Embed Size (px)
Citation preview

©2004 FASEB
The FASEB Journal express article 10.1096/fj.03-1336fje. Published online September 22, 2004. Acute myocardial hypoxia increases BNP gene expression Jens Peter Goetze,*Alicia Gore,* Christian H. Møller,† Daniel A. Steinbrüchel,† Jens F. Rehfeld,* and Lars B. Nielsen* Departments of *Clinical Biochemistry and †Thoracic Surgery, Rigshospitalet, University of Copenhagen, Copenhagen, Denmark
Corresponding author: Lars B. Nielsen, Department of Clinical Biochemistry, KB 3011 Rigshospitalet, University of Copenhagen, 9 Blegdamsvej, DK-2100, Copenhagen, Denmark. E-mail: [email protected]
ABSTRACT
It is well established that cardiac failure increases cardiac B-type natriuretic peptide (BNP) expression due to myocardial stretching. However, patients with ischemic heart disease also display increased plasma BNP and proBNP concentrations despite preserved cardiac function. In this study, we examined whether acute myocardial hypoxia increases cardiac BNP expression. Surgical reduction of the blood flow to an area of the anterior ventricular wall in pigs reduced the myocardial oxygen tension from 46 ± 4 to 13 ± 5 mmHg. The tissue contents of VEGF and BNP mRNA increased 1.8-fold and 3.5-fold, respectively (n=10, P<0.005) in hypoxic compared with normoxic ventricular myocardium after 2.2 ± 0.2 h; the magnitude of the increase in BNP mRNA expression was positively correlated with that of VEGF in hypoxic myocardium (r=0.66, P<0.05). In support of a hypoxia-induced increase of BNP gene transcription, the content of a premature BNP mRNA was increased in hypoxic myocardium (4.8-fold, P<0.005) and in freshly harvested ventricular myocytes when kept in culture flasks and oxygen-deprived for 3 h (2.2-fold, P=0.002). ProBNP peptide accumulated in the medium of freshly harvested ventricular myocyte cultures but was undetectable in ventricular myocardium, indicating rapid release of the newly synthesized proBNP peptide. Accordingly, the plasma proBNP concentration increased after 2 h of myocardial hypoxia (P=0.028). Cumulatively, the data suggest that acute hypoxia stimulates cardiac BNP expression.
Key words: ANP • B-type natriuretic peptide • heart failure • ischemic heart disease
ardiac secretion of B-type natriuretic peptide (BNP) is increased in congestive heart failure (1, 2). Accordingly, increased plasma concentrations of BNP and its biosynthetic precursor, proBNP, are markers of left ventricular systolic dysfunction (3). The main
stimulus for the increased BNP expression is ventricular dilatation and concomitant myocyte stretching, which stimulate p38 mitogen-activated protein kinase (MAPK). p38 MAPK activates the transcription factor NF-κB and BNP gene transcription (4). Vasoactive substances like catecholamines and angiotensin-II also increase BNP gene transcription in a p38 MAPK-dependent manner (5–7). Moreover, myocyte stretching increases the intracellular calcium concentration (8) and modulates calcium-binding proteins that in turn regulate downstream modulators, including calcineurin, which in turn stimulates myocyte BNP gene expression (9). Thus, cardiac BNP gene expression can be modulated in vitro by blocking the p38 MAPK and
C
Page 1 of 18(page number not for citation purposes)

the calcineurin pathways (4, 10, 11) and in vivo by pharmacological intervention in heart failure patients (12–14).
The diagnostic specificity of increased plasma BNP and proBNP concentrations is nevertheless remarkably low in heart failure (15–17). This implies that other stimuli besides myocyte stretching and neurohormonal activation may be involved in regulating cardiac BNP gene expression. Plasma BNP and proBNP concentrations in patients with acute coronary syndromes and myocardial infarction are also increased (18–24). Interestingly, the increased plasma concentrations precede and predict later development of heart failure (20, 21). Moreover, ventricular BNP gene expression is associated with increased plasma BNP and proBNP concentrations in stable ischemic heart disease patients without ventricular dysfunction (25). A recent report disclosed that hypoxia stimulates the atrial natriuretic peptide gene promoter and that this effect is mediated by the heterodimeric transcription factor hypoxia-inducible factor HIF-1 (26). HIF-1 is stabilized by low oxygen tension (27) and enhances transcription of several genes, including the vascular endothelial growth factor (VEGF) gene (28). The BNP promoter region also has a HIF-1 binding site, and cultured myocytes transfected with a stable hybrid form of HIF-1 display increased BNP mRNA expression (29). Moreover, hypobaric hypoxia stimulates both ANP and BNP gene transcription (30) and peptide release in vitro from perfused rat heart preparations (31). Interestingly, ANP and BNP mediate vasodilatation of the coronary arteries, which suggest a local regulatory effect on the myocardial blood supply (32–35). We therefore hypothesized that myocardial hypoxia per se may increase ventricular BNP expression in vivo.
To test this idea, we established a porcine model of reduced blood flow to an area of the left ventricle and compared the BNP expression in hypoxic and normoxic ventricular myocardium. The results suggest that acute myocardial hypoxia increases cardiac BNP gene transcription and raises the plasma proBNP concentration.
MATERIALS AND METHODS
Myocardial hypoxia
Twenty-one Danish-bred pigs were used in this study. To examine myocardial hypoxia, 14 pigs were anesthetized with a mixture of zolazepam (11.9 mg/mL), tiletamine (11.9 mg/mL), xylazine (12.38 mg/mL), ketamine (14.29 mg/mL), and methadone (2.38 mg/mL). The mixture was injected intramuscularly (0.1 mL/kg), and the pigs were intubated and ventilated with 1 L oxygen and 2 L of atmospheric air/min. The anesthesia was maintained on isoflurane (3%) and intravenous haldide (400 µg/h). Amiodarone (300 mg) was administered intravenously before sternotomy to minimize later arrhythmic complications. After sternotomy, the left internal mammary artery was mobilized and anastomosed to the left ascending interventricular vein. Ligation bands were placed without closing around the left anterior descending interventricular artery (LAD). The LAD was closed, and blood flow from the anastomozed left internal mammary artery to the interventricular vein was established. In this way, a limited arterial blood supply to the anterior ventricular wall was ensured. The intramyocardial oxygen tension was continuously monitored with a Revoxode pO2 probe (Licox CMP instruments, Plainsboro, NJ; 36). The central venous pressure and heart rate were also monitored continuously, and dobutamine was administered when necessary to maintain normal hemodynamics. Four pigs
Page 2 of 18(page number not for citation purposes)

developed ventricular fibrillation and were excluded from the study. Control pigs (n=4) underwent the same procedures, except that the anastomosis between the mammary artery and the interventricular vein was omitted and the LAD was not closed. The pigs were killed with pentobarbital 2.2 ± 0.2 h after induction of myocardial hypoxia. The experimental animal protocol was approved by the local ethics committee for animal research.
Myocardial tissue and plasma
Transmural biopsies (0.5–1.0 g) were collected immediately after sacrifice from the normoxic inferior left ventricular wall, from a hypoxic region adjacent to the pO2 probe in the anterior left ventricular wall, and from the appendage of the left atrium. The biopsies were immediately frozen in liquid nitrogen and stored at –80°C. Blood was collected before induction of myocardial hypoxia and 2 h after in Na2-EDTA (1.5 mg/mL) containing tubes and the plasma was stored at –80°C.
Freshly harvested ventricular cells
To access myocyte gene expression and peptide release in the absence of mechanical stimulation, three pigs were killed with pentobarbital and transmural biopsies (~2.0 g) were obtained from the anterior left ventricular wall and the appendage of the left atrium (~2 g tissue/biopsy). The tissues were rapidly minced with a scalpel into 0.5 × 0.5 mm pieces and immersed in 37°C serum-free culture medium (Dulbecco’s MEM with L-alanyl-L-glutamine and 4.5 g/L glucose, Invitrogen, Taastrup, Denmark) supplemented with protease inhibitors (Complete EDTA-free, Roche, Hvidovre, Denmark). The minced tissue was washed twice in medium by centrifugation (1500 rpm, 10 min), resuspended in fresh medium, and transferred to culture flasks (~200 mg tissue in 5 mL medium). The flasks were kept at 37°C for 3 h in atmospheric air or in an anaerobic workstation (Concept 400, Ruskinn Technology, Leeds, UK). Subsequently, the cells and culture medium were separated by centrifugation and stored at –80°C. The medium oxygen tension was measured in an ABL 700 (Radiometer, Copenhagen, Denmark).
mRNA Purification and cDNA amplification
Total RNA was isolated with TRIzol (Invitrogen), and the integrity of RNA was ensured by 1% agarose gel electrophoresis or a RNA Nano LabChip kit on an Agilent 2100 Bioanalyzer (Agilent Technologies, Frankfurt, Germany). The contribution of contaminating DNA in the RNA preparations was negligible as judged from real-time PCR analysis of heart total RNA (i.e., prior to cDNA synthesis) with primers that amplify premature BNP. First-strand cDNA was synthesized from 1 µg RNA with M-MULV reverse transcriptase (40 U, Roche). We have previously reported that the porcine heart contains both mature BNP mRNA and a premature BNP mRNA (37). The mature BNP mRNA was measured with primers: 5′-CCCGCAGTAGCATCTTCCA-3′; and 5′-TTGCTTTGAAGGGGAGCAG-3′ or as described previously (37). The immature BNP pre-mRNA was amplified with primers: 5′-GGGACCAAACTCCGAGAAA-3′; and 5′-CAGCCAGGCAATGACAGAAT-3′. VEGF mRNA was measured with primers: 5′-TGAGCTTCCTACAGCACAACA-3′; and 5′-ACCGCCTCGGCTTGTC-3′. For normalization, β-actin mRNA was amplified as described previously (37) and GADPH mRNA with primers: 5′-TGGAAATCCCATCACCATCT-3′; and 5′-TTCATGCCCATCACAAACAT-3′ (Sigma
Page 3 of 18(page number not for citation purposes)

Genosys, Haverhill, UK). Agarose gel electrophoresis and DNA sequencing of polymerase chain reaction (PCR) products ensured the specificity of each reverse-transcription PCR reaction.
Real-time mRNA quantification
Quantitative real-time PCR analyses were performed with the Lightcycler and Faststart DNA Master SYBR Green I mix (Roche). Each PCR reaction contained cDNA synthesized from 20 or 200 ng total RNA. Parallel analyses of dilutions of heart cDNA pooled from normal pigs (n=5) were used to determine the relation between the time point of the log-linear increase of the fluorescence signal and the concentration of an mRNA transcript. The content of each transcript was always quantitated in duplicate: The intraassay coefficients of variation varied from 7% (BNP pre-mRNA) to 11% (BNP mRNA). The VEGF, BNP, and proBNP mRNA results were normalized to the contents a housekeeping gene (β-actin or GADPH) mRNA. The entire experimental procedures for mRNA quantification from RNA purification to real-time quantification was performed twice with similar results.
To assess the contribution of genomic DNA contamination of RNA samples to the BNP pre-mRNA measurements, we also performed real-time PCR quantification using total RNA rather than cDNA template. Those studies showed that the contribution of genomic DNA to the present results was 0% in most samples and always <20%. No difference was found in genomic contamination between samples from normoxic and hypoxic regions.
BNP and proBNP radioimmunoanalysis
Frozen myocardial biopsies were minced with a scalpel and boiled in H2O for 10 min. The suspension was centrifuged (10,000 g, 30 min) and stored at –80°C before radioimmunoanalysis (RIA). Plasma samples were extracted with ethanol as described previously (38). BNP in tissue extracts and cell culture medium was measured with a commercial RIA for porcine BNP (Phoenix Peptides, Karlsruhe, Germany) that detects porcine BNP-26 and BNP-32 without cross-reactivity to atrial natriuretic peptide (39). In our hands, this assay was not useful for plasma BNP measurements in the pigs without severe cardiac failure. ProBNP in tissue extracts, cell medium, and plasma was measured with a RIA specific for the N-terminus of porcine proBNP, which detects both the intact precursor and the N-terminal proBNP-derived fragments (37).
Chromatography
The BNP and proBNP immunoreactivity in culture medium from atrial cells was characterized by gel filtration chromatography. Culture medium (2 ml) was applied to a 1000 × 10 mm Sephadex G-50 Superfine column (Pharmacia, Uppsala, Sweden) and eluted at 4°C in a Tris buffer containing 2 g human serum albumin/L (flow rate: 4 mL/h). Void and total volume was determined by the elution of 125I-labeled albumin and 22Na, respectively.
Immunohistochemistry
Paraformaldehyde-fixed sections (4 µm) of porcine myocardium were incubated with anti-proBNP antiserum (1:100) or preimmune rabbit antiserum (control) (37). Bound antibodies were visualized with antirabbit antiserum conjugated with fluorescein isothiocyanate (Dako,
Page 4 of 18(page number not for citation purposes)

Copenhagen, Denmark) and a confocal laser-scanning microscope (LSM510, Zeiss, Göttingen, Germany).
Statistics
Data are expressed as mean ± SE. The Wilcoxon-matched pairs and Mann-Whitney tests were used for comparison of data within and between groups, respectively. P-values <0.05 were considered statistically significant.
RESULTS
Surgical reduction of the blood flow to the anterior left ventricular wall lead to slight discoloration of the affected myocardium (Fig. 1A) and a decrease of the oxygen tension in the affected region from 46 ± 4 to 13 ± 5 mmHg after 15 min (Fig. 1B). To see whether the reduced oxygen tension resulted in stimulation of HIF-1α regulated gene transcription, we quantified VEGF mRNA. After 2.2 ± 0.2 h, the expression of VEGF mRNA was increased 1.8-fold in the hypoxic region compared with normoxic myocardium (P<0.001, Fig. 1C). The content of BNP mRNA in the hypoxic myocardium was increased 3.5-fold compared with that in normoxic myocardium (P<0.005, Fig. 2A). A positive correlation was found between the increase in VEGF and BNP mRNA contents in the hypoxic myocardium (r=0.66, P<0.05, data not shown). The porcine myocardium contains significant amounts of immature BNP pre-mRNA (37). Quantification with a BNP pre-mRNA specific real-time PCR assay revealed that the BNP pre-mRNA content was 4.8-fold higher in the hypoxic compared with the normoxic myocardium (P<0.005, Fig. 2B). The VEGF mRNA, BNP mRNA, and BNP pre-mRNA contents were similar in the normoxic anterior and inferior left ventricular walls in control pigs (Figs. 1C, 2A, B).
To assess whether the increased BNP gene expression in hypoxic ventricular myocardium was accompanied by increased BNP peptide production, we analyzed BNP and proBNP peptides in myocardial tissue with radioimmunoassays and immunohistochemistry. However, neither hypoxic nor normoxic left ventricular myocardium contained detectable proBNP concentrations by RIA (Fig. 3A) or on immunohistochemistry (Fig. 3B). In contrast, left atrial tissue contained both BNP and proBNP peptides, with a markedly lower BNP than proBNP concentration (Fig. 3C). Confocal laser-scanning microscopy of immunostained atrial sections showed a granular-like appearance of proBNP extending along the contractile apparatus in the myocytes (Fig. 3D). Gel chromatography of medium from freshly harvested atrial cells kept revealed two forms of proBNP-derived peptides, i.e., the intact precursor and a smaller N-terminal fragment (Fig. 3E), and a small amount of BNP.
The plasma proBNP concentrations in hypoxic pigs increased 2 h after induction of myocardial hypoxia (P=0.028, Fig. 4A). In control pigs without surgical reduction of the blood flow, the proBNP plasma concentration was not significantly different from baseline values (95±10.2%, P=0.62). The absence of BNP and proBNP peptides in the ventricular myocardium may reflect rapid release of newly synthesized peptide from the myocytes. To examine this possibility, we kept freshly harvested ventricular cells for 3 h in serum-free culture medium. Although the cells did not contain detectable amounts of BNP or proBNP peptides before or after incubation, we found proBNP accumulation in the culture medium (P<0.0001, Fig. 4B). The oxygen tension and pH did not change in the culture medium during the experiments (data not shown).
Page 5 of 18(page number not for citation purposes)

Finally, to examine whether acute hypoxia also increases myocyte BNP gene transcription in the absence of mechanical stimulation, freshly harvested ventricular cells were incubated under oxygen-deprived conditions. The oxygen tension in the culture medium decreased from 159 ± 5 to 42 ± 5 mmHg, whereas pH did not change significantly during the 3 h incubation (7.45 ± 0.06 at 0 h vs. 7.32 ± 0.04 at 3 h). The BNP pre-mRNA content was 2.2-fold higher in oxygen-deprived cells compared with control cells (P=0.002, Fig. 5A). The difference in BNP mRNA content between oxygen-deprived and control cells was only borderline significant (P=0.08, Fig. 5B), suggesting that the hypoxic stimulation was less pronounced in the in vitro system than in vivo.
DISCUSSION
To study the effect of acute myocardial hypoxia on cardiac BNP expression, we used a new porcine model with low oxygen delivery to the left ventricular myocardium. Myocardial hypoxia rather than anoxia was achieved by ligation of the left anterior descending interventricular artery and partial restoration of the blood flow through a myocardial vein. The oxygen tension in the affected porcine myocardium was similar to that in patients with chronic ischemic heart disease (36,40). The experimental model therefore allowed us to study the BNP gene and peptide expression in both normoxic and hypoxic myocardium from the same ventricle, thus minimizing possible biases from anesthesia or neurohormonal activation during surgery.
A principal finding of the study was a robust increase in the BNP mRNA content of the left ventricular myocardium after 2 h of hypoxia (Fig. 2A). BNP mRNA contents are regulated at the level of stabilization/degradation, probably due to several AU-rich elements in the 3′- untranslated region (41). However, we suspect that our findings reflect increased BNP gene transcription rather than BNP mRNA stabilization, as the content of immature BNP pre-mRNA was even more increased than that of the mature BNP mRNA in the hypoxic myocardium (Fig. 2B).
It is well established that stretching of the myocyte leads to increased BNP gene transcription (42). Together with the present study, several observations now indicate that the oxygen tension in the myocardium is another regulator of BNP gene expression. First, the magnitude of the difference in BNP mRNA expression levels between the normoxic and hypoxic myocardial regions showed a positive linear correlation with the magnitude of the difference in VEGF mRNA levels between the two regions. VEGF mRNA expression is a principal sensor of the tissue oxygen content (28) and is regulated by HIF-1 (27). Second, keeping ventricular myocytes in a culture medium for 3 h (i.e., eliminating stretch-associated stimuli) under oxygen-deprived conditions also stimulated BNP gene transcription (Fig. 5). Third, recent data suggest that transfection of myocardial cells with a stable hybrid HIF-1 protein increases the BNP gene transcription (29), and in vitro hypoxia stimulates release of both ANP and BNP from perfused rat ventricular myocardium (31). Fourth, the BNP gene promoter region contains a HIF-1 responsive element upstream of the transcription start site (43). Finally, the ANP and BNP genes are closely related, and a recent report has disclosed that low oxygen also stimulates the ANP promoter and expression of the ANP gene in cultured cells (26). It is, however, important to emphasize that hypoxia can also stimulate the cardiac expression of a variety of genes, which in turn could affect the BNP gene transcription.
Page 6 of 18(page number not for citation purposes)

To see whether the increased BNP mRNA contents in hypoxic myocardium resulted in increased BNP peptide synthesis, we measured BNP and proBNP peptides in myocardial biopsies. ProBNP was undetectable in both normal and hypoxic ventricular biopsies (Fig. 3A, B). It has been suggested that normal ventricular myocytes secrete proBNP-derived peptides in a constitutive manner (44). However, this concept has primarily been based on indirect evidence, i.e., the absence of proBNP-derived peptides in tissue samples. To seek further evidence of a constitutive proBNP peptide release, we kept ventricular myocytes in a serum-free medium and measured BNP and proBNP peptides in the cells and medium. Although the ventricular tissue remained devoid of peptides, we found an accumulation of proBNP in the medium (Fig. 4B). In contrast to ventricular myocytes, the atrial myocytes stored proBNP in granular-like organelles (Fig. 3C). Interestingly, recent reports have revealed a cardiac-specific protease (corin) that is dominantly expressed in atrial myocytes and is suggested to cleave the precursor prior to secretion (45, 46). Although atrial cells were found to release both intact proBNP and an N-terminal fragment, only a relatively small amount of BNP was detected in the medium (BNP/proBNP ratio in culture medium = 3.3 ± 0.3%, Fig. 3E). Moreover, only proBNP and not BNP was detectable in medium from ventricular cells (Fig. 4B). This finding may suggest that proBNP could be the most sensitive plasma marker of a rapid rise in cardiac BNP gene expression. Unfortunately, the BNP immunoassay was not useful for plasma measurements. Thus, we could not test this result in vivo, and it cannot be excluded that altered expression or activity of corin may have lead to inefficient proBNP processing in the cultured myocytes.
Nevertheless, the present data suggest that the effect of acute myocardial hypoxia on BNP gene expression may be detectable by measuring proBNP concentrations in plasma. Although we cannot exclude that the rise in plasma proBNP in pigs with acute left ventricular hypoxia, at least in part, reflected stretching of the myocytes in the hypoxic region rather than the hypoxia per se, it is noteworthy that we did not see a rise in plasma proBNP in control pigs. The rise in plasma proBNP concentrations was small but achieved after only 2 h of hypoxia in a small area of the left ventricular wall (Fig. 1A). In patients with chronic myocardial hypoxia due to extensive coronary atherosclerosis, the plasma BNP and proBNP concentrations appeared to be elevated to an extent similar to that in patients with severe systolic dysfunction (25). This may imply that prolonged oxygen-deprivation, despite preserved viability of the myocardium and despite lack of detectable cardiac systolic dysfunction, increases the plasma BNP and proBNP concentrations. In support of this, a recent report confirmed an association between inducible ischemia and plasma BNP concentrations in patients with stable coronary artery disease (47). Interestingly, BNP is a potent vasodilator in the coronary vasculature (33–35). A hypoxia-induced increase in BNP gene expression makes teleological sense, if BNP functions in a paracrine regulatory mechanism that increases the local blood supply to the hypoxic area. Thus, even modest increases in BNP gene expression may have important local functions.
In conclusion, the present study suggests that acute myocardial hypoxia results in a rapid increase in cardiac BNP gene transcription, release of proBNP peptide from the ventricular myocytes, and a significant rise in plasma proBNP concentrations. Further studies are needed to establish the potential value of plasma proBNP measurements in patients with chronic myocardial hypoxia due to coronary atherosclerosis.
Page 7 of 18(page number not for citation purposes)

ACKNOWLEDGMENTS
The expert technical assistance of Lone Olsen, Lis Schütt Nielsen, Annemette Borch, and Karen Rasmussen is gratefully acknowledged. We also thank Letty Klarskov and Mette Olsen for assistance with the animal experiments, and Bo Lindberg for help with the immunohistochemistry. The study was supported by a grant from the Danish Heart Foundation.
REFERENCES
1. Mukoyama, M., Nakao, K., Saito, Y., Ogawa, Y., Hosoda, K., Suga, S., Shirakami, G., Jougasaki, M., and Imura, H. (1990) Increased human brain natriuretic peptide in congestive heart failure. N. Engl. J. Med. 323, 757–758
2. Mukoyama, M., Nakao, K., Hosoda, K., Suga, S., Saito, Y., Ogawa, Y., Shirakami, G., Jougasaki, M., Obata, K., Yasue, H., et al. (1991) Brain natriuretic peptide as a novel cardiac hormone in humans. Evidence for an exquisite dual natriuretic peptide system, atrial natriuretic peptide and brain natriuretic peptide. J. Clin. Invest. 87, 1402–1412
3. de Lemos, J. A., McGuire, D. K., and Drazner, M. H. (2003) B-type natriuretic peptide in cardiovascular disease. Lancet 362, 316–322
4. Liang, F., and Gardner, D. G. (1999) Mechanical strain activates BNP gene transcription through a p38/NF-kappaB-dependent mechanism. J. Clin. Invest. 104, 1603–1612
5. Hanford, D. S., Thuerauf, D. J., Murray, S. F., and Glembotski, C. C. (1994) Brain natriuretic peptide is induced by alpha 1-adrenergic agonists as a primary response gene in cultured rat cardiac myocytes. J. Biol. Chem. 269, 26227–26233
6. Liang, F., and Gardner, D. G. (1998) Autocrine/paracrine determinants of strain-activated brain natriuretic peptide gene expression in cultured cardiac myocytes. J. Biol. Chem. 273, 14612–14619
7. Wiese, S., Breyer, T., Dragu, A., Wakili, R., Burkard, T., Schmidt-Schweda, S., Fuchtbauer, E. M., Dohrmann, U., Beyersdorf, F., Radicke, D., et al. (2000) Gene expression of brain natriuretic peptide in isolated atrial and ventricular human myocardium: influence of angiotensin II and diastolic fiber length. Circulation 102, 3074–3079
8. Frey, N., McKinsey, T. A., and Olson, E. N. (2000) Decoding calcium signals involved in cardiac growth and function. Nat. Med. 6, 1221–1227
9. Kudoh, S., Akazawa, H., Takano, H., Zou, Y., Toko, H., Nagai, T., and Komuro, I. (2003) Stretch-modulation of second messengers: effects on cardiomyocyte ion transport. Prog. Biophys. Mol. Biol. 82, 57–66
10. Molkentin, J. D., Lu, J. R., Antos, C. L., Markham, B., Richardson, J., Robbins, J., Grant, S. R., and Olson, E. N. (1998) A calcineurin-dependent transcriptional pathway for cardiac hypertrophy. Cell 93, 215–228
Page 8 of 18(page number not for citation purposes)

11. Zhu, W., Zou, Y., Shiojima, I., Kudoh, S., Aikawa, R., Hayashi, D., Mizukami, M., Toko, H., Shibasaki, F., Yazaki, Y., et al. (2000) Ca2+/calmodulin-dependent kinase II and calcineurin play critical roles in endothelin-1-induced cardiomyocyte hypertrophy. J. Biol. Chem. 275, 15239–15245
12. Richards, A. M., Doughty, R., Nicholls, M. G., MacMahon, S., Sharpe, N., Murphy, J., Espiner, E. A., Frampton, C., and Yandle, T. G. (2001) Plasma N-terminal pro-brain natriuretic peptide and adrenomedullin: prognostic utility and prediction of benefit from carvedilol in chronic ischemic left ventricular dysfunction. Australia-New Zealand Heart Failure Group. J. Am. Coll. Cardiol. 37, 1781–1787
13. Stanek, B., Frey, B., Hulsmann, M., Berger, R., Sturm, B., Strametz-Juranek, J., Bergler-Klein, J., Moser, P., Bojic, A., Hartter, E., et al. (2001) Prognostic evaluation of neurohumoral plasma levels before and during beta-blocker therapy in advanced left ventricular dysfunction. J. Am. Coll. Cardiol. 38, 436–442
14. Latini, R., Masson, S., Anand, I., Judd, D., Maggioni, A. P., Chiang, Y. T., Bevilacqua, M., Salio, M., Cardano, P., Dunselman, P. H., et al. (2002) Effects of valsartan on circulating brain natriuretic peptide and norepinephrine in symptomatic chronic heart failure: the Valsartan Heart Failure Trial (Val-HeFT). Circulation 106, 2454–2458
15. Davidson, N. C., Naas, A. A., Hanson, J. K., Kennedy, N. S., Coutie, W. J., and Struthers, A. D. (1996) Comparison of atrial natriuretic peptide B-type natriuretic peptide, and N-terminal proatrial natriuretic peptide as indicators of left ventricular systolic dysfunction. Am. J. Cardiol. 77, 828–831
16. McDonagh, T. A., Robb, S. D., Murdoch, D. R., Morton, J. J., Ford, I., Morrison, C. E., Tunstall-Pedoe, H., McMurray, J. J., and Dargie, H. J. (1998) Biochemical detection of left-ventricular systolic dysfunction. Lancet 351, 9–13
17. Yamamoto, K., Burnett, J. C., Jr., Bermudez, E. A., Jougasaki, M., Bailey, K. R., and Redfield, M. M. (2000) Clinical criteria and biochemical markers for the detection of systolic dysfunction. J. Card. Fail. 6, 194–200
18. Morita, E., Yasue, H., Yoshimura, M., Ogawa, H., Jougasaki, M., Matsumura, T., Mukoyama, M., and Nakao, K. (1993) Increased plasma levels of brain natriuretic peptide in patients with acute myocardial infarction. Circulation 88, 82–91
19. Kikuta, K., Yasue, H., Yoshimura, M., Morita, E., Sumida, H., Kato, H., Kugiyama, K., Ogawa, H., Okumura, K., Ogawa, Y., et al. (1996) Increased plasma levels of B-type natriuretic peptide in patients with unstable angina. Am. Heart J. 132, 101–107
20. Richards, A. M., Nicholls, M. G., Yandle, T. G., Frampton, C., Espiner, E. A., Turner, J. G., Buttimore, R. C., Lainchbury, J. G., Elliott, J. M., Ikram, H., et al. (1998) Plasma N-terminal pro-brain natriuretic peptide and adrenomedullin: new neurohormonal predictors of left ventricular function and prognosis after myocardial infarction. Circulation 97, 1921–1929
Page 9 of 18(page number not for citation purposes)

21. de Lemos, J. A., Morrow, D. A., Bentley, J. H., Omland, T., Sabatine, M. S., McCabe, C. H., Hall, C., Cannon, C. P., and Braunwald, E. (2001) The prognostic value of B-type natriuretic peptide in patients with acute coronary syndromes. N. Engl. J. Med. 345, 1014–1021
22. Sabatine, M. S., Morrow, D. A., de Lemos, J. A., Gibson, C. M., Murphy, S. A., Rifai, N., McCabe, C., Antman, E. M., Cannon, C. P., and Braunwald, E. (2002) Multimarker approach to risk stratification in non-ST elevation acute coronary syndromes: simultaneous assessment of troponin I, C-reactive protein, and B-type natriuretic peptide. Circulation 105, 1760–1763
23. Jernberg, T., Stridsberg, M., Venge, P., and Lindahl, B. (2002) N-terminal pro brain natriuretic peptide on admission for early risk stratification of patients with chest pain and no ST-segment elevation. J. Am. Coll. Cardiol. 40, 437–445
24. Deo, R., and de Lemos, J. A. (2003) B-type Natriuretic Peptide in Ischemic Heart Disease. Curr. Cardiol. Rep. 5, 271–277
25. Goetze, J. P., Christoffersen, C., Perko, M., Arendrup, H., Rehfeld, J. F., Kastrup, J., and Nielsen, L. B. (2003) Increased cardiac BNP expression associated with myocardial ischemia. FASEB J. 17, 1105–1107
26. Chun, Y. S., Hyun, J. Y., Kwak, Y. G., Kim, I. S., Kim, C. H., Choi, E., Kim, M. S., and Park, J. W. (2003) Hypoxic activation of the atrial natriuretic peptide gene promoter through direct and indirect actions of hypoxia-inducible factor-1. Biochem. J. 370, 149–157
27. Semenza, G. L. (2003) Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 3, 721–732
28. Semenza, G. L. (2000) Hypoxia, clonal selection, and the role of HIF-1 in tumor progression. Crit. Rev. Biochem. Mol. Biol. 35, 71–103
29. Jiang, C., Lu, H., Vincent, K. A., Shankara, S., Belanger, A. J., Cheng, S. H., Akita, G. Y., Kelly, R. A., Goldberg, M. A., and Gregory, R. J. (2002) Gene expression profiles in human cardiac cells subjected to hypoxia or expressing a hybrid form of HIF-1 alpha. Physiol. Genomics 8, 23–32
30. Perhonen, M., Takala, T. E., Vuolteenaho, O., Mantymaa, P., Leppaluoto, J., and Ruskoaho, H. (1997) Induction of cardiac natriuretic peptide gene expression in rats trained in hypobaric hypoxic conditions. Am. J. Physiol. 273, R344–R352
31. Toth, M., Vuorinen, K. H., Vuolteenaho, O., Hassinen, I. E., Uusimaa, P. A., Leppaluoto, J., and Ruskoaho, H. (1994) Hypoxia stimulates release of ANP and BNP from perfused rat ventricular myocardium. Am. J. Physiol. 266, H1572–H1580
32. Chu, A., Morris, K. G., Kuehl, W. D., Cusma, J., Navetta, F., and Cobb, F. R. (1989) Effects of atrial natriuretic peptide on the coronary arterial vasculature in humans. Circulation 80, 1627–1635
Page 10 of 18(page number not for citation purposes)

33. Okumura, K., Yasue, H., Fujii, H., Kugiyama, K., Matsuyama, K., Yoshimura, M., Jougasaki, M., Kikuta, K., Kato, H., Tanaka, H., et al. (1995) Effects of brain (B-type) natriuretic peptide on coronary artery diameter and coronary hemodynamic variables in humans: comparison with effects on systemic hemodynamic variables. J. Am. Coll. Cardiol. 25, 342–348
34. Supaporn, T., Wennberg, P. W., Wei, C. M., Kinoshita, M., Matsuda, Y., and Burnett, J. C. (1996) Role for the endogenous natriuretic peptide system in the control of basal coronary vascular tone in dogs. Clin. Sci. 90, 357–362
35. Zellner, C., Protter, A. A., Ko, E., Pothireddy, M. R., DeMarco, T., Hutchison, S. J., Chou, T. M., Chatterjee, K., and Sudhir, K. (1999) Coronary vasodilator effects of BNP: mechanisms of action in coronary conductance and resistance arteries. Am. J. Physiol. 276, H1049–H1057
36. Perko, M. J., and Bay-Nielsen, H. (2002) Regional myocardial oxygenation during surgical revascularisation. Cardiovasc. Surg. 10, 590–594
37. Christoffersen, C., Goetze, J. P., Bartels, E. D., Larsen, M. O., Ribel, U., Rehfeld, J. F., Rolin, B., and Nielsen, L. B. (2002) Chamber-dependent expression of brain natriuretic peptide and its mRNA in normal and diabetic pig heart. Hypertension 40, 54–60
38. Rehfeld, J. F. (1998) Accurate measurement of cholecystokinin in plasma. Clin. Chem. 44, 991–1001
39. Qi, W., Kjekshus, H., Klinge, R., Kjekshus, J. K., and Hall, C. (2000) Cardiac natriuretic peptides and continuously monitored atrial pressures during chronic rapid pacing in pigs. Acta Physiol. Scand. 169, 95–102
40. Nielsen, L. B., Perko, M., Arendrup, H., and Andersen, C. B. (2002) Microsomal triglyceride transfer protein gene expression and triglyceride accumulation in hypoxic human hearts. Arterioscler. Thromb. Vasc. Biol. 22, 1489–1494
41. Sachs, A. B. (1993) Messenger RNA degradation in eukaryotes. Cell 74, 413–421
42. Durocher, D., Grepin, C., and Nemer, M. (1998) Regulation of gene expression in the endocrine heart. Recent Prog. Horm. Res. 53, 7–23
43. Luo, Y., Lu, H., Jiang, C., Belanger, A. J., Akita, G. Y., Wadaworth, S., Gregory, R. J., and Vincent, K. A. (2001) Expression of the human B-type natriuretic peptide (BNP) gene is activated by a HIF-1α/VP16 hybrid factor. Circulation 104, Suppl II, 202
44. de Bold, A. J., Ma, K. K., Zhang, Y., de Bold, M. L., Bensimon, M., and Khoshbaten, A. (2001) The physiological and pathophysiological modulation of the endocrine function of the heart. Can. J. Physiol. Pharmacol. 79, 705–714
Page 11 of 18(page number not for citation purposes)

45. Yan, W., Sheng, N., Seto, M., Morser, J., and Wu, Q. (1999) Corin, a mosaic transmembrane serine protease encoded by a novel cDNA from human heart. J. Biol. Chem. 274, 14926–14935
46. Yan, W., Wu, F., Morser, J., and Wu, Q. (2000) Corin, a transmembrane cardiac serine protease, acts as a pro-atrial natriuretic peptide-converting enzyme. Proc. Natl. Acad. Sci. USA 97, 8525–8529
47. Bibbins-Domingo, K., Ansari, M., Schiller, N. B., Massie, B., and Whooley, M. A. (2003) B-type natriuretic peptide and ischemia in patients with stable coronary disease. Circulation 108, 2987–2992
Received January 21, 2004; accepted August 18, 2004.
Page 12 of 18(page number not for citation purposes)
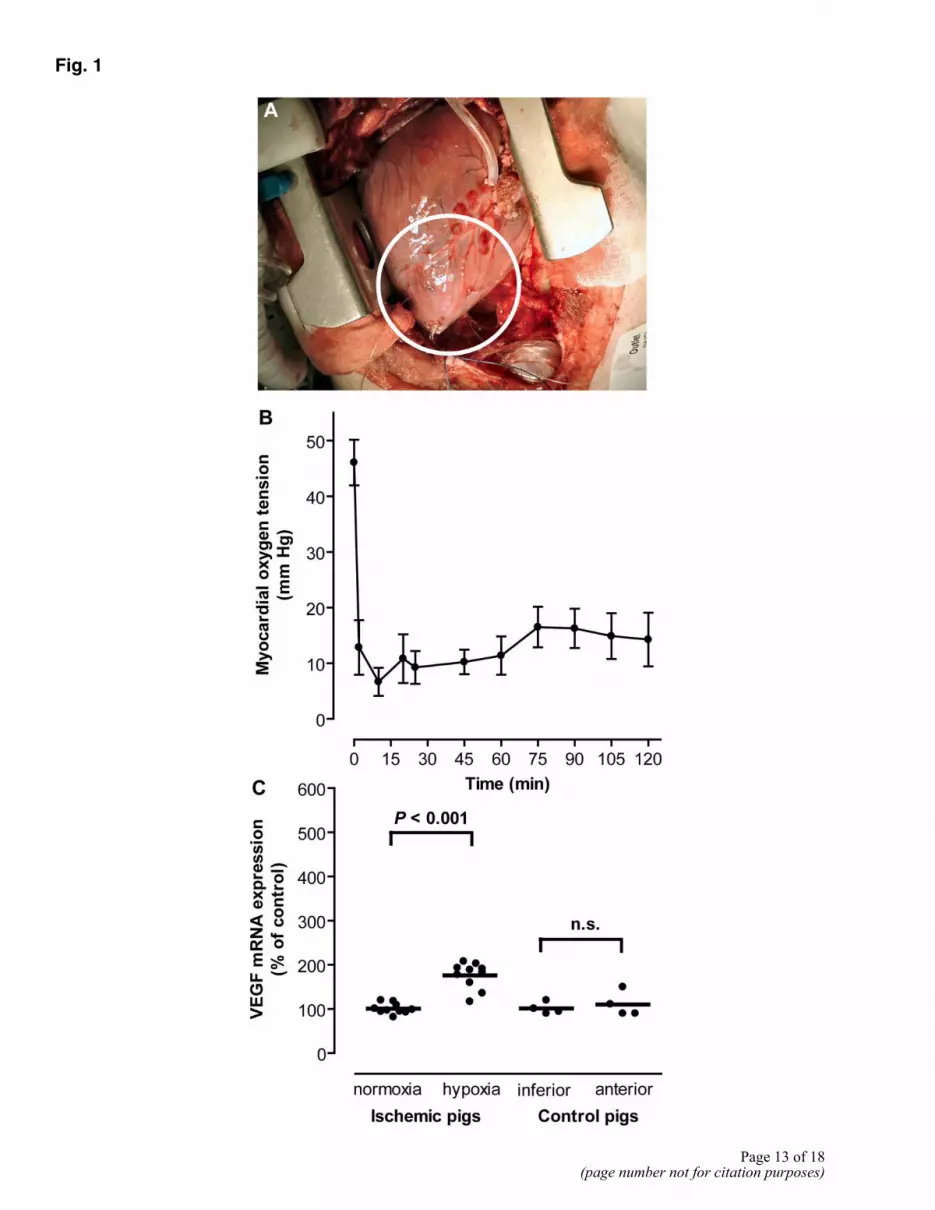
Fig. 1
Page 13 of 18(page number not for citation purposes)

Figure 1. Acute myocardial hypoxia in pigs. A) Shows the anterior left ventricular myocardium 5 min after induction of hypoxia. The white ring encircles the hypoxic myocardium. B) Shows the oxygen tension measured with a pO2-probe placed in myocardium of the affected ventricular region (n=10, mean ± SE). C) Shows the VEGF mRNA expression in a normoxic and the hypoxic region of the left ventricle (n=10). Also, VEGF mRNA expression in the same anatomical regions was determined in four control pigs. Data are expressed as percent of the mean values in the normoxic or inferior ventricular myocardium after normalization to a housekeeping gene, respectively. The horizontal lines indicate mean values, and each point represents data obtained from individual pigs.
Page 14 of 18(page number not for citation purposes)

Fig. 2
Figure 2. Effect of acute myocardial hypoxia on BNP gene transcription. A) Shows the mature BNP mRNA content expressed as percent of the mean values in the normoxic region (ischemic pigs, n=10) or inferior ventricular myocardium (control pigs, n=4). B) Shows the content of BNP pre-mRNA expressed as percent of the mean values in the normoxic (ischemic pigs) or inferior ventricular myocardium (control pigs). Horizontal lines indicate mean values, and each point represents data obtained from individual pigs. The results were normalized to a housekeeping gene.
Page 15 of 18(page number not for citation purposes)

Fig. 3
Figure 3. Myocardial BNP and proBNP peptide contents. A, C) Show the BNP and proBNP contents in ventricular and atrial biopsies, respectively. B) Shows the lack of proBNP contents in both normoxic (upper picture) and hypoxic (lower picture) ventricular myocytes by immunohistochemistry. The corresponding differential contrast image of the basic morphology of the section is shown to the right of each frame. D) Shows the proBNP immunostaining in atrial myocytes, and (E) shows the elution of N-terminal proBNP (two peaks) and BNP (one peak) immunoreactivity after gel filtration chromatography of culture medium from freshly harvested atrial cells kept for 3 h.
Page 16 of 18(page number not for citation purposes)

Fig. 4
Figure 4. Cellular proBNP release. A) Shows the plasma proBNP concentrations from pigs before induction of myocardial hypoxia (0 h) and 2 h after. Values from the same pig are connected by a line. B) Shows the concentration of proBNP (mean ± SE, n=4) in serum-free culture medium from freshly harvested ventricular cells and after a 3 h incubation in atmospheric air.
Page 17 of 18(page number not for citation purposes)

Fig. 5
Figure 5. BNP gene transcription in hypoxic ventricular cells in vitro. A) Shows the BNP pre-mRNA and (B) the mature BNP mRNA contents in ventricular cells incubated under normoxic (control) or anaerobic (hypoxia) conditions for 3 h. The horizontal lines indicate mean values, and each point represents data obtained from individual experiments.
Page 18 of 18(page number not for citation purposes)







![[Biochemical aspects of fetal hypoxia]](https://img.dokumen.tips/doc/110x75/635d79de88f33c6f8200b2b0/biochemical-aspects-of-fetal-hypoxia.jpg)





















