Embed Size (px)
Citation preview

ARTICLE IN PRESS
Microbiological Research 160 (2005) 429—436
KEYWORDChromeazIron chelaMethylobaextorquenXylella fasSideropho
0944-5013/$ - sdoi:10.1016/j.
�CorrespondE-mail addr
www.elsevier.de/micres
Growth and siderophore production of Xylellafastidiosa under iron-limited conditions
Maria Estela Silva-Stenicoa,�, Flavia Tereza Hansen Pachecoa,Jorge Luiz Mazza Rodriguesa, Emanuel Carrilhob, Siu Mui Tsaia
aLaboratorio de Biologia Celular e Molecular, Centro de Energia Nuclear na Agricultura – Universidade de Sao Paulo,Piracicaba, SP, BrazilbLaboratorio de Cromatografia, Instituto de Quımica de Sao Carlos – Universidade de Sao Paulo, Sao Carlos, SP, Brazil
Accepted 21 March 2005
Surol S;tion;cteriums;tidiosa;res
ee front matter & 200micres.2005.03.007
ing author.ess: [email protected].
SummaryIn this study, the production of siderophores by Xylella fastidiosa from the citrusbacteria isolate 31b9a5c (FAPESP – ONSA, Brazil) was investigated. The preliminaryevidence supporting the existence of siderophore in X. fastidiosa was found during theevaluation of sequencing data generated in our lab using the BLAST-X tool, whichindicated putative open reading frames (ORFs) associated with iron-binding proteins.In an iron-limited medium siderophores were detected in the supernatant of X.fastidiosa cultures. The endophytic bacterium Methylobacterium extorquens was alsoevaluated. Capillary electrophoresis was used to separate putative siderophoresproduced by X. fastidiosa. The bacterial culture supernatants of X. fastidiosa wereidentified negative for hydroxamate and catechol and positive for M. extorquens thatsecreted hydroxamate-type siderophores.& 2005 Elsevier GmbH. All rights reserved.
Introduction
Microorganisms need iron, which is an essentialelement for life (Loper and Buyer, 1991) and playsan important role in some host–bacteria interac-tions (Mila et al., 1996). Iron is a cofactor of severalenzymes and acts in transport processes and redoxreactions (Van Vliet et al., 1998). In addition, iron isrequired for a variety of functions in microorgan-
5 Elsevier GmbH. All rights rese
br (M. Estela Silva-Stenico).
isms that grow under aerobic conditions such asreduction of oxygen during ATP synthesis, reductionof ribonucleotide precursors of DNA, formation ofheme (Neilands, 1995), oxidation–reduction incellular reactions, which involve the activities ofcytochromes, hydroperoxidases, non-iron nitro-genases, and ribonucleotide reductases (Robsonet al., 1986). A concentration of, at least, 1 mM ofiron is needed for optimum growth (Neilands,
rved.

ARTICLE IN PRESS
M. Estela Silva-Stenico et al.430
1995); however, this concentration varies fordifferent organisms.
In an iron-limited condition, microorganismsproduce siderophores (low molar mass biomole-cules � 0.5–1.5 kDa) to scavenge ferric ions, whichbind to specific outer membrane receptors withhigh affinity. Some donor groups for the chelationof Fe3+ are hydroxamates, thiohydroxamates andcatecholates (Richardson et al., 1999). Manybacteria can synthesize their own chelator orutilize other microbial- and plant-siderophores foriron acquisition (Grusak et al., 1999).
The mechanism of iron acquisition is known tobe a virulence factor for human and animalpathogenic bacteria (Neilands, 1995). In thecase of plants (e.g. Saintpaulia ionantha leaves),studies conducted by Neema et al. (1993) confirmthat the siderophore chrysobactin producedby Erwinia chrysanthemi was involved in thedisease process.
Xylella fastidiosa is a pathogenic bacteriumin citrus and in a large variety of other hosts. InBrazil, this bacterium causes citrus variegatedchlorosis (CVC), resulting in losses of US $ 100million per year to the citrus industry (Coletta-Filhoet al., 2001). Although the entire genome of X.fastidiosa was sequenced (Simpson et al., 2000),little is known about its genetics, pathogenesis, andecological significance. The presence of putativegenes in X. fastidiosa (Simpson et al., 2000), codingfor iron uptake membrane receptors suggests apotential role for chelating agents during thedevelopment of disease symptoms such as chlorosis(Neema et al., 1993). Pacheco et al. (2001) haveconducted a detailed molecular study relating theproduction of siderophores with the presence ofputative iron uptake receptors. They have screenedover 80 isolates by PCR-specific primers designedfor the ferric enterobactin receptor (PfeA), primersfor the hydroxamate-type ferrisiderophore recep-tor gene (fiuA) (pyoverdin), and also a new set ofprimers for a 300 bp fragment coding for a polyke-tide synthase (PKS). Of the 80 isolates, 70 weresiderophore producers (chemical test) and alsoyielded positive PCR amplifications for all threeputative genes tested; therefore the production ofsecondary metabolites may, in part, be involved invirulence of X. fastidiosa against host plants.
In order to compare siderophores production byother bacteria found in contaminated citrus plantswe included the endophytic bacterium Methylo-bacterium extorquens isolated from healthy andsymptomatic plants of Citrus sinensis (Araujo etal., 2002). The purpose of this work was toinvestigate the effect of iron in the production ofsiderophores by X. fastidiosa, which could be
involved in the iron-stress related virulence of thispathogen.
Materials and methods
Bacterial strains
X. fastidiosa (9a5c – citrus), from the GenomeSequencing Project, was obtained from INRA –
Institut National de la Recherche Agronomique etUniversite Victor Segale, Bordeaux, France. M.extorquens was given by the Department of Geneticsfrom Escola Superior de Agricultura Luiz de Queiroz– Universidade de Sao Paulo (ESALQ/USP), Brazil.
Media and bacterial growth
Strains were cultured three times in an iron-deficient liquid medium (MM9) according to Payne(1994). The medium was composed of 0.3 g l�1
KH2PO4; 0.5 g l�1 NaCl; 1.0 g l�1 NH4Cl; 6.0 g l�1
NaOH, and 30.24 g l�1 PIPES. This solution wasautoclaved and supplemented with 30ml of 10%(m/v) deferrated casamino acids (contaminatingiron was removed with 3% 8-hydroxyquinoline inchloroform), 2.0 g l�1 fructose, 1ml of 1M MgCl2and 1ml of 100mM CaCl2. These solutions wereprepared and sterilized separately.
Glassware was cleaned with 6M HCl prior to use.Growth was measured by monitoring the opticaldensity at 600 nm (OD600) with a Perkin-Elmerspectrophotometer. Bacteria were grown from astandard inoculum (100 ml of a suspension with anabsorbance at 600 nm of 0.5) in 200ml flaskscontaining 50ml MM9 medium and incubated at28 1C on a rotary shaker at 150 rpm.
Iron requirement
To verify iron requirement, X. fastidiosa growthwas examined as a function of iron concentrationadded to the MM9 medium. Iron (FeCl3) was addedto 200ml flasks containing 50ml MM9 medium atthe following concentrations: 0, 0.1, 10, 50 and100 mmol l�1. The flasks were incubated at 28 1C for12 days. Growth rate was measured as above.
Siderophore-type assays
The Chromeazurol S (CAS) assay (Schwyn andNeilands, 1987) was used to detect siderophoresindependent of their structure in the MM9 culturesupernatants. The CAS liquid assay was performedas follows: a 6-ml volume of 10mM hexadecyltri-

ARTICLE IN PRESS
Growth and siderophore production of Xylella fastidiosa under iron-limited conditions 431
methylammonium bromide (HDTMA) solution wasplaced in a 100ml volumetric flask and diluted withwater. A mixture of 1.5ml of 1mM FeCl3 � 6H2O in10mM HCl and 7.5ml 2mM aqueous CAS solutionwas added slowly under stirring. A total of 4.3 g ofanhydrous piperazine was dissolved in water and6.25ml 12M hydrochloric acid was added. The flaskwas completed with distilled water to obtain 100mlof CAS solution. The pH of the solution was 5.6.Cells were removed by centrifugation (12,000g for20min). A 0.5ml aliquot of supernatant was mixedwith 0.5ml CAS assay solution. A reference wasprepared using sterile MM9 medium. After reachingequilibrium the absorbance was measured at630 nm. Positive reactions were recorded by achange in the color of the CAS reagent from blueto yellow or orange. CAS agar plates were alsoused, and positive results were recorded as a haloformation around the colonies. Catechol-type side-rophores were measured in culture supernatants bythe Arnow assay (Arnow, 1937) and hydroxamateswere measured according to Csaky (1948), with2,3-dihydroxybenzoic acid and hydroxylamine hy-drochloride as standards, respectively. Each assaywas performed in triplicate.
Desferrioxamine B equivalent assay
This assay was developed according to Shin et al.(2001). The CAS agar diffusion assay was performedas follows: 60.5mg CAS were dissolved in 50mldeionized water and mixed with 10ml iron (III)solution (1mM FeCl3 � 6H2O, 10mM HCl). Whilestirring, this solution was slowly mixed with72.9mg HDTMA dissolved in 40ml water. Theresultant dark blue solution was autoclaved andmixed with an autoclaved mixture of 900ml water,15 g agarose, 30.24 g PIPES, and 12 g of a solutionwith 50% (m/v) NaOH to raise the pH of the solutionnear the pKa of PIPES (6.8). Wells of 5mm diameterwere punched into agarose plates. Each well wasfilled with 100 mM of the supernatants and thehaloes were measured. A range of desferrioxamineconcentrations (0, 1, 5, 10, 50, 100, 500, 1000,1500, 2000, and 2500 mM) were used to quantifysiderophore production. A blank control of MM9medium was used to identify any interference withthe color development for this CAS agar assay. Theassay was performed in triplicate.
Capillary electrophoresis and samplepreparation
Bacteria were inoculated on agar plates contain-ing MM9 medium plus Chromeazurol S. An agar
block (1 cm2) excised from the halo region was usedfor analysis and a corresponding agar block, inwhich no bacterial growth was observed, was usedas control. Samples of the CAS agar inoculated withX. fastidiosa were extracted as shown in Fig. 3following disruption with 2ml 0.1M NaOH (1minshaking). After centrifugation, the supernatant wasfiltered (0.22 mm membrane) and injected in thecapillary electrophoresis system. Electrophoresisseparation was carried out on a P/ACE Beckman5510 (Palo Alto, CA, USA). A fused-silica capillarywith 75 mm inner diameter and 37 cm total lengthwas used. The samples were injected for 5 s under0.5 psi, the voltage was 30 kV. The temperature washeld constant at 25 1C. The separation was mon-itored at 214 nm using a UV-detector. The capillarywas conditioned after each run by the followingsequence: 1M HCl (1min), 1M NaOH (1min),deionized water (1min), 0.1M NaOH (1min), and0.1M potassium phosphate buffer pH 7 (2min). Thepeaks obtained in the electropherogram wereacquired from the P/ACE System (Beckman) soft-ware.
MALDI-TOF-MS (matrix-assisted laserdesorption–ionization time of flight massspectrometry) analysis
Strains were cultured three times in an iron-deficient liquid medium (MM9) as described earlier.Siderophore purification was carried out as follows:the immobilization of Fe3+ ions in the ChelatingSepharose Fast Flow (CSFF) adsorbent was preparedusing 3ml of resin, which was washed twice with asolution of 0.1mol l�1 iron (FeCl3 � 6H2O), followedby five washes with milli-Q water. The supernatantof each culture was added to the resin andcentrifuged at 1200g for 3min. The supernatantwas discarded and a solution of 1ml of 0.05mMEDTA was added. The complex Fe–resin wasdisrupted and the supernatant containing Fe–sider-ophore complex was lyophilized. Freeze-driedsamples were, then, prepared for MALDI-TOF on aVoyager-DE STR Bioworkstation (PerSeptive Biosys-tems, Framingham, MA, USA). Samples were dis-solved in 1% aqueous trifluoroacetic acid and thematrix sinapinic acid added (a saturated solutiondissolved in acetonitrile/0.1% TFA 1:1, v/v). Thesolution was then vortex mixed and aliquots of 1mlwere applied to the Voyager Bioworkstation sampleplate. Samples were air-dried at room tempera-ture. The spectrometer, equipped with a delayed-extraction system, was operated in linear mode.Sample ions were evaporated by irradiation witha N2 laser at a wavelength of 337 nm, and
ARTICLE IN PRESS
M. Estela Silva-Stenico et al.432
accelerated at 23 kV potential in the ion sourcewith a delay of 150 ns. Samples were ionized with100–200 shots of a 3 ns pulse width laser light. Thesignal was digitized at a rate of 500MHz andaveraged data was presented to a standard Voyagerdata system for manipulation.
Results
Cellular growth in different ironconcentration
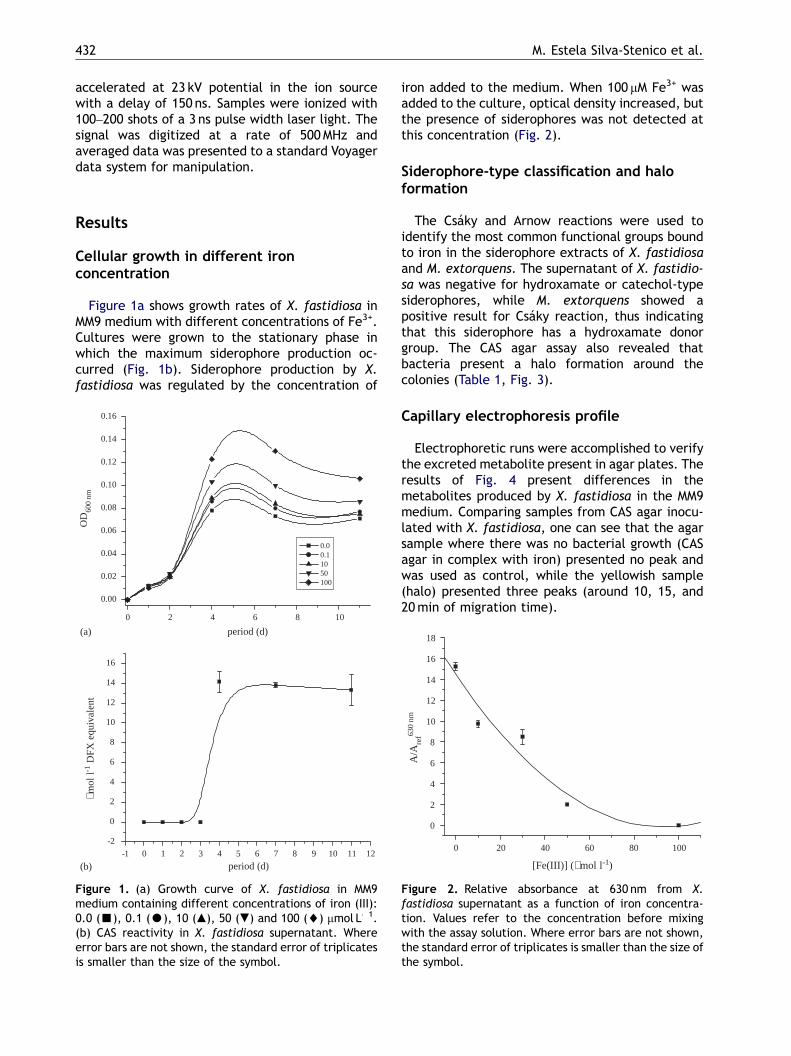
Figure 1a shows growth rates of X. fastidiosa inMM9 medium with different concentrations of Fe3+.Cultures were grown to the stationary phase inwhich the maximum siderophore production oc-curred (Fig. 1b). Siderophore production by X.fastidiosa was regulated by the concentration of
0 2 4 6 8 10
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
OD
600
nm
period (d)
0.0 0.1 10 50 100
-1 0 1 2 3 4 5 6 7 8 9 10 11 12
-2
0
2
4
6
8
10
12
14
16
µmol
l-1
DFX
equ
ival
ent
period (d)
(a)
(b)
Figure 1. (a) Growth curve of X. fastidiosa in MM9medium containing different concentrations of iron (III):0.0 (’), 0.1 (K), 10 (m), 50 (.) and 100 (~) mmol L�1.(b) CAS reactivity in X. fastidiosa supernatant. Whereerror bars are not shown, the standard error of triplicatesis smaller than the size of the symbol.
iron added to the medium. When 100 mM Fe3+ wasadded to the culture, optical density increased, butthe presence of siderophores was not detected atthis concentration (Fig. 2).
Siderophore-type classification and haloformation
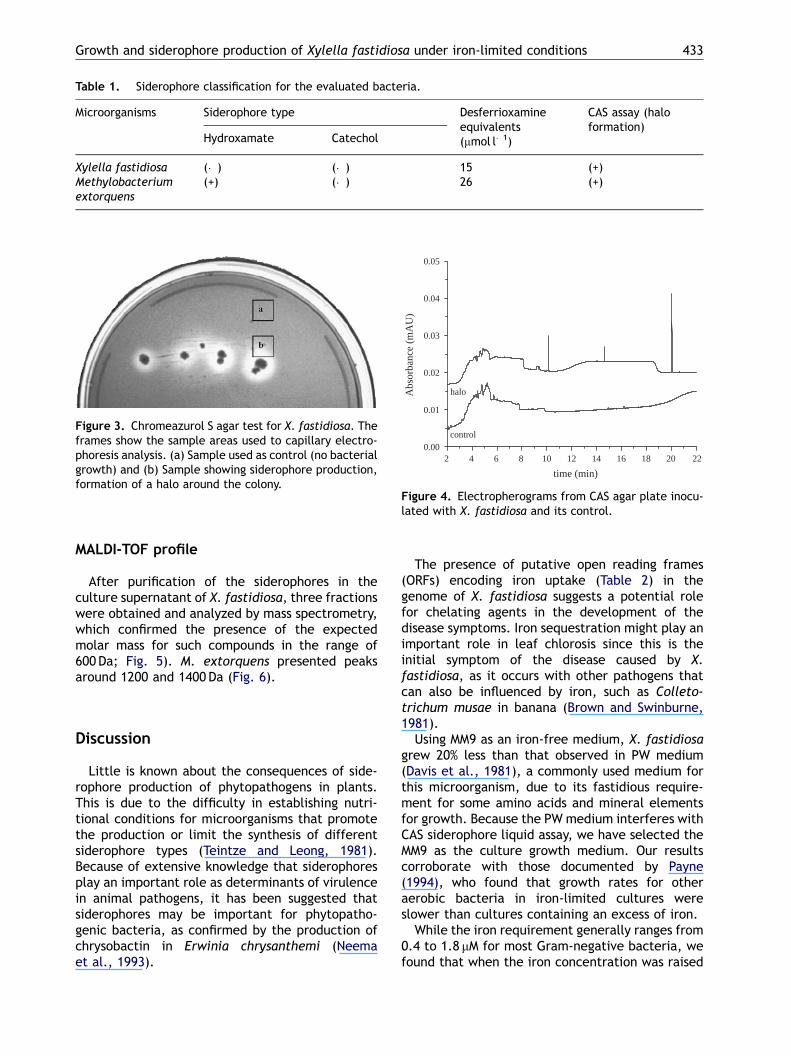
The Csaky and Arnow reactions were used toidentify the most common functional groups boundto iron in the siderophore extracts of X. fastidiosaand M. extorquens. The supernatant of X. fastidio-sa was negative for hydroxamate or catechol-typesiderophores, while M. extorquens showed apositive result for Csaky reaction, thus indicatingthat this siderophore has a hydroxamate donorgroup. The CAS agar assay also revealed thatbacteria present a halo formation around thecolonies (Table 1, Fig. 3).
Capillary electrophoresis profile
Electrophoretic runs were accomplished to verifythe excreted metabolite present in agar plates. Theresults of Fig. 4 present differences in themetabolites produced by X. fastidiosa in the MM9medium. Comparing samples from CAS agar inocu-lated with X. fastidiosa, one can see that the agarsample where there was no bacterial growth (CASagar in complex with iron) presented no peak andwas used as control, while the yellowish sample(halo) presented three peaks (around 10, 15, and20min of migration time).
0 20 40 60 80 100
0
2
4
6
8
10
12
14
16
18
A/A
ref63
0 nm
[Fe(III)] (µmol l-1)
Figure 2. Relative absorbance at 630 nm from X.fastidiosa supernatant as a function of iron concentra-tion. Values refer to the concentration before mixingwith the assay solution. Where error bars are not shown,the standard error of triplicates is smaller than the size ofthe symbol.
ARTICLE IN PRESS
Table 1. Siderophore classification for the evaluated bacteria.
Microorganisms Siderophore type Desferrioxamineequivalents(mmol l�1)
CAS assay (haloformation)
Hydroxamate Catechol
Xylella fastidiosa (�) (�) 15 (+)Methylobacteriumextorquens
(+) (�) 26 (+)
2 4 6 8 10 12 14 16 18 20 220.00
0.01
0.02
0.03
0.04
0.05
halo
control
Abs
orba
nce
(mA
U)
time (min)
Figure 4. Electropherograms from CAS agar plate inocu-lated with X. fastidiosa and its control.
Figure 3. Chromeazurol S agar test for X. fastidiosa. Theframes show the sample areas used to capillary electro-phoresis analysis. (a) Sample used as control (no bacterialgrowth) and (b) Sample showing siderophore production,formation of a halo around the colony.
Growth and siderophore production of Xylella fastidiosa under iron-limited conditions 433
MALDI-TOF profile
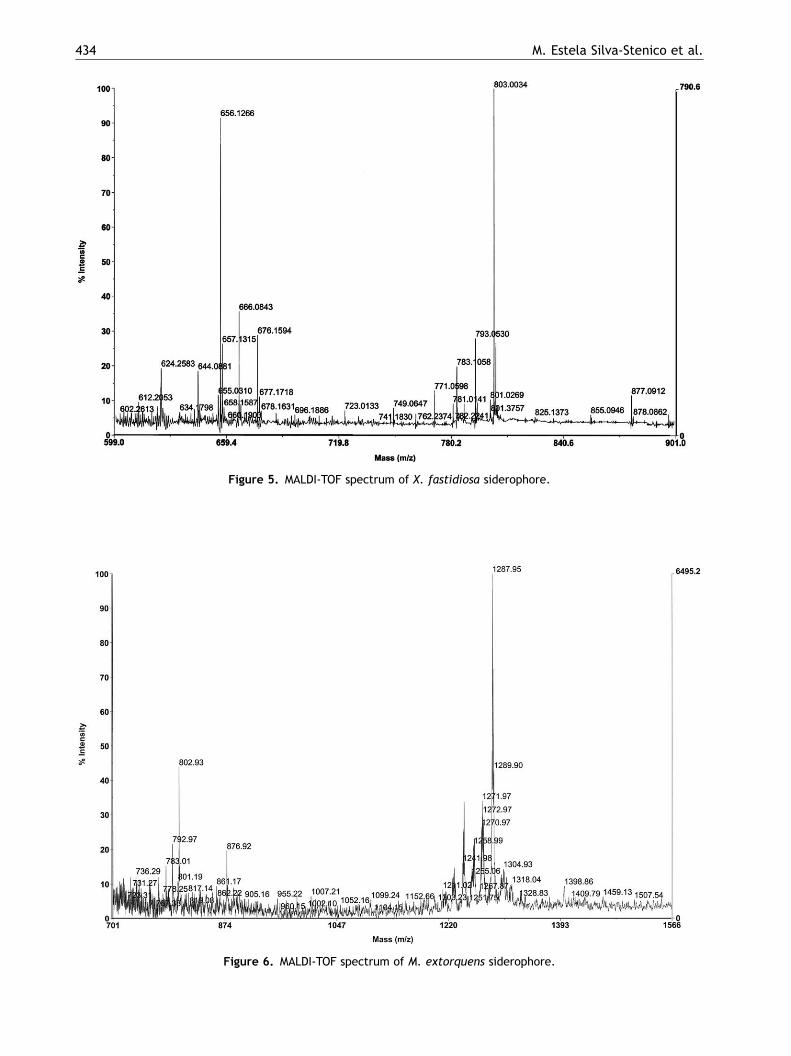
After purification of the siderophores in theculture supernatant of X. fastidiosa, three fractionswere obtained and analyzed by mass spectrometry,which confirmed the presence of the expectedmolar mass for such compounds in the range of600 Da; Fig. 5). M. extorquens presented peaksaround 1200 and 1400 Da (Fig. 6).
Discussion
Little is known about the consequences of side-rophore production of phytopathogens in plants.This is due to the difficulty in establishing nutri-tional conditions for microorganisms that promotethe production or limit the synthesis of differentsiderophore types (Teintze and Leong, 1981).Because of extensive knowledge that siderophoresplay an important role as determinants of virulencein animal pathogens, it has been suggested thatsiderophores may be important for phytopatho-genic bacteria, as confirmed by the production ofchrysobactin in Erwinia chrysanthemi (Neemaet al., 1993).
The presence of putative open reading frames(ORFs) encoding iron uptake (Table 2) in thegenome of X. fastidiosa suggests a potential rolefor chelating agents in the development of thedisease symptoms. Iron sequestration might play animportant role in leaf chlorosis since this is theinitial symptom of the disease caused by X.fastidiosa, as it occurs with other pathogens thatcan also be influenced by iron, such as Colleto-trichum musae in banana (Brown and Swinburne,1981).
Using MM9 as an iron-free medium, X. fastidiosagrew 20% less than that observed in PW medium(Davis et al., 1981), a commonly used medium forthis microorganism, due to its fastidious require-ment for some amino acids and mineral elementsfor growth. Because the PW medium interferes withCAS siderophore liquid assay, we have selected theMM9 as the culture growth medium. Our resultscorroborate with those documented by Payne(1994), who found that growth rates for otheraerobic bacteria in iron-limited cultures wereslower than cultures containing an excess of iron.
While the iron requirement generally ranges from0.4 to 1.8 mM for most Gram-negative bacteria, wefound that when the iron concentration was raised
ARTICLE IN PRESS
Figure 5. MALDI-TOF spectrum of X. fastidiosa siderophore.
Figure 6. MALDI-TOF spectrum of M. extorquens siderophore.
M. Estela Silva-Stenico et al.434

ARTICLE IN PRESS
Table 2. Putative ORFs encoding iron uptake in thegenome of X. fastidiosa.
Genbank accessionnumbers
Gene function
XF0599 TonB-dependent receptorfor iron transport
XF1038 Peptide synthaseXF2135 PKS (polyketide synthase)XF2137 Ferric enterobactin receptorXF2276 Peptide synthase
Growth and siderophore production of Xylella fastidiosa under iron-limited conditions 435
to 100 mM the optical density of X. fastidiosacultures increased 66% (Fig. 1a). Siderophoresynthesis is mainly regulated by iron and can bemaximized under low iron concentration condi-tions, thus improving bacterial growth (Bagg andNeilands, 1987).
The CAS assay, developed by Schwyn and Nei-lands (1987), is based on the chelation of iron byany class of siderophores. Both species X. fastidiosaand M. extorquens were tested positive for thisassay (Table 1). The CAS-reactive peak from X.fastidiosa occurred during the late log to the earlystationary phase of growth, around 4 days (Fig. 1aand b). The CAS assay was not intense for X.fastidiosa (15 mM desferrioxamine equivalents)when compared to animal and human pathogenssuch as Legionella pneumophila (Liles et al., 2000),Helicobacter hepaticus, and H. cianedi (Dhaenenset al., 1999). This could be due to the slow growthof X. fastidiosa, taking up to 5 days for a positiveCAS reaction to occur.
To verify the repression of siderophore produc-tion by iron availability, X. fastidiosa was grown inthe presence of several iron concentrations. Theresults presented in Fig. 2 show that the optimalchelator production was reached when no iron wasadded to the medium. At 50 and 100 mM theproduction was poor or no production occurred.
The supernatant of X. fastidiosa was testednegative for hydroxamate- or catechol-type side-rophores, and that was positive in the Csaky test forM. extorquens. Negative results for X. fastidiosasupernatant suggests that the siderophore type hasno hydroxamate- or catechol-type functionalgroups (Guan et al., 2000).
Capillary electrophoresis provides a powerfultool for the separation of charged analytes andbiomolecules at sub-picomole levels. It is applied asa fast and efficient method for analysis of com-pounds and the technique was chosen due to thelow concentration of these metabolites producedby X. fastidiosa. Mucha et al. (1999) using CEdemonstrated the presence of siderophores in the
subsurface of seawater, which makes CE a good wayto detect such molecules. In the case of X.fastidiosa growing in agar plates, the metaboliteswere analyzed in complex with iron (III) (CAScomplex) and electropherograms showed the pre-sence of three compounds excreted by the bacter-ia. When the same samples were injected in HPLC,we also found three peaks with siderophore activity(data not shown). These compounds have not beenpreviously detected in X. fastidiosa supernatants.
MALDI-TOF spectra indicated a pseudo-molecularion with m/z 656 for X. fastidiosa (Fig. 5) and m/z1287 for M. extorquens (Fig. 6). The appearance ofmajor peaks in both spectra ranging from 790 to876 Da was also detected in the blank (culturemedium with no bacteria), possibly due to theinterference of matrix constituents. For the struc-tural elucidation of such molecules further MS/MSfragmentation should be carried out and it will bepursued next. Such molecular characterization willbe fundamental in the understanding of iron uptakemetabolism and its relationship to virulence ofX. fastidiosa strains.
Acknowledgements
We thank Dr. W.L. Araujo (Department ofGenetics – ESALQ/USP) for providing us with theendophytic bacterium and Dr. C. Bloch Jr. (Massspectrometry laboratory – EMBRAPA) for technicalassistance with MALDI-TOF-MS analysis. This studywas supported by Grants 00/08045-3 and 99/07748-1 from FAPESP (Fundac-ao de Amparo a Pesquisa doEstado de Sao Paulo).
References
Araujo, W.L., Marcon, J., Maccheroni Jr., W., van Elsas,J.D., van Vuurde, J.W.L., Azevedo, J.L., 2002.Diversity of endophytic bacterial populations andtheir interaction with Xylella fastidiosa in citrusplants. Appl. Environ. Microbiol. 68, 4906–4914.
Arnow, L.E., 1937. Colorimetric determination of thecomponents of 3,4-dihydroxyphenylalanine–tyrosinemixtures. J. Biol. Chem. 118, 531–537.
Bagg, A., Neilands, J.B., 1987. Molecular mechanism ofregulation of siderophore-mediated iron assimilation.Microbiol. Rev. 51, 509–518.
Brown, A.E., Swinburne, T.R., 1981. Influence of iron andiron chelators on formation of progressive lesions byColletotrichum musae on banana fruits. Trans. Br.Mycol. Soc. 77, 119–124.
Coletta-Filho, H.D., Takita, M.A., Souza, A.A., Aguilar-Vildoso, C.I., Machado, M.A., 2001. Differentiation ofstrains of Xylella fastidiosa by a variable number of

ARTICLE IN PRESS
M. Estela Silva-Stenico et al.436
tandem repeat analysis. Appl. Environ. Microbiol. 67,4091–4095.
Csaky, T.Z., 1948. On the estimation of bound hydro-xylamine in biological materials. Acta Chem. Scand. 2,450–454.
Davis, M.J., French, W.J., Schaad, N.W., 1981. Axenicculture of the bacteria associated with phony diseaseof peach and plum leaf scald. Curr. Microbiol. 6,309–314.
Dhaenens, L., Szczebara, F., Van Nieuwenhuyse, S.,Husson, M.-O., 1999. Comparison of iron uptake indifferent Helicobacter species. Res. Microbiol. 150,475–481.
Grusak, M.A., Pearson, J.N., Marentes, E., 1999. Thephysiology of micronutrient homeostasis in field crops.Field Crops Res. 60, 41–56.
Guan, L.L., Onuki, H., Kamino, K., 2000. Bacterial growthstimulation with exogenous siderophore and syntheticN-acyl homoserine lactone autoinducers under iron-limited and low-nutrient conditions. Appl. Environ.Microbiol. 66, 2797–2803.
Liles, M.R., Scheel, T.A., Cianciotto, N.P., 2000. Dis-covery of a non-classical siderophore, legiobactin,produced by strains of Legionella penumophila. J.Bacteriol. 182, 749–757.
Loper, J.E., Buyer, J.S., 1991. Siderophores in microbialinteractions on plant surfaces. Mol. Plant MicrobeInteract. 4, 5–13.
Mila, I., Scalbert, A., Expert, D., 1996. Iron withholdingby plant polyphenols and resistance to pathogens androts. Phytochemistry 42, 1551–1555.
Mucha, P., Rekowski, P., Kosakowska, A., Kupryszewski,G., 1999. Separation of siderophores by capillaryelectrophoresis. J. Chromatogr. A 830, 183–189.
Neema, C., Laulhere, J.-P., Expert, D., 1993. Irondeficiency induced by chrysobactin in Saintpaulia
ionantha leaves inoculated with Erwinia chrysanthe-mi. Plant Physiol. 102, 967–973.
Neilands, J.B., 1995. Siderophores: structure and func-tion of microbial iron transport compounds. J. Biol.Chem. 270, 26723–26726.
Pacheco, F.T.H., Silva, M.E., Rodrigues, J.L.M., Tsai, S.M.,2001. Molecular characterization of putative ironreceptor genes in Xylella fastidiosa. In: RhizosphereControl Points: Molecules to Food Webs, June 1–3.University of California, Davis, USA.
Payne, S.M., 1994. Detection, isolation and characteriza-tion of siderophores. Methods Enzymol. 235, 329–344.
Richardson, N., Davies, J.A., Raduchel, B., 1999. Iron(III)-based contrast agents for magnetic resonanceimaging. Polyhedron 18, 2457–2482.
Robson, R.L., Eady, R.R., Richardson, T.H., Miller, R.W.,Hawkins, M., Postgate, J.M., 1986. The alternativenitrogenase of Azotobacter chroococcum. Nature 322,388–390.
Schwyn, B., Neilands, J.B., 1987. Universal chemicalassay for the detection and determination of side-rophores. Anal. Biochem. 160, 47–56.
Shin, S.H., Lim, Y., Lee, S.E., Yang, N.W., Rhee, J.H.,2001. CAS agar diffusion assay for the measurement ofsiderophores in biological fluids. J. Microbiol. Methods44, 89–95.
Simpson, A.J.G., Reinach, F.C., Arruda, P., and 112other authors, 2000. The genome sequence of theplant pathogen Xylella fastidiosa. Nature 406,151–157.
Teintze, M., Leong, J., 1981. Structure of pseudobactinA, a second siderophore from plant growth promotingPseudomonas B10. Biochemistry 20, 6457–6462.
Van Vliet, A.H.M., Wooldridge, K.G., Ketley, J.M., 1998.Iron-responsive gene regulation in a Campylobacterjejuni fur mutant. J. Bacteriol. 180, 5291–5298.





























