Embed Size (px)
Citation preview

Ecotoxicology and Environmental Safety ] (]]]]) ]]]–]]]
Contents lists available at SciVerse ScienceDirect
Ecotoxicology and Environmental Safety
0147-65
http://d
n Corr
Jardim
990, Cu
E-m
PleasEcoto
journal homepage: www.elsevier.com/locate/ecoenv
Genotoxic evaluation of different doses of methylmercury (CH3Hgþ) inHoplias malabaricus
Taynah Vicari a,n, Marcos Vinıcius Mocellin Ferraro a, Wanessa Algarte Ramsdorf a, Maritana Mela b,Ciro Alberto de Oliveira Ribeiro b, Marta Margarete Cestari a
a Departamento de Genetica, Laboratorio de Mutagenese Ambiental, Universidade Federal do Parana (UFPR), Curitiba, PR, Brazilb Departamento de Biologia Celular, Laboratorio de Toxicologia Celular, Universidade Federal do Parana (UFPR), Curitiba, PR, Brazil
a r t i c l e i n f o
Article history:
Received 19 June 2011
Received in revised form
28 April 2012
Accepted 14 May 2012
13/$ - see front matter & 2012 Elsevier Inc. A
x.doi.org/10.1016/j.ecoenv.2012.05.007
esponding author at: Depto de Genetica, S
das Americas, UFPR, Centro Politecnico, Caix
ritiba, PR, Brazil. Fax: þ55 41 3361 1793.
ail address: [email protected] (T. V
e cite this article as: Vicari, T., et al.,xicol. Environ. Saf. (2012), http://dx
a b s t r a c t
The survey of the effects of toxic metals and its organic compounds are of critical importance because
these compounds tend to accumulate in aquatic environments. In the present work, the genotoxic
potential of methylmercury, an organomercurial compound with high toxicity and present in large
amounts in fish of the Amazon due to the mining process, was evaluated using the piscine
micronucleus test and comet assay in fish. Specimens of Hoplias malabaricus (popularly known as
traira), a neotropical specie, was exposed to methylmercury via food web, over 70 days, in two different
concentrations: two groups of fifteen fish were tested with 0.075 mg CH3Hg+/g and 0.75 mg CH3Hg+/g of
methylmercury per gram of fish, at 5-day intervals and over 14 successive intervals whereas control
group, composed by nine fish, was fed by uncontaminated prey fish (Astyanax sp). One of the aims of
this study was to reproduce the contamination found in nature in an attempt to increase our biological
knowledge on the neotropical species. Hoplias malabaricus specimens were then anesthetized for
removal of blood samples and dissected, for cephalic kidney removal. As a result, it was observed that
the piscine micronucleus test showed no significant differences between exposed groups and control
group. The comet assay performed on erythrocyte system cells, showed a significant difference between
controls and contaminated, but there was no difference between doses. In contrast, the kidney cells
comet assay showed no significant difference between groups, probably due to the type of damage
caused by xenobiotics to be related to the tissues of most contact with it, as well as its mode of action
and the chain of bioaccumulation within bodies.
& 2012 Elsevier Inc. All rights reserved.
1. Introduction
The aquatic environment is the ultimate destination of pollu-tants produced by natural sources and by anthropogenic action,and the accumulation and persistence of these substances in theenvironment constitutes a threat to life (Fleeger et al., 2003).Among these pollutants, mercury is considered to be extremelyhazardous to the aquatic environment due to its high toxicity,bioaccumulation, and other deleterious effects on biota, includinggenetic effects and mutagenicity (WHO, 1990). Mercury is aubiquitous contaminant in the environment and is distributedglobally through natural geological emissions as well as throughanthropogenic activities, such as fuel burning, mining, smeltingoperations and the chlor-alkali industry (Wren et al., 1995).
ll rights reserved.
etor de Ciencias Biologicas,
a Postal: 19071, CEP 81531-
icari).
Genotoxic evaluation of dif.doi.org/10.1016/j.ecoenv.2
Inorganic forms of mercury are the most common forms inaquatic ecosystems, and they are converted into organic forms,such as methylmercury (CH3Hgþ), via biogeochemical processes(Guilherme et al., 2008) or via bacterial activity. These organicforms of mercury easily cross the blood–brain barrier (Mela et al.,2007). All forms of mercury are extremely toxic; however,methylmercury is a great concern, particularly as a water con-taminant due to its lipophilicity (Guilherme et al., 2008) and itslarge number of potential molecular targets within the body dueto SH reactive groups in several proteins and membrane consti-tuents (Vassalo et al., 1996).
Mercury is still used in gold mining activities in northernBrazil, primarily in the Amazon region (Pfeiffer et al., 1989;Bidone et al., 1997). Forest fires and soil erosion have also beenadvocated as sources of mercury pollution in the region (Rouletet al., 2001), making the intake of food contaminated withmethylmercury one of the first routes of exposure for fish(Oliveira Ribeiro et al., 1996, 1999; Limke et al., 2004; Drevnicket al., 2006). Several studies have been published on the Amazonrivers basin showing great concern regarding the contamination
ferent doses of methylmercury (CH3Hgþ) in Hoplias malabaricus.012.05.007

T. Vicari et al. / Ecotoxicology and Environmental Safety ] (]]]]) ]]]–]]]2
of the aquatic food chain and the riverside population withmercury and its compounds (Porto et al., 2005; Fostier et al.,2000; Barbosa et al., 1995; Kehrig et al., 1998; Silva-Forsberget al., 1999).
The species Hoplias malabaricus is a neotropical fish with awide distribution ranging from Panama to the south of Argentina.In the Amazon, this fish is widely used in the diet of the riversidepopulation (Grandjean, 1999). The species is essentially piscivor-ous, even in the earlier stages of its development, and it may haveexceptional starvation resistance (Loureiro and Hahn, 1996).Thus, being a top predator carnivorous fish, these animals becomean important route of mercury uptake by human populationsbecause methylmercury is easily biomagnified along the foodchains of aquatic ecosystems (Lebel et al., 1998; Aschner, 2002).Although many studies have been published regarding the effect,accumulation, distribution and speciation of mercury in non-tropical fish species, little information regarding methylmercurycontamination in neotropical fish has been published. Previously,Ferraro and colleagues worked with sub-chronic trophic contam-ination of H. malabaricus (13 trophic doses in a period of 60 day)with Pb2þ and tributyltin (TBT) (Ferraro et al., 2004), andRamsdorf and colleagues used intra-peritoneal injections of twodifferent doses of inorganic lead in H. malabaricus, indicating thatH. malabaricus represents an excellent experimental model forgenotoxicity studies (Ramsdorf et al., 2009).
Current awareness of the potential hazards of metals in theaquatic environment has stimulated much interest in the study offish, which is generally considered to be the most feasibleorganism for pollution monitoring of environmental carcinogens,teratogens and mutagens in aquatic systems (Van der Oost et al.,2003). The DNA molecule is a target site of most, if not all,carcinogenic and mutagenic agents. Growing concern has devel-oped regarding the presence of genotoxic and mutagenic com-pounds in the environment; therefore, it became necessary todevelop methods to detect these effects in aquatic organisms.These approaches include a method to assay the presence of DNAadducts, a chromosome aberration test, a test for DNA breakage, ameasurement of the frequency of micronuclei and other nuclearmorphological changes (Bombail et al., 2001), and more recently,the detection of oxidative damage directly on the DNA molecule(Oliveira et al., 2010).
The micronucleus test is considered to be one of the most usefulmethods for evaluating genotoxicity in aquatic systems. Micro-nuclei are formed by chromosome fragments or whole chromo-somes that lag at cell division due to the lack of a centromere, DNAdamage, or a defect in cytokinesis (Heddle et al., 1991). Thus, theevaluation of micronucleus provides a measure of both theclastogenic and the aneugenic potential of environmental agents.Erythrocytes are the most commonly used cells in the piscinemicronucleus test; however, hepatocytes (C- avas et al., 2005), gillepithelial cells (C- avas et al., 2005), and fin cells (Arkhipchuk andGaranko, 2005) have also been used (C- avas, 2008). Furthermore,the piscine micronucleus test is also used to assess nuclearabnormalities (NAs). Although the mechanisms responsible forNAs have not been fully explained, these abnormalities areconsidered to be indicators of genotoxic damage; therefore, theymay complement the scoring of micronuclei in routine genotoxi-city surveys, which gives greater sensitivity to the analysis ofmicronuclei (C- avas and Ergene-Gozukara, 2005b).
To improve assay sensitivity and to detect nuclear morpholo-gical changes and micronuclei in immature erythrocytes, we usedacridine orange staining in addition to the conventional stainingtechnique (Giemsa). The acridine orange staining technique,commonly used in mammalian micronucleus tests, was firstmodified for use in fish by Ueda et al. (1992) and was successfullyutilized by Hayashi et al. (1998) on fish gill and erythrocyte cells.
Please cite this article as: Vicari, T., et al., Genotoxic evaluation of difEcotoxicol. Environ. Saf. (2012), http://dx.doi.org/10.1016/j.ecoenv.2
In this study, we also used acridine orange staining, to detectmicronuclei in newly formed young erythrocytes to improve thesensitivity of the test system (C- avas and Ergene-Gozukara(2005a)). The frequency of immature erythrocytes was estimatedto evaluate changes in hematological dynamics of copies thatcould be associated with xenobiotics.
Currently, the comet (or single cell gel electrophoresis) assay,is widely used in both the research field and laboratory tests. Thecomet assay detects DNA strand breaks and alkali-labile sites bymeasuring the migration of DNA from immobilized nuclear DNA(Singh et al., 1988). The name comet refers to the formation of along comet-like tail of DNA fragments due to strand breakage(Bombail et al., 2001). The advantages of this assay include therelative ease of application to any tissue of interest as long as it ispossible to separate the cells, the detection of multiple classes ofDNA damage (Hartmann et al., 2003) and sensitivity of DNAdamage detection because a small number of cells are needed toenable the test (o10,000) (Lee and Steinert, 2003). In this study,the comet assay, which is usually performed on blood cells(erythrocytes), was also performed in renal cells due to thehematopoietic role played by the cephalic kidney because thegenotoxic effects of contaminants can be tissue specific(Belpaeme et al., 1998).
This study aimed to evaluate genotoxic damage after exposureto sub-chronic dietary doses of methylmercury (0.075 mgCH3Hgþ/g and 0.75 mg CH3Hgþ/g), which were administereddaily to the fish species Hoplias malabaricus (traıra) through theirdiet for over a period of 14 weeks. For both conditions, we usedthe piscine micronucleus test, conventional staining (giemsa),fluorescent staining (acridine orange), and the comet assay inerythrocytes as well as in kidney cells. The use of these para-meters as sensitive and long-term biomarkers for sublethal, sub-chronic, and trophic exposures to CH3Hgþ was investigated in H.
malabaricus. To increase the biological knowledge regardingtropical species and the toxicity of methylmercury in this fishspecies, which inhabits polluted regions of the Amazon, this studyapplied similar conditions of contamination found in naturaltropical aquatic environments.
2. Materials and methods
2.1. Fish
Hoplias malabaricus (Family–Erythrinidae), popularly known as traıra, was
chosen for this study because it is a Neotropical piscivorous fish with a wide
geographic distribution, primarily in South America, and is largely used in daily
human consumption for riverine populations in the Amazon region (Grandjean,
1999). This fish is advantageous for use in studies because it is easily fed under
laboratory conditions, it devours its prey whole, and it can be easily exposed to
xenobiotics by feeding it prey previously injected with the compounds of interest
(Ferraro et al., 2004).
Overall, 43 specimens of H. malabaricus with an average weight and length of
3075.5 g and 9.571.5 cm, respectively, were supplied by a fish farm in the state
of S~ao Paulo (Southeast-Brazil). Before the experiments, they were acclimated
under laboratory conditions for approximately 40 day at a population density of
1 specimen per 30 l of dechlorinated tap water at 23 1C in an aired aquarium, with
a 12-h light/12-h dark regime cycle, and with a pH of approximately 8. Each study
fish was supplied with one young alive Astyanax sp, a freshwater prey fish species,
which were collected from a fish farm without pollutant sources, for each period
of five days.
2.2. Experimental design
Each study fish (H. malabaricus– traıra) was fed one individual prey fish
(approximately 10% of the wet weight) every 5 day to ensure ingestion. The
contaminant was injected intraperitoneally into the prey fish (Astyanax sp). The
volume of the contaminant (CH3Hgþ) was adjusted according to the individual
weight of each test fish (H. malabaricus) to obtain standardized ingestion doses,
and the doses were diluted in a 1 mN HCl aqueous solution. For methylmercury,
ferent doses of methylmercury (CH3Hgþ) in Hoplias malabaricus.012.05.007

Table 1Frequencies of micronucleated erythrocytes and nuclear alterations in erythro-
cytes of Hoplias malabaricus, with conventional staining (Giemsa), exposed to
different treatments. The results are presented in medians and quartiles (q1–q3).
Treatment N Concentrations
(mg/g)
MN
(medians/
q1–q3)
Nuclear
alterations(NA)
(medians/q1–q3)
Total
Control 13 1/1–2 43/26–64 2000
Methylmercury 15 0.075 1/1–125 36/17.5–43 2000
15 0.75 0/0–0 24/15.5–34.5 2000
T. Vicari et al. / Ecotoxicology and Environmental Safety ] (]]]]) ]]]–]]] 3
the dose administered in our work was very similar to real conditions found in
prey fish from Amazonian rivers impacted by mercury, as reported by Mirlean
et al. (2005). The control group (N¼13) was fed prey fish (Astyanax sp) that
received an intraperitoneal injection of 0.1 ml of distilled water. Of the 30 fish
remaining, half (N¼15) were fed with Astyanax sp. previously intraperitoneally
injected with methylmercury at a concentration of 0.075 mg of CH3Hgþ/g body
weight of H. malabaricus and the other half (N¼15) were fed with Astyanax sp.
previously intraperitoneally injected with methylmercury at a concentration of
0.75 mg of CH3Hgþ/g body weight of H. malabaricus. The prey item was not force-
fed to individual fish. A total of 70 days of infection, accounting for 14 power
cycles were completed. Five days after the last power cycle, the specimens were
anesthetized with 10% benzocaine (Merck, Darmstadt, Germany) (Cabral Gontijo
et al., 2003) and prepared for the removal of samples.
Blood samples were collected from the caudal vein for the piscine micro-
nucleus test and the comet assay using 5 ml heparinized syringes, and the cephalic
kidney was removed for the tissue comet assay.
2.3. Scoring genotoxic damage
Genotoxicity was tested using the piscine micronucleus test performed in
mature peripheral erythrocytes, according to the procedures of Schmid (1975) and
modified by Carrasco et al. (1990). Also, it was tested using the comet assay in
erythrocytes and in kidney cells, according to the procedures of Singh et al. (1988),
with a few modifications by Ferraro et al. (2004).
2.4. Piscine micronucleus test (conventional staining)
After blood samples were obtained, one drop of each sample was immediately
smeared on a clean microscope slide. Then, the smears were left to air-dry at room
temperature and fixed in pure ethanol for 25 min. After they set, the slides were
dried at room temperature and stained with 10% Giemsa in phosphate buffer at pH
6.8 for 15 min. For each fish, a total of 2000 erythrocytes were examined under a
light microscope (1000� magnification) to determine the frequency of micro-
nuclei (MN) and nuclear abnormalities (NA). The frequencies of micronuclei and
nuclear morphological alterations were observed according to Carrasco et al.,
1990, as cited for Ayllon and Garcia-Vazquez (2001), and were summed and
named as MNþNA (micronucleusþnuclear abnormalities). Only nucleated ery-
throcytes with intact cellular and nuclear membranes were scored. Rounded
particles were categorized as MN based on a diameter of 1/5 to 1/20 of the main
nucleus that lacked a connection to the nucleus, a staining characteristic that was
consistent with the main nucleus, and a non-refractory image.
2.5. Piscine micronucleus test (fluorescent staining)
Acridine orange staining of erythrocytes was performed according to Ueda
et al. (1992). Briefly, 10 ml of peripheral blood sample collected from the caudal
vein of the fish was diluted with approximately the same amount of fetal bovine
serum (10 ml-Gibcos) and smeared on clean glass slides. The slides were dried
overnight and fixed with methanol for 10 min. Immediately, using an epifluores-
cence microscope, the cells were stained with 0.003% acridine orange in
Sorenson’s buffer at pH 6.8 for 2–3 min and mounted in the same buffer.
Approximately, 2000 erythrocytes per animal were analyzed by a Leicas epi-
fluorescence microscope under 1000X magnification, classifying them as mature
erythrocytes (ME) and immature erythrocytes (IE) based on selective criteria
established by the stain used. Acridine orange stains the primary nuclei of
erythrocytes (normochromatic) as well as micronuclei with strong yellow–green
fluorescence. In contrast, immature erythrocytes (polychromatic) are easily
identifiable by the presence of RNA in the cytoplasm and red fluorescent staining.
The results are presented as a frequency (%) using the following equation (C- avas
and Ergene-Gozukara, 2005a): IE frequency (%)¼IE/(IEþME)�100; where ME
denotes the mature erythrocytes and IE denotes the immature erythrocytes.
2.6. Comet assay
Clean microscope slides were prepared by covering them with a thin layer of
0.75% NMP agarose (Gibcos) and dried overnight. One slide was prepared for each
fish. For each slide covered with agarose, 10 ml of whole fish blood was diluted in
1 ml of fetal bovine serum, and 10 ml of this mixture was placed in 120 ml of low
melting point agarose (LMP-0.5%-Gibcos). Of the erythrocytes in agarose suspen-
sion, 120 ml were then pipetted onto a fully frosted slide and covered with a
coverslip. Then, the slides were placed on refrigerator for 10 min to allow
complete polymerization of the agarose.
After the agarose polymerized, the coverslips were gently removed, and the
slide was lowered into freshly made lysis solution at 4 1C for 24 h to remove the
cellular proteins. After lysing, the slides were drained of the lysis solution and
placed on a horizontal electrophoresis tray. Freshly made buffer was added to the
tray, and the submerged slides were incubated in alkaline buffer (pH413) for
Please cite this article as: Vicari, T., et al., Genotoxic evaluation of difEcotoxicol. Environ. Saf. (2012), http://dx.doi.org/10.1016/j.ecoenv.2
30 min at 4 1C, allowing the DNA to unwind. For electrophoresis, the power supply
was set at 25 V, and the current was adjusted to 300 mA by slowly changing the
buffer level in the tray. The slides were exposed to this current in the dark at 4 1C
for 25 min. Next, the slides were removed from the tray and neutralized with
neutralizing buffer (0.4 M Tris, pH 7.5) three times in the dark for 5 min/session.
After the last neutralization step, the buffer was drained, and the slides were fixed
in absolute ethanol for 10 min. Then, the slides were stained with 20 ml of
ethidium bromide (10 ml/ml) and covered with a coverslip. The slides were
examined at 400�using a Leicas epifluorescence microscope.
The procedure used for the comet assay in kidney cells is similar to the
protocol used with blood cells; however, differences occur in the method used to
obtain viable cells from kidney tissue, and the protocol was previously described
by Ramsdorf et al. (2009). The kidney tissue cells used for the comet assay were
homogenized (homogenizer Potters type at 1500 rpm) in Saccharose Tris–HCl
buffer solution (pH 8.6). The volume of additional buffer was four times the
collected tissue volume. From the final solution, 50 ml was mixed with 120 ml of
low-melting-point agarose (LMP-0.5%-Gibcos), pipetted onto slides previously
coated with NMP agarose (0.75%-Gibcos) and covered with a coverslip. Next, the
slides were placed on the refrigerator for 10 min to allow complete polymerization
of the agarose. The slides were immersed in lysis solution, and the electrophoresis
protocol also followed Ferraro et al. (2004) as above.
For each fish, 100 nucleoids were visually analyzed according to a method
using visual classification (Collins et al., 1997) based on the migration of DNA
fragments and were categorized into class 0 (no visible damage), class 1 (little
damage), class 2 (medium damage), class 3 (extensive damage) or class 4
(maximally damaged). The score was calculated by multiplying the number of
nuclei found in a class by the class number.
2.7. Statistical analysis
Bio Stats software was used for statistical analyses. Data were tested for
normality using the Kolmogorov–Smirnov test and homogeneity of variance to
meet statistical demand.
Differences between fish treated with two doses of methylmercury and the
untreated control group were tested using the non-parametric Kruskal–Wallis
test, for the micronucleus test stained with conventional staining (Giemsa), the
comet assay for erythrocytes and for the comet assay in kidney tissue. The results
are presented in medians and quartiles (q1–q3). When the data are not symme-
trical, to this method, is the ‘‘average’’ way that provides a better idea of any
general tendency in the data. The Wilcoxon test for paired samples was used for
comparison of the comet assay in erythrocytes and the comet assay of kidney cells.
In case of non-parametric testing, differences between the medians are considered
statistically significant when po0.05.
Only the data for immature erythrocytes (IE) showed normal distribution;
therefore, parametric tests were used to detect differences at the 0.05 level of
significance. ANOVA analysis was used to compare the results between fish
groups, and in this case, all the data are expressed as the median7quartiles.
The Wilcoxon test for paired samples was used for comparison of the results
obtained using the conventional staining method and acridine orange staining, for
all treatment groups (control, 0.075 mg CH3Hgþ/g and 0.75 mg CH3Hgþ/g). In case
of non-parametric testing, differences between the medians are considered
statistically significant when po0.05.
3. Results
3.1. Piscine micronucleus test
3.1.1. Conventional staining (Giemsa)
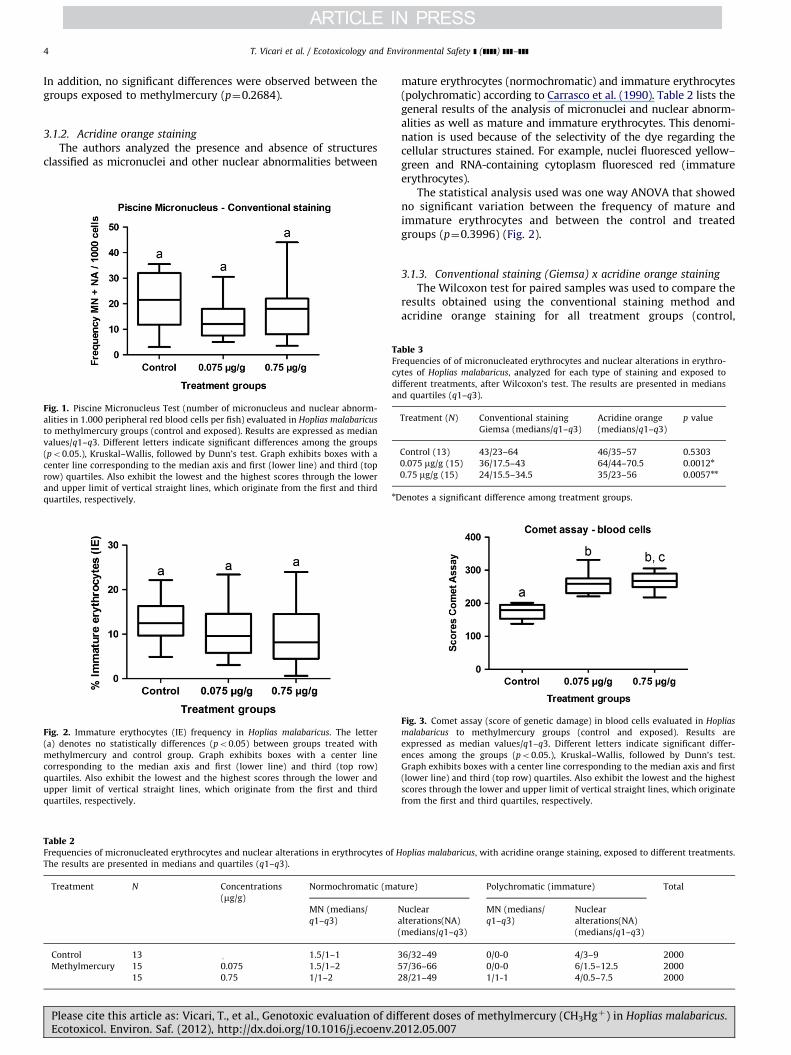
The results summarizing micronuclei and nuclear abnormalitiesas well as normal erythrocytes stained with Giemsa are shown inTable 1 (Fig. 1). No significant differences were found betweenthe control group and the groups treated with methylmercury.
ferent doses of methylmercury (CH3Hgþ) in Hoplias malabaricus.012.05.007
T. Vicari et al. / Ecotoxicology and Environmental Safety ] (]]]]) ]]]–]]]4
In addition, no significant differences were observed between thegroups exposed to methylmercury (p¼0.2684).
3.1.2. Acridine orange staining
The authors analyzed the presence and absence of structuresclassified as micronuclei and other nuclear abnormalities between
Table 2Frequencies of micronucleated erythrocytes and nuclear alterations in erythrocytes of H
The results are presented in medians and quartiles (q1–q3).
Treatment N Concentrations
(mg/g)
Normochromatic (mat
MN (medians/
q1–q3)
Control 13 1.5/1–1
Methylmercury 15 0.075 1.5/1–2
15 0.75 1/1–2
Fig. 2. Immature erythocytes (IE) frequency in Hoplias malabaricus. The letter
(a) denotes no statistically differences (po0.05) between groups treated with
methylmercury and control group. Graph exhibits boxes with a center line
corresponding to the median axis and first (lower line) and third (top row)
quartiles. Also exhibit the lowest and the highest scores through the lower and
upper limit of vertical straight lines, which originate from the first and third
quartiles, respectively.
Fig. 1. Piscine Micronucleus Test (number of micronucleus and nuclear abnorm-
alities in 1.000 peripheral red blood cells per fish) evaluated in Hoplias malabaricus
to methylmercury groups (control and exposed). Results are expressed as median
values/q1–q3. Different letters indicate significant differences among the groups
(po0.05.), Kruskal–Wallis, followed by Dunn’s test. Graph exhibits boxes with a
center line corresponding to the median axis and first (lower line) and third (top
row) quartiles. Also exhibit the lowest and the highest scores through the lower
and upper limit of vertical straight lines, which originate from the first and third
quartiles, respectively.
TaFr
cy
di
an
nD
Please cite this article as: Vicari, T., et al., Genotoxic evaluation of difEcotoxicol. Environ. Saf. (2012), http://dx.doi.org/10.1016/j.ecoenv.2
mature erythrocytes (normochromatic) and immature erythrocytes(polychromatic) according to Carrasco et al. (1990). Table 2 lists thegeneral results of the analysis of micronuclei and nuclear abnorm-alities as well as mature and immature erythrocytes. This denomi-nation is used because of the selectivity of the dye regarding thecellular structures stained. For example, nuclei fluoresced yellow–green and RNA-containing cytoplasm fluoresced red (immatureerythrocytes).
The statistical analysis used was one way ANOVA that showedno significant variation between the frequency of mature andimmature erythrocytes and between the control and treatedgroups (p¼0.3996) (Fig. 2).
3.1.3. Conventional staining (Giemsa) x acridine orange staining
The Wilcoxon test for paired samples was used to compare theresults obtained using the conventional staining method andacridine orange staining for all treatment groups (control,
oplias malabaricus, with acridine orange staining, exposed to different treatments.
ure) Polychromatic (immature) Total
Nuclear
alterations(NA)
(medians/q1–q3)
MN (medians/
q1–q3)
Nuclear
alterations(NA)
(medians/q1–q3)
36/32–49 0/0-0 4/3–9 2000
57/36–66 0/0-0 6/1.5–12.5 2000
28/21–49 1/1-1 4/0.5–7.5 2000
ble 3equencies of of micronucleated erythrocytes and nuclear alterations in erythro-
tes of Hoplias malabaricus, analyzed for each type of staining and exposed to
fferent treatments, after Wilcoxon’s test. The results are presented in medians
d quartiles (q1–q3).
Treatment (N) Conventional staining
Giemsa (medians/q1–q3)
Acridine orange
(medians/q1–q3)
p value
Control (13) 43/23–64 46/35–57 0.5303
0.075 mg/g (15) 36/17.5–43 64/44–70.5 0.0012n
0.75 mg/g (15) 24/15.5–34.5 35/23–56 0.0057nn
enotes a significant difference among treatment groups.
Fig. 3. Comet assay (score of genetic damage) in blood cells evaluated in Hoplias
malabaricus to methylmercury groups (control and exposed). Results are
expressed as median values/q1–q3. Different letters indicate significant differ-
ences among the groups (po0.05.), Kruskal–Wallis, followed by Dunn’s test.
Graph exhibits boxes with a center line corresponding to the median axis and first
(lower line) and third (top row) quartiles. Also exhibit the lowest and the highest
scores through the lower and upper limit of vertical straight lines, which originate
from the first and third quartiles, respectively.
ferent doses of methylmercury (CH3Hgþ) in Hoplias malabaricus.012.05.007

Fig. 4. Comet assay (score of genetic damage) in kidney cells evaluated in Hoplias
malabaricus to methylmercury groups (control and exposed). Results are
expressed as median values/q1–q3. Different letters indicate significant differ-
ences among the groups (po0.05.), Kruskal–Wallis, followed by Dunn’s test.
Graph exhibits boxes with a center line corresponding to the median axis and first
(lower line) and third (top row) quartiles. Also exhibit the lowest and the highest
scores through the lower and upper limit of vertical straight lines, which originate
from the first and third quartiles, respectively.
Table 4Scores of comet assay in erythrocytes (CAE) and comet assay in kidney cells (CAK),
after Wilcoxon’s test. The results are presented in medians and quartiles (q1–q3).
Treatment (N) CAE (medians/q1–q3) CAK (medians/q1–q3) p value
Control (13) 179/157–193 169/122.5–171.5 0.2475
0.075 mg/g (15) 259/235–274 175/167–209 0.0046n
0.75 mg/g (15) 268/251.5–283 189/158.5–236 0.0020nn
nDenotes a significant difference among treatment groups.
T. Vicari et al. / Ecotoxicology and Environmental Safety ] (]]]]) ]]]–]]] 5
0.075 mg CH3Hgþ/g and 0.75 mg CH3Hgþ/g) (Table 3). Among thespecimens subjected to 0.075 mg CH3Hgþ/g and 0.75 mg CH3Hgþ/g, a statistically significant difference between the two types ofstaining used (p¼0.0012 and p¼0.0057, respectively) wasobserved between the two groups. No statistically significantdifference was found between the specimens from the controlgroup (p¼0.5303) when compared to the test specimensdiscussed above.
3.2. Comet assay
3.2.1. Comet assay in erythrocytes
For the comet assay applied to the blood samples, thestatistical test showed that significant differences exist betweenthe control group and the group treated with 0.075 mg CH3Hgþ/g(po0.0001) as well as between the control group and the grouptreated with 0.75 mg CH3Hgþ/g (po0.0001). Among the groupstreated with methylmercury, no significant difference betweenthese groups was found (p¼0.6365) (Fig. 3).
3.2.2. Comet assay in kidney cells
For the comet assay in kidney cells, after statistical analysis, nosignificant differences were observed between the control groupand the group treated with 0.075 mg CH3Hgþ/g as well asbetween the control group and the group treated with 0.75 mgCH3Hgþ/g. Also, no statistically significant differences were foundbetween the groups treated with methylmercury (p¼0.2677)(Fig. 4).
3.2.3. Comet assay in erythrocytes x comet assay in kidney cells
The Wilcoxon test for paired samples was used to compare theresults achieved in the comet assay of erythrocytes and in the
Please cite this article as: Vicari, T., et al., Genotoxic evaluation of difEcotoxicol. Environ. Saf. (2012), http://dx.doi.org/10.1016/j.ecoenv.2
comet assay of kidney cells. Among the samples subjected to0.075 mg CH3Hgþ/g and to 0.75 mg CH3Hgþ/g, the blood wasmore sensitive to contamination by methylmercury compared tothe kidney cells in Hoplias malabaricus (p¼0.0046 and p¼0.0020,respectively). No statistically significant difference was found forspecimens from the control group (p¼0.2475) compared to thetest specimens mentioned above (Table 4).
4. Discussion
Short-term toxicological tests, primarily genetic in nature, areuseful tools to determine the toxicity of certain compounds andtheir effect on the genetic material of cells. These tests generateinformation that will allow the identification of potential carcino-gens and consequently protect the health of the public and theenvironment. In Brazil, mining activity located in the Amazonregion is responsible for high mercury levels found in the muscleof carnivorous fish that inhabit the region (Pfeiffer et al., 1989,1991; Malm et al., 1995; Souza Lima et al., 2000; Santos et al.,2002; Mela et al., 2007). These levels exceed the maximum andrecommended levels safe for human consumption allowed byBrazilian law (CONAMA, 2005) and by international law (WHO,1990). In this case, aquatic predators that are located at the top ofthe food chain are an important link between mercury pollutionand human health (Santos et al., 2002).
The present study investigated the genotoxic potential ofmethylmercury using the species Hoplias malabaricus, popularlyknown as traıra, as a model organism. The species H. malabaricus
is largely used in the daily human consumption in the Amazonianregion (Grandjean, 1999). The species also occupies a place at thetop of the aquatic food chain. Although our results cannot beextended to humans, human communities inhabiting some Ama-zonian areas are still exposed to fish containing similar levels ofmethylmercury as described in work published by Mela et al.(2007).
Mela and colleagues studied similar trophic contaminationusing 0.075 mg/g wet weight of CH3Hgþ and assessed the amountof total mercury in the liver and the muscle of Hoplias malabaricus
specimens after 70 day of exposure (Mela et al., 2007). The resultsshowed that total quantities of mercury in the liver and themuscle increased in both tissues. In their study, very lowbioaccumulation of total mercury was detected in the liver andthe muscle of individuals from the control group, suggesting thatthe control specimens were chronically exposed to trace mercuryconcentrations before the experiment. However, the increase oftotal mercury bioaccumulation in individuals from the test groupwas evident, demonstrating the high trophic bioavailability ofCH3Hgþ in fish as described by Oliveira Ribeiro et al. (1999). Fastuptake of CH3Hgþ due to diet has also been reported for other fishspecies (Oliveira Ribeiro et al., 1999; Berntssen et al., 2003). Thisefficient biomagnification of CH3Hgþ is attributed to the ability ofthis small lipophilic molecule to diffuse through the intestinalbarrier and reach cells and tissues rich in lipids. Organic mercuryforms are more likely to be responsible for the observed aneu-genic and clastogenic action due to its lipophilicity (De Flora et al.,1994).
According to De Flora et al. (1994), the genotoxicity of organicand inorganic mercury compounds are qualitatively comparable,suggesting the involvement of a common genotoxic entity, whichprimarily depends on a differential bioavailability. Usually, thefirst mechanism of action of genotoxic agents is to promote DNAdamage, which can have three possible outcomes: the damagecan be repaired, the damage can become irreversible, or thedamage may lead to cell death. The genetic damage observed inthis study refers to a substance with clastogenic and/or aneugenic
ferent doses of methylmercury (CH3Hgþ) in Hoplias malabaricus.012.05.007

T. Vicari et al. / Ecotoxicology and Environmental Safety ] (]]]]) ]]]–]]]6
action. The most typical damage produced by mercury at thegenetic level was the disturbance of the spindle mechanism,which is due to the affinity of mercury for the sulfhydryl groupspresent in spindle fiber proteins (De Flora et al., 1994).
Porto and colleagues conducted an evaluation of mercurypollution in three Amazonian fish species using the piscinemicronucleus test, and they found that the test was very usefulto indicate genotoxicity of mercury in Amazon rivers. Further-more, they discovered that the species Hoplias malabaricus is anexcellent sentinel species for monitoring the area (Porto et al.,2005). In our study, the results of the piscine micronucleus testwere not statistically significant, potentially because we selectedonly one route of contamination and due to the reduced time ofthe experiment (70 day—sub-chronic experiment). However, thedose used in our work was very similar to the conditions found inprey fish from Amazonian rivers impacted by mercury as recentlyreported by Mirlean et al. (2005).
As previously reported by Das and Nanda (1986), Brunettiet al. (1988), Nepomuceno et al. (1997) and recently byGuilherme et al. (2008), a gradual decrease in the frequency ofmicronuclei with increasing time of exposure and with increas-ing concentrations of xenobiotics can be explained by aninhibitory effect of contaminants on cell division and conse-quently by the hindrance in the passage of the affected cells inperipheral circulation (Das and Nanda, 1986). This possibilitymay explain why we did not find significant differences in thiswork between the control and infected groups when using thepiscine micronucleus test. This effect may either a result frominhibition of DNA synthesis (Williams and Metcalfe, 1992) orfrom direct inhibition of erythropoiesis (Dinnen et al., 1988)caused by several genotoxic agents at certain concentrations,and this effect is regarded as the cytotoxic property of thexenobiotics (Udroiu, 2006). In addition, the appearance ofnuclear lesions is determinately affected by a variety of factors,such as erythropoiesis, the time required for maturation and thelifespan of the erythrocytes (Udroiu, 2006). All of these con-founding factors should be carefully considered because theymay cause a false negative diagnosis.
For discussion regarding the use of nuclear abnormalitiesduring the piscine micronucleus test, we clearly observe thelack of pre-established uniform criteria, allowing an effectivecomparison between the previous studies from different labora-tories. The relationship between nuclear abnormalities and geno-toxic effects of contaminants has not been well established;however, preliminary observations suggest that such morpholo-gical changes may be a manifestation of the effects of xenobiotics.Some authors (Hooftman and Raat, 1982; Hose et al., 1987)suggest that variations in the shape and the size of the nucleusmay represent an alternative approach for the detection ofgenotoxicity. According to Serrano-Garcia and Montero-Montoya(2001), the phenomena of budding cell nuclei and bi-nucleatedcells have a similar origin, and micronuclei are supposed to becaused by genotoxic occurrences. Moreover, a positive relation-ship between micronucleus induction and nuclear buds wasfound by different authors (Bolognesi et al., 2006; Ergene et al.,2007), suggesting that nuclear bud formation in erythrocytes maybe a useful complementary assay for genotoxicity assessmentin fish.
The evaluation of erythropoietic toxicity is a key component ofsafety assessment in the development of new drugs, and thecount of immature (polychromatic) erythrocytes in peripheralblood is one of the most popular and convenient methods ofmonitoring erythropoiesis. The decrease in the proportion ofimmature (polychromatic) erythrocytes compared to matureerythrocytes (normochromatic) is considered to be an indicatorof mutagenicity-induced cytotoxicity (Schmid, 1975; Suzuki et al.,
Please cite this article as: Vicari, T., et al., Genotoxic evaluation of difEcotoxicol. Environ. Saf. (2012), http://dx.doi.org/10.1016/j.ecoenv.2
1989). Routinely included in testing of micronuclei in mammals,the ratio of polychromatic/normochromatic cells is a key compo-nent in the evaluation of cytotoxicity in these animals (Criswellet al. 1998; Celik et al., 2003). Relevant articles are available in theliterature and discuss the relationship between the polychro-matic/normochromatic cell ratio and the micronucleus test in fish(Smith, 1990; Pacheco and Santos, 2002; C- avas and Ergene-Gozukara, 2005a; Guilherme et al., 2008).
Smith (1990) reported a decreased rate of immature (poly-chromatic) erythrocytes in peripheral blood of infected fish inareas with polycyclic aromatic hydrocarbons (PAHs). In anotherstudy, Pacheco and Santos (2002) reported a decrease in thenumber of immature erythrocytes in the peripheral bloodof Anguilla anguilla; however, the frequency of micronucleiincreased as a result of treatments with benzo [a] pyrene,dehidroabietic acid and paper mill effluents. C- avas and Ergene-Gozukara (2005a) examined the peripheral blood of specimensof Oreochromis niloticus and reported a significant decrease inthe rate of immature/mature erythrocytes after treatment withthe antibiotic and antiparasitic agent metronidazole (MTZ).The work performed in Portugal by Guilherme et al. (2008),using a gradient of mercury contamination in this region,analyzed Liza aurata erythrocytes and the relationship betweenthe appearance of nuclear abnormalities, the frequency ofimmature erythrocytes and the temperature of the season.Surprisingly, there was no induction of nuclear abnormalitiesduring the winter when they found the highest mercury con-centrations in fish blood, which is presumably due to a change inhematological dynamics caused by a decrease in the frequencyof immature erythrocytes. In this study, the reduction in thenumber of immature erythrocytes was not statistically signifi-cant, preventing a more detailed conclusion regarding hemato-poietic dynamics. This finding is most likely due to the shorttime of contamination coupled with low doses of methylmer-cury, which were provided every five days to the animals. Inaddition, the determination of IE frequency provides importantinformation regarding hematological dynamics, but its isolatedanalysis does not allow a clear identification of the causebecause it reflects the balance between a variety of factors, suchas immature cell input vs. splenic cell removal and cell matura-tion rate (Guilherme et al., 2008).
Regarding the results obtained in the comet assay, somestudies have shown that the assay is actually able to detectDNA damage caused by different classes of xenobiotics in fish,showing that the test is highly sensitive. For example, Pandrangiet al. (1995) showed an increase in the DNA damage in catfisherythrocytes (Ameiurus nebulosus) and carp erythrocytes (Cyprinus
carpio) after the capture of fish in areas known to be contami-nated with PAHs and PCBs. The comet assay results wereconsistent with the degree of contamination of the sites, provingthe sensitivity of the assay. Another study conducted with fish,more specifically in H. malabaricus, that were subjected to acutecontamination with different concentrations of inorganic lead(Ramsdorf et al., 2009), showed negative results with respect tothe piscine micronucleus test but tested positive for the cometassay in erythrocytes.
In this study, the comet assay in erythrocyte system cellsshowed significant differences between the control and the groupscontaminated with methylmercury (po0.0001) (Fig. 3). In experi-ments performed by Ferraro et al. (2004), working with similarconditions of trophic contamination (13 trophic levels in a periodof 60 day) using a lead (Pb2þ) concentration of 21 mg/g in H.
malabaricus, they found an increased frequency of nuclearabnormalities, chromosomal abnormalities and also a significantincrease in the number of comets in fish treated with Pb2þ ,showing once again that long exposure to xenobiotics, in this
ferent doses of methylmercury (CH3Hgþ) in Hoplias malabaricus.012.05.007

T. Vicari et al. / Ecotoxicology and Environmental Safety ] (]]]]) ]]]–]]] 7
case lead, can cause damage to the genetic material of fish,corroborating our results in the comet assay with the blood of H.
malabaricus exposed to methylmercury. According to Lee andSteinert (2003), interactions between the DNA molecule and thecontaminants to which organisms are exposed occur in fivedifferent ways: either genotoxic agents act directly on the DNAmolecule; the metabolites of this genotoxic compound interactwith DNA; DNA damage is caused by reactive oxygen species,which is formed by the action of the genotoxic compound; thecompounds can inhibit DNA synthesis and DNA repair; andgenotoxic compounds can produce DNA damage through multi-lateral mechanisms. The latter two mechanisms of fundamentalimportance for understanding the action of methylmercury on theDNA molecule, and some studies indicate that these last twomechanisms could be related to the type of interaction thatmethylmercury has with the DNA molecule, causing the damagethat we observed in blood cells of these organisms (Snyder et al.,1989; Cantoni et al., 1984a, 1984b).
Also, regarding the result obtained in the comet assay appliedto kidney cells, we analyzed the dynamics involved in the pathtaken by the xenobiotics into the body of the animal. The liver isresponsible for turning some harmful compounds into metabo-lites that are excreted directly into the bile, continuing thedetoxifying process. When a compound is excreted into bile andenters the small intestine, it has two fates: either it is reabsorbedin the intestine, or it is excreted in feces. Methylmercury isreabsorbed in a process called entero-hepatic recirculation(Gordon and Skett, 1986). When methylmercury undergoes thiscycle, the mercury is retained by the body and has a substantiallyincreased half-life. If the concentration of the organomercurialcompound is high enough, this cyclic metabolism can resultin prolonging the pharmacological activity of mercury (Gordonand Skett, 1986). Methylmercury can cross the blood–brainbarrier and nuclear membranes to react directly with cellularand nuclear components (Boening, 2000). Thus, methylmercuryreactions that act directly on cellular and nuclear componentsmost likely occur only in the kidney. Mela et al. (2007) reportedan increase of melano-macrophages centers (MMCs) in the kidneyof individuals exposed to trophic methylmercury, suggesting thepresence of dead cells or large erythrocyte turnover, which wasalso described by Meseguer et al. (1994) and Ravaglia andMeseguer (1995). In conclusion, Mela et al. (2007) suggests thatthe damages found in the kidney are severe enough to causeirreversible consequences to cell physiology or to organ functionas reported in this paper.
These results may have also been obtained by the comet assayin kidney cells and in blood cells because differences may berelated to the different tissues studied. Different tissues ofdifferent animal species have varying amounts of alkali-labilesites. While conducting the comet assay in different tissues, thedifferences in morphology, staining characteristics and the fre-quency of different types of cells should be checked. Some of thefactors that can influence DNA damage in a particular tissueinclude cell type, heterogeneity, cell cycle, cell turnover frequencyand culture or growth conditions. Different cell types can havevery different background levels of DNA single-strand breaks dueto variations in excision repair activity, metabolic activity, anti-oxidant concentrations and other factors that affect the finaloutcome of the experiments (Lee and Steinert, 2003).
It is important to emphasize the difference between theresponses observed in the comet assay for kidney cells and bloodcells, which showed a relevant association. The blood had a moreacute reaction to the presence of methylmercury, indicating thatthis tissue is more sensitive than kidney tissue, which is mostlikely due to the type of contamination used (trophic); therefore,the kidney would require more exposure time and possibly an
Please cite this article as: Vicari, T., et al., Genotoxic evaluation of difEcotoxicol. Environ. Saf. (2012), http://dx.doi.org/10.1016/j.ecoenv.2
increase in the concentration of the contaminant. Overall, inerythrocytes, the comet assay is an important tool because itproves to be sensitive to agents known to be mutagenic. Thisstudy reinforces the idea by Belpaeme et al. (1998), who suggestthat the animal response may depend on the experimentalconditions of each species, the cell type, the mutagenic agentand the exposure time.
In conclusion, we studied this type of contamination (trophicroute) for the time period of 70 day (sub-chronic) in an attempt tothe mimic environmental conditions, and the piscine micronu-cleus test showed no significant differences when stained withconventional staining; however, when stained with acridineorange, the results obtained for the exposed groups were moreinformative and meaningful, allowing a broader discussionregarding the parameters found. In regards to the comet assay,erythrocytes were more sensitive than kidney cells, most likelybecause contamination was conducted throughout several days insmall doses, and a more immediate response was observed in theblood, followed by damage to kidney cells that were not con-sidered significant in our work. However, more studies areneeded to verify the genotoxic damages of CH3Hgþ for extendedtime periods and with other doses of methylmercury to clarifyoutstanding issues regarding its contamination. Our data indicatethat H. malabaricus represents a good experimental model forgenotoxicity studies.
Acknowledgments
The authors thank CNPq (Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico) and Aquatoxi group. This researchwas supported by CAPES (Coordenac- ~ao de Aperfeic-oamento dePessoal de Nıvel Superior—Brazil).
References
Arkhipchuk, V.V., Garanko, N.N., 2005. Using the nucleolar biomarker and themicronucleus test on in vivo fish fin cells. Ecotoxicol. Environ. Saf. 62,42–52.
Aschner, M., 2002. Neurotoxic mechanisms of fish-borne methylmercury. Environ.Toxicol. Pharmacol. 12, 101–104.
Ayllon, F., Garcia-Vazquez, E., 2001. Micronuclei and other nuclear lesions asgenotoxicity indicators in rainbow trout Oncorhynchus mykiss. Ecotoxicol.Environ. Saf. 49, 221–225.
Barbosa, A.C., Boischio, A.A., East, G.A., Ferrari, I., Gonc-alves, A., Silva, P.R.M., Cruz,T.M.E., 1995. Mercury contamination in the Brazilian Amazon. Environmentaland occupational aspects. Water Air Soil Pollut. 80, 109–121.
Belpaeme, K., Cooreman, K., Kirsch-Volders, M., 1998. Development and validationof the in vivo alkaline comet assay for detecting genomic damage in marineflatfish. Mutat. Res. 415 (3), 167–184.
Berntssen, M.H.G., Aatland, A., Handy, R.D., 2003. Chronic dietary mercuryexposure causes oxidative stress, brain lesions, and altered behaviour inAtlantic salmon (Salmo salar) parr. Aquat. Toxicol. 65, 55–72.
Bidone, E.D., Castilhos, Z.C., Cid de Souza, T.M., et al., 1997. Fish contaminationand human exposure to mercury in the Tapajos River Basin, Para State,Amazon, Brazil: a screening approach. Bull. Environ. Contam. Toxicol. 59 (2),194–201.
Boening, D.W., 2000. Ecological effects, transport, and fate of mercury: a generalreview. Chemosphere 40, 1335–1351.
Bolognesi, C., Perrone, E., Roggieri, P., Pampanin, D.M., Sciutto, A., 2006. Assess-ment of micronuclei induction in peripheral erythrocytes of fish exposed toxenobiotics under controlled conditions. Aquat. Toxicol. 78, S93–S98.
Bombail, V., Aw, D., Gordon, E., Batty, J., 2001. Application of the comet andmicronucleus assays to butterfish (Pholis gunnellus) erythrocytes from theFirth of Forth, Scotland. Chemosphere 44, 383–392.
Brunetti, R., Majone, F., Gola, I., Beltrame, C., 1988. The micronucleus test:examples of application to marine ecology. Mar. Ecol. Prog. Ser. 44, 65–68.
Cantoni, N.T.O., Christie, A., Swann, D.B., Drath, M., Costa, M., 1984a. Mechanismof HgCl2 cytotoxicity in cultured mammalian cells. Mol. Pharmacol. 26,360–368.
Cantoni, N.T.O., Christi, S.H., Robison, M., Costa, M., 1984b. Characterization ofDNA lesions produced by HgCl2 in cell culture systems. Chem. Biol. Interact.49, 209–224.
ferent doses of methylmercury (CH3Hgþ) in Hoplias malabaricus.012.05.007

T. Vicari et al. / Ecotoxicology and Environmental Safety ] (]]]]) ]]]–]]]8
Carrasco, K.R., Tilbury, K.L., Myers, M.S., 1990. Assessment of the piscine micro-nucleus test as an in situ biological indicator of chemical contaminant effects.Can. J. Fish. Aquat. Sci. 47, 2123–2136.
Cabral Gontijo, A.M.M., Barreto, R.E., Speit, G., Reyes, V.A.V., Volpato, G.L.,Salvadori, D.M.F., 2003. Anesthesia of fish with benzocaine does not interferewith comet assay results. Mutat. Res. 534, 165–172.
Celik, A., C- avas, T., Ergene-Gozukara, S., 2003. Cytogenetic biomonitoring in petrolstation attendants: micronucleus test in exfoliated buccal cells. Mutagenesis18, 417–721.
Criswell, K.A., Krishna, G., Zielinski, D., Urda, G.A., Theiss, J.C., Juneau, P., Bleavins,M.R., 1998. Use of acridine orange in flow cytometric: assessment of micro-nuclei induction. Mutat. Res. 414, 63–75.
C- avas, T., Ergene-Gozukara, S., 2005a. Genotoxicity evaluation of metronidazoleusing the piscine micronulceus test by acridine orange fluorescent staining.Environ. Toxicol. Pharmacol. 19, 107–111.
C- avas, T., Ergene-Gozukara, S., 2005b. Induction of micronuclei and nuclearabnormalities in Oreochromis niloticus following exposure to petroleumrefinery and chromium processing plant effluents. Aquat. Toxicol. 74,264–271.
C- avas, T., Garanko, N.N., Arkhipchuk, V.V., 2005. Induction of micronuclei in blood,gill and liver cells of fishes subchronically exposed to cadmium chloride andcopper sulphate. Food Chem. Toxicol. 43, 569–574.
C- avas, T., 2008. In vivo genotoxicity of mercury chloride and lead acetate:micronucleus test on acridine orange stained fish cells. Food Chem. Toxicol.46, 352–358.
Collins, A., Dusinska, M., Franklin, M., Somorovska, M., Petrovska, H., Duthie, S.,Fillion, L., Panayiotidis, M., Raslova, K., Vaughan, N., 1997. Comet assay inhuman biomonitoring studies: reliability, validation, and applications.Environ. Mol. Mutagen. 30 (2), 139–146.
CONAMA, 2005. Conselho Nacional do Meio Ambiente. Resoluc- ~ao 357.Das, R.K., Nanda, N.K., 1986. Induction of micronuclei in peripheral erythrocytes of
the fish, Heteropneustes fossilis, by mitomycin C and paper mill effluent. Mutat.Res. 175, 67–71.
De Flora, S., Bennicelli, C., Bagnasco, M., 1994. Genotoxicity of mercury com-pounds: a review. Mutat. Res. 317, 57–79.
Dinnen, R.D., Tomlinson, S.M., Hart, D., Chopra, C., Heddle, J.A., 1988. Application ofa micronucleus assay to the peripheral blood cells of rainbow trout, Salmogairdneri. Can. Tech. Rep. Fish. Aquat. Sci. 1607, 69–78.
Drevnick, P.E., Sandheinrich, M.B., Oris, J.T., 2006. Increased ovarian follicularapoptosis in fathead minnows (Pimephales promelas) exposed to dietarymethylmercury. Aquat. Toxicol. 79, 49–54.
Ergene, S., C- avas, T., Celik, A., Koleli, N., Kaya, F., Karahan, A., 2007. Monitoring ofnuclear abnormalities in peripheral erythrocytes of three fish species from theGoksu Delta (Turkey): genotoxic damage in relation to water pollution.Ecotoxicology 16, 385–391.
Ferraro, M.V.M., Fenocchio, A.S., Mantovani, M.S., Oliveira Ribeiro, C., Cestari, M.M.,2004. Mutagenic effects of tributyltin and inorganic lead (Pb II) on thefish H. malabaricus as evaluated using the comet assay and the piscinemicronucleus and chromosome aberration tests. Genet. Mol. Biol. 27 (1),103–107.
Fleeger, J.W., Carman, K.R., Nisbet, R.M., 2003. Indirect effects of contaminants inaquatic ecosystems. Sci. Total Environ. 317, 207–233.
Fostier, A.-H., Forti, M.C., Guimar~aes, J.R.D., Melfi, A.J., Boulet, R., Espirito Santo, C.M.,Krug, F.J., 2000. Mercury fluxes in a natural forested Amazonian catchment (Serrado Navio, Amapa State, Brazil). Sci. Total Environ. 260, 201–211.
Grandjean, P., 1999. Mercury risks: controversy or just uncertainty? Publ. HealthRep. 114 (6), 512–515.
Gordon, G., Skett, P., 1986. Introduction to Drug Metabolism. Chapman and Hall,New York, NY.
Guilherme, S., Valega, M., Pereira, M.E., Santos, M.A., Pacheco, M., 2008. Erythro-cytic nuclear abnormatilities in wild and caged fish (Liza aurata) along anenvironmental mercury contamination gradient. Ecotoxicol. Environ. Saf. 70(3), 411–421.
Hartmann, A., Agurell, E., Beevers, C., Brendler-Schwaab, S., Burlinson, B., Clay, P.,Collins, A., Smith, A., Speit, G., Thybaud, V., Tice, R.R., 2003. Fourth interna-tional comet assay workshop. Recommendations for conducting the in vivoalkaline Comet Assay. Mutagenesis 18, 45–51.
Hayashi, M., Ueda, T., Uyeno, K., Wada, K., Kinae, N., Saotome, K., Tanaka, N., Takai,A., Sasaki, Y.F., Asano, N., Sofuni, T., Ojima, Y., 1998. Development ofgenotoxicity assay systems that use aquatic organisms. Mutat. Res. 399,125–133.
Hooftman, R.N., Raat, W.K., 1982. Induction of nuclear anomalies (micronuclei) inthe peripheral blood erythrocytes of the eastern mudminnow Umbra pygmaeaby ethyl methanesulphonate. Mutat. Res. 104, 147–152.
Hose, J.E., Cross, J.N., Smitr, S.G., Diehl, D., 1987. Elevated circulating erythrocytesmicronuclei in fishes from contaminated sites of southern California. Mar.Environ. Res. 22, 167–176.
Heddle, J.A., Cimino, M.C., Hayashi, M., Romagna, M.D., Tucker, J.D., Vanprais, P.H.,Macgregor, J.T., 1991. Micronuclei as an index of Cytogenetic Damage: past,present and future. Environ. Mol. Mutagen. 18, 277–291.
Kehrig, H.A., Malm, O., Moreira, I., 1998. Mercury in a widely consumed fishMicropogonias furnieri (Demarest, 1823) from four main Brazilian estuaries.Sci. Total Environ. 213, 263–271.
Lebel, J., Mergler, D., Branches, F., Lucotte, M., Amorim, M., Larribe, F., Dolbec, J.,1998. Neurotoxic effects of low-level methylmercury contamination in theAmazonian Basin. Environ. Res. 79 (1), 20–32.
Please cite this article as: Vicari, T., et al., Genotoxic evaluation of difEcotoxicol. Environ. Saf. (2012), http://dx.doi.org/10.1016/j.ecoenv.2
Lee, R.F., Steinert, S., 2003. Use of the single cell gel electrophoresis/comet assayfor detecting DNA damage in aquatic (marine and freshwater) animals. Mutat.Res.—Rev. Mutat. Res. 544, 43–64.
Limke, T.L., Heidemann, S.F., Atchison, W.D., 2004. Disruption of intraneuronaldivalent cation regulation by methylmercury: are specific targets involved inaltered neuronal development and cytotoxicity in methylmercury poisoning?Neurosci. Toxicol. 25, 741–760.
Loureiro, V.E., Hahn, N.S., 1996. Dieta e atividade alimentar da traıra, Hopliasmalabaricus (Bloch, 1794) (Osteichthyes, Erythrinidae), nos primeiros anos deformac- ~ao do reservatorio de Segredo-PR. Acta Limnol. Bras. 8, 195–205.
Malm, O., Branches, F.J., Akagi, H., Castro, M.B., Pfeiffer, W.C., Harada, M., Bastos,W.R., Kato, H., 1995. Mercury and methyl mercury in fish and human hair fromthe Tapajos river basin. Braz. Sci. Total Environ. 175, 141–150.
Mela, M., Randi, M.A.F., Ventura, D.F., Carvalho, C.E.V., Pelletier, E., Oliveira Ribeiro,C.A., 2007. Effects of dietary methyl mercury on liver and kidney histology inthe neotropical fish Hoplias malabaricus. Ecotoxicol. Environ. Saf. 68, 426–435.
Meseguer, J., Lopez-Ruiz, A., Esteban, M.A., 1994. Melano-macrophages of theseawater teleosts, sea bass (Dicentrarcus labrax) and gilthead seabream (Sparusaurata): morphology, formation and possible function. Cell Tissue Res. 277, 1–10.
Mirlean, N., Larned, S.T., Nikora, V., Kutter, V.T., 2005. Mercury in lakes and lakefishes on a conservation-industry gradient in Brazil. Chemosphere 60,226–236.
Nepomuceno, J.C., Ferrari, I., Spano, M.A., Centeno, A.J., 1997. Detection ofmicronuclei in peripheral erythrocytes of Cyprinus carpio exposed to metallicmercury. Environ. Mol. Mutagen. 30, 293–297.
Oliveira, M., Maria, V.L., Ahmada, I., Teles, M., Serafim, A., Bebianno, M.J., Pacheco,M., Santos, M.A., 2010. Golden grey mullet and sea bass oxidative DNA damageand clastogenic/aneugenic responses in a contaminated coastal lagoon. Eco-toxicol. Environ. Saf. 73, 1907–1913.
Oliveira Ribeiro, C.A., Guimar~aes, J.R., Pffeiffer, W.C., 1996. Accumulation anddistribution of inorganic mercury in a tropical fish (Trichomycterus zonatus).Ecotoxicol. Environ. Saf. 34, 190–195.
Oliveira Ribeiro, C.A., Rouleau, C., Pelletier, E., Audet, C., Tjalve, H., 1999. Distribu-tion kinetics of dietary methylmercury in the artic charr (Salvelinus alpinus).Environ. Sci. Technol. 33, 902–907.
Pacheco, M., Santos, M.A., 2002. Biotransformation, genotoxic, and histopatholo-gical effects of environmental contaminants in European eel (Anguilla angu-illa). Ecotoxicol. Environ. Saf. 53, 331–347.
Pandrangi, R., Petras, M., Ralph, S., Vrzoc, M., 1995. Alkaline single cell (comet):assay and genotoxicity monitoring using bullhead and carp. Environ. Mol.Mutagen. 26, 345–356.
Pfeiffer, W.C., Malm, O., Souza, C.M.M., Silveira, E.G., Bastos, W.R., 1989. Mercuryconcentration in inland waters of gold mining areas in Rondonia, Brazil. Sci.Total Environ. 87, 33–240.
Pfeiffer, W.C., Malm, O., Souza, C.M.M., Silveira, E.G., Bastos, W.R., 1991. Mercuryin the Madeira river ecosystem Rondonia, Brazil. For. Ecol. Manage. 38,239–245.
Porto, J.I.R., Araujo, C.S.O., Feldberg, E., 2005. Mutagenic effects of mercurypollution as revealed by micronucleus test on three Amazonian fish species.Environ. Res. 97, 287–292.
Ramsdorf, W.A., Ferraro, M.V.M., Oliveira-Ribeiro, C.A., Costa, J.R.M., Cestari, M.M.,2009. Genotoxic evaluation of different doses of inorganic lead (PbII) in Hopliasmalabaricus. Environ. Monit. Assess. 158, 77–85.
Ravaglia, M.A., Meseguer, M.C., 1995. Melano-macrophage centers in thegonads of the swamp eel, Synbranchus marmoratus Bloch, (Pisces, Synbran-chidae): histological and histochemical characterization. J. Fish. Distrib. 18,117–125.
Roulet, M., Lucotte, M., Canuel, R., Farella, N., Goch, Y.G., Peleja, J.R., Guimar~aes,J.R.D., Mergler, D., Amorim, M., 2001. Spatio-temporal geochemistry ofmercury in waters of the Tapajos and the Amazonian rivers. Oceanogr 46,1141–1157.
Santos, E.C.O., Maura de Jesus, I., Brado, E.S., Loureiro, E.B., Mascarenhas, A.F.S.,Weirich, J., Camara, V.C., Cleary, D., 2002. Mercury exposures in riversideAmazon communities in Para, Brazil. Environ. Res. Sect. 84, 100–107.
Serrano-Garcia, L., Montero-Montoya, R., 2001. Micronuclei and chromatid budsare the result of related genotoxic events. Environ. Mol. Mutagen. 38 (1),38–45.
Schmid, W., 1975. The micronucleus test. Mutat. Res. 31, 9–15.Silva-Forsberg, M.C., Forsberg, B.R., Zeidemann, V.K., 1999. Hg contamination in
humans linked to river chemistry in the Amazon basin. Ambio 28, 519–521.Singh, N.P., McCoy, M.T., Tice, R.R., Schneider, E.L., 1988. A simple technique for
quantification of low levels of DNA damage in individual cells. Exp. Cell Res.175, 184–191.
Smith, I.R., 1990. Erythrocytic micronuclei in wild fish from Lakes Superior andOntario that have pollution-associated neoplasias. J. Great Lakes Res. 16,139–142.
Snyder, R.D., David, G.F., Lachmann, P.J., 1989. Inhibition by metals of X-ray andultraviolet-induced DNA repair in human cells. Biol. Trace Elem. Res. 21,389–398.
Souza Lima, A.P., Muller, R.C.S., Sarkis, J.E.S., Alves, C.N., Bentes, M.H.S., Brado, E.,Santos, E.O., 2000. Mercury contamination in fish from Santarem, Para, Brazil.Environ. Res. Sect. 83, 117–122.
Suzuki, Y., Nagae, Y., Li, J., Sabaka, H., Mazowa, K., Takahashi, A., Shimuzu, H., 1989.The micronucleus test and erythropoiesis: effects of erythropoietin anda mutagen on the ratio of polychromatic to normochromatic erythrocytes(P/N ratio). Mutagenesis 4, 420–424.
ferent doses of methylmercury (CH3Hgþ) in Hoplias malabaricus.012.05.007

T. Vicari et al. / Ecotoxicology and Environmental Safety ] (]]]]) ]]]–]]] 9
Udroiu, I., 2006. The micronucleus test in piscine erythrocytes. Aquat. Toxicol. 79,201–204.
Ueda, T., Hayashi, M., Koide, N., Sofuni, T., Kobayashi, J., 1992. A preliminary studyof the micronucleus test by acridine orange fluorescent staining comparedwith chromosomal aberration test using fish erythropoietic and embryoniccells. Water Sci. Technol. 25, 235–240.
Van der Oost, R., Beyer, J., Vermeulen, N.P.E., 2003. Fish bioaccumulation andbiomarkers in environmental risk assessment: a review. Environ. Toxicol.Pharmacol. 13, 57–149.
Vassalo, D.V., Massaroni, L., Oliveira, E.M., Rossoni, L.V., Amaral, S.M.C., Vassalo,P.F., 1996. Ac- ~oes toxicas e agudas do mercurio sobre o aparelho cardiovascular.
Please cite this article as: Vicari, T., et al., Genotoxic evaluation of difEcotoxicol. Environ. Saf. (2012), http://dx.doi.org/10.1016/j.ecoenv.2
Centro Biomedico da UFGS, Vitoria e Hospital Universitario da UFSM, SantaMaria RS-Vitoria ES. Arq. Bras. Cardiol. 67, 54–61.
WHO, 1990. World Health Organization. Environmental Health Criteria 101.
Methylmercury. World Health Organization, Geneva, Switzerland.Williams, R.C., Metcalfe, C.D., 1992. Development of an in vitro hepatic micro-
nucleus assay with rainbow trout. Aquat. Toxicol. 23, 193–202.Wren, C.D., Harris, S., Harttrup, N., 1995. Ecotoxicology of Mercury and Cadmium.
In: Hoffman, D.J., Rattner, B.A., Burton, G.A.J.R., Cairns, J.J.R. (Eds.), Handbook ofEcotoxicology. Lewis, Boca Raton, FL, cap.17.
ferent doses of methylmercury (CH3Hgþ) in Hoplias malabaricus.012.05.007





























