Embed Size (px)
Citation preview

Journal: TPJ Disk used Despatch Date: 15/4/2003Article : npg_tpj_6500176 Pages: 1–12 OP: GNS/Jolly
ORIGINAL ARTICLE
Hydroxyurea downregulates endothelin-1 gene
expression and upregulates ICAM-1 gene
expression in cultured human endothelial cells
M Brun1,2,3
S Bourdoulous1,4,5
PO Couraud1,4,5
J Elion1,5,6
R Krishnamoorthy1,3,6
C Lapoumeroulie1,3,6
1Institut National de la Sante et de la RechercheMedicale (INSERM), France; 2Universite desAntilles et de la Guyane, Faculte de MedecineUMR458, France; 3Institut Claude Bernard,IFR02/UMR458 Biologie du developpement,France; 4Universite Paris 5, Rene DescartesFaculte de Medecine Cochin, Port Royal, France;5Institut Cochin, UMR 567 Maladies Infectieuses,France; 6Universite Paris 7, Denis Diderot Facultede Medecine Xavier Bichat, France
Correspondence:C Lapoumeroulie, UMR 458, Hopital RobertDebre, 48 Bd Serurier, 75019 Paris, France.Tel: +33 1 40 03 19 24, Fax: +33 1 40 03 19 03E-mail: [email protected]
Received: 8 October 2002Revised: 7 March 2003Accepted: 20 March 2003
ABSTRACTThe clinical efficacy of oral hydroxyurea (HU) in adults and children withsickle cell anemia (SCA) cannot solely be explained by its ability to enhancefetal hemoglobin (HbF) expression. Since increased adherence of sickle redblood cells to vascular endothelium is a possible contributing factor to vaso-occlusive crisis (VOC), we explored the effect of HU on human endothelialcell (EC) lines (TrHBMEC and EA-hy 926). We demonstrated that HU, in adose-dependent and reversible manner, significantly decreased (up to three-fold) the release of endothelin-1 (ET-1), a vasoconstrictor peptide throughdownregulation (up to three-fold) of ET-1 gene expression. This finding is oftherapeutic relevance as SCA patients exhibit elevated serum levels of ET-1during episodes of VOC and levels correlate with disease severity.Unexpectedly, HU upregulated (up to three-fold) the expression ofmembrane-bound intercellular cell adhesion molecule 1 (mbICAM-1) andits soluble form (sICAM-1) with a parallel increase in ICAM-1 mRNAexpression. Although ICAM-1 does not appear to be involved in the sicklecell adhesion to vascular endothelium, it may exacerbate vaso-occlusion bypromoting leukocyte adhesion. The HU-induced increase in mbICAM-1 mayappear inconsistent with the clinical benefits confered by HU. However, boththe increase in sICAM-1- and HU-induced leukocyte reduction in patients,may counteract the potentially detrimental effect of elevated mbICAM-1expression. Also HU reduces the expression of vascular cell adhesionmolecule (VCAM-1) on EC. Since HU reduces the very late antigen 4-positivereticulocytes in SCA patients, a ligand for VCAM-1, HU-induceddownregulation of VCAM-1 on EC will very likely decrease the reticulo-cyte–endothelium adhesion. Thus, HU, apart from inducing HbF expressionin the red cell, also affects the expression profile of EC compartment.The Pharmacogenomics Journal (2003) 00, 000–000. doi:10.1038/sj.tpj.6500176
Keywords: sickle cell anemia; hydroxyurea; endothelium
INTRODUCTIONHydroxyurea (HU) is an inhibitor of ribonucleotide reductase, a rate-limitingenzyme, involved in the conversion of ribonucleotides into deoxyribonucleo-tides. Thus, HU affects cells that are actively synthesizing DNA. HU has long beenused to treat a variety of neoplastic disorders including chronic granulocyteleukemia, head and neck cancer and polycythemia vera. In addition to inhibitingribonucleotide reductase, HU affects gene expression, most notably of thehuman fetal globin genes. HU enhances the expression of fetal hemoglobin(HbF) in cell culture and primate models.1,2 As sickle cell anemia (SCA) patients
The Pharmacogenomics Journal (2002) 0, 1–12& 2002 Nature Publishing Group All rights reserved 1470-269X/02 $25.00
www.nature.com/tpj
with high levels of HbF experience a less severe form of thedisease, HU therapy was tested in several clinical trials inpatients with SCA to induce HbF expression.
In clinical trials, HU therapy had many positive clinicaland biological benefits. The clinical benefits include a largereduction in the frequency of hospitalizations, pain epi-sodes, acute chest syndromes (ACS) and blood transfusions.3
Biologically, HU therapy increased HbF levels, F reticulocytecount, F-cell count, mean corpuscular volume, red bloodcell (RBC) survival and deformability, oxygen affinity, cationcontent and hydration. Also, HU decreased the meancorpuscular Hb concentration, hemolysis, number of irre-versibly sickled red cells, KCl cotransport4–8 expression ofred cell adhesion molecules and their in vitro adhesion.9
Although the efficacy of HU therapy in SCA was believed tobe mediated primarily through the induction of HbFexpression, the improvement in clinical symptoms preced-ing any significant increase in HbF raises the possibility thatHU may also act through other mechanisms.10 In SCApatients treated with HU, sickle cell–endothelial cell (EC)adherence decreased prior to any significant increase inHbF.11 This suggests that HU may affect the RBC and/or ECsurface.
Very few studies have explored the effect of HU on ECfunction.9 ECs play a major role in vasoadhesion of a varietyof blood cells via cell surface adhesion molecules, such asintercellular cell adhesion molecule 1 (ICAM-1) and vascularcell adhesion molecule 1 (VCAM-1). RBCs from SCA patients(SS RBCs) have the predilection to adhere to the endothelialsurface as compared to the RBCs from normal subjects (AARBCs).12 Such abnormal RBC–endothelial adherence isbelieved to initiate a painful vaso-occlusive crisis (VOC), aphysiopathologic hallmark of the disease. ECs respond tonumerous extracellular signals such as vasoactive factors(ET-1 and NO), proinflammatory cytokines (TNFa, inter-leukin 1 and IFNg), oxidants and hypoxia.13 Variations inthese signals can affect the EC characteristics and therebycontribute to the unpredictable occurrences of VOC. Theresulting endothelial injury, indicated by the presence ofactivated ECs circulating in the blood of SCA patients,14
could contribute to additional complications. Here, weevaluate the effects of HU on vascular ECs.
RESULTSHU Reduces ET-1 mRNA and Peptide ExpressionTrHBMEC and EA-hy 926 cells were exposed to HU at aconcentration (250 mM) similar to the in vivo HU levels oftreated SCA patients. Experiments were performed both inthe absence and in the presence of 100 U/ml of TNFa andIFNg (a concentration that induces the expression ofadhesion molecules, but does not affect the synthesis andrelease of endothelin-1, ET-1). The presence of proinflam-matory cytokines allowed simulation of the in vivo inflam-matory status of SCA patients.
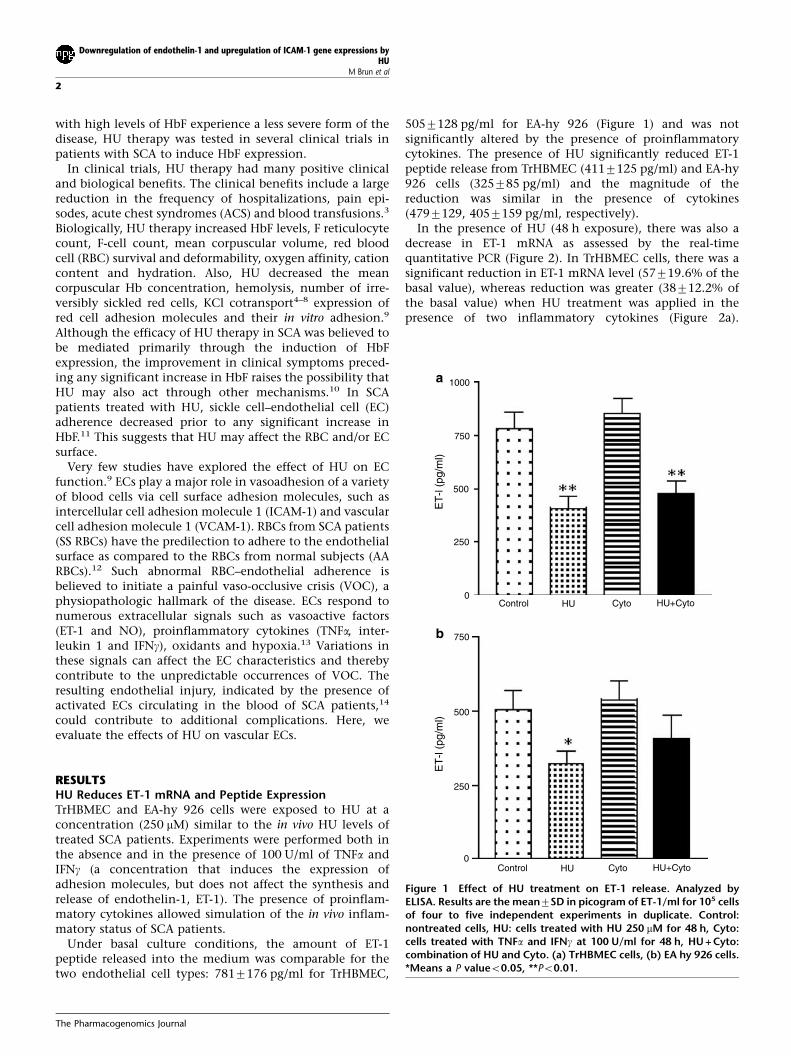
Under basal culture conditions, the amount of ET-1peptide released into the medium was comparable for thetwo endothelial cell types: 7817176 pg/ml for TrHBMEC,
5057128 pg/ml for EA-hy 926 (Figure 1) and was notsignificantly altered by the presence of proinflammatorycytokines. The presence of HU significantly reduced ET-1peptide release from TrHBMEC (4117125 pg/ml) and EA-hy926 cells (325785 pg/ml) and the magnitude of thereduction was similar in the presence of cytokines(4797129, 4057159 pg/ml, respectively).
In the presence of HU (48 h exposure), there was also adecrease in ET-1 mRNA as assessed by the real-timequantitative PCR (Figure 2). In TrHBMEC cells, there was asignificant reduction in ET-1 mRNA level (57719.6% of thebasal value), whereas reduction was greater (38712.2% ofthe basal value) when HU treatment was applied in thepresence of two inflammatory cytokines (Figure 2a).
1000
750
500
250
0Control HU Cyto HU+Cyto
ET
-l (p
g/m
l)
750
500
250
0Control HU Cyto HU+Cyto
ET
-l (p
g/m
l)
a
b
Figure 1 Effect of HU treatment on ET-1 release. Analyzed byELISA. Results are the mean7SD in picogram of ET-1/ml for 105 cellsof four to five independent experiments in duplicate. Control:nontreated cells, HU: cells treated with HU 250 mM for 48 h, Cyto:cells treated with TNFa and IFNg at 100 U/ml for 48 h, HU + Cyto:combination of HU and Cyto. (a) TrHBMEC cells, (b) EA hy 926 cells.*Means a P valueo0.05, **Po0.01.
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions byHU
M Brun et al
2
The Pharmacogenomics Journal
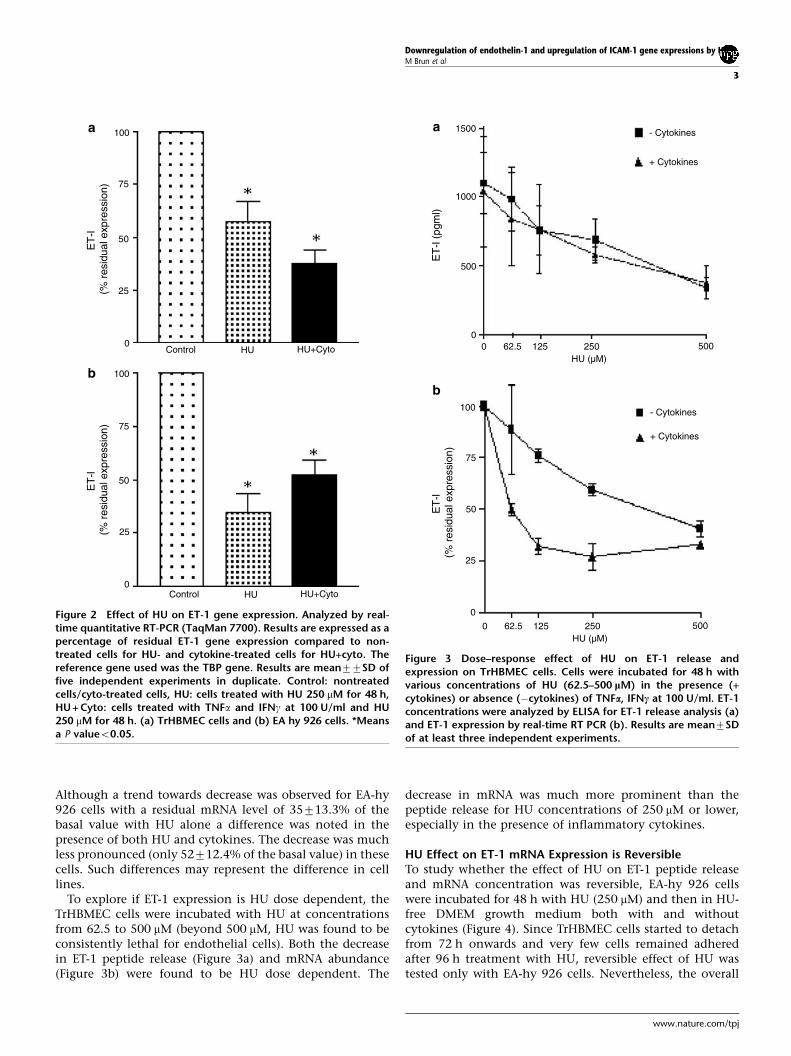
Although a trend towards decrease was observed for EA-hy926 cells with a residual mRNA level of 35713.3% of thebasal value with HU alone a difference was noted in thepresence of both HU and cytokines. The decrease was muchless pronounced (only 52712.4% of the basal value) in thesecells. Such differences may represent the difference in celllines.
To explore if ET-1 expression is HU dose dependent, theTrHBMEC cells were incubated with HU at concentrationsfrom 62.5 to 500 mM (beyond 500 mM, HU was found to beconsistently lethal for endothelial cells). Both the decreasein ET-1 peptide release (Figure 3a) and mRNA abundance(Figure 3b) were found to be HU dose dependent. The
decrease in mRNA was much more prominent than thepeptide release for HU concentrations of 250 mM or lower,especially in the presence of inflammatory cytokines.
HU Effect on ET-1 mRNA Expression is ReversibleTo study whether the effect of HU on ET-1 peptide releaseand mRNA concentration was reversible, EA-hy 926 cellswere incubated for 48 h with HU (250 mM) and then in HU-free DMEM growth medium both with and withoutcytokines (Figure 4). Since TrHBMEC cells started to detachfrom 72 h onwards and very few cells remained adheredafter 96 h treatment with HU, reversible effect of HU wastested only with EA-hy 926 cells. Nevertheless, the overall
100
75
50
25
0Control HU HU+Cyto
Control HU HU+Cyto
100
75
50
25
0
ET
-l (%
res
idua
l exp
ress
ion)
ET
-l (%
res
idua
l exp
ress
ion)
a
b
Figure 2 Effect of HU on ET-1 gene expression. Analyzed by real-time quantitative RT-PCR (TaqMan 7700). Results are expressed as apercentage of residual ET-1 gene expression compared to non-treated cells for HU- and cytokine-treated cells for HU+cyto. Thereference gene used was the TBP gene. Results are mean77SD offive independent experiments in duplicate. Control: nontreatedcells/cyto-treated cells, HU: cells treated with HU 250 mM for 48 h,HU + Cyto: cells treated with TNFa and IFNg at 100 U/ml and HU250 mM for 48 h. (a) TrHBMEC cells and (b) EA hy 926 cells. *Meansa P valueo0.05.
1500
1000
500
00 62.5 125 250 500
HU (µM)
- Cytokines
+ Cytokines
- Cytokines
+ Cytokines
100
75
25
0
50
0 62.5 125 250 500HU (µM)
ET
-l (%
res
idua
l exp
ress
ion)
ET
-l (p
gml)
a
b
Figure 3 Dose–response effect of HU on ET-1 release andexpression on TrHBMEC cells. Cells were incubated for 48 h withvarious concentrations of HU (62.5–500 lM) in the presence (+cytokines) or absence (�cytokines) of TNFa, IFNc at 100 U/ml. ET-1concentrations were analyzed by ELISA for ET-1 release analysis (a)and ET-1 expression by real-time RT PCR (b). Results are mean7SDof at least three independent experiments.
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions by HUM Brun et al
3
www.nature.com/tpj
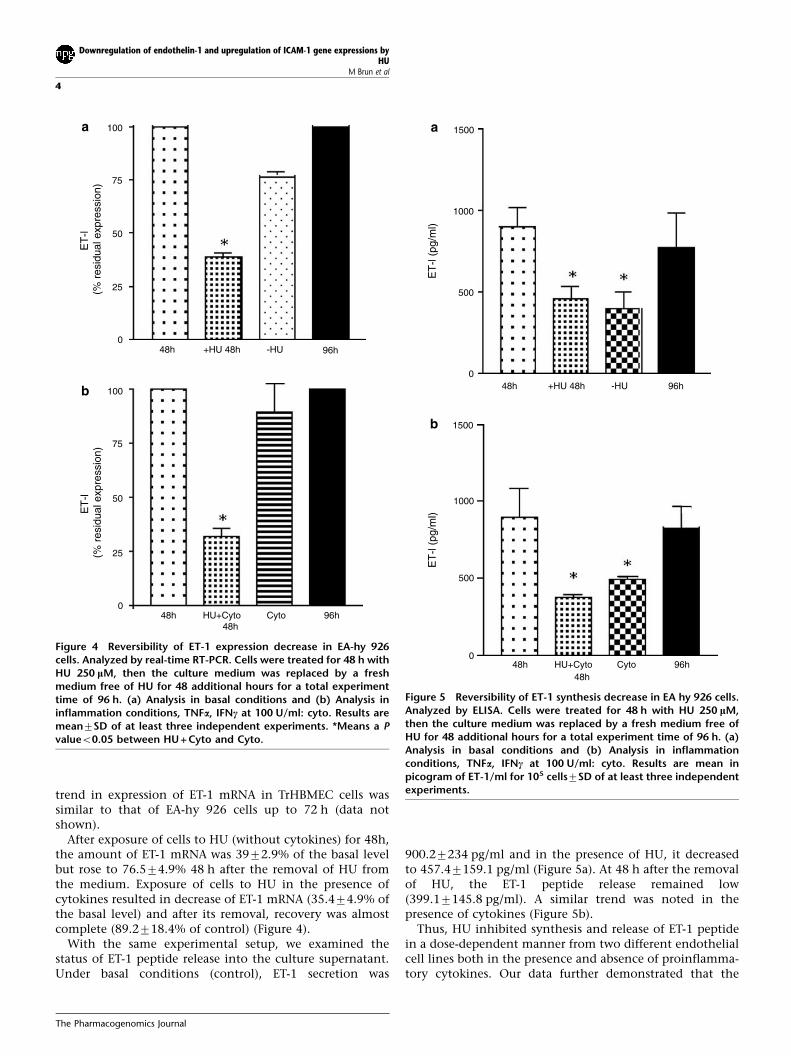
trend in expression of ET-1 mRNA in TrHBMEC cells wassimilar to that of EA-hy 926 cells up to 72 h (data notshown).
After exposure of cells to HU (without cytokines) for 48h,the amount of ET-1 mRNA was 3972.9% of the basal levelbut rose to 76.574.9% 48 h after the removal of HU fromthe medium. Exposure of cells to HU in the presence ofcytokines resulted in decrease of ET-1 mRNA (35.474.9% ofthe basal level) and after its removal, recovery was almostcomplete (89.2718.4% of control) (Figure 4).
With the same experimental setup, we examined thestatus of ET-1 peptide release into the culture supernatant.Under basal conditions (control), ET-1 secretion was
900.27234 pg/ml and in the presence of HU, it decreasedto 457.47159.1 pg/ml (Figure 5a). At 48 h after the removalof HU, the ET-1 peptide release remained low(399.17145.8 pg/ml). A similar trend was noted in thepresence of cytokines (Figure 5b).
Thus, HU inhibited synthesis and release of ET-1 peptidein a dose-dependent manner from two different endothelialcell lines both in the presence and absence of proinflamma-tory cytokines. Our data further demonstrated that the
100
75
50
25
048h +HU 48h -HU 96h
ET
-l (%
res
idua
l exp
ress
ion)
100
75
50
25
0
ET
-l (%
res
idua
l exp
ress
ion)
48h48h
HU+Cyto Cyto 96h
a
b
Figure 4 Reversibility of ET-1 expression decrease in EA-hy 926cells. Analyzed by real-time RT-PCR. Cells were treated for 48 h withHU 250 lM, then the culture medium was replaced by a freshmedium free of HU for 48 additional hours for a total experimenttime of 96 h. (a) Analysis in basal conditions and (b) Analysis ininflammation conditions, TNFa, IFNc at 100 U/ml: cyto. Results aremean7SD of at least three independent experiments. *Means a Pvalueo0.05 between HU + Cyto and Cyto.
1500
1000
500
048h +HU 48h -HU 96h
ET
-l (p
g/m
l)E
T-l
(pg/
ml)
1500
1000
500
048h
48hHU+Cyto Cyto 96h
a
b
Figure 5 Reversibility of ET-1 synthesis decrease in EA hy 926 cells.Analyzed by ELISA. Cells were treated for 48 h with HU 250 lM,then the culture medium was replaced by a fresh medium free ofHU for 48 additional hours for a total experiment time of 96 h. (a)Analysis in basal conditions and (b) Analysis in inflammationconditions, TNFa, IFNc at 100 U/ml: cyto. Results are mean inpicogram of ET-1/ml for 105 cells7SD of at least three independentexperiments.
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions byHU
M Brun et al
4
The Pharmacogenomics Journal
downregulation of ET-1 by HU occurs at the transcriptionallevel.
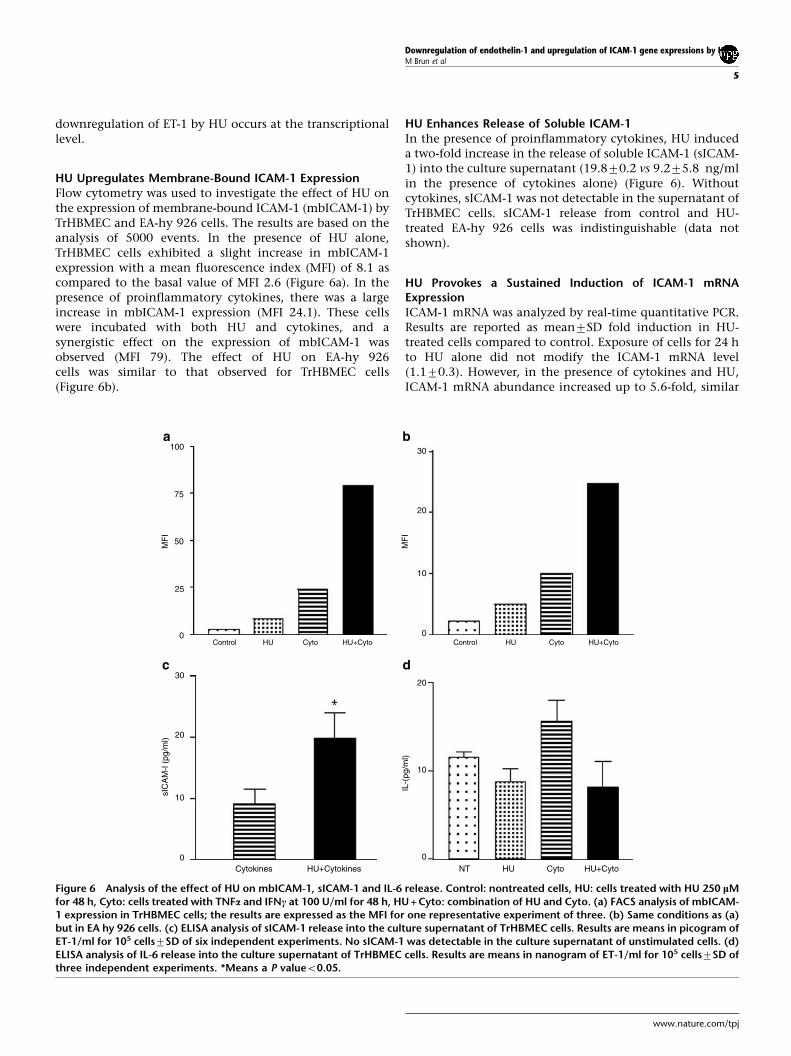
HU Upregulates Membrane-Bound ICAM-1 ExpressionFlow cytometry was used to investigate the effect of HU onthe expression of membrane-bound ICAM-1 (mbICAM-1) byTrHBMEC and EA-hy 926 cells. The results are based on theanalysis of 5000 events. In the presence of HU alone,TrHBMEC cells exhibited a slight increase in mbICAM-1expression with a mean fluorescence index (MFI) of 8.1 ascompared to the basal value of MFI 2.6 (Figure 6a). In thepresence of proinflammatory cytokines, there was a largeincrease in mbICAM-1 expression (MFI 24.1). These cellswere incubated with both HU and cytokines, and asynergistic effect on the expression of mbICAM-1 wasobserved (MFI 79). The effect of HU on EA-hy 926cells was similar to that observed for TrHBMEC cells(Figure 6b).
HU Enhances Release of Soluble ICAM-1In the presence of proinflammatory cytokines, HU induceda two-fold increase in the release of soluble ICAM-1 (sICAM-1) into the culture supernatant (19.870.2 vs 9.275.8 ng/mlin the presence of cytokines alone) (Figure 6). Withoutcytokines, sICAM-1 was not detectable in the supernatant ofTrHBMEC cells. sICAM-1 release from control and HU-treated EA-hy 926 cells was indistinguishable (data notshown).
HU Provokes a Sustained Induction of ICAM-1 mRNAExpressionICAM-1 mRNA was analyzed by real-time quantitative PCR.Results are reported as mean7SD fold induction in HU-treated cells compared to control. Exposure of cells for 24 hto HU alone did not modify the ICAM-1 mRNA level(1.170.3). However, in the presence of cytokines and HU,ICAM-1 mRNA abundance increased up to 5.6-fold, similar
100
75
50
25
0Control HU Cyto HU+Cyto Control HU Cyto HU+Cyto
MF
I
MF
I
30
20
10
0
sIC
AM
-l (p
g/m
l)
IL-(
pg/m
l)
30
20
10
0
20
10
0Cytokines HU+Cytokines NT HU Cyto HU+Cyto
a b
c d
Figure 6 Analysis of the effect of HU on mbICAM-1, sICAM-1 and IL-6 release. Control: nontreated cells, HU: cells treated with HU 250 lMfor 48 h, Cyto: cells treated with TNFa and IFNc at 100 U/ml for 48 h, HU + Cyto: combination of HU and Cyto. (a) FACS analysis of mbICAM-1 expression in TrHBMEC cells; the results are expressed as the MFI for one representative experiment of three. (b) Same conditions as (a)but in EA hy 926 cells. (c) ELISA analysis of sICAM-1 release into the culture supernatant of TrHBMEC cells. Results are means in picogram ofET-1/ml for 105 cells7SD of six independent experiments. No sICAM-1 was detectable in the culture supernatant of unstimulated cells. (d)ELISA analysis of IL-6 release into the culture supernatant of TrHBMEC cells. Results are means in nanogram of ET-1/ml for 105 cells7SD ofthree independent experiments. *Means a P valueo0.05.
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions by HUM Brun et al
5
www.nature.com/tpj
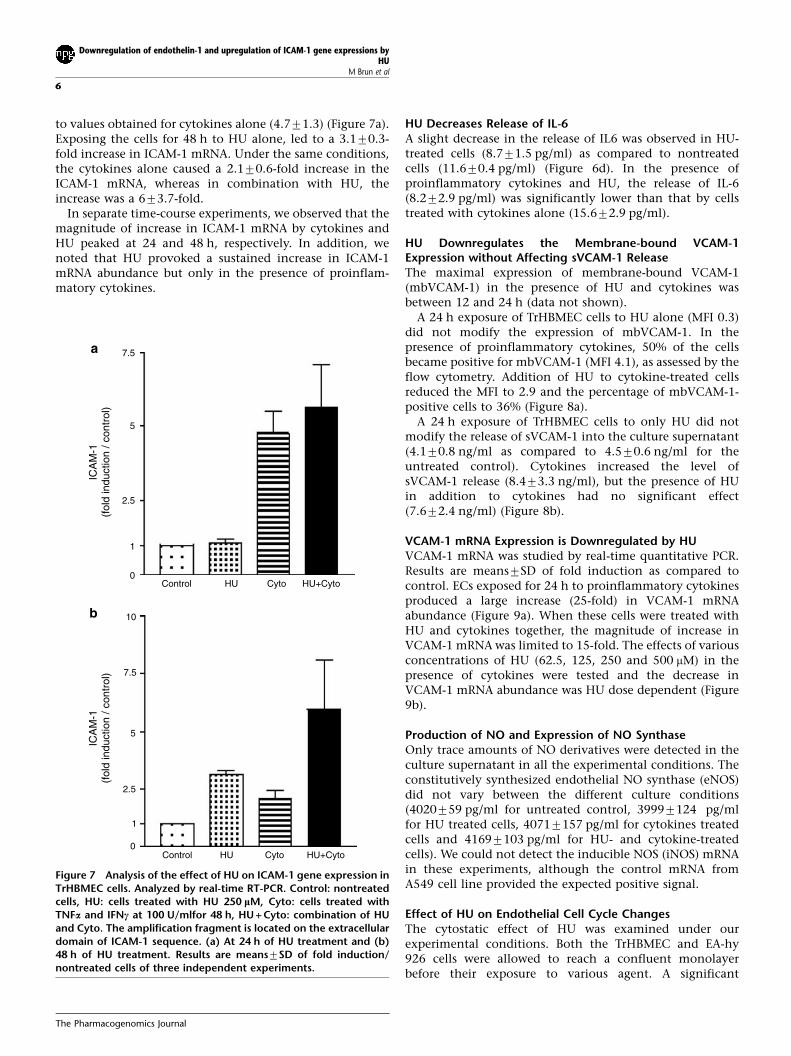
to values obtained for cytokines alone (4.771.3) (Figure 7a).Exposing the cells for 48 h to HU alone, led to a 3.170.3-fold increase in ICAM-1 mRNA. Under the same conditions,the cytokines alone caused a 2.170.6-fold increase in theICAM-1 mRNA, whereas in combination with HU, theincrease was a 673.7-fold.
In separate time-course experiments, we observed that themagnitude of increase in ICAM-1 mRNA by cytokines andHU peaked at 24 and 48 h, respectively. In addition, wenoted that HU provoked a sustained increase in ICAM-1mRNA abundance but only in the presence of proinflam-matory cytokines.
HU Decreases Release of IL-6A slight decrease in the release of IL6 was observed in HU-treated cells (8.771.5 pg/ml) as compared to nontreatedcells (11.670.4 pg/ml) (Figure 6d). In the presence ofproinflammatory cytokines and HU, the release of IL-6(8.272.9 pg/ml) was significantly lower than that by cellstreated with cytokines alone (15.672.9 pg/ml).
HU Downregulates the Membrane-bound VCAM-1Expression without Affecting sVCAM-1 ReleaseThe maximal expression of membrane-bound VCAM-1(mbVCAM-1) in the presence of HU and cytokines wasbetween 12 and 24 h (data not shown).
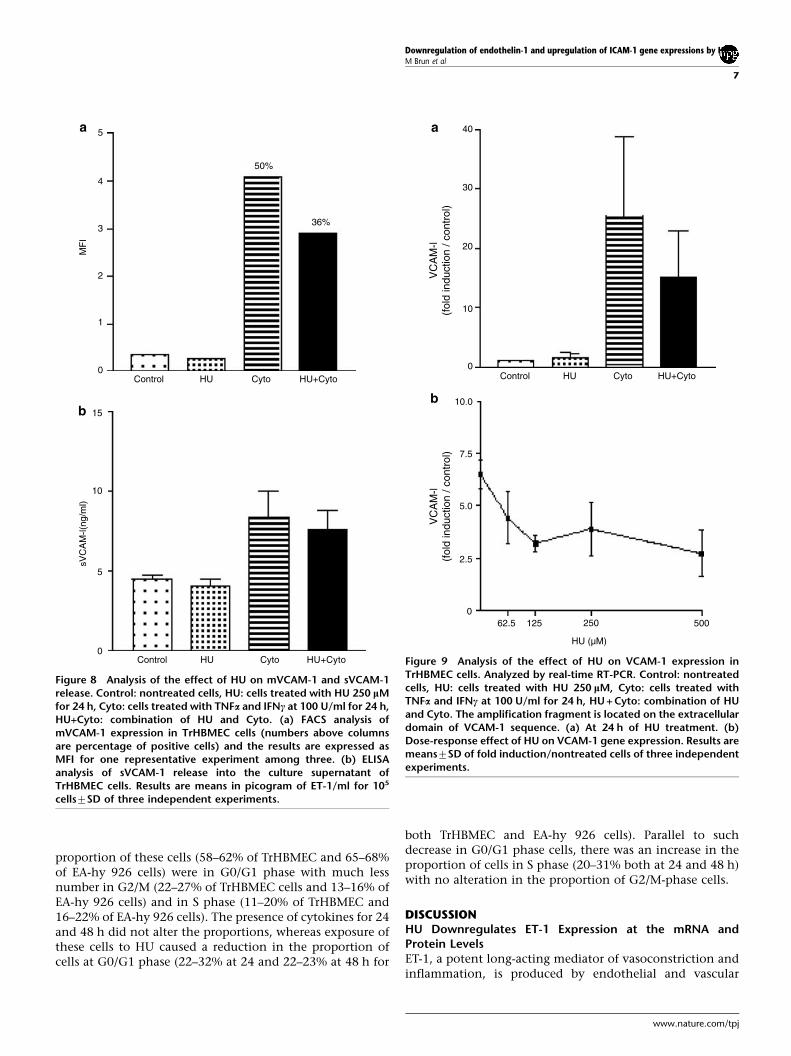
A 24 h exposure of TrHBMEC cells to HU alone (MFI 0.3)did not modify the expression of mbVCAM-1. In thepresence of proinflammatory cytokines, 50% of the cellsbecame positive for mbVCAM-1 (MFI 4.1), as assessed by theflow cytometry. Addition of HU to cytokine-treated cellsreduced the MFI to 2.9 and the percentage of mbVCAM-1-positive cells to 36% (Figure 8a).
A 24 h exposure of TrHBMEC cells to only HU did notmodify the release of sVCAM-1 into the culture supernatant(4.170.8 ng/ml as compared to 4.570.6 ng/ml for theuntreated control). Cytokines increased the level ofsVCAM-1 release (8.473.3 ng/ml), but the presence of HUin addition to cytokines had no significant effect(7.672.4 ng/ml) (Figure 8b).
VCAM-1 mRNA Expression is Downregulated by HUVCAM-1 mRNA was studied by real-time quantitative PCR.Results are means7SD of fold induction as compared tocontrol. ECs exposed for 24 h to proinflammatory cytokinesproduced a large increase (25-fold) in VCAM-1 mRNAabundance (Figure 9a). When these cells were treated withHU and cytokines together, the magnitude of increase inVCAM-1 mRNA was limited to 15-fold. The effects of variousconcentrations of HU (62.5, 125, 250 and 500 mM) in thepresence of cytokines were tested and the decrease inVCAM-1 mRNA abundance was HU dose dependent (Figure9b).
Production of NO and Expression of NO SynthaseOnly trace amounts of NO derivatives were detected in theculture supernatant in all the experimental conditions. Theconstitutively synthesized endothelial NO synthase (eNOS)did not vary between the different culture conditions(4020759 pg/ml for untreated control, 39997124 pg/mlfor HU treated cells, 40717157 pg/ml for cytokines treatedcells and 41697103 pg/ml for HU- and cytokine-treatedcells). We could not detect the inducible NOS (iNOS) mRNAin these experiments, although the control mRNA fromA549 cell line provided the expected positive signal.
Effect of HU on Endothelial Cell Cycle ChangesThe cytostatic effect of HU was examined under ourexperimental conditions. Both the TrHBMEC and EA-hy926 cells were allowed to reach a confluent monolayerbefore their exposure to various agent. A significant
7.5
5
2.5
1
0Control HU Cyto HU+Cyto
Control HU Cyto HU+Cyto
ICA
M-1
(fol
d in
duct
ion
/ con
trol
)IC
AM
-1(f
old
indu
ctio
n / c
ontr
ol)
a
b 10
7.5
5
2.5
0
1
Figure 7 Analysis of the effect of HU on ICAM-1 gene expression inTrHBMEC cells. Analyzed by real-time RT-PCR. Control: nontreatedcells, HU: cells treated with HU 250 lM, Cyto: cells treated withTNFa and IFNc at 100 U/mlfor 48 h, HU + Cyto: combination of HUand Cyto. The amplification fragment is located on the extracellulardomain of ICAM-1 sequence. (a) At 24 h of HU treatment and (b)48 h of HU treatment. Results are means7SD of fold induction/nontreated cells of three independent experiments.
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions byHU
M Brun et al
6
The Pharmacogenomics Journal
proportion of these cells (58–62% of TrHBMEC and 65–68%of EA-hy 926 cells) were in G0/G1 phase with much lessnumber in G2/M (22–27% of TrHBMEC cells and 13–16% ofEA-hy 926 cells) and in S phase (11–20% of TrHBMEC and16–22% of EA-hy 926 cells). The presence of cytokines for 24and 48 h did not alter the proportions, whereas exposure ofthese cells to HU caused a reduction in the proportion ofcells at G0/G1 phase (22–32% at 24 and 22–23% at 48 h for
both TrHBMEC and EA-hy 926 cells). Parallel to suchdecrease in G0/G1 phase cells, there was an increase in theproportion of cells in S phase (20–31% both at 24 and 48 h)with no alteration in the proportion of G2/M-phase cells.
DISCUSSIONHU Downregulates ET-1 Expression at the mRNA andProtein LevelsET-1, a potent long-acting mediator of vasoconstriction andinflammation, is produced by endothelial and vascular
5
4
3
2
1
0Control HU Cyto HU+Cyto
Control HU Cyto HU+Cyto
MF
IsV
CA
M-l(
ng/m
l)
15
10
5
0
50%
36%
a
b
Figure 8 Analysis of the effect of HU on mVCAM-1 and sVCAM-1release. Control: nontreated cells, HU: cells treated with HU 250 lMfor 24 h, Cyto: cells treated with TNFa and IFNc at 100 U/ml for 24 h,HU+Cyto: combination of HU and Cyto. (a) FACS analysis ofmVCAM-1 expression in TrHBMEC cells (numbers above columnsare percentage of positive cells) and the results are expressed asMFI for one representative experiment among three. (b) ELISAanalysis of sVCAM-1 release into the culture supernatant ofTrHBMEC cells. Results are means in picogram of ET-1/ml for 105
cells7SD of three independent experiments.
40
30
20
10
0Control HU Cyto HU+Cyto
VC
AM
-l(f
old
indu
ctio
n / c
ontr
ol)
VC
AM
-l(f
old
indu
ctio
n / c
ontr
ol)
a
b 10.0
7.5
5.0
2.5
062.5 125 250 500
HU (µM)
Figure 9 Analysis of the effect of HU on VCAM-1 expression inTrHBMEC cells. Analyzed by real-time RT-PCR. Control: nontreatedcells, HU: cells treated with HU 250 lM, Cyto: cells treated withTNFa and IFNc at 100 U/ml for 24 h, HU + Cyto: combination of HUand Cyto. The amplification fragment is located on the extracellulardomain of VCAM-1 sequence. (a) At 24 h of HU treatment. (b)Dose-response effect of HU on VCAM-1 gene expression. Results aremeans7SD of fold induction/nontreated cells of three independentexperiments.
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions by HUM Brun et al
7
www.nature.com/tpj

smooth muscle cells in response to hypoxia and shear stress.Other stimuli such as TNFa, a proinflammatory cytokine,induces ET-1 release from a variety of ECs in a time andconcentration-dependent manner accompanied by a corre-sponding increase in the transcriptional rate of the ET-1gene.18 Interestingly, ET-1 mRNA production and ET-1peptide release increase when ECs from bovine pulmonaryartery were exposed to plasma from patients with ACS.19
Similarly sickle erythrocytes specifically stimulate theexpression of the ET-1 gene and production of the peptidein human ECs in culture.20 In animal models ET-1: (i) causesaccumulation of leukocytes in the pulmonary microvascu-lature,21 (ii) increases vascular permeability in the presenceof leukocytes in perfusion22 and (iii) results in a time- anddose-dependent sequential entrapment of platelets andneutrophils in the pulmonary circulation.23 ET-1 activatesthe Gardos channel in mouse erythrocytes.24 This channelactivation, in the context of sickle red cell, is expected todehydrate these cells. Important clinical findings includesignificantly elevated levels of ET-1 in SCA patientsduring the episodes of painful crisis25,26 and ACS.Also, the extent of ET-1 increase correlates with diseaseseverity.19
These experimental and clinical observations suggest thatET-1 might play an important role in the cycle of ischemiaand inflammation that initiates and sustains a painful crisisin sickle cell patients. Local ET-1 levels in the microvascu-lature are probably much higher than those in the systemiccirculation, and may contribute to the prolonged localvasospasm and inflammation and thus to vaso-occlusiveevents, the hallmark of the sickle cell disease. Our findingthat HU significantly suppresses ET-1 gene expression andpeptide release in a dose-dependent manner from ECs(Figure 3a and b) indicates one of the ways in which HUexerts beneficial effects in SCA.
The effect of HU on the downregulation ET-1 expression(further supported by our unpublished observations thatSCA patients under HU have significantly lower plasma ET-1levels) if confirmed in vivo, might offer novel therapeuticstrategies for sickle cell disease. Furthermore, the two-foldreduction in IL-6 production by the HU-exposed ECs (Figure6d) may stem from the suppression of ET-1 expression.Previous studies show that ET-1 can induce IL-6 expressionin human endothelial cells.27 Alternatively, HU may have adirect effect on IL-6 expression. Induced secretion of IL-6 byECs is associated with increased cell permeability, genera-tion of reactive oxygen species and transmigration ofmonocytes through endothelium28 presumptively causalfor vasoendothelial injury. Again the beneficial effects of HUin SCA may be mediated through its effect on IL-6 eitherdirectly or through suppression of ET-1 expression.
We also examined the effect of HU on the expression ofNO, an important component of the vasomotor regulation.HU is believed to be a chemical donor of NO,29,30 a potentvasodilator, and NO seems to provide a variety of beneficialeffects in SCA: (i) low-dose inhalation of NO increases theoxygen affinity of sickle red cells both in vivo and in vitro,31
(ii) NO inhalation at 20 ppm seems to provide a rapid
protection against severe hypoxic stress in a transgenic micemodel for SCD,32 (iii) in SCA patients with ACS, inhaled NOimproves ventilation–perfusion matching33 and (iv) SCApatients with elevated levels of NO metabolites report lowerpain scores.34 NOS tightly regulates the NO expression inendothelial cells. We did not observe any alteration in thelevel of NO metabolites or changes in the endothelialexpression of NOS either at transcriptional or protein levelin HU-treated ECs, which suggests that the HU effect maynot be mediated by this pathway.
HU Upregulates mbICAM-1 Expression and sICAM-1 Release(mbICAM-1 or CD54) is a cell surface glycoprotein of theimmunoglobulin superfamily with multiple immune re-sponse-related functions, which include cell–cell interac-tions and leukocyte adhesion to vascular endothelial cells.The latter event initiates transmigratory egress of leukocytesfrom the vasculature. ICAM-1 performs these functionsthrough its ability to bind b2 integrins such as LFA-1 andMac-1. Expressed constitutively in several cell types includ-ing vascular EC, the level of expression of mbICAM-1 ismarkedly upregulated by inflammatory stimuli such as TNFaand IFNg.35 mbICAM-1 also acts as a major cell-surfacereceptor for rhinovirus and Plasmodium falciparum.36 Inaddition, ICAM-1/LFA-1 interaction seems to promote HIV-1infectivity.37
An sICAM-1 containing most of the extracellular portionof mbICAM-1 is detectable in normal plasma. The sICAM-1level may reflect the expression status of mbICAM-1 onendothelial cells thereby indicating the degree of inflam-matory process and/or EC activation.38 Elevated serumsICAM-1 levels have been noted in various immune andinflammatory disorders including SCA.39 Indeed, sickleRBCs have been shown to induce mbICAM-1 expressionby endothelial cells in culture both in stationary and flowstudies.40 Under flow conditions, upregulation of mbICAM-1 involves both transcriptional and translational control.sICAM-1 levels are higher (two- to three-fold) in childrenwith SCA than in ethnically matched controls.39
Our in vitro experimental finding that HU enhances theexpression of mbICAM-1 by endothelial cells in culture(Figure 6a and b) does not appear to be consistent with thesignificant clinical benefits conferred by this drug in SCApatients. The strongest correlation was found between totalwhite cell count and severity of crisis rather than witherythrocyte-related parameters. Although there is no evi-dence showing that ICAM-1 is directly involved in sickle celladhesion to endothelial cells, it is plausible that leukocyte–endothelium adhesion may initiate the sequestration andentrapment of RBCs resulting in the obstruction of themicrovascular lumen. Accordingly HU-enhanced mb ICAM-1 expression would be expected to aggravate rather than toattenuate the sickle cell crisis. An explanation for thisparadox may come from the HU-induced increase in sICAM-1 (Figure 6c). Indeed sICAM-1 and soluble selectins havebeen shown to suppress significantly the neutrophil–endothelial adhesion in vitro.41 Thus, the beneficial effectof HU may be mediated through enhanced release of soluble
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions byHU
M Brun et al
8
The Pharmacogenomics Journal

adhesive receptors, which compete with the membrane-bound receptor for leukocytes and thereby minimize thevascular adhesion.
How sICAM-1 is generated is still unclear. Two possibilitiesare: (i) differential splicing of ICAM-1 mRNA42 and (ii)proteolytic cleavage of mbICAM-1.43 Distinct mRNA speciesencoding sICAM-1 have been identified in several humancell lines, which support the first possibility. However, andin support of the second possibility, the generation ofsICAM-1 from neutrophil mbICAM-1 by elastase has beendemonstrated.44 Whatever the mechanism, the HU-inducedenhancement of the release/synthesis of soluble ICAM-1may participate in the beneficial effects of this drug. Thus,targeted antiadhesion therapies may become a usefulalternative in the treatment of SCA.
HU Downregulates VCAM-1 ExpressionVCAM-1, like the ICAM-1, belongs to the immunoglobulinsuperfamily and exists in both membrane-bound(mbVCAM-1) and soluble (sVCAM-1) forms. VCAM-1 whoseexpression in ECs is upregulated by proinflammatorycytokines is the receptor for the very late activation antigen4 (VLA-4, a4 b1),45 expressed, only on the sickle erythrocytemembranes.46 ECs stimulated by cytokines exhibit increasedlevels of surface adhesion molecules that could be specifi-cally blocked by antibodies to VLA-4 and VCAM-1. Adhe-sion results in morphological damage to endothelium, theextent of which correlates with the number of adherentsickle erythrocytes.47
Similarly, hypoxia significantly and specifically increasessickle erythrocyte adhesion to aortic and retinal capillaryECs without having any effect on normal erythrocytes.48
The enhanced adhesion of sickle erythrocytes under hypox-ia is accompanied by upregulation of expression of bothVCAM-1 and ICAM-1.40 The expression of these surfaceadhesion molecules is downregulated in vivo in mousemodel by sulfasalazine, a powerful inhibitor of activationof nuclear factor kappa B (NFkB).49 In SCA patients, asignificantly high concentration of sVCAM-1 has beenreported; however, HU treatment does not seem to affectthe sVCAM-1 level.50 Our experimental observation that HUdownregulates VCAM-1 gene expression and mbVCAM-1production in a dose-dependent manner, especially in thepresence of cytokines (Figures 8a, b and 9b and 9b),contrasts with what was observed for ICAM-1 expression.However, HU did not promote the release of sVCAM-1. Suchdownregulation of mbVCAM-1, although modest, couldcontribute to the attenuation of SS RBC–endotheliumadhesion.
The molecular mechanisms by which HU differentiallyregulates the ICAM-1 and VCAM-1 gene expression are notclear, although the promoters of both genes have, bindingsites for several transcription factors including NFkB, AP-1,AP-2, AP-3, and GATA in their 50 regulatory regions.51,52
Interestingly, HU prevented the binding of NFkB to the HIV-1 LTR region, which might explain the downregulation ofVCAM-1, but not the upregulation of ICAM-1.
HU and MalariaICAM-1 is the major cell-surface receptor on vascularendothelial cells for Plasmodium falciparum-infected RBC.53
As HU upregulates mbICAM-1 expression, it may promoteRBC–endothelium adhesion and promote the progression ofmalaria. Hence, the administration of HU to patients fromendemic areas for malaria warrants caution. Alternatively, asdiscussed above for SCA, the enhanced release of sICAM-1may tend to mitigate such effects and eventually bebeneficial. At present, cytoadherence experiments areunderway to clarify the value of using HU in areas endemicfor malaria.
HU and Cell Cycle ChangesGiven the fact that HU is a S-phase blocking agent, theabove-discussed HU-induced changes in EC phenotype andgene expression could be the result of cell cycle changes. Toexplore this possibility, cell cycle studies were performed. Inour experimental conditions around 20–30% of the cells arearrested at the G1/S border and the observed changes in theexpression status of ECs could be because of the accumula-tion of such cell subpopulation. However, the proinflam-matory cytokines did not have any effect on the proportionof cells in S phase, but they do affect the expression status ofcell surface and soluble adhesion molecules (ICAM-1 andVCAM-1) of ECs. This suggests that cell cycle changes alonecannot explain the changes in the expression of thesemolecules. Whatever the mechanism of action of HU, eitherthrough its action on gene expression or on cell cycle orboth, this study provides further insight into the effect ofHU on endothelial cells. Deciphering the transcriptionalsignature of HU in ECs may shed further light on themechanism of action of HU and thereby provide clues toother treatment options for SCA.
METHODSEndothelial Cell CultureTwo types of ECs were studied and were maintained at 371Cwith 5% carbon dioxide in a humidified incubator.
Transformed human bone marrow endothelial cells(TrHBMEC), kindly provided by B Weskler, were culturedon gelatin-coated Petri dishes as previously described.15 Forthis cell line, all experiments were carried out betweenpassage 20 and 24. The cell line EA-hy 926 (epithelial/endothelial hybrid cell line), a gift from CS Edgell, werecultured under the experimental conditions describedpreviously.16 Both cell types were grown until confluenceand then the basal medium was replaced by a mediumcontaining HU at various concentrations (62.5, 125, 250 and500 mM) in the presence or absence of a mixture of TNFa andIFNg (R&D System, Abingdon, UK) each at 100 U/ml andincubated for 48 h, unless otherwise indicated.
Flow Cytometry for Cell Cycle AnalysisTrHBMEC cells and EA-hy 926 were grown as describedabove and treated with 250 mM HU for 24 and 48 h in threeindependent experiments both in the presence and absenceof proinflammatory cytokines (TNFa and IFNb at 100 U/ml).
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions by HUM Brun et al
9
www.nature.com/tpj

Cells were fixed with 100 ml of ice-cold 70% ethanol for10 min at 41C. Cells were spun for 5 min at 200 rpm andthen 100 ml of PBS–RNAse A (Sigma, Saint Louis, MO, USA)at 20 mg/ml was added and cells were incubated for 30 minat room temperature. Then, 200 ml of PBS–propidium iodide(Sigma, Saint Louis, MO, USA) at 75 mg/ml was added andcells were incubated for 15 min at room temperature. A totalof 10 000 cells were analyzed in an Epics-Elite flowcytometer (Coulter Electronic, Hialeah, FL, USA).
ET-1, sICAM-1, IL-6 and sVCAM-1 AssayCulture supernatants were spun for 10 min at 10 000 g at41C, frozen at �201C in several aliquots (500 ml) until assayfor ET-1, sVCAM, IL-6 and sICAM by ELISA (commerciallyavailable kits BBE5, BBE3, D6050, and BBE 1B assay Kit, R&DSystem, Abingdon, UK). The results for the supernatant from5�105 plated cells are expressed in pg/ml for ET-1 and inng/ml for sICAM-1, IL-6 and sVCAM-1.
RNA Extraction and Reverse TranscriptionTotal RNA was extracted from the cultured cells using acommercial kit (RNeasy, Qiagen, Valencia, CA, USA). Briefly,50 ml of the RNA preparation was mixed with 10 U of RQ1RNAse free DNAse (Promega, Madison, WI, USA) andincubated for 30 min at 371C in the presence of DNAsebuffer. Aliquots of 3 mg of RNA were annealed to 80 pg ofrandom primer mixture (Life Technologies, Grand Island,NY, USA) at 701C for 10 min and then the tubes were placedin ice. The reverse transcription reaction in a final volume of50 ml, containing 10 ml of 5� RT buffer, 8 ml of 100 mM
dithiothreitol, 1 ml (40 U) of RNAsin (Promega, Madison,WI, USA) and 4 ml of 25 mM dNTPs, was initiated by adding200 U of Superscript II reverse transcriptase from MoloneyMurine Leukemia Virus (Life Technologies Grand Island, NY,USA) and incubated for 1 h at 371C. The reaction wasstopped by incubating the reaction mixture at 701C for10 min. The tubes were stored at –201C until the quantita-tive PCR assay.
Real-Time Quantitative PCR Assay for ET-1, ICAM-1 andVCAM-1 mRNATo evaluate the expression of ET-1, ICAM-1 and VCAM-1mRNA levels under different experimental conditions, real-time quantitative PCR was performed using an ABI PRISM7700 Sequence Detector (PE Applied Biosystem, Foster City,CA, USA). The fluorogenic Taqman probes and the primerswere designed using the Primer Express Software 1.7 (PEApplied Biosystem, Foster City, CA, USA). The probesconsisted of an oligonucleotide with a 50 FAM (6-carboxy-fluorescein) reporter dye and a 30 TAMRA (6-carboxy-tetramethylrhodamine) quencher dye.
A fragment of 115 bp extending from the untranslated 50
region of the ET-1 mRNA to the 30 end of the coding regionwas amplified using the following primers: forward hET-1(A)50-ACGGCGGGGAGAAACC-30 and reverse hET-1(B) 50-AT-GATGTCCAGGTGGCAGAAG-30. For ICAM-1, a fragment of102 bp corresponding to the extracellular domain wasamplified using the following primers: forward hICAM-1
(A) 50-GCAATGTGCAAGAAGATAGCCA-30 and reverse hI-CAM-1 (B) 50-GGGCAAGACCTCAGGTCATGT-30. A 125 bpfragment of the region encoding extracellular domain ofVCAM-1 was amplified using the following primers:hVCAM-1 (A) 50-GAGTACGCAAACACTTTATGTCAATGT-30
and reverse hVCAM-1 (B) 50-CTCGTCCTTTCGGGACCG-30.The quality of the PCR products was checked by size analysisand by direct nucleotide sequencing.
To normalize the quantitative data, TATA binding protein(TBP) mRNA was used as an internal control (detailed below)with the following primers: forward TBP primer: 50-CAC-GAACCACGGCACTGATT-30 and reverse TBP primer: 50-TTTTCTTGCTGCCAGTCTGGAC-30, which provide an am-plicon of 88 bp.
Sequences of the fluorogenic probes are as follows: ET-1probe 50-(FAM)-TGCTCCCTGCTCGTCCCTGATGGATA-30-(TAMRA); TBP probe 50-(FAM)-TGTGCACAGGAGCCAA-GAGTGAAGA-30-(TAMRA); VCAM-1 probe 50-(FAM)-AACCGTCTTGGTCAGCCCTTCCTCC-30-(TAMRA); ICAM-1probe 50-(FAM)-CAATGTGCTATTCAAACTGCCCTGATGGG-30-(TAMRA). cDNA samples were diluted three-fold with theMaster Mix (PE Applied Biosystem, Foster City, CA, USA)containing MgCl2, PCR buffer, dNTP, AmpErase UNG andAmpliTaq Gold DNA polymerase to a final volume of 25 mland were run for real-time quantitative PCR under thefollowing thermal conditions: initially 2 min at 501C and10 min at 951C and then 45 cycles of 15 s at 951C and 1 minat 601C.
Normalization of the Quantitative PCR DataPrior to quantitative analysis of the samples, a standardcurve for threshold cycle (CT) vs amount of RNA (for eachcell type) was established to account for the differences inPCR efficiency between the test ET-1, ICAM-1, VCAM-1mRNA and the control (TBP) mRNA. For this purpose, RNAconcentrations of 1, 10, 100 pg and 1 ng (diluted with asolution of tRNA to keep the total RNA concentrationconstant) were used. Slopes of the standard curves generatedwere always between 3.2 and 3.6 corresponding to a PCRefficiency of 85–90% with a correlation coefficient consis-tently higher than 0.9. The variations in Ct for TBP mRNAdid not exceed one between experiments. For each sample,the test/TBP mRNA ratio was calculated and the results aregiven as the percentage of relative expression in treated vsnontreated cells.
Flow Cytometry Analysis for mbICAM-1 and mbVCAM-1ExpressionCells (final cell density of 5�105 cells/ml) were incubatedfor 2 h at 41C in 100 ml PBS containing 3% fetal calf serum(FCS) both in the presence and absence of 1 mg of ICAM-1and VCAM-1 antibodies (R&D System, Abingdon, UK). Thenthe cells were washed, resuspended in the same buffer in thepresence of 100 ml FITC-labeled goat anti-mouse Igs andincubated at 41C for 1 h in the dark. After three washes withPBS containing 3% of FCS, the cells were fixed with 500 ml of4% paraformaldehyde and a total of 5000 cells were
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions byHU
M Brun et al
10
The Pharmacogenomics Journal

analyzed in an Epics-Elite flow cytometer (Coulter Electro-nic, Hialeah, FL, USA). Results are expressed as MFI.
NO and NO SynthaseBoth nitrites and nitrates, as indirect measures of NO, wereassessed in the culture supernatants using the Griessreaction.17 The eNOS in cell lysates were determined byELISA using a commercial kit (R&D System, Abingdon, UK).To explore the expression of the iNOS gene, we carried out areal-time quantitative PCR with Syber Green probe using thefollowing primers : forward primer iNOS (c) 50-GGACCA-CATCTACCAGGAGGAG-30 and reverse primer iNOS (d) 50-CCTGAACATAGACCTTGGGCT-30, which allowed amplifi-cation of a 111 bp fragment encompassing the sequence ofexon 25.
StatisticsStatistical analysis was performed with the nonparametricMann–Whitney test using GraphPad Prism (Graph PadSoftware, San Diego, CA, USA) and a difference betweenthe groups was considered significant when Po0.05.
ACKNOWLEDGEMENTSWe are very grateful to Babette B Weskler from Cornell UniversityMedical College, NY, USA and Cora-Jean S Edgell from University ofNorth Carolina, Chapel Hill, USA for providing the TrHBMEC andEA-hy 926 cell lines, respectively. We thank Mary Dominique Hardy-Dessources for IL-6 experiments (Guadeloupe, FWI). We also thankHelene Cave and Cecile Acquaviva (Service de Biochimie Genetique,Hospital Robert Debre, Paris, France) for technical assistance forreal-time quantitative PCR. Manuel Brun is a recipient of Doctoralfellowship from the French Ministere de la recherche et de laTechnologie.DUALITY OF INTERESTNone declared.
REFERENCES1 Dover GJ, Humphries RK, Moore JG et al. Hydroxyurea induction of
hemoglobin F production in sickle cell disease: relationship betweencytotoxicity and F cell production. Blood 1986; 67: 735–738.
2 Miller BA, Platt O, Hope S, Dover G, Nathan DG. Influence ofhydroxyurea on fetal hemoglobin production in vitro. Blood 1987; 70:1824–1829.
3 Charache S, Terrin ML, Moore RD et al. Effect of hydroxyurea on thefrequency of painful crises in sickle cell anemia. Investigators of theMulticenter Study of Hydroxyurea in Sickle Cell Anemia. N Engl J Med1995; 332: 1317–1322.
4 Charache S, Terrin ML, Moore RD et al. Design of the multicenter studyof hydroxyurea in sickle cell anemia. Investigators of the MulticenterStudy of Hydroxyurea. Control Clin Trials 1995; 16: 432–446.
5 Maier-Redelsperger M, Labie D, Elion J. Long-term hydroxyureatreatment in young sickle cell patients. Curr Opin Hematol 1999; 6:115–120.
6 Ballas SK, Dover GJ, Charache S. Effect of hydroxyurea on therheological properties of sickle erythrocytes in vivo. Am J Hematol1989; 32: 104–111.
7 Goldberg MA, Brugnara C, Dover GJ, Schapira L, Charache S, Bunn HF.Treatment of sickle cell anemia with hydroxyurea and erythropoietin. NEngl J Med 1990; 323: 366–372.
8 Ballas SK, Marcolina MJ, Dover GJ, Barton FB. Erythropoietic activity inpatients with sickle cell anaemia before and after treatment withhydroxyurea. Br J Haematol 1999; 105: 491–496.
9 Adragna NC, Fonseca P, Lauf PK. Hydroxyurea affects cell morphology,cation transport, and red blood cell adhesion in cultured vascularendothelial cells. Blood 1994; 83: 553–560.
10 Aquino SL, Gamsu G, Fahy JV et al. Chronic pulmonary disorders insickle cell disease: findings at thin-section CT. Radiology 1994; 193:807–811.
11 Styles LA, Lubin B, Vichinsky E, Lawrence S, Hua M, Test S et al.Decrease of very late activation antigen-4 and CD36 on reticulocytes insickle cell patients treated with hydroxyurea. Blood. 1997; 89: 2554–2559.
12 Gee BE, Platt OS. Sickle reticulocytes adhere to VCAM-1. Blood 1995;85: 268–274.
13 Pober JS, Cotran RS. The role of endothelial cells in inflammation.Transplantation 1990; 50: 537–544.
14 Solovey A, Lin Y, Browne P, Choong S, Wayner E, Hebbel RP. Circulatingactivated endothelial cells in sickle cell anemia. N Engl J Med 1997; 337:1584–1590.
15 Schweitzer KM, Vicart P, Delouis C et al. Characterization of a newlyestablished human bone marrow endothelial cell line: distinct adhesiveproperties for hematopoietic progenitors compared with humanumbilical vein endothelial cells. Lab Invest 1997; 76: 25–36.
16 Edgell CJ, McDonald CC, Graham JB. Permanent cell line expressinghuman factor VIII-related antigen established by hybridization. Proc NatlAcad Sci USA 1983; 80: 3734–3737.
17 Miles AM, Wink DA, Cook JC, Grisham MB. Determination of nitricoxide using fluorescence spectroscopy. Methods Enzymol 1996; 268:105–120.
18 Marsden PA, Brenner BM. Transcriptional regulation of the endothelin-1gene by TNF-alpha. Am J Physiol 1992; 262: C854–C861.
19 Hammerman SI, Kourembanas S, Conca TJ, Tucci M, Brauer M, FarberHW. Endothelin-1 production during the acute chest syndrome in sicklecell disease. Am J Respir Crit Care Med 1997; 156: 280–285.
20 Phelan M, Perrine SP, Brauer M, Faller DV. Sickle erythrocytes,after sickling, regulate the expression of the endothelin-1 gene andprotein in human endothelial cells in culture. J Clin Invest 1995; 96:1145–1151.
21 Sanz MJ, Johnston B, Issekutz A, Kubes P. Endothelin-1 causes P-selectin-dependent leukocyte rolling and adhesion within rat mesentericmicrovessels. Am J Physiol 1999; 277: H1823–H1830.
22 Helset E, Kjaeve J, Hauge A. Endothelin-1-induced increases inmicrovascular permeability in isolated, perfused rat lungs requiresleukocytes and plasma. Circ Shock 1993; 39: 15–20.
23 Knofler R, Urano T, Malyszko J, Takada Y, Takada A. In vitro effectof endothelin-1 on collagen, and ADP-induced aggregation in humanwhole blood and platelet rich plasma. Thromb Res 1995; 77:69–78.
24 Rivera A, Jarolim P, Brugnara C. Modulation of Gardos channel activityby cytokines in sickle erythrocytes. Blood 2002; 99: 357–603.
25 Graido-Gonzalez E, Doherty JC, Bergreen EW, Organ G, Telfer M,McMillen MA. Plasma endothelin-1, cytokine, and prostaglandin E2levels in sickle cell disease and acute vaso-occlusive sickle crisis. Blood1998; 92: 2551–2555.
26 Rybicki AC, Benjamin LJ. Increased levels of endothelin-1 in plasma ofsickle cell anemia patients (letter). Blood 1998; 92: 2594–2596.
27 Stankova J, D’Orleans-Juste P, Rola-Pleszczynski M. ET-1 induces IL-6gene expression in human umbilical vein endothelial cells: synergisticeffect of IL-1. Am J Physiol 1996; 271: C1073–C1078.
28 Ali MH, Schlidt SA, Chandel NS, Hynes KL, Schumacker PT, Gewertz BL.Endothelial permeability and IL-6 production during hypoxia: role ofROS in signal transduction. Am J Physiol 1999; 277: L1057–L1065.
29 Jiang J, Jordan SJ, Barr DP, Gunther MR, Maeda H, Mason RP. In vivoproduction of nitric oxide in rats after administration of hydroxyurea.Mol Pharmacol 1997; 52: 1081–1086.
30 Gladwin MT, Shelhamer JH, Ognibene FP et al. Nitric oxide donorproperties of hydroxyurea in patients with sickle cell disease. Br JHaematol 2002; 116: 436–444.
31 Head CA, Brugnara C, Martinez-Ruiz R et al. Low concentrations of nitricoxide increase oxygen affinity of sickle erythrocytes in vitro and in vivo. JClin Invest 1997; 100: 1193–1198.
32 Martinez-Ruiz R, Montero-Huerta P, Hromi J, Head CA. Inhaled nitricoxide improves survival rates during hypoxia in a sickle cell (SAD)mouse model. Anesthesiology 2001; 94: 1113–1118.
33 Sullivan KJ, Goodwin SR, Evangelist J, Moore RD, Mehta P. Nitric oxidesuccessfully used to treat acute chest syndrome of sickle cell disease in ayoung adolescent. Crit Care Med 1999; 27: 2563–2568.
Q2
Q1
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions by HUM Brun et al
11
www.nature.com/tpj

34 Nahavandi M, Wyche MQ, Perlin E, Tavakkoli F, Castro O. Nitric oxidemetabolites in sickle cell anemia patients after oral administration ofhydroxyurea; hemoglobinopathy. Hematology 2000; 5: 335–339.
35 Hayflick JS, Kilgannon P, Gallatin WM. The intercellular adhesionmolecule (ICAM) family of proteins. New members and novel functions.Immunol Res 1998; 17: 313–327.
36 Ockenhouse CF, Betageri R, Springer TA, Staunton DE. Plasmodiumfalciparum-infected erythrocytes bind ICAM-1 at a site distinct from LFA-1, Mac-1, and human rhinovirus. Cell 1992; 68: 63–69.
37 Most J, Zangerle R, Herold M et al. Elevated concentrations ofcirculating intercellular adhesion molecule 1 (ICAM-1) in HIV-1infection. J Acquir Immune Deficiency Syndrome 1993; 6: 221–226.
38 Gearing AJ, Hemingway I, Pigott R, Hughes J, Rees AJ, Cashman SJ.Soluble forms of vascular adhesion molecules, E-selectin, ICAM-1, andVCAM-1: pathological significance. Ann NY Acad Sci 1992; 667: 324–331.
39 Benkerrou M, Delarche C, Brahimi L et al. Hydroxyurea corrects thedysregulated L-selectin expression and increased H(2)O(2) production ofpolymorphonuclear neutrophils from patients with sickle cell anemia.Blood 2002; 99: 2297–2303.
40 Shiu YT, Udden MM, McIntire LV. Perfusion with sickle erythrocytes up-regulates ICAM-1 and VCAM-1 gene expression in cultured humanendothelial cells (In Process Citation). Blood 2000; 95: 3232–3241.
41 Ohno N, Ichikawa H, Coe L, Kvietys PR, Granger DN, Alexander JS.Soluble selectins and ICAM-1 modulate neutrophil–endothelial adhe-sion and diapedesis in vitro. Inflammation 1997; 21: 313–324.
42 Wakatsuki T, Kimura K, Kimura F et al. A distinct mRNA encoding asoluble form of ICAM-1 molecule expressed in human tissues. Cell AdhesCommun 1995; 3: 283–292.
43 Budnik A, Grewe M, Gyufko K, Krutmann J. Analysis of the production ofsoluble ICAM-1 molecules by human cells. Exp Hematol 1996; 24: 352–359.
44 Champagne B, Tremblay P, Cantin A, St Pierre Y. Proteolytic cleavage ofICAM-1 by human neutrophil elastase. J Immunol 1998; 161: 6398–6405.
45 Dean DC, Iademarco MF, Rosen GD, Sheppard AM. The integrin alpha4 beta 1 and its counter receptor VCAM-1 in development and immunefunction. Am Rev Respir Dis 1993; 148: S43–S46.
46 Swerlick RA, Eckman JR, Kumar A, Jeitler M, Wick TM. Alpha 4 beta 1-integrin expression on sickle reticulocytes: vascular cell adhesionmolecule-1-dependent binding to endothelium. Blood 1993; 82:1891–1899.
47 Lutty GA, Taomoto M, Cao J et al. Inhibition of TNF-alpha-inducedsickle RBC retention in retina by a VLA-4 antagonist. Invest OphthalmolVis Sci 2001; 42: 1349–1355.
48 Setty BN, Stuart MJ. Vascular cell adhesion molecule-1 is involved inmediating hypoxia-induced sickle red blood cell adherence toendothelium: potential role in sickle cell disease. Blood 1996; 88:2311–2320.
49 Solovey AA, Solovey AN, Harkness J, Hebbel RP. Modulation ofendothelial cell activation in sickle cell disease: a pilot study. Blood2001; 97: 1937–1941.
50 Saleh AW, Hillen HF, Duits AJ. Levels of endothelial, neutrophil andplatelet-specific factors in sickle cell anemia patients during hydroxyur-ea therapy. Acta Haematol 1999; 102: 31–37.
51 Degitz K, Li LJ, Caughman SW. Cloning and characterization of the 50-transcriptional regulatory region of the human intercellular adhesionmolecule 1 gene. J Biol Chem 1991; 266: 14024–14030.
52 Iademarco MF, McQuillan JJ, Rosen GD, Dean DC. Characterization ofthe promoter for vascular cell adhesion molecule-1 (VCAM-1). J BiolChem 1992; 267: 16323–16329.
53 Berendt AR, Simmons DL, Tansey J, Newbold CI, Marsh K. Intercellularadhesion molecule-1 is an endothelial cell adhesion receptor forPlasmodium falciparum. Nature 1989; 341: 57–59.
Downregulation of endothelin-1 and upregulation of ICAM-1 gene expressions byHU
M Brun et al
12
The Pharmacogenomics Journal





























