Embed Size (px)
Citation preview

doi:10.1182/blood-2008-12-195941Prepublished online May 19, 2009;2009 114: 844-859
Rius and Claire E. LewisRandall S. Johnson, Hongxia Z. Imityaz, M. Celeste Simon, Erik Fredlund, Florian R. Greten, Jordi Hsin-Yu Fang, Russell Hughes, Craig Murdoch, Seth B. Coffelt, Subhra K. Biswas, Adrian L. Harris, in primary macrophages experiencing hypoxiaHypoxia-inducible factors 1 and 2 are important transcriptional effectors
http://bloodjournal.hematologylibrary.org/content/114/4/844.full.htmlUpdated information and services can be found at:
(214 articles)Phagocytes, Granulocytes, and Myelopoiesis �Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Washington DC 20036.by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

PHAGOCYTES, GRANULOCYTES, AND MYELOPOIESIS
Hypoxia-inducible factors 1 and 2 are important transcriptional effectors inprimary macrophages experiencing hypoxiaHsin-Yu Fang,1 Russell Hughes,1 Craig Murdoch,2 Seth B. Coffelt,1 Subhra K. Biswas,3 Adrian L. Harris,4
Randall S. Johnson,5 Hongxia Z. Imityaz,6 M. Celeste Simon,6 Erik Fredlund,7 Florian R. Greten,8 Jordi Rius,9 andClaire E. Lewis1
1Academic Unit of Inflammation and Tumour Targeting and 2Department of Oral and Maxillofacial Medicine & Surgery, Faculty of Medicine, Dentistry and Health,University of Sheffield, Sheffield, United Kingdom; 3Singapore Immunology Network, Biomedical Sciences Institutes, Agency for Science, Technology &Research (A*STAR), Singapore; 4CRUK Molecular Oncology Laboratory, Weatherall Institute of Molecular Medicine, University of Oxford, Oxford, UnitedKingdom; 5Molecular Biology Section, University of California, San Diego School of Medicine, La Jolla; 6Abramson Family Cancer Research Institute,Philadelphia, PA; 7Center for Molecular Pathology, Lund University, University Hospital MAS, Malmo, Sweden; 8Second Department of Medicine, Klinikum rechtsder Isar, Technical University Munich, Munich, Germany; and 9Department of Pharmacology, University of California, San Diego School of Medicine, La Jolla
Ischemia exists in many diseased tis-sues, including arthritic joints, atheroscle-rotic plaques, and malignant tumors. Mac-rophages accumulate in these sites andup-regulate hypoxia-inducible transcrip-tion factors (HIFs) 1 and 2 in response tothe hypoxia present. Here we show thatthe gene expression profile in primaryhuman and murine macrophages changesmarkedly when they are exposed to hyp-oxia for 18 hours. For example, they wereseen to up-regulate the cell surface recep-
tors, CXCR4 and GLUT1, and the potent,tumor-promoting cytokines, vascular en-dothelial growth factor A, interleukin(IL)-1� and IL-8, adrenomedullin, CXCR4,and angiopoietin-2. Hypoxia also stimu-lated their expression and/or phosphory-lation of various proteins in the nuclearfactor-�B (NF-�B) signaling pathway. Wethen used both genetic and pharmaco-logic methods to manipulate the levels ofHIFs-1� and 2� or NF-�B in primary mac-rophages to elucidate their role in the
hypoxic induction of many of these keygenes. These studies showed that bothHIF-1 and -2, but not NF-�B, are importanttranscriptional effectors regulating the re-sponses of macrophages to such a pe-riod of hypoxia. Further studies usingexperimental mouse models are now war-ranted to investigate the role of suchmacrophage responses in the progres-sion of various diseased tissues, suchas malignant tumors. (Blood. 2009;114:844-859)
Introduction
Cells experience sustained periods of hypoxia in diseased tissues,such as malignant tumors, atherosclerotic plaques, and arthriticjoints.1-3 The predominant transcription factors mediating theeffects of hypoxia on gene expression are hypoxia-induciblefactors (HIFs) 1 and 2.4,5 These consist of distinct, hypoxia-responsive � subunits and an identical, constitutively expressed� subunit. In the presence of oxygen, the � subunits are hydroxy-lated by oxygen-sensitive enzymes called prolyl hydroxylases,which targets them for degradation by a ubiquitin-proteasomalpathway.4 In hypoxia, HIF� subunits accumulate and translocate tothe nucleus, couple with the HIF-1� subunit, and bind to hypoxicresponse elements (HREs) in the promoters of various genes,activating their transcription.4,5
Macrophages accumulate in most ischemic diseased sites,including tumors,6-9 where they accumulate both HIF-1� and2�10,11 and up-regulate HIF target genes, such as the potentproangiogenic growth factor, vascular endothelial growth factor A(VEGFA).12 There are conflicting views of the relative contributionof each HIF to the regulation of hypoxic gene expression in thesecells. Some studies suggest that the main form of HIF up-regulatedby tumor-associated macrophages (TAMs) is HIF-2,11,13 and over-expression of HIF-2� in normoxic human macrophages up-regulatesvarious proangiogenic genes.14 However, human macrophages also
markedly up-regulate HIF-1� when exposed to hypoxia in vitroand in tumors,10 and HIF-1�–deficient murine macrophages expresslower levels of such HIF-regulated genes as VEGF and the glucosereceptor GLUT1 in hypoxia than their wild-type counterparts.15
Interestingly, the exact contribution of HIFs-1 and -2 to theregulation of hypoxic gene expression appears to vary betweendifferent cell types. HIF-1, for example, mediates the induction ofvirtually all hypoxia-activated genes in mouse embryonic fibro-blasts and human breast tumor cells,16,17 whereas HIF-2 performsthis function in renal tumor cells.17 This depends partly on thecell type–specific expression of other transcription factors, such asElk-l, which bind to the promoters of some genes conferring HIF-2target specificity on them.18,19
Hypoxia may also employ another transcription factor, nuclearfactor-�B (NF-�B), as 2 major components of canonical NF-�Bsignaling, �B kinase � (IKK-�) and p65 (RelA) are activated whenmurine macrophages experience short-term (� 4 hours) hypoxia.This then up-regulates their expression of both HIF-1� and variousHIF target genes.20-22
In the present study, we show that exposure to hypoxia for18 hours markedly up-regulates a broad array of tumor-promotinggenes in primary macrophages, and then investigated the role ofHIFs-1 and -2 and NF-�B in this phenomenon.
Submitted December 22, 2008; accepted April 22, 2009. Prepublished onlineas Blood First Edition paper, May 19, 2009; DOI 10.1182/blood-2008-12-195941.
The publication costs of this article were defrayed in part by page charge
payment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 USC section 1734.
© 2009 by The American Society of Hematology
844 BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

Methods
Cells
Two forms of primary macrophages were used in this study: macrophagesdifferentiated in vitro from human peripheral blood (monocyte-derivedmacrophages [MDMs]) and bone marrow–derived macrophages (BMDMs)derived from bone marrow progenitors isolated from control (ie, wild-typeor �/�) mice or mice bearing myeloid cell-targeted deletions in either theHIF-1� or HIF-2� gene. Mouse studies were approved and conducted bythe Abramson Family Cancer Research Institute.
Isolation and culture of human MDMs. Monocytes were isolatedfrom Buffy coats (National Blood Service, Sheffield, United Kingdom) aspreviously described.10 A total of 50 � 106 mononuclear cells were seededin Iscove Modified Dulbecco Media (BioWhittaker UK Ltd) with 5%human AB serum (neat AB serum contains � 1 ng/mL human CSF-1) and2 mM L-glutamine (all from Sigma-Aldrich) and incubated at 37°C, 5%CO2. After 2 hours, adherent cells were washed and cultured for 7 days toallow differentiation into MDMs.
Isolation and culture of murine BMDMs. As previously described,22
BMDMs were isolated from the bones of control mice or mice bearing atargeted deletion of (1) the HIF-1� gene in myeloid cells (2loxP/1loxP,LysM Cre/� mice15) or (2) the HIF-2� gene in myeloid cells (2loxP/1loxP,LysM Cre/� mice; H.Z.I. and M.C.S., manuscript submitted, April 2009).
Bone marrow aspirates were washed and resuspended in medium with10% heat-inactivated fetal calf serum (BioWhittaker UK Ltd), 2 mML-glutamine (Sigma-Aldrich), 100 IU/mL penicillin and 100 g/mL strep-tomycin (BioWhittaker UK Ltd), and murine macrophage colony-stimulating factor (M-CSF; PeproTech Ltd), and cultured at 37°C, 5% CO2
for 7 days to allow macrophage differentiation. Their purity was assessedafter 7 days using an F4/80 antibody. Only BMDM cultures of more than90% purity were used in subsequent experiments.
Successful deletion of HIFs-1 or 2� has been demonstrated previouslyusing Southern and/or immunoblotting assays of extracts from hypoxicBMDMs from the HIF-1� LysM-Cre mice23 and HIF-2� LysM-Cre (H.Z.I.and M.C.S., manuscript submitted April 2009) mice used in this study.
Normoxic and hypoxic cell cultures
Human MDMs or murine BMDMs were subjected to severe hypoxia(� 0.5% O2) or normoxia (20.9% O2) in 5% CO2 humidified multigasincubators (Heto) for 18 hours.
siRNA treatment of human MDMs in vitro
siRNA duplexes for HIF-1� or HIF-2� were synthesized by EurogentecLaboratories. A randomly scrambled duplex was synthesized as a negativecontrol. The HIF-1� siRNA duplex sequences were composed of: sense,5-CUGAUGACCAGCAACUUGAdTdT-3; and antisense, 5-UCAAG-UUGCUGGUCAUCAGdTdT-3. The HIF-2� siRNA duplex sequences were:sense, 5-CAGCAUCUUUGAUAGCAGUdTdT-3; and antisense, 5-ACUGC-UAUCAAAGAUGCUGdTdT-3. The scrambled nonspecific duplex sequenceswere: sense, 5-AGUUCAACGACCAGUAGUCdTdT-3; and antisense, 5-GAC-UACUGGUCGUUGAdTdT-3. Transient siRNA transfections were carried outusing RNAifect as described by the manufacturer’s instructions (QIAGEN).Five-day human MDMs were washed and incubated in 100 L siRNA complexfor 48 hours. Cells were then washed, fresh media added, and cells incubated innormoxia or hypoxia for 18 hours as described earlier.
RNA and protein extraction from human MDMs
Total RNA was prepared using RNeasy kit (QIAGEN) according to themanufacturer’s instructions and stored at �80°C. For protein extraction,cells were lysed with lysis buffer (50 mM Tris-HCl, pH 8.0, 150 mM NaCl,1% Triton X-100, and 1 protease inhibitor tablet, Roche). Protein levelswere measured using the bicinchoninic acid (BCA) protein assay(Sigma-Aldrich).
RNA and protein extraction from murine BMDMs
Total RNA and protein isolation was prepared using NucleoSpin RNA/Protein kit (Macherey-Nagel) and stored at �80°C for RNA and �20°C forprotein. For HIF-2�–deficient BMDMs, whole cell extracts were preparedusing radioimmunoprecipitation assay (RIPA) buffer (50 mM Tris, pH 8.0,150 mM NaCl, 1% NP40, 0.1% sodium dodecyl sulfate, 0.25% deoxy-cholate, 1 mM ethylenediaminetetraacetic acid) containing phosphotaseinhibitors (0.1 mM sodium fluoride, 1 mM sodium orthovanadate, 2 mMsodium pyrophosphate, and 10 mM �-glycerophosphate). Again, proteinextracts were stored at �20°C until used for immunoblotting.
Transcriptional profile analysis
Human Genome U133A plus 2.0 gene chip arrays (Affymetrix UK) thatdetect 47 000 transcripts were used. Total RNA was reverse-transcribed togenerate cDNA libraries using oligo dT and superscript II (Invitrogen).cDNA was amplified using MEGscript T7 kit and cleaned using GeneChipCleanup (both Affymetrix). Labeled cRNA was synthesized using Gene-Chip IVT kit and then hybridized to the arrays after the manufacturer’sinstructions (Affymetrix). Gene chips were processed using an AffymetrixGeneChip scanner 3000.
To verify the results obtained using Affymetrix arrays, total RNA wasextracted from 2 separate experiments, reverse-transcribed, amplified, andhybridized to Sentrix HumanRef-8_V2 Bead Chip from Illumina accordingto the manufacturer’s protocols. After washing and drying, the Beadarraywas scanned using an Illumina Bead Station 500X, which uses SentrixScanApplication, Version 2.7.2 software. Illumina BeadStudio software wasused for quality control assessment and normalization of data using theLOESS normalization method from BioConductor R packages.
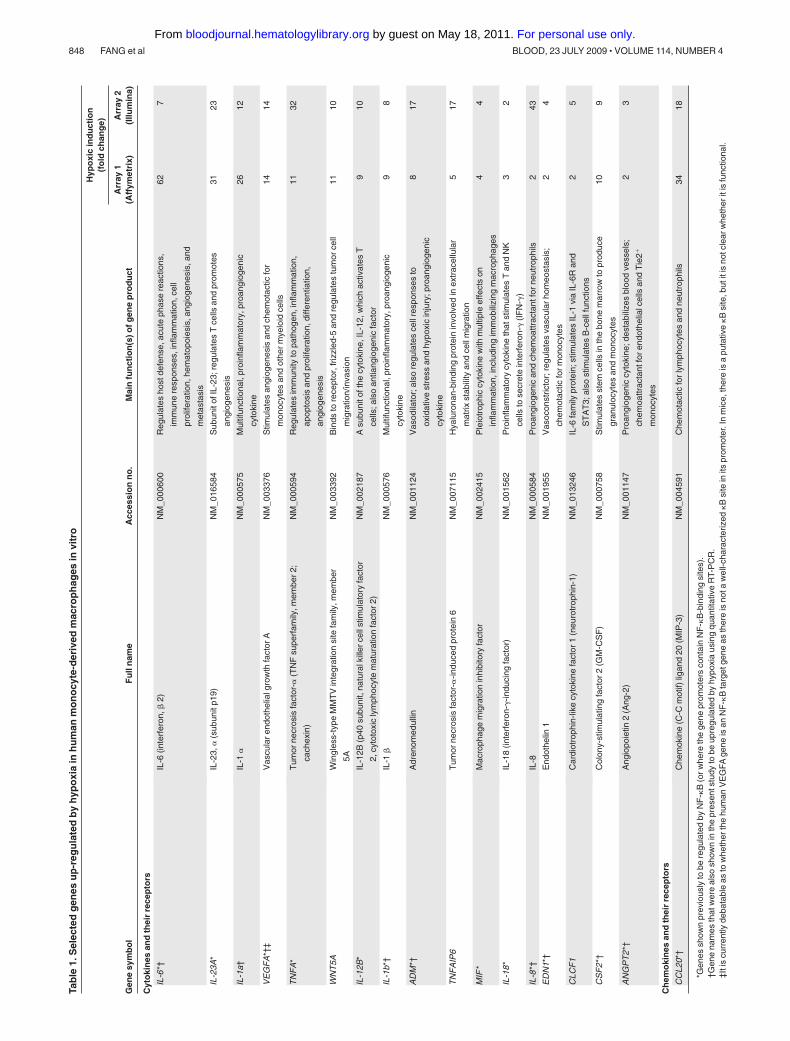
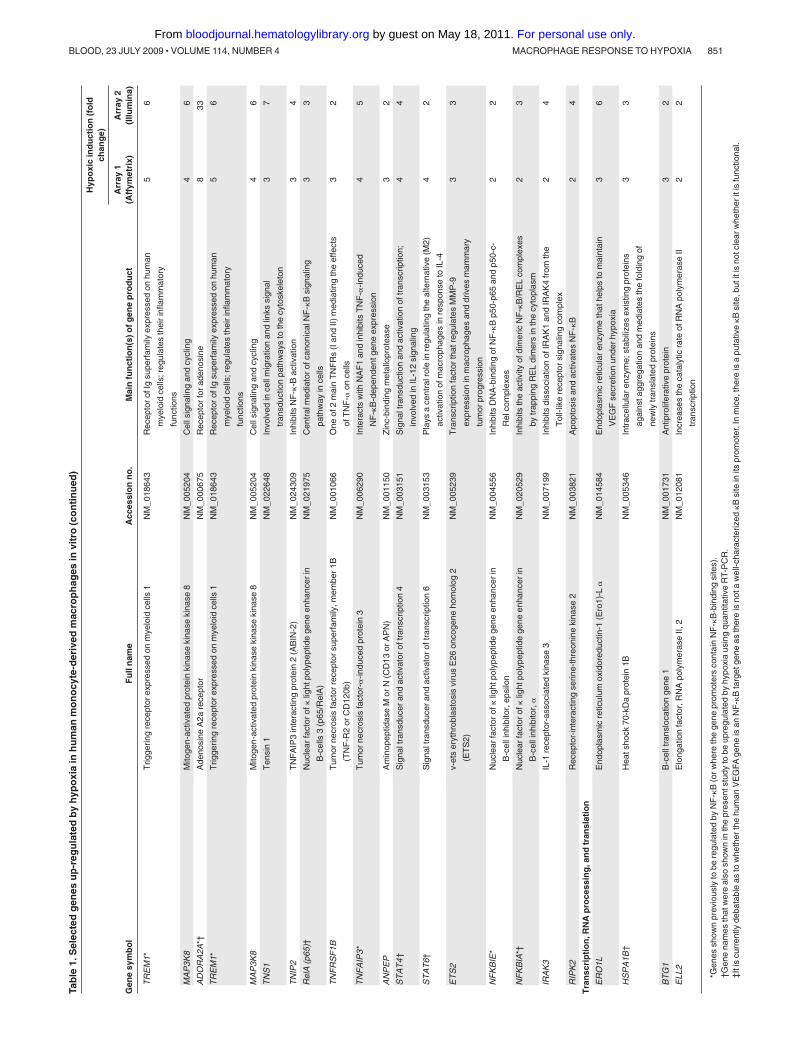
Genes that were up-regulated in both arrays by more than 1.5-fold ordown-regulated by less than 0.67-fold in hypoxia relative to normoxia wereconsidered differentially expressed. One Affymetrix and an Illuminamicroarray were conducted on RNA isolated from separate experiments.Their combined use was considered to be the first level of screening for themost robust hypoxia robust genes in human macrophages. Only mRNAspecies regulated by hypoxia on all arrays were considered to be reproduc-ibly regulated by hypoxia and worthy of further study. Using this criterion,148 genes were up-regulated and 60 genes down-regulated by hypoxia.A panel of selected genes was then further analyzed using real-timepolymerase chain reaction (PCR).
Real-time PCR
cDNAs was prepared from 1 g total RNA using SuperScript Synthesis kit(Invitrogen) and amplified with TaqMan gene expression Master Mix andpredesigned gene probes using a ABI 7900HT Sequence DetectionSystem (Applied Biosystems). The human TaqMan gene expression assayprobes used were VEGF, interleukin-1� (IL-1�), IL-6, CXCL8, CXCR4(chemokine C-X-C receptor 4), adrenomedullin (ADM), STAT4, adenosinereceptor 2A (ADORA2A), intercellular adhesion molecule 1 (ICAM1),heme oxygenase 1 (HMOX1), prolyl hydroxylase 2 (PHD2), CITED2, heatshock 70-kDa protein 1B (HSPA1B), ADAM metallopeptidase domain 8(ADAM8), ERO1-like (ERO1L), matrix metalloproteinase 7 (MMP7),glucose transporter 1 (GLUT-1), and �-2-microglobulin as the endogenouscontrol (Applied Biosystems). The murine TaqMan probes used for murinehomologs of these were also supplied by Applied Biosystems. Real-timePCR cycling conditions for both human and murine samples were2 minutes at 50°C and then 95°C for 10 minutes followed by 40 cycles of15 seconds at 95°C followed by 1 minute at 60°C. In addition, the humanNF-�B signaling genes were analyzed using SyBr green real-time PCR. Theprimer sequences used were as follows: NFKBIA, forward, TCGCAGTG-GACCTGCAAAAT; reverse, TGAGCTGGTAGGGAGAATAGC; IKK-�,forward, CACCATCCACACCTACCCTG; reverse, CTTATCGGGGAT-CAACGCCAG; IKK-, forward, CGTACTGGGCGAAGAGTCTC; re-verse, GGCTGGCTTGGAAATGCAG; NFKB1 (p50), forward, TGCCAA-CAGATGGCCCATAC; reverse, TGTTCTTTTCACTAGAGGCACCA; andRel A, forward, TTGAGGTGTATTTCACGGGACC; reverse, GCACAT-CAGCTTGCGAAAAGG. Real-time PCR was done using SyBr GreenPCR Master Mix, detected by ABI-Prism 5700 Sequence Detector, and dataprocessed using GeneAmp software (Applied Biosystems) The murine
MACROPHAGE RESPONSE TO HYPOXIA 845BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

TaqMan probes used for murine homologs of Rel A and IKK-� were alsosupplied by Applied Biosystems. The threshold cycle (Ct) of all human andmurine data was normalized against their respective endogenous controls(unaltered by hypoxia). Real-time PCR was analyzed in RNA extractsgenerated in 3 to 5 independent experiments and then fold changes inexpression relative to normoxic cells calculated with �Ct values of thesample and reference gene using the formula 2���Ct.
Immunoblotting studies
Immunoblotting for human HIFs-1� and 2� was conducted as describedpreviously10,11 using 1:1000 antihuman HIF-1� monoclonal antibodysupplied by BD Biosciences or 1:1000 antihuman HIF-2� monoclonalantibody from Novus. Both blots were incubated with horseradish peroxidase-conjugated antimouse antibody (Dako Denmark) and protein bands visualizedusing an enhanced chemilluminescence detection system (GE Healthcare). In allcases, expression of �-actin was used as a loading control. For NF-�Bimmunoblotting assays, an antihuman phospho-NF-�B p65, total NF-�B p65,phospho-IKK-�/IKK-�, or total IKK-�/� (Cell Signaling Technology) was usedat a dilution of 1:500 or 1:1000 and incubated overnight at 4°C.
Cytokine release assay
Cell supernatants were centrifuged for 5 minutes at 400g and filtered toeliminate cell debris and then stored at �20°C. The levels of VEGF, IL-8,and IL-1� in these supernatants were measured using a BD FACSArraybioanalyzer (BD Biosciences).
Role of NF-�B in hypoxic gene regulation in primarymacrophages
This was investigated in 2 ways. First, human MDMs were exposed to aspecific NF-�B inhibitor, 4-methyl-N1-(3-phenyl-propyl)-benzene-1,2-diamine (JSH-23; Merck Chemicals), which blocks translocation of phopho-rylated NF-�B (p65) to the nucleus of cells and its subsequent activation ofNF-�B gene targets.24 MDMs were exposed to medium alone or mediumcontaining 40 M JSH-23 (or the equivalent amount of the vehicle forJSH-23, dimethyl sulfoxide [DMSO]) for 1.5 hours, washed, and incubatedin normoxia or hypoxia for 18 hours. Normoxic MDMs were also exposedto 10 ng/mL recombinant human tumor necrosis factor-� (TNF-�; Pepro-Tech) for 18 hours as a positive control for NF-�B activation. RNA andnuclear proteins were then extracted from parallel cultures of MDMs afterthese treatments for real-time reverse-transcribed (RT) PCR and immuno-blot analysis, respectively. Some cells were also fixed in 3% formaldehydein phosphate-buffered saline for 15 minutes, washed and permeabilizedwith ice-cold 100% methanol for 10 minutes, and blocked with 5% goatserum in 0.3% Triton X-100/phosphate-buffered saline solution for 1 hour.NF-�B p65 was detected using a rabbit anti–mouse antibody (1:25, CellSignaling Technology) followed by addition of goat anti–rabbit Alexa-488secondary antibody (Invitrogen; 1:250 dilution). Cells were counterstainedwith 300 nM 4,6-diamidino-2-phenylindole (DAPI; Invitrogen) and thenphotographed on a confocal fluorescent microscope (original magnifica-tion �400). Twelve areas of cells were photographed for each treatmentgroup and the degree of nuclear p65 immunofluorescence (ie, Alexa-488labelling) in each DAPI-stained nuclei quantified using Analysis D software(Olympus). The proportion of green fluorescence per nuclei was thencalculated for all nuclei in 5 fields of view/treatment. The number of allMDMs in each field of view containing Alexa-488–labeled (p65�) nucleiwas also counted. To confirm JSH-23 inhibition of NF-�B activity inhypoxic MDMs, electrophoretic mobility shift assays for NF-�B binding toan NF-�B DNA consensus site were conducted as described previously byus25 on lysates from MDMs exposed to nomoxia, hypoxia, or hypoxia plusJSH-23 (all in the presence of DMSO as the vehicle for JSH-23). Proteinextracts from parallel cultures of MDMs were also immunoblotted forHIFs-1 and -2� (as described above in “Immunoblotting studies”).
The second approach was to infect MDMs with an adenovirusexpressing a dominant negative inhibitor of IKK-� to block phosphorylation/activation of p65/RelA. After 4 days in culture, MDMs were exposed to50 ng/mL recombinant human M-CSF for 24 hours to stimulate up-
regulation of integrin �V�5 (required for adenovirus infection of macro-phages26). The adv-IKK-�DN and control adv (Adv-GFP; a gift from DrThorsten Hagemann, Barts and The London Cancer Centre, London, UnitedKingdom) were E1/E3-deleted, of the Ad5 serotype, and used to transfectMDMs as described previously.27 MDMs were infected for 2 hours with100 multiplicity of infection of either adenovirus in serum-free medium.The adenovirus was then removed and fresh medium containing 2%antibody serum added. MDMs were maintained for a further 2 days inculture and then exposed to hypoxia or normoxia for 18 hours. Thisinfection protocol markedly reduces the activity of p65/RelA in humanMDMs27 and human endothelial cells.28
Immunofluorescent labeling of IL-1� expressed by TAMs inhypoxic areas of murine 4T1 mammary tumors
Frozen sections of 4T1 murine mammary tumors were generated in aprevious study.29 These had been grown in the mammary fat pads of femaleBALB/c mice and removed and snap frozen 2 hours after injection of micewith the hypoxic cell marker, pimonidazole.29 Sections (7 M) wereblocked with FcR Blocking Reagent (Miltenyi Biotec) in Tris-bufferedsaline-0.05% Tween 20 for 30 minutes at room temperature and thenincubated with rat anti–F4/80-Alexa 488 (1 g/mL, clone CI:A3-1; AbDSerotec), goat anti–mouse IL-1� (15 g/mL; R&D Systems), and rabbitanti-PIMO (1:4000, a gift from James Raleigh) for 30 minutes at roomtemperature. Negative controls included substitution of primary antibodieswith species-matched, nonspecific antibodies. Sections were then washedtwice and incubated in donkey anti–goat-Alexa 568 (8 g/mL; Invitrogen)or Alexa 647–conjugated goat anti–rabbit (8 g/mL; Invitrogen) secondaryantibodies for 30 minutes at room temperature in the dark and 30 nM DAPI(Invitrogen) for 2 minutes.
Gene set enrichment analysis
Gene set enrichment analysis (GSEA) was performed as described previ-ously30 on gene lists ranked by level of hypoxic gene induction (hypoxia/normoxia fold induction) separately for both the Affymetrix and Illuminagene expression datasets. Correlations to the predefined curated andtranscription factor target gene set collections were analyzed with theGSEA preranked tool using 1000 permutations. Further informationregarding the gene sets used in these analyses is available in the MolecularSignatures Database (MSigDB; www.broad.mit.edu/gsea/msigdb).
Statistics
All experiments were repeated 3 to 6 times. Statistical analyses wereperformed using the 1- or 2-tailed Student t test to determine statisticalsignificance after checking the data for normality (as appropriate). P valuesless than .05 were considered statistically significant. All data are expressedas mean plus or minus SEM.
Results
Evidence of distinct transcriptional signaling in primary humanmacrophages experiencing hypoxia
Hypoxic MDMs up-regulated both HIF-1� and HIF-2�, and thiswas markedly inhibited by prior treatment with siRNA to eitherHIF� (Figure 1A). As in previous publications,31,32 genes weredefined as being differentially regulated in hypoxia if they exhib-ited more than 1.5-fold increase in gene expression (Table 1) ordown-regulated if they showed less than 0.67-fold change (Table 2)compared with normoxic cultures. A comparison of our humanMDM microarray results (Tables 1-2) with those obtained previ-ously for related human myeloid cell types exposed to hypoxia(monocytes and monocyte-derived dendritic cells31,32) shows thatsome genes were seen to be regulated by all 3 cell types(up-regulated: VEGFA, CXCR4, TNF-�, TIMP1, PHD3, aldolases
846 FANG et al BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

A and C, enolase 2, TREM1, NCF1; down-regulated: cathepsin C).However, some genes regulated by hypoxia in MDMs are notsimilarly regulated by hypoxia in these other 2 cell types, such asIL-1�, IL-12p40, Ang-2, endothelin 1, STATS 4 and 6, CCLs 3 and5, CCR7, HMOX1, and hsp70 (up-regulated) and CD36, PECAM1(CD31), HIF-2�, and MHCII DM�; down-regulated; Tables 1-2).
The 2 full sets of array data (Affymetrix and Illumina) havebeen deposited in NCBI’s Gene Expression Omnibus and areaccessible through GEO Series accession numbers GSE15949 andGSE16099, respectively.33,34
Several key genes were selected and their up-regulation confirmedusing quantitative RT-PCR (Table 1). Macrophages were also shown toexpress abundant IL-1� protein in pimonidazole-stained (hypoxic) areasof murine 4T1 mammary tumors (Figure 2C).
Genetic manipulation of HIFs-1 and -2� demonstrates thecoregulation of genes in primary human macrophagesexperiencing hypoxia
The hypoxic accumulation of both HIFs-1 and -2� was ablatedafter transfection with siRNA for either � subunits. Both VEGFAmRNA and protein were markedly increased by hypoxia, and thiswas significantly inhibited by siRNA for either HIF� subunit
(Figure 1B left and middle panels). It may appear that the hypoxicinduction of VEGFA mRNA is higher in hypoxic macrophagestreated with the scrambled control siRNA than in the “no siRNA”group. However, this failed to reach statistical significance. Thiswas also the case for these 2 groups in Figure 3E,G,I, and K.
CXCL8 mRNA and protein release were also up-regulated inhypoxic MDMS (Figure 1C); and although both HIF� siRNAtreatments reduced hypoxia-induced CXCL8 mRNA, only theeffect of HIF-2� siRNA reached significance. However, both HIF�siRNA species significantly reduced CXCL8 protein release (Fig-ure 1C). The inhibitory effect of HIF siRNA on the hypoxicinduction of both VEGFA and CXCL8 appeared to be slightlygreater at the protein than the mRNA level.
Hypoxia also up-regulated IL-1� mRNA and protein, and thiswas significantly inhibited by exposure to siRNA for either HIF �subunit (Figure 2A). We then investigated the role of HIFs-1 and -2in the hypoxic regulation of several other genes listed in Table 1.The hypoxic up-regulation of mRNA for CXCR4, GLUT1, ad-renomedulin (ADM), and STAT-4 was significantly (P � .05)reduced by HIF-1� or 2� siRNA (Figure 3A,C,E,G). In contrast tothe other genes investigated, the hypoxic induction of adenosineA2a receptor (ADORA2A) and ICAM1 mRNA was significantly(P � .05) inhibited only by HIF-2� siRNA (Figure 3I,K).
Figure 1. Role of HIF-1� and -2� in the hypoxicinduction of VEGFA and CXCL8: insights from siRNAknockdown studies and use of macrophages bearinga deletion in the HIF-1� gene. (A) Immunoblots ofHIF-1� or HIF-2 � in MDM lysates after their exposure tonormoxia (20.9% O2; N) or hypoxia (0.1% O2; H) for18 hours, or hypoxia for 18 hours after exposure tosiRNA for HIF-1� (1�), HIF-2 (2�), both HIFs-1� and 2�together (1� � 2�), or a scrambled control (Scr). Load-ing controls were �-actin. Vertical lines have been in-serted to indicate repositioned lanes from the same gel.Below each gel picture is the densitometric analysis ofHIF expression relative to its �-actin loading control.(B-C) Effects of HIF-1� and -2� knockdown on thehypoxic induction of VEGFA (B) and CXCL8 (IL-8; C)mRNA and protein. In the case of VEGF, gene expres-sion was also assayed in normoxic and hypoxic BMDMsfrom mice bearing a myeloid cell-specific knockout of theHIF-1 � gene (HIF-1��/�) in vitro by quantitative RT-PCR(B right panel). It was not possible to do this for CXCL8,as this gene is not expressed in mice. Pooled datafrom 6 replicate experiments are shown. *P � .05 com-pared with corresponding normoxic group. ^P � .05compared with the scr siRNA/hypoxia group. �P � .05compared with macrophages from wild-type mice ex-posed to hypoxia.
MACROPHAGE RESPONSE TO HYPOXIA 847BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom
Tab
le1.
Sel
ecte
dg
enes
up
-reg
ula
ted
by
hyp
oxi
ain
hu
man
mo
no
cyte
-der
ived
mac
rop
hag
esin
vitr
o
Gen
esy
mb
ol
Fu
lln
ame
Acc
essi
on
no
.M
ain
fun
ctio
n(s
)ofg
ene
pro
du
ct
Hyp
oxi
cin
du
ctio
n(f
old
chan
ge)
Arr
ay1
(Aff
ymet
rix)
Arr
ay2
(Illu
min
a)
Cyt
oki
nes
and
thei
rre
cep
tors
IL-6
*†IL
-6(in
terf
eron
,�2)
NM
_000
600
Reg
ulat
esho
stde
fens
e,ac
ute
phas
ere
actio
ns,
imm
une
resp
onse
s,in
flam
mat
ion,
cell
prol
ifera
tion,
hem
atop
oies
is,a
ngio
gene
sis,
and
met
asta
sis
627
IL-2
3A*
IL-2
3,�
(sub
unit
p19)
NM
_016
584
Sub
unit
ofIL
-23;
regu
late
sT
cells
and
prom
otes
angi
ogen
esis
3123
IL-1
a†IL
-1�
NM
_000
575
Mul
tifun
ctio
nal,
proi
nflam
mat
ory,
proa
ngio
geni
c
cyto
kine
2612
VE
GF
A*†
‡V
ascu
lar
endo
thel
ialg
row
thfa
ctor
AN
M_0
0337
6S
timul
ates
angi
ogen
esis
and
chem
otac
ticfo
r
mon
ocyt
esan
dot
her
mye
loid
cells
1414
TN
FA
*T
umor
necr
osis
fact
or-�
(TN
Fsu
perf
amily
,mem
ber
2;
cach
exin
)
NM
_000
594
Reg
ulat
esim
mun
ityto
path
ogen
,infl
amm
atio
n,
apop
tosi
san
dpr
olife
ratio
n,di
ffere
ntia
tion,
angi
ogen
esis
1132
WN
T5A
Win
gles
s-ty
peM
MT
Vin
tegr
atio
nsi
tefa
mily
,mem
ber
5A
NM
_003
392
Bin
dsto
rece
ptor
,friz
zled
-5an
dre
gula
tes
tum
orce
ll
mig
ratio
n/in
vasi
on
1110
IL-1
2B*
IL-1
2B(p
40su
buni
t,na
tura
lkill
erce
llst
imul
ator
yfa
ctor
2,cy
toto
xic
lym
phoc
yte
mat
urat
ion
fact
or2)
NM
_002
187
Asu
buni
toft
hecy
toki
ne,I
L-12
,whi
chac
tivat
esT
cells
;als
oan
tiang
ioge
nic
fact
or
910
IL-1
b*†
IL-1
�N
M_0
0057
6M
ultif
unct
iona
l,pr
oinfl
amm
ator
y,pr
oang
ioge
nic
cyto
kine
98
AD
M*†
Adr
enom
edul
linN
M_0
0112
4V
asod
ilato
r;al
sore
gula
tes
cell
resp
onse
sto
oxid
ativ
est
ress
and
hypo
xic
inju
ry;p
roan
giog
enic
cyto
kine
817
TN
FA
IP6
Tum
orne
cros
isfa
ctor
-�-in
duce
dpr
otei
n6
NM
_007
115
Hya
luro
nan-
bind
ing
prot
ein
invo
lved
inex
trac
ellu
lar
mat
rixst
abili
tyan
dce
llm
igra
tion
517
MIF
*M
acro
phag
em
igra
tion
inhi
bito
ryfa
ctor
NM
_002
415
Ple
iotr
ophi
ccy
toki
new
ithm
ultip
leef
fect
son
infla
mm
atio
n,in
clud
ing
imm
obili
zing
mac
roph
ages
44
IL-1
8*
IL-1
8(in
terf
eron
--in
duci
ngfa
ctor
)N
M_0
0156
2P
roin
flam
mat
ory
cyto
kine
that
stim
ulat
esT
and
NK
cells
tose
cret
ein
terf
eron
-(I
FN
-)
32
IL-8
*†IL
-8N
M_0
0058
4P
roan
giog
enic
and
chem
oattr
acta
ntfo
rne
utro
phils
243
ED
N1*
†E
ndot
helin
1N
M_0
0195
5V
asoc
onst
ricto
r;re
gula
tes
vasc
ular
hom
eost
asis
;
chem
otac
ticfo
rm
onoc
ytes
24
CLC
F1
Car
diot
roph
in-li
kecy
toki
nefa
ctor
1(n
euro
trop
hin-
1)N
M_0
1324
6IL
-6fa
mily
prot
ein;
stim
ulat
esIL
-1vi
aIL
-6R
and
ST
AT
3;al
sost
imul
ates
B-c
ellf
unct
ions
25
CS
F2
*†C
olon
y-st
imul
atin
gfa
ctor
2(G
M-C
SF
)N
M_0
0075
8S
timul
ates
stem
cells
inth
ebo
nem
arro
wto
prod
uce
gran
uloc
ytes
and
mon
ocyt
es
109
AN
GP
T2
*†A
ngio
poie
tin2
(Ang
-2)
NM
_001
147
Pro
angi
ogen
iccy
toki
ne;d
esta
biliz
esbl
ood
vess
els;
chem
oattr
acta
ntfo
ren
doth
elia
lcel
lsan
dT
ie2�
mon
ocyt
es
23
Ch
emo
kin
esan
dth
eir
rece
pto
rs
CC
L20*
†C
hem
okin
e(C
-Cm
otif)
ligan
d20
(MIP
-3)
NM
_004
591
Che
mot
actic
for
lym
phoc
ytes
and
neut
roph
ils34
18
*Gen
essh
own
prev
ious
lyto
bere
gula
ted
byN
F-�
B(o
rwhe
reth
ege
nepr
omot
ers
cont
ain
NF
-�B
-bin
ding
site
s).
†Gen
ena
mes
that
wer
eal
sosh
own
inth
epr
esen
tstu
dyto
beup
regu
late
dby
hypo
xia
usin
gqu
antit
ativ
eR
T-P
CR
.‡I
tis
curr
ently
deba
tabl
eas
tow
heth
erth
ehu
man
VE
GFA
gene
isan
NF
-�B
targ
etge
neas
ther
eis
nota
wel
l-cha
ract
eriz
ed�
Bsi
tein
itspr
omot
er.I
nm
ice,
ther
eis
apu
tativ
e�
Bsi
te,b
utit
isno
tcle
arw
heth
erit
isfu
nctio
nal.
848 FANG et al BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

Tab
le1.
Sel
ecte
dg
enes
up
-reg
ula
ted
by
hyp
oxi
ain
hu
man
mo
no
cyte
-der
ived
mac
rop
hag
esin
vitr
o(c
on
tin
ued
)
Gen
esy
mb
ol
Fu
lln
ame
Acc
essi
on
no
.M
ain
fun
ctio
n(s
)ofg
ene
pro
du
ct
Hyp
oxi
cin
du
ctio
n(f
old
chan
ge)
Arr
ay1
(Aff
ymet
rix)
Arr
ay2
(Illu
min
a)
CX
CL2
*C
hem
okin
e(C
-X-C
mot
if)2
(MIP
-2a)
NM
_002
089
Che
mot
actic
for
neut
roph
ilsan
dhe
mat
opoi
etic
stem
cells
234
CX
CL1
*C
hem
okin
e(C
-X-C
mot
if)1
(MS
GA
-�)
NM
_001
511
Che
mot
actic
for
neut
roph
ils20
27
CC
R7
*C
hem
okin
e(C
-Cm
otif)
rece
ptor
7N
M_0
0183
8R
ecep
tor
for
chem
okin
es,C
CL1
9an
dC
CL2
110
52
CC
L5*
Che
mok
ine
(C-C
mot
if)5
(RA
NT
ES
)N
M_0
0298
5C
hem
otac
ticfo
rT
cells
,eos
inop
hils
,and
baso
phils
;
recr
uits
leuk
ocyt
esto
infla
mm
ator
ysi
tes
910
CC
L3*
Che
mok
ine
(C-C
mot
if)3
(MIP
-1a)
NM
_002
983
Rec
ruitm
enta
ndac
tivat
ion
ofne
utro
phils
36
CX
CR
4*†
Che
mok
ine
(C-X
-Cm
otif)
rece
ptor
4N
M_0
0346
7R
ecep
tor
for
SD
F-1
(CX
CL1
2),w
hich
regu
late
s
hem
atop
oiet
icst
eman
dm
yelo
idce
llre
crui
tmen
t
bytis
sues
;inv
olve
din
prol
ifera
tion
and
met
asta
sis
oftu
mor
cells
;cor
ecep
tor
for
entr
yof
HIV
into
T
cells
26
Intr
acel
lula
ren
zym
esan
dm
etab
olis
m
EG
LN3
†H
IFpr
olyl
hydr
oxyl
ase
3(P
HD
3)N
M_0
2207
3O
neof
3P
HD
enzy
mes
that
hydr
oxyl
ates
HIF
s,
resu
lting
inth
eir
bind
ing
toV
HL
and
degr
adat
ion;
regu
late
sH
IF-2
�m
ore
than
HIF
-1�
5412
CA
12C
arbo
nic
anhy
dras
eX
IIN
M_0
0121
8E
nzym
eth
atca
taly
zes
the
reve
rsib
lehy
drat
ion
of
carb
ondi
oxid
e;ac
idifi
esex
trac
ellu
lar
mili
euof
tum
orce
lls,s
timul
atin
gth
eir
grow
th/in
vasi
on
3281
ALD
OC
Ald
olas
eC
,fru
ctos
e-bi
spho
spha
teN
M_0
0516
5G
lyco
lytic
enzy
me;
cata
lyze
sth
ebr
eakd
own
of
fruc
tose
1,6-
bisp
hosp
hate
2323
SLC
2A5
*S
olut
eca
rrie
rfa
mily
2(f
acili
tate
dgl
ucos
e/fr
ucto
se
tran
spor
ter)
,mem
ber
5(G
LUT
-5)
NM
_003
039
Tra
nspo
rts
fruc
tose
and
gluc
ose
into
the
cell
1211
NC
F1
Neu
trop
hilc
ytos
olic
fact
or1
NM
_000
265
Sup
erox
ide
prod
uctio
n8
6
SLC
2A1†
Sol
ute
carr
ier
fam
ily2
(fac
ilita
ted
gluc
ose
tran
spor
ter)
,
mem
ber
1(G
LUT
-1)
NM
_006
516
Tra
nspo
rts
gluc
ose
into
the
cell
723
HM
OX
1*†
Hem
eox
ygen
ase
(dec
yclin
g)1
NM
_002
133
Ess
entia
lenz
yme
inhe
me
cata
bolis
m;c
leav
es
hem
eto
form
biliv
erdi
n
74
SLC
2A6
Sol
ute
carr
ier
fam
ily2
(fac
ilita
ted
gluc
ose
tran
spor
ter)
,
mem
ber
6(G
LUT
-6)
NM
_017
585
Tra
nspo
rts
gluc
ose
into
the
cell
63
SLC
2A3
Sol
ute
carr
ier
fam
ily2
(fac
ilita
ted
gluc
ose
tran
spor
ter)
,
mem
ber
3(G
LUT
-3)
NM
_006
931
Tra
nspo
rts
gluc
ose
into
the
cell
514
PF
KF
B3
6-ph
osph
ofru
cto-
2-ki
nase
/fruc
tose
-2,6
-bip
hosp
hata
se3
NM
_004
566
Syn
thes
isan
dde
grad
atio
nof
fruc
tose
2,6-
bisp
hosp
hate
47
SLC
7A5
Sol
ute
carr
ier
fam
ily7
(cat
ioni
cam
ino
acid
tran
spor
ter,
y�sy
stem
),m
embe
r5
NM
_003
486
Invo
lved
ince
llula
ram
ino
acid
upta
ke3
3
PF
KP
Pho
spho
-fru
ctok
inas
eN
M_0
0262
7G
lyco
lytic
enzy
me
23
HK
1H
exok
inas
e1
NM
_000
188
Com
mits
gluc
ose
toth
egl
ycol
ytic
path
way
22
ALD
OA
Ald
olas
eA
NM
_000
034
Gly
coly
ticen
zym
e;ca
taly
zes
the
brea
kdow
nof
fruc
tose
1,6-
bisp
hosp
hate
22
*Gen
essh
own
prev
ious
lyto
bere
gula
ted
byN
F-�
B(o
rwhe
reth
ege
nepr
omot
ers
cont
ain
NF
-�B
-bin
ding
site
s).
†Gen
ena
mes
that
wer
eal
sosh
own
inth
epr
esen
tstu
dyto
beup
regu
late
dby
hypo
xia
usin
gqu
antit
ativ
eR
T-P
CR
.‡I
tis
curr
ently
deba
tabl
eas
tow
heth
erth
ehu
man
VE
GFA
gene
isan
NF
-�B
targ
etge
neas
ther
eis
nota
wel
l-cha
ract
eriz
ed�
Bsi
tein
itspr
omot
er.I
nm
ice,
ther
eis
apu
tativ
e�
Bsi
te,b
utit
isno
tcle
arw
heth
erit
isfu
nctio
nal.
MACROPHAGE RESPONSE TO HYPOXIA 849BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

Tab
le1.
Sel
ecte
dg
enes
up
-reg
ula
ted
by
hyp
oxi
ain
hu
man
mo
no
cyte
-der
ived
mac
rop
hag
esin
vitr
o(c
on
tin
ued
)
Gen
esy
mb
ol
Fu
lln
ame
Acc
essi
on
no
.M
ain
fun
ctio
n(s
)ofg
ene
pro
du
ct
Hyp
oxi
cin
du
ctio
n(f
old
chan
ge)
Arr
ay1
(Aff
ymet
rix)
Arr
ay2
(Illu
min
a)
EG
LN1*
†H
IFpr
olyl
hydr
oxyl
ase
2(P
HD
2)N
M_0
2205
1O
neof
3P
HD
enzy
mes
that
hydr
oxyl
ates
HIF
s,
resu
lting
inth
eir
bind
ing
toV
HL
and
degr
adat
ion;
regu
late
sH
IF-1
�m
ore
than
HIF
-2�
25
Ext
race
llula
ren
zym
es/m
ole
cule
s
SE
RP
INE
1†S
erpi
npe
ptid
ase
inhi
bito
rN
M_0
0060
2R
egul
atio
nof
fibrin
olys
is7
5
AD
AM
8†
AD
AM
met
allo
pept
idas
edo
mai
n8
NM
_001
109
Mem
bran
e-an
chor
edpr
otei
nin
volv
edce
ll-ce
llan
d
cell-
mat
rixin
tera
ctio
ns
65
CF
BC
ompl
emen
tfac
tor
BN
M_0
0171
0A
com
pone
ntof
the
alte
rnat
ive
path
way
of
com
plem
enta
ctiv
atio
n
65
F3
*†C
oagu
latio
nfa
ctor
III(t
hrom
bopl
astin
,tis
sue
fact
or;
CD
142)
NM
_001
993
Cel
lsur
face
glyc
opro
tein
that
clea
ves
prot
hrom
bin
to
thro
mbi
n,pr
omot
ing
coag
ulat
ion
37
TIM
P1
TIM
Pm
etal
lope
ptid
ase
inhi
bito
r1
NM
_003
254
Inhi
bits
activ
ityof
mos
tkno
wn
MM
Ps;
stim
ulat
es
prol
ifera
tion
ina
wid
era
nge
ofce
llty
pes;
also
antia
popt
otic
23
MM
P7
†M
atrix
met
allo
pept
idas
e7
(mat
rilys
in,u
terin
e)N
M_0
0242
3E
nzym
ew
ithbr
oad
subs
trat
esp
ecifi
city
in
extr
acel
lula
rm
atrix
;pro
mot
esw
ound
heal
ing,
angi
ogen
esis
,tum
orin
vasi
on,a
ndm
etas
tasi
s
22
Cel
lvia
bili
ty
SE
RP
INB
2*
Ser
pin
pept
idas
ein
hibi
tor,
clad
eB
(ova
lbum
in),
mem
ber
2(P
AI-
2)
NM
_002
575
Inhi
bits
serin
epr
otea
se,t
issu
e-ty
pe-
and
urok
inas
e-
type
plas
min
ogen
activ
ator
;tP
A,u
PA
;als
o
regu
late
sge
neex
pres
sion
,cel
lpro
lifer
atio
n,
diffe
rent
iatio
n,an
dap
opto
sis
102
4
PT
GS
2(C
OX
2)*†
Pro
stag
land
in-e
ndop
erox
ide
synt
hase
2
(cyc
lo-o
xyge
nase
-2)
NM
_000
963
Pro
infla
mm
ator
y;st
imul
ates
expr
essi
onof
pros
tano
ids
278
EN
O2
Eno
lase
2N
M_0
0197
5G
lyco
lytic
enzy
me
810
IGF
BP
6In
sulin
-like
grow
thfa
ctor
bind
ing
prot
ein
6N
M_0
0217
8B
inds
and
prol
ongs
the
half-
life
ofth
eIG
Fs
42
BN
IP3L
BC
L2/a
deno
viru
sE
1B19
-kD
ain
tera
ctin
gpr
otei
n3-
like
NM
_004
331
Pro
apop
totic
Bcl
fam
ilypr
otei
n3
5
IER
3*
Imm
edia
teea
rlyre
spon
se3
(IE
X-2
orD
IF-2
)N
M_0
0389
7R
egul
ates
grow
than
dap
opto
sis
(inhi
bits
NF
-�B
-
indu
ced
apop
tosi
s)
213
NR
G1*
Neu
rore
gulin
1N
M_0
0609
6G
row
thfa
ctor
that
regu
late
sce
llap
opto
sis
and
prol
ifera
tion
(pro
tect
sce
llsin
isch
emia
)
24
Rec
epto
rsan
dce
llad
hes
ion
/sig
nal
ing
DD
IT4
DN
A-d
amag
e-in
duci
ble
tran
scrip
t4(D
IG2
orR
ED
D-1
)N
M_0
1905
8A
stre
ssre
spon
sege
ne,a
nes
sent
ialr
egul
ator
ofth
e
chec
kpoi
ntki
nase
,mT
OR
103
54
HIG
2H
ypox
ia-in
duci
ble
prot
ein
2N
M_0
0109
8786
Gro
wth
fact
orth
atst
imul
ates
tum
orce
llgr
owth
1221
NF
KB
2*
Nuc
lear
fact
orof
�lig
htpo
lype
ptid
ege
neen
hanc
erin
B-c
ells
2(p
49/p
100)
NM
_002
502
Cen
tral
med
iato
rof
nonc
anon
ical
NF
-�B
sign
alin
g
path
way
ince
lls
113
TR
AF
1*T
NF
rece
ptor
-ass
ocia
ted
fact
or1
NM
_005
658
One
ofth
eT
NF
Rm
embe
rs;f
orm
sa
com
plex
with
TR
AF
2to
activ
ate
MA
PK
8/JN
Kan
dN
F-�
B;
med
iate
sth
ean
tiapo
ptot
icsi
gnal
sfr
omT
NF
rece
ptor
s
98
AD
OR
A2A
*†A
deno
sine
A2a
rece
ptor
NM
_000
675
Rec
epto
rfo
rad
enos
ine
833
*Gen
essh
own
prev
ious
lyto
bere
gula
ted
byN
F-�
B(o
rwhe
reth
ege
nepr
omot
ers
cont
ain
NF
-�B
-bin
ding
site
s).
†Gen
ena
mes
that
wer
eal
sosh
own
inth
epr
esen
tstu
dyto
beup
regu
late
dby
hypo
xia
usin
gqu
antit
ativ
eR
T-P
CR
.‡I
tis
curr
ently
deba
tabl
eas
tow
heth
erth
ehu
man
VE
GFA
gene
isan
NF
-�B
targ
etge
neas
ther
eis
nota
wel
l-cha
ract
eriz
ed�
Bsi
tein
itspr
omot
er.I
nm
ice,
ther
eis
apu
tativ
e�
Bsi
te,b
utit
isno
tcle
arw
heth
erit
isfu
nctio
nal.
850 FANG et al BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom
Tab
le1.
Sel
ecte
dg
enes
up
-reg
ula
ted
by
hyp
oxi
ain
hu
man
mo
no
cyte
-der
ived
mac
rop
hag
esin
vitr
o(c
on
tin
ued
)
Gen
esy
mb
ol
Fu
lln
ame
Acc
essi
on
no
.M
ain
fun
ctio
n(s
)ofg
ene
pro
du
ct
Hyp
oxi
cin
du
ctio
n(f
old
chan
ge)
Arr
ay1
(Aff
ymet
rix)
Arr
ay2
(Illu
min
a)
TR
EM
1*T
rigge
ring
rece
ptor
expr
esse
don
mye
loid
cells
1N
M_0
1864
3R
ecep
tor
ofIg
supe
rfam
ilyex
pres
sed
onhu
man
mye
loid
cells
;reg
ulat
esth
eir
infla
mm
ator
y
func
tions
56
MA
P3K
8M
itoge
n-ac
tivat
edpr
otei
nki
nase
kina
seki
nase
8N
M_0
0520
4C
ells
igna
ling
and
cycl
ing
46
AD
OR
A2A
*†A
deno
sine
A2a
rece
ptor
NM
_000
675
Rec
epto
rfo
rad
enos
ine
833
TR
EM
1*T
rigge
ring
rece
ptor
expr
esse
don
mye
loid
cells
1N
M_0
1864
3R
ecep
tor
ofIg
supe
rfam
ilyex
pres
sed
onhu
man
mye
loid
cells
;reg
ulat
esth
eir
infla
mm
ator
y
func
tions
56
MA
P3K
8M
itoge
n-ac
tivat
edpr
otei
nki
nase
kina
seki
nase
8N
M_0
0520
4C
ells
igna
ling
and
cycl
ing
46
TN
S1
Ten
sin
1N
M_0
2264
8In
volv
edin
cell
mig
ratio
nan
dlin
kssi
gnal
tran
sduc
tion
path
way
sto
the
cyto
skel
eton
37
TN
IP2
TN
FA
IP3
inte
ract
ing
prot
ein
2(A
BIN
-2)
NM
_024
309
Inhi
bits
NF
-�-B
activ
atio
n3
4
Rel
A(p
65)†
Nuc
lear
fact
orof
�lig
htpo
lype
ptid
ege
neen
hanc
erin
B-c
ells
3(p
65/R
elA
)
NM
_021
975
Cen
tral
med
iato
rof
cano
nica
lNF
-�B
sign
alin
g
path
way
ince
lls
33
TN
FR
SF
1BT
umor
necr
osis
fact
orre
cept
orsu
perf
amily
,mem
ber
1B
(TN
F-R
2or
CD
120b
)
NM
_001
066
One
of2
mai
nT
NF
Rs
(Ian
dII)
med
iatin
gth
eef
fect
s
ofT
NF
-�on
cells
32
TN
FA
IP3
*T
umor
necr
osis
fact
or-�
-indu
ced
prot
ein
3N
M_0
0629
0In
tera
cts
with
NA
F1
and
inhi
bits
TN
F-�
-indu
ced
NF
-�B
-dep
ende
ntge
neex
pres
sion
45
AN
PE
PA
min
opep
tidas
eM
orN
(CD
13or
AP
N)
NM
_001
150
Zin
c-bi
ndin
gm
etal
lopr
otea
se3
2
ST
AT
4†
Sig
nalt
rans
duce
ran
dac
tivat
orof
tran
scrip
tion
4N
M_0
0315
1S
igna
ltra
nsdu
ctio
nan
dac
tivat
ion
oftr
ansc
riptio
n;
invo
lved
inIL
-12
sign
alin
g
44
ST
AT
6†S
igna
ltra
nsdu
cer
and
activ
ator
oftr
ansc
riptio
n6
NM
_003
153
Pla
ysa
cent
ralr
ole
inre
gula
ting
the
alte
rnat
ive
(M2)
activ
atio
nof
mac
roph
ages
inre
spon
seto
IL-4
42
ET
S2
v-et
ser
ythr
obla
stos
isvi
rus
E26
onco
gene
hom
olog
2
(ET
S2)
NM
_005
239
Tra
nscr
iptio
nfa
ctor
that
regu
late
sM
MP
-9
expr
essi
onin
mac
roph
ages
and
driv
esm
amm
ary
tum
orpr
ogre
ssio
n
33
NF
KB
IE*
Nuc
lear
fact
orof
�lig
htpo
lype
ptid
ege
neen
hanc
erin
B-c
elli
nhib
itor,
epsi
lon
NM
_004
556
Inhi
bits
DN
A-b
indi
ngof
NF
-�B
p50-
p65
and
p50-
c-
Rel
com
plex
es
22
NF
KB
IA*†
Nuc
lear
fact
orof
�lig
htpo
lype
ptid
ege
neen
hanc
erin
B-c
elli
nhib
itor,
�
NM
_020
529
Inhi
bits
the
activ
ityof
dim
eric
NF
-�B
/RE
Lco
mpl
exes
bytr
appi
ngR
EL
dim
ers
inth
ecy
topl
asm
23
IRA
K3
IL-1
rece
ptor
-ass
ocia
ted
kina
se3
NM
_007
199
Inhi
bits
diss
ocia
tion
ofIR
AK
1an
dIR
AK
4fr
omth
e
Tol
l-lik
ere
cept
orsi
gnal
ing
com
plex
24
RIP
K2
Rec
epto
r-in
tera
ctin
gse
rine-
thre
onin
eki
nase
2N
M_0
0382
1A
popt
osis
and
activ
ates
NF
-�B
24
Tra
nsc
rip
tio
n,R
NA
pro
cess
ing
,an
dtr
ansl
atio
n
ER
O1L
End
opla
smic
retic
ulum
oxid
ored
uctin
-1(E
ro1)
-L�
NM
_014
584
End
opla
smic
retic
ular
enzy
me
that
help
sto
mai
ntai
n
VE
GF
secr
etio
nun
der
hypo
xia
36
HS
PA
1B†
Hea
tsho
ck70
-kD
apr
otei
n1B
NM
_005
346
Intr
acel
lula
ren
zym
e;st
abili
zes
exis
ting
prot
eins
agai
nsta
ggre
gatio
nan
dm
edia
tes
the
fold
ing
of
new
lytr
ansl
ated
prot
eins
33
BT
G1
B-c
ellt
rans
loca
tion
gene
1N
M_0
0173
1A
ntip
rolif
erat
ive
prot
ein
32
ELL
2E
long
atio
nfa
ctor
,RN
Apo
lym
eras
eII,
2N
M_0
1208
1In
crea
ses
the
cata
lytic
rate
ofR
NA
poly
mer
ase
II
tran
scrip
tion
22
*Gen
essh
own
prev
ious
lyto
bere
gula
ted
byN
F-�
B(o
rwhe
reth
ege
nepr
omot
ers
cont
ain
NF
-�B
-bin
ding
site
s).
†Gen
ena
mes
that
wer
eal
sosh
own
inth
epr
esen
tstu
dyto
beup
regu
late
dby
hypo
xia
usin
gqu
antit
ativ
eR
T-P
CR
.‡I
tis
curr
ently
deba
tabl
eas
tow
heth
erth
ehu
man
VE
GFA
gene
isan
NF
-�B
targ
etge
neas
ther
eis
nota
wel
l-cha
ract
eriz
ed�
Bsi
tein
itspr
omot
er.I
nm
ice,
ther
eis
apu
tativ
e�
Bsi
te,b
utit
isno
tcle
arw
heth
erit
isfu
nctio
nal.
MACROPHAGE RESPONSE TO HYPOXIA 851BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

Tab
le2.
Sel
ecte
dg
enes
do
wn
-reg
ula
ted
by
hyp
oxi
ain
hu
man
mo
no
cyte
-der
ived
mac
rop
hag
esin
vitr
o
Gen
esy
mb
ol
Fu
lln
ame
Acc
essi
on
no
.M
ain
fun
ctio
n(s
)ofg
ene
pro
du
ct
Hyp
oxi
cd
ow
n-r
egu
lati
on
(fo
ldch
ang
e)
Arr
ay1
(Aff
ymet
rix)
Arr
ay2
(Illu
min
a)
Cel
lad
hes
ion
and
cell
jun
ctio
nm
ole
cule
s
CD
36C
D36
mol
ecul
e(t
hrom
bosp
ondi
nre
cept
or)
NM
_000
072
Am
ultif
unct
iona
lcla
ssB
scav
enge
rre
cept
or;
bind
sth
rom
bosp
ondi
n,ap
opto
ticce
lls,a
nd
LDLs
0.16
0.17
VC
LV
incu
linN
M_0
1400
0A
cyto
skel
etal
prot
ein
asso
ciat
edw
ithce
ll-ce
llan
d
cell-
mat
rixju
nctio
ns;i
nvol
ved
ince
llad
hesi
on,
cell
mor
phol
ogy,
and
loco
mot
ion
0.42
0.65
PE
CA
M1
Pla
tele
t/end
othe
lialc
ella
dhes
ion
mol
ecul
e(C
D31
antig
en)
NM
_000
442
Sur
face
rece
ptor
expr
esse
dby
endo
thel
ialc
ells
,
plat
elet
s,an
dva
rious
othe
rce
lls;h
elps
mac
roph
ages
tore
mov
eag
edne
utro
phils
0.62
0.52
Cel
lmet
abo
lism
AC
AT
1A
cety
l-coe
nzym
eA
acet
yltr
ansf
eras
e1
NM
_000
019
Pla
ysa
role
inlip
opro
tein
asse
mbl
yan
ddi
etar
y
chol
este
rola
bsor
ptio
n
0.25
0.38
NM
E1
Non
met
asta
ticce
lls1
prot
ein
(NM
23A
)N
M_0
0026
9A
nucl
eosi
dedi
phos
phat
eki
nase
linke
dto
met
asta
sis
supp
ress
ion
inso
me
cell
type
s
0.29
0.58
PD
HB
Pyr
uvat
ede
hydr
ogen
ase
(lipo
amid
e)�
NM
_000
925
Cat
alyz
esth
eov
eral
lcon
vers
ion
ofpy
ruva
teto
acet
yl-C
oAan
dC
O(2
)
0.37
0.49
ST
3GA
L5S
T3
�-g
alac
tosi
de�
-2,3
-sia
lyltr
ansf
eras
e5
NM
_003
896
Pro
mot
esce
lldi
ffere
ntia
tion,
mod
ulat
ion
ofce
ll
prol
ifera
tion,
sign
altr
ansd
uctio
n,an
din
tegr
in-
med
iate
dce
llad
hesi
on
0.40
0.54
LYP
LA3
Lyso
phos
phol
ipas
e3
(lyso
som
alph
osph
olip
ase
A2)
NM
_012
320
Reg
ulat
esth
em
ultif
unct
iona
llys
opho
spho
lipid
sin
cell
mem
bran
es
0.49
0.48
Intr
acel
lula
rtr
ansp
ort
TO
MM
22T
rans
loca
seof
oute
rm
itoch
ondr
ialm
embr
ane
22
hom
olog
(yea
st)
NM
_020
243
Mito
chon
dria
lmem
bran
epr
otei
n;im
port
s
cyto
solic
prep
rote
ins
into
the
mito
chon
drio
n
0.26
0.53
HLA
-DM
BM
ajor
hist
ocom
patib
ility
com
plex
,cla
ssII,
DM
�N
M_0
0211
8P
lays
ace
ntra
lrol
ein
the
pept
ide
load
ing
ofM
HC
clas
sII
mol
ecul
es
0.29
0.60
SLC
17A
5S
olut
eca
rrie
rfa
mily
17(a
nion
/sug
artr
ansp
orte
r),
mem
ber
5
NM
_012
434
Prim
ary
solu
tetr
ansl
ocat
orfo
ran
ioni
csu
bsta
nces
0.34
0.57
ST
6GA
L1*
ST
6�
-gal
acto
sam
ide
�-2
,6-s
ialy
l-tra
nsfe
rase
1N
M_0
0303
2T
rans
fers
sial
icac
idfr
omth
edo
nor
ofsu
bstr
ate
CM
P-s
ialic
acid
toga
lact
ose-
cont
aini
ng
acce
ptor
subs
trat
es
0.42
0.31
MR
PL3
Mito
chon
dria
lrib
osom
alpr
otei
nL3
NM
_007
208
Hel
psw
ithpr
otei
nsy
nthe
sis
with
inth
e
mito
chon
drio
n
0.47
0.50
Rec
epto
rsan
dce
llsi
gn
alin
g
TN
FS
F13
B*
Tum
orne
cros
isfa
ctor
(liga
nd)
supe
rfam
ily,m
embe
r
13b
NM
_006
573
Pro
mot
esce
llpr
olife
ratio
n0.
390.
39
RA
SG
RP
3R
AS
guan
yl-r
elea
sing
prot
ein
3(c
alci
uman
d
DA
G-r
egul
ated
)
NM
_015
376
Gua
nine
nucl
eotid
eex
chan
gefa
ctor
(GE
F)
for
Ras
and
Rap
1
0.13
0.38
TF
RC
Tra
nsfe
rrin
rece
ptor
(p90
,CD
71)
NM
_003
234
Reg
ulat
esce
llula
rup
take
ofiro
nan
diro
n
met
abol
ism
0.25
0.19
HIF
-1A
Hyp
oxia
-indu
cibl
efa
ctor
-�su
buni
t(H
IF-2
�)
NM
_001
530
Reg
ulat
esre
spon
seof
cells
tohy
poxi
a0.
360.
21
*Gen
essh
own
prev
ious
lyto
bere
gula
ted
byN
F-�
B(o
rwhe
reth
ege
nepr
omot
ers
cont
ain
NF
-�B
bind
ing
site
s).
852 FANG et al BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

Tab
le2.
Sel
ecte
dg
enes
do
wn
-reg
ula
ted
by
hyp
oxi
ain
hu
man
mo
no
cyte
-der
ived
mac
rop
hag
esin
vitr
o(c
on
tin
ued
)
Gen
esy
mb
ol
Fu
lln
ame
Acc
essi
on
no
.M
ain
fun
ctio
n(s
)ofg
ene
pro
du
ct
Hyp
oxi
cd
ow
n-r
egu
lati
on
(fo
ldch
ang
e)
Arr
ay1
(Aff
ymet
rix)
Arr
ay2
(Illu
min
a)
EP
AS
1E
ndot
helia
lPA
Sdo
mai
npr
otei
n1
(HIF
-2�
)N
M_0
0143
0R
egul
ates
resp
onse
ofce
llsto
hypo
xia
0.33
0.21
TLR
4T
oll-l
ike
rece
ptor
4N
M_0
0326
6C
ells
urfa
cere
cept
orth
atbi
nds
man
ylig
ands
,
incl
udin
gba
cter
ialL
PS
and
fibrin
ogen
0.26
0.63
MA
PR
E2
Mic
rotu
bule
-ass
ocia
ted
prot
ein,
RP
/EB
fam
ily,
mem
ber
2
NM
_014
268
Invo
lved
inm
icro
tubu
lepo
lym
eriz
atio
n,ce
ll
mig
ratio
n
0.35
0.52
CT
SC
Cat
heps
inC
NM
_001
814
Aly
soso
mal
cyst
eine
prot
eina
seth
atac
tivat
es
man
yse
rine
prot
eina
ses
in
imm
une/
infla
mm
ator
yce
lls
0.41
0.48
PR
CP
Pro
lylc
arbo
xype
ptid
ase
(ang
iote
nsin
ase
C)
NM
_005
040
Aly
soso
mal
prol
ylca
rbox
ypep
tidas
e,w
hich
clea
ves
C-t
erm
inal
amin
oac
ids
linke
dto
prol
ine
inpe
ptid
es
0.43
0.48
SP
AR
CS
ecre
ted
prot
ein,
acid
ic,c
yste
ine-
rich
(ost
eone
ctin
)N
M_0
0311
8A
calc
ium
bind
ing
glyc
opro
tein
that
also
bind
s
EC
Mco
nstit
uent
s,su
chas
colla
gen
0.46
0.26
PE
PD
Pep
tidas
eD
NM
_000
285
Col
lage
nm
etab
olis
m0.
600.
58
RT
N3
Ret
icul
on3
NM
_006
054
May
indu
ceca
spas
e-8
casc
ade
and
apop
tosi
s0.
610.
48
PS
EN
2P
rese
nilin
2(A
lzhe
imer
dise
ase
4)N
M_0
0044
7C
ytop
lasm
icpa
rtiti
onin
gof
prot
eins
0.61
0.57
Cel
lvia
bili
ty
CA
TC
atal
ase
NM
_001
752
Pro
tect
sce
llsfr
omth
eto
xic
effe
cts
ofhy
drog
en
pero
xide
0.48
0.47
PR
DX
6P
erox
iredo
xin
6N
M_0
0490
5In
volv
edin
redo
xre
gula
tion
ofth
ece
ll,pr
otec
tion
agai
nsto
xida
tive
inju
ry
0.61
0.57
Tra
nsc
rip
tio
n,R
NA
pro
cess
ing
and
tran
slat
ion
PO
LE3
Pol
ymer
ase
(DN
Adi
rect
ed),
�3
(p17
subu
nit)
NM
_017
443
Inte
ract
sw
ithot
her
hist
one-
fold
prot
eins
tobi
nd
DN
Ain
ase
quen
ce-in
depe
nden
tman
ner
0.47
0.66
LSM
7LS
M7
hom
olog
,U6
smal
lnuc
lear
RN
Aas
soci
ated
(S.c
erev
isia
e)
NM
_016
199
Pre
-mR
NA
splic
ing
0.48
0.53
SR
MS
perm
idin
esy
ntha
seN
M_0
0313
2In
volv
edin
num
erou
spr
e-m
RN
Apr
oces
sing
even
ts
0.48
0.28
TC
EA
L4T
rans
crip
tion
elon
gatio
nfa
ctor
A(S
II)-li
ke4
NM
_024
863
Anu
clea
rph
osph
opro
tein
that
may
mod
ulat
e
gene
tran
scrip
tion
0.62
0.54
*Gen
essh
own
prev
ious
lyto
bere
gula
ted
byN
F-�
B(o
rwhe
reth
ege
nepr
omot
ers
cont
ain
NF
-�B
bind
ing
site
s).
MACROPHAGE RESPONSE TO HYPOXIA 853BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

Transcriptional signaling in primary human MDMs experiencinghypoxia for 18 hours is independent of NF-�B
Gene set enrichment analysis. To assess the likelihood of NF-�Bplaying a role in hypoxic signal transduction in human macrophages, wefirst searched our data for correlations with several published gene setsrelating to hypoxia-regulated regulated genes in other cell types (eg, the“HYPOXIA_REVIEW” gene set35; Figure 4 top panels). This high-lighted a significant degree of enrichment of known hypoxia-regulatedgenes in our array data, showing that hypoxia-induced gene expressionchanges in MDMs follow a consensus hypoxia gene expression profile(Figure 4 top panels). This was evident for both the Affymetrix arraydata (Normalized Enrichment Score [NES]: 2.2; False Discovery Rate,q � 0.001) and the Illumina data (NES: 2.24; q � 0.001). Table 1 showsthat many genes up-regulated by hypoxic MDMs have previously beenidentified as NF-�B target genes. In the GSEA analysis, the hypoxicMDMS array data also correlated significantly with several NF-�B-related gene sets (eg, the V$NFKAPPAB_01 gene set36; Figure 4 bottompanels).Again, this was evident for both theAffymetrix data (NES: 1.69,q 0.02) and the Illumina data (NES: 1.67; q 0.12).
Hypoxic up-regulation of NF-�B signaling in human macro-phages: role of HIF-1 and -2. The effect of exposure of humanMDMs to hypoxia for 18 hours on NF-�B signaling was thenassessed. IKK-� and , I�B�, NF-�B1 (p50), and p65/RelAmRNA levels were up-regulated (and IKK-� mRNA slightlydown-regulated) in MDMs exposed to 0.1% O2 for 18 hours. Thishypoxic regulation (with the exception of IKKs � and ) wasinhibited using siRNA to knock down either HIF-1 or 2� (Figure5A). Figure 5C shows that, although there was a small hypoxicinduction of total IKK-� protein, the hypoxic up-regulation of
p65/RelA mRNA was not mirrored by a similar up-regulation oftotal p65/RelA protein, suggesting a differential effect of hypoxiaon mRNA vs protein expression for p65/RelA. By contrast, thephosphorylation of both IKK-�/� and p65/RelA was up-regulatedin hypoxic human MDMs (Figure 5C).
Role of NF-�B in the transcriptional responses of humanmacrophages to hypoxia. Figure 6A and B illustrates the effects ofthe NF-�B inhibitor JSH-23 on the hypoxic induction of various genesin human MDMs. This shows that immunoreactive p65 was cytoplas-mic in normoxic MDMs but transported to the nucleus on exposure toTNF-� or 18 hours of hypoxia. In both cases, this was significantly(P � .05) inhibited by prior exposure to JSH-23. EMSA assays con-firmed the induction of NF-�B DNAbinding in hypoxic MDMs and theinhibition of this by JSH-23 (Figure 6C). JSH-23–treated cells alsoexhibited slightly lower levels of HIFs-1 and -2� (particularly HIF-1�)than MDMs exposed to hypoxia alone (Figure 6C). We then investi-gated the effect of JSH-23 inhibition of NF-�B activity on the inductionof 8 hypoxia-regulated genes listed in Table 1. Exposure to TNF-� for18 hours significantly (P � .05) increased the expression of VEGFA,CXCL8, and STAT4, in a manner that was inhibited by JSH-23 (Figure6). Hypoxia significantly (P � .05) up-regulated all 8 genes studied(Figure 6D) in a manner that was not reduced by prior exposure ofcells to JSH-23.
Infection of MDMs with adv-IKK-�DN significantly (P � .05)inhibited their TNF-�–induced expression of CXCL8 mRNA (Figure7A) as well as the nuclear accumulation of phospho-p65/RelA byMDMS after 18 hours of exposure to hypoxia (Figure 7B). The controladv vector had no such effect. However, adv-IKK-�DN blockade ofhypoxia-induced phospho-p65/RelAfailed to reduce the hypoxic induc-tion of VEGFA, CXCL8, GLUT-1, CXCR4, or ADM mRNA.
Figure 2. Hypoxic up-regulation of IL-1� by humanMDMs in vitro and by TAMs in hypoxic areasof murine mammary tumors: role of HIF-1 and -2.(A) IL-1� mRNA levels and protein release by humanMDMs after their exposure to normoxia (20.9% O2; N) orhypoxia (0.1% O2; H) for 18 hours, or hypoxia for 18 hoursafter exposure to siRNA for HIF-1 � (1 �), HIF-2� (2 �),both HIFs-1� and -2� together (1� � 2�), or a scrambledcontrol (Scr). (B) Hypoxic induction of IL-1� mRNA byBMDMs from wild-type mice and mice bearing a myeloidcell-specific knockout of the HIF-1 � gene. (C) Up-regulated expression of IL-1� by F4/80� macrophages inpimonodazole-stained (ie, hypoxic; H) compared withpimonodazole-unstained (ie, normoxic; N) areas of mu-rine mammary (4T1) tumors (yellow arrows on the mergedH image). Pooled data from 3 replicate experimentsare shown. *P � .05 compared with corresponding nor-moxic group. ^P � .05 compared with the Scr siRNA/hypoxia group. �P � .05 compared with macrophagesfrom wild-type mice exposed to hypoxia.
854 FANG et al BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

Hypoxic regulation of genes in primary murine macrophages:role of HIFs-1 and -2
HIF-1–deficient BMDMs were only able to mount partial VEGFAand IL-1� responses to hypoxia (Figures 1B right panel, 2B). Thehypoxic up-regulation of CXCR4 and STAT4 was lost in BMDMslacking HIF-1� (Figure 3B,H). This contrasts with our aforemen-tioned human MDMs data showing that these were regulated byboth HIFs-1 and -2 (Figure 3A,G). The fact that GLUT1 and ADMwere reduced but not ablated in HIF-1� null BMDMs (Figure3D,F) agrees with our finding that these 2 genes are coregulated byHIFs-1 and -2 in human MDMs (Figure 3C,E). In addition, inagreement with the human MDM data (Figure 3I,K), the hypoxicup-regulation of neither the ADORA2A nor ICAM1 genes wasinhibited in hypoxic HIF-1� null BMDMs (Figure 3J,L).
Figure 5B shows that hypoxic up-regulation of IKK-� andp65/RelA mRNA levels was lost in HIF-1�–deficient BMDMs.Moreover, the hypoxic induction of phosphorylated p65/RelA was
lost in HIF-2�–deficient BMDMs (Figure 5D). Similar results wereseen for HIF-1�–deficient BMDMs (data not shown).
Discussion
Our data show that exposure to hypoxia activates a distinct transcrip-tional profile in primary human macrophages, including the up-regulation of VEGFA, ILs-1� and �, IL-8, STAT4, and ADM; thereceptors, glucose transporter, GLUT1, CXCR4, and the adenosinereceptor 2A (ADORA2A). Some were seen to also be regulated byhypoxia in monocytes and immature dendritic cells (VEGFA, GLUT1,and CXCR4).31,32 However, others, such as IL-1�, ADORA2A, andSTAT4, were only altered in hypoxic macrophages. These differencescould be the result of variations in the severity and/or duration ofhypoxia applied to cells31,32 and/or may reflect differences in thetranscription factors used by these 3 cell types in hypoxia. For example,
0
20
40
60
*
Normoxia Hypoxia
0
0.5
1
1.5
2
2.5
Fo
ld c
han
ge
A
Fo
ld c
han
ge
^+
GLUT-1
0
2
4
6
8
Fo
ld c
han
ge
B
0
2
4
6
8
10
E
Fo
ld c
ha
ng
e
ICAM1
CXCR4
Fo
ld c
han
ge
ADM
0
5
10
15
Fo
ld c
ha
ng
e
G
+
STAT4
0
1
2
3
4
5
Fo
ld c
ha
ng
e
WT HIF-1α -/-
0
2
4
6
8
10
^
*
*
*
*
0
0.5
1.0
1.5
2.0
2.5
Fo
ld c
ha
ng
e
*
*
*
*
^
**
^ ^ ^*
*
* * *
*^ ^* *
* * *
0
10
20
30
40
Fo
ld c
ha
ng
ed
*
*
^ ^
^*
+
ADORA2A
0.5
1
1.5
2
2.5
0
Fo
ld c
ha
ng
e
*
+
C D
F
H
J
L
I
0
10
20
30
40
Fo
ld c
han
ged
*
*
^
*
*
*
Fo
ld c
han
ge
K
- - Scr Scr 1α 2α 1α+2α0
2
4
6
*
*
* * *
8
^ ^
Figure 3. Role of HIFs-1� and -2� in the hypoxic induction ofother key genes by macrophages. (A,C,E,G,I,K) Hypoxic induc-tion of mRNA for CXCR4, GLUT-1, ADM, STAT4, ADORA2A, andICAM1 (as measured by quantitative RT-PCR) after exposure ofprimary human macrophages to normoxia (20.9% O2; N) orhypoxia (0.1% O2; H) for 18 hours, or hypoxia after exposure tosiRNA for HIF-1 � (1 �), HIF-2 � (2 �), both HIFs-1 � and -2 �together (1 � � 2 �), or a scrambled control (Scr). (B,D,F,H,J,L)Hypoxic induction of the same genes in wild-type or HIF-1�–deficient murine BMDMs. Pooled data from 3 replicate experi-ments are shown. *P � .05 compared with corresponding nor-moxic group. ^P � .05 compared with the Scr siRNA/hypoxiagroup. �P � .05 compared with macrophages from wild-typemice exposed to hypoxia.
MACROPHAGE RESPONSE TO HYPOXIA 855BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

hypoxic human monocytes exposed to a similar level and duration ofhypoxia as in the current study failed to up-regulate HIFs-1 and -2� butrather other transcription factors, such as ATF-4 and Egr-1.37 Moreover,the ability to regulate hypoxic gene expression via HIFs is maturation-linked in macrophages.38 Although dendritic cells accumulate HIF-1� inhypoxia,39 immature forms of this cell type up-regulate other hypoxia-responsive genes, such as CCL20 via up-regulated p50/p50 NF-�Bhomodimers rather than HIFs.40 A study of the responses of such related
myeloid cell types to identical hypoxic conditions would be interestingbut beyond the remit of this study.
As macrophages are known to express receptors for bothVEGF41 and IL-1,42 it is possible that that, during such exposure tohypoxia, their hypoxia-induced release might have then stimulatedthe expression of other genes in macrophages (making it look as ifthey are also directly up-regulated by hypoxia when, indeed, theeffect is indirect). However, hypoxic gene expression by human
Array 1 Array 2
Hyp
oxi
a-re
gu
late
d
gen
eset
Nf-
kBta
rget
g
enes
etFigure 4. Gene enrichment analysis to compare key genesup-regulated by hypoxia in human MDMs and known NF-�B–regulated genes. Hypoxia up-regulated genes identified in2 separate macrophage cultures using Affymetrix or Illuminamicroarrays were ranked by level of hypoxia-mediated induction.The ranked gene lists were then compared with both a previouslypublished gene set for hypoxia-regulated genes in tumor cells(the hypoxia-regulated gene set, top row) or genes previouslyshown to be NF-�B target genes (the NF-�B target gene set,bottom row) by GSEA. The hypoxia-regulated gene set (top row)was significantly enriched in the hypoxic macrophage gene setidentified on both the Affymetrix (Array 1; NES 2.2, q � 0.001)and the Illumina (Array 2; NES 2.24, q � 0.001) arrays. TheNF-�B target gene set (lower row) was also enriched in thehypoxic macrophage gene set on both Affymetrix (NES 1.69,q 0.02) and Illumina (NES 1.67, q 0.12) arrays.
Figure 5. Effect of hypoxia on the expression and/orphosphorylation of components of the canonicalNF-�B signaling pathway in MDMs: regulation byHIF-1� and -2�. (A) Fold induction (hypoxia, 0.1%O2/normoxia, 20.9% O2) of mRNA levels for individualNF-�B signaling proteins in primary human MDMs. Thecontribution of both HIFs-1 and -2 to the regulation ofmany of these genes was also assessed using siRNA toknock down the expression of each � subunit in MDMs.(B) Effect of normoxia (N) or hypoxia (0.1% O2; H) for18 hours on the expression of IKK-� and p65 mRNA bymurine BMDMs from wild-type or HIF-1–deficient mice.*P � .05 compared with corresponding normoxic group.�P � .05 compared with macrophages from wild-typemice exposed to hypoxia. (C) Immunoblots showing theeffect of exposure to normoxia or hypoxia (0.1% O2) for18 hours on the levels of total and phosphorylated IKK-�and p65/RelA in primary human MDMs. Vertical lineshave been inserted to indicate repositioned lanes fromthe same gel. (D) Effects of normoxic (N) or hypoxic(0.5% O2; H) culture on the level of total or phosphory-lated p65 protein in control (�/�) or HIF-2�–deficientmurine BMDMs. Similar results were obtained usingwild-type or HIF-1�–deficient murine BMDMs (data notshown).
856 FANG et al BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

MDMs is not reduced in the presence of either a neutralizing VEGFantibody or an IL-1 receptor antagonist (H.-Y.F., C.M., R.H., andC.E.L., unpublished observations, March 2009).
We also show, for the first time, that genes encoding the2 transcription factors, STATs (signal transducers and activators oftranscription) 4 and 6, are up-regulated by hypoxia in macro-phages. STATs 4 and 6 are known to mediate the marked effects of2 central immunomodulatory cytokines, IL-12 and IL-4, respec-tively.43,44 It remains to be seen whether their hypoxic inductioncould in this way “prime” macrophages to the effects of thesecytokines.
Our HIF siRNA studies showed that both HIFs play a part inregulating the hypoxic induction of the known HIF target genes,VEGFA, GLUT1, CXCR4, IL-8, and ADM by MDMs. Further-more, hypoxic induction of these genes was reduced but not lost inHIF-1�–deficient murine macrophages. Similar results were ob-tained for the hypoxic up-regulation of VEGFA and ADM inmurine HIF-2�–deficient BMDMs (H.Z.I. and M.C.S., manuscriptsubmitted, April 2009).
The pluripotent cytokine, IL-1�, stimulates many steps in tumorprogression45 and was up-regulated by hypoxic MDMs. We showthat TAMs express abundant IL-1� in hypoxic areas of murinemammary (4T1) tumors. The IL-1� gene promoter bears multiple
HREs and is transactivated by HIF-1.46,47 Our HIF siRNA knock-down studies show that HIFs-1 and -2 coregulate the hypoxicinduction of IL-1� in macrophages, a finding confirmed by thehypoxic up-regulation of IL-1� being only partially diminished inHIF-1�–deficient (Figure 1) and HIF-2�–deficient BMDMs (H.Z.I.and M.C.S., manuscript submitted April 2009).
It remains to be seen whether HIFs-1 and -2 bind to differentHREs on the promoters of the aforementioned coregulated genes orwhether other, unknown mechanisms underpin the phenomenon ofdual HIF responsiveness. Furthermore, as mentioned previously,this may vary between cell types as HIF-1 has been shown to be theprimary regulator of various genes in some cell types,4,16 whereasother cells use HIF-2 or both HIFs in their regulation.18,19,48,49
Interestingly, when just one HIF was inhibited using siRNA, theother did not appear to compensate for its loss and maintainmaximal hypoxic induction. It is known that many HIF-targetgenes have multiple HREs. If, once HIFs-1 and -2 have bound todifferent HREs in a given promoter they then cooperate, both mightbe required for maximal gene transcription. This may be similar tothe molecular “cooperation” that takes place between HIF-2 andElk-1 on some gene promoters.18,19
The knockdown of both HIFs-1� and 2� in MDMs failedto completely block the hypoxic up-regulation of most of the
05
101520
200
400
600
800
ADM
0
10
20
30
40
50
**
**
A
C
N
H
N+TNF+JSHN +TNF (+ DMSO)
H+JSHH +DMSO
i iii
iv v vi
B
VEGFA
0
20
40
60
80
*
IL-8
0
5
10
15
20100
200
300
400
*
* * **
IL1β
STAT4
ADORA2A
0
10
20
30
40
50
*
*
*
*
*
*
CXCR4
0
5
10
15
20
* *
**
GLUT-1
0
20
40
60
80
100
*
*
**
0
5
10
152030
45
60
75
* * ** *
*
*
*
*
*
*
**
JSH-23:- + +
TNF:
- + -
+
++
+- + - +
- - --
DMSO:
-
-
+
-
-
+
Normoxia Hypoxia
**
JSH-23:
- + +
TNF:
- + -
+
++
+- + - +
- - --
DMSO:
-
-
+
-
-
+
JSH-23:
- + +
TNF:
- + -
+
++
+- + - +
- - --
DMSO:
-
-
+
-
-
+
**
**
**
****
******
**
****
**
**
****
****
**
**
- + +
TNF:
- - -
JSH-23:
+++- + - +
- - --
DMSO:
Normoxia Hypoxia
0
3
9
12
5
% n
ucle
ar p
65
6*
+
*
^*
50 31 33 32 16
+
*
$
$
$
D
N H H+JSH
Bound NF-κB
HIF-1α
HIF-2α
β-actin
N H H+JSH
$$
$
Figure 6. Inhibition of nuclear translocation of p65has no effect on the hypoxic induction of variousgenes in human MDMs in vitro. (A-B) Effect of the p65inhibitor JSH-23 (or its vehicle, DMSO) on the nucleartranslocation of p65 induced by TNF-� or hypoxia byhuman MDMs. N indicates normoxia; H, hypoxia (0.1%O2); % nuclear p65 immunofluorescence, the percentageof the total, DAPI-stained (blue) area of MDM nuclei thatwas GFP� (green). The figures at the base or just aboveeach bar represent the average percentage of all MDMnuclei immunofluorescent for p65 (B). *P � .05 withrespect to normoxia alone group. �P � .05 with respectto TNF-� � DMSO group. ^P � .05 with respect tohypoxia � DMSO group. (C) Effect of JSH-23 on NF-�Bbinding and accumulation of HIFs-1 and -2� in hypoxicMDMs. N indicates normoxia; H, hypoxia, H � JSH-23,hypoxia after JSH.23 treatment. All 3 groups received thevehicle for JSH-23, DMSO. A vertical line has beeninserted to indicate repositioning of lanes from the samegel; (1) left panel: EMSA showing NF-�B binding to a DNAconsensus sequence, and (2) right panel: immunoblotsfor HIFs-1 and -2�. (D) Effect of JSH-23 blockade of p65function on the fold induction of VEGFA, CXCL8, IL-1�,CXCR4, GLUT-1, STAT4, ADM, and ADORA2A by TNF-�or hypoxia. *P � .05 with respect to normoxia with DMSOalone. **P � .05 with respect to group indicated. $P � .05with respect to TNF � DMSO group. Pooled data from3 replicate experiments are shown.
MACROPHAGE RESPONSE TO HYPOXIA 857BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

HIF-target genes shown in Figures 1 through 3. This suggests thatother transcription factors may also be involved in regulating theirhypoxic induction. The transcription factor, NF-�B, may be onesuch factor. This has been shown recently to be activated inmacrophages by short-term (2-4 hours) exposure to hypoxia, withthe expression and/or phosphorylation of IKK-�, IKB�, andp65/RelA, as well as the nuclear translocation and DNA-bindingactivity of p65 being up-regulated.20-22 There also appears to be aclose interplay between NF-�B and HIF-1 as p65/p50 heterodimersbind to the HIF-1� gene promoter and drive its expression underhypoxia. Interestingly, HIF-2� is not up-regulated by NF-�B inmurine macrophages during short-term hypoxia (4 hours).22 Thepresent report shows that p65 protein is phosphorylated and bindsDNA in the nuclei of MDMs in hypoxia. Furthermore, we showthat both HIFs-1 and -2 contribute to the maintenance of high levelsof p65 expression and phosphorylation in such cells.
As many of the genes we found to be markedly up-regulated inhuman macrophages by hypoxia had previously been identified aspotential NF-�B target genes (Table 1; Figure 4), we examined therole of p65 in the hypoxic up-regulation of the most highlyup-regulated ones. Studies using the synthetic inhibitor of nucleartranslocation of p65, JSH-23,24 or an adenoviral inhibitor of IKK-�showed that NF-�B is not essential for their induction during an18-hour exposure to hypoxia. The up-regulation of HIF-1� inmacrophages exposed to short-term hypoxia (4 hours) is partiallydependent on NF-�B,22 so the fact that both HIFs-1 and -2�continued to be up-regulated in JSH-treated MDMs after hypoxiain our study suggests that either p65 inhibition was incomplete orthe accumulation of these subunits during a more sustained period(18 hours) of hypoxia is independent of NF-�B.
However, although both forms of NF-�B inhibition resulted in themarked inhibition of TNF-�-induced CXCL8 (as well as other genesexamined with JSH-23), it had no detectable effect on the hypoxicexpression of any of the genes examined. These data are supported bythe recent finding that that the hypoxic induction of several such NF-�Btarget genes in murine BMDMs does not involve activation of NF-�B(H.Z.I. and M.C.S., manuscript submitted April 2009).
Our data indicate then that NF-�B signaling may not contributeto the induction of these genes by macrophages in response to an18-hour exposure to hypoxia. At first glance, this appears tocontrast with the finding mentioned earlier: that hypoxic induction
of HIF-1� and various HIF target genes was diminished in IKK-�–deficient BMDMs after exposure to short-term hypoxia (ie, 4 hours).22
It may be that there is a switch from acute, NF-�B-dependent hypoxicresponses in macrophages that are critical for innate immunity (eg,bacterial infection) to an NF-�B-independent, HIF-driven response tothe chronic hypoxia present at sites such as tumors. Clearly, a detailedinvestigation of the role of NF-�B in the temporal and gene-specific responses of macrophages to hypoxia is now warranted.
Taken together, our data show that, when macrophages experi-ence hypoxia for 18 hours, it elicits a profound change in theirexpression of various tumor-promoting genes. Although this studyprovides invaluable insights into the basic repertoire of suchmacrophage responses, it should be remembered that macrophagesin hypoxic areas of complex tissues such as tumors are aheterogeneous mix of cells, including immature, monocyte-likecells and mature macrophages.6 Moreover, the responses of thesecells to hypoxia will also be influenced by a host of tumor-derivedsignals, such as cytokines and enzymes. Further in vivo studies arenow warranted to investigate the role of hypoxic macrophageresponses within the complex milieu of tumors.
Acknowledgments
The authors thank Drs Bosco and Varesio (G. Gaslini Institute,Genoa, Italy) and Michael Karin (United States) for their helpfulcomments in the preparation of this manuscript, Dr ThorstenHagemann (Institute of Cancer, Barts and The London School ofMedicine and Dentistry, London, United Kingdom) for providingthe control and IKKBdn adenoviruses, and Drs Sheila Francis andSarah Walmsley (University of Sheffield Medical School, Shef-field, United Kingdom) for their help with the adenoviral infectionsand HIF-1�/� BMDM isolations, respectively.
This work was supported by Yorkshire Cancer Research, UnitedKingdom and Breast Cancer Campaign, United Kingdom.
Authorship
Contribution: H.-Y.F., R.H., C.M., S.B.C., S.K.B., E.F., F.R.G., andJ.R. designed and performed research; A.L.H. and C.E.L. contrib-uted vital new reagents or analytical tools, analyzed data, and wrote
0
20
CA
Phospho-p65/RelA
Normoxia Hypoxia
IKKBdnadv
Controladv
Normoxia HypoxiaCXCL8
Control a
dv
IKKBdn a
dv
Control a
dv
IKKBdn a
dv
B
No TNFα
+ TNFα
No adv
GLUT-1
No adv
Control a
dv
adv-
IKKβD
N
No adv
Control a
dv
0
10
20
30
*
*
*
Fol
d ch
ange
adv-
IKKβD
N
CXCL8
No adv
Control a
dv
No adv
Control a
dv
0
10
20
30
40
*
*
*
Fol
d ch
ange
adv-
IKKβD
N
adv-
IKKβD
N
VEGFA
0
10
20
30
*
*
*
No adv
Control a
dv
No adv
Control a
dv
Fol
d ch
ange
adv-
IKKβD
N
adv-
IKKβD
N
ADM
0
5
10
15*
**
No adv
Control a
dv
No adv
Control a
dv
Fol
d ch
ange
adv-
IKKβD
N
adv-
IKKβD
N
CXCR4
0
1
2
3
4
5
**
*
No adv
Control a
dv
No adv
Control a
dv
Fol
d ch
ange
adv-
IKKβD
N
adv-
IKKβD
N
+15
10
5Fo
ld c
han
ge
Controladv
IKKBdnadv
Figure 7. IKK-� inhibition has no effecton the hypoxic induction of various genesin human MDMs in vitro: use of a recom-binant adenovirus expressing a domi-nant negative inhibitor of IKK-� (adv-IKK-�DN). MDM infection with adv-IKK-�DN (butnot control adv) significantly inhibited bothTNF-�–induced gene expression of CXCL8(A) and hypoxia-induced nuclear accumula-tion of phospho-p65/RelA (B) by humanMDM. (C) Hypoxia significantly increasedthe expression of VEGFA, CXCL8, CXCR4,GLUT-1, and ADM mRNA in untreated andadenovirally infected MDM compared withrespective normoxic MDM controls. How-ever, there was no significant difference inthe expression of these genes betweenhypoxic MDM infected with adv-IKK-�DN orthe control adenovirus. N 3. *P � .05 withrespect to respective normoxic group.�P � .05 with respect to TNF-� � controladenovirus group.
858 FANG et al BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom

the paper; R.S.J. and M.C.S. contributed vital new reagents oranalytical tools and analyzed data; and H.Z.I. performed research.
Conflict-of-interest disclosure: The authors declare no compet-ing financial interests.
Correspondence: Claire E. Lewis, Academic Unit of Pathology,Faculty of Medicine, Dentistry and Health, University of Sheffield,Beech Hill Rd, Sheffield, S10 2RX, United Kingdom; e-mail:[email protected].
References
1. Vaupel P, Kelleher DK, Hockel M. Oxygen statusof malignant tumors: pathogenesis of hypoxiaand significance for tumor therapy. Semin Oncol.2001;28:29-35.
2. Taylor PC, Sivakumar B. Hypoxia and angiogen-esis in rheumatoid arthritis. Curr Opin Rheumatol.2005;17:293-298.
3. Bjornheden T, Levin M, Evaldsson M, Wiklund O.Evidence of hypoxic areas within the arterial wall invivo. Arterioscler Thromb Vasc Biol. 1999;19:870-876.
4. Bertout JA, Patel SA, Simon MC. The impact ofO2 availability on human cancer. Nat Rev Cancer.2008;8:967-975.
5. Semenza GL. Hypoxia-inducible factor 1: oxygenhomeostasis and disease pathophysiology.Trends Mol Med. 2001;7:345-350.
6. Lewis CE, Pollard JW. Distinct role of macro-phages in different tumor microenvironments.Cancer Res. 2006;66:605-612.
7. Murdoch C, Giannoudis A, Lewis CE. Mecha-nisms regulating the recruitment of macrophagesinto hypoxic areas of tumors and other ischemictissues. Blood. 2004;104:2224-2234.
8. Lewis C, Murdoch C. Macrophage responses to hyp-oxia: implications for tumor progression and anti-cancer therapies. Am J Pathol. 2005;167:627-635.
9. Murdoch C, Lewis CE. Macrophage migrationand gene expression in response to tumor hyp-oxia. Int J Cancer. 2005;117:701-708.
10. Burke B, Tang N, Corke KP, et al. Expression of HIF-1alpha by human macrophages: implications for theuse of macrophages in hypoxia-regulated cancergene therapy. J Pathol. 2002;196:204-212.
11. Talks KL, Turley H, Gatter KC, et al. The expres-sion and distribution of the hypoxia-inducible fac-tors HIF-1alpha and HIF-2alpha in normal humantissues, cancers, and tumor-associated macro-phages. Am J Pathol. 2000;157:411-421.
12. Lewis JS, Landers RJ, Underwood JC, Harris AL,Lewis CE. Expression of vascular endothelialgrowth factor by macrophages is up-regulated inpoorly vascularized areas of breast carcinomas.J Pathol. 2000;192:150-158.
13. Leek RD, Talks KL, Pezzella F, et al. Relation ofhypoxia-inducible factor-2 alpha (HIF-2 alpha)expression in tumor-infiltrative macrophages totumor angiogenesis and the oxidative thymidinephosphorylase pathway in human breast cancer.Cancer Res. 2002;62:1326-1329.
14. White JR, Harris RA, Lee SR, et al. Genetic am-plification of the transcriptional response to hyp-oxia as a novel means of identifying regulators ofangiogenesis. Genomics. 2004;83:1-8.
15. Cramer T, Yamanishi Y, Clausen BE, et al. HIF-1alpha is essential for myeloid cell-mediated in-flammation. Cell. 2003;112:645-657.
16. Park SK, Dadak AM, Haase VH, Fontana L, GiacciaAJ, Johnson RS. Hypoxia-induced gene expressionoccurs solely through the action of hypoxia-induciblefactor 1alpha (HIF-1alpha): role of cytoplasmic trap-ping of HIF-2alpha. Mol Cell Biol. 2003;23:4959-4971.
17. Sowter HM, Raval RR, Moore JW, Ratcliffe PJ,Harris AL. Predominant role of hypoxia-inducibletranscription factor (Hif)-1alpha versus Hif-2alphain regulation of the transcriptional response tohypoxia. Cancer Res. 2003;63:6130-6134.
18. Hu CJ, Sataur A, Wang L, Chen H, Simon MC.
The N-terminal transactivation domain conferstarget gene specificity of hypoxia-inducible fac-tors HIF-1alpha and HIF-2alpha. Mol Biol Cell.2007;18:4528-4542.
19. Elvert G, Kappel A, Heidenreich R, et al. Cooperativeinteraction of hypoxia-inducible factor-2alpha (HIF-2alpha) and Ets-1 in the transcriptional activation ofvascular endothelial growth factor receptor-2 (Flk-1).J Biol Chem. 2003;278:7520-7530.
20. Van Uden P, Kenneth NS, Rocha S. Regulation ofhypoxia-inducible factor-1alpha by NF-kappaB.Biochem J. 2008;412:477-484.
21. Cummins EP, Berra E, Comerford KM, et al. Prolylhydroxylase-1 negatively regulates IkappaB kinase-beta, giving insight into hypoxia-induced NFkappaBactivity. Proc Natl Acad Sci U S A. 2006;103:18154-18159.
22. Rius J, Guma M, Schachtrup C, et al. NF-kappaBlinks innate immunity to the hypoxic responsethrough transcriptional regulation of HIF-1alpha.Nature. 2008;453:807-811.
23. Peyssonnaux C, Datta V, Cramer T, et al. HIF-1alphaexpression regulates the bactericidal capacity ofphagocytes. J Clin Invest. 2005;115:1806-1815.
24. Shin HM, Kim MH, Kim BH, et al. Inhibitory ac-tion of novel aromatic diamine compound onlipopolysaccharide-induced nuclear transloca-tion of NF-kappaB without affecting IkappaBdegradation. FEBS Lett. 2004;571:50-54.
25. Greten FR, Arkan MC, Bollrath J, et al. NF-kappaB isa negative regulator of IL-1beta secretion as re-vealed by genetic and pharmacological inhibition ofIKKbeta. Cell. 2007;130:918-931.
26. Bondeson J, Browne KA, Brennan FM, Foxwell BM,Feldmann M. Selective regulation of cytokine induc-tion by adenoviral gene transfer of IkappaBalpha intohuman macrophages: lipopolysaccharide-induced,but not zymosan-induced, proinflammatory cytokinesare inhibited, but IL-10 is nuclear factor-kappaB inde-pendent. J Immunol. 1999;162:2939-2945.
27. Hagemann T, Lawrence T, McNeish I, et al. “Re-educating” tumor-associated macrophages bytargeting NF-kappaB. J Exp Med. 2008;205:1261-1268.
28. Oitzinger W, Hofer-Warbinek R, Schmid JA,Koshelnick Y, Binder BR, de Martin R. Adenovirus-mediated expression of a mutant IkappaB kinase 2inhibits the response of endothelial cells to inflamma-tory stimuli. Blood. 2001;97:1611-1617.
29. Ryan RM, Green J, Williams PJ, et al. Bacterialdelivery of a novel cytolysin to hypoxic areas ofsolid tumors. Gene Therapy. 2009;16:329-339.
30. Subramanian A, Tamayo P, Mootha VK, et al.Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wideexpression profiles. Proc Natl Acad Sci U S A.2005;102:15545-15550.
31. Bosco MC, Puppo M, Santangelo C, et al. Hyp-oxia modifies the transcriptome of primary humanmonocytes: modulation of novel immune-relatedgenes and identification of CC-chemokine ligand20 as a new hypoxia-inducible gene. J Immunol.2006;177:1941-1955.
32. Ricciardi A, Elia AR, Cappello P, et al. Transcrip-tome of hypoxic immature dendritic cells: modula-tion of chemokine/receptor expression. Mol Can-cer Res. 2008;6:175-185.
33. National Center for Biotechnology Information.GEO: Gene Expression Omnibus. http://www.
ncbi.nlm.nih.gov/geo/query/acc.cgi?acc.GSE15949. Accessed May 5, 2009.
34. National Center for Biotechnology Information.GEO: Gene Expression Omnibus. http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc.GSE1099. Accessed May 14, 2009.
35. Harris AL. Hypoxia: a key regulatory factor in tu-mour growth. Nat Rev Cancer. 2002;2:38-47.
36. Xie X, Lu J, Kulbokas EJ, et al. Systematic dis-covery of regulatory motifs in human promotersand 3� UTRs by comparison of several mammals.Nature. 2005;434:338-345.
37. Elbarghati L, Murdoch C, Lewis CE. Effects ofhypoxia on transcription factor expression in hu-man monocytes and macrophages. Immunobiol-ogy. 2008;213:899-908.
38. Oda T, Hirota K, Nishi K, et al. Activation of hypoxia-inducible factor 1 during macrophage differentiation.Am J Physiol Cell Physiol. 2006;291:C104-C113.
39. Elia AR, Cappello P, Puppo M, et al. Human dendriticcells differentiated in hypoxia down-modulate antigenuptake and change their chemokine expression pro-file. J Leukoc Biol. 2008;84:1472-1482.
40. Battaglia F, Delfino S, Merello E, et al. Hypoxiatranscriptionally induces macrophage-inflammatoryprotein-3alpha/CCL-20 in primary human mono-nuclear phagocytes through nuclear factor (NF)-kap-paB. J Leukoc Biol. 2008;83:648-662.
41. Lee MC, Wei SC, Tsai-Wu JJ, Wu CH, Tsao PN.Novel PKC signaling is required for LPS-inducedsoluble Flt-1 expression in macrophages. J Leu-koc Biol. 2008;84:835-841.
42. Dickensheets HL, Donnelly RP. IFN-gamma andIL-10 inhibit induction of IL-1 receptor type I and typeII gene expression by IL-4 and IL-13 in human mono-cytes. J Immunol. 1997;159:6226-6233.
43. Cho SS, Bacon CM, Sudarshan C, et al. Activation ofSTAT4 by IL-12 and IFN-alpha: evidence for the in-volvement of ligand-induced tyrosine and serinephosphorylation. J Immunol. 1996;157:4781-4789.
44. Hou J, Schindler U, Henzel WJ, Ho TC, BrasseurM, McKnight SL. An interleukin-4-induced tran-scription factor: IL-4 Stat. Science. 1994;265:1701-1706.
45. Apte RN, Dotan S, Elkabets M, et al. The involve-ment of IL-1 in tumorigenesis, tumor invasive-ness, metastasis and tumor-host interactions.Cancer Metastasis Rev. 2006;25:387-408.
46. Hirani N, Antonicelli F, Strieter RM, et al. Theregulation of interleukin-8 by hypoxia in humanmacrophages: a potential role in the pathogene-sis of the acute respiratory distress syndrome(ARDS). Mol Med. 2001;7:685-697.
47. Zhang W, Petrovic JM, Callaghan D, et al. Evi-dence that hypoxia-inducible factor-1 (HIF-1) me-diates transcriptional activation of interleukin-1beta (IL-1beta) in astrocyte cultures.J Neuroimmunol. 2006;174:63-73.
48. Carroll VA, Ashcroft M. Role of hypoxia-inducible fac-tor (HIF)-1alpha versus HIF-2alpha in the regulationof HIF target genes in response to hypoxia, insulin-like growth factor-I, or loss of von Hippel-Lindau func-tion: implications for targeting the HIF pathway. Can-cer Res. 2006;66:6264-6270.
49. Meade ES, Ma YY, Guller S. Role of hypoxia-inducible transcription factors 1alpha and 2alphain the regulation of plasminogen activator inhibi-tor-1 expression in a human trophoblast cell line.Placenta. 2007;28:1012-1019.
MACROPHAGE RESPONSE TO HYPOXIA 859BLOOD, 23 JULY 2009 � VOLUME 114, NUMBER 4
For personal use only. by guest on May 18, 2011. bloodjournal.hematologylibrary.orgFrom





























