Embed Size (px)
Citation preview

Fresh insights into long-term changes in flora,
vegetation, land use and soil erosion in the karstic
environment of the Burren, western Ireland
Ingo Feeser* and Michael O’Connell
Palaeoenvironmental Research Unit, Department of Botany, National University of Ireland Galway, Galway, Ireland
Summary
1. The study focuses on species-rich, upland, heathy vegetation with arctic-alpine floristic affinities
and Sesleria grasslands in the karstic Burren region, western Ireland. The investigations aimed at
reconstructing the long-term development of these high conservation-value communities and the
role of farming in their formation and long-term survival.
2. The methods used included pollen analysis and 14C-dating of short monoliths and investigation
of grykes (fissures in karstic limestone) for evidence of soil erosion. Special attention was paid to
fossil, coprophilous fungal spores as indicators of local grazing. The strong local character of the
pollen records facilitated identification of inter-site differences as well as regional patterns. It is
shown that open pine woodland characterized the Cappanawalla uplands between c. 1500 BC and
500 BC. It is proposed that such woodlands, with floristic affinities to Scandinavian open pine
woodlands on calcareous soils, provided a suitable environment for the present-day, open heath
vegetation with species such as Dryas octopetala, Arctostaphylos uva-ursi, Geranium sanguineum
andEmpetrum nigrum.
3. Burning of vegetation as a management tool was important in the uplands over most of the last
two millennia. Firing seems to have ceased with the onset of more intensive grazing regimes in the
18th century.
4. Synthesis. Upland palaeoarchives, derived from shallow peaty deposits, show that the upland
Burren supported mainly plagioclimax Corylus-dominated woody vegetation and grasslands from
c. 1500 BC (beginning of present record), until possibly as late as the 17th century AD. In the
uplands of the north-western Burren, open, species-rich pinewoods with hazel dominated. The
northern-arctic elements in the present-day upland flora survived clearances, involving initially
Pinus sylvestris (c. 500 BC) and subsequently Corylus avellana (c. AD 1600). Colluvial material
retrieved from grykes supports the idea of considerable soil loss occurring as late as the first and
early 2nd millennium AD. The investigations highlight the potential of upland palaeoarchives,
consisting of short sequences, for elucidating vegetation and land-use dynamics in karstic environ-
ments such as the Burren.
Key-words: biogeography, coprophilous fungal spores, environmental change, karst, land use,
late Holocene, non-pollen palynomorphs, pollen analysis, soil erosion, vegetation dynamics
Introduction
The Burren, a mainly upland region extending over some
300 km2 in north-west County Clare, western Ireland (Fig. 1),
is internationally recognized for its karstic geology, an excep-
tional flora that includes arctic-alpine and northern elements
often growing in great abundance and in close proximity to
southern elements, and a rich archaeology that testifies to
settlement and farming extending back to the early Neolithic
(c. 4000 cal. BC; dates are given in calibrated years BC ⁄AD,
except 14C dates which are cited as BP, i.e. before AD 1950).
Research by Irish and visiting scientists has elucidated several
aspects of this region including the geology and soils (Finch
1971; Pracht et al. 2004), vegetation (Ivimey-Cook & Proctor
1966; Kirby 1981) and environmental plant physiology
(Osborne et al. 2003). More recently, the role of management
— essentially extensive farming as influenced by the EU Com-
mon Agricultural Policy (CAP) and specific instruments of
CAP such as the Rural Environment Protection Scheme*Correspondence author. E-mail: [email protected]
Journal of Ecology 2009, 97, 1083–1100 doi: 10.1111/j.1365-2745.2009.01533.x
� 2009 The Authors. Journal compilation � 2009 British Ecological Society

(REPS) — has received considerable attention (Sheehy Skeff-
ington & Gormally 2001; Dunford 2002). Much of this infor-
mation has been collated and synthesized in general
overviews (D’Arcy & Hayward 1992; O’Connell & Korff
2001; Jones 2004) and detailed accounts (e.g. Webb & Scan-
nell 1983; Osborne & Jones 2003). Despite these advances,
however, the impression remains of a ‘‘false sense of security’’
provided by the wealth of ‘‘descriptive biological detail’’
whereas ‘‘the truth is we don’t … understand the factors that
have shaped the present communities associated with the Bur-
ren’’ (Osborne & Jeffrey 2003, p. 107).
The present paper is conceived as a contribution to
improved understanding of the Burren and, specifically, the
plant communities and soils of the Burren uplands (altitude c.
300 m a.s.l.). These areas are home to some of the most dis-
tinctive plant communities and habitats, such as the Arcto-
staphylos uva-ursi—Dryas octopetala heaths (cf. Ivimey-Cook
& Proctor 1966) on the Cappanawalla–Gleninagh–Black
Head uplands in the north-western Burren and the Calluna
heaths and species-rich limestone grasslands on Gortaclare
and Slieve Rua–Mullach Mor uplands in the eastern Burren.
The particular aim of these investigations is the recon-
struction of environmental change in the areas referred to,
and, specifically, to determine if farming played a key role
in the establishment and long-term maintenance of these
species-rich communities or if, on the other hand, these com-
munities are best regarded as representing ‘‘a true climax ...
within a certain range of altitude and exposure’’, as proposed
by Ivimey-Cook & Proctor (1966, p. 251) for the Arctostaph-
ylos uva-ursi–Dryas octopetala heaths. Attention is also
focussed on possible widespread erosion and hence large-
scale loss of soil as a result of early farming. Techniques
employed include pollen analysis and AMS 14C dating and
probing of grykes, i.e. fissures in the karstic limestone, for
evidence of former soils. Particular attention is given to
identification of critical pollen taxa and also non-pollen
palynomorphs (NPP) including coprophilous fungal spores
(CFS) which are particularly useful for the reconstruction of
grazing activity (cf. van Geel et al. 2003; Blackford & Innes
2006; Davis & Shafer 2006; Feeser & O’Connell 2010).
Site descriptions
CAPPANAWALLA
The plateau-like upland of Cappanawalla (c. 300 m a.s.l.;
Fig. 1) is characterized by extensive limestone pavement with
deep grykes and patches of shallow rendzina-like soil. Shallow
Fig.1. Contoured maps showing the extent of the Burren region (eastern lowlands excluded) and details of the three study areas, Cappanawalla
(CPW), Gortaclare (GTC) and Sliabh Rua (RUA) (the squares are 10 · 10 km). Location of sampling sites, including Lios LairthınMor (profile
LLM II; Jelicic &O’Connell 1992), is indicated. Archaeological features are after Robinson (1999) and OSDiscovery Seriesmaps, with additions
andminormodifications.
1084 I. Feeser & M. O’Connell
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

depressions with peat deposits, mainly developed on minero-
genic, probably drift-derived soils several decimetres thick, are
a feature of the area.
Cappanawalla is one of five upland areas in the Burren of
exceptionally high floristic interest (Webb & Scannell 1983).
The heath communities are definedmainly byCalluna vulgaris,
Dryas octopetala, Arctostaphylos uva-ursi and Empetrum
nigrum (higher plant nomenclature follows Stace 1997). Erica
cinerea, Succisa pratensis, Carex panicea and C. binervis are
common; other wet heath and bog species include Narthecium
ossifragum,Trichophorum cespitosum andParnassia palustris.
Species associated with the upland limestone, i.e. pavement,
grykes and small patches of shallow soils, include Sesleria
caerulea, Thymus polytrichus, Achillea millefolium, Phyllitis
scolopendrium, Polystichum setiferum, Cystopteris fragilis and
the neophyte Mycelis muralis. Like most Burren uplands, the
Cappanawalla plateau is extensively grazed, mainly by cattle,
in winter as part of a transhumance-type practice known as
winterage (Dunford 2002).
On the slopes of Cappanawalla, the vegetation changes
with altitude, bedrock and soil characteristics, and land use.
The eastern slopes, which have little soil cover, harbour the
more floristically interesting vegetation. Dryas octopetala is
locally abundant above c. 100 m a.s.l. and Empetrum nigrum
occurs regularly above c. 200 m a.s.l. Calluna-dominated
heath with Molinia caerulea is locally common along the
mid-slopes and Filipendula ulmaria is frequent. At lower
elevation, above the intensively managed valley-floor
pastures, open scrubby vegetation with Corylus avellana,
Crataegus monogyna, Prunus spinosa, Ilex aquifolium, Rosa
pimpinellifolia is common, while in open rocky parts Solidago
virgaurea, Campanula rotundifolia, Geranium sanguineum and
Pteridium aquilinum are common.
GORTACLARE
Gortaclare, together with Slievecarran, forms the most exten-
sive upland in the eastern Burren (Fig. 1). Calluna heath and
short-sward Sesleria-rich limestone grassland characterizes the
southern part of the uplands. Heath is common here, its devel-
opment being favoured by the frequent outcropping of dark
grey chert that forms isolated nodules, ridges and
semi-continuous beds. Apart from the occasional, wind-bent
specimen of C. monogyna, the uplands are bare of tall woody
vegetation. At lower elevations, e.g. the eastern side of
Slievecarran and the Glen of Clab, Corylus scrub, with
Fraxinus excelsior, is extensive and largely dominant. The
uplands, like at Cappanawalla, are important for winterage
(Keane & Sheehy Skeffington 1995). The valley floors, in con-
trast to the uplands, have substantial, drift-derived soil cover
that supports rather intensive, pastoral-based farming.
SLIABH RUA
Sliabh Rua lies at the centre of the Knockanes–Sliabh Rua–
Mullach Mor uplands (Fig. 1) which, together with the sur-
rounding lowlands, constitute the Burren National Park. The
soils, which are very patchy, show considerable small-scale var-
iation in mineralogical composition and glacial origin, but
apparently do not include loess deposits (Moles & Moles
2002), as has been postulated (Moles et al. 1995; Jeffrey 2003).
The uplands carry mainly Sesleria-dominated, limestone
grassland on shallow soils intermingled with bare limestone
pavement often with well-defined grykes. There are extensive
patches ofCalluna-dominated heath, especially between Sliabh
Rua andMullachMor and on the side of Knockanes. Cultiva-
tion ridges, indicative of arable farming (probably potato
growing), occur on drift on the north-eastern flank ofMullach
Mor (Moles &Moles 2002).
ARCHAEOLOGICAL F IELD EVIDENCE FOR PAST HUMAN
ACTIV ITY
A rich archaeological field assemblage points to substantial
human activity from Neolithic times onwards (last c.
6000 years) in the Burren, but, surprisingly, little has been
published on the upland areas that were investigated here
(Fig. 1). Features noted on Cappanawalla include a ringfort
(diameter c. 20 m), a cairn and several stone-wall enclosures
including possible hut sites. Several low walls, including
so-called mound, tumble and slab walls (possibly of Bronze
Age, Iron Age, and late Iron Age or mediaeval period, respec-
tively), are present, superimposed on which are high, so-called
single walls that are probably nomore than a few centuries old
(dating of walls after Plunkett Dillon, 1985, and Jones, 2004).
Small U-shaped stone structures, presumably of relatively
recent origin and used for management of sheep and ⁄or goatsand possibly also for drying sods and cow pats as a fuel source
(Dunford 2001), are common. Stone walls, c. 5–10 m long and
about 20 in all, serve as windbreaks for cattle on the exposed
hilltop. The surrounding lowlands, on the other hand, are
much richer in archaeological features, indicative of a substan-
tial human presence from the Bronze Age onwards and espe-
cially during themediaeval period (Fig. 1).
Field evidence for past farming on Gortaclare consists
mainly of stone walls. Conspicuous single walls and also
mound, tumble and slab walls suggest active landmanagement
extending back to at least the Bronze Age. There is the occa-
sional stone-wall windbreak, presumably of rather recent ori-
gin (Fig. 2). Cairns, including a particularly large and
prominent cairn on Slievecarran, suggest substantial pre-his-
toric human presence (cf. also Turlough Hill where there is a
large enclosure, a very substantial cairn and c. 150 hut sites;
undated but possiblyNeolithic).
In the Knockanes–Sliabh Rua–Mullach Mor uplands, the
main archaeological features are cairns on Mullach Mor and
Knockanes (Fig. 1).
Materials and methods
FIELDWORK
Cappanawalla.A small depression, c. 5 · 4 m, on a shelf to the south
of the highest part of the plateau, was selected for sampling (Fig. 1;
Long-term environmental change in the Burren 1085
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

Table 1). The monolith, CPW1, was taken in August 2006 where the
deepest sediments were noted using a gouge corer (Fig. 2). Common
species at the sampling site included Calluna vulgaris (dominant),
Erica cinerea, Succisa pratensis, Carex spp., Lathyrus linifolius and
Potentilla erecta.
Gortaclare. The main sampling location, GTC7, lies towards the
southern end of the plateau (Fig. 1; Table 1) where there is a patch-
work of partially bare limestone, grassland and peat-covered chert
overlying limestone. Species of note include Dryas octopetala and
occasionally Rosa pimpinellifolia and Gentiana verna. Heathy vege-
tation with Calluna and a well-developed bryophyte layer domi-
nates where peat forms the substrate. The monolith, GTC7,
was taken in May 2006 from peat on a pronounced chert ridge,
c. 10 m long and elevated c. 50 cm above the surrounding lime-
stone grassland (Fig. 2).
On a c. 40-m wide terrace — referred to as the Gortaclare terrace
— that lies c. 5 m below the main plateau on the north-western side,
sediments were found in grykes (Fig. 1, GTC1–5). Here a c. 5-m
wide strip of jointed limestone pavement with patchy grassy cover
formed a zone, distinct from otherwise densely vegetated areas
(grassland and Calluna-dominated heath) with less jointing and
fewer grykes.
Sliabh Rua. In the Knockanes–Sliabh Rua–Mullach Mor uplands,
grykes and heath-covered surfaces were probed using a gouge corer
and spade. Potentially suitable sites were identified only in the saddle
between Sliabh Rua and Mullach Mor where Calluna on shallow
peat dominates over a relatively extensive area. Otherwise, the area
is characterized by jointed limestone pavement with patchy grass-
land and heath. The peat deposits, resting on up to several-decimetre-
thick minerogenic sediments, proved to be very shallow (maximum
Fig.2. Photographs of monoliths and sam-
pling sites. At top, monolith Cappanawalla
(CPW1) (left) and removing the monolith
(right); limestone grassland ⁄ heath domi-
nates in the foreground and the background
consists mainly of bare limestone pavement
(view to SW; 21 August 2006). At bottom,
monolithGortaclare (GTC7) (left; scale divi-
sions = 2 cm), and view of sampling area
(right) with Calluna-dominated heath and
stone-wall windbreak in the background (12
May 2006).
Table 1. Geographical location and other details of sites investigated
Location and
monolith ⁄ core number Latitude ⁄ longitudeAltitude
(m a.s.l.) Comments
Cappanawalla
(CPW1)
53�7.062¢ N, 9�11.812¢ W 296 Monolith from a Calluna-dominated heath
on shallow peat that rested on a silty-sandy soil
Gortaclare
(GTC1 and GTC5)
53�4.931¢ N, 9�2.111¢ W 285 Cores GTC1–5 from grykes on terrace below main
plateau (co-ordinates relate to the end points,
GTC1 and GTC5). Calluna was common at GTC5
53�4.945¢ N, 9�2.041¢ W
Gortaclare
(GTC7)
53�4.646¢ N, 9�2.651¢ W 272 Monolith from peat on cherty limestone
RUA1–3 53�0.884¢ N, 8�59.672¢ W 167 Cores from grykes in grassland ⁄ heath;co-ordinates given for core RUA2
RUA4 53�0.881¢ N, 8�59.690¢ W 170 Shallow peat deposit that rested on silty-sandy soil;
examined only in field
1086 I. Feeser & M. O’Connell
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

depth noted: 18 cm in core RUA4) and were not investigated in
detail. Corings of grykes (RUA1–3; gryke depths 180–200 cm)
yielded c. 70 cm of minerogenic sediment with charcoal flecks near
the base.
LABORATORY INVESTIGATIONS
Samples of 2 cm3 from 1-cm thick slices were taken from monoliths
CPW1 and GTC7 for pollen analysis. Lycopodium clavatum spores,
in tablet form, were added to facilitate calculation of pollen concen-
trations (Stockmarr 1971; tablets supplied by Department of Geol-
ogy, Lund University, Sweden). Standard treatments were given that
involved use of KOH, HCl and HF, and acetolysis (cf. Moore et al.
1991). At the end of the preparation procedure, the pellets were sieved
through a 5-lm mesh sieve in an ultrasonic water bath to remove
unwanted fine particles. The samples were mounted in glycerol and
counted using a Leica DM 4000B microscope (Leica Microsystems,
Wetzlar, Germany) at ·400magnification; phase contrast ·1000mag-
nification was used in the case of critical grains.
Counting, data calculations and plotting of pollen diagrams were
carried out using CountPol 3.2, a java program by I. Feeser. A
pollen sum (PS) of about 1000 terrestrial pollen grains, excluding
heath taxa, was generally achieved. For spectra with exceptionally
large numbers of heath taxa or exceedingly poor pollen concentra-
tion, a lower PS was accepted. Percentage representation was calcu-
lated on the basis of total terrestrial pollen (TTP, including fern
spores), but excluding heath taxa which are expressed relative to
TTP.
Pollen and spore identification follows mainly Moore et al.
(1991) and Beug (2004). As regards cereal-type pollen, the pollen
and annulus-size criteria of Beug (2004) were followed, but modi-
fied as follows. Four pollen-size categories were distinguished (max-
imum length cited), i.e. 37–39 lm, 40–44 lm, 45–49 lm and
>50 lm. Secale pollen were noted separately. The smallest size cat-
egory was subsequently included in the Poaceae (undifferentiated)
because of the high probability that at least some, and probably
several, of the grains were from non-cultivated grasses (see Inter-
pretation of data). Ericaceous pollen were distinguished using
Moore et al. (1991) and modern reference material. Non-pollen
palynomorphs (NPP), i.e. microfossils other than pollen, were
identified using published photomicrographs and descriptions (e.g.
van Geel (1978); Pals et al. (1980); Bakker & van Smeerdijk (1982);
van der Wiel (1982); van Geel et al. (1989, 2003)). Conspicuous
NPP that were regularly observed but not identified with certainty
were assigned a type number, e.g. Type BRN1 (BRN to indicate
Burren).
Material retained after initial sieving of the pollen samples
(>100 lm) was scanned under a stereo-microscope for macrofossil
remains and other matter including charcoal fragments (referred to as
macro-charcoal as distinct from micro-charcoal recorded on the pol-
len slides). Abundances were noted using a four-point scale (rare, +;
occasional, 1; frequent, 2; abundant, 3).
To obtain material suitable for AMS 14C dating, 1-cm thick slices
were taken from the monoliths, sieved and washed with distilled
water, and scanned for macrofossils. As far as possible, terrestrial
macrofossils of known origin were submitted for AMS 14C dating.
Loss-on-ignition at 550 �C (LOI) was determined using c. 2-cm3
samples taken from 1-cm thick slices. Procedures recommended by
Heiri et al. (2001) were adhered to.
In addition to the above investigations, charcoal fragments (at
most a few millimetres long, i.e. too small to be identified) were
picked from basal sediments retrieved from grykes on Gortaclare
and Sliabh Rua (cores GTC1 and RUA2 and 3) and submitted
for dating.
Results
DATA PRESENTATION
Stratigraphical descriptions of the monoliths are presented in
Table 2. Photographs showing monoliths and sampling in the
field are provided in Fig. 2. Pollen and NPP curves, LOI and
macrofossil (pollen sievings) data relating to profiles CPW1
and GTC7 are presented in Figs 3 and 4. Local pollen assem-
blage zone (PAZ) boundaries are indicated where at least one,
and usually several, pollen curves show substantial changes.
Results from constrained incremental sum of squares cluster-
ing (CONISS), carried out using ZONE (ver. 1.2; program by
S. Juggins), supported the zone boundaries as distinguished by
eye. An overview of the CFS and indeterminate NPP is pre-
sented in Table 3 and photomicrographs are presented in
Fig. 5.
Results from the 14C dating are given in Table 4. The 14C
dates were calibrated using the program OxCal 4.0 (Ramsey
2001) and the IntCal04 calibration curve (Reimer et al. 2004).
Table 2. Stratigraphical descriptions for monoliths CPW1
(Cappanawalla) andGTC7 (Gortaclare)
Depth (cm) Description
Monolith CPW1
0–6 Poorly decomposed peat with many
roots. Brownish black (7.5 YR 2 ⁄ 2)*6–22 Well-decomposed peat with fibres.
Dark reddish brown (5 YR 3 ⁄ 2)22–29 Black organic-rich clayey silt with
few fibres. Black (7.5 YR 2 ⁄ 1)29–32 Silty clay with c. 1% small stones
up to 3 mm in diameter. Brownish
black (7.5 YR 3 ⁄ 1)32–36 Silty clay. Brownish grey (5 YR 5 ⁄ 1)
with greyish brown (7.5 YR 4 ⁄ 2) toorange (7.5 YR 6 ⁄ 8) mottling
36–45 Clayey silt. Orange (7.5 YR 6 ⁄ 8)
Monolith GTC7
0–15 Dark brown, well decomposed peat
with roots (brownish black, 7.5 YR 2 ⁄ 2)15–20 Poorly decomposed peat with moss
remains and a minerogenic component
(dark reddish brown, 5 YR 3 ⁄ 2)20–35 Dark brown highly decomposed peat
with some rootlets (brownish
black, 7.5 YR 2 ⁄ 2)35–40 Dark grey brown peaty silt; more
minerogenic and sandy towards the bottom
(brownish black, 7.5 YR 3 ⁄ 1)40–42 Sandy minerogenic deposit with a substantial
organic component; small stones
(brownish black, 7.5 YR 3 ⁄ 1)
*Colour chart by Oyama & Takehara (1970) was used for assign-
ing colours.
Long-term environmental change in the Burren 1087
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100
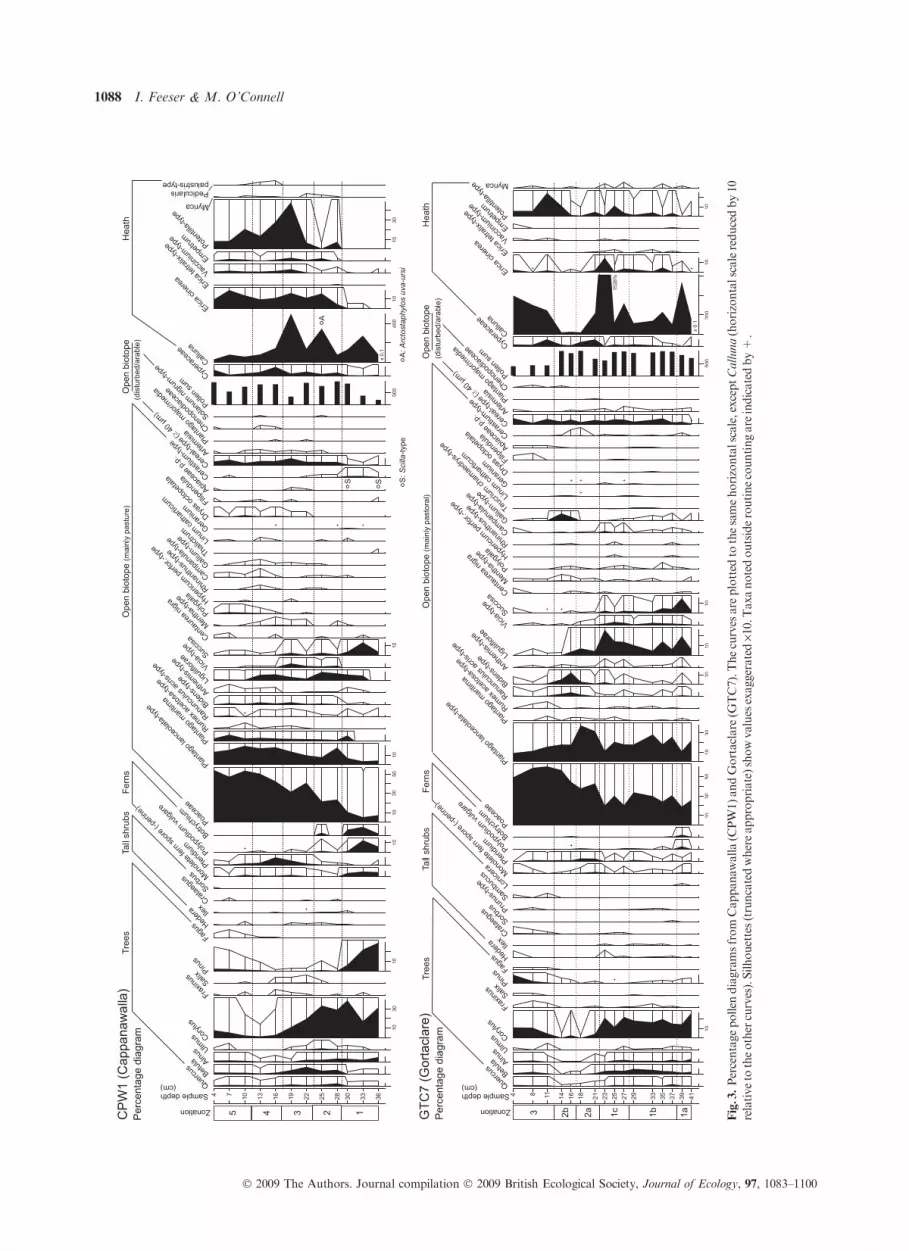
Fig.3.Percentagepollen
diagramsfrom
Cappanawalla(C
PW1)an
dGortaclare
(GTC7).Thecurves
are
plotted
tothesamehorizontalscale,exceptCalluna(horizontalscalereducedby10
relative
totheother
curves).Silhouettes(truncatedwhereappropriate)showvaluesexaggerated
·10.T
axanotedoutsideroutinecountingare
indicatedby+.
1088 I. Feeser & M. O’Connell
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

Fig.4.Plotsofthefollowingdata,p
rofilesCappan
awalla(C
PW1)andGortaclare
(GTC7):non-pollen
palynomorphs(N
PPs)andmicro-charcoalcurves
(percentagevalues),LOI,
14Cdates;m
a-
crofossils(abundance),andconcentrationandpollen
accumulationrate(PAR)curves.S
ilhouettesand‘+
’values
havesameconnotationsasinFig.3.
Long-term environmental change in the Burren 1089
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

INTERPRETATION OF DATA
Interpretation of pollen analytical data is seldom straightfor-
ward and this is particularly true when the material under
investigation derives from sources that are rather unconven-
tional, as in the present instance. Estimating the pollen source
area is crucial, i.e. the relative contribution of pollen of local,
regional and extra-regional or long-distance origin (Jacobson
& Bradshaw 1981; Court-Picon et al. 2006). In this instance,
the local component is regarded as deriving from the upland
vegetation and, in particular, the area close to the sampling
site. As regards the regional component, its contribution is
expected to be strongly influenced by local conditions such as
wind speeds and direction, and dispersal capacity, which for
non-arboreal pollen (NAP) generally and especially for sev-
eral critical Burren taxa is low. In the case of percentage
curves, the inter-dependence of the curves must also be borne
in mind. In the present instance, Calluna (also other heath
taxa) is regarded as deriving mainly from vegetation at or
near the sampling sites and hence will tend to be strongly
over-represented (cf. Moore et al. 1986; Mulder & Janssen
1999). Calluna and other heath taxa are therefore excluded
from the pollen sum. To overcome the disadvantages inherent
in percentage data, concentration and pollen accumulation
rate (PAR) curves are also presented (Fig. 4; selected taxa
only).
Reconstruction of the history of typical Burren upland
species such as Gentiana verna, Dryas octopetala and
Arctostaphylos uva-ursi is difficult because of the severe
under-representation of most of the species in the pollen
record. Indeed, some of the more interesting species are com-
pletely silent, e.g. the orchids including Neotinea maculata.
Another hurdle is that some of the pollen taxa may include
pollen of several species; e.g. Mentha-type which, in the
present context, most likely consists more or less exclusively
of pollen of Thymus polytrichus. On the positive side, the
deposits used here are particularly well suited to archiving
the pollen of under-represented species, because many of the
species in question can be expected to have been present at or
near the point of sampling. Furthermore, the overall low
Table 3. Overview of the coprophilous fungal spore types (CFS) (Groups A and B) and indeterminate non-pollen palynomorphs (NPP)
(Group C)
Type Classification ⁄ description ⁄ ecology
Group A: identified coprophilous fungal types*
Sordaria-type (T. 55A and T. 205) Includes various species of the Sordariales (van Geel et al. 2003). Sordariales are mainly
coprophilous, but occur also on rotting wood and soil (cf. Lundqvist 1972)
Sporormiella-type (T. 113) Widely recognized coprophilous spore type indicative of grazing (e.g. Blackford & Innes 2006;
Davis & Shafer 2006; Raper & Bush 2009). The common species, S. intermedia, occurs on
the dung of most herbivores (Dennis 1978)
Tripterospora-type (T. 169) This type derives probably from a coprophilous Sordariales species and possibly the genus
Tripterospora (van Geel et al. 2003)
Arnium-type (T. 261) Spores of this type derive from the Sordariales and probably the genus Arnium
(van Geel et al. 2003). According to Dennis (1978), most Arnium species occur on dung,
but some species are also abundant on rotting herbaceous stems and wood
Podospora-type (T. 368) This type is regularly recorded in samples from archaeological sites (van Geel et al. 1981, 2003)
and appears to be connected with grazing (cf. Blackford & Innes 2006). Kirk et al. (2001)
regard Podospora species as coprophilous
Group B: additional possible coprophilous fungal types recorded in these investigations
BRN4 (cf. Delitschia sp.) Brown spore c. 29 · 13 lm with truncated base and an apical pore (c. 1.5 lm). The spore
as illustrated (Fig. 7) was probably originally two-celled (possibly a Delitschia species†)
BRN6 (cf. Podospora) Elliptical, non-septate, brown fungal spore, c. 23 · 45 lm with truncated base and a bent
tip with a small pore; resembles spores of Podospora (cf. Lundqvist 1972)
BRN7 (cf. Schizothecium conicum) Elliptical, non-septate, fungal spore, c. 16 · 25 lm with protruding apical pore. It resembles
spores of S. conicum, a common coprophilous species†
BRN8 (cf. Hypocorpa stercoraria) Ellipsoidal, dark brown spore, c. 45 · 21 lm, with longitudinal slit. It resembles
H. stercoraria, a Xylariales fungus that produces perithecia in stromata embedded in dung†
BRN9 (cf. Podospora ⁄Cercophora) Brown, ellipsoidal spore, c. 12 · 20 lm, truncated at one end and a slightly subapical pore
opposite; resembles spores of Podospora and Cercophora (cf. Lundqvist 1972)
Group C: regularly encountered indeterminate NPPs‡
BRN1 Globose microfossil (c. 20 lm diameter) with c. 1.5 lm long processes
BRN2 Spindle-shaped fungal spore (ascospore?), c. 18 · 37 lm, with pores at both ends; one end
tapered
BRN3 Bi-septate fungal spore c. 17 · 33 lm, with truncated pore-like endings at one or both ends.
It may derive from a dematiaceous hyphomycete (includes plant parasitic, saprophytic and
soil fungi)
BRN5 Brown, 2–3 septate fungal spore, c. 23 · 16 lm. This type probably derives from a
dematiaceous hyphomycete
*T. = Type as described in van Geel et al. 2003; †M. Richardson, pers. comm.; ‡Provisional type numbers with prefix BRN (= Burren).
1090 I. Feeser & M. O’Connell
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

input of arboreal pollen (AP), combined with substantial
pollen counts, increases the probability of recording critical
pollen taxa that have low representation.
Regarding the NPP and especially the CFS, these taxa often
represent plants and animals that were present at the coring
site. CFS are regarded as deriving from species that grew
mainly on herbivore dung. Spore production in these fungi is
usually high but dispersal is comparatively poor because of the
location of fruiting bodies close to the ground. CFS are there-
fore regarded as indicative of local grazing (cf. Blackford &
Innes 2006;Mazier et al. 2009).
The diversity and occurrence of coprophilous fungi is
influenced by various factors. Generally, they show prefer-
ences for dung of particular herbivore species (Richardson
1972, 2001). Changes in the CFS assemblage are known to
be influenced by the grazers, local environmental factors
and the local vegetation, i.e. the diet of the animal produc-
ing the dung can be important. Although it cannot be auto-
matically assumed that all the spore types classified as CFS
derive exclusively from dung, good agreement of the avail-
able records with NAP indicators of pastoral activity sup-
ports such an interpretation.
CHRONOLOGY — MONOLITHS
Age–depth curves for profiles CPW1 and GTC7 are presented
in Figs 6 and 7, respectively.
Three 14CAMSdates are used in constructing the age–depth
curve for CPW1. The 14C date, 440 ± 35 BP, was considered
to be unreliable by the 14C laboratory due to small sample size
and so was not used. At 10 cm depth, the so-called secondary
rise in Pinus is evident and AP shows some recovery; these fea-
tures suggest that this level dates to c.AD 1850. The top of the
monolith is assigned to AD 2005. The basal 14C date,
3290 ± 40 BP, derives from fine particulate charcoal. The
charcoal particles are probably broadly contemporaneouswith
the deposition of the minerogenic matrix in which the charcoal
occurs so the date is regarded as indicative of the age of deposi-
tion. Alternatively, the charcoal (also the pollen) may have
been worked into an existing deposit. In either scenario, the14C date suggests that the base of the pollen profile relates to
the mid Bronze Age (c. 1550 BC). The basal lithological unit
(36–45 cm) probably represents glacial drift.
The age–depth curve for profile GTC7 relies on the three
available 14C AMS dates, the date AD 1850, which marks the
Fig.5. Photomicrographs of selected non-
pollen palynomorphs (NPPs) including
coprophilous fungal spores (CFS). 1: Sorda-
ria-type (Type 55A), 2: Sordaria-type (Type
205), 3: Tripterospora-type (Type 169), 4:
Sporormiella-type (Type 113), 5: Podospora-
type (Type 368), 6: Arnium-type (Type 261),
7: Type BRN4 (cf. Delitschia sp.), 8: Type
BRN6 (cf. Podospora), 9: Type BRN7 (cf.
Schizothecium conicum), 10: Type BRN8 (cf.
Hypocorpa stercoraria), 11: Type BRN9
(cf. Podospora ⁄Cercophora), 12: Type
BRN2 (unknown fungal spore), 13: Type
BRN1 (unknown NPP), 14: Type BRN5
(unknown conidia), 15: Type BRN3
(unknown conidia).
Long-term environmental change in the Burren 1091
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100
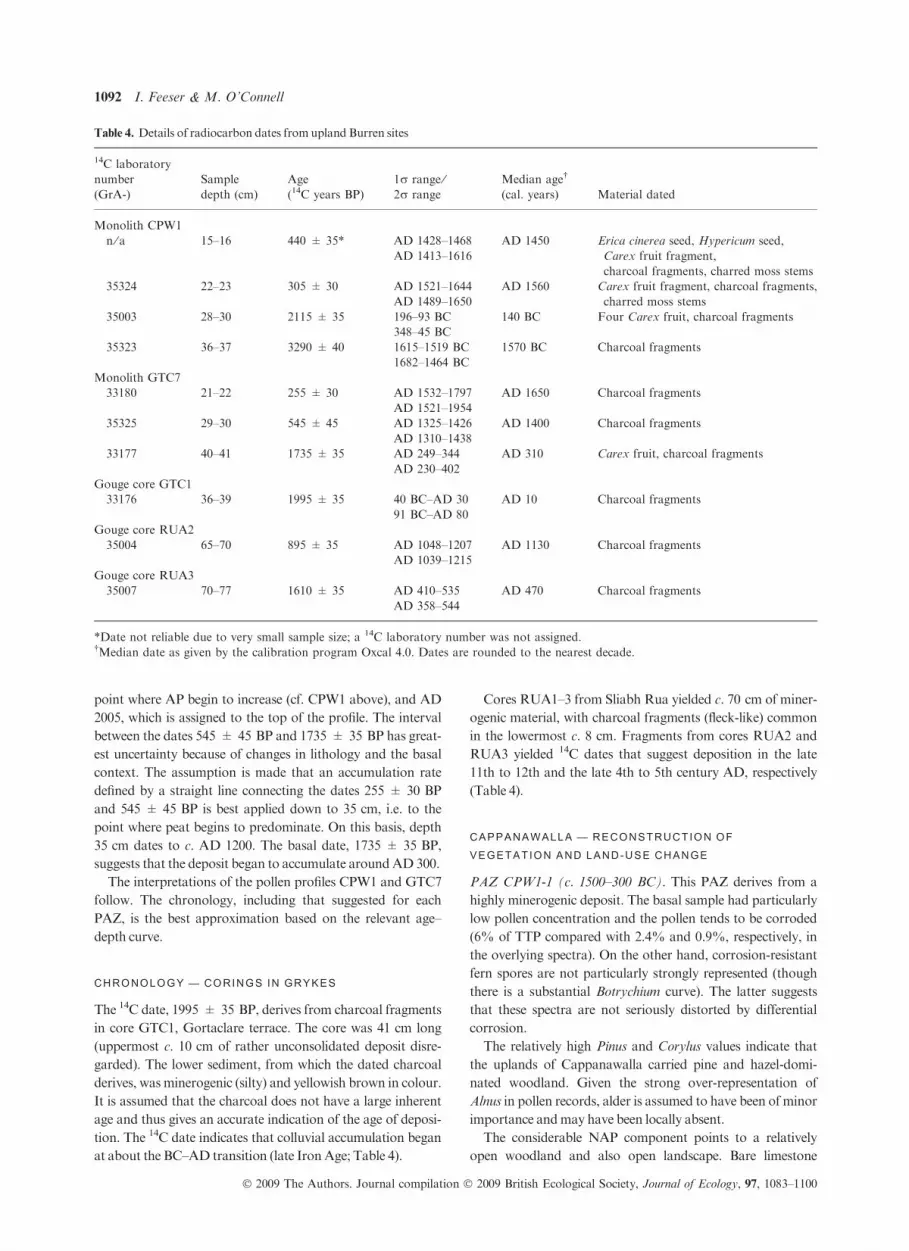
point where AP begin to increase (cf. CPW1 above), and AD
2005, which is assigned to the top of the profile. The interval
between the dates 545 ± 45 BP and 1735 ± 35 BP has great-
est uncertainty because of changes in lithology and the basal
context. The assumption is made that an accumulation rate
defined by a straight line connecting the dates 255 ± 30 BP
and 545 ± 45 BP is best applied down to 35 cm, i.e. to the
point where peat begins to predominate. On this basis, depth
35 cm dates to c. AD 1200. The basal date, 1735 ± 35 BP,
suggests that the deposit began to accumulate aroundAD300.
The interpretations of the pollen profiles CPW1 and GTC7
follow. The chronology, including that suggested for each
PAZ, is the best approximation based on the relevant age–
depth curve.
CHRONOLOGY — CORINGS IN GRYKES
The 14C date, 1995 ± 35 BP, derives from charcoal fragments
in core GTC1, Gortaclare terrace. The core was 41 cm long
(uppermost c. 10 cm of rather unconsolidated deposit disre-
garded). The lower sediment, from which the dated charcoal
derives, wasminerogenic (silty) and yellowish brown in colour.
It is assumed that the charcoal does not have a large inherent
age and thus gives an accurate indication of the age of deposi-
tion. The 14C date indicates that colluvial accumulation began
at about the BC–AD transition (late IronAge; Table 4).
Cores RUA1–3 from Sliabh Rua yielded c. 70 cm of miner-
ogenic material, with charcoal fragments (fleck-like) common
in the lowermost c. 8 cm. Fragments from cores RUA2 and
RUA3 yielded 14C dates that suggest deposition in the late
11th to 12th and the late 4th to 5th century AD, respectively
(Table 4).
CAPPANAWALLA — RECONSTRUCTION OF
VEGETATION AND LAND-USE CHANGE
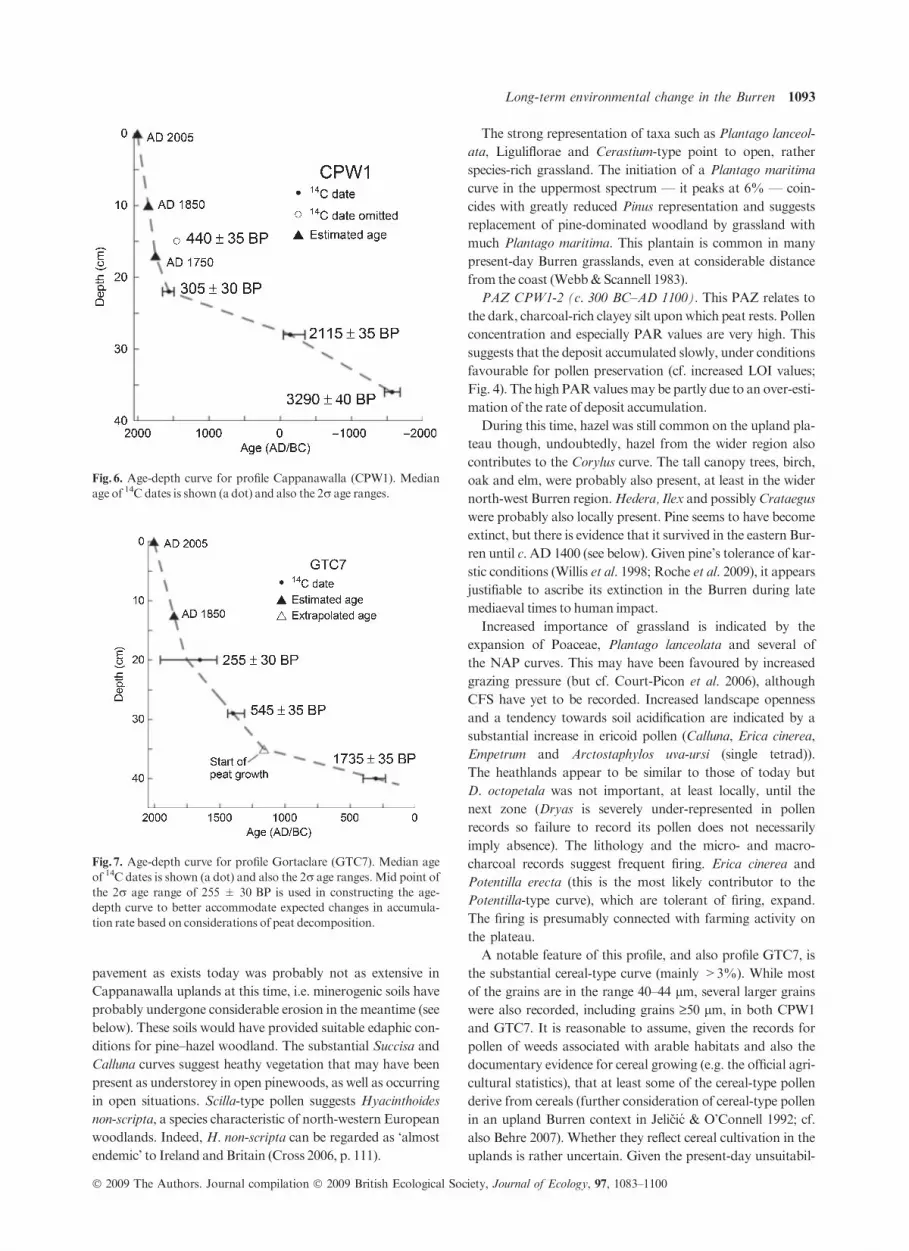
PAZ CPW1-1 (c. 1500–300 BC). This PAZ derives from a
highly minerogenic deposit. The basal sample had particularly
low pollen concentration and the pollen tends to be corroded
(6% of TTP compared with 2.4% and 0.9%, respectively, in
the overlying spectra). On the other hand, corrosion-resistant
fern spores are not particularly strongly represented (though
there is a substantial Botrychium curve). The latter suggests
that these spectra are not seriously distorted by differential
corrosion.
The relatively high Pinus and Corylus values indicate that
the uplands of Cappanawalla carried pine and hazel-domi-
nated woodland. Given the strong over-representation of
Alnus in pollen records, alder is assumed to have been of minor
importance andmay have been locally absent.
The considerable NAP component points to a relatively
open woodland and also open landscape. Bare limestone
Table 4. Details of radiocarbon dates fromupland Burren sites
14C laboratory
number
(GrA-)
Sample
depth (cm)
Age
(14C years BP)
1r range ⁄2r range
Median age†
(cal. years) Material dated
Monolith CPW1
n ⁄ a 15–16 440 ± 35* AD 1428–1468 AD 1450 Erica cinerea seed, Hypericum seed,
Carex fruit fragment,
charcoal fragments, charred moss stems
AD 1413–1616
35324 22–23 305 ± 30 AD 1521–1644 AD 1560 Carex fruit fragment, charcoal fragments,
charred moss stemsAD 1489–1650
35003 28–30 2115 ± 35 196–93 BC 140 BC Four Carex fruit, charcoal fragments
348–45 BC
35323 36–37 3290 ± 40 1615–1519 BC 1570 BC Charcoal fragments
1682–1464 BC
Monolith GTC7
33180 21–22 255 ± 30 AD 1532–1797 AD 1650 Charcoal fragments
AD 1521–1954
35325 29–30 545 ± 45 AD 1325–1426 AD 1400 Charcoal fragments
AD 1310–1438
33177 40–41 1735 ± 35 AD 249–344 AD 310 Carex fruit, charcoal fragments
AD 230–402
Gouge core GTC1
33176 36–39 1995 ± 35 40 BC–AD 30 AD 10 Charcoal fragments
91 BC–AD 80
Gouge core RUA2
35004 65–70 895 ± 35 AD 1048–1207 AD 1130 Charcoal fragments
AD 1039–1215
Gouge core RUA3
35007 70–77 1610 ± 35 AD 410–535 AD 470 Charcoal fragments
AD 358–544
*Date not reliable due to very small sample size; a 14C laboratory number was not assigned.†Median date as given by the calibration program Oxcal 4.0. Dates are rounded to the nearest decade.
1092 I. Feeser & M. O’Connell
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100
pavement as exists today was probably not as extensive in
Cappanawalla uplands at this time, i.e. minerogenic soils have
probably undergone considerable erosion in the meantime (see
below). These soils would have provided suitable edaphic con-
ditions for pine–hazel woodland. The substantial Succisa and
Calluna curves suggest heathy vegetation that may have been
present as understorey in open pinewoods, as well as occurring
in open situations. Scilla-type pollen suggests Hyacinthoides
non-scripta, a species characteristic of north-western European
woodlands. Indeed, H. non-scripta can be regarded as ‘almost
endemic’ to Ireland and Britain (Cross 2006, p. 111).
The strong representation of taxa such as Plantago lanceol-
ata, Liguliflorae and Cerastium-type point to open, rather
species-rich grassland. The initiation of a Plantago maritima
curve in the uppermost spectrum — it peaks at 6% — coin-
cides with greatly reduced Pinus representation and suggests
replacement of pine-dominated woodland by grassland with
much Plantago maritima. This plantain is common in many
present-day Burren grasslands, even at considerable distance
from the coast (Webb& Scannell 1983).
PAZ CPW1-2 (c. 300 BC–AD 1100). This PAZ relates to
the dark, charcoal-rich clayey silt uponwhich peat rests. Pollen
concentration and especially PAR values are very high. This
suggests that the deposit accumulated slowly, under conditions
favourable for pollen preservation (cf. increased LOI values;
Fig. 4). The high PARvaluesmay be partly due to an over-esti-
mation of the rate of deposit accumulation.
During this time, hazel was still common on the upland pla-
teau though, undoubtedly, hazel from the wider region also
contributes to the Corylus curve. The tall canopy trees, birch,
oak and elm, were probably also present, at least in the wider
north-west Burren region.Hedera, Ilex and possiblyCrataegus
were probably also locally present. Pine seems to have become
extinct, but there is evidence that it survived in the eastern Bur-
ren until c.AD 1400 (see below). Given pine’s tolerance of kar-
stic conditions (Willis et al. 1998; Roche et al. 2009), it appears
justifiable to ascribe its extinction in the Burren during late
mediaeval times to human impact.
Increased importance of grassland is indicated by the
expansion of Poaceae, Plantago lanceolata and several of
the NAP curves. This may have been favoured by increased
grazing pressure (but cf. Court-Picon et al. 2006), although
CFS have yet to be recorded. Increased landscape openness
and a tendency towards soil acidification are indicated by a
substantial increase in ericoid pollen (Calluna, Erica cinerea,
Empetrum and Arctostaphylos uva-ursi (single tetrad)).
The heathlands appear to be similar to those of today but
D. octopetala was not important, at least locally, until the
next zone (Dryas is severely under-represented in pollen
records so failure to record its pollen does not necessarily
imply absence). The lithology and the micro- and macro-
charcoal records suggest frequent firing. Erica cinerea and
Potentilla erecta (this is the most likely contributor to the
Potentilla-type curve), which are tolerant of firing, expand.
The firing is presumably connected with farming activity on
the plateau.
A notable feature of this profile, and also profile GTC7, is
the substantial cereal-type curve (mainly >3%). While most
of the grains are in the range 40–44 lm, several larger grains
were also recorded, including grains ‡50 lm, in both CPW1
and GTC7. It is reasonable to assume, given the records for
pollen of weeds associated with arable habitats and also the
documentary evidence for cereal growing (e.g. the official agri-
cultural statistics), that at least some of the cereal-type pollen
derive from cereals (further consideration of cereal-type pollen
in an upland Burren context in Jelicic & O’Connell 1992; cf.
also Behre 2007). Whether they reflect cereal cultivation in the
uplands is rather uncertain. Given the present-day unsuitabil-
Fig.6. Age-depth curve for profile Cappanawalla (CPW1). Median
age of 14C dates is shown (a dot) and also the 2r age ranges.
Fig.7. Age-depth curve for profile Gortaclare (GTC7). Median age
of 14C dates is shown (a dot) and also the 2r age ranges. Mid point of
the 2r age range of 255 ± 30 BP is used in constructing the age-
depth curve to better accommodate expected changes in accumula-
tion rate based on considerations of peat decomposition.
Long-term environmental change in the Burren 1093
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

ity of the Cappanawalla uplands for cereal growing, the exten-
sive fertile soils of the nearby valley floors where there is also
strong evidence for settlement, and the frequency of strong
winds and open landscape that inevitably favour pollen trans-
port to the uplands, it seems reasonable to assume that the
cereal-type pollen recorded in CPW1 derive, probably
exclusively, from arable farming in the lowlands.
As regards the single record of Solanum nigrum-type in
zone CPW1-2, it is doubtful if this derives from S. tuberosum
as potato growing did not become widespread in Ireland
until, at earliest, the 17th century (Fagan 2000; Bell &
Watson 2008). A native species such as S. dulcamara is a
more likely source.
PAZ CPW1-3 (c. AD 1100–1700). During this interval,
hazel declines so that as the zone ends it is no longer of signifi-
cance. There is a corresponding expansion of grassland and
heath. Notable are the high values for Poaceae and Plantago
lanceolata, and good representation of Bidens-type pollen —
this potentially includes pollen of not only common species,
e.g.Bellis perennis, Solidago virgaurea and Senecio vulgaris, but
alsoAntennaria dioica, a typical andwidespread Burren species
of both limestone grasslands and heaths (Webb & Scannell
1983). Vicia-type pollen is well represented, particularly given
the poor pollen production and dispersal in the Fabaceae. This
taxon may include not only common species such as Vicia
cracca and V. sepium, but also Lathyrus linifolius, which is
common today on heathy Burren uplands. Curves forMentha-
type (probably exclusively Thymus polytrichus) and Dryas are
initiated in this zone. Overall, the heathy communities seem to
have been comparable to those of today. Local presence of
Corylus, Betula and Salix (probably S. repens) cannot, how-
ever, be ruled out. Heathers and Potentilla erecta dominated
at the sampling site and Calluna probably acted as host to
Meliola cf. niessleana (Type 14).
High values for akinetes ofZygnema (Type 314) in the upper
spectrum suggest moist conditions, which is also supported by
records for the rhizopod, Assulina. Calluna probably provided
favourable, moist micro-climate (cf. Mulder & Janssen 1999).
A general shift towards cooler and wetter conditions is also
plausible at about this time, which corresponds with the more
severe phase of the Little Ice Age climatic deterioration (Fagan
2000).
Increased grazing on the uplands is supported by sub-
stantial representation of several CFS in the upper spec-
trum. If arable farming was important in the lowlands (as
seems to be the case; cf. cereal-type curve), then upland
summer grazing, if only to facilitate cereal-growing in the
lowlands, is highly plausible. Heavy grazing pressure would
have led to the decline in hazel.
PAZCPW1-4 (c. AD 1700–1850).The decline in AP to 4%
suggests lack of woody vegetation at both local and regional
levels. By the latter part of the 18th century the population in
Ireland had begun to expand rapidly, and this expansion con-
tinued at an accelerated rate until severely checked and ulti-
mately reversed by the Great Famine (1845–47). As a
consequence of increased human impact, the 19th century saw
the final demise of woody vegetation in the Burren, with recov-
ery beginning only towards the end of that century (Kirby
1981, Jelicic & O’Connell 1992). These developments are
reflected in this zone and they also help to secure the chronol-
ogy. Re-introduction of pine and planting of beech, which was
well underway by the late 18th century in western Ireland
(Molloy & O’Connell 2004), is reflected in a rise in Pinus
(so-called secondary rise) and initiation of a curve for Fagus.
Grasslands have expanded, both locally and regionally, at
the expense of hazel (cf. Poaceae, Plantago lanceolata and Lig-
uliflorae). Other noteworthy NAP curves include Anthemis-
type (potentially includes pollen of Achillea millifolium and
Leucanthemum vulgare), Campanula-type (presumably
C. rotundifolia; C. trachelium seems unlikely given its present-
day rarity in the Burren (Roden 2001) and in Ireland gener-
ally), Rhinanthus-type, Linum cartharticum and Polygala (may
include pollen of P. serpyllifolia and P. vulgaris). The species
mentioned are intimately associated with dry, semi-open grass-
land and ⁄or heathy grasslands in the present-day Burren
(Ivimey-Cook & Proctor 1966; Keane & Sheehy Skeffington
1995).
Calluna declines presumably in response to high grazing
pressure. Foot (1864) remarks that wood was so scarcity in the
Burren that Dryas and Pteridium were collected as firewood.
The woody stems of Calluna would also have been valuable in
this context and so may have been used for fuel. Firing in the
uplands seems to be at its minimum, probably because
Calluna, which is often the incentive to fire so that fresh growth
is stimulated, was no longer important.
There is an overall decline in CFS, although Arnium-type
(Type 261) and Podospora-type (Type 368) spores assume
importance. These changes may reflect a shift in pasture man-
agement, including changes in the grazing regime. A shift in
spore representation may also have come about as a result of
the decline in Calluna, which would have resulted in a change
in dung composition and hence, indirectly, the coprophilous
fungal flora (cf. Richardson 2001).
Type BRN1, which achieves 27% at 13 cm, seems to be
associated with complete clearance and an intensive grazing
regimewhich is also true for profileGTC7.
The slender curve for Chenopodiaceae probably reflects the
importance,withina regional context, of potato cultivationand
the associated weed flora which is invariably characterized by
Chenopodiaceae species (cf. alliance Polygono-Chenopodion,
White & Doyle 1982). Solanum-type pollen (a single record)
most likelyderives, in this instance, fromS. tuberosum.
PAZ CPW1-5 (c. AD 1850–1950). This zone reflects the
regeneration of hazel in response to a decline in population
and farming pressures, and the planting of pine and beech in
the wider region. The modest expansion of Corylus to 5%
reflects the regeneration of hazel at a regional level.
The NAP and heath taxa suggest that the upland vegetation
remained largely unchanged. There is, however, less Mentha-
type and Rhinanthus-type pollen, and Polygala representation
increases. These pollen taxa probably represent mainly local
vegetation at the sampling site and so the changes may not be
of major significance as regards regional vegetation and land-
use dynamics. On the other hand, the Artemisia curve is inter-
1094 I. Feeser & M. O’Connell
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

rupted, suggesting a decline in tillage which characterized the
20th century.
Coprophilous fungi are poorly represented which suggests
further changes in the grazing regime, including possibly less
emphasis on active management. Interestingly, micro-charcoal
is well represented. Firing has not been a feature of land-man-
agement in the Burren in recent times. This, and lack of both
macro-charcoal and indications of burning in the upper part of
the monolith, suggest that the micro-charcoal is of regional
origin and hence not indicative of local firing as in PAZ 2 (see
above).
GORTACLARE – VEGETATION AND LAND-USE CHANGE
PAZ GTC7-1 (c. AD 100–1650). Corylus at c. 18% suggests
the presence of hazel scrub, at least in the region and possibly
locally on the plateau. Slender curves for Betula, Quercus and
Alnus suggest local or at least regional presence of these trees.
Pine was a veryminor component, probably at a regional level,
until c.AD1400 (base of subzone 1c) when it became extinct.
As organic matter began to accumulate, heathy vegetation
with much Calluna and Succisa dominated locally (subzone
1a). Throughout the period, grasslands with much Plantago
lanceolata, Liguliflorae (the latter is expected to include Leon-
todon, Pilosella and Taraxacum), and also Vicia and Lathyrus
spp. (cf.Vicia-type)were important.TypicalBurrenplants such
as Campanula rotundifolia, Galium spp. (most likely G. verum;
possibly also G. boreale, G. saxatile and Rubia peregrina) were
well represented in theuplandflora.
In subzone 1c, Poaceae and Plantago lanceolata play a less
important role than hitherto (cf. percentage and concentration
curves) while the Liguliflorae andCampanula rotundifoliawere
favoured. This is presumably due to changes in farming activ-
ity. Whereas CFS are rather poorly represented at the begin-
ning of the zone, improved representation in subzone 1c (c.
AD 1400–1650) suggests an increase in local grazing. Greater
representation of the testate amoeba Hyalosphenia subflava
points to peat disturbance (van Geel 1978) and dry conditions
(Charman 2001). Micro- and macro-charcoal are important
throughout the zone, and Gelasinospora spores and Cenocco-
cum sclerotia, both from fungi that have a strong association
with firing (van Geel 1978), are well represented. It seems that
firing, which is most likely related to farming activity, was
important on the plateau, especially as the zone ends (cf. sharp
peak in micro-charcoal). Firing would also have favouredCal-
luna andErica cinerea.
A substantial cereal-type curve (average 4.2% in subzones
1b and 1c) suggests considerable emphasis on cereal growing.
Amoreor less continuousArtemisia curve supports this.Cereal
growing was probably confined to the fertile lowlands but the
uplands, even if rather unpromising for cereal growing from a
present-day perspective, cannot be completely excluded.
PAZ GTC7-2 (c. AD 1650–1850). Low AP representation
(average 3.5%) suggests a fully open landscape, more or less
devoid of trees and shrubs. Low AP, high NAP and increased
CFS values suggest intensive farming. The distinct reduction
in Calluna is probably attributable to high grazing pressure
and possibly use of the woody stems as firewood (cf. CPW1-4
above). Firing of vegetation during this time, however, was
greatly reduced or ceased (cf. charcoal data).
During subzone 2a, Plantago lanceolata was exceptionally
well represented in the local grasslands. Increased representa-
tion of Pteridium, coinciding with a pronounced decline in
Corylus, suggests local clearance of hazel scrub during the late
17th century. In subzone 2b, the grassland composition
changed so that Poaceae, Ranunculus and Rubiaceae spp.
(cf. Galium-type curve) were favoured. Interestingly
also tubular composites were favoured over ligulates
(cf. Bidens-type and Liguliflorae curves). As observed in
CPW1-4, Type BRN1 peaks when grazing pressures were
high. Mineral matter was conspicuous in the peat (cf. also
lower LOI) which points to increased local disturbance.
These changes are probably attributable to severe grazing
pressures associated with a sharp rise in population
(subzone 2b spans c. AD 1800–1850).
PAZ GTC7-3 (c. AD 1850–1950). This zone reflects the
regeneration of Corylus and other woody species as a result
of decline in farming that began in the mid 19th century.
Evidence from other sources (e.g. Kirby 1981) indicates that
woodland and scrub regeneration was largely confined to
steeply sloping parts such as the eastern side of Slievecarran
and the Glen of Clab. There is no evidence to support
regeneration on the plateau. Grazing, however, seems to
have declined; cf. decline in NAP and especially Plantago
lanceolata, Liguliflorae and Galium-type, and increase in
Calluna and other ericoids. The NPP records consist mainly
of Assulina and Meliola cf. niessleana which are probably
associated with Calluna (see CPW1). Hyalosphenia subflava,
which is indicative of disturbance, is also well represented.
On the other hand, CFS are poorly represented. Casual
observations over recent years suggest that grazing density
is relatively low on Gortaclare compared with Cappanawal-
la. This seems to have been also true during the late 19th
and earlier part of the 20th century. A peak in micro-char-
coal at 11 cm (also dark, charcoal-enriched peat) suggests a
phase of heath firing, a practice that seems to have been
completely abandoned in recent times.
PALAEOENVIRONMENTAL INFORMATION DERIVED
FROM GRYKES
Grykes on Gortaclare terrace. Though several grykes were
probed on the plateau and surrounding area, substantial min-
erogenic deposits in grykes were noted only on Gortaclare ter-
race (see Fieldwork inMethods and materials). These deposits
are regarded as a colluvium that has arisen through soil loss
from nearby sloping ground. The charcoal fragments from
near the base of sequences were presumably deposited with
eroding soil. The 14C date 1995 ± 35 BP, which derives from
the charcoal, provides a reliable indication that the erosion
dates to about the BC–AD transition. It is assumed that it was
triggered by woodland clearance or intensive grazing. The ero-
sionwas probably contemporaneous with, or shortly preceded,
the beginning of theGTC7 record.
Long-term environmental change in the Burren 1095
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

Grykes at Sliabh Rua. The two available dates indicate
erosion taking place in the early and mid to late mediaeval
periods (late 4th to 5th and late 11th to 12th centuries
AD), respectively, i.e. periods noted for increased levels of
human activity in western Ireland. Again, it is assumed that
the erosion resulted from disturbance of the vegetation
cover due to farming.
Discussion and conclusions
OVERVIEW OF ENVIRONMENTAL CHANGE AND LAND
USE IN THE BURREN UPLANDS
The palaeoecological evidence presented above provides fresh
insights into the developmental history of two important and
species-rich Burren upland habitats, and, furthermore, new
information on past soil erosion. To get a better overall per-
spective, the new data are considered in the context of results
from another upland investigation, namely that at Lios
Lairthın Mor, which lies south-west of the Ballyvaughan low-
lands (grid ref. 53�04.8¢ N, 09�13.26¢ W; 270 m a.s.l.; Jelicic &
O’Connell 1992; Fig. 1). The profile (LLM II; 192 cm long)
derives from a peat mound (diameter <50 m) that lies at the
junction of karstic limestone and a Namurian shale outlier.
Summary diagrams of the three profiles, drawn to an age
rather than a depth scale, are presented in Fig. 8. In LLM II,
the bog and heath pollen are presumed to arise mainly from
the sampling site and to a lesser extent from the large adjoining
bog to the east, while the terrestrial component is expected to
arise mainly from the local karst, which has minimal drift
cover. The main relevant archaeological evidence for land use
at Lios Lairthın Mor are the many ancient field boundaries
and also ringforts, the nearest situated within 80 m of the cor-
ing location, LLM II (Fig. 1).
Profiles CPW1 and LLM II span the interval mid Bronze
Age tomodern times while profile GTC7 opens in the late Iron
Age. The basal part of CPW1 suggests open pine–hazel wood-
land with grassland and heath in the interval c. 1500–300 cal.
BC, i.e. mid Bronze Age to mid Iron Age. At Lios Lairthın
Mor, on the other hand, the landscape was open, and hazel
was the main woody species. The relevant pollen spectra from
CPW1 are, however, from highly minerogenic sediments, so
rather different taphonomic processes are at play and also the
dating may not be as secure as LLM II. These considerations
aside, the overall evidence points to contrasting woodland
composition and cover (at Lios Lairthın Mor, pine unimpor-
tant or absent and landscape more open), presumably largely
due to different farming regimes and especially grazing
intensity.
In CPW1, strong human impact registers first in zone 2
(decline in Pinus and charcoal-enriched minerogenic sedi-
ments) and then increases distinctly at c.AD 1700 (top of zone
3; decline in Corylus and further expansion of Poaceae and
Plantago lanceolata). This sets a pattern that is maintained
until the mid 19th century when regeneration of woody vegeta-
tion commences (zones 4 and 5). Changes over the correspond-
ing intervals, i.e. from c. 300 BC to the present day, are
recorded in greater detail in LLM II. A distinctive feature is
the regeneration of woody vegetation, involving mainly hazel,
ash and birch as a response to a steep decline in farming, i.e.
the so-called Late Iron Age Lull (cf. Molloy & O’Connell
2004), which, in this instance, is dated to c.AD 300–600 (zone
LLM II-2). Human impact, including arable farming, is partic-
ularly strong from about AD 1100, but the first intensive clear-
ances in the partially wooded landscape begin at about AD
1650, and final clearance of hazel scrub begins a century later
(see below).
Profile GTC7, although covering a shorter interval (c. AD
100–1950), has the advantage of a higher sampling resolution
than CPW1. The Gortaclare uplands carried hazel scrub until
c. AD 1650 or, alternatively, the nearby lowlands had more
scrub than today.
Near the base of GTC7 (zone 1a, 39 cm; c.AD 500), there is
a decline in Plantago lanceolata and high Calluna values. This
may be an expression, albeit in subdued form, of a Late Iron
Age Lull. The abandonment of active farming may have
resulted in conditions suitable for peat initiation (cf. Molloy &
O’Connell 1993).
At c.AD 1500, there is evidence for increased farming activ-
ity in both GCT7 (subzone 1c) and LLM II. This may be part
of a general recovery subsequent to the Black Death, which
reached Ireland in AD 1348 (Kelly 2001; Yeloff & van Geel
2007).
The charcoal records from GTC7 and CPW1 indicate that
burning of upland heaths, most likely connected with pastoral
farming, was important until at least the 17th century. Further
research into fire history and the long-term impact of firing
upland heaths in the Burren is desirable.
The evidence from all three sites points to major change,
particularly from the 17th century onwards, that finds expres-
sion mainly in the decline of Corylus in the context of strong
farming pressures (GTC7-2, top of CPW1-3 and CPW1-4,
LLM II-5). This coincides with the laying out of large field sys-
tems in the Burren uplands between c.AD1650–1750, possibly
in connectionwith a shift from subsistence farming to amarket
economy (Plunkett Dillon, 1985). During this time sheep rear-
ing became increasingly important. Dutton (1808), for
instance, reports that large flocks of sheep were being reared
on a year-around basis in the western Burren uplands. High
Potentilla-type values at about this time in profile CPW1 point
to increased abundance and flowering of Potentilla erecta, a
development probably favoured by intensive sheep grazing (cf.
Moore et al. 1986).
Exceptionally low Corylus representation at about c. AD
1800 in all three profiles highlights the extreme scarcity
of shrubby vegetation (even Calluna was adversely affected).
This corresponds with high representation of CFS which
emphasizes the importance of upland grazing in the farming
economy.
The relatively strong recovery in woody vegetation, and
especially hazel, at a regional level, as farming pressures
declined and tree-planting became more common, is
reflected in the uppermost zone of all three profiles. The
phenomenon of hazel-scrub expansion of the last few dec-
1096 I. Feeser & M. O’Connell
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

Fig.8.Selectedpercentagecurves
from
Gortaclare
(GTC7),Cappanawalla(C
PW1)an
dLiosLairthın
Mor(profileLLM
II)plotted
toatimescale.Ulm
usispoorlyrepresentedin
GTC7andso
hasbeenomitted.T
hecoprophilousfungalspore
curves
includethose
taxaclassified
ingroupsAandBin
Table3.
Long-term environmental change in the Burren 1097
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

ades (cf. ERA-Maptec 2006) is probably not captured as
sampling stopped short of the uppermost deposits. A shift
in the composition of the upland grassland communities is
also notable (cf. NAP curves and especially Plantago
lanceolata), presumably as a result of changes in grazing
regimes (cf. decrease in CFS).
EVIDENCE FOR LOSS OF SOIL COVER IN THE BURREN
Apart from most of the lowlands, extensive smooth pavement
and ⁄or shattered limestone terrain with little or no soil cover
characterize much of the Burren. As a result of research in
recent decades, past soil erosion, once largely speculative, is
now underpinned by a considerable body of evidence. This
includes soil remnants sealed by archaeological structures
(Drew 1982, 1983; Plunkett Dillon, 1985; Lynch 1994; Jones
2004), colluvial deposits in grykes (Drew 1983) and karren fea-
tures, preserved on limestone slaps used in megalithic-tomb
construction, that suggest an acidic soil cover in nowadays
soil-free areas (Plunkett Dillon 1983). Drew (1982) reported
preliminary results from U ⁄Th dating of speleothems from
Burren caves that suggested cessation of calcite deposition in
the 2nd millennium BC as a result of loss of soil cover. Mean-
while, a speleothem from Polnagollum cave, south-western
Burren, has yielded an end-of-deposition date of
3230 ± 50 cal. BP (F. McDermott, pers. comm.). This points
to soil cover on the overlying bedrock until at least the mid to
late BronzeAge.
The present investigations have demonstrated the presence
of minerogenic soil in shallow peat-covered depressions on the
Cappanawalla upland and under shallow peat at Sliabh Rua.
In the latter Knockanes–Sliabh Rua area, mass soil movement
has been documented and buried charcoal has yielded the
AMS 14C date 3070 ± 35 BP, which suggests substantial soil
erosion in the late Bronze Age (Moles et al. 1999; Moles &
Moles 2002).
The data presented here indicate that soil erosion may
not be confined to the Neolithic and Bronze Age as is
sometimes postulated (e.g. Gibson 2007), but continued to
be of significance well into the mediaeval period and proba-
bly beyond. Erosion has probably characterized Burren soils
since the ice sheets retreated (c. 12 000 BC), with intense
erosion taking place at particular times, e.g. during climatic
downturns (cf. Younger Dryas; c. 10.6–9.5 BC) or during
phases of woodland clearance associated with intensive
farming.
It is puzzling, however, that lake sediments have failed to
yield conclusive evidence for soil erosion (cf. Watts 1984;
Schettler et al. 2006), even allowing for the possibility that
eroding soils may not readily reach lake basins where karst-
type, sub-surface hydrology prevails. Sediments from Molly’s
Lough, however, show elevated magnetic susceptibility values
that have been interpreted as indicative of sustained erosion
during the Neolithic and Bronze Age (c. 6000–3000 cal. BP;
Lamb&Thompson 2005). The lake, however, may not be typ-
ical for the Burren, lying, as it does, near the karst–shale geo-
logical divide.
While it is clear that soil erosion on a significant scale has
occurred in the Burren since the last glaciation and that the link
with human activity is strong (but cf. Wilson et al. (2008) for a
contrasting soil-erosion history from limestone terrain in
north-west England), details regarding extent, timing and
quantification of erosion, and its consequences for vegetation
dynamics, flora history and past human settlement patterns
have still to be elaborated.
FRESH INSIGHTS INTO THE DEVELOPMENT AND
ORIGIN OF THE UPLAND BURREN FLORA
The late Holocene history of key upland Burren species and
plant communities is considered in the light of the newly avail-
able evidence (see also Interpretation of data).
Dryas was not recorded in LLM II, there is only a single
record in GTC7 (33 cm) while, in CPW1, a continuous
curve is initiated at the base of zone 3 (c. AD 1100), i.e.
the record begins as peat accumulation starts. At CPW1,
the decrease in hazel and the beginning of peat accumula-
tion clearly provided locally favourable conditions for
expansion of Dryas in the mediaeval period. Failure to
record pollen of Dryas prior to this cannot be regarded as
proof of absence. The species appears to be so under-repre-
sented in pollen diagrams as to require local and consider-
able presence before it registers in pollen records (cf. Jelicic
& O’Connell 1992).
Records for Gentiana (cf. verna) are extremely scarce in all
profiles (CPW1-4, 16 cm: two pollen; and LLM II-1, 168 cm;
c. 400 BC: a single pollen). This is presumably due to poor pol-
len production and dispersal rather than rarity so that the
records, though sparse, are of some significance.
As regards Arctostaphylos uva-ursi, there is only a single
record (zone CPW1-2, 25 cm; c. AD 1500), prior to the
decline in Corylus on Cappanawalla. Arctostaphylos uva-ursi
seems to be very much under-represented and so no firm
conclusions regarding presence or absence in the past can be
drawn. Absence, however, is unlikely given the species’
frequency today in the north-west Burren uplands. The
CPW1 profile shows that other heath species such as Erica
cinerea (seed also recorded) and Empetrum nigrum were
important for at least the last two millennia at Cappanawalla
but are much less well represented at Gortaclare (especially
E. nigrum), which again corresponds to the present-day
situation. Interestingly, E. nigrum is abundant on nearby
Turlough Hill which is at similar elevation, but more compa-
rable to the north-west Burren uplands in being near the
coast.
The importance of Plantago maritima at Cappanawalla and
Lios Lairthın Mor, and rather poor representation in GTC7
are noteworthy. Plantago maritima is, however, frequent in the
eastern Burren and is locally abundant in limestone grasslands
on the lower slopes of Gortaclare, but it was not noted at or
near the sampling site, GTC7, which serves to emphasize the
local character of the pollen profiles.
Inter-site differences, as elaborated above, suggest that fac-
tors other than grazing and overall levels of human impact
1098 I. Feeser & M. O’Connell
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

have a major influence on distribution and composition of the
upland plant communities. Differences in wind exposure, sub-
strate (e.g. shallow minerogenic soils bearing heath on Cap-
panawalla; chert locally extensive on Gortaclare), humidity
levels and atmospheric nutrient input as determined by dis-
tance from the ocean probably all play an important role. To
what degree these are plagioclimax communities, especially the
arctic-alpine-like heaths as on Cappanawalla, remains to be
demonstrated.
The survival of arctic-alpine species in an eu-oceanic region
during the post-glacial, when woodlands largely dominated, is
another intriguing aspect of the Burren flora. The evidence pre-
sented here for open pine woodland with hazel on the Cappan-
awalla uplands until relatively late in the Holocene is
noteworthy, but not altogether surprising, given the role
played by pine during much of the Holocene in the Burren and
Aran Islands (Watts 1984; Molloy & O’Connell 2004; Feeser
2009). A modern-day analogue of these former plant commu-
nities is the open pine woodlands common on shallow alkaline
soils in Scandinavia (cf. Dierssen 1996; Kelly & Connolly
2000). TheMelico nutantis–Pinetumwoodlands on calcareous
rendzinas in southern Scandinavia include not only Sesleria
caerulea, Galium boreale, Geranium sanguineum and Campan-
ula rotundifolia,but alsoheathy species suchasCalluna vulgaris,
Arctostaphylos uva-ursi, Succisa pratensis and Potentilla erecta
(cf. Dierssen 1996). From mid Norway northwards, Dryas
octopetala is an important component in open birch–pine
woodlands on shallow alkaline rendzinas (Epipacto
atrorubentis-Betuletum). Given these striking floristic and
environmental similarities, we suggest that open Pinus
sylvestris-dominated woodland provided an important and
extensive habitat for the arctic-alpine flora of the Burren dur-
ing prolonged periods of more or less full woodland cover.
This would also help explain why some 39 species of putative
ectomycorrhizal fungi, normally intimately associated with
pine woodland, are confined in the Burren to these upland arc-
tic-alpine plant communities (Harrington & Mitchell 2005;
also Harrington 2003). Both the understorey and the charac-
teristic ectomycorrhizal flora of these open pine woodlands
have presumably survived, in situ, the demise of pine.
Acknowledgements
A. Bird and N. Higgins assisted with fieldwork, and A. Bird carried out pre-
liminary analyses on CPW1. J. van Leeuwen and M. Richardson advised on
identification of coprophilous fungal spores. F. McDermot provided informa-
tion on unpublished speleothem analyses. The paper has benefited from helpful
comments by referees (two anonymous and M.-J. Gaillard). The research was
financially supported by an EMBARK Initiative Postgraduate Research
Fellowship (IRCSET) to I.F.
References
Bakker, M. & van Smeerdijk, D.G. (1982) A palaeoecological study of a late
Holocene section from ‘‘Het Ilperveld’’, western Netherlands. Review of
Palaeobotany and Palynology, 36, 95–163.
Behre, K.-E. (2007) Evidence for Mesolithic agriculture in and around Central
Europe?VegetationHistory and Archaeobotany, 16, 203–219.
Bell, J. & Watson, M. (2008) A History of Irish Farming 1750–1950. Four
Courts Press, Dublin.
Beug, H.-J. (2004) Leitfaden der Pollenbestimmung fur Mitteleuropa und
angrenzende Gebiete. VerlagDr Friedrich Pfeil,Munchen.
Blackford, J.J. & Innes, J.B. (2006) Linking current environments and
processes to fungal spore assemblages: surface NPM data from woodland
environments.Review of Palaeobotany and Palynology, 141, 179–187.
Charman, D.J. (2001) Biostratigraphic and palaeoenvironmental applications
of testate amoebae.Quaternary Science Reviews, 20, 1753–1764.
Court-Picon, M., Buttler, A. & de Beaulieu, J.-L. (2006) Modern pollen ⁄ vege-tation ⁄ land-use relationships in mountain environments: an example from
theChampsaur valley (FrenchAlps).VegetationHistory and Archaeobotany,
15, 151–168.
Cross, J.R. (2006) The potential natural vegetation of Ireland. Biology and
Environment (Proc. R. Ir. Acad.), 106B, 65–116.
D’Arcy, G. & Hayward, J. (1992) The Natural History of the Burren. Immel
Publishing, London.
Davis, O.K. & Shafer, D.S. (2006) Sporormiella fungal spores, a palynological
means of detecting herbivore density. Palaeogeography, Palaeoclimatology,
Palaeoecology, 237, 40–50.
Dennis, R.W.G. (1978)British Ascomycetes. Cramer, Vaduz.
Dierssen, K. (1996)VegetationNordeuropas. Ulmer Verlag, Stuttgart.
Drew, D.P. (1982) Environmental archaeology and karstic terrains: the exam-
ple of the Burren, Co. Clare, Ireland. BAR International Series, 146, 115–
127.
Drew, D.P. (1983) Accelerated soil erosion in a karst area: the Burren, western
Ireland. Journal of Hydrology, 61, 113–124.
Dunford, B. (2001) Agricultural practices and natural heritage.The Book of the
Burren, 2nd edn (eds J.W. O’Connell & A. Korff), pp. 234–238. Tır Eolas,
Kinvara.
Dunford, B. (2002) Farming and the Burren. Teagasc, Dublin.
Dutton, J. (1808) Statistical Survey of the County of Clare. Royal Dublin
Society, Dublin.
ERA-Maptec (2006) Assessment of Landscape Change and Effects on Archaeol-
ogy and an Assessment of Habitat Survey in the Burren, Co. Clare. The Heri-
tage Council, Kilkenny.
Fagan, B. (2000) The Little Ice Age. How Climate Made History 1300-1850.
Basic Books, NewYork.
Feeser, I. (2009) Palaeocological Investigations Towards Reconstruction of
Holocene Environmental Change in the Burren, Co. Clare, with Particular
Reference to Mullach Mor and Selected Burren Uplands. PhD thesis,
National University of IrelandGalway,Galway.
Feeser, I. & O’Connell, M. (2010) Late Holocene farming and vegetation
dynamics in upland karst based on pollen and coprophilous fungal spore
analyses: an example from the Burren, western Ireland. Vegetation History
and Archaeobotany, 19, in press.
Finch, T.F. (1971) Soils of County Clare. Soil Survey Bulletin No. 23. An Foras
Taluntais, Dublin.
Foot, F.J. (1864) On the distribution of plants in Burren, County of Clare.
Transactions of the Royal Irish Academy, 24, 143–160.
van Geel, B. (1978) A palaeoecological study of Holocene peat bog sections in
Germany and The Netherlands. Review of Palaeobotany and Palynology, 25,
1–120.
van Geel, B., Bohncke, S.J.P. & Dee, H. (1981) A palaeoecological study of an
upper late glacial and Holocene sequence from ‘‘De Borchert’’, The Nether-
lands.Review of Palaeobotany and Palynology, 31, 367–448.
van Geel, B., Coope, G.R. & van der Hammen, T. (1989) Palaeoecology and
stratigraphy of the lateglacial type section at Usselo (The Netherlands).
Review of Palaeobotany and Palynology, 60, 25–129.
vanGeel, B., Buurman, J., Brinkkemper, O., Schelvis, J., Aptroot, A., van Ree-
nen, G. et al. (2003) Environmental reconstruction of a Roman Period settle-
ment site in Uitgeest (The Netherlands), with special reference to
coprophilous fungi. Journal of Archaeological Science, 30, 873–883.
Gibson, D.B. (2007) The hill-slope enclosures of the Burren, Co. Clare. Pro-
ceedings of the Royal Irish Academy, 107C, 1–29.
Harrington, T.J. (2003) Relationships between macrofungi and vegetation
in the Burren. Biology and Environment (Proc. R. Ir. Acad.), 103B, 147–
159.
Harrington, T.J. & Mitchell, D.T. (2005) Ectomycorrhizas associated with a
relict population ofDryas octopetala in the Burren, western Ireland. I. Distri-
bution of ectomycorrhizas in relation to vegetation and soil characteristics.
Mycorrhiza, 15, 425–433.
Heiri, O., Lotter, A.F. & Lemcke, G. (2001) Loss on ignition as a method for
estimating organic and carbonate content in sediments: reproducibility and
comparability of results. Journal of Paleolimnology, 25, 101–110.
Ivimey-Cook, R.B. & Proctor, M.C.F. (1966) The plant communities of the
Burren, Co. Clare.Proceedings of the Royal Irish Academy, 64B, 211–301.
Long-term environmental change in the Burren 1099
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100

Jacobson,G.L. & Bradshaw,R.H.W. (1981) The selection of sites for paleoveg-
etational studies.Quaternary Research, 16, 80–96.
Jeffrey, D.W. (2003) Grasslands and heath: a review and hypothesis to explain
the distribution of Burren plant communities. Biology and Environment
(Proc. R. Ir. Acad.), 103B, 111–123.
Jelicic, Lj. & O’Connell, M. (1992) History of vegetation and land use from
3200 B.P. to the present in the north-west Burren, a karstic region of western
Ireland.VegetationHistory and Archaeobotany, 1, 119–140.
Jones, C. (2004) The Burren and the Aran Islands. Exploring the Archaeology.
Collins Press, Cork.
Keane, S. & Sheehy Skeffington, M. (1995) Vegetation in the east Burren
uplands in relation to land use and conservation. Irish Grasslands. Their Biol-
ogy and Management (eds D.W. Jeffrey, M.B. Jones & J.H. McAdam). pp.
253–266, Royal IrishAcademy,Dublin.
Kelly, D.L. & Connolly, A. (2000) A review of the plant communities associ-
ated with Scots Pine (Pinus sylvestris L.) in Europe, and an evaluation
of putative indicator ⁄ specialist species. Investigacion Agraria, Sistemas y
Recursos Forestales, Fuera de Series No. 1, 15–39.
Kelly,M. (2001)AHistory of the Black Death in Ireland. Tempus, Stroud.
Kirby, E.N. (1981) An Ecological and Phytosociological Study of Corylus
Avellala L. in the Burren western Ireland. PhD thesis, National University of
IrelandGalway (University College Galway), Galway.
Kirk, P.M., Cannon, P.F., David, J.C. & Staplers, J.A. (eds) (2001) Ainsworth
andBisby’sDictionaryof the Fungi, 9th edn.OxfordUniversityPress,Oxford.
Lamb, H.F. & Thompson, A.D. (2005) Unusual mid-Holocene abundance of
Ulmus in western Ireland – human impact in the absence of a pathogen? The
Holocene, 15, 447–452.
Lundqvist, G. (1972) Nordic Sordariaceae s. lat. Symbolae Botanicae Upsalien-
ses, 20, 1–374.
Lynch, A. (1994) Poulnabrone portal tomb. Burren, Co. Clare. IQUA Field
GuideNo. 18 (edM.O’Connell). pp. 18–20, Irish Association forQuaternary
Studies, Dublin.
Mazier, F., Galop, D., Gaillard, M.J., Rendu, C., Cugny, C., Legaz, A. et al.
(2009) Multidisciplinary approach to reconstructing local pastoral activities:
an example from the Pyrenean Mountains (Pays Basque). The Holocene, 19,
171–188.
Moles, N. & Moles, R. (2002) Influence of geology, glacial processes and land
use on soil composition and Quaternary landscape evolution in the Burren
National Park, Ireland.Catena, 47, 291–321.
Moles, N., Moles, R. & O’Donovan, G. (1995) Evidence for the presence of
Quaternary loess-derived soils in the Burren karstic area, western Ireland.
Irish Geography, 28, 48–63.
Moles, R., Moles, N. & Leahy, J.J. (1999) Radiocarbon dated episode of
BronzeAge slope instability in the south-eastern Burren, County Clare. Irish
Geography, 32, 52–57.
Molloy, K. & O’Connell, M. (1993) Early land use and vegetation history at
Derryinver Hill, Renvyle Peninsula, Co. Galway, Ireland. Climate Change
and Human Impact on the Landscape (ed F.M. Chambers). pp. 185–199,
Chapman andHall, London.
Molloy, K. & O’Connell, M. (2004) Holocene vegetation and land-use dynam-
ics in the karstic environment of Inis Oırr, Aran Islands, western Ireland:
pollen analytical evidence evaluated in the light of the archaeological record.
Quaternary International, 113, 41–61.
Moore, P.D., Evans, A.T. & Chater, M. (1986) Palynological and stratigraphic
evidence for hydrological changes in mires associated with human activity.
Anthropogenic Indicators in Pollen Diagrams (ed K.-E. Behre). pp. 209–220,
Balkema, Rotterdam.
Moore, P.D., Webb, J.A. & Collinson, M.E. (1991) Pollen Analysis, 2nd edn.
Blackwell Scientific, London.
Mulder, C. & Janssen, C.R. (1999) Occurrence of pollen and spores in relation
to present-day vegetation in a Dutch heathland area. Journal of Vegetation
Science, 10, 87–100.
O’Connell, J.W. &Korff, A. (2001)The Book of the Burren, 2nd edn. Tır Eolas,
Kinvara.
Osborne, B. & Jeffrey, D.W. (2003) Introduction and overview. Understanding
the Burren. Special Issue, Biology and Environment (Proc. R. Ir. Acad), vol.
103B (eds B. Osborne&M. Jones), pp. 107–109.Royal Irish Academy,Dub-
lin.
Osborne, B. & Jones, M. (eds) (2003) Understanding the Burren. Special Issue,
Biology and Environment (Proc. R. Ir. Acad), vol. 103B. Royal Irish Acad-
emy,Dublin.
Osborne, B., Black, K., Lanigan, G., Perks, M. & Clabby, G. (2003) Survival
on the exposed limestone pavement in the Burren: photosynthesis and water
relations of three co-occurring plant species.Biology and Environment (Proc.
R. Ir. Acad.), 103B, 125–137.
Oyama, M. & Takehara, H. (1970) Standard Soil Color Charts, 2nd edn. Fuji-
hara, Tokyo.
Pals, J.P., van Geel, B. & Delfos, A. (1980) Paleoecological studies in the
Klokkeweel bog near Hoogkarspel (Prov. of Noord-Holland). Review of
Palaeobotany and Palynology, 30, 371–418.
Plunkett Dillon, E.C. (1983) Karren analysis as an archaeological technique.
Landscape Archaeology in Ireland, pp. 81–94 (eds T. Reeves-Smyth &
F.Hamond). p. 116, Series 116, Oxford.
Plunkett Dillon, E.C. (1985) The Field Boundaries of the Burren, County Clare.
PhD thesis, University ofDublin (Trinity College), Dublin.
Pracht, M., Lees, A., Leake, B., Feely, M., Long, B., Morris, J. et al. (2004)
Geology of Galway Bay: A Geological Description to Accompany the Bedrock
Geology 1:100,000 Scale Map Series, Sheet 14, Galway Bay. Geological Sur-
vey of Ireland,Dublin.
Ramsey, C.B. (2001) Development of the radiocarbon calibration program
OxCal.Radiocarbon, 43, 355–363.
Raper, D. & Bush, M. (2009) A test of Sporormiella representation as a predic-
tor of megaherbivore presence and abundance. Quaternary Research, 71,
490–496.
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Bertrand,
C.J.H. et al. (2004) IntCal04 terrestrial radiocarbon age calibration, 0–26 cal
kyr BP.Radiocarbon, 46, 1029–1058.
Richardson, M.J. (1972) Coprophilous ascomyctes on different dung types.
Transactions of the BritishMycological Society, 58, 37–48.
Richardson, M.J. (2001) Diversity and occurrence of coprophilous fungi.
Mycological Research, 105, 387–402.
Robinson, T. (1999) The Burren. A Two-inchMap of the Uplands of North-west
Clare. Folding Landscapes, Roundstone.
Roche, J.R., Mitchell, F.J.G. & Waldren, S. (2009) Plant community ecology
of Pinus sylvestris, an extirpated species reintroduced to Ireland. Biodiversity
and Conservation, DOI 10.1007/s10531-009-9582-x
Roden, C. (2001) The Burren flora. The Book of the Burren, 2nd edn (eds J.W.
O’Connell &A.Korff). pp. 243–244, Tır Eolas, Kinvara.
Schettler, G., Romer, R.L., O’Connell, M. & Molloy, K. (2006) Holocene cli-
matic variations and postglacial sea-level rise geochemically recorded in the
sediments of the brackish karst lake An Loch Mor, western Ireland. Boreas,
35, 674–692.
Sheehy Skeffington, M. & Gormally, M. (2001) Introduction to the Burren.
Recognising European Pastoral Farming Systems and Understanding Their
Ecology (eds. L. Lysaght, M. Sheehy Skeffington & M. Gormally), pp. 14–
16. European Forum for Nature Conservation and Pastoralism, Bridgend
(Argyll).
Stace, C.. (1997) New Flora of the British Isles, 2nd edn. Cambridge University
Press, Cambridge.
Stockmarr, J. (1971) Tablets with spores used in absolute pollen analysis.Pollen
et Spores, 13, 615–621.
Watts, W.A. (1984) The Holocene vegetation of the Burren, western Ireland.
Lake Sediments and Environmental History (eds E.Y. Haworth & J.W.G.
Lund). pp. 359–376, Leicester University Press, Leicester.
Webb, D.A. & Scannell, M.J.P. (1983) Flora of Connemara and the Burren.
Royal Dublin Society and Cambridge University Press, Dublin and
Cambridge.
White, J. & Doyle, G. (1982) The vegetation of Ireland. A catalogue raisonne.
Studies in Irish Vegetation (ed J. White). pp. 289–368, Royal Dublin Society,
Dublin.
van der Wiel, A.M. (1982) A palaeoecological study of a section from the foot
of Hazendonk (Zuid-Holland, The Netherlands), based on the analysis of
pollen, spores and macroscopic plant remains. Review of Palaeobotany and
Palynology, 38, 35–90.
Willis, K.J., Bennett, K.D. & Birks, H.J.B. (1998) The late Quaternary dynam-
ics of pines in Europe.Ecology and Biogeography of Pinus (edD.M.Richard-
son). pp. 107–121, CambridgeUniversity Press, Cambridge.
Wilson, P., Vincent, P.J., Telfer, M.W. & Lord, T.C. (2008) Optically stimu-
lated luminescence (OSL) dating of loessic sediments and cemented scree in
northwest England.TheHolocene, 18, 1101–1112.
Yeloff, D. & van Geel, B. (2007) Abandonment of farmland and vegetation
succession following the Eurasian plague pandemic of AD 1347–1352.
Journal of Biogeography, 34, 575–582.
Received 3 February 2009; accepted 9 June 2009
Handling Editor: Richard Bradshaw
1100 I. Feeser & M. O’Connell
� 2009 The Authors. Journal compilation � 2009 British Ecological Society, Journal of Ecology, 97, 1083–1100



![[FRESH (FOR ADMISSION) - CRIMINAL CASES]](https://img.dokumen.tips/doc/110x75/63362e47e687b37e4f0607f0/fresh-for-admission-criminal-cases.jpg)

























