrent address: Sea Mammal Ecology Group, Department of Zoology, La Trobe University, Melbourne, Victoria, Australia.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

122
errors decrease the future reproductive success of the offspring (Scherrer and Wilkinson 1993).
Although a range of cues (olfactory, visual and tactile) is available to mediate reunions between parents and offspring, vocalisations are the primary cues used by many species (Stoddard and Beecher 1983). To achieve individual recognition by vocalisations, variation within an individual's repertoire must be less than the variation between individuals. There are many examples where individually distinct calls have been shown (e.g. pinnipeds (Van Parijs 2000), deer (Reby et al. 1998), canids (Frommolt et al. 1997), bats (Scherrer and Wilkinson 1993), cetaceans (Smolker et al. 1993), pigs (Blackshaw et al. 1996), primates (Hammerschmidt and Fischer 1998), anurans (Nairns et al. 1998), birds (Wanker et al. 1998), insects (Schatral 1990)), suggesting that recognition is often mediated by vocal behaviour. Individual recognition of vocalisations can be demonstrated when animals recognise and react to different animals' vocalisations (e.g. pinnipeds (Trillmich 1981), birds (Wanker et al. 1998), primates (Hammerschmidt and Fischer 1998)). While olfactory, visual and tactile cues are undoubtedly important in parent-offspring recognition, the most practical sense to locate an individual over large distances in crowded colonies is through auditory signals (Trillmich 1981, Aubin and Jouventin 1998).
Most otariids (fur seals and sea lions) breed in densely populated colonies where many thousands of pups may be born in a season. As males do not assist with parental care, recognition between mother and pup is a critical aspect of maternal care in these species. Mothers begin calling to their pups (pup attraction call: PAC) during or shortly after birth and pups respond with their female attraction call (FAC). Calling is very intense following birth and is likely driven by the need for mother and pup to imprint on each others' calls (Charrier et al. 2001). Pups remain with their mother for seven to 10 days before the mother leaves the colony to forage. When mothers return from foraging trips they come ashore and begin to make PACs. Playback studies confirm that it is generally the mother's pup that responds to her PACs (Trillmich 1981). Mothers and offspring locate each other in this way many times over the course of lactation (Insley 1992).
Macquarie Island provides a unique opportunity to study fur seal vocalisations as two species breed there: antarctic Arctocephalus gazella and subantarctic A. tropicalis fur seals. Also male New Zealand fur seals A. forsteri haul out at Macquarie Island and some challenge for territories, while females give birth there occasionally (Goldsworthy unpublished data). Inter-specific matings involving all three species have been observed and hybrid fur seals of all age and sex classes are common (Goldsworthy et al. 1999).
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

123
Following the expectation that vocalisations are important in individual recognition for mother-pup pairs, this study tested the hypothesis that variation in PACs and FACs between individuals (within each species) would be greater than within individuals. As such, the aims of this study were firstly to determine whether calls of mothers and pups show sufficient stereotypy to enable individual recognition and ultimately to compare individual recognition potential between species and offer suggestions to investigate recognition in species with extreme levels of call stereotypy.
METHODS
Classification of species and hybrids
Animals were classified as A. gazella, hybrids, A. tropicalis or A. forsteri before analyses commenced. Mitochondrial genotype (where known) and a range of phenotypic characters were used to identify species (Goldsworthy et al. 1999, Wynen et al. 2000).
Recording techniques
Recordings were made on Sony DT-120RA digital audio tapes using a Sennheiser MKH 815T directional microphone powered by an external 12 volt battery pack and a Sony Digital Audio Tape-Corder (model TCD-DS). Vocalisations were sampled at 44.1 kHz by the DATCorder. Digitised vocalisations have been deposited at the Sound Library of the Australian National Wildlife Collection, CSIRO, in Canberra.
Recordings were made at two locations. On Macquarie Island (54°49'30"S, 158°94'48"E) recordings of A. gazella, hybrids and A. tropicalis were made on the North Head peninsula at Secluded Beach, Goat Bay and Aerial Cove. Arctocephalus forsteri calls were recorded at Victory Beach on the Otago Peninsula in New Zealand (45°49'03"S, 173°44'20"E).
Recordings of mothers and pups were made while they vocalised to locate each other. The recordings were made between one and ten metres from the vocalising animal without affecting their behaviour. Animals' tag numbers were noted while recording. Up to forty vocalisations were recorded from each animal to obtain sufficient vocalisations with high signal to noise ratios for acoustic analyses.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

124
Acoustic analyses
Spectral analyses were carried out using Canary V. 1.2.1 (Bioacoustics Research Program, Cornell Laboratory of Ornithology, USA). Calls were played into the computer at the same tape speed they were recorded, sampled at 44.1 kHz (frequency range 0-22 kHz). Temporal and spectral characteristics were examined on spectrograms and power spectra after performing 512-point Fast Fourier Transforms (FFT) (time resolution = 5.805 msec, frequency resolution = 86.13 Hz). Noise below -75 dB was not represented in spectrums. Contrast and brightness controls in Canary were adjusted to examine structural features on spectrograms, such as frequency modulation. Up to ten vocalisations per individual were randomly selected for analysis.
The acoustic features (variables) measured and the criteria used in measuring each variable are described in Phillips and Stirling (2000). Variation in cursor placement was evaluated by repeating three measurements ten times on structural components of waveforms, spectrograms and power spectra. The ranges of cursor errors were: 1) waveforms (DUR): ±1 msec, 2) spectrograms (INF, MAXF, HI, ENDF, FMR and FMP): ±1 msec and ±13 Hz, 3) power spectra (RAMP, PEAK): ±0.3 dB and ±3 Hz.
Statistical analyses
The number of parts per call (NP) was square root transformed to improve normality before analyses (Sakal & Rohlf, 1981, Phillips & Stirling, 2000). To control for interdependence a maximum of five calls were analysed from a single bout. The categorical variables TONAL, PULSE and FM were combined into a new variable describing call SHAPE, consisting of five possible categories: 1) TONAL, 2) PULSE, 3) TONAL+ FM, 4) TONAL+ PULSE, 5) TONAL+ PULSE+ FM. The variables used in the multivariate analyses of female and pup calls were SHAPE, NP, DUR, INF, MAXF, ENDF, PEAK1, PEAK2, PEAK3, RAMP1 and RAMP2.
Semi-strong-hybrid multi-dimensional scaling (MDS) was employed for multivariate analyses in three dimensions using the PATN Analysis Package (Division of Wildlife & Ecology, CSIRO, Canberra, Australia). The Bray and Curtis association measure was used for the analysis (Beals 1984). Principal axes correlation was used to determine the most influential variables in MDS analyses. Principal axes correlation takes the acoustic variables and finds the location of the best-fitted vector in the ordination space. Variables with significant correlation coefficients, r, were recorded with the a level of statistical significance set at 0.05. Stress values were calculated in two dimensions, to give an accurate indication of how well the data were represented during transformation. The stress in an MDS is a measure
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

125
of how distorted (or scattered) the data are after being confined to a limited number of vectors. The lower the stress value, the better the MDS representation of the data. Generally, stress values of less than 0.10 are regarded as being unlikely to result in misinterpretation of the data (Schiffman et al. 1981). Half of the individuals' calls were removed from each MDS plot to improve the clarity of the 2D figures.
Discriminant function analyses (DFA) were used to test for differences between individuals' vocalisations. DFA indirectly indicates the variation between individuals by showing the proportion of calls that are correctly allocated back to individuals based on the acoustic variables. To facilitate comparison with Phillips and Stirling (2000), a stepwise (Jack-knifed) DFA was used with minimisation of Wilk's A (F1N=3.84; F ouT=2. 71) as the selection criterion for variables to be included in a function.
A one-way ANOVA was used on each acoustic feature to examine absolute differences in structures between individuals' PACs and F ACs for each species. After a Bonferonni correction to the significance level (P value/number of comparisons) the a level of significance was set for A. gazella, hybrids, A. tropicalis and A. forsteri: PACs: 0.005, 0.004, 0.005, 0.008; and FACs: 0.006, 0.004, 0.005, 0.005.
To examine intra-individual variation, coefficients of variation (CV) were calculated for each acoustic structure per individual (Sokal and Braumann 1980, Insley 1992, Phillips and Stirling 2000). Variation between individuals was explored by calculating the added variance component (s2A = group mean squares minus error sum of squares) between groups (Sokal and Rohlf 1981, Phillips and Stirling 2000). Levels of inter-individual variation (s2 A) were compared with levels of intra-individual variation (CV) using Wilcoxon signed-ranks tests to determine whether inter-individual variability was greater for PACs, and FACs, of each species, than intra-individual variability.
RESULTS
Description of pup attraction calls (PACs)
Up to 10 PACs from 10 A. gazella (N = 100), 12 hybrid (N = 117), 10 A. tropicalis (N = 100) and six A. forsteri (N = 34) females (overall N = 351) were analysed (Table 3) out of the total 891 PACs recorded. Only six A. forsteri females were recorded and four of these individuals were analysed with less than 10 PACs (Table 3). Although a smaller sample decreases the level of variation shown for these females (Figure 2) the multivariate analyses were chosen as they provide robust measures of similarities despite the unequal samples. The additional 450 calls were not analysed as preliminary MDS and DFA
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

126
analyses indicated that 10 calls per individual were adequate to indicate statistical significance. Summary statistics and representative spectrograms of A. gazella, hybrids, A. tropicalis and A. forsteri PACs are presented in Table 1 and Figure 1 respectively.
PACs of the three species recorded in this study were similar to those previously described for A. tropicalis (Roux and Jouventin 1987, Goldsworthy et al. 1999), A. gazella (Shaughnessy and Fletcher 1987, Goldsworthy et al. 1999) and A. forsteri (Stirling 1970 and 1971, Stirling and Warneke 1971, Brown 1974). Arctocephalus gazella calls (Figure 1) all contained tonal regions and had relatively high fundamental frequencies. Most A. gazella contained more than one part, largely as a result of the presence of PULSED or FM regions. PULSED and FM regions commonly made new parts of the calls and made the calls sound like high pitched trills. Hybrids' calls (Figure 1) showed characteristics of A. gazella, A. tropicalis and A. forsteri in categorical, frequency and amplitude measurements. Most hybrids' calls had few parts (one female skewed mean values) and FM was present in very few calls. Arctocephalus tropicalis calls (Figure 1) tended toward a purely tonal structure often consisting of one tonal part overlain by PULSES that did not constitute new parts. Arctocephalus tropicalis calls had relatively low fundamental frequencies and FM was not present in many calls, making them sound like low frequency moans. Arctocephalus forsteri (Figure 1) calls were made up of many parts, often composed of several pulsed regions followed by a louder tonal region similar to those described by Phillips and Stirling (2000) for South American fur seal, A. australis, females. Arctocephalus forsteri calls had relatively low fundamental frequencies and one female gave some entirely PULSED calls.
Description of female attraction calls (FACs)
Ten FACs from nine A. gazella (N = 90), 10 A. tropicalis (N = 100), 10 A. forsteri (N = 100) and 12 hybrid (N = 120) pups (Overall N = 410) were analysed out of 994 FACs recorded. The additional 584 calls were not analysed, as initial multi-variate analyses using 10 calls per individual were adequate. FACs of the three species recorded in this study were similar to those previously described for A. gazella (Stirling and Warneke 1971) and A. forsteri (Stirling and Warneke 1971, Brown 197 4) though more variation was evident, possibly due to the larger samples.
Summary statistics and representative spectrograms of A. gazella, hybrids, A. tropicalis and A. forsteri FACs are presented in Table 2 and Figure 1, respectively. Arctocephalus gazella calls consisted of many parts. Most A. gazella calls were made up of PULSED+TONAL regions or PULSED+TONAL+FM. Many A. gazella calls
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

TA
BL
E 1
Su
mm
ary
sta
tist
ics
of
PA
Cs
from
10
A.
ga
zell
a,
12 h
yb
rid
s,
10 A
. tr
op
ica
lis
and
six
A.
fors
teri
. A
ll v
alu
es a
re i
n H
z ex
cep
t N
P (
n).
DU
R a
nd
FM
P (
sec)
an
d R
AM
P (
prop
orti
on).
A.
gaze
lla
Hyb
rids
A
. tr
opic
alis
A
. fo
rste
ri
Fea
ture
M
ean
S.E
. M
in.
Max
. #c
alls
M
ean
S
.E.
Min
. M
ax.
#cal
ls M
ean
S.E
. M
in.
Max
. #c
alls
M
ean
S.E
. M
in.
Max
. #c
alls
NP
2.
8 0.
2 1.
0 10
.0
100
3.2
0.4
1.0
25.0
11
7 1.
4 0.
1 1.
0 3.
0 10
0 4.
7 0.
8 1.
0 18
.0
34
DU
R
1.2
0.0
0.7
2.1
100
1.5
0.0
0.7
2.6
117
1.4
0.0
0.7
2.4
100
1.6
0.1
1.0
2.2
34
PE
AK
1 77
9 12
49
1 10
75
100
665
13
374
1122
11
7 52
4 16
22
2 81
8 10
0 49
6 61
14
1 12
68
34
PE
AK
2 14
67
26
795
2081
10
0 14
85
34
655
2689
11
7 10
28
30
508
1677
10
0 97
3 84
31
0 22
79
34
PE
AK
3 21
90
46
1029
40
21
99
2329
73
10
75
5330
10
9 15
55
47
690
2934
10
0 15
65
152
471
3460
32
R
AM
P1
1.31
0.
01
1.21
1.
54
100
1.30
0.
02
0.86
1.
92
117
1.15
0.
03
0.71
2.
12
100
0.92
0.
02
0.72
1.
11
34
RA
MP2
1.
38
0.02
1.
04
1.77
99
1.
35
0.02
1.
01
2.10
10
9 1.
18
0.03
0.
73
2.20
10
0 0.
96
0.05
0.
72
1.32
32
IN
F
750
15
426
1161
10
0 55
2 13
23
3 12
25
117
415
14
171
746
100
475
56
112
1097
34
M
AX
F 82
2 16
42
6 13
17
100
624
13
233
1225
11
7 54
4 16
26
5 88
4 10
0 54
8 68
11
2 13
40
34
HI
760
14
308
1076
10
0 68
5 23
96
13
98
106
506
14
192
748
100
530
61
151
1221
34
E
ND
F
658
15
308
1174
10
0 51
4 17
17
1 12
53
117
399
14
151
732
100
877
90
128
1599
32
F
MP
0.
02
0.00
0.
01
0.05
55
0.
05
0.01
0.
01
0.27
19
0.
02
0.00
0.
00
0.04
22
0.
03
0.01
0.
01
0.04
4
FM
R
296
18
110
726
55
330
23
179
516
19
245
20
102
392
22
176
29
107
224
4
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

128
(A)
4
! ~~~
,-._ ,_ ~,; 'r, , N
~ = '~ ~ '-' ~ <:.1 - ' =
"' ~
~~ = a' I ~
""' ~ ~~~~I 1
WW./r, 0
0 A. gazella A. gazella Hybrid Hybrid 20
4
.I
,, ,-._ N = I ~ '-' II' I ~ = II ~ = a' ~
""' ~
Hybrid Hybrid A. tropicalis A. tropical is A. forsteri A.forsteri 0 20
Time (sec)
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

129
(B)
8
0
0 A. gazella A. gazella Hybrid Hybrid Hybrid 12
Hybrid A. tropicalis A. tropicalis A. forsteri A.forsteri 0 12
Time (sec)
Figure 1. Spectrograms showing intra-individual and inter-individual variation of A. gazella, hybrid, A. tropicalis and A. forsteri PACs (A) and FACs (B). Two calls per individual. Frequency bandwidth= 86Hz.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

130
A.
B.
SHAPE NPARTS PEAK
C') ... 0 -u ~
RAMP PEAK2
PEAK3
ENDF NPARTS SHAPE
ouR Vector 1 SHAPE
OUR PEAK3 RAMP
MAXF INF
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

c.
D.
PEAK1 RAMP2 RAMP1
DUR
ENDFL--------_.;:""'~~\'
DUR ENDF PEAK2
Vector 2
DUR PEAK1 RAMP
C"') ... 0
~
ENDF SHAPE
RAMP1 NPARTS ENDF
RAMP2
Vector 1
DUR ENDF
PEAK2 PEAK3 PEAK1
ector 2
INF MAXF SHAPE
DUR
131
Figure 2. Intra-individual and inter-individual variation in (A) 49 PACs by five A. gazella females (the area enclosed by the polygon shows variation in 99 PACs of 10 A. gazella females with 2D stress= 0.12), (B) 55 PACs by six hybrid females (polygon shows variation in 110 PACs of 12 hybrid females with 2D stress= 0.12), (C) 50 PACs by five A. tropicalis females (polygon shows variation in 100 PACs of lOA. tropicalis females with 2D stress= 0.06) and (D) 32 PACs by six A. forsteri females (polygon shows variation in 32 PACs of six A. forsteri females with 2D stress = 0.06). Each call is represented as a single point. Individuals are represented as different symbols. Variables that had significant correlation coefficients are shown for each end of each axis.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

132
began with PULSED regions and ended with TONAL regions. Arctocephalus gazella calls had relatively high fundamental frequencies and FM was present in many calls. Some A. gazella calls were made up of many pulses separated by distinct silences making a staccato sound. Hybrids' calls showed characteristics of A. gazella, A. tropicalis and A. forsteri in categorical, frequency and amplitude measurements. Arctocephalus tropicalis calls were characterised by long constant TONAL regions and FM was present in many calls and made up the entirety of some calls. Arctocephalus tropicalis calls had relatively low fundamental frequencies resulting in a low-pitched scream sound. Arctocephalus forsteri calls were similar to those of A. gazella although PULSED regions made up relatively more of each call than TONAL regions. The PULSED regions present in most A. forsteri calls made them sound like high pitched guttural screams.
Intra- and inter-individual pup attraction call (PAC) variation
The results of MDS analyses for each species show the similarity of one call to each other call (by the same and different individuals, Figure 2). The relatively tight groups formed by the individuals' calls indicate low levels of intra-individual variability. The relatively distinct groups of calls indicate high levels of inter-individual variability. Up to three significantly correlated variables are shown for each end of each axis.
Individuality was also quantified using three further statistical analyses:
(1) DFA indicated a high level of call individuality within species. Table 3 shows the percentage of PACs assigned correctly based on the DFA for A. gazella, hybrids, A. tropicalis and A. forsteri. The DFA was most successful at assigning A. forsteri calls and least successful at assigning hybrid calls (Table 3). On average the DFA assigned 74%, 66%, 84% and 88% of calls to the correct individual for A. gazella, hybrids, A. tropicalis and A. forsteri, respectively, which is greater than random expectation (10% (Z=3.16 P=0.002), 8% (Z=3.46 P=0.001), 10% (Z=3.16 P=0.002) and 17% (Z=2.50 P=0.014), respectively).
(2) Analyses of variance showed that 13, 12, 11 and 11 of the 13 acoustic features were significantly different among individual::; for A. gazella, hybrids, A. tropicalis and A. forsteri, respectively. Frequency modulation range and period (FMP and FMR) were both significantly different among A. gazella and FMP varied significantly among hybrids, possibly because few other individuals gave frequency-modulated calls.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13
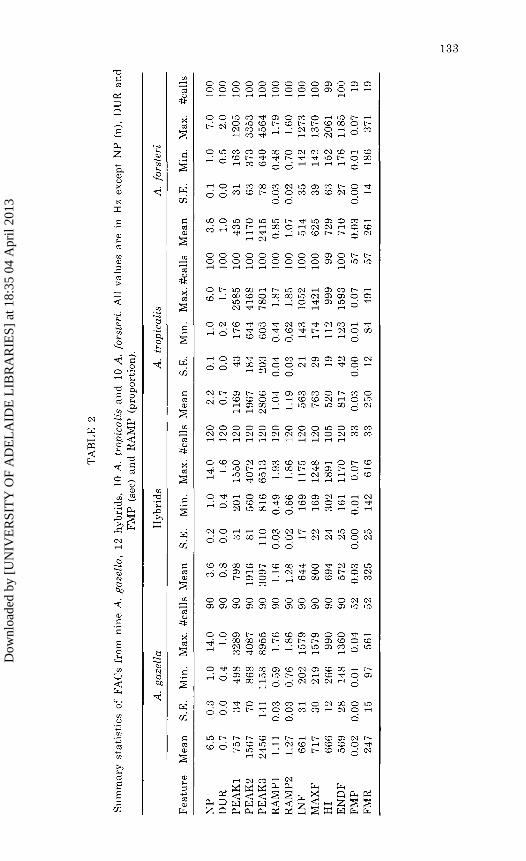
TA
BL
E 2
Sum
mar
y st
atis
tics
of
FAC
s fr
om n
ine
A.
gaze
lla.
12
hyb
rids
, 10
A.
trop
ical
is a
nd
10
A.
fors
teri
. A
ll va
lues
are
in
Hz
exce
pt N
P (
n),
DU
R a
nd
FM
P (
sec)
an
d R
AM
P (
prop
orti
on).
A.
ga
zell
a
Hyb
rids
A
. tr
opic
alis
A
. fo
rste
ri
Fea
ture
M
ean
S.E
. M
in.
Max
. #c
alls
M
ean
S.E
. M
in.
Max
. #c
alls
M
ean
S.E
. M
in.
Max
. #c
alls
M
ean
S.E
. M
in.
Max
. #c
alls
NP
6.
5 0.
3 1.
0 14
.0
90
3.6
0.2
1.0
14.0
12
0 2.
2 0.
1 1.
0 6.
0 10
0 3.
8 0.
1 1.
0 7.
0 10
0 D
UR
0.
7 0.
0 0.
4 1.
0 90
0.
8 0.
0 0.
4 1.
6 12
0 0.
7 0.
0 0.
2 1.
7 10
0 1.
0 0.
0 0.
5 2.
0 10
0 P
EA
K1
757
34
498
3289
90
79
8 31
20
1 15
50
120
1169
43
17
6 25
85
100
435
31
163
1205
10
0 P
EA
K2
1567
70
86
8 40
87
90
1916
81
56
0 40
72
120
1967
18
4 64
4 41
68
100
1170
63
37
3 33
53
100
PE
AK
3 24
56
141
1158
89
55
90
3097
11
0 81
6 65
13
120
2806
20
3 60
3 78
01
100
2415
78
64
0 45
64
100
RA
MP1
1.
11
0.03
0.
59
1.76
90
1.
16
0.03
0.
49
1.93
12
0 1.
04
0.04
0.
44
1.87
10
0 0.
85
0.03
0.
48
1.79
10
0 R
AM
P2
1.27
0.
03
0.76
1.
86
90
1.28
0.
02
0.66
1.
86
120
1.19
0.
03
0.62
1.
85
100
1.07
0.
02
0.70
1.
60
100
INF
66
1 31
20
2 15
79
90
644
17
169
1175
12
0 56
3 21
14
3 10
52
100
514
35
142
1273
10
0 M
AX
F 71
7 30
21
9 15
79
90
800
22
169
1248
12
0 76
3 29
17
4 14
21
100
625
39
142
1370
10
0 H
I 66
6 12
26
6 99
0 90
69
4 24
30
2 18
91
105
520
19
112
999
99
729
63
152
2061
99
E
ND
F
569
28
148
1360
90
57
2 25
16
1 11
70
120
817
42
123
1593
10
0 71
0 27
17
6 11
85
100
FM
P
0.02
0.
00
0.01
0.
04
52
0.03
0.
00
0.01
0.
07
33
0.03
0.
00
0.01
0.
07
57
0.03
0.
00
0.01
0.
07
19
FM
R
247
15
97
561
52
325
25
142
616
33
250
12
84
491
57
261
14
186
371
19
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13
B.
134
A.
INF
(") MAXF .. s u Q)
> RAMP1
SHAPE
Vector 3
PEAK3 PEAK2 ENDF
MAXF DUR IN F
Vector 1
ENDF
PEAK1
END F L-------------~NNPP-;A R T s ENDF IN F PEAK3
Vector 2 RAMP 1 SHAPE
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

c.
D.
('I) ... 0 -() ~
DUR PEAK 1
tv'AXF lf\F
Vecta 1
Vector3
ENDF MAXF PEAK3 SHAPE
NPARTS PEAK 1
Vector2
RAM P1
RAMP2L----------------------8HA; PEAK2 SHAPE PEAK3 MAXF IN F
Vector1
135
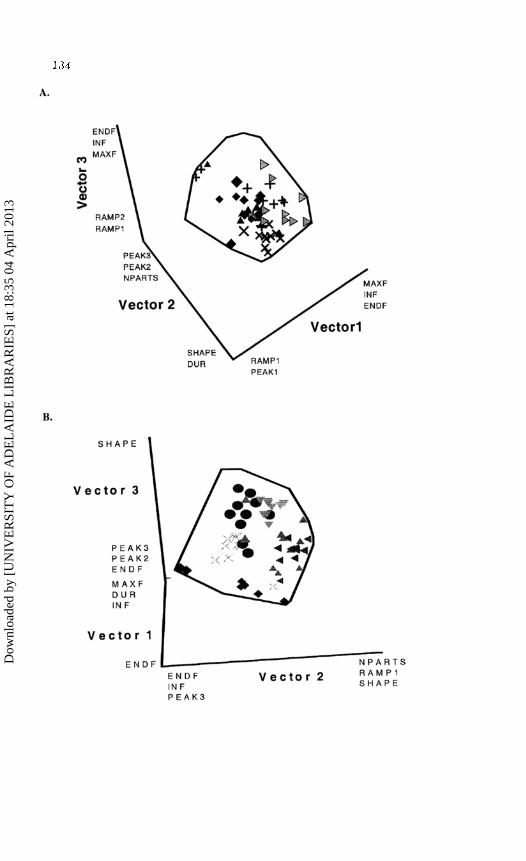
Figure 3. Intra-individual and inter-individual variation in (A) 50 FACs by five A. gazella pups (the area enclosed by the polygon shows variation in 90 FACs of nine A. gazella pups with 2D stress = 0.09), (B) 60 FACs by six hybrid pups (polygon shows variation in 120 FACs of 12 hybrid pups with 2D stress= 0.10), (C) 50 FACs by five A. tropical is pups (polygon shows variation in 100 FACs of 10 A. tropical is pups with 2D stress= 0.0 11) and (D) 50 FACs by five A. forsteri pups (polygon shows variation in 100 FACs of lOA. forsteri pups with 2D stress= 0.10). Each call is represented as a single point. Individuals are represented as different symbols. Variables that had significant correlation coefficients are shown for each end of each axis.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

TA
BL
E 3
Th
e p
erce
nta
ge
of
PA
Cs
assi
gn
ed c
orr
ectl
y a
nd
th
e ra
nd
om
pro
bab
ilit
y o
f as
sig
nin
g t
he
call
s co
rrec
tly
to
ind
ivid
ual
fem
ales
b
ased
on
the
dis
crim
inan
t fu
nct
ion
an
aly
ses
A.
gaze
lla
Hyb
rid
A
. tr
opic
alis
A
. fo
rste
ri
Fem
ale
n %
co
rrec
t R
and
om
n
% c
orre
ct
Ran
do
m
n %
co
rrec
t R
and
om
n
%
corr
ect
Ran
do
m
Fem
ale
1 10
10
0 10
10
10
0 s
10
100
10
1 0
17
Fem
ale
2 10
50
10
10
so
s
10
so
10
10
90
17
Fem
ale
3 10
so
10
s
63
s 10
90
10
5
so
17
Fem
ale
4 10
so
10
10
60
s
10
100
10
10
100
17
Fem
ale
5 10
10
0 10
7
29
s 10
so
10
4
75
17
Fem
ale
6 10
60
10
10
90
s
10
so
10
2 10
0 17
F
emal
e 7
10
90
10
10
30
s 10
so
10
F
em
ale
S
10
70
10
10
30
s 10
40
10
F
emal
e 9
10
40
10
10
60
s 10
90
10
F
emal
e 10
9
67
0 6
100
s 10
10
0 10
F
emal
e 11
9
67
s F
emal
e 1
2
10
90
s
N,
mea
n
99
74
10
110
67
s 10
0 S4
10
32
ss
17
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

137
(3) Comparing CV to s2A showed there was significantly less variation, for a given acoustic structure, within one individual than there was among individuals for each species (A gazella: Z=3.18 P=0.001; hybrids: Z=3.11 P=0.002; A. tropicalis: Z=3.11 P=0.002; A. forsteri: Z=3.11 P=0.002).
Intra- and inter-individual female attraction call (FAC) variation
The results of MDS analyses are presented for A. gazella, hybrids, A. tropicalis and A. forsteri in Figure 3, showing equivalent results for FACs that Figure 2 shows for PACs.
Individuality was further quantified using the same three statistical analyses as for FACs:
(1) DFA indicated a high level of call individuality within species. Table 4 shows the percentage of F ACs assigned correctly based on the DFA for A. gazella, hybrids, A. tropicalis and A. forsteri. The DFA was most successful at assigning A. tropicalis and hybrid calls and least successful at assigning A. gazella calls (Table 4). On average the DFA assigned 52%, 83%, 83% and 79% of calls to the correct individual for A. gazella, hybrids, A. tropicalis and A. forsteri, respectively, which is greater than random expectation (11% (Z=2.50 P=0.014), 8% (Z=3.46 P=0.001), 10% (Z=3.16 P=0.002) and 10% (Z=3.16 P=0.002), respectively).
(2) Analyses of variance showed that 13, 12, 12 and 11 of the 13 acoustic features were significantly different among individuals for A. gazella, hybrids, A. tropicalis and A. forsteri, respectively. Frequency modulation range and period (FMP and FMR) both varied significantly among A. gazella individuals and the variable FMR
was significantly different among hybrids and A. tropicalis. (3) Comparing CV to s2 A showed there was significantly less variation,
for a given acoustic structure, within one individual than there was among individuals for each species (A gazella: Z = 3.18 P = 0.001; hybrids: Z = 3.11 P = 0.002; A. tropicalis: Z = 3.11 P = 0.002; A. forsteri: Z = 3.11 P = 0.002).
Comparison of individual vocal traits between species
The data on individual variation suggests A. forsteri and A. tropicalis PACs would result in less recognition errors than A. forsteri and A. tropicalis PACs. Multi-dimensional scaling and discriminant function analyses both indicated higher inter-individual variation for A. forsteri and A. tropicalis PACs. The spread of each individuals' symbols in the
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

TA
BL
E 4
Th
e p
erce
nta
ge
of F
AC
s as
sig
ned
co
rrec
tly
an
d t
he
ran
do
m p
rob
abil
ity
of
assi
gn
ing
th
e ca
lls
corr
ectl
y t
o
ind
ivid
ual
pu
ps
bas
ed o
n t
he
dis
crim
inan
t fu
nct
ion
an
aly
ses
(n =
10 f
or e
ach
in
div
idu
al).
A.
ga
zell
a
Hyb
rid
A
. tr
opic
alis
A
. fo
rste
ri
Pu
p
% c
orr
ect
Ran
do
m
%
corr
ect
Ran
do
m
%
corr
ect
Ran
do
m
%
corr
ect
Ran
do
m
Pu
p 1
40
11
10
0 8
80
10
90
10
Pu
p 2
80
11
80
8
100
10
70
10
Pu
p 3
30
11
90
8
100
10
90
10
Pu
p 4
70
11
70
8
70
10
70
10
Pu
p 5
3
0
11
90
8 90
10
90
10
P
up
6
80
11
70
8 10
0 10
10
0 10
P
up
7
40
11
90
8 60
10
70
10
P
up
8
60
11
70
8 60
10
70
10
P
up
9
40
11
50
8 10
0 10
90
10
P
up
10
100
8 70
10
50
10
P
up
11
80
8 P
up
12
100
8
N,
mea
n
52
11
83
8 83
10
79
10
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

139
MDS (Figure 2) shows a pattern of A. tropicalis and A. forsteri individuals' PACs being relatively discrete, which is confirmed by the lower stress values (Figure 2). The same pattern is reflected in the DFA, which assigned PACs most accurately to A. forsteri and A. tropicalis (Table 3). Finally, an increased number of parts -changing from tonal to pulsed to frequency-modulated - in an attraction call may make a species' attraction calls more complex and therefore different to other species' calls. If this assumption were correct we would expect A. forsteri (highest number of parts) and hybrid individuals to have more stereotyped PACs. However, contrary to data from MDS and DFA we would then expect A. tropicalis (fewest parts) to have less stereotyped PACs (Table1).
The patterns from FACs were not conclusive for interspecific comparisons as MDS grouped individuals from each species with similar stress values (Figure 3) and the DFA attributed FACs of hybrids, A. tropicalis and A. forsteri with similar accuracy. However, the DFA attributed A. gazella pups relatively inaccurately, with five of the nine pups having just 40% (or less) of their calls correctly assigned (Table 4). Contrary to data from the DFA, the FACs with the highest number of parts were made by A. gazella while A. tropicalis individuals had the fewest parts (Table 1).
DISCUSSION
For the attraction calls of mothers and pups to be useful as signatures, they must be variable among individuals and repeatable within individuals. This study demonstrated that the level of intra-individual stereotypy in PACs and F ACs (in three Arctocephalus species and their hybrids) is high enough that these calls most likely convey information to enable individual recognition.
Strong vocal recognition is likely to be advantageous to both parent and offspring in colonial species for two reasons. Firstly, accurate recognition of filial offspring should enhance the lifetime reproductive success of parents by ensuring that resources are directed only to offspring and not to unrelated young (McMahon et al. 1999). Secondly, parents often behave agonistically to non-filial offspring that stray too close (Jones et al. 1987, Balcombe and McCracken 1992, Phillips and Stirling 2000). Therefore, reduced recognition errors by dependent young are likely to reduce potential injuries and enhance survival.
Other colonial breeding species employ vocalisations for individual recognition in an extreme manner. For example, King penguins Aptenodytes patagonicus breed in colonies numbering from a few hundred to 300,000 pairs (Aubin and Jouventin 1998). Unlike fur seals, which may also use visual, olfactory and vocal cues for
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

140
recognition, identification in penguins occurs using vocalisations alone (Aubin and Jouventin 1998). King and Emperor Aptenodytes forsteri penguins encode vocal individuality using brief frequencymodulated syllables separated by silences (Bremond et al. 1990, Aubin and Jouventin 1998). The highly frequency-modulated calls of king penguins exemplify the result of extreme selection for individual recognition. For their attraction calls to convey individuality, amidst thousands of calls from conspecifics, frequency varies markedly within calls. In comparison, frequency modulation was not present in all fur seal calls. When frequency modulation was present, its range and period were small. Although frequency modulation further stereotypes calls of individuals, it does not seem to be necessary for individual recognition in fur seals. Phillips and Stirling (2000) suggested that frequency modulation and pulses in A. australis PACs and F ACs aided in identifying and locating individuals but their calls were most likely discerned on the suite of frequency, temporal and amplitude-related characteristics.
Comparing call stereotypy between species may reveal differing selective pressure on vocal individuality (Miller 1991). The results of this study suggest A. forsteri and A. tropicalis PACs are more stereotyped than those of other fur seals (including A. australis; Phillips and Stirling (2000)) and that F ACs vary similarly between species, with the exception of A. gazella that showed lower levels of individual variation. This study did not compare traits nor behaviours that may help explain the observed differences in PACs nor FACs. However, it is worth discussing aspects of species biology that may have influenced PAC variability. Phillips and Stirling, (2000) suggested A. australis mothers had highly stereotyped PACs because mothers regularly separated from their pups to cool off in the water. As a result, mother-pup reunifications were more frequent compared with other fur seals, leading to selection for highly stereotyped attraction calls in A. australis. Similarly, A. forsteri and A. tropicalis breed in warm regions and on hot days females swim to cool off, so it is possible selection may operate at a similar level. Secondly, typical A. forsteri and A. tropicalis breeding colonies are found on jumbled rocky coastline whereas A. gazella breed on open beaches (Goldsworthy et al. 1999). If jumbled rocks obscure vision then vocalisations would most likely be used to a greater extent (even from close range) by mother and pup A. forsteri and A. tropicalis. Finally, lactation length may affect selection for highly stereotyped vocalisations. Both A. forsteri and A. tropicalis lactate for about 200 days longer than A. gazella (Kovacs and Lavigne 1992), so mother-pup reunions take place many more times in A. forsteri and A. tropicalis before pups are weaned. Stereotyped PACs may be strongly selected in all species, but in species that rely less on visual cues and reunite more frequently over an extended period, selection for more efficient PACs may result in
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

141
increased fitness to both mother and pup. A comparison of mother-pup reunion behaviour in A. tropicalis (high variation) and A. gazella (lower variation) at Macquarie Island may elucidate whether vocalisations are more important in A. tropicalis compared with A. gazella.
In summary, female A. forsteri and A. tropicalis appear to have more stereotyped PACs than other fur seals. This may be a result of differing levels of selective pressure on call stereotypy that minimise recognition errors during extended mother-pup relations. Such recognition errors represent breakdowns in the mother's ability to discern an unrelated pup from her own and are rare in otariids (Bowen 1991). The importance of individual recognition based on vocalisations is therefore best visualised in terms of maximising fitness.
ACKNOWLEDGMENTS
We thank Snowy, Peter Fullagar, Sue Robinson, John van den Hoff and Mal Page for supplying the recording gear and computers used in this project. Thanks to Ian Preston for helping transport the recording gear. We thank Alana Phillips and Daryl Boness for reading drafts and reviewing this manuscript and Alana for her help throughout the project. We thank Corey Bradshaw and Dion Burns for help in New Zealand. Louise Wynen provided genotypes for many of the individual seals used in this study. We also acknowledge the CSIRO Wildlife and Sound Library, Australian Antarctic Advisory Committee, Zeagrahm Expeditions and ABC Radio Tasmania.
REFERENCES
Aubin, T. & Jouventin, P. (1998). Cocktail-party effect in king penguin colonies. Proceedings of the Royal Society of London - Series B: Biological Sciences, 265, 1665-1673.
Balcombe, J.P. & McCracken, G.F. (1992). Vocal recognition in Mexican free-tailed bats: do pups recognise mothers? Animal Behaviour, 43, 79-87.
Beals, E.W. (1984). Bray curtis ordination: an effective strategy for analysis of multivariate ecological data. Advances in Ecological Research, 14, 1-55.
Blackshaw, J.K., Jones, D.N. & Thomas, F.J. (1996). Vocal individuality during suckling in the intensively housed domestic pig. Applied Animal Behaviour Science, 50, 33-41.
Bowen, W.D. (1991). Behavioural ecology of pinniped neonates. In: The Behaviour of Pinnipeds (D. Renouf, ed.). Chapman and Hall; Cambridge.
Bremond, J-C., Aubin, T., Nyamsi, R.M. & Robisson, R. (1990). The song of the emperor penguin (Aptenodytes forsteri): research of parameters likely to be used for individual recognition. Animal Psychology, 311, 31-35.
Brown, D.L. (1974). Vocal communication of the New Zealand fur seal on Open Bay Islands. Fisheries Technical Report No. 130. New Zealand Ministry of Agriculture and Fisheries. Wellington, New Zealand.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

142
Charrier, I., Mathevon, N., & Jouventin, P. (2001). Mother's voice recognition by seal pups - Newborns need to learn their mother's call before she can take off on a fishing trip. Nature, 412, 873.
Frommolt, K-H., Kruchenkova, E.P. & Russig, H. (1997). Individuality of territorial barking in Arctic foxes, Alopex lagopus (L., 1758). Proceedings of the 1st International Symposium on Physiologie and Ethology of Wild and Zoo Animals, Supplement II, 66-70.
Goldsworthy, S.D., Boness, D. & Fleischer, R. (1999). Mate-choice among sympatric fur seals: female preference for con-phenotypic males. Behavioural Ecology and Sociobiology, 45, 253-267.
Hammerschmidt, K. & Fischer, J. (1998). Maternal discrimination of offspring vocalisations of barbary macaques (Macaca sylvanus). Primates, 39, 231-236.
Insley, S.J. (1992). Mother-offspring separation and acoustic stereotypy: a comparison of call morphology in two species of pinnipeds. Behaviour, 120, 103-121.
Jones, I.L., Falls, J.B. & Gaston, A.J. (1987). Vocal recognition between parents and young of ancient murrelets, Synthliboramphlus antiquus (Aves: Alcidae). Animal Behaviour, 35, 1405-1415.
Kovacs, K.M., & Lavigne, D.M. (1992). Maternal investment in seals and walruses. Canadian Journal of Zoology, 70, 1953-1964.
Marler, P. (1955). Characteristics of some animal calls. Nature, 176, 6-8. McMahon, C.R., Burton, H.R., & Bester, M.N. (1999). First-year survival of southern
elephant seals, Mirounga leonina, at subantarctic Macquarie Island. Polar Biology, 21, 279-284.
Miller, E.H. (1991). Communication in pinnipeds, with special reference to nonacoustic signalling. In: The Behaviour of Pinnipeds (D. Renouf, ed.). Chapman and Hall; Cambridge.
Mitani, J.C., Gros-Louis, J. & Macedonia, J.M. (1996). Selection for acoustic individuality within the vocal repertoire of wild chimpanzees. International Journal of Primatology, 17, 569-583.
Nairns, P.M., Feng, A.S., Yong, H-S., Christensen-Dalsgaard, J.C. (1998). Morphological, behavioural and genetic divergence of sympatric morphotypes of the treefrog, Polypedates leucomystax, in Peninsular Malaysia. Herpetologica, 54, 129-142.
Phillips, A.V. & Stirling, I. (2000). Vocal individuality in mother and pup South American fur seals, Arctocephalus australis. Marine Mammal Science, 16, 592-616.
Reby, D., Joachim, J., Lauga, J., Lek, S. & Alagnier, S. (1998). Individuality in the groans of fallow deer (Dama dama) bucks. Journal of Zoology, 245, 79-84.
Roux, J-P. & Jouventin, P. (1987). Behavioural cues to individual recognition in the subantarctic fur seal, Arctocephalus tropicalis. In: Status Biology and Ecology of Fur Seals. (J.P. Croxall and R.L. Gentry, eds.). National Oceanic and Atmospheric Administration; USA, pp 95-102.
Schatral, A. (1990). Interspecific acoustic interactions in bush crickets. In: The Tettigoniidae: Biology, Systematics and Evolution. (W.J. Bailey and D.C.F. Rentz, eds.). Crawford House Press; Australia, pp 152-165.
Scherrer, J.A. & Wilkinson, G.S. (1993). Evening bat isolation calls provide evidence for heritable signatures. Animal Behaviour, 46, 84 7-860.
Schiffman S., Reynolds M.L., & Young FW. (1981). Introduction to multidimmensional scaling. Academic Press; New York.
Shaughnessy, P.D. & Fletcher, L. (1987). Fur seals, Arctocephalus spp., at Macquaric Island. In: Status Biology and Ecology of Fur Seals (J.P. Croxall and R.L. Gentry, eds.). National Oceanic and Atmospheric Administration; USA, pp 177-188.
Smolker, R.A., Mann, J. & Smuts, B.B. (1993). Use of signature whistles during separations and reunions by wild bottlenose dolphin mothers and infants. Behavioural Ecology and Sociobiology, 33, 393-402.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13

143
SakaL R.R. & Braumann, C.A. (1980). Significance tests for coefficients of variation and variability profiles. Systematic Zoology, 29, 50-66.
Sakal, R.R. & Rohlf, F.J. (1981). Biometry. (2nd Ed) W.H. Freeman and Co.; San Francisco, pp 191-198.
Stirling, I. (1970). Observations on the behaviour of the New Zealand fur seal (Arctocephalus forsteri). Journal of Mammalogy, 51, 766-778.
Stirling, I. (1971). Studies on the behaviour of the South Australian fur seal, Arctocephalus forsteri (Lesson). Australian Journal of Zoology, 19, 243-266.
Stirling, I. & Warneke, R.M. (1971). Implications of a comparison of the airborne vocalisations and some aspects of the behaviour of the two Australian fur seals, Arctocephalus spp., on the evolution and present taxonomy of the genus. Australian Journal of Zoology, 19, 227-241.
Stoddard, P.K. & Beecher, M.D. (1983). Parental recognition of offspring in the cliff swallow. Auk, 100, 795-799.
Trillmich, F. (1981). Mutual mother-pup recognition in Galapagos fur seals and sea lions: cues used and functional significance. Behaviour, 78, 21-42.
Van Parijs, S.M. (2000). Individual and geographical variation in display behaviour of male harbour seals in Scotland. Animal Behaviour, 59, 559-568.
Wanker, R., Apein, J., Jennerjahn, B. & Waibel, B. (1998). Discrimination of different social companions in spectacled parrotlets (Forpus conspicillatus): evidence for individual vocal recognition. Behavioural Ecology and Sociobiology, 43, 197-202.
Wynen, L.P., Goldsworthy, S.D., Guinet, C., Bester, M.N., Boyd, I.L., Gjertz, I., Hofmeyr, G.J.G., White, R.G. & Slade, R. (2000). Postsealing genetic variation and population structure of two species of fur seal (Arctocephalus gazella and A. tropicalis). Molecular Ecology, 9, 299-314.
Received 10 May 2000, revised 12 December 2001 and accepted 17 January 2002.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
8:35
04
Apr
il 20
13





























