Embed Size (px)
Citation preview

Fertility in barley flowers depends on Jekyll functions in male andfemale sporophytes
Volodymyr Radchuk1, Jochen Kumlehn1, Twan Rutten1, Nese Sreenivasulu1, Ruslana Radchuk1, Hardy
Rolletschek1, Cornelia Herrfurth2, Ivo Feussner2 and Ljudmilla Borisjuk1
1Leibniz Institute of Plant Genetics and Crop Plant Research (IPK), Corrensstrasse 3, D-06466 Gatersleben, Germany; 2Georg August University, Albrecht von Haller Institute, Department of
Plant Biochemistry, Justus-von-Liebig Weg 11, 37077 Gottingen, Germany
Author for correspondence:Ljudmilla Borisjuk
Tel: +49 039482 5 687Email: [email protected]
Received: 19 August 2011
Accepted: 30 November 2011
New Phytologist (2012) 194: 142–157doi: 10.1111/j.1469-8137.2011.04032.x
Key words: cereals, fertility, flower,gametophyte–sporophyte interactions, geneexpression, sexual reproduction.
Summary
• Owing to its evolutional plasticity and adaptability, barley (Hordeum vulgare) is one of the
most widespread crops in the world. Despite this evolutionary success, sexual reproduction of
small grain cereals is poorly investigated, making discovery of novel genes and functions a
challenging priority. Barley gene Jekyll appears to be a key player in grain development; how-
ever, its role in flowers has remained unknown.
• Here, we studied RNAi lines of barley, where Jekyll expression was repressed to different
extents. The impact of Jekyll on flower development was evaluated based on differential gene
expression analysis applied to anthers and gynoecia of wildtype and transgenic plants, as well
as using isotope labeling experiments, hormone analysis, immunogold- and TUNEL-assays
and in situ hybridization.
• Jekyll is expressed in nurse tissues mediating gametophyte–sporophyte interaction in
anthers and gynoecia, where JEKYLL was found within the intracellular membranes. The
repression of Jekyll impaired pollen maturation, anther dehiscence and induced a significant
loss of fertility. The presence of JEKYLL on the pollen surface also hints at possible involve-
ment in the fertilization process.
• We conclude that the role of Jekyll in cereal sexual reproduction is clearly much broader
than has been hitherto realized.
Introduction
Small grain cereals (wheat, barley and rye) form the foundationof most agricultural systems in temperate climates. Among these,barley (Hordeum vulgare) is one of the most widespread crops inthe world (Morrell & Clegg, 2007). The evolutional plasticityand adaptability of barley stimulate interest to sexual reproduc-tion in this cereal crop.
The plant life cycle alternates between the haploid and the dip-loid state. In barley and other angiosperms, the diploid sporo-phyte predominates, while the haploid gametophyte is typicallyreduced to a structure consisting of a few cells only. The gameto-phytes develop within specialized male (anther) and female(ovule) sporophytic organs.
The establishment of the same specialized cell types in theanther is rather conserved in monocotyledonous and dicotyle-donous species (Scott et al., 2004; Wilson & Zhang, 2009)despite some considerable differences (Zhang et al., 2011). Thegametophyte develops within a microsporangium derived fromarchesporial cells, which divide periclinally to form an inner pri-mary sporogenous and an outer primary parietal cell (Feng &
Dickinson, 2010). Meiotic division generates a tetrad of haploidmicrospores, which, after separation, undergo microsporogenesis,resulting in the production of mature pollen grains (Ma, 2005).The surrounding tissues, consisting of the tapetum, middle layer,endothecium and epidermis, are derived from the outer primaryparietal cells (Goldberg et al., 1993; Feng & Dickinson, 2010).The essential role of the tapetum for microspore developmentand maturation has been demonstrated (Scott et al., 2004; Ma,2005; Wilson & Zhang, 2009; Parish & Li, 2010; Chang et al.,2011). Control of tapetal cell identity, specification and tapetalprogrammed cell death (PCD) involves the interplay of manygenes (Feng & Dickinson, 2007, 2010; Chang et al., 2011;Zhang et al., 2011). Tapetal degeneration is especially importantfor late male reproductive development and fertility in higherplants (Ma, 2005; Zhang et al., 2011). The rice PERSISTENTTAPETAL CELL 1 (PTC1) and its Arabidopsis ortholog MALESTERILITY1 (MS1) control programmed tapetal cell degradation(Li et al., 2011). APOPTOSIS INHIBITOR 5 (API5; Li et al.,2011) and TAPETUM DEGENERATION RETADRATION(TDR; Li et al., 2006) are other regulators of tapetal PCD. Incereals, degeneration of the tapetal layer contributes to the release
Research
142 New Phytologist (2012) 194: 142–157
www.newphytologist.com� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust

of cell wall materials and other nutrients to the microspore viaformation of Ubisch bodies or orbicules (Zhang et al., 2011).The endothecium is thought to be essential for anther dehiscenceand pollen release (Ma, 2005; Yang et al., 2007). MYB26 plays acritical role in the regulation of endothecial maturation in a path-way distinct from that defined for the jasmonic acid (JA)-controlled dehiscence pathway (Yang et al., 2007). Mutations inbasic leucine-zipper transcription factors TGA9 and TGA10 leadto male sterility by abnormal stability of the middle layer and lackof endothecium and epidermis degeneration, resulting in non-dehiscent anthers (Murmu et al., 2010).
The female gametophyte differentiates from the tip cell of thenucellus and gives rise to the embryo sac (Yang et al., 2010).Early steps of anther and gynoecium development are likelypartially overlapping. As in the anthers, the SPOROCYTE-LESS ⁄ NOZZLE (SPL ⁄ NZZ) plays an essential role in definingthe identity of sporogenous cells in gynoecia (Schiefthaler et al.,1999; Yang et al., 1999). In rice, MULTIPLE SPOROCYTE 1(MSP1) forms a receptor complex with TAPETUM DETERMI-NANT1-LIKE (TDL1A; Zhao et al., 2008), similar to theTPD1-EMS1 ⁄ EXS1 complex present in the Arabidopsis anther.While the development of the female gametophyte has attractedmuch recent attention (Sundaresan & Alandete-Saez, 2010; Yanget al., 2010), the fate of its surrounding nucellus during gameto-genesis has hardly been explored, with only few reports available(Rangan & Rangaswamy, 1999; Luigi et al., 2006). The genesregulating nucellus development and degradation and the pro-cesses involved in the nucellar–embryo sac interaction are largelyunknown (Brambilla et al., 2008). Male and female sterility is animportant trait in plant breeding, and the study of its molecularbasis is of high priority (Johnson-Brousseau & McCormick,2004; Fuji & Toriyama, 2008; Watanabe, 2008; Borg et al.,2009).
A few years ago we cloned and characterized the new geneJekyll, encoding a small cysteine-rich protein which plays a keyrole in the sexual reproduction of barley (Radchuk et al., 2006).The gene is highly expressed in maternal seed tissues and isinvolve in sucrose allocation during early grain filling (Melkuset al., 2011). Jekyll transcripts are also present in flowers but atmuch lower abundances. However, the reduction of the grain setin transgenic lines with RNAi-mediated down-regulation of Jekyllexpression was more severe than anticipated, raising questions asto whether and how JEKYLL might be involved in the develop-ment of functional floral organs or in fertilization.
Materials and Methods
Plant material
Wildtype and transgenic RNAi-mediated Jekyll down-regulatedplants of barley (Hordeum vulgare L.) were grown under standardglasshouse conditions at 18�C with 16 h of light and a relativeair humidity of 60%. Transgenic plants with weak (line N18),moderate (line N61) and strong decreases (lines N91) in JekyllRNA levels were selected for the study (Radchuk et al., 2006).The determination of stages of anther development was
performed by evaluation of microspores under a confocal micro-scope as described in the Supporting Information (Fig. S1). Oneof three anthers of a flower was used for examination of thedevelopmental stage and fitness of microspores by confocalmicroscopy, while the other two anthers and a gynoecium werecollected and frozen in liquid nitrogen. At least six flowers fromsix separate spikelets were collected for each developmental stageand used for quantitative reverse transcription polymerase chainreaction (RT-PCR). For cDNA arrays and quantitative RT-PCRof the transgenic plants, anthers and gynoecia from at least 50different flowers of the same plant line were collected. For eachmicroscopic analysis, 40–60 microspores were used. Probe wasclassified as ‘aberrant phenotype’ if at least 10 microspores wereaberrant.
For the controlled crosses, the wildtype (WT) or transgenicflowers from the middle region of an isolated spike were emascu-lated while the other flowers were discarded. Emasculated andisolated flowers were hand-pollinated with transgenic or WTpollen and plants were further grown for grain maturation.Between five (for WT · WT) and 20 spikes were tested for eachcrossing combination. Fertilization rates were calculated as therelative ratio of the number of developed grains to the number ofpollinated flowers in a spike.
Histochemical techniques and microscopy
Histochemical techniques applied to flower organs as well as insitu hybridization, immunostaining and green fluorescent protein(GFP) visualization were performed as described previously(Radchuk et al., 2006). The 33P-labeled Jekyll DNA fragmentderived by PCR with the gene-specific primers 5¢-CGTGGATCCGATCTCCACAAGTGCTTCTG-3¢ and 5¢-GAAGAAGCTTAATTCTCGGCCTATACCG-3¢ was used asa probe for in situ hybridization. Immunolocalization of theJEKYLL protein was performed by polyclonal anti-Jekyll anti-bodies, which were derived previously (Radchuk et al., 2006)and additionally tested on Escherichia coli cells expressingJEKYLL (Fig. 6i). TUNEL assay was performed essentially asdescribed (Radchuk et al., 2011). For transient expression ofJEKYLL-yellow fluorescent protein (JEKYLL-YFP) in Arabidop-sis protoplasts, the coding region of Jekyll was fused in frame withthe Yellow Fluorescence Protein using primers 5¢-GCAACTC-GAGATGGCGGCTCGCGGTGGGAA-3¢ and 5¢- GAAGCTCGAGGCGACATTGAACTCGCCGTG-3¢ (the cloning sitefor Xho I is underlined), and cloned under the control of theCaMV 35S promoter. Arabidopsis protoplasts were isolated fromsuspension culture and transiently transformed with theJEKYLL-YFP construct as previously described (Ellerstrom et al.,2005). The expression of YFP alone driven under the samepromoter was used as the control. YFP signals were measured invivo with a Zeiss LSM510 META confocal laser scanning micro-scope. Preparation for electron microscopy and immunogoldlabeling was carried out according to Rutten et al. (2003), andscanning electron probes were examined in a Hitachi S4100SEM (Hisco Europe, Ratingen, Germany) at 5 kV accelerationvoltage.
NewPhytologist Research 143
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com

Biochemical procedures
For measurement of 14C-sucrose uptake, stems with spikelets werecut at the pollen developmental stage I and placed into 100 ml ofsolution containing 10 mM sucrose, 5 mM glutamine, 5 mMasparagine, 5 mM KH2PO4, 10 mM 2-(N-morpholino)ethane-sulfonic acid (MES), pH 7.0, and 500 ll [U-14C] sucrose(7.4 MBq ml)1; Amersham-Buchler, Braunschweig, Germany).Incubation was performed in the light (400 lmol m)2 s)1) dur-ing stages I–III (according to Fig. S1). After incubation, theanthers and gynoecia of WT and transgenic flowers were collectedseparately and frozen in liquid nitrogen. Subsequently, the plantmaterial was homogenized in 2 ml methanol (60%, v ⁄ v).Radioactivity was determined by a liquid scintillation counter typeWallac 1409 (Wallac, Germany) using Rotiszint eco plus (Roth,Karlsruhe, Germany). Counts were corrected for background andquenching by external standards.
Determination of phytohormones
Concentrations of phytohormones were measured as previouslydescribed (Luo et al., 2009). Plant material (500 mg) wasextracted with 0.75 ml of methanol containing 10 ng of D6-JA,30 ng of D5-oxo-phytodienoic acid (D5- oPDA), 10 ng of D4-JA-Leu each as internal standard (all three provided by OttoMiersch, Institute of Plant Biochemistry, Halle, Germany).Further procedures are described in detail in Methods S1.
cDNA array and data analysis
Total RNA was extracted from anthers and gynoecia of WT andJekyll down-regulated lines N61 and N91 (Radchuk et al., 2006)using the Gentra RNA isolation kit (Biozyme, Oldendorf,Germany). The material was collected at stages III–IV (Fig. S1).The isolated RNA was treated with RNase-free DNase, purifiedusing an RNeasy Plant Mini Kit (Qiagen) and used for the syn-thesis of 33P-dCTP (2¢-deoxycytidine 5-triphosphate)-labeledprobes. Probe preparation, hybridization and processing of12K barley seed cDNA array was done as previously described(Sreenivasulu et al., 2006) and annotations were refined based onMapMan functional categories (Sreenivasulu et al., 2008). Imagesof hybridized nylon membranes were subjected to automatic spotdetection using the MATLAB program and scored the signalintensities of 11 787 genes from the double spots, enabling us toassess two technical replications. Additionally, two biologicalrepetitions were performed using RNA from independentlygrown plants to check the biological reproducibility. Quantilenormalization was carried out on the complete data set (Bolstadet al., 2003). Fold changes between WT and transgenic probeswere calculated from two technical and two biological replicates.P-values were calculated based on a moderated t-test to detect falsepositives. Only the twofold and higher expression differences withstatistically significant regulated genes in anthers ⁄ gynoecia oftransgenic lines were selected for further analyses. The detailedset of the normalized values, fold difference, and P-values ofdifferentially expressed genes are provided in Tables S1 and S2
and the complete normalized data of all 12K genes are provided inTable S3.
Quantitative RT-PCR analyses
For quantitative RT-PCR analysis of the developing barleyanthers, poly(A) RNA was isolated from anthers and gynoecia atstages I–IV (Fig. S1) of pollen development using the DynabeadsmRNA Direct Kit (Invitrogen). A quantity of 8 ng of mRNA wastaken for linear amplification and cDNA synthesis using theExpressArt mRNA Amplification Nano Kit (Amptec GmbH,Hamburg, Germany). For quantitative RT-PCR of the transgenicanthers, 5 lg of the total RNA isolated from the same stage as thatused for cDNA arrays was used for reverse transcription by Super-Script III reverse transriptase (Invitrogen) with the oligo(dT) 20primer. The resulting cDNAs were used as a template for quanti-tative RT-PCR analyses, which were performed as described previ-ously (Radchuk et al., 2011). The efficiencies of PCRs wereestimated using the LinRegPCR software (Ramakers et al., 2003).Primer sets for each gene are listed in Table S4. All samples wererun in biological triplicates for each experiment. Dissociationcurves confirmed the presence of a single amplicon in each PCR.The Ct of each gene of interest (GOI) from each sample was nor-malized against the endogenous reference gene actin (GenBankaccession number AY145451) using the formula DCt = CtGOI –Ctactin and calculated as an arithmetic mean of the replicates.
For qRT-PCR of the developing barley anthers, DDCt valueswere calculated in comparison to the lowest signal value observedin a tissue for each gene. Fold changes were calculated accordingto Livak & Schmittgen (2001). In order to point out the relativeexpression levels in transgenic vs WT anthers, the gene expressionvalues are presented as (1 + E))DCt according to Czechowskiet al., 2005. All statistical data are presented as means ± SD andtransformed to relative gene expression (%) values, where thehighest gene expression is equal to 100%.
Accession numbers
Sequence data from this article can be found in the GenBank ⁄EMBL databases under the following accession numbers: Jekyll(AM261729), HvSERK1 (CK566938), HvMYB26 (BU998112),HvTDR (AK375074), HvDUO3 (AV924028), HvGAMYB(X87690), HvAPI5 (BU984683), HvMADS3 (BU995744) andHvPTC1 (AK373836).
Results
Down-regulation of Jekyll reduces grain set and alterscarbon uptake but does not impair jasmonate metabolism
Development of transgenic barley caryopses expressing RNAi-mediated Jekyll down-regulation (e.g. lines N18, N61 and N91)is largely disturbed, resulting in reduction of yield (Radchuket al., 2006). Reciprocal crosses of Jekyll-repressed plants withWT were performed here to investigate whether down-regulationof Jekyll expression might impair seed set. First, we performedreciprocal crosses between WT and Jekyll down-regulated plant
144 Research
NewPhytologist
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com
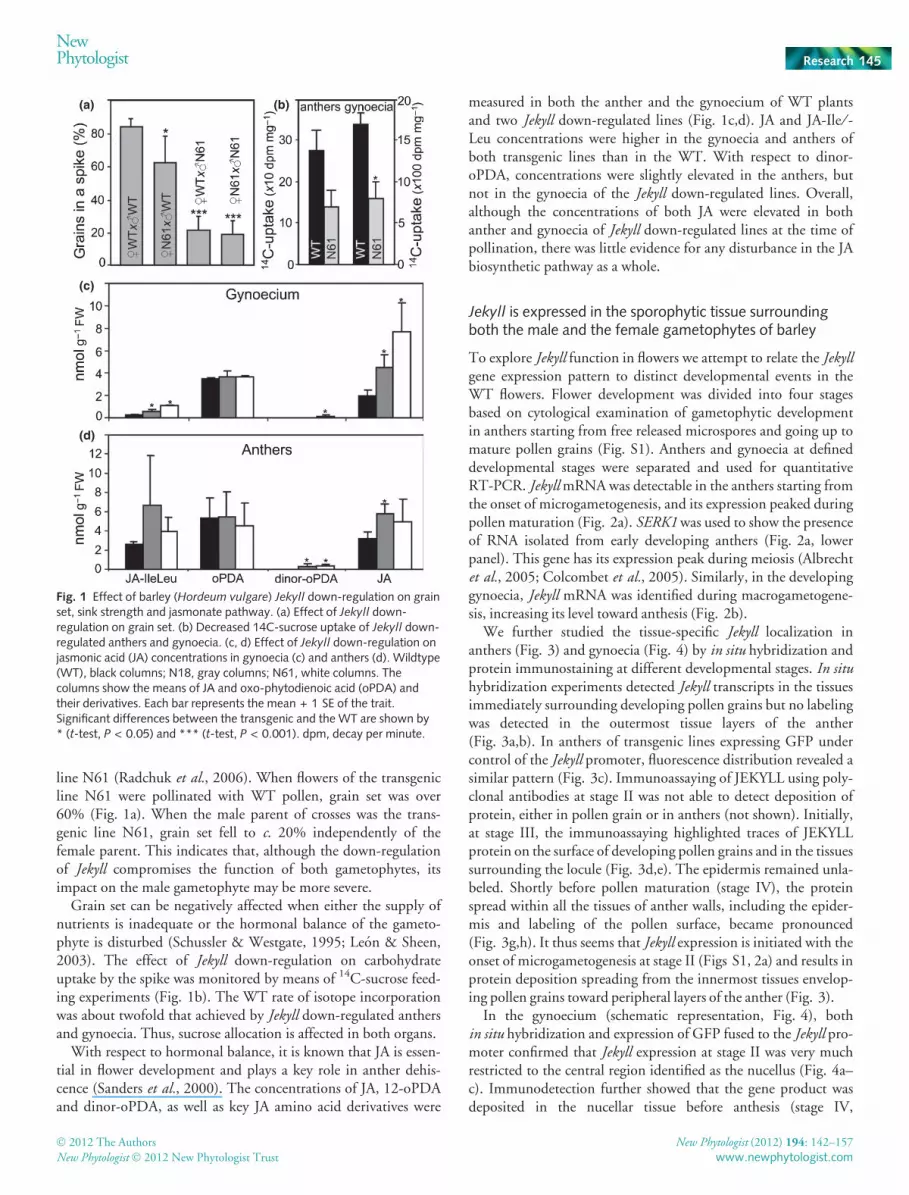
line N61 (Radchuk et al., 2006). When flowers of the transgenicline N61 were pollinated with WT pollen, grain set was over60% (Fig. 1a). When the male parent of crosses was the trans-genic line N61, grain set fell to c. 20% independently of thefemale parent. This indicates that, although the down-regulationof Jekyll compromises the function of both gametophytes, itsimpact on the male gametophyte may be more severe.
Grain set can be negatively affected when either the supply ofnutrients is inadequate or the hormonal balance of the gameto-phyte is disturbed (Schussler & Westgate, 1995; Leon & Sheen,2003). The effect of Jekyll down-regulation on carbohydrateuptake by the spike was monitored by means of 14C-sucrose feed-ing experiments (Fig. 1b). The WT rate of isotope incorporationwas about twofold that achieved by Jekyll down-regulated anthersand gynoecia. Thus, sucrose allocation is affected in both organs.
With respect to hormonal balance, it is known that JA is essen-tial in flower development and plays a key role in anther dehis-cence (Sanders et al., 2000). The concentrations of JA, 12-oPDAand dinor-oPDA, as well as key JA amino acid derivatives were
measured in both the anther and the gynoecium of WT plantsand two Jekyll down-regulated lines (Fig. 1c,d). JA and JA-Ile ⁄ -Leu concentrations were higher in the gynoecia and anthers ofboth transgenic lines than in the WT. With respect to dinor-oPDA, concentrations were slightly elevated in the anthers, butnot in the gynoecia of the Jekyll down-regulated lines. Overall,although the concentrations of both JA were elevated in bothanther and gynoecia of Jekyll down-regulated lines at the time ofpollination, there was little evidence for any disturbance in the JAbiosynthetic pathway as a whole.
Jekyll is expressed in the sporophytic tissue surroundingboth the male and the female gametophytes of barley
To explore Jekyll function in flowers we attempt to relate the Jekyllgene expression pattern to distinct developmental events in theWT flowers. Flower development was divided into four stagesbased on cytological examination of gametophytic developmentin anthers starting from free released microspores and going up tomature pollen grains (Fig. S1). Anthers and gynoecia at defineddevelopmental stages were separated and used for quantitativeRT-PCR. Jekyll mRNA was detectable in the anthers starting fromthe onset of microgametogenesis, and its expression peaked duringpollen maturation (Fig. 2a). SERK1 was used to show the presenceof RNA isolated from early developing anthers (Fig. 2a, lowerpanel). This gene has its expression peak during meiosis (Albrechtet al., 2005; Colcombet et al., 2005). Similarly, in the developinggynoecia, Jekyll mRNA was identified during macrogametogene-sis, increasing its level toward anthesis (Fig. 2b).
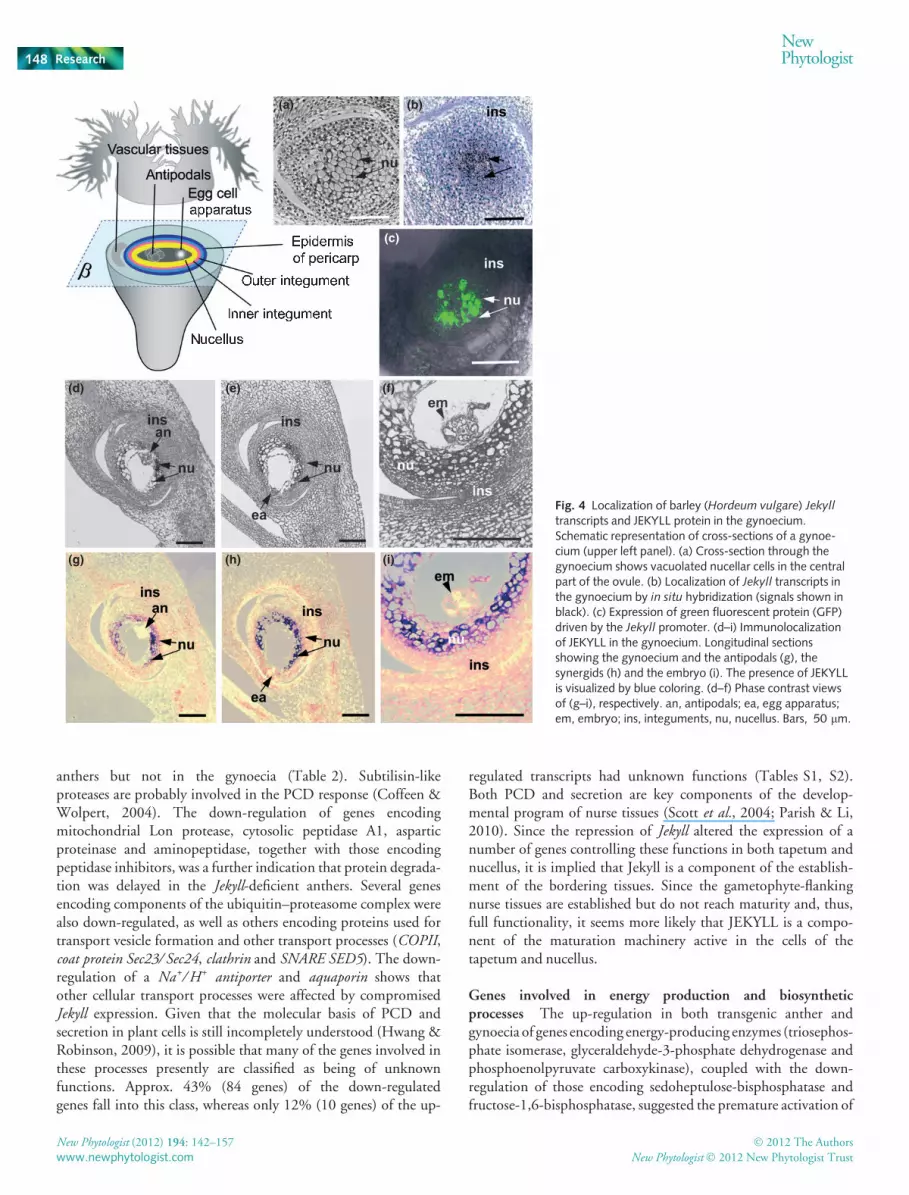
We further studied the tissue-specific Jekyll localization inanthers (Fig. 3) and gynoecia (Fig. 4) by in situ hybridization andprotein immunostaining at different developmental stages. In situhybridization experiments detected Jekyll transcripts in the tissuesimmediately surrounding developing pollen grains but no labelingwas detected in the outermost tissue layers of the anther(Fig. 3a,b). In anthers of transgenic lines expressing GFP undercontrol of the Jekyll promoter, fluorescence distribution revealed asimilar pattern (Fig. 3c). Immunoassaying of JEKYLL using poly-clonal antibodies at stage II was not able to detect deposition ofprotein, either in pollen grain or in anthers (not shown). Initially,at stage III, the immunoassaying highlighted traces of JEKYLLprotein on the surface of developing pollen grains and in the tissuessurrounding the locule (Fig. 3d,e). The epidermis remained unla-beled. Shortly before pollen maturation (stage IV), the proteinspread within all the tissues of anther walls, including the epider-mis and labeling of the pollen surface, became pronounced(Fig. 3g,h). It thus seems that Jekyll expression is initiated with theonset of microgametogenesis at stage II (Figs S1, 2a) and results inprotein deposition spreading from the innermost tissues envelop-ing pollen grains toward peripheral layers of the anther (Fig. 3).
In the gynoecium (schematic representation, Fig. 4), bothin situ hybridization and expression of GFP fused to the Jekyll pro-moter confirmed that Jekyll expression at stage II was very muchrestricted to the central region identified as the nucellus (Fig. 4a–c). Immunodetection further showed that the gene product wasdeposited in the nucellar tissue before anthesis (stage IV,
(a)
(c)
(d)
(b)
Fig. 1 Effect of barley (Hordeum vulgare) Jekyll down-regulation on grainset, sink strength and jasmonate pathway. (a) Effect of Jekyll down-regulation on grain set. (b) Decreased 14C-sucrose uptake of Jekyll down-regulated anthers and gynoecia. (c, d) Effect of Jekyll down-regulation onjasmonic acid (JA) concentrations in gynoecia (c) and anthers (d). Wildtype(WT), black columns; N18, gray columns; N61, white columns. Thecolumns show the means of JA and oxo-phytodienoic acid (oPDA) andtheir derivatives. Each bar represents the mean + 1 SE of the trait.Significant differences between the transgenic and the WT are shown by* (t-test, P < 0.05) and *** (t-test, P < 0.001). dpm, decay per minute.
NewPhytologist Research 145
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com
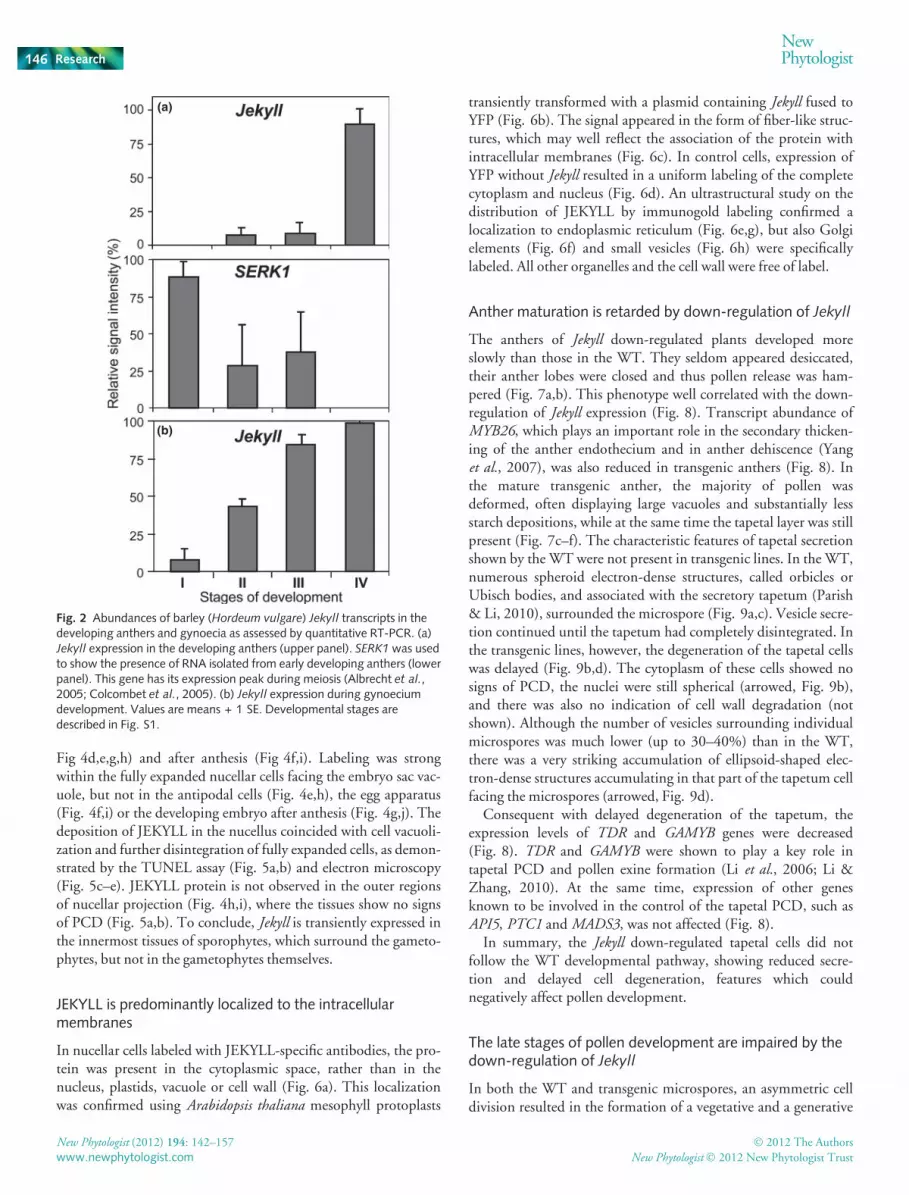
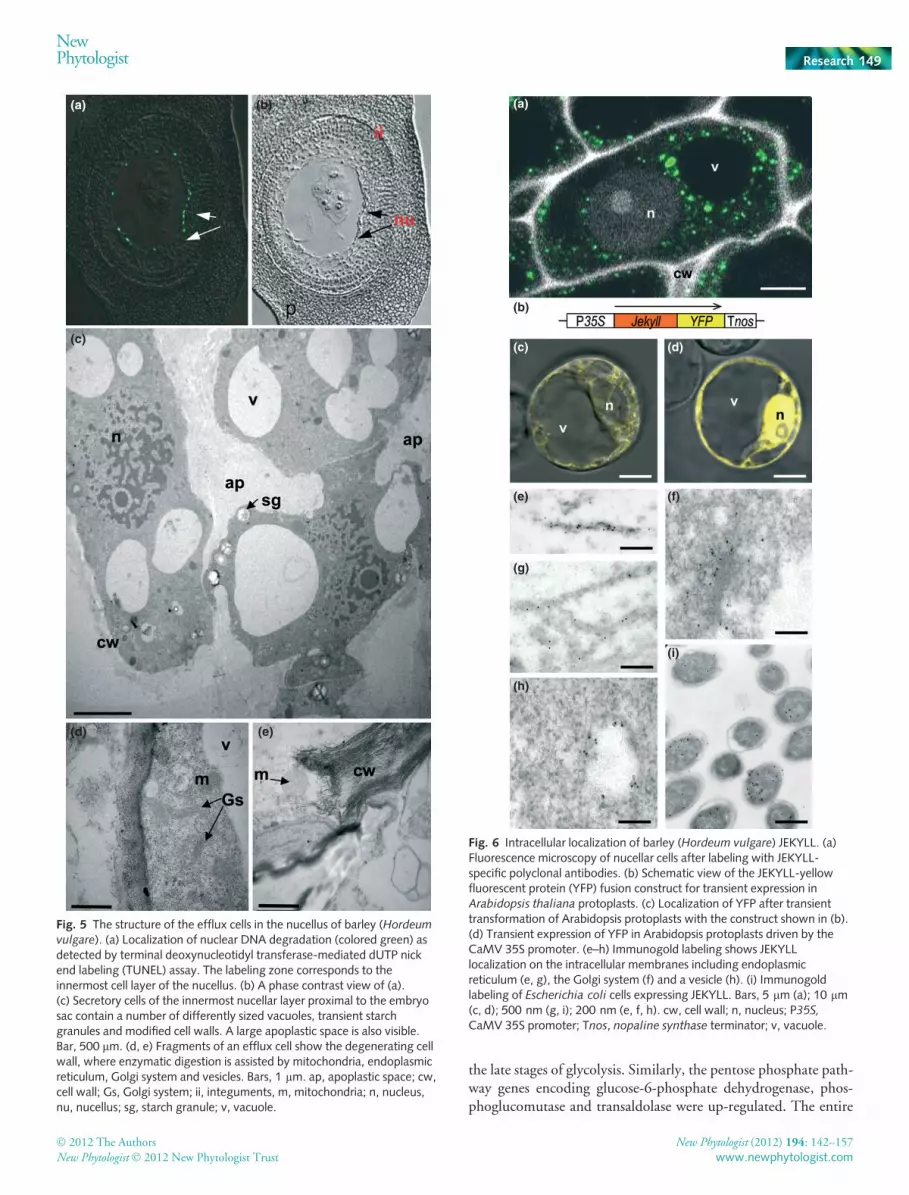
Fig 4d,e,g,h) and after anthesis (Fig 4f,i). Labeling was strongwithin the fully expanded nucellar cells facing the embryo sac vac-uole, but not in the antipodal cells (Fig. 4e,h), the egg apparatus(Fig. 4f,i) or the developing embryo after anthesis (Fig. 4g,j). Thedeposition of JEKYLL in the nucellus coincided with cell vacuoli-zation and further disintegration of fully expanded cells, as demon-strated by the TUNEL assay (Fig. 5a,b) and electron microscopy(Fig. 5c–e). JEKYLL protein is not observed in the outer regionsof nucellar projection (Fig. 4h,i), where the tissues show no signsof PCD (Fig. 5a,b). To conclude, Jekyll is transiently expressed inthe innermost tissues of sporophytes, which surround the gameto-phytes, but not in the gametophytes themselves.
JEKYLL is predominantly localized to the intracellularmembranes
In nucellar cells labeled with JEKYLL-specific antibodies, the pro-tein was present in the cytoplasmic space, rather than in thenucleus, plastids, vacuole or cell wall (Fig. 6a). This localizationwas confirmed using Arabidopsis thaliana mesophyll protoplasts
transiently transformed with a plasmid containing Jekyll fused toYFP (Fig. 6b). The signal appeared in the form of fiber-like struc-tures, which may well reflect the association of the protein withintracellular membranes (Fig. 6c). In control cells, expression ofYFP without Jekyll resulted in a uniform labeling of the completecytoplasm and nucleus (Fig. 6d). An ultrastructural study on thedistribution of JEKYLL by immunogold labeling confirmed alocalization to endoplasmic reticulum (Fig. 6e,g), but also Golgielements (Fig. 6f) and small vesicles (Fig. 6h) were specificallylabeled. All other organelles and the cell wall were free of label.
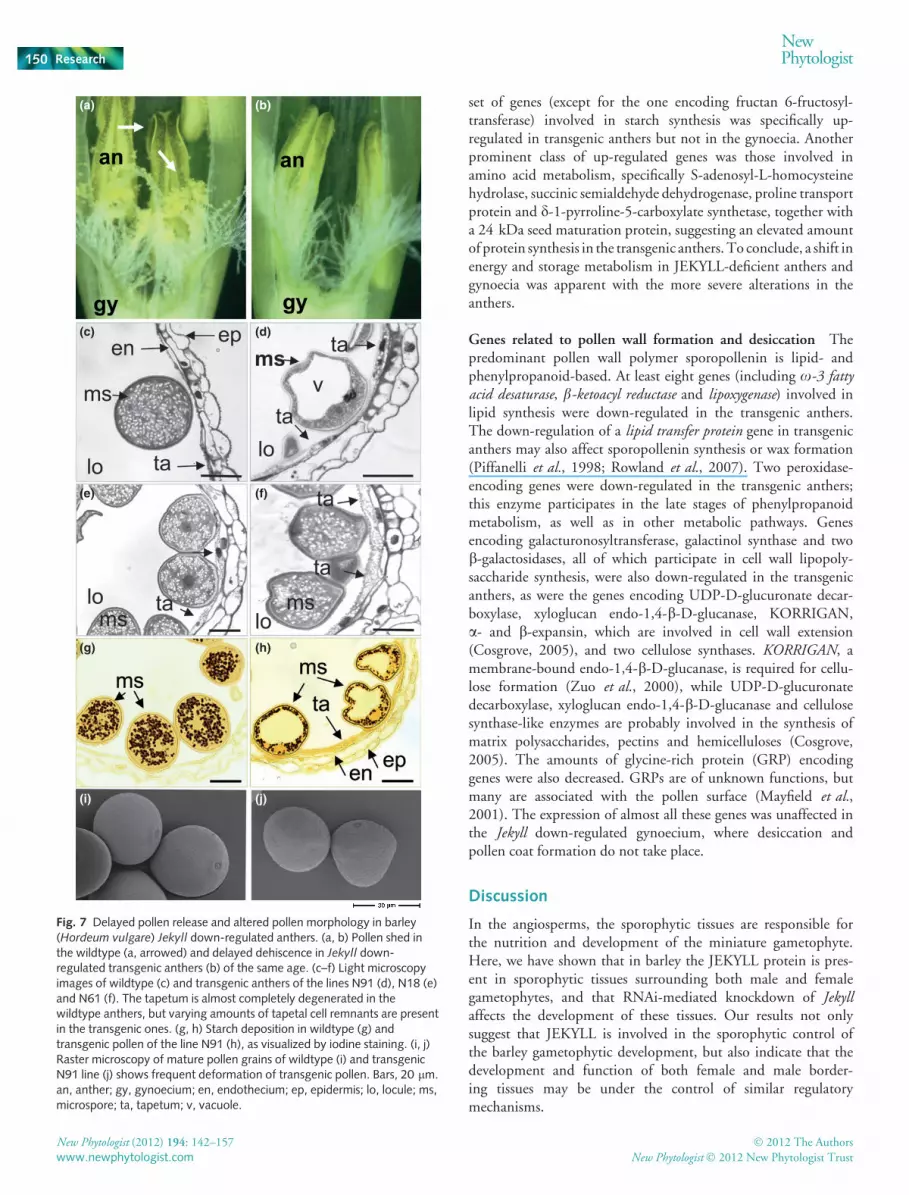
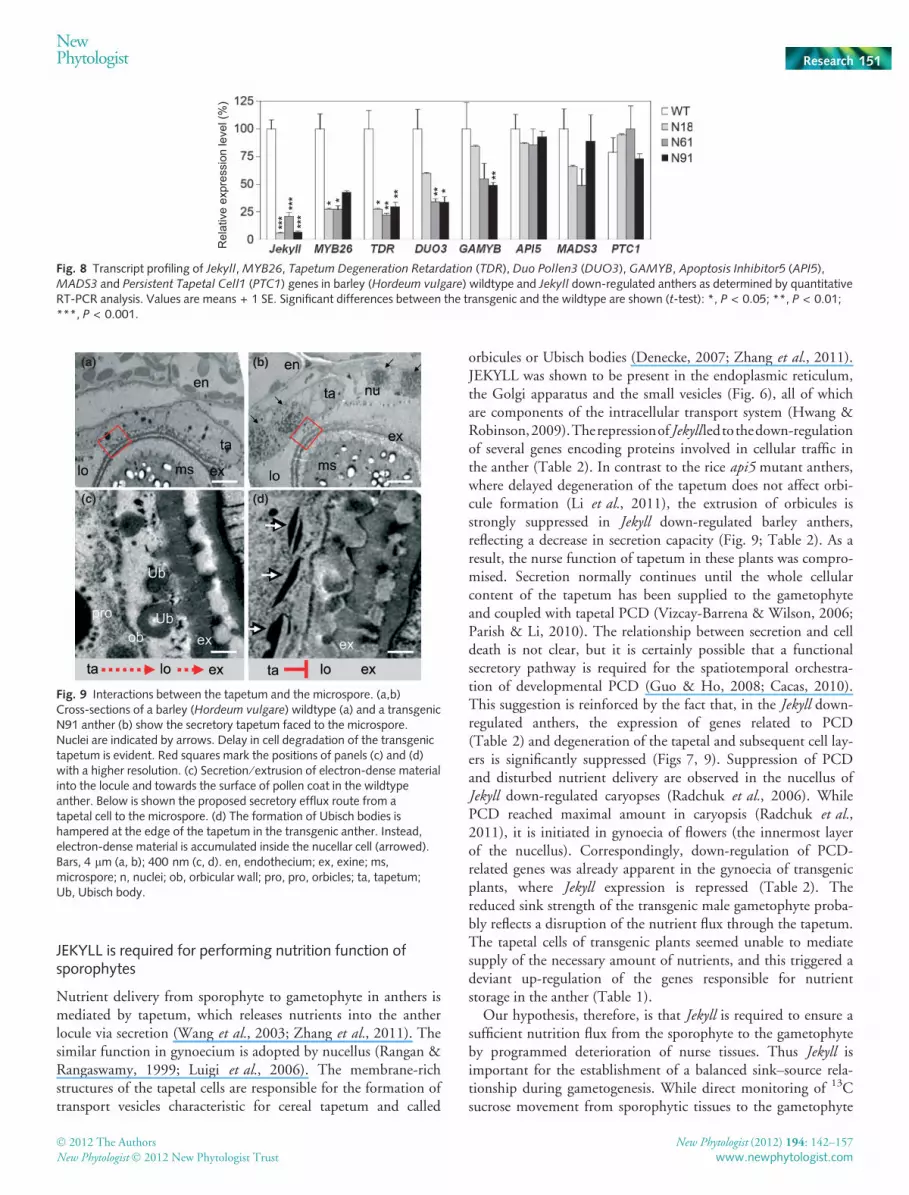
Anther maturation is retarded by down-regulation of Jekyll
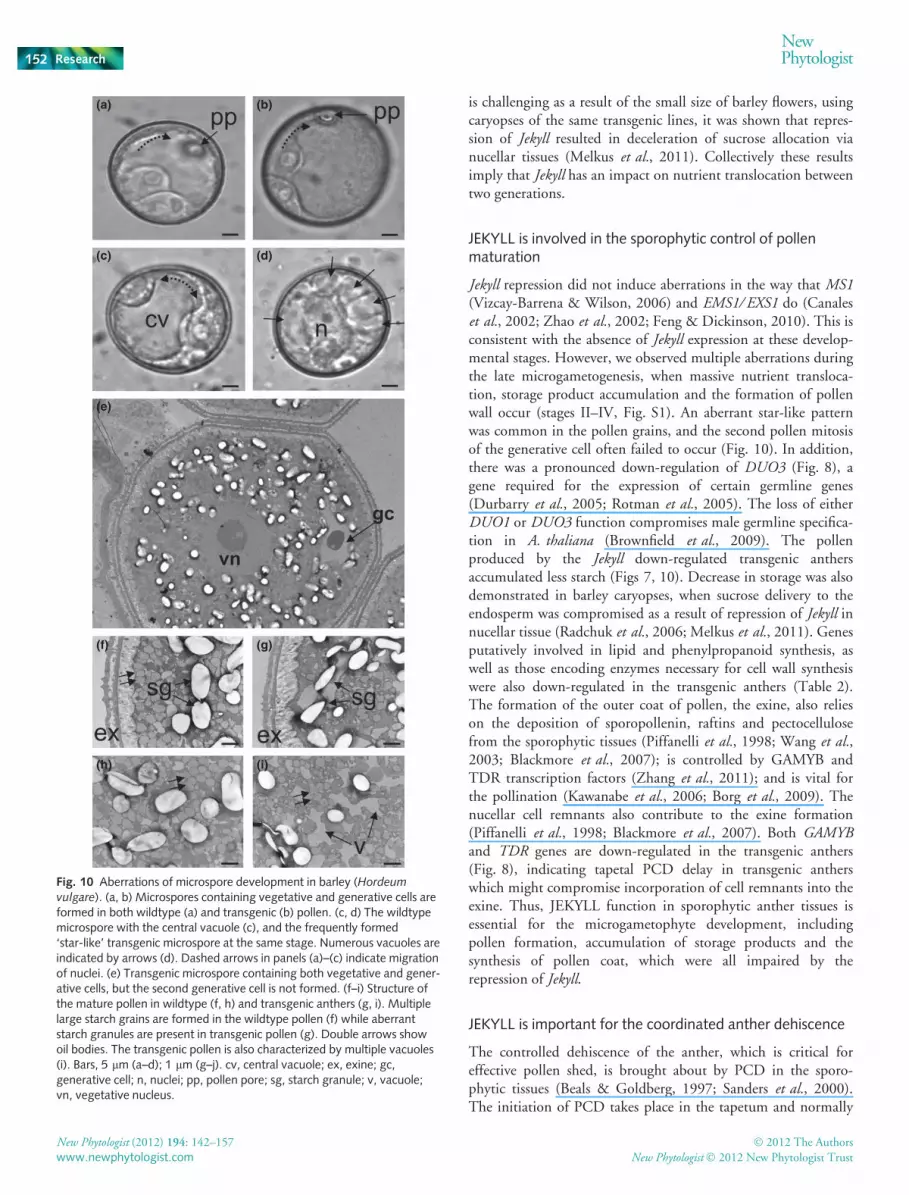
The anthers of Jekyll down-regulated plants developed moreslowly than those in the WT. They seldom appeared desiccated,their anther lobes were closed and thus pollen release was ham-pered (Fig. 7a,b). This phenotype well correlated with the down-regulation of Jekyll expression (Fig. 8). Transcript abundance ofMYB26, which plays an important role in the secondary thicken-ing of the anther endothecium and in anther dehiscence (Yanget al., 2007), was also reduced in transgenic anthers (Fig. 8). Inthe mature transgenic anther, the majority of pollen wasdeformed, often displaying large vacuoles and substantially lessstarch depositions, while at the same time the tapetal layer was stillpresent (Fig. 7c–f). The characteristic features of tapetal secretionshown by the WT were not present in transgenic lines. In the WT,numerous spheroid electron-dense structures, called orbicles orUbisch bodies, and associated with the secretory tapetum (Parish& Li, 2010), surrounded the microspore (Fig. 9a,c). Vesicle secre-tion continued until the tapetum had completely disintegrated. Inthe transgenic lines, however, the degeneration of the tapetal cellswas delayed (Fig. 9b,d). The cytoplasm of these cells showed nosigns of PCD, the nuclei were still spherical (arrowed, Fig. 9b),and there was also no indication of cell wall degradation (notshown). Although the number of vesicles surrounding individualmicrospores was much lower (up to 30–40%) than in the WT,there was a very striking accumulation of ellipsoid-shaped elec-tron-dense structures accumulating in that part of the tapetum cellfacing the microspores (arrowed, Fig. 9d).
Consequent with delayed degeneration of the tapetum, theexpression levels of TDR and GAMYB genes were decreased(Fig. 8). TDR and GAMYB were shown to play a key role intapetal PCD and pollen exine formation (Li et al., 2006; Li &Zhang, 2010). At the same time, expression of other genesknown to be involved in the control of the tapetal PCD, such asAPI5, PTC1 and MADS3, was not affected (Fig. 8).
In summary, the Jekyll down-regulated tapetal cells did notfollow the WT developmental pathway, showing reduced secre-tion and delayed cell degeneration, features which couldnegatively affect pollen development.
The late stages of pollen development are impaired by thedown-regulation of Jekyll
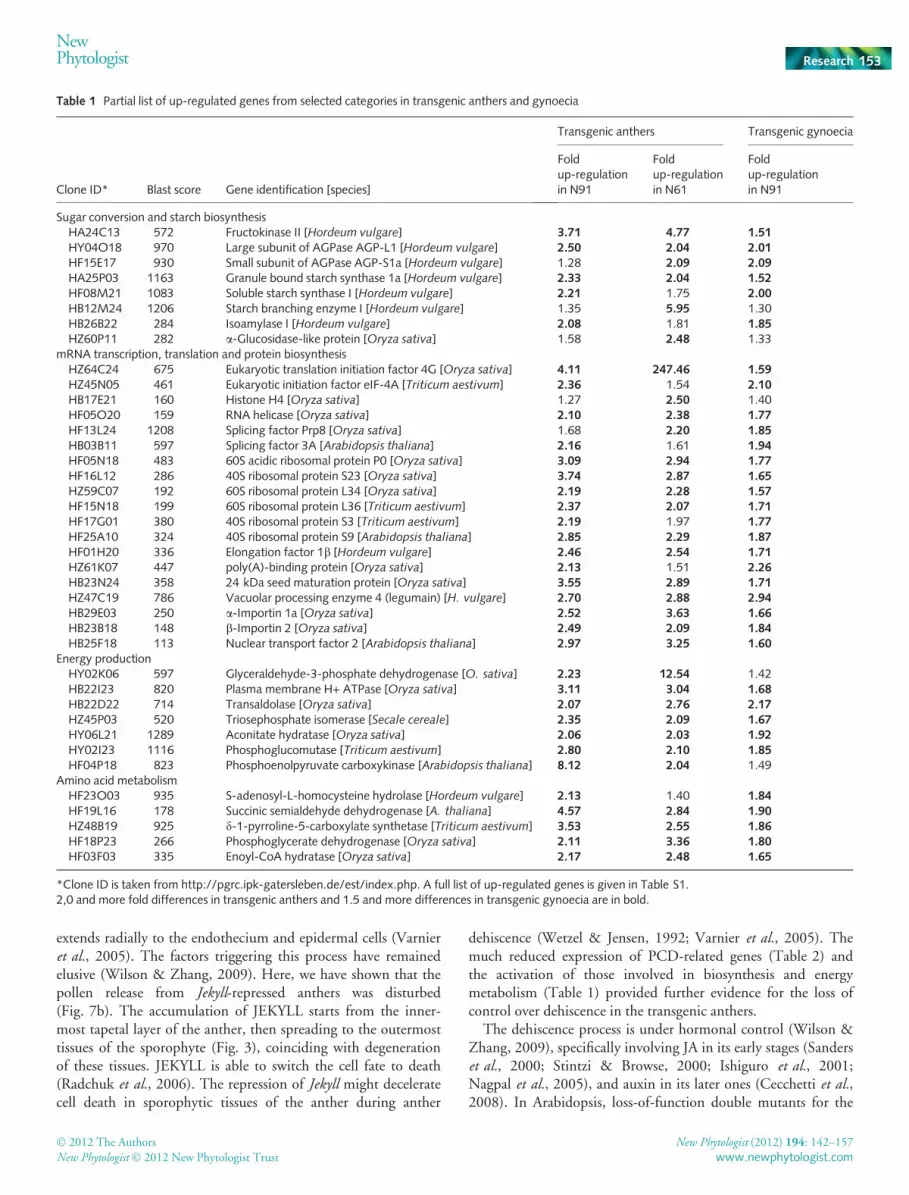
In both the WT and transgenic microspores, an asymmetric celldivision resulted in the formation of a vegetative and a generative
(a)
(b)
Fig. 2 Abundances of barley (Hordeum vulgare) Jekyll transcripts in thedeveloping anthers and gynoecia as assessed by quantitative RT-PCR. (a)Jekyll expression in the developing anthers (upper panel). SERK1 was usedto show the presence of RNA isolated from early developing anthers (lowerpanel). This gene has its expression peak during meiosis (Albrecht et al.,2005; Colcombet et al., 2005). (b) Jekyll expression during gynoeciumdevelopment. Values are means + 1 SE. Developmental stages aredescribed in Fig. S1.
146 Research
NewPhytologist
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com

cell. The nucleus of the vegetative cell eventually migrated to thepollen pore. In Jekyll down-regulated plants, however, the vegeta-tive and generative nuclei tended to remain in close proximity,and some pollen grains acquired a star-like pattern (Fig. 10a–d)indicative of stress and ⁄ or the acquisition of embryogenic compe-tence (Maraschin et al., 2005). In the WT, the generative cellunderwent a mitotic division to form two sperm cells, but in thetransgenic plants, some pollen grains retained a single generativecell rather than producing two sperms (Fig. 10e). In Jekyll down-regulated anthers, the expression of the barley homolog of theDUO3 transcription factor, known to be a regulator of generativecell division and sperm cell specification in A. thaliana (Brown-field et al., 2009), was reduced (Fig. 8). Mature WT pollengrains contained starch granules and oil bodies (Fig. 9f,h), butthese were sparse in the transgenic pollen (Fig. 9g,i) with thestarch granules being smaller and seemingly deformed (Fig. 10g).Furthermore, transgenic pollen contained more vacuoles(Fig. 10i) and Golgi bodies (not shown), which are characteristicof a less advanced stage of development. As a result, the trans-genic pollen was not adapted to the desiccation process takingplace in the context of anthesis.
Transcript profiling of the Jekyll-deficient anthers andgynoecia
A 12K handmade cDNA array (Sreenivasulu et al., 2006) wasused to characterize the transcriptome in the anthers of WT andthe two transgenic lines N61 and N91, while only the line N91was used to compare the expression profiles of the gynoecium ofWT and transgenic lines (Tables 1, 2, S1 and S2). Anthers andgynoecia from stages III–IV (which correspond to the maximumof Jekyll expression) were collected and two biological replicates
with two technical replicates each were performed. Transcriptabundances are given as ratios between WT and correspondingtransgenic tissues. Of 12 000 cDNA clones present on the cDNAarray filter, 199 were down-regulated and 91 were up-regulated(Tables S1, S2). The largest group of up-regulated genes withknown function belongs to the transcription and translationgroups (37 genes) followed by sugar conversion and starch bio-synthesis (11 genes), as well amino acid metabolism (eight genes)and energy production (eight genes) (Fig. S1). Down-regulatedgenes were predominantly enriched by PCD-related genes (14genes), cell wall synthesis and modification (11 genes), lipidsynthesis (nine genes), and others (Table S2).
Genes regulating transcription and translation The largestgroup of up-regulated transcripts in both transgenic anthers andgynoecia was formed by genes involved in general transcriptionand translation (Table 1). This group includes genes encoding H4histones, RNA helicase and splicing factors, a- and b-importins,translation initiation factors, various ribosomal proteins and aputative nuclear transport protein. By contrast, DNA methyltrans-ferase and methionine synthase genes were down-regulated(Table 2). Most of the methionine produced by methionine syn-thase is used for DNA and histone transmethylation (Ravanelet al., 1998), suggesting that general patterns of gene expressionwere modified in the Jekyll down-regulated organs, a conclusionsupported by the down-regulation of JmjC domain-containinghistone lysine demethylase and histone acetyl transferase. In rice,histone lysine demethylase is important for the development of thefloral organs (Sun & Zhou, 2008).
Genes involved in secretion and PCD Subtilisin-like serineprotease encoding genes were down-regulated in the transgenic
(a)
(f)
(b) (c)
(d) (e)
(g) (h)
Fig. 3 Localization of barley (Hordeum vulgare) Jekyll
transcripts and JEKYLL protein in the anther.(a) Schematic representation of longitudinal sections andcross-sections of an anther. (b) Localization of Jekyll
transcripts in the anther by in situ hybridization(longitudinal section of anther, stage II; signals shown inblack). (c) Expression of green fluorescent protein (GFP)driven by the Jekyll promoter (longitudinal section ofanther, stage II). (d, e) Immunolocalization of JEKYLL inanthers (stage III) and longitudinal tissue section usedfor immunoassaying (phase contrast views: d).(f) Cross-section of locule showing microspores attachedto the tapetum and surrounded by endothecium, middlelayer and epidermis. (g, h) Immunolocalization of JEKYLLin anthers (stage IV) appears blue (g, arrowed) andcross-section shows tissue used for immunoassaying (h,phase contrast views). Bars, 30 lm (b–e, h); 20 lm (f).en, endothecium; ep, epidermis; ms, microspore; md,middle layer; ta, tapetum.
NewPhytologist Research 147
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com
anthers but not in the gynoecia (Table 2). Subtilisin-likeproteases are probably involved in the PCD response (Coffeen &Wolpert, 2004). The down-regulation of genes encodingmitochondrial Lon protease, cytosolic peptidase A1, asparticproteinase and aminopeptidase, together with those encodingpeptidase inhibitors, was a further indication that protein degrada-tion was delayed in the Jekyll-deficient anthers. Several genesencoding components of the ubiquitin–proteasome complex werealso down-regulated, as well as others encoding proteins used fortransport vesicle formation and other transport processes (COPII,coat protein Sec23 ⁄ Sec24, clathrin and SNARE SED5). The down-regulation of a Na+ ⁄ H+ antiporter and aquaporin shows thatother cellular transport processes were affected by compromisedJekyll expression. Given that the molecular basis of PCD andsecretion in plant cells is still incompletely understood (Hwang &Robinson, 2009), it is possible that many of the genes involved inthese processes presently are classified as being of unknownfunctions. Approx. 43% (84 genes) of the down-regulatedgenes fall into this class, whereas only 12% (10 genes) of the up-
regulated transcripts had unknown functions (Tables S1, S2).Both PCD and secretion are key components of the develop-mental program of nurse tissues (Scott et al., 2004; Parish & Li,2010). Since the repression of Jekyll altered the expression of anumber of genes controlling these functions in both tapetum andnucellus, it is implied that Jekyll is a component of the establish-ment of the bordering tissues. Since the gametophyte-flankingnurse tissues are established but do not reach maturity and, thus,full functionality, it seems more likely that JEKYLL is a compo-nent of the maturation machinery active in the cells of thetapetum and nucellus.
Genes involved in energy production and biosyntheticprocesses The up-regulation in both transgenic anther andgynoecia of genes encoding energy-producing enzymes (triosephos-phate isomerase, glyceraldehyde-3-phosphate dehydrogenase andphosphoenolpyruvate carboxykinase), coupled with the down-regulation of those encoding sedoheptulose-bisphosphatase andfructose-1,6-bisphosphatase, suggested the premature activation of
(a) (b)
(c)
(d) (e) (f)
(g) (h) (i)
Fig. 4 Localization of barley (Hordeum vulgare) Jekyll
transcripts and JEKYLL protein in the gynoecium.Schematic representation of cross-sections of a gynoe-cium (upper left panel). (a) Cross-section through thegynoecium shows vacuolated nucellar cells in the centralpart of the ovule. (b) Localization of Jekyll transcripts inthe gynoecium by in situ hybridization (signals shown inblack). (c) Expression of green fluorescent protein (GFP)driven by the Jekyll promoter. (d–i) Immunolocalizationof JEKYLL in the gynoecium. Longitudinal sectionsshowing the gynoecium and the antipodals (g), thesynergids (h) and the embryo (i). The presence of JEKYLLis visualized by blue coloring. (d–f) Phase contrast viewsof (g–i), respectively. an, antipodals; ea, egg apparatus;em, embryo; ins, integuments, nu, nucellus. Bars, 50 lm.
148 Research
NewPhytologist
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com
the late stages of glycolysis. Similarly, the pentose phosphate path-way genes encoding glucose-6-phosphate dehydrogenase, phos-phoglucomutase and transaldolase were up-regulated. The entire
(a)
(c)
(d) (e)
(b)
Fig. 5 The structure of the efflux cells in the nucellus of barley (Hordeumvulgare). (a) Localization of nuclear DNA degradation (colored green) asdetected by terminal deoxynucleotidyl transferase-mediated dUTP nickend labeling (TUNEL) assay. The labeling zone corresponds to theinnermost cell layer of the nucellus. (b) A phase contrast view of (a).(c) Secretory cells of the innermost nucellar layer proximal to the embryosac contain a number of differently sized vacuoles, transient starchgranules and modified cell walls. A large apoplastic space is also visible.Bar, 500 lm. (d, e) Fragments of an efflux cell show the degenerating cellwall, where enzymatic digestion is assisted by mitochondria, endoplasmicreticulum, Golgi system and vesicles. Bars, 1 lm. ap, apoplastic space; cw,cell wall; Gs, Golgi system; ii, integuments, m, mitochondria; n, nucleus,nu, nucellus; sg, starch granule; v, vacuole.
(a)
(b)
(c) (d)
(e) (f)
(g)
(h)
(i)
Fig. 6 Intracellular localization of barley (Hordeum vulgare) JEKYLL. (a)Fluorescence microscopy of nucellar cells after labeling with JEKYLL-specific polyclonal antibodies. (b) Schematic view of the JEKYLL-yellowfluorescent protein (YFP) fusion construct for transient expression inArabidopsis thaliana protoplasts. (c) Localization of YFP after transienttransformation of Arabidopsis protoplasts with the construct shown in (b).(d) Transient expression of YFP in Arabidopsis protoplasts driven by theCaMV 35S promoter. (e–h) Immunogold labeling shows JEKYLLlocalization on the intracellular membranes including endoplasmicreticulum (e, g), the Golgi system (f) and a vesicle (h). (i) Immunogoldlabeling of Escherichia coli cells expressing JEKYLL. Bars, 5 lm (a); 10 lm(c, d); 500 nm (g, i); 200 nm (e, f, h). cw, cell wall; n, nucleus; P35S,
CaMV 35S promoter; Tnos, nopaline synthase terminator; v, vacuole.
NewPhytologist Research 149
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com
set of genes (except for the one encoding fructan 6-fructosyl-transferase) involved in starch synthesis was specifically up-regulated in transgenic anthers but not in the gynoecia. Anotherprominent class of up-regulated genes was those involved inamino acid metabolism, specifically S-adenosyl-L-homocysteinehydrolase, succinic semialdehyde dehydrogenase, proline transportprotein and d-1-pyrroline-5-carboxylate synthetase, together witha 24 kDa seed maturation protein, suggesting an elevated amountof protein synthesis in the transgenic anthers. To conclude, a shift inenergy and storage metabolism in JEKYLL-deficient anthers andgynoecia was apparent with the more severe alterations in theanthers.
Genes related to pollen wall formation and desiccation Thepredominant pollen wall polymer sporopollenin is lipid- andphenylpropanoid-based. At least eight genes (including x-3 fattyacid desaturase, b-ketoacyl reductase and lipoxygenase) involved inlipid synthesis were down-regulated in the transgenic anthers.The down-regulation of a lipid transfer protein gene in transgenicanthers may also affect sporopollenin synthesis or wax formation(Piffanelli et al., 1998; Rowland et al., 2007). Two peroxidase-encoding genes were down-regulated in the transgenic anthers;this enzyme participates in the late stages of phenylpropanoidmetabolism, as well as in other metabolic pathways. Genesencoding galacturonosyltransferase, galactinol synthase and twob-galactosidases, all of which participate in cell wall lipopoly-saccharide synthesis, were also down-regulated in the transgenicanthers, as were the genes encoding UDP-D-glucuronate decar-boxylase, xyloglucan endo-1,4-b-D-glucanase, KORRIGAN,a- and b-expansin, which are involved in cell wall extension(Cosgrove, 2005), and two cellulose synthases. KORRIGAN, amembrane-bound endo-1,4-b-D-glucanase, is required for cellu-lose formation (Zuo et al., 2000), while UDP-D-glucuronatedecarboxylase, xyloglucan endo-1,4-b-D-glucanase and cellulosesynthase-like enzymes are probably involved in the synthesis ofmatrix polysaccharides, pectins and hemicelluloses (Cosgrove,2005). The amounts of glycine-rich protein (GRP) encodinggenes were also decreased. GRPs are of unknown functions, butmany are associated with the pollen surface (Mayfield et al.,2001). The expression of almost all these genes was unaffected inthe Jekyll down-regulated gynoecium, where desiccation andpollen coat formation do not take place.
Discussion
In the angiosperms, the sporophytic tissues are responsible forthe nutrition and development of the miniature gametophyte.Here, we have shown that in barley the JEKYLL protein is pres-ent in sporophytic tissues surrounding both male and femalegametophytes, and that RNAi-mediated knockdown of Jekyllaffects the development of these tissues. Our results not onlysuggest that JEKYLL is involved in the sporophytic control ofthe barley gametophytic development, but also indicate that thedevelopment and function of both female and male border-ing tissues may be under the control of similar regulatorymechanisms.
(a) (b)
(c) (d)
(e) (f)
(g) (h)
(i) (j)
Fig. 7 Delayed pollen release and altered pollen morphology in barley(Hordeum vulgare) Jekyll down-regulated anthers. (a, b) Pollen shed inthe wildtype (a, arrowed) and delayed dehiscence in Jekyll down-regulated transgenic anthers (b) of the same age. (c–f) Light microscopyimages of wildtype (c) and transgenic anthers of the lines N91 (d), N18 (e)and N61 (f). The tapetum is almost completely degenerated in thewildtype anthers, but varying amounts of tapetal cell remnants are presentin the transgenic ones. (g, h) Starch deposition in wildtype (g) andtransgenic pollen of the line N91 (h), as visualized by iodine staining. (i, j)Raster microscopy of mature pollen grains of wildtype (i) and transgenicN91 line (j) shows frequent deformation of transgenic pollen. Bars, 20 lm.an, anther; gy, gynoecium; en, endothecium; ep, epidermis; lo, locule; ms,microspore; ta, tapetum; v, vacuole.
150 Research
NewPhytologist
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com
JEKYLL is required for performing nutrition function ofsporophytes
Nutrient delivery from sporophyte to gametophyte in anthers ismediated by tapetum, which releases nutrients into the antherlocule via secretion (Wang et al., 2003; Zhang et al., 2011). Thesimilar function in gynoecium is adopted by nucellus (Rangan &Rangaswamy, 1999; Luigi et al., 2006). The membrane-richstructures of the tapetal cells are responsible for the formation oftransport vesicles characteristic for cereal tapetum and called
orbicules or Ubisch bodies (Denecke, 2007; Zhang et al., 2011).JEKYLL was shown to be present in the endoplasmic reticulum,the Golgi apparatus and the small vesicles (Fig. 6), all of whichare components of the intracellular transport system (Hwang &Robinson,2009).TherepressionofJekyll ledtothedown-regulationof several genes encoding proteins involved in cellular traffic inthe anther (Table 2). In contrast to the rice api5 mutant anthers,where delayed degeneration of the tapetum does not affect orbi-cule formation (Li et al., 2011), the extrusion of orbicules isstrongly suppressed in Jekyll down-regulated barley anthers,reflecting a decrease in secretion capacity (Fig. 9; Table 2). As aresult, the nurse function of tapetum in these plants was compro-mised. Secretion normally continues until the whole cellularcontent of the tapetum has been supplied to the gametophyteand coupled with tapetal PCD (Vizcay-Barrena & Wilson, 2006;Parish & Li, 2010). The relationship between secretion and celldeath is not clear, but it is certainly possible that a functionalsecretory pathway is required for the spatiotemporal orchestra-tion of developmental PCD (Guo & Ho, 2008; Cacas, 2010).This suggestion is reinforced by the fact that, in the Jekyll down-regulated anthers, the expression of genes related to PCD(Table 2) and degeneration of the tapetal and subsequent cell lay-ers is significantly suppressed (Figs 7, 9). Suppression of PCDand disturbed nutrient delivery are observed in the nucellus ofJekyll down-regulated caryopses (Radchuk et al., 2006). WhilePCD reached maximal amount in caryopsis (Radchuk et al.,2011), it is initiated in gynoecia of flowers (the innermost layerof the nucellus). Correspondingly, down-regulation of PCD-related genes was already apparent in the gynoecia of transgenicplants, where Jekyll expression is repressed (Table 2). Thereduced sink strength of the transgenic male gametophyte proba-bly reflects a disruption of the nutrient flux through the tapetum.The tapetal cells of transgenic plants seemed unable to mediatesupply of the necessary amount of nutrients, and this triggered adeviant up-regulation of the genes responsible for nutrientstorage in the anther (Table 1).
Our hypothesis, therefore, is that Jekyll is required to ensure asufficient nutrition flux from the sporophyte to the gametophyteby programmed deterioration of nurse tissues. Thus Jekyll isimportant for the establishment of a balanced sink–source rela-tionship during gametogenesis. While direct monitoring of 13Csucrose movement from sporophytic tissues to the gametophyte
***
******
* * * **** ** *
**
Rel
ativ
e ex
pres
sion
leve
l (%
)
Fig. 8 Transcript profiling of Jekyll, MYB26, Tapetum Degeneration Retardation (TDR), Duo Pollen3 (DUO3), GAMYB, Apoptosis Inhibitor5 (API5),MADS3 and Persistent Tapetal Cell1 (PTC1) genes in barley (Hordeum vulgare) wildtype and Jekyll down-regulated anthers as determined by quantitativeRT-PCR analysis. Values are means + 1 SE. Significant differences between the transgenic and the wildtype are shown (t-test): *, P < 0.05; **, P < 0.01;***, P < 0.001.
(a) (b)
(c) (d)
Fig. 9 Interactions between the tapetum and the microspore. (a,b)Cross-sections of a barley (Hordeum vulgare) wildtype (a) and a transgenicN91 anther (b) show the secretory tapetum faced to the microspore.Nuclei are indicated by arrows. Delay in cell degradation of the transgenictapetum is evident. Red squares mark the positions of panels (c) and (d)with a higher resolution. (c) Secretion ⁄ extrusion of electron-dense materialinto the locule and towards the surface of pollen coat in the wildtypeanther. Below is shown the proposed secretory efflux route from atapetal cell to the microspore. (d) The formation of Ubisch bodies ishampered at the edge of the tapetum in the transgenic anther. Instead,electron-dense material is accumulated inside the nucellar cell (arrowed).Bars, 4 lm (a, b); 400 nm (c, d). en, endothecium; ex, exine; ms,microspore; n, nuclei; ob, orbicular wall; pro, pro, orbicles; ta, tapetum;Ub, Ubisch body.
NewPhytologist Research 151
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com
is challenging as a result of the small size of barley flowers, usingcaryopses of the same transgenic lines, it was shown that repres-sion of Jekyll resulted in deceleration of sucrose allocation vianucellar tissues (Melkus et al., 2011). Collectively these resultsimply that Jekyll has an impact on nutrient translocation betweentwo generations.
JEKYLL is involved in the sporophytic control of pollenmaturation
Jekyll repression did not induce aberrations in the way that MS1(Vizcay-Barrena & Wilson, 2006) and EMS1 ⁄ EXS1 do (Canaleset al., 2002; Zhao et al., 2002; Feng & Dickinson, 2010). This isconsistent with the absence of Jekyll expression at these develop-mental stages. However, we observed multiple aberrations duringthe late microgametogenesis, when massive nutrient transloca-tion, storage product accumulation and the formation of pollenwall occur (stages II–IV, Fig. S1). An aberrant star-like patternwas common in the pollen grains, and the second pollen mitosisof the generative cell often failed to occur (Fig. 10). In addition,there was a pronounced down-regulation of DUO3 (Fig. 8), agene required for the expression of certain germline genes(Durbarry et al., 2005; Rotman et al., 2005). The loss of eitherDUO1 or DUO3 function compromises male germline specifica-tion in A. thaliana (Brownfield et al., 2009). The pollenproduced by the Jekyll down-regulated transgenic anthersaccumulated less starch (Figs 7, 10). Decrease in storage was alsodemonstrated in barley caryopses, when sucrose delivery to theendosperm was compromised as a result of repression of Jekyll innucellar tissue (Radchuk et al., 2006; Melkus et al., 2011). Genesputatively involved in lipid and phenylpropanoid synthesis, aswell as those encoding enzymes necessary for cell wall synthesiswere also down-regulated in the transgenic anthers (Table 2).The formation of the outer coat of pollen, the exine, also relieson the deposition of sporopollenin, raftins and pectocellulosefrom the sporophytic tissues (Piffanelli et al., 1998; Wang et al.,2003; Blackmore et al., 2007); is controlled by GAMYB andTDR transcription factors (Zhang et al., 2011); and is vital forthe pollination (Kawanabe et al., 2006; Borg et al., 2009). Thenucellar cell remnants also contribute to the exine formation(Piffanelli et al., 1998; Blackmore et al., 2007). Both GAMYBand TDR genes are down-regulated in the transgenic anthers(Fig. 8), indicating tapetal PCD delay in transgenic antherswhich might compromise incorporation of cell remnants into theexine. Thus, JEKYLL function in sporophytic anther tissues isessential for the microgametophyte development, includingpollen formation, accumulation of storage products and thesynthesis of pollen coat, which were all impaired by therepression of Jekyll.
JEKYLL is important for the coordinated anther dehiscence
The controlled dehiscence of the anther, which is critical foreffective pollen shed, is brought about by PCD in the sporo-phytic tissues (Beals & Goldberg, 1997; Sanders et al., 2000).The initiation of PCD takes place in the tapetum and normally
(a) (b)
(c)
(e)
(d)
(f) (g)
(h) (i)
Fig. 10 Aberrations of microspore development in barley (Hordeum
vulgare). (a, b) Microspores containing vegetative and generative cells areformed in both wildtype (a) and transgenic (b) pollen. (c, d) The wildtypemicrospore with the central vacuole (c), and the frequently formed‘star-like’ transgenic microspore at the same stage. Numerous vacuoles areindicated by arrows (d). Dashed arrows in panels (a)–(c) indicate migrationof nuclei. (e) Transgenic microspore containing both vegetative and gener-ative cells, but the second generative cell is not formed. (f–i) Structure ofthe mature pollen in wildtype (f, h) and transgenic anthers (g, i). Multiplelarge starch grains are formed in the wildtype pollen (f) while aberrantstarch granules are present in transgenic pollen (g). Double arrows showoil bodies. The transgenic pollen is also characterized by multiple vacuoles(i). Bars, 5 lm (a–d); 1 lm (g–j). cv, central vacuole; ex, exine; gc,generative cell; n, nuclei; pp, pollen pore; sg, starch granule; v, vacuole;vn, vegetative nucleus.
152 Research
NewPhytologist
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com
extends radially to the endothecium and epidermal cells (Varnieret al., 2005). The factors triggering this process have remainedelusive (Wilson & Zhang, 2009). Here, we have shown that thepollen release from Jekyll-repressed anthers was disturbed(Fig. 7b). The accumulation of JEKYLL starts from the inner-most tapetal layer of the anther, then spreading to the outermosttissues of the sporophyte (Fig. 3), coinciding with degenerationof these tissues. JEKYLL is able to switch the cell fate to death(Radchuk et al., 2006). The repression of Jekyll might deceleratecell death in sporophytic tissues of the anther during anther
dehiscence (Wetzel & Jensen, 1992; Varnier et al., 2005). Themuch reduced expression of PCD-related genes (Table 2) andthe activation of those involved in biosynthesis and energymetabolism (Table 1) provided further evidence for the loss ofcontrol over dehiscence in the transgenic anthers.
The dehiscence process is under hormonal control (Wilson &Zhang, 2009), specifically involving JA in its early stages (Sanderset al., 2000; Stintzi & Browse, 2000; Ishiguro et al., 2001;Nagpal et al., 2005), and auxin in its later ones (Cecchetti et al.,2008). In Arabidopsis, loss-of-function double mutants for the
Table 1 Partial list of up-regulated genes from selected categories in transgenic anthers and gynoecia
Clone ID* Blast score Gene identification [species]
Transgenic anthers Transgenic gynoecia
Foldup-regulationin N91
Foldup-regulationin N61
Foldup-regulationin N91
Sugar conversion and starch biosynthesisHA24C13 572 Fructokinase II [Hordeum vulgare] 3.71 4.77 1.51HY04O18 970 Large subunit of AGPase AGP-L1 [Hordeum vulgare] 2.50 2.04 2.01HF15E17 930 Small subunit of AGPase AGP-S1a [Hordeum vulgare] 1.28 2.09 2.09HA25P03 1163 Granule bound starch synthase 1a [Hordeum vulgare] 2.33 2.04 1.52HF08M21 1083 Soluble starch synthase I [Hordeum vulgare] 2.21 1.75 2.00HB12M24 1206 Starch branching enzyme I [Hordeum vulgare] 1.35 5.95 1.30HB26B22 284 Isoamylase I [Hordeum vulgare] 2.08 1.81 1.85HZ60P11 282 a-Glucosidase-like protein [Oryza sativa] 1.58 2.48 1.33
mRNA transcription, translation and protein biosynthesisHZ64C24 675 Eukaryotic translation initiation factor 4G [Oryza sativa] 4.11 247.46 1.59HZ45N05 461 Eukaryotic initiation factor eIF-4A [Triticum aestivum] 2.36 1.54 2.10HB17E21 160 Histone H4 [Oryza sativa] 1.27 2.50 1.40HF05O20 159 RNA helicase [Oryza sativa] 2.10 2.38 1.77HF13L24 1208 Splicing factor Prp8 [Oryza sativa] 1.68 2.20 1.85HB03B11 597 Splicing factor 3A [Arabidopsis thaliana] 2.16 1.61 1.94HF05N18 483 60S acidic ribosomal protein P0 [Oryza sativa] 3.09 2.94 1.77HF16L12 286 40S ribosomal protein S23 [Oryza sativa] 3.74 2.87 1.65HZ59C07 192 60S ribosomal protein L34 [Oryza sativa] 2.19 2.28 1.57HF15N18 199 60S ribosomal protein L36 [Triticum aestivum] 2.37 2.07 1.71HF17G01 380 40S ribosomal protein S3 [Triticum aestivum] 2.19 1.97 1.77HF25A10 324 40S ribosomal protein S9 [Arabidopsis thaliana] 2.85 2.29 1.87HF01H20 336 Elongation factor 1b [Hordeum vulgare] 2.46 2.54 1.71HZ61K07 447 poly(A)-binding protein [Oryza sativa] 2.13 1.51 2.26HB23N24 358 24 kDa seed maturation protein [Oryza sativa] 3.55 2.89 1.71HZ47C19 786 Vacuolar processing enzyme 4 (legumain) [H. vulgare] 2.70 2.88 2.94HB29E03 250 a-Importin 1a [Oryza sativa] 2.52 3.63 1.66HB23B18 148 b-Importin 2 [Oryza sativa] 2.49 2.09 1.84HB25F18 113 Nuclear transport factor 2 [Arabidopsis thaliana] 2.97 3.25 1.60
Energy productionHY02K06 597 Glyceraldehyde-3-phosphate dehydrogenase [O. sativa] 2.23 12.54 1.42HB22I23 820 Plasma membrane H+ ATPase [Oryza sativa] 3.11 3.04 1.68HB22D22 714 Transaldolase [Oryza sativa] 2.07 2.76 2.17HZ45P03 520 Triosephosphate isomerase [Secale cereale] 2.35 2.09 1.67HY06L21 1289 Aconitate hydratase [Oryza sativa] 2.06 2.03 1.92HY02I23 1116 Phosphoglucomutase [Triticum aestivum] 2.80 2.10 1.85HF04P18 823 Phosphoenolpyruvate carboxykinase [Arabidopsis thaliana] 8.12 2.04 1.49
Amino acid metabolismHF23O03 935 S-adenosyl-L-homocysteine hydrolase [Hordeum vulgare] 2.13 1.40 1.84HF19L16 178 Succinic semialdehyde dehydrogenase [A. thaliana] 4.57 2.84 1.90HZ48B19 925 d-1-pyrroline-5-carboxylate synthetase [Triticum aestivum] 3.53 2.55 1.86HF18P23 266 Phosphoglycerate dehydrogenase [Oryza sativa] 2.11 3.36 1.80HF03F03 335 Enoyl-CoA hydratase [Oryza sativa] 2.17 2.48 1.65
*Clone ID is taken from http://pgrc.ipk-gatersleben.de/est/index.php. A full list of up-regulated genes is given in Table S1.2,0 and more fold differences in transgenic anthers and 1.5 and more differences in transgenic gynoecia are in bold.
NewPhytologist Research 153
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com

Table 2 Partial list of down-regulated genes from selected categories in transgenic anthers and gynoecia
Clone ID* Blast score Gene identification [species]
Transgenic anthers Transgenic gynoecia
Folddown-regulationin N91
Folddown-regulationin N61
Folddown-regulationin N91
Protein degradation and programmed cell death relatedHF04C05 337 Ubiquitin carboxyl-terminal hydrolase [Oryza sativa] 2.77 9.92 0.67HA27F15 382 20S proteasome alpha subunit C [Oryza sativa] 3.94 5.76 1.67HB03A23 639 Aspartic proteinase [Oryza sativa] 4.71 4.72 1.50HA30N09 162 Trypsin inhibitor BTICMc (Cystatin) [Hordeum vulgare] 2.36 4.08 1.71HB19J04 382 Aminopeptidase [Oryza sativa] 3.61 4.05 1.24
HZ59L08 174 Ubiquitin-conjugating enzyme domain protein [Oryza sativa] 2.02 3.12 0.80HA30C15 396 Lon protease [Triticum aestivum] 2.52 2.61 1.44HA03C18 327 Subtilisin-like serine protease [Oryza sativa] 2.15 2.51 0.89HB03H12 367 Ubiquitin carboxyl-terminal hydrolase [Oryza sativa] 2.98 3.12 1.97
Cell wall synthesis and related proteinsHF01L09 378 Lichenase [Hordeum vulgare] 5.14 2.54 1.32HB21G01 154 UDP-D-glucuronate decarboxylase [Hordeum vulgare] 3.63 2.35 1.49HB11L06 308 Cellulose synthase A3 [Oryza sativa] 2.07 2.26 1.88HZ56P15 399 Cellulose synthase CSLC9 [Oryza sativa] 2.76 2.03 1.32HB03P20 360 KORRIGAN [Hordeum vulgare] 3.83 2.00 1.72HZ52D12 492 a-Expansin A11 [Triticum aestivum] 2.19 2.20 0.80HZ42I19 292 b-Expansin B3 [Triticum aestivum] 2.39 2.19 1.23
Lipid metabolismHB11I06 689 x-3-Fatty acid desaturase [Triticum aestivum] 2.04 44.72 1.26HZ36L02 523 b-Keto-acyl-reductase [Hordeum vulgare] 2.09 12.29 0.60HY10P14 293 Galacturonosyltransferase 9 [Arabidopsis thaliana] 4.14 21.08 1.38HF12G23 445 Galactinol synthase 1 [Zea mays] 3.32 3.94 2.48HB25A02 484 b-Galactosidase [Oryza sativa] 2.25 2.81 0.99HF12O20 347 b-Galactosidase [Oryza sativa] 4.28 3.21 0.92HZ36H04 438 Lipoxygenase [Hordeum vulgare] 4.62 6.90 1.20HA01G02 61 Lipid transfer protein [Arabidopsis thaliana] 4.44 8.26 0.78HZ36L09 563 Xyloglucan endo-1,4-beta-D-glucanase [Hordeum vulgare] 3.21 2.06 0.54
Desiccation and dehydrationHB04N05 145 Water-stress protein [Zea mays] 13.94 34.52 0.96HA02G14 127 Desiccation-related protein [Arabidopsis thaliana] 7.61 7.46 2.71HB12M08 252 Dehydration-responsive protein [Arabidopsis thaliana] 7.54 6.79 2.05HF16I03 383 Early responsive to dehydration protein [Oryza sativa] 1.99 2.92 1.16HA02A15 220 WD-40 repeat family protein [Arabidopsis thaliana] 2.76 2.20 1.74HA22N22 240 Prolyl-4-hydroxylase [Arabidopsis thaliana] 3.08 17.48 1.49HF01E01 86 Proline-rich protein [Arabidopsis thaliana] 3.22 4.61 1.09HF16C21 142 Proline-rich protein [Arabidopsis thaliana] 2.03 2.73 1.28
Sugar metabolismHF11K03 201 Fructan 6-fructosyltransferase [Lolium perenne] 2.21 2.49 1.03HA03N06 462 Fructose-bisphosphatase [Triticum aestivum] 2.48 2.06 2.94HA03O24 742 Sedoheptulose-1,7-bisphosphatase [Triticum aestivum] 5.54 3.60 2.32
Chromatin structureHF22L15 56 DNA methyltransferase 2 [Oryza sativa] 2.92 4.41 1.31HA09D07 1445 Methionine synthase I [Hordeum vulgare] 2.21 3.04 2.18HB16E21 69 JmjC-containing histone lysine demethylase [A. thaliana] 10.52 8.35 0.93HB11G08 337 Histone acetyltransferase [Oryza sativa] 4.02 3.92 1.32
Hormone-responsibleHF15B23 308 Auxin response transcription factor ARF6 [Oryza sativa] 3.07 5.67 1.55HA01N04 106 Auxin-regulated protein [Arabidopsis thaliana] 2.01 1.95 1.51
OthersHF17M04 291 Peroxidase 1 [Triticum aestivum] 3.84 6.08 1.26HF24J20 468 Peroxidase III [Oryza sativa] 2.50 4.73 2.58HZ42L21 50 Glycine-rich protein [Oryza sativa] 1.97 2.41 0.65HY09E11 39 Glycine-rich protein [Arabidopsis thaliana] 2.08 2.27 1.14HZ44C15 168 Glycine-rich protein [Oryza sativa] 2.17 2.03 1.35
*Clone ID is taken from http://pgrc.ipk-gatersleben.de/est/index.php. A full list of up-regulated genes is given in Table S2.2,0 and more fold differences in transgenic anthers and 1.5 and more differences in transgenic gynoecia are in bold.
154 Research
NewPhytologist
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com

auxin response factors ARF6 and ARF8 produce indehiscentanthers and short filaments (Nagpal et al., 2005), and the barleyARF6 homolog was strongly down-regulated in both Jekyll RNAilines (Table 2). The MYB26 factor was also down-regulated inboth RNAi lines (Fig. 8). In A. thaliana, MYB26 is involved inthe regulation of anther dehiscence and, along with MYB24, is acomponent of the JA response (Mandaokar & Browse, 2009)during the late stages of anther development. The JA and JA-Ile-Leu (the active form of JA) content of the transgenic anthersduring the period when Jekyll expression was at its peak washigher than in the WT anthers (Fig. 1). JA is known to be able toactivate its own synthesis via a positive feedback loop (Farmer,2007; Wasternack, 2007). However, despite the high concentra-tion of JA present in the transgenic anthers, they still sufferedfrom delayed dehiscence. The data clearly indicate that JEKYLLis involved in the regulation of pollen release, but further studyfocused to a greater extent on JEKYLL interplay with other hor-mones governing the late stages of anther development (e.g. GA,auxin) is required to understand the mechanism of JEKYLL inanther dehiscence.
The impact of JEKYLL on fertility
Defects in the generative nuclei, reductions in the amount ofstorage products available and imperfect pollen walls all impairthe success of pollination. All these aberrations occurred in thepollen produced by Jekyll down-regulated plants, and so canexplain the major reduction in seed set experienced when theirpollen was used for fertilization (Fig. 1a). A more direct role forJEKYLL is also imaginable. The protein composition of thepollen wall is important for adhesion, signaling and sexual com-patibility (Edlund et al., 2004). JEKYLL protein was detectableon the pollen grain surface (Fig. 3g). The down-regulation ofJekyll would therefore likely reduce the amount of JEKYLLpresent on or within the pollen wall. During the course of theinteraction between the germinating pollen grain and thestigma, PCD is triggered as a means of incompatibility (Bosch &Franklin-Tong, 2008), but it is also required for the normalgrowth of the pollen tube (Wu & Cheun, 2000; Crawford &Yanofsky, 2008). JEKYLL has structural similarity (see Radchuket al., 2006) to pollen wall GRPs in the Brassicaceae (Fiebig et al.,2004). Mutation of GRP17 in Arabidopsis has been shown todelay pollen hydration and to decrease the rate of effectivepollination (Mayfield & Preuss, 2000).
Genes involved in the reproductive system are prone to rapidevolutionary change, which provides a means of curtailing fertil-ization by nonselfs (Fiebig et al., 2004). The Jekyll sequence ispresent as a single copy in barley, with homologous sequencesknown in wheat and rye but not in more distant species(Radchuk et al., 2006). In Arabidopsis and some related species, afamily of pollen-specific GRPs appears to have diverged markedlythrough a series of duplications and deletions, as well as throughmutations in their C-terminal repetitive sequences (Fiebig et al.,2004). A comparison of Jekyll homologs has also identified diver-gence in the central and C terminal repetitive regions, while the Ntermini have remained rather well conserved (Radchuk et al.,
2006). Such divergence in genes involved in sexual reproductioncan promote speciation by preventing wide crossing and ⁄ or byreducing the extent of self-incompatibility expressed (Edlundet al., 2004; Wheeler et al., 2009). While the evolutionarysignificance of JEKYLL remains to be explored, its role in thesexual reproduction of barley is clearly much broader than hasbeen hitherto realized.
Acknowledgements
We thank U. Wobus, T. Altmann, J. Thiel and D. Weier forvaluable discussions. Special thanks to M. Strickert and S. Friedelfor support in quantitative normalization and statistics. Wegreatly appreciate U. Siebert, A. Stegmann, G. Einert, and E.Fessel for excellent technical assistance. This work was supportedin part by the German Ministry of Education and Researchwithin the German Plant Genome Initiative (GABI-sysSEED,FKZ 0315044A and GABI-GRAIN, FKZ0315041A).
References
Albrecht C, Russinova E, Hecht V, Baaijens E, de Vries S. 2005. The Arabidopsisthaliana SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASES 1 and 2control male sporogenesis. Plant Cell 17: 3337–3349.
Beals TP, Goldberg RB. 1997. A novel cell ablation strategy blocks tobacco
anther dehiscence. Plant Cell 9: 1527–1545.
Blackmore S, Wortley AH, Skvarla JJ, Rowley JR. 2007. Pollen wall
development in flowering plants. New Phytologist 174: 483–498.
Bolstad BM, Irizarry RA, Astrand M, Speed TP. 2003. A comparison of
normalization methods for high density oligonucleotide array data based on
bias and variance. Bioinformatics 19: 185–193.
Borg M, Brownfield L, Twell D. 2009. Male gametophyte development: a
molecular perspective. Journal of Experimental Botany 60: 1465–1478.
Bosch M, Franklin-Tong VE. 2008. Self-incompatibility in Papaver: signalling to
trigger PCD in incompatible pollen. Journal of Experimental Botany 59:
481–490.
Brambilla V, Kater M, Colombo L. 2008. Ovule integument identity
determination in Arabidopsis. Plant Signaling and Behavior 3: 246–247.
Brownfield L, Hafidh S, Durbarry A, Khatab H, Sidorova A, Doerner P, Twell
D. 2009. Arabidopsis DUO POLLEN3 is a key regulator of male germline
development and embryogenesis. Plant Cell 21: 1940–1956.
Cacas J. 2010. Devil inside: does plant programmed cell death involve the
endomembrane system? Plant, Cell & Environment 33: 1453–1473.
Canales C, Bhatt AM, Scott R, Dickinson H. 2002. EXS, a putative LRR
receptor kinase, regulates male germline cell number and tapetal identity and
promotes seed development in Arabidopsis. Current Biology 12: 1718–1727.
Cecchetti V, Altamura MM, Falasca G, Costantino P, Cardarelli M. 2008.
Auxin regulates Arabidopsis anther dehiscence, pollen maturation, and filament
elongation. Plant Cell 20: 1760–1774.
Chang F, Wang Y, Wang S, Ma H. 2011. Molecular control of
microsporogenesis in Arabidopsis. Current Opinion in Plant Biology 14: 66–73.
Coffeen WC, Wolpert TJ. 2004. Purification and characterization of serine
proteases that exhibit caspase-like activity and are associated with programmed
cell death in Avena sativa. Plant Cell 16: 857–873.
Colcombet J, Boisson-Dernier A, Ros-Palau R, Vera CE, Schroeder JI. 2005.
Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASES 1 and 2 are
essential for tapetum development and microspore maturation. Plant Cell 17:
3350–3361.
Cosgrove DJ. 2005. Growth of the plant cell wall. Nature Reviews Molecular CellBiology 6: 850–861.
Crawford BC, Yanofsky MF. 2008. The formation and function of the female
reproductive tract in flowering plants. Current Biology 18: R972–R978.
NewPhytologist Research 155
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com

Czechowski T, Stitt M, Altmann T, Udvardi MK, Scheible WR. 2005.
Genome-wide identification and testing of superior reference genes for
transcript normalization in Arabidopsis. Plant Physiology 139: 5–17.
Denecke J. 2007. Endoplasmic reticulum. In: Roberts K, ed. Handbook of plantscience. Hoboken, NJ, USA: Wiley, 223–227.
Durbarry A, Vizir I, Twell D. 2005. Male germ line development in Arabidopsis:duo pollen mutants reveal gametophytic regulators of generative cell cycle
progression. Plant Physiology 137: 297–307.
Edlund AF, Swanson R, Preuss D. 2004. Pollen and stigma structure and
function: the role of diversity in pollination. Plant Cell 16: S84–S97.
Ellerstrom M, Reidt W, Ivanov R, Tiedemann J, Melzer M, Tewes A, Moritz T,
Mock HP, Sitbon F, Rask L et al. 2005. Ectopic expression of EFFECTOROF TRANSCRIPTION perturbs gibberellin-mediated plant developmental
processes. Plant Molecular Biology 59: 663–681.
Farmer EE. 2007. Plant biology: jasmonate perception machines. Nature 448:
659–660.
Feng X, Dickinson HG. 2007. Packaging the male germline in plants. Trends inGenetics 23: 503–510.
Feng X, Dickinson HG. 2010. Tapetal cell fate, lineage and proliferation in the
Arabidopsis anther. Development 137: 2409–2416.
Fiebig A, Kimport R, Preuss D. 2004. Comparisons of pollen coat genes across
Brassicaceae species reveal rapid evolution by repeat expansion and
diversification. Proceedings of the National Academy of Sciences, USA 101:
3286–3291.
Fuji S, Toriyama K. 2008. Genome barriers between nuclei and mitochondria
exemplified by cytoplasmic male sterility. Plant and Cell Physiology 49:
1484–1494.
Goldberg RB, Beals TP, Sanders PM. 1993. Anther development: basic
principles and practical applications. Plant Cell 5: 1217–1229.
Guo WJ, Ho TH. 2008. An abscisic acid-induced protein, HVA22, inhibits
gibberellin-mediated programmed cell death in cereal aleurone cells. PlantPhysiology 147: 1710–1722.
Hwang I, Robinson DG. 2009. Transport vesicle formation in plant cells.
Current Opinion in Plant Biology 12: 660–669.
Ishiguro S, Kawai-Oda A, Ueda J, Nishida I, Okada K. 2001. The DEFECTIVEIN ANTHER DEHISCIENCE gene encodes a novel phospholipase A1
catalyzing the initial step of jasmonic acid biosynthesis, which synchronizes
pollen maturation, anther dehiscence, and flower opening in Arabidopsis. PlantCell 13: 2191–2209.
Johnson-Brousseau SA, McCormick S. 2004. A compendium of methods useful
for characterizing Arabidopsis pollen mutants and gametophytically-expressed
genes. Plant Journal 39: 761–775.
Kawanabe T, Ariizumi T, Kawai-Yamada M, Uchimiya H, Toriyama K. 2006.
Abolition of the tapetum suicide program ruins microsporogenesis. Plant andCell Physiology 47: 784–787.
Leon P, Sheen J. 2003. Sugar and hormone connections. Trends in Plant Science8: 110–116.
Li H, Yuan Z, Vizcay-Barrena G, Yang C, Liang W, Zong J, Wilson ZA, Zhang
D. 2011a. PERSISTENT TAPETAL CELL1 encodes a PHD-finger protein
that is required for tapetal cell death and pollen development in rice. PlantPhysiology 156: 615–630.
Li H, Zhang D. 2010. Biosynthesis of anther cuticle and pollen exine in rice.
Plant Signaling & Behavior 5: 1121–1123.
Li N, Zhang DS, Liu HS, Yin CS, Li X, Liang W, Yuan Z, Xu B, Chu HW,
Wang J et al. 2006. The rice Tapetum Degeneration Retardation gene is
required for tapetum degradation and anther development. Plant Cell 18:
2999–3014.
Li X, Gao X, Wei Y, Deng L, Ouyang Y, Chen G, Li X, Zhang Q, Wu C.
2011b. Rice APOPTOSIS INHIBITOR5 coupled with two DEAD-box
adenosine 5’-triphosphate-dependent RNA helicases regulates tapetum
degeneration. Plant Cell 23: 1416–1434.
Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using
real-time quantitative PCR and the 22)DDCT method. Methods 25: 402–408.
Luigi B, Papini A, Milocani E, Vesprini JL. 2006. Programmed cell death in the
nucellus of Tillandsia (Bromeliaceae). Caryologia 59: 334–339.
Luo ZB, Janz D, Jiang X, Gobel C, Wildhagen H, Tan Y, Rennenberg H,
Feussner I, Polle A. 2009. Upgrading root physiology for stress tolerance
by ectomycorrhizas: insights from metabolite and transcriptional profiling
into reprogramming for stress anticipation. Plant Physiology 151:
1902–1917.
Ma H. 2005. Molecular genetic analyses of microsporogenesis and
microgametogenesis in flowering plants. Annual Review of Plant Biology 56:
393–434.
Mandaokar A, Browse J. 2009. MYB108 acts together with MYB24 to regulate
jasmonate-mediated stamen maturation in Arabidopsis. Plant Physiology 149:
851–862.
Maraschin SF, de Priester W, Spaink HP, Wang M. 2005. Androgenic switch:
an example of plant embryogenesis from the male gametophyte perspective.
Journal of Experimental Botany 56: 1711–1726.
Mayfield JA, Fiebig A, Johnstone SE, Preuss D. 2001. Gene families from the
Arabidopsis thaliana pollen coat proteome. Science 292: 2482–2485.
Mayfield JA, Preuss D. 2000. Rapid initiation of Arabidopsis pollination requires
the oleosin-domain protein GRP17. Nature Cell Biology 2: 128–130.
Melkus G, Rolletschek H, Fuchs J, Radchuk V, Grafahrend-Belau E,
Sreenivasulu N, Rutten T, Weier D, Heinzel N, Schreiber F et al. 2011.
Dynamic 13C ⁄ 1H NMR imaging uncovers sugar allocation in the living seed.
Plant Biotechnology Journal 9: 1022–1037.
Morrell PL, Clegg MT. 2007. Genetic evidence for a second domestication of
barley (Hordeum vulgare) east of the Fertile Crescent. Proceedings of theNational Academy of Sciences, USA 104: 3289–3294.
Murmu J, Bush MJ, DeLong C, Li S, Xu M, Khan M, Malcolmson C, Fobert
PR, Zachgo S, Hepworth SR. 2010. Arabidopsis basic leucine-zipper
transcription factors TGA9 and TGA10 interact with floral glutaredoxins
ROXY1 and ROXY2 and are redundantly required for anther development.
Plant Physiology 154: 1492–1504.
Nagpal P, Ellis CM, Weber H, Ploense SE, Barkawi LS, Guilfoyle TJ, Hagen G,
Alonso JM, Cohen JD, Farmer EE et al. 2005. Auxin response factors ARF6
and ARF8 promote jasmonic acid production and flower maturation.
Development 132: 4107–4118.
Parish RW, Li SF. 2010. Death of a tapetum: a programme of developmental
altruism. Plant Science 178: 73–89.
Piffanelli P, Ros JHE, Murphy DJ. 1998. Biogenesis and function of the lipidic
structures of pollen grains. Sexual Plant Reproduction 11: 65–80.
Radchuk V, Borisjuk L, Radchuk R, Steinbiss HH, Rolletschek H, Broeders S,
Wobus U. 2006. Jekyll encodes a novel protein involved in the sexual
reproduction of barley. Plant Cell 18: 1652–1666.
Radchuk V, Weier D, Radchuk R, Weschke W, Weber H. 2011. Development
of maternal seed tissue in barley is mediated by regulated cell expansion and
cell disintegration and coordinated with endosperm growth. Journal ofExperimental Botany 62: 1217–1227.
Ramakers C, Ruijter JM, Deprez RH, Moorman AF. 2003. Assumption-free
analysis of quantitative real-time polymerase chain reaction (PCR) data.
Neuroscience Letters 13: 62–66.
Rangan TS, Rangaswamy NS. 1999. Nucellus – a unique embryologic system.
Phytomorphology 49: 337–376.
Ravanel S, Gakiere B, Job D, Douce R. 1998. The specific features of
methionine biosynthesis and metabolism in plants. Proceedings of the NationalAcademy of Sciences, USA 95: 7805–7812.
Rotman N, Durbarry A, Wardle A, Yang WC, Chaboud A, Faure JE, Berger F,
Twell D. 2005. A novel class of MYB factors controls sperm-cell formation in
plants. Current Biology 15: 244–248.
Rowland O, Lee R, Franke R, Schreiber L, Kunst L. 2007. The CER3 wax
biosynthetic gene from Arabidopsis thaliana is allelic to WAX2 ⁄ YRE ⁄ FLP1.
FEBS Letters 581: 3538–3544.
Rutten T, Kruger C, Melzer M, Stephan UW, Hell R. 2003. Discovery of an
extended bundle sheath in Ricinus communis L. and its role as a temporal
storage compartment for the iron chelator nicotianamine. Planta 217:
400–406.
Sanders PM, Lee PY, Biesgen C, Boone JD, Beals TP, Weiler EW,
Goldberg RB. 2000. The Arabidopsis DELAYED DEHISCENCE1 gene
encodes an enzyme in the jasmonic acid synthesis pathway. Plant Cell 12:
1041–1061.
Schiefthaler U, Balasubramanian S, Sieber P, Chevalier D, Wisman E,
Schneitz K. 1999. Molecular analysis of NOZZLE, a gene involved in
156 Research
NewPhytologist
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com

pattern formation and early sporogenesis during sex organ development in
Arabidopsis thaliana. Proceedings of the National Academy of Sciences, USA 96:
11664–11669.
Schussler JR, Westgate ME. 1995. Assimilate flux determines kernel set at low
water potential in maize. Crop Science 35: 1074–1080.
Scott RJ, Spielman M, Dickinson HG. 2004. Stamen structure and function.
Plant Cell 16: S46–S60.
Sreenivasulu N, Radchuk V, Strickert M, Miersch O, Weschke W, Wobus U.
2006. Gene expression patterns reveal tissue-specific signaling networks
controlling programmed cell death and ABA-regulated maturation in
developing barley seeds. Plant Journal 47: 310–327.
Sreenivasulu N, Usadel B, Winter A, Radchuk V, Scholz U, Stein N, Weschke
W, Strickert M, Close TJ, Stitt M et al. 2008. Barley grain maturation and
germination: metabolic pathway and regulatory network commonalities and
differences highlighted by new MapMan ⁄ PageMan profiling tools. PlantPhysiology 146: 1738–1758.
Stintzi A, Browse J. 2000. The Arabidopsis male-sterile mutant, opr3, lacks the
12-oxophytodienoic acid reductase required for jasmonate synthesis.
Proceedings of the National Academy of Sciences, USA 97: 10625–10630.
Sun Q, Zhou DX. 2008. Rice jmjC domain-containing gene JMJ706 encodes
H3K9 demethylase required for floral organ development. Proceedings of theNational Academy of Sciences, USA 105: 13679–13684.
Sundaresan V, Alandete-Saez M. 2010. Pattern formation in miniature: the
female gametophyte of flowering plants. Development 137: 179–189.
Varnier AL, Mazeyrat-Gourbeyre F, Sangwan RS, Clement C. 2005.
Programmed cell death progressively models the development of anther
sporophytic tissues from the tapetum and is triggered in pollen grains during
maturation. Journal of Structural Biology 152: 118–128.
Vizcay-Barrena G, Wilson ZA. 2006. Altered tapetal PCD and pollen wall
development in the Arabidopsis ms1 mutant. Journal of Experimental Botany57: 2709–2717.
Wang A, Xia Q, Xie W, Datla R, Selvaraj G. 2003. The classical Ubisch bodies
carry a sporophytically produced structural protein (RAFTIN) that is essential
for pollen development. Proceedings of the National Academy of Sciences, USA100: 14487–14492.
Wasternack C. 2007. Jasmonates: an update in biosynthesis, signal transduction
and action in plant stress response, growth and development. Annals of Botany100: 681–697.
Watanabe M. 2008. Towards a comprehensive understanding of molecular
mechanisms of sexual reproduction in higher plants. Plant and Cell Physiology49: 1404–1406.
Wetzel CLR, Jensen WA. 1992. Studies of pollen maturation in cotton: the
storage reserve accumulation phase. Sexual Plant Reproduction 5: 117–127.
Wheeler MJ, de Graaf BH, Hadjiosif N, Perry RM, Poulter NS, Osman K,
Vatovec S, Harper A, Franklin FC, Franklin-Tong VE. 2009. Identification of
the pollen self-incompatibility determinant in Papaver rhoeas. Nature 459:
992–995.
Wilson ZA, Zhang DB. 2009. From Arabidopsis to rice: pathways in pollen
development. Journal of Experimental Botany 60: 1479–1492.
Wu HM, Cheun AY. 2000. Programmed cell death in plant reproduction. PlantMolecular Biology 44: 267–281.
Yang C, Xu Z, Song J, Conner K, Vizcay-Barrena G, Wilson ZA. 2007.
Arabidopsis MYB26 ⁄ MALE STERILE35 regulates secondary thickening in the
endothecium and is essential for anther dehiscence. Plant Cell 19: 534–548.
Yang WC, Shi DQ, Chen YH. 2010. Female gametophyte development in
flowering plants. Annual Review of Plant Biology 61: 89–108.
Yang WC, Ye D, Xu J, Sundaresan V. 1999. The SPOROCYTELESS gene of
Arabidopsis is required for initiation of sporogenesis and encodes a novel
nuclear protein. Genes and Development 13: 2108–2117.
Zhang D, Luo X, Zhu L. 2011. Cytological analysis and genetic control
of rice anther development. Journal of Genetics and Genomics 38:
379–390.
Zhao X, de Palma J, Oane R, Gamuyao R, Luo M, Chaudhury A, Herve P, Xue
Q, Bennett J. 2008. OsTDL1A binds to the LRR domain of rice receptor
kinase MSP1, and is required to limit sporocyte numbers. Plant Journal 54:
375–387.
Zhao DZ, Wang GF, Speal B, Ma H. 2002. The EXCESS MICROSPORO-CYTES 1 gene encodes a putative leucine-rich repeat receptor protein kinase
that controls somatic and reproductive cell fates in the Arabidopsis anther.
Genes and Development 16: 2021–2031.
Zuo J, Niu QW, Nishizawa N, Wu Y, Kost B, Chua NH. 2000.
KORRIGAN, an Arabidopsis endo-1,4-beta-glucanase, localizes to the cell
plate by polarized targeting and is essential for cytokinesis. Plant Cell 12:
1137–1152.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1 Description of the stages of barley anther developmentused in the study.
Table S1 Complete list of up-regulated genes in the transgenicbarley anthers and gynoecia compared to the wildtype plants
Table S2 Complete list of down-regulated genes in the transgenicbarley anthers and gynoecia compared to the wildtype plants
Table S3 Complete list of quantile normalized expression data of12K cDNA array from transgenic barley anthers ⁄ gynoecia andthe respective wildtypes
Table S4 Primers used in quantitative RT-PCR analyses
Methods S1 Determination of phytohormones.
Please note: Wiley-Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
NewPhytologist Research 157
� 2012 The Authors
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 194: 142–157
www.newphytologist.com





























