Embed Size (px)
Citation preview

Plant Biotechnology Journal
(2006)
4
, pp. 251–261 doi: 10.1111/j.1467-7652.2005.00178.x
© 2006 Blackwell Publishing Ltd
251
tBlackwell Publishing, Ltd.Oxford, UKPBIPlant Biotechnology Journal1467-7644© 2005 Blackwell Publishing Ltd? 20052?Original Article
Transformation of barley pollen cultures
Jochen Kumlehn
et al.
Genetic transformation of barley (
Hordeum vulgare
L.) via infection of androgenetic pollen cultures with
Agrobacterium tumefaciens
Jochen Kumlehn
1,2,
*, Liliya Serazetdinova
2,
†, Goetz Hensel
1
, Dirk Becker
2
and Horst Loerz
2
1
Institute of Plant Genetics and Crop Plant Research Gatersleben, Plant Reproductive Biology, Corrensstr. 3, 06466 Gatersleben, Germany
2
University of Hamburg, Biocentre Klein Flottbek, Developmental Biology and Biotechnology, Ohnhorststr. 18, 22609 Hamburg, Germany
Summary
A novel genetic transformation method for barley (
Hordeum vulgare
L.), based on infection
of androgenetic pollen cultures with
Agrobacterium tumefaciens
, is presented. Winter-type
barley cv. ‘Igri’ was amenable to stable integration of transgenes mediated by
A.
tumefaciens
strain LBA4404 harbouring a vector system that confers hypervirulence, or by
the non-hypervirulent strain GV3101 with a standard binary vector. The efficacy of gene
transfer was substantially influenced by pollen pre-culture time, choice of
Agrobacterium
strain and vector system,
Agrobacterium
population density, medium pH and the
concentrations of acetosyringone, CaCl
2
and glutamine. After co-culture, rapid removal of
viable agrobacteria was crucial for subsequent development of the pollen culture. To this
end, the growth of agrobacteria was suppressed by the concerted effects of appropriate
antibiotics, low pH, reduced level of glutamine and high concentrations of CaCl
2
and
acetosyringone. Following infection with LBA4404 and GV3101, about 31% and 69%,
respectively, of the primary transgenic (T
0
) plants carried a single copy of the sequence
integrated. The use of hypervirulent
A. tumefaciens
and hygromycin resistance as a
selectable marker resulted in 3.7 T
0
plants per donor spike. About 60% of the primary
transgenic plants set seed, indicating spontaneous genome doubling. An analysis of 20 T
1
populations revealed that four progenies did not segregate for reporter gene expression.
This indicates that the approach pursued enables the generation of instantly homozygous
primary transgenic plants. The method established will be a valuable tool in functional
genomics as well as for the biotechnological improvement of barley.
Received 12 May 2005;
revised 21 October 2005;
accepted 27 October 2005.
*
Correspondence
(fax +49394825692;
e-mail [email protected])
†
Present address
: Sainsbury Laboratory,
John Innes Centre, Colney Lane,
Norwich NR4 7UH, Norfolk, UK
Keywords:
Agrobacterium
,
androgenesis, barley, genetic
transformation,
Hordeum vulgare
,
pollen embryogenesis.
Introduction
Reliable and efficient techniques to genetically transform
important experimental model plant species, as well as crops,
are required for comprehensive functional gene analyses and
molecular breeding. As an important food and feed crop grown
worldwide, barley has been used as a model species in cereal
genetic research for decades. The barley genetic resources
generated to date (e.g. Close
et al
., 2004; Zhang
et al
., 2004)
are now being intensively employed in diverse research
approaches and, for many sequences, a detailed functional
characterization via stable genetic transformation is desirable.
Stable transformation of barley has been performed by
direct DNA transfer to immature embryos (Wan and Lemaux,
1994), androgenetic pollen (Jaehne
et al
., 1994), shoot mer-
istematic cultures (Zhang
et al
., 1999), protoplasts (Funatsuki
et al
., 1995) or isolated zygotes (Holm
et al
., 2000). Tingay
et al
. (1997) were the first to establish a protocol for
Agro-
bacterium
-mediated transformation of barley. Later, basically
the same method, which is based on the infection of imma-
ture embryos with agrobacteria, was also employed in other
laboratories (Horvath
et al
., 2000; Patel
et al
., 2000; Trifonova
et al
., 2001; Wang
et al
., 2001; Fang
et al
., 2002; Stahl
et al
.,
2002). In more recently published reports, transformation

252
Jochen Kumlehn
et al.
© Blackwell Publishing Ltd,
Plant Biotechnology Journal
(2006),
4
, 251–261
efficiencies higher than 10%, based on the number of in-
fected embryos, were achieved (Matthews
et al
., 2001; Hensel
and Kumlehn, 2004; Murray
et al
., 2004).
Agrobacterium
-
mediated transformation of cereals is still confined to a few
selected genotypes. For example, in barley, only a few spring-
type lines have been found to be useful to some extent (Tingay
et al
., 1997; Wang
et al
., 2001; Murray
et al
., 2004), whereas,
so far, there is no example of genetic transformation of a
barley winter-type line. In particular, the spring-type cv. ‘Golden
Promise’ is extraordinarily amenable to
Agrobacterium
-
mediated transformation of immature embryos.
In comparison with direct gene transfer methods,
Agrobacterium
-mediated transformation is basically governed
by a naturally evolved mechanism (Tinland, 1996; Zupan and
Zambryski, 1997). It predominantly results in the integration
of single or low copy numbers of transferred DNA per
transformed cell, which supports expression stability in
subsequent generations. Barakat
et al
. (2000) revealed
a predisposition of agrobacteria to integrate T-DNAs into
transcriptionally active regions of the rice genome. Further
advantages of
Agrobacterium
-mediated transformation
include the high potential for independent integration of
co-transformed DNA fragments, the transfer of large DNA
sequences (Hamilton, 1997) and the exclusion of backbone
sequences from the DNA introduced into the plant genome.
Immature barley pollen at the vacuolated microspore or
early bicellular stage can deviate from the normal process of
pollen formation to undergo androgenesis (Sunderland
et al
.,
1978). The most interesting aspect of genetic transformation
of haploid target cells, such as microspores, is that transgene
integration followed by genome doubling opens up the unique
opportunity to immediately generate plants homozygous for
the gene transferred. In addition to their haploid nature, the
incomparable potential for mass regeneration makes immature
pollen cultures an attractive target for
Agrobacterium
-based
transformation. Sangwan
et al
. (1993) showed the generation
of transgenic
Datura innoxia
and
Nicotiana tabacum
plants
following
Agrobacterium
infection of cotyledonary-stage
androgenetic embryos; however, no transformation was
obtained using freshly isolated immature pollen or andro-
genetic pro-embryos. Successful
A. tumefaciens
-mediated
transformation of barley pollen-derived cell suspension
cultures, obtained by Wu
et al
. (1998), did not result in the
regeneration of transgenic plants. Reports on the generation
of transgenic
Brassica napus
plants following co-culture of
androgenetic pollen with
A. tumefaciens
, published by Pechan
(1989) and disclosed by Dormann
et al
. (2001), lacked com-
pelling evidence for the integration of transgenes in the plant
genome, their inheritance in following generations and the
instant homozygosity of the genes transferred. In the present
study, we have identified and optimized parameters influenc-
ing gene transfer and target cell survival to eventually estab-
lish an efficient and reproducible method of generating
transgenic barley following infection of androgenetic pollen
cultures with agrobacteria. We show that the transgenes are
stably integrated into the barley genome, that these genes
are expressed and inherited in the following generations, and
that some of the primary transgenic plants obtained are
homozygous for the gene transferred.
Results and discussion
Co-culture of androgenetic barley pollen with
A. tumefaciens
Preliminary experiments were carried out with varying pollen
pre-culture times prior to co-culture with the
A. tumefaciens
strain LBA4404 carrying the co-integrative vector pSB1:UG35PAT.
Infection of pollen within the first 5 days after isolation
resulted in no, or extremely rare, transformation events. The
optimal temporal window of efficient infection with agro-
bacteria was determined to be after 6–11 days of pre-culture
(Figure 1A,B).
At least three classes of environmental stimuli act syner-
gistically to induce the transformation machinery of
A. tume-
faciens
, namely monocyclic aromatic hydrocarbons, such as
acetosyringone, monosaccharides, such as glucose, and
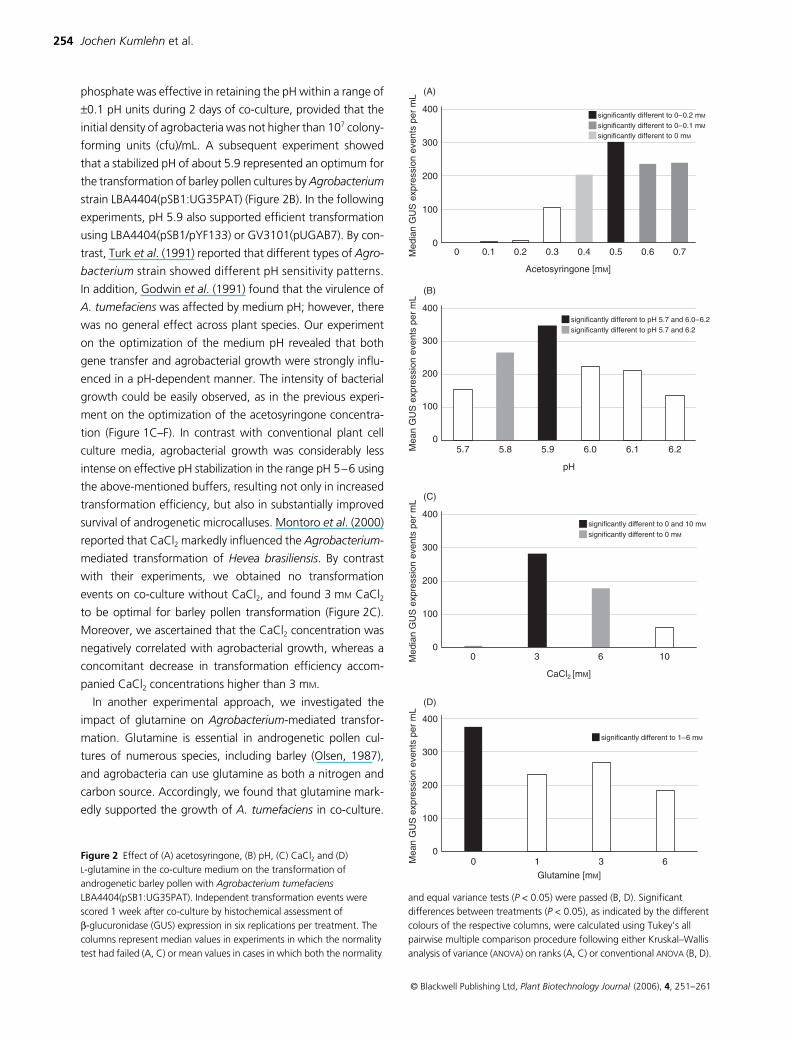
acidic pH (Winans, 1991). Acetosyringone plays a dual role in
Agrobacterium
-mediated transformation. First, it serves as a
chemotacticum for the spatial orientation of agrobacteria to
wounded sites of plant tissue (Ashby
et al
., 1987). Second, it
elicits the agrobacterial virulence machinery (Stachel
et al
.,
1985). In addition, we have shown that acetosyringone not
only enhances the transformation activity of
A. tumefaciens
(Figure 2A), but can also substantially limit its multiplication.
Figure 1C displays the overgrowth of the pollen culture in the
absence of acetosyringone, whereas the bacterial growth in
direct contact with the Petri dish, as well as the aggregation
of the androgenetic pollen grains, markedly decreased with
increasing concentration of acetosyringone (Figure 1D–F).
However, concentrations beyond 0.5 m
M
did not further
improve the transformation efficiency (Figure 1E,F).
The moderately acidic pH in the vicinity of wounded plant sites
has been found to activate
A. tumefaciens Vir
genes (Li
et al
.,
2002). Our preliminary attempts to effectively stabilize the
medium pH as a prerequisite for subsequent optimization
revealed that a combination of 10 m
M
2-morpholinoethan-
esulphonic acid (MES) and 50 m
M
potassium hydrogen
Transformation of barley pollen cultures
253
© Blackwell Publishing Ltd,
Plant Biotechnology Journal
(2006),
4
, 251–261
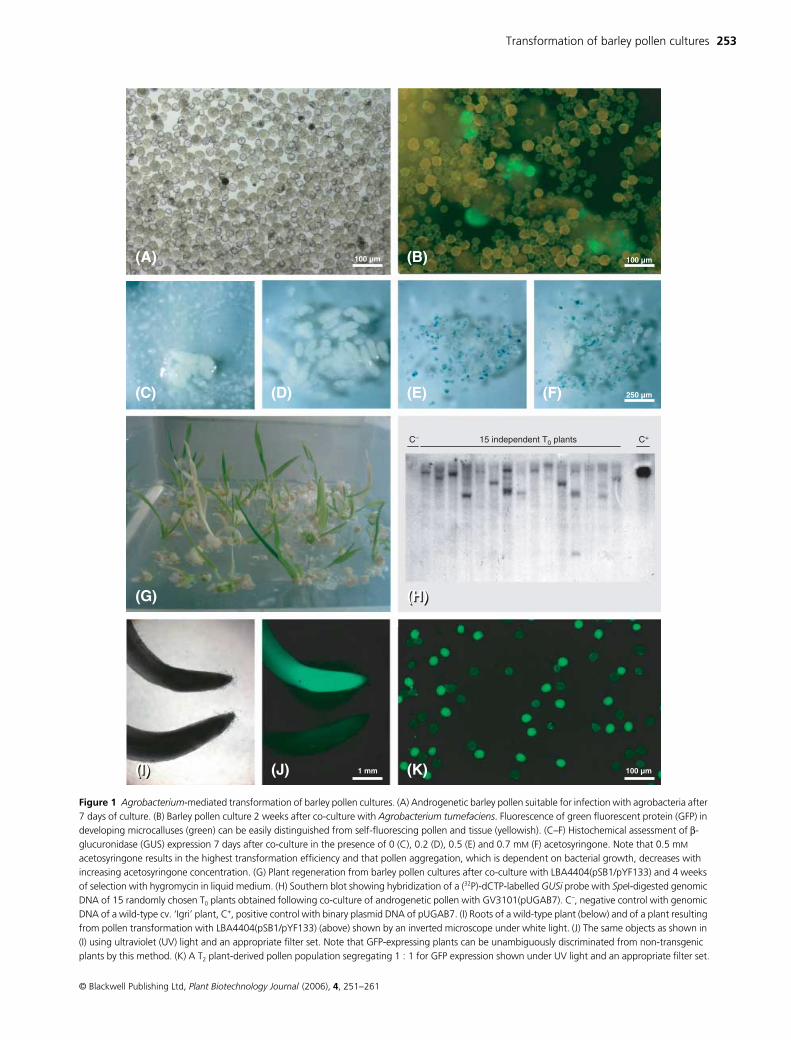
Figure 1 Agrobacterium-mediated transformation of barley pollen cultures. (A) Androgenetic barley pollen suitable for infection with agrobacteria after 7 days of culture. (B) Barley pollen culture 2 weeks after co-culture with Agrobacterium tumefaciens. Fluorescence of green fluorescent protein (GFP) in developing microcalluses (green) can be easily distinguished from self-fluorescing pollen and tissue (yellowish). (C–F) Histochemical assessment of β-glucuronidase (GUS) expression 7 days after co-culture in the presence of 0 (C), 0.2 (D), 0.5 (E) and 0.7 mM (F) acetosyringone. Note that 0.5 mM acetosyringone results in the highest transformation efficiency and that pollen aggregation, which is dependent on bacterial growth, decreases with increasing acetosyringone concentration. (G) Plant regeneration from barley pollen cultures after co-culture with LBA4404(pSB1/pYF133) and 4 weeks of selection with hygromycin in liquid medium. (H) Southern blot showing hybridization of a (32P)-dCTP-labelled GUSi probe with SpeI-digested genomic DNA of 15 randomly chosen T0 plants obtained following co-culture of androgenetic pollen with GV3101(pUGAB7). C–, negative control with genomic DNA of a wild-type cv. ‘Igri’ plant, C+, positive control with binary plasmid DNA of pUGAB7. (I) Roots of a wild-type plant (below) and of a plant resulting from pollen transformation with LBA4404(pSB1/pYF133) (above) shown by an inverted microscope under white light. (J) The same objects as shown in (I) using ultraviolet (UV) light and an appropriate filter set. Note that GFP-expressing plants can be unambiguously discriminated from non-transgenic plants by this method. (K) A T2 plant-derived pollen population segregating 1 : 1 for GFP expression shown under UV light and an appropriate filter set.
254
Jochen Kumlehn
et al.
© Blackwell Publishing Ltd,
Plant Biotechnology Journal
(2006),
4
, 251–261
phosphate was effective in retaining the pH within a range of
±
0.1 pH units during 2 days of co-culture, provided that the
initial density of agrobacteria was not higher than 10
7
colony-
forming units (cfu)/mL. A subsequent experiment showed
that a stabilized pH of about 5.9 represented an optimum for
the transformation of barley pollen cultures by
Agrobacterium
strain LBA4404(pSB1:UG35PAT) (Figure 2B). In the following
experiments, pH 5.9 also supported efficient transformation
using LBA4404(pSB1/pYF133) or GV3101(pUGAB7). By con-
trast, Turk
et al
. (1991) reported that different types of
Agro-
bacterium
strain showed different pH sensitivity patterns.
In addition, Godwin
et al
. (1991) found that the virulence of
A. tumefaciens
was affected by medium pH; however, there
was no general effect across plant species. Our experiment
on the optimization of the medium pH revealed that both
gene transfer and agrobacterial growth were strongly influ-
enced in a pH-dependent manner. The intensity of bacterial
growth could be easily observed, as in the previous experi-
ment on the optimization of the acetosyringone concentra-
tion (Figure 1C–F). In contrast with conventional plant cell
culture media, agrobacterial growth was considerably less
intense on effective pH stabilization in the range pH 5–6 using
the above-mentioned buffers, resulting not only in increased
transformation efficiency, but also in substantially improved
survival of androgenetic microcalluses. Montoro
et al
. (2000)
reported that CaCl
2
markedly influenced the
Agrobacterium
-
mediated transformation of
Hevea brasiliensis
. By contrast
with their experiments, we obtained no transformation
events on co-culture without CaCl
2
, and found 3 m
M
CaCl
2
to be optimal for barley pollen transformation (Figure 2C).
Moreover, we ascertained that the CaCl
2
concentration was
negatively correlated with agrobacterial growth, whereas a
concomitant decrease in transformation efficiency accom-
panied CaCl
2
concentrations higher than 3 m
M
.
In another experimental approach, we investigated the
impact of glutamine on
Agrobacterium
-mediated transfor-
mation. Glutamine is essential in androgenetic pollen cul-
tures of numerous species, including barley (Olsen, 1987),
and agrobacteria can use glutamine as both a nitrogen and
carbon source. Accordingly, we found that glutamine mark-
edly supported the growth of
A. tumefaciens
in co-culture.
Figure 2
Effect of (A) acetosyringone, (B) pH, (C) CaCl
2
and (D) L-glutamine in the co-culture medium on the transformation of androgenetic barley pollen with Agrobacterium tumefaciens LBA4404(pSB1:UG35PAT). Independent transformation events were scored 1 week after co-culture by histochemical assessment of β-glucuronidase (GUS) expression in six replications per treatment. The columns represent median values in experiments in which the normality test had failed (A, C) or mean values in cases in which both the normality
and equal variance tests (P < 0.05) were passed (B, D). Significant differences between treatments (P < 0.05), as indicated by the different colours of the respective columns, were calculated using Tukey’s all pairwise multiple comparison procedure following either Kruskal–Wallis analysis of variance (ANOVA) on ranks (A, C) or conventional ANOVA (B, D).
Transformation of barley pollen cultures 255
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2006), 4, 251–261
Furthermore, our study demonstrated that glutamine did not
enhance the transformation activity of agrobacteria and, sur-
prisingly, it appeared not to be required during the 2 days of
co-culture for androgenetic development to proceed (Figure 2D).
Effect of Agrobacterium strain and vector system
Transgenic barley plants were produced using three different
Agrobacterium strain–vector combinations. The generation
of a superbinary vector, as suggested by Komari et al. (1996),
was comparatively intricate and we were unable to maintain
the resultant strain LBA4404(pSB1:UG35PAT) for more than
a few months. As a consequence, we pursued an alternative
approach based on the introduction of the non-integrative
binary vector pYF133 (Fang et al., 2002) into the hypervirulent
strain LBA4404(pSB1). The resultant LBA4404(pSB1/pYF133)
was the most efficient with regard to the generation of trans-
genic plants (Table 1). The substantially lower transformation
efficiency obtained with LBA4404(pSB1:UG35PAT) can prob-
ably be attributed to the PAT gene used as a selectable
marker. However, effects associated with the different vector
construction cannot be ruled out. In addition to hypervirulent
agrobacteria, we tested the non-hypervirulent A. tumefa-
ciens strain GV3101 containing the standard binary vector
pUGAB7. As a result, transformation of barley pollen cultures
was obtained with an efficiency comparable with that of
LBA4404(pSB1:UG35PAT) (Table 1).
Removal of agrobacteria and selective development of
transgenic tissue and plants
Based on the results from the co-culture experiments, the ASt
medium composition (see Table 3) was optimized in prelim-
inary small-scale experiments in order to effectively suppress
agrobacterial growth immediately after transformation, as
well as to ensure ongoing androgenetic development. In
particular, the pH was adjusted to 5.0 and stabilized using
10 mM MES together with 25 mM potassium hydrogen phos-
phate, the CaCl2 concentration was increased to 10 mM, ace-
tosyringone was added to obtain a concentration of 0.5 mM,
and glutamine was added to obtain a concentration of 1 mM.
These treatments markedly contributed to the control of
agrobacteria, whereas the androgenetic development was
apparently not adversely affected (data not shown). Further-
more, a combination of the antibiotics amoxicillin (100 mg/L)
and cefotaxime (125 mg/L) was successfully used to remove
viable agrobacteria. After 2 weeks of culture in ASt medium,
we checked the presence of viable agrobacteria by transfer-
ring some pollen-derived calluses on to solid Luria–Bertani
(LB) medium without the addition of antibiotics. Not a single
case of outgrowing agrobacteria was detected, indicating their
effective removal under the conditions used after co-culture.
Hygromycin (50 µM) or Bialaphos (75 µM), added immediately
after co-culture, was useful to prevent substantial formation
of non-transgenic tissue and plants. One week after co-culture,
the transgenic microcalluses tolerated even further increased
concentrations of selective agent. Accordingly, S medium
(see Table 3) was supplemented with 100 µM of either hygro-
mycin or Bialaphos.
A considerable delay in the generation of plantlets from
transformed pollen cultures was observed when selective
pressure was applied during regeneration in preliminary
experiments. The experiments presented here demonstrate
that plantlet formation is more efficient and the proportion
of non-transgenic ‘escapes’ remains low when the selective
agent is entirely omitted from the regeneration medium
(Figure 1G, Table 1). This observation can be explained by the
strong selective pressure during previous liquid culture.
Integration of transgenes into the barley genome and
expression of recombinant protein
All candidate primary transgenic plants were analysed using
Southern blotting to reveal stable transgene integration within
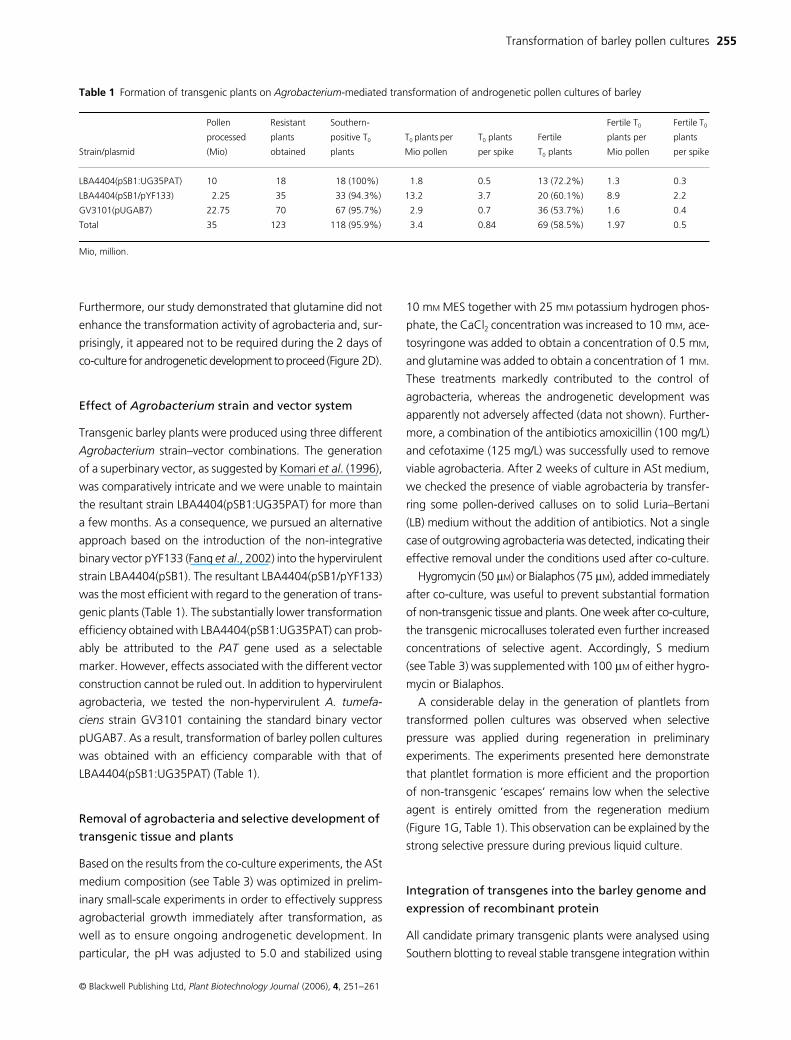
Table 1 Formation of transgenic plants on Agrobacterium-mediated transformation of androgenetic pollen cultures of barley
Strain/plasmid
Pollen
processed
(Mio)
Resistant
plants
obtained
Southern-
positive T0
plants
T0 plants per
Mio pollen
T0 plants
per spike
Fertile
T0 plants
Fertile T0
plants per
Mio pollen
Fertile T0
plants
per spike
LBA4404(pSB1:UG35PAT) 10 18 18 (100%) 1.8 0.5 13 (72.2%) 1.3 0.3
LBA4404(pSB1/pYF133) 2.25 35 33 (94.3%) 13.2 3.7 20 (60.1%) 8.9 2.2
GV3101(pUGAB7) 22.75 70 67 (95.7%) 2.9 0.7 36 (53.7%) 1.6 0.4
Total 35 123 118 (95.9%) 3.4 0.84 69 (58.5%) 1.97 0.5
Mio, million.
256 Jochen Kumlehn et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2006), 4, 251–261
their genome (Figure 1H). In the case of transformation using
LBA4404(pSB1/pYF133), followed by selection based on
hygromycin, transgene integration was missing in only two of
35 regenerated plants (Table 1). Compared with hygromycin,
the effect of Bialaphos was markedly less efficient in vitro.
Using GV3101(pUGAB7), a total of 174 plantlets was
regenerated, only 70 (40.2%) of which survived spraying
with BASTA solution. Yet, about 96% and 100% of the
surviving plants derived from co-culture using GV3101(pSB1/
UGAB7) or LBA4404(pSB1:UG35PAT), respectively, showed
transgene integration according to Southern hybridization
(Table 1), indicating that spraying with BASTA can be
considered as a fairly reliable method. Southern blotting
analyses further revealed that 16 of 51 (31.4%) and 46 of 67
(68.6%) T0 plants obtained following co-culture with
LBA4404 and GV3101, respectively, carried a single copy of
the transferred T-DNA. Expression of green fluorescent
protein (GFP) was unambiguously shown by fluorescence
microscopy of root tips in 93.9% of Southern-positive T0
plants transformed with LBA4404(pSB1/pYF133) (Figure 1I,J ).
Generative transgene transmission to T1 plants was also
revealed by Southern blotting (not shown). Fluorescence
microscopy further demonstrated one-to-one segregation of
GFP expression in pollen populations of hemizygous single-
copy T2 plants, which provides evidence for sustainable trans-
mission, segregation and expression of the integrated GFP
gene (Figure 1K).
Considering that spontaneous genome doubling appears
to preferentially occur early in androgenetic development,
and that transformation was feasible only after a consider-
able period of pollen pre-culture time, it was surprising that
four of 20 T1 populations did not segregate for marker
gene expression (Table 2), thus indicating that spontaneous
genome doubling must have taken place in some cases
subsequent to the transformation event. Such late spontaneous
genome doubling events have recently been observed by
González-Melendi et al. (2005).
Conclusions
A novel technology has been established for Agrobacterium-
based genetic transformation of barley. Compared with
immature embryo-based transformation, the system pre-
sented here has several advantages. The spikes are harvested
at a markedly earlier developmental stage, i.e. a higher
throughput of donor plants can be obtained with a given
glasshouse capacity, and, as the donor plants can be dis-
carded prior to anthesis, phytopathogens can be controlled
more easily. Moreover, the microspores from 30 spikes can
be isolated and transferred to culture within about 1 h. Given
Table 2 Transgene segregation in 20 T1 populations derived from Agrobacterium-mediated transformation of barley pollen cultures
T0 plants
Reporter gene
integrated
No. of copies
according to
Southern blot
Number of T1
plants analysed
Reporter gene
expression/no
expression
Segregation
observed
(assumed)
χ2
value
Likelihood (P)
according to
χ2 test
1 GUSi 1 20 14 : 6 2.33 : 1 (3 : 1) 0.24 > 0.5
2 GUSi 1 24 18 : 6 3.0 : 1 (3 : 1) 0 > 0.99
3 GUSi 2 43 37 : 6 6.17 : 1 (15 : 1) 1.9 > 0.1
4 GUSi 1 26 17 : 9 1.89 : 1 (3 : 1) 1.06 > 0.3
5 GUSi 2 39 34 : 5 6.8 : 1 (15 : 1) 1.32 > 0.2
6 GUSi 1 22 22 : 0 22 : 0 (22 : 0) 0 > 0.99
7 GUSi 1 51 40 : 9 3.64 : 1 (3 : 1) 0.35 > 0.5
8 GUSi 1 51 34 : 17 2.0 : 1 (3 : 1) 1.59 > 0.2
9 GUSi 1 50 32 : 18 1.78 : 1 (3 : 1) 2.63 > 0.1
10 GUSi 1 53 42 : 11 3.82 : 1 (3 : 1) 0.58 > 0.3
11 GUSi 1 16 16 : 0 16.0 : 0 (16 : 0) 0 > 0.99
12 GFP 1 46 35 : 11 3.18 : 1 (3 : 1) 0.03 > 0.8
13 GFP 2 43 37 : 6 6.17 : 1 (15 : 1) 1.9 > 0.1
14 GFP 2 27 23 : 4 5.75 : 1 (15 : 1) 1.42 > 0.2
15 GFP 1 20 20 : 0 20.0 : 0 (20 : 0) 0 > 0.99
16 GFP 1 18 12 : 6 2.0 : 1 (3 : 1) 0.56 > 0.3
17 GFP 2 21 19 : 2 9.5 : 1 (15 : 1) 0.2 > 0.5
18 GFP 1 24 24 : 0 24.0 : 0 (24 : 0) 0 > 0.99
19 GFP 2 14 12 : 2 6.0 : 1 (15 : 1) 0.66 > 0.3
20 GFP 2 19 16 : 3 5.3 : 1 (15 : 1) 1.19 > 0.2
Transformation of barley pollen cultures 257
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2006), 4, 251–261
10 transgenic plants per 100 embryos and 15 caryopses used
per donor spike in Agrobacterium-mediated transformation
using barley immature embryos, the efficiency of 2.2 fertile
transgenic plants obtained per spike in this study compares
well with that obtained by embryo transformation. The par-
ticular value of the method established in this work is that a
considerable number of plants which are homozygous for
the gene transferred can be found amongst the primary
transgenics. Although segregation analysis is also necessary
in this approach, homozygous plants can be identified one
generation earlier in comparison with conventional transfor-
mation methods.
There are several reasons why further improvements of the
newly developed technology can be expected. The majority
of spring- and winter-type genotypes are amenable to
doubled haploid formation via pollen culture. This appears to
be promising with regard to further transformation experi-
ments of diverse barley breeding lines. The surprisingly high
proportion of haploid primary transformants, together with
the amenability of barley to induced genome doubling
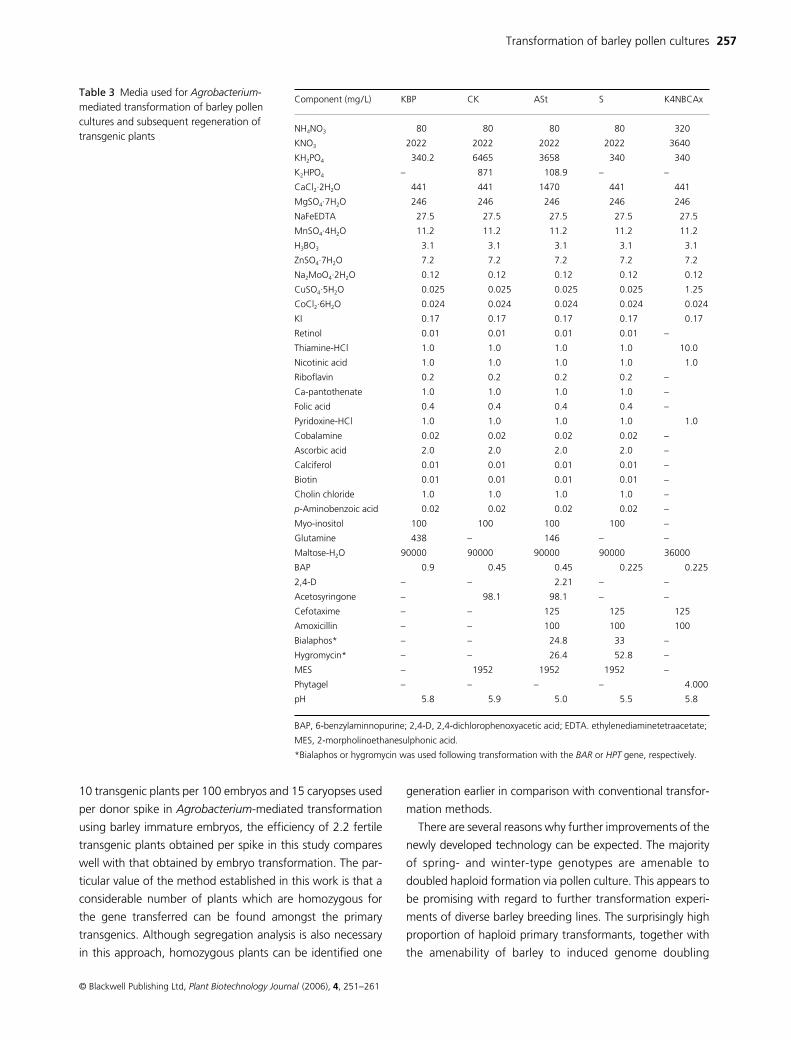
Component (mg/L) KBP CK ASt S K4NBCAx
NH4NO3 80 80 80 80 320
KNO3 2022 2022 2022 2022 3640
KH2PO4 340.2 6465 3658 340 340
K2HPO4 – 871 108.9 – –
CaCl2·2H2O 441 441 1470 441 441
MgSO4·7H2O 246 246 246 246 246
NaFeEDTA 27.5 27.5 27.5 27.5 27.5
MnSO4·4H2O 11.2 11.2 11.2 11.2 11.2
H3BO3 3.1 3.1 3.1 3.1 3.1
ZnSO4·7H2O 7.2 7.2 7.2 7.2 7.2
Na2MoO4·2H2O 0.12 0.12 0.12 0.12 0.12
CuSO4·5H2O 0.025 0.025 0.025 0.025 1.25
CoCl2·6H2O 0.024 0.024 0.024 0.024 0.024
KI 0.17 0.17 0.17 0.17 0.17
Retinol 0.01 0.01 0.01 0.01 –
Thiamine-HCl 1.0 1.0 1.0 1.0 10.0
Nicotinic acid 1.0 1.0 1.0 1.0 1.0
Riboflavin 0.2 0.2 0.2 0.2 –
Ca-pantothenate 1.0 1.0 1.0 1.0 –
Folic acid 0.4 0.4 0.4 0.4 –
Pyridoxine-HCl 1.0 1.0 1.0 1.0 1.0
Cobalamine 0.02 0.02 0.02 0.02 –
Ascorbic acid 2.0 2.0 2.0 2.0 –
Calciferol 0.01 0.01 0.01 0.01 –
Biotin 0.01 0.01 0.01 0.01 –
Cholin chloride 1.0 1.0 1.0 1.0 –
p-Aminobenzoic acid 0.02 0.02 0.02 0.02 –
Myo-inositol 100 100 100 100 –
Glutamine 438 – 146 – –
Maltose-H2O 90000 90000 90000 90000 36000
BAP 0.9 0.45 0.45 0.225 0.225
2,4-D – – 2.21 – –
Acetosyringone – 98.1 98.1 – –
Cefotaxime – – 125 125 125
Amoxicillin – – 100 100 100
Bialaphos* – – 24.8 33 –
Hygromycin* – – 26.4 52.8 –
MES – 1952 1952 1952 –
Phytagel – – – – 4.000
pH 5.8 5.9 5.0 5.5 5.8
BAP, 6-benzylaminnopurine; 2,4-D, 2,4-dichlorophenoxyacetic acid; EDTA. ethylenediaminetetraacetate;
MES, 2-morpholinoethanesulphonic acid.
*Bialaphos or hygromycin was used following transformation with the BAR or HPT gene, respectively.
Table 3 Media used for Agrobacterium-mediated transformation of barley pollen cultures and subsequent regeneration of transgenic plants

258 Jochen Kumlehn et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2006), 4, 251–261
techniques, opens up the opportunity for directed and exclusive
formation of doubled haploid T1 seeds instantly homozygous
for the transgene. This could be obtained by flow-cytometric
selection of haploid primary transgenic plants at an early
stage of development, followed by treatment with colchi-
cine. As haploid plants are sterile, seeds will be produced only
from doubled haploid spikes or flowers, resulting in exclusive
formation of homozyous transgenic offspring. However, this
approach requires experimental testing.
Because androgenesis and Agrobacterium-mediated trans-
formation are feasible for many angiosperms, the method
described here is anticipated to have potential for other plant
species.
The method established here has been shown to be repro-
ducible in both laboratories that contributed to this study. Based
on the protocol presented here, several hundred transgenic
barley plants have been produced within the framework of
current studies on promoter characterization, over-expression
and knock-down of genes of interest. Moreover, the method was
successfully employed in a mutant complementation approach
using the barley EIF4E gene, which was convincingly proven
to be associated with Bymovirus resistance (Stein et al., 2005).
Experimental procedures
Plant material
Barley (Hordeum vulgare L.) plants cv. ‘Igri’ (Saatzucht Acker-
mann, Irlbach, Germany) were germinated in a growth
chamber (14/12 °C day/night, 16 h light cycle), followed by
8 weeks vernalization treatment (2 °C, 9 h light cycle) and
cultivation in a controlled glasshouse (18/14 °C day/night, 16 h
light cycle). Artificial illumination was provided by SON-T-
Agro lamps (Philips, Hamburg, Germany) at about 200 W/m2.
Bacterial strains and vectors
The A. tumefaciens octopine strain LBA4404 and the nopaline
strain GV3101, carrying the disarmed Ti plasmids pAL4404 and
pMP90RK, respectively, were used (Hellens et al., 2000). The
LBA4404 strain employed in this study additionally contains
the acceptor vector pSB1 (kindly provided by Japan Tobacco
Inc., Higashibara, Japan). This plasmid harbours homologous
recombination sites for the introduction of a T-DNA region of
choice (Komari et al., 1996) and confers hypervirulence as a
result of the presence of accessory alleles of VirB, VirC and VirG.
The Escherichia coli β-glucuronidase gene (GUS), containing
the first intron of the Solanum tuberosum LS1 gene (Vancanneyt
et al., 1990), was introduced as a BamHI/SacI fragment
between the maize ubiquitin-1 promoter with first intron
(UBIp) and the A. tumefaciens nopaline synthase terminator
(NOSt) into the vector pUBI.cas (D. Becker, unpublished). The
resultant UBIp::GUSi::NOSt expression cassette obtained by
HindIII/EcoRI digestion, as well as an EcoRI fragment from the
vector p35SPAT (kindly provided by P. Eckes, Bayer Crop
Science, Frankfurt, Germany) harbouring the Streptomyces
viridochromogenes phosphinothricin acetyltransferase gene
(PAT) flanked by the promoter and the terminator of the cau-
liflower mosaic virus (CaMV) 35S gene, were introduced into
the T-DNA region of the shuttle vector pSB11 (Komari et al.,
1996; kindly provided by Japan Tobacco Inc.). The tail-to-tail
orientation of the expression cassettes within the generated
vector pUG35PAT was confirmed by mini-preparations and
restriction digests. Unlike pUC, pSB11 retains mob function
of pBR322 (Sambrook et al., 1989), and can be mobilized
by conjugal helper plasmids, such as pRK2013. The pSB11-
derived vector pUG35PAT was introduced via triparental
mating to LBA4404(pSB1). LBA4404 strains with co-integration
of pUG35PAT and pSB1 via homologous recombination were
selected on LB medium (Sambrook et al., 1989) containing
30 µg/mL spectinomycin and 10 µg/mL tetracycline.
The T-DNA of pYF133 includes a synthetic GFP gene driven
by the maize ubiquitin-1 promoter with first intron and an HPT
selectable marker gene under the control of an enhanced CaMV
35S promoter (Fang et al., 2002). The T-DNA of pUGAB7 (DNA
Cloning Service, Hamburg, Germany) contains the GUSi gene
driven by the maize ubiquitin-1 promoter with first intron and
the Streptomyces hygroscopicus BAR gene downstream of
the CaMV 35S promoter.
Aliquots (1 µg) of plasmid DNA from the binary vectors
pUGAB7 and pYF133 were added to 100 µL of frozen compe-
tent cells of LBA4404(pSB1) and GV3101, respectively. The cells
were thawed by incubation for 5 min at 37 °C and then placed
on ice for 30 min. Subsequently, the cells were spread on LB
plates containing the appropriate antibiotics and incubated for
2 days at 28 °C. The presence of a binary vector in the colonies
obtained was confirmed by isolation of plasmid DNA and
restriction analysis. Agrobacteria were maintained on stock
plates containing Agrobacterium broth (AB) medium (Chilton
et al., 1974) supplemented with appropriate antibiotics. The
plates were incubated for 2 days at 28 °C and then stored for
up to 4 weeks at 4 °C.
Prior to infection of androgenetic pollen, a loopful of agro-
bacteria was cultured overnight in 8 mL of liquid AB medium
supplemented with appropriate antibiotics. Grown bacteria were
spun down at 3220 g for 10 min, the pellet was resuspended
in 5 mL of AB6 medium (liquid AB medium containing 50 mM
potassium hydrogen phosphate and adjusted to pH 6) and

Transformation of barley pollen cultures 259
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2006), 4, 251–261
incubated in a 9-cm Petri dish at 21 °C with shaking at
50 r.p.m. for 1–5 h.
Microspore isolation and culture
Spikes were harvested when the tips of the awns had just
emerged from the sheath of the flag leaf. At this develop-
mental stage, the central two-thirds of the florets mainly con-
tain highly vacuolated microspores immediately before the
first pollen mitosis. After removal of the leaf sheaths under
sterile conditions, the awns were removed with forceps. The
spikes were then incubated on a wet filter paper disc in 9-cm
sealed Petri dishes at 4 °C for 3–5 weeks to stimulate andro-
genetic development. To isolate immature pollen, 10 spikes
were chopped down to about 1-cm segments and trans-
ferred to a Waring blender (Eberbach, Ann Arbor, MI, USA)
pre-cooled in a refrigerator. Tissue homogenization was
carried out with 18 mL of ice-cold 0.4 M mannitol twice for
5 s at ‘low’ speed. The homogenate was poured through
a 100-µm nylon mesh (Wilson, Nottingham, UK) into a plastic
container on ice. The debris retained on the mesh was gently
squashed with forceps, rinsed with 10 mL of mannitol solu-
tion and transferred back into the blender. Homogenization
was repeated using 12 mL of 0.4 M mannitol solution and the
debris was washed again with 10 mL. The pollen suspension
collected in the container was transferred into a 50-mL tube
and centrifuged at 100 g for 10 min at 4 °C. After removal of
the supernatant, the pellet was resuspended in 8 mL of ice-
cold 0.55 M maltose. This volume was distributed into two
15-mL tubes and each aliquot was cautiously overlain with
1.5 mL of 0.4 M mannitol. After density gradient centrifuga-
tion at 4 °C and 100 g in a swing-out rotor for 10 min, the
nearly pure population of highly vacuolated microspores was
drawn up from the interphase and suspended in a final vol-
ume of 20 mL filled up with 0.4 M mannitol in a 50-mL tube.
Prior to final pelleting of the immature pollen at 100 g for
10 min, two 15-µL samples were taken to assess the total
number of pollen using a haemocytometer (Paul Marienfeld,
Lauda-Königshofen, Germany). The pellet was diluted in an
appropriate volume of KBP medium (Table 3) to obtain a final
population density of 2.5 × 105 pollen per millilitre. One-
millilitre aliquots were cultured in 35-mm sealed Petri dishes
at 26 °C in the dark until further processing.
Agrobacterium-mediated transformation of pollen
cultures
Six to eleven days after pollen isolation, KBP medium was
removed from the pollen cultures using a disposable pipette
and replaced by 1 mL of CK medium (Table 3). The first
experiments on the optimization of the co-culture conditions
were carried out using a preliminary composition of CK
medium including 0.3 mM acetosyringone, pH 5.8 and 3 mM
glutamine. According to the experimental results, a step-by-
step optimization of the co-culture medium was conducted
to eventually yield the composition of CK medium as given
in Table 3. The population density of the Agrobacterium
pre-culture was determined using a spectrophotometer,
and an appropriate volume was added to a 1-mL culture of
androgenetic pollen to obtain a bacterial density of 108 cfu/
mL, if not stated otherwise. The dishes were incubated at
21 °C with shaking at 65 r.p.m. After 48 h of co-culture, the
medium was removed by means of a disposable pipette,
the pollen was washed in 0.4 mL of ASt medium and sub-
sequently cultured in 1.1 mL of ASt medium supplemented
with the appropriate selective agents (Table 3). After an incu-
bation period of 7 days at 26 °C with shaking at 65 r.p.m.,
ASt medium was replaced either by staining buffer according
to Jefferson (1987) for the assessment of GUS expression, or
by 1.5 mL of S medium (Table 3) for further development.
Sealed culture dishes were kept at 26 °C with further shaking
at 65 r.p.m. S medium was renewed every 7 days. Four
weeks after co-culture, the grown calluses were plated on to
K4NBCAx medium (Table 3) for regeneration. After 1 week
of incubation at 26 °C in the dark, the plates were transferred
into the light. After 4 weeks on K4NBCAx, the developing
structures were transferred to plastic containers with the
same medium and cultured for another 4 weeks. If necessary,
structures were cultured for an additional 4 weeks on fresh
medium prior to the plantlets being potted in soil.
The experiments on the optimization of the co-culture con-
ditions were based on six dishes per treatment. For each A.
tumefaciens strain–binary vector combination used for stable
transformation, at least four independent experiments, each
comprising several 1-mL co-cultures, were performed. GUS
activity was assayed histochemically as described by Jefferson
(1987). GFP expression in callus, root tips and pollen was
visualized by an Axiovert 200M inverted microscope equipped
with epifluorescence (Carl Zeiss, Oberkochen, Germany).
Regenerated plants putatively carrying the selectable marker
genes PAT or BAR were sprayed twice with a 150 mg/L solution
of the herbicide BASTA (AgrEvo, Düsseldorf, Germany) with
an interval of about 7 days.
DNA isolation and Southern blotting analysis
Total genomic DNA was isolated from young leaves as
described by Palotta et al. (2000). Genomic DNA (15–20 µg)

260 Jochen Kumlehn et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2006), 4, 251–261
was digested at 37 °C for 5–6 h with enzymes that had unique
restriction sites in the corresponding T-DNA region. Genomic
DNA of non-transformed cv. ‘Igri’ plants digested with the
same enzymes served as negative control, and 80 pg of the
corresponding restricted plasmid DNA served as positive con-
trol. The digested DNA was size-fractionated by gel electro-
phoresis using 0.8% (w/v) agarose and capillary blotted as
essentially described by Sambrook et al. (1989). For radioactive
blots, DNA was transferred on to Hybond NX nylon membranes
(Amersham, Braunschweig, Germany) which were then
hybridized to (32P)-dCTP-labelled probes synthesized from
polymerase chain reaction (PCR) fragments using a Prime-It II
Random Primer Labelling Kit (Stratagene, Heidelberg, Germany).
Non-incorporated nucleotides were removed with HR S300
microspin columns (Amersham Pharmacia Biotech, Freiburg,
Germany). A GFP probe (0.54 kb) was PCR amplified using the
forward primer 5′-GCGACGTAAACGGCCACAAGTTCA-3′and the reverse primer 5′-TAGTGGTTGTCGGGCAGCAGC-3′.A GUS-intron probe (0.63 kb) was amplified with the primer
pair 5′-GCCAGCGTATCGTGCTGCGTTTCGATGCGG-3′ and
5′-CCAGTTGCAACCACCTGTTGATCCGC-3′. Hybridization was
carried out in dextransulphate buffer as described by Sauter
(1997). Membranes were pre-hybridized at 65 °C for 2–3 h,
radioactively labelled probe [(1.0–1.5) × 106 counts per minute
and millilitre of hybridization buffer] was added and hybrid-
ization was continued overnight. Membranes were washed
successively at 65 °C for 10 min in SSC (0.15 M sodium chloride,
0.015 M sodium citrate), and twice for 5 min in 0.5 × SSC
with 0.1% sodium dodecylsulphate (SDS, w/v). Hybridized filters
were exposed to Hyperfilm MP (Amersham Pharmacia Biotech)
at −80 °C for 1–21 days on an eightfold Cronex intensifier
screen (DuPont, Wilmington, DE, USA). For non-radioactive
blots, the digested DNA was transferred on to a Hybond N+
membrane under alkaline conditions as described by the
manufacturer (Amersham). The membrane was hybridized
with an HPT probe labelled by DIG-dUTP using a PCR DIG
(Digoxigenin) Probe Synthesis Kit (Roche Diagnostics,
Mannheim, Germany), the primers 5′-GATCGGACGATT-
GCGTCGCA-3′ and 5′-TATCGGCACTTTGCATCGGC-3′ as
well as plasmid pYF133 (Fang et al., 2002) as template DNA.
Hybridization and DIG detection using CDP-Star were per-
formed as described in the DIG Application Guide for Filter
Hybridization (Roche Diagnostics). The signals were visualized
using Chemiluminescent Detection Film (Roche Diagnostics).
Acknowledgements
We thank P. Eckes (Bayer Crop Science, Frankfurt, Germany)
for providing the p35SPAT plasmid, G. Vancanneyt (Bayer
Crop Science, Gent, Belgium) for the vector p35SGUSINT
and Japan Tobacco Inc. (Higashibara, Japan) for providing
the plasmids pSB1 and pSB11. We are grateful to A. Bam-
mann, B. Betty, M. Gohra, M. Nissen, C. Marthe and I. Otto
for excellent technical assistance. J.K. and L.S. were sup-
ported by the German Bundesministerium für Bildung und
Forschung (grant 0312281A), which we gratefully acknowl-
edge. We are indebted to T. Sharbel for critical reading of
the manuscript.
References
Ashby, A.M., Watson, M.D. and Shaw, C.H. (1987) A Ti-plasmiddetermined function is responsible for chemotaxis of Agrobacte-rium tumefaciens towards the plant wound product acetosyrin-gone. FEMS Microbiol. Lett. 41, 189–192.
Barakat, A., Gallois, P., Raynal, M., Mestre-Ortega, D., Salland, C.,Guiderdoni, E., Delseny, M. and Bernardi, G. (2000) The distribu-tion of T-DNA in the genomes of transgenic Arabidopsis and rice.FEBS Lett. 471, 161–164.
Chilton, M.D., Currier, T.C., Farrand, S.K., Bendich, A.J., Gordon,M.P. and Nester, E.W. (1974) Agrobacterium tumefaciens DNAand PS8 bacteriophage DNA not detected in crown gall tumors.Proc. Natl. Acad. Sci. USA, 71, 3672–3676.
Close, T.J., Wanamaker, S.I., Caldo, R.A., Turner, S.M., Ashlock,D.A., Dickerson, J.A., Wing, R.A., Muehlbauer, G.J., Kleinhofs, A.and Wise, R.P. (2004) A new resource for cereal genomics: 22KBarley GeneChip comes of age. Plant Physiol. 134, 960–968.
Dormann, M., Wang, H.M. and Oelck, M. (2001) Transformedembryogenic microspores for the generation of fertilehomozygous plants. US Patent, 6,316,694 B1.
Fang, Y.-D., Akula, C. and Altpeter, F. (2002) Agrobacterium-mediatedbarley (Hordeum vulgare L.) transformation using green fluorescentprotein as a visual marker and sequence analysis of the T-DNA:barley genomic DNA junctions. J. Plant Physiol. 159, 1131–1138.
Funatsuki, H., Kuroda, H., Kihara, M., Lazzeri, P.A., Mueller, E.,Loerz, H. and Kishinami, I. (1995) Fertile transgenic barley gener-ated by direct DNA transfer to protoplasts. Theor. Appl. Genet.91, 707–712.
Godwin, I., Todd, G., Ford-Lloyd, B. and Newbury, H.J. (1991) Theeffects of acetosyringone and pH on Agrobacterium-mediatedtransformation vary according to plant species. Plant Cell Rep. 9,671–675.
González-Melendi, P., Ramírez, C., Testillano, P.S., Kumlehn, J. andRisueno, M.C. (2005) Three dimensional confocal and electronmicroscopy imaging define the dynamics and mechanisms ofdiploidisation at early stages of barley microspore-derived embry-ogenesis. Planta, 222, 47–55.
Hamilton, C.M. (1997) A binary-BAC system for plant transforma-tion with high-molecular-weight DNA. Gene, 200, 107–116.
Hellens, R., Mullineaux, P. and Klee, H. (2000) A guide to Agrobac-terium binary Ti vectors. Trends Plant Sci. 5, 446–451.
Hensel, G. and Kumlehn, J. (2004) Genetic transformation of barley(Hordeum vulgare L.) by co-culture of immature embryos withAgrobacteria. In Transgenic Crops of the World – Essential Proto-cols (Curtis, I.S., ed.), pp. 35–45. Dordrecht: Kluwer AcademicPublishers.

Transformation of barley pollen cultures 261
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2006), 4, 251–261
Holm, P.B., Olsen, O., Schnorf, M., Brinch-Pedersen, H. and Knud-sen, S. (2000) Transformation of barley by microinjection into iso-lated zygote protoplasts. Transgenic Res. 9, 21–32.
Horvath, H., Huang, J.T., Wong, O., Kohl, E., Okita, T., Kannangara, C.G.and von Wettstein, D. (2000) The production of recombinantproteins in transgenic barley grains. Proc. Natl. Acad. Sci. USA, 97,1914–1919.
Jaehne, A., Becker, D., Brettschneider, R. and Loerz, H. (1994)Regeneration of transgenic, microspore-derived, fertile barley.Theor. Appl. Genet. 89, 525–533.
Jefferson, R.A. (1987) Assaying chimeric genes in plants by the GUSfusion system. Plant Mol. Biol. Rep. 5, 387–405.
Komari, T., Hiei, Y., Saito, Y., Murai, N. and Kumashiro, T. (1996)Vectors carrying two separate T-DNAs for co-transformation ofhigher plants mediated by Agrobacterium tumefaciens and seg-regation of transformants free from selection markers. Plant J. 10,165–174.
Li, L.P., Jia, Y.H., Hou, Q.M., Charles, T.C., Nester, E.W. and Pan, S.Q.(2002) A global pH sensor: Agrobacterium sensor protein ChvGregulates acid-inducible genes on its two chromosomes and Tiplasmid. Proc. Natl. Acad. Sci. USA, 99, 12 369–12 374.
Matthews, P.R., Wang, M.B., Waterhouse, P.M., Thornton, S., Fieg, S.J.,Gubler, F. and Jacobsen, J.V. (2001) Marker gene eliminationfrom transgenic barley, using co-transformation with adjacent‘twin T-DNAs’ on a standard Agrobacterium transformation vector.Mol. Breed. 7, 195–202.
Montoro, P., Teinseree, N., Rattana, W., Kongsawadworakul, P. andMichaux-Ferriere, N. (2000) Effect of exogenous calcium onAgrobacterium tumefaciens-mediated gene transfer in Heveabrasiliensis (rubber tree) friable calli. Plant Cell Rep. 19, 851–855.
Murray, F., Brettell, R., Matthews, P., Bishop, D. and Jacobsen, J.(2004) Comparison of Agrobacterium-mediated transformationof four barley cultivars using the GFP and GUS reporter genes.Plant Cell Rep. 22, 397–402.
Olsen, F.L. (1987) Induction of microspore embryogenesis incultured anthers of Hordeum vulgare. The effects of ammoniumnitrate, glutamine and asparagine as nitrogen sources. CarlsbergRes. Commun. 52, 393–404.
Palotta, M.A., Graham, R.D., Langridge, P., Sparrow, D.H.B. andBarker, S.J. (2000) RFLP mapping of manganese efficiency inbarley. Theor. Appl. Genet. 101, 1100–1108.
Patel, M., Johnson, J.S., Brettell, R.I.S., Jacobsen, J. and Xue, G.P.(2000) Transgenic barley expressing a fungal xylanase gene in theendosperm of the developing grains. Mol. Breed. 6, 113–123.
Pechan, P. (1989) Successful cocultivation of Brassica napus micro-spores and proembryos with Agrobacterium. Plant Cell Rep. 8,387–390.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular Clon-ing: a Laboratory Manual, 2nd edn. Cold Spring Harbor: ColdSpring Harbor Laboratory Press.
Sangwan, R.S., Ducrocq, C. and Sangwan-Norreel, B. (1993) Agro-bacterium-mediated transformation of pollen embryos in Daturainnoxia and Nicotiana tabacum – production of transgenic haploidand fertile homozygous dihaploid plants. Plant Sci. 95, 99–115.
Sauter, M. (1997) Differential expression of a CAK (cdc2-activatingkinase)-like protein kinase, cyclins and cdc2 genes from rice duringthe cell cycle and in response to gibberellin. Plant J. 11, 181–190.
Stachel, S.E., Messens, E., Van Montague, M. and Zambryski, P.
(1985) Identification of the signal molecules produced bywounded plant cells that activate T-DNA transfer in Agrobacte-rium tumefaciens. Nature, 318, 624–629.
Stahl, R., Horvath, H., Van Fleet, J., Voetz, M., von Wettstein, D. andWolf, N. (2002) T-DNA integration into the barley genome fromsingle and double cassette vectors. Proc. Natl. Acad. Sci. USA, 99,2146–2151.
Stein, N., Perovic, D., Kumlehn, J., Pellio, B., Stracke, S., Streng, S.,Ordon, F. and Graner, A. (2005) The eukaryotic translation initia-tion factor 4E confers multiallelic recessive Bymovirus resistance inHordeum vulgare L. Plant J. 42, 912–922.
Sunderland, N., Roberts, M., Evans, L.J. and Wildon, D.C. (1978)Multicellular pollen formation in cultured barley anthers. J. Exp.Bot. 30, 1133–1144.
Tingay, S., McElroy, D., Kalla, R., Feig, S., Wang, M., Thornton, S.and Brettell, R. (1997) Agrobacterium tumefaciens-mediatedbarley transformation. Plant J. 11, 1369–1376.
Tinland, B. (1996) The integration of T-DNA into plant genomes.Trends Plant Sci. 1, 178–184.
Trifonova, A., Madsen, S. and Olesen, A. (2001) Agrobacterium-mediated transgene delivery and integration into barley under arange of in vitro culture conditions. Plant Sci. 162, 871–880.
Turk, S.J., Melchers, L.S., Dendulkras, H., Regensburgtuink, A.J.G.and Hooykaas, P.J.J. (1991) Environmental conditions differentiallyaffect VIR gene induction in different Agrobacterium strains –role of the VirA sensor protein. Plant Mol. Biol. 16, 1051–1059.
Vancanneyt, G., Schmidt, R., O’Connor-Sanchez, A., Willmitzer, L.and Rocha-Sosa, M. (1990) Construction of an intron-containingmarker gene: splicing of the intron in transgenic plants and its usein monitoring early events in Agrobacterium-mediated transfor-mation. Mol. Gen. Genet. 220, 245–250.
Wan, Y. and Lemaux, P.G. (1994) Generation of large numbers ofindependently transformed fertile barley plants. Plant Physiol.104, 37–48.
Wang, M.B., Abbott, D.C., Upadhyaya, N.M., Jacobsen, J.V. andWaterhouse, P.M. (2001) Agrobacterium tumefaciens-mediatedtransformation of an elite Australian barley cultivar with virusresistance and reporter genes. Australian J. Plant Physiol. 28, 149–156.
Winans, S.C. (1991) An Agrobacterium 2-component regulatorysystem for the detection of chemicals released from plantwounds. Mol. Microbiol. 5, 2345–2350.
Wu, H., McCormac, A.C., Elliott, M.C. and Chen, D.-F. (1998)Agrobacterium-mediated stable transformation of cell suspensioncultures of barley (Hordeum vulgare). Plant Cell Tiss. Organ Cult.54, 161–171.
Zhang, S., Cho, M.J., Koprek, T., Yun, R., Bregitzer, P. and Lemaux,P.G. (1999) Genetic transformation of commercial cultivars of oat(Avena sativa L.) and barley (Hordeum vulgare L.) using shoot mer-istematic cultures derived from germinated seedlings. Plant CellRep. 18, 959–966.
Zhang, H., Sreenivasulu, N., Weschke, W., Stein, N., Rudd, S.,Radchuk, V., Potokina, E., Scholz, U., Schweizer, P., Zierold, U.,Langridge, P., Varshney, R.K., Wobus, U. and Graner, A. (2004)Large-scale analysis of the barley tanscriptome based onexpressed sequence tags. Plant J. 40, 276–290.
Zupan, J. and Zambryski, P. (1997) The Agrobacterium DNA transfercomplex. Crit. Rev. Plant Sci. 16, 279–295.










![Association analysis for traits associated with powdery mildew tolerance in barley [Hordeum vulgare L.] using AFLP markers](https://img.dokumen.tips/doc/110x75/634e1eb0940e97c6be037b98/association-analysis-for-traits-associated-with-powdery-mildew-tolerance-in-barley.jpg)


















