Embed Size (px)
Citation preview

R
H
Ba
b
c
a
ARRA
KGSRHACAA
C
1
ntfhappp
u
0d
Scientia Horticulturae 127 (2010) 119–126
Contents lists available at ScienceDirect
Scientia Horticulturae
journa l homepage: www.e lsev ier .com/ locate /sc ihor t i
eview
ormonal signaling in rootstock–scion interactions
. Alonia,∗, R. Cohenb, L. Karnia, H. Aktasc, M. Edelsteinb
Department of Vegetable Research, Institute of Plant Science, Agricultural Research Organization (A.R.O.), The Volcani Center, Bet-Dagan 50250, IsraelDepartment of Vegetable Research, Institute of Plant Science, Agricultural Research Organization, Newe Ya’ar Research Center, P.O. Box 1021, Ramat Yishay 30-095, IsraelSuleyman Demirel University, Agriculture Faculty, Horticulture Department, East Campus, 32260 Isparta, Turkey
r t i c l e i n f o
rticle history:eceived 1 March 2010eceived in revised form 2 September 2010ccepted 6 September 2010
eywords:
a b s t r a c t
The use of grafting technology in vegetable crops, represents a significant component of the vegetableindustries throughout the world. Most, if not all rootstocks and successful grafting combinations areselected on the basis of empirical testing. Obviously, there is a need for better understanding of theendogenous factors which control rootstock scion communication and processes which lead to the ben-eficial effects of grafting. The huge progress in understanding signaling processes and the involvementof phyto-hormones in all aspects of plant development and crop productivity may be utilized for more
raftingcionootstockormonal signalinguxinytokinins
profound probing into rootstock–scion communication in grafted plants. This review summarizes someof the research results in four aspects of hormonal signaling in rootstock–scion interactions: (1) for-mation of the rootstock–scion union; (2) rootstock–scion communication; (3) improvement of graftinginteractions by hormonal manipulations; (4) hormonal influence on growth, flowering, and fruit quality.
BAbiotic stress
© 2010 Elsevier B.V. All rights reserved.
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1192. Formation of the rootstock–scion union . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1203. Rootstock–scion communication . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1204. Improvement of grafting by hormonal manipulations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1215. Hormonal influence on growth, yield, flowering and fruit quality . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1246. Future prospects. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 125
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 125
. Introduction
Grafted vegetable transplants represent a significant compo-ent of the vegetable industries throughout the world. However,he use of grafted plants in the vegetable sector has not reached itsull potential mostly due to high pricing of the grafted seedlings,igh frequency of incompatibility occurrence between the scionnd the rootstock, especially at the late stages of fruit bearinglants, and lack of adequate information about the factors and the
mones as signaling and control factors, we can anticipate enhancedprogress in understanding grafting communication and thereforeimprovement of grafting technology and an increase in graftingimplementation.
In their last review on vegetable grafting Leonardi and Romano(2004) have presented the difficulties in forming an acceptable for-mula for defining grafting compatibility and success in vegetables.
rocesses that determine the communication between the graftedartners under diverse environmental conditions.
Nevertheless, due to the progress made in the recent years innderstanding signaling processes in plants and the impact of hor-
∗ Corresponding author.E-mail address: [email protected] (B. Aloni).
304-4238/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.scienta.2010.09.003
They have stated that vegetable grafting has been improved overthe last two decades, due to the implementation of new techniquesand materials. However, despite the recent advance, the percentageof grafted plants in the overall vegetable production is still rela-
tively low. The high cost of grafting, problems with soil-borne pestsand diseases control, and adaptation of the grafted seedlings to abi-otic stresses are the major constraints (Cohen et al., 2005, 2007).Although the importance of plant growth regulators for improv-ing the performance of grafted vegetable seedlings have long been
1 rticulturae 127 (2010) 119–126
rrh
pihf
2
obcotwsHfttitpdttfrtwblbnp
rYvopxs(cwc(wO1aicso1tg
gmr
Fig. 1. Transport rates of indol acetic acid (IAA) and zeatin riboside (ZR) in the xylemsap of ungrafted and grafted rootstocks of three Prunus genotypes. The invigoratingrootstocks change the IAA/ZR balance in favor of ZR thereby inducing higher shoot
20 B. Aloni et al. / Scientia Ho
ecognized, their implementation in the field has not been mate-ialized, mostly because the risks vs. benefits to the grafted cropsave not been defined yet.
In the following review we will present the work that has beenublished on the major phyto-hormones as it relates to graft-
ng signaling in conjunction with scion–rootstock compatibility,ormonal factors affecting rootstock contribution to the scion per-
ormance and rootstock–scion interactions under abiotic stresses.
. Formation of the rootstock–scion union
The success of grafting depends primarily on the identificationf stress and pathogen-resistant rootstocks, and on the compati-ility of the graft union in terms of fast formation of the vascularonnections between the rootstock and the scion and fast renewalf root and canopy growth (Cohen et al., 2007). In grafted plants,he vascular regeneration is reestablished by complex processes,hich include structural differentiation of the parenchymatous tis-
ue from both sides of the graft union into xylem and phloem tubes.artmann et al. (2002) overviewed the sequence of events in the
ormation of the graft union: “(1) Cut scion tissue capable of meris-ematic activity is brought into contact with similarly cut stockissue in a manner that the cambial regions of both are in prox-mity in order to interconnect through the callus bridge. Once thewo components of the graft, stock and scion, are in contact, newarenchymatous cells proliferate from both stock and scion, pro-ucing a callus tissue that intermingles and interlocks, filling uphe spaces between the two components connecting the scion andhe rootstock. (2) New cambial cells differentiate from the newlyormed callus, forming a continuous cambial connection betweenootstock and scion. Prior to the binding of vascular cambium acrosshe callus bridge, initial xylem and phloem are differentiated. Theound-repair xylem is generally the first differentiated tissue to
ridge the graft union, followed by wound-repair phloem. (3) In theast step of the graft establishment process, the newly formed cam-ial layer in the callus bridge begins typical cambial activity formingew vascular tissues. Production of new xylem and phloem thusermits the vascular connection between the scion and rootstock”.
For the majority of authors, this is considered as the basicequirement for a successful graft (Moore and Walker, 1981a,b;eoman, 1984; Tiedemann, 1989). In his review on regulation ofascular development, Dengler (2001) stated that vascular devel-pment includes: (1) formation of the longitudinal pattern ofrimary vascular strands; (2) formation of the radial pattern ofylem and phloem within vascular strands; (3) differentiation ofpecialized cell types from xylem and phloem precursors; and4) cell proliferation and cell differentiation within the vascularambium. Vascular regeneration experiments in which hormonesere applied exogenously to stem segments, indicate that low con-
entrations (0.1%, w/w, applied in lanolin) of indolic acetic acidIAA) stimulate phloem differentiation, whereas higher levels (1.0%,/w) induce xylem differentiation (Aloni, 1980, 1987, 1995, 2001).ther phytohormones, particularly cytokinins at a concentration of0 �g/ml, have been shown to induce vascular elements differenti-tion in wounded stems (Aloni et al., 1990; Aloni, 1993). Likewise,n grafting an important substance involved in the development ofompatible graft union is auxin, which is released from vasculartrands of the stock and the scion and induces the differentiationf vascular tissues, functioning as morphogenic substances (Aloni,987; Mattsson et al., 2003). Auxin translocation from the scion tohe rootstock was found to accelerate the formation of a successful
raft in Cactus (Shimomura and Fujihara, 1977).Collectively, the formation of vascular bridges across therafting zone is a primary need for grafting establishment butaintenance of the rootstock–scion communication is the second
equirement.
growth rates.
Adopted from Sorce et al. (2002).
3. Rootstock–scion communication
Plant hormones are important endogenous factors which regu-late all aspects of plant vegetative and reproductive developmentand thus are believed to be important player in root–shoot commu-nication. The “hormone message concept” dictates that hormonesare produced in one part of the plant and affect a remote part.According to that concept auxin which is produced in shoot apexesis translocated to the root where it affects root development, mor-phology and functioning. Auxin has also been shown to affect theproduction and activity of cytokinins which are known to be pro-duced in the root and translocated to the shoot where they controlimportant developmental processes such as shoot growth and pro-ductivity (Lockard and Schneider, 1981; Jones, 1986; Albacete etal., 2008). Bangerth (1994) has demonstrated that a feedback loopexists in which a decrease in basipetal flow of IAA from the shootstimulate the synthesis and export of cytokinins from the root. Therise of cytokinin concentrations in the xylem sap would increasethe synthesis and translocation of IAA out of the shoot apex whichin turn reduce cytokinin levels in the xylem sap. Working withpeach (Prunus persica L.) rootstocks, Sorce et al. (2002) have demon-strated that in non-grafted plants, auxin/cytokinin is well balanced.However, in grafted plants the auxin/cytokinin balance is upset,therefore the invigorating properties of the rootstocks induce ahigher growth rate in the scion, possibly by increasing the sup-ply of cytokinin to the shoot and decreasing that of IAA (Fig. 1).Van Hooijdonk et al. (2010) have recently shown that apple root-stock clones of different vigour could modify the architecture ofthe commercial cultivar, ‘Royal Gala’ scions following grafting. Thedwarfing (‘M.9’) rootstock significantly decreased the mean totalshoot length and node number of the scion compared with invigo-rating rootstocks. Similarly, N-1-naphthylphthalamic acid (NPA),an auxin transport inhibitor, applied to the stem of invigorat-ing rootstocks, decreased the total shoot growth of the scion andimposed architectural changes most closely resembled those of‘M.9’. In scions grafted 6-benzylaminopurine (BPA)-treated root-stocks or ‘M.9’, BAP re-instated the formation of secondary shoots,while application of GA4+7 reduced the proportion of primary and
secondary shoots that terminated growth early, thereby increasingtheir final length and node number. It is therefore proposed thatdwarfing rootstocks reduce the basipetal transport of IAA to theroot, thereby decreasing the amount of root-produced cytokininand gibberellin transported to the scion. At the scion, a limited
rticult
sb
(tnmBcbdnsettaaatcsozdat
bistatsgoatilt(scdttc(iNggoatcttttsiTi
B. Aloni et al. / Scientia Ho
upply of cytokinin may modify the architecture by decreasingranching.
Omid et al. (2007) have identified, in the phloem-sap of melonCucumis melo L.), transcripts which are associated with signalransduction and part of them were associated with auxin sig-aling. In this study it was found that numerous proteins andRNA molecules are present in the phloem translocation stream.
y genomic approach they found high proportion of genes asso-iated with biotic stimulus, response to stress, and metal–ioninding. Over 40% of the transcripts were related to stress andefense responses, while over 15% of them were related to sig-al transduction. In grafting of pumpkin (Cucurbita maxima Duch.)cion on melon rootstock they have established that 6 out of 43xamined transcripts in the phloem are capable of long-distancerafficking from melon stocks to pumpkin scions. Annotation ofhese six transcripts revealed that three were associated withuxin-signal transduction. The significance of these transcripts touxin signaling is still not known. Taking an interesting and uniquepproach, Kuroha et al. (2005) found that squash xylem sap con-ains cytokinins, which stimulates the elongation of tobacco BY-2ell protoplasts, grown in suspension system (Fig. 2). The xylemap also showed inhibitory activity of auxin induced elongationf excised cucumber hypocotyles, suggesting that the cytokinin,eatin riboside (ZR) and probably additional substances are pro-uced in the root and affect above ground organs by modulatinguxin activity. Such an approach would be most valuable for iden-ification rootstock derived hormones.
In vegetables, most grafting studies have been done on cucurbitsut only few have provided insight into the hormonal involvement
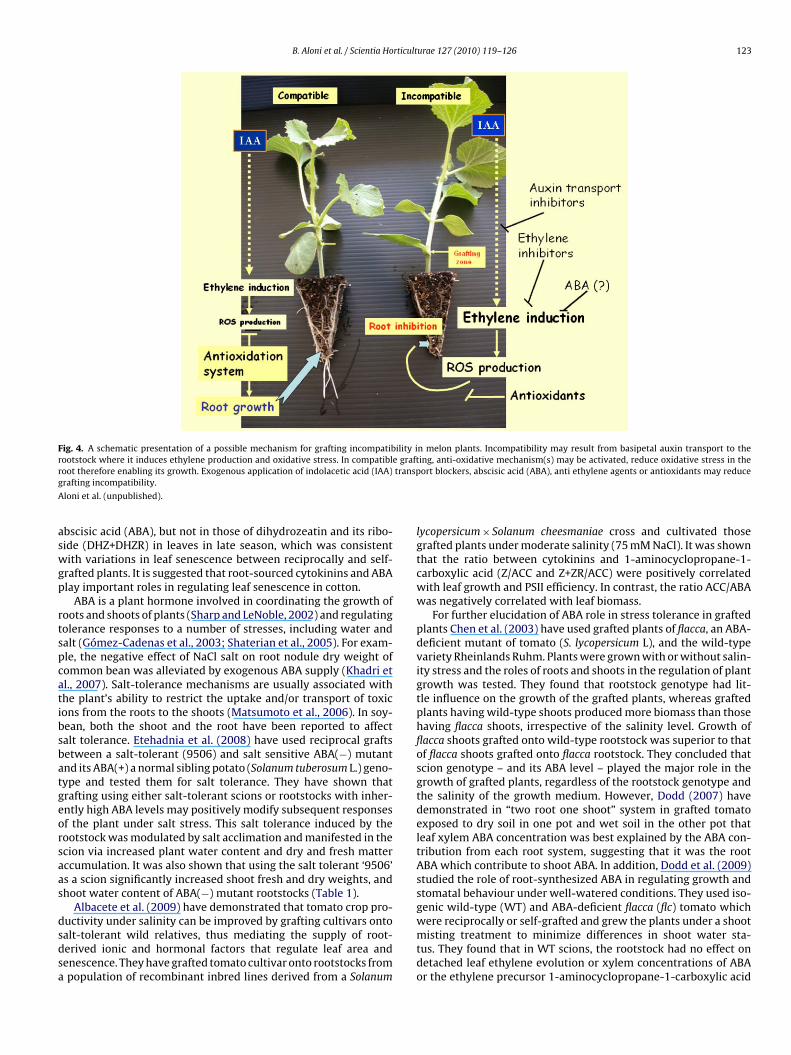
n grafting success. Melon (C. melo L.) can be grafted onto Cucurbitapp. The Cucurbita rootstock provides non-specific but efficient pro-ection from a wide range of soil-borne diseases and against somebiotic stresses. However, the compatibility between the scion andhe rootstock varies. Edelstein et al. (2004) have selected inter-pecific Cucurbita rootstocks with differential compatibilities whenrafted with melon. Aloni et al. (2008) found that the disruptionf rootstock–scion connections in incompatible grafting occurredpproximately 25 days after grafting. However, they proposed thathe main cause for incompatibility is the occurrence of hormonalmbalance, primarily, of auxin and ethylene in the root system fol-owing the establishment of the grafting connections. They showedhat exposure of the grafted transplants to high temperature regimeday/night, 32/28 ◦C), after grafting establishment, resulted in sub-tantial inhibition of root and shoot development and seedlingollapse of the incompatible grafted plants as compared to theevelopment of the compatible grafts. Morphologically, it appearedhat transplant collapse was initiated in the rootstock part whilehe scion part remained intact. Exogenous application of high con-entrations (≥10−7 M) of synthetic auxin [naphthalene acetic acidNAA)] to roots, caused inhibition of root growth. Root growth ofncompatible grafts was more affected than the compatible ones.AA was also applied directly to the root in hydroponically-grownrafted transplants which caused root decay in the incompatiblerafts and was less harmful to compatible ones. The involvementf auxin in the incompatibility mechanism emerged from threedditional observations: (A) endogenous IAA analysis revealed thathe roots and stem of incompatible seedlings contained higher IAAoncentrations than that tissues of compatible ones. (B) Applica-ion of the auxin transport inhibitor, 2,3,5-triiodobenzoic acid, onhe stem of the grafted transplants, negated root degradation inhe incompatible seedlings whereas it had only slight effect on
he compatible grafting combinations (Fig. 3) and (C) root andhoot development of incompatible grafts was normal after block-ng basipetal IAA transport by partial stem girdling in these plants.hese results support the notion that auxin, produced in the scion,s translocated downwards to the root after grafting connectionurae 127 (2010) 119–126 121
is established, and when reaching a threshold concentration, auxintriggers degradative processes, causing root decay. It was suggestedthat the incompatible pumpkin rootstock, is either more sensitiveto auxin or accumulates more auxin than the compatible root-stocks, therefore, the rate of root decay in this genotypes is high.
One of the possible mechanisms that trigger degradation pro-cesses in the rootstock might be oxidative stress activities which areknown to induce cell death symptoms in plant tissue (Yamamoto etal., 2003; Tamas et al., 2006). Reactive oxygen species (ROS) such assuperoxide radical (O2
•), hydrogen peroxide (H2O2) and hydroxylradical (•OH) are produced under normal conditions (Mittler, 2002;Foyer and Noctor, 2003). However, under abiotic and biotic stressesthey are produced in elevated concentrations and become harm-ful, resulting in lipid peroxidation, protein oxidation and damage tonucleic acids. Aloni et al. (2008) have shown that following appli-cation of high NAA concentration to the roots of grafted melonseedlings H2O2 concentration in the root increased most notablyin the incompatible grafted transplants. The compatible graftedseedlings accumulated significantly less H2O2 in their roots. On theother hand, the activities of peroxidase and SOD in the root sys-tem of compatible transplants were increased by NAA treatmentand to higher levels than in the root of incompatible ones. Over-all, this study supports the notion that incompatibility may resultfrom auxin-induced oxidative stress in the rootstock. In compatiblegrafting, anti-oxidative mechanism(s) may be activated, thereforereducing oxidative stress in the root and enabling its growth asproposed in Fig. 4.
Several reports suggest that root inhibition by high auxin con-centrations may be caused by auxin-induced ethylene production(Mulkey et al., 1982; Rahman et al., 2001). Aloni et al. (2008) haveshown that ethylene production in the root is increased several foldfollowing 3 days of maintenance in NAA. In addition, the roots ofincompatible grafted seedlings treated with NAA, produced moreethylene than the roots of compatible grafts. It was therefore pro-posed that in incompatible grafts, basipetally-transported auxinmay reach high concentration in the rootstock were it enhanceethylene production and subsequently causes inhibition of rootgrowth. In addition, ethylene may trigger ROS production whichmay lead to root degradation of these grafted transplants.
Ruzicka et al. (2007) proposed a model for ethylene dependentroot growth that accounts for auxin effects on root growth. Ethy-lene stimulates auxin biosynthesis in different plant organs viaits known signaling pathway. In addition, ethylene increases thecapacity of auxin transport by regulating the transcription of auxintransport components. The produced auxin is redistributed by polarauxin transport toward the root tip and, from there, through outercell layers of the root meristem into the elongation zone. Themajor components of auxin transport in these tissues, AUX1 andPIN2, mediate auxin delivery into cells of the elongation zone,where auxin accumulates and induces local auxin responses thatinhibit overall root growth. Whether such a model is applicable torootstock–scion relationship is worth investigating.
4. Improvement of grafting by hormonal manipulations
With the concept that hormones are involved in rootstock–scioninteractions, in many studies attempt has been made to use vari-ous plant growth regulators for grafting improvement. Köse andGüleryüz (2006) have tested the effect of cytokinin (Kinetin) andindole-3-butryic acid (IBA) by dipping the cut surface of both scion
and rootstock of grafted cuttings of four different grapevine (Vitisvinifera L.). It was shown that kinetin and benzyladenine (BA) stim-ulated rapid proliferation of callus between the scion and rootstockwhereas NAA and IBA increased root formation at the basal end ofgrafted cuttings compared with the control. Kinetin and BA showed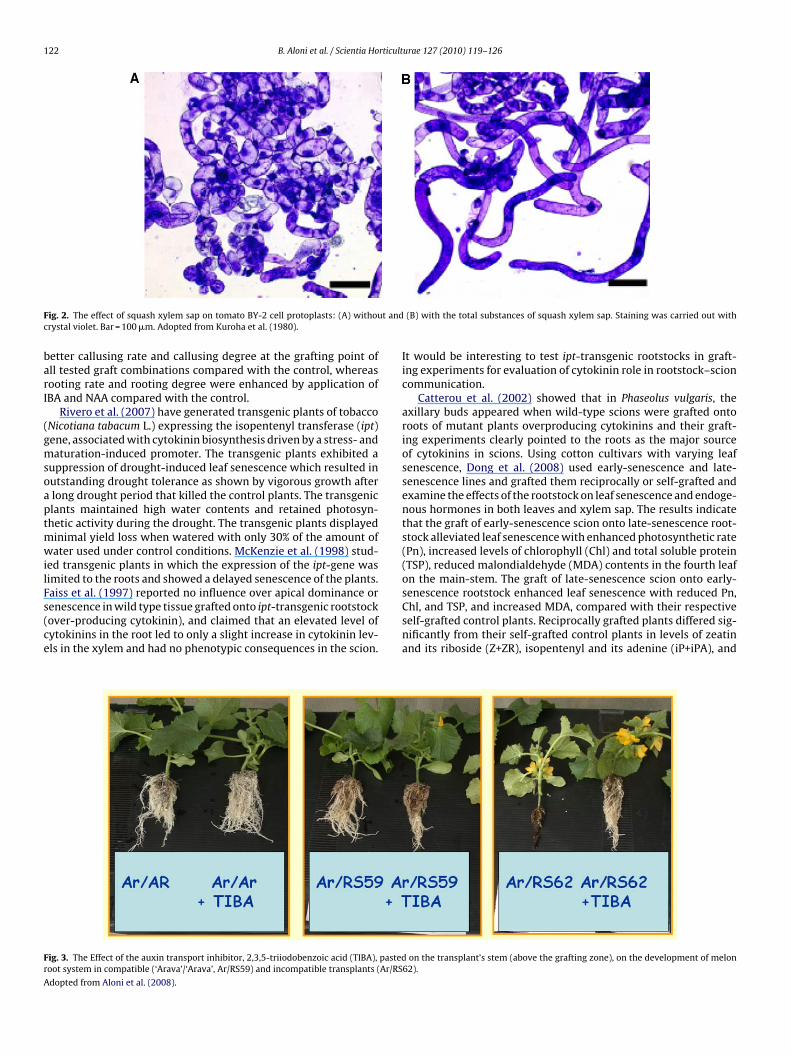
122 B. Aloni et al. / Scientia Horticulturae 127 (2010) 119–126
F t andc
barI
(gmsoaptmwilFs(ce
Fr
A
ig. 2. The effect of squash xylem sap on tomato BY-2 cell protoplasts: (A) withourystal violet. Bar = 100 �m. Adopted from Kuroha et al. (1980).
etter callusing rate and callusing degree at the grafting point ofll tested graft combinations compared with the control, whereasooting rate and rooting degree were enhanced by application ofBA and NAA compared with the control.
Rivero et al. (2007) have generated transgenic plants of tobaccoNicotiana tabacum L.) expressing the isopentenyl transferase (ipt)ene, associated with cytokinin biosynthesis driven by a stress- andaturation-induced promoter. The transgenic plants exhibited a
uppression of drought-induced leaf senescence which resulted inutstanding drought tolerance as shown by vigorous growth afterlong drought period that killed the control plants. The transgeniclants maintained high water contents and retained photosyn-hetic activity during the drought. The transgenic plants displayed
inimal yield loss when watered with only 30% of the amount ofater used under control conditions. McKenzie et al. (1998) stud-
ed transgenic plants in which the expression of the ipt-gene wasimited to the roots and showed a delayed senescence of the plants.aiss et al. (1997) reported no influence over apical dominance or
enescence in wild type tissue grafted onto ipt-transgenic rootstockover-producing cytokinin), and claimed that an elevated level ofytokinins in the root led to only a slight increase in cytokinin lev-ls in the xylem and had no phenotypic consequences in the scion.ig. 3. The Effect of the auxin transport inhibitor, 2,3,5-triiodobenzoic acid (TIBA), pastedoot system in compatible (‘Arava’/‘Arava’, Ar/RS59) and incompatible transplants (Ar/RS
dopted from Aloni et al. (2008).
(B) with the total substances of squash xylem sap. Staining was carried out with
It would be interesting to test ipt-transgenic rootstocks in graft-ing experiments for evaluation of cytokinin role in rootstock–scioncommunication.
Catterou et al. (2002) showed that in Phaseolus vulgaris, theaxillary buds appeared when wild-type scions were grafted ontoroots of mutant plants overproducing cytokinins and their graft-ing experiments clearly pointed to the roots as the major sourceof cytokinins in scions. Using cotton cultivars with varying leafsenescence, Dong et al. (2008) used early-senescence and late-senescence lines and grafted them reciprocally or self-grafted andexamine the effects of the rootstock on leaf senescence and endoge-nous hormones in both leaves and xylem sap. The results indicatethat the graft of early-senescence scion onto late-senescence root-stock alleviated leaf senescence with enhanced photosynthetic rate(Pn), increased levels of chlorophyll (Chl) and total soluble protein(TSP), reduced malondialdehyde (MDA) contents in the fourth leafon the main-stem. The graft of late-senescence scion onto early-senescence rootstock enhanced leaf senescence with reduced Pn,
Chl, and TSP, and increased MDA, compared with their respectiveself-grafted control plants. Reciprocally grafted plants differed sig-nificantly from their self-grafted control plants in levels of zeatinand its riboside (Z+ZR), isopentenyl and its adenine (iP+iPA), andon the transplant’s stem (above the grafting zone), on the development of melon62).
B. Aloni et al. / Scientia Horticulturae 127 (2010) 119–126 123
Fig. 4. A schematic presentation of a possible mechanism for grafting incompatibility in melon plants. Incompatibility may result from basipetal auxin transport to therootstock where it induces ethylene production and oxidative stress. In compatible grafting, anti-oxidative mechanism(s) may be activated, reduce oxidative stress in ther transpg
A
aswgp
rtspcatibsbatgeorsaas
dsdsa
oot therefore enabling its growth. Exogenous application of indolacetic acid (IAA)rafting incompatibility.
loni et al. (unpublished).
bscisic acid (ABA), but not in those of dihydrozeatin and its ribo-ide (DHZ+DHZR) in leaves in late season, which was consistentith variations in leaf senescence between reciprocally and self-
rafted plants. It is suggested that root-sourced cytokinins and ABAlay important roles in regulating leaf senescence in cotton.
ABA is a plant hormone involved in coordinating the growth ofoots and shoots of plants (Sharp and LeNoble, 2002) and regulatingolerance responses to a number of stresses, including water andalt (Gómez-Cadenas et al., 2003; Shaterian et al., 2005). For exam-le, the negative effect of NaCl salt on root nodule dry weight ofommon bean was alleviated by exogenous ABA supply (Khadri etl., 2007). Salt-tolerance mechanisms are usually associated withhe plant’s ability to restrict the uptake and/or transport of toxicons from the roots to the shoots (Matsumoto et al., 2006). In soy-ean, both the shoot and the root have been reported to affectalt tolerance. Etehadnia et al. (2008) have used reciprocal graftsetween a salt-tolerant (9506) and salt sensitive ABA(−) mutantnd its ABA(+) a normal sibling potato (Solanum tuberosum L.) geno-ype and tested them for salt tolerance. They have shown thatrafting using either salt-tolerant scions or rootstocks with inher-ntly high ABA levels may positively modify subsequent responsesf the plant under salt stress. This salt tolerance induced by theootstock was modulated by salt acclimation and manifested in thecion via increased plant water content and dry and fresh matterccumulation. It was also shown that using the salt tolerant ‘9506’s a scion significantly increased shoot fresh and dry weights, andhoot water content of ABA(−) mutant rootstocks (Table 1).
Albacete et al. (2009) have demonstrated that tomato crop pro-
uctivity under salinity can be improved by grafting cultivars ontoalt-tolerant wild relatives, thus mediating the supply of root-erived ionic and hormonal factors that regulate leaf area andenescence. They have grafted tomato cultivar onto rootstocks frompopulation of recombinant inbred lines derived from a Solanumort blockers, abscisic acid (ABA), anti ethylene agents or antioxidants may reduce
lycopersicum × Solanum cheesmaniae cross and cultivated thosegrafted plants under moderate salinity (75 mM NaCl). It was shownthat the ratio between cytokinins and 1-aminocyclopropane-1-carboxylic acid (Z/ACC and Z+ZR/ACC) were positively correlatedwith leaf growth and PSII efficiency. In contrast, the ratio ACC/ABAwas negatively correlated with leaf biomass.
For further elucidation of ABA role in stress tolerance in graftedplants Chen et al. (2003) have used grafted plants of flacca, an ABA-deficient mutant of tomato (S. lycopersicum L), and the wild-typevariety Rheinlands Ruhm. Plants were grown with or without salin-ity stress and the roles of roots and shoots in the regulation of plantgrowth was tested. They found that rootstock genotype had lit-tle influence on the growth of the grafted plants, whereas graftedplants having wild-type shoots produced more biomass than thosehaving flacca shoots, irrespective of the salinity level. Growth offlacca shoots grafted onto wild-type rootstock was superior to thatof flacca shoots grafted onto flacca rootstock. They concluded thatscion genotype – and its ABA level – played the major role in thegrowth of grafted plants, regardless of the rootstock genotype andthe salinity of the growth medium. However, Dodd (2007) havedemonstrated in “two root one shoot” system in grafted tomatoexposed to dry soil in one pot and wet soil in the other pot thatleaf xylem ABA concentration was best explained by the ABA con-tribution from each root system, suggesting that it was the rootABA which contribute to shoot ABA. In addition, Dodd et al. (2009)studied the role of root-synthesized ABA in regulating growth andstomatal behaviour under well-watered conditions. They used iso-genic wild-type (WT) and ABA-deficient flacca (flc) tomato which
were reciprocally or self-grafted and grew the plants under a shootmisting treatment to minimize differences in shoot water sta-tus. They found that in WT scions, the rootstock had no effect ondetached leaf ethylene evolution or xylem concentrations of ABAor the ethylene precursor 1-aminocyclopropane-1-carboxylic acid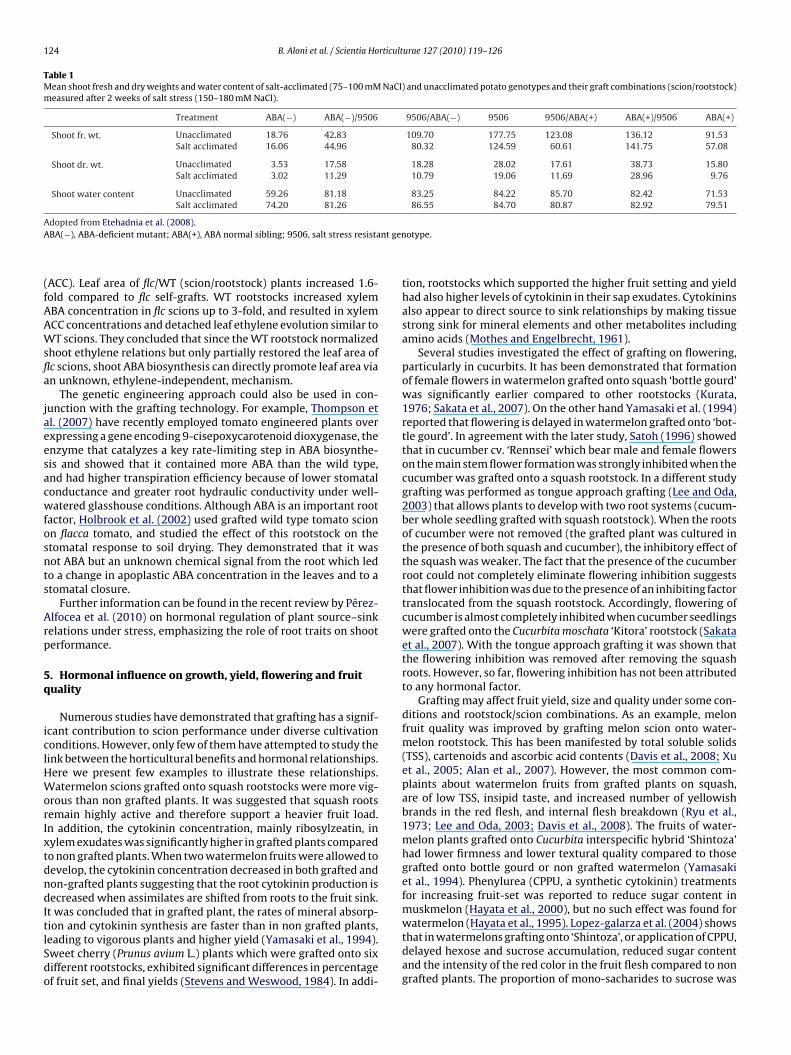
124 B. Aloni et al. / Scientia Horticulturae 127 (2010) 119–126
Table 1Mean shoot fresh and dry weights and water content of salt-acclimated (75–100 mM NaCl) and unacclimated potato genotypes and their graft combinations (scion/rootstock)measured after 2 weeks of salt stress (150–180 mM NaCl).
Treatment ABA(−) ABA(−)/9506 9506/ABA(−) 9506 9506/ABA(+) ABA(+)/9506 ABA(+)
Shoot fr. wt. Unacclimated 18.76 42.83 109.70 177.75 123.08 136.12 91.53Salt acclimated 16.06 44.96 80.32 124.59 60.61 141.75 57.08
Shoot dr. wt. Unacclimated 3.53 17.58 18.28 28.02 17.61 38.73 15.80Salt acclimated 3.02 11.29 10.79 19.06 11.69 28.96 9.76
Shoot water content Unacclimated 59.26 81.18 83.25 84.22 85.70 82.42 71.53Salt acclimated 74.20 81.26 86.55 84.70 80.87 82.92 79.51
AA nt gen
(fAAWsfla
jaeesacwfosnts
Arp
5q
iclHWorIxtdndItlSdo
dopted from Etehadnia et al. (2008).BA(−), ABA-deficient mutant; ABA(+), ABA normal sibling; 9506, salt stress resista
ACC). Leaf area of flc/WT (scion/rootstock) plants increased 1.6-old compared to flc self-grafts. WT rootstocks increased xylemBA concentration in flc scions up to 3-fold, and resulted in xylemCC concentrations and detached leaf ethylene evolution similar toT scions. They concluded that since the WT rootstock normalized
hoot ethylene relations but only partially restored the leaf area ofc scions, shoot ABA biosynthesis can directly promote leaf area vian unknown, ethylene-independent, mechanism.
The genetic engineering approach could also be used in con-unction with the grafting technology. For example, Thompson etl. (2007) have recently employed tomato engineered plants overxpressing a gene encoding 9-cisepoxycarotenoid dioxygenase, thenzyme that catalyzes a key rate-limiting step in ABA biosynthe-is and showed that it contained more ABA than the wild type,nd had higher transpiration efficiency because of lower stomatalonductance and greater root hydraulic conductivity under well-atered glasshouse conditions. Although ABA is an important root
actor, Holbrook et al. (2002) used grafted wild type tomato scionn flacca tomato, and studied the effect of this rootstock on thetomatal response to soil drying. They demonstrated that it wasot ABA but an unknown chemical signal from the root which ledo a change in apoplastic ABA concentration in the leaves and to atomatal closure.
Further information can be found in the recent review by Pérez-lfocea et al. (2010) on hormonal regulation of plant source–sinkelations under stress, emphasizing the role of root traits on shooterformance.
. Hormonal influence on growth, yield, flowering and fruituality
Numerous studies have demonstrated that grafting has a signif-cant contribution to scion performance under diverse cultivationonditions. However, only few of them have attempted to study theink between the horticultural benefits and hormonal relationships.ere we present few examples to illustrate these relationships.atermelon scions grafted onto squash rootstocks were more vig-
rous than non grafted plants. It was suggested that squash rootsemain highly active and therefore support a heavier fruit load.n addition, the cytokinin concentration, mainly ribosylzeatin, inylem exudates was significantly higher in grafted plants comparedo non grafted plants. When two watermelon fruits were allowed toevelop, the cytokinin concentration decreased in both grafted andon-grafted plants suggesting that the root cytokinin production isecreased when assimilates are shifted from roots to the fruit sink.
t was concluded that in grafted plant, the rates of mineral absorp-
ion and cytokinin synthesis are faster than in non grafted plants,eading to vigorous plants and higher yield (Yamasaki et al., 1994).weet cherry (Prunus avium L.) plants which were grafted onto sixifferent rootstocks, exhibited significant differences in percentagef fruit set, and final yields (Stevens and Weswood, 1984). In addi-otype.
tion, rootstocks which supported the higher fruit setting and yieldhad also higher levels of cytokinin in their sap exudates. Cytokininsalso appear to direct source to sink relationships by making tissuestrong sink for mineral elements and other metabolites includingamino acids (Mothes and Engelbrecht, 1961).
Several studies investigated the effect of grafting on flowering,particularly in cucurbits. It has been demonstrated that formationof female flowers in watermelon grafted onto squash ‘bottle gourd’was significantly earlier compared to other rootstocks (Kurata,1976; Sakata et al., 2007). On the other hand Yamasaki et al. (1994)reported that flowering is delayed in watermelon grafted onto ‘bot-tle gourd’. In agreement with the later study, Satoh (1996) showedthat in cucumber cv. ‘Rennsei’ which bear male and female flowerson the main stem flower formation was strongly inhibited when thecucumber was grafted onto a squash rootstock. In a different studygrafting was performed as tongue approach grafting (Lee and Oda,2003) that allows plants to develop with two root systems (cucum-ber whole seedling grafted with squash rootstock). When the rootsof cucumber were not removed (the grafted plant was cultured inthe presence of both squash and cucumber), the inhibitory effect ofthe squash was weaker. The fact that the presence of the cucumberroot could not completely eliminate flowering inhibition suggeststhat flower inhibition was due to the presence of an inhibiting factortranslocated from the squash rootstock. Accordingly, flowering ofcucumber is almost completely inhibited when cucumber seedlingswere grafted onto the Cucurbita moschata ‘Kitora’ rootstock (Sakataet al., 2007). With the tongue approach grafting it was shown thatthe flowering inhibition was removed after removing the squashroots. However, so far, flowering inhibition has not been attributedto any hormonal factor.
Grafting may affect fruit yield, size and quality under some con-ditions and rootstock/scion combinations. As an example, melonfruit quality was improved by grafting melon scion onto water-melon rootstock. This has been manifested by total soluble solids(TSS), cartenoids and ascorbic acid contents (Davis et al., 2008; Xuet al., 2005; Alan et al., 2007). However, the most common com-plaints about watermelon fruits from grafted plants on squash,are of low TSS, insipid taste, and increased number of yellowishbrands in the red flesh, and internal flesh breakdown (Ryu et al.,1973; Lee and Oda, 2003; Davis et al., 2008). The fruits of water-melon plants grafted onto Cucurbita interspecific hybrid ‘Shintoza’had lower firmness and lower textural quality compared to thosegrafted onto bottle gourd or non grafted watermelon (Yamasakiet al., 1994). Phenylurea (CPPU, a synthetic cytokinin) treatmentsfor increasing fruit-set was reported to reduce sugar content inmuskmelon (Hayata et al., 2000), but no such effect was found for
watermelon (Hayata et al., 1995). Lopez-galarza et al. (2004) showsthat in watermelons grafting onto ‘Shintoza’, or application of CPPU,delayed hexose and sucrose accumulation, reduced sugar contentand the intensity of the red color in the fruit flesh compared to nongrafted plants. The proportion of mono-sacharides to sucrose was
rticult
afltfifi
6
fstrstptm
smatbosrnTlctsssghvittaragc
dYfsig
nsgAoph
ft
B. Aloni et al. / Scientia Ho
lso increased, and this may be associated with alteration of fruitavor. It is likely the squash rootstock produces some cytokininshat are transported to the scion and induce a negative effect onruit quality. However, no reduction in fruit quality has been foundn an experiment conducted with the same cultivars in the openeld (Miguel, 2004).
. Future prospects
The full understanding of hormonal control of grafting success isar from being complete. As demonstrated above, different resultsometime, even contradictory, are published, reflecting probably,he variation in experimental setups and the origin of plant mate-ial. Hormonal signaling may have different impacts on differenttages of the establishment of grafting communication. Needlesso say that nowadays the research tools for investigating basicrocesses in rootstock–scion communication are available andherefore pinpointing the important limiting factors which deter-
ine the obtainment of successful grafting in any crop is possible.Great benefits can stem from the ability to graft Arabidopsis
eedlings (Turnbull et al., 2002). The ability to graft such seedlingsay facilitate experiments to investigate long-distance signaling
t almost any stage of development. Single grafts enable simpleests for shoot-to-root and root-to-shoot signaling, which haveeen implicated in a number of processes, such as co-ordinationf nutrition deficiencies, regulation of hormonal transport andtress responses. Grafting of Arabidopsis mutants should enableapid assignment of genes for regulation of transmissible sig-als involved in these developmental and physiological processes.here is the potential for detection of other, currently unrecognized,ong-distance signals with roles in developmental and physiologi-al processes. For this purpose, similar approach can also be appliedo crop species. For example, using tomato interspecific introgres-ion lines (Fridman et al., 2004), with increasingly defined genomicequences, as rootstocks. These tomato lines contain chromosomeegments of wild species (Solanum pennellii Corell.) in the back-round of the cultivated tomato (S. lycopersicum L.). Fita et al. (2008)ave recently identified quantitative trait loci (QTLs) controllingariation in melon root growth and architecture by using near-sogenic lines (NILs), constructed by introgressing the genome ofhe exotic Korean accession Shongwan Charmi [SC (PI161375)] intohe genetic background of commercial cultivar. Similarly, Asins etl. (2010) have recently demonstrated the existence of QTLs in theootstocks of wild tomato which partially contributed to salt toler-nce of salt sensitive genotype. Such genetic sources can be used inrafting experiments for identification of genomic sequences whichontribute to grafting success.
Gene transformation has been recently used in developingrought and salinity stress-tolerant genotypes (Rivero et al., 2007;amaguchi and Blumwald, 2005). Transferring desirable hormonal
actors from the root of salt-resistant rootstocks to the shoot of saltensitive scions through gene transcription in the root and actionn the shoot is than an alternative way of overcoming salinity byrafting.
Plant growth regulators (PGR) either natural or synthetic areot in common use in practical propagation of grafted vegetableeedlings. However, there is a good prospect of utilizing PGR inrafting technology, especially the use of auxins, ethylene-blockers,BA or its analogues (which are currently available), gibberellinsr compounds with anti-oxidation properties (Johkan et al., 2008)
rovided that the temporal and concentration specificity of theseormones will be defined.Finally, the often observations of negative impact of grafting onruit quality and its association with unbalanced hormonal rela-ions also deserve profound investigation.
urae 127 (2010) 119–126 125
References
Alan, O., Zdemir, N., Nen, Y., 2007. Effect of grafting on watermelon plant growth,yield and quality. J. Agron. 6, 362–365.
Albacete, A., Ghanem, M.E., Martínez-Andújar, C., Acosta, M., Sánchez-Bravo, J.,Martínez, V., Lutts, S., Dodd, I.C., Pérez-Alfocea, F., 2008. Hormonal changesin relation to biomass partitioning and shoot growth impairment in salinizedtomato. J. Exp. Bot. 59, 4119–4131.
Albacete, A., Martínez-andújar, C., Ghanem, M.E., Acosta, M., Sánchez-bravo, J., Asins,M.J., Cuartero, J., Lutts, S., Dodd, I.C., Pérez-Alfocea, F., 2009. Rootstock-mediatedchanges in xylem ionic and hormonal status are correlated with delayed leafsenescence, and increased leaf area and crop productivity in salinized tomato.Plant Cell Environ. 32, 928–938.
Aloni, B., Karni, L., Deveturero, G., Levin, Z., Cohen, R., Kazir, N., Lotan-Pompan,M., Edelstein, M., Aktas, H., Turhan, E., Joel, D.M., Horev, C., Kapulnic, Y., 2008.Physiological and biochemical changes at the rootstock–scion interface in graftcombinations between Cucurbita rootstocks and a melon scion. J. Hortic. Sci.Biotechnol. 83, 777–783.
Aloni, R., 1980. Role of auxin and sucrose in the differentiation of sieve and trachearyelements in plant tissue cultures. Planta 150, 255–263.
Aloni, R., 1987. Differentiation of vascular tissues. Ann. Rev. Plant Physiol. 38,179–204.
Aloni, R., 1995. The induction of vascular tissues by auxin and cytokinin. In:Davies, P.J. (Ed.), Plant Hormones. Kluwer Academic Publishers, Netherlands,pp. 531–546.
Aloni, R., 2001. Foliar and axial aspects of vascular differentiation–hypotheses andevidence. J. Plant Growth. Regul. 20, 22–34.
Aloni, R., 1993. The role of cytokinin in organized differentiation of vascular tissues.Aust. J. Plant Physiol. 20, 601–608.
Aloni, R., Baum, S.F., Peterson, C.A., 1990. The role of cytokinin in sieve tube regen-eration and callose production in wounded Coleus internodes. Plant Physiol. 93,982–989.
Asins, M.J., Bolarín, M.C., Pérez-Alfocea, F., Estan, M.T., Martínez-Andújar, C.,Albacete, A., Villalta, I., Bernet, G.P., Dodd, I.C., Carbonell, E.A., 2010. Geneticanalysis of physiological components of salt tolerance conferred by Solanumrootstocks. What is the rootstock doing for the scion? Theor. Appl. Genet. 121,105–115.
Bangerth, F., 1994. Response of cytokinin concentration in the xylem exudate ofbean (Phaseolus vulgaris L.) plants to decapitation and auxin treatment, andrelationship to apical dominance. Planta 194, 439–442.
Catterou, M., Dubois, F., Smets, R., Vaniet, S., Kichey, T., Van Onckelen, H., Sangwan-Norreel, B.S., Sangwan, R.S., 2002. Hoc: an Arabidopsis mutant overproducingcytokinins and expressing high in vitro organogenic capacity. Plant J. 30,273–287.
Chen, G., Fu, X., Lips, H., Sagi, M., 2003. Control of plant growth resides in the shoot,and not in the root, in reciprocal grafts of flacca and wild-type tomato (Lysoper-sicon esculentum), in the presence and absence of salinity stress. Plant Soil 256,205–215.
Cohen, R., Burger, Y., Horev, C., Porat, A., Edelstein, M., 2005. Performance of Galia-type melons grafted on to Cucurbita rootstock in Monosporascus cannonballus-infested and non-infested soils. Ann. Appl. Biol. 146, 381–387.
Cohen, R., Burger, Y., Koern, A., Edelstein, M., 2007. Introducing grafted cucurbitstomodern agriculture; The Israeli experience. Plant Dis. 91, 916–923.
Davis, A.R., Perkins-Veazie, P., Levi, A., King, S.R., CohenZhang, X., 2008. Graftingeffects on vegetable quality. HortScience 43, 1670–1672.
Dengler, N.G., 2001. Regulation of Vascular Development. J. Plant Growth Regul. 20,1–13.
Dodd, I.C., 2007. Soil moisture heterogeneity during deficit irrigation alters root-to-shoot signalling of abscisic acid. Funct. Plant Biol. 34, 439–448.
Dodd, I.C., Theobald, J.C., Richer, S.K., Davies, W.J., 2009. Partial phenotypic reversionof ABA-deficient flacca tomato (Solanum lycopersicum) scions by a wild-typerootstock: normalizing shoot ethylene relations promotes leaf area but doesnot diminish whole plant transpiration rate. J. Exp. Bot. 60, 4029–4039.
Dong, H., Niu, Y., Liand, W., Zhang, D., 2008. Effects of cotton rootstock on endoge-nous cytokinins and abscisic acid in xylem sap and leaves in relation to leafSenescence. J. Exp. Bot. 59, 1295–1304.
Edelstein, M., Burger, Y., Horev, C., Porat, A., Meir, A., Cohen, R., 2004. Assessing theeffect of genetic and anatomic variation of Cucurbita rootstocks on vigor, survivaland yield of grafted melons. J. Hortic. Sci. Biotechnol. 79, 370–374.
Etehadnia, M., Waterer, D., De Jong, H., Tanino, K.K., 2008. Scion and rootstock effectson ABA-mediated plant growth regulation and salt tolerance of acclimated andunacclimated potato genotypes. J. Plant Growth Regul. 27, 125–140.
Faiss, M., Zalubı‘lova, J., Strnad, M., Schmulling, T., 1997. Conditional transgenicexpression of the ipt gene indicates a function for cytokinins in paracrine sig-naling in whole tobacco plants. Plant J. 12, 401–415.
Fita, A., Picó, B., Monforte, A.J., Nuez, F., 2008. Genetics of root system architectureusing near-isogenic lines of melon. J. Am. Soc. Hortic. Sci. 133, 448–458.
Foyer, C.H., Noctor, G., 2003. Redox sensing and signaling associated in plants: a re-evaluation of the concept of oxidative stress response in leaves. J. Exp. Bot. 53,1249–1254.
Fridman, E., Carrari, F., Liu, Y., Fernie, A.R., Zamir, D., 2004. Zooming in on a quan-titative trait for tomato yield using interspecific introgressions. Science 305,1786–1789.
Gómez-Cadenas, A., Arbona, V., Jacas, J., Primo-Millo, E., Talon, M., 2003. Abscisicacid reduces leaf abscission and increases salt tolerance in citrus plants. J. PlantGrowth Regul. 21, 234–240.

1 rticult
H
H
H
H
J
J
K
K
K
K
L
L
L
L
M
M
M
M
M
M
M
M
M
Yamasaki, A., Yamashita, M., Furuya, S., 1994. Mineral concentrations and cytokinin
26 B. Aloni et al. / Scientia Ho
ayata, Y., Nimi, Y., Iwasaki, N., 1995. Synthetic cytokinin 1-(2-chloro-4-pyridyl)-3-phenylurea (CPPU) promotes fruit setand induced parthenocarpy inwatermelon. J. Am. Soc. Hortic. Sci. 120, 997–1000.
ayata, Y., Nimi, Y., Inoue, K., Kondo, S., 2000. CPPU and BA with and without pol-lination affect set, growth, and quality of muskmelon fruit. HortScience 35,868–870.
artmann, H.T., Kester, D.E., Davies, F.T., Geneve, R.L., 2002. Plant Propagation. Prin-ciples and Practices, seventh ed. Prentice Hall, Upper Saddle River, NJ, ISBN0-13-679235-9, pp. 849.
olbrook, N.M., Shashidhar, V.R., James, R.A., Munns, R., 2002. Stomatal control intomato with ABA-deficient roots: response of grafted plants to soil drying. J. Exp.Bot. 53, 1503–1514.
ohkan, M., Oda, M., Mori, G., 2008. Ascorbic acid promotes graft-take in sweetpepper plants (Capsicum annuum L.). Sci. Hortic. 116, 343–347.
ones, O.P., 1986. Endogenous growth regulators and rootstock/scion interactions inapple and cherry trees. Acta Hortic. 179, 177–184.
hadri, M., Tejera, N.A., Lluch, C., 2007. Sodium chloride-ABA interaction in two com-mon bean (Phaseolus vulgaris) cultivars differing in salinity tolerance. Environ.Exp. Bot. 60, 211–218.
öse, C., Güleryüz, M., 2006. Effects of auxins and cytokinins on graft union ofgrapevine (Vitis vinifera) New Zealand. J. Crop Hortic. Sci. 34, 145–150.
urata, H., 1976. Studies on the sex expression of flowering by day-length andtemperature in pumpkin and watermelon. Mem. Fac. Agric., Kagawa Univ. 29,1–49.
uroha, T., Sakurai, M., Satoh, S., 2005. Squash xylem sap has activities that inhibitproliferation and promote the elongation of tobacco BY-2 cell protoplasts. PlantPhysiol. Biochem. 43, 465–471.
ee, J.M., Oda, M., 2003. Grafting of herbaceous vegetable and ornamental crops.Hortic. Rev. 28, 61–124.
eonardi, C., Romano, D., 2004. Recent issues on vegetable grafting. Acta Hortic. 631,163–174.
ockard, R.G., Schneider, G.W., 1981. Stock and scion growth relationships and thedwarfing mechanism in apple. Hortic. Rev. 3, 315–375.
opez-galarza, S., San Bautista, A., Perez, D.M., Miguel, A., Baixauli, C., Pascual, B.,Maroto, J.V., Guardiola, J.L., 2004. Effects of grafting and cytokinin-induced fruitsetting on colour and sugar-content traits in glasshouse-grown triploid water-melon. J. Hortic. Sci. Biotechnol. 79, 971–976.
attsson, J., Ckurshumova, W., Berleth, T., 2003. Auxin signaling in Arabidopsis leafvascular development. Plant Physiol. 131, 1327–1339.
atsumoto, K., Tamura, F., Chun, J.P., Tanabe, K., 2006. Native Mediterranean Pyrusrootstock, P. amygdaliformis and P. elaeagrifolia, present higher tolerance to salin-ity stress compared with Asian natives. J. Jpn. Soc. Hortic. Sci. 75, 450–457.
cKenzie, M.J., Mett, V., Reynolds, P.H.S., Jameson, P.E., 1998. Controlled cytokininproduction in transgenic tobacco using a copper-inducible promoter. Plant Phys-iol. 116, 969–977.
iguel, A., 2004. Use of grafted cucurbitsin the Mediteranean region as an alternativeto methyl bromide. In: Proc. Fifth International Conference on Alternatives toMethyl Bromide, Lisbon, pp. 151–156.
ittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends PlantSci. 7, 405–410.
oore, R., Walker, D.B., 1981a. Studies on vegetative compatibility-incompatibilityin higher plants. I. A structural study of a compatible autograph in Sedum tele-phoides (Crassulaceae). Am. J. Bot. 68, 820–830.
oore, R., Walker, D.B., 1981b. Studies on vegetative compatibility-incompatibilityin higher plants. II. A structural study of an incompatible heterograft betweenSedum telephoides (Crassulaceae) and Solanum pennelli (Solanaceae). Am. J. Bot.68, 831–842.
othes, K., Engelbrecht, L., 1961. Kinetin-induced directed transport of substancesin excised levels in the dark. Phytochemistry 1, 58–62.
ulkey, T.J., Kuzmanoff, K.M., Evans, M.L., 1982. Promotion of growth and shift inthe auxin dose/response relationship in maize roots treated with the ethylenebiosynthesis inhibitors aminoethoxyvinylglycine and cobalt. Plant Sci. Lett. 25,43–48.
urae 127 (2010) 119–126
Omid, A., Keilin, T., Glass, A., Leshkowitz, D., Wolf, S., 2007. Characterization ofphloem-sap transcription profile in melon plants. J. Exp. Bot. 58, 3645–3656.
Pérez-Alfocea, F., Albacete, A., Ghanem, M.E., Dodd, I.C., 2010. Hormonal regulationof source–sink relations to maintain crop productivity under salinity: a casestudy of root-to-shoot signalling in tomato. Funct. Plant Biol. 37, 592–603.
Rahman, A., Amakawa, T., Goto, N., Tsurumi, S., 2001. Auxin is a positive regulatorof ethylene-mediated response in the growth of Arabidopsis roots. Plant CellPhysiol. 42, 301–307.
Rivero, R.M., Kojima, M., Gepstein, A., Sakakibara, H., Mittler, R., Gepstein, S.,Blumwald, E., 2007. Delayed leaf senescence induces extreme drought tolerancein a flowering plant. Proc. Natl. Acad. Sci. U.S.A. 104, 19631–19636.
Ruzicka, K., Ljung, K., Vanneste, S., Podhorska, R., Beeckman, T., Friml, J., Benkova, E.,2007. Ethylene regulates root growth through effects on auxin biosynthesis andtransport-dependent auxin distribution. Plant Cell 19, 2197–2212.
Ryu, J.S., Choi, K.S., Lee, S.S., 1973. Effect of grafting stock on growth, quality, andyield of watermelon. J. Kor. Soc. Hortic. Sci. 13, 45–49.
Sakata, Y., Takayoshi, O., Mitsuhiro, S., 2007. The history and present state of thegrafting of cucurbitaceous in Japan. Acta Hortic. 731, 159–170.
Satoh, S., 1996. Inhibition of flowering of cucumber grafted on rooted squash stocks.Physiol. Plant. 97, 440–444.
Sharp, R.E., LeNoble, M.E., 2002. ABA, ethylene and the control of shoot and rootgrowth under water stress. J. Exp. Bot. 53, 33–37.
Shaterian, J., Waterer, D., De Jong, H., Tanino, K.K., 2005. Differential stress responsesto NaCl salt application in early- and late maturing diploid potato (Solanum sp.)clones. Environ. Exp. Bot. 54, 202–212.
Shimomura, T., Fujihara, K., 1977. Physiological study of graft union formation inCactus. II. Role of auxin on vascular connection between stock and scion I. Jpn.Soc. Hortic. Sci. 45, 397–406.
Sorce, C., Massai, R., Picciarelli, P., Lorenzi, R., 2002. Hormonal relationships inxylem sap of grafted and ungrafted Prunus rootstock. Sci. Hortic. 93, 333–342.
Stevens, G., Weswood, M.N., 1984. Fruit set and cytokinin-like activity in the xylemsap of sweet cherry (Prunus avium) as affected by rootstock. Physiol. Plant. 61,464–468.
Tamas, L., Huttova, J., Mistrik, I., Simonovicova, M., Siroka, B., 2006. Aluminium-induced drought and oxidative stress in barley roots. J. Plant Physiol. 163,781–784.
Thompson, A.J., Andrews, J., Mulholland, B.J., McKee, J.M.T., Hilton, H.W., Hjorridge,J.S., Farquhar, G.D., Smeeton, R.C., Smillie, I.R.A., Black, C.R., Taylor, I.B., 2007.Overproduction of abscisic acid in tomato increases transpiration efficiency androot hydraulic conductivity and influences leaf expansion. Plant Physiol. 143,1905–1917.
Tiedemann, R., 1989. Graft union development and symplastic phloem contact in theheterograft Cucumis sativus on Curcubita ficifolia. J. Plant Physiol. 134, 427–440.
Turnbull, C.G.N., Booker, J.P., Leyser, H.M., 2002. Micrografting techniques for testinglong-distance signalling in Arabidopsis. Plant J. 32, 255–262.
Van Hooijdonk, B.M., Woolley, D.J., Warrington, I.J., Tustin, D.S., 2010. Initialalteration of scion architecture by dwarfing apple rootstocks may involveshoot–root–shoot signalling by auxin, gibberellin, and cytokinin. J. Hortic. Sci.Biotechnol. 85, 59–65.
Xu, C.Q., Li, T.L., Qi, H.Y., Wang, H., 2005. Effects of grafting on growth and develop-ment, yield, and quality of muskmelon. China Veg. 6, 12–14.
Yamaguchi, T., Blumwald, E., 2005. Developing salt-tolerant crop plants: challengesand opportunities. Trends Plant Sci. 10, 615–620.
Yamamoto, Y., Kobayashi, Y., Devi, S.R., Rikiishi, S., Matsumoto, H., 2003. Oxidativestress triggered by aluminum in plant roots. Plant Soil 255, 239–243.
activity in the xylem exudates of grafted watermelons as affected by rootstockand crop load. J. Jpn. Soc. Hortic. Sci. 62, 817–826.
Yeoman, M.M., 1984. Cellular recognition systems in grafting. In: Linkskens, H.F.,Heslop-Harrison, I. (Eds.), Cellular Interaction, Encyclopaedia of Plant Physiol-ogy, New Series, vol. 17. Springer-Verlag, Berlin, pp. 453–472.




























