Embed Size (px)
Citation preview
DeutscherTanmay Dutta, Arun Malhotra and Murray P. Zof the Processing Enzyme RNase BN/RNasetRNAs and tRNA Precursors from Action How a CCA Sequence Protects MatureRNA:
doi: 10.1074/jbc.M113.514570 originally published online September 10, 20132013, 288:30636-30644.J. Biol. Chem.
10.1074/jbc.M113.514570Access the most updated version of this article at doi:
.JBC Affinity SitesFind articles, minireviews, Reflections and Classics on similar topics on the
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/288/42/30636.full.html#ref-list-1
This article cites 33 references, 20 of which can be accessed free at
at UN
IV O
F MIA
MI M
ILL
ER
SCH
OO
L O
F ME
DIC
INE
on Decem
ber 23, 2013http://w
ww
.jbc.org/D
ownloaded from
at U
NIV
OF M
IAM
I MIL
LE
R SC
HO
OL
OF M
ED
ICIN
E on D
ecember 23, 2013
http://ww
w.jbc.org/
Dow
nloaded from

How a CCA Sequence Protects Mature tRNAs and tRNAPrecursors from Action of the Processing Enzyme RNaseBN/RNase Z*
Received for publication, August 28, 2013, and in revised form, September 9, 2013 Published, JBC Papers in Press, September 10, 2013, DOI 10.1074/jbc.M113.514570
Tanmay Dutta, Arun Malhotra, and Murray P. Deutscher1
From the Department of Biochemistry and Molecular Biology, University of Miami Miller School of Medicine, Miami, Florida 33136
Background: 3�-Terminal CCA-containing tRNAs and precursors are resistant to action of RNase BN/RNase Z.Results: Arg274 and the two C residues are required for protection by the CCA sequence.Conclusion: Presence of Arg274 and CC sequence prevents RNA substrate from moving into the RNase catalytic site.Significance: This mechanism explains how mature tRNAs are protected from removal of the CCA sequence by a processingRNase.
In many organisms, 3� maturation of tRNAs is catalyzed bythe endoribonuclease, RNase BN/RNase Z, which cleaves afterthe discriminator nucleotide to generate a substrate for additionof the universal CCA sequence. However, tRNAs or tRNA pre-cursors that already contain a CCA sequence are not cleaved,thereby avoiding a futile cycle of removal and readditionof theseessential residues.We show here that the adjacent C residues ofthe CCA sequence and anArg residue within a highly conservedsequencemotif in the channel leading to the RNase catalytic siteare both required for the protective effect of the CCA sequence.When both of these determinants are present, CCA-containingRNAs in the channel are unable to move into the catalytic site;however, substitution of either of the C residues by A or U ormutation of Arg274 to Ala allows RNA movement and catalysisto proceed.These data define anovelmechanism for how tRNAsare protected against the promiscuous action of a processingenzyme.
Cells contain a large repertoire of RNases that are involved ineither maturation or targeted degradation of cellular RNAs, aprocess collectively known as RNA metabolism (1–4). Inas-much as many of these RNases are also destructive enzymeswith the ability to degrade or damage important functionalRNAs (5, 6), various regulatory mechanismsmust have evolvedto protect cellular RNAs from unwanted degradation by theseenzymes, although very little information is currently availablein this area. Some RNAs such as tRNA and rRNA are stable incells because they have extensive secondary structure or con-tain protective nucleotide sequences or because they aresequestered in RNP particles. In addition, certain RNases mayalso have built-in specificities that limit their action on func-tional RNAs (7). In Escherichia coli, which contains more than20RNaseswith different characteristics, it is likely thatmultiplemechanisms operate to protect cellular RNAs.
One clear example of such a protective mechanism involvesthe enzyme, RNase BN (8–11). RNase BN is the E. colimemberof the RNase Z family of enzymes that participate in 3� matura-tion of tRNA precursors in many organisms (12). FunctionaltRNAs require the universal CCAsequence at their 3�-terminusfor amino acid attachment and for aminoacyl-tRNA action onthe ribosome. In some bacteria such as E. coli, this sequence isalready present in all the tRNA precursors, and 3� maturationinvolves removal of extra residues following theCCA sequence,a process catalyzed by any one of a number of exoribonucleases(13–15). In organisms lacking an encoded -CCA sequence in allor some of their tRNA precursors, RNase Z makes an endonu-cleolytic cleavage following the discriminator nucleotide, andCCA is added by tRNA nucleotidyltransferase (16, 17). How-ever, in no case does RNase Z remove the CCA sequence,resulting in protection of mature tRNAs (10, 11, 18).In contrast to other members of the RNase Z family, which
act solely as endoribonucleases (16, 17), RNase BN also displaysa 3�-5� exoribonuclease activity onmodel RNAs and tRNA pre-cursor substrates in vitro (9, 10), and it acts as a dual functionnuclease on tRNA precursors in vivo (11). RNase BN action onE. coli tRNA precursors that already contain a CCA sequenceinvolves either a single endonucleolytic cleavage after the CCAsequence or exonucleolytic trimming to remove extra 3� resi-dues up to the CCA sequence, depending on the metal ioncofactor present (10). Like other RNase Z family members, italso does not remove the CCA sequence from mature tRNAs(10, 11, 18). Thus, RNase BN and other members of the RNaseZ family have built-in specificity to avoid action on alreadyCCA-containing precursors and on 3� mature tRNAs, therebyavoiding a futile cycle of removal and resynthesis of CCAsequences. The mechanism of how this is accomplished is ofconsiderable interest because it results in protection of maturetRNA molecules from unwanted damage.In this study, we examined the mechanism of how the CCA
sequence protects against RNase BN activity. Based onsequence alignments and structural model building, we identi-fied a conserved amino acid residue, Arg274, located in thechannel leading to the catalytic site of RNase BN that isrequired for discrimination against mature tRNAs. In addition,
* This work was supported, in whole or in part, by National Institutes of HealthGrant GM16317 (to M. P. D.).
1 To whom correspondence should be addressed: Dept. of Biochemistry andMolecular Biology, University of Miami Miller School of Medicine, P. O. Box016129, Miami, FL 33101. Tel.: 305-243-3150; Fax: 305-243-3955; E-mail:[email protected].
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 288, NO. 42, pp. 30636 –30644, October 18, 2013© 2013 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
30636 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
at UN
IV O
F MIA
MI M
ILL
ER
SCH
OO
L O
F ME
DIC
INE
on Decem
ber 23, 2013http://w
ww
.jbc.org/D
ownloaded from

we determined the contribution of the CCA sequence itself tothe protective effect. We found that two adjacent C residuescompletely block the action of RNase BN. Furthermore, wewere able to show that the presence of both Arg274 and the twoC residues preventsCCA-containingRNAs in the channel frommoving to the catalytic site. Replacement of Arg274 or either ofthe twoC residues in theCCA sequence allows RNA to proceedto the catalytic site. These data indicate that Arg274 interactswith the C residues of the CCA sequence leading to inhibitionof RNase BN action. Based on these findings, we conclude thatRNases can have built-in specificities that prevent their indis-criminate action onRNA substrates, thus explaining onemech-anism that maintains the stability of mature tRNAs.
EXPERIMENTAL PROCEDURES
Materials—RNA oligonucleotides were synthesized byDharmacon, Inc. ExpressHyb hybridization solution was pur-chased from Clontech. DpnI was from Fermentas. [�-32P]ATP,5�-[32P]pCp, andGeneScreen Plus hybridization transfermem-brane were obtained from PerkinElmer Life Sciences. T4 poly-nucleotide kinase was obtained from New England Biolabs. T4RNA ligase, calf intestine alkaline phosphatase, NucwayTM spincolumns, and the MEGAshortscriptTM kit were purchasedfrom Ambion Inc. The GeneEluteTM PCR clean-up kit andbis(p-nitrophenyl) phosphate were from Sigma. The KOD HotStart DNA polymerase was purchased fromNovagen. Sequagelfor denaturing urea-polyacrylamide gels was obtained fromNational Diagnostics. The His-Trap HP column was obtainedfrom GE Healthcare. All other chemicals were reagent grade.Site-directed Mutagenesis, Overexpression, and Purification
of the Mutant Protein—The wild type rbn gene was cloned intoplasmid pET15b, and its overexpression and purification weredescribed previously (12). Site-directed mutagenesis withprimers R1 (CGTCAGCTCGGCATATGATGACAAAGGTT-GTCA) and R2 (TGTCATCATATGCCGAGCTGACGTGG-GTAATGATTAG) was used to introduce themutation R274Ainto a plasmid-encoded rbn gene. KOD Hot Start DNApolymerase was used for PCR. Wild type rbn-containingtemplate plasmid was then digested by DpnI treatment at 37 °Cfor 2 h. Plasmids containing the mutant rbn gene were purifiedusing a gel extraction kit (Qiagen) and transformed into E. coliBL21(DE3)I�II�. Mutant protein was overexpressed in E. colistrain BL21(DE3)I�II�/pLys and purified using the same proc-edure used previously for purification of wild type His-taggedRNase BN (9, 12). The purity ofwild type andmutant RNase BNproteins was determined on an overloaded SDS-polyacryla-mide gel (�3.0 �g of the purified protein). For all of the prot-eins, a single band at �35 kDa was observed without any de-tectable contaminating bands.Synthesis and 3�-End Labeling of Mature tRNA—E. coli
genomic DNA was used as the template for PCR using KODHot StartDNApolymerase to synthesize full-length tRNAPhe inan in vitro transcription reaction using the MEGAshort-scriptTM transcription kit as described (10). The forwardprimer in the PCR contained theT7RNApolymerase promotersequence. PCR products were purified using the GeneEluteTM
PCR clean-up kit. Mature tRNA was purified by phenol/chlo-
roform/isoamylalcohol (25:24:1) extraction followed by etha-nol precipitation as described (10).Transfer-RNAPhe was labeled at its 3�-end with 5�-[32P]pCp
using T4 RNA ligase in the presence of unlabeled ATP at 4 °Cfor 16 h as described previously (10). NucwayTM spin columnwas used to remove unincorporated 5�-[32P]pCp, and the3�-terminal phosphate from 3�-[32P]pCp-labeled tRNAPheV
was removed by calf intestine alkaline phosphatase. Afterdephosphorylation, 3�-end-labeled tRNAPheV was purified asdescribed (10).RNase BN/Z Assay—A typical 30-�l reaction mixture con-
tained 10 mM Tris-HCl, pH 7.5, 200 mM potassium acetate, 5mM MgCl2, and 3�-[32P]pC-labeled tRNAs (�0.05 �M) or5�-32P-labeled model RNA substrates (10 �M) and 0.14 �M
purified wild type or mutant RNase BN, except as otherwisestated in the figure legends. Reaction mixtures were incubatedat 37 °C. Portions were taken at the indicated times, and thereaction was terminated by the addition of 2 volumes of gelloading buffer (90% formamide, 20 mM EDTA, 0.05% SDS,0.025% bromphenol blue, and 0.025% xylene cyanol). Reactionproductswere resolved on 20%denaturing 7.5Murea polyacryl-amide gels and visualized using a STORM 840 phosphorimag-ing device (GE Healthcare). ImageQuant (GE Healthcare) wasused to quantitate the bands.Phosphodiesterase Assay—Standard reaction conditions for
the determination of the phosphodiesterase activity of RNaseBN were 20 mM Tris-HCl (pH 7.4), 2 mM bis(p-nitrophenyl)phosphate, 0.6 �M of purified His-tagged wild type or mutantRNase BN, and 5 mM MgCl2. Release of p-nitrophenol (� �11,500 M�1 cm�1 at pH 7.4) was continuously monitored for 2min at 405 nm. One unit of activity corresponds to 1 �mol ofp-nitrophenol liberated/min at 37 °C.Northern Blot Analysis—tRNAPheV samples digested by
either wild type or mutant RNase BN were resolved on a 6%denaturing polyacrylamide gel in 0.5� Tris borate/EDTAbuffer and transferred to a nitrocellulose membrane by hori-zontal transfer for 1.5 h at 150 mA using 0.5� Tris borate/EDTA as the transfer solution. DNA oligonucleotide probescomplementary to the 5�-end of the tRNA were 32P-labeled attheir 5�-ends with T4 polynucleotide kinase. Probes wereallowed to anneal to the transferred RNA by overnight incuba-tion in ExpressHyb hybridization solution (Clontech Laborato-ries Inc.), and the detected bands were visualized by Phosphor-Imager analysis.
RESULTS
In early work from our laboratory, it was found that RNaseBN could remove the 3�-terminal residue from tRNA-CU ortRNA-CA in vitro, whereas tRNA-CC and tRNA-CCA wereessentially inactive as substrates (8). Subsequent work showedthat RNase BN can remove residues following a CCA sequencebut that it stops at the CCA sequence in amodel RNA substrate(9).We also found that in vivoRNase BN can process the 3�-endof E. coli tRNA precursors without removing their encodedCCA sequence (11). Likewise, the CCA sequence has a protec-tive effect on other members of the RNase Z family (17, 19). Allof these data raise the interesting question of what structuralfeatures of RNase BN/RNase Z and of the CCA sequence make
Mechanism of the CCA Inhibition of RNase BN
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30637
at UN
IV O
F MIA
MI M
ILL
ER
SCH
OO
L O
F ME
DIC
INE
on Decem
ber 23, 2013http://w
ww
.jbc.org/D
ownloaded from

mature tRNA and CCA-containing precursors resistant to theactions of this RNase, thereby avoiding a futile cycle of removaland resynthesis of the 3�-CCA terminus.Each C Residue of the CCA Sequence Affects Resistance to
RNase BN Action—Because tRNA-CCA and tRNA-CC wereessentially resistant to RNase BN, whereas tRNA-CU andtRNA-CAwere substrates (8), it suggested that two consecutiveC residues are sufficient to prevent RNase BN action. To exam-ine this point in more detail and to evaluate the role of each Cresidue, we constructed a series of model oligonucleotide RNAsubstrates with the sequence G5A12NNA, where N representsC, U, or A. Each of the substrates was 32P-labeled at its 5�-end.The action of RNase BN on these substrates was assessed byacrylamide gel analysis, and the disappearance of the startingmaterial was quantified (Table 1). In agreement with earlierwork (9), the substrate terminating with a CCA sequence waslargely resistant to RNase BN under these assay conditions. Incontrast, replacement of either C residue with a U or A res-idue rendered the oligonucleotide sensitive to RNase BN.Substitution of the C residue closer to the 3�-end had agreater effect than replacing the more 5� C residue, but it isclear that each provides a major protective effect, indicatingthat two adjacent C residues are required to afford essen-tially complete protection.Although the first series of model substrates was designed to
mimic mature tRNAs in which the CCA sequence is at the3�-end of the RNA, we also examined a second group of sub-strates designed to mimic tRNA precursors. In these modelsubstrates, the CCA sequence or its derivatives are followed byfive additional A residues. As shown in Fig. 1, RNase BNremoves the A residues following the CCA sequence, but thenstops (lanes 7–9). In contrast, when the CCA sequence isreplaced by CAA (lanes 2–4), ACA (lanes 10–12), or UUA(lanes 14–16), RNase BN action continues through the resi-dues. Although there is a slight pause in the CAA- and ACA-containing substrates that is not seenwith theUUAsubstrate, itis clear that only the presence of two adjacent C residues as isfound in the CCA sequence results in almost a complete stop.These data indicate that RNase BN is essentially blocked by aCCA sequence when it resides in either a mature or a tRNAprecursor-like substrate. Based on these findings, we concludethat RNaseBNspecifically recognizes the adjacentC residues in
the CCA sequence, resulting in its inability to remove thesenucleotides from an RNA molecule.Identification of AminoAcid Residues in RNase BNThatMay
Interact with the CCA Sequence—It is known from the crystalstructures of E. coli RNase BN (20) and Bacillus subtilis RNaseZ (21–23) that each member of the RNase Z family is a dimerwith a core zinc-dependent�-lactamase domain that contains aHXHXDH metal binding motif as well as additional His andAsp residues that also coordinate to the two metal ions. Eachsubunit possesses a protruding flexible arm that is believed tohave a role in tRNA binding. The metal-binding site is thoughtto be the catalytic site of the protein. A channel leading to themetal-binding site, which we term the catalytic channel, isresponsible for the stable binding of the RNA substrate to facil-itate the catalytic process. A channel leading from the metal-binding site, called the exit channel, binds the 3�-extra residuesof tRNA precursors that are removed by endonucleolyticcleavage.The fact that RNase BN exoribonucleolytically removes res-
idues following the CCA sequence (Fig. 1) but then stops sug-gests that the terminal A residue of the CCA sequence is unableto move into the catalytic site. Taken together with the conclu-sion that the enzyme specifically recognizes the adjacent C res-idues of the CCA sequence, these data imply that interaction ofone ormore amino acidswithin the catalytic channel with theCresidues is responsible for the inability of RNase BN to act onCCA-containing RNA substrates. This idea was re-enforced byour findings that mutations of amino acids in the exit channelhad no effect on the protective function of the CCA sequence(data not shown). Thus, we focused our attention on aminoacids in the catalytic channel that were positioned such thatthey could interact with the two C residues of the CCA
TABLE 1Contribution of C residues to protection against the action of RNaseBNModel RNAs (10�M)with the sequenceG5A12Xwere labeledwith 32P at their 5� endas described under “Experimental Procedures” and treated with RNase BN (1.7 �M)for 20 and 40min. Digestion products were analyzed by 20% denaturing PAGE. Theamount of substrate remaining after 20 and 40min of digestionwas quantified usingImageQuant (GE Healthcare). The data are expressed as the percentages of radio-activity present at 0 min for each model RNA. The values shown are the averages ofthree independent experiments.
Substrate (G5A12NNN)Substrate remaining
20 min 40 min
%NNN � CCA 96 � 2 94 � 3
UCA 25 � 3 6 � 1ACA 21 � 3 7 � 2CUA 7 � 1 5 � 1CAA 8 � 2 4 � 1
FIGURE 1. Action of RNase BN on CCA-containing and various CCA-lessmodel RNA substrates. Reactions were carried out as described under“Experimental Procedures” using 5�-32P-labeled single-stranded model RNAsubstrates with the sequence G5A12XA5 (10 �M). Digestion was carried outwith purified RNase BN (1.7 �M) in the presence of Mg2�, and portions werewithdrawn at 0, 20, and 40 min. Digestion products were analyzed by 20%denaturing PAGE. The numbers at the bottom of the gel are the lane numbers,and the numbers on the side are the chain lengths of standards (25 or 18) or ofa product (6).
Mechanism of the CCA Inhibition of RNase BN
30638 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
at UN
IV O
F MIA
MI M
ILL
ER
SCH
OO
L O
F ME
DIC
INE
on Decem
ber 23, 2013http://w
ww
.jbc.org/D
ownloaded from
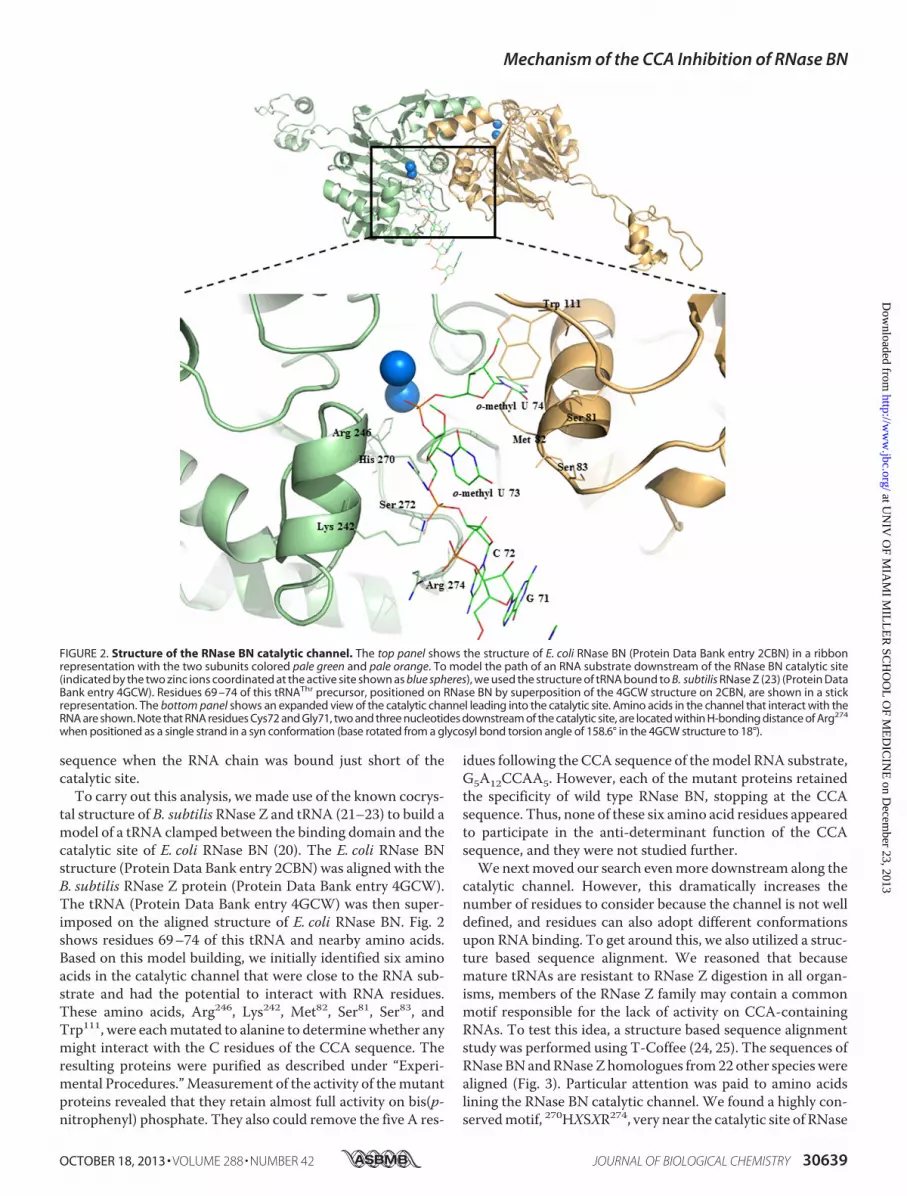
sequence when the RNA chain was bound just short of thecatalytic site.To carry out this analysis, we made use of the known cocrys-
tal structure of B. subtilis RNase Z and tRNA (21–23) to build amodel of a tRNA clamped between the binding domain and thecatalytic site of E. coli RNase BN (20). The E. coli RNase BNstructure (Protein Data Bank entry 2CBN) was aligned with theB. subtilis RNase Z protein (Protein Data Bank entry 4GCW).The tRNA (Protein Data Bank entry 4GCW) was then super-imposed on the aligned structure of E. coli RNase BN. Fig. 2shows residues 69–74 of this tRNA and nearby amino acids.Based on this model building, we initially identified six aminoacids in the catalytic channel that were close to the RNA sub-strate and had the potential to interact with RNA residues.These amino acids, Arg246, Lys242, Met82, Ser81, Ser83, andTrp111, were eachmutated to alanine to determinewhether anymight interact with the C residues of the CCA sequence. Theresulting proteins were purified as described under “Experi-mental Procedures.”Measurement of the activity of themutantproteins revealed that they retain almost full activity on bis(p-nitrophenyl) phosphate. They also could remove the five A res-
idues following the CCA sequence of themodel RNA substrate,G5A12CCAA5. However, each of the mutant proteins retainedthe specificity of wild type RNase BN, stopping at the CCAsequence. Thus, none of these six amino acid residues appearedto participate in the anti-determinant function of the CCAsequence, and they were not studied further.We nextmoved our search evenmore downstream along the
catalytic channel. However, this dramatically increases thenumber of residues to consider because the channel is not welldefined, and residues can also adopt different conformationsupon RNA binding. To get around this, we also utilized a struc-ture based sequence alignment. We reasoned that becausemature tRNAs are resistant to RNase Z digestion in all organ-isms, members of the RNase Z family may contain a commonmotif responsible for the lack of activity on CCA-containingRNAs. To test this idea, a structure based sequence alignmentstudy was performed using T-Coffee (24, 25). The sequences ofRNase BNandRNaseZhomologues from22 other specieswerealigned (Fig. 3). Particular attention was paid to amino acidslining the RNase BN catalytic channel. We found a highly con-servedmotif, 270HXSXR274, very near the catalytic site of RNase
FIGURE 2. Structure of the RNase BN catalytic channel. The top panel shows the structure of E. coli RNase BN (Protein Data Bank entry 2CBN) in a ribbonrepresentation with the two subunits colored pale green and pale orange. To model the path of an RNA substrate downstream of the RNase BN catalytic site(indicated by the two zinc ions coordinated at the active site shown as blue spheres), we used the structure of tRNA bound to B. subtilis RNase Z (23) (Protein DataBank entry 4GCW). Residues 69 –74 of this tRNAThr precursor, positioned on RNase BN by superposition of the 4GCW structure on 2CBN, are shown in a stickrepresentation. The bottom panel shows an expanded view of the catalytic channel leading into the catalytic site. Amino acids in the channel that interact with theRNA are shown. Note that RNA residues Cys72 and Gly71, two and three nucleotides downstream of the catalytic site, are located within H-bonding distance of Arg274
when positioned as a single strand in a syn conformation (base rotated from a glycosyl bond torsion angle of 158.6° in the 4GCW structure to 18°).
Mechanism of the CCA Inhibition of RNase BN
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30639
at UN
IV O
F MIA
MI M
ILL
ER
SCH
OO
L O
F ME
DIC
INE
on Decem
ber 23, 2013http://w
ww
.jbc.org/D
ownloaded from

BN that has the potential to interact with residues of tRNAlocated in the catalytic channel. In fact, His270 in this conservedmotif is a known essential amino acid that coordinates to one ofthe twometal ions. His270 is present in a loop that extends awayfrom the metal-binding site toward the catalytic channel. Thetwo other conserved residues in the motif, Ser272 and Arg274,are also present in this loop. The extensive sequence conserva-tion of this motif, as well as that of nearby amino acids, made itan interesting candidate for additional mutational analysis.Effect of Mutations in the Conserved HXSXR Motif on RNase
BN Activity—To examine the role of amino acids in the con-served HXSXRmotif on RNase BN activity and specificity, sev-eral mutations were made within the motif, and the resultingproteins were purified. These included conversion of His270,Ser272, or Arg274 to alanine. The activity of these mutant pro-teins on both bis(p-nitrophenyl) phosphate and onmodel RNAsubstrates was measured. His270 is part of the active site, and asmight be expected, its conversion to alanine caused a loss ofRNase BN activity on both bis(p-nitrophenyl) phosphate, asmall molecule that is the chromogenic substrate for the phos-phodiesterase activity of RNase BN, andmodel RNAs (data notshown).Mutant S272Awas also inactive onmodel RNAs, and itshowed greatly reduced activity on the chromogenic substrate(data not shown). Consequently, we were unable to studyeither of these mutant proteins. In contrast, mutation ofArg274 to alanine only reduced phosphodiesterase activity�25% compared with the wild type protein, and activityagainst model RNAs was reduced to �30% of wild typeRNase BN. Most importantly, this mutation changed thespecificity of RNase BN.To explore how the R274A mutation affected the specificity
of RNase BN, we made use of the model substrates G5A12CCAand G5A12CCAA5. As shown above and in Fig. 4A, wild type
RNase BN is unable to act onG5A12CCA. (The small amount ofproduct generated is due to contamination of the substrate byshorter molecules lacking a CCA sequence because the full-length substrate is essentially unaltered over a period of 2 h.)Onthe other hand, the R274A mutant protein digests G5A12CCAeffectively such that �90% is gone in 2 h, generating a range ofshorter products. (RNase BN is known to slow down consider-ably when the product length is reduced to �10 nucleotides(9).) On G5A12CCAA5 (Fig. 4B), wild type RNase BN removesthe five A residues but stops at the CCA sequence. The R274Amutant protein also removes the A residues, but it continuesthrough the CCA sequence, generating the same shorter prod-ucts as in Fig. 4A. These data show that the R274A mutant
FIGURE 3. Members of RNase Z family contain a conserved HXSXR motif located near the catalytic site. A structure-based sequence alignment wasgenerated by T-Coffee using sequences of RNase Z proteins from 22 different species. ESPript was used to present the sequence alignment (26). The sequencesare from the Uniprot database. The sequences included are: RBN_ECOLI, E. coli RNase BN (entry no. P0A8V0); UniRef90_C4BUZ1, Enterobacteriaceae RNase BN;F7RBB1_SHIFL, Shigella flexneri RNase BN (F7RBB1); B7LM62_ESCF3, Escherichia fergusonii RNase BN (B7LM62); RBN_SALA4, Salmonella agona RNase BN(B5EZJ2); C1M711_9ENTR, Citrobacter sp. RNase BN (C1M711); D6DRB4_ENTCL, Enterobacter cloacae RNase BN (D6DRB4); RBN_KLEP3, Klebsiella pneumoniaRNase BN (B5XNW7); H5UXS8_ESCHE, Escherichia hermannii RNase BN (H5UXS8); RBN_ERWT9, Erwinia tasmaniensis RNase BN (B2VHF1); RNZ_BACSU, B. subtilisRNase Z (P54548); NZ_BACAN, Bacillus anthracis RNase Z (Q81M88); NZ_BACC2, Bacillus cereus RNase Z (B71WQ5); RNZ_ENTFA, Enterococcus faecalis RNase Z(Q834G2); RNZ_LACLA, Lactococcus lactis RNase Z (Q9CHT8); RNZ_STRZJ, Streptococcus pneumonia RNase Z (C1CD41); RNZ_SYNY3, Synechocystis sp. RNase Z(Q55132); NZ_MICAN, Microcystis aeruginosa RNase Z (B0JGG3); RNZ_STAEQ, Staphylococcus epidermidis RNase Z (Q5HP47); RNZ_LACAC, Lactobacillus aci-dophilus RNase Z (Q5FKH3); RNZ2_MOUSE, Mus musculus ElaC2 protein (Q80Y81); and RNZ2_MOUSE, Homo sapiens ElaC2 (Q9BQ52). The stars indicate aconserved motif present in the catalytic channel. Other conserved residues are highlighted including the catalytic site residues Asp212 and His270.
FIGURE 4. Activity of wild type and R274A mutant RNase BN on CCA-con-taining mature and precursor model tRNA substrates. 5�-32P-labeled sin-gle-stranded model RNA substrates (10 �M) with the sequence G5A12CCA (A)or G5A12CCAA5 (B) were digested with wild type or mutant RNase BN (13.6 �M)as described under “Experimental Procedures.” Portions were withdrawn atthe indicated times and analyzed by denaturing PAGE. The values at the bot-tom of each lane indicate the percentage of the initial substrate remaining.Note that the band labeled 6 is the limit product generated by the WTenzyme.
Mechanism of the CCA Inhibition of RNase BN
30640 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
at UN
IV O
F MIA
MI M
ILL
ER
SCH
OO
L O
F ME
DIC
INE
on Decem
ber 23, 2013http://w
ww
.jbc.org/D
ownloaded from

RNase BN can act on model RNA substrates that mimic bothmature and precursor tRNAs. The findings also indicate thatthe Arg274 residue in the catalytic channel of RNase BN plays acritical role in the CCA inhibition of RNase BN action, suggest-ing that Arg274 interacts with the C residues of the CCAsequence.Action of R274A Mutant RNase BN on tRNA—RNase BN
does not act onmature tRNAeither in vitro or in vivo because ofthe presence of aCCA sequence (10, 11). From the above exper-iments, it is evident that the R274A mutant protein is notaffected by a CCA sequence onmodel RNA substrates. Thus, itwas of considerable interest to determine whether the mutantRNase BN protein can also act on tRNA substrates particularlybecause RNase BN functions primarily as an exoribonucleaseonmodel RNA substrates, but as an endoribonuclease on tRNAprecursors in the presence of Mg2�, cleaving after the CCAsequence (10, 11). It does not work at all on mature tRNAs (10,11). To examine how the R274Amutant protein acts on a tRNAsubstrate, full-length tRNAPhe fromE. coli (Fig. 5A) was synthe-sized as described under “Experimental Procedures” and wasdigested with either wild type or mutant RNase BN. The origi-nal substrate and cleavage products were detected by Northernblot analysis using a labeled DNA oligonucleotide complemen-
tary to the 5�-end of tRNAPhe. As shown in Fig. 5B, full-lengthtRNA was resistant to wild type RNase BN; however, mutantRNase BN removes the CCA sequence from the 3�-end of thetRNA to generate a three-nucleotide shorter product. Thesedata indicate that the inhibitory effect of the CCA sequence oftRNA substrates on RNase BN activity is relieved by mutationof Arg274.To obtain additional information on how the Arg274 mutant
protein removes the CCA sequence, tRNAPhe was labeled at its3�-endwith 5�-[32P]pCp andT4RNA ligase. Calf intestine alka-line phosphate was then used to remove the 3�-terminal phos-phate, and the resulting tRNA-CCA-[32P]C was used as thesubstrate in reactions with wild type and mutant RNase BN.The gel in Fig. 5C shows that the wild type protein generatesCMP, because it can remove the single extra nucleotide but isunable to remove any part of the CCA sequence. In contrast, atetranucleotide product was observed usingmutant RNase BN,and noCMPwas generated. The additional bands seenwith theR274A mutant have not been identified but may representsome nonspecific cleavages. These data demonstrate that theR274A mutant protein removes the CCA sequence from full-length tRNA endonucleolytically, cleaving after the discrimina-tor nucleotide as is usual for members of the RNase Z familywhen the CCA sequence is absent. Thus, the CCA sequencedoes not display its usual inhibitory properties in the presenceof the mutant RNase BN.Use of Phosphodiesterase Assay to Explain How the CCA
Sequence Inhibits RNase BN—The data presented to this pointindicate that both the substrate (CC sequence) and the enzyme(Arg274) contribute to the inability of RNase BN to act on CCA-containing RNA molecules. To explain the mechanism of thisinhibition, we hypothesized that Arg274, located in the catalyticchannel, interacts with the two adjacent C residues of the CCAsequence and prevents the 3� terminus of the RNA moleculefrom moving into the catalytic site (Fig. 6). This hypothesispredicts that when both Arg274 and the CC sequence are pres-ent, the catalytic site will remain empty, whereas when eitherdeterminant is absent, the RNA substrate will move into the
FIGURE 5. Action of wild type and R274A mutant RNase BN on tRNAPhe. A,structure of tRNAPheV is shown with the 3�-terminal CCA sequence in boldtype. The 3�-terminal [32P]C residue used in C is in parentheses. B, wild type andmutant RNase BN (1.12 �M) were used to digest full-length tRNAPhe (�0.05�M). Portions were withdrawn at 0, 1.5, and 3 h. Cleavage products wereanalyzed by 6% denaturing PAGE, followed by Northern blotting using a5�-probe. M, full-length tRNA. C, full-length tRNA (�0.05 �M), labeled with[32P]pC at its 3�-end, was treated with wild type and mutant RNase BN (1.12�M) for 0, 1.5, and 3 h. Digestion products were analyzed by 20% denaturingPAGE. P, precursor tRNA.
FIGURE 6. Schematic representation of the cleavage of CCA-containingand CCA-less tRNAs at the catalytic site of RNase BN. cat site is the catalyticsite of RNase BN, and N represents either A or U residues replacing C residuesin the CCA sequence. The arrows indicate the sites of RNA cleavage after thediscriminator nucleotide (D) of tRNA. 1, Arg274 of wild type RNase BN interactswith adjacent C residues at the 3�-end of tRNA and prevents tRNA from mov-ing into the catalytic site. 2 and 3, mutation of Arg274 to Ala (step 2) or substi-tution of adjacent C residues with A or U residues (step 3) allows the 3�-end oftRNA to move into the catalytic site enabling cleavage to occur after thediscriminator nucleotide. Bis(p-nitrophenyl) phosphate, the phosphodiester-ase (PDase) substrate, binds to the catalytic site when Arg274 and CCA arepresent (1) but is unable to bind when either one of these two determinantsis absent (steps 2 and 3).
Mechanism of the CCA Inhibition of RNase BN
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30641
at UN
IV O
F MIA
MI M
ILL
ER
SCH
OO
L O
F ME
DIC
INE
on Decem
ber 23, 2013http://w
ww
.jbc.org/D
ownloaded from
catalytic site, and RNase actionwill proceed (Fig. 6). To test thisprediction, we developed an assay using bis(p-nitrophenyl)phosphate. When the catalytic site is unoccupied because theRNA substrate is prevented from entering, bis(p-nitrophenyl)phosphate is able to enter, and phosphodiesterase activity willbe observed. However, when the RNA substrate can move intothe catalytic site, phosphodiesterase activity will be inhibited.This assay can distinguish between an empty and an occu-
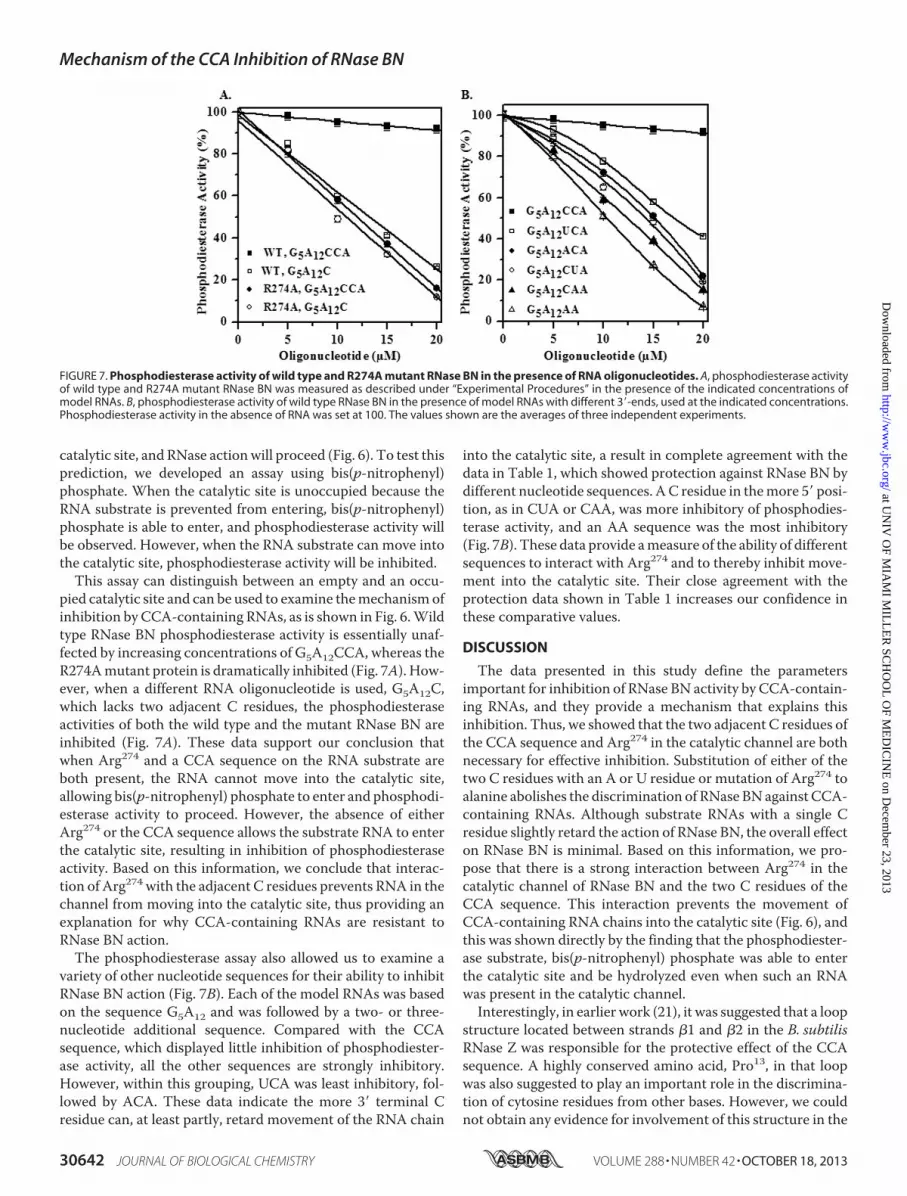
pied catalytic site and can be used to examine themechanismofinhibition by CCA-containing RNAs, as is shown in Fig. 6.Wildtype RNase BN phosphodiesterase activity is essentially unaf-fected by increasing concentrations of G5A12CCA, whereas theR274Amutant protein is dramatically inhibited (Fig. 7A). How-ever, when a different RNA oligonucleotide is used, G5A12C,which lacks two adjacent C residues, the phosphodiesteraseactivities of both the wild type and the mutant RNase BN areinhibited (Fig. 7A). These data support our conclusion thatwhen Arg274 and a CCA sequence on the RNA substrate areboth present, the RNA cannot move into the catalytic site,allowing bis(p-nitrophenyl) phosphate to enter and phosphodi-esterase activity to proceed. However, the absence of eitherArg274 or the CCA sequence allows the substrate RNA to enterthe catalytic site, resulting in inhibition of phosphodiesteraseactivity. Based on this information, we conclude that interac-tion of Arg274 with the adjacent C residues prevents RNA in thechannel from moving into the catalytic site, thus providing anexplanation for why CCA-containing RNAs are resistant toRNase BN action.The phosphodiesterase assay also allowed us to examine a
variety of other nucleotide sequences for their ability to inhibitRNase BN action (Fig. 7B). Each of the model RNAs was basedon the sequence G5A12 and was followed by a two- or three-nucleotide additional sequence. Compared with the CCAsequence, which displayed little inhibition of phosphodiester-ase activity, all the other sequences are strongly inhibitory.However, within this grouping, UCA was least inhibitory, fol-lowed by ACA. These data indicate the more 3� terminal Cresidue can, at least partly, retard movement of the RNA chain
into the catalytic site, a result in complete agreement with thedata in Table 1, which showed protection against RNase BN bydifferent nucleotide sequences. AC residue in themore 5� posi-tion, as in CUA or CAA, was more inhibitory of phosphodies-terase activity, and an AA sequence was the most inhibitory(Fig. 7B). These data provide ameasure of the ability of differentsequences to interact with Arg274 and to thereby inhibit move-ment into the catalytic site. Their close agreement with theprotection data shown in Table 1 increases our confidence inthese comparative values.
DISCUSSION
The data presented in this study define the parametersimportant for inhibition of RNase BN activity by CCA-contain-ing RNAs, and they provide a mechanism that explains thisinhibition. Thus, we showed that the two adjacent C residues ofthe CCA sequence and Arg274 in the catalytic channel are bothnecessary for effective inhibition. Substitution of either of thetwo C residues with an A or U residue or mutation of Arg274 toalanine abolishes the discrimination of RNase BN against CCA-containing RNAs. Although substrate RNAs with a single Cresidue slightly retard the action of RNase BN, the overall effecton RNase BN is minimal. Based on this information, we pro-pose that there is a strong interaction between Arg274 in thecatalytic channel of RNase BN and the two C residues of theCCA sequence. This interaction prevents the movement ofCCA-containing RNA chains into the catalytic site (Fig. 6), andthis was shown directly by the finding that the phosphodiester-ase substrate, bis(p-nitrophenyl) phosphate was able to enterthe catalytic site and be hydrolyzed even when such an RNAwas present in the catalytic channel.Interestingly, in earlier work (21), it was suggested that a loop
structure located between strands �1 and �2 in the B. subtilisRNase Z was responsible for the protective effect of the CCAsequence. A highly conserved amino acid, Pro13, in that loopwas also suggested to play an important role in the discrimina-tion of cytosine residues from other bases. However, we couldnot obtain any evidence for involvement of this structure in the
FIGURE 7. Phosphodiesterase activity of wild type and R274A mutant RNase BN in the presence of RNA oligonucleotides. A, phosphodiesterase activityof wild type and R274A mutant RNase BN was measured as described under “Experimental Procedures” in the presence of the indicated concentrations ofmodel RNAs. B, phosphodiesterase activity of wild type RNase BN in the presence of model RNAs with different 3�-ends, used at the indicated concentrations.Phosphodiesterase activity in the absence of RNA was set at 100. The values shown are the averages of three independent experiments.
Mechanism of the CCA Inhibition of RNase BN
30642 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
at UN
IV O
F MIA
MI M
ILL
ER
SCH
OO
L O
F ME
DIC
INE
on Decem
ber 23, 2013http://w
ww
.jbc.org/D
ownloaded from

case of E. coli RNase BN. In fact, we have found that removal ofamino acid residues Ser9 to Arg17 in this loop has no effect onthe inability of RNase BN to remove a CCA sequence from aCCA-containing model RNA. Likewise, the loop mutant actssimilarly to wild type RNase BN on another model RNA sub-strate in removing five A residues following a CCA sequencebut then stopping (unpublished observations). Therefore, it isunlikely that either residues in the loop or Pro13 are involved inthe CCA inhibition of RNase BN. However, further work isneeded to clarify the apparent differences between the E. coliand B. subtilis enzymes.Arginine is one of the most prevalent amino acids present in
nucleic acid-binding domains of proteins. The reasons behindthis fact include the length of its side chain, its capacity to inter-act in different conformations, and its ability to generate favor-able hydrogen-bonding geometries (27). We attempted toexamine whether Lys could replace Arg in position 274. Unfor-tunately, the derivative was inactive (unpublished observation).Other amino acids that are not as efficient as arginine in inter-actingwith nucleic acids either lack long side chains or can onlyinteract in one configuration (27). The importance of Arg forinteraction with C residues in RNA molecules has beenobserved in several systems. In one example, arginine is part ofthe RNA-binding motif of an evolved PUF protein and is spe-cifically involved in recognition of cytosine residues (28, 29).Arginine also forms direct hydrogen bonds to cytosine duringthe selection of adenine over cytosine by CCA-adding enzymeat position 76 of tRNA (30, 31).One point that remains to be resolved is whether Arg274 is
the sole amino acid interacting with the two C residues of theCCA sequence or whether other amino acid residues might beinvolved. Clearly, removal of Arg274 is sufficient for eliminatingprotection by the CCA sequence, and the bidentate structure ofArg may enable it to interact with both C residues. In fact, suchbidentate interactions are seen in DNA-protein interaction(27). On the other hand, because mutations of other possibleinteracting amino acids, such as His270 and Ser272, led to loss ofactivity, it was not possible to assess their roles. Also some otherinteracting amino acid might not have been examined. Cocrys-tal structures of RNase BN and CCA-containing RNA will benecessary to settle this question.Mature tRNAs are resistant to digestion by RNases both in
vitro and in vivo. Their complex secondary structures, the pres-ence of an amino acid that protects the 3� terminus, and, asshown here, the universal 3�-CCA sequence of tRNA, all con-tribute to the stability of tRNAs. Members of the RNase Z fam-ily, which are responsible for 3�-end maturation of tRNA pre-cursors in many organisms, are important for survival in mostof these organisms. However, these enzymes had to evolve amechanism to distinguish mature, CCA-containing tRNAsfrom tRNA precursors that lack this sequence. As we haveshown, members of the RNase Z family contain a strictly con-served motif, HXSXR, near the catalytic site whose Arg residuein RNase BN is responsible for the lack of action on maturetRNAs. We presume that a similar mechanism operates inother members of the RNase Z family. In the absence of Arg274,RNase BN removes residues after the discriminator nucleotidein a process identical to the removal of 3�-extra residues from
CCA-less tRNA precursors by RNase Z. Thus, this family ofRNases has a built-in specificity that avoids indiscriminateaction on CCA-containing RNAs.However, this is not the only mechanism for avoiding action
on CCA-containing RNAs. Another RNase, termed RNase T,which is involved in the exonucleolytic 3�-end maturation ofE. coli tRNA precursors, is also inhibited by the presence of aCC sequence (32). In this case, the mechanism of inhibition iscompletely different. Four phenylalanine and one glutamic acidresidue of RNase T form a “C-filter” before the catalytic site ofRNase T that screens out C residues (7). This leads to inhibitionof RNase T on RNA substrates with C residues at their 3�-end,although the 3� terminal A residue can be removed (33). Itwould not be surprising if additional mechanisms have evolvedamong the many RNases present in cells to avoid action onmature tRNAs as distinguished from tRNA precursors. Thestudy presented here adds considerably to our knowledge of theRNase Z family of enzymes and of how they avoid acting onCCA-containing RNAs.
Acknowledgments—We thank Dr. T. K. Harris and members of ourlaboratory for critical comments on the manuscript.
REFERENCES1. Deutscher, M. P. (2006) Degradation of RNA in bacteria. Comparison of
mRNA and stable RNA. Nucleic Acids Res. 34, 659–6662. Andrade, J. M., Pobre, V., Silva, I. J., Domingues, S., and Arraiano, C. M.
(2009) The role of 3�-5� exoribonucleases in RNA degradation. Prog. Mol.Biol. Transl. Sci. 85, 187–229
3. Deutscher, M. P. (2003) Degradation of stable RNA in bacteria. J. Biol.Chem. 278, 45041–45044
4. Arraiano, C.M., Andrade, J.M., Domingues, S., Guinote, I. B.,Malecki,M.,Matos, R. G., Moreira, R. N., Pobre, V., Reis, F. P., Saramago,M., Silva, I. J.,and Viegas S. C. (2010) The critical role of RNA processing and degrada-tion in the control of gene expression. FEMSMicrobiol. Rev. 34, 883–923
5. Mackie, G. A., (2013) RNase E. At the interface of bacterial RNA process-ing and decay. Nat. Rev. Microbiol. 11, 45–57
6. Deutscher, M. P. (1988) The metabolic role of RNases. Trends Biochem.Sci. 13, 136–139
7. Hsiao, Y. Y., Yang, C. C., Lin, C. L., Lin, J. L., Duh, Y., andYuan,H. S. (2011)Structural basis for RNA trimming by RNase T in stable RNA 3� endmaturation. Nat. Chem. Biol. 7, 236–243
8. Asha, P. K., Blouin, R. T., Zaniewski, R., andDeutscher,M. P. (1983) RNaseBN. Identification and partial characterization of a new tRNA processingenzyme. Proc. Natl. Acad. Sci. 80, 3301–3304
9. Dutta, T., and Deutscher M. P. (2009) Catalytic Properties of RNase BN/RNase Z from Escherichia coli. RNase BN is both an exo- and an endori-bonuclease. J. Biol. Chem. 284, 15425–15431
10. Dutta, T., andDeutscher,M. P (2010)Mode of action of RNase BN/RNaseZ on tRNA precursors. RNase BN does not remove the CCA sequencefrom tRNA. J. Biol. Chem. 285, 22874–22881
11. Dutta, T., Malhotra, A., and Deutscher, M. P. (2012) Exoribonuclease andendoribonuclease activities of RNase BN both function in vivo. J. Biol.Chem. 287, 35747–35755
12. Ezraty, B., Dahlgren, B., and Deutscher, M. P. (2005) The RNase Z homo-logue encoded by Escherichia coli elaC gene is RNase BN. J. Biol. Chem.280, 16542–16545
13. Li, Z., and Deutscher, M. P. (1996) Maturation pathway for E. coli tRNAprecursors. A random multienzyme process in vivo. Cell 86, 503–512
14. Li, Z., andDeutscher,M. P. (1994) The role of individual exoribonucleasesin processing at the 3�-end of Escherichia coli tRNA precursors. J. Biol.Chem. 269, 6064–6071
15. Reuven, N. B., and Deutscher, M. P. (1993) Multiple exoribonucleases are
Mechanism of the CCA Inhibition of RNase BN
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30643
at UN
IV O
F MIA
MI M
ILL
ER
SCH
OO
L O
F ME
DIC
INE
on Decem
ber 23, 2013http://w
ww
.jbc.org/D
ownloaded from

required for the 3� processing of Escherichia coli tRNA precursors in vivo.FASEB J. 7, 143–148
16. Redko, Y., Li de la Sierra-Gallay, I., and Condon, C. (2007) When all’s zedand done. The structure and function of RNase Z in prokaryotes.Nat. Rev.Microbiol. 5, 278–286
17. Pellegrini, O., Nezzar, J., Marchfelder, A., Putzer, H., and Condon, C.(2003) Endonucleolytic processing of CCA-less tRNA precursors byRNase Z in Bacillus subtilis. EMBO J. 22, 4534–4543
18. Minagawa, A., Takaku, H., Takagi, M., and Nashimoto, M. (2004) A novelendonucleolytic mechanism to generate the CCA 3�termini of tRNAmol-ecules in Thermotoga maritima. J. Biol. Chem. 279, 15688–15697
19. Mohan, A., Whyte, S., Wang, X., Nashimoto, M., and Levinger, L. (1999)The 3�-end CCA of mature tRNA is an antideterminant for eukaryotic3�-tRNase. RNA. 5, 245–256
20. Kostelecky, B., Pohl, E., Vogel, A., Schilling, O., and Meyer-Klaucke, W.(2006) The crystal structure of the zinc phosphodiesterase from Esche-richia coli provides insight into function and cooperativity of tRNase Zfamily protein. J. Bacteriol. 188, 1607–1614
21. Li de la Sierra-Gallay, I., Pellegrini, O., and Condon, C. (2005) Structuralbasis for substrate binding cleavage and allostery in the tRNA maturaseRNase Z. Nature. 433, 657–661
22. Li de la Sierra-Gallay, I., Mathy, N., Pellegrini, O., and Condon, C. (2006)Structure of the ubiquitous 3� processing enzymeRNaseZ bound to trans-fer RNA. Nat. Struct. Mol. Biol. 13, 376–377
23. Pellegrini, O., Li de la Sierra-Gallay, I., Piton, J., Gilet, L., and Condon, C.(2012) Activation of tRNA maturation by downstream uracil residues inBacillus subtilis. Structure. 20, 1769–1777
24. Notredame, C., Higgins, D. G., and Heringa, J. (2000) T-Coffee. A novelmethod for fast and accurate multiple sequence alignment. J. Mol. Biol.
302, 205–21725. Di Tommaso, P., Moretti, S., Xenarios, I., Orobitg, M., Montanyola, A.,
Chang, J. M., Taly, J. F., andNotredame, C. (2011). T-Coffee. A web serverfor the multiple sequence alignment of protein and RNA sequences usingstructural information and homology extension. Nucleic Acids Res. 39,W13–W17
26. Gouet, P., Courcelle, E., Stuart, D. I., and Métoz, F. (1999) ESPript. Multi-ple sequence alignments in PostScript. Bioinformatics 15, 305–308
27. Luscombe, N. M., Laskowski, R. A., and Thornton, J. M. (2001) Aminoacid-base interactions. A three dimensional analysis of protein DNA in-teractions at an atomic level. Nucleic Acids Res. 29, 2860–2874
28. Filipovska, A., Razif, M. F., Nygård, K. K., and Rackham, O. (2011) Auniversal code for RNA recognition by PUF proteins. Nat. Chem. Biol. 7,425–427
29. Dong, S.,Wang, Y., Cassidy-Amstutz, C., Lu, G., Bigler, R., Jezyk,M. R., Li,C., Hall, T. M., and Wang, Z. (2011) Specific and molecular binding codefor cytosine recognition in Pumilio/FBF (PUF) RNA-binding domain.J. Biol. Chem. 286, 26732–26742
30. Xiong, Y., and Steitz, T. A. (2006) A story with a good ending. tRNA 3� endmaturation by CCA-adding enzymes. Curr. Opin. Struct. Biol. 16, 12–17
31. Pan, B., Xiong, Y., and Steitz, T. A. (2010) How the CCA-adding enzymeselects adenine over cytosine at position 76 of tRNA. Science. 330,937–940
32. Zuo, Y., andDeutscher,M. P. (2002) The physiological role of RNaseT canbe explained by its unusual substrate specificity. J. Biol. Chem. 277,29654–29661
33. Deutscher, M. P., Marlor, C. W., and Zaniewski, R. (1985) RNase T isresponsible for the end-turnover of tRNA in Escherichia coli. Proc. Natl.Acad. Sci. U.S.A. 82, 6427–6430
Mechanism of the CCA Inhibition of RNase BN
30644 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
at UN
IV O
F MIA
MI M
ILL
ER
SCH
OO
L O
F ME
DIC
INE
on Decem
ber 23, 2013http://w
ww
.jbc.org/D
ownloaded from





























