Embed Size (px)
Citation preview

Ž .Comparative Biochemistry and Physiology Part B 129 2001 407�417
Review
Gonadotropins, their receptors, and the regulation oftesticular functions in fish
R.W. Schulza,�, H.F. Vischer a, J.E.B. Cavacoa, E.M. Santosb, C.R. Tylerb,H.J.Th. Goosa, J. Bogerda
aResearch Group Endocrinology, Department of De�elopmental Biology, Faculty of Biology, Utrecht Uni�ersity, Padualaan 8,3584 CH Utrecht, The Netherlands
bDepartment of Biological Sciences, Brunel Uni�ersity, Uxbridge, Middlesex UB8 3PA, UK
Received 4 August 2000; received in revised form 20 November 2000; accepted 8 December 2000
Abstract
Ž . Ž .The pituitary gonadotropins luteinizing hormone LH and follicle-stimulating hormone FSH regulate steroidogene-Ž . Ž .sis and spermatogenesis by activating receptors expressed by Leydig cells LH receptor and Sertoli cells FSH receptor ,
respectively. This concept is also valid in fish, although the piscine receptors may be less discriminatory than theirmammalian counterparts. The main biological activity of LH is to regulate Leydig-cell steroid production. Steroidogene-
Žsis is moreover modulated in an autoregulatory manner by androgens. The male sex steroids testosterone in higher.vertebrates, 11-ketotestosterone in fish are required for spermatogenesis, but their mode of action has remained
obscure. While piscine FSH also appears to have steroidogenic activity, specific roles have not been described yet in thetestis. The feedback of androgens on gonadotrophs presents a complex pattern. Aromatizable androgens�estrogensstimulate LH synthesis in juvenile fish; this effect fades out during maturation. This positive feedback on LH synthesis isbalanced by a negative feedback on LH release, which may involve GnRH neurones. While the role of GnRH as LHsecretagogue is evident, we have found no indication in adult male African catfish for a direct, GnRH-mediatedstimulation of LH synthesis. The limited available information at present precludes a generalized view on the testicularfeedback on FSH. � 2001 Elsevier Science Inc. All rights reserved.
Keywords: Teleost fish; African catfish; Luteinizing hormone; Follicle-stimulating hormone; Luteinizing hormone receptor; Follicle-stimulating hormone receptor; Androgen production; Spermatogenesis; Steroid feedback
� Corresponding author. Tel.: �31-30-2533046; fax: �31-30-2532837.Ž .E-mail address: [email protected] R.W. Schulz .
1096-4959�01�$ - see front matter � 2001 Elsevier Science Inc. All rights reserved.Ž .PII: S 1 0 9 6 - 4 9 5 9 0 1 0 0 3 3 9 - 6

( )R.W. Schulz et al. � Comparati�e Biochemistry and Physiology Part B 129 2001 407�417408
1. Scope of the review
Spermatogenesis and hormone production areregulated extrinsically by pituitary hormones andintrinsically by testicular signalling systems. Thiscontribution deals with both aspects of the regu-lation of testicular functions. The duality of go-nadotropins will first be discussed, followed by apresentation on the recent findings on go-nadotropin receptors. Attention will then be di-rected to the regulation of the production ofandrogens by androgens, before we turn to theirrole in spermatogenesis. Aspects of the androgenfeedback on brain and pituitary will then be dis-cussed. In addition to reviewing recent informa-tion from different species, we will present some
Žoriginal data from male African catfish Clarias.gariepinus .
2. Gonadotropins
The gonadotropins are heterodimeric glycopro-teins composed of a common �-subunit and ahormone-specific �-subunit. While teleost go-nadotropins belong to the FSH or the LH groupfrom a structural point of view, discussion on thefunctional homology is ongoing, at least with re-gard to FSH. In this review, however, we willrefer to GTH I-like hormones as FSH, and toGTH II-like hormones as LH, as has been pro-
Ž .posed earlier by others e.g. Prat et al., 1996 . Theduality of gonadotropins has now been es-tablished for all teleost orders examined. Whilewe have previously expressed a more cautious
Ž .view Schulz et al., 1995a , recent work in theŽJapanese eel Anguilla japonica, Yoshiura et al.,
.1999 and in our laboratory on African catfishŽ .Vischer and Bogerd, unpublished results hasconfirmed the presence of FSH �-subunit mRNAin the pituitary of these species. Hence, anotherattempt was made to purify catfish FSH. To this
Žend, a protocol was adopted Swanson et al.,.1991 that has been successful for a number of
Žother fish species e.g. Weltzien et al., 1999; San-.tos et al., 2000 .
Proteins from 800 male catfish pituitaries wereethanol-extracted and fractionated by gel filtra-tion chromatography and the fractions corre-sponding to the molecular weight of the go-
Ž .nadotropins approx. 40 000 Da were pooled and
further fractionated by DE-52 ion exchange chro-matography, using an ammonium bicarbonate
Ž .buffer pH 7.8 with a step-wise increasing molar-Ž .ity 0.05�0.5 M . The resulting peak fractions,
containing intact LH and its subunits, were iden-Ž .tified by RIA Schulz et al., 1995b . Column frac-
tions eluted at 0.2 M ammonium bicarbonate thatdisplaced LH and�or its subunits in the RIAswere further fractionated by gel filtration chro-matography on a Sephadex G75 column. Theascending part of the resulting peak fraction was
Ž .analysed by HPLC Fig. 1 . The HPLC profilesŽshowed a number of �-subunit peaks ca. 25-min
.elution time and two peaks corresponding to theŽelution positions of �-subunits ca. 35-min elution
.time . Our original thoughts were that the lattermight correspond to FSH and LH �-subunits.However, analysis of the amino acid compositionand partial amino acid sequence analysis identi-
Žfied them both as catfish LH �-subunit by com-parison with the sequence obtained by Koide et
.al., 1992 . The difference in elution time in HPLCmay be due to different glycan compositions ofthe �-subunits. Together, these results demon-strate the absence of other �-subunits in theisolated LH preparation, and in this way, vali-dated this preparation as a standard for use inimmunoassays, receptor-binding assays and physi-ological studies.
The �-subunits eluted in HPLC as two groupsof peaks that differed in their amino acid se-quence and�or glycan composition, as found in
Žother fish species Swanson et al., 1991; Gou-.voroun et al., 1997 . It remains unclear what the
biological significance of two distinct �-subunitsis, and how the composition of intact LH varieswith the reproductive stage of the fish. In theanalysis of this evidence, the possibility that thedifferent peaks may correspond to immatureforms of LH subunits during their productionprocess within the gonadotrophs must be con-sidered, and more studies are needed to elucidateif different isoforms are released into the circula-tion. In mammals, a number of studies have de-
Žmonstrated that the glycan composition and.therefore, the acidity of the gonadotropins in-
Žfluences their biological activity reviewed in Tho-.takura and Blithe, 1995 . In fish, however, no data
are available on the presence of different go-nadotropin isoforms in the circulation and�or
Ž .their biological function if any . We are now
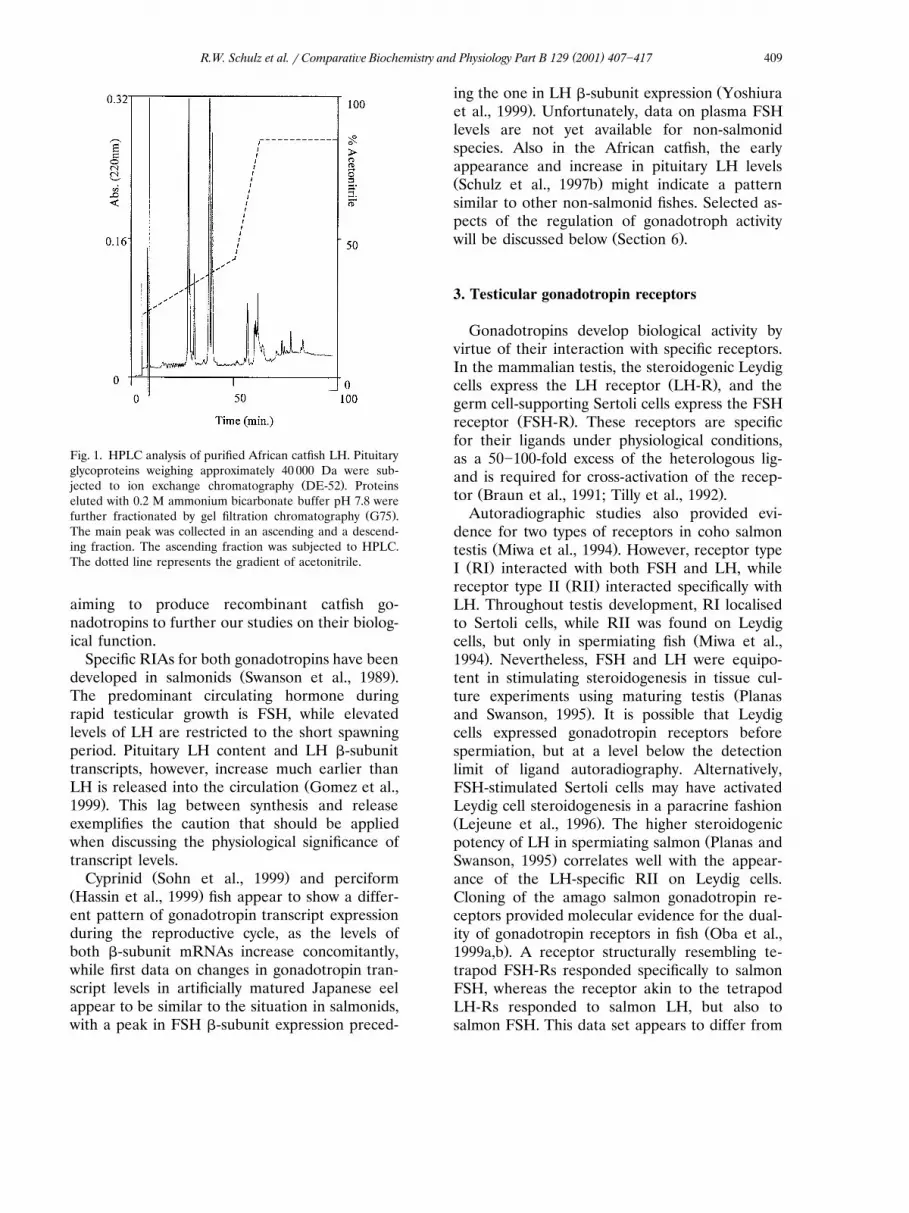
( )R.W. Schulz et al. � Comparati�e Biochemistry and Physiology Part B 129 2001 407�417 409
Fig. 1. HPLC analysis of purified African catfish LH. Pituitaryglycoproteins weighing approximately 40 000 Da were sub-
Ž .jected to ion exchange chromatography DE-52 . Proteinseluted with 0.2 M ammonium bicarbonate buffer pH 7.8 were
Ž .further fractionated by gel filtration chromatography G75 .The main peak was collected in an ascending and a descend-ing fraction. The ascending fraction was subjected to HPLC.The dotted line represents the gradient of acetonitrile.
aiming to produce recombinant catfish go-nadotropins to further our studies on their biolog-ical function.
Specific RIAs for both gonadotropins have beenŽ .developed in salmonids Swanson et al., 1989 .
The predominant circulating hormone duringrapid testicular growth is FSH, while elevatedlevels of LH are restricted to the short spawningperiod. Pituitary LH content and LH �-subunittranscripts, however, increase much earlier than
ŽLH is released into the circulation Gomez et al.,.1999 . This lag between synthesis and release
exemplifies the caution that should be appliedwhen discussing the physiological significance oftranscript levels.
Ž .Cyprinid Sohn et al., 1999 and perciformŽ .Hassin et al., 1999 fish appear to show a differ-ent pattern of gonadotropin transcript expressionduring the reproductive cycle, as the levels ofboth �-subunit mRNAs increase concomitantly,while first data on changes in gonadotropin tran-script levels in artificially matured Japanese eelappear to be similar to the situation in salmonids,with a peak in FSH �-subunit expression preced-
Žing the one in LH �-subunit expression Yoshiura.et al., 1999 . Unfortunately, data on plasma FSH
levels are not yet available for non-salmonidspecies. Also in the African catfish, the earlyappearance and increase in pituitary LH levelsŽ .Schulz et al., 1997b might indicate a patternsimilar to other non-salmonid fishes. Selected as-pects of the regulation of gonadotroph activity
Ž .will be discussed below Section 6 .
3. Testicular gonadotropin receptors
Gonadotropins develop biological activity byvirtue of their interaction with specific receptors.In the mammalian testis, the steroidogenic Leydig
Ž .cells express the LH receptor LH-R , and thegerm cell-supporting Sertoli cells express the FSH
Ž .receptor FSH-R . These receptors are specificfor their ligands under physiological conditions,as a 50�100-fold excess of the heterologous lig-and is required for cross-activation of the recep-
Ž .tor Braun et al., 1991; Tilly et al., 1992 .Autoradiographic studies also provided evi-
dence for two types of receptors in coho salmonŽ .testis Miwa et al., 1994 . However, receptor type
Ž .I RI interacted with both FSH and LH, whileŽ .receptor type II RII interacted specifically with
LH. Throughout testis development, RI localisedto Sertoli cells, while RII was found on Leydig
Žcells, but only in spermiating fish Miwa et al.,.1994 . Nevertheless, FSH and LH were equipo-
tent in stimulating steroidogenesis in tissue cul-Žture experiments using maturing testis Planas
.and Swanson, 1995 . It is possible that Leydigcells expressed gonadotropin receptors beforespermiation, but at a level below the detectionlimit of ligand autoradiography. Alternatively,FSH-stimulated Sertoli cells may have activatedLeydig cell steroidogenesis in a paracrine fashionŽ .Lejeune et al., 1996 . The higher steroidogenic
Žpotency of LH in spermiating salmon Planas and.Swanson, 1995 correlates well with the appear-
ance of the LH-specific RII on Leydig cells.Cloning of the amago salmon gonadotropin re-ceptors provided molecular evidence for the dual-
Žity of gonadotropin receptors in fish Oba et al.,.1999a,b . A receptor structurally resembling te-
trapod FSH-Rs responded specifically to salmonFSH, whereas the receptor akin to the tetrapodLH-Rs responded to salmon LH, but also tosalmon FSH. This data set appears to differ from
( )R.W. Schulz et al. � Comparati�e Biochemistry and Physiology Part B 129 2001 407�417410
Žprevious findings in coho salmon Miwa et al.,.1994 . The use of more sensitive detection sys-
tems and recombinant, species-specific go-nadotropins will help in clarifying the questionsrelating to ligand specificity.
A gonadotropin receptor from the African cat-Ž .fish testis Bogerd et al., 1999 shows structural
similarities to the FSH-R family. We have testedŽpurified piscine gonadotropins courtesy of Dr B.
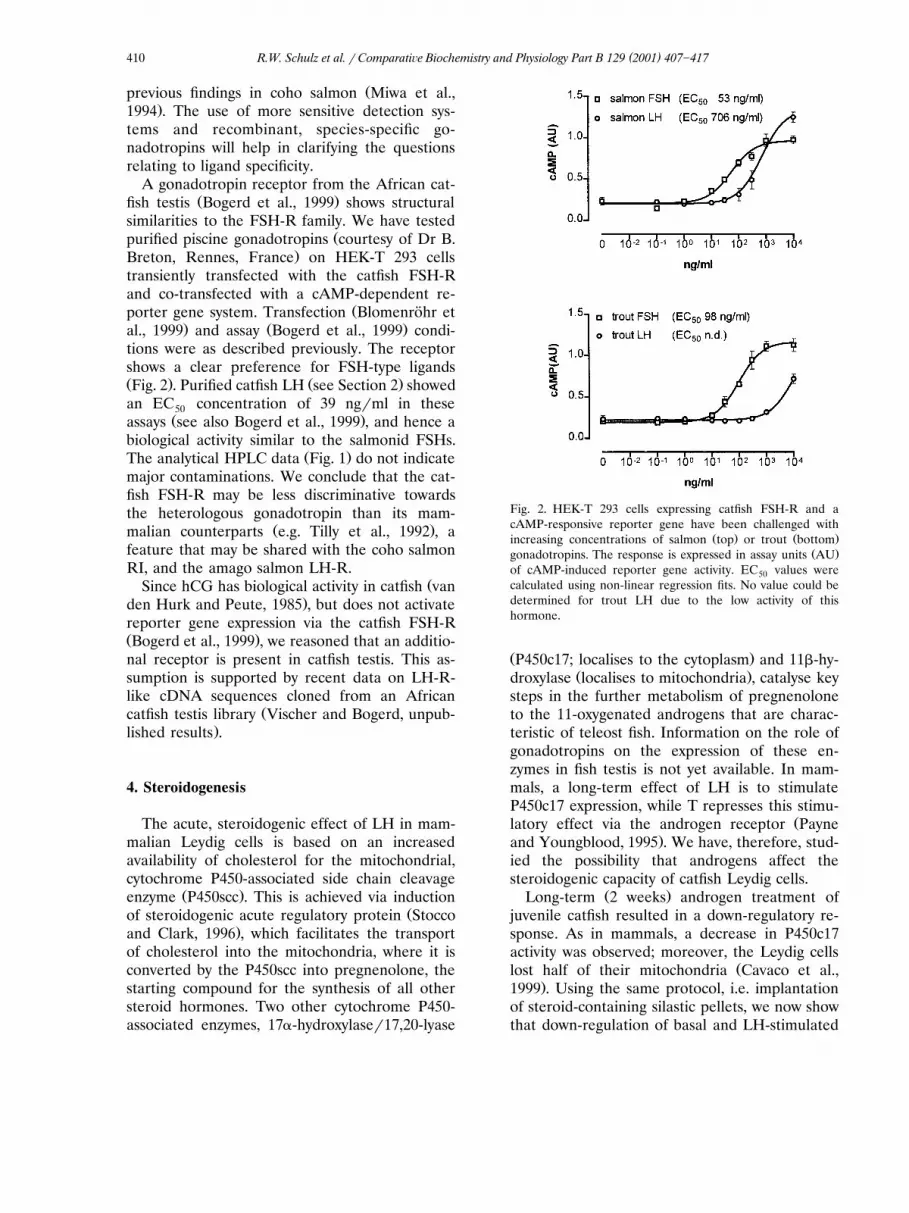
.Breton, Rennes, France on HEK-T 293 cellstransiently transfected with the catfish FSH-Rand co-transfected with a cAMP-dependent re-
Žporter gene system. Transfection Blomenrohr et¨. Ž .al., 1999 and assay Bogerd et al., 1999 condi-
tions were as described previously. The receptorshows a clear preference for FSH-type ligandsŽ . Ž .Fig. 2 . Purified catfish LH see Section 2 showedan EC concentration of 39 ng�ml in these50
Ž .assays see also Bogerd et al., 1999 , and hence abiological activity similar to the salmonid FSHs.
Ž .The analytical HPLC data Fig. 1 do not indicatemajor contaminations. We conclude that the cat-fish FSH-R may be less discriminative towardsthe heterologous gonadotropin than its mam-
Ž .malian counterparts e.g. Tilly et al., 1992 , afeature that may be shared with the coho salmonRI, and the amago salmon LH-R.
ŽSince hCG has biological activity in catfish van.den Hurk and Peute, 1985 , but does not activate
reporter gene expression via the catfish FSH-RŽ .Bogerd et al., 1999 , we reasoned that an additio-nal receptor is present in catfish testis. This as-sumption is supported by recent data on LH-R-like cDNA sequences cloned from an African
Žcatfish testis library Vischer and Bogerd, unpub-.lished results .
4. Steroidogenesis
The acute, steroidogenic effect of LH in mam-malian Leydig cells is based on an increasedavailability of cholesterol for the mitochondrial,cytochrome P450-associated side chain cleavage
Ž .enzyme P450scc . This is achieved via inductionŽof steroidogenic acute regulatory protein Stocco
.and Clark, 1996 , which facilitates the transportof cholesterol into the mitochondria, where it isconverted by the P450scc into pregnenolone, thestarting compound for the synthesis of all othersteroid hormones. Two other cytochrome P450-associated enzymes, 17�-hydroxylase�17,20-lyase
Fig. 2. HEK-T 293 cells expressing catfish FSH-R and acAMP-responsive reporter gene have been challenged with
Ž . Ž .increasing concentrations of salmon top or trout bottomŽ .gonadotropins. The response is expressed in assay units AU
of cAMP-induced reporter gene activity. EC values were50calculated using non-linear regression fits. No value could bedetermined for trout LH due to the low activity of thishormone.
Ž .P450c17; localises to the cytoplasm and 11�-hy-Ž .droxylase localises to mitochondria , catalyse key
steps in the further metabolism of pregnenoloneto the 11-oxygenated androgens that are charac-teristic of teleost fish. Information on the role ofgonadotropins on the expression of these en-zymes in fish testis is not yet available. In mam-mals, a long-term effect of LH is to stimulateP450c17 expression, while T represses this stimu-
Žlatory effect via the androgen receptor Payne.and Youngblood, 1995 . We have, therefore, stud-
ied the possibility that androgens affect thesteroidogenic capacity of catfish Leydig cells.
Ž .Long-term 2 weeks androgen treatment ofjuvenile catfish resulted in a down-regulatory re-sponse. As in mammals, a decrease in P450c17activity was observed; moreover, the Leydig cells
Žlost half of their mitochondria Cavaco et al.,.1999 . Using the same protocol, i.e. implantation
of steroid-containing silastic pellets, we now showthat down-regulation of basal and LH-stimulated

( )R.W. Schulz et al. � Comparati�e Biochemistry and Physiology Part B 129 2001 407�417 411
ŽFig. 3. Testicular secretion ng�mg testis tissue incubated;.mean � S.E.M., n � 10 of 11�-hydroxyandrostenedione
Ž . Ž . ŽOHA . Basal open columns and LH-stimulated black.columns secretion was quantified using tissue from 10-week-
Ž . Ž .old fish that were implanted 2 top or 4 bottom days earlierŽ .with empty capsules C , or with capsules containing a dose of
Ž . Ž .30 �g of 11-ketotestosterone 11-KT or testosterone T �gbody wt. All experimental procedures were as described in
Ž .Cavaco et al. 1999 . Asterisk indicates a significant differenceŽP�0.05; ANOVA followed by Fisher’s least significance
.difference test to the group implanted with empty capsules.
androgen release is evident after 2 days in juve-Ž .nile males Fig. 3 . The main androgen produced
by African catfish testis tissue is 11�-hydroxy-Ž .androstendione OHA , which is rapidly con-
Ž .verted to 11-ketotestosterone 11-KT in vivo, themain androgen in the blood of African catfish and
Ž .most other male teleosts Cavaco et al., 1997 .In 6�7-month-old adolescent catfish, a differ-
ent type of androgen treatment, namely a single,Žintraperitoneal injection of 11-KT 20 �g�kg body
.wt. that transiently elevated circulating 11-KTlevels for a few hours, up-regulated testicularandrogen production, as tested 1 week laterŽ .Schulz et al., 1997a . Using this short-term treat-ment protocol, the dose of 11-KT was varied. At 1week after treatment with 4 �g 11-KT�kg, andro-gen release did not differ from the control groupŽ .Fig. 4a . A dose of 20 �g of 11-KT�kg resultedin an up-regulation as observed previously, whilea dose of 100 �g of 11-KT�kg led to a down-reg-ulation of androgen release in response to the
Žhigher concentrations of LH used in vitro Fig.
.4a . As mentioned above, the major androgenproduced by catfish testis is OHA, which is rapidly
Ž .converted to 11-KT in vivo Cavaco et al., 1997 .We tested if OHA and 11-KT would have similareffects. As expected, no major differences wereobserved 7 days after a single injection of 20
Ž .�g�kg of OHA or 11-KT Fig. 4b . The time-course of the up-regulatory response was investi-gated further by studying the response to 20 �gof 11-KT�kg at 1 and 14 days after the singleinjection. Shortly after injection, both basal andLH-stimulated androgen outputs were up-regu-
Ž .lated Fig. 4c ; this response was still partiallyŽ .visible after 14 days Fig. 4d . We propose that
the 11-KT-triggered, up-regulatory response inadolescent fish reflects their capacity to augmentthe steroidogenic capacity towards the adult state,which is three-fold higher than in adolescent
Ž .males Schulz et al., 1994a . This response, how-ever, seems to be blunted by high doses of 11-KT.
A short-term elevation of LH plasma levels,which resulted in similarly elevated circulating11-KT levels as injection with 20 �g of 11-KT�kg,had a rather different effect: down-regulation of
Ž .the steroidogenic response Schulz et al., 1997a .Apparently, LH treatment triggers an entirelydifferent response mechanism that overrides thattargeted by 11-KT. It therefore seems possiblethat the rather low plasma LH levels found
Žthroughout puberty and adolescence Schulz et.al., 1994b may be required to prevent a down-
regulation.At present we are investigating if the response
to long- and short-term androgen treatment showsdevelopmental changes during puberty. More-over, in the light of the long-term effects ofrather brief changes in hormone blood levels, itwill be interesting to study the dynamics of LHand androgen plasma levels, which may also mod-ulate gonadotropin receptor expression.
5. Spermatogenesis, gonadotropins, andandrogens
Spermatogenesis is a developmental processwith a mitotic, a meiotic, and a spermiogenicphase. Stem cell mitosis provides spermatogonia,which then pass through a species-specific, fixednumber of mitotic cell cycles. During the subse-quent two meiotic cell cycles with only one roundof DNA duplication, the genetic information is
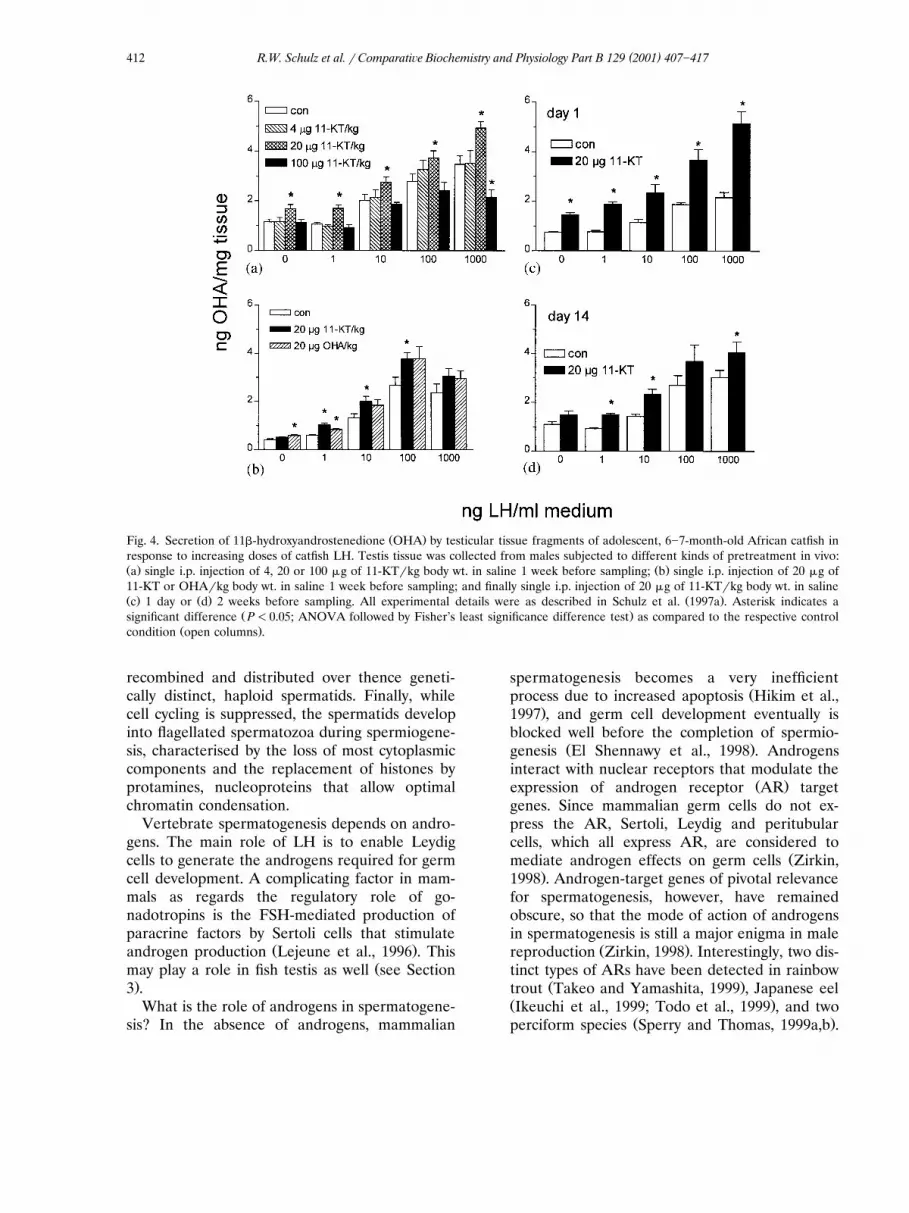
( )R.W. Schulz et al. � Comparati�e Biochemistry and Physiology Part B 129 2001 407�417412
Ž .Fig. 4. Secretion of 11�-hydroxyandrostenedione OHA by testicular tissue fragments of adolescent, 6�7-month-old African catfish inresponse to increasing doses of catfish LH. Testis tissue was collected from males subjected to different kinds of pretreatment in vivo:Ž . Ž .a single i.p. injection of 4, 20 or 100 �g of 11-KT�kg body wt. in saline 1 week before sampling; b single i.p. injection of 20 �g of11-KT or OHA�kg body wt. in saline 1 week before sampling; and finally single i.p. injection of 20 �g of 11-KT�kg body wt. in salineŽ . Ž . Ž .c 1 day or d 2 weeks before sampling. All experimental details were as described in Schulz et al. 1997a . Asterisk indicates a
Ž .significant difference P�0.05; ANOVA followed by Fisher’s least significance difference test as compared to the respective controlŽ .condition open columns .
recombined and distributed over thence geneti-cally distinct, haploid spermatids. Finally, whilecell cycling is suppressed, the spermatids developinto flagellated spermatozoa during spermiogene-sis, characterised by the loss of most cytoplasmiccomponents and the replacement of histones byprotamines, nucleoproteins that allow optimalchromatin condensation.
Vertebrate spermatogenesis depends on andro-gens. The main role of LH is to enable Leydigcells to generate the androgens required for germcell development. A complicating factor in mam-mals as regards the regulatory role of go-nadotropins is the FSH-mediated production ofparacrine factors by Sertoli cells that stimulate
Ž .androgen production Lejeune et al., 1996 . ThisŽmay play a role in fish testis as well see Section
.3 .What is the role of androgens in spermatogene-
sis? In the absence of androgens, mammalian
spermatogenesis becomes a very inefficientŽprocess due to increased apoptosis Hikim et al.,
.1997 , and germ cell development eventually isblocked well before the completion of spermio-
Ž .genesis El Shennawy et al., 1998 . Androgensinteract with nuclear receptors that modulate the
Ž .expression of androgen receptor AR targetgenes. Since mammalian germ cells do not ex-press the AR, Sertoli, Leydig and peritubularcells, which all express AR, are considered to
Žmediate androgen effects on germ cells Zirkin,.1998 . Androgen-target genes of pivotal relevance
for spermatogenesis, however, have remainedobscure, so that the mode of action of androgensin spermatogenesis is still a major enigma in male
Ž .reproduction Zirkin, 1998 . Interestingly, two dis-tinct types of ARs have been detected in rainbow
Ž .trout Takeo and Yamashita, 1999 , Japanese eelŽ .Ikeuchi et al., 1999; Todo et al., 1999 , and two
Ž .perciform species Sperry and Thomas, 1999a,b .

( )R.W. Schulz et al. � Comparati�e Biochemistry and Physiology Part B 129 2001 407�417 413
Moreover, androgens appear to be present inŽtrout and rat germ cells Schulz and Blum, 1988;¨
.Schulz et al., 1989 , and the presence of andro-gen-binding, germ cell membrane-anchored gan-
Žgliosides is required for fertility Takamiya et al.,.1998 . These selected findings illustrate that more
than one androgen receptive system may be oper-ating in the germinal epithelium.
Ž .In immature European Colombo et al., 1987Ž .and Japanese eel Miura et al., 1991a , treatment
with the LH-like hCG resulted in Leydig andthen Sertoli cell activation, followed by spermato-gonial proliferation, meiosis and spermiogenesis.In the Japanese eel, hCG also stimulates sper-matogenesis under tissue culture conditions, 11-KT being capable of replacing hCG as regards
Žthe induction of full spermatogenesis Miura et. Žal., 1991b . In the rainbow trout Oncorhynchus
.mykiss , on the contrary, 11-KT or other steroidŽ .hormones had no such effect Loir, 1999 , al-
though both gonadotropins stimulated germ cellproliferation in isolated spermatogonia�Sertoli
Ž .cell clusters spermatocysts . This suggests that amore complex stimulus than 11-KT alone may berequired in the trout.
In mammals, FSH ensures the survival andstimulates the proliferation of differentiatingspermatogonia, which often involves Sertoli cell-derived growth factor production, e.g. activinŽ . ŽMather et al., 1990 or stem cell factor Loveland
.and Schlatt, 1997 . Moreover, FSH-R-dependentsignalling plays a significant role in nuclear con-
Ždensation during spermiogenesis Krishnamurthy.et al., 2000 . A specific role for FSH in fish
spermatogenesis has not yet been found, butgrowth factor production may also be an FSH-regulated aspect of Sertoli cell function in fish. Inthe Japanese eel, a stimulatory effect on sper-matogonial proliferation may be mediated by Ser-
Ž .toli cell-derived activin Miura et al., 1995, 1999 .In rainbow trout, insulin-like growth factor-IŽ .IGF-I stimulates the proliferation of spermato-
Ž .gonia Loir, 1999 , which express IGF-I receptorsŽ .Loir and LeGac, 1994 . In the Japanese eel,IGF-I is a necessary, permissive factor for 11-KTaction, which otherwise fails to stimulate sper-
Ž .matogenesis in tissue culture Nader et al., 1999 .In another salmonid, and in the medaka, fibro-
Ž .blast growth factor FGF may play a role forŽspermatogonial proliferation Watanabe and Oni-
.take 1995; Watanabe et al., 1997 ; the FGF familyof growth factors and their receptors are promi-
Žnently expressed in the mammalian testis Cancilla.et al., 2000 . In all these cases, the possible role of
FSH seems unexplored in the piscine testis.It appears that testicular physiology is not fully
accounted for by traditional endocrine paradigms.Rather, spermatogenesis depends on regulatory
Ž .inputs from distant sources classical endocrine ,Žfrom local sources e.g. growth factors and
.steroids , as well as from the germ cell’s genomeŽ .Franca et al., 1998 , which � in mammals �dominates the timing of the developmental stepstaken by the germ cells. This setting results in adynamic microenvironment that is adjusted ac-cording to the developmentally changing require-ments of germ cells undergoing spermatogenesis.
6. Steroid feedback, gonadotrophs and GnRHneurones
Sex steroids and gonadotropin-releasing hor-Ž .mones GnRHs are important factors regulating
the activity of pituitary gonadotrophs. In addition,sex steroids also modulate the activity of GnRHneurones, and thus regulate gonadotrophs bothdirectly and indirectly.
A distinct cellular source for the two go-nadotropins, such as that found in a number of
Ž .fish species e.g. Nozaki et al., 1990 , may facili-tate the distinct regulation of gonadotropin syn-thesis and secretion. While a large body of infor-mation is available with respect to LH, assays toquantify FSH are restricted to salmonids, and
Ž . Ž .testosterone T and estradiol-17 E reduce FSH2Žplasma levels in immature coho Larsen and
.Swanson, 1997; Dickey and Swanson, 1998 andŽ .Atlantic salmon Borg et al., 1998 . Although
information on FSH �-subunit mRNA levels isavailable from more species, it seems too early todiscuss the physiological significance of these data.This is exemplified by the above-mentioned stud-ies on coho salmon, where FSH transcript andprotein levels have been quantified in parallel.Both T and E reduced plasma FSH, but only E2 2reduced FSH transcript levels. Moreover, the ef-fect on transcript levels faded out during matura-tion, while the negative feedback on FSH plasmalevels was maintained.
Aromatisable androgens and estrogens havebeen shown to activate LH �-subunit gene ex-pression and to increase pituitary LH proteinlevels in juveniles of several species. The Euro-

( )R.W. Schulz et al. � Comparati�e Biochemistry and Physiology Part B 129 2001 407�417414
pean eel is a noteworthy exception, since andro-Ž .gens including non-aromatisable ones , but not
ŽE , stimulated LH gene expression Huang et al.,2.1997 . The molecular mechanism in salmonids
involves the oestrogen receptor and anothermember of the nuclear receptor family, the or-
Ž .phan receptor SF-1 Le Drean et al., 1996 . How-´ever, the steroid-dependent stimulation fades outduring maturation. In African catfish, a strongincrease in pituitary LH content is found after E2
Ž .treatment of juvenile males Cavaco et al., 1995 .In adult males, E has no such effect, whereas2the response to E is again found after castration2Ž .Teves et al., unpublished data , or in primary
Ž .pituitary cell cultures Rebers et al., 2000a , bothconditions depriving the adult gonadotroph ofgonadal influences. It is possible that LH geneexpression is already stimulated maximally in ad-vanced males and�or that testicular factors inter-fere with the stimulation of LH synthesis in adult,but not in juvenile, males.
Aromatisable androgens and�or 11-KT havebeen shown to stimulate GnRH neurones in dif-ferent teleost species. In juvenile African catfish,steroid effects were rather selective, in that onlyT, but neither E nor 11-oxygenated androgens,2induced a precocious maturation of the catfishŽ .cf GnRH-producing neurones in the ventralforebrain, while the mesencephalic chickenGnRH-II-producing neurones did not appear to
Žbe affected by these treatments Dubois et al.,.1998 . In addition to an increased number of
immunoreactive cfGnRH neurones and a moreintense staining of perikarya and axons, treatmentwith T reduced circulating LH levels in bothjuvenile and adult males. This may indicate thatpart of the increased immunoreactivity in theGnRH neurones is related to accumulation ofcfGnRH when assuming a reduced release of thepeptide.
Next to its namesake function as gonadotropinsecretagogue, GnRH is a major stimulator of
Žgonadotropin synthesis in mammals e.g. Kaiser.et al., 1997 . We have been unable, however, to
find evidence for a direct stimulation of LH pro-duction by GnRH in mature male African catfishŽ .Rebers and Schulz, unpublished observations .In tilapia, however, GnRH prolongs, via cAMP,
Žthe half-life of the LH �-subunit Melamed et al.,.1996 , and other stimulatory mechanisms may
operate in other teleost species. It seems that, atleast in the African catfish, GnRH has no direct
role in the stimulation of LH synthesis. Steroidhormones dominate this area, while the role ofGnRH as secretagogue seems fully developed. Inthis role, the two endogenous GnRHs modulate
Žeach other’s LH release activity in catfish Bosma.et al., 2000 . Moreover, a GnRH-triggered mech-
anism appears to be present that amplifiesŽ .cfGnRH release Rebers et al., 2000b . This may
be of relevance for the natural dynamics of circu-lating reproductive hormones. In female rainbowtrout, a regular pulsatility, such as recorded inmammals, was not found, although gonadotropin
Ž .secretion bursts can occur Zohar et al., 1986 .Finally, GnRH release bursts may be of relevancefor the pituitary’s GnRH sensitivity in contextwith the homologous down-regulation of the pitu-
Ž .itary GnRH receptor Blomenrohr et al., 1999 ,¨such that a strong stimulation may result in apartial refractory condition.
Acknowledgements
JEBC and EMC gratefully acknowledge thesupport from ‘PRAXIS XXI’ J.N.I.C.T.-Portugalgrants.
References
Blomenrohr, M., Heding, A., Sellar, R. et al., 1999.¨Pivotal role for the cytoplasmic carboxyl-terminaltail of a non-mammalian gonadotropin-releasinghormone receptor in cell surface expression, ligandbinding, and receptor phosphorylation and internal-ization. Mol. Pharmacol. 56, 1229�1237.
Bogerd, J., Andersson, E., Blomenrohr, M. et al., 1999.¨Discrepancy between structural characteristics andligand specificity of an African catfish gonadotropinreceptor. In: Norberg, B., Kjesbu, O.S., Taranger,
Ž .G.L., Andersson, E., Stefansson, S.O. Eds. ,Proceedings of the Sixth International Symposiumon the Reproductive Physiology of Fish, FishSymp99, Bergen, pp. 167�169.
Borg, B., Antonopoulou, E., Mayer, I., Andersson, E.,Berglund, I., Swanson, P., 1998. Effects of gonadec-tomy and androgen treatments on pituitary andplasma levels of gonadotropins in mature male At-lantic salmon, Salmo salar, parr � positive feedbackcontrol of both gonadotropins. Biol. Reprod. 58,814�820.
Bosma, P.T., Rebers, F.E.M., van Dijk, W., Willems,P.H.G.M., Goos, H.J.Th., Schulz, R.W., 2000. Inhibi-tory and stimulatory interactions between en-dogenous gonadotropin-releasing hormones in the

( )R.W. Schulz et al. � Comparati�e Biochemistry and Physiology Part B 129 2001 407�417 415
Ž .African catfish Clarias gariepinus . Biol. Reprod. 62,731�738.
Braun, T., Schofield, P.R., Sprengel, R., 1991. Amino-terminal leucine-rich repeats in gonadotropin recep-tors determine hormone selectivity. EMBO J. 10,1885�1890.
Cancilla, B., Davies, A., Ford-Perriss, M., Risbridger,G.P., 2000. Discrete cell- and stage-specific localisa-tion of fibroblast growth factors and receptor expres-sion during testis development. J. Endocrinol. 164,149�159.
Cavaco, J.E.B., Lambert, J.G.D., Schulz, R.W., Goos,H.J.T., 1995. Will puberty be regulated by sexual
Ž .steroids in male African catfish Clarias gariepinus ?Ž .In: Goetz, F.W., Thomas, P. Eds. , Proceedings of
the Fifth International Symposium on the Reproduc-tive Physiology of Fish. FishSymp 95, Austin, p. 360.
Cavaco, J.E.B., Vischer, H.F., Lambert, J.G.D., Goos,H.J.T., Schulz, R.W., 1997. Mismatch between pat-terns of circulating and testicular androgens inAfrican catfish, Clarias gariepinus. Fish Physiol.Biochem. 17, 155�162.
Cavaco, J.E.B., van Blijswijk, B., Leatherland, J.F.,Goos, H.J.Th., Schulz, R.W., 1999. Androgen-in-duced changes in Leydig cell ultrastructure andsteroidogenesis in juvenile African catfish, Clariasgariepinus. Cell Tissue Res. 297, 291�299.
Colombo, G., Grandi, G., Romeo, A. et al., 1987. Testiscytological structure, plasma sex steroids, and gonadcytosol free steroid receptors of heterologous go-
Ž .nadotropin hCG -stimulated silver eel, Anguilla an-guilla L. Gen. Comp. Endocrinol. 65, 167�178.
Dickey, J.T., Swanson, P., 1998. Effects of sex steroidsŽ .on gonadotropin FSH and LH regulation in coho
Ž .salmon Oncorhynchus kisutch . J. Mol. Endocrinol.21, 291�306.
Dubois, E.A., Florijn, M.A., Zandbergen, M.A., Peute,J., Goos, H.J.Th., 1998. Testosterone accelerates thedevelopment of the catfish GnRH system in the
Ž .brain of immature African catfish Clarias gariepinus .Gen. Comp. Endocrinol. 112, 383�393.
El Shennawy, A., Gates, R.J., Russell, L.D., 1998. Hor-monal regulation of spermatogenesis in the hypophy-sectomized rat: cell viability after hormonal replace-ment in adults after intermediate periods of hy-pophysectomy. J. Androl. 19, 320�334.
Franca, L.R., Ogawa, T., Avarbock, M.R., Brinster,R.L., Russell, L.D., 1998. Germ cell genotype con-trols cell cycle during spermatogenesis in the rat.Biol. Reprod. 59, 1371�1377.
Gomez, J.M., Weil, C., Ollitrault, M., LeBail, P.Y.,Ž .Breton, B., LeGac, F., 1999. Growth hormone GH
and gonadotropin subunit gene expression and pitu-itary and plasma changes during spermatogenesis
Žand oogenesis in rainbow trout Oncorhynchus.mykiss . Gen. Comp. Endocrinol. 113, 413�428.
Gouvoroun, M.S., Huet, J.-C., Pernollet, J.-C., Breton,B., 1997. Use of immobilized metal ion affinity chro-matography and dye�ligand chromatography for theseparation and of rainbow trout pituitary go-nadotropins, GTH I and GTH II. J. Chromatogr. B698, 35�46.
Hassin, S., Holland, M.C.H., Zohar, Y., 1999. Ontogenyof follicle-stimulating hormone and luteinizing hor-mone gene expression during pubertal development
Žin the female striped bass, Morone saxatilis Tele-.ostei . Biol. Reprod. 61, 1608�1615.
Hikim, A.P.S., Rajavashisth, T.B., Hikim, I.S. et al.,1997. Significance of apoptosis in the temporal andstage-specific loss of germ cells in the adult rat aftergonadotropin deprivation. Biol. Reprod. 57,1193�1201.
Huang, Y.S., Schmitz, M., Le Belle, N., Chang, C.F.,Querat, B., Dufour, S., 1997. Androgens stimulategonadotropin-II �-subunit in eel pituitary cells invitro. Mol. Cell. Endocrinol. 131, 157�166.
Ikeuchi, T., Todo, T., Kobayashi, T., Nagahama, Y.,1999. cDNA cloning of a novel androgen receptorsubtype. J. Biol. Chem. 274, 25205�25209.
Kaiser, U.B., Jakubowiak, A., Steinberger, A., Chin,W.W., 1997. Differential effects of gonadotropin-
Ž .releasing hormone GnRH pulse frequency on go-nadotropin subunit and GnRH receptor messengerribonucleic acid levels in vitro. Endocrinology 138,1224�1231.
Koide, Y., Noso, T., Schouten, G. et al., 1992. Matura-tional gonadotropin from the African catfish, Clariasgariepinus. Purification, characterisation, localisationand biological activity. Gen. Comp. Endocrinol. 87,327�341.
Krishnamurthy, H., Danilovich, N., Morales, C.R.,Sairam, M.R., 2000. Qualitative and quantitative de-cline in spermatogenesis of the follicle-stimulating
Ž .hormone receptor knockout FORKO mouse. Biol.Reprod. 62, 1146�1159.
Larsen, D.A., Swanson, P., 1997. Effects of gonadec-tomy on plasma gonadotropins I and II in cohosalmon, Oncorhynchus kisutch. Gen. Comp. En-docrinol. 108, 152�160.
Le Drean, Y., Liu, D., Wong, A.O.L., Xiong, F., Hew,´C.L., 1996. Steroidogenic factor 1 and estradiol re-ceptor act in synergism to regulate the expression ofthe salmon gonadotropin II � subunit gene. Mol.Endocrinol. 10, 217�229.
Lejeune, H., Chuzel, F., Thomas, T. et al., 1996.Paracrine regulation of Leydig cells. Ann. En-docrinol. 57, 55�63.
Loir, M., LeGac, F., 1994. Insulin-like growth factor-Iand -II binding and action on DNA synthesis inrainbow trout spermatogonia and spermatocytes.Biol. Reprod. 51, 1154�1163.

( )R.W. Schulz et al. � Comparati�e Biochemistry and Physiology Part B 129 2001 407�417416
Loir, M., 1999. Spermatogonia of rainbow trout: II. Invitro study of the influence of pituitary hormones,growth factors and steroids on mitotic activity. Mol.Reprod. Dev. 53, 434�442.
Loveland, K.L., Schlatt, S., 1997. Stem cell factor andc-kit in the mammalian testis: lessons originatingfrom Mother Nature’s gene knockouts. J. En-docrinol. 153, 337�344.
Mather, J.P., Attie, K.M., Woodruff, T.K., Rice, G.C.,Phillips, D.M., 1990. Activin stimulates spermatogo-nial proliferation in germ-Sertoli cell cocultures fromimmature rat testis. Endocrinology 127, 3206�3214.
Melamed, P., Gur, G., Elizur, A. et al., 1996. Differen-tial effects of gonadotropin-releasing hormone, do-pamine and somatostatin and their second messen-gers on the mRNA levels of gonadotropin II �subunit and growth hormone in the teleost fish,Tilapia. Neuroendocrinology 64, 320�328.
Miura, T., Yamauchi, K., Nagahama, Y., Takahashi, H.,1991a. Induction of spermatogenesis in maleJapanese eel, Anguilla japonica, by a single injectionof human chorionic gonadotropin. Zool. Sci. 8, 63�73.
Miura, T., Yamauchi, K., Takahashi, H., Nagahama, Y.,1991b. Hormonal induction of all stages of sper-matogenesis in vitro in the male Japanese eelŽ .Anguilla japonica . Proc. Natl. Acad. Sci. USA 88,5774�5778.
Miura, T., Miura, C., Yamauchi, K., Nagahama, Y.,1995. Human recombinant activin induces prolifera-tion of spermatogonia in vitro in the Japanese eelAnguilla japonica. Fish. Sci. 61, 434�437.
Miura, C., Miura, T., Kudo, N., Yamashita, M., Ya-mauchi, K., 1999. cDNA cloning of a stage-specificgene expressed during HCG-induced spermatogene-sis in the Japanese eel. Develop. Growth Differ. 41,463�471.
Miwa, S., Yan, L., Swanson, P., 1994. Localization oftwo gonadotropin receptors in the salmon gonad byin vitro ligand autoradiography. Biol. Reprod. 50,629�642.
Nader, M.R., Miura, T., Ando, N., Miura, C., Ya-mauchi, K., 1999. Recombinant human insulin-likegrowth factor I stimulates all stages of 11-keto-testosterone-induced spermatogenesis in theJapanese eel, Anguilla japonica, in vitro. Biol. Re-prod. 61, 944�947.
Nozaki, M., Naito, N., Swanson, P. et al., 1990. Sal-monid pituitary gonadotrophs. II. Ontogeny of GTH
ŽI and GTH II cells in the rainbow trout Salmo.gairdneri irideus . Gen. Comp. Endocrinol. 77,
358�367.Oba, Y., Hirai, T., Yoshiura, Y., Yoshikuni, M.,
Kawauchi, H., Nagahama, Y., 1999a. The duality offish gonadotropin receptors: Cloning and functionalcharacterization of a second gonadotropin receptorcDNA expressed in the ovary and testis of amago
Ž .salmon Oncorhynchus rhodurus . Biochem. Biophys.Res. Commun. 265, 366�371.
Oba, Y., Hirai, T., Yoshiura, Y., Yoshikuni, M.,Kawauchi, H., Nagahama, Y., 1999b. Cloning, func-tional characterization, and expression of a go-nadotropin receptor cDNA in the ovary and testis of
Ž .amago salmon Oncorhynchus rhodurus . Biochem.Biophys. Res. Commun. 263, 584�590.
Payne, A.H., Youngblood, G.L., 1995. Regulation ofexpression of steroidogenic enzymes in Leydig cells.Biol. Reprod. 52, 217�225.
Planas, J.V., Swanson, P., 1995. Maturation-associatedchanges in the response of the salmon testis to the
Žsteroidogenic actions of gonadotropins GTH I and.GTH II in vitro. Biol. Reprod. 52, 697�704.
Prat, F., Sumpter, J.P., Tyler, C.R., 1996. Validation ofradioimmunoassays for two salmon gonadotropinsŽ .GTH I and GTH II and their plasma concentra-tions throughout the reproductive cycle in male and
Ž .female rainbow trout Oncorhynchus mykiss . Biol.Reprod. 54, 1375�1382.
Rebers, F.E.M., Hassing, G.A.M., Zandbergen, M.A.,Goos, H.J.T., Schulz, R.W., 2000a. Regulation ofsteady-state luteinizing hormone messenger ribonu-cleic acid levels, de novo synthesis, and release bysex steroids in primary pituitary cell cultures of maleAfrican catfish, Clarias gariepinus. Biol. Reprod. 62,864�872.
Rebers, F.E.M., Bosma, P.T., van Dijk, W., Goos,H.J.Th., Schulz, R.W., 2000b. GnRH stimulates LHrelease directly via inositol phosphate and indirectlyvia cAMP in African catfish. Am. J. Physiol. 278,R1572�R1578.
Santos, E.M., Rand-Weaver, M., Tyler, C.R., 2000. FSHand its subunits in rainbow trout: purification, char-acterization, radioimmunoassays and plasma and pi-tuitary seasonal profiles in females. Comp. Biochem.Physiol., in press.
Schulz, R., Blum, V., 1988. Testosterone immunoreac-¨Ž .tivity in rainbow trout Salmo gairdneri Richardson
testis. Gen. Comp. Endocrinol. 72, 80�89.Schulz, R., Paris, F., Lembke, P.J., Blum, V., 1989.¨
Testosterone immunoreactivity in the seminiferousepithelium of rat testis: effect of treatment withethane dimethanesulfonate. J. Histochem. Cy-tochem. 37, 1667�1673.
Schulz, R.W., van der Corput, L., Janssen-Dommerholt,J., Goos, H.J.T., 1994a. Sexual steroids during pu-
Ž .berty in male African catfish Clarias gariepinus :serum levels and gonadotropin-stimulated testicular
� �secretion in vitro. J. Comp. Physiol. B 164, 195�205.Schulz, R.W., van der Sanden, M.C.A., Bosma, P.T.,
Goos, H.J.T., 1994b. Effects of gonadotrophin-re-leasing hormone during the pubertal development of
Ž .the male African catfish Clarias gariepinus : go-

( )R.W. Schulz et al. � Comparati�e Biochemistry and Physiology Part B 129 2001 407�417 417
nadotrophin and androgen levels in plasma. J. En-docrinol. 140, 265�273.
Schulz, R.W., Bogerd, J., Bosma, P.T. et al., 1995a.Physiological, morphological, and molecular aspectsof gonadotropins in fish with special reference to theAfrican catfish, Clarias gariepinus. In: Goetz, F.W.,
Ž .Thomas, P. Eds. , Proceedings of the Fifth Interna-tional Symposium on the Reproductive Physiology ofFish, FishSymp 95, Austin, pp. 2�6.
Schulz, R.W., Renes, I.B., Zandbergen, M.A., van Dijk,W., Peute, J., Goos, H.J.Th., 1995b. Pubertal devel-
Ž .opment of male African catfish Clarias gariepinus .Pituitary ultrastructure and responsiveness to go-nadotropin-releasing hormone. Biol. Reprod. 53,940�950.
Schulz, R.W., van der Wind, F., Janssen-Dommerholt,C. et al., 1997a. Modulation of testicular androgen
Žproduction in adolescent African catfish Clarias.gariepinus . Gen. Comp. Endocrinol. 108, 56�66.
Schulz, R.W., Zandbergen, M.A., Peute, J., Bogerd, J.,van Dijk, W., Goos, H.J.T., 1997b. Pituitary go-nadotrophs are strongly activated at the beginning ofspermatogenesis in African catfish, Clarias gariepi-nus. Biol. Reprod. 57, 139�147.
Sohn, Y.C., Yoshiura, Y., Kobayashi, M., Aida, K.,1999. Seasonal changes in mRNA levels of go-nadotropin and thyrotropin subunits in the goldfish,Carassius auratus. Gen. Comp. Endocrinol. 113,436�444.
Sperry, T.S., Thomas, P., 1999a. Identification of twoŽnuclear androgen receptors in kelp bass Paralabrax
.clathratus and their binding affinities for xenobi-Žotics: comparison with Atlantic croaker Micro-
.pogonias undulatus androgen receptors. Biol. Re-prod. 61, 1152�1161.
Sperry, T.S., Thomas, P., 1999b. Characterization oftwo nuclear androgen receptors in Atlantic croaker:comparison of their biochemical properties andbinding specificities. Endocrinology 140, 1602�1611.
Stocco, D.M., Clark, B.J., 1996. Regulation of the acuteproduction of steroids in steroidogenic cells. En-docrinol. Rev. 17, 221�244.
Swanson, P., Bernard, M., Nozaki, M., Suzuki, K.,Kawauchi, H., Dickhoff, W.W., 1989. GonadotropinsI and II in juvenile coho salmon. Fish Physiol.Biochem. 7, 169�176.
Swanson, P., Suzuki, K., Kawauchi, H., Dickhoff, W.W.,1991. Isolation and characterization of two cohosalmon gonadotropins, GTH I and GTH II. Biol.Reprod. 44, 29�38.
Takamiya, K., Yamamoto, A., Furukawa, K. et al.,1998. Complex gangliosides are essential in sper-matogenesis of mice: possible roles in the transport
of testosterone. Proc. Natl. Acad. Sci. USA 95,12147�12152.
Takeo, J., Yamashita, S., 1999. Two distinct isoforms ofcDNA encoding rainbow trout androgen receptors.J. Biol. Chem. 274, 5674�5680.
Thotakura, N.R., Blithe, D.L., 1995. Glycoprotein hor-mones � glycobiology of gonadotropins, thyrotropin
Ž .and free �-subunit. Glycobiology 5 1 , 3�10.Tilly, J.L., Aihara, T., Nishimori, K. et al., 1992. Expres-
sion of recombinant human follicle-stimulating hor-mone receptor: species-specific ligand binding, signaltransduction, and identification of multiple ovarianmessenger ribonucleic acid transcripts. Endo-crinology 131, 799�806.
Todo, T., Ikeuchi, T., Kobayashi, T., Nagahama, Y.,1999. Fish androgen receptor: cDNA cloning, steroidactivation of transcription in transfected mammaliancells, and tissue mRNA levels. Biochem. Biophys.Res. Commun. 254, 378�383.
van den Hurk, R., Peute, J., 1985. Functional aspects ofthe postovulatory follicle in the ovary of the Africancatfish, Clarias gariepinus, after induced ovulation.Cell Tissue Res. 240, 199�208.
Watanabe, A., Onitake, K., 1995. Changes in the dis-tribution of fibroblast growth factor in the teleosteantestis during spermatogenesis. J. Exp. Zool. 272,475�483.
Watanabe, A., Hatakeyama, N., Yasuoka, A., Onitake,K., 1997. Distributions of fibroblast growth factorand the mRNA for its receptor, MFR1, in the devel-oping testis of the medaka, Oryzias latipes. J. Exp.Zool. 279, 177�184.
Weltzien, F.-A., Andersen, O., Kobayashi, T., Swanson,P., Norberg, B., 1999. FSH and LH from pituitary
Žglands of Atlantic halibut Hippoglossus hippoglossus.L. . In: Norberg, B., Kjesbu, O.S., Taranger, G.L.,
Ž .Andersson, E., Stefansson, S.O. Eds. , Proceedingsof the Sixth International Symposium on the Repro-ductive Physiology of Fish, FishSymp 99, Bergen, pp.475�477.
Yoshiura, Y., Suetake, H., Aida, K., 1999. Duality ofgonadotropin in a primitive teleost, Japanese eelŽ .Anguilla japonica . Gen. Comp. Endocrinol. 114,121�131.
Zirkin, B.R., 1998. Spermatogenesis: its regulation bytestosterone and FSH. Semin. Cell. Dev. Biol. 9,417�421.
Zohar, Y., Breton, B., Fostier, A., 1986. Short-termprofiles of plasma gonadotropin and estradiol-17�levels in the female rainbow trout, from early ovar-ian recrudescence and throughout vitellogenesis.Gen. Comp. Endocrinol. 64, 172�188.





























