Embed Size (px)
Citation preview

This article was published in an Elsevier journal. The attached copyis furnished to the author for non-commercial research and
education use, including for instruction at the author’s institution,sharing with colleagues and providing to institution administration.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Fine structure of Mytella falcata (Bivalvia) gill filaments
Jose Augusto de Oliveira David a,*, Renato B. Salaroli b, Carmem S. Fontanetti a
a Departamento de Biologia, Instituto de Biociencias, Universidade Estadual Paulista, Av. 24A, no. 1515, CP 199,
CEP 13506-900, Rio Claro, SP, Brazilb NAP/MEPA, ESALQ, Universidade de Sao Paulo, Av. Padua Dias, no. 11, CP 9, CEP 13418-900, Piracicaba, SP, Brazil
Received 4 April 2007; received in revised form 9 June 2007; accepted 10 June 2007
Abstract
Bivalve filter feeders are sessile animals that live in constant contact with water and its pollutants. Their gill is an organ highly exposed to these
conditions due to its large surface and its involvement in gas exchanges and feeding. The bivalve Mytella falcata is found in estuaries of Latin
America, on the Atlantic as well as the Pacific Coast. It is commonly consumed, and sometimes is the only source of protein of low-income
communities. In this study, gill filaments of M. falcata were characterized using histology, histochemistry and transmission electron microscopy for
future comparative studies among animals exposed to environmental pollutants. Gill filaments may be divided into abfrontal, intermediate and
frontal zones. Filaments are interconnected by ciliary discs. In the center of filaments, haemocytes circulate through a haemolymph vessel
internally lined by an endothelium and supported by an acellular connective tissue rich in polysaccharides and collagen. The abfrontal zone
contains cuboidal cells, while the intermediate zone consists of a simple squamous epithelium. The frontal zone is composed of five columnar cell
types: one absorptive, mainly characterized by the presence of pinocytic vesicles in the apical region of the cell; one secretory, rarely observed; and
three ciliated with abundant mitochondria. All cells lining the filament exhibit numerous microvilli and seem to absorb substances from the
environment. PAS staining was observed in mucous cells in the frontal and abfrontal zones. Bromophenol blue allowed the distinction of
haemocytes and detection of a glycoprotein secretion in the secretory cells of the frontal region. The characteristics of M. falcata gill filaments
observed in this study were very similar to those of other bivalves, especially other Mytilidae, and are suitable for histopathological studies on the
effect of water-soluble pollutants.
# 2007 Elsevier Ltd. All rights reserved.
Keywords: Histochemistry; Polysaccharides; Protein; Collagen; Transmission electron microscopy; Gill morphology; Mussel
1. Introduction
The bivalve Mytella falcata is found on the Atlantic Coast,
from Venezuela to Argentina, as well as on Pacific shores and
the Galapagos Islands (Narchi and Galvao-Bueno, 1983).
Considered an important food item, this species is widely
consumed in several northeastern regions of Brazil and is
sometimes the only source of protein of lower-income
communities (Boffi, 1979). Species of the genus Mytella are
of great interest in biomonitoring studies, since they are found
partially buried in estuaries and thus provide data on the
conditions of the substrate (Narchi and Galvao-Bueno, 1983).
In bivalve mollusks such as the mussel Mytilus gallopro-
vincialis, the gills are key organs involved in nutrient uptake,
digestion and respiration (Gomez-Mendikute et al., 2005). The
gills of suspension-feeding bivalves are organs with a large
surface that create water currents by moving their ciliated
surface. M. falcata gills consist of two ctenidia, each formed by
two V-shaped demibranchs and the latter are composed by a
group of numerous gill filaments that can be divide into frontal
zone, intermediate zone and abfrontal zone (David and
Fontanetti, 2005).
According to Dumouhtsidou and Dimitriadis (2004), M.
galloprovincialis gill filament consists mainly of a single
layer of various types of epithelial cells (ciliated and non-
ciliated columnar cells and mucous cells) and endothelial
cells surrounding a central lumen and resting on a basement
membrane. Gomez-Mendikute et al. (2005) described, in the
same species, the cell types present in gill filaments, as
follows: in the frontal zone five cell types (columnar frontal
cells with short cilia, large eu-latero-frontal cells with cirri,
post-latero-frontal cells with no cilia, large lateral cells with
www.elsevier.com/locate/micron
Micron 39 (2008) 329–336
* Corresponding author. Tel.: +55 19 35264135; fax: +55 19 35264136.
E-mail address: [email protected] (J.A. de Oliveira David).
0968-4328/$ – see front matter # 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.micron.2007.06.002

Author's personal copy
large nuclei and long cilia and small non-ciliated post-lateral
cells); flattened endothelial cells in the intermediate zone and
ciliated and non-ciliated cells in the abfrontal zone.
Mucocytes were also identified by those authors in the
frontal and abfrontal zones, as well as lipofuscin granules in
cells of the three zones.
Due to the importance of this organ to the bivalve mollusks
and the fact that gills have a large surface constantly exposed to
water and its substances (harmful or not), the gill filaments
morphology has been extensively used as an indicator of
aquatic pollution (Gregory et al., 2002). According to Sunila
(1988), because of their simple epithelium with very
specialized cells, gill filaments are suitable for histopatholo-
gical analysis, in which the effects of water-soluble pollutants
are easily observed.
In this study, M. falcata gill filaments were characterized
for the first time using histology, histochemistry and
transmission electron microscopy to provide the framework
for future histopathological studies to detect the effect of
pollutants in the gills of this bivalve broadly distributed in
Brazilian estuaries.
2. Materials and methods
Specimens of M. falcata with approximately 4.0 cm were
collected in Santos estuary (Brazil) (23855.0520S;
46826.9750W) and transported to the laboratory, where they
were maintained in 5 L aquaria containing water of the
collecting site with constant aeration. The area where the
specimens were collected is not under the influence of
industries and can be considered as an unpolluted site. Small
gill fragments from 19 individuals were excised and fixed for
histological and histochemical analysis; for the ultrastructural
analysis gill fragments from 10 individuals were used.
Fig. 1. Histological sections of Mytella falcata gill filaments (H/E). (A) General view of gill filaments with ciliary discs; (B) view of a gill filament showing its zones
and the haemolymph vessel; (C) detail of the haemolymph vessel and the abfrontal zone; (D) detail of the frontal zone of the filament. Abbreviations: az, abfrontal
zone; cd, ciliary discs; ed, endothelium; ep, epithelium; fc, frontal cells; fz, frontal zone; g, granulocyte; iz, intermediate zone; lc, lateral cells; lfc, eu-latero-frontal
cells; v = haemolymph vessel. Scale bar in A = 20 mm and in B, C and D = 10 mm.
J.A. de Oliveira David et al. / Micron 39 (2008) 329–336330

Author's personal copy
2.1. Histology
Gill fragments were fixed in 4% paraformaldehyde in 0.1 M
phosphate buffer solution pH 7.4. The material was submitted
to the same buffer for 24 h and dehydrated in an ascending
series of ethanol baths. The material was then embedded (24 h)
in JB4 historesin with a catalyst at 4 8C to avoid premature
polymerization and placed in an incubator at 37 8C for
polymerization. Blocs were sectioned with 5 mm using the
Sorvall JB4 (Bio Rad) microtome with glass knives. Sections
were stained with haematoxylin and eosin (H/E) according to
histology routine.
2.2. Histochemistry
The material fixed in aqueous Bouin solution, was processed
according to the same histological procedures described
previously; however, to remove the picric acid residues, the
material was washed four times (5 min each) with phosphate
buffer pH 7.4 prior to the dehydration. Sections were submitted
to histochemical tests for detection of polysaccharides—PAS
(Junqueira and Junqueira, 1983), total proteins—bromophenol
blue (Pearse, 1985) and collagen—picrosirius-hematoxylin
(Junqueira and Junqueira, 1983) with some modifications, as
follows: the sections were rehydrated in water for 1 min and
submitted to the picrosirius solution at 60 8C for 60 min in an
incubator at 60 8C, the slides were washed in water, counter-
stained with hematoxylin for 5 min, washed in water for
15 min, air dried and mounted with Canada balsam.
2.3. Transmission electron microscopy
Small gill fragments were fixed in 2.5% glutaraldehyde in
0.1 M cacodylate buffer pH 7.2 at 4 8C and post-fixed in 1%
osmium tetroxide for 2 h. The material was contrasted with 2%
uranyl acetate in 10% ethanol for 4 h, dehydrated in a ascending
series of acetone, embedded in Epon-araldite with catalyst for
24 h and then placed in an incubator at 70 8C for 24 h. Ultrathin
sections were, placed in grids and the material was examined
and photographed using a transmission electron microscope
(TEM) Phillips CM 100.
3. Results
Ciliary discs composed of ciliated columnar cells inter-
connect gill filaments of M. falcata. Cilia from cells of a
filament intercalate between cilia from cells of the following
filament, thus connecting each other (Fig. 1A). The filament
may be divided into frontal, intermediate and abfrontal zones
(Fig. 1B). The frontal zone differs from other zones by the
presence of ciliated cells (Fig. 1B and D). A haemolymph
vessel, located in the central portion of the filament (Fig. 1A–
C), is internally lined by an endothelium composed of cells with
very flat nucleus. In these cells, part of the chromatin is
condensed and mainly located in the periphery of the nucleus.
Also, in the cytoplasm, mitochondria and a well-developed
Golgi complex are observed (Fig. 2A).
The intermediate zone consists of a simple squamous
epithelium (Fig. 1C) formed by epithelial cells exhibiting a flat
nucleus, abundant microvilli in their apical surface (Fig. 2B)
and many vesicles near the surface of cells (arrowhead in
Fig. 2. Electron micrographs of the intermediate zone of Mytella falcata gill
filament. (A) Endothelial cell; (B) epithelial cell; detail, vesicle; (C) detail of
junctions observed in all cells forming the gill surface. Abbreviations: aj,
adhesive junction; ct, connective tissue; ed, endothelium; ep, epithelium; gc,
Golgi complex; mv, microvilli; n, nucleus; rer, rough endoplasmic reticulum; sj,
septate junction; v, haemolymph vessel; arrowheads, vesicle. Scale bar in A and
B = 3 mm and in the detail and C = 0.3 mm.
J.A. de Oliveira David et al. / Micron 39 (2008) 329–336 331

Author's personal copy
Fig. 2B and in detail). Adhesive junctions and septate junctions
connect all cells lining gill filaments (Fig. 2C).
Between the epithelium and endothelium, there is a structure
that gives support to the haemolymph vessel and the gills. This
structure is stained with PAS (asterisk in Fig. 3A) and
picrossirius (asterisk in Fig. 3D), indicating the presence of
collagen and polysaccharides, and under TEM, this structure
exhibits fibrous elements (Fig. 2A and B). Due to its
composition this structure will be called in this study as
‘‘connective tissue’’ as discussed latter.
Inside the haemolymph vessel, haemocytes are commonly
seen and may be distinguished between granulocytes and
agranulocytes. Granulocytes are strongly stained by eosin
(Fig. 1C) and bromophenol blue (Fig. 3B), demonstrating the
presence of protein granules in the cytoplasm. Agranulocytes
are weakly stained by eosin and bromophenol blue (Fig. 3B).
Fig. 3. Mytella falcata gill filaments stained with PAS (A), bromophenol blue (B and C) and pricrosirius-haematoxylin (D). (A) General view of gill filaments
showing the position of mucous cells (strongly stained) and the connective tissue; (B) note the prominent staining for proteins in the region of basal bodies and
granulocytes, and weak stained agranulocytes; (C) detail of the frontal zone showing a secretory cell with granules stained with bromophenol blue; (D) note the
presence of collagen (red staining) in the connective tissue. Abbreviations: a, agranulocyte; az, abfrontal zone; fz, frontal zone; g, granulocyte; iz, intermediate zone;
v, haemolymph vessel; *, connective tissue; arrows, secretory cells of the frontal zone; arrowhead, basal bodies. Scale bars = 30 mm.
J.A. de Oliveira David et al. / Micron 39 (2008) 329–336332
Author's personal copy
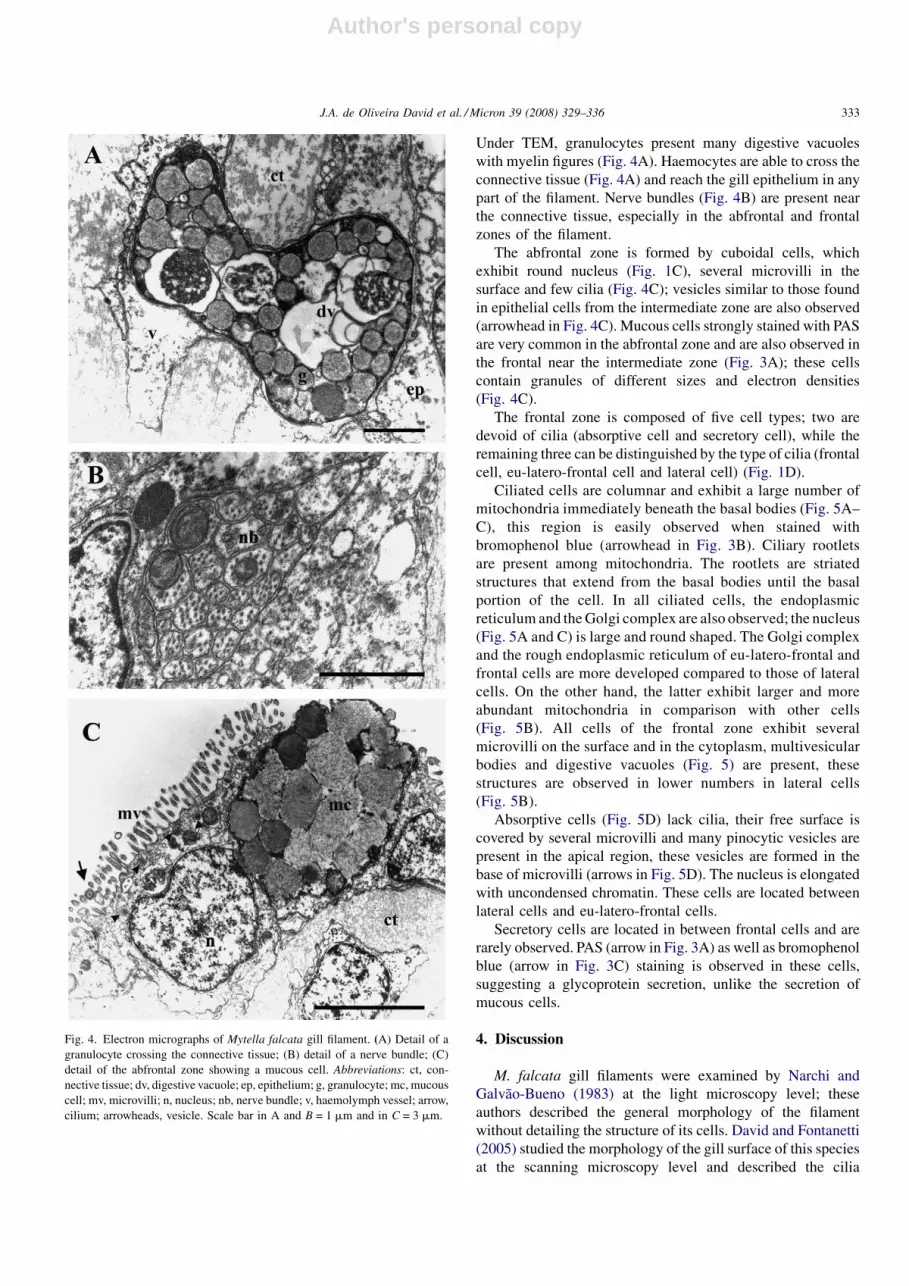
Under TEM, granulocytes present many digestive vacuoles
with myelin figures (Fig. 4A). Haemocytes are able to cross the
connective tissue (Fig. 4A) and reach the gill epithelium in any
part of the filament. Nerve bundles (Fig. 4B) are present near
the connective tissue, especially in the abfrontal and frontal
zones of the filament.
The abfrontal zone is formed by cuboidal cells, which
exhibit round nucleus (Fig. 1C), several microvilli in the
surface and few cilia (Fig. 4C); vesicles similar to those found
in epithelial cells from the intermediate zone are also observed
(arrowhead in Fig. 4C). Mucous cells strongly stained with PAS
are very common in the abfrontal zone and are also observed in
the frontal near the intermediate zone (Fig. 3A); these cells
contain granules of different sizes and electron densities
(Fig. 4C).
The frontal zone is composed of five cell types; two are
devoid of cilia (absorptive cell and secretory cell), while the
remaining three can be distinguished by the type of cilia (frontal
cell, eu-latero-frontal cell and lateral cell) (Fig. 1D).
Ciliated cells are columnar and exhibit a large number of
mitochondria immediately beneath the basal bodies (Fig. 5A–
C), this region is easily observed when stained with
bromophenol blue (arrowhead in Fig. 3B). Ciliary rootlets
are present among mitochondria. The rootlets are striated
structures that extend from the basal bodies until the basal
portion of the cell. In all ciliated cells, the endoplasmic
reticulum and the Golgi complex are also observed; the nucleus
(Fig. 5A and C) is large and round shaped. The Golgi complex
and the rough endoplasmic reticulum of eu-latero-frontal and
frontal cells are more developed compared to those of lateral
cells. On the other hand, the latter exhibit larger and more
abundant mitochondria in comparison with other cells
(Fig. 5B). All cells of the frontal zone exhibit several
microvilli on the surface and in the cytoplasm, multivesicular
bodies and digestive vacuoles (Fig. 5) are present, these
structures are observed in lower numbers in lateral cells
(Fig. 5B).
Absorptive cells (Fig. 5D) lack cilia, their free surface is
covered by several microvilli and many pinocytic vesicles are
present in the apical region, these vesicles are formed in the
base of microvilli (arrows in Fig. 5D). The nucleus is elongated
with uncondensed chromatin. These cells are located between
lateral cells and eu-latero-frontal cells.
Secretory cells are located in between frontal cells and are
rarely observed. PAS (arrow in Fig. 3A) as well as bromophenol
blue (arrow in Fig. 3C) staining is observed in these cells,
suggesting a glycoprotein secretion, unlike the secretion of
mucous cells.
4. Discussion
M. falcata gill filaments were examined by Narchi and
Galvao-Bueno (1983) at the light microscopy level; these
authors described the general morphology of the filament
without detailing the structure of its cells. David and Fontanetti
(2005) studied the morphology of the gill surface of this species
at the scanning microscopy level and described the cilia
Fig. 4. Electron micrographs of Mytella falcata gill filament. (A) Detail of a
granulocyte crossing the connective tissue; (B) detail of a nerve bundle; (C)
detail of the abfrontal zone showing a mucous cell. Abbreviations: ct, con-
nective tissue; dv, digestive vacuole; ep, epithelium; g, granulocyte; mc, mucous
cell; mv, microvilli; n, nucleus; nb, nerve bundle; v, haemolymph vessel; arrow,
cilium; arrowheads, vesicle. Scale bar in A and B = 1 mm and in C = 3 mm.
J.A. de Oliveira David et al. / Micron 39 (2008) 329–336 333

Author's personal copy
distribution along the filament. The present study is the first to
use histochemical and ultrastructural techniques to analyze the
gill filaments of M. falcata.
The shape and distribution of cilia present in the frontal
zone, observed in this study, are very similar to those observed
in other species. These cilia have an essential role in capturing
and transporting particles during feeding and, therefore, are
apparently highly conserved in the group (Owen, 1974;
Jorgensen, 1996; Gregory and George, 2000).
The ciliary movement is ATP-dependent and mitochondria
are the organelles capable of converting energy from nutrients
into ATP, providing the energy source necessary to this
process. In this study, larger and more abundant mitochondria
were observed in lateral cells. This might be associated with
their role in the surface of the gill filament. According to Owen
(1974), lateral cells are mainly responsible for water
circulation inside the bivalve body, causing its passage
between two filaments. On the other hand, Jones et al.
(1990) suggests that this role, in Mytilus edulis, may be divided
with cilia found in the abfrontal surface. When comparing the
abfrontal ciliation of M. falcata and other bivalvia species, it
was concluded that M. falcata abfrontal cilia were not detected
Fig. 5. Electron micrographs of cells in the frontal zone of Mytella falcata gill filament. (A) Eu-latero-frontal cells; (B) lateral cell; (C) frontal cell; (D) absorptive
cell. Abbreviations: bb, basal body; gc, Golgi complex; m, mitochondria; mv, microvilli; n, nucleus; pv = pinocytic vesicles; rer, rough endoplasmic reticulum; *,
multivesicular bodies and digestive vacuoles; arrow, formation of pinocytic vesicles. Scale bars = 3 mm.
J.A. de Oliveira David et al. / Micron 39 (2008) 329–336334

Author's personal copy
in numbers high enough to generate water circulation inside
the bivalve body. This idea agrees with the study of David and
Fontanetti (2005) that concluded that M. falcata abfrontal
ciliation was similar to that found in Modiolus modiolus by
Dufour and Beninger (2001) and was not sufficient to promote
water circulation.
The cytoplasm of lateral cells exhibited the least amount of
organelles, except by mitochondria, which are present in large
numbers. On the surface of these cells, few microvilli were
observed. This might be associated with the high specialization
of these cells, which play a major role in water circulation
inside the body of the organism and thus are vital for the
animals survival. Other cell types, in the frontal as well as other
zones of the filament, exhibit a cytoplasm with small pinocytic
vesicles that reflect the absorptive nature of the entire gill
epithelium. This fact is in agreement with Junqueira and
Carneiro (2005) that suggested that the role of microvilli might
be associated with the absorption of substances from the
environment into cells, as they increase surface area. Fawcett
and Porter (1954) discussed the presence and role of microvilli
on cells from the frontal surface of gill filaments and concluded
that microvilli might function as a mechanical support for an
amorphous mucous material and together, microvilli and
amorphous material, would strengthen the anchorage of cilia.
In this study, a moderate PAS staining was observed in the
surface of gill epithelium (Fig. 4A) suggesting the presence of a
polysaccharide layer that may act as a protection against
potentially harmful agents, as well as a connecting point for
particles to be ingested.
The absorptive cell between frontal cells and eu-latero-
frontal cells was called by some authors as post-latero-frontal
cell and considered without a known function (Gregory et al.,
2002; Gomez-Mendikute et al., 2005). Some authors suggested
that the absence of ciliation in these cells was important to crate
a space where lateral cilia and eu-latero-frontal cilia could carry
through their movement (Aiello and Sleigh, 1972). In the
present study, the term absorptive cell was used because this
cell presented many microvilli and pinocytic vesicles suggest-
ing material uptake from the environment.
In epithelial and abfrontal cells, the vesicles near the cell
membrane have a specific role that still needs clarification.
However, the content of these vesicles presents more electron
density when compared to mucous cells granules, excluding the
idea of a small secreting mucous vesicle.
The junctions that connect the different cell types of gill
filaments of M. falcata are structures commonly found in
invertebrates with specific roles and morphological character-
istics. The adhesive junction located in the apical portion of
cells form a ring that encircles the entire cell and acts as a
connecting point to the cytoskeleton network, increasing the a
stiffness of the brush border. The septate junctions also form a
ring around the entire cell and play a role in occlusion, not
allowing the passage of material from the environment through
intercellular spaces, and in communication and adhesion
among cells of the tissue (Satir and Gilula, 1970).
The structure supporting the haemolymph vessel has been
observed by other authors, but its constitution is not well
defined. Most authors regard it as a chitinous-like structure
(Gregory et al., 2002), but others name the structure as a
basement membrane (Dumouhtsidou and Dimitriadis, 2004). In
the present study, the characteristics observed with PAS and
picrosirius staining and the ultrastructural analysis, demon-
strated that this structure exhibits polysaccharides associated
with collagen. It was also observed that haemocytes are capable
of cross this structure and since haemocytes are not able to cross
chitinous-like layers, the term connective tissue was applied in
this study. Thus this study classified the structure as an acellular
connective tissue rich in polysaccharides and collagen that
gives sustentation to gill and the haemolymph vessel. The
thickness of this connective tissue is variable according to
friction that is subjected to gill filaments, caused by constantly
water circulation in the interfilamentar space; this is observed in
the ciliary discs region where the connective tissue is thicker
(Fig. 1A).
Acknowledgements
The authors are thankful to CAPES, CNPq and FUNDU-
NESP for financial support, to ‘‘Nautica da Ilha’’ and Marcia
M. Hoshina for helping during collecting trips, to Cristiane M.
Mileo, for the illustrations, and to Gerson Mello Souza, Monika
Iamonte and Antonio T. Yabuki for the support during technical
procedures.
References
Aiello, E., Sleigh, M.A., 1972. The metachronal wave of lateral cilia of Mytilus
edulis. J. Cell Biol. 54, 493–506.
Boffi, A.V., 1979. Moluscos brasileiros de interesse medico e economico.
HUCITEC, Sao Paulo.
David, J.A.O., Fontanetti, C.S., 2005. Surface morphology of Mytella falcata
gill filaments from three regions of the Santos Estuary. Braz. J. Morphol.
Sci. 22, 203–210.
Dufour, S.C., Beninger, P.G., 2001. A functional interpretation of cilia and
mucocyte distributions on the abfrontal surface of vivalve gills. Mar. Biol.
138, 295–309.
Dumouhtsidou, G.P., Dimitriadis, V.K., 2004. Lysosomal, tissue and cellular
alterations in the gills, palps and intestine of the mussel Mytilus gallopro-
vincialis, in relation to pollution. Mar. Biol. 145, 109–120.
Fawcett, D.W., Porter, K.R., 1954. A study of the fine structure of ciliated
epithelia. J. Morphol. 94, 221–281.
Gomez-Mendikute, A., Elizondo, M., Venier, P., Cajaraville, M.P., 2005.
Characterization of mussel gill cells in vivo and in vitro. Cell Tiss. Res.
321, 131–140.
Gregory, M.A., George, R.C., 2000. The structure and surface morphology of
gill filaments in the brown mussel Perna perna. Zool. Afr. 35, 121–129.
Gregory, M.A., Marshall, D.J., George, R.C., Anandraj, A., McClurg, T.P.,
2002. Correlations between metal uptake in the soft tissue of Perna perna
and gill filament pathology after exposure to mercury. Mar. Pollut. Bull. 45,
114–125.
Jones, H.D., Richards, O.G., Hutchinson, S., 1990. The role of ctenidial
abfrontal cilia in water pumping in Mytilus edulis L. J. Exp. Mar. Biol.
Ecol. 143, 15–26.
Jorgensen, C.B., 1996. Bivalve filter feeding revisited. Mar. Ecol. Prog. Ser.
142, 287–302.
Junqueira, L.C., Carneiro, J., 2005. Biologia Celular e Molecular. Editora
Guanabara Koogan, Rio de Janeiro.
Junqueira, L.C.U., Junqueira, M.M.S., 1983. Tecnicas basicas de citologia e
histologia. Livraria Editora Santos, Sao Paulo.
J.A. de Oliveira David et al. / Micron 39 (2008) 329–336 335

Author's personal copy
Narchi, W., Galvao-Bueno, M.S., 1983. Anatomia funcional de Mytella char-
ruana (D’Orbigny, 1846) (Bivalvia: Mytilidae). Bolm. Zool. Univ. S. Paulo
6, 113–145.
Owen, G., 1974. Studies on the gill of Mytilus edulis: the eu-latero-frontal cirri.
In: Proceedings of the Royal Society of London, vol. 187. pp. 83–91.
Pearse, A.G.E., 1985. Histolchemical. Theoretical and Applied, Churchill.
Satir, P., Gilula, N.D., 1970. The cell junction in a lamellibranch gill ciliated
epithelium: localization of pyroantimonate precipitate. J. Cell Biol. 47,
468–487.
Sunila, I., 1988. Acute histological responses of the gill of the mussel, Mytilus
edulis, to exposure by envrironmental pollutants. J. Invertebr. Pathol. 52,
137–141.
J.A. de Oliveira David et al. / Micron 39 (2008) 329–336336





























