Embed Size (px)
Citation preview

Gastropods from the Lower Cenomanian of Koraćica (Kosmaj Mountain, central Serbia)
Wagih Ayoub-Hannaa, Barbara V. Radulović, Vladan J. Radulović, and Franz Theodor Fürsich
With 17 figures and 19 tables
Abstract: Twenty Early Cenomanian gastropod taxa belonging to 14 genera and 10 families are systematically described from reddish coarse-grained bioclastic packstones to rudstones of the Grab well section (Kosmaj Mountain) of the Vardar Zone of central Serbia. The fauna is strongly dominated by Ampullinidae and Pleurotomariidae followed by Tylostomatidae. This paper describes three new species: Anodomaria kosmajae sp. nov., Latiala? callositae sp. nov., and Pseudonerinea jovankae sp. nov. In addition, seven species, Conotomaria folkestonensis Cox 1960b, Astele gurgitis (Brongniart, 1822), Pictavia pungens (J. de C. SowerBy in Fitton, 1836), Pseudamaura subbulbiformis (d’orBigny, 1850), Tylostoma elevatum (Shumard, 1853), Tylostoma cf. minimum white, 1887, and Avellana subincrassata d’orBigny, 1850 are recorded from the Lower Cenomanian of Serbia for the first time. Two species are described in open nomenclature. The identified gastropod species come from a half-a-meter-thick and approximately 2-m-long lens with hundreds of reworked fossils. The faunal elements of this assemblage are mostly disarticulated, fragmented, and lack preferred orientation. Taphonomic and sedimentological data suggest that this lens is the result of a storm event (storm-wave concentration) and that the fossils are parautochthonous. The species lived in a shallow-marine setting above the storm wave-base, in warm-temperate waters within the photic zone.
Key words: Lower Cenomanian, gastropods, taxonomy, palaeoecology, Kosmaj Mountain, central Serbia.
1. Introduction
The Lower Cenomanian succession of the Grab well section (Kosmaj Mountain) of the Vardar Zone of central Serbia yields a rich macroinvertebrate fauna composed of gastropods, ammonites, bivalves, echinoids, corals, and brachiopods. The earliest studies on the Albian-Cenomanian macrofauna of Serbia were carried out by Jovanović (1938). The latter author described two Upper Albian (Vraconian)-Lower Cenomanian ammonite species from thin-bedded sandy limestones at the quarry on the left bank of the Alinac spring. Later, from the same locality, but at the right bank of the spring, andJelković (1953) listed five ammonite
and three gastropod species from the conglomerates of the upper part of the succession, which overlie the sandy limestones. During the 1970’s Jovanka Mitrović-Petrović and Milodrag andJelković from Belgrade University collected macrobenthic fauna and ammonites from the fossiliferous lens of this locality. They listed 142 Upper Albian-Lower Cenomanian species, which were dominated by gastropods (45 species) followed by ammonites (41), bivalves (36), echinoids (13), brachiopods (4), and corals (3) (Mitrović-Petrović & andJelković 1991: 583-584). Their plan to monograph the fauna, however, was never realized. Therefore, the collection was given to the present authors for further processing and revising. Recently, ayoub-Hannaa et
©2015 E. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart, Germany www.schweizerbart.de
DOI: 10.1127/njgpa/2015/0474 0077-7749/2015/0474 $ 8.75
N. Jb. Geol. Paläont. Abh. 276/1 (2015), 27–62 ArticleStuttgart, April 2015E
eschweizerbart_xxx

28 W. Ayoub-Hannaa et al.
al. (2014) provided detailed taxonomic descriptions and revision of brachiopods (two species) and echinoids (eight species, five of them recorded for the first time from Serbia) from the above mentioned collection.
Albian-Cenomanian gastropods from central Serbia are poorly documented in the literature. Mitrović-Petrović & andJelković (1991: 583) listed 45 gastropod taxa, which have neither been described nor figured and whose names are not valid. Therefore, a detailed taxonomic description and revision of these Early Cenomanian gastropods were carried out in the present study. In addition, palaeoecological and taphonomic aspects of the macrofaunal assemblage, with special reference to gastropods, are discussed.
2. Geological and stratigraphic setting
In central Serbia, Albian-Cenomanian rocks crop out along the southern slopes of the Kosmaj Mountain. The best outcrop containing abundant macroinvertebrates is located near the village of Koraćica, at the Grab well, on the right bank of the Alinac spring (Fig. 1; GPS co-ordinates: 44°26’59.7”N; 20°35’56.4”E). The outcrop area is situated within the Main Vardar Zone (roBertSon et al. 2009). The Albian-Cenomanian succession, about
41-m-thick, is bounded by faults at the base and top, by serpentinites in the west and Upper Cretaceous flysch in the east. The lower 20 m of the succession, which is exposed in the quarry on the left bank of the Alinac stream, consists of Upper Albian thin-bedded, rarely medium-bedded, silty marlstones and marly siltstones. These marlstones and siltstones are intercalated with thinly bedded, fine-grained wackestones to packstones (Fig. 2). A latest Albian (Vraconian) age of this part of the succession is confirmed by the presence of the ammonite Puzosia planulata (J. de C. SowerBy) (Jovanović 1938) and the planktonic foraminifer Thalmanninella appenninica (renz) (ayoub-Hannaa et al. 2014). The contact, approximately 10-m-thick, between the uppermost Albian and lowermost Cenomanian rocks is covered by debris, and could not be observed. The Lower Cenomanian rocks are exposed on the right bank of the stream. They attain a thickness of 7.1 m and are composed mainly of limestone breccia-conglomerates with coarse-grained calcarenite matrix. The lower 2.2 m of this succession contain the larger foraminifer Orbitolina (Conicorbitolina) conica (d’arChiaC), which indicates an Early Cenomanian age. Up-section, orbitolinids are lacking, while the coralline alga Archaeolithothamnium amphiroaeforme rothpletz rapidly increases in abundance. The upper
Fig. 1. A. Locality map of the investigated section (black star) in the Kosmaj Mountain, central Serbia.
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 29
5.1 m of the succession are also assigned to the Lower Cenomanian based on the brachiopods Capillithyris capillata (d’arChiaC) and Terebrirostra lyra (J. SowerBy), which characterize the earliest Cenomanian Mantelliceras mantelli ammonite Zone (gaSpard 1997). These two brachiopod species, like the material studied, have been collected from a lens consisting of reddish coarse-grained bioclastic packstones to rudstones, which lie above algal grainstones (0.7 m) and are overlain by sandy grainstones (3 m). The lens attains a thickness of half a meter and is approximately two meters long. For more details about the litho- and biostratigraphy see ayoub-Hannaa et al. (2014).
3. Material and methodsThis paper deals with 62 specimens collected from the Lower Cenomanian rocks of the Kosmaj Mountain, central Serbia. Most of the specimens are preserved as internal and composite moulds. A few of them occur in shell preservation. In fact, internal moulds show so few features that a precise identification was not possible. Therefore, 17 specimens from the present collection are not further discussed due to their poor preservational state and lack of diagnostic features (e.g., outer lip, protoconch, ornamentation). Specimens were prepared mechanically and cleaned carefully. Subsequently, they have been whitened with magnesium oxide dust, which accentuates finer ornamental details. In addition, axial cross-sections of some specimens (e.g., nerineids) have been produced in order to investigate internal structures such as columellar and outer lip folds.Morphological terminology follows the glossary presented by Cox (1960a). The classification used here is that of BouChet & rocroi (2005).
Linear measurements (taken with a Vernier caliper) are in millimetres and their abbreviations are as follows: H – height; D – diameter; HL – height of last whorl; HA – height of aperture; WA – width of aperture; NW – number of whorls; Sa – spire angle (in degrees); HW– height of whorl; DW– diameter of whorl. Orientations of measurements are shown in Fig. 3A. In addition, terms used in the text to describe taxa of the family Pleurotomariidae are shown in Fig. 3B. These terms are: Sz – selenizone; Lu – lunulae; S – suture; Sr – sub-sutural ramp; Ra – ramp angle; St – spiral threads; Ct – collabral threads; Sl – slit; A – aperture; B – base; P – periphery; U – umbilicus.
The material is housed in the Department of Palaeontology, Faculty of Mining and Geology, University of Belgrade, collection RGF JMP.
Fig. 2. Litho- and biostratigraphic section of the Upper Albian–Lower Cenomanian deposits in the Grab well section, Kosmaj Mountain, central Serbia (modified after ayoub-Hannaa et al. 2014). Upper Albian: 1, dark grey marlstones, silty marlstones, marly siltstones with fine-grained sandy wackstones/packstones intercalations, 2, limestone breccia-conglomerates with calcarenite matrix; 3, algal grainstones; 4, coarse-grained bioclastic packstones to rudstones containing the described fauna; 5, weathered yellowish sandy grainstones.
eschweizerbart_xxx

30 W. Ayoub-Hannaa et al.
4. Systematic palaeontology
Class Gastropoda Cuvier, 1797Clade Vetigastropoda Salvini-Plawen, 1980
Superfamily Pleurotomarioidea SwainSon, 1840Family Pleurotomariidae SwainSon, 1840
Genus Anodomaria SzaBó, 1980
Type species: Pleurotomaria scacchi gemmellaro, 1874.
Remarks: The genus Anodomaria SzaBó, 1980 is characterized by having (1) a gradate-trochoform spire, (2) weakly convex whorls, (3) strong spiral threads, crossed by collabral threads and forming a reticulate pattern and producing small tubercles at intersection, (4) a concave
selenizone, which is broad in latter whorls and situated above mid-whorl, (5) a broad and slightly convex phaneromphalous base, and (6) a narrow subsutural ramp. The genus Leptomaria e. eudeS-deSlongcHaMPS, 1864 differs from Anodomaria in having a very narrow selenizone, which runs slightly to distinctly below mid-whorl, a narrow slit, a convex base, strongly convex whorls (not angular), a rounded periphery of last whorl, and a reticulate ornamentation, which is restricted to the early whorls. According to Monari & gatto (2014: 12), spiral threads predominate on the fully adult shell.
The genus Conotomaria Cox, 1959 carries some similarities to Leptomaria but differs in having a flat or slightly sigmoidal whorl flank, gradate outline, a nearly flat base, an angulated periphery of the last whorl, and a selenizone which lies above the mid-whorl height, and which is wider. Pleurotomaria deFranCe, 1826 differs from the above mentioned genera by having a large-sized, discoidal to high trochoform shell, nodes (large tubercles) at the shoulder,
Fig. 3. A – Abbreviations of measured parameters of the gastropods. H: height; D: diameter; HL: height of last whorl; HA: height of aperture; WA: width of aperture; Sa: spire angle (in degree). B – Apertural view of pleurotomariid showing descriptive terms used in text. Sz: selenizone; Lu: lunulae; S: suture; Sr: sub-sutural ramp; Ra: ramp angle; St: spiral threads; Ct: collabral threads; Sl: slit; A: aperture; B: base; P: periphery; U: umbilicus.
Fig. 4. A-L – Anodomaria kosmajae sp. nov. from the Lower Cenomanian, Kosmaj Mountain, central Serbia. A: Apertural view; B: abapertural view; C: close-up showing moderately broad and deep selenizone (Sz), which lies at mid-whorl height; D: close-up showing well developed lunulae (Lu) and details of reticulate ornamentation; E: apical view; F: basal view showing strong spiral threads adaxially; RGF JMP 138 (holotype); G: apertural view; H: abapertural view showing graded whorls and deep suture; I: close-up showing a moderately broad selenizone (Sz) (located at the mid-whorl height); J: basal view showing small umbilicus (U); K: apical view showing a wide sub-sutural ramp; RGF JMP 129 (paratype); L: abapertural view showing deep suture and graded outline of whorls; RGF JMP 119 (paratype). For abbreviations see Fig. 3.
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 31
Fig. 4.
eschweizerbart_xxx

32 W. Ayoub-Hannaa et al.
and its selenizone is located below the ramp angle (Cataldo & lazo 2012: 80). The Serbian specimens differ from some Cretaceous species of Bathrotomaria Cox, 1985 (e.g., Kiel & bandel 2004: 104-106, fig. 3) in having a narrow sub-sutural ramp, a high conical shell, slightly convex whorl sides (angular in Bathrotomaria), and strong reticulate ornamentation.
Based on ornamentation, outline, and position of selenizone, the Serbian specimens strongly resemble Anodomaria. The latter genus is restricted to Early and Middle Jurassic rocks (HaraSewycH & kiel, 2007: 78, fig. 3). According to vöröS et al. (2003: 60), Anodomaria is a typically Mediterranean gastropod genus with only a few species (SzaBó 1988). Recently, some species have been recorded from younger strata (Late Bathonian–Callovian) of western India by Jaitly et al. (2000). In the present work, Anodomaria extends to the Early Cenomanian. For more discussion on some other pleurotomariid genera and subgenera see Jaitly et al. (2000), HaraSewycH & kiel (2007), haraSewyCh et al. (2009), and Monari & gatto (2013, 2014).
Anodomaria kosmajae sp. nov.Figs. 4A-L, 5, 6A-C
Etymology: This species is named after the Kosmaj Mountain, located in central Serbia, 50 km south of Belgrade.
Holotype: Specimen no. RGF JMP 138 (Fig. 4A-F), Lower Cenomanian, Kosmaj Mountain, central Serbia.
Paratypes: Four specimens, RGF JMP 119, 129, 130, 140 (Figs. 4G-L, 5, 6A-C).
Type locality: Grab well, Koraćica village, Kosmaj Mountain, central Serbia.
Type horizon: Lens of Lower Cenomanian bioclastic coarse-grained packstones to rudstones.
Diagnosis: Shell medium-sized; outline conical; spire angle on average ~ 57°; whorl flank slightly convex; base broad and slightly convex; selenizone moderately broad and deep, lying at mid-whorl height and covered by strong crescentic linear ridges (lunulae); a canaliculate suture, well-developed
collabral threads prosocline-prosocyrt between selenizone and adapical suture, and opisthocline-straight between selenizone and abapical suture; aperture ovate (elongated obliquely to coiling axis); ornamentation strong cancellate with tubercles at the intersection between collabral and spiral threads.
Description: Medium-sized, slightly higher than wide (D/H = 0.93 on average; Table 1), outline conical, spire moderately high and gradate. Spire consisting of 5-7 whorls and accounting about 59% of total shell height. Spire angle ~ 57°. Whorl flank of spire slightly convex, separated by deep suture (canaliculate). Convexity of whorls gradually decreasing adapically. Base slightly convex with narrow umbilicus. Sub-sutural ramp narrow to moderately wide, slightly concave in the early teleoconch whorls, wide and slightly convex in last whorl, and covered by small and prosocline axial nodes. Last whorl wide, with rounded-angular periphery. Selenizone moderately broad (1.4 mm on the last whorl; selenizone width/suture to suture distance ~0.12) (Fig. 4C, D, I), moderately deep, lying at mid-whorl height, and covered by well developed lunulae, which are concave towards aperture (Fig. 4D). Aperture irregularly ovate and elongated obliquely to coiling axis (Figs. 5, 6C). Each whorl covered with well developed spiral cords, 4-10 between selenizone and abapical suture and 3-8 between selenizone and adapical suture (Table 1). These spiral cords are more prominent and closely spaced near suture, finer and widely spaced close to the selenizone, and are crossed by collabral threads that produce a cancellate pattern with tuberculate appearance (Fig. 4H). Collabral threads prosocline-prosocyrt between selenizone and adapical suture, and opisthocline-prosocyrt between selenizone and periphery. Base covered with 18-25 spiral threads, which are well developed adaxially and separated by relatively deep and narrow interspaces (Fig. 4F, J). Axial threads extending slightly below the periphery of last whorl and disappearing close to umbilicus.
Discussion: Anodomaria kosmajae sp. nov. resembles Leptomaria loricatula (StoliCzKa, 1868: 385, pl. 25, fig. 4) from the Aptian-Turonian Uttatur Group of India in having a conical shell, a convex whorl flank, and a rounded periphery of the last whorl but the latter differs in having faint collabral threads between the selenizone and periphery, a narrow selenizone which lies above mid-whorl height, a quadrangular and large aperture, and in being higher than the
Table 1. Dimensions (in mm) of Anodomaria kosmajae sp. nov.
Specimen H D HL HA WA SA° nw D/H HL/HN cords N cords of
baseSz→P Sz→SRGF JMP 119 28 25 12 -- -- 57º 6 0.89 0.42 5-8 3-8 ?20RGF JMP 129 26 27 10 7 -- 55º 7 1.04 0.38 4-6 4-8 18RGF JMP 130 27 25 11 10 16 55 5 0.92 0.41 6-10 3-6 ?20RGF JMP 138 30 26 13 11 17 61 7 0.87 0.43 5-7 4-7 25
Range 27-30 25-27 11-13 7-11 16-17 55-61 5-7 0.87-1.04 0.38-0.43 4-10 3-8 ?18-25Mean 27.75 25.75 11.5 9.33 16.5 57 6 0.93 0.41 4-10 3-8 ?21
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 33
present species. Conotomaria falcata (d’orBigny, 1843: 263, pl. 200, figs. 9-12) from the Cenomanian of France differs in having a narrower and deeper selenizone, non-tuberculated axial ribs, a moderately deep suture, and a more acute shell (D/H = 0.82 as opposed to 0.93 on average). Kollmann (2005: 112) considered C. matheroniana (d’orBigny, 1843: 264, pl. 201, figs. 1-4) from the Cenomanian of France as a junior synonym of C. falcata. Despite the strong similarities between the latter two species in general outline and ornamentation, C. matheroniana differs in each whorl having a bulging periphery, in an angulate periphery of the last whorl, numerous faint collabral threads, and a selenizone that lies at mid-whorl height. Leptomaria indica (ForBeS, 1846: 121, pl. 13, fig. 13) from the Upper Cretaceous of India differs from A. kosmajae sp. nov. in being wider (D/H = 1.19 as opposed to 0.93 on average), larger (H = 38, D = 44 mm as opposite to H = 28, D = 26 mm on average), and in having an angulate periphery of last whorl, an angulate whorl flank, a narrow selenizone, and a broad and flat base. L. gibsi (J. SowerBy, 1821) as figured and described by Cox (1960b: 397, pl. 49, figs. 1-3) from the lower Middle Albian of England resembles the present species in having a cancellate ornamentation but differs in having a low conical shell, a flush selenizone, numerous fine collabral threads, an angulate periphery of the last whorl, and in being smaller (H = 17 mm as opposed to 27.75 mm on average). Similarly, L. brongniartiana (d’orBigny, 1843) as figured by Kollmann (2005: 114, pl. 12, fig. 9) from the Cenomanian of France differs in having a low conical shell, a wide umbilicus, a strongly convex whorl flank, a shallow
and smooth selenizone, and widely spaced collabral threads, which cover the areas above and below the selenizone. A smooth base, narrow and flush selenizone, and widely spaced collabral threads (opisthocline below the selenizone) distinguish L. simplex (d’orBigny, 1843: 252, pl. 194, figs. 1-3) from the Cenomanian of France from A. kosmajae sp. nov. L. retectus (white, 1887: 199, pl. 10, fig. 8) from the Albian of the Sergipe Basin, Brazil, differs in having fewer spiral cords, wide interspaces, and a small and sub-ovate aperture smaller than the present species. C. percevali Cox, 1960 as figured and described by cleevely & MorriS (2002: 169, pl. 28, fig. 6) and kiel & bandel (2004: 106, fig. 3L-N) from the Cenomanian of England and Germany, respectively differs in being larger and in having a sub-angular periphery of the last whorls, broad flat spiral cords, a wide umbilicus, and a selenizone that lies in the upper half of whorls. C. swinnertoni (Cox, 1960b: 401, pl. 45, figs. 2, 3) from the Lower Cretaceous of England differs in being smaller (H = 10, D = 11.2 mm as opposed to H = 28, D = 26 mm on average) and in having a raised selenizone, no umbilicus, angulate whorls, and irregularly and widely spaced spiral cords. C. elegans (d’orBigny, 1843: 242, pl. 90, figs. 1-4) from the Lower Cretaceous of France resembles the present species in having a moderately wide selenizone, located adapically of mid-whorl height, a cancellate ornamentation, and slightly convex whorl flank. It differs, however, in having a wide and deep umbilicus, a convex base, a moderately deep suture with no shoulder below, and less developed tubercles.
Genus Conotomaria Cox, 1959
Type species: Pleurotomaria mailleana d’orBigny, 1843, Cenomanian of France.
Conotomaria folkestonensis Cox, 1960bFig. 6D-I
1960b Conotomaria folkestonensis sp. nov. – Cox, p. 410, pl. 49, fig. 4.
2002 Leptomaria gibsi (J. SowerBy). – Kollmann, p. 27, pl. 5, figs. 70-71, non 72-75.
Material: One specimen from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 1).
Description: Shell small, outline low-conical, diameter exceeding height (D/H = 1.31; Table 2); moderately high-spired. Spire consisting of four well preserved teleoconch whorls, separated by a moderately deep suture. Protoconch and early teleoconch whorls not preserved. Whorl sides more inclined and slightly convex abapically and strongly convex adapically. Sub-sutural ramp narrow, convex, and moderately inclined. Selenizone narrow, moderately deep, positioned abapically of mid-whorl, and delimited by strong ribs (Fig. 6F). Base broad, moderately convex with small and rounded umbilicus (Fig. 6G). Periphery of last whorl acute and bulging. Outer lip not complete. The area between selenizone and adapical suture (upper half of whorl) covered
Fig. 5. Axial sketch of Anodomaria kosmajae sp. nov. from the Lower Cenomanian of Kosmaj Mountain showing elongated whorls oblique to the coiling axis; RGF JMP 140 (paratype).
eschweizerbart_xxx

34 W. Ayoub-Hannaa et al.
Fig. 6.
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 35
with well-developed prosocline collabral threads (Fig. 6E), while area between selenizone and abapical suture covered by tuberculated spiral threads (Fig. 6F). Base covered with numerous faint spiral threads, separated by relatively deep and narrow interspaces.
Discussion: Conotomaria folkestonensis Cox, 1960b can be easily recognized by a low-conical shell, an angulated periphery of last whorl, broad and slightly convex base, a narrow selenizone (located abapically of mid-whorl), and well developed prosocline collabral threads in the area between selenizone and adapical suture, and tuberculated spiral threads in the area between selenizone and abapical suture (Fig. 6E, F).
Leptomaria gibsi (J. SowerBy, 1821: 139, pl. 287, fig. 1) from the middle Albian of England and L. gurgitis (d’orBigny, 1843: 249, pl. 192, figs. 4-6; non Brongniart 1822: pl. 9, fig. 7) from the Albian of France resemble the present species in having a low-conical shell, a narrow selenizone (located abapically of mid-whorl), and a wide last whorl with an acute and bulging periphery but differs in having angulated whorls, a very convex base, a narrow sub-sutural ramp, and reticulate ornamentation which covers the entire whorls. Kollmann (2002, 2005) assigned SowerBy’s species gibsi to the genus Leptomaria. He placed this species in Leptomaria because of the position of the selenizone abapically of the carination of the whorl. However, apart from features characteristic of Leptomaria, the species has also a very prominent periphery and low base which are unusual features of that genus and are more consistent with characters of Conotomaria. Therefore, some specimens of L. gibsi as figured by Kollmann (2002: 27, pl. 5, figs. 71, 72) from the Albian of Austria are considered as synonyms of C. folkestonensis.
C. cassisiana (d’orBigny, 1843: 266, pl. 202, figs. 1-4) from the Cenomanian of France differs in having flat whorl sides, reticulated ornamentation above and below the selenizone zone and in being much wider (D/H = 1.60 as opposed to 1.31) and larger (D = 55 mm as opposed to 25 mm). The present species resembles also L. royana (d’orBigny, 1843: 269, pl. 203, figs. 5, 6) from the Campanian of France in having a low conical shell but the latter differs in having a strongly convex whorl flank, a wide selenizone, a strongly convex base, a wide umbilicus, and fewer, widely spaced, collabral threads.
Occurrence: Albian of England (Cox 1960b), Austria (Kollmann 2002), Lower Cenomanian of the Kosmaj Mountain, central Serbia (this study).
Conotomaria cf. pricei (Cox, 1960b)Fig. 6J-M
cf. 1960b Leptomaria pricei n. sp. – Cox, p. 398, pl. 49, fig. 8.
cf. 1991 Leptomaria pricei Cox. – Mitrović-Petrović & andJelković, p. 583.
Material: One distorted specimen from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 105).
Description: Shell medium in size (Table 3), gradate-conical, wider than high (D/H = 1.67), moderately high and broadly trochiform. Protoconch and first few teleoconch whorls not preserved. Spire consisting of four teleoconch whorls which
Table 2. Dimensions (in mm) of Conotomaria folkestonensis Cox, 1960b.
Specimen H D HL HA WA SA° nw D/H HL/HN cords N cords of
baseSz→P Sz→SRGF JMP 1 19 25 10 8 -- 82 4 1.31 0.52 ?14 8-11 ?20
Table 3. Dimensions (in mm) of Conotomaria cf. pricei (Cox, 1960b).
Specimen H D HL HA WA SA° nw D/H HL/HN cords N cords of
baseSz→P Sz→SRGF JMP 105 24 40 12 ?9 -- 85 4 1.67 0.50 12-22 6-11 24
Fig. 6. A-C – Anodomaria kosmajae sp. nov. from the Lower Cenomanian, Kosmaj Mountain, central Serbia. A: Apertural view showing the ovate aperture; B: abapertural view showing moderately convex whorl flank; RGF JMP 130 (paratype); C: axial section showing oblique, ovate whorls; RGF JMP 140 (paratype). D-I – Conotomaria folkestonensis Cox, 1960b. D: Apertural view; E: close-up showing prosocline collabral cords above the selenizone (Sz); F: close-up showing a narrow selenizone, the deep suture, and spiral threads (St); G: basal view showing small and rounded umbilicus (U); H: apical view; I: abapertural view; RGF JMP 1. J-M – Conotomaria cf. pricei (Cox, 1960) from the same section. J: Abapertural view showing an angular periphery of last whorl; K: close-up showing a narrow selenizone (Sz), which lies above the mid-whorl height and the reticulate ornamentation above and below selenizone; L: apertural view of distorted specimen; M: basal view showing wide and rounded umbilicus; RGF JMP 105. For abbreviations see Fig. 3.
eschweizerbart_xxx
36 W. Ayoub-Hannaa et al.
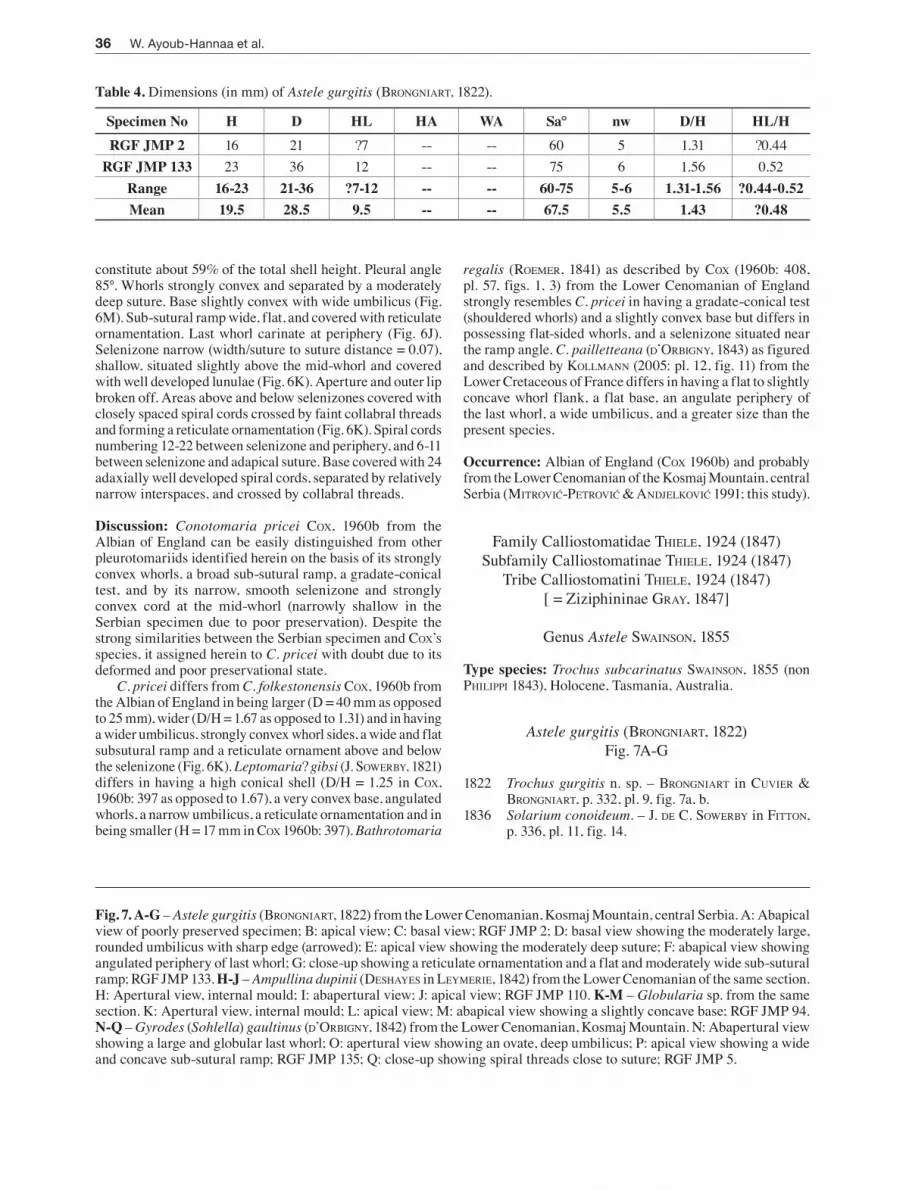
constitute about 59% of the total shell height. Pleural angle 85°. Whorls strongly convex and separated by a moderately deep suture. Base slightly convex with wide umbilicus (Fig. 6M). Sub-sutural ramp wide, flat, and covered with reticulate ornamentation. Last whorl carinate at periphery (Fig. 6J). Selenizone narrow (width/suture to suture distance = 0.07), shallow, situated slightly above the mid-whorl and covered with well developed lunulae (Fig. 6K). Aperture and outer lip broken off. Areas above and below selenizones covered with closely spaced spiral cords crossed by faint collabral threads and forming a reticulate ornamentation (Fig. 6K). Spiral cords numbering 12-22 between selenizone and periphery, and 6-11 between selenizone and adapical suture. Base covered with 24 adaxially well developed spiral cords, separated by relatively narrow interspaces, and crossed by collabral threads.
Discussion: Conotomaria pricei Cox, 1960b from the Albian of England can be easily distinguished from other pleurotomariids identified herein on the basis of its strongly convex whorls, a broad sub-sutural ramp, a gradate-conical test, and by its narrow, smooth selenizone and strongly convex cord at the mid-whorl (narrowly shallow in the Serbian specimen due to poor preservation). Despite the strong similarities between the Serbian specimen and Cox’S species, it assigned herein to C. pricei with doubt due to its deformed and poor preservational state.
C. pricei differs from C. folkestonensis Cox, 1960b from the Albian of England in being larger (D = 40 mm as opposed to 25 mm), wider (D/H = 1.67 as opposed to 1.31) and in having a wider umbilicus, strongly convex whorl sides, a wide and flat subsutural ramp and a reticulate ornament above and below the selenizone (Fig. 6K). Leptomaria? gibsi (J. SowerBy, 1821) differs in having a high conical shell (D/H = 1.25 in Cox, 1960b: 397 as opposed to 1.67), a very convex base, angulated whorls, a narrow umbilicus, a reticulate ornamentation and in being smaller (H = 17 mm in Cox 1960b: 397). Bathrotomaria
regalis (roemer, 1841) as described by Cox (1960b: 408, pl. 57, figs. 1, 3) from the Lower Cenomanian of England strongly resembles C. pricei in having a gradate-conical test (shouldered whorls) and a slightly convex base but differs in possessing flat-sided whorls, and a selenizone situated near the ramp angle. C. pailletteana (d’orBigny, 1843) as figured and described by Kollmann (2005: pl. 12, fig. 11) from the Lower Cretaceous of France differs in having a flat to slightly concave whorl flank, a flat base, an angulate periphery of the last whorl, a wide umbilicus, and a greater size than the present species.
Occurrence: Albian of England (Cox 1960b) and probably from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (Mitrović-Petrović & andJelković 1991; this study).
Family Calliostomatidae thiele, 1924 (1847)Subfamily Calliostomatinae thiele, 1924 (1847)
Tribe Calliostomatini thiele, 1924 (1847)[ = Ziziphininae gray, 1847]
Genus Astele SwainSon, 1855
Type species: Trochus subcarinatus SwainSon, 1855 (non philippi 1843), Holocene, Tasmania, Australia.
Astele gurgitis (Brongniart, 1822)Fig. 7A-G
1822 Trochus gurgitis n. sp. – Brongniart in Cuvier & Brongniart, p. 332, pl. 9, fig. 7a, b.
1836 Solarium conoideum. – J. de C. SowerBy in Fitton, p. 336, pl. 11, fig. 14.
Fig. 7. A-G – Astele gurgitis (Brongniart, 1822) from the Lower Cenomanian, Kosmaj Mountain, central Serbia. A: Abapical view of poorly preserved specimen; B: apical view; C: basal view; RGF JMP 2; D: basal view showing the moderately large, rounded umbilicus with sharp edge (arrowed); E: apical view showing the moderately deep suture; F: abapical view showing angulated periphery of last whorl; G: close-up showing a reticulate ornamentation and a flat and moderately wide sub-sutural ramp; RGF JMP 133. H-J – Ampullina dupinii (deShayeS in leymerie, 1842) from the Lower Cenomanian of the same section. H: Apertural view, internal mould; I: abapertural view; J: apical view; RGF JMP 110. K-M – Globularia sp. from the same section. K: Apertural view, internal mould; L: apical view; M: abapical view showing a slightly concave base; RGF JMP 94. N-Q – Gyrodes (Sohlella) gaultinus (d’orBigny, 1842) from the Lower Cenomanian, Kosmaj Mountain. N: Abapertural view showing a large and globular last whorl; O: apertural view showing an ovate, deep umbilicus; P: apical view showing a wide and concave sub-sutural ramp; RGF JMP 135; Q: close-up showing spiral threads close to suture; RGF JMP 5.
Table 4. Dimensions (in mm) of Astele gurgitis (Brongniart, 1822).
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 2 16 21 ?7 -- -- 60 5 1.31 ?0.44
RGF JMP 133 23 36 12 -- -- 75 6 1.56 0.52Range 16-23 21-36 ?7-12 -- -- 60-75 5-6 1.31-1.56 ?0.44-0.52Mean 19.5 28.5 9.5 -- -- 67.5 5.5 1.43 ?0.48
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 37
Fig. 7.
eschweizerbart_xxx

38 W. Ayoub-Hannaa et al.
1842 Solarium conoideum Fitton. – d’orBigny, p. 198, pl. 179, figs. 12, 14, non fig. 13 (non Solarium conoideum J. SowerBy, 1813).
1847 Solarium conoideum J. de C. SowerBy. – Pictet & roux, p. 223, pl. 21, fig. 10.
?1931 Trochus waterloti. – Collignon, p. 58 (18), pl. 2, figs. 4-5.
2005 Astele gurgitis (Brongniart). – Kollmann, p. 79, pl. 9, figs. 11-13.
Material: Two specimens from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 2, 133).
Description: Shell medium-sized to moderately large (Table 4), trochiform, wide (D/H = 1.43 on average), and moderately high-spired. Spire consisting of five whorls accounting for ~ 52% of the total shell height. Protoconch and first few teleoconch whorls not preserved. Subsequent teleoconch whorls slightly concave below, slightly convex above, separated by a moderately deep suture. Base broad and slightly convex. Umbilicus moderately large (30% of total shell diameter) and round, with sharp edge (Fig. 7D). Last whorl carinate at periphery with rounded crest (Fig. 7F). Aperture and outer lip broken off. Ornamentation consisting of numerous, closely spaced, fine spiral cords crossed by collabral threads, and forming a reticulate pattern (Fig. 7G). Intersection between collabral and spiral threads with fine nodes, which are well developed close to the suture.
Discussion: Kollmann (2005: 79) assigned Brongniart’s species to the genus Astele on the basis of (1) a trochiform shell, (2) a wide and rounded umbilicus with sharp edge, and (3) a moderately oblique labrum. Astele gurgitis (Brongniart) can be easily distinguished by having a low trochiform shell, concavo-convex whorls, an angulate periphery of the last whorl, a slightly convex base, a large and rounded umbilicus with sharp edge, and reticulated ornamentation.
With respect to general outline, size and shape of umbilicus, and a deep suture, Trochus waterloti Collignon, 1931 from the Cenomanian of Madagascar is very similar to A. gurgitis (Brongniart) as figured and described by Kollmann (2005). Therefore, Collignon’S species is probably a synonym of the present species. T. cherbensis tHoMaS & peron in peron (1889: 19, figs. 1-3) from the Upper Albian of Tunisia resembles the present species in having concave whorls, numerous faint spiral threads, and last whorls with an angulate periphery but differs in having a less convex base, a very elevated spire, and a small umbilicus. T. regalis roemer (1841: pl. 12, fig. 7) from the Upper Cretaceous of North Germany differs in having angular whorls, a wide sub-sutural ramp, and a last whorl with rounded periphery.
Occurrence: Albian of France (brongniart 1822; Pictet & roux 1847; kollMann 2005), Upper Albian of England (J.
de C. SowerBy in Fitton 1836), Lower Cenomanian of the Kosmaj Mountain, central Serbia (this study).
Clade Sorbeoconcha Ponder & lindberg, 1996Superfamily Campaniloidea douvillé, 1904
Family Ampullinidae CoSSmann, 1919[ = Ampullospiridae Cox, 1930; = Gyrodinae wenz,
1938Globulariinae wenz, 1941; = Pseudamauridae
kowalke & bandel, 1996]
Genus Ampullina BowdiCh, 1822
Type species: Ampullaria depressa lamarCK, 1804, Eocene of Paris Basin, France.
Ampullina dupinii (deShayeS in leymerie, 1842)Fig. 7H-J
1842 Natica Dupinii n. sp. – leymerie, p. 13, pl. 16, fig. 7. 1842 Natica Dupinii leymerie. – d’orBigny, p. 158, pl. 173,
figs. 5-6.1991 Natica dupinii deShayeS in leymerie. – Mitrović-
Petrović & andJelković, p. 583.2005 Ampullina dupinii (deShayeS in leymerie). –
Kollmann, p. 60, pl. 8, fig. 7.2011 Ampullina dupinii (deShayeS in leymerie). – ayoub-
Hannaa & FürSicH, p. 126, fig. 8A-C.
Material: One specimen from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 110).
Description: Shell medium-sized (Table 5), low-spired, outline ovate. Spire conical and consisting of three overlapping whorls, separated by a moderately deep suture and forming 25% of total shell height. Whorls with moderately convex flank, abruptly decreasing in convexity adapically. Sub-sutural ramp narrow, slightly convex, and oblique in the early whorls, wide and slightly concave in subsequent whorls. Last whorl large, forming the main part of shell (75% of total shell height), sub-globose, with rounded periphery. Base broad and slightly convex. Aperture large (77% of total shell height), elongated tear-shaped, narrow posteriorly, and strongly rounded anteriorly with strongly convex out lip. Inner lip not callous. Umbilicus narrowly elongated and shallow (Fig. 7H). No ornament preserved.
Discussion: Ampullina dupinii (deShayeS in leymerie) from the Albian of France is characterized by a large tear-shaped aperture (strongly rounded anteriorly), a low-spired shell, a
Table 5. Dimensions (in mm) of Ampullina dupinii (deShayeS in leymerie, 1842).
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 110 28 25 21 22 17 115 3 0.89 0.75
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 39
narrowly elongated umbilicus, and a sub-globose last whorl with rounded flank.
A. ervyna (d’orBigny, 1842: 159, pl. 173, fig. 7) from the Albian of France resembles the present species in having a low-spired shell and smooth whorls but differs in having an ovate aperture, wider whorls, and a narrow umbilicus. A. umbilicoplicata Kollmann (1978: 176, pl. 1, fig. 15; pl. 2, figs. 16-22) from the Middle Albian-Lower Cenomanian of Austria differs in having a wider and concave sub-sutural ramp, a larger and deeper umbilicus with a well developed umbilical margin, and wide and shouldered whorls. A.? multistriata (Baily, 1855) as figured by kiel & bandel (2003: 454, fig. 2/11) from the Santonian-Campanian of South Africa differs from A. dupinii by having an elongated and ovate aperture.
Occurrence: Albian of France (leyMerie 1842; d’orbigny 1842; kollMann 2005), Cenomanian of Egypt (ayoub-Hannaa & FürSicH 2011), Lower Cenomanian of the Kosmaj Mountain, central Serbia (Mitrović-Petrović & andJelković 1991; this study).
Genus Globularia SwainSon, 1840
Type Species: Ampullaria sigaretina lamarCK 1804, Eocene of Europe.
Remarks: The genus Globularia can be easily distinguished from other ampullinid genera by having a narrow umbilicus, a wide and oblique aperture (laterally expanded), a rounded base, and a low-spired globose shell.
Globularia sp.Fig. 7K-M
Material: One specimen from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 94).
Description: Shell medium-sized, globose, higher than wide (D/H = 0.90; Table 6), and moderately high-spired. Spire consisting of three whorls, separated by moderately deep suture, rapidly decreasing in diameter adapically, and
accounting for 13% of the total shell height. Whorl sides evenly and slightly convex. Sub-sutural ramp flat and narrow in the early teleoconch whorls; wide, and concave in the last whorl. Umbilicus narrow, open. Last whorl large (HL/H = 0.87) and elongated, globose with slightly concave base (Fig. 7M). Aperture moderately wide, elongated, and sub-ovate, slightly oblique. Outer lip broken off. Inner lip concave and covered by callus (Fig. 7K). Whorl surface smooth.
Discussion: The specimen is similar to Gyrodes (Sohlella) gaultina (d’orBigny, 1842: 156, pl. 173, figs. 3-4) from the Albian of France in having a wide concave sub-sutural ramp and a less elevated spire but differs in being very elongated, higher than wide (D/H = 0.90 as opposed to 1.14; Table 6), and in having a narrow umbilicus and an elongated-ovate last whorl. Ampullina lyrata (J. de C. SowerBy, 1831) as figured by d’orBigny (1842: 161, pl. 172, fig. 5) from the Turonian of France differs in being larger, ovoid, and in having a large and elongated-ovate aperture. Globularia (Ampullella) umbilica (wade, 1926) as figured and described by Sohl (1960: 125, pl. 18, figs. 39-40) from the Maastrichtian of Tennessee differs in having a high spire, slightly convex to flat whorl flank, and a straight inner lip. It is also smaller (H = 10.4; D = 9.3 mm as opposed to H = 30, D = 27 mm; Table 6).
Occurrence: Lower Cenomanian of the Kosmaj Mountain, central Serbia.
Genus Gyrodes Conrad, 1860
Type species: Natica (Gyrodes) crenata Conrad, 1860 ( = Rapa supraplicata Conrad, 1858), Late Cretaceous of the Owl Creek Formation, Mississippi.
Subgenus Sohlella PoPenoe, Saul & SuSuki, 1987
Type species: Gyrodes canadensis whiteaveS, 1903, Campanian, Vancouver Island, Canada.
Remarks: The genus Gyrodes has been subdivided into different types and subgenera on the basis of (1) its sub-
Table 6. Dimensions (in mm) of Globularia sp. indet.
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 94 30 27 26 24 ?15 105 3 0.90 0.87
Table 7. Dimensions (in mm) of Gyrodes (Sohlella) gaultinus (d’orBigny, 1842).
Specimen No H D HL HA WA Sa° nw D/H HL/H
RGF JMP 93 30 ?35 23 22 19 117 3 1.17 0.77
RGF JMP 135 33 37 27 26 18 115 3 1.12 0.82
Range 30-33 35-37 23-27 22-26 18-19 115-11 3 1.12-1.17 0.77-0.82
Mean 31.5 36 25 24 18.5 116 3 1.14 0.80
eschweizerbart_xxx

40 W. Ayoub-Hannaa et al.
sutural ramp (e.g., concave, flat, narrow or wide), (2) umbilical margin (angulated or rounded), (3) suture, (4) growth lines (prosocline or opisthocline), and (5) presence or absence of crenulations or nodes near the suture. For instance, Sohl (1960: 116) suggested the subdivision of Gyrodes into three main types, based upon characters of the umbilical margin and suture. Subsequently, popenoe et al. (1987) subdivided the genus Gyrodes into the subgenera Gyrodes, Sohlella, Hypterita, and the group of G. americanus.
The subgenus Sohlella is characterized by a sub-globose shell, asymmetrically inflated and smooth whorls, narrow, slightly concave ramp, and a biangulate to sub-rounded umbilical margin (free of callus). Bandel (2000: 87) pointed out that the growth lines close to the suture are notched in the subgenus Gyrodes and not notched in the subgenus Sohlella. Stratigraphically, G. (Sohlella) ranges from the Cenomanian to Middle Maastrichtian. popenoe et al. (1987) assigned G. quercus PoPenoe, Saul & SuSuki, 1987, G. canadensis whiteaveS, 1903, G. pacificus PoPenoe, Saul & SuSuki, 1987, and G. expansus gaBB, 1864 to the subgenus Sohlella. Kollmann (2005) assigned Gyrodes gaultinus (d’orBigny, 1842) to the same subgenus. Bandel (1999) erected another new subgenus, Dockeryella, to distinguish low-spired Gyrodes species. For more details about the other subgenera see popenoe et al. (1987). The Lower Cenomanian specimens from Serbian fit perfectly into the subgenus Sohlella by their narrow, concave sub-sutural ramp and biangulate to sub-rounded umbilicus.
Gyrodes (Sohlella) gaultina (d’orBigny 1842)Fig. 7N-Q
1842 Natica Gaultina n. sp. – d’orBigny, p. 156, pl. 173, figs. 3-4.
?1847 Natica gaultina d’orBigny. – Pictet & roux, p. 184, pl. 18, fig. l.
1978 Gyrodes gaultinus (d’orBigny). – Kollmann, p. 174, pl. 1, figs. 1-6.
1991 Natica gaultina d’orBigny. – Mitrović-Petrović & andJelković, p. 583
2002 Gyrodes gaultinus (d’orBigny). – Kollmann, p. 47, pl. 6, figs. 97-98.
2005 Gyrodes (Sohlella) gaultinus (d’orBigny). – Kol-lmann, p. 59, pl. 8, figs. 10-11.
2006 Ampullina gaultina (d’orBigny). – Kiel, p. 457, fig. 3.4-3.5.
Material: Three specimens from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 5, 93, 135).
Description: Shell medium-sized (Table 7), globular, wider than high (D/H = 1.14 on average), low-spired. Spire consisting of three shouldered teleoconch whorls, rapidly decreasing in diameter adapically, and accounting for 20% of total test height. Teleoconch whorls smooth with strongly convex flank. Protoconch and early teleoconch whorls not preserved. Last whorl globular and slightly pointed apically. Sub-sutural ramp wide, slightly concave, and bearing faint growth lines (Fig. 7Q). Suture accompanied by a sharply tabulate shoulder from early to last whorl. Aperture large, slightly oblique, and elongated-ovate with straight inner lip. Outer lip strongly convex. Umbilicus moderately deep, sub-ovate, and steeply sloping. Ornament consisting of faint spiral threads.
Discussion: A wide and concave sub-sutural ramp, a moderately wide and sub-ovate umbilicus, nearly smooth and strongly convex whorls, and a wide, slightly concave base are the most diagnostic features of Gyrodes (Sohlella) gaultina (d’orBigny, 1842).
G. hebertianus (Cotteau, 1854: 29) from the Albian of France strongly resembles the present species in having a globular shell, a low spire, and shouldered whorls but differs in having a strongly umbilicated shell and height equaling the diameter. According to Cotteau (1854), the whorls of G. hebertianus are ornamented with faint collabral threads, crossed by spiral lines [smooth in G. (S.) gaultina]. G. (S.?) yolensis popenoe et al. (1987: 79, fig. 5.2, 5.11, 5.16, 5.21, 5.25, 5.30) from the Turonian of California resembles the present species in having a narrow and concave sub-sutural ramp and wide, globular shell but differs in the umbilical margin having two rounded angulations, a wide and deep umbilicus, less wide whorls, and well developed crenulations close to the suture. G. subexcavatus (tHoMaS & Peron in peron, 1889: 19, figs. 19-21) and G. (Dockeryella) renniei kiel & Bandel (2003: 458, fig. 4.3-4.4) from the Cenomanian of Tunisia and Santonian-Campanian of Cape Province, South Africa, respectively differ in having a depressed spire, strongly rounded whorls, a flat to slightly convex sub-sutural ramp (not shouldered), a wide, deep umbilicus, and a smaller height-width ratio than the present species. A narrow, deep umbilicus, and a slightly higher shell of G. (S.) woodsi Kiel & bandel (2003: 458, fig. 4.5-4.6) from the Campanian of Cape Province, South Africa, distinguish their species from G. (S.) gaultinus. G. (S.) tenella StoliCzKa, 1868 as figured and described by Bandel (2000: 88, pl. 4, figs. 1-2) from
Fig. 8. A-F – Pictavia pagoda (ForBeS, 1846) from the Lower Cenomanian, Kosmaj Mountain, central Serbia. A: Apertural view, internal mould showing a small, slit-like umbilicus; RGF JMP 111; B: abapertural view, internal mould; C: apertural view; D: apical view; RGF JMP 112; E: apical view, internal mould showing the deep suture; RGF JMP 90; F: abapical view with shell remains showing the canaliculated suture; RGF JMP 103. G-L – Pictavia pungens (J. de C. SowerBy in Fitton, 1836) from the Cenomanian of the same area. G: Apertural view with shell remains; H: abapertural view; I: apical view; RGF JMP 133B; J: Apertural view, internal mould; K: abapertural view; L: apical view; RGF JMP 123. M-O – Pseudamaura subbulbiformis (d’orBigny, 1850) from the Lower Cenomanian, Kosmaj Mountain; M: Apertural view, partly in shell preservation; N: abapertural view showing the canaliculated suture (arrowed); O: apical view; RGF JMP 112B. P-R – Littorina cf. strenua StoliCzKa 1868 from the Cenomanian of the same section. P: Apical view, incomplete with shell remains; Q: abapertural view showing an ovate aperture with callus on inner lip; R: abapertural view; RGF JMP 127.
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 41
Fig. 8.
eschweizerbart_xxx

42 W. Ayoub-Hannaa et al.
the Coniacian-Santonian of India differs in having a deeper umbilicus, less wide whorls of the spire, and a sub-globose last whorl. G. (S.) excavata (miChelin 1836) as figured and described by Kollmann (2005: 58, pl. 8, fig. 9) from the Albian of France differs from G. (S.) gaultina in having a wide and moderately deep umbilicus, and a very depressed spire with less wide and overlapping whorls.
Occurrence: Albian of France (d’orBigny 1842; Pictet & roux 1849; kollMann 2005), Eastern Alps (Kollmann 1978), northwestern Madagascar (Kiel 2006), late Lower Aptian of Austria (Kollmann 2002), and Lower Cenomanian of the Kosmaj Mountain, central Serbia (Mitrović-Petrović & andJelković 1991; this study).
Genus Pictavia CoSSmann, 1925
Type species: Natica pictaviensis d’orBigny, 1850, by original designation; Middle Jurassic (Bathonian) of south-eastern France.
Remarks: According to SquireS & Saul (2004: 29), the most diagnostic features of the genus Pictavia are (1) an elongated-oval shell, (2) a high spire, (3) a small slit-shaped umbilicus, (4) convex whorl sides, (5) prosocline growth lines, and (6) a folded columellar lip covering the umbilical area.
Pictavia pagoda (ForBeS, 1846)Fig. 8A-F
1846 Natica pagoda n. sp. – ForBeS, p. 136, pl. 12, fig. 14.
non 1887 Euspira pagoda ForBeS. – white, p. 185, pl. 17, figs. 11-12.
1991 Natica (Ampullina) cf. pagoda ForBeS. – Mitrović-Petrović & andJelković, p. 583.
Material: Six specimens from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 89-91, 103, 111, 112).
Description: Shell medium-sized, elongated-ovate, higher than wide (D/H = 0.68 on average; Table 8), moderately high-spired (spire: 28% of total shell height). Spire consisting of four convex whorls. Suture deep, canaliculated in some specimens (Fig. 8F). Protoconch and early teleoconch whorls not preserved. Last whorl rounded with slight sub-tabulate shoulder, and accounting for the main part of the shell. Umbilicus small and deep. Outer lip strongly convex. Inner lip strongly concave, smooth, and covered with thin callus. Aperture elliptical in outline, narrow posteriorly and widely rounded anteriorly. Whorl surface covered by prosocline growth lines.
Discussion: Pictavia pagoda (ForBeS) can be easily distinguished by having a deep to canaliculated suture, a small umbilicus, a moderately high spire, a rounded last whorl, and a spire with slightly convex whorl flank.
P. pagoda (ForBeS) as figured and described by white (1887) from the Middle Albian of the Sergipe Basin, Brazil differs from Indian material described by ForBeS in having an acute spire with a sharp apex, a very globose last whorl with a broad and flat sub-sutural ramp, and with an angular base. Therefore, maury (1937: 191) re-identified white’s specimens and considered them as synonyms of her new species P. sergipensis (1937: 191, pl. 14, figs. 3-4). P. santana SquireS & Saul (2004: 29, figs. 11-13) from the Upper Turonian of Santa Ana Mountains, Orange County, southern California, is similar to the present species in having an elliptical aperture, a rounded last whorl, and convex whorl flank but differs in
Table 8. Dimensions (in mm) of Pictavia pagoda (ForBeS, 1846).
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 91 40 31 31 31 14 75 4 0.78 0.78RGF JMP 103 46 31 32 28 12 64 5 0.67 0.69RGF JMP 111 48 30 32 29 16 70 5 0.63 0.67
RGF JMP 112A 53 34 40 30 17 69 5 0.64 0.75Range 40-53 30-34 31-40 28-31 12-17 64-75 4-5 0.63-0.78 0.67-0.78Mean 46.75 31.50 33.75 29.5 14.75 69.5 4 0.68 0.72
Table 9. Dimensions (in mm) of Pictavia pungens (J. de C. SowerBy in Fitton, 1836).
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 123 34 23 26 23 -- 75 6 0.68 0.74
RGF JMP 133B 25 17 19 15 12 82 5 0.68 0.76Range 25-34 17-23 16-26 15-23 12 75-82 5-6 0.68 0.74-0.76Mean 29.5 20 22.5 19 12 78.5 5 0.68 0.75
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 43
having an acute shell, a slightly impressed to indented suture, no umbilicus, and in being very elongated. P. pungens (J. de C. SowerBy in Fitton, 1836) from the Upper Albian of England differs in being smaller, less elongated, and in having a rounded small aperture, a moderately deep suture, and its whorl surface carries a reticulate ornamentation.
Occurrence: Upper Cretaceous of India (ForBeS 1846), Lower Cenomanian of the Kosmaj Mountain, central Serbia (Mitrović-Petrović & andJelković 1991; this study).
Pictavia pungens (J. de C. SowerBy in Fitton, 1836)Fig. 8G-L
1836 Littorina pungens n. sp. – J. de C. SowerBy in Fitton, p. 241, pl. 18, fig. 5.
1842 Natica Clementina (leymerie). – d’orBigny, p. 154, pl. 172, fig. 4.
1978 Pictavia pungens (J. de C. SowerBy). – Kollmann, p. 175, pl. 1, figs. 8-10.
2005 Pictavia pungens (J. de C. SowerBy). – Kollmann, p. 57, pl. 7, figs. 9-10.
Material: Two specimens from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 123, 133B).
Description: Shell medium-sized (Table 9), oviform, moderately thick-shelled, and moderately high-spired. Pleural angle approximately 78°. Spire consisting of five whorls, accounting for about 25% of total shell height, separated by a moderately deep suture, and with moderately convex sides. Last whorl large, rounded, with slight shoulder, separated from the spire whorls above by a deep suture, and accounting for 75% of total shell height. Outer lip convex. No umbilicus. Aperture moderately large, sub-elliptical, broadly rounded at base, and angular posteriorly. Whorl surface smooth except for numerous faint, collabral threads on the whorls of the spire, seen with a hand-lens.
Discussion: With respect to shell outline, size, and shape of aperture, Natica clementina d’orBigny from the Albian of France is identical to Pictavia pungens (J. de C. Sow-erBy) from the Upper Albian of England. In agreement with Kollmann (1978, 2005), N. clementina is therefore consid-ered herein as a junior synonym of P. pungens. N. laevigata d’orBigny (1842: 170, figs. 4, 6) from the Albian of France resembles the present species in having an oviform shell, a
strongly convex outer lip, and a rounded base with no umbi-licus but differs in having an elongated, ovate aperture and in being wider and larger than the present species. P. neoco-miensis (d’orBigny, 1843) as figured by Kollmann (2005: pl. 7, fig. 11) from the Hauterivian of France differs in having a narrow umbilicus, a wider aperture, and a thick inner lip.
Occurrence: Upper Albian of England (J. de C. SowerBy in Fitton 1836), Albian of France (d’orBigny 1842; Kollmann 2005), Austria (Kollmann 1978), Lower Cenomanian of the Kosmaj Mountain, central Serbia (this study).
Genus Pseudamaura FiSCher, 1885
Type species: Natica bulbiformis J. de C. SowerBy, 1823 from the Upper Cretaceous of the Northern Alps.
Remarks: The genus Pseudamaura FiSCher, 1885 includes species, which are medium- to large-sized and have a high-spired naticoid shell, a canaliculated suture, a small umbilicus (a small slit, occasionally lacking), a large and egg-shaped last whorl, and an ovate aperture.
Pseudamaura subbulbiformis (d’orBigny, 1850)Fig. 8M-O
1842 Natica bulbiformis SowerBy. – d’orBigny, p. 162, pl. 174, figs. 3, 3’ [non Natica bulbiformis J. de C. SowerBy in Sedgewick & MurcHiSon, 1835].
1850 Natica subbulbiformis n. sp. – d’orBigny, p. 191.2005 Pseudamaura subbulbiformis (d’orBigny). –
Kollmann, p. 63, pl. 8, figs. 12-15.
Material: Five specimens from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 71A, 88, 99, 112, 128).
Description: Shell large (Table 10), elongated-naticoid, and moderately high-spired. Spire conical and consisting of five whorls with slightly convex sides which account for 29% of the total shell height. Protoconch and early teleoconch whorls not preserved. Whorls of spire shouldered, wide, and gradually decreasing adapically. Suture channeled and sub-horizontal (Fig. 8N-O). Sub-sutural ramp narrow, flat to slightly concave, and smooth. Last whorl large, egg-shaped with rounded periphery, and accounting for 71% (on average)
Table 10. Dimensions (in mm) of Pseudamaura subbulbiformis (d’orBigny, 1850).
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 71 68 40 47 45 22 60 5 0.59 0.69RGF JMP 88 69 42 53 44 24 58 ?6 0.61 0.76
RGF JMP 112B 65 40 44 40 21 67 5 0.61 0.68Range 65-69 40-42 44-53 40-45 21-24 58-67 5-6 0.59-0.61 0.68-0.76Mean 67.33 40.67 48 43 22.33 61.67 5 0.60 0.71
eschweizerbart_xxx

44 W. Ayoub-Hannaa et al.
of the total shell height (Table 10). Basal part slightly rounded. No umbilicus. Aperture large, elongated-ovate (WA/HA = 0.52 on average), and slightly oblique. Outer lip strongly convex and thick. Inner lip slightly concave, thick, and covered by callus. Whorls smooth.
Discussion: d’orBigny (1842) identified Natica bulbiformis J. de C. SowerBy, 1832 from the Turonian of France. In 1850, he revised his collection and stated that SowerBy’s species, which came from the Upper Cretaceous of the Northern Alps, differs in having a more slender shell and a less elongated aperture. In addition, N. bulbiformis J. de C. SowerBy has thin outer and inner lips and a less rounded last whorl. Therefore, d’orBigny (1850: 191) ascribed his specimens to a new spe-cies, N. subbulbiformis.
Pseudamaura nobilis (StoliCzKa, 1868: 43, pl. 5, figs. 2, 4) from the Upper Cretaceous of India differs from the present species in having a relatively high spire, strongly convex whorls flank without shoulder, a small and sub-rounded last whorl, thin inner and outer lips, and a smaller and tear-shaped aperture. P. nobilis (StoliCzKa) as figured and described by Bandel (2000: 80, pl. 2, figs. 6-7) from the Coniacian-Santonian of India is closer to N. subbulbiformis than to P. nobilis. P. bulimoides (StoliCzKa, 1868: 42, pl. 5, fig. 5) from the Campanian-Maastrichtian Ariyalur Group of the same area differs in being smaller and in having a narrow, small aperture, a high spire, and a strongly convex whorl flank without shoulder. Phasianella? incerta ForBeS (1846: 123, pl. 13, fig. 8) from the Upper Cretaceous of India differs in having much more whorls with convex flank, a deep suture, and with a smaller and lunulated aperture. P. requieniana (d’orBigny, 1842: 161, pl. 174, fig. 2) from the Turonian of France strongly resembles the present species in having a large and ovate aperture, shouldered whorls with a slightly concave sub-sutural ramp, thick inner and outer lips, and a rounded base but differs in having a deep umbilicus with a thick umbilical margin and in being slightly wider than the present species and much more globular. P. lirata (wade, 1926: p. 165, pl. 57, figs. 5, 8) from the Campanian(?) to
Maastrichtian of Tennessee, USA, differs in having a very elevated spire, a large D-shaped aperture (flaring anteriorly), and in being smaller ( H = 30.6, D = 20.20, HA = 19.33 mm as opposed to H = 67.33, D = 40.67, HA = 43 mm on average; Table 10).
Occurrence: Turonian of France (d’orbigny 1842; kollMann 2005), Lower Cenomanian of the Kosmaj Mountain, central Serbia (this study).
Table 11. Dimensions (in mm) of Littorina cf. strenua StoliCzKa, 1868.
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 127 20 16 15 13 8 75 5 0.80 0.75
Fig. 9. Sketch of Littorina cf. strenua StoliCzKa, 1868 from the Lower Cenomanian, Kosmaj Mountain, central Serbia; RGF JMP 127.
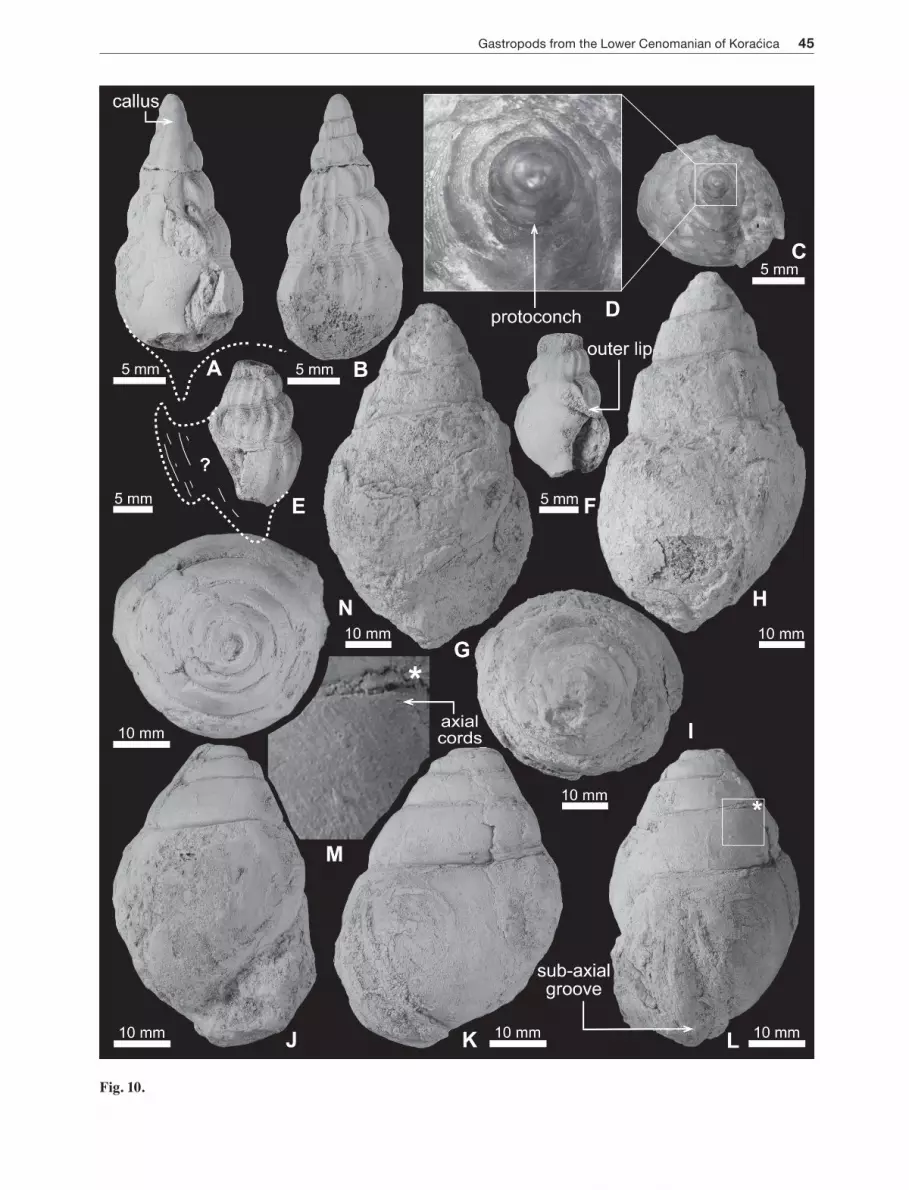
Fig. 10. A-F – Latiala? callositae sp. nov. from the Lower Cenomanian, Kosmaj Mountain, Serbia. A: Incomplete apertural view showing a band of callus extending to cover the protoconch and teleoconch whorls (arrowed); B: abapertural view showing irregular axial ribs, wider interspaces, and spiral threads close to suture; C: apical view; D: close-up of protoconch showing smooth and convex protoconch whorls; RGF JMP 108 (holotype); E: abapertural view, fragmented; F: apertural view showing an attachment of outer lip with penultimate whorl and smooth inner lip; RGF JMP 139 (paratype). G-I – Pterodonta elongata d’orBigny, 1843 from the Lower Cenomanian of the same locality. G: Apertural view, internal mould; H: abapertural view; I: apical view; RGF JMP 115. J-N – Pterodonta inflata d’orBigny, 1843 from the Lower Cenomanian of the Kosmaj Mountain. J: Apertural view, internal mould; K: abapertural view showing slightly extended outer lip; L: side view showing a sub-axial groove close to outer lip (arrowed); M: close-up showing faint and numerous axial cords; N: apical view; RGF JMP 125.
eschweizerbart_xxx
Gastropods from the Lower Cenomanian of Koraćica 45
Fig. 10.
eschweizerbart_xxx

46 W. Ayoub-Hannaa et al.
Clade Littorinimorpha golikov & Starobogatov, 1975
Superfamily Littorinoidea Children, 1834Family Littorinidae Children, 1834
Subfamily Littorininae Children, 1834[ = Echinininae roSewater, 1972 = Tectariinae
roSewater, 1972 =Melaraphidae Starobogatov & Sitnikova, 1983]
Genus Littorina FéruSSaC, 1822
Type species: Turbo littoreus linnaeuS, 1758, Western Europe, Recent.
Littorina cf. strenua StoliCzKa, 1868Figs. 8P-R, 9
cf. 1868 Littorina strenua n. sp. – StoliCzKa, p. 267, pl. 20, fig. 8.
cf. 1991 Littorina strenua StoliCzKa. – Mitrović-Petrović & andJelković, p. 583.
Material: One specimen from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 127).
Description: Shell medium-sized (Table 11), outline turbinate, and moderately high-spired (25% of total shell height). Spire consisting of five whorls separated by a deep suture. Pleural angle approximately 75°. Whorls strongly convex and abruptly increasing in diameter abapically without shoulder. Last whorl much inflated with strongly rounded periphery. Base rounded. Aperture ovate, and much higher than wide (WA/HA = 0.61). Inner lip thick, smooth (Figs. 8Q, 9), and covered with callus. Ornamentation consisting of faint spiral threads near the suture.
Discussion: The present specimen is very similar in general outline and size to Littorina strenua StoliCzKa, 1868 from the Aptian-Turonian Uttatur Group of India. However, the Serbian specimen is poorly preserved, and therefore it is assigned herein only tentatively to StoliCzKa’s species.
L. crassitesta StoliCzKa (1868: 265, pl. 20, figs. 16-17) from the Campanian-Maastrichtian Ariyalur Group of India resembles the present material in having a smooth and thick inner lip but differs in having an elongated-ovate last whorl with a distinct shoulder and in being more elongated than L. strenua.
Occurence: Aptian-Turonian Uttatur Group of India (StoliCzKa 1868), Lower Cenomanian of the Kosmaj
Mountain, central Serbia (Mitrović-Petrović & andJelković 1991; this study).
Superfamily Stromboidea raFineSque, 1815Family Aporrhaidae gray, 1850
Subfamily Aporrhainae gray, 1850( = Chenopidae deShayeS, 1865)
Genus Latiala Sohl, 1960
Type species: Anchura lobata wade, 1926 from the Maastrichtian Ripley Formation, North America.
Remarks: The genus Latiala can be easily distinguished from other aporrhaid genera in having a high-spired shell with strongly convex whorls flank, a thick outer lip, slightly callous inner lip, and well developed, irregular axial ribs, which are separated by wide and deep interspaces and disappear close to the anterior siphonal canal. According to kiel & bandel (2002: 84), the outer lip of Latiala is bilobed at the terminus with one lobe directed upward and a second, blunter lobe directed downward. Drepanochilus meeK, 1964 resembles Latiala in having a high-spired shell and well developed axial ribs, which disappear on the lower half of the body whorl but differs in having two or three spiral keels along the lower half of the body whorl. Anchura Conrad, 1860 differs from Drepanochilus in having only one keel on the last whorl. The Serbian specimens are not complete (outer lip and basal part are broken off), lack spiral keels on the last whorl, and their axial ribs are well developed, slightly sloping, irregular, and disappear on the lower half of the body whorl. Despite its similarities with the genus Latiala, the specimens from Serbia cannot be ascribed with confidence to this genus since the upper and outer margins of the wing are broken off. Therefore, they are here questionably assigned to Latiala.
Latiala? callositae sp. nov.Figs. 10A-F, 11
Etymology: Named for the callus (Latin: callositae), which extends across the protoconch and teleoconch whorls.
Holotype: Specimen RGF JMP 108 (Fig. 10A-C).
Paratypes: Two incomplete specimens RGF JMP 139, 140 (Fig. 10E, F).
Type locality: Grab well, Koraćica village, Kosmaj Mountain, central Serbia.
Table 12. Dimensions (in mm) of Latiala? callositae sp. nov.
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 108 27 ?18 12 16 -- 45 7 ?0.66 0.44
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 47
Type horizon: Lens of Lower Cenomanian bioclastic coarse-grained packstones to rudstones.
Diagnosis: Small-sized aporrhaid gastropod with a pleural angle of 45º; spire and last whorl of nearly equal height; protoconch and teleoconch whorls covered by a band of callus; aperture lenticular, small with small anterior siphonal canal; strong, opisthocline, and irregular axial ribs with wide and deep interspaces; well developed spiral threads close to suture; protoconch with three smooth and conical volutions.
Description: Shell small (Table 12), fusiform, and high-spired (spire: 56% of total shell height). Protoconch consisting of three conical and smooth whorls (H = 2.2; max D = 2.8 mm), the initial one very small, the following increasing rapidly in diameter abapically (Fig. 10C-O). Teleoconch high-spired, turreted, consisting of four whorls, which gradually decrease in diameter adapically and are separated by a moderately deep suture. Pleural angle 45º. Whorl sides of spire convex with a small sub-sutural ramp. Aperture small, oblique, lenticular in outline, and ending with a small anterior siphonal canal. Inner lip slightly concave, smooth, and highly callous. Outer lip broken off but some remains attached to the penultimate whorl (Fig. 10E, F). Whorls covered with 16 strong, opisthocline, and irregular-axial ribs separated by deep interspaces. The two strongest ribs with rounded crests
situated on the penultimate and last whorls. Upper third of last whorl with axial ribs; lower two-thirds with growth lines only. Faint spiral cords well developed close to the suture (sub-sutural spiral cords) and on the basal part of shell. A band of callus present on the ventral side of spire and extending across the protoconch whorls (Figs. 10A, 11).
Discussion: Latiala? callositae sp. nov. is similar to L.? ponsi kiel & bandel (2002: 85, fig. 1H-J) from the Upper Cretaceous of the Puimanyons Olistostrome, Spain, in having a band of callus on the spire and strong, irregular axial ribs but the latter differs in having a higher spire with ten whorls (four in the present species), irregular varices, a last whorl with less strong axial ribs, and smaller protoconch whorl (1.5 mm wide, 1.35 mm high). Moreover, L.? ponsi has been recorded from a younger stratigraphic level (Campanian).
L. lobata (wade, 1926: 150, pl. 52, figs. 11, 12) from the Upper Cretaceous of Tennessee resembles the present species in having well developed axial ribs and a moderately high-spired shell but differs in having more numerous axial ribs (30 as opposed to 16) and its wing is not attached to the penultimate whorl but only to the last whorl. L. calcaris (wade, 1926: 152, pl. 53, figs. 5, 8, 13) from the Upper Cretaceous of the same area differs in having a higher last whorl, sharp base, stronger axial ribs with rounded crests and deep and narrow interspaces, and in lacking a callus band.
Fig. 11. A, B – Sketches of Latiala? callositae sp. nov. from the Lower Cenomanian, Kosmaj Mountain, central Serbia. A: Apertural view showing a band of callus extending to protoconch and teleoconch whorls; B: abapertural view showing irregular axial ribs, spiral threads close to suture, smooth protoconch whorls, and hypothetical outline of outer lip; RGF JMP 108 (holotype).
eschweizerbart_xxx

48 W. Ayoub-Hannaa et al.
L.? tegulata (StoliCzKa, 1868) as figured and described by kiel & bandel (2002: 85, fig. 1F) from the Upper Cretaceous of Southern India differs in having a higher spire (slightly higher than the last whorl), a last whorl with stronger axial ribs which continue onto the rostrum, and a larger shell. L. besairiei (Collignon, 1949) as figured and described by Kiel (2006: 466, fig. 5/12-16) from the Albian, Mahajanga Basin of Madagascar is similar to the present species in having strongly convex whorls, a high-spired teleoconch, and a wing attached to the penultimate whorl but differs in having 24 thin, opisthocyrt to opisthocline ribs per whorl, a deep suture, and a protoconch that is smaller and consists of four whorls.
Family Colombellinidae FiSCher, 1884( = Columbellariidae zittel 1895 = Zitteliidae,
SChilder, 1936)
Genus Pterodonta d’orBigny, 1843
Type species: Pterodonta inflata d’orBigny, 1843 from the Cenomanian-Turonian of France.
Remarks: The genus Pterodonta is characterized by a large to very large shell, a globose adult last whorl, a deep suture, an axial groove, which extends nearly to the outer lip, strongly convex whorls, an ovate aperture, and a short
anterior siphonal canal (d’orBigny 1843; ayoub-Hannaa & FürSicH 2011: 135). According to the latter authors Pterodonta differs from Tylostoma Sharpe, 1849 in having a less plump last whorl, a projected and incurved outer lip, a deep suture, and sinuous growth lines. In addition, Tylostoma has an ovate or globose smooth shell, a moderately elevated spire, and an ovate-lunate aperture.
Pterodonta elongata d’orBigny, 1843Fig. 10G-I
1843 Pterodonta elongata n. sp. – d’orBigny, p. 316, pl. 218, fig. 2.
1843 Pterodonta intermedia. – d’orBigny, p. 319, pl. 220, fig. 1.
?1843 Pterodonta ovata. – d’orBigny, p. 317, pl. 218, fig. 3.?1974 Pterodonta ovata d’orBigny. – albaneSi & buSSon,
pl. 24, fig. 1.1985 Pterodonta elongata d’orBigny. – Kollmann, p. 97,
fig. 5f, g.1991 Pterodonta intermedia d’orBigny. – Mitrović-Pet-
rović & andJelković, p. 583, pl. 1, fig. 5.
Material: One specimen from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 115).
Fig. 12. A-F – Tylostoma elevatum (Shumard, 1853) from the Lower Cenomanian, Kosmaj Mountain, central Serbia. A: Apertural view, internal mould; B: close-up showing spiral cords next to suture (arrowed); C: axial section with, strongly oblique oval whorls; D: side view showing an axial groove on the penultimate whorl (arrowed); E: abapertural view showing a sub-axial groove on last whorl (arrowed); F: apical view showing the last two grooves (arrowed); RGF JMP 126. G-J – Tylostoma cf. minimum white, 1887 from the Lower Cenomanian of the same section. G: Apertural view, internal mould; H: abapertural view showing the slight shoulder of the last whorl and a large penultimate whorl; I: axial section showing oblique ovate whorls; J: apical view; RGF JMP 137. K-M – Tylostoma sp. from the Kosmaj Mountain. K: Apical view, internal mould; L: apertural view; M: Abapertural view; RGF JMP 6. N-Q – Vanikoropsis cassisiana (d’orBigny, 1842) from the Lower Cenomanian of the same area. N: abapertural view, internal mould showing slightly concave base; O: apertural view showing strongly convex, thick outer lip, and small umbilicus; P: apical view; Q: close-up showing tuberculated spiral threads; RGF JMP 117.
Table 13. Dimensions (in mm) of Pterodonta elongata d’orBigny, 1843.
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 115 75 40 50 42 25 45 5 0.53 0.67
Table 14. Dimensions (in mm) of Pterodonta inflata d’orBigny, 1843.
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 86 66 43 47 40 ?20 63 4 0.65 0.71RGF JMP 87 45 33 35 30 16 65 4 0.73 0.78RGF JMP 102 57 37 43 38 -- 60 4 0.65 0.75RGF JMP 125 55 35 40 33 19 60 5 0.64 0.73
Range 45-66 33-43 35-47 30-40 16-?20 60-95 4-5 0.64-0.73 0.71-0.78Mean 55.75 37 41.25 35.25 18.33 62 4 0.67 0.74
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 49
Fig. 12.
eschweizerbart_xxx

50 W. Ayoub-Hannaa et al.
Description: Shell large-sized (H = 75, D = 40 mm; Table 13), elongated biconical (D/H = 0.53), and moderately high-spired. Spire consisting of five whorls, separated by a moderately deep suture. Pleural angle 45º. Flank of whorls strongly convex, gradually decreasing adapically. Last whorl large, with rounded periphery, elongated, slightly inflated, and accounting for 67% of the total shell height. Base narrow and slightly rounded. Aperture narrow-ovate (WA/HA = 0.59) with small anterior notch. Outer lip strongly convex. Inner lip slightly concave.
Discussion: The present material is an internal mould without sign of ornamentation except for faint tuberculated spiral cords on earlier whorls. With respect to general outline, convexity, and number of whorls, Pterodonta intermedia d’orBigny, 1843 is identical to P. elongata. d’orBigny separated the two species on the basis of the small size of P. elongata. Kollmann (1985: 99) noted that moulds of different size from the Charente collection (France) have the same whorl shape. Following Kollmann, P. intermedia is therefore regarded as a junior synonym of P. elongata. Similarly, P. ovata d’orBigny, 1843 strongly resembles the present species in general outline but differs merely in having a much extended sub-axial groove running parallel to the outer lip. P. deffisi tHoMaS & Peron (1889: 20, figs. 17-18) and P. gigantica aBBaSS (1963: 83, pl. 7, figs. 1-3) from the Cenomanian of Tunisia and Egypt, respectively differ in having fewer whorls, which abruptly decrease in diameter adapically, and a spiral groove on the last whorls.
Occurrence: Cenomanian-Upper Santonian of France (d’orBigny 1843; Kollmann 1985; 2005: 136), probably from the Upper Cretaceous (Cenomanian-Turonian) of Algeria (albaneSi & buSSon 1974), Cenomanian of central Serbia (Mitrović-Petrović & andJelković 1991; this study).
Pterodonta inflata d’orBigny, 1843Fig. 10J-N
1843 Pterodonta inflata n. sp. – d’orBigny, p. 318, pl. 219.1862 Pterodonta subinflata. – Coquand, p. 179, pl. 6, fig.
1.1971 Pterodonta inflata d’orBigny. – Collignon, p. 153,
pl. A, fig. 4.1991 Pterodonta inflata d’orBigny. – Mitrović-Petrović
& andJelković, p. 583, pl. 1, fig. 4.1985 Pterodonta inflata d’orBigny. – Kollmann, p. 99, fig.
5d, e.2005 Pterodonta inflata d’orBigny. – Kollmann, p. 137,
pl. 14, fig. 18a, b.
Material: Four specimens from the Lower Cenomanian of
the Kosmaj Mountain, central Serbia (RGF JMP 86, 87, 102, 125).
Description: Shell medium- to large-sized (H = 45-66 mm; Table 14), broadly ovate and moderately high-spired. Spire consisting of four slightly convex whorls separated by a moderately deep suture. Protoconch and first few teleoconch whorls not preserved. Pleural angle approximately 62º (on average). Last whorl large, globose, and accounting for 74% (on average; Table 14) of total shell height. Base broadly rounded. Aperture narrowly elliptical with small anterior notch. Outer lip convex and slightly expanded with deep sub-axial groove close to outer lip (Fig. 10K, L). Inner lip straight to slightly concave.
Discussion: The present specimens are internal moulds with no sign of ornamentation except for faint axial threads on the penultimate whorl (Fig. 10M). Pterodonta inflata d’orBigny, 1843 can be easily distinguished by its very globose and large last whorl with a sub-axial groove, rounded base, and smooth and slightly convex whorls, which gradually increase abapically.
P. elongata d’orBigny (1843: pl. 218) from the Cenomanian of France resembles the present species in having a sub-vertical groove and moderately convex and smooth whorls but differs in having an angular base, a narrow-conical last whorl, and in being very elongated (D/H = 0.53 as opposed to 0.67 on average). P. subinflata Coquand, 1862 from the Santonian of Algeria is identical to the type species P. inflata as figured and described by d’orBigny (1843) from the Cenomanian-Turonian of France in having convex whorls, a rounded base, a globose last whorl, and a narrow-ovate aperture (see Kollmann 2005: pl. 14, fig. 18). P. subinflata is therefore regarded as a junior synonym of P. inflata. P. deffisi (tHoMaS & Peron in peron, 1889: 83, pl. 20, figs. 17-18) from the Cenomanian of Tunisia differs in having a very elevated last whorl, a sharp base, fewer, strongly convex whorls, a spiral groove on the last whorl, and a shell more acute than the present species.
Occurrence: Cenomanian-Turonian and probably Coniacian of France (d’orBigny 1843; Kollmann 1985, 2005), Turonian of Algeria (Collignon 1971), and Lower Cenomanian of the Kosmaj Mountain, central Serbia (Mitrović-Petrović & andJelković 1991: 583; this study).
Family Tylostomatidae StoliCzKa, 1868Genus Tylostoma Sharpe, 1849
Type species: Tylostoma torrubiae Sharpe, 1849, by subsequent designation (white 1880: 142); Cretaceous, Portugal.
Table 15. Dimensions (in mm) of Tylostoma elevatum (Shumard, 1853).
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 126 46 30 32 28 -- 72 4 0.65 0.69
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 51
Remarks: As stated by SquireS & Saul (2004), there is some confusion about the type species of the genus Tylostoma. Most palaeontologists followed wenz (1941: 1026) and used T. globosum Sharpe, 1849 as the type species. In fact, Sharpe (1849) did not designate a type species for the genus Tylostoma. KaBat (1991: 437) correctly reported the type species to be T. torrubiae Sharpe, 1849. For a detailed history of designation of the type species of Tylostoma, see KaSe (1984: 156) and SquireS & Saul (2004: 24-25).
Tylostoma can be easily distinguished by having an elongated naticiform and anomphalous shell, a low to high spire, and a thick outer lip, which forms internal varices.
Tylostoma elevatum (Shumard, 1853)Figs. 12A-F, 13
1853 Globiconcha? elevata n. sp. – Shumard, p. 208, pl. 4, fig. 4.
1947 Tylostoma elevatum (Shumard). – Stanton, p. 68, pl. 53, figs. 1-5.
Material: Two specimens from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 118, 126).
Description: Shell moderately large (Table 15), outline elongated-ovate, moderately high-spired (spire: 31% of total
shell height). Spire conical and consisting of four whorls, abruptly decreasing in diameter adapically, separated by a moderately deep suture. Protoconch whorls not preserved. Whorl flank convex with rounded periphery and a slight shoulder. Last whorl ovoid, large, with rounded periphery, and accounting for 69% of total shell height (Table 15), base rounded. Penultimate and last whorl carrying two varices, which left deep grooves on the internal moulds (Figs. 12D, E, 13A, B). Last whorl groove wide, V-shaped in cross-section, deep, sub-axial, which indicates that outer lip formed a thickened varix. Penultimate groove narrower and axial, which indicates that outer lip slightly extended onto the spire (Fig. 12D). Aperture oval and oblique (Figs. 12C, 13C). Whorl surface smooth, except for faint spiral threads close to suture of last whorl (Fig. 12B).
Discussion: The Serbian specimens strongly resemble Tylostoma elevatum (Shumard, 1853) as figured and described by Stanton (1947) from the Upper Cretaceous of Texas in having the same size, and general outline, a wide and deep sub-axial groove on the last whorl, wide and convex whorls, an oblique, ovate aperture, and in being smooth. However, the outer lip of the latter extends onto earlier whorls (only on penultimate and last whorl in the Serbian specimens; Fig. 12F).
T. materinum white (1887: 189, pl. 17, fig. 1) from the Albian of the Sergipe Basin (Brazil) resembles the present species in having smooth and strongly convex whorls and a
Fig. 13. A-C – Sketches of Tylostoma elevatum (Shumard, 1853) from the Lower Cenomanian, Kosmaj Mountain, central Serbia. A: Abapertural view showing a subaxial varix (leaving groove on internal mould) on last whorl; B: apical view showing the two varices of the penultimate and last whorl; C: longitudinal whorl section showing strongly oblique ovate whorls; RGF JMP 126.
Table 16. Dimensions (in mm) of Tylostoma cf. minimum white, 1887.
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 137 39 26 23 ?19 -- 65 5 0.67 0.59
eschweizerbart_xxx

52 W. Ayoub-Hannaa et al.
deep suture but differs in being larger (H = 150, D = 80 mm as opposed to H = 46, D = 30 mm; Table 15) and in having less wide whorls and a high spire. T. fleuriausa (d’orBigny, 1842: 144, pl. 169, fig. 18) from the Campanian of France differs in having a narrow-lunate aperture, narrower whorls, and a conical and high spire. T. tumidum (Shumard, 1853) as figured and described by Stanton (1947: 70, pl. 53, figs. 6-9) from the Upper Cretaceous of Texas strongly resembles the present species in being smooth and in having sub-axial grooves on the penultimate and last whorls, an ovate, oblique aperture, and strongly convex whorls. It differs in having a more and conical elevated spire, more whorls, which gradually decrease in diameter adapically, and a large shell (H = 67, D = 44 mm as opposed to H = 46, D = 30 mm).
Occurrence: Upper Cretaceous of Texas (SHuMard 1853; Stanton 1947), Lower Cenomanian of central Serbia (this study).
Tylostoma cf. minimum white, 1887Fig. 12G-J
cf. 1887 Tylostoma minimum n. sp. – white, p. 188, pl. 17, figs. 6-9.
cf. 1937 Tylostoma minimum white. – maury, p. 195, pl. 14, figs. 9, 16-17.
Material: One specimen from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 137).
Description: Shell medium-sized (Table 16), outline elongated-ovate (D/H = 0.67), and high-spired (spire: 41% of total shell height). Spire conical and consisting of five whorls, separated by a moderately impressed suture, and gradually decreasing in diameter adapically. Whorls of spire moderately convex with rounded periphery. Last whorl accounting for 59% of total shell height, ovate in outline with slight shoulder (Fig. 12H) and with a very narrow axial varix. Penultimate whorl much higher and wider than earlier whorls (Fig. 12H). Axial whorl section displaying obliquely ovate whorls (sharply pointed adapically, strongly rounded abapically; Fig. 12I). Outer lip convex and slightly extended anteriorly. Base narrow and rounded with no umbilicus. Whorl surface smooth.
Discussion: The Serbian specimen strongly resembles Tylostoma minimum white, 1887 from the Middle Albian
of the Sergipe Basin, Brazil in having the same size (H = 39, D = 26 mm; Table 16), a large penultimate whorl, a narrow axial varix along peripheral margin of last whorl, and smooth, convex whorls, but differs in having a slightly acute shell.
T. minimum is very close to Tylostoma allisoni SquireS & Saul (2004: 25, figs. 2-6) from the Late Aptian of California in having an ovate-elongated shell, smooth and rounded whorls, and a narrow axial varix along the peripheral margin of the last whorl but the latter species differs in being very large (height up to 123 mm as opposed to 39 mm), very elongated (D/H = 0.50 as opposed to 0.67; Table 16), and in having a deep suture. T. cossoni tHoMaS & Peron in peron (1889: 57, pl. 19, figs. 24-25) from the Turonian of Tunisia differs in being very globose, large, and in having a large, elongated aperture, a less elevated spire, and a strongly convex outer lip. The Serbian specimen is also similar to T. pallaryi (peron & Fourtau, 1904) as figured and described by greCo [1916: 150 (92), pl. 18 (10)] from the Turonian of the Eastern Desert, Egypt but the latter differs in having a wide and sub-ovate aperture, less elevated last whorl with strongly convex flank, and in lacking an axial varix.
Tylostoma sp.Fig. 12K-M
Material: One internal mould from the Lower Cenomanian of the Kosmaj Mountain (RGF JMP 6).
Description: Internal mould medium-sized (Table 17), ovate, and moderately high-spired. Spire conical and consisting of four whorls, separated by a moderately deep suture, and accounting for 22% of total shell height. Whorls of spire convex, with rounded periphery, and gradually decreasing in diameter adapically. Last whorl large, ovoid, with a sub-axial varix, large, rounded periphery, and representing 78% of total shell height. Base strongly rounded. Aperture large (HA/H = 67% of total shell height), semi-lunate, and oblique with a strongly convex, thick outer lip as indicated by the sub-axial varix on the left side of the last whorl. No ornamentation preserved.
Discussion: The Serbian specimen strongly resembles Tylostoma cossoni tHoMaS & Peron in peron (1889: 57, pl. 19, figs. 24-25) from the Turonian of Tunisia in having a semi-lunate, oblique aperture, convex and smooth whorls, a thick outer lip, and a well developed varix on the last whorl but differs in being smaller (H = 27 mm as opposed
Table 17. Dimensions (in mm) of Tylostoma sp. indet.
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 6 27 23 21 18 12 79 4 0.85 0.78
Table 18. Dimensions (in mm) of Vanikoropsis cassisiana (d’orBigny, 1842).
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 117 34 26 24 ?20 -- 75 4 0.76 0.70
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 53
Fig. 14. A-F – Pseudonerinea jovankae sp. nov. from the Lower Cenomanian, Kosmaj Mountain, central Serbia. A: Abapertural view showing high and slightly concave whorls with deep, sinuous suture; B: apertural view showing deep suture; C: longitudinal whorl section showing smooth columellar and outer lip folds (fold formula: 0000); D: close-up of the earlier whorl showing faint columellar folds; RGF JMP 124 (holotype); E: apertural view with shell remains showing smooth whorls; F: longitudinal whorl section lacking columellar folds; RGF JMP 114. G-J – Avellana subincrassata d’orBigny, 1850 from the Lower Cenomanian of the same section. G: Apertural view, composite mould, H: abapertural view; I: close-up showing details of ornamentation (well developed spiral ribs with numerous, faint axial striae in-between); J: apical view; RGF JMP 73.
eschweizerbart_xxx

54 W. Ayoub-Hannaa et al.
to H = 84 mm) and in having a very globose last whorl. T. globosum Sharpe (1849: 379, pl. 9, figs. 5-6) from the Turonian of Portugal differs in having highly overlapping whorls, a very low spire, a broadly ovate shell, and in being larger. T. fleuriausa (d’orBigny, 1842: 169, fig. 18) from the Campanian of France differs in being very elongated, with a less globose last whorl and a slightly concave base, the latter being strongly rounded and wide in the Serbian specimen. Moreover the whorls are less convex.
Superfamily Vanikoroidea gray, 1840Family Vanikoridae gray, 1840
[ = Naricidae réCluz, 1845 = Merriidae hedley, 1918= Lyocyclinae thiele, 1925 = Caledoniellidae roSe-
water, 1969= Conradiinae goliKov & StaroBogatov, 1987]
Genus Vanikoropsis meeK, 1876
Type species: Natica toumeyana Meek & Hayden, 1856 (by original designation), Upper Cretaceous, mouth of the Judith River, Fox Hills Group, USA.
Remarks: meeK (1876) erected the genus Vanikoropsis on the basis of thick, non-umbilicate shells with strong growth rugae and placed it in the family Vanikoridae. CoSSmann (1925) and wenz (1942) assigned the genus to the family Naticidae due to the lack of an umbilicus. Sohl (1967: B22) pointed out that the type species Natica toumeyana Meek & hayden, 1856 is incomplete (last whorl partly broken off). Apart from being poorly preserved (Meek & Hayden, 1856: pl. 5, figs. 15-16) the holotype has a narrow umbilical slit (for more details, see Sohl 1967: B22).
The genus Vanikoropsis is characterized by having a thick naticiform shell, a moderately high spire, a small umbilicus, an ovate aperture with strongly convex outer lip, and strong spiral cords crossed by strong sub-axial costae.
Vanikoropsis cassisiana (d’orBigny, 1842)Fig. 12N-Q
1842 Natica Cassisiana n. sp. – d’orBigny, p. 166, pl. 175, figs. 1-4.
1991 Vanikoropsis (Natica) cassisiana (d’orBigny). – Mitrović-Petrović & andJelković, p. 583, pl. 1, fig. 3.
2005 Vanikoropsis? cassisiana (d’orBigny). – Kollmann, p. 65, pl. 8, fig. 17.
Material: One specimen from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 117).
Description: Shell medium-sized (Table 18), biconical, and moderately high-spired. Protoconch and early teleoconch whorls not preserved. Apical angle 75º. Spire consisting of four whorls, separated by moderately deep suture and accounting for 30% of total shell height. Whorl sides of spire
convex with no shoulder. Last whorl ovate in outline, large, and representing 70% of total shell height. Aperture ovate, rounded anteriorly and angulated posteriorly. Base slightly broadened and concave (Fig. 12N). Outer lip strongly convex. Whorl surface covered with faint axial ribs, crossed by spiral cords. Intersection of axial ribs and spiral cords produce a reticulated pattern with tuberculate appearance (Fig. 12N, Q).
Discussion: Vanikoropsis cassisiana (d’orBigny, 1842) can be easily distinguished by having a biconical shell, convex whorls, a large, ovoid last whorl, a slightly concave to flat base, and a wide and ovate aperture.
V. demipleurus Stilwell & HenderSon (2002: 459, fig. 5.1-5.2, 5.4, 5.7) from the Cenomanian of northern Australia differs in being smaller (less than 10 mm in height) and in having a broad apical angle (94º as opposed to 75º), a less elevated spire with three to four whorls, and a weakly channeled suture. V. nebrascensis (Meek & Hayden, 1860) as described and figured by Sohl (1967: B22, pl. 5, figs. 1, 5-10, 12, 14, 17; pl. 6, figs. 1-4, 11) from the Upper Cretaceous of the Western United States (Wyoming) differs in having a thicker outer lip, stronger ornamentation, a sub-rounded aperture, a narrow umbilicus, and in being smaller (H = 13.2-19.3 mm as opposed to 34 mm).
Occurrence: Cenomanian of France (d’orbigny 1842; Kollmann 2005), Lower Cenomanian of central Serbia (Mitrović-Petrović & andJelković 1991; this study).
Clade Heterobranchia gray, 1840[Informal Group “Lower Heterobranchia” ( = Allo-
gastropoda)]Superfamily Nerineoidea zittel, 1873
Family Nerineidae zittel, 1873Subfamily Nerineinae zittel, 1873
( = Phaneroptyxidae pChelintSev, 1965= Fibuloptygmatididae haCoBJan, 1973)
Genus Pseudonerinea de loriol, 1890
Type species: Pseudonerinea blauenensis de loriol, 1890, by subsequent designation of dietriCh (1925), Lower Jurassic-Lower Cretaceous, Germany.
Remarks: Nerineids vary in shape and ornamentation, but the most important elements for their generic classification are the internal features of the shell. Most nerineids are characterized by the presence of internal folds on the columella and outer lip [parietal fold (P), columellar fold (C), labial fold (L), and basal fold (B)]. The number of folds ranges from one to seven (e.g., Neoptyxis pChelintSev, 1934, Eunerinea Cox, 1949, Ptygmatis BoSe, 1906, Pseudonerinea de loriol, 1890). For details about the internal structure of nerineids, see delpey (1939), Cox (1949), wieCzoreK (1979), BarKer (1990), Sirna (1995), and ayouB-hannaa & FürSicH (2011).
Pseudonerinea de loriol, 1890 is characterized by having smooth and slightly concave whorls, a narrow anterior siphonal canal an ovate, small aperture, no whorl folds (P = 0, C = 0, L
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 55
= 0, B = 0; fold formula: 0000), a deep suture and a flat sutural band. Pseudonerinea resembles Aptyxiella FiSCher, 1885 in lacking columellar folds and outer lip folds (wieCzoreK 1979: 301, fig. 2). Pseudonerinea differs, however, in having a more elongated turriculate shell, a narrower aperture, and in being smooth. The Serbian specimens are assigned to Pseudonerinea with respect to their internal and external features.
Pseudonerinea jovankae sp. nov.Figs. 14A-F, 15
Etymology: In honour of Jovanka Mitrović-Petrović, pro-fessor of the Belgrade University, who collected the material from the Grab well locality during the 70’s of the last century and for her contributions to the Mesozoic and Cenozoic pa-laeontology and geology of Serbia.
Holotype: Composite mould of specimen RGF JMP 124 (Fig. 14A-D).
Paratypes: Two incomplete internal moulds RGF JMP 114 and 121 (Fig. 14E, F).
Type locality: Grab well, Koraćica village, Kosmaj Mountain, central Serbia.
Type horizon: Lens of Lower Cenomanian coarse-grained bioclastic packstones to rudstones.
Diagnosis: Very large nerineid (L = 205, W = 38 mm) with high and slightly concave whorls having a slight bulge adapically and abapically, a deep suture, a small, oblique elongated-ovate aperture, a small and narrow anterior siphonal canal, and smooth whorls. Neither columellar nor outer lip folds are developed (fold formula: 0000).
Description: Shell large- to very large-sized (H = 205 mm, W = 38 mm), widely turriculate, elongated, and high-spired. Spire angle ~10°. Spire consisting of ?4-6 whorls, separated by a deep suture. Teleoconch whorls wide, with concavo-convex flank (concave in the middle part, with slight bulge close to adapical and abapical sutures; Fig. 15A), very high [i.e., the height of whorl above the last whorl (HW/H) = 13% of the total shell height; Fig. 15A], and gradually decreasing adapically. Protoconch and early teleoconch whorls not preserved. Last whorl moderately rounded with sub-angular base. Suture canaliculated and locally faintly sinuous (Fig. 14A). Aperture small, oblique, elongated-ovate, narrowing
Fig. 15. A-B – Pseudonerinea jovankae sp. nov. from the Lower Cenomanian, Kosmaj Mountain, central Serbia. A: Apertural view showing deep suture, high and slightly concave whorls, and small anterior notch; B: longitudinal whorl section showing smooth, ovate whorls (no columellar and outer lip folds) except for an early whorl which shows faint columellar folds; RGF JMP 124 (holotype).
eschweizerbart_xxx

56 W. Ayoub-Hannaa et al.
anteriorly and posteriorly, and with small anterior siphonal canal. Axial whorl section without internal columellar and outer lip folds (P = 0, C = 0, L = 0, B = 0; fold formula: 0000; Figs. 14C, 15B), except for the earlier teleoconch whorls of some specimens which probably possess small columellar folds (Fig. 14D). Shell smooth.
Discussion: The species is similar to Pseudonerinea? presidiensis Stanton (1947: 95, pl. 63, figs. 26-27) from the Upper Cretaceous of Texas in having smooth whorls, a narrowly ovate aperture, a narrow base, a short siphonal anterior canal, and in lacking internal folds but the latter differs in being smaller (L = 59, W = 24 mm as opposed to L = 205; W = 38 mm) and has a more slender shell, a slightly higher apical angle (pleural angle > 15° as opposed to 10°), and well developed sutural band. In addition, the teleoconch whorls of P.? presidiensis are lower with convex sides and separated by a moderately deep suture. P. vaceki Kollmann (2002: 50, pl. 3, fig. 32) from the Lower Barremian-Lower Aptian of Austria resembles the present species in having high whorls and a deep suture. It differs in having strongly concave whorls, in which the adapical half is bulging (very weak in the present species) whereas the abapical one is angulated; and a narrow, high, tear-shaped aperture. Moreover, it is narrow-turriculate and smaller (H = 70, W = 20 mm). P. proctori (Cragin, 1863) as figured and described by Stanton (1947: 95, pl. 63, figs. 22-23, 28-30) from the Upper Cretaceous of Arkansas differs in having a more slender and more acute shell with an angular base, a greater number of slightly convex teleoconch whorls, well developed spiral tubercles above and below the suture and in being smaller. The axial cross-section of P.? arkansensis Stanton (1947: 95, pl. 63, figs. 9, 10) from the Upper Cretaceous of the same area is very similar to the present species (oblique-ovate whorls) but the shell is much smaller. In addition, P.? arkansensis has a very acute, slender shell, numerous, very short, and moderately convex whorls (20), a deep suture, a narrow aperture and a distinctly angulated base. P. stirtoni alliSon (1955: 423, pl. 43, figs. 1-2) from the Mid-Cretaceous of China resembles the present species in having high whorls but differs in being smaller and in having slightly convex, acute whorls, a narrow-ovate aperture, and a shallow suture. In addition, the latter species has been described based on incomplete internal moulds. Morphologically, the material ascribed by d’orBigny (1842: 77, pl. 158, figs. 1-2) to Nerinea gigantea d’HoMbre-FirMaS from the Albian of France resembles the present species in having a large shell and wide whorls but differs internally in having columellar and outer lip folds (fold formula: 1110 as opposed to 0000). Therefore, Kollmann (2005: 27, pl. 3, fig. 6) assigned the latter species to the genus Eunerinea Cox, 1949. N. dubia BlanCKenhorn [1927: 150, pl. 7 (3), figs. 55-56] from the Cenomanian of Syria resembles P. jovankae in having high and smooth whorls but these whorls are narrower
with flanks that are slightly more convex than in the present species.
Superfamily Ringiculoidea philippi, 1853Family Ringiculidae philippi, 1853[ = Avellaninae haCoBJan, 1976]
Genus Avellana d’orBigny, 1842
Type species: Avellana avellana (Brogniart, 1822) from the Cenomanian of France.
Remarks: The genus Avellana strongly resembles the genus Eriptychia meeK, 1876 but differs in columellar structures. According to kiel & bandel (2001) the bifurcate basal columellar folds of Eriptychia might be a further element of distinction.
Avellana subincrassata d’orBigny, 1850Fig. 14G-J
1842 Avellana incrassata [SowerBy]. – d’orBigny, p. 133, pl. 168, figs. 13-16.
1850 Avellana subincrassata. – d’orBigny, p. 128.2002 Avellana subincrassata d’orBigny. – Kollmann, p.
54, pl. 6, figs. 99-100.2002 Avellana incrassata (SowerBy). – cleevely & MorriS,
p. 175, pl. 29, figs. 6-8.2005 Avellana subincrassata d’orBigny. – Kollmann, p.
49, pl. 6, figs. 17-18.
Table 19. Dimensions (in mm) of Avellana subincrassata d’orBigny, 1850.
Specimen No H D HL HA WA Sa° nw D/H HL/HRGF JMP 73 20 16 17 16 10 105 3 0.80 0.85
Fig. 16. Species diversity (number of taxa) of the gastropod families from the Lower Cenomanian of the Kosmaj Mountain, central Serbia.
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 57
Fig. 17. Autecology of some characteristic taxa of the Lower Cenomanian storm-wave concentration.
eschweizerbart_xxx

58 W. Ayoub-Hannaa et al.
Material: One specimen from the Lower Cenomanian of the Kosmaj Mountain, central Serbia (RGF JMP 73).
Description: Shell small, globular, and low-spired. Spire consisting of three rapidly expanding whorls, separated by a moderately deep suture with no distinct ramp, and accounting for 15% of total shell height (Table 19). Whorl flank convex. Protoconch whorls not preserved. Last whorl large and sub-rounded with slightly rounded base. Aperture oblique, lunate, long, narrow posteriorly and broadly rounded abapically. Columellar folds not completely preserved. Inner lip with a well developed columellar fold close to the base. Outer lip incomplete. Ornament consisting of numerous, equally spaced, well developed spiral ribs, separated by wide interspaces. These interspaces are covered with many axial striae (Fig. 14H, I).
Discussion: Despite the poor preservational state of the columellar folds and incomplete outer lip, the present specimen closely resembles Avellana subincrassata d’orBigny in having a globular shell, numerous equally spaced spiral furrows covered with a dense pattern of axial striae, and a slightly rounded base.
d’orBigny (1842: 133) considered Auricula incrassata mantell (1822: 110, pl. 19, fig. 33) as a synonym of A. incrassata. Kollmann (2002: 55) stated that the specimens of d’orBigny are globular in outline and possess a flat adapical area, whereas A. incrassata (mantell) is more oviform. The internal folds of the latter two species are similar. d’orBigny (1850: 128) correctly changed the species name into A. subincrassata but did not discuss it (see Kollmann 2002).
A. intermedia Kollmann (1976: 191, pl. 7, figs. 63-66) from the Lower Cenomanian of Austria strongly resembles the present species in general outline and ornamentation but differs in being smaller (H = 7.8-10.0, D = 6.0-8.2 mm as opposed to H = 20, D = 16 mm; Table 19), more globose, and in having a different columellar structure (Kollmann 1976: 192). A. hugardiana d’orBigny (1842: 135, pl. 168, figs. 17-19) from the Albian of France differs in being more oviform and in having a strongly convex, thick outer lip, an egg-shaped, large last whorl, and less developed spiral threads.
Occurrence: Albian of France (d’orbigny 1842; kollMann 2005), Austria (Kollmann 2002), Turonian of England (cleevely & MorriS 2002), Lower Cenomanian of the Kosmaj Mountain, central Serbia (this study).
5. Palaeoecological and taphonomic remarks
The identified gastropods come from a lens of reddish coarse-grained bioclastic packstones, which overlies algal grainstones and is, in turn, overlain by sandy grainstones (Fig. 2). The lens is half a meter thick and approximately two meters long and contains a rich and diverse assemblage of gastropods, ammonites, bivalves, echinoids, brachiopods, and corals (142 taxa; see Mitrović-Petrović & andJelković 1991: 583). Among the benthic fauna, gastropods and bivalves
dominate in term of taxa (51% of the total macrofauna) followed by echinoids. Ammonites are also common and represent 37% of the total number of species. Other groups such as brachiopods and corals are comparatively rare. Faunal elements with an originally aragonitic shell (e.g., gastropods and infaunal bivalves) are usually preserved as internal or composite moulds. Originally calcitic taxa (e.g., brachiopods and echinoids), in contrast, are preserved with their shells.
The taxa of the fossiliferous lens belong to several trophic/life habit groups. Epifaunal elements account for 89 per cent (90% of the gastropods, 78% of the echinoids, and all brachiopods). Grazers are common (71% of the gastropods, 33% of the echinoids), followed by carnivores and deposit-feeders. With respect to gastropods, the majority of the identified species (62%; Fig. 16) belong to the families Ampullinidae, Pleurotomariidae, and Tylostomatidae. Pleurotomariids account for 19.5% of the total number of species. Recent representatives are epifaunal mobile forms found mainly at bathyal depths exceeding 200 m (HickMan 1976; HaraSewycH & kiel 2007: 87). For the Cenomanian, kiel & bandel (2004: 122) demonstrated that Pleurotomaria still inhabited rocky-shore environments.
According to Kollmann (1985: 106), nerineids, which represent 5% of the total number of species, are typical shallow-water elements of the Tethyan realm, while pleurotomariids are restricted to the temper-ate realm or may have inhabited greater depth within the Tethyan realm. Recently, cataldo & lazo (2012: 91) interpreted Pleurotomaria gerthi weaver, 1931 to inhabit storm-influenced inner shelf and middle car-bonate ramp environments in the Lower Cretaceous of Argentina.
With respect to echinoids, 56% of the identified echinoid taxa are regular (ayoub-Hannaa et al. 2014). According to Smith (1984, 1995), Smith et al. (1988, 1995) and el qot et al. (2009), all regular echinoids are epifaunal, shallow-water, rocky bottom inhabit-ants wandering as grazers and browsers over the sea floor in search of food. Thus, the taxa discussed so far clearly represent a hard-bottom fauna. In contrast, an-other group of taxa represents a soft-bottom fauna, as indicated by 64% shallow-infaunal bivalves and 44% irregular echinoids (86% of these taxa are suspension-feeders; see Mitrović-Petrović & andJelković 1991: 583). Bivalves include the shallow-infaunal suspen-sion-feeders Anisocardia, Granocardium, Isocardia, and Opis (Mitrović-Petrović & andJelković, 1991). Thus the faunal composition of the lens is a mixture of hard- and soft-substrate elements implying either
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 59
transport of one of the major guilds or else a differen-tiated habitat.
The high species richness of this lens is connect-ed with early transgressive conditions in the study area during Early Cenomanian times (Fig. 17). Dur-ing transgression, little sedimentation took place and reworking was common, leading to mixing of shells from different time planes and environments. FürSicH (1995: 646) pointed out that a low rate of sedimenta-tion can lead to concentration of skeletal elements even when the production of biogenic hardparts is low. Ac-cording to Mitrović-Petrović & andJelković (1991) most of the shells in this assemblage were damaged, disarticulated, fragmented, without preferred orienta-tion or grading, and poorly sorted (fossils representing a wide range size). These taphonomic data suggest a shallow-water setting well above the storm wave-base (storm-wave concentration of FürSicH & oScHMann 1993, fig. 5). The fauna is clearly reworked but did not necessarily suffer significant lateral transport. Steno-haline echinoids, ammonites, and corals indicate fully marine, warm-temperate waters, and a position within the photic zone.
6. Conclusions
A detailed taxonomic description and revision of the Early Cenomanian gastropod fauna from the Grab well section (Kosmaj Mountain) of the Vardar Zone, central Serbia, earlier listed by Mitrović-Petrović & andJelković (1991), yielded 20 taxa belonging to 14 genera and 10 families. The fauna is strongly domi-nated by Ampullinidae and Pleurotomariidae followed by Tylostomatidae.
Three of the species, Anodomaria kosmajae sp. nov., Latiala? callositae sp. nov., and Pseudonerinea jovankae sp. nov., are new, and seven species are recorded from the Lower Cenomanian of Serbia for the first time. Two species are described in open nomenclature.
Palaeoecological, taphonomic, and sedimentologi-cal data suggest that the gastropods and associated fauna lived in a shallow marine environment above storm wave-base and are parautochthonous. Most like-ly, they were concentrated by storm events during a transgressive setting.
Acknowledgements
Great thanks are due to Professor Jovanka Mitrović-Petrović (Belgrade) for the kind loan of her collection. We are also grateful to the editor-in-chief günter ScHweigert, and
reviewers Prof. Dr. SteFano monari (Padova) and Prof. Dr. FranK riedel (Berlin) for their useful comments and remarks that helped to improve the original manuscript. The research was supported by the Ministry of Education, Science and Technological Development of the Republic of Serbia, Project No. 176015 (grant to BR and VR).
References
aBBaSS, h.l. (1963): A monograph on the Egyptian Cretaceous gastropods. – Geological Survey and Mineral Research Department, Monographs of the Geological Museum, Palaeontological Series, 2: 1-146.
albaneSi, c. & buSSon, g. (1974): Gastéropodes du Crétacé Supérieur de l’extrème-sud tunisien et de la région du Tinrhert (Sahara Algérien). – Rivista Italiana di Paleontologia e Stratigrafia, 80: 251-342.
alliSon, e.C. (1955): Middle Cretaceous gastropoda from Punta China, Baja California, Mexico. – Journal of Paleontology, 29: 400-432.
andJelković, M. (1953): Quelques espèces caractéristique d’ammonites et de gastéropodes de l’albien-cénomanien et de sénonien à Kosmaj. – Geološki anali Balkanskoga poluostrva, 21: 125-132 (in Serbian with French summary).
ayoub-Hannaa, w.S. & FürSicH, F.t. (2011): Revision of Cenomanian-Turonian (Upper Cretaceous) gastropods from Egypt. – Zitteliana, (A), 51: 115-152.
ayoub-Hannaa, w.S., radulović, b.v., radulović, v.J., Für-SicH, F.t. & vaSić, n. (2014): Brachiopods and echinoids from the Lower Cenomanian of Koraćica (Kosmaj Moun-tain, central Serbia). – Cretaceous Research, 51: 121-147.
Bandel, K. (1999): On the origin of the carnivorous gastropod group Naticoidea (Mollusca) in the Cretaceous with description of some convergent but unrelated groups. – Greifswalder Geowissenschaftliche Beiträge, 6: 143-175.
Bandel, K. (2000): Some gastropods from the Trichinopoly Group Tamil Nadu, India and their relation to those from the American Gulf Coast. – Memoirs of the Geological Society of India, 46: 65-111.
BarKer, m.J. (1990): The palaeobiology of nerineacean gastropods. – Historical Biology, 3: 249-264.
BlanCKenhorn, m. (1927): Die fossilen Gastropoden und Scaphopoden der Kreide von Syrien-Palästina. – Palaeontographica, 69: 111-186.
boucHet, P. & rocroi, J.P. (2005): Classification nomenclator of gastropod families. – Malacologia, 47: 1-397.
Brongniart, A. (1822): Description géologique des couches des environs de Paris, parmi se trouvent les gypses à ossements. – In: Cuvier, G. (Ed.): Recherches sur les ossements fossiles, 2 (2): 239-648; Paris.
cataldo, c.S. & lazo, d.g. (2012): Redescription of Pleurotomaria gerthi weaver, 1931 (Gastropoda, Vetigastropoda), from the Early Cretaceous of Argentina: New data on its age, associated palaeoenvironments and palaeobiogeographic affinities. – Ameghiniana, 49: 75-95.
cleevely, r.J. & MorriS, n.J. (2002): Introduction to molluscs and bivalves. – In: Smith, A.B. & Batten, D.J. (Eds.): Fossils of the Chalk, revised and enlarged, 161-175; London (Palaeontological Association).
eschweizerbart_xxx

60 W. Ayoub-Hannaa et al.
Collignon, m. (1931): Paléontologie de Madagascar. XVI. La faune du Cénomanien à fossiles pyriteux du nord de Madagascar. – Annales de Paléontologie, 20: 43-104 (1-64).
Collignon, M. (1971): Gastéropodes et Lamellibranches du Sahara. – Annales de Paléontologie (Invertébrés), 57: 143-202.
Coquand, m.h. (1862): Géologie et paléontologie de la région Sud de la province de Constantine. – Mémoires de la Société d’Emulation de la Provence, 2: 5-342.
CoSSmann, m. (1925): Essais de paléochonchologie comparée, 13: 1-345; Paris (Presses Universitaires de France).
Cotteau, g. (1853-1857): Études sur les mollusques fossiles du département de l’Yonne. Premier fascicule comprenant l’introduction et le prodrome. – 1-141; Paris.
Cox, l.r. (1949): On the genotype of Nerinea; with a new subgeneric name Eunerinea. – Proceedings of the Malacological Society of London, 27: 248-250.
Cox, l.r. (1960a): General characteristics of gastropods. – In: moore, r.C. (Ed.): Treatise on Invertebrate Paleontology, Part L, Mollusca 1: L249-L251; New York & Lawrence (Geological Society of America & University of Kansas Press).
Cox, l.r. (1960b): The British Cretaceous Pleurotomariidae. – Bulletin of the British Museum (Natural History), Geology, 4: 388-423.
delpey, g. (1939): Les gastéropodes mésozoiques de la région libanaise. – Notes et Mémoires de la Section d’Études géologiques du Haut-Commissariat de la République Française en Syrie et au Liban, 3: 5-292.
dietriCh, w.o. (1925): Gastropoda mesozoica. Fam. Nerineidae. – Fossilium Catalogus 1, Animalia, 31: 164 pp.; Berlin.
el qot, g.M., FürSicH, F.t., abdel-gawad, g.i. & ayoub-hannaa, w.S. (2009): Taxonomy and palaeoecology of Cenomanian-Turonian (Upper Cretaceous) echinoids from eastern Sinai, Egypt. – Beringeria, 40: 55-98.
ForBeS, e. (1846): Report on the fossil Invertebrata from Southern India, collected by Mr. Kaye & Mr. CunliFFe. – Transactions of the Geological Society, London, (2) 7: 97-174.
FürSicH, F.t. (1995): Shell concentrations. – Eclogae Geologicae Helvetiae, 88: 643-655.
FürSicH, F.t. & oScHMann, W. (1993): Shell beds as tools in basin analysis: the Jurassic of Kachchh, Western India. – Journal of the Geological Society of London, 150: 169-185.
gaSpard, D. (1997): Distribution and recognition of phases in Aptian-Turonian (Cretaceous) brachiopod development in NW Europe. – Geologica Carpathica, 48: 145-161.
greCo, B. (1916): Fauna cretacea dell’Egitto raccolta dal Figari Bey. Parte seconda. Gasteropoda. – Palaeontographica Italica, 22: 103-170.
HaraSewycH, M.g. & kiel, S. (2007): Upper Jurassic Pleurotomariidae (Gastropoda) from southwestern Madagascar. – The Nautilus, 121 (2): 76-89.
HaraSewycH, M.g., oleinik, a. & zinSMeiSter, w. (2009): The Cretaceous and Paleocene pleurotomariid (Gastropoda: Vetigastropoda) fauna of Seymour Island, Antarctica. – Journal of Paleontology, 83: 750-766.
hiCKman, C.S. (1976): Pleurotomaria (Archaeogastropoda) in the Eocene of the northeastern Pacific: a review of
Cenozoic biogeography and ecology of the genus. – Journal of Paleontology, 50: 1090-1102.
Jaitly, a.k., Szabô, J. & FürSicH, F.t. (2000): Contributions to the Jurassic of Kachchh, western India. VII. The gastropod fauna. Part I. Pleurotomarioidea, Fissurelloidea, Trochoidea and Eucycloidea. – Beringeria, 27: 31-61.
Jovanović, r. (1938): Beitrag zur Geologie des Kosmaj. – Vesnik Geološkog instituta kraljevine Jugoslavije, 6: 135-154 (in Serbian with German summary).
KaBat, a.R. (1991): The classification of the Naticidae (Mollusca: Gastropoda): review and analysis of the supraspecific taxa. – Bulletin of the Museum Comparative Zoology, 152: 417-449.
KaSe, t. (1984): Early Cretaceous marine and brackish-water Gastropoda from Japan. – 263 pp.; Tokyo (National Science Museum).
Kiel, S. (2006): New and little-known gastropods from the Albian of the Mahajanga Basin, northwestern Madagascar. – Journal of Paleontology, 80: 455-476.
kiel, S. & bandel, k. (2001): Archaeogastropoda from the Campanian of Torallola, northern Spain: Trochidae. – Acta Geologica Polonica, 51: 137-154.
kiel, S. & bandel, k. (2002): About some aporrhaid and strombid gastropods from the Late Cretaceous. – Paläontologische Zeitschrift, 76: 83-97.
kiel, S. & bandel, k. (2003): New taxonomic data for the gastropod fauna of the Umzamba Formation (Santonian-Campanian, South Africa) based on newly collected material. – Cretaceous Research, 24: 449-475.
kiel, S. & bandel, k. (2004): The Cenomanian Gastropoda of the Kassenberg quarry in Mülheim (Germany, Late Cretaceous). – Paläontologische Zeitschrift, 78: 103-126.
Kollmann, h.a. (1976): Gastropoden aus den Losensteiner Schichten der Umgebung von Losenstein (Oberösterreich). 1. Teil: Euthyneura und Prosobranchia 1 (Neogastropoda). – Annalen des Naturhistorischen Museums Wien, 80: 163-206.
Kollmann, h.a. (1978): Gastropoden aus Losensteiner Schichten der Umgebung von Losenstein (Oberösterreich). 2. Teil: Naticidae, Colombellinidae, Aporrhaidae, Ceritellidae, Epitoniidae (Mesogastropoda). – Annalen des Naturhistorischen Museums Wien, 81: 173-201.
Kollmann, h.a. (1985): Upper Cretaceous gastropods from excavations for the highways A10 (Charente, France). – Cretaceous Research, 6: 85-111.
Kollmann, h.a. (2002): Gastropods from the Lower Cretaceous of Vorarlberg, Austria. A systematic review. – Annalen des Naturhistorischen Museums in Wien, (A), 103: 23-73.
Kollmann, h.a. (2005): Gastropodes Crétacés. – In: FiSCher, J.C. (Ed.): Révision critique de la Paléontologie française d’alCide d’orBigny (incluant la réédition de l’original), 3: 1-239; Leiden (Backhuys).
leymerie, m.a. (1842): Suite du mémoire sur le terrain Crétacé du département de l’Aube. 2ème partie: Partie paléontologique. – Mémoires de la Société géologique de France, 5: 1-34.
mantell, g. (1822): The fossils of the South Downs; Illustrations of the Geology of Sussex, xvii + 327 pp.; London.
maury, C.J. (1937): O Cretaceo de Sergipe. – Serviço Geolo-gico e Mineralogico do Brasil, Monographia, 11: 1-283.
eschweizerbart_xxx

Gastropods from the Lower Cenomanian of Koraćica 61
meeK, F.B. (1876): A report on the invertebrate Cretaceous and Tertiary fossils of the upper Missouri Country. – Report of the United States Geological Survey of the Territories, Washington, 9: 1-629.
Mitrović-Petrović, J. & andJelković, M. (1991): Biostratigraphic and paleoecologic characteristics of Alb-Cenomanian sediments of Kosmaj, Serbia. – Bulletin of the Geological Society of Greece, 25: 581-590.
Monari, S. & gatto, r. (2013): Pleurotomaria deFranCe, 1826 (Gastropoda, Mollusca) from the Lower Bajocian (Middle Jurassic) sediments of Luxembourg, with considerations on its systematics, evolution and palaeobiogeographical history. – Palaeontology, 56: 751-781.
Monari, S. & gatto, r. (2014): The genus Leptomaria e. eudeS-deSlongChampS, 1864 (Gastropoda, Pleurotomariidae) from the Early Bajocian of Luxembourg: systematics and paleobiogeography. – Historical Biology, 26: 810-826.
orBigny, a. de (1842-1843): Paléontologie française. Déscription zoologique et géologique de tous les animaux mollusques et rayonnés fossiles de France. Terrains Crétacés. T. 2. Ptéro- et Gastéropodes, 1-224 (1842), 225-456 (1843); Paris (Masson).
orBigny, a. de (1850): Prodrome de Paléontologie stratigraphique universelle des animaux mollusques et rayonnés, 2: 1-427; Paris (Masson).
paCaud, J.m. (2004): Révision des mollusques du Danien (Paléocène inférieur) du Bassin de Paris. 1. Gastropoda: Patellogastropoda et Vetigastropoda (pro parte). – Geodiversitas, 26: 577-629.
Peron, a. (1889-1891): Déscription des mollusques fossiles des terrains crétacés de la région sud des Hauts-Plateaux de la Tunisie recueillis en 1885 et 1886 par thomaS, m. p. – In: Exploration Scientifique de la Tunisie. – 405 pp.; Paris (Imprimerie Nationale).
Pictet, F.J. & roux, w. (1847): Déscription des mollusques fossiles qui se trouvent dans les Grés Verts des environs de Genève. – Mémoires de la Société de physique et d’histoire naturelle de Genève, 2: 257-412.
PoPenoe, w.P., Saul, l.r. & SuSuki, t. (1987): Gyrodiform gastropods from the Pacific coast Cretaceous and Paleocene. – Journal of Paleontology, 61: 70-100.
robertSon, a., karaMata, S. & Šarić, K. (2009): Overview of ophiolites and related units in the Late Palaeozoic–Early Cenozoic magmatic and tectonic development of Tethys in the northern part of the Balkan region. – Lithos, 108: 1-36.
roemer, F.a. (1841): Die Versteinerungen des norddeutschen Kreidegebirges, 2: 49-145; Hannover (Hahn).
Sharpe, d. (1849): On Tylostoma, a proposed genus of gasteropodous mollusks. – Quarterly Journal of the Geological Society, London, 5: 376-380.
SHuMard, b.F. (1853-1854): Paleontology. Description of the species of Carboniferous and Cretaceous fossils collected. – In: marCy, r.B. (Ed.): Exploration of the Red River of Louisiana, in the year 1852 with reports on the natural history of the country. – 32nd Congress, 2nd Session. Senate Executive Documents, 8: 54; Washington.
Smith, a.B. (1984): Echinoid palaeobiology. – 190 pp.; London (George Allen & Unwin).
Smith, a.B. (1995): Late Campanian-Maastrichtian echinoids from the United Arab Emirates-Oman border region. – Bulletin of Natural History Museum, 51: 121-240.
SMitH, a.b., Paul, c.r.c., gale, a.S. & donovan, S.k. (1988): Cenomanian and Lower Turonian echinoderms from Wilmington, south-east Devon, England. – Bulletin of the British Museum (Natural History), 42: 1-245.
SMitH, a.b., MorriS, n.J., gale, a.S. & roSen, b.r. (1995): Late Cretaceous (Maastrichtian) echinoid-mollusc-coral assemblages and palaeoenvironments from a Tethyan carbonate platform succession, northern Oman Mountains. – Palaeogeography, Palaeoclimatology, Palaeoecology, 119: 155-168.
Sirna, g. (1995): The nerineids: taxonomy, stratigraphy and palaeoecology with particular references to Italian examples. – Geologica Romana, 31: 285-305.
Sohl, n.F. (1960): Archaeogastropoda, Mesogastropoda and stratigraphy of the Ripley, Owl Creek, and Prairie Bluff formations. – United States Geological Survey, Professional Papers, (A), 133: 1-151.
Sohl, n.F. (1967): Upper Cretaceous gastropods from the Pierre Shale at Red Bird, Wyoming. – United States, Geological Survey, Professional Papers, (B), 393: 1-43.
SowerBy, J. (continued by J. de C. SowerBy) (1812-1846): The mineral conchology of Great Britain, 7 vols. – 1812-1822, J. SowerBy (pl. 1-383), 1823-1846, J. de c. Sowerby (pl. 384-648); London (meredith).
SowerBy, J. (1835): In: Sedgwick, a. & MurcHiSon, R.A. (Eds.): A sketch of the structure of the Eastern Alps; with sections through the newer formations on the northern flank of the chain, and through the Tertiary deposits of Styria. – Transactions of the Geological Society of London, 2: 301-420.
SowerBy, J. de C. (1836): Descriptive notes respecting the shells figured in Plates XI to XXIII. – In: Fitton, w.h. (Ed.): Observations on some of the strata between the Chalk and the Oxford Oolite, in the south-east of England. – Transactions of the Geological Society of London, Second Series, 4: 335-349.
SquireS, r.l. & Saul, l.r. (2004): Uncommon Cretaceous naticiform gastropods from the Pacific Slope of North America. – The Veliger, 47: 21-37.
Stanton, t.w. (1947): Studies of some Comanche pelecypods and gastropods. – United States Geological Survey, Professional Papers, 211: 1-256.
Stilwell, J.d. & HenderSon, r.a. (2002): Description and paleobiogeographic significance of a rare Cenomanian molluscan faunule from Bathurst Island, Northern Australia. – Journal of Paleontology, 76: 447-471.
StoliCzKa, F. (1868): Cretaceous fauna of southern India. Vol. 2, The Gastropoda. – Memoirs of the Geological Survey of India, Palaeontologica Indica, 5 (2): 1-498.
wade, B. (1926): The fauna of the Ripley Formation on Coon Greek, Tennessee. – United States Geological Survey, Professional Papers, 137: 1-272.
wenz, w. (1938–1944). Teil 1: Allgemeiner Teil und Prosobranchia. – In: SChindewolF, o.h. (Ed.): Handbuch der Paläozoologie, 6, Gastropoda. – 1639 pp.; Berlin (Borntraeger).
white, C.a. (1887): Contribuições à paleontologia do Brazil. – Archivos do Museu Nacional do Rio de Janeiro, 7: 1-273.
wieCzoreK, J. (1979): Upper Jurassic nerineacean gastropods from the Holy Cross Mts (Poland). – Acta Palaeontologica Polonica, 24: 299-350.
eschweizerbart_xxx

62 W. Ayoub-Hannaa et al.
Manuscript received: August 26th, 2014Revised version accepted by the Stuttgart editor: December 16th, 2014
Addresses of the authors:
wagiH ayoub-Hannaa, Department of Geology, Faculty of Sciences, Minufiya University, El-Minufiya, Shibin El Kom, Egypt. Current address: GeoZentrum Nordbayern, Fachgruppe Paläoumwelt der Friedrich-Alexander-Universität Erlangen-Nürnberg, Loewenichstr. 28, D-91054 Erlangen, Germany;e-mail: [email protected] v. radulović, vladan J. radulović, Department of Palaeontology, Faculty of Mining and Geology, University of Belgrade, Kamenička 6, 11000 Belgrade, Serbia; e-mails: [email protected], [email protected] t. FürSicH, GeoZentrum Nordbayern, Fachgruppe Paläoumwelt der Friedrich-Alexander-Universität Erlangen-Nürnberg, Loewenichstr. 28, D-91054 Erlangen, Germany; e-mail: [email protected]
eschweizerbart_xxx





























