Embed Size (px)
Citation preview

1 23
ProtoplasmaAn International Journal of Cell Biology ISSN 0033-183X ProtoplasmaDOI 10.1007/s00709-014-0664-6
Identification of inhibitors of Plasmodiumfalciparum RuvB1 helicase usingbiochemical assays
Moaz Ahmad, Mohammed Tarique,Farhat Afrin, Narendra Tuteja & RenuTuteja

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Wien. This e-offprint is for personal
use only and shall not be self-archived
in electronic repositories. If you wish to
self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

ORIGINAL ARTICLE
Identification of inhibitors of Plasmodium falciparumRuvB1 helicase using biochemical assays
Moaz Ahmad & Mohammed Tarique & Farhat Afrin &
Narendra Tuteja & Renu Tuteja
Received: 5 March 2014 /Accepted: 6 June 2014# Springer-Verlag Wien 2014
Abstract Human malaria is a major parasitic infection, andthe situation has worsened mainly due to the emergence ofresistant malaria parasites to several anti-malarial drugs. Thus,an urgent need to find suitable drug targets has led to thedevelopment of newer classes of anti-malarial drugs.Helicases have been targeted to develop therapeutics for viral,bacterial, and other microorganism infections. Recently, Plas-modium falciparum RuvB ATPases/helicases have been char-acterized and proposed as a suitable antimalarial drug target.In the present study, the screening of various compounds wasdone and the results suggest that PfRuvB1 ATPase activity isinhibited considerably by the novobiocin and partially bycisplatin and ciprofloxacin. Helicase assay of PfRuvB1 inthe presence of various compounds suggest novobiocin, acti-nomycin, and ethidium bromide as potent inhibitors. Novobi-ocin inhibits the helicase activity of PfRuvB1 possibly byblocking the ATPase activity of PfRuvB1. This study isunique in respect to the identification of novobiocin as inhib-itor of PfRuvB1, partially by competing with ATP binding at
its active site and provides evidence for PfRuvB1 as target ofnovobiocin after DNA gyrase-B and HSP90. These studieswill certainly help the pharmacologist to design and developsome novel inhibitor specific to PfRuvB1, whichmay serve assuitable chemotherapeutics to target malaria.
Keywords RuvB like protein . RuvB1 . ATPase . Helicase .
Plasmodium falciparum
Introduction
Malaria is the protozoan parasitic disease caused byPlasmodium species and responsible for over 0.66 millionhuman death globally every year (Murray et al. 2012;World-Malaria-Report 2012). There are certainly limiteddrugs available for the treatment of malaria, including chloro-quine, sulphadoxine pyremethamine, and derivatives. Effortsbeing made to identify suitable drug targets led to the devel-opment of newer classes of drugs, as malaria parasite hasshown resistance to the currently available drugs. Thus, it isimportant to explore the underlying cause of the emergence ofdrug-resistant parasite and develop novel therapeutics to treatthe resistant malaria parasite infections. Although many puta-tive therapeutic targets for malaria have been reported inrecent studies (Augagneur et al. 2013; Bullard et al. 2013;Canduri et al. 2007; Doerig and Meijer 2007; Flannery et al.2013; Jirage et al. 2010; Maloney et al. 2011; Mpangase et al.2013; Ramdhave et al. 2013; Spitzmuller and Mestres 2013),screening with small molecule is required in order to get theinsight for the development of suitable drugmolecule. Varioushelicases have been proposed as suitable drug targets forhuman cancers as well as many pathogens including viruses,bacteria, and plasmodium (Aggarwal et al. 2013; Ahmad et al.2012a; Field and Biswas 2011; Frick 2003; Kwong et al.2005; Li et al. 2012; Seow et al. 2005; Shadrick et al. 2013;
Moaz Ahmad and Mohammed Tarique contributed equally to this work.
Handling Editor: Reimer Stick
Electronic supplementary material The online version of this article(doi:10.1007/s00709-014-0664-6) contains supplementary material,which is available to authorized users.
M. Ahmad :M. Tarique :N. Tuteja : R. Tuteja (*)Malaria Group, International Centre for Genetic Engineering andBiotechnology, P.O. Box 10504, Aruna Asaf Ali Marg, NewDelhi 110067, Indiae-mail: [email protected]
R. Tutejae-mail: [email protected]
F. AfrinDepartment of Biotechnology, Jamia Hamdard, Hamdard Nagar,New Delhi, India
ProtoplasmaDOI 10.1007/s00709-014-0664-6
Author's personal copy

Sharma et al. 2005; Tuteja 2007, 2013; Tuteja and Pradhan2006; Weller and Kuchta 2013; Xi 2007).
During genome-wide analysis of Plasmodium falciparum,it was discovered that parasite contains three different putativeRuvB helicases and later their characterization revealed thatpontin-related (human RuvB1) PfRuvB1 and PfRuvB2 areactive as ATPase as well as DNA helicase. Conversely,reptin-related (human RuvB2) PfRuvB3 is active only asATPase and did not show appreciable helicase activity withrecombinant protein (Ahmad et al. 2012b; Ahmad and Tuteja2013a, b). Considering the characteristic ATPase/helicase ac-tivity of PfRuvB family of proteins and their crucial interplayfor robust helicase activity during intraerythrocytic mitosis ofP. falciparum, we speculate PfRuvB family of protein aspossible therapeutic targets. Additionally, RuvB homolog inyeast is essential for the survival and cell cycle progression.Overexpression of RuvB-like proteins from various humancancers has been reported, and through further studies, it wasproposed as a suitable drug target for chemotherapeutics formany carcinomas (Rosenbaum et al. 2013). Later, it wasreported that ATPase activity of RuvB2 (reptin) is crucial forthe growth of hepatocellular carcinomas (Grigoletto et al.2013), thus it seems that targeting ATPase or its coupledhelicase activity can result into some successful finding to-ward the development of suitable therapeutics. Thus, based onthe biochemical role of PfRuvB1 (Ahmad and Tuteja 2013b)and essential requirement of RuvB, homolog in yeast reflectsthe importance of this family of proteins, and it seems thatPfRuvB1 can serve as a drug target for malaria control.
Here, in this study, we report the effect of commerciallyavailable different molecules such as actinomycin,camptothecin, ciprofloxacin, cisplatin, cyclophosphamide,DAPI, etoposide, ethidium bromide (EtBr), genistein, morin,and novobiocin on the helicase and ATPase activities ofPfRuvB1 protein. Out of these molecules tested, only novo-biocin was found to be the potent inhibitor for the PfRuvB1enzymatic activities. It is interesting to note that althoughactinomycin does not inhibit ATPase activity, it showed con-siderable inhibition of DNA unwinding activity. This studywill help to understand the mechanism of inhibition of theactivities of PfRuvB1 in order to develop it as a novel drugtarget.
Materials and methods
Materials and their source
γ-32P ATP was purchased from Perkin Elmer (Boston, MA,USA), and M13mp19 ssDNAwas purchased from Invitrogen(Carlsbad, CA, USA). Synthetic DNA oligonucleotides weresynthesized chemically. The DNA-interacting compounds,ciprofloxacin, camptothecin, etoposide, cisplatin, and morin
were purchased from Topogene Inc. (Columbus, OH, USA).Novobiocin, cyclophosphamide, and genistein were obtainedfrom Sigma Chemical Co. (St. Louis, MO, USA). EtBr waspurchased from BDH (E. Merck, Mumbai, India), and actino-mycin was purchased from Boehringer Mannheim (Indianap-olis, IN, USA). All of these compounds were dissolved indimethylsulfoxide (DMSO) and stored at 4 °C in the dark.
ATPase assay
The hydrolysis of ATP catalyzed by PfRuvB1 was assayed bymeasuring the formation of Pi from [γ-32P] ATP. Standardprotocol of ATPase assay was followed as described in theprevious report (Ahmad et al. 2012a; Ahmad and Tuteja2013b). The reaction mixture of 10 μl contained [γ-32P]ATP (specific activity 222 TBq mmol−1) and cold ATP(1 mM), ATPase buffer (20 mM Tris–HCl, pH 8.0, 8 mMDTT, 1.0 mM MgCl2, 20 mM KCl, and 16 μg/ml BSA),purified PfRuvB1 and 50 ng of M13 mp19 ssDNA. Reactionmixtures were incubated at 37 °C for 60 min. This wasfollowed by thin-layer chromatography (TLC), and the platewas scanned on phosphoimager. The quantitation was doneusing AlphaImager-EP/Image-J software (http://rsbweb.nih.gov/ij/). For enzyme kinetics analysis, a single time-pointmeasurement at 30 min was performed to determine the Km
and Vmax values with various ATP concentrations. The kineticparameters were calculated using GraphPad Prism version 5.V is the rate of reaction in micromolars per minute, S is thesubstrate concentration in millimolars, Vmax is the maximumreaction rate of the enzyme, and Km is the Michaelis-Mentonconstant. In order to study the effect of DNA-interactingcompounds on ssDNA-dependent ATPase activity, differentcompounds were added into the reaction mixture prior to theaddition of the PfRuvB1 protein.
Preparation of substrate and DNA helicase assay
Helicase assay was performed using the purified fraction ofPfRuvB1 protein. The specially designed partial duplex sub-s t ra te cons i s ted of a 32P- labe led 47-mer DNAoligodeoxynucleotide annealed to M13mp19 phage ssDNA.This oligodeoxynucleotide consisted of the nucleotide se-quence 5′-(T)15GTTTTCCCAGTCACGAC(T)15-3′, and itcontains 15 base-pairs of noncomplementary region (T)15 atboth the 5′- and 3′-ends. The oligodeoxynucleotide was la-beled at 5′-end with T4 polynucleotide kinase (PNK; 5 U;New England Biolabs) using the poly nucleotide kinase buffer(New England Biolabs) and 1.85 MBq of [γ-32P] ATP (spe-cific activity, 222 TBq mmol−1) at 37 °C for 1 h. The reactionwas stopped by heating at 95 °C for 1 min. This labeledoligodeoxynucleotide was then annealed using standard an-nealing buffer (20 mM Tris–HCl, pH 7.5, 10 mM MgCl2,100 mM NaCl, and 1 mM DTT) with 0.5 μg of single-
M. Ahmad et al.
Author's personal copy

stranded circular M13mp19 (+) phage DNA by heating at95 °C for 1 min, and then transferring immediately to 65 °Cfor 2 min and then slow cooling to room temperature. Usinggel filtration through a Sepharose 4B column (Pharmacia,Sweden), nonhybridized oligodeoxynucleotide was removed(Ahmad et al. 2012a). The reaction volume of 10 μl contain-ing the 32P-labeled helicase substrate (1,000 cpm/10 μl) inappropriate buffer (20 mM Tris–HCl, pH 8.0, 8 mM DTT,1.0 mM MgCl2, 20 mM KCl, and 16 μg/ml BSA), and thepurified PfRuvB1 protein was incubated at 37 °C for 60 min.The substrate and products were separated by electrophoresison a nondenaturing 12 % polyacrylamide gel electrophoresis(PAGE), and the gel was scanned on phosphoimager. Thequantitation of both the substrate and unwound DNA bandswas done using AlphaImager-EP/Image-J software (http://rsbweb.nih.gov/ij/). In order to study the effect of DNA-interacting compounds on helicase activity, different com-pounds were added to the helicase assay mixture prior to theaddition of the PfRuvB1 protein.
Immunodepletion assay
To confirm that the enzyme activities are specific to PfRuvB1protein, we performed immunodepletion study. An aliquot ofthe purified recombinant PfRuvB1 protein was incubated withIgG purified from preimmune and anti-PfRuvB1 antiseraseparately at 0 °C for 60 min, and the antigen–antibodycomplex was removed by the addition of equilibrated proteinA sepharose beads. The supernatants of both the anti-PfRuvB1 and preimmune IgG-depleted fraction were usedfor the ATPase and helicase activity assays in the same wayas described above for ATPase and helicase assay.
Results
PfRuvB1 purification and enzyme kinetics
PfRuvB1 protein was purified using Ni-NTA affinity chroma-tography, and purity of the protein was confirmed byCoomassie brilliant blue and silver staining of the sodiumdodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) using the method described previously (Ahmad andTuteja 2013a). Previously, we have reported that PfRuvB1 isactive as ATPase as well as DNA helicase. Here, in order toconfirm that the ATPase activity is specific to PfRuvB1 pro-tein, we performed immunodepletion assay in similar way asdescribed previously (Ahmad and Tuteja 2013a). The resultsclearly show that anti-PfRuvB1-depleted purified PfRuvB1protein fraction was unable to show the ATP hydrolysis whilepreimmune IgGs-depleted fraction showed ATP hydrolysis(Supplementary Fig. 1a). Thus, this study confirms that the
purified PfRuvB1 protein we used for this study is free fromany contaminating protein or ATPase.We performed the effectof increasing substrate (ATP) concentration in order to esti-mate the kinetics of ATPase activity. The estimation of percentATP hydrolysis vs. substrate concentration shows thatPfRuvB1 follows typical sigmoid curve (Fig. 1a). The esti-mation of Pi released vs. substrate concentration showed thatthe maximum release of Pi is ~1.3 μM/min (Fig. 1b). Thekinetics of ATP hydrolysis by PfRuvB1 showed that Km andVmax values are 0.2281±0.03860 mM and 1.346±0.057 μM/min, respectively (Fig. 1b).
Effects of various compounds on ATPase activity
The effect of various compounds (50 μM) on ATPase activityof PfRuvB1 (0.5 μM) was analyzed using the standard proto-col explained in the “Materials and methods.” The resultsclearly show that only novobiocin inhibit the ATP hydrolysissignificantly at 50 μM (Fig. 1c, lanes 3). The other com-pounds which inhibit the ATPase activity at moderate levelunder in vitro conditions are ciprofloxacin and cisplatin(Fig. 1c, lanes 5 and 6, respectively). Among these threeidentified compounds, which inhibit the ATPase activity; no-vobiocin inhibited the ATPase activity at low concentrationcompared with ciprofloxacin and cisplatin. We further per-formed assay in the presence of increasing concentration ofnovobiocin in order to obtain the kinetics of inhibition ofATPase activity. The concentration of novobiocin used inthe ATPase reaction ranged from 2.5 to 15 μM (Fig. 2a, lanes3–6). The results of the concentration curve show that the IC50
value for novobiocin is 8.0723 μM (Fig. 2b). Other inhibitorslike cisplatin and ciprofloxacin did not show significant inhi-bition at 2.5–15 μM (data not shown) while they inhibited~50 % activity at 50 μM (Fig. 1c, lanes 5 and 6). It isinteresting to note that although novobiocin has been testedfor inhibition of ATPase activity in some previous studies, ithas not been reported to be inhibitor of ATPase activity(Table 1). It has been reported previously that novobiocindid not inhibit the ATPase activity of human helicase II(Tuteja et al. 1997).
The other compounds like camptothecin, cyclophospha-mide, etoposide, actinomycin, genistein, morin, and EtBr(Fig. 1c, lanes 4, 7, and 8–12, respectively) were not able toinhibit the ssDNA-dependent ATPase activity of Pf RuvB1 at50 μM concentration under in vitro condition. Thus, in thepresent study, we have identified three compounds (novobio-cin, cisplatin, and ciprofloxacin) which showed potential toinhibit the ATPase activity of PfRuvB1 helicase. Among thesethree compounds, novobiocin is well known to bind at theATPase active site of DNA gyrase and Hsp90 protein(Donnelly and Blagg 2008; Jackson and Maxwell 1993;Raghu Ram et al. 2007; Xiao et al. 2006). Thus, further studywas performed to obtain an insight of this inhibition. To know
Identification of inhibitors of P. falciparum RuvB1 helicase
Author's personal copy

if novobiocin binds at the ATPase active site of PfRuvB1protein, the assay was performed with the fixed concentration(0/5/7.5/10 μM) of novobiocin in the presence of increasingconcentration of ATP (Fig. 2c). The results clearly indicatethat novobiocin partially competes with the ATP for binding atthe ATPase active site (Fig. 2c). Thus, this study indicates thatinhibition observed due to novobiocin is mixed type ofinhibition.
Effect of various compounds on DNA helicase activity
To confirm the purity of PfRuvB1 preparation, theimmunodepletion assay was used. The result clearly showthat anti-PfRuvB1-depleted PfRuvB1 protein fraction did
not show the helicase activity while preimmune IgG-depleted fraction showed helicase activity (SupplementaryFig. 1b). Thus, this purified PfRuvB1 protein, which is freefrom any contaminating protein and activities was used in thisstudy. The DNA helicase activity of PfRuvB1 (0.5 μM) in thepresence of 50 μM of different compounds was tested byusing 1 ng of partially duplex helicase substrate. The resultsobtained clearly show that the compounds, which inhibit theATPase, i.e., novobiocin and ciprofloxacin also inhibit theDNA unwinding activity of PfRuvB1 with different strength(Fig. 3, lanes 2 and 4, respectively). Interestingly, actinomycinand EtBr, which did not inhibit the ATPase activity, inhibitedthe DNA unwinding activity considerably (Fig. 3, lanes 8 and11, respectively). Whereas, the other compounds like
+C 0.1 0.2 0.5 1 2 3
Km: 0.2281 0.0386 mM
Vmax: 1.346 0.057 µM/min
ATP (mM)ATP (mM)
Pi
rele
ase
d (μM
/min
)
0
10
20
30
40
50
1 2 3 4 5 6 7 8 9 10 11 12
EtB
r
Morin
Gen
estin
Actin
om
ycin
Eto
po
side
Cy
clop
ho
s.
Cis- p
latin
Cip
roflo
xa
cin
Cam
pto
thecin
Novob
iocin
Ru
vB
1
Con
trol
% A
TP
ase
act
ivit
y
Pi
ATP
A B
C
0 1 2 3 0 1 2 3
40
30
20
10
0
1.5
1.0
0.5
0.0
% A
TP
hy
dro
lysi
s
+_
+_
Fig. 1 PfRuvB1 ATPase kineticsand screening of variouscompounds. a ATPase assay for30’ with 0.5 μM PfRuvB1 in thepresence of differentconcentration of ATP. The graphof percent ATP hydrolysis vs.ATP concentration was preparedcorresponding to theautoradiogram. b The graph ofrate of reaction vs. ATPconcentration was preparedcorresponding to theautoradiogram in (a). c The effectof various compounds (50 μM)on the ATPase activity. Thecompounds are mentioned belowthe autoradiogram. PercentATPase activity of PfRuvB1 inthe presence of variouscompounds are presented in thebar diagram corresponding to theautoradiogram. Lane 1, noenzyme control; lane 2, controlreaction of PfRuvB1 without anycompound; lanes 3–12, theATPase reactions with enzyme inthe presence of differentcompounds labeled below theautoradiogram
M. Ahmad et al.
Author's personal copy

camptothecin, cisplatin, cyclophosphamide, etoposide, genis-tein, and morin were not able to inhibit the DNA unwindingactivity of PfRuvB1 in in vitro condition (Fig. 3, lanes 3, 5, 6,7, 9, and 10, respectively). The three identified inhibitormolecules (novobiocin, actinomycin, and EtBr) were furtherused for study of inhibition kinetics of helicase activity ofPfRuvB1 protein. The concentration of inhibitors used in
unwinding reaction of PfRuvB1 ranged from 2.5 to 20 μM(Fig. 4a–c), and the results clearly show that the effectiveinhibitor is novobiocin with an IC50 value of 7.6 μM(Fig. 4a); other inhibitors are actinomycin and EtBr withIC50 values of 15.1 and 17.8 μM, respectively (Fig. 4b, c).Thus, these results clearly show that at least these three com-pounds have potential to inhibit the helicase activity of
0
10
20
30
40
50
% A
TP
hy
dro
lysi
s
0 5 10 15 20
IC50: 8.0723 (µM)
Pi
ATP
C 0 2.5 5 10 15
0.0µM
5.0 µM
7.5 µM
10 µM
1 2 3 4 5 6
A B
C
Novobiocin (µM) Novobiocin (µM)
0.0 0.5 1.0 1.5 2.0 2.5
40
30
20
10
0
% A
TP
hy
dro
lysi
s
ATP concentration (mM)
Fig. 2 Analysis of ATPaseinhibition kinetics. aConcentration curve ofnovobiocin. The position of ATPand released Pi is shown on theleft side of the autoradiogram.Lane 1, control without enzyme;lane 2, reaction with enzymewithout any compound. Lanes 3–6, reactions with enzyme in thepresence of 2.5, 5, 10, and 15 μMnovobiocin labeled below theautoradiogram. b Graphicalrepresentation of inhibition ofATPase activity by differentconcentration of novobiocin. cAnalysis of mechanism ofATPase inhibition by novobiocin.Sets of ATPase assay in presenceof fixed concentration ofnovobiocin (5/7.5/10 μM) withincreasing concentration of ATPin the reaction. In the controlreaction, no novobiocin andincreasing concentration of ATPwas added. Graph was preparedusing the Graph Pad Prismversion 5
Table 1 Comparison of inhibito-ry potential for ATPase activity
ND not determined, NI noinhibition
Compounds PfRuvB1 PfUDN PfH45 PfD66/PfDDX19 PfDH60 Human DNAhelicase II
Ciprofloxacin ND 9.9 ND ND ND ND
Cisplatin ND NI ND ND ND ND
Ethidium bromide ND 3.1 1.5 1.5 2.5 ND
Daunorubicin ND 4.4 5.0 ND 3.0 ND
Netropsin ND 3.1 1.5 1.0 3.0 ND
Nogalamycin ND 3.3 0.8 3.2 0.5 ND
Novobiocin 8.0 NI NI NI NI NI
Identification of inhibitors of P. falciparum RuvB1 helicase
Author's personal copy

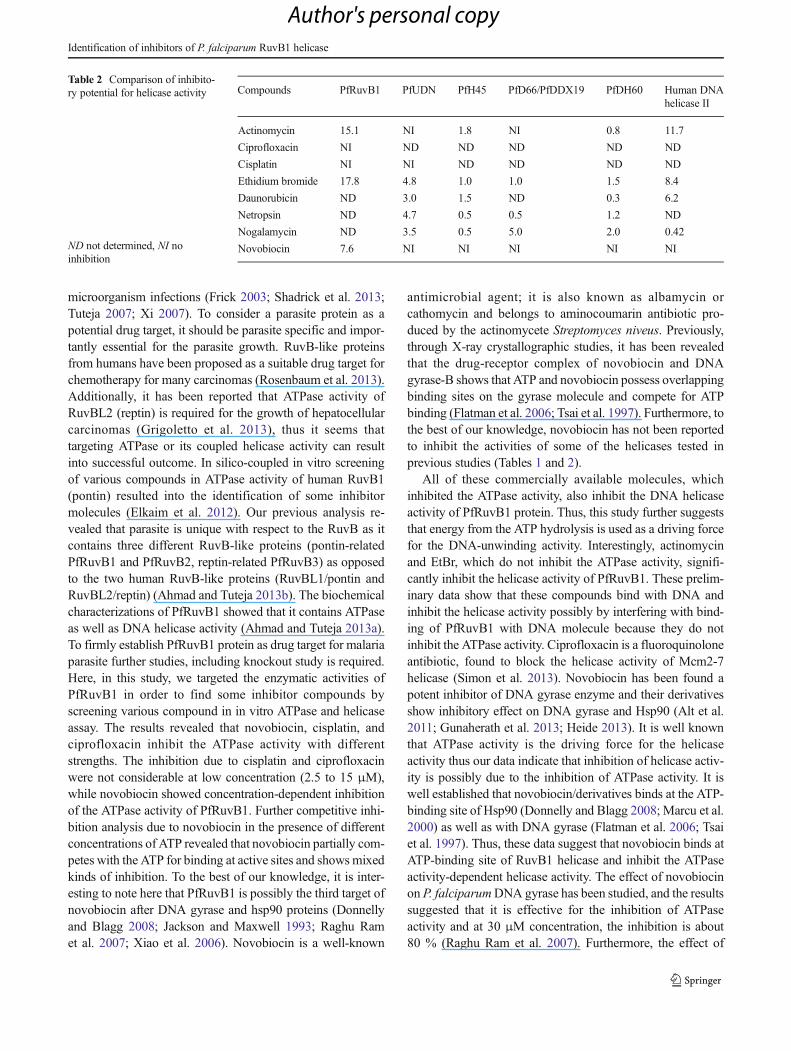
PfRuvB1 protein. A comparison of the inhibitor potential ofsome of these inhibitors for various helicases shows that foractinomycin and EtBr, the values are highly variable (Table 2).It is interesting to further note that although novobiocin hasbeen tested for inhibition of helicase activity in some previousstudies also, it has not been reported to be an inhibitor ofhelicase activity (Table 2). It has been reported previously thatnovobiocin did not inhibit the helicase activity of humanhelicase II (Tuteja et al. 1997). Therefore, these data collec-tively suggest that novobiocin might be a specific inhibitor forPfRuvB1 helicase activities.
Discussion
The identification and characterization of suitable chemother-apeutic target and development of newer class of antimalarialdrug has been a challenging task for the parasitologist. In therecent past, many drug targets have been proposed but furtherextensive studies to screen the drug molecules are required inorder to obtain an insight to design and develop novel inhib-itor molecules with high specificity. Helicases have been anattractive target to fight with viral, bacterial, and many other
0
20
40
60
80
100
B C 1 2 3 4 5 6 7 8 9 10 11
EtB
r
Morin
Gen
estin
Actin
om
ycin
Eto
po
side
Cyclo
ph
os.
Cisp
latin
Cip
roflo
xa
cin
Cam
pto
thecin
No
vo
bio
cin
Ru
vB
1
Co
ntro
l
Bo
iled
% D
NA
un
win
din
g
Fig. 3 Effect of various compounds on the DNA unwinding activity ofPfRuvB1 helicase. The structure of the DNA helicase substrate is shownon the left side of the autoradiogram of the gel. Lane C, the controlwithout enzyme; lane B, heated substrate. Lane 1, the reaction withPfRuvB1 helicase without any compound. Lanes 2–11, the reactions withenzyme in the presence of different compounds (50 μM) labeled belowthe autoradiogram. Percent helicase activity of PfRuvB1 in the presenceof various compounds are presented in the bar diagram corresponding tothe autoradiogram
0
20
40
60
80
100
% D
NA
un
win
din
g
C B 0 2.5 5 7.5 10 15 20
Novobiocin ( M)
IC50:7.645 0.5523 ( M)
0 5 10 15 20
A
1 2 3 4 5 6 7
0
20
40
60
80
100
% D
NA
un
win
din
g IC50: 15.17 0.4889 ( M)
0 5 10 15 20 Actinomycin ( M)
B C 0 2.5 5 7.5 10 15 20
B
1 2 3 4 5 6 7
0
20
40
60
80
100
% D
NA
un
win
din
g IC50: 17.8409 0.9272 ( M)
0 5 10 15 20
C
Ethidium Bromide ( M)
C B 0 2.5 5 7.5 10 15 20
1 2 3 4 5 6 7
µ
µ
µ
µ
µ
µ
+_
+_
+_
Fig. 4 Analysis of RuvB1helicase inhibition kinetics.Concentration curves ofcompounds: a novobiocin, bactinomycin, and c ethidiumbromide. In each panel, theautoradiogram of the assay isshown on the left side and thequantitative data on the right side.a–c Lane C, control withoutenzyme; lane B, reaction isboiled; lane 1, helicase reactionswith PfRuvB1 enzyme alone;lanes 2–7, reaction in thepresence of 2.5, 5, 7.5, 10, 15, and20 μM compounds (as labeledbelow the autoradiogram)
M. Ahmad et al.
Author's personal copy
microorganism infections (Frick 2003; Shadrick et al. 2013;Tuteja 2007; Xi 2007). To consider a parasite protein as apotential drug target, it should be parasite specific and impor-tantly essential for the parasite growth. RuvB-like proteinsfrom humans have been proposed as a suitable drug target forchemotherapy for many carcinomas (Rosenbaum et al. 2013).Additionally, it has been reported that ATPase activity ofRuvBL2 (reptin) is required for the growth of hepatocellularcarcinomas (Grigoletto et al. 2013), thus it seems thattargeting ATPase or its coupled helicase activity can resultinto successful outcome. In silico-coupled in vitro screeningof various compounds in ATPase activity of human RuvB1(pontin) resulted into the identification of some inhibitormolecules (Elkaim et al. 2012). Our previous analysis re-vealed that parasite is unique with respect to the RuvB as itcontains three different RuvB-like proteins (pontin-relatedPfRuvB1 and PfRuvB2, reptin-related PfRuvB3) as opposedto the two human RuvB-like proteins (RuvBL1/pontin andRuvBL2/reptin) (Ahmad and Tuteja 2013b). The biochemicalcharacterizations of PfRuvB1 showed that it contains ATPaseas well as DNA helicase activity (Ahmad and Tuteja 2013a).To firmly establish PfRuvB1 protein as drug target for malariaparasite further studies, including knockout study is required.Here, in this study, we targeted the enzymatic activities ofPfRuvB1 in order to find some inhibitor compounds byscreening various compound in in vitro ATPase and helicaseassay. The results revealed that novobiocin, cisplatin, andciprofloxacin inhibit the ATPase activity with differentstrengths. The inhibition due to cisplatin and ciprofloxacinwere not considerable at low concentration (2.5 to 15 μM),while novobiocin showed concentration-dependent inhibitionof the ATPase activity of PfRuvB1. Further competitive inhi-bition analysis due to novobiocin in the presence of differentconcentrations of ATP revealed that novobiocin partially com-petes with the ATP for binding at active sites and shows mixedkinds of inhibition. To the best of our knowledge, it is inter-esting to note here that PfRuvB1 is possibly the third target ofnovobiocin after DNA gyrase and hsp90 proteins (Donnellyand Blagg 2008; Jackson and Maxwell 1993; Raghu Ramet al. 2007; Xiao et al. 2006). Novobiocin is a well-known
antimicrobial agent; it is also known as albamycin orcathomycin and belongs to aminocoumarin antibiotic pro-duced by the actinomycete Streptomyces niveus. Previously,through X-ray crystallographic studies, it has been revealedthat the drug-receptor complex of novobiocin and DNAgyrase-B shows that ATP and novobiocin possess overlappingbinding sites on the gyrase molecule and compete for ATPbinding (Flatman et al. 2006; Tsai et al. 1997). Furthermore, tothe best of our knowledge, novobiocin has not been reportedto inhibit the activities of some of the helicases tested inprevious studies (Tables 1 and 2).
All of these commercially available molecules, whichinhibited the ATPase activity, also inhibit the DNA helicaseactivity of PfRuvB1 protein. Thus, this study further suggeststhat energy from the ATP hydrolysis is used as a driving forcefor the DNA-unwinding activity. Interestingly, actinomycinand EtBr, which do not inhibit the ATPase activity, signifi-cantly inhibit the helicase activity of PfRuvB1. These prelim-inary data show that these compounds bind with DNA andinhibit the helicase activity possibly by interfering with bind-ing of PfRuvB1 with DNA molecule because they do notinhibit the ATPase activity. Ciprofloxacin is a fluoroquinoloneantibiotic, found to block the helicase activity of Mcm2-7helicase (Simon et al. 2013). Novobiocin has been found apotent inhibitor of DNA gyrase enzyme and their derivativesshow inhibitory effect on DNA gyrase and Hsp90 (Alt et al.2011; Gunaherath et al. 2013; Heide 2013). It is well knownthat ATPase activity is the driving force for the helicaseactivity thus our data indicate that inhibition of helicase activ-ity is possibly due to the inhibition of ATPase activity. It iswell established that novobiocin/derivatives binds at the ATP-binding site of Hsp90 (Donnelly and Blagg 2008; Marcu et al.2000) as well as with DNA gyrase (Flatman et al. 2006; Tsaiet al. 1997). Thus, these data suggest that novobiocin binds atATP-binding site of RuvB1 helicase and inhibit the ATPaseactivity-dependent helicase activity. The effect of novobiocinon P. falciparumDNA gyrase has been studied, and the resultssuggested that it is effective for the inhibition of ATPaseactivity and at 30 μM concentration, the inhibition is about80 % (Raghu Ram et al. 2007). Furthermore, the effect of
Table 2 Comparison of inhibito-ry potential for helicase activity
ND not determined, NI noinhibition
Compounds PfRuvB1 PfUDN PfH45 PfD66/PfDDX19 PfDH60 Human DNAhelicase II
Actinomycin 15.1 NI 1.8 NI 0.8 11.7
Ciprofloxacin NI ND ND ND ND ND
Cisplatin NI NI ND ND ND ND
Ethidium bromide 17.8 4.8 1.0 1.0 1.5 8.4
Daunorubicin ND 3.0 1.5 ND 0.3 6.2
Netropsin ND 4.7 0.5 0.5 1.2 ND
Nogalamycin ND 3.5 0.5 5.0 2.0 0.42
Novobiocin 7.6 NI NI NI NI NI
Identification of inhibitors of P. falciparum RuvB1 helicase
Author's personal copy

novobiocin (10 μM) on P. falciparum culture shows that itinhibits the parasite growth and the inhibitory effect wasobserved during conversion of trophozoite to schizont stage.Similar study with 30 μM novobiocin showed greater reduc-tion in parasetemia (Raghu Ram et al. 2007). Similar effectwith ciprofloxacin has also been reported for the inhibition ofmalaria parasite growth (Goodman et al. 2007).
EtBr is a well-known DNA-interacting compound; thus, itseems that after binding with DNA, this compound perhapsblocks the interaction of PfRuvB1 with DNA and inhibits theDNA unwinding activity of PfRuvB1 helicase. Although EtBris a well-known mutagen, it cannot be used for therapeuticpurposes. Additionally, the concentration at which novobiocinand actinomycin inhibit the helicase activity of PfRuvB1 iscertainly not suitable for the therapeutics purpose, but thisstudy provides preliminary data to develop novel derivativesin order to achieve significant inhibition of ATPase andhelicase activity of PfRuvB1 at much lower concentration.Thus, this study will certainly help pharmacologists to devel-op novel derivative of these identified compounds specific tothe PfRuvB1, which may work at much lower concentrationwith the hope to be useful to fight with malaria.
Acknowledgments This work is partially supported by the Departmentof Biotechnology and Department of Science and Technology grants.Infrastructural support from the Department of Biotechnology, Govern-ment of India is gratefully acknowledged.
Conflict of interest The authors declare that they have no conflict ofinterest.
References
AggarwalM, Banerjee T, Sommers JA, Brosh RM Jr (2013) Targeting anAchilles’ heel of cancer with a WRN helicase inhibitor. Cell Cycle12:3329–3335
Ahmad M, Ansari A, Tarique M, Satsangi AT, Tuteja R (2012a)Plasmodium falciparumUvrD helicase translocates in 3′ to 5′ direc-tion, colocalizes with MLH and modulates its activity throughphysical interaction. PLoS One 7:e49385
Ahmad M, Singh S, Afrin F, Tuteja R (2012b) Novel RuvB nuclearATPase is specific to intraerythrocytic mitosis during schizogonyof Plasmodium falciparum. Mol Biochem Parasitol 185:58–65
Ahmad M, Tuteja R (2013a) Plasmodium falciparum RuvB1 is an activeDNA helicase and translocates in the 5′-3′ direction. Gene 515:99–109
AhmadM, Tuteja R (2013b) Plasmodium falciparumRuvB2 translocatesin 5′-3′ direction, relocalizes during schizont stage and its enzymaticactivities are up regulated by RuvB3 of the same complex. BiochimBiophys Acta 1834:2795–2811
Alt S, Mitchenall LA, Maxwell A, Heide L (2011) Inhibition of DNAgyrase and DNA topoisomerase IV of Staphylococcus aureus andEscherichia coli by aminocoumarin antibiotics. J AntimicrobChemother 66:2061–2069
Augagneur Y, Jaubert L, Schiavoni M, Pachikara N, Garg A, Usmani-Brown S, Wesolowski D, Zeller S, Ghosal A, Cornillot E et al(2013) Identification and functional analysis of the primary
pantothenate transporter, PfPAT, of the human malaria parasitePlasmodium falciparum. J Biol Chem 288:20558–20567
Bullard KM, DeLisle RK, Keenan SM (2013) Malarial kinases: noveltargets for in silico approaches to drug discovery. Methods Mol Biol993:205–229
Canduri F, Perez PC, Caceres RA, de Azevedo WF Jr (2007) Proteinkinases as targets for antiparasitic chemotherapy drugs. Curr DrugTargets 8:389–398
Doerig C, Meijer L (2007) Antimalarial drug discovery: targeting proteinkinases. Expert Opin Ther Targets 11:279–290
Donnelly A, Blagg BS (2008) Novobiocin and additional inhibitors of theHsp90 C-terminal nucleotide-binding pocket. Curr Med Chem 15:2702–2717
Elkaim J, Castroviejo M, Bennani D, Taouji S, Allain N, Laguerre M,Rosenbaum J, Dessolin J, Lestienne P (2012) First identification ofsmall-molecule inhibitors of pontin by combining virtual screeningand enzymatic assay. Biochem J 443:549–559
Field HJ, Biswas S (2011) Antiviral drug resistance and helicase-primaseinhibitors of herpes simplex virus. Drug Resist Updat 14:45–51
Flannery EL, Chatterjee AK, Winzeler EA (2013) Antimalarial drugdiscovery—approaches and progress towards new medicines. NatRev Microbiol 12:849–862
Flatman RH, Eustaquio A, Li SM, Heide L, Maxwell A (2006) Structure-activity relationships of aminocoumarin-type gyrase and topoisom-erase IV inhibitors obtained by combinatorial biosynthesis.Antimicrob Agents Chemother 50:1136–1142
Frick DN (2003) Helicases as antiviral drug targets. Drug News Perspect16:355–362
Goodman CD, Su V, McFadden GI (2007) The effects of anti-bacterialson the malaria parasite Plasmodium falciparum. Mol BiochemParasitol 152:181–191
Grigoletto A, Neaud V, Allain-Courtois N, Lestienne P, Rosenbaum J(2013) The ATPase activity of reptin is required for its effects ontumor cell growth and viability in hepatocellular carcinoma. MolCancer Res 11:133–139
Gunaherath GM, Marron MT, Wijeratne EM, Whitesell L, GunatilakaAA (2013) Synthesis and biological evaluation of novobiocin ana-logues as potential heat shock protein 90 inhibitors. Bioorg MedChem 21:5118–5129
Heide L (2013) New aminocoumarin antibiotics as gyrase inhibitors. Int JMed Microbiol 304:31–36
Jackson AP, Maxwell A (1993) Identifying the catalytic residue of theATPase reaction of DNA gyrase. Proc Natl Acad Sci U S A 90:11232–11236
Jirage D, Keenan SM, Waters NC (2010) Exploring novel targets forantimalarial drug discovery: plasmodial protein kinases. InfectDisord Drug Targets 10:134–146
Kwong AD, Rao BG, Jeang KT (2005) Viral and cellular RNA helicasesas antiviral targets. Nat Rev Drug Discov 4:845–853
Li B, Pai R, Di M, Aiello D, Barnes MH, Butler MM, Tashjian TF, PeetNP, Bowlin TL, Moir DT (2012) Coumarin-based inhibitors ofBacillus anthracis and Staphylococcus aureus replicative DNAhelicase: chemical optimization, biological evaluation, and antibac-terial activities. J Med Chem 55:10896–10908
Maloney, P., Hedrick, M., Peddibhotla, S., Hershberger, P., Milewski, M.,Gosalia, P., Li, L., Preuss, J., Sugarman, E., Hood, B., et al. (2011).A selective inhibitor of Plasmodium falciparum glucose-6-phosphate dehydrogenase (PfG6PDH). Probe Reports from theNIH (http://www.ncbi.nlm.nih.gov/entrez/queryfcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=23762930)
Marcu MG, Chadli A, Bouhouche I, Catelli M, Neckers LM (2000) Theheat shock protein 90 antagonist novobiocin interacts with a previ-ously unrecognized ATP-binding domain in the carboxyl terminusof the chaperone. J Biol Chem 275:37181–37186
Mpangase PT, Szolkiewicz MJ, le Grange M, Smit JH, Burger PB,Joubert F (2013) Discovery-2: an interactive resource for the rational
M. Ahmad et al.
Author's personal copy

selection and comparison of putative drug target proteins in malaria.Malar J 12:116
Murray CJ, Rosenfeld LC, Lim SS, Andrews KG, Foreman KJ, HaringD, Fullman N, Naghavi M, Lozano R, Lopez AD (2012) Globalmalaria mortality between 1980 and 2010: a systematic analysis.Lancet 379:413–431
Raghu Ram EV, Kumar A, Biswas S, Kumar A, Chaubey S, Siddiqi MI,Habib S (2007) Nuclear gyrB encodes a functional subunit of thePlasmodium falciparum gyrase that is involved in apicoplast DNAreplication. Mol Biochem Parasitol 154:30–39
Ramdhave AS, Patel D, Ramya I, Nandave M, Kharkar PS (2013)Targeting heat shock protein 90 for malaria. Mini Rev Med Chem13:1903–1920
Rosenbaum J, Baek SH, Dutta A, Houry WA, Huber O, Hupp TR, andMatias PM (2013) The emergence of the conserved AAA+ATPasespontin and reptin on the signaling landscape. Sci Signal 6, mr1
Seow F, Sato S, Janssen CS, Riehle MO, Mukhopadhyay A, Phillips RS,Wilson RJ, Barrett MP (2005) The plastidic DNA replication en-zyme complex of Plasmodium falciparum. Mol Biochem Parasitol141:145–153
Shadrick WR, Ndjomou J, Kolli R, Mukherjee S, Hanson AM, Frick DN(2013) Discovering new medicines targeting helicases: challengesand recent progress. J Biomol Screen 18:761–781
Sharma S, Doherty KM, Brosh RM Jr (2005) DNA helicases as targets foranti-cancer drugs. Curr Med Chem Anticancer Agents 5:183–199
Simon N, BochmanML, Seguin S, Brodsky JL, Seibel WL, Schwacha A(2013) Ciprofloxacin is an inhibitor of the Mcm2-7 replicativehelicase. Biosci Rep 33(5):e00072
Spitzmuller A, Mestres J (2013) Prediction of the P falciparum targetspace relevant to malaria drug discovery PLoS Comput Biol 9:e1003257
Tsai FT, Singh OM, Skarzynski T, Wonacott AJ, Weston S,Tucker A, Pauptit RA, Breeze AL, Poyser JP, O'Brien Ret al (1997) The high-resolution crystal structure of a 24-kDa gyrase B fragment from E. coli complexed with one ofthe most potent coumarin inhibitors, clorobiocin. Proteins 28:41–52
Tuteja N, Phan TN, Tuteja R, Ochem A, Falaschi A (1997) Inhibition ofDNA unwinding and ATPase activities of human DNA helicase IIby chemotherapeutic agents. Biochem Biophys Res Commun 236:636–640
Tuteja R (2007) Helicases—feasible antimalarial drug target forPlasmodium falciparum. FEBS J 274:46994704
Tuteja R (2013) In silico analysis of Plasmodium species specific UvrDhelicase. Communicative and Integrative Biology 6(2):e23125
Tuteja R, Pradhan A (2006) Unraveling the ‘DEAD-box’ helicases ofPlasmodium falciparum. Gene 376:1–12
Weller SK, Kuchta RD (2013) The DNA helicase-primase complex as atarget for herpes viral infection. Expert Opin Ther Targets 17:1119–1132
World-Malaria-Report (2012). http://www.who.int/malaria/publications/world_malaria_report_2012/en/index.html
Xi XG (2007) Helicases as antiviral and anticancer drug targets. CurrMed Chem 14:883–915
Xiao L, Lu X, Ruden DM (2006) Effectiveness of hsp90 inhibitors asanti-cancer drugs. Mini Rev Med Chem 6:1137–1143
Identification of inhibitors of P. falciparum RuvB1 helicase
Author's personal copy





























