Embed Size (px)
Citation preview

EXOCRINOLOGY
THE SCIENCE OF LOVE
Human Pheromones in Criminology, Psychiatry, and Medicine
B. Nicholson


Exocrinology The Science of
LoveHuman Pheromones in Criminology, Psychiatry,
and Medicine
B. Nicholson
EXOCRINOLOGY

Nicholson Science2604 W JETTON AVE
TAMPA, FL [email protected]
Published in the United States of America
By Nicholson Science, all rights reserved
Nicholson Science 2604 W JETTON AVE TAMPA FLORIDA 33629-5325
© 2011 Ross Nicholson
Library of Congress Control Number: not yet assigned
ISBN 978-1461187288
(via createspace.com)

Dedicated to Iraneus Eibl-Eibesfeldt and E.O. Wilson
EXOCRINOLOGY


Preface to Exocrinology The Science of Love A new perspective in science may be difficult to appreciate. This one particularly. The facts are these: male facial skin surface lipid has cured delinquency and runaway behavior hundreds of times. A few dozen times, homosexuality has been transmuted to heterosexuality and none want to return to perversion. Several times, heroin and crack cocaine addicted young women have, with one dose less than 250 mg, abstained from the drug and rebuilt their lives with family and pregnancy. Sexual ideation can be stimulated with a human pheromone, too. This new perspective has allowed accurate predictions of human pheromone to be found in emotional tears, the molecular basis for chemosensory transduction, and pheromonal epigenetic influences. Given these fabulous anecdotal successes, this exemplary record of accurate prediction, and with the astounding reluctance of human beings to examine the very ends of their own noses, drawing this new map of the logic seems prudent. If this glacial pace is all we can expect, then we will all die of something we should have cured long before substantial progress is made. Suppose we take this new perspective and consider what if? If you lack the economic, chemistry, and neuroanatomy skills to follow all the logic, just chump along. Do not abandon the gold mine because you cannot tell ore from dirt. The nuggets may be buried, but they are here. What if modern science believed in fairy tales? Our confidence for scientific triumph over baseless nonsense is decidedly misplaced. How could that be? When one strips untested consensus from our knowledge of human behavior, very little remains. The so-called explanations of psychology, sociology, criminology and most of psychiatry suffer acutely from insufficient evidence. Paring it down is easy. If it doesn’t have mass, energy, or value; it doesn’t exist. It is only when our body of knowledge is completely free of smug half truths, mis-appellation, and consensus of opinion that we can get down to business. Today may indeed be Monday, 21 May 2011 by the accepted calendar, but there is nothing intrinsically Monday about today. People can agree it is Monday, but that does not make it so. For instance, modern science has ridiculed the broad application of economic theory to human behavioral evolution as patently absurd. Of what possible use could the assumptions of economic man, perfect information (in geologic time), unlimited wants, equations of optimality (e.g. linear programming), marginal value, kinetic homeostasis, and dedication of resources for the same marginal profit, bring to understanding evolution? Some wit suggested a funny name (Dr. Pangloss) and bingo, everyone fell into line. What happened to testing hypotheses? The watchword has been reductio ad absurdum vs the maxim of “equations don’t lie” and, sadly for
EXOCRINOLOGY

science, equations lost. This author predicated his investigations on reversal of the outcome of that unfair fight. Absurdity is over-rated anyway. Next, today’s science despises religion, rejecting its sacred testimony not as a helpful lay history of honest witness, but as, again, mere hermeneutics. Whoa! Equations AND God? Is there any wonder why we die from so many unknown causes? This author suggests that in this volume can be found the tools of Christ, Mohammed, and the Olympians: exocrinology. OK, it sounds weird, but sometimes truth is just strange. Nearly thirty years ago, this author’s early economic “equations don’t lie” logic foretold: that human pheromones would prove vital to our understanding of human physiology and behavior, that tears would have dissolved pheromone receptor protein “primer pheromone”, that sexual orientation would be pheromonal; that crime and insanity would be devices of human pheromones as well. That same atomic evaluation of the organism as optimizing evolution of parts with physiology and behavior suggested that organs, cells, even molecules would be purposed in concert to fulfill evolved function. Does it sound like “The Force? As chance would have it, this author was once young and creative enough to have invented “The Force” as a metaphor for monotheism. Steve Spielberg carried it off to his Hollywood friends. Spielberg’s friends came back for more. Taking a wider view let this author contribute more than a hundred more moralistic movies to the current popular vernacular: Up, Inception, Avatar, Titanic, The Matrix, Phantom Menace, Forrest Gump, Braveheart are all this author’s stories. Star Wars and E.T. were among my first attempts and yes, my Spielberg show business association has indeed been cause for significant distraction, but it has always been fun. Would you expect science from a ‘Hollywood-type’? Alas, my character is assassinated by my experiences. Unfortunately for those whose perspicacity is skin deep, my character is further assassinated by my youthful political associations. An intellect capable of unraveling most modern medical mysteries mathematically should also prove useful for sleuthing out great unsolved crimes, constructing public policy, as well as in movie entertainment. My helps to successful politicians, (e.g. the Bushes (unfortunately villainous fiends who murdered JFK, MLK, RFK, LBJ, George Wallace, and many more), the Clintons, Julian Bond, Zell Miller, Ann Richards, Strobe Talbot, Khalid Sheikh Mohammed: “Privatization”, “Deadbeat Dad Laws”, “Inclusive Democracy”, “HOPE Scholarships”, “Freedom of Religion Rules in the Federal Workplace” have helped our country, but serve only to disparage me among the various science tribes. The popular meaning of “inclusive” -- at least its non-mathematical, political definition -- came to me whilst sitting in back of my English class at Richardson High School in 1971. And does anyone blame me for helping our

Federal Reserve realize that they were not out of options at 0% that they could buy negotiable instruments to combat deflation (quantitative easing) and even had the power to do it.
Why couldn’t I stop whilst ahead? The JFK assassination and almost all following American assassinations were conducted to appease Barbara Bush’s sexual lusts. Her vengeful husband, the cuckold George H.W. Bush, carried it off. What on earth is the most famous un-caught serial assassin being made mention of here in a science book? I had to open my big mouth and solve the greatest unsolved crime of my time, right? The shooting of Congresswoman Giffords is the latest in the string b the plumbers. Unfortunately, the same insight that serves so well in scientific elucidation also serves here. Still, we didn’t get all dressed up for nuthin’. Rest assured that we will now leave all these distractions behind, and get to the meat of the matter at hand: saving humanity with the cool tools of Jesus Christ.
EXOCRINOLOGY

Preface to the 2nd Edition This book, OF LOVE, textbook of medical science: exocrinology, is a revision of an earlier work OF LOVE kisses pass epigenetic pheromones in the pathogenesis of sociopathy, ‘mental illness’, and disease. The cure for crime. The cure for drug addiction. The cure for sexual perversion. This 2nd edition with its abbreviated title, incorporates changes suggested by colleagues and scholars from around the world. Chapters 8 and 9 were switched in their places to introduce the detailed and better understood sociopathies of crime, addiction, and perversion in anticipation of the less easily understood autoimmune diseases. Low and behold, human sexual orientation does indeed depend upon pheromone recognition, just as the author predicted in 1983. (Indeed, human pheromone can change sexual orientation as my open trials have tentatively demonstrated.) There, as the author expected, is the convolutional change in pheromone receptor (binding) proteins exactly as predicted. The pheromone receptor proteins are indeed essential to chemosensory transduction just as predicted. Little scientific headway has been made in the intervening two years since first publication of this book despite its wide dissemination. The sexual dimorphic tears of women were found for the first time to have the pheromonal effect of reducing masculine testosterone while hugs were shown to change the concentration of pheromone receptor proteins in the blood. The latter idea was unanticipated but welcome in economic explanation for that affectionate behavior. The former was suggested by this author nearly thirty years ago. Hopefully, at least human pheromone reception will be traced to the microvillar “brush border” cells that line the human upper-respiratory system before the next two orbits of the sun shine on the next edition, but time will tell. America has a startling inability to fund double-blind testing of the masculine facial skin surface lipids cure for crime. Hopefully, the preponderance of charlatans will abate with application of these ideas overseas. In other realms, with fewer psychologists, criminals, mischievous anorexics, and offended sexual perverts, perhaps progress can be made unhindered. Of course, the economic and scientific explanations for human romantic and parental love behaviors will be just as fresh and new to those who completely ignored them decades ago. This time around we provide much needed therapeutic options. Finally, in medical school, sooner or later your professors will admit that most of us suffer and die from illnesses for which no one knows cause or cure. 55% is a conservative estimate for deaths from atherosclerosis and its complications. Up to 55% of women suffer from hypoactive sexual desire syndrome. 55% of some

populations will spend some time incarcerated, addicted, unloved or perverted. This book is for that 55%.
EXOCRINOLOGY


TABLE OF CONTENTSIntroduction ! ! ! ! ! ! ! ! 1Chapter 1: Kissing ! ! ! ! ! ! ! 9Chapter 2: Parental Love of Offspring ! ! ! 49Chapter 3: Chemical Characteristics of ! Human Pheromones ! ! ! ! ! 77Chapter 4: More about Bonding: ! Neglect and Abuse ! ! ! ! ! ! 119Chapter 5: Eating Disorders ! ! ! ! ! 149Chapter 6: Crying Chemoreception and Love ! ! 157Chapter 7: Human Pheromone Transduction ! ! 191Chapter 8: Pheromone Control of Crime and ! Drug Addiction! ! ! ! ! ! ! 223Chapter 9: Pheromone Control of Sexual ! Desire Syndromes Chapter 10: Pheromone Reception & Disease ! 233Chapter 11: Atherosclerosis ! ! ! ! ! 277Chapter 12: The Function of Myelin ! ! ! 297Chapter 13: The Future ! ! ! ! ! ! 311Appendix! ! ! ! ! ! ! ! ! 317
EXOCRINOLOGY


Introduction
Pheromones regulate the human hierarchy that determines rates of infection, sociopathy, disease, accidents, and death. Passionate pheromone feed-back systems maintain criminality, insanity, and all sorts of perversions that we can now modify to our liking. Pheromones regulate fertility and with other inputs (hormonal and neural) choose the best candidate oocytes for fertilization and just the right sperm.
A policeman just called your cell phone. They have your suddenly gone wild and gone missing puberal daughter down at the police station. You are her father, now what do you do? The proposed cure for runaway behavior is in the Appendix. You can cure your daughter with one pack of Wrigley’s Rain chewing gum (15 pieces) and the grease on your face.
First of all, it is not her fault, so do not blame your daughter. Bad thinking does not cause misbehavior problems. Certain easily avoidable biological transgressions or sins can, but so can air pollution. Such epigenetic transgressions can lay blame with you, or your spouse, your parents, even grandparents or great grand parents, just like Exodus 34:7. Next, you can alleviate your daughter’s immediate difficulty with a pheromonal communion. Follow the instructions at the very end of this book: collection of healthy adult male facial skin surface lipid, and give your daughter the pheromone chewing gum to chew. You need not be present, but your daughter’s misbehavior requires your male, fatherly facial chemical signature, the shine on the end of your nose, to be put upon her saliva, a big dose just one time. Your daughter needs your ‘blessing’ to behave, to perform well in school, and now to stay out of prison. So give your returning prodigal the shine off your nose, and celebrate. It actually is the right thing to do.
OF LOVE is a science book for scientists and best appreciated by those
physicians and medical students familiar with the rather severe microeconomic principles of Milton Freedman. A familiarity with neuroanatomy and first semester organic chemistry could help too, but the few necessary concepts are introduced in the text so a review should be unnecessary. Indeed, finding relevance in the science
EXOCRINOLOGY
1

of love for organic chemistry sufferers may even stimulate students to master those subjects more thoroughly.
Discovering the pheromonal basis of disease, the clinical relevance of this book for practicing physicians is clear, but awaiting clinical trials. So this is a book of anecdotes and theory, but it is compelling on both counts. All open trials have succeeded, although the pheromone has little effect on alcoholism. [Please see the appendix for collection protocol details. The cure for drug addiction is 150-250 mg of healthy adult male facial skin surface lipid taken in one dose on a chewing gum vehicle by mouth. The cure for juvenile delinquency is also 150-250 mg of adult male facial skin surface lipid taken in one dose by mouth. The cure for homosexual ideation and behavior is 150-250 mg of adult male facial skin surface lipid taken in one dose by mouth. The cure for teenage runaways is 150-250 mg of adult male facial skin surface lipid taken in one dose by mouth.] Similarly successful treatments for autoimmune diseases and cancers are likely. For instance, that same wonderful dose has stopped the progression and reversed some symptoms of Alzheimer’s disease within a few weeks on four occasions so far. Refined and better understood with double-blind cross-over protocols and repetition in other labs, will these anecdotes stand the test of time? Science will decide.
This book begins the end of important human institutions that are even now outliving their usefulness. Jails and prisons will simply dry up and cease to exist. There will be an end to widespread police presence, an end to the ‘homosexual community’, an end to the oxymoronic criminal justice system, an end to the drug wars, an end to jihad and an end to terror. The world will ultimately save a net present value estimated in the hundreds of trillions of dollars. Unfortunately, to be studied, this book must be read, and sadly, your author is a thinker, not a writer. Composing this volume has taken decades after the first insights from 1982. It was 1987 before the first tentative remedies were effected. Presently, a demonstration is underway in Hollywood among desperately troubled ladies. Apparent successes so far are Leelee Sobieski, Jessica Simpson, Mila Kunis, Lindsay Lohan, Paris Hilton, and Ellen Page. More will follow.
2

Science can be truly useful to humanity only when knowledge is unencumbered by legend, mythology, and untestable hypothesis. Arriving at truth can be a formidable process, too rococo for comprehensive disclosure. Chain thinking, concentration for many hours did help, as did (and one can almost hear legions of government-paid colleagues gasp) a deep and abiding faith in God. Do you see? Automatically, here thinking diverges from common notions. This is a book in the van of many modern medical miracles and there is no harm in giving credit where it is due.
Most human sociopathy stems from lack or pollution of family care. The primary pheromone deficiency driving sociopathy is of the father, but symptoms of paternal pheromone deficiency vary by other pheromone receptions, choices of self-medication, and behavioral self-stimulation. Human pheromones are here implicated in addictions of substances and behaviors as well as the immune & autoimmune diseases, and cancer. The eating disorders of obesity, anorexia nervosa, bulimia nervosa, are treated together. Excitement or adrenal addictions include afflictions such as gambling addiction, criminal thrill addiction, shopping addiction, all are substance addictions, just endocrine substances, and these are reachable by paternal pheromone therapy. Addictions to other pheromones suggest etiologies for homosexuality, pedophilia, habits of torture (sadism and masichism), the rapist’s addiction to female alarm pheromone, foot & hand fetishes for those pheromones, nymphomania and satiriasis. Meanwhile overt pheromone poisonings suggest etiologies for blighted ovum, sudden infant death syndrome (SIDS), diabetes, Alzheimer’s disease, multiple sclerosis (MS), differential susceptibility to infection, and much more. Pheromones regulate population fertility by adjusting perversion, criminality, drug addiction, insanity, etc. to hobbled population resources and needs.
The larger issues aside, this book is also full of tiny controversies, most of which go unresolved. Suffice it to say that whole books have been written about many of these. Obligatory mention merely indicates one’s familiarity with the literature, an unfortunate necessity in 2011.
Economic modeling, especially that of classical microeconomics, rules here axiomatically. This will foster charges of teleological thinking. While that cannot be
EXOCRINOLOGY
3

helped, it most certainly should not be deplored at least until these several fruits are tasted first.
Just as a grasp of organic chemistry rewards fresh beginnings, discovering truth means starting from a blank slate and rejecting unprovable propositions. Out went the self-concepts, the egos, the states of denial, psychological states, complexes, ideation and motives. If it didn’t have mass, value, or energy, then it did not exist. The author has striven to maintain his strictly empirical economic bias out of an abundance of caution. Surely, even the most indoctrinated psychological establishmentarian can admit that it is better to check out the physical & economic causes for behavior first. Resorting to some willy-nilly consensus of psychological opinion should be the last resort. Thus in this book the author has no personality and neither does anybody else. Of course, there is no convincing passionate defenders of air castles, but in America at least, it is a shame that our public authorities support and maintain ‘psychology’. The United States has a constitutional prohibition against establishing religions, but there they have managed it anyway. The psychological fiction is extremely well intrenched, even among otherwise staid and sober chemical ecologists. One thoughtful and incompetent chemist once offered this tidbit: ‘every blade of grass has its own personality.’ What is next, fellow? Rocks with egos?
Human pheromones are not always airborne. Indeed, most are not. Human pheromones tend not to be single chemicals, either. The kissing pheromone alone contains more than 735 different chemical species. Chapters 5, 9 & 10 deal with some human behavioral & other illness of presumed airborne etiology.
Pheromones have been subdivided dubiously into releaser and primer pheromones as explained in Chapters 6 & 7. Water soluble proteinaceous primer pheromones are extracellular receptors for the much smaller non-protein releaser chemical mixtures. Protein pheromone receptors can be self-produced and/or acquired in social behavior.
Generally, human pheromones affect only human beings and perhaps our parasites (dogs & cats) for which our pheromones are their allomones. There appear to be four basic human pheromones: love, anger, fear-stress-alarm, and the dimorphic
4

sexual aromas. Love, by far the most complex, takes up most of the pages of this volume, hence the title. There also appear to be roughly three basic receptivities: happiness, sadness, and romance. All three are accompanied by emotional lacrimation. No one should be surprised if every human inkling is laced with pheromones.
Love transmits the sebaceous skin surface and mucosal lipids via kissing of ‘kissable’ skin surfaces (inside the mouth and on skin where you normally kiss). The various human emission glands are the largest and most active of any species. (Heavy breathing airborne pheromones will be largely left to my gasping colleagues.)
Sebaceous secretions, these are the “king” pheromones that can controvert the effects of most others. Generally, these hundreds of different skin surface lipids together compose a surface pheromone set that is passed in kissing and differentiates us chemically to keep us kissing the right person and not kissing the wrong person or animal. Thus the pheromone may contain ‘yummy’ secretions and aversive pheromone secretions depending upon your age and station in life. Girls (and doubtless young men) are ‘yummiest’ in their late teens and early twenties when strange long chain strange wax esters accumulate on kissable skin surfaces with melting points that make those chemicals ‘melt in your mouth’. Anyone too young for reproduction will have an aversive pheromone component, probably the odd cholesterols peculiar to childhood that make kissing a child so boring and uninteresting or even nauseous. Childish pheromones passed among children, taints kin to us in adulthood, as a dispersal mechanism seen in tadpoles. The human skin surface lipid has unique human-only components, but pets and horses emit chemicals that come close, endearing them to us chemically. Rats and mice, too, have fatty acid and ester pheromone components that probably account for their seeking our human proximity for their habitat, endearing us chemically--to them.
Anger emits primarily from the top of the head (blowing one’s top, hot head), mostly from younger adult males and goes airborne at the slightly raised scalp temperature of anger. Anger’s emission appears poisonous, causing most of the autoimmune diseases and Alzheimer’s disease. The reactive long-chain conjugated
EXOCRINOLOGY
5

dienoic free fatty acids peculiar to the hair atop of the head of young men, ‘head’ the list of potentially dangerous human anger pheromone chemicals.
The fear pheromone has been observed in Austria and is now being synthesized at SUNY by DARPA at the author’s suggestion as a potent fear-producing pheromone that could reduce war’s lethality. The cold fear of panic is thermo-graphically observable on an infrared camera. Stress seems akin to fear, except that the lacrimation is pheromone-receptive. Stress and fear odors likely emit from the axillae, perhaps with a dorsal element (yellow streak down his back), but perhaps not. The fear-stress pheromone probably diminishes natural immunity and likely lowers T-cell counts as it does in laboratory animals. Diminished immunity caused by pheromones likely accounts for cancer reaching a clinically noticeable stage, and for dangerous susceptibility to infectious disease. Of course on the battlefield, fleeing cowards are shot down like partridges, often by both sides.
The body odor of happy, healthy young people appears conducive to good general health. This sexually dimorphic pheromone set should be reproduced synthetically and piped into hospitals and retirement homes. Perhaps visits by young people themselves can be dispensed with to avoid the potentially toxic anger emissions so poisonous to the hospitalized elderly and infirm. Basically, gruff young male doctors really should wear space-suits when they are angry.
Happiness is a receptive state of pheromonal contentment. It is having a recognized, unchallenged place in the local hierarchy, with pheromonal appetites sated. Producing this state artificially allows a ‘reset’ or perhaps a ‘rebirth’ that allows withdrawal from addictive drug cravings, along with criminal and perverse sexual appetites as well. Thus artificial human pheromone manipulation portends to revolutionize human society to cast aside most sociopathic ills. Of course the biological functions that those sociopathies had in population control will eventually have to be managed artificially. It is a dismal but likely prospect for the distant future.
Sadness or grief evokes psychogenic lacrimation and grief-specific postures and behaviors (boo-hoo-hoo), which set up chromatographic solvent chambers in
6

pheromone sensory cell (microvillar or brush-border cells) receptive tissues and structures. Emotional tears are the solvent and contain the first receptors in solution. The upper respiratory system is covered with pheromone sensory microvillar cells. These cells together comprise the largest pheromone reception organ of any animal species. The same cells inhabit the gut (Peyer’s patches) and thus might account for emotional nausea in response to perceived local hierarchical changes, as when spouses cheat.
Exocrinology is such a robust explanation for human behavior that we can dispense with some nonsense. Thus as stated for our purposes, emotional or ‘psychological’ states do not exist, per se. These are instead, emissional states and receptive states with specific empirical functions that glue us together in pair bonds, families, nations, and as a species. Even human pet ownership can be explained as allomonal parasitism.
EXOCRINOLOGY
7


Chapter 1: Kissing Kissing compares to crying, or laughing, yawning or hiccuping. We all do it. We always have. Nobody really knows why.
What is kissing for? Why do we put so much effort into the physical act? Why do our brains reward the behavior with pleasure? Kissing, especially passionate kissing between a man and a woman, can be a rather untidy affair. It involves risk, too. Saliva, passing back and forth, passes germs back and forth as well. Kissing couples share a risk of periodontal disease and cold sores (Herpes Type I).1 Herpes Type II and gonorrhea can also find their way to the pharynx. How is no mystery. The contacts are sexual.
Kissing also incurs what economists describe as opportunity costs. Humans have other things to do besides kissing. In simple terms, kissing takes our attention from food production and avoiding predators. Our effort and time in kissing costs us, so why do it?
Human fertilization requires no mouth to mouth contact at all; so for reproduction's sake, we could dispense with kissing altogether. The increased risk of disease infection alone would auger well for our avoiding the behavior.
EXOCRINOLOGY
9

Assume, quite reasonably, that time and energy are scarce. We should suspect that any non-kissing couple would improve their chances of reproductive success in a world of inefficient kissers. Yet, all human beings, everywhere, kiss.2 Indeed, kissers seem to reproduce far better in families than non-kissers. Do they not?
Kissing made it all the way into the present from prehistoric beginnings. For kissing to have gotten here, evolving over eons or arriving in one of Gould’s punctuated equilibria,3 kissing must justify its existence. It just costs too much. We've mentioned energy, time, infection risks, and lost opportunities. Moreover, the body's responses to and preparations for kissing, develop with unusual and costly intricacy. From the histology of skin to circulation and respiration, kissing transpires in coordinated effort. We will delve into this further on, economics, "satisficing" spandrels and all.
Let us look at kissing in detail. Kiss someone else's skin, the uppermost surface called the epidermis. Your mouth picks up skin oils, waxes and alcohols in a complex mixture from that skin you just kissed. The Romans named the mixture "sebum". The glands producing sebum go by the name of "sebaceous". Hundreds of thousands of microscopic sebaceous glands embedded in the lips, skin and interior mouth surfaces (mucosa) of the person you are kissing secrete the sebaceous skin chemical mix. When she kisses you back, your skin secretions get into her mouth, too. Perhaps a primed, mutual addiction to face and body greases binds couples in physical love?4 Dimorphism and perversion occur in humans, functioning every bit as kissing does. [For the time being, please assume the human male, heterosexual author's point of view, as it eases his writing. If the reader falls outside those categories, please try to understand at least until reading technology improves enough to adjust author viewpoint to reader tastes.]
If addiction joins us together, then we are not consciously aware of it. Non-conscious perception and recognition of human odors has been demonstrated.5 A mouse model is instructive, since pheromone reception is biologically ubiquitous. Since the hippocampus is required for learning one ordinary smell from another in mice,6 and since the infertility mouse-behavior-altering chemical, a pheromone, still works well when mice no longer have hippocampi,6 then conscious learning appears
10

not to be involved in pheromone recognition in mice either. The hippocampus is important in conscious learning, yet conditioning can be effected out of the consciousness of the subject.7 Similar experiments have seen similar results in hundreds of species.
Any chemical responsible for human love would have to be colorless, odorless, and tasteless. Human sebum is.8 Any chemical love would have to vary from kissed skin to kissed skin in amount and composition, so that different kisses would function differently. That way kissing someone on the forehead and kissing the same person’s tonsils with your tongue would perform different natural functions. Human sebum does differ, both chemically and in amount, from place to place on the human body.9
Similarly that variability would need to vary consistently from person to person. That way kissing your sister would be no fun, but kissing your "honey" would be. Human sebum has just that variability.10 There would have to be a whole lot of it at birth when only love holds death from exposure at bay. There is.11,7 There would have to be a lot of it on the skin around the time most people get married. There is.12 It would even have to stick to lips like nothing else in nature. It does.13 Finally, delivery of a bonding chemical would need to be highly specific. Yes, kisses are quite specific.14
Because each person's skin secretion mixture is unique, your sebaceous chemical signature is distinguishable from anyone else's.10 Those secretions mark your uniqueness over long periods, perhaps from fetus to grave. Primed, with reciprocal receptivity and exposure, the love in your mate-mate relationship might logically depend upon a non-conscious mutual addiction to each other's unique biochemical secretions of the dermis and its glands.
The more social birds kiss after a sort, too. Like those birds, human beings rub our beaks together most when pair bonds first form and at the reunification of lovers.15 Kiss your returning loved one and you feel so much better! Perhaps a
EXOCRINOLOGY
11

chemical obtained by the kiss drugs us into the feeling? The author's casual anecdotal experiments suggest this is so.
The heat of sexual passion is no generalized fever.16 Only some skin areas take part in passionate behavior. Overall body temperature remains stable. During intercourse some skin heats up, other skin cools down.17 Because hot skin moves sebum up and out of sebaceous glands,18,19 and because saliva dilutes and carries it so well,20 expression of sexual passion transfers skin chemicals quickly and efficiently.
(though love be a day and life be nothing,
it shall not stop kissing).
E.E. CUMMINGS
Perhaps compulsions for skin secretions compel people to linger together just like birds and bees? This author’s hypothesis for the case of birds21 has gained little acceptance. The avian case seems to be due to inertia set up by chains of citations from misguided Nobel laureates, but for the bees we may know. Blum22 found cutaneous surface secretion transfers and suggested from his experiments that they improved nest cohesion. Bravo Blum.
The chemical structure of the sebaceous chemicals compels us to believe they carry information, epigenetic information, and lots of it. (See Chapter 3) Such information carrying chemicals are called "semiochemicals" and sometimes "pheromones." Sebum contains semiochemicals. We know that much because dogs find sebum helpful to distinguish humans, even twins.23 Twins are identical genetically, but differ epigenetically with increasing age. Semiochemicals
12

communicate a biochemical signal (Greek semeion-- a sign) from one organism to another.24
Sebum may more narrowly be classified as a pheromone. Pheromones are chemicals that one organism emits into air, onto skin or onto environmental surfaces which influences the behavior or physiology of another organism of the same species.25 Conspecific specificity defines a pheromonal signal. Arguably these sebaceous chemicals and other suspected human pheromones may assuage the cravings that people have for parental approval, mates and offspring.14
People and animals have physical bodies with physical appetites. We see with eyes, like other animals. We digest food like other animals, we smell, hear, touch and taste all like other animals. Virtually every human physical function has an animal counterpart. Logic insists we assume physical human love does not differ fundamentally from animal bonding. Granted, parasitized people insist that their cat or dog loves them alone even with the obvious lack of conspecificity and despite the knowledge that dogs and cats mate promiscuously. The biological fact is that human beings support canine and feline parasites. Dogs and cats are parasites of human beings.
However, if humans form love-bonds in a way entirely different from the way animals form them, you would be surprised. Most other animals do bond if only for a short time. (Asexual reproduction, where the same individual is both "male" and "female," occurs in many animals: worms, some reptiles, many microscopic species and in much plant reproduction.) Many species mate for life. Other animals mate the opposite sex promiscuously.15 Observing the behavior of newly introduced animals calls to mind similar human behavior. Strange conspecifics are not allowed too close. Many marriages begin with a certain amount of distance, too. Closing that distance may require successful exchange of semiochemicals.
Some psychological writers insist human physical love differs fundamentally from the animal behavior, and that this defines humanity itself.26,27,28 Without evidence on either side, who can tell? Psychology must now become a failed paradigm belonging to the 20th century. Personalities, psychological states, egos, self
EXOCRINOLOGY
13

concepts, witches, spirits, ids, ghosts, and self-actualizations, these present cultural manikins lacking physical existence and should lose scientific credibility.
Much psychologist-collected data support critical periods in human pheromone recognition. Much of the psychological rationalizations remains suspect. However, almost certainly classical and Pavlovian conditionings take place behind each untasted morsel of sebum and behind each un-detected whiff of airborne pheromone.29 Unfortunately, psychologists manage to control for pheromone contamination or semiochemical sepsis only inadvertently. It is as though they have yet to detect the air around them, oblivious to the thousand or so chemicals and microbes with which they mark their own homes, their lives, and their loved-ones. Psychologists have been amazingly witless.
From available evidence, for birds and bees and human beings, falling in love means falling in love. Getting hooked on the addictive bloom of the partner's face, mouth and body must be similar for all bonding species. The biological devices thought to cause bonding look most alike in animals nurturing young and forming mating pairs or groups.
"Mutual licking and grooming sequences occur very commonly in mammals. The male hunting dog sniffs and licks the female vulva increasingly as estrus is approached and will even vigorously nibble at the female's skin from the ear to the groin (van Heerden, 1981). The asiatic elephant transfers chemical information from its mate to its mouth using the tip of its trunk (Eisenberg et al., 1971), black-tailed deer sniff and lick each other's tarsal scent organs (MüllerSchwarze, 1977) and the male golden hamster is motivated to lick its mate's vaginal secretion (Johnston, 1974)".30
A passionate kiss synchronizes the bodies' cardiovascular, exocrine, respiratory, and neuromuscular systems. Of course, behavior of two people must also be synchronized (as courtship). Consider this. Both kissing mouths contain soft tissues and sharp teeth that cut. Moving all these structures around so close together requires considerable instinctive orchestration. The physiology and ethology of kissing needs increased scientific attention. All the sharp teeth and vulnerable tissues never cause serious injury.
14

Inarguably, one kiss moves sebum from one person to one other person. Transfers of pheromone from skin to mouth have advantages and disadvantages over pheromone dispersal into the air. Kissing allows specific reception of an individual's chemical signals. You can be smelled by a whole room-full of people all at the same time, but kissing needs permission. That distinction signifies something important in many mammalian species.
"Males of nearly all mammalian species attempt to sniff the genital region of potential sexual partners. ...In many species females that are in estrus or approaching it allow males to investigate their genital region, whereas females that are not in this state avoid or repel such attempts. ... Thus, permission to investigate can be considered a subtle but important courtship display since it indicates an approaching readiness to mate".31
A mouth-suction applied to the skin typifies human kissing. Suction helps to get skin sebum, the putative pheromone, into the kissing mouth. However, the slight sucking in such kisses seem insufficient for efficient inhaling or swallowing. At least some receptors must be either in the mouth or close by in the upper respiratory system and that is where they were found. Eureka! The human vomeronasal organ, a pheromone reception device in all other species, seemed a likely candidate for a low volatility pheromone reception in humans.32,33 Sebum has low volatility.20 However, the sensory brush border or microvillar cells that line most of the upper respiratory system and into the digestive tract with Peyer’s patches (that ‘kicked in the gut’ pain of betrayal?) must certainly be pheromone receptors. They are identical to known specialized pheromone receptor cells in all other species of animals. The narrowness of medical education perpetuates an absurd debate about the function of these cells that should have closed long ago. Physician cultures abhor labeling incompetence so much that the pheromone reception system has gone unobserved for generations. As my character Forrest Gump decried, “Sometimes there are just not enough rocks” with which to stone ineptitude.
EXOCRINOLOGY
15

Long periods of passionate osculation in mate-mate bonding stimulate salivation and frequent swallowing. Pheromone reception and immune response might also take place via the alimentary tract by absorption through the duodenum. Relatively small amounts are involved, but pheromones are certainly the most potent drugs ever discovered.
Could a single kiss convey information? Clearly, yes it can. Insects communicate with exceedingly small quantities, far less than the amount of sebum transferred in a single kiss. Pheromones signal efficiently.25 Insects communicate at the ultra-trace level (PPB, parts per billion air molecules).34 Rodents communicate well with pheromones emitted at the trace level (PPT parts per thousand molecules of air).35 Very small quantities of supposed pheromones of human origin have elicited electrical responses from the human vomeronasal organ.33 The physiological evidence is there, if one only looks.
LIPS
For about the past thirty years, a brouhaha stewed among ethologists about why people have lips on their faces. (This is one among many petty controversies.) Other primates copulate with their females from behind and in some of these species genital coloration acts as a sexual display. Women's lips attract men, especially vaginal ones.
16

Perhaps human lips played counterpart to the gaily colored "sexual skins" of primate genitalia? Lips do have a distinctive red color, at least among light skinned people. Maybe lips evolved as bait to bring the male around to the front?36 The counter-argument ran that white men also have red lips. Furthermore, the poor chaps who couldn't differential between vaginal lips and facial lips (and male lips and female lips) were poor candidates for evolution anyway,37 and back and forth. Very funny debate, it was, if a bit tedious.
Perhaps lips evolved to do what they do? Lips puckering-up for a kiss do three things. First, they bring the most available surface area to bear. Second, they maintain and only expose their moist inside surface just before contact. Both devices improve transfer. Third, lips act like a flexible gasket around osculating mouths. (Kissing may get sloppy from so much saliva, but kisser's drool may be excitedly lapped up, posing no contradiction to the gasketing hypothesis.) If kissing executes a valuable function which improves by having fleshy glabrous facial lips, then the purpose for their natural selection might be inferred.
EXOCRINOLOGY
17

RITUALIZED FEEDING?
Scientists, ethologists, have observed a step-by-step progression of behavior from eating through kissing in humans and other species. Admitting they possessed no hard evidence, human ethologists suggested that our kissing must have evolved from feeding behavior. "All animals eat and not so many kiss," they reasoned, "Look at all the cases where mothers chew food before passing it to their little ones by mouth. Humans aren't the only ones." They termed instinctive osculation, ritualized feeding behavior.2,37,38
Eibl hypothesizes that kissing resembles eating. Indeed, the eating of small quantities of an extensive mixture glues us together by bonds of addictive craving. Mothers in many cultures do pre-chew their baby's food.2,37,38 However, perhaps babies have difficulty chewing while toothless? Pheromonal emissions transferred in kissing reward caregiving well. However, the weight of evidence to this writer suggests that kissing evolved from allogrooming.
Kissing behavior changes somewhat from culture to culture, but the feeding on sebaceous pheromone represents the universal and constant idea. The evolution of pheromone exchanges must have started with the first colony of bacteria, for colonies require pheromones to communicate well enough to become colonial. Many of the pheromones of colonial species served later as hormones, neurotransmitters, cytokines and the like. In at least one species, the pheromone that signals a school of organisms to scatter is the same chemical that signals pain when it is released inside the body: bradykinin.39 Indeed, alarm pheromones generally are associated with immune function.40 It should come as no surprise that NSAIDs -- aspirin-like drugs -- affect pheromone reception -- and therefore the diseases affected by NSAIDs have pheromone, at least alarm pheromone, components.
Mind, the ethological hypothesis of kiss as ritualized feeding has appeal. Who calls you "Honey," "Peaches," "Sweetheart," "Sugar," "Cupcake," "Dumplings, "Sweetiepie," "Lollipop," "Honeybun," or "Lambchop?" Why should tasty food be so
18

commonly used for romantic pet names in all cultures? The association may have significance if a feeding of some sort takes place. On the other hand, loving a "dried-up old prune," may present problems. One should expect low sebum secretions in people difficult to begin loving and that has generally proven true, if only anecdotally.
ATTRACTION
Attraction is a separate issue and will receive little attention in this book. Attraction in human beings may be another process dominated by visual cues. You will find fuller discussions of issues of attraction elsewhere. Airborne pheromones seem to be important for sexual orientation,14, 41, 42 conviviality and sexual attraction in humans and to moderate43 or synchronize female menstrual cyclicity.44, 45 Airborne pheromones by worried people or people in pain sitting in cancer ward waiting rooms may diminish immune response by diminishing natural killer cell immunity and T cell counts.46 The human alarm or fear pheromone47 emitted by frightened people may help cause generalized panic while addiction to it may perpetuate human kidnapping, rape, and torture behavior.
The relative size of the bed nucleus of the stria terminalis is associated with sexual orientation. Hormonal manipulations and neuron tracking have shown that information concerning pheromone emissions is carried there.48
SEBUM
As we've said, "sebum" in Latin means "grease." Calling sebum a face-grease does injustice. What a marvelous material! Does sebum speak the language of love? The evidence, reviewed in these pages, suggests so. This skin-produced, soothingly addictive aphrodisiac makes love possible, and medical miracles like dramatic improvement in Alzheimer’s cases, cures for drug addiction, perversion, and criminal behavior.
EXOCRINOLOGY
19

Sebum seems to be tailored for exchange by kisses. Sebum is extremely hydrophilic (water loving);20 indeed, it was once considered the most hydrophilic natural organic mixture known to science.49 Plainly, sebum rides atop saliva, which is mostly water. Sebum's affinity for moisture insures its transfer from the relatively dry skin surface of its origin to saliva covered, 'moistened' lips kissing that surface.
Good reason may exist for wet kisses. In females, salivary sulfur-containing breath volatiles of microbial origin vary cyclically with circulating hormone level, as do vaginal levels of lactic acid. Physicians exploit the effect to detect female ovulation. Other chemical signaling may be going on in saliva exchange kisses. Rates of intercourse vary cyclically with peaks at ovulation and premenstrually,50 although these may have more to do with menstrual period discomfort. One of two cyclical secretion peaks might portend the period of peak fertility and intercourse.14
Beasts have their own distinctive sebums, too.51,52 Do animals exchange unique but still similar substances in their own kissing-type behaviors? They "groom." They "preen." They "give nuptial gifts" to each other. Are these behaviors their kiss-equivalents? Nuptial gifts get covered with the animal's own sebum and no mating takes place without this excited transfer.14 Of course it is exactly the same thing.
PHEROMONES
Those readers already familiar with pheromones tend to recall them as chemical attractants. Chemical attraction was first observed in butterflies. In the fifteenth century, chemical attraction in moth and butterflies was first noticed.53 In 1690, John Ray reported seeing several Biston betularia (L.) flying around a visibly obscured caged female. Attempts to figure out the chemicals responsible began in the nineteen twenties as an adjunct of agricultural research.54 Many tried. For years they had no luck. At last in 1959, Butenandt et al.55 identified the first sex pheromone, E,Z10,12-hexadecadieneol. They named it "bombykol." Getting enough crude extract
20

took twenty years to collect and the sacrifice of the pheromone producing glands of around half a million female moth sex organs.56
Human sebum functions similarly to other animal's fur and skin pheromones but can be easily distinguished from them. Lions respond to lion sebum.57 Moles respond to mole sebum.58 Mice respond to mouse sebum.59 Tamarins respond to tamarin sebum.60 Geese respond to goose sebum.61, 62 Gerbils respond to gerbil sebum.63,64,65 Goats respond to goat sebum.66 Even yeast cells respond to tested unsaturated fatty acids of the type seen in sebum.67 Ordinarily no confusion presents difficulty.
Allelochemicals (or allomones) are semiochemicals that communicate across species. The skunk's defensive spray holds all too familiar allelochemicals that dogs (and people) soon learn to avoid. Hunted animals may warn comrades with an alarm pheromone scent that becomes an allelochemical trail for the predator.68 From economics, the author expects that crowding should increase the detectability of the spoor. Similarly uncharacterized human alarm substances may lower our disease immunity, while conspecific sebum consumption may improve it.69
Dogs smell well. Fleeing prey lay down chemical trails as pheromones to remind themselves and their fellow conspecifics to avoid the area for a time where a predator was detected. Dogs smell these and other scents left by the withdrawing animal as allelochemicals. Canine olfactory versatility trades off against the economic efficiency of specialization. The compromise produces a device less efficient and more expensive to maintain than the more specific recognition devices used for pheromone recognition. Bloodhounds on the chase compensate. They must keep their nostrils close to the scent and training for the hunt is both time consuming and costly.
Dogs recognize humans easily, detecting sebaceous fatty acids at concentrations of one percent of the lowest conscious human perception. They can distinguish among family members and even between identical twins.70 Sebum might even be utilized by pets to synchronize their behavior to ours. Economics predicts
EXOCRINOLOGY
21

that disease organisms, particularly more sophisticated ones like malaria and worms, should do this too.
Various sebums are semiochemical in many mammalian species, including at least one other primate species.71 In the past, scientists differentiated semiochemicals from insect pheromones. Mammals respond to semiochemical stimuli differently depending upon the behavioral context, while pheromones (as thought back then) had robot-like effects on insects. Further study revealed more similarities than differences, so the distinction lost favor.
SEBACEOUS GLANDS
Sebaceous glands differ morphologically from animal species to animal species. The glands look a bit different.72 They stain a bit different on the microscope slide.27 Different glands do different things. Generally animals' sebum marks territorial hunting grounds, nests and trails5, as well as individual group members.25 This need for persistence may account for its being among the less volatile organic excretions.9
Sea birds such as heron gulls take sebum from their sebaceous rictus and preen glands to mark twigs, grass, and pebbles over their nesting territories.21 Recognition of these pheromone deposits give birds confidence or timorousness depending upon whether the deposit tells them they are standing their own ground or intruding. "Most animals use their own odor to permeate their environment, not so much to repel intruders but rather to tend to their own comfort.21 Similarly, deposits and emissions inside our dwellings may make us feel safe and secure living there. Who cannot remember a sleepless first night in a new home?
Thousands of ornithologists observed, filmed, and analyzed avian border marking behavior only to agree with Tinbergin that the processes were totally meaningless. Nikko Tinbergin (winner of the 1973 Nobel prize in physiology or medicine with Karl von Frisch and Konrad Lorenz) was among the first to apply ethological tools in ornithology. He was one of those who coined "displacement
22

activity" for supposedly meaningless behavior. Naming differs fundamentally from explaining. Careful catalogs compiling millions of hours of watch time were to no avail against scientific inertia. Tinbergin's untestable hypothesis stood unchallenged in our modern age for 50 years, a testimony to the enduring influence of this great pioneer. Of course, he was totally wrong. Marking territories with chemicals that humans cannot smell well during ‘displacement activity’ is the basis for most nesting territoriality and behavior.21
Skin sebaceous glands are ubiquitous in mammals, have essentially the same metabolic characteristics, and produce secretions of complex composition. Specialized sebaceous glands constitute a major proportion of the presumptive mammalian marking and scent glands. Although their secretions are, apparently, even more complex they are almost certainly variations on the common sebaceous metabolic theme. Since this metabolic capacity is sufficiently adaptable to provide scent glands for multiple species, it is not unreasonable to assume that it can also provide several odor cues within a species.73
Tiny sebaceous glands embedded in and opening onto the skin and hair produce the tiny amounts of sebum needed to keep people together (and coming back for more). Roughly two grams of sebum get secreted onto skin, clothing, and bedclothes every day.74 These glands, the sebaceous glands, appear on skin that human beings kiss most.4
Think of skin we humans kiss often. Does kissing there vary from culture to culture, from climate to climate? With no one measuring how many kisses go where for how long, the question goes unanswered.
Ordinary human experience allows construction of a likely scenario even without hard data. Some skin gets few kisses. For instance, who kisses elbows? You might expect the elbow to have a dearth of glands, and that is so.8 There are almost no glands on the elbow. Likewise, there are few sebaceous glands on the palms of the hands and on the soles of the feet.
In animals other than humans, variations in gland prevalence and morphology show corresponding differences in gland secretion, composition, and function. Different gland shapes generally mean different secretions. Sebaceous glands
EXOCRINOLOGY
23

fulfilling different functions differ in size, shape, number and location. They produce secretions at differing rates and compositions. Glands vary anatomically. Different glands do different jobs. Glands that open onto virtually all hair follicles and secrete human sebum onto clothing and bed linens differ from those that open directly onto skin surfaces. Both of those are larger and less symmetrical than those invading mucosae inside the mouth or elsewhere.
Some specialized sebaceous-type glands probably have mundane functions. The cerumenous glands probably just make it easy to shake the water out of our ears. The Meibomian glands of the eyelid seem to contribute to maintenance of tear-film stability over the cornea. Who knows? Some insist even today, that the specialized glands in the human integument or skin may contribute nothing to kissing, but the preponderance of the evidence, much of which is anecdotal, says kissing is biologically important. Economics insists that it is.
However, surface-opening capillary-encapsulated sebaceous glands do clump together in large numbers. Such gland aggregations appear on the scalp, forehead, cheeks, nose, at the outside edges of the lips, on the neck, and the female nipples and areolae. Face and scalp sebaceous glands can be differentiated on a histochemical basis.75 Also, please see figure in Albone’s book. It shows that differentiation can also be made from the presence or absence of certain androgenic steroid metabolizing enzymes.30 The glands in and around the female mammary spots (the vermilion areolae & nipples) differ physiologically from their kin. Note that sebum glands of unusual bi-lobular shape also open onto skin surfaces all around inside the mouth on the buccal and oral mucosae. Others open onto the contact surfaces of the mature sex organs, as still others coat pubic hair.
Put Figure 3.5 from page 58 of E.S. Albone, 1984 about here. It must be reproduced with permission from Baillie, Ferguson and Hart (1966), Developments in Steroid Histochemistry, Academic Press, Inc. (London) Ltd.
24

Although no one knows the chemical composition of the various oral sebum glands' secretion, it probably differs from those found on the face. This would follow the rule observed in other animals where gland physiology and chemistry vary together with function. Specialized use would help to explain why these specialized mate-mate bonding glands are, like female areolar glands, hidden away and protected from indiscriminate licking in our clothed cultures.
"In man, the largest and most numerous glands are in the skin of the head, forehead, cheeks, and chin 400 to 900 glands per 1 cm2 as compared with less than 100 per 1 cm2 in some parts of the body76 and none at all on palms and soles".73
SEXUAL GLANDS
Skin surface sebaceous glands of varying morphology thrive where most kissing takes place: face and head, lips, inside the mouth on mucosal surfaces within tongue-reach of an osculation partner, female nipples and areolae. Specialized sebaceous glands line the sexual contact surfaces of the uncircumcised penis tip (the preputial glands) and vaginal labia and are also known as Fordyce spots. Their appearance with sexual maturity is no coincidence.
The sebaceous glands of the female sexual organ (Bartholin's glands) secrete significantly only during the plateau phase of sexual intercourse. Thus they are worthless for lubricant.77 Very few glands inhabit the clitoris,78 which swells with excitement but does not heat.79 That means heat increased sebaceous secretion can not be related to tumescence, except where increased circulation suffuses the peri-glandular vasculature.80 We must strongly suspect mutual pheromone exchanges between an uncircumcised man and a woman during sexual intercourse, owing to the extreme heat and immediate contact of gland filled skins. The unusually lengthy plateau phase in human intercourse35 may facilitate communication of both Bartholin's secretion to the penis and the smegma of the preputial glands & Fordyce spots of the penis to female vagina.
EXOCRINOLOGY
25

At sexual intromission, too many sebaceous-type glands contribute to semen for simple nutrition's sake.34 One hormone found in semen and elsewhere, prostaglandin, incidentally released by female goldfish as a pheromone, stimulates male spawning behavior. With other similar findings it has been established that hormones and their metabolites may commonly serve as reproductive pheromones in fish.81
From the author's examination of sebaceous chemistry (see Chapter 3), these glands can move large amounts of information back and forth. Note that urinary pheromones affect oocyte selection in ovaries and they also can block pregnancy in rodents.82,83,84 Information brought by frequent human intercourse could epigenetically select sperm and perhaps even influence chromosomal selection after syngamy, an epigenetic function. Fetal development and subsequent behavior of the partners may also be affected in humans making economic use of available biological information. While mechanisms for pheromone exchange were hypothetical in 1991; human glands, human secretion and animal examples did exist, even then. We know that pheromones effect changes in the quaternary structure of DNA and the proteins surrounding DNA epigenetically.
A pheromonal use might help to economically justify the high frequency and low fertility of human intercourse. Polyandrous and promiscuous cultures are less fertile than monogamous ones.2,85 Perhaps a destructive semiochemical sperm competition explains this? Competition may be overemphasized. Instead perhaps a pheromonal cooperation reduces fertility? Semen from more than one man may either compete or cooperate in the female reproductive tract. First sex after a period of separation excites a greater volume of semen.
FACIAL GLANDS
As already mentioned, saliva moves sebum around. Lips may conserve mouth moisture when not kissing. Lips maintain a pheromone-grabbing, wet, sucking surface always available. Additionally, excited facial lips become hot lips. They heat in passionate courtship behavior. Hot lips further encourage sebaceous transfers.
26

Honeycomb drips from her barbarous lips And her palate is smoother than oil: But her tongue is as bitter as wormwood, And sharp! Like a two edged foil. Proverbs Chapter 5, Verses 3 and 4.
Puberal onset sebaceous glands line the soft, moist, glabrous mucosae that together make up the inside surfaces of the mouth.7 Glands also invade the gingival mucosa in the interior cheek mucosa. These "adult osculation glands" form a rough line with the molars and incisors in the oral mucosa.
When visible these unusually shaped sebaceous osculation glands of the sexual contact surfaces and inside the mouth on the oral, buccal and gingival mucosa are called Fordyce spots. Most of those in the mouth lie within a tongue-length of another human being kissing that mouth. The unknown rhythms of Fordyce spots deserve study.
What does a passionate "French" kiss accomplish? Are we trying to lick and suck those glands clean of sebum with our tongues in each other's mouth? The adult develops a specialized surface on the tongue in puberty38 arriving with the glands just in time for this behavior. Perhaps it is a mechanically lipophilic surface?
Most body tissues develop from fetal precursors inside our mothers' wombs, but the mucosal sebum glands seem to pop out of nowhere, de novo, right about the time open-mouth kissing starts in earnest: at puberty.8 Sebaceous glands of pubertal genesis number among the so-called "accessory sexual" glands.12 The sebum glands lining the mouth (and elsewhere) are especially active during the late teens and early twenties.8 Early adulthood marks peak human fertility. Also in early adulthood, kissing frequency peaks and people choose mates for themselves.10 Of course, the peak of sebum secretion is shared with peak rape frequency, too. The pinnacle of reproductive fitness86 sexual behavior10 and the highest sebaceous secretion8 are simultaneous.
EXOCRINOLOGY
27
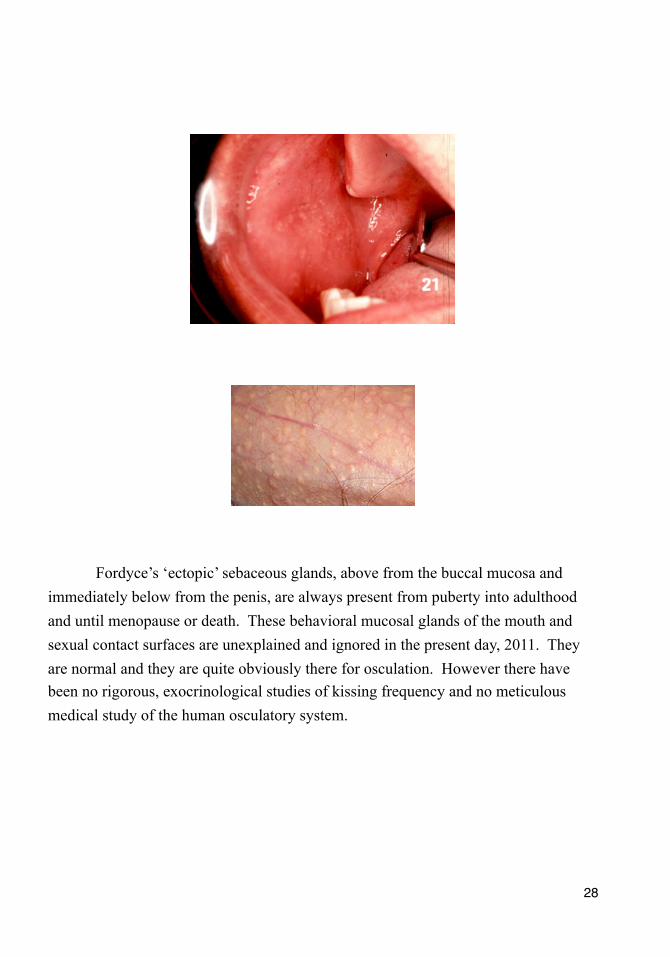
Fordyce’s ‘ectopic’ sebaceous glands, above from the buccal mucosa and immediately below from the penis, are always present from puberty into adulthood and until menopause or death. These behavioral mucosal glands of the mouth and sexual contact surfaces are unexplained and ignored in the present day, 2011. They are normal and they are quite obviously there for osculation. However there have been no rigorous, exocrinological studies of kissing frequency and no meticulous medical study of the human osculatory system.
28
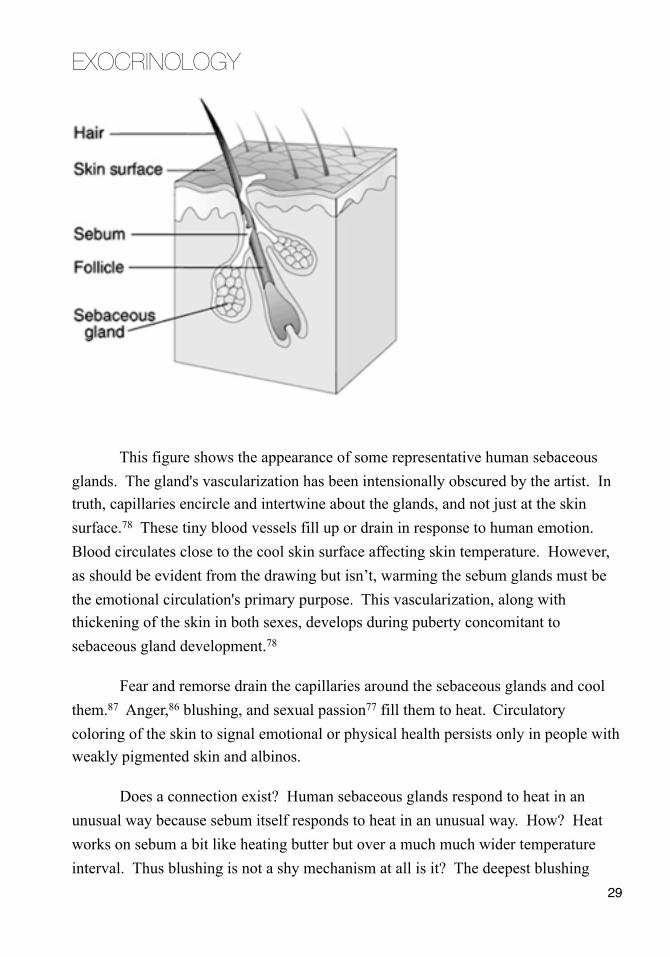
This figure shows the appearance of some representative human sebaceous glands. The gland's vascularization has been intensionally obscured by the artist. In truth, capillaries encircle and intertwine about the glands, and not just at the skin surface.78 These tiny blood vessels fill up or drain in response to human emotion. Blood circulates close to the cool skin surface affecting skin temperature. However, as should be evident from the drawing but isn’t, warming the sebum glands must be the emotional circulation's primary purpose. This vascularization, along with thickening of the skin in both sexes, develops during puberty concomitant to sebaceous gland development.78
Fear and remorse drain the capillaries around the sebaceous glands and cool them.87 Anger,86 blushing, and sexual passion77 fill them to heat. Circulatory coloring of the skin to signal emotional or physical health persists only in people with weakly pigmented skin and albinos.
Does a connection exist? Human sebaceous glands respond to heat in an unusual way because sebum itself responds to heat in an unusual way. How? Heat works on sebum a bit like heating butter but over a much much wider temperature interval. Thus blushing is not a shy mechanism at all is it? The deepest blushing
EXOCRINOLOGY
29

pumps the most sebum up onto the very skin at the very time and place where important kisses are most likely. If you like someone a lot and want them to like you more, and if that person is about to kiss you: blush! Would anyone insist that blushing is merely a color signal just in white people?
PASSION
A functional basis probably exists for the heat of "hot dates" and the "sex flush" of sexual intercourse noted by Masters and Johnson in Human Sexual Response.16 During passionate heterosexual courtship behavior, increases and decreases in skin temperature can be recorded,77 and heat releases torrents of sebum.18
Passionate heat makes sebum secretion available in the right place and time to gush sebum under the kisses of lovers. The warmth of affection then, would be functional. The hot surfaces so described and the areas of highly vascularized sebaceous gland aggregation are one and the same.4
As already noted the heat of a passion produces no generalized fever. Although some would say their brains burn with passion (in rats the hypothalamic circulation actually cools88), only some skin areas warm. When kissing, the skin temperature increases become noticeable. (Pheromones produce hyperthermic responses in laboratory animals89 suggestive of the fever response to chemicals of pathogenic origin, and helping to explain the anti-pheromonal effects of NSAIDs. Some children with autism spectrum disorders (ASD) exhibit improved behaviors and enhanced communication during febrile episodes90) The brain responds instinctively to romantic signals and sends the blood to be circulated there.
Usually the capillary network just under the skin helps regulate body temperature. Is lovemaking so strenuous? During a hot date, some skin heats up while other skin areas, like the belly button, cool down (although kissing the bellybutton of a passionate, aroused female might be considered inappropriate and less rewarding than kissing her elsewhere.) The hot skin is the kissable skin saturated with furiously 'pumping' sebum glands. Glands secrete continuously and passively.
30

They have no muscles, so no sphincters. Afferent (inbound toward the brain sensory) neurons innervate some glands. We know not why. Perhaps it is for tingles?
The passionate heat increases sebaceous (sebum) secretion abruptly for a short period. The relationship between skin temperature and sebum secretion rate is highly significant. Heating and cooling produce secretion rate changes of about 10% per 1o C.18
Human adult kissable skin seems to have a ready reservoir of sebum.91 Does the reservoir in the pores of the skin hold sebum for moments of romantic ecstasy? The highly elastic sebum secretion rate with respect to temperature changes and the heat of passion work in concert to encourage timely sebaceous secretion, kissing, and bonding.
A LITTLE MORE ABOUT GLANDS
Since about the time of Frederick the Great, scientists have wondered what these glands were doing on the skin in so many strange places.92 There were just too many glands and they were too active to be dismissed as trivial. Nor did they seem to be mere vestiges from earlier stages of evolution when they were more important to the body.12
While many dermatologists finally suspected that the glands communicated some biological message, that message escaped them.93 Dermatologists as a group seem determined to avoid behavior issues.
OTHER IDEAS ABOUT SEBUM
Was sebum also a lubricant? Was it some sort of moisture barrier? Perhaps an emollient? Or was it merely a nuisance? One by one, those ideas were tested and dismissed.76
EXOCRINOLOGY
31

Does sebum enhance water resistance in other species? Water birds come to mind. The feathers of waterfowl need no waterproofing.59 Deposits of preening sebum onto them do nothing other than impart a characteristic bird stink. The arrangement of the feathers is important, however.94
Is sebum a natural waterproofer? As mentioned, even in waterfowl, feathers do not depend on sebum to shed water. Does sebum help the body retain moisture? Maybe it does a little. Unfortunately for that hypothesis, an effective sebaceous moisture barrier needs a thickness of sebum an order of magnitude thicker than the normal sebaceous film. According to both Kligman76 and Rothman13 only a sebaceous layer at least ten times normal would be required to impede moisture transmission even slightly. Skin moisture does not need sebum as dry skin may or may not be sheathed in it.95 Only at birth sebum might reduce water loss significantly. The newborn's covering of greasy pheromone may be thick enough to contain some moisture.
Medical texts make sebum out to be fungicidal or bactericidal. That is an unjustified belief that must stop. Nothing in sebum goes out of its way to kill fungus or bacteria. Whether sebum is on the skin or not, commensal bacterial growth does not respond to its presence.96 Cove et al90 should have ended the controversy, but people still publish nonsense.
Bacteria and fungi find the skin a mechanically dangerous place. The skin is like a table top. Nothing much holds moisture there. Bacteria and fungi falling there dry up (desiccate) and just blow away.76
Does sebum act like "false food" to keep bacteria or fungi populations static? Some evidence supports this view.10 However, consider this. Would being appetizing to fungi or bacteria recommend itself for a pheromone constantly exposed on the skin? Pheromones of at least one species do have some anti-fungal properties.97 One does not preclude the other indeed, all naturally occurring antibiotics are probably semiochemicals in their own right.
32

Lastly, Guthrie98 thought sebum helped make unwashed faces menacing somehow. Of course, the facial skin surface lipids have no color, no odor, and are virtually undetectable except as a slight shininess.
SEBUM SECRETION AND SWEET KISSES
Rates of secretion onto the skin surface also vary. They yield variation both in composition and abundance.9 Maybe different sebums effect different bonds depending upon anatomical origin of the sebum? Human beings form a variety of love bonds. We kiss other human beings with varying techniques and at differing body locations depending upon sex, kinship, age, and even time of day. “Time of day?” you ask, try scheduling a prom dance at 8:30 am! While culture differences are found, kissing remains fairly consistent from culture to culture.83 We have one kiss for brother or sister and another kiss entirely for a lover. You do not "French" kiss your baby or your grandmother. The distinctive anatomical sebaceous compositions with kissing technique may foster distinctive bonds for social functioning.
When women kiss it always reminds one of prize fighters shaking hands. H. L. MENCKEN
The sebum secretion rate increases during the last trimester of pregnancy, despite high levels of estrogen in the blood, a hormone which normally inhibits sebum excretion.99 The same elevated sebum secretion rate stays high during the time a mother nurses from her breasts.100 Are pregnant and lactating women more "kissable" than non-pregnant, non-lactating women? Enhanced bonding effort during the period when reproducing females are most vulnerable seems both logical and economical.
EXOCRINOLOGY
33

GETTING THE CHEMISTRY RIGHT
Romantic kissing does not bring on true love in every case. Why not? Even though mutual attraction usually precedes romantic kissing, kissing does not reliably predict reproductive success. Primed human kissing (from circulating fetoproteins, lochia, and/or colostrum), may effect parent-infant bonding, and primed kissing (from previous sexual experience) predicts continued successful mating in humans. Other influences affect pheromone receptivity, however, including endocrine state and even air pollution.
The chemical distinctions in sebum developing at or near puberty may hold at least part of the answer. Indeed, such changes distinguish mature from immature sebum.101
The sebaceous wax ester component increases around puberty.73 The same esters distinguish the vernix caseosa of full term neonates from premature ones.102 Among all pheromones known, esters are pheromones more often than any other chemical functional group.103
Wax esters are just what they sound like. They are waxy substances. Wax esters in sebum "melt" at slightly different temperatures in their turns as gland temperature rises. In adults, particularly young adults, these sequential liquefactions give the rush of sebaceous pheromone that comes with the hot skin of passionate kissing.
Once their parents are hooked, offspring have no need for such heady stuff as passion, so wax proliferation awaits puberty. Does that sound reasonable? Free fatty acid information maintains some consistency,104 but the composition of newborn waxes vary greatly from those of puberty and of adulthood (Pochi, Peter E., et al., 1979; Nicolaides, N., et al., 1972). Thus very young couples with still developing sebaceous compositions may suddenly find themselves poorly matched as they grow older. Marriages of very young teens do tend to end in divorce. Changing sebums, changing love: have they something in common?
34

The sebaceous composition of the adult differs from that of the infant, the child and the pubescent.96 Perhaps aversive pheromones, which protect the young in other species, may also linger into human puberty? Thus when one mate is very young, chemical compatibility difficulties may also be encountered?
However, most couples are of comparable ages. Perhaps genotypic or environmental information codes into sebum105 and vice versa? A chemical compatibility may be a prerequisite for love. A mismatch might signal a fizzle. If so that might explain why even some mating kissers fail to love. The chemistry just wasn't right. This suggestion that pubertal pheromone changes, with regular seasonal fluctuations, encode genotypic or other information into our skin secretions to influence mate choice appears logical. A selectively attractive secretion might enhance "hybrid vigor".97
Hybrid vigor results from a cross between two members of the same species that have very different genotypes (or gene makeups). An example may serve to explain. Take three groups of pea plants, "A," "B," and "Hybrid AB." One group, group "A," had parents that were closely similar genetically; say they have group "A" typical traits.
Take another similar group of pea plants having different characteristics from "A." They are group "B." Now take the offspring of cross-fertilization of both groups "A" and "B," the "Hybrid AB" group. They have had one parent from the "A" and one parent from the "B." Expose all three groups to a destructive pea plant virus. Which of the three groups remains healthiest? "A?" "B?" or "Hybrid AB?"
We are talking about hybrid vigor, right? The "Hybrid AB" group with hybrid vigor has dissimilar parents. Axiomatic among animal and plant breeders, hybrids remain strong, vigorous, and prolific while purebreds are weak, spent and infertile.
What is more, inbred populations are inherently riskier. We know that inbreeders carry more deleterious genetically inherited traits, and biological devices to avoid inbreeding might be expected.
EXOCRINOLOGY
35

I wasn't kissing her, I was whispering in her mouth.
CHICO MARX
Hybrid vigor does not follow automatically from sebum uniqueness. Merely encoding genotypic information into our kissing sebum would not do the trick. Preferring non-kin secretions must complete the picture. How?
An elegant murine experiment was set up as follows. A female mouse could follow either of two aromas. She could follow the scent of a male from her own identical strain or she could approach a male from a strain differing from her own by only one gene. She invariably chose to approach the non-twin.106
While the hybrid vigor scenario appears to make a tidy argument, there are counter examples that support another hypothesis, which may fit the facts a bit better. What about the Kibbutz children? Israel has Kibbutzim, or communal farms, where all the children live together. Adults share child-rearing. Such kibbutz children live with unrelated children as if they were all one family. Children raised together, related or not, tend not to marry each other. Instead they marry outside the group.
Consider also those living in isolated communities on islands, in high mountain valleys, or under constraint of religious proscriptions, who suffer a risk of inbreeding. Such people might be expected (at least non-consciously) to avoid genetic illness, provided they had the information to do so.
Remember Oedipus and Electra? The existence of incest in ignorance shows counter-evidence. While inbreeding might be answered economically as making the best of a bad situation, incest in ignorance cannot be.
Perhaps the genotypic information idea is a bit too facile. Genotypic information may not necessarily be the signal conveyed at all. Perhaps the appetite is just for something strange? This chemical recognition from association may be immunologically determined from prior inoculation. An appetite for alien sebum
36

would assure that hybrid vigor would be achieved most often. Tit for tat, hybrid vigor assures greater success of offspring inheriting genes for genotypic pheromone signals and alien appetites.
A love pheromone of non-self and non-family (or non-kibbutz family) may serve for attaining hybrid vigor. No genetic encoding of sebum would be necessary beyond randomness. Kissing your sister may be distasteful only because you chemically recognize her from association as something to avoid. Kissing children romantically must be distasteful for another reason: an aversive pheromone. The best candidate seems to be an unusual, childhood only, skin surface emission of a specialized cholesterol.
Finally, anecdotes suggest that the brain needs evidence of paternal blessing to maintain vigorous and deeply satisfying heterosexual behavior. The father’s (or other adult male) facial skin surface lipid taken by mouth in sufficient quantity without sexual receptivity effectively strips lesbians of all homosexuality. Loving your father seems well worth the effort.
SEBUM IN ANIMALS AND US?
Scientists have watched the behavior of social animals and tracked gland masses. They have followed exchanges from animal to animal with dyes and radioactive tracers.107 They have exposed animals to pheromonal chemicals under controlled conditions, too (e.g. Roelofs, W.L., 1979108; Müller-Schwarze, D., 1969109). From such experiments, scientists have learned that sebum is a semiochemical in many species.110
As in mammals generally, the male of our species has more sebaceous glands than the female. Dimorphic differences in appetites for pheromones drive human sexual preference14 There is no ‘gay gene’ however, since homosexuality can be alleviated by oral intake of 150 mg to 250 mg of healthy adult male facial skin surface lipid p.o. Pheromone appetites determine fruit fly sexual preference.111 Indeed, it is easy to alter sexual preference from homosexual to heterosexual as this
EXOCRINOLOGY
37

author has done eleven times now, beginning in 1989. Providing 150-250 mg of adult male facial skin surface lipid by mouth forces the transition.
A "pecking order" may be pecking for pheromones. In social birds pecking sebaceous glands (in the cocks comb, rictus perimeter and preen gland at the base of the tail) is strictly hierarchical.112 Human behavior suggests the importance of human hierarchies. The wearing of judicial robes and seating behind high court benches may prejudice court decisions just as business suits and desk real estate widen interpersonal space for the safety of deciders.
In laboratory animals, specialized sebaceous gland size varies with social status within the local behavioral hierarchy. Top rats have heavy, active glands while low ranking rats (who "don't exist socially") have glands that are dried up and atrophied.70 Lorenz noted that low ranking rats climbed trees--a decidedly dangerous behavior for a rat--in order to escape the scent marks of higher ranking rats. Dangerous behavior and hen-pecked fins are typical of hatchery raised fish because of crowded unnatural conditions. Fish hatcheries presumably destroyed the salmon runs by releasing aversive warning pheromones from hatchlings that washed downstream warning prospective salmon parents of poor breeding conditions.
Who has access to whose fur defines troop sociality exclusively in other primate species. Sometimes only the top banana has access to all troop members' fur, sometimes the reverse.15
In humans, overactive sebaceous glands seem to cause the skin condition known as acne. Acne is a skin disease that has had it own effect upon hierarchical status in pubescent teens. It causes disfiguring eruptions of face and body pustules, diminishing status temporarily.
Human beings do not groom one another as apes do. We kiss instead. The repertoire of human kissing technique appears to exceed the range of primate grooming devices. Some primatologists will disagree. Doubtless some apes will, too. Furthermore kissing should play little part in work-related hierarchical struggles. So what governs work-related social interaction? Perhaps--and to a surprising extent--airborne pheromones do.
38

SEBUM CHEMICAL COMPOSITION AND ROMANTIC FAILURE
While the various chemical components of sebum are characteristically human, our sebums are still just as distinctive as our faces. Every adult human being has his own personal biochemical recipe because each human being carries a unique sebum. We share most of the same human-typical components, their relative abundances in the mix change from person to person. Such mixture variations serve to distinguish one person's sebum from another's.10
So your pheromones set you apart. Does your semiochemical chemical composition change with emotion? This may be plausible because different passions vary in their skin heats. Sebums of the passions would have varying components in their melts. If these expectations are met then sebum is "personal mood slime."
In humans sebum composition varies from place to place on our bodies as well.9 The sebum on your forehead has different components from that of your chin. Both of those differ from that inside of your mouth. And all differ from the sebum around your nipples (if you are a reproductive female).16 Does uptake of the female's breast sebum affect subsequent male behavior?
Pheromones explain much, perhaps even the whole story. Human beings retain the free will to love or not to love, until they decide to draw neigh. Then what follows may not be entirely in their hands, as all true lovers know.
EXOCRINOLOGY
39

Marriage, like a Trap set for flies, may possibly be ointed, at the Entrance, with a little Voluptuousness, under which is contained a draught of deadly wine, more
pricking and tedius than the Passions it pretends to cure, leaving the Patient in little quieter condition in the morning, than him that hath overnight kill'd a man to gratifie
his revenge.
Advice to a Son Page 7.II
II,. Love and Marriage 1683
40
1 Birt D, Main J Oral manifestations of herpes simplex virus infections. Laryngoscope 1977;87(6):872-878.
2 Eibl-Eibesfeldt, Irenäus. (1970a). Ethology the Biology of Behavior, translated by Erich Klinghammer New York: Holt, Rinehart and Winston 530 p.
3 Gould SJ, Eldredge N. Punctuated equilibria: the tempo and mode of evolution reconsidered. Paleobiology 1977;3:115-51.
4 Nicholson, B. 1984; Does kissing aid human bonding by semiochemical addiction? British Journal of Dermatology 111(5):623-627.
5 Schleidt, M. 1980; Personal odor and nonverbal communication. Ethology and Sociobiology 1:225.
6 Selway, R, and E.B. Keverne 1990; Hippocampal lesions are without effect on olfactory memory formation in the context of pregnancy block. Physiol. Behav. 47(2):249-252.
7 Kandel, Eric R, and James H. Schwartze 1985; Principles of Neural Science. New York: ElsevieR 979 pages. pp 21-22.
8 Montagna, William, and Paul F. Parakkal 1974; The Structure and Function of Skin. New York: Academic Press. 433 p.
9 Greene RS, Downing DT, Pochi PE, Strauss JS. Anatomical variation in the amount and composition of human skin surface lipid. Journal of Investigative Dermatology 1970;54(3):240-47.
10 Nicolaides, N. 1974; Skin lipids: their biochemical uniqueness. Science. 186:19-26.
11 Agache, P., D. Blanc, C. Barrand, and R Laurent R 1980; Sebum levels during the first year of life. British Journal of Dermatology. 103:643-649.
12 Montagna, William (1965). The skin. Scientific American 212:56.
13 Rothmann, Stephen (1959). The Human Integument. Washington, D.C.: American Association for the Advancement of Science, Publication No. 54

EXOCRINOLOGY
41
14 Nicholson B. Love and kisses: a semiochemical addiction model for human bonding. 1983. Paper presented to the International Society for Human Ethology and Animal Behaviour Society biannual international meeting at Bucknell University, Bucknell PA.
15 Wilson, Edward O. 1975; Sociobiology The New Synthesis. Cambridge, Mass.: Belknap Press of Harvard University Press. 697p.
16 Masters, W.H., and Virginia E. Johnson (1966). Human Sexual Response. Boston: Little, Brown and Company.
17 Seeley, T., T. Talbot, P.R Abramson, L.B. Perry, A.B. Rothblatt, and D.M. Seeley 1980; Thermographic measurement of sexual arousal: a methodological note. Archives of Sexual Behavior 9(2):77-86.
18 Cunliffe WJ, Burton JL, Shuster S. The effect of local temperature variations on the sebum excretion rate. British Journal of Dermatology 1970;83:650-4.
19 Pierard-Franchimont, C., G.E. Pierard and A.M. Kligman 1990; Seasonal modulation of sebum excretion. Dermatologica 181(1):21-2, 1990.
20 Burton, J L. (1970). The physical properties of sebum in acne vulgaris. Clinical Science 39:757-767.
21 Nicholson, B. 1985; An economic model of phermone transmission in avians. Paper presented to the Animal Behavior Society biannual meeting, Raliegh, N.C.
22 Blum M. Pheromonal bases of social manifestations in insects. In: Birch MC, editor Pheromones. Amsterdam: North-Holland Publishing Company, 1974.
23 Moulton, David G. 1977; Minimum odorant concentrations detectable by the dog and their implications for olfactory receptor sensitivity. pp. 455-464. In: Chemical Signals in Vertebrates Müller-Schwarze, D and Maxwell M. Mozell editors. New York: Plenum Press 610 p.
24 Albone, Eric S. 1990; Mammalian semiochemistry and its applications. pp 77-83. Chapter I.9 In: Chemical Signals in Vertebrates 5. MacDonald, D.W., D Müller-Schwarze, and Steven E. Natynczuk editors. Oxford: Oxford Science Press. 659 p.
25 Wilson, Edward O. (1963). Pheromones. Scientific American 208:100.
26 Finck HT. Romantic Love and Personal Beauty; Their Development, Causal Relations, Historic and Natural Peculiarities. London: MacMillan. (1891).
27 Harlow, Harry F. (1958). The nature of love. Amer Psychol. 13:673-685.
28 Murstein, B.I. (1971). Theories of Attraction and Love. New York: Springer Publishing Company, Incorporated.
29 Hold-Cavell, Barbara, and Margret Schleidt 1981; A Cross-Cultural Study on the Attitude towards Personal Odors. Journal of Chemical Ecolology 7(1):1
30 Albone, Eric S. 1984; Mammalian Semiochemistry The Investigation of Chemical Signals Between Mammals. Chichester, England: J Wiley & Sons Limited. 360 pages p. 13

42
31 Johnston, Robert E. (1983a). Chemical signals and reproductive behavior Chapter 1. pp. 3-37 In: Pheromones and Reproduction in Mammals. Bandenbergh, J G. editor. New York: Academic Press, Inc. 298 p.
32 Garcia-Velasco J, Mondragon M. The incidence of the vomeronasal organ in 1000 human subjects and its possible clinical significance. The Journal of Steroid Biochemistry and Molecular Biology. (Proceedings of the International Symposium on Recent Advances in Mammalian Pheromone Research.) 1991; 39(4B):561-563.
33 Monti-Bloch, L., and B.I. Grosser 1991; Effect of putative pheromones on the electrical activity of the human vomeronasal organ and olfactory epithelium. The Journal of Steroid Biochemistry and Molecular Biology. (Proceedings of the International Symposium on Recent Advances in Mammalian Pheromone Research.) 39(4B):573-582.
34 Morrison, Robert Thornton, and Robert Neilson Boyd 1987; Organic Chemistry Fifth Edition. Boston: Allyn and Bacon, Inc. 1486 p. (p. 347)
35 Drickamer LC. Puberty-influencing chemosignals in house mice: ecological and evolutionary considerations. In: Duvall D, Müller-Schwarze D, Silverstein RM. editors. Chemical Signals in Vertebrates 4. Plenum Press: New York. 1986;441-56.
36 Morris, D. (1967). The Naked Ape. New York: McGraw-Hill.
37 Eibl-Eibesfeldt, Irena us. (1970b). Love and Hate: On the Natural History of Behavior Patterns. New York: Methuen and Company.
38 Eibl-Eibesfeldt, Irenäus 1983; Patterns of parent-child interaction in cross-cultural perspective. In: The Behavior of Human Infants. Oliverio Albert and Michele Zappella editors. New York: Plenum Publishing Corporation pp. 177-217.
39 Verheggen FJ, Haubruge E, Mescher MC. Alarm pheromones-chemical signaling in response to danger. Vitam Horm. 2010;83:215-39.
40 Chivers DP, Wisenden BD, Hindman CJ, Michalak TA, Kusch RC, Kaminskyj SG, Jack KL, Ferrari MC, Pollock RJ, Halbgewachs CF, Pollock MS, Alemadi S, James CT, Savaloja RK, Goater CP, Corwin A, Mirza RS, Kiesecker JM, Brown GE, Adrian JC Jr, Krone PH, Blaustein AR, Mathis A.Epidermal 'alarm substance' cells of fishes maintained by non-alarm functions: possible defense against pathogens, parasites and UVB radiation. Proc Biol Sci. 2007 Oct 22;274(1625):2611-9.
41 Muscarella F, Fink B, Grammer K, Kirk-Smith M. Homosexual orientation in males: evolutionary and ethological aspects. Neuroendocrinology Letters 2001;22:393–400.
42 Kohl JV, Atzmueller M, Fink B, Grammer K. Human pheromones: integrating neuroendocrinology and ethology. Neuroendocrinology Letters 2001;22:309–321.
43 Preti G, Cutler WB, Christensen CM, Lawley H, Huggins GR, Garcia C-R. Human axillary extracts: Analysis of compounds from samples which influence menstrual timing. Journal of Chemical Ecology 13(4): 717-731.
44 McClintock MK, Menstrual synchrony and suppression. Nature 1971;229:244-245.

EXOCRINOLOGY
43
45 Cutler WB, Preti G, Krieger A, Huggins GR, Garcia CR, Lawley HJ. Human axillary secretions influence women's menstrual cycles: the role of donor extract from men. Hormones and Behavior 1986;20:463-73.
46 Cocke R, Thiessen D, Nicholson B. Alarm chemosignals suppress the immune system. In: MacDonald DW, Müller-Schwarze D, Natynczuk SE, editors. Chemical signals in vertebrates 5. Oxford: Oxford University Press; 1990. p.125-31.
47 Ackerl K, Atzmueller M, Grammer K. The scent of fear. Neuroendocrinology Letters 2002; 23:79-84.
48 Arakawa H, Arakawa K, Blandino P Jr, Deak T. The role of neuroinflammation in the release of aversive odor cues from footshock-stressed rats: Implications for the neural mechanism of alarm pheromone. Psychoneuroendocrinology. 2011 May;36(4):557-568.
49 Rothman, Stephen (1954). Physiology and Biochemistry of the Skin, Chicago: University of Chicago Press Chapter 12 Sebaceous-Gland Excretion p. 285.
50 Udry, JR, and Naomi M. Morris (1968). Distribution of coitus in the menstrual cycle. Nature 220:563.
51 Wheatley, Victor R (1956). Sebum: Its chemistry and biochemistry. Am. Perfumer 68:37-47.
52 Nikkari, Tapio 1974; Comparative chemistry of sebum. Journal of Invesigative Dermatology 62(3):257-267.
53 Inscoe, M. 1977; Chemical communication in insects. J Wash. Acad. 67:16-33.
54 Collins, C.W., and S.F. Potts (1932). Attractants for flying gypsy moths as an aid in locating new infestations. U.S. Dept. Agric. Tech. Bull. 336:1-43.
55 Butenandt A, Beckmann R, Stamm D, Hecker E. Über den Sexual-Lockstoff des Seidenspinners Bombyx mori. Reindarstellung und Konstitution. Z Naturforsch 1959;14:283-4.
56 Tumlinson, James H., and P.E.A. Teal 1987; Pheromone Biosynthesis and its regulation. pp 3-26. Chapter 1. In: Pheromone Biochemistry. Prestwich, Glenn D., and Gary J. Blomquist editors. Orlando: Academic Press, Inc. 565 p.
57 Albone, Eric S., Pauline E. Gosden, and Georges C. Ware 1977; Bacteria as a source of chemical signals in mammals. pp. 35-43 In: Chemical Signals in Vertebrates (Müller-Schwarze, D, and Maxwell M. Mozell eds.) New York: Plenum Press 610 p.
58 Gorman, ML, Stone RD. Mutual avoidance by European moles Talpa europaea. In: MacDonald DW, Müller-Schwarze D, Natynczuk SE, editors. Chemical Signals in Vertebrates 5. Oxford: Oxford Science Press. 1990; 367-77.
59 Marchlewska-Koj, Anna 1983; Priming pheromones in mice. pp. 135-151. In: Chemical Signals in Vertebrates 3. (Müller-Schwarze, D, and Robert M. Silverstein eds.) New York: Plenum Press. 368 p.
60 Epple G. Relationships between aggression, scent marking, and gonadal state in a primate, the tamarin saguinus fuscicollis. In Chemical Signals Vertebrates and Aquatic Invertebrates. Mu ller-Schwarze D. and RM. Silverstein editors. New York: Plenum Press. 1980.

44
61 Paris, Paul (1913). Recherches sur la glande urophgienne des oiseaux. Archives de Zoologie Experimentale et Ge'ne'rale. 53:139-276.
62 Hou, H.C. (1928). Studies on the glandula uropygialis of birds. Chinese Journal of Physiology. 2:345-380.
63 Idris, M.D., and I. Prakash 1988; Perception of dominance status through conspecific scent in the desert gerbil Meriones hurrianae. Indian Journal of Experimental Biology 26(6):479-480.
64 Soni, G.R and I. Prakash 1988; Mitigation of poison shyness in desert gerbil by adding conspecific sebum of ventral scent marking gland and urine in poison baits. Indian Journal of Experimental Biology 26(6):476-478.
65 Albone, Eric S. 1990; Mammalian semiochemistry and its applications. pp 77-83. Chapter I.9 In: Chemical Signals in Vertebrates 5. MacDonald, D.W., D Müller-Schwarze, and Steven E. Natynczuk editors. Oxford: Oxford Science Press. 659 p.
66 Sugiyama, T., and H. Sasada 1988; Chemistry and biochemistry of unusual branched fatty acids with pheromonal activity from mature male goat. Seikagaku - Journal of Japanese Biochemical Society. 60(2):118-122.
67 Doi S, Tanabe K, Yoshimura M. Induction of sexual agglutinability by unsaturated fatty acids in Saccharomyces cervisiae. FEMS Microbiol Lett 1990;55(1-2):89-91.
68 Weldon, Paul J 1983; The evolution of alarm pheromones. pp. 309-312 In: Chemical Signals in Vertebrates 3. Müller-Schwarze, D and Robert M. Silverstein editors. New York: Plenum Press 368 p.
69 Nicholson, B. 1988; Pheromones in the pathogenesis of autoimmune and behavioral illness. Paper presented informally to the International Society of Chemical Ecology, Hull, England.
70 Moulton, David G. 1977; Minimum odorant concentrations detectable by the dog and their implications for olfactory receptor sensitivity. pp. 455-464. In: Chemical Signals in Vertebrates Müller-Schwarze, D and Maxwell M. Mozell editors. New York: Plenum Press 610 p.
71 Mykytowycz, R 1973; Reproduction of mammals in relation to environmental odours. Journal of Reproductive Fertility. [Suppliment] 19:431-444.
72 Bell M. A comparative study of the ultrastructure of the sebaceous glands of man and other primates. Journal of Investigative Dermatology 1974;62(3):132-43.
73 Brown JC, Williams JD. The rodent preputial gland. Mammal Review 1972;2(4):105-51.
74 Albone, Eric S. 1984; Mammalian Semiochemistry The Investigation of Chemical Signals Between Mammals. Chichester, England: J Wiley & Sons Limited. 360 pages p. 13
75 Sansone, Gail, and RM. Reisner (1971). Differential rates of conversion of testosterone to dihydrotestosterone in acne and in normal human skin--a possible pathogenic factor in acne. Journal of Investigative Dermatology 56:366-372.
76 Benfenati A, Brillanti F. Sebaceous glands distribution in skin of human body Arch. Italdi. Dermat., sif. 1939;15:33-42.

EXOCRINOLOGY
45
77 Masters, W.H., and Virginia E. Johnson (1966). Human Sexual Response. Boston: Little, Brown and Company.
78 Kligman, A.M. (1963). The uses of sebum? pp. 110-124. In: Advances in Biology of Skin. Vol. 4. the Sebaceous Glands (ed. by Montagna, W., RA. Ellis, and A. Silver) Oxford: Pergamon Press.
79 Seeley, T., T. Talbot, P.R Abramson, L.B. Perry, A.B. Rothblatt, and D.M. Seeley 1980; Thermographic measurement of sexual arousal: a methodological note. Archives of Sexual Behavior 9(2):77-86.
80 Hafez, E.S.E. 1976; Parameters of sexual maturity in man. pp. 105-122 In: Volume 3 Sexual Maturity: Physiological and clinical Parameters E.S.E. Hafez and JJ Peluso editors. Ann Arbor, Michigan: Ann Arbor Science Publishers Inc. 283p.
81 Sorensen, P.W., T.J Hara, N.E. Stacey and F.W. Goetz 1988; F prostaglandins function as potent olfactory stimulants that comprise the postovulatory female sex pheromone in goldfish. Biol Reprod 39(5):1039-1050.
82 Bruce HM. An exteroceptive block to pregnancy in the mouse. Nature 1959;184:105.
83 Bruce HM. A block to pregnancy in the mouse caused by proximity to strange males. Journal of Reproductive Fertility 1960;1:96-103.
84 Drickamer, L.C. 1986. Puberty-influencing chemosignals in mice: ecological and evolutionary considerations. In, D. Duvall, D. Müller-Schwarze and R.M. Silverstein (eds.), Chemical Signals in Vertebrates, Vol. 4. New York: Plenum Publishing. Pp. 441-455.
85 Eibl-Eibesfeldt, Irenäus 1975; Ethology: The Biology of Behavior, second edition. New York: Holt, Rinehart and Winston.
86 Wilson, Edward O. and William H. Bossert (1971). A Primer of Population Biology. Sunderland, Massachusetts: Sinauer Associates, Inc. Publishers 192 p.
87 Lorenz, T.H., D.T. Graham, and S. Wolf (1953). The relation of life stress and emotions to human sebum secretion and to the mechanism of acne vulgaris. Journal of Laboratory and Clinical Medicine 41:11.
88 Mennella, Julie A, and Howard Moltz 1989; Pheromonal emission by pregnant rats protects against infanticide by nulliparous conspecifics. Physiology and Behavior 46:591- 595.
89 Kikusui T, Takigami S, Takeuchi Y, Mori Y. 2001; Alarm pheromone enhances stress-induced hyperthermia in rats. Physiol Behav. Jan;72(1-2):45-50.
90 Mehler MF, Purpura DP. Autism, fever, epigenetics and the locus coeruleus. Brain Res Rev. 2009 Mar;59(2):388-92. Epub 2008 Nov 24.
91 Downing DT, Stewart ME, Strauss JS. Estimation of sebum production rates in man by measurement of the squalene content of skin biopsies. Journal of Investigative Dermatology 1981; 77(4):358-60.
92 Pochi, Peter E. 1982; Commentary: androgen effects on human sebaceous glands. Archives of Dermatology 118(10):803.

46
93 Strauss, JS., D.T. Downing and F.J Ebling 1983; Sebaceous glands. In: Biochemistry and Physiology of the Skin. Goldsmith L.A. editor. New York: Oxford University Press.
94 Hou, H.C. (1929). Relation of the preen gland (glandula uropygialis) of birds to rickets. Chinese Journal of Physiology. 3:171-182.
95 Frantz RA, Kinney CK. Variables associated with skin dryness in the elderly. Nursing Research 1986;35(2):98-100.
96 Cove JH, Holland KT, Cunliffe WJ. An analysis of sebum excretion rate, bacterial population and the production rate of free fatty acids on human skin. Br J Dermatol 1980;103:383.
97 Kuwahara, Y., W.S. Leal, T. Suzuki, M. Maeda and T. Masutani 1989; Antifungal activity of Caloglyphus polyphyllae sex pheromone and other mite exudates. Pheromone study on astigmatid mites, XXIV. Naturwissenschaften 76(12):578-579.
98 Guthrie, RD. (1970). Evolution of human threat display organs. In: Evolutionary Biology, vol. 4. Dobzhansky, T., M.K. Hecht, and W.C. Steere editors. New York: Appleton-Century-Crofts.
99 Burton JL, Shuster S, Cartlidge M. The sebotrophic effect of pregnancy. Acta Dermato-vernereologica (Stockholm). 1975; 55:11-3.
100 Burton, JL., Sam Shuster, M. Cartlidge, L.J Libman, and U. Martell (1973a). Lactation, sebum excretion, and melanocyte stimulating hormone. Nature 243:349-350.
101 Downing DT, Strauss JS. Synthesis and composition of surface lipids of human skin. Journal of Investigative Dermatology 1974;62(3):228-44.
102 Wysocki, S.J, A. Grauaug, G. O'Neill, and R Hähnel 1981; Lipids in forehead vernix from newborn infants. Biol. Neonate. 39:300-304.
103 Silverstein, Robert M. 1984; Chemistry of insect communication. Chapter 5, pp 105-121. In: Insect Communication. Lewis, Trevor editor. Academic Press, London. 414 p.
104 Sansone-Bazzano, Gail, B. Cummings, A.K. Seeler, and RM. Reisner 1979; Differences in the lipid constituents of sebum from pre-pubertal and pubertal subjects. British Journal of Dermatology 103:131.
105 Wysocki, Charles J Chuck 1983; You must remember this, a kiss isn't just a kiss. Paper presented to the Animal Behavior Society and the International Society for Human Ethology, Lewisburg, PA.
106 Beauchamp GK, Yamazaki K, Boyse EA. The chemosensory recognition of genetic individuality. Scientific American 1985;253(1):86-92.
107 Adachi, K., 1984 personal communication.
108 Roelofs, W.L. 1979; Pheromone perception in Lepidoptera. In: Neurotoxicology of Insecticides and Pheromones. New York: Plenum Press.
109 Müller-Schwarze, Dietland (1969). Complexity and relative specificity in a mammalian pheromone. Nature (Lond.) 23:525-526.

EXOCRINOLOGY
47
110 Shorey, H.H. 1976; Animal Communication by Pheromones. New York: Academic Press.
111 Grosjean Yael, Grillet Micheline, Hrvoje Augustin, Ferveur Jean-François Ferveur & David E Featherstone A glial amino-acid transporter controls synapse strength and courtship in Drosophila Nature Neuroscience 11, 54-61 (2008).
112 Lucas, A.M. 1980; Lipoid secretion of the body epidermis in avian skin. In: The Skin of Vertebrates. Spearman RI.C. and P.A. Riley. editors. New York: Academic Press.


Chapter 2: Parental Love of Offspring Before describing the love of adults for their children, the concept of children itself should be examined. (These petty biological controversies bore everyone, but some contemporary readers expect this.) What are children for? Loving young immature forms and caring for them into their reproductive adulthood denominates a life strategy. Besides mammals, some species of fish, reptile, insect, and amphibian also practice rudimentary forms of it. The predominant reproductive strategy of rearing young suits the economic constraint set of human beings better than any other alternative.
What does parent-infant bonding accomplish for the human species? Any argument should hold for all other bonding species. This means we should find similar emission chemistry, pheromone emissions, pheromone receptors, gland physiology, as well as similar parental and offspring behaviors. Other immature mammals require nurture but human neonates require more. Human immaturity extends longer than any other animal species1 requiring the greatest nurturing of all. That nurture depends upon parental love. Therefore assuming an altricial childhood strategy must be less effective without bonding, nurtured, pheromone-triggered, life strategies should also develop in childhood.
Lengthy immaturity is not the only drawn-out characteristic of human ontogeny. Humans have one of the longest gestation periods (not the longest because this is influenced by body size,2 among the longest courtship periods, and the most frequent intercourse (continual inseminations throughout adulthood).3 These must economically contribute to human flexibility as a species. Presumably, a longer time for relationships enables "fine tuning." Adjustments can be made to new information. Long-loved children can be molded gradually to best fit a changing world, but so can un-loved children as we shall see.
Actually, childhood (or immaturity) may be dispensed with altogether. Many animals give birth to little copies of themselves. Many more species lay eggs which
EXOCRINOLOGY
49

hatch miniature adults. Species with the miniature adult (precocial) strategy usually abandon their offspring or tend rudimentary nests. For instance, many a reptile mom lays her eggs to fend for themselves. Others find metamorphosing larval stages more appropriate, which parents ignore completely. Human beings and other species, which love and care for their offspring give birth to feeble immature forms. Indeed, our babies have been described as the most immature form of all. Even though marsupials bear less viable young, no offspring take so long to rear as human children. We must bear this question in mind: why?
DOES CUTENESS WORK?
It has to work, to do something. The shape of the head and face mark an obvious distinction between abandoned and loved offspring. Abandoning species leave behind young with small but "adult-shaped" heads. Loving species have smashed-in faces. (And of course, larvae rarely resemble adults).
Innate mental preferences bear upon parent-infant bonding just as upon mate-mate bonding. Men just naturally like curves. Women admire phalluses. The German ethologist, Konrad Lorenz (1943) suggested that chubby cheeks and a large forehead associated with infancy have universal appeal among human beings and inspire parental care. Many cartoon characters supposedly exploit this innate fondness as can be seen in the accompanying illustration.
50

Figure 277. "Baby schema" of man. Left: head proportions that are generally considered to be "cute"; right: adult forms, which do not activate the drive to care for the young (brood care). (From K. Lorenz [1943].)
Notice that the foregoing illustrations are "cute." Where do we find "cuteness?" In mammals and avians that nurture their young, infants and juveniles have pushed-in snouts when compared to adults. Remember reptile offspring carry adult-shaped heads when first entering the world. Pushed-in snouts and the accouterments of parental care (nests) are seen in some fossilized dinosaur remains.
Dinosaur species that apparently cared for their young may have found their offspring cute, too. Given the probable intelligence of those beasts, such an untestable explanation may be a bit too facile.
We can educe the following. Pushed-in immature faces evolved earlier. Human recognition of "cuteness" came later. Dinosaurs preceded us here, after all. Keep that in mind.
Consider the pain of human childbirth. Alone among the females of the world, human females must distort their pelvic skeletons to give birth. Would that pain and death risk be entirely compensated by "cuter" yet feeble, big-headed babies? The baby’s brain develops precociously, of course, but these distortions of gestation
EXOCRINOLOGY
51

keep more of the baby’s brain keyed into the mother’s perceptions longer allowing maternal chemosensory insights that would be otherwise unavailable to the neonate.
Physiologically adult-shaped faces vary from the pushed-in snouts of infantile forms because the skull grows to include the sinuses. Our sinuses are hollow organs carved during infancy, childhood, and puberty into the growing bones around nasal passages. When we are born, we are born with no sinuses. A lack of sinuses coincides with "cute" infantile features (see Netter, F.H. 1989)4. Do sinuses form just to reduce cuteness? With what we are bearing in mind, this is not likely. Perhaps cute heads get through narrower-the-better-for-running birth canals more easily? Only the dinosaurs would know for sure.
Sinuses have no known function.5 However, sinuses may function passively in pheromone recognition, perhaps as semiochemical storage or separation vessels. Chemists call similar vessels condensers or retorts. When most modern reptiles hatch, they have their full complement of sinuses in their miniature, adult-shaped heads. Apparently no bio-economic need for building sinuses arises during reptile immaturity. Reptile young do not depend precariously upon their mothers as mammals do. Young mammals and birds confined to the nest or burrow may not require keen pheromone discrimination. Only adults maintain extended territories. Such territories might require sinuses in perception of faint trail marking pheromone the animals use to exploit their territories efficiently. Keeping ‘just the right distance’ from an adjacent conspecific (or auto-emitted) pheromone trails allows economical resource harvesting, with wild animals moving through their personal territories methodically just like we harvest a crop.6
As we will see in Chapter 6, lacrimation or crying, carries pheromone receptor proteins in tears which inhabit the sinuses. In human beings, sinuses develop in phases. Sinuses may be involved, somehow, with pheromone recognition, perception, or maintenance. Economically, if pheromone recognition by the mother requires a set of sinuses at a certain age, then that set should develop just before the need. The development of a sinus set in puberty before mate-mate bonding might be expected, too. Sequences of sinus development does proceed in such phases. They do reach final completion at reproductive maturity. Too bad for this idea, we have no
52

indications of especial activity in parental sinuses in the period leading up to birth, or immediately after marriage. We do have lacrimation.
New pheromone receptor proteins--the feto proteins--develop during pregnancy and may be transmitted sexually or by proximity in maternal saliva, skin lipids, lochia and/or colostrum. And if those sources are insufficient, the neonate is crawling with fetoproteins in his or her blood. Alpha fetoprotein is essentially albumin which acts exactly like a pheromone binding protein under antennagraphic analysis. Since the injection of maternal blood into virgin female rats stimulates maternal behavior in them, this suggests that some blood plasma borne factor stimulates maternal bonding.7 Much more will be said about them in Chapter 6.
Sinus growth (and the "adult typical" head shape) should behave economically and await its need according to the economic model of evolution. "Cute" heads are characterized by relatively large head sizes, small bodies, juvenile furs/plumage (soft skin in the human case), helplessness, and perhaps infant calls (tearless crying in the human case). These and other traits release parental responses alone. Furthermore releaser recognition of infantile traits varies with species. A single spot of color in the open mouths of young or on the beaks of adults is sufficient stimulus to release feeding behavior in many birds, for instance.8
A parental preference might change easier than baby head circumference. Appearance usually subserves function. Which came first here, the chicken or the egg? Do we like cuteness, soft skin, baby noises and so on just because of its physically constrained functional utility? It is a likely prospect as it seems to be the simplest explanation for now.
DO PHEROMONES AFFECT PARENT TO INFANT BONDING?
The pheromone hypothesis suggests that humans bond to their neonates (newborns) in a way corresponding to mate-mate bonding. Gestation seems to accentuate semiochemical receptivity in both parents, but especially so in the mother. As birth nears, the economic theory predicts that the new arrival will have prepared
EXOCRINOLOGY
53

well to make a big splash on birth day. For the last five and one half months inside the womb, baby has been secreting and stockpiling sebum within the placenta.9 When the newborn appears, he wears that unctuous stuff, sebum, from head to toe, and it goes everywhere as anyone who has delivered one knows. What a slippery mess!
Elevated rates of new sebum secretion from the newborn baby crest the fourth day after birth and then gradually abate.10
[T]here is a marked postnatal sebum secretion. Our findings revealed that the levels obtained on the 4th day of life were comparable to those of young adults. This is an especially interesting fact which should be emphasized.
Bolstered by the mother's critical period receptivity, does addiction to baby's pheromone "birthday suit" and to newly secreted sebum precipitate parental love?
A pheromone model of parental love will help answer many questions. Parent-infant bonding: what are characteristics of this phenomenon? Bonding has remarkably low risk of failure. This seems reasonable since a baby may quickly starve or die of exposure without love. Love must work reliably and effectively; it must not fail.
This author watched bonding failure result in the death of a neonate when he was in medical school. The ‘too small to be viable’ infant, vigorously crying his lungs out, lay on a Mayo table for seven and one half hours, just twenty feet from his mortified mother. My objections were heard and acted upon as I did not receive academic credit that term. So if you are pregnant and of African descent, chose your doctor wisely because your baby’s life may depend upon your choice.
54

There may be different types of parental love; each must have an accounting. There must be adequate identification of the loved offspring to reduce mimicry. It must not err.
"Human beings can discriminate between their own offspring and unfamiliar children by means of olfactory cues and are even able to discriminate between one of their own children and their other child using olfactory cues (Porter and Moore, 1981). Human odors vary with diet (Wallace, 1977), a factor that may allow familial identification, given a common family diet. Identical twins are more difficult to discriminate between by their odor than unrelated adults or siblings that are not twins (Kalmus, 1955; Wallace, 1977), and the olfactory discriminability of the identical twins can be magnified if they have very different diets (Wallace, 1977)" (Leon, M., 1983).11
Recognizable individual odor differences and/or sebum differences among children may be the encouragement for a parent to "miss" an absent child. If own-child odors were monolithic or parental in origin, one son or daughter would be as good as any other. A woman with thirty children may grieve unconsolably at a single loss.
Watch human parental behavior toward returning offspring at an airport. See the kissing and hugging? Hugging may trigger a non-kiss pheromone recognition mechanism. Indeed, warm partner contact (hugging & kissing) influences peripheral circulating proteins with chemosensory functions, beta2-microglobulin (11.7 k Da), serum albumin (66-k Da), and an unidentified 5.9 k Da serum protein.12
Also seen in resumption of mate-mate bonding, these affectionate airport behavior may be pheromone replenishment behaviors. Interestingly enough, something similar can be observed in birds:
EXOCRINOLOGY
55

Besides being associated with bodily contact and sociality, allopreening is most frequent in species that are sexually dimorphic, form persistent pair bonds, or both; and it reaches its highest intensity within species when birds are first brought together or reunited after a prolonged absence.13
Adjustments, changes, or outright cessation of the parent-infant bond happen as young grow up. Can the pheromone hypothesis explain all of these characteristics? An acceptable semiochemical explanation must do just that.
LOW FAILURE RISK
An axiom of good design holds that providing backup systems enhances system reliability. If one breaks down, the backup functions in its place. A mechanical device for lowering airplane landing gear backs up hydraulic systems. A gas generator backs up the electrical power grid at the hospital. Making such systems dissimilar reduces risk further, an axiom of portfolio theory. More than one such reserve device reduces risk further, but against diminishing returns on investment. The more reliable the system per total expenditure, the more successful that system should be, all else equal. The same logical process holds true working in reverse. For instance, extremely ‘expensive’ biochemistry indicates tissue and emission importance.
There appears to be at least two devices which separately prime and stimulate bonding in humans, one spiritual, the other kissing. Spiritual love may be signaled by the spirits, or off-gases, of the female's vagina during delivery. If the physiological basis for kissing involves sebum, a skin secretion, then parent-infant kissing would have its own special features. The feto proteins that mark phases of pregnancy are pheromone receptor proteins just like their cousin molecule albumin, more about them later.
56

DIFFERENTIAL PARENTAL SOLICITUDE
Loving parents choose favorites in all animal families (Daly, M. and M. Wilson, 1980). They love some young more than others. Could there be a biological basis for this differential parental solicitude? If so then differences in love suggest we should see differences in the signals stimulating that love.
Do love signals differ? One would expect that there would be recognizable differences in bonding pheromones from species to species, between kin and non-kin, and among individual family members. Odors allow parental discrimination of young in humans and other species.10 All such individual differences exist in human and other mammalian sebums. (For a fuller review of the uniqueness of our exocrine secretions, please see Chapter 3 on pheromone chemistry).
A physical imprinting upon pheromone receptors by individually unique pheromones would accomplish this purpose. Our so-called immune system, our reproductive system, our endocrine system, our nervous systems may help fill this role as they all evolved from pheromone and allomone recognition systems clearly observable in lowly colonial bacteria.
A moistness of the eyes seems to help imprint the pheromones of children. Indeed, those families that bond to a survivor most successfully grieve for critically ill infants born prematurely. A dearth of grief by parents marks families leaning toward abuse of their recovered premie.14 Such observations appear to support the pheromone reception bonding hypothesis.15,16
JUST IN TIME LOVE
Bonding speed varies from species to species. The time the neonate can survive without care ultimately constrains. In human beings, a baby might survive alone only a few hours or less time depending upon environment. Without love, the
EXOCRINOLOGY
57

patience of an un-bonded caregiver might be gone in only a few hours or days depending upon food abundance, predation risk, crowding and other factors. The neonate must use that precious time. A baby must succeed and influence, if not control, parental behavior.
Bonding time being so often short and fraught with dangers, we would expect no nonsense about it. Lurking sabre-tooth tigers saw to that. Evolution pressures against inefficiency. Any trait causing pre-reproductive death would get no second chance. Likewise any trait tending to hurt early survival must contribute more to reproductive success later in ontogeny. Just how much more, may be a product of the prevailing internal rate of return and actuarial probability of survival. Compressing bonding time would require evolving more efficient behavior.
One device common to most mammalian parent-infant bonding is "neonatal grooming." Is it an oral transfer of pheromone from the skin of the neonate to the mouth of the mother? How to test this? Preventing neonatal grooming and separating parent from infant until the pheromone on the neonate's body has evaporated should stop bonding. So rejection of offspring should result from shampooing the newborn before the mother can nuzzle the infant, perhaps even within the sight of the mother. Withholding all neonatal exocrine emissions from the parent might maintain pheromone receptivity longer than normally necessary, too.
In some ungulate species, each female isolates herself from the protection of the herd to bear her young. Because she puts herself outside the safety of the group, the predators have a field day. Indeed, predator reproduction is often keyed to such feast days.
To minimize predation, some seasonal signal makes mothers calf simultaneously. Mother-infant bonding generally proceeds as rapidly as possible. Simultaneous, quick births mean that the lions gorge themselves once only. Because lions take a while to get hungry again, most of the new babies escape. Consider the situation and its bonding behavior: The defenseless cow is completely exhausted. The newborn can not stand and predators lurk all about. (As expected, predators come from miles around for the occasion.) Is that the time for the mother to spend her
58

scarce energy and time to "eagerly clean" her newborn? Obviously it is not. Is this then the time for crossed signals to foul up and cause nonsensical ‘displacement activity’? Get real.
Indeed, with predators literally watching and by all accounts, the herbivorous mother cow eats the amniotic substance "ravenously." The intense appetite for the neonatal goo comes whether or not she is hungry. Well fed mothers eat it up just like emaciated mothers. The substance has little nutritional value. Only about 30% of the sebaceous fraction can conceivably be used for food at all. Some of the rest may even be poisonous if it were taken in dietary doses. Plus the new ungulate mom is generally satisfied with just the skin/fur birth coverings of her own young.
An old expression has it that "there is no accounting for taste." There actually is an accounting for taste. If provided a minimally prepared smorgasbord of healthy foods, infants will choose a nutritious diet.17 Similarly, oddball food preferences of pregnant humans have an economic metabolic basis.18,19 Admittedly, reducing lion-attracting aromas may have something to do with it, but then why emit them? Furthermore, species which do not isolate themselves out of the herd, such as sheep, are just as greedy for afterbirth’s vernix.20
Given economic assumptions, we proceed teleologically. What purpose does isolation serve at time of delivery? The cows might isolate themselves because their birth pheromones are indiscriminate to avoid costs or to encourage lion populations with bigger, slower bellies. The self-imposed isolation would avoid accidental adoptions and paternity/maternity disputes. Remember, isolation from herd pheromones occurs during a particularly chemosensitive time.
Following the suggestion of this author, Levy and Poindron (1987) found that something in ewe amniotic fluid seems to encourage bonding. Amniotic fluid has attracted some scientific attention (Montagna, W., 1964; Nicolaides, N., 1974). We will examine it more closely shortly. This author has observed plant-eating mule deer does eating fawn afterbirth and exhibiting the pheromone-only Flehmen response observed in horses and in moose.
EXOCRINOLOGY
59

Meanwhile, what other characteristics should a love manufacturing process have? Love should be cheap. It should be possible to shut it off for those species opting for a strategy of seasonal parenthood. Finally, it should accommodate transition from generation to generation.
CHEAP LOVE
Pheromones cost little to produce and recognize; even bacteria can detect their own pheromones. Pheromone effects may last a long time, too. Shorey21 and others22 recount how pheromone exposure strictly determines adult behavior. If left alone, human love spans a lifetime. (Divorce, a contemporary and temporary scourge, may be remedied by medicinal use of human pheromones. See Chapter 8.) This seems consistent with the pheromone hypothesis of bonding.
However, seasonal,23 even daily24 changes in human skin surface lipid expression have unstudied effects on bonding. Humans may even have a "breeding season" in the fall. The autumn temperature decrease affects exocrine emissions and may signal the peculiar increased ejaculations observed.25
Birds shutdown their sebaceous uropygial gland secretions, along with their reproductive organs and even their pheromone reception systems after nesting season. Their sebaceous glands atrophy, often completely, and chemical simplification of the pheromone takes place.26 Humans do not mate intermittently. The small autumnal changes seen in human male apocrine secretions (Preti, G., et al., 1987) remain enigmatic. Does cool air or a shorter day bring on Preti's secretion to induce sexiness?
Addiction sustains a potent human chemical appetite. What more logical appetite would gratify parents enough to compensate them for caring for young? 10 From economic analysis, the mechanism of love seems so basic, so foolproof, with backups for backups, that its mechanisms should be straightforward to elucidate. Perhaps the offspring's recognition pheromone itself stimulates a reward for addictive craving? The physiology of the mammalian appetite and reward system, the limbic system, attracts a particular interest. Mice will brave hunger, thirst, even electric
60

shock in exchange for hypothalamic stimulation. The hypothalamus is reached by pheromone reception neuro-pathways.
Theoretical games people, notably Trivers,27 have thought that describing parental payoffs in terms of investment for the "reward" of having offspring survive to adulthood is sufficient explanation. Offspring pheromones, and not offspring themselves, may drive this device. That idea would explain its failures. The evolved economic necessity would, however, drive the pheromones driving the behavior. Do you see?
Analogous appetites are hunger, thirst, breath, sexual desire, and elimination. Take food as an example. Food seeking behavior generally improves an organism's survivability. Any stimulus to food seeking behavior, particularly one that may be regulated in tandem with other appetites would improve survivability (closely approximating optimum division of behavioral resources among competing appetites). Evolving appetites for essential behavior slaked for rewarding essential accomplishment may be basic to life. Perhaps crafting both the appetite and the pleasure of the reward erbkoordinates love.
The most efficient recognition devices are chemosensory. Short, trigeminal recognition of dangerous natural vapors, carbon dioxide and ammonia, consists of exposed nerve endings in mucous. Appetite efficiency should be expected as well. Other mammals besides humans have addictive appetites for chemicals which vary from species to species.28 Keeping error rates low requires semiochemical species-specificity. As there are many times more chemicals possible than species, sex and individual sub-specificity are possible and would be useful. These advantages proceed from dermal biosyntheses of the unique stereochemistries of pheromones. The uniqueness is critical only within the ecosystem experienced by the emitting organism. Other non-chemical checks should discourage predator lure secretions, but these are seen. Human hunters use this tactic frequently. Think about this.
Reproductive conditions affect the reliability of the pheromone addiction process. Ungulates avoid bearing young in the herd where paternity/maternity might be confused. That confusion possibility indicates a valued pheromonal recognition.
EXOCRINOLOGY
61

Pheromone must move from infant to mother. Weather conditions may influence bonding success by influencing pheromone transfer success. Accordingly, birth times may so correlate with low winds or lack of precipitation (seeking shelter).
A competing addiction to drugs reduces bonding success and may prevent it altogether. Data has strongly suggested that neoparous women taking drugs, some of which are addictive, bond less effectively to their infants than neoparous women taking fewer drugs or no drugs.29 Drug addicted men and women are notoriously poor parents, with much higher recorded rates of abuse and neglect.30 Thus the pheromone must itself have a drug-like effect, or so the logic goes. One pheromone can counteract and overwhelm another, such as in the case of the puberty acceleration and puberty delay pheromones of mice which work in opposition:
At rations of 1020 parts (puberty) acceleratory chemosignal to 1 part (puberty) delay chemosignal the acceleratory effect overrides the delay chemosignal, and the mice attain first estrus at earlier ages than controls. Ratios of about 4 to 1 up to 7 to 1 result in mean ages for puberty that are not accelerated or delayed relative to controls.31
Yin and Yang? The opposition of pheromone against pheromone may even reify these anecdotal oriental hypotheses. An addictive effect would be counteracted and overwhelmed only by another drug, explaining the neglectful misbehavior of drug addicted mothers. Contemplate this.
BIRTHDAY LOVE
Humans are not pretty at birth. We are gray-blue, slithery little things, dripping with a combination of amniotic fluid, mucous, blood, and vernix caseosa. This quarter inch of coating does little to enhance appearance. To believe that these "decorate" the newborn is unreasonable. Just what is this thick greasy layer, the vernix caseosa? It does not lubricate exit from the womb, the maternal mucous does that. Recent German research shows a greasy skin enhances warmth retention for long distance swimmers. Since human beings swim innately if born in the water,
62

vernix may have evolved from a prior aquatic adaptation.32 Given neonatal metabolism, high surface to volume ratio, and exposure, it should conserve warmth somewhat by reducing evaporative water loss. It is, however, completely inadequate by itself.
When a mother gives birth to a child, he/she comes into the world wearing a slippery chemical coating with the consistency of vegetable shortening. If the hypothesis proves correct, then when primed for it, that greasy coating contains a combination of what must be one of the most addictive mixtures extant. Obstetricians call the observed mix vernix caseosa. More Latin, this time for cheesy varnish.
The slippery, greasy vernix is mostly sebaceous. It is produced by the fetal sebaceous glands. These same glands are the first to function in the newly forming human being. Secreting sebum well ahead of eccrine (perspiration) glands, lacrimal (tear) glands, and reproductive secretion glands have been noted by embryologists.33
The large number of fatty chemicals composing the vernix impart a chemical signature to the mix making it personally unique.34 In practice, forensic identification of individuals from their vernix caseosa have only been successful to species and blood group.35
Interestingly, much of the vernix caseosa flakes off from the skin of the maturing fetus about a week before birth. The milky appearance of the vernix-amniotic fluid mix can be detected by sonograph. In addition, the gradual changes in the composition of the vernix accelerate at about the 37th week of gestation.36
The neonate comes into the world owning nothing but the gummy substance of oily pheromone covering him completely. Theoretically someone must be willing to provide urgently needed care and attention in exchange for the new baby's sole possession of value: that skin secretion.15 This readiness to nurture for pheromone would provide the same evidence for bonding in all mammals and all bonding species generally. Civilized human beings appear quite prepared to do anything for love of their children, perhaps, especially for men if primed by pre-pregnancy oral, lacrimal, skin, vaginal, areolar and nipple secretions of their wives.
EXOCRINOLOGY
63

The sebum of the vernix may alone be responsible for a mother's love, or it may have companion secretions as back-up. For instance, the shorter chain chemical fatty acid and ester chemical species released during labor and after birth, remain unmeasured. These pervade the atmosphere of the birth room, and may affect the behavior of any of those present. The supposed receptivity must be primed; mothers do love while attending medical personnel do not.
Another likely possibility, pregnant females may begin to secrete colostrum upon the husband's breast stimulation. The behavior presents a perfect opportunity for transmission of a bonding-receptivity-enhancing pheromone, most likely the protein pheromone receptors themselves (see Chapter 6). Lipid carrier proteins of colostrum may be egregiously mislabeled and function as pheromone receptor proteins.
The sebum excretion rate increases during the last trimester of pregnancy, despite high levels of estrogen, a hormone normally inhibiting sebum excretion.37 The elevated sebum secretion rate typical of pregnancy is also maintained during lactation.38 Perhaps pregnant and lactating women are more 'kissable' than nonpregnant, nonlactating women?39 Extra bonding effort during the period when a reproducing female is most vulnerable seems logical.
Another private mate-mate exchange opportunity for fetal-specific pheromone receptivity comes from vaginal discharges in the last trimester of pregnancy. Birch (1956) suggested that rat dams may become familiar with the odor of their own young during pregnancy by perceiving the odors of pre-birth vaginal secretions. To 1983, there had been no test of his proposal.40 Pre-birth and post-birth vaginal secretions ooze into the vaginas as lochia, and may also serve some pheromonal priming function.
Researchers following this author’s suggestion13,14 noticed that some unknown maternal pheromone primes bonding to offspring in Norway rats. They found that three days before delivery of her pups, the pregnant dam releases a priming pheromone onto her bedding. The scientists placed soiled bedding into cages of other rats and watched their behavior toward the dam's neonates. Something in the soiled
64

bedding primed a change in rat behavior. Three days after being exposed, the primed rats of both sexes would no longer cannibalize newborn pups. Instead they behave "maternally".41,42 It should not surprise anyone if mother vaginal secretions were cued and timed to prime fathers as well since human intercourse is so frequent.
Unanalyzed vaginal secretions of sexual readiness accompany even innocent kissing in unmarried human females. Perhaps it is primer pheromone for mate-mate bonding? The effect diminishes (unneeded) after marriage. Sexual exchanges are likely, especially during the last trimester of pregnancy. Marriage bed smells associated only with pregnancy have been reported anecdotally. Nocturnal pheromone exchanges between husband and wife might explain some of the pleasure of sleeping together. Sleeping together has some unexplained cohesive effects.43
At the first hour of the first child's life, inexperienced fathers interact less enthusiastically than second time fathers.44 Similar differential solicitude for inexperienced vs. experienced males pervades the literature. This exactly parallels primiparous maternal behavior and supports the addictive pheromone bonding hypothesis, as well as pheromone receptor protein mediated ‘memory’.
Labor and birth bring on secretions which are characteristic of pregnancy. Some secretions, such as vernix caseosa, are chemically unique and identifiable to the baby being born.33 Mucous plug expulsion mixed with bloody show may also enhance bonding success. This is suggested by the effect of forewaters amniotomy, a minor operation to induce labor, being associated with reduced bonding success. The researchers found a link between forewaters amniotomy and a delayed onset of maternal affection that was unexpected.45 We expect it though. The chemical composition of the mucoid plug damming the fetal sack within the uterus has not been carefully studied. Its natural deterioration in labor may release a maternal pheromone to enhance bonding receptivity. Surgical rupture would prevent or diminish pheromone releases as found, and should thus be minimized obstetrically. The possibility also exists of a semiochemical role for amniotic fluid naturally released during labor.13,14
EXOCRINOLOGY
65

Inducing labor suggests obstetric need. Amniotomy is also associated with long and difficult labor which is also associated in turn with reduced bonding success. So which, forewaters amniotomy or long & difficult labor, causes reduced bonding success? Compositions, concentrations, variations, intra-specific comparisons; we know very little.
Rupture of the amniotic sac (breaking the bag of waters), has important semiochemical consequences in other species. In sheep, the small area marked by released amniotic fluid is exclusive to the marking ewe. If removed from the site, she returns to it to bear her lamb.
The gush of warm fluid heralding a human baby's arrival may simply be for waste removal. It has no known function in humans aside from warning. Mucous glands already line the path of the emerging neonate and provide all necessary lubrication. However, human amniotic fluid does have a substantial sebaceous fraction. This provides hope that human adoption might be artificially stimulated one day. Perhaps such a process might be similar to a practice know to shepherds for millennia (and to shepherd King David of ancient Israel but apparently not to his son, Solomon) putting a ewe's amniotic fluid onto an adoptive lamb to enhance adoptive bonding success.46
Human females bearing young release profuse perspiration and may involuntarily urinate. Again, these are unappreciated specialized secretions. Their most probable functions may be to make way for heat dissipation and for the descending baby's mass, respectively. We will learn with analysis and testing.
Although sudodiferous (perspiration producing) eccrine glands are prominent in humans, the chemistry of pure heat or exertion perspiration is simple, varying with metabolism. (You sweat what you eat, see Leon.10 Mice perspiration is similar.47) The odd perspirations of intercourse48 needs careful examination and chemical analysis as well.
During labor and after the birth, the shorter chain volatile lipid and other emitted chemical species evaporate. These chemicals have not been analyzed either.23
66

If these are pheromones, they can be captured, identified, synthesized and used medically.
Upon the birth of the baby, the mother may be oblivious to all others for a short time. Typically she first inspects the body of her new love. She usually, but not always, kisses her new infant.22
Addiction, like love, can last many decades. If we judge by the strength of the resulting bond, any addictive power of neonatal sebum for receptive females would be awesome. Human sebum would not be any more potent than some insect pheromones, however. Some of those are effective in doses at the ultra-trace level: a few molecules of pheromone in a billion molecules of atmosphere.
Full term neonatal vernix caseosa fundamentally resembles young adult sebum. Wax esters are better represented during these bond-forming times of life. Wax esters "melt" roughly in the sequence of their chain lengths as the skin temperature rises. Sequential meltings are useful for secretions of passion during courtship behavior. Such wax esters are among the most commonly pheromonal.49
The about-to-be mother in the delivery room undergoes poorly understood physiological changes. Blood serum chemistry changes as labor proceeds. The pelvis bones unhinge in birth giving under the influence of progesterone. Human females birthing pains are unknown among other primates. (The painful movement passes the relatively large human head.) For some still obscure reason, long or difficult labors correlate more often with bonding failure. Perhaps maternal pheromone receptivity might erode during labor, or birth canal off-gasing might be less dense?
The bonding effort in those moments right after birth results in immediate expressed love of the just-born infant in about 40% of first time mothers.24 As discussed, like other mammalian species, the human mother examines her baby. (Are all the fingers and toes in place?) If all checks out, bonding to the neonate begins. Deformities are associated with decreased bonding success.50 What physiological doors open with such a realization? Recreating the physiology and pheromone
EXOCRINOLOGY
67

receptivity and pheromone receptions at birth and during the first week could be used to fight or cure physical child abuse and to improve adoption bonding success.
CRITICAL PERIODS
There are two critical periods of parent-to-infant bonding. The first falls during the first hours after birth (Klaus M.H. and J.H. Kennel, 1982).51 Extra contact between mother and neonate for the first hour may even improve the quality of the maternal bond:
Primiparous mothers and their infants who had an extra 15-20 minutes' suckling and skin to skin contact during the first hour after delivery, behaved differently at 36 hours post partum compared with a control group without this extra contact. The present study is a 3 month followup of these mothers and infants by means of direct observation of mother-infant free play and a personal interview with the mothers. Mothers in the extra contact group spent more time kissing and looking en face at their infants; these infants smiled more often and cried less frequently. A greater proportion of the mothers with extra contact were still breast feeding at 3 months. The influence of extra contact on behaviour was more pronounced in boy-mother than in girl-mother pairs (De Chateau, P. and B. Wiberg, 1977).52
This sexually dimorphic maternal behavior may have a pheromonal explanation as we
will soon see.
The 2nd critical period comes during the first days of the neonate's life.30
In about 70% of human cultures, anthropologists recognize a period of 7 to 10 days after birth when the mother and baby are isolated together.30 This isolation may have evolved to enhance and insure mother-infant bonding13,14 It also may serve to protect the bonding mother in her highly receptive state from possible semiochemical aggression of other human beings, or perhaps even them from her? So, DARPA should try to find and weaponize that pheromone, too, then?
68

Since the nineteen-thirties dermatologists have known about a marked postnatal sebum secretion in the neonate.53,54 Better measures have been made more recently.9 Sebaceous secretion levels obtained on the fourth day of life are as high as those of young adults.9 In the mother, the "baby blues" or the "third day blues" precedes the fourth day sebum surge of the baby. These two phenomena may work in tandem to insure mother-to-infant love in the first isolation week.
In one study of first-time mothers, researchers asked them what was their predominant emotional reaction as they held their newborn in their arms for the first time. Forty percent recalled that their predominant emotional reaction was one of indifference. 25 percent of 40 second-time mothers reported the same indifference. Forty percent of both groups felt affection immediately when holding their babies for the first time. Most mothers in both groups had developed affection for their babies within the first week.24 Unfortunately, as its authors begrudge, this study's design is retrospective, its data are subjective, and the work is, to this author’s knowledge, un-replicated.
Perhaps the new baby's vernix caseosa forms the vanguard while the fourth day sebum peak, under the mother's emotional pheromone-receptive tears, is a backup system? Backup systems that chronologically follow up other systems and would be expected.
Charles Darwin first noticed the flushing of the face of newborns when crying (without tears) in his The Expression of the Emotions in Man and Animals (1872). “If the parent does not respond to the infant's satisfaction, a flushed face and loud crying follow. The response to this? The baby, brought close, is kissed.”55 Perhaps that kiss takes the last step in a managed process?
Under this author’s paradigm, the baby must gauge the caregiver's addiction to its pheromonal secretions. One way might be by noticing the pitch of the voice (or a musical slur to higher pitch), the animation of the face, and the gentleness of handling.18 A failing mark on any score stimulates the head and face to flush and produce the kiss me cry. The increased blood circulation to the well-developed sebaceous glands in skin surfaces of the face and scalp instantly increases secretion of
EXOCRINOLOGY
69

sebaceous pheromone. When the caretaker's kiss arrives on the skin secreting the pheromone, the cry is muffled, the increased circulation withdrawn, and the gland cooled. The sebaceous pheromone congeals with the drop of only a degree or so of temperature at the skin surface, conserving pheromone for the next transfer event. The infant's innate instinct again examines the face and sounds it hears, determining if another dose is warranted. "Pheromoning" caregivers might be the newborn's number one job, particularly if staying alive depends on it.
Snatching up and running with an infant will silence a baby. Survival has favored this startle adaptation for a stealthy retreat. Capture by an enemy frequently meant death for babies. Mass infanticides by victors brought young captive mothers back to reproductive fertility the quickest.56 The effect diminishes in older infants, who may require more than motion cues to sense danger.
Assuming loving infants to be a pheromonal process, then only a fraction of the sebaceous pheromone required to addict, maintains addictive love for older children. This must be so because sebum secretion declines unsteadily and sexually dimorphically until the age of about one year. There, secretion remains at a lower level until puberty when the bonding needs of approaching adulthood bring back the higher levels once again.8 We might suspect that high sebaceous pheromone secretion levels make new bonding to the secreting party possible, while low levels maintain such bonds. Paralleling these changes through ontogeny are changes in skin surface pH.57 Such changes may affect semiochemical secretion or reception as well. pH changes can affect soluble pheromone binding protein function,58 improving the pheromone receptivity described in Chapters 6 & 7, when the pheromone binding protein functions as a extracellular pheromone receptor protein.59
Human cultural groups treat public kissing of children in a variety of ways. It ranges from the outright social prohibition among oriental societies to the vulgar "sexual child abuse" of a few primitive tribal societies.60 (Oriental tolerance for crowding and tea taste sensitivity are suggestive as we will soon see.)
In between there may be other pheromone releases influencing maternal behavior. Male nipples function briefly in the perinatal period. The enigmatic
70

"witch's milk" of both sexes and other secretions of the "genital crises" remain unanalyzed and quite obviously, completely misunderstood.
Oriental people have fewer sebaceous pheromone producing glands (Rothmann, S., 1959) compensated, perhaps, by increased receptivity. (Oriental people also seem to have a lower olfactory perception threshold to body odor.61 People of European ancestry living in the Orient might profit by frequent baths.) Sebaceous pheromone receptivity remains unmeasured anywhere, however a greater sensitivity in Oriental people might be expected. Oriental infants are kissed, but kissing of children, especially in public, is discouraged. Vaporous uptake may even suffice. Even the eye folds may improve pheromone receptivity as a more advanced adaptation.
Interestingly enough, depending upon sex of the infant, there are differences in a mother's love early on.
Mother-child interaction was studied on the 2nd and 4th days after delivery. Mothers with girls showed more distal contact behaviour such as talking, smiling, and 'en face' responses. They also displayed more skin-to-skin contact behaviour (patting, rubbing, kissing, and touching) toward girl babies. Mothers with boys, on the other hand, showed more types of behaviours directed to clothed parts of the infant such as patting and adjusting clothes. These findings were statistically significant only on the 2nd day after delivery. On day 4 differences between male and female infants both in the frequency and in the pattern of sucking were observed (Hwang, C.P., 1978).62
Similar slight differences in maternal affection might be explained by differences in sebaceous secretion of baby girls vs. baby boys. For instance, one group of scientists watched mothers kissing, talking and rocking their babies at the age of one month. That group found that male infants got more attention than did female infants.63,64
Another researcher65 had data showing that female infants got more attention than boys during the second six months of life. She also pointed out that females tended to be weaned later that baby boys. Dermatological data bearing on these points are indefinite, but indications are that at one month, male infants will have
EXOCRINOLOGY
71

been secreting more sebum than females.9 The same dermatological report also shows that during the sixth through twelfth months after birth, female infants secrete about half again as much as their male counterparts. Although by no means settled, kissing frequency does seem on first blush to depend upon harvestable quantity of sebaceous pheromone, following economic expectations. Thus parental kissing of babies may indeed depend upon neonatal sebaceous secretion.
Adults use the "blowkiss" as described by Eibl-Eibesfeldt39 on the umbilicus or "belly buttons" of older children in play. Here, too, lurks a sebaceous aggregation in the skin around the infant navel. The navel area's steroid metabolism is similar to other kissable skin.66 The intensity of the pleasure of the youngster suggests the importance of such innocent contact.
Eibl-Eibesfeldt13 also presents an interesting example of genital kissing of children by parents in a primitive South American Indian tribe. Among the Yanomami of the upper Orinoco river in South America, mothers and fathers alike "blowkiss," (The term "blowkiss" for this adult to child kissing is descriptive and should supplant the vulgar British and Australian usage of this term for fellatio.) lick or manually rub the vaginal orifices of baby girls and stroke the scrotums of boys or mouth their penises until about the age of three. The genital surfaces observed have large active clusters of unusually shaped sebaceous glands perinatally.8 These are active in the neonate. Sebaceous glands respond to manual stimulation with an increased secretion rate.67
The parental bonding by pheromone reception hypothesis has great power. Even the savage practice of gumming babies' genitals can be specifically accounted for by non-conscious sebaceous secretion seeking! The Yanomami should have the thanks of all humanity now for their conservation of their natural customs.
72
1 Comfort Alex Nature and human nature. London: Weidenfeld and Nicolson, 1966.
2 Morse, Douglass H. 1980; Behavioral Mechanisms in Ecology. Cambridge, Massachusetts: Harvard University Press 383 p.

EXOCRINOLOGY
73
3 Eibl-Eibesfeldt, Irenäus (1970a). Ethology the Biology of Behavior, translated by Erich Klinghammer New York: Holt, Rinehart and Winston 530 p.
4 Netter, Frank H. 1989; Paranasal Sinuses: Changes with Age. Plate 44. In: Atlas of Human Anatomy Summit, New Jersey: Ciba-Geigy Corporation. 585 p.
5 Drettner B. The paranasal sinuses. In: The nose: upper airway physiology and the atmospheric environment. Proctor A.editor. Amsterdam: Elsevier Biomedical Press. 1982;145-62.
6 Nicholson, B. 1985; An economic model of phermone transmission in avians. Paper presented to the Animal Behavior Society biannual meeting, Raliegh, N.C.
7 Terkel J, Rosenblatt JS. Maternal behavior induced by maternal blood plasma injected into virgin rats. J Comp Physiol Psychol. 1968 Jun;65(3):479-482.
8 Tinbergen, Niko (1960). The Herring Gull's World a study of the social behaviour of birds New York: Basic Books.
9 Montagna, William, and Paul F. Parakkal 1974; The Structure and Function of Skin. New York: Academic Press. 433 p.
10 Agache, P., D. Blanc, C. Barrand, and R Laurent R 1980; Sebum levels during the first year of life. British Journal of Dermatology. 103:643-649.
11 Leon, Michael 1983; Chemical Communication in Mother-Young Interactions. pp. 39-77. Chapter 2. In: Pheromones and Reproduction in Mammals. J G. Vandenbergh, editor. New York; Academic Press, Inc. 298 p.
12 Matsunaga M, Sato S, Isowa T, Tsuboi H, Konagaya T, Kaneko H, Ohira H. 2009; Profiling of serum proteins influenced by warm partner contact in healthy couples. Neuro Endocrinol Lett. 2009;30(2):227-236.
13 Wilson, Edward O. 1975; Sociobiology The New Synthesis. Cambridge, Mass.: Belknap Press of Harvard University Press. 697p.
14 Benfield, D. Gary, Susan A. Leib, and Jeanette Reuter 1976; Grief response of parents after referral of the critically ill newborn to a regional center New England Journal of Medicine, 294(18):975.
15 Nicholson B. Love and kisses: a semiochemical addiction model for human bonding. 1983. Paper presented to the International Society for Human Ethology and Animal Behaviour Society biannual international meeting at Bucknell University, Bucknell PA.
16 Nicholson, B. 1984; Does kissing aid human bonding by semiochemical addiction? British Journal of Dermatology 111(5):623-627.
17 Tamborlane William V (ed). 1997; The Yale Guide to Children’s Nutrition. Newhaven Connecticut: Yale University Press.
18 Dickens G, Trethowan WH. Cravings and aversions in pregnancy. Journal of Psychosomatic Research (1971).15:259-65.
19 Profet, Margie 1992; (In) The Adapted Mind New York: Oxford University Press.

74
20 Vandenbergh, J G. editor Pheromones and Reproduction in Mammals. New York: Academic Press, Inc. 298 p.
21 Shorey, H.H. 1976; Animal Communication by Pheromones. New York: Academic Press.
22 Fillion, TJ, Blass EM. Infantile experience with suckling odors determines adult sexual behavior in male rats. Science 1986;231(4739):729-731.
23 Preti, George, Winnifred Berg Cutler, Carol M. Christensen, Henry Lawley, George R Huggins, and Celso-Ramon Garcia 1987; Human axillary extracts; an analysis of compounds from samples which influence menstrual timing. Journal of Chemical Ecology 13(4):717-732.
24 Greene RS, Downing DT, Pochi PE, Strauss JS. Anatomical variation in the amount and composition of human skin surface lipid. Journal of Investigative Dermatology 1970;54(3):240-47.
25 Reinberg, Alain, and Michel Lagoguey 1978; Circadian and circannual rhythms in sexual activity and plasma hormones (FDS, LH, Testosterone) of five human males. Archives of Sexual Behavior 7(1):13-30.
26 Kolattukudy, P.E., and L. Rogers 1987; Biosynthesis of 3-hydroxy fatty acids, the pheromone components of female mallard ducks, by cell free preparations of the uropygial gland. Arch. Biochem. Biophys. 252(1):121-129.
27 Trivers, R. L. (1972) Parental investment and sexual selection. In B. Campbell (Ed.) Sexual selection and the descent of man, 1871-1971 (pp 136–179). Chicago, Aldine.
28 Garland T Jr, Schutz H, Chappell MA, Keeney BK, Meek TH, Copes LE, Acosta W, Drenowatz C, Maciel RC, van Dijk G, Kotz CM, Eisenmann JC. The biological control of voluntary exercise, spontaneous physical activity and daily energy expenditure in relation to obesity: human and rodent perspectives. J Exp Biol. 2011 Jan 15;214(Pt 2):206-229.
29 Hollenbeck, A.R, JL. Gewirtz, S.L. Sebris, and JW. Scanlon 1984; Labor and delivery medication influences parent-infant interaction in the first post-partum month. Infant Behavior and Development. 7:201-209.
30 Silver, H., R Wapner, M. Vega, and L.P. Finnegan 1987; Addiction in pregnancy: high risk intrapartum management and outcome. J Perinatol 7(3):178-184.
31 Drickamer LC. Acceleration and delay of sexual maturation in female house mice (Mus domesticus) by urinary chemosignals: mixing urine sources in unequal proportions. Journal of Comparative Psychology 1988;102(3):215-21.
32 Hardy, A. C. (1963). Man and the beneficent sea. Evening discourse delivered on 3 September 1962, at the Manchester Meeting of the British Association for the Advancement of Science. Report. British Association for the Advancement of Science. 19: 533-544.
33 Von Eggeling, H. (1940). Origin of cutaneous glands of mammals. Anat. Anz. 90:149-157.
34 Nicolaides, N. 1974; Skin lipids: their biochemical uniqueness. Science. 186:19-26.
35 Tesar J. 1986; [Identification of vernix caseosa.] Soud Lek. 1986 Nov;31(4):54-5. Czech.

EXOCRINOLOGY
75
36 Wysocki SJ, Grauaug A, O'Neill G, Hähnel R. 1981; Lipids in forehead vernix from newborn infants. Biol Neonate. 1981;39(5-6):300-4.
37 Burton JL, Shuster S, Cartlidge M. The sebotrophic effect of pregnancy. Acta Dermato-vernereologica (Stockholm). 1975;55:11-3.
38 Burton, JL., Sam Shuster, M. Cartlidge, L.J Libman, and U. Martell (1973a). Lactation, sebum excretion, and melanocyte stimulating hormone. Nature 243:349-350.
39 Nicholson B. Does kissing aid human bonding by semiochemical addiction? Br J Dermatol 1984 Nov;111(5):623-627.
40 Leon, Michael 1983; Chemical Communication in Mother-Young Interactions. pp. 39-77. Chapter 2. In: Pheromones and Reproduction in Mammals. J G. Vandenbergh, editor. New York; Academic Press, Inc. 298 p.
41 Mennella, Julie A, and Howard Moltz 1988; Infanticide in rats: male strategy and female counter-strategy. Physiology & Behavior 42:19-28.
42 Mennella, Julie A, and Howard Moltz 1989; Pheromonal emission by pregnant rats protects against infanticide by nulliparous conspecifics. Physiology and Behavior 46:591-595.
43 Montagu, Ashley (1944). Some factors in family cohesion Psychiatry 7:349-52.
44 Dunn, D.M., and D.G. White 1981; Interactions of mothers with their newborns in the first half-hour of life. Journal of Advanced Nursing 6(4):271-275.
45 Robson, K.M., and R Kumar 1980; Delayed onset of maternal affection after childbirth. British Journal of Psychiatry 136:347-353.
46 Levy, F., and P. Poindron 1987; The importance of amniotic fluids for the establishment of maternal behaviour in experienced and inexperienced ewes. Animal Behavior 35:1188-1192.
47 Schellinck, H.M., A.M. West, and RE. Brown 1992; Rats can discriminate between the urine odors of genetically identical mice maintained on different diets. Physiology & Behavior 51(5):1079-1082.
48 Masters, W.H., and Virginia E. Johnson (1966). Human Sexual Response. Boston: Little, Brown and Company.
49 Silverstein, Robert M. 1984; Chemistry of insect communication. Chapter 5, pp 105-121. In: Insect Communication. Lewis, Trevor editor. Academic Press, London. 414 p.
50 Cady E, Cady F. How to adopt a child. New York: Whiteside, Inc. and William and Morrow Company, 1956.
51 Klaus, M.H., and JH. Kennell 1982; Parent-Infant Bonding. St. Louis: The C.V. Mosby Company.
52 De Chateau P, Wiberg B. Long-term effect on mother-infant behaviour of extra contact during the first hour post partum. II. A follow-up at three months. Acta Paediatrica Scandinavica 1977;66(2):145-51.
53 Benfenati A, Brillanti F. Sebaceous glands distribution in skin of human body Arch. Italdi. Dermat., sif. 1939;15:33-42.

76
54 Rothman, Stephen (1954). Physiology and Biochemistry of the Skin, Chicago: University of Chicago Press Chapter 12 Sebaceous-Gland Excretion p. 285.
55 Darwin C. The Expression of the Emotions in Man and Animals. London: Murray. 1872;162
56 Eibl-Eibesfeldt, Irena us. (1970b). Love and Hate: On the Natural History of Behavior Patterns. New York: Methuen and Company.
57 Behrendt H, Marvin Green M. Patterns of skin pH from birth through adolescence. Springfield: Charles C. Thomas. 1971.
58 Damberger, Fred, Nikonova Larisa, Horst Reto, Peng Guihong, Leal Walter Soares, and Wüthrich Kurt, NMR characterization of a pH-dependent equilibrium between two folded solution conformations of the pheromone-binding protein from Bombyx mori. Protein Science (2000), 9:1038-1041.
59 Nicholson B., Pheromones cause disease: pheromone/odourant transduction. Med Hypotheses. 2001 Sep;57(3):361-77.
60 Eibl-Eibesfeldt, Irenäus 1983; Patterns of parent-child interaction in cross-cultural perspective. In: The Behavior of Human Infants Oliverio Albert and Michele Zappella editors. New York: Plenum Publishing Corporation pp. 177-217.
61 Schleidt, M. 1980; Personal odor and nonverbal communication. Ethology and Sociobiology 1:225.
62 Hwang, C.P. 1978; Mother-infant interaction; effects of sex of infant on feeding behaviour. Early Human Development 2(4):341-349.
63 Moss, H.A. (1967). Sex, age and state as determinants of mother-infant interaction. Merrill-Palmer Quarterly 13:19-36.
64 Moss, H.A, K.S. Robson, and F. Pedersen (1969). Determinants of Maternal Stimulation of Infants and Consequences of Treatment for Later Reactions to Strangers. Developmental Psychology, 1:239-46.
65 Clay VS. The effect of culture on mother-child tactile communication. (Ph.D. Dissertation. Teachers College, Columbia University, NY: 1966.)
66 Albone, Eric S. 1984; Mammalian Semiochemistry The Investigation of Chemical Signals Between Mammals. Chichester, England: J Wiley & Sons Limited. 360 pages p. 13
67 Mills, JW., and P.M. Quinton 1981; Formation of stimulus-induced vacuoles in serous cells of tracheal submucosal glands. American Journal of Physiology, 241(1):C-18.

Chapter 3: Chemical Characteristics of Human Pheromones
Up to this point, we have examined kissing, the sebaceous glands, their behavioral correlations, and the physical characteristics of sebum. But pheromones carry information, so the chemicals in sebum must also be capable of carrying information.
Now consider this. Identifiable compounds in the human skin surface lipid set number about 700.1 Although ultra-trace detection of pheromone concentration has been observed in other animals, perhaps twenty different detectable concentrations of a single human pheromone component could be considered a reasonable approximation for human reception. For simplicity’s sake, we shall diminish twenty to one. Let us presume that all seven hundred components are equally detectable. Then we have 700 detectable elements, n. If perhaps, say, six elements can be detected as a single signal, the number of detectable combinations of k = 6 elements from a group of size n = 700 elements
so why do we need a hundred quadrillion possible signals when people are acquainted with only about 300 different other human beings in a lifetime? My point here is that we are only scratching the surface and we must suspect more information transfer rather than less.
Since the pheromone mixture set can change anatomically for an entirely different reception. (Presumably we kiss grandma differently from our spouses with different receptivities.) We multiply by those changes again, and since k can be any component compared to itself in another location, another component in the same or in a different anatomical location, or any group up to seven hundred (up to twenty necessary components in differing necessary concentrations had been seen in animal studies), well that is a big, big number, too. Why is it such a big number?
EXOCRINOLOGY
77

Why should there be such complexity? Perhaps, at least, it exists to assure unique individual recognition by pheromone? Did we evolve these chemical signaling molecules over billions of years to give a few microbes tummy aches, when the dry surface of the skin is more effective? Economics predicts that we don’t invent vast alphabets, nay whole languages, for nothing. “Junk” DNA is as economically absurd as “displacement activity” and “nuptial gifting”. (Anyone trained in accounting will recognize that DNA serves double duty as a double-entry accounting system.)
Sebaleic acid, 5Z,8Z-octadecadienoic acid (both unsaturations, one between carbons number 5 and 6, and between carbons number 8 and 9 with counting starting from the functional group (acid) end of the aliphatic chain, are -cis) is unique in all nature and found only in human sebum according to Nicolaides (1974).1 (Thus, we only need sebaleic acid to distinguish a human ‘scent’ as human even though sebaleic acid is odorless, colorless, and tasteless.)
Sebaleic acid, a major component of human sebum, has extremely interesting characteristics. Sebaleic acid is metabolized down the same pathway as omega-six (counting from the non-functional group end of the aliphatic chain) free fatty acids found in fish oil. Fish oils have beneficial health effects in human beings which are both species-specific and synergistic, just like chemical components of pheromones have long been known to be both species-specific (in our case effective only in people) and synergistic (greater effect than the sum of its parts). Metabolites of sebaleic acid breakdown are found in plants said to have fertility-enhancing and even aphrodisiac effects along with other pharmacological properties such as an attractant for human neutrophils.2 Pheromones are chemical signals, after all. Since pheromones work synergistically, analysis of a single chemical’s effects is largely worthless as a predictor of biological activity.
The question becomes "Do sebum's chemicals look like pheromones?" While they may, if sebum carries information, only testing can so determine. Appearance may economically suggest function. The latter requires proof, while the former (appearance) unfolds below.
78

This chapter shows chemical and physical characteristics typical of known pheromones and how they compare to some suspected human pheromones. Human sebum does look suspiciously like known aliphatic pheromones. Human sebum is uniquely human in the same ways that other species differentiate themselves chemically.
EXOCRINOLOGY
79

Use the following chart in deciphering the figures in the text.
80

Figure 1. One of thousands of human-typical triglycerides. Is it a pheromone? Possibly.
We will look at insect pheromones mainly, although avian and vertebrate pheromones appear. Understand, the people who study insects have had advantages over their vertebrate-studying colleagues. (Insects cost little and nobody minds what twisted things you do with them.) The insect people also have a methodological advantage. In particular, they have a simple device known as the electroantennagraph (or EAG) to detect and see direct electrical responses to chemical stimulations on an oscilloscope. Entomological pheromone chemists might suggest they have merely been smarter. Similar apparatus to the EAG, the EOG for electro-olfactograph, has been available for many years.
THE ELECTRO-ANTENNAGRAPH
The EAG can be represented by the drawing in Figure 2. Experimenters pipe a known concentration of a suspected pheromone under the glass with an insect
EXOCRINOLOGY
81
antenna stump. A pheromone evokes a typical frequency pattern of responses that show up on the voltmeter/strip chart. Some similar chemicals also stimulate a response on the meter/chart. EAGs show us which are pheromone-typical chemical characteristics.
Figure 2. EAG or electro-antennagraph.3 The severed antenna is placed under glass and its poles are wired to the positive and negative lead of a voltmeter/strip chart or CRT oscilloscope. Scientists pass an odor under the glass into contact with the antenna and observe the voltmeter to see if the odor chemical generated a biological generated electrical response.
82

Chemists test pheromones and chemicals slightly altered from the pheromone. Using the EAG, subtle changes can show substantial differences in detectability. As experience was built up, the same stereochemical oddities kept showing up to differentiate one pheromone from another. Examining such subtle oddities in putative human pheromones reveals interesting information. Human sebum is a pheromone flexible enough to automate most human behavior. Let us examine the known pheromone-like chemical quirks of sebum one at a time.
STEREOCHEMISTRY ACROSS A DOUBLE BOND
Double bonds (or unsaturations-(footnote for chemists: here reference to an unsaturation will always mean a double bond, never the presence of a ring)) are stiff, flat places between two carbons on a single file, aliphatic carbon chain. A double bond holds its two carbons rigidly. Each end carbon can hold onto (or bond) at most two other atoms. The result resembles two letters Y glued flat together base to base. In a chain, only two of the three prongs of the Y bond to carbons. Most of our double bonds exhibit either an S shape or C shape across them.
Figure 3. shows two stereochemical isomers of 3-heptene, one C shaped (the Z isomer on the left) and the other S shaped (the E isomer on the right). (Hydrogens ignored for clarity.)
These S and C shapes are called "E" or "Z." "E" is for the German word, Entgegen, meaning opposite sided. "Z" is for another German word, Zusammen,
EXOCRINOLOGY
83

meaning same sided. To confuse things, the C shaped, Z chemical may be called "cis" and the S shaped, E chemical may be called "trans."
Figure 4. Sex attractant pheromones of the European pine shoot moth (E)9-Dodecen1-yl acetate and of the grape berry moth (Z)9-Dodecen1-yl acetate. (Adapted from Morrison, R.T., and R.N. Boyd, 19874)
Figure 4 shows two chemicals, pheromones for two different species. They share the same chemical formula, only the fixed shapes or stereochemistry differs. Individual carbon and oxygen atoms attach in the same order for each. They are both constitutional isomers (with the same elements) and stereoisomers (with the same order, but with different shapes).
84

Figure 5. #2. is the silk moth pheromone, Bombykol, called (10E,12Z)10,12Hexadecadien1ol in the language of chemistry. Its three stereoisomers are #1., (10E,12E)10,12Hexadecadien1ol, #3. (10Z,12Z)10,12Hexadecadien1ol, and #4., (10Z,12E)10,12Hexadecadien1ol. (Adapted from Morrison, R.T., and R.N. Boyd, 19874).
As seen here in Figure 5, we show an extreme example of the shape of a molecule affecting its communicating power. Chemical #'s 1, 2, 3, and 4 are stereoisomers. They have the same number of carbons bonded together with exactly the same types of covalent bonds, single bonds represented by a dash ( - ) or slash ( \ or / ) and double bond represented by an equals sign ( = ). Single bonds are flexible and allow rotation, whilst double bonds hold adjoining carbons flat and rigidly locked in the same plane. These four molecules have very similar chemical characteristics. #2 is the pheromone sex attractant (called Bombykol) of the silk moth. It is a billion times more attractive to the male than the other three isomers. The sole distinction among them is the "Z" or "E" shapes around the two rigid double bonds.4
Exclusive mixes of one configuration or the other appear frequently in nature. One or a few related lipids with usually the Zusammen, (zamesided, "Z" "cis" or C shaped) configuration across the double bond show up most commonly. This remains true for most non-pheromone lipids. Suspected human pheromones, like the pheromones of other vertebrates, have this common "Z" (or "cis") configuration. Z-E
EXOCRINOLOGY
85

isomerism has been found in human sebum only once to date5 and that in lipids taken from scalp hair secret haunt of the ‘anger’ or aggression pheromone as seen later.
DOUBLE BOND POSITION & MULTICOMPONENT
COMPOSITION
Double bonds may replace single bonds between any two carbons on a carbon chain. However, in nature most aliphatic chains have consistent unsaturations. Pheromonal lipids differ from lipids found in fatty tissue, however. While most pheromone receptors carefully tune to the position of the double bond.6,7
Figure 6. The main component of the tarsal scent gland of black-tailed deer, cis-4-hydroxydodec-6-enoic acid lactone.
Figure 6 shows the main component of a pheromone for the blacktailed deer.8,9 Although only a few other chemicals were checked, the results were consistent. If the carbon-carbon double bond (C=C) moves from the shown location, the pheromonal power of this chemical falls precipitously. Here, this time in ungulates, double bond (or unsaturation) position determines pheromonal character. Not surprisingly, similar species-specific dienoic lactones have been found in horse sebum, but behavioral assays were neglected.10
86

Figure 7. The sex attractant pheromone of the oriental fruit moth consists of these two stereoisomers in a ratio of 7% the (E) isomer and 93% of the (Z) isomer. The two isomers must be present in precisely these concentrations. The pure (Z) isomer is completely inactive. (Adapted from Morrison, R.T., and R.N. Boyd, 19874). Compare to Figure 8.
The pair of chemicals in Figure 7. illustrateS two characteristics typical of pheromones. First note how these two chemicals resemble those of Figure 4. They have the same molecular formula, except that the double bond has moved over to the eighth (8) carbon from the ninth (9). That puts three carbons on the end beyond the double bond rather than only two.
Secondly, and importantly for our purposes, note that the proportions of the chemical present determine whether an effect is observed. This requirement of a precise mixture of stereoisomers typifies pheromones.4 Mixture precision explains persistent and time-sensitive human sebaceous mixtures as pheromonal.
EXOCRINOLOGY
87

Figure 8. Two stereoisomers of a volatile short chain free fatty enoic acid thought, perhaps, to function as human pheromones moderating menstrual cyclicity in some women with irregular cyclicity. Compare to Figure 7.
Specific concentrations of Z and E isomers have been identified in the isolates of a probable human pheromonal extract. According to Leyden et al., (1990),11 a concentration of roughly ten to one of the Z to E isomers appears. See Figure 8.
The concentration ratio has not been firmly established, though. The author's laboratory submitted for analysis axillary extracts taken from American German-speaking primitive farmers to Leyden's group. In the farmer samples the trans isomer component diminished to 20:1 rather than 10:1. Differences in collection method may explain our results, however. If the difference is real, it could explain Amish social cohesion as pheromonal.
For awhile, many thought that one chemical led robotically to only one behavior.12 Then, Silverstein et al. (1966)13 discovered in insects the first multi-component blend of chemicals having a pheromone effect. (The author had the honor to take a class in spectrometric identification from Silverstein before he retired).
88

Over the years from the sixties, many other examples have been seen.14 That suggested that many chemicals would trigger many components of behavior independently.15 Current belief has it that sometimes pheromones act independently and sometimes not. In insects, precise mixtures of as many as twenty (20) chemical components show up,16 particularly among more social species. Indeed, pheromone component mixture quantity increases with sociality, placing humans on top of that list. We are the most social species on this planet, certainly not ants, as any economist can demonstrate.
Exhibiting another characteristic of semiochemicals, blends of many diverse unsaturated lipids cover human kissable skins. A wide range of unsaturated lipids typifies plant and animal semiochemicals.17
With so many more components possible in an individual's mix, each human being actually has a unique chemical signature in pheromone-type chemicals.1 In crime laboratories human sebum identifies human beings forensically.18 A mature individual's precise combinations of sebaceous lipids remain stable through time.19,20 The ability of dogs to tell twins apart derives from otherwise identical twins having different sebums.21 Since genes are identical in identical twins, epigenetics explains the difference. Epigenetics, the ability of genes to acquire and shed expressions with experience, is pheromonal. Indeed, chromosomal aberrations are attributable to both natural and synthetic pheromone reception in laboratory mice.22
METHYLATION
Unusual methylation marks another chemical characteristic typical of pheromones. A methyl group presents a single, hydrogen-saturated carbon sticking out alone from the carbon chain, a non-reactive chemical ‘bump’. To methylate a carbon chain, you add a methyl to any carbon in the chain but the last (which would just make the chain one carbon longer).
DNA methylation (adding a methyl group to form 5-methylcytosine where cytosine is followed immediately by a guanine in the DNA sequence) and protein
EXOCRINOLOGY
89

methylation of histones are epigenetic. It is logical that these post-translational modifications are pheromonal because it is biologically economical, it explains known pheromonal influences on genetic inheritance, but most importantly, methylation is key to the language of pheromones. Methylation in the nucleus and elsewhere suggests it as an early evolutionary adaptation. From the point of evolution, pheromone-sensing and chemosenses must be our earliest sense of perception. Pheromones have been with us, and with us among our cells, since colonial times: colonial bacteria.
Methyl group being added or "methylated" onto a carbon chain puts a carbon "bump" on it, changing the chain's shape slightly. Can such bumps be read, like Braille, by chemical pheromone receptors in animals? Yes, they can.
Jones et al. (1971) evaluated several positional isomers of Xmethylhentriacontaines as kairomones eliciting antennation by M. croceipes. Bioassays produced the following order of activity (scores in parenthesis) to 150 ng of chemical: 13methyl analogue (2.32), 15methyl analogue (1.68), 12methyl analogue (0.48), 11methyl analogue (0.22), 9methyl analogue (0.00). The data indicate an acceptor site that is very specific for the methyl position (Jones, R.L., 1986).23 [BN emphasis]
Jones' group established that methyl group placement on a carbon chain affects reception. Similarly, such differences in the way drugs are structured affect enzymatic action. Whether a methyl group (or multi-carbon hydrogen-saturated alkyl group) adorns a drug's carbon skeleton may make or break its efficacy. M.I. Kabachnik et al. (1970)24 found that differences in the placement of alkyl groups (such as methyl groups) on drugs affect various human enzymes differently. If a proteinaceous enzyme can distinguish methyl group position, then human pheromone receptor proteins might, too. Recall from above that differences in methyl group placement on a carbon chain can affect insect reception in both chemical and electrical responses.
"[I]n [the ant] A. texana the closer the methyl group was to the fourth carbon atom in 7carbon ketones, the greater the potency (Table I) (Moser, John C., 1970).15
90

EXOCRINOLOGY
91

We shall see that interior methylation of sebaceous lipids, waxes and alcohols takes place more frequently on one particular, species-specific, carbon. Humans sebum shares that characteristic with ant pheromones.
There is evidence in the data that there might be a constancy in optimum methyl position from the other end of the molecule, but more precise studies would be needed to clarify this. For example, the 12-methyl-dotriacontane is very active and the relative position of the methyl group from the other end of the molecule in 11-methyl-hentriacontane, 12-methyl-dotriacontane, or 13-methyl-tritriacontane is the same. (Jones, R.L., 1986).22
Observing this characteristic requires counting carbons from the "wrong" or omega end of the chain. Data has since been gathered to establish this with more certainty.25 Chemists count carbons to name chemicals. Chemists start counting carbons from the end of the chain closest to "the functional group"the area most subject to reaction. Unfortunately pheromone receptors seem to count the carbons the other way, from the end furthest from the functional group.
CHAIN LENGTH and MIXTURES OF CHAIN LENGTHS
Vinson et al. (1975) provide a more detailed study with the response of Cardiochiles nigriceps to a series of X-methyl hen, do and tri-triacontaines. These data provide information on chain lengths as well as positional isomers and support those of Jones et al. (1971) with respect to non-synergism by chemicals with methyl groups in variable positions. They also show a bell-shaped dose-response curve, with optimum responses at 5 micrograms of chemical. The optimum methyl positions did vary with chain length and were 11-methyl-hentriacontane, 16-methyl-do-triacontaine, and 13-methyl-tri-triacontane.
The most interesting feature of the study was a marked synergism observed when chemicals of varying chain lengths were mixed. Mixtures of several methyl isomers of hen, do, and tritriacontanes exhibited synergistic activity, with dose-
92

response curves having optimum activity at the 500 ng level and very good activity at the 50 ng level. These results provide strong evidence that the parasitoid perceives hydrocarbons of varying chain lengths at different acceptor sites and that the different acceptor sites have different optima for methyl positions (Jones, R.L., 1986).22
Human sebum contains one of the largest biological collections of chain lengths science has ever seen, except in semiochemicals.
[In the ant, A. texana] Seven-carbon chains evoked stronger responses than shorter or longer chains (Table II.) (Moser, J.C., 1970).15
EXOCRINOLOGY
93

That chain length also plays a role in pheromone reception also might be presumed from varying drugs prompting enzymatic action. (Likewise the drug action might be suspected to mimic intracellular or other pheromonal action, as well, remembering our ‘colonial’ phylogeny lesson.)
94

Dixon and Webb (1964) have shown rate profiles of 17 enzymes in which the chain lengths of alkyl and acyl groups attached to substrates were successively varied by the addition of methylene groups. In general, as the length of the linear hydrocarbon chain increased, a regular increase in enzyme activity was observed up to a maximal value, followed by a decrease in activity as the chain length was further increased. The optimal chain length varied widely among the enzymes, ranging between 3 and 17 carbon atoms.26
Note that Dixon and Webb27 purified only one enzyme for examination at a time in their beautiful systematic experiment. Enzymatic activity, such as metabolizing or synthesizing hormones, may depend upon recognition of chain length.
FUNCTIONAL GROUP
The sensitivity to functional group varies widely in pheromones. (An alcohol has one functional group, a ketone has another.) Moser (1970)15 held everything constant, but changed the functional group on his pheromone to an alcohol. Table III shows that the alcohol, 4-methyl-3-heptanol, produced alarm in A. texana. The effect just like the real pheromone, needed a concentration 100,000 times higher than the related [pheromonal] ketone.15 Moser was another brilliantly systematic chemist.
EXOCRINOLOGY
95

The chemical activity of the functional group does not enter into things. Recognition of pheromones does not require any chemical reactions where new bonds form.6 Another brilliant scientist, Prestwich (1987)28shows us still another example of functional group specificity in a pheromone.
3. Agrotis segetum
The responses of sensory cells from male antennae of the turnip moth, A. segetum (Noctuidae), were measured upon stimulation with analogs of Z712:Ac (I, the pheromone) in which the structure of the acetate was varied (Liljefors et al., 1984) (Figure 9.). Replacement of the alcohol oxygen with a methylene to obtain the methyl ketone (LXIII) afforded a weakly active stimulus. In contrast, the formate (LXIV), the propionate (LXV), the trifluoroacetate (LXVI), and the ethyl ether (LXVII) analogs were all essentially inactive.
96
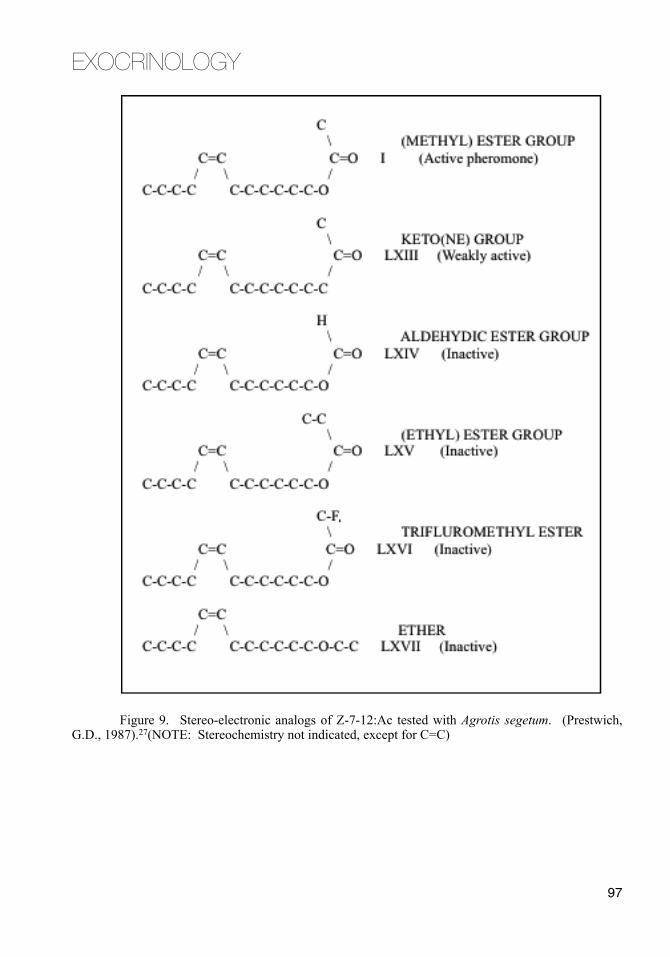
Figure 9. Stereo-electronic analogs of Z-7-12:Ac tested with Agrotis segetum. (Prestwich, G.D., 1987).27(NOTE: Stereochemistry not indicated, except for C=C)
EXOCRINOLOGY
97

FUNCTIONAL GROUP POSITION
In all but the aldehydes and carboxylic acids, the simpler functional groups can appear anywhere along the carbon chain. As noted, functional groups fail to react chemically in pheromone recognition by animals.4,6 The pheromone's shape seems the most important factor. Where the functional group appears along the carbon chain may have as much relevance to pheromone specificity as the group itself.
[In the ant, A texana] the ketone group was most effective on carbon atom 3; 4-methyl-3-heptanone was 10,000 times more potent than 4-methyl-2-heptanone. ... When the ketone and methyl positions were reversed (3-methyl-4-heptanone), sensitivity was decreased by 10,000 times. [See Table I.]
SAME CHEMICAL, TWO SPECIES
Two or more species may react to a single chemical. For example, 4-methyl-3-heptanone does not excite Trachymyremex septentrionalis (McCook), a leaf-cutting ant closely related to A. texana, but it does alarm Pogonomyrmex comanche (Wheeler), a distant relative. Nests of both species may be adjacent to those of A. texana (Moser, 1960) (Moser, John C., 1970).15
Figure 10. Identical chemical excites two different ant species.
98

The same chemicals have been shown to be pheromones in more than one species. How can this be? Won't the two species get all mixed up? After all, in the lab, we have seen that pheromone stimulation can result in unnatural behavior29 such as interspecific mating. More likely, the same chemical, pheromone to one, allomone to the other, communicates information to both species simultaneously depending upon the needs of the species. Ordinary dogs, common mammalian parasites of human beings, lick human skin. Animal to human bonding may or may not take place. The beasts may deposit allomone to direct human behavior, or to mark the human host as parasitized for “other pets”.
Pheromones rarely become confused in nature. When two species do use the same chemical pheromone for similar purposes, you see chemical emissions that take place in different seasons, at different times of the day or night, or in different habitats.30
DOUBLE BONDS IN HUMAN SEBUM
Let us now look more narrowly at human sebum. Human skin surface lipids have highly unusual patterns of unsaturation. Human sebum contains more than 200 examples of such double bonded carbon chains. These fatty acid chains do not show up in body fat in any quantity.1 The patterns of double bonds distinguish them from those seen in sebums of other animals.1 Here we see evidence of information bearing capacity.
The presence of double bonds at delta 6 [i.e. between the 6th and 7th carbons counting from the functional group end] in the mono-enoic fatty acids of human skin surface lipid is a striking feature (A.W. Weitkamp, et al., 1947; N. Nicolaides, et al., 1964; N. Nicolaides and T. Ray, 1965). In most naturally occurring mono-enoic acids the double bond position is at delta 9. Mono-enoic acids with the delta 6 double bond are extremely rare, occurring only in human skin, the sebaceous tissues of some other animals [with different chain lengths & functional groups-Nicholson] and the seed fats of the parsley family and some other rare plants (G.F. Spenser, et al.,1971) (Nicolaides, N., 1974).1
EXOCRINOLOGY
99

FUNCTIONAL GROUPS IN SEBUM
The tri-acyl glycerols constitute 60 percent of the lipids of human sebum (Table IV). "Interestingly the triacyl glycerols and their products derived from hydrolysis occur only in the skin surface lipids of human beings".1 (Hydrolysis, or water cutting, splits a molecule, capping off the ends with an H and OH from H2O. Hydrolysis may be performed on human triglycerides by bacterial action, or environmental action.1 Thus the presence or absence of various combinations of skin surface microbes affects sebaceous composition (but see Cove, J.H., et al., 1980).) We know of only so many functional groups in carbon chemistry. To find not one, but several apparently reserved for human use among mammalian sebums was unexpected. However, free fatty acids do function as pheromones in several species. We do see free fatty acids similar to human sebum's (differing only in unsaturation position and methylation) in bumble bee pheromones31 and wood tick pheromones32 Peculiar branching fatty acids characterize some goat pheromones33 and hydroxylated, maybe even duck pheromones.34
Perhaps other studied mammals have found their best interests in reducing the chances of being mistaken for human? Chemical differentiation of pheromones probably evolved in the dim mists of unrecorded time. Perhaps, an ancient evolution accounts for their relative simplicity, chemical recognition35 and pheromone secretion.36
Again, other ideas may explain human sebum's uniqueness.37,38 Is sebum excreted as a waste product? After all, only about a third of the triacyl glycerols or triglycerides of human sebum could profitably be used internally.1 A waste disposal task for sebum can be ruled out, however. Squalene, a biologically useful chemical, is a common precursor. Sebum is generally manufactured in the skin from squalene with the exclusion of common precursor fatty acids for relatively rarer ones.
Perhaps as Nicolaides suggests1 different sebums fool bacteria or other pathogens by being difficult to digest? In light of Kligman's29 cogent and lasting
100

analysis, that possibility remains doubtful. Since skin bacteria ignore sebum,39 and with so many other sebums function as pheromones,40,41 maybe that misleading idea will fall from persistent medical acceptance error? Hopefully, the dramatic changes in human behavior brought about by oral intake of skin surface pheromones (loss of criminality, addiction, and sexual perversion) will eventually convince everyone.
Table IV shows major components of human sebum. The long-chain wax esters in the mix probably maintain the wide melting point characteristics of sebum. That heat-variable viscosity,42 you will recall, looks important for human passionate behavior. Esters, the second most numerous set in sebum, form the most numerous class of functional groups among known pheromones.6 Some economic functionality for the complexity seems likely in light of the trouble and expense the skin glands apply in making sebum.1
The wax ester component of human sebum expands so quickly just before birth at the end of gestation that physicians can spot it on sonographs to gauge labor.43 Wax esters also expand during puberty but more slowly until an equivalent mix is
EXOCRINOLOGY
101

obtained at reproductive maturity.44 Both ester percentage increases precede peak bonding periods of birth and young adulthood. An economic "just in time" logic suggests, along with the chemical evidence of the prevalence of ester groups in pheromones, that these human wax esters may stimulate or enhance human bonding pheromonally. Do you see? This is all fitting together too well to allow psychological ‘explanations’ to continue to exist.
METHYLATION OF HUMAN SEBUM
In sebum, all but the iso chains have methyl groups only on the even numbered carbons (with carbon atom of the carboxyl group being counted as #1).1 While methyl groups do show up on the much commoner penultimate and antepenultimate carbons; however, in his own words:
[...] Maximum amounts of methyl branching appear on the fourth C-atom for all chain lengths. The significance of this is not known.1
As it turns out, methyl branching on a particular carbon in the chain characterizes many pheromones. Those methyl bumps may work like the teeth on an old brass key. Return to Table I again. Recall how A. texana liked the 4th carbon for its lone methyl group? Recall how M. croceipes could detect that 13-methyl analogue? Finding peculiarly consistent methylation on fourth carbons of human sebaceous fatty chains represents chemical evidence for our sebum being pheromonal.
Funny methylation changes in sebum can be seen with respect to age.45 Kosugi and Ueta44 investigated sebum of children and old people versus young adults. They noted that a methyl group appeared predominantly on the third carbon from the end of the chain in children and old people, while fertile adults had their carbon on the second carbon from the end. Kisses for children and old people differ from those
102

of fertile adults, as do differential solicitude and patterns of abuse and criminal behavior. The chemistry is, indeed, different!
Human sebum goes further with its peculiarities. Dimethyl (two methyl groups) carbon chains are common, but again, only on the even carbons.1 While this may suggest some sort of synthesis anomaly, we have seen that receptors exist in other animals which can distinguish methylation from methylation.22,24 Indeed, only a methyl group differentiates hormone adrenalin from its nor-adrenalin semi-antithesis, suggesting early evolution of this sensing ability. Recall hormones were once pheromones among colonial microbes before the organization of organism bodies.
CHAIN LENGTH and MIXTURES OF CHAIN LENGTHS IN
HUMAN SEBUM
Sebum's components exhibit an "extremely wide range of chain lengths".1 A few strictly even or a few strictly odd chain lengths usually compose lipid accumulations in plants and animals.4 A mixing, as here in sebum, typifies pheromones. The mix stretches from fewer than 6 carbons to as many as 38, in vernix caseosa. Recall what Jones (1986) reported above. In receptive tissues, a synergism, a more than additive increased sensitivity, can result where chain lengths mix.
Very long chain lengths make the pheromone stay stuck longer. Longer chain and therefore heavier and less volatile pheromones tend to persist in the environment longer and thus tend to be utilized as territory marking pheromones. Long chain species wash off with water less easily. They evaporate less readily, also. Vernix caseosa's persistence suggests that carrying a little one home to mark the bed might improve family life. We should try, the divorce rate remains too high.
EXOCRINOLOGY
103
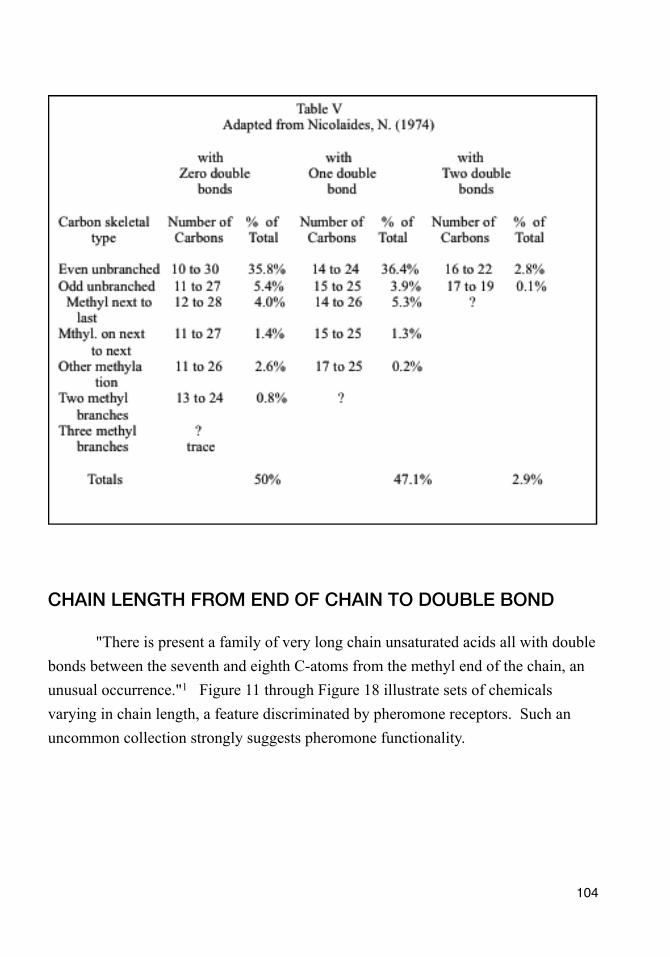
CHAIN LENGTH FROM END OF CHAIN TO DOUBLE BOND
"There is present a family of very long chain unsaturated acids all with double bonds between the seventh and eighth C-atoms from the methyl end of the chain, an unusual occurrence."1 Figure 11 through Figure 18 illustrate sets of chemicals varying in chain length, a feature discriminated by pheromone receptors. Such an uncommon collection strongly suggests pheromone functionality.
104
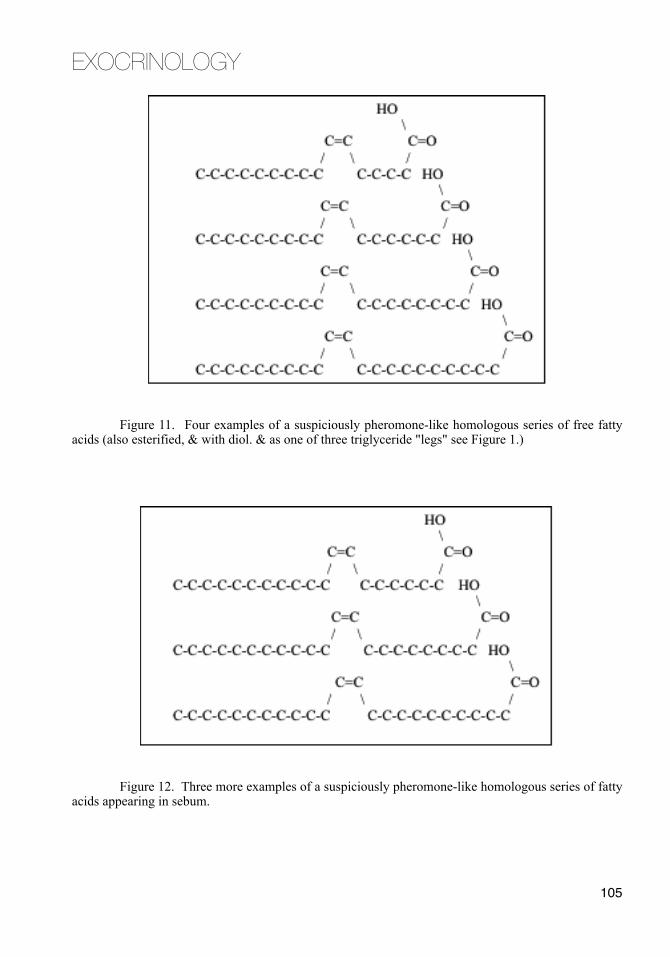
Figure 11. Four examples of a suspiciously pheromone-like homologous series of free fatty acids (also esterified, & with diol. & as one of three triglyceride "legs" see Figure 1.)
Figure 12. Three more examples of a suspiciously pheromone-like homologous series of fatty acids appearing in sebum.
EXOCRINOLOGY
105

Figure 13. Three more examples of a suspiciously pheromone-like homologous series of fatty acids appearing in sebum.
Figure 14. Four more examples of a suspiciously pheromone-like homologous series of fatty acids found in human sebum.
106
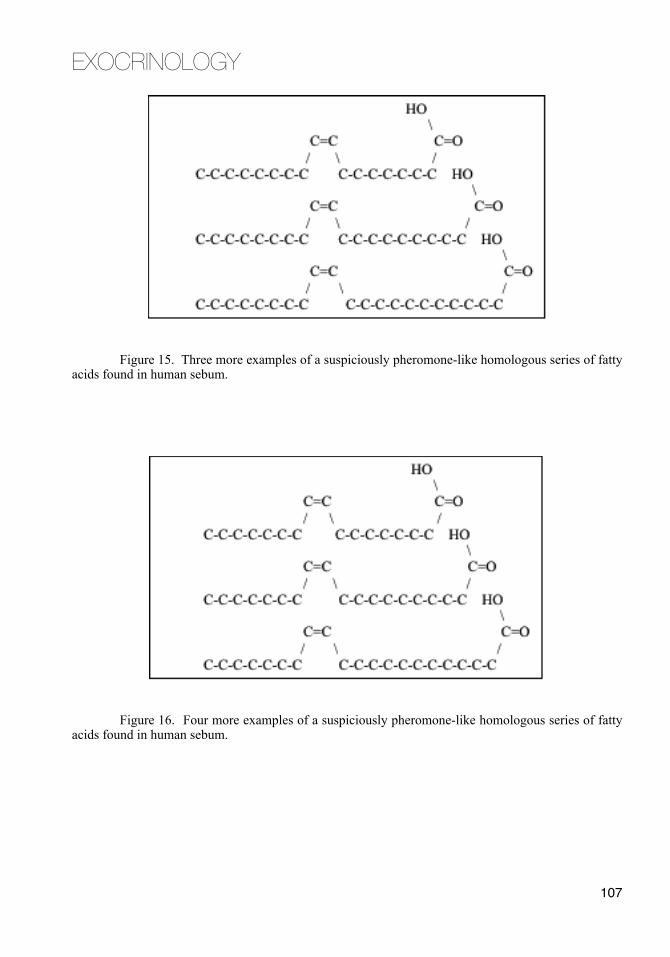
Figure 15. Three more examples of a suspiciously pheromone-like homologous series of fatty acids found in human sebum.
Figure 16. Four more examples of a suspiciously pheromone-like homologous series of fatty acids found in human sebum.
EXOCRINOLOGY
107

Figure 17. Three examples of a suspiciously pheromone-like homologous series of iso-free fatty acids.
Figure 18. Three examples of a suspiciously pheromone-like homologous series of iso-free fatty acids.
108

ESTER PRIMER PHEROMONES
Also included must be mono-esters of the Figures 11 through 18 fatty acids (and shorter ones that Nicolaides1 declined to identify, such as 3-methyl-2-hexenoic acid) esterified to related long chain fatty alcohols, and at least two types of diesters. The first diester type would be formed from these fatty acids with an alpha-hydroxy esterification to another fatty alcohol and a third group of fatty acids. The second would consist of two subtypes 1,2 alkane diols esterified to two groups of fatty acids; or 2,3 alkane diols esterified to two groups of these fatty acids (see Nicolaides, N., 19741). The numbers indicate order on the carbon chain. A 1,2 alkane diol is an alkane (all Hydrogen saturated carbons with only single bonds) with two alcohol, or OH, groups located on the first and second carbons. Diol means a chemical species with two alcohol groups on it, generally.
As we've said, esters proliferate in the vernix abruptly just before the baby delivers and much later more gradually during puberty, peaking in early adulthood. Because bonding reaches its highest frequency in these periods, the esters in Figure 19 through Figure 21 and their homologues, may together stimulate or reward bonding onset.
Cholesterol esters are found more commonly in childhood and neonatal sebums, differentiating them from adult-typical compositions.46 Higher concentrations of linoleate are found in childish sebum, too.47 These, and other childhood-typical chemical species, should be investigated as candidates for a human child-aversion or child recognition pheromone. A human child-aversion pheromone could explain the Kibbutz aversion data and find a place in semiochemical medical practice for treatment of sexual child abuse.
Pheromonal aversion can involve nausea, probably involving the Peyer’s patches full of microvillar cells in the alimentary tract. Since disgust sensitivity predicts intuitive disapproval of sexually perverted people,48 this seems a reasonable conclusion to draw.
EXOCRINOLOGY
109

What makes esters the most frequently pheromonal functional group? Perhaps esters are merely easier to transfer in kisses? Chemically, esters are among the least reactive functional groups.
Figure 19. Methylated enoic monoester typical of those thought to induce bonding. Derived from the first fatty acid in Figure 11 and from a methylated straight chain from an iso C 20 carboxylic acid commonly found in sebum. See Table V for more examples.
Figure 20. One of two main types of di-esters found in human sebum. It is similar, except in chain lengths and 4th carbon methylation, to esters found in scent glands of other animals.
110

Figure 21. The second main type of di-esters found in human sebum. It is similar, except in chain lengths, 4th carbon methylation and unsaturation position, to esters found in sebaceous scent glands of other animals.
NON-PHEROMONE COMPONENTS IN SEBUM
Why should obvious non-pheromones show up in sebum? Why should Palmitic acid, Myristic acid, Stearic acid, and Oleic acid comprise 25.33%, 6.88%, 2.89%, and 1.87% respectively of the fatty acids in sebum? Wouldn't you expect to find sebum to be completely pheromonal? No, you would not. (Palmitic acid is found in palms, butter, cheese, milk and meat. Myristic acid is named after nutmeg. Stearic acid is found in many animal and vegetable fats and oils, like cocoa. Oleic acid is the most abundant fatty acid in human adipose tissue.)
Well, a fatty acid mix containing palmitic and oleic acids stimulates sexual behavior in male ticks.31 These same two chemicals stimulate emigration for two genetically different strains of common fruit flies.49
EXOCRINOLOGY
111

First, you might expect to find a very few "intermediates." Nicolaides1 errs to suggest that "intermediate compounds accumulate in the skin." Batch processing, where intermediates do accumulate, typifies digestion not secretion. However, since cells burst to deliver sebum, something minimal might remain of that cellular machinery.
Second, other chemicals play supporting roles in any environmental chemical. Unsaturations lower the melting point of a fatty acid. A few unsaturations are all that separate lard-like shortening from vegetable oil. For a semisolid that can hold on in skin pores, but which will melt in the heat of a lover's embrace, a balance must be struck.
Taken into consideration must be opposing costs of manufacture to the body, what component dissolves what other, and what affects the melting point, hydrophilicity, and so on of the entire mixture. A test of this straightforward hypothesis might be modeled with a matrix of equations for each constraint in a linear program, but the thesis seems logical. Chemicals released into the air also might dispense with co-products.
Take another look at Table IV. Squalene is an unctuous vehicle commonly used by pharmaceutical companies. The pharmaceutical use for squalene (from shark liver) is as a vehicle for ointments. Might it function similarly here? Squalene's low melting point <20o C or 4o F) suggests that it supplies the egg-and-milk in the batter. Perhaps it dissolves some of the otherwise solid fats? Interestingly squalene itself reportedly has pheromonal properties, at least in aliphatic nonvolatile sebaceous secretions of garter snakes.50
OTHER HUMAN PHEROMONES[?]
Ellin's group,51 followed by Dubinin et al. (1985),52 examined volatile human skin effluents. "The authors remark (Ellin, R.I. et al., 1974) that although they had expected to observe a large number of [relatively volatile] alcohols and ketones in the effluvia of man, they had not expected the large number of unsaturated and branched-
112

chain hydrocarbons they observed nor had they expected isoprene to be one of the most abundant human volatiles."53 Dubinin's group43 found an unexplained increase in palmitoleic acid (another of the few components of adipose tissue) in the facial sebum of men over a short time.
Terpinoid pheromones (see Wheeler, J.W.54) originate from those abundant isoprenoic volatiles. As has been shown, unusual unsaturated and branched-chain hydrocarbons would be expected in volatile human pheromones. Short term changes in whole human volatile emissions might reflect impatience, anger, or merely the progress of digestion. Localization of emissions, controlling and testing for psychogenic cause (if any) should take high priority in any systematic study of volatile human semiochemicals.
Some of these studies were stimulated by astronauts’ space suit design. Now, one of their own, poor Lisa Nowak’s fate may depend upon it. Her exposure to high concentrations of ozone (space laundry) doubtless caused her violent outburst against her competitor for the favor of astronaut William Oefelein. Atmospheric ozone concentration correlates nicely with human violence.55
CHEMICAL CONCLUSIONS
We must avoid the trap of thinking too soon we have solved the puzzle. Even based upon clearcut chemical and behavioral evidence, sound judgment can not substitute for demonstration. Diesters of 3-hydroxy C8, C10, and C12 acids, the female mallard duck pheromones, were found as the major products of the uropygial [preen] glands only during the breeding season.33
"Take a gander" at Figure 22. Notice that varied chain lengths, and consistent hydroxylation are consistent with these chemicals being pheromones, too. Production only during the breeding season, also provides evidence of pheromonal activity. Doubtless as a marking pheromone to post territories during ‘displacement activity’ where the birds gather these chemicals from their preen gland and spread them around their nests on objects they pick up and drop once they are covered with the
EXOCRINOLOGY
113

species-specific chemicals. It leads one to suspect that there must be no such thing as ‘displacement activity’ but instead just territory marking activity. Similar behavior by hundreds of species of birds, the so called ‘displacement activity’ concerning ‘nuptial gifts’ is going to be better understood as pheromone receptivity lacrimation and pheromone-induced bonding activities. More later.
Figure 22. 3-hydroxyoctanoic acid, 3-hydroxydecanoic acid, and 3-hydroxyundecanoic acid. Pheromone (Kolattukudy, P.E., and L. Rogers, 1987) secretions of the female mallard duck preen gland.
It may look like a duck. It may quack like a duck. But is it really a duck? Before we describe any chemical as a pheromone, it must be isolated, bio-assayed, synthesized, bio-assayed and then the whole must be replicated elsewhere.56
This chapter has introduced some of the chemicals jumbled together in human sebum. Those peculiar things setting sebum apart from other lipids put it in a class
114

with known pheromonal chemicals. Pheromone receptors in diverse species know and use those same chemical oddities, as well. Conservation of this ability is not out of the question at all. Together with social, dermatological and physiological data presented earlier, the case for human pheromones improves.
EXOCRINOLOGY
115
1 Nicolaides, N. 1974; Skin lipids: their biochemical uniqueness. Science. 186:19-26.
2 Cossette Chantal, Patel Pranav, Anumolu Jaganmohan R., Sivendran Sashikala S, Lee Gue Jae, Gravel Sylvie, Graham Francois D, Lesimple Alain, Mamer Orval A, Rokach Joshua, and Powell William S. Human neutrophils convert the sebum-derived polyunsaturated fatty acid sebaleic acid to a potent granulocyte chemoattractant. April 25, 2008. The Journal of Biological Chemistry, 283:11234-11243.
3 Boeckh J, Kaissling KE, Schneider D. Sensillen und Bau der Antennengeissel von Telea polyphemus (Vergleiche mit weiteren Saturniden: Antheraea, Platysamia, und Philosamia). Zool J Anat 1960;78:559-84.
4 Morrison, Robert Thornton, and Robert Neilson Boyd 1987; Organic Chemistry Fifth Edition. Boston: Allyn and Bacon, Inc. 1486 p. (p. 347)
5 Weitkamp, A.W., A. Smiljanic, and S. Rothman (1947). The free fatty acids of human hair fat. Journal of the American Chemical Society Aug. 69:1936-1939.
6 Silverstein, Robert M. 1984; Chemistry of insect communication. Chapter 5, pp 105-121. In: Insect Communication. Lewis, Trevor editor. Academic Press, London. 414 p.
7 Roelofs, W.L. 1979; Pheromone perception in Lepidoptera. In: Neurotoxicology of Insecticides and Pheromones. New York: Plenum Press.
8 Müller-Schwarze, D (1969). Complexity and relative specificity in a mammalian pheromone. Nature (Lond.) 223:525-526
9 Brownlee RG, Silverstein RM, Müller-Schwarze D, Singer AG. Isolation, identification and function of the chief component of the male tarsal scent in black-tailed deer Nature 1969;221:284-285.
10 Frost ML, Colton SW the 6th, Wertz PW, and Downing DT. Structures of the dienoic lactones of horse sebum. Comparative Biochemistry & Physiology - B: Comparative Biochemistry. 1984;78(3):549-52.
11 Leyden, JJ, X.-N. Zeng, K. McGinley, H.J Lawley, and George Preti 1990; Characterization of pungent axillary odors. Journal of Investigative Dermatology. 94(4):549.
12 Wilson, Edward O. (1963). Pheromones. Scientific American 208:100.
13 Silverstein, Robert M., JO. Rodin, and D.L. Wood (1966). Sex attractants in frass produced by male Ips confusus in ponderosa pine. Science 154:509-510.

116
14 Linn, C.E. Jr., L.B. Bjostad, JW. Du, and W.L. Roelofs (1986a). Redundancy in a chemical signal: behavioral responses of male Trichoplusia ni to a 6-component sex pheromone blend. Journal of Chemical Ecology. 10(11):659-668.
15 Moser, J C. (1970). Pheromones of social insects. pp. 161- 171. (In) Control of Insect Behavior by Natural Products. Wood, David L., Robert M. Silverstein, and Minoru Nakajima editors. New York: Academic Press 345p
16 Linn, C.E. Jr., M.G. Campbell, and W.L. Roelofs (1986b). Male moth sensitivity to multicomponent pheromones: critical role of female-released blend in determining the functional role of components and active space of the pheromone. Journal of Chemical Ecology. 12(3):659-668.
17 Handel, Steven N., and Andrew J Beattie 1990; Seed dispersal by ants. Scientific American 263(2):76-83a.
18 Tesar, J 1986; Indentifikace mazu syroveho (vernix caseosa). soud Lek. 31(4):54-55.
19 Montagna, William 1974; An introduction to sebaceous glands. Journal of Investigative Dermatology. 62(3):120-123.
20 Downing DT, Strauss JS. Synthesis and composition of surface lipids of human skin. Journal of Investigative Dermatology 1974;62(3):228-44.
21 Moulton, David G. 1977; Minimum odorant concentrations detectable by the dog and their implications for olfactory receptor sensitivity. pp. 455-464. In: Chemical Signals in Vertebrates Mu ller-Schwarze, D and Maxwell M. Mozell editors. New York: Plenum Press 610 p.
22 Daev EV, Surinov BP, Dukel'skaia AV, Marysheva TM. Chromosomal abnormalities and spleenocyte production in laboratory mouse males after exposure to stress chemosignals. Tsitologiia. 2007;49(8):696-701.
23 Jones RL. Orientation by insect parasitoids. pp. (In) Mechanisms in Insect Olfaction Payne, T.L., Martin C. Birch, and C.E.J Kennedy editors. Oxford: Clarendon Press. 1986;149-156.
24 Kabachnik, M.I., Brestkin, A.P., Godovikov, N.N., Michelson, M.J., Rosengart, E.V. and V.I. Rozengart (1970) Pharmacol. Rev. 22, 355-388.
25 Jones, RL., W.J Lewis, M.C. Bowman, M. Beroza, and B.A. Bierl (1971). Host-seeking stimulant for parasite of corn earworm: isolation, identification and synthesis. Science .c 173:842-843.
26 Foye WO. editor. Physical-chemical properties and biologic activity. In: Principles of medicinal chemistry 3rd ed. Philadelphia: Lea & Feibiger 1989;18.
27 Dixon M, Webb E. Enzymes. New York: Academic Press, Chapter 6. (1964).
28 Prestwich, Glenn D. 1987; Chemical studies of pheromone reception and catabolism. Chapter 14, pp. 473-527 (In) Pheromone Biochemistry Prestwich, Glenn D., and Gary J Blomquist editors. Orlando: Academic Press, Inc. 565p.
29 Tumlinson, James H., and P.E.A. Teal 1987; Pheromone Biosynthesis and its regulation. pp 3-26. Chapter 1. In: Pheromone Biochemistry. Prestwich, Glenn D., and Gary J Blomquist editors. Orlando: Academic Press, Inc. 565 p.

EXOCRINOLOGY
117
30 Morgan, ED 1984; Chemical words and phrases in the language of pheromones for foraging and recruitment. Chapter 8, pp 169-194. In: Insect Communication. Lewis, Trevor editor. Academic Press, London. 414 p.
31 Boch, R, and D.A. Shearer (1965). Alarm in the beehive. American Bee Journal 105:206-207.
32 Allan S.A, JS. Phillips and D.E. Sonenshine 1989; Species recognition elicited by differences in composition of the genital sex pheromone in Dermacentor variabilis and D. andersoni (Acari: Ixodidae). Journal of Medical Entomology 26(6):539-546
33 Sugiyama, T., and H. Sasada 1988; Chemistry and biochemistry of unusual branched fatty acids with pheromonal activity from mature male goat. Seikagaku - Journal of Japanese Biochemical Society. 60(2):118-122.
34 Kolattukudy, P.E., and L. Rogers 1987; Biosynthesis of 3-hydroxy fatty acids, the pheromone components of female mallard ducks, by cell free preparations of the uropygial gland. Arch. Biochem. Biophys. 252(1):121-129.
35 Gould J. Ethology the Mechanism and evolution of behavior. New York: Norton Press 1982
36 Von Eggeling, H. (1940). Origin of cutaneous glands of mammals. Anat. Anz. 90:149-157.
37 Guthrie, RD. (1970). Evolution of human threat display organs. In: Evolutionary Biology, vol. 4. Dobzhansky, T., M.K. Hecht, and W.C. Steere editors. New York: Appleton-Century-Crofts.
38 Kligman, A.M. (1963). The uses of sebum? pp. 110-124. In: Advances in Biology of Skin. Vol. 4. the Sebaceous Glands (ed. by Montagna, W., RA. Ellis, and A. Silver) Oxford: Pergamon Press.
39 Cove JH, Holland KT, Cunliffe WJ. An analysis of sebum excretion rate, bacterial population and the production rate of free fatty acids on human skin. Br J Dermatol 1980;103:383.
40 Mykytowycz, R (1970). The role of skin glands in mammalian communication. In: Advances in Chemoreception, Vol. I. Johnston, JW., D.G. Moulton, and A. Turk editors. New York: Appleton-Century-Crofts.
41 Mykytowycz, R, and B.S. Goodrich 1974; Skin glands as organs of communication in mammals. Journal of Investigative Dermatology 62(3):124-131.
42 Burton, J L. (1970). The physical properties of sebum in acne vulgaris. Clinical Science 39:757-767.
43 Wysocki, S.J, A. Grauaug, G. O'Neill, and R Hähnel 1981; Lipids in forehead vernix from newborn infants. Biol. Neonate. 39:300-304.
44 Sansone-Bazzano, Gail, B. Cummings, A.K. Seeler, and RM. Reisner 1979; Differences in the lipid constituents of sebum from pre-pubertal and pubertal subjects. British Journal of Dermatology 103:131.
45 Kosugi, Hiroko, and Nobuo Ueta 1977; The structure of triglyceride in human sebum. Japanese Journal of Experimental Medicine 47(5):335-340.

118
46 Stewart, Mary Ellen, and Donald T. Downing 1991; Unusual cholesterol esters in the sebum of young children. Journal of Investigative Dermatology 95(5):603-606.
47 Stewart, Mary Ellen 1992; Sebaceous Stewart, gland lipids. Seminars in Dermatology Jun; 11(2):100-105.
48 Inbar Y, Pizarro DA, Knobe J, Bloom P. 2009; Disgust sensitivity predicts intuitive disapproval of gays. Emotion. 2009 Jun;9(3):435-9.
49 Narise, S., and T. Narise 1991; Chemical communication of emigration behavior of Drosophila melanogaster II. identification of chemical substances. Idengaku Zasshi - Japanese Journal of Genetics 66(4):411-420.
50 Mason, RT., H.M. Fales, T.H. Jones, L.K. Pannell, JW. Chinn and D. Crews 1989; Sex pheromones in snakes. Science 245(4915):290-293.
51 Ellin, RI., RL. Farrand, F.W. Oberst, C.L. Crouse, N.B. Billups, W.S. Koon, N.P. Musselman, and F.E. Sidell 1974; An apparatus for the detection and quantitation of volatile human effluents. J Chromatog, 100:137-152.
52 Dubinin, D.M., V.P. Naidina, and S.N. Zaloguev 1985; [Evaluation of human skin function in a sealed room by chromatographic method.] Kosm. Biol. Aviakosm Med. 19(6):69-73.
53 Albone, Eric S. 1984; Mammalian Semiochemistry The Investigation of Chemical Signals Between Mammals. Chichester, England: J Wiley & Sons Limited. 360 pages p. 13
54 Wheeler, James W. 1977; Properties of compounds used as chemical signals. pp 61-70 In Chemical Signals in Vertebrates. Müller-Schwarze, D and Maxwell Maxwell M. Mozell New York: Plenum Press 609 pages p. 66
55 Rotton, J, and J Frey 1985; Air pollution, weather, and violent crimes: concomitant time-series analysis of archival data. Journal of Pers Soc. Psychol. 49(5):1207-1220.
56 Silverstein Robert M. 1981. Pheromones: background and potential for use in insect pest control. Science 213(4514):1326-1332.

Chapter 4: More about Bonding: Neglect and Abuse
Because the ground is chapt, For there was no rain in the earth The plowmen were ashamed, They covered their heads.
Yea the hind also calved in the field, And forsook it, Because there was no grass.
Jeremiah 14:45
Human beings commit infanticide with greater regularity than any other species. This disproportion may be due at least in part to our human-specific hygiene practices. Bad thinking or bad thoughts probably takes no part, but there is no way to tell, is there?
Who is a typical child abuser? The male or less often female abuser is of reproductive age.1 Frequently he has never committed a crime in his life, although if he is a criminal he is more likely to abuse his children. He has low socioeconomic status, he is a victim of unemployment, poverty, and Medicaid usage.2 He feeds, clothes, shelters, protects and loves his entire family except for the one child being abused. He is often new to the family. He has no close-knit extended family on which the family can lean for "supportive services," i.e. baby sitting, talk, and financial assistance.
The typical physical child abusive parent does not believe that he is a child abuser. He, like the other members of the family, feels that the victim deserves what he gets and has been treated quite fairly. On the average he is just as likely to
EXOCRINOLOGY
119

physically abuse children of either sex. The siblings of the abused, both brothers and sisters, are treated normally, doubtless in the very same way other parents live with their children.
The abuse takes the form of horrible burns, ("The kid just pulled the pot off the stove onto his head") bruises and bones broken from blows ("he ran into a wall"). ("He fell. He is just an unusually clumsy child.") The abuser may grab the poor child by the arms and shake them so severely that radial fractures in the bones of little arms will appear in x-rays.
Perhaps the physical child abuser tells the truth? Biologically speaking, like the hind in the fields of ancient Israel, they are innocent of any wrongdoing. Plausibly, they merely suffer symptoms of a pheromone deprivation, or even an anti-pheromone poisoning.
New boyfriends are over-represented among physical child abusers. New boyfriends typically become acquainted with infants outside of critical pre-delivery primer pheromone secretion periods. They therefore, have reduced bonding success as opposed to natural parents present at the birth. Infanticide can be seen in many species,3 even birds.4 According to Eibl-Eibesfeldt,3 infanticidal males improve their reproductive fitness by bringing females into earlier sexual receptivity.
The acts of physical child abusers range from "forgetting to feed them," to inflicting slow death by the most horrible mutilations imaginable.5 Parent-infant eye contact is lacking.6
This chapter builds a theoretical model of physical child abuse and suggests to physicians how a medical treatment for the disorder could be devised. The model includes explanations drawn from the ethological literature for factors associated with abuse. Additionally many physiological, behavioral, and biochemical anomalies gain a single rationale.
Love might be made artificially. A love-creating technology, long the lore of shepards and long under development for rats, may be useful in pheromonal medical treatments for human physical child abuse syndrome, for sexual child abuse
120

syndrome, for child neglect syndrome, and to enhance the parental experience in adoptions of older children.7
If either failed sebum secretion or failed sebum consumption contributes to physical child abuse, then it may be repaired. By replicating the physiology of bonding between mother and baby at birth, artificial love bonds will be possible.
Dermatologists associated sebum with skin acne.8 Accordingly since the 1940's they have sought control of the sebaceous pheromone producing glands without knowing or even guessing they were pheromone producing glands. Dermatologists pioneered manipulating these glands' secretions, thanks to acne's significance. Importantly for our purpose, they learned to restart the secretion of the glands of toddlers and children up to the age of about twelve.9,10
Simply put, if the glands make the pheromonal secretions and if the secretion makes the wanted behavior, then making the glands secrete will make the wanted behavior. Given that child abuse and neglect are truly exocrine disorders, then a failed technology for treating acne vulgaris may find use in semiochemical/pheromonal medical treatments.
Why develop artificial love? Extensive examination of moral, ethical, and legal concerns remains a matter for institutional review boards. Few would object to strengthening the bond between an orphan and adopting parents. However, such technology used in treatment of physical child abuse, sexual child abuse, and child neglect would supplant legal authority bent on punishment and misguided revenge.
Other applications of pheromone administration by mouth and of skin topical hormonal stimulations might treat divorce and settle interpersonal disputes and complaints amicably. Crime will become a chief complaint of the past. The very idea of runaway children will become stuff of legends. Having no more orphans, no more neglected and no more physically abused children presents a worthy medical goal. Getting rid of most lawyers and courts has been a worthy if previously preposterous goal of civilization since Shakespeare:
EXOCRINOLOGY
121

"The first thing we do, let's kill all the lawyers!" -
(Act IV, Scene II).
King Henry the Sixth, Part II
Sebum acts as a semiochemical pheromone in many other species. Animals send out pheromones, chemicals sent out by one animal which affect the behavior or physiology of another of the same kind of animal.11
Pheromones may be categorized incorrectly as releasers or primers. Releaser pheromones get an immediate response from the receiving animal. An example might be the sexually receptive male silk moth, Bombyx mori. A tiny whiff of pheromone (just three parts per billion of air) motivates him. He criss-crosses the scent trail or pheromone plume upwind (chemotaxis) to the emitting female just as quickly as his little wings will get him there. Releaser pheromones stimulate behavior right away, with no lag time. If determination indicates the reward, then getting releaser pheromone must feel better than slaking just about any other appetite. Getting a male to do anything else but follow the scent engenders only frustration. Pheromone reception should follow the economic law of diminishing marginal returns. After getting so much sex attractant, the insect should become indifferent between chasing more pheromone and some other behavior. The function should vary from species to species. It reminds one of hypothalamic auto-stimulation. Pheromone reception and association pathways do reach the hypothalamus.
Another recognized type of semiochemical, primer pheromones, cause long term changes in behavior. Often primer-caused changes may not manifest themselves for a season, or even years. Additionally, repeated exposure may affect the final behavioral expression. For a case in point, in the moose, exposure to dominant male
122

scent will determine the adult social status of a colt.12 A similar phenomenon is seen in mice.13 Most of these primer pheromones will be the pheromone receptor proteins needed for pheromone recognition in the species.
A primer-type pheromone receptor protein inoculation might explain the tendency of abused children to abuse their own children. A vaccination analogy might be appropriate. Such a primer pheromone might alter pheromone reception to evoke "inappropriate" responses to subsequent semiochemical stimuli. This might be most understandable if the primer were an antibody to the pheromone itself. Similar ability to recognize chemicals for bodily reaction is conferred from mother to neonate via breast milk.14 Breast milk probably contains fatty acid pheromones active in neural development from sebaceous glands of Montgomery ringing the areolae. Oddly enough, these fatty acids could function similarly to deoxycholic acid or other acids detected in maternal horse droppings15 and maternal rat feces.16,17,18,19 Offspring ingest these essential chemicals from feces for their own neural development. Of course, bad habits like social isolation run in families, too.
Declining levels of sebum shortly after birth coupled with the pheromone addiction hypothesis of human bonding offer a logical, physical reason for mother-infant bond failure, child neglect, and physical child abuse. Sebum acts, perhaps, as a releaser-type pheromone, whose receptivity begins upon earlier or continuing pheromone priming.20,11 As such, repeated synchronized exposures and some time would obtain the best effect. However, proteins of the pheromone receptor type have been found in various sebaceous emissions and their presence there and delivery with the releaser sebaceous pheromone suggest the mechanism for ‘experienced’ sexual and maternal behavior in many species, including man. The inoculation of pheromone receptor proteins must enhance innate receptivity, molding sexual identity.
Thinking critically about the hypothesis leads to some reasonable expectations. We would expect that the behavior of sebum in neonates, infants and children could account for our differential feelings for these little ones. Differences in sebum composition are particularly evident when the sebum of prepubertal children compares with that of young adults. One such difference is the higher concentrations of linoleate found on the skin of children.21 Another is the unusual
EXOCRINOLOGY
123

cholesterol esters found in the sebum of neonates and young children. Cholesterol esters having fatty acids of more than 18 carbons are a prominent feature of fetal skin surface lipid (vernix caseosa), but are a minor component of adult lipid.22 These may be pheromones that stimulate aversion/avoidance or maternal feelings in non-perverted adults. The lack of receptivity for these or other child-specific pheromones suggests the biological cause for sexual child abuse. Increasing receptivity under experimental conditions resembling those which usually occur naturally will correct physical child abuse and neglect.
From experimental evidence in sheep, individual olfactory signatures are major determinants of early maternal discrimination between twin lambs.23 Much of the time, humans can distinguish people by smell but often with consistent errors of identification.24,25
Babies of first-time mothers suffer disproportionately more of the beatings, burns and starvations of physical child abuse5 in both humans and animals.26 Experienced mothers bond more readily to subsequent neonates. These observations indicates that some sort of learning or immunity plays a role in bond formation. Perhaps new mothers must learn non-consciously, via uptake of pheromone receptor proteins, the pleasure of neonatal pheromone receptions?
The phenomenon of "uninfected" siblings baffles psychological physical child abuse researchers. How can an abusive parent love and care normally for "uninfected" brothers and sisters? How can younger and older children in the family receive perfectly normal parental attention while the child victim of physical abuse suffers so pathetically? Chance may play a large part in a pheromonal bonding.
Infants expose their parents to the most sebum when parental love most serves survival: at birth. About 40 % of first time mothers love their newborns immediately.27 Most of the rest bond successfully by about the third or fourth day with all forming the love bond within the first week of life.28 The first week, with a peak on day four, sees high rates of sebum secretion not seen again until early adulthood.29
124

That babe's 4th day surge of sebum follows the day 3 perinatal lacrimal episodes and post partum depression in many mothers.30 Sebum secretion and lacrimation may synchronize to assure semiochemical pheromone receptivity in the mother.7,31 We have touched on this already and more will be said in Chapter 6.
Some neonatal behavior in the arms of parents may reinforce parental addiction to sebaceous pheromone. The attention getting behavior, alternately smiling and crying, depends upon the attentions of the mother.32 If the holding parent gives insufficient attention, the baby objects with a crying episode. Insufficient attention may be indicated by lack of squeaky-voice, making faces, and so on. We have discussed this, too. With abuse, this ritualized behavior ceases. Abused babes stare vacantly while waiting--like their mothers.
"The link between forewater amniotomy and a delayed onset of maternal affection was unexpected" (Robson, K.M., and R. Kumar, 1980).27 Bringing on labor artificially may disrupt the programmed ritual of pheromone secretion and pheromone reception. The operation itself indicates prolonged or difficult labor, indicative of still more risk factors.
Economically, why should parents neglect their children? Within the context of the present paradigm, the disorder may result from a lack of sebaceous secretion on the part of the neonate, a lack of transfers to the parents, or a failure of parental reception.
EXOCRINOLOGY
125
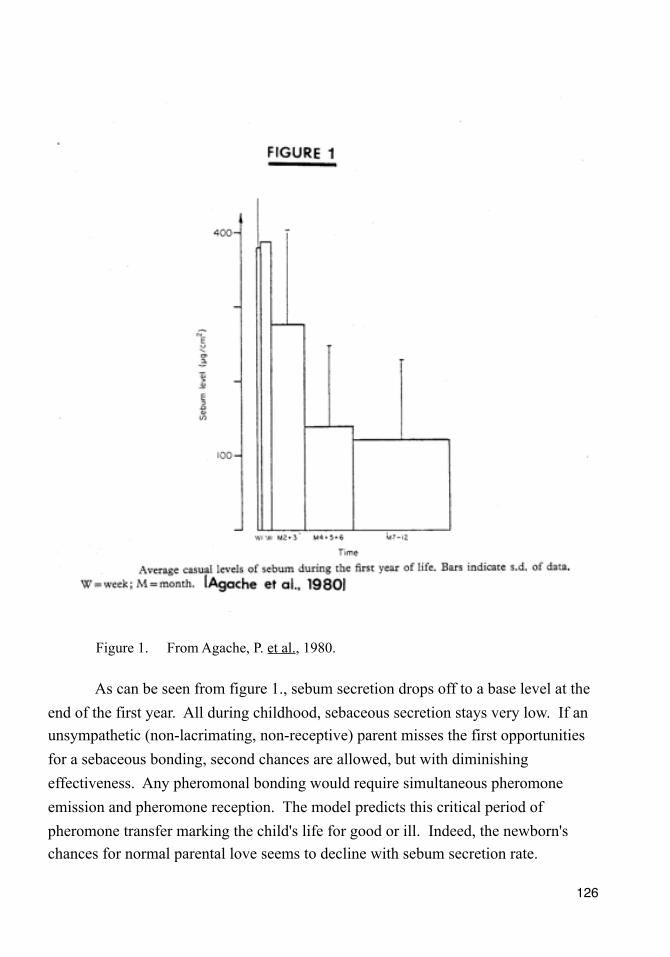
Figure 1. From Agache, P. et al., 1980.
As can be seen from figure 1., sebum secretion drops off to a base level at the end of the first year. All during childhood, sebaceous secretion stays very low. If an unsympathetic (non-lacrimating, non-receptive) parent misses the first opportunities for a sebaceous bonding, second chances are allowed, but with diminishing effectiveness. Any pheromonal bonding would require simultaneous pheromone emission and pheromone reception. The model predicts this critical period of pheromone transfer marking the child's life for good or ill. Indeed, the newborn's chances for normal parental love seems to decline with sebum secretion rate.
126

A plausible explanation for parent-neonate bonding has just been put forward, but that poses another question: Why should it be so much harder to fall in love with an older child? Older orphans the world over exist with no hope for real parental love. Adoption agencies try to place babies with adoptive parents as soon after birth as possible. Their rule from decades of clinical experience: "the earlier, the better." Placing infants into the arms of new parents improves the chance that the adoption will "take" and decreases the likelihood of child abuse. Experience has also taught us to pity the plight of the older orphan doomed to grow up at arms length from his caretakers.33
Perhaps if one or both of the parents do not get secretion to satisfy their semiochemical appetites, increased risk of physical child abuse results? A source for appeasing those parental appetites might be vernix or infantile sebaceous secretion. Sebaceous secretion falls off after the first month. Declining secretion levels after the first year or so may wean the parents, making way for the adult appetites to bring on a little sister or brother, recall.
Occasionally, parents get separated from their new baby for extended periods. Sickness, jail, or war may keep them apart. Over time, less and less sebaceous pheromone remains for parental kisses. Any physical separation during the first year precludes parents getting the fullest portion of infantile sebum. From whatever the cause, parents risk insufficient exposure to their little one's sebum and other pheromones in the first year. Insufficient uptake of baby's addictive pheromone may jeopardize the parent-infant bonding process. Perhaps a "pheromone deficiency" jeopardizes appropriate bonding?
This may answer why adopting couples prefer babies, too. Older orphans must surely be easier to manage? Older babies and infants have less sebum than their juniors.29 Perhaps the adopting parents' marked preference for babies is, in fact, a marked preference for baby sebum and its reputation for bonding success.
Science can change low childhood sebaceous secretion. We can stimulate sebum back up to perinatal levels if we must.34,35 Hope now exists for a
EXOCRINOLOGY
127

semiochemical medical treatment for physical child abuse by recreating the pheromone receptivity in the parents and providing the child’s pheromone.
The model of pheromone stimulated parental love for a child must explain several peculiarities of physical and sexual child abuse. Let us look at them each in turn.
PREMATURITY AND SEBUM
Babies born prematurely suffer more than their share of parental abuse and neglect.5 Two facts might explain the common abuse of premature babies. Premature infants may require a longer stay in the hospital. Longer hospital stays reduce parent-infant time together during the critical first hours and first week.28,36,37 With prematurity, the opportunity for synchronized pheromonal bonding under optimum conditions of high secretion, high receptivity and isolation is lost. Remember the mother-infant isolation for the first week of life? More extensive stays in the hospital, for either infant or mother, are associated with reduced bonding success.5 The pheromone transfer break-off of parent-infant separation due to the prematurity would not be the only hazard. The sebaceous glands of premature infants are smaller and less well developed than those of full term infants and of adults, premature babies' sebaceous glands vary from term newborn and adult skin with "the lipid ... preserved to greater advantage in term and adult skin."38 Logically, insufficient sebum excretion must be considered as a medical reason that premature infants suffer disproportionate abuse. Hopefully, it will soon become a forgivable offense.
The vernix collected from preterm (<37 weeks) infants differed markedly from that collected from near-term (> or = 37 weeks) infants. The principle difference was an increase in the content of squalene relative to other lipids in premature sebum. This change indicated a surge in fetal sebaceous gland function at about 37 weeks gestation just prior to birth.39 Squalene forms an intermediate in the synthesis of human sebum, including shorter chain molecules.40 Those short chain molecules have high vapor pressures, so they evaporate quickly after a birth.
128

Unfortunately these potentially pheromonal off-gases remain unstudied.41 Amoore (1974) first proposed a pheromonal function for human short chain fatty acids.42 They are produced in human glands at sites essentially identical to scent glands of many mammals.43
Another difference between premature and full term sebums is that wax esters in the sebum increase just prior to birth.39 Wax esters of our triglycerides and free fatty acids also increase as puberty progresses.44 Chemically then, sebums emitted at birth and marriage have wax esters in common. Children and premature neonates have different sebums than full term infants and young adults. Esters are the chemical functional group most utilized for pheromone communication among the hundreds of insect and vertebrate chemistries known.45 These associations are suggestive.
Not only the vernix caseosa differs in premature births. The glands of prematurely born human infants also differ significantly from those of full term infants.46 Such differences in gland maturity typically affect the chemical composition of the sebum secreted. The backup 4th day sebum surge found in the skin of normal infants may be lacking, absent, or dysfunctional in premies. If the hypothesis is correct, then perhaps semiochemical medical monitoring of pheromonal emissions may help cue parents for their best receptivity efforts.
Twins and other multiple births, on the average tend to suffer a disproportionate share of physical child abuse.5 Twins and other multiple births tend to be born prematurely. Perhaps prematurity's risks alone can account for the correlation? Alternatively, perhaps identical babies have diminishing marginal utility to drive the correlation. Epigenetic sebaceous differentiation increases with age.
Sebaceous secretion can be measured a number of ways.47 For instance in a technique useful in neonates sebum secretion can be measured photometrically by use of sintered glass pressed on the skin.48 A simpler method, more accurate for greater quantities, might be by gravimetric comparison.49 Easier still might be either the Sebutape or the Lipometre/Sebumeter method.50,51 (Nicholson Science uses the gravimetric method learned at Cunliffe’s knee at the Royal Infirmary at Leeds.)
EXOCRINOLOGY
129

Exocrinologists cringe on hearing unlovable infants described as "dry babies." Abusive or neglectful parents may also describe the babies as "dry." Note the cross-cultural consistency of appellation. Elderly spinsters may also be described as "dried up" old ladies. Sebaceous secretion declines precipitously with menopause.40 However, the look, feel, and moistness of elderly skin does not fall off with menopause.52 Low levels of sebaceous gland activity do not correlate to occurrences of dry skin.53
Perhaps unlovable "dry babies", as some workers describe them, are merely dry of sebum? The lack may even be temporary. Note the wide standard deviations in the French team's data (figure 1). These data allow enough variability to accommodate temporarily "dry" skin.
Overall males and females share an equivalent perinatal sebum secretion. An identical 1 to 1 sex ratio is seen for victims of physical child abuse.54 If a semiochemical sexual dimorphism existed, then one would expect an inverse relationship to skin surface lipid output. Cases of physical abuse are about equally distributed between boys and girls. While female and male infants may have sex-typical patterns of excretion, overall they have equivalent sebum excretion rates. Expectation of a broad consistency between rates of abuse and rates of excretion is fulfilled.
The same study found a different ratio for sexual child abuse, with an overwhelming preponderance of female victims. An exocrinological epidemiologist would suspect other than a sebaceous etiology for sexual child abuse. Perhaps perpetrators have a receptivity problem? Perhaps the lopsided victimization stems from some non-recognition of some child-secreted aversion-inducing pheromone as rats have?
When virgin female Norway rats (Rattus norvegicus) initially encounter rat pups, they begin to alternate tentative approaches, at which time they sniff the pups, with an active avoidance of the pups.55,56 Two lines of evidence indicate that strange pups emit an odor that females find aversive.57 Perhaps the Israeli
130

kibutzim children's strong disinclination to marry their on-campus childhood playmates as adults may also result from a similar primer aversion pheromone?
According to this new model, if the sebaceous pheromone does not get through to the adult, then abuse becomes much more likely, but not automatic. Self-control can exert itself. Human appetites can be overcome by the will. A semiochemical bonding disfunction may be a "dry baby" secretion problem, a reception problem on the part of "cold" parents, or some evil combination.
PHYSICAL EVIDENCE
If these glands are key to acceptability, then animal experiments should show this too. Since similar glands are small and diffuse in most mammals, cutting them all out, or extirpating them, poses problems. However in birds, adults seem to bond semiochemically to their young and each other,7,58 and most sebum secretes from one big preen gland (Lucas, A.M., 1980).59 The preen gland also goes by the term uropygial gland. Gland extirpations have been tried with predictable results.60,61,62 Hou found that the gland-extirpated bird lost social acceptance and stole sebum from unwilling conspecifics. That might also indicate an auto-pheromone deficiency and a persistent appetite for the substance gotten by the act of preening. Balthazart and Schoffeniels (1979)63 correctly found that mating was disrupted without the main gland.
The secretion rate of the sebaceous uropygial gland also declines precipitously after hatching in tested avian species.59 One should expect a similar difficulty for adopted immature birds. Indeed, Lorenz63 noted a similar declining readiness to bond after hatching among geese.
EXOCRINOLOGY
131

If one makes the rather cruel experiment of transplanting a gosling into a strange family, it will be found that the later it is removed from its family the more difficulty the poor baby will have in finding acceptance by the strangers' triumph community (Lorenz, K., 1963).63
Primatologists recount episodes of newborn death among chimpanzees. For a few days, the mother may drag the dead body of her infant about where ever she goes, "grooming it." If she treasures the carcass for its precious groomed-for pheromone, then these illogical anthropomorphic attributions of ape sentimentality might finally be called into question. After all, animals do not bury their dead. Comparing preferences for sebum smeared furs vs. ether, ethanol or detergent cleaned controls might shed some light on this. Placing synthetic pheromone on synthetic fur and testing opposed to controls would help more. Primatologists need to run more experiments to test hypotheses, to be scientists, to stop pandering to a gullable public and get out of the storytelling business altogether. Animals do not have personalities and neither do we.
A sebaceous vernix caseosa covering at birth and neonatal secretion surge have been found in most mammals and birds tested (e.g. Yager, J.A., et al., 198864; c.f. Montagna, W., and P.F. Parakkal, 197440) These vary in chemical composition species-specifically.65,40,41 These facts support a hypothesis of pheromonally induced (or cued) parent bonding to offspring. One would expect an absence, or aversion-inducing perinatal secretions, in nonbonding species.
Animal evidence exists that pheromones may play a role in offspring abuse and neglect. "Failure to thrive" can be symptomatic of neglectful parental behavior among many other ills. Benuck and Rowe66 found that olfactory bulb-ectomized Norway rat dams had a higher proportion of pups dying early in the perinatal period. Perhaps the pups suffered from neglect (or from sudden infant death syndrome?) This is suggested because pup urine has been found to be attractive to adult rats, and bulb-ectomized dams tend to lick the perianal region of their pups less than control dams (Charten et al, 1971, cited in Leon, M., 1983).67
132

Bulb-ectomized dams spent less time in contact with their pups and their pups grew less than pups of sham-operated controls. [However] "females made anosmic by means of zinc sulfate did not have any more deaths than control dams (Benuck and Rowe, 1975). (Leon, M., 1983)67
Bulb-ectomy can cut out pheromonal sensation as well as olfactory. Given the difference of behavior seen, zinc sulfate seems to target olfaction rather than pheromone reception. However, pleasant smells probably still have their place, more so for some species than others. The licking behavior observed also suggests that a relatively nonvolatile pheromone may have been responsible for the differences in maternal behavior. Rat sebum remains a logical candidate. It should be noted that experiments with animals involve other than chemosensory variables. Controlling for them can be difficult or counter-intuitive.
In the brain of a rat, ablation of the septal region causes permanent loss of parental behavior. The septal area makes up a part of the limbic system in both the rat brain and the human brain. Phylogenetically, the limbic system extends the old rhinencephalon, or nosebrain. It routes and integrates appetites, including chemical appetites like addiction, and serves other functions.
Ablation and stimulation experiments have shown us much about nasal-origin pheromone sensing in the brain. Pheromonal inputs travel distinct from normal smells. Inputs move from mucosal neural detectors in each nasal passage to the pheromone sensing accessory olfactory bulbs (where present).68 They go: nasal passage to olfactory pathway/accessory olfactory bulb to cortical amygdala to entorhinal cortex to subiculum to the rest of the hippocampus.73,69 Incidentally, from the amygdala out, the areas described are the areas hit hardest by Alzheimers Disease, with earlier cases showing more lesions rostrally (noseward). A pheromonal pathology for Alzheimers may be indicated.31 More recently, intranasal insulin, which affects chemoreception, improves memory in Alzheimers patients.70 Another pathway takes off at the amygdala for the hypothalamus,69 the brain center for emotional and other appetites.
EXOCRINOLOGY
133

DRUGS AND PHEROMONES DON'T MIX
Dopaminergic-active drugs, such as amphetamine, can modulate the regular odor detection performance of male rats.71 It appears that some alarm pheromone (or pheromones) shares an affinity for brain benzodiazepine receptors.72,73 Naturally, if a pheromone (or pheromones) were responsible for bonding in animals, then you would see inverse consumption of pheromone vs. addictive drug. Higley et al.73 performed such an experiment and the results were as predicted by the pheromone-addiction bonding hypothesis. With greater pheromone exchange opportunity, less sweetened ethanol solution was consumed, while with less pheromone exchange opportunity, more booze was consumed. Microeconomically, this substitutability defines substitutes. Human pheromones, possibly simple facial sebum, might be used as a substitute for addictive drugs.
Twenty-two 50-month old rhesus monkeys were provided concurrent free access to an aspartame-sweetened 7% ethanol solution and an aspartame-sweetened vehicle before, during, and after social separation. Subjects had been reared for their first 6 months of life either without access to adults but with constant access to age mates (peer reared), a condition producing reduced exploration and increased fear-related behaviors, or as controls with their mothers; thereafter, all subjects received identical treatment. During home-cage periods, for 1 hr each day, 4 days a week, when the ethanol solution and vehicle were freely available, peer-reared subjects consumed significantly more alcohol than mother-reared subjects. When stress was increased via social separation, mother-reared animals increased their alcohol consumption to a level nearly as high as that of peer-reared monkeys. [...] Social separation increases alcohol consumption to levels producing intoxication even in monkeys not particularly vulnerable to stress. (Higley, J.D., et al., 1991)73
Higley's experiment must be repeated with the chemical pheromone alone vs. alcohol consumption. Similarly, Harry Harlow's famous studies of wire and terrycloth "motherless monkeys"74,75 must be rerun without the psychological mumbo jumbo. Higley's team continues a dismal tradition of psychological incompetence and a pathetic readiness to project human mental processes onto other species.
134

Perhaps semiochemical-controlled experiments would demonstrate an underlying pheromonal effect?
Very young monkeys were reared singly in comfortable cages which had two mother figures: a "wire mother" and a "cloth mother." Behind each mother there was a light bulb providing warmth and each mother could be equipped with a nursing bottle attached to its "breast." One group of monkeys had their cloth mothers equipped with nursing bottles and another group of monkeys had their wire mothers similarly equipped. . . Both groups spent relatively more time with the cloth mother than with the wire mother in spite of the fact that only one group secured its nourishment from the cloth mother.78
The preference for the cloth mother was rationalized as an instinctive desire for "contact comfort." "[B]oth mothers provided the basic known requirements for adequate nursing, only the cloth mother provided an additional variable of contact comfort."77 Wire contains less surface area than terry cloth. Films of these experiments show wire mothers were made of chicken fencing. Perhaps the monkey's own confidence building skin secretions were better deposited upon the cloth dummy than the wire one? Harlow did not control for pheromone deposition, thus his conclusions (and most of the "conclusions" deriving from this work continuing today under Soumi and others) must be treated with skepticism if not outright derision. Better controlled tests must be run.
Robson and Kumar27 found that during labor, doses of more than 125 mg. of meperidine, (or Demerol) a powerful analgesic, delayed the onset of bonding. Hollenbeck and his coworkers76 found that among other factors observed during feeding in the first month, higher doses of all types of medication, even aspirin, received by mothers during the first four postpartum days were associated with mothers kissing their infants less often. If kissing picks up an addiction-assuaging pheromone, these puzzling behaviors would gain logical explanations. NSAIDs allay pain and swelling, both are alarm pheromone responses of the organism and the colony. NSAIDs affect the biochemical metabolism of arachidonic acid, which closely resembles dozens of sebum’s chemical components.
EXOCRINOLOGY
135

Drug and alcohol addicted women more frequently make poor mothers.77 Addicted mothers tend to neglect and physically abuse their offspring.78,79 A physiological competition for biological receptor sites between drug and pheromone, its metabolites, and/or stimulated opiate releases may explain the phenomenon.
Interestingly, family therapy, rather than friend therapy, has a greater effect upon successful recovery from simultaneous alcoholism and street drug abuse. Patients in therapy living with family members had significant improvement while those living with close friends and acquaintances had none.80 What differs?
Here is an explanation with some appeal. Friends kiss less than families. When less of a skin surface pheromone substitutes for an addictive drug, more of the addictive drug is desired. Human beings need pheromonal love. If we do not get it, we tend to substitute something else.
Willpower may have its place. Perhaps defective reception of the pheromone may simply be willed by the drunken mother because alcohol satisfies or masks her craving for the addictive product of her baby's sebaceous glands?
TESTING
Two protocols, or roadmaps for an experiment, suggest themselves. One would involve use of sebum or vernix caseosa donated by a healthy, loved neonate. The other method would stimulate secretion of much larger quantities of native sebum onto the kissable skin of the abused child. Advantages and disadvantages accrue to each.
The sebum donation experiment may be the first tried. This utilizes a methology similar to that already tried and proven successful in sheep.81 The protocol would have you put vernix caseosa/amniotic fluid onto the kissable skin of the abused child. Then you have the now receptive, abusive parent kiss the child, taking up some of the donated fluid/sebum, and hopefully, loving the youngster for the first time.
136

Getting the abusive parent pheromonally receptive may require closely approximating the conditions at birth. One group, picking up on what shepherds have done for millennia, has stressed vagino-cervical stimulation to encourage ewe acceptance of an orphan lamb.82 These researchers do not seem to have controlled adequately against semiochemical sepsis, however. Because of the potency of pheromones (remember these chemicals can and do work at the ultra-trace level) extreme precautions must be taken. Manipulation of the ewe vagina may induce more adoption for another reason. Enthusiastic researchers may pull out more priming pheromone with their agitating arms, vibrating balloons and such. Males accept and care for offspring without benefit of delivery, so the Kendrick group must be wrong?
Despite these reservations, any increase in semiochemical receptivity would be welcome. Using a vibrator device to stimulate human female genitalia, especially within some period of a successive birth, may have a place here. It may prove useful in recreating biological conditions favorable for initiating a new, fresh parental bond. Genital stimulation just might biologically excite an abusive mother to receptivity for the sebum of her abused child.
Other methods may be utilized to induce an addictive pheromone receptivity. Obtaining the most potent donated semiochemical fraction chromatographically may be tried. Amniotic fluid in both man and sheep has a large sebaceous fraction.40 Synthetic components of human amniotic fluid, vernix caseosa, and other semiochemical candidates should be tried singly and together in various, dosages, concentrations, and administration regimens to achieve best effect. Logically the closest approximation to natural exposures should prove optimal.
Obtaining blood plasma from the natural mother and injecting it into the virgin adoptive mother may work. The technique works well in rats.83 Waves of fetoproteins, found in gravid plasma, may be pheromone receptor proteins that accumulate in the mother and are transferred to the father to enable human parental bonding to neonates. If so, the fractions responsible for bonding receptivity might be isolated and synthesized and used medically to stimulate adoption and the treat
EXOCRINOLOGY
137

physical child abuse and neglect. Similar blood proteins encouraged by hugs84 may also prove therapeutic.
Providing a vernix caseosa donation over kissable skins of the abused child has several advantages over attempting to stimulate new skin surface lipid in the abused child's kissable skins. Applying a sebotrophic agent, such as dehydroepiandrosterone, increases sebum secretion a week or so after topical application.85,40 Waiting may be inconvenient. Furthermore, steroid uptake and resulting sebum secretion rate vary from gland to sebaceous gland,86 so consistency might be a problem. However, the sebaceous stimulation approach, mimicking the naturally gradual decline of sebaceous sebum over a period of months, might be most practical. No gradual shift over of the maternal recognition from the donated to the child's native sebum would need management. Injection of birth associated fetoproteins (likely pheromone receptor proteins), or the natural mother’s hug-stimulated plasma may also stimulate pheromonal receptivity in abusive adoptive parents.
In a large herd of domestic sheep, ewes lamb together simultaneously, facilitating experimentation. In the human case, we might utilize abusive mothers recently giving birth or suffering a stillborn delivery. That would recreate actual bonding physiology and optimize fostering with either method, donated or stimulated pheromone.
Maternal secretions a set period prior to birth stimulate maternal behavior in rats.87,88 Accordingly, we should at least suspect a similar primer pheromone in humans. A pheromone binding protein of the lipid-carrier class would be a prime suspect for pheromone receptor proteins. Emissions might be vaginal, colostrual, locial or mucosal. We must search.
Maternal sebaceous secretion remains high during the third trimester of pregnancy89 as well as during lactation.90 Because these periods correlate to the period of enhanced maternal receptivity, autoemission and autoreception of a sebaceous secretion must also be considered as a possible primer pheromone in
138

humans. Indeed, proteins have been found in sebaceous glands91 and these, too, appear of the pheromone binding type.92
The sebum secretion stimulation method might utilize dehydroepiandrosterone, a fairly benign steriod.93 The effect of topical application has local effect, while going without measured systemic effects. However, because steroids may have potent effects upon development, their use should be carefully monitored.94
Otherwise, is the stimulated sebum of say, a four year old, the same as vernix caseosa, or even neonatal sebum? If the steroid produces an adult-type secretion, then the inadvertent unsavory stimulation of sexual attraction or even mate-mate bonding may be expected. A longitudinal study ascertaining the similarity of stimulated immature sebum to that of the same child's vernix caseosa, and other sebums through development would insure against such surprises.
Pheromone receptivity enhancements, such as pheromone receptor protein injections or tear substitution, must stimulate the right receptivity. We do not want friendships to form. We need no mate-mate bonding between parent and child.
Neither do we seek some extra secretion of a childhood avoidance pheromone. However, it might be sought for use in a similar protocol in the treatment of the sexually abusive father or mother. If a human pheromone drives kibbutzim to seek outside mates, say some anti-romantic priming pheromone, it might find significant use in the treatment of child molesters. Realizing it may be there is the first step. We must find it, isolate it, and synthesize it, if it exists at all. Of course none of this is easy since the expected chemicals are colorless, odorless, tasteless, imperceptible and ephemeral, but we chemists have our ways.
Immunosuppressive agents may affect bonding success. Pheromone and tissue recognition are similar enough and close enough genetically that they may be the same system. Indeed, following the author's suggested protocol to the letter, Cocke and Thiessen95 found that alarm pheromones do reduce immune response. It would be interesting to observe immune system effects in rats exposed to the maternal behavior inducing pheromones of Menella and Moltz.85,86 Testing effects of
EXOCRINOLOGY
139

such agents on pheromone secretion directly may be a useful, needless to say medicinal, line of research. Ingestion of sebum may benefit people suffering from immune deficiencies.
Immune dysfunction itself may be present as a result of drug abuse or other illness. The reverse may also be the case as self-medication.
Sickness in the mother, neonate or family can affect immunity. Likewise illness itself indicates a weakened or vulnerable immune system. Immune pheromone recognition might then also be weakened or vulnerable to failure. Additionally, while mother or newborn lie ill, opportunities for pheromone transmission and uptake are foregone.
Along with illness in either the mother or the newborn and prematurity, prolonged or difficult labor has also been correlated with increased physical child abuse.5 A long and difficult birthing process may be indicative of some illness in mother or newborn. Problems in the bearing of a child may also be caused by drug addiction96:
Drug-dependent women have an increased incidence of medical and obstetrical complications. ... The incidence of premature delivery, abruptio placentae, breech presentation, and intrauterine growth retardation were significantly greater in drug-dependent women. The average duration of the first, second, and third stages of labor compared well with the normal course of labor and matched the results of the comparison group. Labor abnormalities and cesarean sections were of no greater incidence, but there were more than twice as many forceps deliveries, which coincides with the 40 per cent increased use of epidural anesthesia. Analgesia and anesthesia were in excess of that which is given to the average patient (Silver et al., 1987).94
Alcohol and drug addiction, as we have seen, may interfere with or substitute for human pheromone reception. Given this, there should be no surprise that pheromone reception should affect the perception of pain, allowing pheromone manipulations to ease chronic and other pain as analgesics. Oral sebum should be tried.
140

Crowded or difficult living conditions also correlate to suppressed immunity.93 Crowded conditions at home seems to increase the probability of abuse5,97 A pheromone secretion and reception mechanism can account for these. Ultra-crowded fish hacheries probably account for the loss of salmon runs similarly.
A friendly female nearby during delivery eases the birthing process.37 So having no female "helper" close-by during delivery may be an indicator of greater bonding difficulty. Thus pheromone reception of the friendly female would affect pheromone receptivity of the mother for her own offspring. Similar progressions are seen in insects commonly from long-distance reception to short-distance second pheromone reception and behavior.
The child abuser might be dissuaded by a consistent dosage regimen of addictive skin surface pheromone. This idea might lead his limbic system to perceive the presence of extended family. (Absence of an extended family has also been associated with increased risk of abusive behavior.)
Adoption agencies place babies with adopting parents as soon after birth as possible, to increase bonding success. The probability of obtaining a successful bond appears to decline with sebum excretion by the neonate.
If a mother's love is addiction to sebum produced by her infant, then any competition with or interruption of her critical consumption should reduce bonding success. Such a competition might be due to consumption of other addictive substances affecting the same receptor site or sites. Likewise, an interruption might result from defective excretion by the infant or physical separation during the period of high neonatal secretion.
Perhaps together with pre-delivery colostrum and vaginal secretions timely applied or with feto receptor proteins injected, extracts or raw sebaceous secretions from kissable skin surfaces of the abused child might cause an appetite for the abused child’s skin secretions? If so, such skin secretions themselves might be stimulated with topical application of dehydroepiandrosterone and other local sebotrophic stimulants.83
EXOCRINOLOGY
141

Once adopting or abusive parents can reliably be induced to love with extracts, analyses should yield the ingredients of the chemical mixture. With those results the pheromones for love could be manufactured de novo. Double-blinded, crossed over, assays of the artificial chemical mix, repeated and replicated by other laboratories, could prove pheromones prime, then assuage our love appetites. Those would be the same criteria for scientific proof for any pheromone,98 difficult steps rarely taken in practice.
142
1 Stark E, Flitcraft AH Women and children at risk: a feminist perspective on child abuse. Int J Health Serv. 1988;18(1):97-118.
2 Zielinski DS. Child maltreatment and socioeconomic well-being. Child Abuse Negl. 2009 Oct; 33(10):661-665.
3 Eibl-Eibesfeldt, Irenäus. (1970b). Love and Hate: On the Natural History of Behavior Patterns. New York: Methuen and Company.
4 Shields, W.M., J.R. Crook, M.L. Hebblethwaite, and S.S. Wiles-Ehmann. 1988. Ideal free coloniality in the swallows. In, Ecology of Social Behavior, C. N. Slobodchikoff, ed., Academic Press, NY, pp 189-228.5 Lynch, M.A. 1975; Ill health and child abuse. Lancet, 2(7929):317-319.
6 Jason, Janine, Sandra L. Williams, Anthony Burton, Roger Rochat 1982; Epidemiologic differences between sexual and physical child abuse. Journal of the American Medical Association 247(24):3344-3348.
7 Nicholson, B. 1983; A semiochemical addiction model for human bonding. Paper presented to the Animal Behavior Society and the International Society for Human Ethology, Lewisburg, PA.
8 Pochi, Peter E., 1985; Sebum: its nature and physiopathologic responses. pp. 75-78 Section II In: Dermatology Volume 1/Second Edition Moschella, Samuel L. and Harry J Hurley editors. Philadelphia, PA: W.B. Saunders Company.
9 Pochi, Peter E. 1982; Commentary: androgen effects on human sebaceous glands. Archives of Dermatology 118(10):803
10 Pochi, Peter E., and J S. Strauss (1969). Sebaceous gland response in man to the administration of testosterone delta-4-androstenedione, and dehydroisoandrosterone. Journal of Investigative Dermatology 52(1):32-36.
11 Wilson, Edward O. (1963). Pheromones. Scientific American 208:100.

EXOCRINOLOGY
143
12 Shorey, H.H. 1976; Animal Communication by Pheromones. New York: Academic Press.
13 Fillion, TJ, Blass EM. Infantile experience with suckling odors determines adult sexual behavior in male rats. Science 1986;231(4739):729-731.
14 Anderson P. The reproductive role of the human breast. Current Anthropology 1983;24(1):25-45.
15 Crowell-Davis, S.L. and K.A. Houpt Coprophagy by foals: effect of age and possible functions. Equine Veterinary Journal 1985;17(1):17-19.
16 Kilpatrick, S.J, T.M. Lee, and H. Moltz 1983; The maternal pheromone of the rat: testing some assumptions underlying a hypothesis. Physiology & Behavior 30(4):539-543.
17 Lee, T.M., and H. Moltz (1984a). The maternal pheromone and brain development in the preweanling rat. Physiology & Behavior 33(3):385-390.
18 Lee, T.M., and H. Moltz (1984b). The maternal pheromone and deoxycholic acid in relation to brain myelin in the preweanling rat. Physiology & Behavior 33(3):391-395.
19 Schumacher, S.K., and H. Moltz 1985; Prolonged responsiveness to the maternal pheromone in the postweanling rat. Physiology & Behavior 34(3):471-473.
20 Pietras, RJ 1981; Sex pheromone production by the preputial gland. Chemical Senses 6(4):391.
21 Stewart, Mary Ellen 1992; Sebaceous gland lipids. Seminars in Dermatology 11(2):100-105.
22 Stewart, Mary Ellen, and Donald T. Downing 1991; Unusual cholesterol esters in the sebum of young children. Journal of Investigative Dermatology 95(5):603-606.
23 Porter, RH., F. Le'vy, P. Poindron, M. Litterio, B. Schaal and C. Beyer 1991; Individual olfactory signatures as major determinants of early maternal discrimination in sheep. Developmental Psychobiology. 24(3):151-158.
24 Hold-Cavell, Barbara, and Margret Schleidt 1981; A Cross-Cultural Study on the Attitude towards Personal Odors in the Journal of Chemical Ecolology 7(1):1
25 Porter, Richard H., Rene D. Balogh, Jennifer M. Cernoch and Christie Franchi 1986; Recognition of kin through characteristic body odors. Chemical Senses 11:389-395.
26 Wray, Herbert 1982; The Evolution of Child Abuse. Science News 122:24.
27 Robson, K.M., and R Kumar 1980; Delayed onset of maternal affection after childbirth. British Journal of Psychiatry 136:347-353.
28 Klaus, M.H., and JH. Kennell 1982; Parent-Infant Bonding. St. Louis: The C.V. Mosby Company.
29 Agache, P., D. Blanc, C. Barrand, and R Laurent R 1980; Sebum levels during the first year of life. British Journal of Dermatology. 103:643-649.
30 Coppen A, Stein G, Wood K. Postnatal depression and tryptophan metabolism. In: Sandler, M. editor. Mental illness in pregnancy and the puerperium. Oxford: Oxford University Press, 1978.

144
31 Nicholson B., Pheromones cause disease: pheromone/odourant transduction. Med Hypotheses. 2001 Sep;57(3):361-77.
32 Brazelton TB, Tronick E, Adamson L, Als H, Wise S. Early mother-infant reciprocity. Ciba Found Symp. 1975;(33):137-154.
33 Cady E, Cady F. How to adopt a child. New York: Whiteside, Inc. and William and Morrow Company, 1956.
34 Pochi, Peter E. 1982; Commentary: androgen effects on human sebaceous glands. Archives of Dermatology 118(10):803.
35 Shuster, Sam, and Anthony J Thody 1974; The control and measurement of sebum secretion. Journal of Investigative Dermatology 62(3):172- 190.
36 Anisfeld E, Curry MA, Hales DJ, Kennell JH, Klaus MH, Lipper E, et al. Maternal-infant bonding: a joint rebuttal. Pediatrics 1983;72(4):569-572.
37 Klaus, Marshall H., and J H. Kennell 1983; Parent to infant bonding: Setting the record straight. The Journal of Pediatrics (St. Louis) 102(4):575-576
38 Holbrook, Karen A. 1986; A Histological Comparison of Infant and Adult Skin pp. 3-31 In: Neonatal Skin Structure and Function. Maibach, Howard, and Edward K. Boisits editors. New York: Marcel Dekker, Inc.
39 Wysocki, S.J, A. Grauaug, G. O'Neill, and R Hähnel 1981; Lipids in forehead vernix from newborn infants. Biol. Neonate. 39:300-304.
40 Montagna, William, and Paul F. Parakkal 1974; The Structure and Function of Skin. New York: Academic Press. 433 p.
41 Nicolaides, N. 1974; Skin lipids: their biochemical uniqueness. Science. 186:19-26.
42 Amoore JE. Four primary odour modalities of man: experimental evidence and possible significance. In: Denton DA, Coughlan P, editors. Olfaction and taste, V. New York: Academic Press, 1974; 283-9.
43 Comfort A. 1974; The likelihood of human pheromones. In: Pheromones. Birch, M.C. editor. Amsterdam: North-Holland Publishing Company.
44 Pochi, Peter E., J S. Strauss, and Donald T. Downing 1979; Age-related changes in sebaceous gland activity. Journal of Investigative Dermatology 73:108-111.
45 Silverstein, Robert M. 1984; Chemistry of insect communication. Chapter 5, pp 105-121. In: Insect Communication. Lewis, Trevor editor. Academic Press, London. 414 p.
46 Bell M. A comparative study of the ultrastructure of the sebaceous glands of man and other primates. Journal of Investigative Dermatology 1974;62(3):132-143.
47 Shuster, Sam, and Anthony J Thody 1974; The control and measurement of sebum secretion. Journal of Investigative Dermatology 62(3):172- 190.

EXOCRINOLOGY
145
48 Simpson, N.B., et al. 1983; A more reliable photometric technique for the measurement of scalp sebum excretion. British Journal of Dermatology 109(6):647-652.
49 Lookingbill, D.P., and W.J. Cunliffe 1986; A direct gravimetric technique for measuring sebum excretion rate. British Journal of Dermatology. 114:75-81.
50 Leveque, JL., C. Pierard-Franchimont, J de Rigal, D. Saint- Leger and G.E. Pierard 1991; Effect of topical corticosteroids on human sebum production assessed by two different methods. Archives of Dermatological Research 283(6):372-376.
51 Serup, J 1991; Formation of oiliness and sebum output--comparison of a lipid-absorbant and occlusive-tape method with photometry. Clinical & Experimental Dermatology 16(4):258-263.
52 Frantz RA, Kinney CK. Variables associated with skin dryness in the elderly. Nursing Research 1986; 35(2):98-100.
53 Downing DT, Stewart ME, Wertz PW, Colton SW, Abraham W, Strauss JS. Skin lipids: an update. Journal of Investigative Dermatology 1987; 88(3 Suppl):2s-6s.
54 Jason, Janine, Sandra L. Williams, Anthony Burton, Roger Rochat 1982; Epidemiologic differences between sexual and physical child abuse. Journal of the American Medical Association 247(24):3344-3348.
55 Fleming AS, Luebke C. Timidity prevents the virgin female rat from being a good mother: emotionality differences between nulliparous and parturient females. Physiol Behav. 1981 Nov; 27(5):863-868.
56 Fleming AS, Rosenblatt JS. Olfactory regulation of maternal behavior in rats I&II. J Comp Physiol Psychol. 1974 Feb; 86(2):221-246.
57 Leon, Michael 1983; Chemical Communication in Mother-Young Interactions. pp. 39-77. Chapter 2. In: Pheromones and Reproduction in Mammals. J G. Vandenbergh, editor. New York; Academic Press, Inc. 298 p.
58 Nicholson, B. 1985; An economic model of phermone transmission in avians. Paper presented to the Animal Behavior Society annual meeting, Raliegh, N.C.
59 Lucas, A.M. 1980; Lipoid secretion of the body epidermis in avian skin. In: The Skin of Vertebrates. Spearman RI.C. and P.A. Riley . editors. New York: Academic Press.
60 Hou, H.C. (1928). Studies on the glandula uropygialis of birds. Chinese Journal of Physiology. 2:345-380.
61 Hou, H.C. (1929). Relation of the preen gland (glandula uropygialis) of birds to rickets. Chinese Journal of Physiology. 3:171-182.
62 Balthazart, J, and E. Schoffeniels pheromonal action of preen gland secretion in ducks. Naturwiss. 1979;66:55.
63 Lorenz, Konrad (1963). On Agression. Translated by Marjorie Kerr Wilson (1966). New York: Bantam Books Inc. 306 p.

146
64 Yager, JA, D.B. Hunter, M.R Willson, and O.B. Allen 1988; A source of cutaneous maternal semiochemicals in the mink? Experientia 44(1):79-81.
65 Mykytowycz, R (1970). The role of skin glands in mammalian communication. In: Advances in Chemoreception, Vol. I. Johnston, JW., D.G. Moulton, and A. Turk editors. New York: Appleton-Century-Crofts.
66 Benuck I, Rowe FA. Centrally and peripherally induced anosmia: influences on maternal behavior in lactating female rats. Physiol Behav. 1975 Apr;14(04):439-447.
67 Leon, Michael 1983; Chemical Communication in Mother-Young Interactions. pp. 39-77. Chapter 2. In: Pheromones and Reproduction in Mammals. J G. Vandenbergh, editor. New York; Academic Press, Inc. 298 p.
68 Devor M. Central processing of odor signals: lessions from adult and neonatal olfactory tract lesions. In: Müller-Schwarze D. Mozell MM, editors. Chemical Signals in Vertebrates (I) 1977;529-45.
69 Shepard, Gordon M. 1977; Central processing of olfactory signals. pp.489-497 In (Mu^H ller-Schwarze, D and Maxwell M. Mozell, eds.) New York: Plenum Press 609 pages page 490.
70 Benedict C, Hallschmid M, Hatke A, Schultes B, Fehm HL, Born J, Kern W. Intranasal insulin improves memory in humans. Psychoneuroendocrinology. 2004 Nov;29(10):1326-34.
71 Doty RL. Modulation of the odour detection performance of male rats by castration and the administration of dopaminergic active drugs: a summary of recent findings. In: Macdonald DW, Müller-schwarze D, and Natynczuk SE, editors. Chemical signals in vertebrates 5 Oxford: Oxford University Press, 1990;100-108.
72 Rehnberg, Bradley G, Emily H. Bates, R Jan F. Smith, B. Duff Sloley, and J Steven Rson. 1989; Brain benzodiazepine receptors in fathead minnows and the behavioral response to alarm pheromone. Pharmacology Biochemistry & Behavior, 33:435-442.
73 Rehnberg, Bradley G, and RJF. Smith 1990; Behavioural and physiological responses to alarm pheromone by ostariophsan fishes and a possible role for brain benzodiazepine receptors. In: Chapter II.5 In: Chemical Signals in Vertebrates 5. D.W., Dietland Müller-Schwarze, and Steven E. Natynczuk editors. Oxford: Oxford Science Press. 659 p.
74 Harlow, Harry F., and Zimmerman, RR (1956) Affectional responses in infant monkeys. Science, 130:421-432.
75 Harlow, Harry F. (1958). The nature of love . Amer Psychol. 13:673-685.
76 Hollenbeck, A.R, JL. Gewirtz, S.L. Sebris, and JW. Scanlon 1984; Labor and delivery medication influences parent-infant interaction in the first post-partum month. Infant Behavior and Development 7:201-209.
77 Cuskey WR, Wathey RB. Female addiction. Lexington: D.C. Heath and Company, 1982.
78 Silver, H., R Wapner, M. Vega, and L.P. Finnegan 1987; Addiction in pregnancy: high risk intrapartum management and outcome. J Perinatol 7(3):178-184.

EXOCRINOLOGY
147
79 Balal, A.M. Correlates of addiction-related problems in Kuwait: a cross-cultural view. Acta Psychiatr Scand. 1988;789(4):414-416.
80 Crowley P. Family therapy approach to addiction. Bulletin on Narcotics 1988;40(1):57-62.
81 Levy, F., E.B. Kevern, V. Piketty and P. Poindron 1990; Physiological determinism of olfactory attraction for amniotic fluids in sheep. pp. 162-165, Chapter II.9 In: Chemical Signals in Vertebrates V. MacDonald, D.W., Dietland Müller-schwarze, and S.E. Natynczuk editors. Oxford: Oxford Science Publications 649p.
82 Kendrick, K.M., F. Le'vy, and E.B. Keverne 1991; Importance of vaginocervical stimulation for the formation of maternal bonding in primiparous and multiparous parturient ewes. Physiology & Behavior 50(3):595-600.
83 Terkel J, Rosenblatt JS. Maternal behavior induced by maternal blood plasma injected into virgin rats. J Comp Physiol Psychol. 1968 Jun;65(3):479-482.
84 Matsunaga M, Sato S, Isowa T, Tsuboi H, Konagaya T, Kaneko H, Ohira H. 2009; Profiling of serum proteins influenced by warm partner contact in healthy couples. Neuro Endocrinol Lett. 2009;30(2):227-236.
85 Pochi, Peter E., and J S. Strauss (1969). Sebaceous gland response in man to the administration of testosterone delta-4-androstenedione, and dehydroisoandrosterone. Journal of Investigative Dermatology 52(1):32-36.
86 Sansone-Bazzano, Gail, and Ronald M. Reisner 1974; Steroid pathways in sebaceous glands. Journal of Investigative Dermatology 62(3):211-216.
87 Mennella, Julie A, and Howard Moltz 1988; Infanticide in rats: male strategy and female counter-strategy. Physiology & Behavior 42:19-28.
88 Mennella, Julie A, and Howard Moltz 1989; Pheromonal emission by pregnant rats protects against infanticide by nulliparous conspecifics. Physiology and Behavior 46:591-595.
89 Burton JL, Shuster S, Cartlidge M. The sebotrophic effect of pregnancy. Acta Dermato-vernereologica (Stockholm). 1975;55:11-3.
90 Burton, JL., Sam Shuster, M. Cartlidge, L.J Libman, and U. Martell (1973a). Lactation, sebum excretion, and melanocyte stimulating hormone. Nature 243:349-350.
91 Mancini, M.A, D. Majumdar, B. Chatterjee and A.K. Roy 1989; Alpha 2u-globulin in modified sebaceous glands with pheromonal functions: localization of the protein and its mRNA in preputial, meibomian, and perianal glands. J Histochem Cytochem Feb 37(2):149-157.
92 Singer, A.G. 1991; A chemistry of mammalian pheromones. Journal of Steroid Biochemistry & Molecular Biology. 39(4B):627-632.
93 Downing DT, Strauss JS, Ramasastry P, Abel M, Lees CW, Pochi PE. Measurement of time between synthesis and surface excretion of sebaceous lipids in sheep and man. Journal of Investigative Dermatology 1975;46:444.
94 Pochi, Peter E. 1982; Commentary: androgen effects on human sebaceous glands. Archives of Dermatology 118(10):803.

148
95 Cocke R, Thiessen D. Alarm chemosignals suppress the immune system. In: Chemical sgnals in vertebrates. MacDonald, D.W., D.Mu eller-Schwarze D, and S.E. Natynczuky editors. Oxford: Oxford University Press. 1990; 125-138
96 Silver, H., R Wapner, M. Vega, and L.P. Finnegan 1987; Addiction in pregnancy: high risk intrapartum management and outcome. J Perinatol 7(3):178-184.
97 Jason, Janine, Sandra L. Williams, Anthony Burton, Roger Rochat 1982; Epidemiologic differences between sexual and physical child abuse. Journal of the American Medical Association 247(24):3344-3348.
98 Silverstein, Robert M. 1984; Chemistry of insect communication. Chapter 5, pp 105-121. In: Insect Communication. Lewis, Trevor editor. Academic Press, London. 414 p.

Chapter 5: Eating Disorders
This chapter deals with the exocrinology of anorexia nervosa (AN) and speculates for cures for anorexia nervosa, bulimia nervosa, and obesity.
Diet, psychotherapy, and drugs have sometimes alleviated symptoms of AN patients but the root cause and the best treatment have eluded a century of research.1
The symptoms of Anorexia Nervosa (AN) in human beings are largely the symptoms of pheromone-induced puberty delay in many species of animals. AN maintains mature scent glands in its victims. The axillary and pubic hair tufts are secondary sex characteristics that are associated with passing the age of puberty in humans. Those mature sexual hairy areas cover and are part of scent glands which are among the largest in nature despite the presence of a pre-puberal endocrine set in anorexic sufferers. Indeed, axillary and pubic scent glands appear to develop and function normally in AN sufferers.2,3,4 Pheromone-induced puberty delay (PIDP) seen in animals also maintains mature, functioning scent glands5 in the presence of a pre-pubertal endocrine set6 just like AN! Surely that alone must be more convincing than psychological speculations.
Both AN and PIPD occur at the same time in the life cycle, when puberty is first attained, under similarly stressful conditions. Immatures without secondary sex characteristics lack the ability to develop AN and PIPD, as well.
Humans might seem to differ in that we have another incidence peak at entry into college. However, if little lab animals went to college, they would have an entry into college peak, too. Crowded dormitory grouping of puberal females readily stimulates pheromone induced puberty delay in animals.5 An
EXOCRINOLOGY
149

effective puberty delay obviously named the pheromone set in animals. In AN, poor Fischer finally noted a developmental lag after eleven years of psychoanalysis.7
The sex ratios are nearly identical, too. Both AN and PIPD have far more female sufferers than males. The ratios of female to male runs 50:1 in PIPD.8,9 The ratio of female to male human sufferers in AN is between 10:1 and 20:1.10
Both AN and PIPD share having a stressed fertile female nearby. In the animals, the stressed and dominant fertile female emits a puberty delay pheromone that sends the puberal female back into pre-puberty.5 Of course, the dominant fertile female can be the mother of the PIPD victim. Psychologist report AN onset associated with a mother/daughter or female teacher/female student ‘dyad’11 so the mature female need not be a family member.
In both PIPD and in AN, the condition continues after the victim is removed from the shared atmosphere with the stressed fertile female.7 In PIPD the glands of the victim’s intact adult scent glands emit the same pheromone chemical signal as the original stressed female’s scent glands emitted to begin her puberty delay.5 In AN, remember the intact large scent glands? In AN, the girl taken away from her mother continues with anorexia nervosa,12 just like PIPD.
Both AN in humans and PIPD in animals occur to daughters of gentry. Both AN and PIPD sufferers are raised with food available ad libitum. Having plenty of food available is a necessary condition for PIPD and exactly the same phenomenon is observed in AN. Food is always available to anorexics,15 just as it a necessary condition for PIPD to develop.5
Both AN and PIPD result in periodic peregrination. AN sufferers have altered metabolisms and take long walks.13 Pheromonally induced puberty
150

delay animals also have altered metabolisms and take long walks--just the PIPD animals walk on the cage exercise wheels.5
The pheromone induced puberty delay in laboratory animals is typically about 30 days or so while the human anorexia nervosa can last for decades. However, the period of puberty delay as a percentage of lifespan is about the same.
Both PIPD and AN see victim avoidance of sexually mature conspecific males. PIPD animals avoid males and even the odor of males.8 AN sufferers generally avoid dating men.12 In animals where mature males are unavoidable or welcome, ‘[t]he presence of a conspecific male was effective in maintaining normal oestrous cycles in underfed females to some extent’14 which suggested to Drickamer (1988) that acceleration pheromones and delay pheromones worked in opposition. Indeed, here is what Drickamer found:
At ratios of 10-20 parts acceleratory chemosignal to 1 part delay chemosignal, the acceleratory effect overrides the delay chemosignal, and the mice attain first oestrus at earlier ages than controls. Ratios of about 4 to 1 up to 7 to 1 result in mean ages for puberty that are not accelerated or delayed relative to controls.5
The return of female reproductive cyclicity marks recovery in both the human AN and the animal PIPD. When menstrual cyclicity does appear, the prognosis is excellent for both AN15 and in PIPD the delay is merely over.
Given that return of cyclicity bodes well for recovery from anorexia nervosa and that ‘having a boyfriend seems to help’ anorexic girls,12 a human axillary pheromone extract which encourages regular menstruation in women16 was prepared. Unfortunately it was never tried and the opportunity was wasted. The author has seen only one complete recovery from AN after providing only one delinquent young woman with male skin surface
EXOCRINOLOGY
151

pheromone from the adult male face, but that may have been happenstance. AN is so rare that psychologists hoard them away from exocrinologists. Publication of an article in a peer-reviewed medical journal by this author describing the likely exocrine origin of anorexia nervosa17 was ignored and even ridiculed.17
Smelling the axillary pheromone component, androstadienone, increases cortisol levels in women.18
This book may open new avenues of scientific cooperation and discovery. However this may be difficult to accomplish for a number of reasons. First, human pheromones usually come in sets that are nearly undetectable by human senses. This insidious nature of pheromones can be disconcerting, and the detection difficulty is exacerbated by unusually long detection times on the gas chromatograph, with many components taking longer than 30 minutes. Second, researchers are human, so working with human pheromones can be dangerous. Working on this research can be destructive to ones professional career, too, since hierarchical position colors pheromone perception in the Pharisaic effect.
Evidence recently has been found to support this authors contention of a human pheromone which causes anorexia nervosa syndrome19 when a pheromone was shown in the laboratory to cause loss of appetite, or anorexia, in male Syrian hamsters. Morgan et al.20 found that anorexia by pheromone can be observed in Syrian hamsters, three years following publication of the article written by this author describing pheromonal anorexia in humans (Nicholson B, 2000). Morgan et al. labeled pheromonal anorexia a ‘novel phenomenon’ and suggested it would ‘provide a model to study neural and endocrine mechanisms that underlie eating disorders.’ The flank gland of the female, the same gland associated with puberty delay emissions in that species, emits the anorexia causing pheromone.
From Chapter 3, recall that different species mean different pheromones. Human anorexia-causing pheromones are most likely from axillary and pubic scent
152

glands. Hamsters use flank glands with a sebaceous fraction and mouse pheromones are urinary. And oddly enough, axillary scent glands may perpetuate obesity as well. This author found he could control his weight if his axillae were kept covered with a thick layer of petroleum jelly.
Bulimia nervosa is a related eating disorder of bingeing and purging, with normal weight maintained. In bulimia, appetitive fellatio obtains pubic and penile pheromones to self-medicate the disorder.
The cure for all eating disorders should be 150 mg p.o. of healthy ‘father age’ adult male facial skin surface lipid kissing pheromone in a single dose. The cure might effected by the anorexic/bulimic/obese person chewing 15 pieces of regular Wrigley’s chewing gum rubbed on the face of the person’s father over a three day period utilizing good laboratory practices and good sanitation. Covering the axillae with petroleum jelly may help effect immediate change as the author discovered by personal experience.
Fat people see themselves in photographs and rationalize that they are unphotogenic, because photographs appear to exaggerate their size. After two days of walking around uncomfortably with his armpits’ axillary scent glands covered with petroleum jelly, the obese author’s mirror image changed to show his true size. The abrupt self-perception change was imperceptibly insidious. Plates of food only days before observed to be small became too much to eat at one sitting, perception of size in the horizontal direction changed. Dramatic weight loss followed the perception change, but once the interruption of the axiallary chemosignals ceased, the weight was imperceptibly regained.
Since the axillary chemosignal set depends upon the skin’s microflora for processing, 21 antiseptic interventions were tried, but failed. Some weight loss was finally maintained by a rigorous regimen of long distance hiking and bicycle exercise. Others22 have begun similar investigations with some hope of progress.
A derivative of fish oils has helped one severely anorexic patient.23 Given the
similar tertiary stereochemistry of fish oils to components of human skin surface
EXOCRINOLOGY
153

lipid, treatment of AN with the far more complex synergistic facial skin surface lipids
of the human adult male should be of benefit. Injection of pheromone binding
proteins and epigenetic chemotherapy may also find usefulness treating anorexia
nervosa.
Of course, bulimia nervosa sufferers famously find solace in fellatio. At the
very least, male sexual skin surface lipids collected in the lab or synthesized de novo
should sate that appetite without most of the risk of sexually transmitted disease.
154
1 Murray J B Psychological aspects of anorexia nervosa. Genet Soc Gen Psychol Monogr 1986; 112(1): 5-40.
2 Treasure JL, Gordon PA, King EA, Wheeler M, Russell GF. Cystic ovaries: a phase of anorexia nervosa. Lancet 1985; 2(8469-70): 1379-1382.
3 Treasure JL, Wheeler M, King EA, Gordon PA, Russell GF. Weight gain and reproductive function: ultrasonographic and endocrine features in anorexia nervosa. Clin Endocrinol (Oxf) 1988; 29(6): 607-616.
4 Robbins SL, Cotran RS, Kumar V. Robbins pathologic basis of disease. Philadelphia: W.B. Saunders, 1986.
5 Drickamer LC. Acceleration and delay of sexual maturation in female house mice (Mus domesticus) by urinary chemosignals: mixing urine sources in unequal proportions. J Comp Psychol 1988; 102(3): 215-221.
6 Keverne EB. Pheromonal influences on the endocrine regulation of reproduction. Trends Neurosci 1983; 6: 381-384.
7 Fischer N. Anorexia nervosa and unresolved rapprochement conflicts: a case study. Int J Psychoanal 1989; 70(Pt1): 41-54.
8 Vandenbergh JG Pheromonal Regulation of Puberty. In: Vandenbergh JG, editor. Pheromones and reproduction in mammals. New York: Academic Press, 1983: 95-112.
9 Drickamer LC Puberty-influencing chemosignals in house mice: ecological and evolutionary considerations. In: Duvall D, Müller-Schwarze D, Silverstein RM, editors. Chemical signals in vertebrates 4. New York: Plenum Press, 1986: 441-456.
10 Dambro MR, Griffth JA Griffith’s 5 minute clinical consult. Baltimore: Williams & Wilkins 1997: 60.

EXOCRINOLOGY
155
11 Ehrensing RH, Weitzman EI The mother-daughter relationship in anorexia nervosa. Psychosom Med 1970; 32(2): 4201-4211.
12 Dally P, Gomez J, Isaacs AJ Anorexia Nervosa. London: William Heinemann Medical Books Ltd. 1979.
13 Russell GFM Metabolic aspects of anorexia nervosa. Proc R Soc Med 1965; 58: 811-814.
14 Gangrade BK, Dominic CJ Influence of conspecific males on the oestrous cycle of underfed female mice. Exp Clin Endocrinol 1985; 86(1): 35-40.
15 Halmi KA Anorexia nervosa: recent investigations. Annu Rev Med 1978; 29: 137-148.
16 Cutler WB, Preti G, Krieger A, Huggins GR, Garcia CR, Lawley HJ Human axillary secretions influence women’s menstrual cycles: the role of donor extract from men. Horm Behav 1986; 20: 463-473.
17 Gatward N. Because it feels good (smells funny!) European Eating Disorders Review 2001; 9(3): 210-212(3).
18 Wyart C, Webster WW, Chen JH, Wilson SR, McClary A, Khan RM, Sobel N. Smelling a single component of male sweat alters levels of cortisol in women. J Neurosci. 2007 Feb 7; 27(6):1261-1265.
19 Nicholson B Pheromones cause disease: the exocrinology of anorexia nervosa Med Hypotheses 2000; 54(3): 438-443.
20 Morgan C, Urbanski HF, Fan W, Akil H, and Cone RD. Pheromone-induced anorexia in male Syrian hamsters. Am J Physiol Endocrinol Metab 2003; 285: E1028-E1038.
21 Albone, Eric S., Pauline E. Gosden, and Georges C. Ware 1977; Bacteria as a source of chemical signals in mammals. pp. 35-43 In: Chemical Signals in Vertebrates (Müller-Schwarze, D, and Maxwell M. Mozell eds.) New York: Plenum Press 610 p.
22 McKenzie VJ, Bowers RM, Fierer N, Knight R, Lauber CL. Co-habiting amphibian species harbor unique skin bacterial communities in wild populations. ISME J. 2011 Sep 29. doi: 10.1038/ismej.2011.129. [Epub ahead of print]
23 Ayton AK, Azaz A, Horrobin DF. Rapid improvement of severe anorexia nervosa during treatment with ethyl-eicosapentaenoate and micronutrients. Eur Psychiatry. 2004 Aug;19(5):317-9.


Chapter 6: Crying Chemoreception and Love
Erbkoordinated lacrimation, or emotional crying, is ubiquitous throughout the cultures of man.1 Why?
This chapter presents a pheromone reception model for emotional tears or lacrimation.2,3 Bond breaking, bond establishment, and bond stress bring tears to our eyes. If pheromone addiction is responsible for human bonding, a flush mechanism would enhance flexibility of the process. It would allow old bonds to expire and new ones to form under appropriate circumstances. Crying is released under just such biologically appropriate conditions. Indeed, recent work has found tears themselves to be human pheromones.
EXPECTATIONS OF HUMAN LACRIMAL PHEROMONE
RECOGNITION
What might one reasonably expect of any pheromone recognition system? Any acceptable explanation should explain diverse human behaviors associated with pheromone reception situations. Human actions during a pheromone reception should be directly comparable to "ritualized" or “displacement” (actually chemoreception) behaviors of other animals. Pheromone reception behavior should remain similar for all pheromone receptive species from yeast to human beings.
The courtship dances of birds, involving much swaying and ducking of the head, serve to get tears full of dissolved pheromone receptor protein and special pheromone reception ions into position for chemo-analysis. It will be found, prediction here, that the plumbing of the pheromone reception system determines each movement of the courtship dance, each sway of the displacement activity, every twitch of the erbkoordination. Flight restrains against carrying pheromone emission apparatus out of season, so pheromone emission glands atrophy away from the nest. The same should be found for pheromone reception apparatus in flighted birds.
EXOCRINOLOGY
157

When we humans cry, lean forward, and cover our faces with our hands, we are getting the angle of our naso-lacrimal ducts and our hiatus semilunaris pheromone reception structures in proper alignment, and in proper sequence, for human pheromone reception. Even the male behavior of lifting her lovely chin, kissing her, and holding her in your arms (her head in your armpit), must be economically justified, likely as assuring transmission of episodically released human pheromone.
In human sexual behavior, the engorged turgid erectile tissues of the nasal passages are evacuated at the very instant of climax. Our ears pop and our eyes water. Anguished facial expressions at the joyful event of intromission must also be economically necessary for timely human pheromone perception, just as pheromonal emissions take place. Economics rule instinctive human behavior and nothing is for nothing, TANSTAAFL. (There ain’t no such thing as a free lunch.) This axiom should be assumed for all human behavior, and for all cells of all organs of all organ systems.
Economically, pheromone reception similarity should extend from gross behavior even to cells and cellular events. Nerve cells for humans look like nerve cells for other mammals, reptiles, and birds. Likewise cells carrying oxygen in the blood, look alike. Thus cells specialized for pheromone reception in other animals should appear in humans, too, and they should look and work the same. Cellular activity should proceed from pheromone exposures similarly in all species, from bacterium to butterfly to human.
Pheromone recognition in people should function to conserve biological resources. Receptivity should be more when needed more and less when needed less. Such flexibility should be observable in other animal species. Similar glands, ducts, secretions, and behaviors should typify responses among all animals. Chemo-recognition's dissimilarities among species also must be accounted for physiologically.
Lacrimal glands secrete special ion and protein concentrations in emotional tears4 which may be involved in some human pheromone recognition.2,3 The
158

secretion of these glands takes place within the choreographed sequence of behaviors we call "crying."
Crying identifies a special behavior. Within it ionic concentrations vary (as do peculiar lipids (self-affecting pheromones?), antimicrobial enzymes, and other proteins specific only to emotional tears in the nose and mouth) in response to emotion.5 These concentrations of ions and proteins are much smaller in irritant induced tears.4,6 Onion fumes irritate. Irritant-induced tears are dilute tears which help flush irritating particles out of the eyes and nose.
LACRIMATION AND MATE-MATE LOVE
Desmond Morris7 observed an increase in tear production during mate-mate bonding. He thought it responsible for "the shining eyes of love" and considered it merely a visual signal. The increased lacrimal secretion might be expected during episodes when humans fall in love pheromonally. "These are the shining eyes of love, and combine with pupil dilation to leave no doubt about the condition of the signaler." However, visual signals may be less important in the semidarkness romantic couples prefer. Illegal drugs and belladonna can produce pupil dilation, too, and drug-takers are generally less ready to form long-lasting bonds than more innocent people.
Typical human romantic scenarios may involve dim lighting. Dim light allows easier pupil dilation. Pupil dilation expands the visual fields of the eyes to include more of the phylogenetically old rod photoreceptors. The retina eventually maps onto the brain's occipital lobe, but before reaching it, parallel nerve fibers course in broad arcs through the two temporal lobes. Expanding the visual fields expands the lines of transmission through the temporal lobes where face recognition takes place. In romantic pupil dilation, face and pheromone reception are being associated. Human male axillary pheromone emissions stimulate cortisol in women close enough to take the pheromone from the air or by contact,8 and cortisol
EXOCRINOLOGY
159

cooperates with adrenaline to create vivid memories. So 4,16-adrostadien-3-one stimulates women to remember us males.
Ever notice how kissing lovers close their eyes? Sack et al. (1992)9 noticed increased tear proteins in closed eye tears versus open eye tears. For kissers on the other hand, the view at 10 centimeters lacks something. Alternatively, perhaps nocturnal semiochemical reception and auto-reception plays some role?
Typically, the scenario plays as follows. In the dim light of romantic acceptance, young faces close together. You start telling her how much you love her, about your hopes and dreams. Eyes grow misty. Her eyes are like limpid pools. You kiss. Your lacrimal fluid's proteins, accompanying a peculiarly high manganese cation concentration from specialized glands above your eyes, go to work recognizing her sebaceous lipids. Those lipids are now, due to her excitation, oozing hotly from mucosal glands only recently matured out of nothing.10 You easily reach and scrape out of her mouth's sebaceous glands those nice waxy goodnesses with your tongue. Repeatedly you hunt her wax esters down with your tongue's specialized lipid harvesting surface, also developing just in time11 as your limbic reward. Your own odd-looking sebaceous glands specific to the oral, gingival, and buccal mucosa also come to heat. Out flows your interesting, chemically unique, possibly addictive sebaceous greases. The sebum floats away high on her saliva, just as hers does on yours. The greases you've just licked from her mouth reach the nasal passages where lipid triglycerides have been found,12 and/or they reach the upper-respiratory system where sensory microvillar ‘brush border’ cells line fifty percent of the surface area, and/or they fall into the alimentary canal for digestion and possibly perception by Peyer’s patches again, full of brush-border pheromone sensory cells. The microvillar cells of the human vomeronasal organ may sense something,13 but since we do not carry tails to tell us where we have been and need to go today to recover resources from our territories, the vast fields of microvillar cells visible under surface electron microscopy bunched at the end of the naso-lacrimal ducts in the inferior meati and elsewhere lining the entire upper respiratory system and in patches along the gut serve to recognize the gathered human lipid pheromone.
160

Then there are those tears all over her cheeks. Kissing those tears away may encourage a sharing of pheromone receptivity. Placing one's tears into the saliva of another, with appropriate pheromone accompaniment, would, if recognized, encourage bonding, and perhaps some shared immune resistance as well. Female tears diminish masculine testosterone, slowing the males‘ sex drive. Lacrimal episodes are usually of short duration, and of small consequence to victorious warriors with new widows of slain enemies.
Kissing with lacrimal receptivity might be most efficacious in bond formation. Tears may be elicited by other than romantic situations. This would bring the lacrimal reception pheromone addiction hypothesis into surprising agreement with Finck14:
Love can only be excited by strong and vivid emotion, and it is almost immaterial whether these emotions are agreeable or disagreeable. The Cid wooed the proud heart of Diana Ximene, whose father he had slain, by shooting one after another of her pet pigeons (Finck, H.T., 1891).
Would this help explain why some women remain with abusive husbands?
Another tearful human behavior has been noted by grooms of innocent brides. The behavior has been called a brides "wedding night hysterics." Apparently such unreasonable sobbing by the bride on the marriage bed may encourage pheromonal fealty to her new husband. The newly married woman cries, sometimes all night long, at the prospect and conclusion of first intercourse. Why defloration has been accompanied by "seemingly unmotivated tears" has baffled billions of grooms. Other signs of emotionality of the intact bride remain mysterious. One down, one million to go.
Women's tears and bonding function differ from men's. Women bond somewhat more suddenly. Perhaps on her wedding night an unbonded female bonds herself firmly to the unfamiliar nocturnal exocrine secretions of her new husband? Experiencing husbandly pheromones for the first time would be a logical occasion for dramatic efforts of pheromonal receptivity. Perhaps her defloration lacrimation
EXOCRINOLOGY
161

functions to begin receptivity to the new signals of her husband and to reduce the importance of old family signals. Loyalties may adjust pheromonally in the marriage bed just as at the graveside.
More evidence of sex differences in bonding exist. Sexual dimorphisms, physical differences between the sexes, have been detected in agreement with this idea. Women have a greater frequency15 ionic concentration16 and proteinaceous makeup of adult human tears.17 Men counter with a greater output of dermal sebaceous secretions.9
There were significant differences in the amounts of LF [LactoFerrin] and two [of five] kinds of STPs [Specific Tear Proteins] in the different sexes. The amounts of these proteins were larger in females (Mii S., et al., 1992).16
The Mii16 team's specific tear proteins appear suspiciously similar to some pheromone binding proteins which can act as primer pheromones. The Mii group took non-emotion-produced samples from the conjunctival sacs, a part of the naso-lacrimal duct. However, our interest is emotional tears. Because the total protein content of human tears differs as to stimulus15 and with duration of stimulus18 and because women cry more frequently than men14 more information is needed. However, the field looks pregnant with discovery.
Lacrimation frequency declines with age.4 Concentrations of one tear glycoprotein declines after age thirty.19 Perhaps falling in love with spouse and children is more of a young person's pastime? With longer life-spans, perhaps rejuvenating our tears by adding more or different receptor proteins would be appropriate? Such therapy might benefit older adopting parents or give acceptance and renewed capacity for love to criminals, perverts, old maids and old bachelors. Yet ‘recreational use’ of pheromone reception is too dangerous to indulge.
LACRIMATION AND PARENT-INFANT BONDING
Among the most peculiar of human behaviors, postpartum depression, more commonly "the third day blues", is "normal" in mothers shortly after childbirth.20
162

Mild disturbances of mood occur in 50-70 percent of women during the postpartum period.21
Three to four days after the birth of their child, some women begin to cry for no apparent reason. The crying may be intense and prolonged and may often, but not always, be associated with a depressed mood (Coppen, et al., 1978).22
The neonatal sebum secretion surge on the fourth day after birth found by dermatologist Agache and his associates23 closely follows parental postpartum lacrimation. Evidence exists of a parent-infant bonding effort during this same critical period.24 Is the purpose for the mother's lacrimation sebaceous-pheromone reception enhancement?
Benfield and his associates indicate that the absence of grieving behavior in mothers of ill and absent premature infants can lead to "disintegration" of the family.25 Similar difficulties encounter stillbirth.26 Perhaps continued lacrimation maintains receptivity and the flexibility required for adaptability? In which case should not psychogenic lacrimation be encouraged, even cinematically with the expectation of improved outcomes? We need sad movies in these obstetric waiting rooms, (but upbeat flicks in cancer wards, of course.)
Brazelton videotaped mother and neonate interaction simultaneously, displaying the two faces on a split CRT screen. When the mother put on a blank expression (no "funny" faces or squeaky speech) the baby's face turned sanguine and he began to cry.27 This neonatal behavior flushes the face and scalp,28 wherein lie about 600,000 sebaceous glands,9 heating them (e.g. Seeley, T., et al.29), which stimulates abrupt, phasic sebum flow directly onto the surface 30 as it is being kissed. Crying, especially in babies, elicits an affectionate response.31 The neonate manipulates the parent's pheromone reception to strengthen the mother to infant bond. Seeing these heros of science miss this natural and now obvious sequence does not diminish their contributions, so essential to our eventual understanding of insidious pheromone addiction as the operative process involved in how human beings fall in love.
EXOCRINOLOGY
163

Forming the new maternal loving bond to the infant may affect other preexisting ones. Receptivity at the prospect of reception of the offspring's semiochemicals may renew her bond to the father and other family members allowed in close. This readiness to lacrimate, and the chemical makeup of maternal lacrimation itself, may be stimulated by a primer pheromone. Since plasma from maternal rats injected into virgin rats stimulates maternal behavior,32 perhaps human fetoproteins released into maternal circulation during pregnancy are pheromone receptor proteins? The fetus itself may emit feto-proteins as pheromone receptor proteins into maternal circulation for emission or for concentration onto receptor surfaces. Perhaps her pheromone receptivity is by a nocturnal autoemission and autoreception of a priming pheromone receptor protein? Lochia, a pregnancy and postpartum secretion of the vagina, or the breast's pre-milk colostrum are candidate sources along with the neonate's milky emissions of the 'genital crisis' (see Agache, P., et al.23) Such a protein primer scenario would approximate the conditions of rodent maternal priming seen by Mennella & Moltz.33 Excessive doses of priming pheromone might conceivably even contribute to instances of post partum depression in the mother. It is tempting to lay post partum depression at the feet of anti-pain pharmacology.
Reported stimuli to lacrimation vary, but time and length of crying behavior hold consistent. Female crying bouts usually occur in the evening.14 Family members draw together about that time of the day. Long episodes of crying happen more commonly in females.14 Lacrimation’s bonding effort is also suggested by the observation of post partum-like depression in adoptive parents.34
The female's husband-bonding and baby-bonding behaviors have been described with respect to lacrimation. Perhaps the wife and mother keeps her nose in the wind, so to speak. Is she the family member most sensitive to change? Perhaps the mother's being the bulwark of the home involves specialized semiochemical duties for which she is uniquely equipped?
164

MOURNING
Every human knows that emotional tears appear at funerals. A perceived probability of the necessity of a bond break (such as having word of the grave illness or distress of a loved one) evokes lacrimation. As a practical matter, the breaking of obsolete love-bonds and new biological bonding should both be undertaken during mourning. These functions are proposed for the emotional crying induced by such social situations.
Pheromone receptivity in mourning might adjust to compensate grief with new and renewed love. Crying would flush away a bond no longer useful. No economic logic justifies loving the inanimate or even for going to their funerals, for that matter. The dead no longer need our love, the living do. So in saying goodbye with tears, perhaps the chemistry of forgetting engages? We attend funerals for the living, and it is why families cling together at such times.
Reproductive fitness (survival of our genes in our offspring, or our relative's offspring) advances none at all by our loving any but the living. Replacing a useless bond with renewed closeness to those left alive makes sense. Tears, then, would be real and practical medicine for emotional hurts.
Suppose crying removes pheromone addiction and enhances receptivity to new pheromones in human beings. We would then mourn the loss of a loved one because of a biological economic reality. It is a practical matter. A malleability of affection at times of grief would be biologically expedient. It would also be economical.
If human emotional tears help insure pheromone addiction, then a biological reason explains why human beings are brought closer when they share a common grief. A crying person would seek out another individual with whom a bond should be established or strengthened.
EXOCRINOLOGY
165

After loyalties are established, casualties of war engender little sympathy from fellow and opposing soldiers. Light or early combat losses, however, may establish unusually close comradeship.
FAREWELLS, AND WELCOMES
Human tears can accompany welcomes and goodbyes. While animals, such as songbirds35 may anxiously welcome their fellows after an absence, only people say goodbye with foreknowledge, so only people part emotionally. Wars see much of this behavior, as well. As the troopship steams away into danger, wives and girlfriends shed (and absorb) tears. Does this chemoreception ripen criers for new love, domestic and/or romantic? Flushing out the old would open receptivity for the new semiochemical signal. Is biology so pragmatic as to start the flushing even before the boat pulls away from the dock? Sadly for soldiers, it would seem so. Don’t cry for me, Argentina!
Tears of welcome for our returning fighting men would function to open a place in our circle for our heroes. Recall the tears of parting, which are posited to work to assure that unmarried girls on shore need not wait? All shed more tears on the boat's return.
Tears precede and accompany a kissing frenzy at the reunion of loved-ones.34 Tearful kissing frequency/duration may be proportional to length of absence and the strength of the previous bond. If erbkoordinated lacrimation aids pheromone reception, its excretion during such emotional behavioral situations is logical.
TEARS AND THE HUMAN HIERARCHY
Tears and pheromone recognition may play some role in adjustments of our social hierarchy set. When one's position shifts, such as on winning a beauty pageant, or with election to high office, episodes of lacrimation often come with the news.
166

(The author anticipates the opportunity to self-observe upon news that he has won Nobel prizes for Peace, Medicine, Chemistry, and Economics.) The greater the increase (or decrease) in status, the greater the predicted lacrimal response to accommodate one to the change. Do you recall a young team captain's tears after winning the impossible football triumph? Try remembering a national beauty contest without the winner’s tears. Lacrimation and hierarchical change go hand in hand. The higher the assent and the lower the descent, i.e. the bigger the shift, the greater should be the lacrimation.
But, you may say, "I saw that proud moment, and I cried a little, too." Shifting of others in a hierarchy also requires a bit of our own lacrimation in observation. The "beautiful wedding" that brought tears to everyone's eyes is an excellent example. Welcome to your new hierarchical status, newly married couple!
Lacrimation-inducing entertainments, such as sad movies and plays, would employ our innate hierarchy adjustment device. Suspending disbelief may engage this instinct. Television and moving picture industries are new to human evolution. These stimuli would stimulate unnecessary semiochemical receptivity within the present paradigm. So audiences should become more jaded with experience.
Modern males, ever the opportunists, might exploit such situations by exposing females to axillary scents during such entertainments. Anecdotes exist of unwarranted improvement of mood in unfamiliar females following such exposures. A controlled study of such effects and those upon population semiochemistry might be in order.
Crying presents both visual and aural signals which instinctively provoke sympathy.1 Indeed, perhaps these signals stimulate reciprocal receptivity for chemosignals in all of us?
Crying can help people adjust to new political realities, too. Who of us alive at the time did not weep on hearing of a young president's death? (Aside from the umbrella-man killer of JFK, George H.W. Bush, of course.)36 Human beings may have done most of their social evolving in small groups.37 In small groups, submission to leadership would involve physical contact and personal demonstrations
EXOCRINOLOGY
167

of fealty. Although the new leadership in a country may not be able to meet everybody, the populace may prepare to accept them semiochemically.
French people crying as Nazi columns roll through Paris July, 1940.
Joyous celebrations of victory may be accompanied by tears. In such a case the entire population may need to adjust to its new supremacy just as in a small band of hunter-gatherers. Likewise a popular defeat might require similar adjustment.
Disgrace, right or wrong, may bring on lacrimation. Tears in disgrace reflect one's diminished hierarchical status. Such pheromonal receptivity before angry authority may be hazardous to one’s health by virtue of one’s exposure to aggression pheromone, a logical autoimmunity poison. Alternatively tears might flush out acceptance of the disgracers and ready the disgraced for another hierarchy down the road of life.
Pain of injury or disease may stimulate soothing tears in adults. During human evolution, injury may have once foretold death or permanently lowered status within the group. However, natural opiates in tears18 could have anesthetic qualities as well.
Psychological depression can have onset at a change or perceived change of the victim's hierarchical status. Life events such as the loss of a limb or the loss of a
168

loved one typify onset conditions in many cases of depression. An increase in the number and duration of lacrimal episodes accompany depression.14 This would indicate a super-receptivity for human pheromone. Depressed people more often become addicted to nicotine and smokers more often become depressed.38 Some beneficial effects of family therapy can be seen in depressed patients. Perhaps these all share unassuaged appetites for human pheromones? Semiochemical medical treatment of receptives with donated paternal facial skin surface pheromones under conditions of pheromone asepsis should be investigated to remedy depression. A standard dose of 150 mg of adult male facial skin surface lipid taken by mouth would be a good starting point.
In addition, as heterosexual men attest, a female partner can exhibit an unbridled joy upon exposure to male semen. It appears that semen may have antidepressant pheromonal properties from hormones transmitted in ejaculate.39,40 Seminal pheromones and pheromone binding proteins as well as pubic tuft and penile skin surface lipids so anecdotally adept at inducing sexual desire might also improve mood. Of course, these are tiny transmissions likely to escape scientific attention into the foreseeable future. Hormonal injections to establish post-coital serum levels compared against these findings might be instructive, however unless some synergism of the full complement exists, serum levels of testosterone alone do not seem to improve mood while levels of dehydroepiandrosterone--a sebaceous skin surface emission stimulant--do.41
Another time for crying comes when change appears likely. Periods of family stress increase lacrimal episodes. More frequent lacrimation would enhance the sensitivity of chemoanalyses to keep us ready for when the change finally did come.
Crying alone and "finding a shoulder to cry on" may find new meaning within the pheromone reception hypothesis of psychogenic lacrimation. Crying alone may not improve family cohesiveness, and, depending upon the pheromone ensemble of the crier, may even trigger pathology. "Finding a shoulder to cry on" might be taken to mean "finding an armpit to sniff while I lacrimate." Men would do well to "comfort" their families at such times by allowing lacrimation on the shoulder (near the armpit). Reducing stress might also influence results.
EXOCRINOLOGY
169

We should observe little crying in only children of stable households. On the other hand, we should also see frequent tears in closely spaced children living in unstable families.
EMOTIONAL LACRIMATION OF INFANTS AND CHILDREN
Crying is innate, although "infants do not commonly weep until they have attained the age of from two to three or four months".27 The reason for emotional tears in infants and children is unknown.
Crying signals the mother;42 however, tears are not necessary for that purpose because neonates have no tears.27 Economically, if tears were superfluous in childhood they would have sunk under the pressure of natural selection.
If emotional lacrimation enhances semiochemical reception then such reception must be very important in infancy and childhood. Small children weep frequently; typically seeking their primary caretaker for close contact and kissing. Here are semiochemical analysis opportunities on a more or less regular basis.
What could be the purpose of such continuing semiochemical analyses? Semiochemical communication has been speculated to have a role in psychosexual development.43 Physical separation from parents during critical periods has an affect upon the development of delinquent behavior in males.44 Physical separation is ipso facto semiochemical deprivation. Perhaps delinquency is pheromone deprivation? Testing of adult male facial sebum to treat juvenile delinquency has been quite successful in open trials. Modern pheromone deprivation may result from parental hygiene practices. After all, parents did not evolve with RightGuard, hot showers, and feminine hygiene protection! Similarly, criminal behavior is associated with ozone concentration. Ozone irritates mucosal pheromone-receptive surfaces.
Sparing the rod's stimulation to lacrimation in the parent's presence may indeed spoil the child. How appropriate for the love-bond between parent and child
170

to be strengthened when the parent discovers misbehavior and applies negative conditioning therapy sufficient only to stimulate lacrimation.
OTHER EXPLANATIONS
FOR HUMAN TEARS
There are alternative hypotheses about why we cry. One may be among the oldest hypotheses in medicine. Do we weep to remove unknown toxins or "ill humors" as an excretory function?4 Frey reluctantly reached this tentative conclusion by process of elimination. There are proteins with no known function in tears.16 Another45 (Montagu, A., 1959), suggested some sort of mucous soothing function for lacrimation. Montagu wrote that because crying involves inhaling and exhaling a lot of air, tears must replace lost moisture. If tears are there to moisturize the lungs of crying people, then a few questions arise. Why have lungs that desiccate? According to Frey,4 these membranes are not so delicate as to need tears to protect them. Why not just breath less emphatically in lacrimation? Or, if crying was just to signal grief, why have one signal for all humanity? Shouldn't different cultures grieve differently? Frey4 discounts Montagu's hypothesis. He notes that not all weeping involves increases in respiration.
Speculating psychological writers insist human beings cry to "let it out," (e.g. Lipe46). It being some emotion-phlogiston perhaps? No tangible evidence supports their hypothesis. There are no poisons in human tears and no waste products.4 Human beings do not lacrimate to express themselves. Human lacrimation does not relieve tension psychologically, but we usually feel better after doing it.
How does human lacrimation soothe? Lacrimal glands are the only place outside the CNS and adrenal medulla where the natural opiate, leucine-enkephalin, resides.4 When we cry, we also dose ourselves with that endorphin. Note the precision with which this device engages. The more pain, the more crying, the more a soothing balm is laid on. And absurd atheists ask why heaven gives tears to crippled children!
EXOCRINOLOGY
171

Tears are hard-wired to the pheromone recognition hardware of the brain. Human emotional tears are induced by activation of the medial prefrontal cortex.47 The medial prefrontal cortex is a projection area of the pheromone sensing accessory olfactory bulb in the cat.48 Humans have no accessory olfactory bulb, per se. People have fields of pheromone-sensing microvillar brush border cells lining the entire upper respiratory system as all physicians learn in first year histology. However, the human prefrontal cortex (which does many things) is among the brain structures with dissimilarities that differentiate heterosexual men and homosexual women from homosexual men and heterosexual women.49 Right next door, the anterior part of the inferior lateral prefrontal cortex is activated on PET scans after stimulation with androstadienone, an airborne human pheromone.50
ANIMAL BEHAVIOR
Are tears for pheromone recognition in all bonding vertebrates? Humans walk upright. We alone shed tears onto the face at pheromone reception appropriate times.4 This fact would be important if tears were purely visual signals. Why aren't tears purple or green? They would be better visual signals if they were not so small and transparent.
Because most species have nasolacrimal ductwork from eye to nasal passage, lacrimal pheromone reception may not be externally obvious. Most land vertebrates walk on all fours and other two-legged beasts have beaks. If animals utilize lacrimal chemosensory reception for bonding it might be expected episodically like ours. Unfortunately direct measurements of simultaneous pheromone reception and internal lacrimal secretion in any species are lacking.
However, ethological observations support this hypothesis of lacrimal involvement in pheromone recognition. In geese Konrad Lorenz51 observed physiological changes around the eyes after behavioral experience akin to human bereavement in pair-bonding avians. In some species of sea birds, inconsistently observed portions of the courtship display utilize behavior seen during excretory
172

lacrimation of sea salts.52 Clearly this and other courtship displays and courtship dances of birds forming pair bonds, are mechanically distributing pheromone reception fluids (tears) to chromatographic pheromone reception surfaces. Construction of lucite models of avian nasal passages, including nasolacrimal ducts with tear origin at lacrimal glands will demonstrate the necessity for the gyrations of courtship in avian bonding.
Tear-secretion behaviors can appear at pheromone-reception-appropriate times. Many avians "bill shake" and "dance" when they fall in love. Perhaps their bills are flung from side to side to bring down lacrimal pheromone-reception fluids. Bill shaking looks like behavior observed as concentrated brine is excreted from the lacrimal glands through the nasolacrimal ducts and out through the nostrils in the beaks of oceangoing birds deprived of fresh water. Indeed, you can inject salt water in some species and see the same type of head shaking behavior as a salt elimination device.53 If glands concentrate ions for salt elimination in sea birds, might the same glands also concentrate ions for pheromone recognition? Since the behavior appears in the "courtship dance," the secretion may be there as well.46
Why should birds gyrate so to fall in love? Birds have beaks. Hard beaks can not utilize peristalsis or mucosal cilia to transport tears internally. Those mechanisms are available to non-beak mammalian species. The funny movements of birds get tears where they need to go inside the beak for pheromonal bonding. Indeed, economically, every dip and twist of the head should be accountable for some internal avian nasolacrimal obstacle.
Ornithologists have termed what they consider functionless behaviors, "displacement activity."54 This unfortunate appellation has been applied to a number of avian behaviors which appear pheromone-receptive. Among them are territory marking behavior of avians (utilizing the "preen" gland), and pheromone exchange behavior.
Pheromone exchange behavior would be mostly the male's depositing of the preen gland lipid onto food or nest and signaling its location to the prospective mate with nesting or courtship displays. [Do hen pupils dilate seeing a tom turkey spread
EXOCRINOLOGY
173

his tail feathers? No, the avian eye is a special case.] Pheromone exchange behavior would also include marking a "nuptial gift" with pheromone for the prospective mate to swallow or sniff, and also beak rubbing. Beak rubbing is seen in more social birds.33 The sebaceous glands of the rictuses55 come in contact during the behavior. Perhaps it is for bonding pheromone exchange?
Jouventin's46 report typifies some ornithological thinking. He has done a particularly good job of observation, however.
Sea birds have a supraorbital gland which concentrates excess sodium chloride, as shown by McFarland (1959) in Aptenodytes forsteri, A. patagonicus and other species. They regularly and vigorously shake their heads from side to side to get rid of the concentrated sodium chloride solution, sometimes visible hanging in droplets on the edge of the bill. Billshaking can be induced experimentally by injecting NaCl (SchmidtNielsen and Sladen, 1958). Billshaking can often be observed, repeated several times without apparent need, in conflict situations, as a typical displacement activity. One step further, this movement would even have become part of the courtship display of the King penguin, according to Stonehouse (1960, p. 27), who considers it similar to the "headflagging" described in black headed gulls (Tinbergen and Moynihon, 1952, p. 2122). However, when the head is lowered, e.g. while singing, excess liquid probably flows to the bill and is expelled by headshaking. Yet many King penguins were not observed to shake their heads while singing [Perhaps because the fluid is used internally?BN]. Moreover, the encounter with a future partner is fraught with such emotion that displacement activities frequently accompany it. In the Emperor penguin, which lowers its head to sing, headshaking was recorded in 21% of the cases (n=108) just after singing, while van Zinderen Bakker Jr. (1971, p 256) observed it in the Gentoo penguin in 95% of observations just after chicks were fed, which also involves lowering the head. [Just in time for an addictive pheromone from the chick as reward for bringing foods.BN] In 86% of Emperor observations (39 out of 45), billshaking was followed by a swallowing movement; according to Isenmann (1971) this restores the inner ear pressure, but we interpret it (1971) as elimination of the liquid left after headshaking. In any case, bill shaking and swallowing form a miniature stereotyped sequence which often appears during courtship display: then, about 18% of billshaking was followed by swallowing in the Emperor penguin according to our results (n=50). [Note frequent swallowing is seen in humans kissing to form a long lasting bond.BN]
174

Even though the sequence cannot be considered an element of courtship display, as it is absent four times out of five, and as long-lasting couples can do without it, it is not only a basic movement but a displacement activity [...] (Jouventin, P., 1982).46
This is similar to human experience where heavy shared lacrimation may be a rare occurrence. Long married couples can comfortably do without much of youth's passion. The same reduced passions appear in long-matched songbirds.33 The expectation of lacrimation with formation of long-lasting relationships in animals may be met with further study using current techniques. In the few cases where tears or tearing behavior can be seen, bonding and tears go together. Other anecdotal accounts exist.56
Many animal tears carry pheromones. Associating a pheromone reception fluid with pheromone would both prime and release simultaneously.
Pheromonal tears exit the glands around the eye in marsupials, insectivores, armadillos, rabbits, rodents, dolphins, walruses, seals, pigs, and deer.57 Instead of smearing tears around on their faces like we do, animals may smear the tears all over their bodies, along with saliva. These sebaceous chemicals in gerbil Harderian gland secretion have yet to be analyzed.51 In contrast the numerous sebaceous glands around the human eyes are mostly tiny, but some are big enough to be seen. They may function in maintaining the tear film;9 however, some role in the semiochemical commerce can not be ruled out.
DOGS
Working with canine tears, Barrera R., et al.58 found no differences in protein concentrations between sexes. In dogs pheromone reception probably cycles with estrous and the Barrera group may not have tested bitches in heat. Perhaps dimorphisms exist elsewhere to accomplish canine sexual orientation; however, the complex protein composition of dog tears52 may simply reflect a repertoire of binding proteins useful for tracking semiochemicals of other species. Dogs have excellent noses.59
EXOCRINOLOGY
175

To explain their devotion to humankind, a similarity between, or even a duplication of, some human specific tear protein should be expected in canine lacrimal or other pheromone receptor secretion. Pheromone recognition sharing may be seen in other parasite-host pairings60 and may follow from shared receptor proteins.
NASOLACRIMAL VOMERONASAL APPARATUS
In humans, behind and above each nostril, three nasal passages, or meatuses, carry air from the nose. From these meatuses blind sinuses extend. The nasolacrimal duct channels tear fluid through the nasolacrimal bone from the inner corner of one eye to the bottom nasal passage on the same side. The bottom nasal passage is called the inferior meatus ("lower passage" in Latin).
A pair of vomeronasal, or Jacobsen's, organs detect low volatility pheromones in the naso-pharygial area of diverse species (Beauchamp, G.K., et al., 1983; Halpern, M. and J.L. Kubie, 1983; Halpern, M., 1987). Human beings might have functioning vomeronasal organs located above and forward of the inferior meatus (Moran, D.T. et al., 1991; GarciaVelasco, J. and M. Mondragon, 1991), in which chemoreception has been detected in response to putative human pheromones (MontiBloch, L. and B.I. Grosser, 1991). The receptor cells of vomeronasal organs are sensory microvillar cells, the same cells that line the entire upper-respiratory system in human beings. Nasolacrimal ducts open variably in different people. Like the nasal passages and sinuses, only rough bilateral symmetry prevails from one side to the other as well. Generally the duct from the eye opens into one end of a vomeronasal cleft, with the vomeronasal organ of Jacobsen at the other end (Schaeffer, J.P., 1912; 1916; 1920). A sexual dimorphism in vomeronasal organ pheromone reception has been recorded (MontiBloch, L. and B.I. Grosser, 1991) supporting sex-defined functions discussed earlier.
176

MICROVILLAR CELLS IN PHEROMONE RECEPTION
It was once thought that the machinery for human pheromone reception did not exist or, where it could be found, it was merely a vestige (Comfort, A., 1974). A functionless, evolved vestige would be an economic impossibility, a contradiction in terms. Ignorance of function was really the preferred explanation. Happily, microvillar cells of the type that senses pheromones in all animals examined were recently consistently detected in olfactory and nasal mucosa. One of the places where these microvillar cells were found was at the opening of the nasolacrimal duct to the inferior meatus. These pheromone receptor tissues are in the right place to be bathed in receptor proteins and the high ionic concentration necessary to that reception. Olfactory pheromones as well as nonvolatiles hydrophilics such as sebum must reach these tissues readily.
Clumps of pheromone sensing microvillar cells lie at the terminus of the nasolacrimal duct. Down the same duct flow out over microvillar cells the manganese ions and receptor proteins binding to lipid and other pheromones in the lacrimal fluid.
A drawing depicting microvillar cells from the vomeronasal organ of elephants (Rasmussen, L.E.L. and B. Hultgren, 1990) are seen below.
EXOCRINOLOGY
177

Fig. 4, page 155 Chemical Signals in Vertebrates 5, D.W. MacDonald, D. MllerSchwarze, and S.E. Natynczuk, 1990. Oxford Science Publications London.
The human vomeronasal organ's microvillar cells differ only slightly. Microvillar cells resemble olfactory cells but specialize for pheromone reception with shorter receptor cilia.
178

Microvillar cells belong to a class of chemoreceptor cells which normally inhabit a discreet organ in most other animal species. The organ, Jacobsen's organ or the vomeronasal organ, specializes in recognizing pheromones. This knowledge was acquired only recently. Indeed, until about 1970 or so, the mammalian vomeronasal organ was a structure in search of a function.61 In human beings, the microvillar cells are not so concentrated, and were thought until lately62 to clump into a recognizable organ only in about 15% of humans.63 Of course, as every medical student learns, sensory non-motile microvillar ‘brush border’ cells line the upper half of the human respiratory system, a phenomenon which is unique to the human species. Eureka, these broad swathes of pheromone sensing microvillar cells must be those that recognize human pheromones. It is altogether right that we have the largest area devoted to pheromone reception, since humans have the largest scent glands and most active skin surface lipid glands by far.
NASAL PASSAGES
The nasal passages run from the exterior vestibule of the nose into three chambers running one above the other and thence into one again at the choana to the pharynx. The function of the six nasal passages and their attendant sinuses passages remains a mystery. However a function in pheromone recognition may be hypothesized.
A few clear lucite models of the nasal passages taken from cadaver impressions were provided to the author by previous researchers.64 The Hornung group followed radioactive airstreams to detect spatial consistencies in them. However, by passing various fluids and smoke through these clear models, it was transparent that parallel lines were inaccurate.
Smoke sniffed into the models displayed a consistent roiling in agreement with the Hornung depiction. However, erectile tissues pervade the nasal passages, opening and closing regularly and in response to emotion and sexual releases.65 These erections direct or divert incoming and outgoing air. A dynamic nuclear
EXOCRINOLOGY
179

magnetic resonance imaging (dMRI) of these structures during emotional situation changes should prove interesting. Such motion pictures might improve our understanding of the physiology involved and allow better modeling for more precise laboratory experimentation.
Surprisingly, passing smoke through these models established that airflow taken into the nose is treated much differently than exhaled air. Nasal passages (at least those made of lucite) route the incoming airflow through the upper meati in a consistent turbulence. This induced turbulence may ensure more complete chemoreception.66 However, because air borne odor plumes are discontinuous,67 turbulence may also randomize samples. Obtaining an adequately randomized sample is the first essential step in analytical chemical analysis,68 accordingly biological chemo-analyses may proceed similarly.
Randomized incoming airflow reaches the olfactory mucosa of the superior meatus for olfactory perception. Odorants concentrate by dissolving into the Bowman's secretion69 where, in all species, specialized Odorant Binding Proteins (OBPs) in the class of lipid binding proteins are also dissolved.70 Much less volatile sebaceous lipids reach this area through mucosal surface diffusion.
The extreme "spreadability" of sebum has been utilized for precise measurements of sebaceous secretion.71 The precision of the technique is demonstrated by one of their figures. In it the data show menstrual variations in a female subject. Identical data on menstrual variability of sebaceous secretion appeared many years later.72
In the smoke observations of the clear lucite nasal passage model, outgoing airflow behaved unexpectedly, too. Outgoing, lung-cleansed air73 converged upon the olfactory mucosa and the semilunar hiatus from both directions at once. Perhaps this distinction allows discrimination of interior odors, which make up most of our perception of food, from exterior odors. Since exhaled air is so clean, the inward flow of exhaled air back through the upper two meati may reset olfactory reception.
180

The sensory stretch receptors of the maxillary sinuses probably inform the direction of air in inferior meatus, allowing such a reset from local sensation.
However crude, Lucite models do allow us to hypothesize. Perhaps the three nasal passages work together to form an ingenious passive valve for liquids as well? For fluids arising from the rear as from the stomach, the valve diverts all liquids into the inferior meatus or "low passage". Valving refluxing gastric juices away from sensitive olfactory tissues seems a logical function for this maze of "functionless" structures near the choana. From the models, the best effect seems to come when the head is bent over about fifteen degrees from vertical. Interestingly, this leaning forward position appears to be taken instinctively by humans about to throw-up. Maybe that stance keeps the vomitus off of one's tunic, too?
The mechanics of lacrimation include some gross behavioral characteristics which any casual observer can identify. With the head bent down, criers maintain a forward posture in stance or sitting. Again from observations of liquid passing through transparent Lucite model of the nasal passages, this posture appears to keep that lacrimal fluid collecting in the inferior meatus. Perhaps this posture protects the olfactory mucosa from wetting by tears, but more likely it keeps the reservoir at the base of the hiatus semilunaris full of lacrimal fluid for chromatographic solvent.
Tears run onto face from the eyes after filling the reservoirs in the nasolacrimal duct. The tears wet the face, and some roll down the cheeks. A few enter the mouth at the corners. (Is this compensation for our lack of a nasopalatine duct between mouth and inferior meatus? Only about 10% of the population retains a hole between palate and nasal passage.) No, it probably is not, but speculating in a book like this is harmless enough if you also halt the progression of a few incurable diseases (Alzheimer’s) and cure sociopathies (crime, perversion, drug-addiction), eh?
SOBS
Ever notice the breath effects of sobbing and the holding of the hands over the face in severe lacrimal episodes? Sobs appear to vary somewhat like sniffs do. Sobs
EXOCRINOLOGY
181

may function analogously to sniffs, but for pheromone reception in humans. Sobs may impart the chromatographic effect of sending many solvent fronts of tears over the pheromone sample being analyzed.
In regular olfaction chromatographic resolution may be enhanced by a "sniff".74 Any cooling and warming of a chromatographic substrate improves resolution. This seems to be what happens to the olfactory mucosa by the sudden evaporative inhalation characteristic of sniffs. Animals, with more sensitive olfactory apparatuses, regularly vary sniffs accelerating to a climax.75 These may represent a whole series of temperature programs, familiar to chromatographers,76 lasting but a very short time.
Who has failed to note the trembling hands held tightly over the face of a person engaged intensely in lacrimal pheromone-receptivity activity? Why do we instinctively cup our hands to our face in intense psychogenic lacrimation? Perhaps the association of reception enhancing tears and a hypothesized addictive facial pheromone prime and release bonding for anyone kissing those tears away? Alternatively, we may be forming a developing chamber similar in function to those of thin layer chromatography.
Thin layer chromatography resembles the paper chromatography most of us did in elementary school. Put a dot of a chemical mixture near the bottom of an absorptive plate. Put the plate into a covered pot with some liquid solvent at the bottom. A mysterious "solvent front" climbs onto the plate and past the dot of mixture. When it passes, different components of the mixture go with it and then drop off one at a time as the front continues up the plate. You may have to "develop" the plate (kind of like film) in order to see that a number of faded dots mark where parts of the mixture dropped off. The distance from the dot of mixture down at the bottom to the spot where the chemical stopped climbing with the solvent front and dropped off usually differs for different chemicals. You can even scrape the plate always at the same distance and get a pure sample.
Thin layer chomatographers content themselves with hanging around until a single front physically shifts one chemical away from another. Their eyes and rulers
182

take the measurements. In contrast gas chromatographers use a detector, about the size of a fist, to measure the time course over a constant distance instead of different distances in a common time. The detector burns or zaps the chemical passing by in a gas stream and marks a computer screen with a bump when one does. Time from the start to detection usually differs characteristically from chemical to chemical.
The one big detector at the end of the column in gas chromatography is replaced by many tiny microvillar and/or olfactory nerve cells. These cells may perform the same job in Jacobsen's organ and olfactory tissues. Instead of a single big detector, an array of chemosensory cells do the detecting. Remember how components stopped migrating up the place as a characteristic of the chemical?
Now, remember from Chapter 3 that humans may have many hundreds, possibly thousands, of different pheromones to be detected? Some chromatographic separations would be a logical device to have behind our noses to aid human chemo-analyses.
Why use an array of detectors? After all, one detector is enough for sophisticated chemical analysis.70 The answer is speed. Instead of a set starting point and time, and then a series of detections across a single detector, chemosensory cells work in parallel. Starting point and time are sensed in the surface lymph by the dissolved pheromone receptor protein. Follow-on times from differing pheromone receptor proteins, either dissolved in the lymph or on the chemosensory cell surface, are compared in accessory olfactory bulbs and differences plotted to the brain into a chaotic pattern (reception points as the new origin). It is the reproducible chaotic pattern which the brain recognizes as a scent.
Olfactory sensations are turned into information at the glomeruli of the olfactory bulb, where the timing of the first, extracellular Bowman’s secretion soluble protein conformational charge release on odorant contact, and subsequent dendrite surface sensations are sorted out. These differences are all projected in parallel simultaneously to the brain. Unknown chromatographic forces impart identifying delays for chemicals moving through the Bowman's secretion and these are detected by simple comparison in the bulb. Intensity of the signal, imparted by the odorant's
EXOCRINOLOGY
183

affinity to the OBP, is a less identifiable portion. These signals map into a chaotic pattern recognizable to the brain as a fragrance.
If pheromone reception during lacrimation maintains a chromatographic component, as seems indicated, then perhaps the peculiarities of the sinuses serve as well. Sinuses have no known function and no reasonable explanation for them has ever been found.11 The sinuses form a column of retorts opening off of the canalis semilunaris or semilunar hiatus. Ostia (or openings) of almost all of the sinuses connect with the semilunar hiatus. Recall, the lack of sinuses might well be responsible for the babyish features, noted by Lorenz45 and others,77 of enlarged forehead, small face, and chubby cheeks. The gradual development of more and more sinuses during childhood may be correlated to sensing one's relationship and status within a community.
OTHER HUMAN PHEROMONE RECOGNITION SYSTEMS?
This chapter has introduced one model of pheromone reception. Mattes78 contends there is increasing evidence supporting an oral chemosensory detection system for free fatty acids with multiple transduction mechanisms.
Other pheromone reception mechanisms should be expected, as backups or in parallel. For instance, some research suggests that the long-chain triglycerides, a major component of human sebum capable carrying information, are not digested but hydrolysed taken whole into the lymph. Micelles transport hydrolized triglycerides from gut to lymph to ovary and beyond while shorter chain cousins are relegated to digestion in the liver.79 Thus an entirely different system exists by which probable human lipid pheromones may have direct, unmodulated access to the reproductive system and beyond.
184

EXOCRINOLOGY
185
1 Eibl-Eibesfeldt, Irenäus (1970a). Ethology the Biology of Behavior, translated by Erich KlinghammeR New York: Holt, Rinehart and Winston 530 p.
2 Nicholson, B. 1983; A semiochemical addiction model for human bonding. Paper presented to the Animal Behavior Society and the International Society for Human Ethology, Lewisburg, PA.
3 Nicholson B., Pheromones cause disease: pheromone/odourant transduction. Med Hypotheses. 2001 Sep;57(3):361-377.
4 Frey WH. the II. Crying the mystery of tears. Minneapolis, Minnesota: Winston Press, Inc. 1985;174.
5 Feksi A, et al. 'Maternity Blues' and hormone levels in saliva. J Affective Disord. 1984;6(3-4):351-355.
6 Fullard RJ, Tucker DL. Changes in human tear protein levels with progressively increasing stimulus. Investigative Ophthalmology & Visual Science. 1991;32(8):2290-301.
7 Morris, Desmond 1972; Intimate Behavior New York: Random House.
8 Wyart C, Webster WW, Chen JH, Wilson SR, McClary A, Khan RM, Sobel N. Smelling a single component of male sweat alters levels of cortisol in women. J Neurosci. 2007 Feb 7;27(6):1261-1265.
9 Sack, RA, K.O. Tan and A. Tan 1992; Diurnal tear cycle: evidence for a nocturnal inflammatory constitutive tear fluid. Investigative Ophthalmology & Visual Science 33(3):626-640.
10 Montagna, William; Parakkal, Paul F. The structure and function of skin 3rd edition. New York: Academic Press 1974.
11 Hafez, E.S.E. 1976; Parameters of sexual maturity in man. pp. 105-122 In: Volume 3 Sexual Maturity: Physiological and clinical Parameters E.S.E. Hafez and JJ Peluso editors. Ann Arbor, Michigan: Ann Arbor Science Publishers Inc. 283p.
12 Drettner B. The paranasal sinuses. In: The nose: upper airway physiology and the atmospheric environment. Proctor A.editor. Amsterdam: Elsevier Biomedical Press. 1982;145-62.
13 Monti-Bloch, L., and B.I. Grosser 1991; Effect of putative pheromones on the electrical activity of the human vomeronasal organ and olfactory epithelium. The Journal of Steroid Biochemistry and Molecular Biology. (Proceedings of the International Symposium on Recent Advances in Mammalian Pheromone Research.) 39(4B):573-582.
14 Finck HT. Romantic Love and Personal Beauty; Their Development, Causal Relations, Historic and Natural Peculiarities. London: MacMillan. (1891).
15 Frey WH the II, Hoffman-Ahern C, Jason RA, Lykken DT, Tuason VB. 1983; Crying Behavior in the Human Adult. Integrative Psychiatry September-October:94-98.
16 Frey WH. the II, Desota-Johnson D, Hoffman C, McCall JT. Effect of stimulus on the chemical composition of human tears. American Journal of Opthalmology 1981;92:559-567.
17 Mii, S., K. Nakamura, K. Takeo and S. Kurimoto 1992; Analysis of human tear proteins by two-dimensional electrophoresis. Electrophoresis 13(6):379-382.

186
18 Fullard RJ, Tucker DL. Changes in human tear protein levels with progressively increasing stimulus. Investigative Ophthalmology & Visual Science. 1991;32(8):2290-301.
19 Bjerrum K, Halken P, Prause JU. The normal human tear glycoprotein profile detected with lectin probes. Experimental Eye Research 1991;53(4):431-5.
20 Pitt, B. 1978; Introduction. In Mental Illness in Pregnancy and the Puerperium. Sandler, M. editor. Oxford: Oxford University Press.
21 Yalom ID, Lunde DT, Moos RH, Hamburg DA. “Postpartum blues” syndrome. A description and related variables. Arch Gen Psychiatry. 1968 Jan;18(1):16-27.
22 Coppen A, Stein G, Wood K. Postnatal depression and tryptophan metabolism. In: Sandler, M. editor. Mental illness in pregnancy and the puerperium. Oxford: Oxford University Press, 1978.
23 Agache, P., D. Blanc, C. Barrand, and R Laurent R 1980; Sebum levels during the first year of life. British Journal of Dermatology. 103:643-649.
24 Klaus, M.H., and JH. Kennell 1982; Parent-Infant Bonding. St. Louis: The C.V. Mosby Company.
25 Benfield, D. Gary, Susan A. Leib, and Jeanette Reuter 1976; Grief response of parents after referral of the critically ill newborn to a regional centeR New England Journal of Medicine, 294(18):975.
26 Lewis E, Page A. Failure to mourn at stillbirth: an overlooked catastrophe. Br J Med Psychol. 1978 Sep;51(3):237-241.
27 Brazelton TB, Tronick E, Adamson L, Als H, Wise S. Early mother-infant reciprocity. Ciba Found Symp. 1975;(33):137-154.
28 Darwin Charles The Expression of the Emotions in Man and Animals. London: Murray. 1872;162
29 Seeley, T., T. Talbot, P.R Abramson, L.B. Perry, A.B. Rothblatt, and D.M. Seeley 1980; Thermographic measurement of sexual arousal: a methodological note. Archives of Sexual Behavior 9(2):77-86.
30 Cunliffe WJ, Burton JL, Shuster S. The effect of local temperature variations on the sebum excretion rate. British Journal of Dermatology 1970;83:650-4.
31 Lorenz, T.H., D.T. Graham, and S. Wolf (1953). The relation of life stress and emotions to human sebum secretion and to the mechanism of acne vulgaris. Journal of Laboratory and Clinical Medicine 41:11.
32 Terkel J, Rosenblatt JS. Maternal behavior induced by maternal blood plasma injected into virgin rats. J Comp Physiol Psychol. 1968 Jun;65(3):479-482.
33 Mennella, Julie A, and Howard Moltz 1989; Pheromonal emission by pregnant rats protects against infanticide by nulliparous conspecifics. Physiology and Behavior 46:591-595.
34 Foli KJ. 2010; Depression in adoptive parents: a model of understanding through grounded theory. West J Nurs Res. 2010 Apr;32(3):379-400.
35 Wilson, Edward O. 1975; Sociobiology The New Synthesis. Cambridge, Mass.: Belknap Press of Harvard University Press. 697p.

EXOCRINOLOGY
187
36 Nicholson, Ross (2008). Hillary’s Angel Tampa, FL: Nicholson Science, 2008.
37 Wilson Edward O. Biology and the social sciences. Daedalus. 1977 Fall;106(4):127-140.
38 Breslau N. Daily cigarette consumption in early adulthood: age of smoking initiation and duration of smoking. Drug Alcohol Depend. 1993 Oct;33(3):287-291.
39 Ney PG. The intravaginal absorption of male generated hormones and their possible effect on female behaviour. Med Hypotheses. 1986 Jun;20(2):221-231.
40 Gallup GG Jr, Burch RL, Platek SM. Does semen have antidepressant properties? Arch Sex Behav. 2002 Jun;31(3):289-93.
41 Basson R, Brotto LA, Petkau AJ, Labrie F. Role of androgens in women's sexual dysfunction. Menopause. 2010 Sep-Oct;17(5):962-971.
42 Lester, B., 1984; A biosocial model of infant crying. In: Lewis P. Lipsitt and Carolyn K. Rovee-Collier editors. Advances in Infancy Research. Norwood, N.J: ABLEX Publishing.
43 Kalogerakis, M.G. (1963). The role of olfaction in sexual development. Psychosomatic Medicine 25:420-432.
44 Clarke AM, Clarke ADB. Early Experience: Myth and Evidence. New York: Free Press, 1976.
45 Montagu, Ashley (1959). Natural selection and the origin and evolution of weeping in man. Science 130:1572-1573.f
46 Lipe, Hillary P. 1980; The function of weeping in the adult. Nursing Forum. 19(1):26-44.
47 Arita H. Emotional tears are induced by activation of medial prefrontal cortex. Nippon Yakurigaku Zasshi. 2007 Feb; 129(2):99-103.
48 Staples LG, McGregor IS, Apfelbach R, Hunt GE. Cat odor, but not trimethylthiazoline (fox odor), activates accessory olfactory and defense-related brain regions in rats. Neuroscience. 2008 Feb 19;151(4):937-47.
49 Savic I, Lindström P. PET and MRI show differences in cerebral asymmetry and functional connectivity between homo- and heterosexual subjects. Proc Natl Acad Sci U S A. 2008 Jul 8;105(27):9403-9408.
50 Gulyás B, Kéri S, O'Sullivan BT, Decety J, Roland PE. The putative pheromone androstadienone activates cortical fields in the human brain related to social cognition. Neurochem Int. 2004 Jun;44(8):595-600.
51 Lorenz, Konrad (1963). On Agression. Translated by Marjorie Kerr Wilson (1966). New York: Bantam Books Inc. 306 p.
52 Jouventin, Pierr 1982; Visual and Vocal Signals in Penguins, their Evolution and Adaptive Characteristics. West Berlin: Verlag Paul Parey.
53 Sturkie, P.D. 1976; Kidneys, extrarenal salt excretion,and urine. pp. 275-279 Chapter 14 Avian Physiology third edition P.D. Sturkie editor. New York: Springer-Verlag.

188
54 Sevenster, P., (1977) Displacement activities. In: Grzimek's Encyclopedia of Ethology. (Ed. by B. Grzimek). Van Nostrand Reinhold Company, New York.
55 Lucas, A.M. 1980; Lipoid secretion of the body epidermis in avian skin. In: The Skin of Vertebrates. Spearman RI.C. and P.A. Riley . editors. New York: Academic Press.
56 Hopson, Janet L. 1979; Scent Signals, the Silent Language of Sex. New York: William Morrow and Company, Inc.
57 Thiessen, Delbert D. 1988; Body temperature and grooming in the Mongolian gerbil. Annals of the New York Academy of Sciences. 525:27-39.
58 Barrera R, A. Jimenez, R Lopez, M.C. Mane, JF. Rodriguez and JM. Molleda Evaluation of total protein content in tears of dogs by polyacrylamide gel disk electrophoresis. American Journal of Veterinary Research 1992;53(4):454-456
59 Moulton, David G. 1977; Minimum odorant concentrations detectable by the dog and their implications for olfactory receptor sensitivity. pp. 455-464. In: Chemical Signals in Vertebrates Müller-Schwarze, Dietland and Maxwell M. Mozell editors. New York: Plenum Press 610 p.
60 Tumlinson, James H., W. Joe Lewis, and Louise E.M. Vet (1993). How parasitic wasps find their hosts. Scientific American 266(3):100-106.
61 Wysocki, Charles J Chuck 1989; Vomeronasal chemoreception: its role in reproductive fitness and physiology. pp. 545-566. In: Neural Control of Reproductive Function: proceedings of the Fifth Galveston Neuroscience Symposium held in Galveston, Texas, May 10-13, 1988. New York: Liss.
62 Garcia-Velasco J, Mondragon M. The incidence of the vomeronasal organ in 1000 human subjects and its possible clinical significance. The Journal of Steroid Biochemistry and Molecular Biology. (Proceedings of the International Symposium on Recent Advances in Mammalian Pheromone Research.) 1991;39(4B):561-563.
63 Moran, David T., Bruce W. Jafek and J Carter Rowley III 1991; The vomeronasal (Jacobson's) organ in man: Ultrasructure and frequency of occurrence. The Journal of Steroid Biochemistry and Molecular Biology. (Proceedings of the International Symposium on Recent Advances in Mammalian Pheromone Research.) 39(4B):545-552.
64 Hornung, David E., Donald A. Leopold, Steven L. Youngentob, Paul R Sheehe, George M. Gagne, F. Deaver Thomas, and Maxwell M. Mozell 1987; Airflow patterns in a human nasal model. Archives of Otolaryngology 113:169-172.
65 Holmes, Thomas H., et al. (1950). The Nose; an experimental study of reactions within the nose in human subjects during varying life experiences with a foreward by Warfield T. Lonscope. Springfield, Ill: Thomas.
66 Mozell M Mozell (1977). Processing of Olfactory Stimuli at Peripheral Levels. pp 465-482. In: Chemical Signals in Vertebrates Müller-Schwarze, Dietland and Maxwell M. Mozell editors. New York: Plenum Press 610 p.
67 Murlis, J 1986; The structure of odour plumes. Chapter 3, pp. 27-38 In: Mechanisms of Insect Olfaction. Payne, T.L., Martin C. Birch, and C.E.J Kennedy editors. Oxford: Clarendon Press, 364 p.

EXOCRINOLOGY
189
68 Kennedy, J H. 1984; Analytical Chemistry Principles. San Diego, California: Harcourt Brace Javanovich, Publishers. p. 3. (753 pages).
69 Hornung DE, Mozell MM. Factors influencing the differential sorption of odorant molecules across the olfactory mucosa. J Gen Phsiol. 1977 Mar;69(3):343-361.
70 Pevsner, J, P.M. Hwang, P.b. Sklar, V.J Clark and Soloman H. Snyder 1988; Odorant-binding protein and its mRNA are localized to lateral nasal gland implying a carrier function. Procedings of the National Academy of Sciences 85:2383-2387.
71 Lorenz, T.H., D.T. Graham, and S. Wolf (1953). The relation of life stress and emotions to human sebum secretion and to the mechanism of acne vulgaris. Journal of Laboratory and Clinical Medicine 41:11.
72 Burton, J.L., M. Cartilidge, and Sam Shuster (1973b). Variations in sebum excretion during the menstrual cycle. Acta Dermato. Vener (Stockholm) 53:81.
73 Mozell Maxwell M. (1992). Personal communication.
74 Sobel N, Prabhakaran V, Desmond JE, Glover GH, Goode RL, Sullivan EV Gabrieli JD. Sniffing and smelling: separate subsystems in the human olfactory cortex. Nature 1998 Mar 19;392(6673):282-6.
75 Youngentob SL, Mozell MM, Sheehe PR, Hornung DE. A quantitative analysis of sniffing strategies in rats performing odour detection tasks. Physiol Behav 1987;41(1):59-69.
76 McNair, Harold M. and Ernest J Bonelli (1969). Basic Gas Chromatography. Lithographed by Consolidated Printers, Berkeley, California. Varian Instrument.
77 Eibl-Eibesfeldt, Irena us 1975; Ethology: The Biology of Behavior, second edition. New York: Holt, Rinehart and Winston.
78 Mattes RD. Oral detection of short-, medium-, and long-chain free fatty acids in humans. Chem Senses. 2009 Feb;34(2):145-50. Epub 2008 Dec 16.
79 Clément J. Digestion and absorption of dietary triglycerides. J Physiol (Paris). 1976;72(2):137-170.


Chapter 7: Human Pheromone Transduction The human upper respiratory system is simply covered with pheromone receptors. We have more pheromone receptor cells than any other species, as might be expected from the beast that carries more pheromone-emitting glands than any other species. The pheromone sensory microvillar cells, histologists call them brush border cells, line most of the human upper-respiratory system. Every physician remembers memorizing their odd appearance because that is the key to differentiating upper from lower respiratory mucosa under the microscope.
In vertebrates generally, most such cells with short, non-motile cilia and their secretory companions can be found in a specialized pair of vomeronasal organs (VNO) located in the animal’s nasal passages. Sometimes the VNO is called Jacobsen’s organ. Vomeronasal organs generally work with the many sebaceous scent glands on an animal’s tail. Wagging or dragging tails lay down the animal’s own scent and the VNO detects the scent laid previously to keep the animal on course to utilize its territorial resources optimally. It is much like mowing a lawn because you detect where you have been and stay close. All one need do is maintain a constant concentration at the VNO.1
Of course, the microvillar cells of the VNO recognize territory edges, too, (often scent marked with the egregiously mis-named ‘displacement activity’), and perhaps recognize mates and potential mates. Humans have tiny VNOs if we have them at all, since we have no tails. In humans, the microvillar cells escaped the tiny organ and can be found ectopically all over the place in the upper respiratory system, in Peyer’s patches in the digestive system, and elsewhere whenever chemosensory work is needed, like the pituitary. There will be more about them later.
This chapter presents an electrical model of chemoreception, or chemosensory transduction. (Three more models appear as sketches in the next chapter: extracellular chemoreception and pheromone emission in some synaptic and hormonal transmissions, myelin/neural function in propagating excitatory post-synaptic potential differences (EPSPs), and ionic particle ATP-ase pumps being membrane-bound proteins functioning as linear accelerators pumping charged
EXOCRINOLOGY
191

particles against concentration gradients. These long proteins have the potential to carry information or even power as plasma conduits in much the same way as metal wires conduct electricity in 2010.)
CHEMORECEPTION AND TRANSDUCTION
The author’s electrical reception model posits a mucous-based extracellular mechanism for most chemosensory transduction. The mechanism functions much like a common gas chromatography detector, the flame ionization detector, with considerable improvements in the biology to greatly diminish identification time.
Chemoreception builds upon ordinary biochemistry. Our cells distinguish stereochemical subtleties easily, like unusual chain lengths, odds and even aliphatic chains, unusual unsaturation positions, and functional groups. We have learned already that these features distinguish pheromones.
For instance, consider enantiomers. The chemical representation for a pair of enantiomers shows two non-superimposable mirror images. Think of a pair of gloves, with a left (L) and a right hand (D for dextro-rotary). Enantiomeric gloves exhibit identical physicochemical characteristics, so chemists have had a devil of a time telling them apart down through the years. (Pasteur is famous for being the first to succeed. He noticed tiny crystals had two distinct shapes, so he sat down under the microscope and manually separated them with tweezers!) Blue algae, E. coli, paramecia, hydra, lions, tigers and bears have no difficulty at all differentiating enantiomers. We know insects recognize enantiomeric pheromones.2 Obviously, living things found out billions of years ago how to recognize enantiomers. Cells accept D-sugars while discarding L-sugars; similarly, only L-amino acids make it into proteins.3,4,5 Where does such subtle recognition take place? It takes place within the solution of the cytosol or protoplasm of the cell. If recognizing chemicals outside the cell, look first in an exterior mucus for dissolved globular protein receptors. We will see in a bit, how physiologists have been seeing just such receptors for many years,
192

without understanding. Thus our suggestion how pheromone/odorant transduction works has to be able to detect such things.
Physical logic suggests the model, too. Given that neural transmission outpaces odor transmission through air, an efficient chemo-detector would appear as near the exterior of the organism's body as other constraints allow. From economic analysis we might expect all olfactory tissues on the exterior of the organism. This was once the case, as indicated by ontogeny's recapitulation of phylogeny and comparative physiology. Microbes appear to smell only at their cellular membrane. Many modern multicellular organisms maintain exterior chemoreceptors. Witness those in the dermis of frogs or on the feet of houseflies, seastars, spiders and butterflies. Humans, too, maintain exposed chemoreceptor cells, such as those in the eyes which sense irritation to redden the eyes and stimulate irritant-induced tears. Although interior vertebrate olfactory tissues lie in permanent darkness, they maintain ancestral camouflage patterns.
Additionally, non-motile olfactory "cilia" exhibit a strange microtubular pattern in cross section. The pattern is of two tubulin tubes encircled by nine other pairs.6 This demonstrates still another link with an early exterior surface function; this time, motility. The extremely long "cilia" of mammalian olfactory chemoreceptor neurons and the short ones of microvillar cells never move. They float motionless within the Bowman's secretion of the olfactory mucosa yet they resemble real cilia that can move. Cilia first evolved on the outside surface to move the cell itself. There cilia must have gained the chemoreception and mechano-reception capabilities evidenced by modern microbes. Incidentally, the cilia only enhance chemoreception. Just as many unicellular creatures can smell without cilia, so can receptor neurons function without the reception enhancement cilia provide.
With premiums for both sensitivity and speed, concentration of odorant chemicals at the sensor might also make sense. Liquid sorbs and concentrates airborne odorants.7 This explains why receptor tissues have moist surfaces to meet smells.7 However, because liquid slows odorant movement relative to the gaseous state, detectors would be expected to lie as close to the surface of the liquid film as other constraints allow. Having receptors on the surface where the odourant or
EXOCRINOLOGY
193

pheromone molecules are caught, instead of being buried behind a mucous obstruction, makes intrinsic sense.
Cilia reach out from the microvillar cell into the liquid film, but usually not all the way to the odor laden surface, thus propagation of signal across the liquid film at this depth must be at least as quick as neural propagation or the neuron’s dendrites would simply evolve closer. Odorants need ride no glacial ferry to dendritic chemoreceptors when an electrical representation of the odorant may serve. If that were the case and odorant chemicals were carried on the backs of much larger molecules, the trouble and expense of maintaining partially depolarized dendrites would be dispensable. Partially depolarized membranes are very near EPSP neural pulse generation, so they’re easy to trigger electrically.
Dissolved proteinaceous receptors lie within the moist film covering olfactory tissues, and the dissolved receptors transmit to underlying dendrites by affecting the ion concentration and conductivity of the liquid film. Dendrites sense or produce an EPSP (excitatory post synaptic response) based upon voltage: potential difference. (Potential difference defines voltage, a bit of physics my colleagues forgot.) Thus dendrites can easily electrically sense odorant reception in the overlying sensory lymph film or receive increased electrical stimulation as free ion concentration increases. Free ion concentration increases changing the potential difference, i.e. the voltage changes, and the dendrite reception fires the axon. That may seem pretty simple, but sensory physiologists have had a devil of a time trying to understand that simplicity. They still think huge chemicals like proteins have minds and personalities so that they can recognize a pheromone, grab it, and transport it over large distances to leave it touching the cilia’s surface chemoreceptors. Never mind that chemicals can’t move against a concentration gradient sans propulsion.
DISSOLVED RECEPTOR PROTEINS ARE EXTRACELLULAR
In preparation for odorant reception, unbound dissolved protein receptors lie in the receptor lymph, slowly sequestering ions from the surrounding film of mucous.
194

In higher animals, the receptor lymph film is pumped full of strange ions of one sort or another. Once ions are captured by the sensory proteins (still called pheromone binding proteins or odorant binding proteins in the literature), reception can proceed.
Concentrated by the surface of mucous covering the non-moving "ciliated" olfactory nerve endings, the stimulator odor chemical binds loosely to the dissolved ion-holding proteins. Binding differs from permanent bonding. Binding seems more like a key in a lock while bonding would be like welding.
In 1998, when this author submitted his breakthrough article on pheromone transduction8 such dissolved receptor protein and pheromone binding had already been observed by several investigators9,10,11 without anyone noticing any conformational change in the protein, much less one that would release sequestered cations to alter the voltage and create an EPSP. Eventually, the conformational change of the pheromone binding protein and pheromone complex as hypothesized by this author was observed12 although this author’s lone prediction had gone unnoticed. It is frustrating that with the book open before them, colleagues cannot abandon the ferry protein phlogiston, a concept clearly impossible. Do they have a Charon fixation? There is no need to ferry the pheromone to the dendrite for reception. The pheromone changes the conformation of the pheromone binding protein right there at the surface and that change in shape causes release of charged particles that locally change the potential difference and that is enough. So basically, pheromone hits the PBP, the PBP changes shape, releasing both pheromone molecule for continued duty and the numerous sequestered ions, changing potential difference, setting off the dendrite electrically.
The dissolved proteins’ source appears to be cells at the base of the sensillum13 or perhaps the dendritic membrane itself.14 On binding to the pheromone, in a sudden conformational change, the receptor protein releases its previously sequestered ions and repels the pheromone away again. Think of a lock opening when the key is turned. Such conformational changes have also been observed15 and within the context of neurotransmitter reception.16
EXOCRINOLOGY
195

Similarly, small soluble proteins, such as calmodulin, bind to cations in solution.3 Anti-calmodulin drugs might be expected to counter pheromone binding proteins binding calcium. In fact, the "alpha factor pheromone" looks and behaves like a pheromone binding protein. Anti-calmodulin drugs, trifluoperazine and chlorpromazine, counter its expected effects in yeasts,17 exactly as would be expected.
Do odorant-protein complexes release any ions when the change in shape takes place (conformation change)? Lymph ionic concentration changes have been observed upon odorant stimulation.18 More details will follow.
TRANSDUCTION
The released ions themselves may be of sufficient charge and density to alter the potential difference across the nearby partially depolarized sensory dendrite. From their surfaces dendrites have a limit of about 5 or 10 microns into the surrounding medium for ionic changes to optimally effect excitation.19 Olfactory dendrites are never too far away from the chemoreceptive action.
Ion releases from a discharging pheromone binding protein (or PBP) may also encourage further ion releases from other PBPs in a chain reaction. A voltage sensitivity for ion sequestering proteins could help account for easier depolarization with prior sub-threshold excitement. In addition ion release would improve solution conductivity, varying resistance in the receptor circuit and allowing a graded flow of current, as has been observed.20 Due to their hyperpolarization, the unusually electrosensitive dendrites of chemosensory mucosae can perceive and conduct currents in the low picoamp range,21 three orders of magnitude less than other dendrites. If not for electrical reception, why else would that be the case?
It is thought that chaotic patterns recognized by the brain as scents or pheromonal signals result from the processing of the resulting neural inputs.22 Inhibitory circuits in peripheral processor centers meet incoming sequential snapshots from olfactory dendrites stimulated by ion releases from odorant-stimulated receptor
196

proteins. Higher centers detect changes (the ordinate becomes the new abscissa), with differences transmitted as recognizable chaotic patterns.
Pattern recognition may be enabled by shared capacitance switching among proximal inter-oligodendritic axons as proposed in the author’s model of saltatory conduction.
MODEL IN GREATER DETAIL:
The author apologizes for inserting so much detail in this monograph designed for practicing and training physicians. This is testimony to the language barrier that exists among those who should have already been able to follow this line of reasoning from the author’s previously published articles. The unfortunate practice of using graduate students to write one’s papers makes contacts with authors another exercise in futility, since the people who really know what is going on--the young people who have just finished the coursework--rarely communicate for the group. Of course, it never hurts to brush up, even for general readers.
Trans-specific similarities in behavior during pheromone receptivity were examined in the preceding chapter. Microeconomic theory predicts that reception of chemical signals should be essentially similar at the molecular level as well. We might expect chemoreception to vary from species to species something on a par with photo-reception (sight) or audition (hearing) systems.
Finding so many chemical resemblances among pheromone molecules (see Chapter 3) suggests that molecular mechanisms of pheromone reception should be comparable, too. Vogt and his colleagues10 have found corresponding pheromone binding protein (PBP) matches among several insect species detecting closely related aliphatic pheromones. Thus both pheromones and pheromone receptor proteins (PBPs) share similarities. For olfactory and somatic chemo-recognition the proposed receptor proteins are drawing from an entire class of so-called "carrier proteins" and antibodies. Vought and colleagues’10 narrower classification of pheromone binding proteins vary with known interspecific and intra-specific (dimorphic) variations of
EXOCRINOLOGY
197

pheromone chemistry recognition. Clones of some pheromone-binding proteins became available in 199223,24 as did an odorant binding protein.25,14 Sexual chemosensory dimorphism has been grossly observed in humans where the immune response of males and females are not identical.26 Of course more recently, dimorphic pheromone recognition27 has been broadly recognized as the physiological basis for sexual orientation, as this author first proposed in 1983.28 This author’s paper caused a sensation and outbursts of laughter--this from primatologists who had sat silent and stone-faced during films on primate copulation. While he read his paper alone, ridiculed with catcalls as he spoke, his paper was plagiarized in print in the ‘joke journal’ “The Journal of Irreproducible Results.” He who laughs last, laughs best.
THE PROCESS OF INSECT CHEMORECOGNITION
Entomologists and insect physiologists maintain a lead on their vertebrate chemosensory transduction colleagues. Part of that is because insects rarely complain much and criminals addicted to canine and feline semiochemicals see fit not to vandalize entomology laboratories. Thus the results from insect studies guide this science and color its presentation here.
Olfaction and pheromone recognition take place on the antennae protruding from the heads of most insects. Attached to the antennae surface in regular order are tiny sensory hairs. Afferent axons travel through the antenna back to the head to report any sensations.
The sensory hairs are hollow, cuticular structures, each about 300 micrometers long and 6 micrometers in diameter,29 with an inner lumen volume of about 10 to the minus twelfth liter30,31 This lumen contains the sensory dendrites, whose somatic cell bodies are buried among the epithelial cells at the base of the hair. A proteinaceous fluid, the sensory hair lymph, bathes these dendrites (see De Kramer and Hemberger [1987]32)33.
198

The electrical model actually follows much of the old mechanical model34,35,36 closely. A pheromone molecule enters the sensory hair lymph,33 a liquid holding our dissolved receptor proteins (pbps) that surrounds the receptor dendrite. Concentrated the pheromone attaches to dissolved Pheromone Binding Proteins (or PBP's) sequentially, until inactivated by sensory esterase.32 The pheromone stays in the lymph and rarely goes anywhere near the sensory dendrite32,13 because it does not need to physically go anywhere. Electricity does the work.
BIOCHEMISTRY OF PHEROMONE RECEPTION
This model predicts that emotional tears hold human pheromone receptor protein for episodic chemoreception. Like insect lymph PBP, lipid carrier-type proteins lie dissolved in the liquid Bowman's secretion covering the olfactory mucosa37 and in emotional tears.38 Their functions in tears and olfactory mucosal films are unknown. The higher proteinaceous component of emotional lacrimation brings tear protein concentrations37 to levels comparable to that of various Bowman's secretions39 and some insect sensory lymphs32 and perhaps vomeronasal mucosal secretions. In addition to five tear-specific proteins, an unusual high molecular weight protein, greater than 200 kD, resides in human tears.40 (A kiloDalton (kD) equals 1000 Daltons. One Dalton is about the size of a hydrogen atom.) They may be pheromone binding receptor proteins.
The proteinaceous secretion covering the pheromone sensing vomeronasal organ differs from the Bowman's secretion of the olfactory mucosa. A 70 kDa glycoprotein vomeromodulin is abundant in the lateral nasal glands and is also present in the posterior septal and vomeronasal glands. It is highly concentrated in the mucus of the vomeronasal organ of Jacobson but is not detectable in the mucus of the main olfactory neuroepithelium.41 This species specific "vomeromodulin" will be a pheromone receptor protein within this model, functioning as previously described.
EXOCRINOLOGY
199

Pheromone binding proteins (or PBPs) are proposed to be pheromone receptor proteins in solution. Some PBPs are known to interact with aliphatic pheromones32 while still others are specific to steroid pheromones.42
DIFFERENCES FROM CURRENT MODELS
Good evidence for G-protein mediated pheromone receptors on cell surfaces abounds (e.g. Boekhoff, I. et al.,43). However excellent counter evidence against a transduction role for G-proteins exists, at least for insects:
Female sex pheromones applied to freshly isolated, living antennae [one antenna, two antennae-BN] of male Antheraea polyphemus and Bombyx mori [a big silk moth and the silk mothBN] led to an increase of cGMP. A 1:1 mixture of 2 pheromone components of Antheraea polyphemus blown for 10 sec. in physiological concentrations over their antennal branches raised cGMP levels about 1.34fold (+/ 0.08 SEM, n = 23) from a basal level of 3.0 +/ 0.6 (SEM, n = 20) pmol/mg protein. Similarly, bombykol [the pheromone for the silk moth, Bombyx mori, identified back in 1959-BN] elicited a 1.29fold (+/ 0.13 SEM, n = 23) cGMP increase in antennae of male Bombyx mori from a basal level of 2.7 +/ 0.5 (SEM, n = 24) pmol/mg protein. No cross-sensitivity was found with respect to pheromones from either species [In line with their not responding to each others pheromones under the EAG. -BN]. In antennae of female silk moths, the cGMP response was missing upon stimulation with their own respective pheromones according to the known lack of pheromone receptor cells in the female. cAMP levels in the male antennae of 14.2 +/ 2.9 (SEM, n = 4) pmol/mg protein in A. polyphemus and 15.0 +/ 3.0 (SEM, n = 5) pmol/mg protein in B. mori were not affected by pheromone stimulation. Within 160 sec, the extent of cGMP increase in B. mori was independent of the duration of pheromone exposure. The levels of cGMP in pheromone-stimulated antennae of both species remained elevated for at least 10 min., i.e., much longer than the duration of the receptor potential measured in single-cell recordings (Ziegelberger, G., et al., 1990).44
The G-protein pathway may have something to do with the metabolism of receptor cells,45 but reaction-time and persistence outlive transmitted results of the stimulus.
Previous models classified pheromone-associated proteins dissolved in the receptor lymphs, tears, and Bowman's secretions as "carrier proteins.”46 Supposedly pheromonal lipids attach to proteins for transport through the aqueous film barrier to the receptor dendrite. On the dendrite, it has been thought, surface membrane-bound protein binds to pheromone which opens a hole in the membrane. The tiny hole, or channel, allows ions inside to depolarize the dendrite. Similar chemically signaled channels have been demonstrated for virtually all cells with reliable patch-clamp
200

techniques.47 However, experimental difficulties have prevented identification of protein-bound, odorant-stimulated channels in olfactory dendrites under the sensory lymph film. The entire complement of membranous proteins for olfactory cilia have been separated and are being sequenced.48 This model accommodates dendritic receptors for chromatographic diffusion timing of odorants through the film.
Poor evidence for such a carrier function for PBP does exists.42 A portion of lipid pheromone adhering to glass can be coaxed off by adding some protein to the brew. 13 This is much like using "enzymes" in laundry detergent. Solubility does improve, but only at unrealistically high concentrations. The same lab saw an impoverished pheromone reception signal set in a dendrite surrounded and wetted by a protein-less pheromone/ionic blend. That seems to be indisputable evidence for at least some dendritic surface receptors. But does it? Proteins of this class can turn over in the receptor lymph at a rate of 2 X 104 proteins/second/sensillum.9 Indeed, mRNA specific to ciliary membrane encodes a secretory protein with significant homology to odorant binding protein (OPB). It shows significant homology to the VEG protein, thought to be involved in taste transduction. The cloned membrane protein is a member of the lipophilic molecule carrier protein family.14 If the membrane does not secrete receptor proteins, some sorption of PBP to those rough dendritic surfaces seems probable.
Finally, without any PBP, an excessive 10,000 pheromone molecules were needed at the dendritic surface to fire the neuron. Recall that 99.96% of the pheromone becomes attached to PBP. With PBP concentration at physiological levels, calculation shows that under the mechanical scenario, 25 million pheromone molecules would be needed in the lymph to fire dendritic receptors. Boding ill for the ‘ferry’ idea of old, only one or a few molecules of pheromone stimulates a transduction event.32 Oops!
Getchell and Getchell,49 dispute the practical necessity of carrier proteins. They note the improbability of gaining any speed by attaching most tiny odorants to large slow-moving proteins. Calculating diffusion coefficients through mucous they show that carry time would exceed the speed of chemoreception by several orders of
EXOCRINOLOGY
201

magnitude. An exposed receptor dendrite fires sooner than a carrier protein can cross the five to ten microns from the mucous surface to the dendrite wall.
The long time periods ranging from 5.4 to 16 seconds, required for OBP to transport odorants from the mucous surface to receptor sites on the proximal cilia are, in general, not consistent with the onset latencies of odorant-evoked excitatory discharges (Getchell, 1986) or membrane currents (Firestein and Weblin, 1989) recorded from olfactory receptor neurons (Getchell, T.V. and M.I. Getchell, 1990b).48
If the proteins hypothesized here to be receptor proteins are carrier proteins as is accepted thinking, then they do a terrible transportation job. Not only do they move too slowly to cover the required distances as indicated above, but 99.96% of pheromone molecules stay in and among the dissolved proteins of the receptor lymph and never reach the dendrite surface. 13
Van den Berg and Ziegelberger13 note yet another failure of the mechanical hypothesis. "Ferried pheromones" remain in the lymph several minutes after the electrical signal ceases.50 These difficulties along with a clearly inefficient mechanism, pose insurmountable problems for the mechanical model, yet still it holds on, inertia from the nineteen seventies.
The present new hypothesis satisfies all of these difficulties, although as Max Planck so elegantly put it, we must wait for all the old men to expire. If pheromone receptor proteins change conformation to release cations (or anions in some cases) on binding to pheromone and sequester more only upon disassociation, then strongly bound pheromone/protein pairs would not contribute to further stimulation. This accounts for the signal latency and the intact pheromone presence observations.13
Thinking of this idea took up the author’s first 20 minutes study of the subject of chemoreception (and the difficulty his colleagues have with it baffles him even today). Half a dozen tests jumped to mind that would support or settle the issue. A literature search indicated that every one of those tests had been done, often elaborately, with results as predicted usually to the surprise of investigators. Outsiders find it easier to notice the obvious.51
202

The 99.96% of pheromone perpetually "in transit" can be explained better by the new model. Since the "carrier proteins" are actually "receptor proteins" under this new electrical model, 99.96% represents an efficiency, not a deficiency.
Meanwhile, a carrier role for fetching pheromones through mucous can also be discounted by the kinetics associated with pheromone and pheromone binding protein.
[T]here is a temporally weak association between individual binding protein and pheromone molecules. Even at 109 M pheromone, a 10,000 fold excess of PBP was required to reduce the availability of pheromone to another protein species, the SE [sensory esterase which digests pheromone for removalBN] (Vogt, R.G. 1987).32
If the pheromone rider won't stay on the PBP horse, the rider is not being carried.
The preceding observation by Vogt32 explains the signal amplification observed by one or a few molecules setting off a reception event from a much larger structure, the sensory dendrite. A pheromone molecule binds to many PBPs in quick succession.32 Because ion release is so much faster than sequestration, depolarization (or hyperpolarization depending on the charge of the ions sequestered) could proceed from the rapid buildup of de-sequestered ion concentration changing the relative potential difference across the nearby dendritic membrane.
Getchell21 noticed "false positives" leading sensory dendrite firing responses to some odorants. Such would be consistent with the new electrical method of initial excitation. Where positives do not lead, increased lymph conductance by released ions could let current outrun a buried "false" response.
The false positive phenomenon was described as a small-amplitude initial positive voltage transient.21 Such small positive voltage transients would be the residues of actual binding protein ion releases, rather than the much larger responses of neural origin.
Getchell's calculation,48 observed sensory latencies, and known physical location of pheromone suspended in lymph support the new model. Because pheromone triggers cation release in the lymph that bathes and surrounds the
EXOCRINOLOGY
203

dendrite, stimulus and receptors join in plenty of time. Having receptors effectively at the receptor surface and immediately beneath it, instead of being buried behind an aqueous mucous barrier when most pheromones for most species are hydrophobic, does make intrinsic sense. Otherwise would not faster-moving hydrophilic pheromones dominate the semiochemical lexicon eliminating the ‘need’ for carrier proteins in the first place?
The puzzle of concentration dependent onset latency is explained by the present model. The electro-olfactograph, or EOG, is similar in concept to the electro-antennagraph previously described in chapter 3 of this book. The EOG is characterized by a concentration dependent onset latency.21 This model explains varying slack time nicely. Contained within the mucous or sensory lymph we find binding proteins such as OBP, PBP, or antibodies. In the mucous/sensory lymph, bound binding proteins release ions on contact with odorant. With more odorant sorbed into the mucous, more binding proteins are bound at the beginning, therefore the firing thresholds of nearby dendrites are attained sooner. Hence a concentration dependent variation in onset latency becomes observable. There is no way to explain concentration-dependent variation in onset latency in the old model.
Another riddle is the definable threshold observable in EOGs for different odorants or pheromones.21 The threshold is that of the odorant concentration, not the electrical threshold of the dendrite--that stays constant. This model predicts that such odor concentration thresholds should vary widely for different odorant species. Such is the case.52 Variability would follow binding affinity, mainly, but other chromatographic variables include miscibility with the mucous, diffusion rate through the mucous, ion-odorant binding affinity (polarity), and others. Since the old model holds that the pheromone binding protein or odorant binding protein does all the swimming (across the receptor lymph to release the odorant or pheromone at the dendrite), variability due to chromatographic and intrinsic chemical variables would play no or almost no role.
The preceding discussion utilizes the concept of concentration. Actually, olfactory tissues measure, not concentration, but something closely related. The information transferred to the brain is that of mass flow rate (dm/dt), unaffected by
204

molar concentration (dn/dv) (see Mozell, et al., 198453 and Kurtz, D.B. and M.M. Mozell, 198551). Some chromatographic detectors detect mass flow rate. The mass flow rate detecting flame ionization detector (or FID) responds to ionization released by burning a passing sample and is unaffected by concentration fluctuations just like the case in olfaction. It should not be surprising that peripheral olfactory systems respond to mass flow rate if they rely on similar ionic triggers as hypothesized.
Holding total moles constant and varying carrier gas volume with proportional increases in stimulation time, researchers showed constant stimulation.51 When total moles are held constant, mass is held constant, too. Proportionality constants are required in both cases (olfaction and gas chromatography) to account for odorant-specific chromatographic variables. The corresponding gc measure then, is mass flow rate, which also does not depend upon carrier gas volume either.54
IONIC CONCENTRATION
Extra-cellular depolarisation by released ions is the mechanism of ordinary pain sensation on cell trauma. Escaping cell contents (K+ ions) change potential difference at exposed pain receptors.55 (24) In odourant or pheromone reception, the ions are released outside the cell by desequestering receptor proteins on binding to odorant aligning polar molecules and setting up the depolarising oscillation.
In ion substitution experiments, one ion substitutes for a naturally occurring lymph ion concentration. In vitro and in vivo these hold Na+ and K+ ions primarily responsible for negative voltage transients recorded from the olfactory mucosa. Oddly enough, olfaction is tetrodotoxin (TTX) insensitive, and relatively unaffected by local anesthetics at concentrations high enough to block action potentials in the olfactory nerve!21 The author proposed that an evaginated ion permeability of the ciliary dendrites could explain this phenomenon in insect sensilla in his article on transduction a decade ago. The logic went that on the antenna’s dendrites the negatively charged "exterior" of Na+ channels would be interior, hence unavailable to TTX (and TTX-insensitive). Local anesthetics block normal membrane receptor dendrite stimulation not signal propagation. Apparently the ion-bloated binding
EXOCRINOLOGY
205

proteins "bursting" on odorant contact in the mucous could stimulate even a receptor-less dendrite. The poor old pbp ferry model just throws up its hands in abject failure, again. Recently, some membrane structures have been identified to be reversed from what is considered normal.56 Well, what do you know? Another correct prediction ignored.
Electrical transients can only be recorded from epithelial regions of the nasal chamber containing olfactory mucosa.21 Only such mucosal areas harbor Bowman's glands, secreting the dissolved odorant binding proteins amid unusual cation concentrations.
Transients of the olfactory mucosa cannot be evoked by antidromic (against flow) electrical stimulation of the olfactory nerve.21 Olfactory nerves, plain C-fibers, lack the myelin associated with one-way-only axons. Some dendritic cilia of the olfactory mucous may have evaginated excitability as the following photo suggest. An odd constriction between the dendrite and the soma, called the ciliary root,31 may be the membrane twisting inside-out. However, Russian researchers57 propose a shunt circuit to account for this phenomenon.
PHASE ANGLE
Most of the new crop of researchers accept previous researchers’ work uncritically. Where theretofore unheard of phenomena are detected, it is merely ignored as irrelevant. Getchell21 cites one of these, another puzzle explicable by the tired old odorant-ferrying model.
Gesteland and his colleagues studied differential impedance changes by continuously passing a sinusoidal current [2050 cycles (cps)] to the olfactory mucosa through an injection electrode. When odorants were delivered to the olfactory mucosa, the negative-driving process was associated with a change in the phase angle that tentatively was identified with an inhibitory process.21
The phase angle indicates the direction of a vector sum of capacitive reactance and inductive reactance, with resistance. A purely resistive alternating current circuit has a phase angle of 0o. Only physical changes in resistance or capacitance* during
206

the course of the cycle could explain these data. Binding and releasing ions in the aqueous mucous would change its resistance and deliver the responses found. Indeed, changes in lymph resistance on odorant stimulation have been shown in insects54
*or inductance Physiologically, there is nothing in the peripheral olfactory sensory system resembling a solenoid, so perhaps inductance can be ignored as it has been.
Others have shown that changes in potassium cation concentration in the sensory lymph follow pheromonal stimulation of the receptor.
"A high potassium content of about 200 mM has been measured in isolated receptor lymph (Kaissling and Thorson, 1980) and also in situ by Xray micro-analysis of ultrathin cryosections (Steinbrecht and Zierold, 1982, 1985, 1987). The later method allows the quantitative analysis of electrolyte elements at the resolution of the electron microscope, and preliminary data indicate that changes in the electrolyte concentrations due to stimulation of the receptors can be monitored (Kaissling and Thorson, 1980; Steinbrecht and Zierold, 1985; R.A. Steinbrecht and K. Zierold, unpublished results)" (Steinbrecht, R.A., 1987). (BN emphasis)
Does this observation cinch the question? More evidence supports pheromone binding protein to be a receptor protein. De Kramer and Hemberger31 demonstrated protein sequestration and de-sequestration of cations to equilibrium with test electrodes definitively and elegantly. (Please refer to the graph of R1 vs. recording time, in figure below.)
EXOCRINOLOGY
207
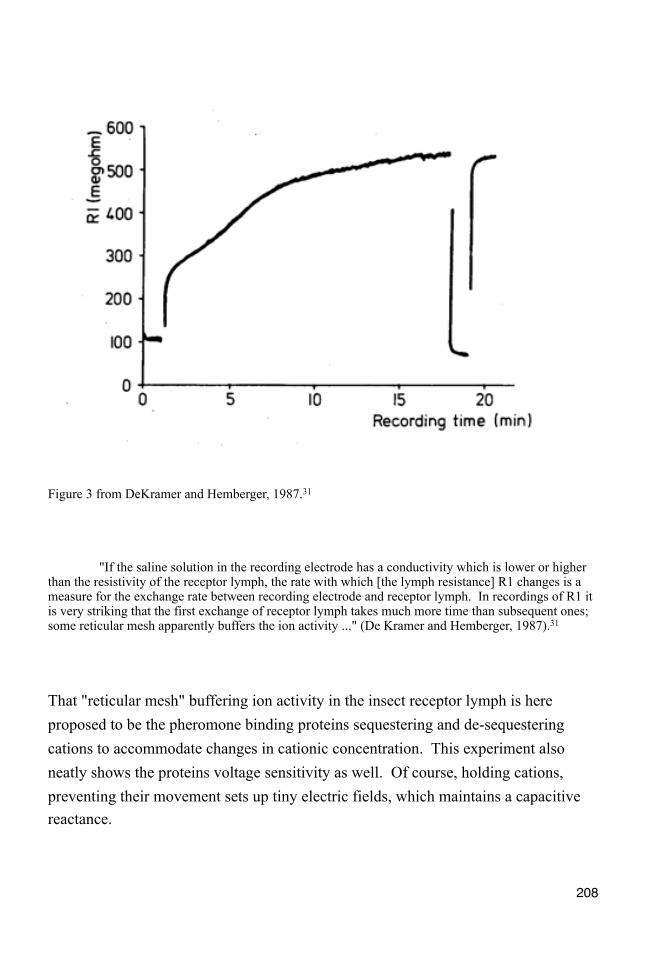
Figure 3 from DeKramer and Hemberger, 1987.31
"If the saline solution in the recording electrode has a conductivity which is lower or higher than the resistivity of the receptor lymph, the rate with which [the lymph resistance] R1 changes is a measure for the exchange rate between recording electrode and receptor lymph. In recordings of R1 it is very striking that the first exchange of receptor lymph takes much more time than subsequent ones; some reticular mesh apparently buffers the ion activity ..." (De Kramer and Hemberger, 1987).31
That "reticular mesh" buffering ion activity in the insect receptor lymph is here proposed to be the pheromone binding proteins sequestering and de-sequestering cations to accommodate changes in cationic concentration. This experiment also neatly shows the proteins voltage sensitivity as well. Of course, holding cations, preventing their movement sets up tiny electric fields, which maintains a capacitive reactance.
208

Is potassium the ion bound up in the lymph? DeKramer and Hemberger31 reviewed the composition of the sensory lymph filling insect sensilla. They drew attention to the difference between total and free K+ (potassium cation) concentrations citing Kaissling and Thorson (1980). That lab had used Xray and flame photometric spectroscopy to confirm concentrations of 200 mM K+ (free and bound) while the K+ activity was only 145 mM. A high potassium concentration in sensillum lymph was later confirmed by Steinbrecht (1992). Potassium pumps into the receptor lymph would be required for the model and these, too, have been observed:
Another important feature of the auxiliary cells [at the base of the sensory hair lumenBN] is their apparent resemblance to the cells of ion-transporting epithelia (Fig. 14), e.g., in insect midgut and salivary glands (Harvey, 1980). Their elaborately folded apical membranes are thought to be the site of an electrogenic cation pump, which mainly pumps potassium into the receptor lymph and thereby creates a trans-epithelial voltage. ... In Bombyx mori the trans-epithelial voltage amounts to 33 mV (Thurm and Wessel, 1979), but its influence on receptor function is still uncertain" (Steinbrecht, R.A., 1987).
Once pumped into the lymph, K+ (potassium) ions must be contained. Holding the evaginated ion concentration requires an electrically insulated container and the hollow sensory hair is one.
Tight-junctional complexes join the epithelial cells and neuronal cell bodies, establishing a high resistance barrier between the sensory hair lymph and the hemolymph (see De Kramer and Hemberger [1987]) (Vogt, 1987).32
A strong electrical resistance exists between sensory hair lymph of the sensory hair (or sensillum) and the hemolymph, or insect "blood."
"Beside their ontogenetic function, the auxiliary cells connect the receptor cells to the epidermis so that the epithelial organization is maintained. Septate junctions seal the intercellular clefts between all cells so that the trans-epithelial resistance is high. In pheromone-sensitive sensilla of moths, but not in mechanoreceptors of crickets and flies, septate junctions are also observed where the axon originates and the thecogen cell borders the glia cell around the axon (Keil and Steinbrecht, 1983, 1987; Steinbrecht and Gnatzy, 1984). A very close contact between the tormogen cell and the cuticle of the hair base is the morphological correlate of a high electrical resistance between neighboring sensilla (Keil, 1984c)" (Steinbrecht, R.A., 1987).
EXOCRINOLOGY
209

Next, a sink for K+ ions (or a source of Na+) separate from the sensory hair lymph and accessible to the interior of the dendrite might be required. Might this be the work of the "specialized junction of hitherto unknown function" between thecogen cell and receptor cell identified by Steinbrecht in 1980 (cited in Steinbrecht, R.A., 1987)? Such an open hole could do the job. Indeed, Kuppers, Josef and Ulrich Thurm (1979) observed an inward equilibrium continuous current of K+ ions flowing like a bucking voltage (see figure from Basic Gas Chromatography) and a second loop of current.
A second loop of current circuit is controlled by adequate stimulation of the sensory cell (Thurm, 1974). The actively generated current flows back in this circuit [Actually, the continuous bucking current is the reverse flow. It helps account for the high electro-sensitivity of these dendrites-BN], entering the receptor cell via its stimulus-increased apical conductance, leaving the cell via its baso-lateral membrane regions, and returning into the tormogen cell via its basolateral membrane. . . . This second current loop in some sensilla increases receptor sensitivity (Erler and Thurm, 1978; Thurm and Gödde, in prep.). [Actually, the bucking current increases the sensitivity, just like in GC detector circuitry-BN]
210

“It is helpful to think of the electrode gap as a variable resistor R1 whose resistance value is determined by the number of charged particles within the gap. With carrier gas flowing a constant concentration of charged particles (due to carrier gas impurities, liquid phase bleeding, etc.) will be present in the gap, thus causing a constant current, I, to flow. This constant current is called the “background current”. It is desirable to minimize the background current so small changes in current can be more easily measured (it is easier to measure a fixed change in a small quantity than the same change in a larger quantity). The background current is reduced to zero by opposing it with a “bucking voltage”. Thus, under “no signal” conditions (carrier gas flow), no net current flows, and the recorder traces a straight baseline. When a sample component passes through the electrode gap, molecules of the component are ionized. This increases the number of charged particles and decreases the value of R1. This decrease permits current to flow which produces a signal which is registered as a peak on the recorder.”
EXOCRINOLOGY
211

The extreme sensitivity of the dendrite to polarization by very slight currents in the picoamp range is analogous to the homeostasis of the background current against the bucking voltage in the flame ionization detector of gas chromatography. The few ions released or exposed by the conformational change of the pheromone receptor proteins in the ‘gap’ are sufficient to set off the chain reaction among the calmodulin proteins inside the dendrite electrically. In some insects with evaginated membranes and ‘pumped’ receptor lymph, ionic concentration just beneath the threshold of conductivity until PBP conformational change allows the passage of an electric ‘spark‘ explains the electrical characteristics of antennae.
There are certainly plenty of voltage gated sodium channels in olfactory neuron dendrites58,59 and voltage sensitive calmodulin is stuffed into dendrites of pheromone receptor dendrites to amplify depolarization.60
Not all chemosensory antennae work as insect antennae do. The electrical model of chemosensory transduction accommodates these, too.
b) Terrestrial Crustacea and Spiders. Measurements on representatives of Crustacea and spiders revealed electrical properties of their sensilla clearly different from those found in insects. At tarsal and antennal chemo and mechanoreceptive sensilla of the isopod Armadillidium (woodlouse) a trans-epidermal voltage in the range of +15 to +35 mV exists. This voltage is not acutely O2-dependent; no voltage change at all occurs during 6 minutes of anoxia. The TEP [trans epithelial potential] at sites far from sensilla is only a few mV positive. The nature as well as the location of the epidermal voltage source at these sensilla remain unknown at present. ... (Thurm, U. and G. Wessel, 1979).61
Apparently, woodlice have PBP (pheromone binding protein), but lack the (O2 sensitive) potassium pumping. PBP would gather ion from the relatively low K+ concentration of the hemolymph, resulting in relatively long latency periods when compared to insects. Perhaps inferior chemo-sensing ability limit these organisms to their more sedentary life strategies?
Unlike the mechanical case, for the electrical model, a simultaneous excitation of the entire dendrite would take place on chemo-stimulation. If a mechanical dendrite receptor depolarizes the neuron, the point of excitement would be detected by examining firing latencies.
212
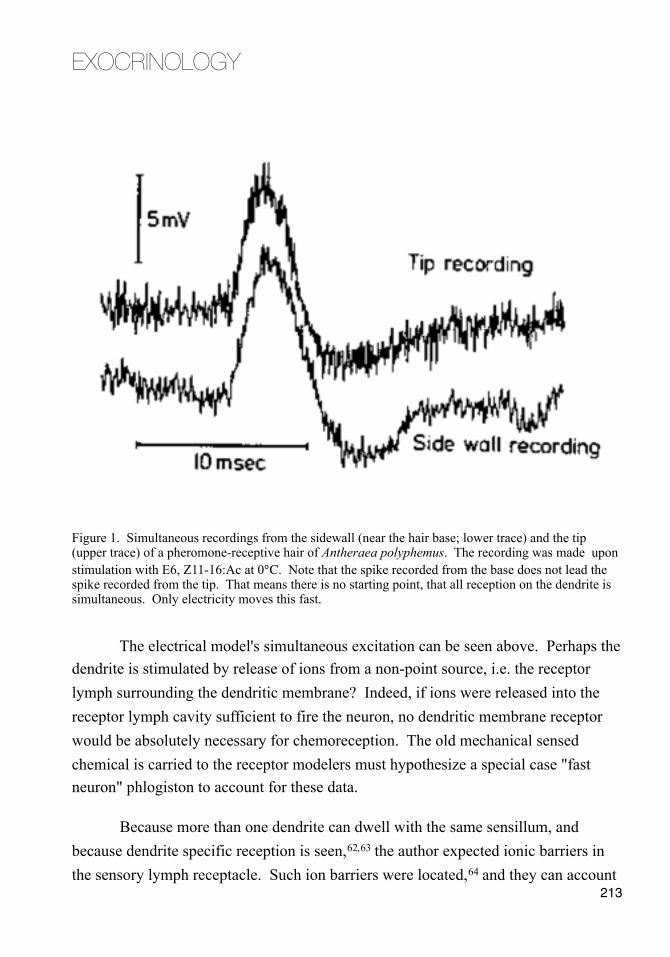
Figure 1. Simultaneous recordings from the sidewall (near the hair base; lower trace) and the tip (upper trace) of a pheromone-receptive hair of Antheraea polyphemus. The recording was made upon stimulation with E6, Z11-16:Ac at 0℃. Note that the spike recorded from the base does not lead the spike recorded from the tip. That means there is no starting point, that all reception on the dendrite is simultaneous. Only electricity moves this fast.
The electrical model's simultaneous excitation can be seen above. Perhaps the dendrite is stimulated by release of ions from a non-point source, i.e. the receptor lymph surrounding the dendritic membrane? Indeed, if ions were released into the receptor lymph cavity sufficient to fire the neuron, no dendritic membrane receptor would be absolutely necessary for chemoreception. The old mechanical sensed chemical is carried to the receptor modelers must hypothesize a special case "fast neuron" phlogiston to account for these data.
Because more than one dendrite can dwell with the same sensillum, and because dendrite specific reception is seen,62,63 the author expected ionic barriers in the sensory lymph receptacle. Such ion barriers were located,64 and they can account
EXOCRINOLOGY
213

for much differential dendritic receptivity. In other cases, different PBPs one for each pheromone sensed in the sensillum,10 might fire sequentially, with the second PBP2 being voltage dependent upon previous PBP1 de-sequestration/conductance. This is life, so there will be great variability, but all variations on the same theme.
The threshold for impulse initiation to depolarizing current pulses for olfactory dendrites is in the pico-amp (pA) range. "These are very low current densities for impulse initiation".21 Yet the currents generated by these nerves are no different from those of most other nerves and occur in the milliamp range. The electrical model of chemoreception transduction accommodates the extreme sensitivity to electric shock shown in ciliary olfactory dendrites. Super-sensitivity allows detection of small changes in local ion concentrations in the sensory mucous. Local ion concentration changes, as hypothesized, have been observed.54,18 Q.E.D.
Van den Berg and Ziegelberger's data13 prove that the few receptors they propose to be on dendritic membranes cannot alone account for chemoreception. They found that a 180 nMolar concentration of pheromone in ringer solution (or about 10,000 pheromone molecules) was necessary to initiate a response vs. zero for ringer. With pheromone binding 99.96% to carrier protein, then 25 million in the lymph would be equivalent. Pheromone uptake efficiency means an exposure to about a billion pheromone molecules would be needed to get a response from dendritebound receptors alone. However, only a few molecules set the receptor off.
Getchell's calculation (1988), observed sensory latencies, and known physical location of pheromone suspended in lymph support the new model. Because pheromone triggers cation release in the lymph, stimulus and receptors join in plenty of time. Having receptors at the receptor surface, instead of being buried behind a mucous barrier, makes intrinsic sense.
CONFORMATIONAL CHANGES IN RECEPTOR PROTEINS
What about predictions of the new model? For instance receptor proteins are proposed to have a conformational change to release or expose ions to effect a change
214

in potential difference, voltage, across the pheromone receptive dendritic membrane. Does pheromone binding (receptor?) protein have two conformations in vivo? Yes it does.
In the early sixties, Maxwell M. Mozell65 first noticed an odd phenomenon in fresh Bowman's secretion both in vivo and in vitro. He noted a change in the viscosity of the secretion with odorant stimulation. It was this remarkable observation that stimulated submission of this model for publication in 1998.
You can see the binding proteins changing their conformations! At X60 magnification, the mucus surface changes its reflectance.66 This is straightforward evidence for conformational change. Mozell's viscosity changes in chemo-stimulated mucous64 were visible to the unaided eye.
These characteristics might be expected upon conformational changes of some odorant binding protein. In fact different conformations have even been distinguished using electrophoretic techniques:
In acryl-amid gels under nonreducing conditions, the binding protein appears as a double band (Kaissling et al., 1985; Klein, 1987; 32); the two populations might reflect two different functional states. They might differ in their disulfide bridges, since the binding protein contains six conserved cysteins, where more than one stable tertiary structure seems possible (Gyrgyi et al., 1988; Raming et al., 1989; Raming et al., 1990). It is conceivable that the binding [...] induces a conformational change [...] of the involved pheromone binding protein (van den Berg, M.J., and G. Ziegelberger, 1991). 13
Such conformation changes, which have finally been admitted,67 amaze the scientific establishment, those adherents of the old mechanical chemical ferrying to dendrite receptor model. It is too bad that they did not notice the author’s article.8
Another characteristic of the new model is a changing electrical conductance on odorant stimulation. Does resistance change in the sensory lymph (Bowman's secretion, tears?) Apparently so. A Russian team found a changing electrical resistance in insect sensilla upon odorant stimulation,54 essentially similar to the R1 of the Flame Ionization Detector of the gas chromatography, leaving a logical consequence of the release of sequestered ions upon binding of receptor proteins.
EXOCRINOLOGY
215

The electrical ability to affect depolarization of a membrane is distance dependent. Sufficient potential differences to depolarize must be within about 5 to 10 microns of the dendritic membrane.19 These measures typify the width of sensory lymph cavities within the sensillum, or sensory hair.31
DIMORPHIC RECEPTOR PROTEIN AND GLANDULAR
MORPHOLOGY
If pheromone binding proteins (PBPs) and odorant binding proteins (OBPs) really are pheromone receptor proteins and odorant receptor proteins as hypothesized, respectively, then PBPs must bind only to one pheromone, while OBPs must bind to many odorants. PBPs must differ in their receptor area enough to account for that specificity while OBPs should remain very similar. Both PBP and OBP would have to explain their ability to sequester cations such as maintaining a negative charge. All three conditions are met.
Odorant binding protein subfamilies associate with distinct classes of olfactory receptor neurons in insects.10 Vogt's group10 compared fourteen OBPs and PBPs from six species. They found PBPs to be highly variable in their sequences when compared among species. In addition, sex specific PBPs were found in species with sexually dimorphic pheromone discrimination. OBPs were found in both sexes in general odor discrimination sensilla. The highly negative charge of PBPs found was consistent with the pheromone ion sequestration and release function here proposed. Discovery of highly positively charged PBPs, for sequestration of anions, might plausibly be expected as well.
What information have we concerning the human case? A sexual dimorphism in human vomeronasal organ response has been recorded for identical putative pheromone stimulants.68 Sexual dimorphisms have been detected in adult human tears69 and in gross immune function.70 There were significant differences in the amounts of LF [lactoferrin] and two kinds of STPs [of five specific tear proteins] in the different sexes. The amounts of these proteins were larger in females.66
216

Dimorphic pheromone receptor proteins have been discovered in human tears and in pheromone emitting glands.71 Some relationship must exist here. Differences in reception reflect in differences in pheromone secretion. Sexual dimorphisms have been detected in dermal sebaceous secretion,72 and axillary secretions.73,74
EXOCRINOLOGY
217

Is it logical that nature would emplace an aqueous barrier to a hydrophobic
pheromone requiring a larger carrier protein to slowly traverse? Kinetic studies
suggest this is impossible.
No such to and fro P-PBP traffic has ever been observed, despite numerous attempts
and approaches.
The association between pheromone and pbp is too short by orders of magnitude to
allow any transport.
Simultaneous excitement of tip and base by EAG prevent entertaining this point-
source step.
(The use of bovine serum albumin (BSA) as a control for PBP is invalid because
BSA is itself a chemosensory protein, changing conformation on contact just like
PBP.)
218

Given the availability of electric communication within the body elsewhere (e.g. pain
reception on cell trauma releasing K+ ions) would not the observed voltage changes
resulting upon PBP conformational change be more easily accommodated as part of
the dendritic excitement process? After all, the sensory lymph is only as wide as the
limits of electrical excitement of a dendrite, any dendrite. Also, why else would the
dendritic membrane be maintained in a constant state of hyperpolarization if not as a
bucking voltage? Why would that membrane be so electrically sensitive, responding
to electric currents in the picoamp range? The phenomenon of "false positives"
correlates well to individual PBP conformational changes at the surface of the sensory
lymph.
If this is not a simple electrical excitation, why is the system so insulated from the
hemolymph? Why so electrically insulated from other identical systems adjacent?
Why does the lymph conductance change on odorant stimulation? The change in
potassium cation concentration observed on pheromone stimulation defines voltage
change. By calculation, the change is [K+] is enough to stimulate electrically.
EXOCRINOLOGY
219

High social standing among one's colleagues should not be sufficient to drive a
fraudulent scientific inertia, a driven mass hysteria. Thousands of scientists with
limited resources are being allocated to a solved conundrum, depriving society of
their valuable services.
220
1 Nicholson, B. 1985; An economic model of phermone transmission in avians. Paper presented to the Animal Behavior Society annual meeting, Raleigh, N.C.
2 Tumlinson, James H., and P.E.A. Teal 1987; Pheromone Biosynthesis and its regulation. pp 3-26. Chapter 1. In: Pheromone Biochemistry. Prestwich, Glenn D., and Gary J Blomquist editors. Orlando: Academic Press, Inc. 565 p.
3 Stryer, Lubert 1988; Biochemistry, Third Edition. New York: W.H. Freeman and Company. 1089p.
4 Lehninger, Albert L. 1982; Principles of Biochemistry. New York: Worth Publishers, Inc. 1011p.
5 Champe PC, Harvey RA. Lippincott's Illustrated Reviews: Biochemistry. Philadelphia: JB.Lippincott Company 441p. 1987.
6 Burkitt H George, Young Barbara, Heath John W, Wheater Paul R Wheater’s functional histology: a text and colour atlas. New York: Churchill Livingstone, 1993.
7 Hornung DE, Mozell MM. Factors influencing the differential sorption of odourant molecules across the olfactory mucosa. J Gen Physiol 1977;69(3):343-61.
8 Nicholson B. Pheromones cause disease: pheromone/odourant transduction. Med Hypotheses. 2001 Sep;57(3):361-377.
9 Vogt, RG, A.C. Kohne, JT. Dubnau and G.D. Prestwich 1989; Expression of pheromone binding proteins during antennal development in they gypsy moth Lymantria dispar Journal of Neuroscience. 9(9):3332
10 Vogt, RG, G.D. Prestwich and M.R Lerner 1991; Odorant-binding-protein subfamilies associate with distinct classes of olfactory receptor neurons in insects. Journal of Neurobiology. 22(1):74-84.
11 Bocskei Z, Groom CR, Flower DR, Wright CE, Phillips SE, Cavaggioni A, et al. Pheromone binding to two rodent urinary proteins revealed by X-ray crystallography. Nature 1992;360(6400):186-188.

EXOCRINOLOGY
221
12 Damberger Fred, Nikonova Larisa, Horst Reto, Peng Guihong, Leal Walter Soares, and Kurt Wüthrich. NMR characterization of a pH-dependent equilibrium between two folded solution conformations of the pheromone-binding protein from Bombyx mori. Protein Science (2000), 9:1038-1041.
13 Steinbrecht, Rudolf Alexander 1992; Experimental morphology of insect olfaction: tracer studies, X-ray microanalysis, autoradiography, and immunocytochemistry with silkmoth antennae. Microscopy Research & Technique 22(4):336-350.
14 Dear TN, Campbell K, Rabbitts TH. Molecular cloning of putative odorant-binding and odorant-metabolizing proteins. Biochemistry 1991;30(43):10376-82.
15 Van den Berg MJ & Ziegelberger G (1991) On the function of the pheromone binding protein in the olfactory hairs of Antheraea polyphemus. J Insect Physiol 37: 79-85.
16 Kanner, Baruch I. (1993). Structure and function of Sodium-coupled neurotransmitter transporters. pp. 243-250, Chapter 19 IN: Molecular Biology and Function of Carrier Proteins Society of General Physiologists Series, Vol 48, 46th Annual Symposium (Reuss, Luis, J M. Russell, JR, and Michael L. Jennings, eds) New York: Rockefeller University Press 324 pages.
17 Ruiz, T., and L. Rodriguez 1986; Effect of anticalmodulin drugs on the action of yeast alpha factor pheromone. Archives of Microbiology 145(1):104-106.
18 Steinbrecht, Rudolf Alexander 1987; Functional morphology of pheromone-sensitive sensilla. pp 353-384. In: Pheromone Biochemistry. Prestwich, Glenn D., and Gary J Blomquist, (eds) Orlando: Academic Press, Inc. 565 p.
19 Ranck, James B., JR 1987; Brain stimulation, electrical, in mammals, principles and consequences P. 170-171. In Encyclopedia of Neuroscience. George Adelman ed. Boston: Birkhauser.
20 Krapitskii, S.V., and F.G. Gribakin 1992; Electroantennogram of the American cockroach: effect of oxygen and an electrical model. Journal of Comparative Physiology-A, Sensory, Neural, & Behavioral Physiology 170(5):651-663.
21 Getchell TV. Functional properties of vertebrate olfactory receptor neurones. Physiol Rev 1986;66(3):772-818.
22 Freeman WJ. The physiology of perception. Scientific American 1991 Feb;264(2):78-85.
23 Raming K, Krieger J, Breer H. Molecular cloning of an insect pheromone-binding protein. Febs Lett 1989;256(1-2):215-8.
24 Krieger J, Raming K, Prestwich GD, Frith D, Stabel S, Breer H. Expression of a pheromone-binding protein in insect cells using a baculovirus vector. Eur J Biochem 1992;203(1-2):161-6.
25 Dal, Monte M., I. Andreini, R. Revoltella, P. Pelosi. Purification and characterisation of two odourant-binding proteins from nasal tissue of rabbit and pig. Comp Biochem Physiol [B] 1991;99(2):445-51.
26 Grossman CJ, Roselle GA, Mendenhall CL. Sex steroid regulation of autoimmunity. J Steroid Biochemistry & Molecular Biology 1991;40(4-6):649-59.

222
27 Bekaert KM, Tuyttens FA, Duchateau L, De Brabander HF, Aluwé M, Millet S, Vandendriessche F, Vanhaecke L. The sensitivity of Flemish citizens to androstenone: influence of gender, age, location and smoking habits. Meat Sci. 2011 Jul;88(3):548-52. Epub 2011 Feb 18.
28 Nicholson B. A semiochemical addiction model for human bonding. Paper presented to the Animal Behaviour Society and the International Society for Human Ethology biannual meeting, Bucknell University Lewisburg, PA. June 12, 1983.
29 Boeckh J, Kaissling KE, Schneider D. Sensillen und Bau der Antennengeissel von Telea polyphemus (Vergleiche mit weiteren Saturniden: Antheraea, Platysamia, und Philosamia). Zool J Anat 1960;78:559-84.
30 Vogt, RG, L.M. Riddiford, and G.P. Prestwich 1985; Kinetic properties of a sex pheromone-degrading enzyme: The sensillar esterase of Antheraea polyphemus. Proc. Natl. Acad. Sci U.S.A. 82:8827-8831.
31 Keil, Thomas A. (1984a). Reconstruction and morphometry of silkmoth olfactory hairs: A comparative study of sensilla trichodea on the antennae of male Antheraea polyphemus and Antheraea pernyi (Insecta, Lepidoptera). Zoomorphology 104:147-156.
32 DeKramer JJ, Hemberger J. The Neurobiology of pheromone reception. In: Prestwich GD, Blomquist GJ, editors. Pheromone biochemistry. Orlando: Academic Press 1987;433-72.
33 Vogt, R G. 1987; The Molecular Basis of Pheromone Reception: Its Influence on Behavior pp. 385-431 In: Pheromone Biochemistry Prestwich, Glenn D. and Gary J Blomquist, editors. Orlando: Academic Press, Inc. 565 p.
34 Kaissling, K.-E. 1974; Sensory transduction in insect olfactory receptors. In: Biochemistry of Sensory Functions. Jaenicke, L. editor., pp. 243-273. Springer-Verlag, New York.
35 Dickens JC, Callahan FE, Wergin WP, Murphy CA, Vogt RG. Intergeneric distribution and immunolocalization of a putative odourant-binding protein in true bugs (Hemiptera, Heteroptera). J Exp Biol 1998 Jan;201(Pt1):33-41.
36 Vogt RG, Riddiford LM. Pheromone binding and inactivation by moth antennae. Nature 1981;293:161-3.
37 Pevsner J, Sklar PB, Snyder SH. Odorant-binding protein: localization to nasal glands and secretions. Proc Natl Acad Sci U S A. 1986 Jul;83(13):4942-4946.
38 Frey WH the II, Desota-Johnson D, Hoffman C, McCall JT. Effect of stimulus on the chemical composition of human tears. Am J Opthalmol 1981;92:559-67.
39 Pevsner J, Hwang PM, Sklar PB, Clark VJ, Snyder SH. Odourant-binding protein and its mRNA are localised to lateral nasal gland implying a carrier function. Proc Nat Acad Sci 1988;85:2383-7.
40 Kuizenga A, van Haeringen NJ, Kijlstra A. Identification of lectin binding proteins in human tears. Investigative Ophthalmol Vis Sci 1991;32(13):3277-84.
41 Khew-Goodall Y, Grillo M, Getchell ML, Danho W, Getchell TV, Margolis FL. Vomeromodulin, a putative pheromone transporter: cloning, characterisation, and cellular localisation of a novel glycoprotein of lateral nasal gland. Faseb J 1991;5(14):2976-82.

EXOCRINOLOGY
223
42 Booth WD, von Glos KI. Pheromaxein, the pheromonal steroid-binding protein, is a major protein synthesised in porcine submaxillary salivary glands. J Endocrinol 1991;128(2):205-12.
43 Boekhoff I, Strotmann J, Raming K, Tareilus E, Breer H. Odourant-sensitive phospholipase C in insect antennae. Cellular Signaling 1990;2(1):49-56.
44 Ziegelberger G, van den Berg MJ, KaisslingK-E, Klumpp S, Schultz JE. Cyclic GMP levels and guanylate cyclase activity in pheromone-sensitive antennae of the silkmoths Antheraea polyphemus and Bombyx mori. J Neurosci 1990;10(4):1217-25.
45 Ronnett GV, Snyder SH. Molecular messengers of olfaction. Trends Neurosci 1992;15(12):50813.
46 Singer AG. A chemistry of mammalian pheromones. -32. Proceedings of the International Symposium on Recent Advances in Mammalian Pheromone Research. J Steroid Biochem Mol Biol 1991;39(4B):627.
47 Kandel ER., Schwartze JH. Principles of Neural Science. New York: Elsevier. 1985:21-2.
48 Ronnett GV, Snyder SH. Molecular messengers of olfaction. Trends Neurosci 1992;15(12):50813.
49 Getchell Thomas V and Marilyn L Getchell Regulatory factors in the vertebrate olfactory mucosa. Chemical Senses 1990;15:223-231.
50 Kasang G, Proff L, Nichols M. Enzymatic conversion and degradation of sex pheromones in antennae of the male silk worm Antheraea polyphemus. Z Naturforsch 1988;43c:275-284.
51 Kuhn, Thomas (1970). The Structure of Scientific Revolutions. Chicago: University of Chicago Press.
52 Kurtz DB, Mozell MM. Olfactory stimulation variables. which model best predicts the olfactory nerve response? J Gen Physiol 1985;86(3):329-52.
53 Mozell Maxwell M, Sheehe PR, Swieck Jr SW, Darts DB, Hornung DE. A parametric study of the stimulation variables affecting the magnitude of the olfactory nerve response. J Gen Physiol 1984;83(2):233-67.
54 McNair HM, Bonelli EJ. Basic Gas Chromatography. Lithographed by Consolidated Printers, Berkeley, California. Varian Instrument. 1969.
55 Guyton AC. Textbook of Medical Physiology Eighth Edition. Philadelphia: WB Saunders Company. 1991.
56 L. Stowers, D. Logan LUSH Shapes Up for a Starring Role in Olfaction Cell, 2008 Jun 27;133(7):1137-1139.
57 Krapitskii, S.V., and F.G. Gribakin 1992; Electroantennogram of the American cockroach: effect of oxygen and an electrical model. Journal of Comparative Physiology-A, Sensory, Neural, & Behavioral Physiology 170(5):651-663.
58 Dubin AE, Dionne VE.Action potentials and chemosensitive conductances in the dendrites of olfactory neurons suggest new features for odor transduction. J Gen Physiol. 1994 February 1;103(2):181-201.

224
59 Choi JS, Hudmon A, Waxman SG, Dib-Hajj SD. Calmodulin regulates current density and frequency-dependent inhibition of sodium channel Nav1.8 in DRG neurons. J Neurophysiol. 2006 Jul;96(1):97-108. Epub 2006 Apr 5.
60 Zhainazarov AB, Doolin RE, Ache BW. Sodium-gated cation channel implicated in the activation of lobster olfactory receptor neurons. J Neurophysiol. 1998 Mar;79(3):1349-1359.
61 Thurm, U., and G. Wessel 1979; Metabolism-dependent trans-epithelial potential differences at epidermal receptors of Arthropods. I. Comparative data. Journal of Comparative Physiology 134:119-130.
62 O'Connell, RJ 1985; Responses to pheromone blends in insect olfactory receptor neurons. Journal of Comparative Physiology. 156:747-761.
63 O'Connell, RJ, JT. Beauchamp, and A.J Grant 1986; Insect olfactory receptor responses to components of pheromone blends. Journal of Chemical Ecology. 12:451-467.
64 Keil, Thomas A, and R Alexander Steinbrecht 1987; Diffusion barriers in silkmoth sensory epithelia: Application of lanthanum tracer to olfactory sensilla of Antheraea polyphemus and Bombyx mori. Tissue & Cell 19:119-134.
65 Mozell Maxwell M. 1992. Personal communication
66 Joshi H, Getchell ML, Zielinski B, Getchell TV. Spectrophotometric determination of cation concentrations in olfactory mucus. Neurosci Lett. 1987 Dec 4;82(3):321-326.
67 Laughlin JD, Ha TS, Jones DN, Smith DP. Activation of pheromone-sensitive neurons is mediated by conformational activation of pheromone-binding protein. Cell. 2008 Jun 27;133(7):1255-65.
68 Monti-Bloch, L., and B.I. Grosser 1991; Effect of putative pheromones on the electrical activity of the human vomeronasal organ and olfactory epithelium. The Journal of Steroid Biochemistry and Molecular Biology. (Proceedings of the International Symposium on Recent Advances in Mammalian Pheromone Research.) 39(4B):573-582.
69 Mii S, Nakamura K, Takeo K, Kurimoto S. Analysis of human tear proteins by two-dimensional electrophoresis. Electrophoresis 1992;13(6):379-382.
70 Grossman CJ, Roselle GA, Mendenhall CL. Sex steroid regulation of autoimmunity. J Steroid Biochemistry & Molecular Biology 1991;40(4-6):649-659.
71 Spielman AI, Zeng XN, Leyden JJ, Preti G. Proteinaceous precursors of human axillary odor: isolation of two novel odor-binding proteins. Experientia. 1995 Jan 15;51(1):40-47.
72 Montagna, William, and Paul F. Parakkal 1974; The Structure and Function of Skin. New York: Academic Press. 433 p.
73 Preti, George, Winnifred Berg Cutler, Celso Ramon Garcia, George R Huggins, and Henry J Lawley 1986; Human axillary secretions influence women's menstrual cycles: the role of donor extract of females. Hormones and Behavior 20:474-482.
74 Cutler WB, Preti G, Krieger A, Huggins GR, Garcia CR, Lawley HJ. Human axillary secretions influence women's menstrual cycles: the role of donor extract from men. Hormones and Behavior 1986;20:463-73.

Chapter 8: Pheromone Control of Crime and Drug Addiction There is a natural distribution of criminal behavior on a continuum with filial behavior that runs from the most malicious industrial killers to the dutiful, devoted, respectful, affectionate and loving child at home asleep, apparently due to differential parental solicitude. But why?
Given estimates of life spans for long ago, being orphaned has been more common in the past, enough to affect life strategy choice on an instinctive level. On the streets of ancient Babylon, the life of a child without parents would have been precarious indeed. Only by stealing, robbing, selling oneself, literally doing anything to survive, would street urchins live to reach reproductive age and have children of their own.
How would an innocent child be persuaded by nature to adopt this highly adaptive criminal behavior? The behavior would depend upon the absence of the parents because filial behavior, following the wishes of one’s parents, would be more advantageous and appropriate in the presence of parents looking out for you. Absence of the mother and/or father, especially during critical periods, has been associated with the appearance of juvenile delinquency to the criminal justice system and other adolescent risk-taking. Upper respiratory changes, particularly sinus development in infancy and childhood as discussed in chapters 2, 6, & 7 differentiate such critical periods.
How would one positively recognize the presence of their parents and adult relatives? (Positively because recognizing crocodiles as one’s parents has dangerous implications for survival.) The answer, Q.E.D. is that parental pheromones stimulate
EXOCRINOLOGY
225

filial behavior and that the absence of detection of parental pheromones stimulates delinquency and other risky antics. Yes, this is easy to test. Yes, the author has tested it dozens of times and providing parental, in particular paternal, pheromones to criminals cures the criminal with a single large 150 mg dose for his or her lifetime. Double-blind, cross-over, replicated clinical trials of healthy adult male facial skin surface lipid pheromone taken by mouth have yet to receive official support. It is hoped that this book’s publication will remedy that difficulty.
Criminal behavior is natural and useful for the advancement of humanity for another reason beyond mere survival. A willingness to engage in what is generally considered to be criminal behavior is important because much behavior, truth-seeking in particular, has been mis-classified as detrimental to society and therefore criminal. Scientific investigations and religious worship have shared official and popular condemnation and punishment as antisocial. Human ethology’s need for blinds from which to study all human behavior might serve as example. Indeed, the information of this tome has been suspected of criminality with the author’s home being rifled by searching officials in the performance of their duty, not once, but on several occasions. One takes one’s life in one’s hands to persevere for truth, and you can just forget about your reputation, too. Fear is a constant companion, but so is Pasteur, and with his great example, the prospect of redemption and success. Indeed, until this author’s research obtains wide public acceptance, his life and livelihood remain precarious.
Established empirically from many open trial anecdotes, paternal pheromone sufficiency allows long term maintenance of non-criminal behavior by criminals. Criminal behaviors: murder, theft, neglect and abuse of elders or children, sexual perversions of rape, homosexuality, bestiality, wild stunts, and drug addictions for heroine & crack cocaine, all of these appear to involve pheromone recognition, as all respond to 150 mg p.o. of healthy adult male facial skin surface lipid kissing pheromone in a single dose. Again, this is merely the author’s contention based upon his many anecdotes from 1987 to 2011.
Proof of the born again experience received from the pheromone tardily awaits appropriate clinical trials, however, much can be gleaned from the literature as
226

it now stands, plus there is no choice but to review the literature. First and foremost, the non-conscious pheromone recognition pathways of the human brain1, 2 correspond to areas of the brain that differ from normal when associated with criminal behavior, with sexual perversion,3 and with drug addiction. Animals and human beings share pheromone sensing hardware.456789 Many articles have demonstrated that pheromone recognition pathways in human beings involve the prefrontal cortex, insula, amygdala, hippocampus, hypothalamus & anterior cingulate areas of the human brain.
Noting that lower I.Q., shorter attention spans, and antisocial behavior typify criminals, Carpenter and Nevin (2009)10 suggest that environmental exposures known to lower IQ and shorten attention spans among children might be considered environmental causes of violence. As such, the causes Carpenter & Nevin discuss have clear effects on pheromone perception and olfaction.
Better resolution of medical imaging technologies has led to better structure identification, allowing better observation of the alignment with rarely studied pheromone pathways. Of course, heat, air pollution (particularly high ozone concentrations) and low wind speeds have also been associated with violent human behavior.11 Think about this.
The author published an article in the British Journal of Dermatology in November of 1984,12 following from a paper he presented at the International Society for Human Ethology in June of the previous year13 in which he theorized that skin surface pheromones would hold the key to understanding mate-mate and parental love, criminal behavior, sexual orientation and sexual perversion. This chapter will attempt to explain in lay terms how such a then preposterous idea could take root and become a proscription for world-wide social change.
Microeconomic theory predicts the social and personal profitability of all behavior, including criminal behavior. The first stumbling block was the microeconomic axiom of perfect information, that all buyers and sellers in a market know everything instantly. Well, in geologic time, that was true, given the human appetite for speech. Then the human light speed communication networks were an
EXOCRINOLOGY
227

evolution beyond an underlying pre-existing biological information sharing system which must exist, too. It was hypothesized that as communicable viruses transmit genetic biological information to all receptive beasts as fast as the fastest ship that sails, basically all disease and vectors of disease were information sharing and population culling devices that worked optimally.
This sounds just like Dr. Pangloss, but equations don’t lie. The next difficulty is the economic man axiom of microeconomics. Surely human and animal and for that matter plant and microbial appetites were optimized as well, as they had to be. Unlimited wants was a straight up line, but the closest thing would be a climbing logarithmic asymptote. Such functions were observable in nature as unrestrained growth of bacterial colonies provided unlimited nutrients and space for growth and waste disposal. And given the second law of thermodynamics, the increasing complexity of life was only organized to drive perfect dispersal of everything. Life itself evolves toward ever more perfect air pollution! So we could detect higher forms of life on other planets by looking for dust clouds expanding from them at near the speed of light.
Yes, this sounds more and more panglossian. However, in a world of perfect information (all know all always) populated by economic men with unlimited wants and with the free-rider problem solved (a gene is a gene is a gene), no uneconomic behavior could exist ever. Yet in every direction, human beings wasted time on apparently unprofitable pursuits. The epiphany came in physics class: equations don’t lie. A primer of population biology14 confirmed my thesis. Thus the discovery of the underlying evolutionary worth of each human motion was discoverable by dissection and socratic examination. I decided to begin with instinctive behaviors such as kissing and crying.
Kissing, crying, and emotion in general had a significant literature, most of it highly speculative and unsystematic. From ethological observation and participation, it dawned that kissing’s biological worth must be as a transfer mechanism. Kissing must be picking something up, so I looked on the skin where people kissed people and found sebum. Next comparisons to similar behavior of other species beginning with the physiology of the mistakenly mis-labeled ‘grooming’ and ‘allogrooming’
228

behaviors in apes, ungulates, avians, insects … All of the ‘allogrooming’, ‘allopreening’, and ‘displacement activity’ being observed by primatologists, ornithologists, ethologists, and entomologists was being misunderstood. All those behaviors had to be pheromone transferring or depositing devices, too.
Furthermore, the glands under the claws, beaks, antennae, snouts, and kissing human lips would be optimized to function at their highest levels in breeding seasons. The microeconomic theory insisted that even the circulation to the glands would be optimized, down to the cellular level, even down to the nuclear and genetic level: epigenetics! So for instance, it was no surprise to discover that sea birds lacked fire detection olfactory apparatus,15 since oceans and sea beaches don’t burn.
About one hundred and fifty milligrams of healthy adult human male facial skin surface lipid is sufficient to alleviate criminal behavior, juvenile delinquency, and drug addiction. Between 150 and 250 milligrams will have the effect of ending perverse sexual behavior. Injections of pheromone receptor proteins and/or epigenetic therapy may be needed for the very worst monsters in our prisons. Testing will tell.
The paternal facial skin surface lipids cure for juvenile delinquency came from empirical investigations. To the present, one hundred and two open anecdotal trials of paternal skin surface lipid kissing pheromone on chewing gum vehicle have resulted in complete remissions of all symptoms of delinquency, all symptoms of criminal inclination, all symptoms of runaway behavior, all symptoms of borderline personality disorder, all symptoms of homosexual perversion (in twelve female lesbians and gay men), and all symptoms in one case of a perversion so problematic to the physician with the disorder that this researcher could not be informed of its true nature.
In 1987, a family member’s difficulties resulted in the first cure of borderline personality disorder with criminal behavior in a fourteen year old relative. Incarcerated and physically restrained, the family member chewed the gum and instantly felt relief from ‘the monkey’ on his back. Removing the monkey would become a familiar metaphor as it would be used by dozens of pheromone recipients in
EXOCRINOLOGY
229

the years to come. Further doses led to graduation from law school at the top of his class and many promotions--a pattern of ambition repeatedly seen later.
The next family member in trouble was thwarted trying to complete his medical degree. After complaining of ‘mitral valve prolapse’ and displaying symptoms of agoraphobia, the pheromone gum was provided, again in desperation. Immediate relief of symptoms began the anecdote, ending with achievement of a tenured faculty position at an accredited medical school.
The author began solicitations to attempt double-blind clinical trials at state and local levels in four states: New York under Cuomo, Texas under Richards, Georgia under Miller, and Florida under Bush. Having had past associations with all four, the author believed that obtaining cooperation from them would be easy. “I provided Cuomo with the ‘wearing of the green’ broccoli bit famous at the 1992 Democratic convention. I was the first person outside Ann Richard’s family to suggest that she run for governor and together with Bill Clinton the three of us had come up with the Dead-beat Dad laws for her. I proposed the HOPE Scholarship Program to Miller when his aide for student & university affairs, and I had been a family friend of the Bushes, proposing back in Texas that Johnny come to Florida in the first place. I was far wrong.” All four failed even to grant a meeting on the subject of a cure for criminal behavior. Moreover, the doors were slammed shut by their prison systems as well.
Bill Clinton and I had been brothers in service in Texas during the McGovern campaign. That only infuriated the US Bureau of Prisons who dismissed my Institutional Review Board submission out of hand. And even though this author had personally saved the life and career of George W. Bush for his family the year before his Clinton experience,1 that pushed the Bureau of Prisons to an uninhibited frenzy, and my submission for a trial of the pheromone was again refused with zeal, this time with prejudice by the bureaucracy of Bush administration. Perhaps other medically trained people can read this monograph and recommend trials with better result? Steal your nerve, ladies and gentlemen. Humanity does love its prisons. Getting rid of them will be no easy task.
230

The hypothesis to be tested is: Does healthy adult male facial skin surface lipid kissing pheromone alleviate symptoms of criminal behavior and juvenile delinquency? The method is to watch behavior records of experimental vs. control groups and see a statistically significant decline in the experimental group receiving pheromone gum over the control group; then with the experimental pheromone gum supplied to the control group, to see that group recover normal behavior as well.
The materials and methods would be to select from volunteers two similar groups from the same population of incarcerated criminals. (Some believe that sick people lack the capacity to volunteer. The author disagrees suggesting that clinical trials within a prison population in order to treat the malady from which the prisoners suffer would not be exploitation. Indeed, helping prisoners recover from criminal, perverse, and drug-addicted inclinations should be the main mission of any prison system.) Using intermediary technicians unaware of the identity of each dose or control, indistinguishable doses of pheromone-laced chewing gum and canola-oil laced chewing gum would be provided to experimental and control populations, respectively. Once a predetermined period of behavioral observation has passed, the control population would be provided the pheromone-laced chewing gum and their behavior assessed by observation. A statistically significant difference between un-dosed and pheromone-dosed prisoners should be seen and pheromone-dosed prisoners should have statistically significant decreases in recidivism. Similar protocols in combination with epigenetic therapy, direct injection of pheromone receptor proteins, or tear transfers (like Clockwork Orange) should also be tried. Doing nothing is negligence.
Pheromone collection must be made from volunteer pheromone donor males of good reputation of about ‘father age’ to the prisoner pheromone recipient. This is the type of donation that has achieved success in previous open trials. Maternal skin surface lipids in similar large unnatural dosage seem to stimulate homosexual perversion as observed anecdotally in one case. However, given the ease with which homosexuality can be cured with the paternal pheromone (eight successful open trials), further investigations using female-donated facial skin surface pheromones may be allowed to go forward, provided there is no sexually perverted motive by researchers.
EXOCRINOLOGY
231

PERIGRINATION
Leaving the old life and circumstances behind has been seen consistently in those criminals, delinquents, and runaways who chewed the paternal pheromone gum. There is little wonder why.
The very real and immediate prospect of pheromonal rehabilitation of criminals will demand close review and careful ethological experimentation. What is the natural history of criminal behavior? In other words, what benefit does the existence of crime hold for mankind? Given the difficulty and expense of controlled experiments on our fellow human beings, we will utilize animal experimentation, famous volunteers, and exocrionological case reporting to hone the methology. 150 mg of a man’s face grease p.o. is just an approximation. Given the multiple devices controlling human bonding, duplication of the paternal pheromone deficiency mechanism to produce criminal behavior seems likely. Observers of criminals16 and criminal-type pheromone deficiencies will vary across the board.
PHEROMONES SUBSTITUTION ILLNESS: DRUG ADDICTION
Addiction played a main theme in the author’s initial reports.17,18 However, the discovery that healthy adult human male facial skin surface lipids in a single sub-gram dose could actually resolve heroine addiction and then crack cocaine addiction in heavy users in the field permanently, came as pleasant surprises. It makes sense that it would be so, at least now it does. The instantaneous, complete remission of all symptoms surprisingly without withdrawal effects, followed by adoption of healthy, reproductive, and socially appropriate lifestyles, exceeded all expectations and is beyond the ken of colleagues who would rather see their patients suffer than to perform the experiment ready and waiting in the appendix to this book.
232

Love-deficient, presumably pheromone deficient, people commit all the crimes of passion, do they not? Certainly drug addicts commit regular criminal acts in support of their addiction. The protocol in the appendix has been used to treat dozens of criminals and delinquents, all successfully. 150 - 250 mg of healthy adult male (over 30 years of age) facial skin surface lipid taken by mouth on a chewing gum vehicle completely eliminates all symptoms of drug addiction instantly, apparently at the first ten milligram exposure.
Sadly, despite several attempts, human pheromones have had little effect upon alcoholism. Experimental exposures to ethanol had no effect on reproductive fertility or behavior, including response to pheromones, of intact experimental animals.19 In humans, alcohol promotes dopamine release in the brain’s nucleus accumbens, with a preferential effect in the ventral striatum.20 The ventral striatum is unexcited in pheromone studies,21, but it does receive tertiary projections from the olfactory and vomeronasal amygdalae.22 Willed ceasing of alcohol consumption cures symptoms of alcoholism. Perhaps alcohol consumption could be stopped by replacement with heroine addiction or crack cocaine, which are more amenable to pheromone intervention?
BORDERLINE PERSONALITY DISORDER
Pheromones are processed in the medial amygdalae and the memory-associated hippocampi, but not the parahippocampal gyri, nor the uncuses. All four areas are associated with abnormalities in BPD. It is asserted with fMRI evidence that dorsolateral prefrontal cortex and the hippocampuses sustain symptoms of borderline personality disorder.23 Lesions of the hippocampus or prefrontal cortex change behavior stimulated by chemoreceptions in cats.24 The medial prefrontal cortex is a projection area for the pheromone-sensing accessory olfactory bulb25 again, in cats. In humans, under fMRI the anterior part of the inferior lateral prefrontal cortex is directly activated by the human pheromone, androstadienone.26 That is close enough for association.
EXOCRINOLOGY
233

A derivative of Omega-3 fatty acids (fish oils), which have some human pheromonal stereochemistry, seems to improve BPD ever so slightly.27 Recall that human sebum taken up in kissing works synergistically and species specifically just like these variously helpful free fatty acids, so this slight dietary effect suggests a much more potent effect from the pheromone.
Borderline sufferers share behavioral similarities to juvenile delinquents who have responded famously in open trials of healthy adult male facial skin surface lipids, so the prognosis for full and rapid recovery with paternal pheromone treatment is excellent.
234
1 Zhou W, Chen D Encoding human sexual chemosensory cues in the orbitofrontal and fusiform cortices. J. Neurosci. 2008 Dec 31; 28(53): 14416-11421.
2 Treyer V, Koch H, Briner HR, Jones NS, Buck A, Simmen DB Male subjects who could not perceive the pheromone 5a-androst-16-en-3-one, produced similar orbito-frontal changes on PET compared with perceptible phenylethyl alcohol (rose). Rhinology. 2006 Dec; 44(4):278-282.
3 Savic I, Berglund H, Lindström P. Brain response to putative pheromones in homosexual men. Proc Natl Acad Sci U S A. 2005 May 17;102(20):7356-7361. Epub 2005 May 9.
4 Moncho-Bogani J, Martinez-Garcia F, Novejarque A, Lanuza E. Attraction to sexual pheromones and associated odorants in female mice involves activation of the reward system and basolateral amygdala. Eur J Neurosci. 2005 Apr;21(8):2186-2198.
5 Sobel N, Prabhakaran V, Hartley CA, Desmond JE, Glover GH, Sullivan EV, Gabrieli JD. Blind smell: brain activation induced by an undetected air-borne chemical. Brain. 1999 Feb;122 (Pt 2):209-217.

EXOCRINOLOGY
235
6 Tirindelli R, Dibattista M, Pifferi S, Menini A. From pheromones to behavior. Physiol Rev. 2009 Jul; 89(3):921-956.
7 Touhara K, Vosshall LB. Sensing odorants and pheromones with chemosensory receptors. Annu Rev Physiol. 2009; 71:307-332.
8 Zufall F, Leinders-Zufall T. Mammalian pheromone sensing. Curr Opin Neurobiol. 2007 Aug;17(4)483-489. Epub 2007 Aug 20.
9 Tizzano M, Sbarbati A. Hormone fatty acid modifications: gram negative bacteria and vertebrates demonstrate common structure and function. Med Hypotheses. 2006; 67(3):513-516. Epub 2006 Apr 27.
10 Carpenter DO, Nevin R. Environmental causes of violence. Physiol Behav. 2009 Sept 14; [epub ahead of print].
11 Rotton J, Frey J. Air pollution, weather, and violent crimes: concomitant time-series analysis of archival data. J Pers Soc Psychol. 1985 Nov;49(5):1207-1220.
12 Nicholson B. Does kissing aid human bonding by semiochemical addiction? 1984 Br J Dermatol. Nov; 111(5):623-627.
13 Nicholson B. Love and kisses: a semiochemical addiction model for human bonding. 1983. Paper presented to the International Society for Human Ethology and Animal Behaviour Society biannual international meeting at Bucknell University, Bucknell PA.
14 Wilson EO, Bossert WH (1971). A Primer of Population Biology Stamford: Sinauer Associates.
15 Tinbergen N The Herring Gull’s World 1989 Gilford, CT: Globe Pequot Press.
16 Clinton H. It Takes a Village: And Other Lessons Children Teach Us. (1996) New York: Simon & Schuster.
17 Nicholson, B. 1983; A semiochemical addiction model for human bonding. Paper presented to the Animal Behavior Society and the International Society for Human Ethology, Lewisburg, PA.
18 Nicholson, B. 1984; Does kissing aid human bonding by semiochemical addiction? British Journal of Dermatology 111(5):623-627.
19 Fadem BH. Effects of postnatal exposure to alcohol on reproductive physiology and sexually dimorphic behavior in a marsupial, the gray short-tailed opossum (Monodelphis domestica). Alcohol Clin Exp Res. 1993 Aug;17(4):870-876.
20 Boileau I, Assaad JM, Pihl RO, Benkelfat C, Leyton M, Diksic M, Tremblay RE, Dagher A. Alcohol promotes dopamine release in the human nucleus accumbens. Synapse. 2003 Sep 15;49(4):226-231.
21 Martínez-Hernández J, Lanuza E, Martínez-García F. Selective dopaminergic lesions of the ventral tegmental area impair preference for sucrose but not for male sexual pheromones in female mice. Eur J Neurosci. 2006 Aug;24(3):885-893.
22 Martinez-Marcos A. On the organization of olfactory and vomeronasal cortices. Prog Neurobiol. 2009 Jan 12;87(1):21-30. Epub 2008 Sep 25.

236
23 Sala M, Caverzasi E, Lazzaretti M, Morandotti N, De Vidovich G, Marraffini E, Gambini F, Isola M, De Bona M, Rambaldelli G, d'Allio G, Barale F, Zappoli F, Brambilla P. Dorsolateral prefrontal cortex and hippocampus sustain impulsivity and aggressiveness in borderline personality disorder. J Affect Disord. 2011 Jan 4; [Epub ahead of print]
24 Nonneman AJ, Kolb BE. Lesions of hippocampus or prefrontal cortex alter species-typical behaviors in the cat. Behav Biol. 1974 Sep;12(1):41-54.
25 Staples LG, McGregor IS, Apfelbach R, Hunt GE. Cat odor, but not trimethylthiazoline (fox odor), activates accessory olfactory and defense-related brain regions in rats. Neuroscience. 2008 Feb 19;151(4):937-947. Epub 2007 Dec 4.
26 Gulyás B, Kéri S, O'Sullivan BT, Decety J, Roland PE. The putative pheromone androstadienone activates cortical fields in the human brain related to social cognition. Neurochem Int. 2004 Jun;44(8):595-600.
27 Zanarini M C, Frankenburg F R. Omega-3 Fatty acid treatment of women with borderline personality disorder: a double-blind, placebo-controlled pilot study. Am J Psychiatry. 2003;160(1):167–169.

EXOCRINOLOGY
237

Chapter 9: Pheromone Control of Sexual Desire
Syndromes
HOMOSEXUALITY
Addictions to other pheromones suggest etiologies for homosexuality, pedophilia, habits of torture (sadism and masichism), the rapist’s addiction to female alarm pheromone, foot & hand fetishes for those pheromones, nymphomania and satiriasis.
Homosexuality differs from crime and drug addiction in that maternal affection pheromone reception is adequate or compensating in the presence of paternal pheromone deficiency. Vitamin deficiencies occur naturally, too, and can be just as easily treated. Homosexuals obtain pleasures of the flesh to no reproductive benefit and this has been accorded respect, properly so. The recovery of heterosexual ideation and behavior by life-long homosexuals was, indeed, an unintended side-effect of the paternal skin surface lipid pheromone gum. However, none of the twelve former homosexuals expressed any interest in returning to the homosexual lifestyle, preferring heterosexuality to homosexuality with definite certainty. Again, though, all were taken aback by the transition and all would have rejected taking the pheromone gum if they had truly believed that it would effect the change they experienced. Homosexuals should therefore be excluded from trials or provided cooling off periods before being taken into trials to give them an appetite for reproduction.
Homosexuals found themselves at sea and uncertain once their perversion was discarded. Jessica Simpson, a movie star/singer who had appropriated the pheromone gum from a trial participant, was able to marry only to leave her spouse after a few years for a series of dancers and football players. Another movie star, Leelee Sobieski, who took part in the open pheromone trials moved through several relationships and is now engaged and expecting her first child. Kunis, Portman, Page, Hilton, Lohan, Charlie Sheen and others are responding to therapy even now.
238

Hollywood knows a good thing. Actors are willing Guinea pigs and their behavior is easy to observe from a great distance.
According to Sokolov,1 on grip surfaces of the hands, feet, and prehensile tails of mammals, eccrine (perspiration) glands tend to predominate. The eccrine glands produce the watery film that works with the serrations we call fingerprints to improve traction and grip. Eccrine perspiration helps to move sebum and other skin gland secretions over our skins. It may contribute to pheromone transmission and secretion. Perspiration, as in intercourse, exaggerates and changes the distribution of sebum and apocrine secretions. At the same time, recognizable chemical differences in hand and foot secretions must be the physiological basis for compulsive hand and foot fetishes.
DISEASES OF ERSATS HUMAN PHEROMONES: LATAH
Latah is a shock-like condition in which the affected individual, a woman [or a male homosexual-BN], mimics the movements and sounds of those nearby against her [/his] will.2 Latah is characterized by an exaggerated motor startle response, often with associated involuntary vocalizations, echolalia, echopraxia, and forced obedience. Exaggerated startle to touch and often to frightening words (similar to BOO!) gets worse or deeper with repeated stimuli. Latah sufferers are quickly enticed to the Latah syndrome by relatives worried upon observing symptoms of depression and/or exaggerated grieving after the passing of a husband. The intense teasing replaces depression with Latah, but life-expectancy does improve, plus the entire family is entertained! Latah patients should see full and complete recovery upon oral administration of the healthy adult male facial skin surface lipid pheromone as described in the appendix to this volume. Latah should resolve once the pheromone deficiency is alleviated.
Recall how cholesterol esters are found more commonly in childhood and neonatal sebums, differentiating them from adult-typical compositions?3 Higher concentrations of linoleate are seen in child sebum, too.4 These, and other childhood-
EXOCRINOLOGY
239

typical chemical species, should be investigated as candidates for a human child-aversion or child recognition pheromone. A human child-aversion pheromone explains the Kibbutz aversion data and find a place in semiochemical medical practice for treatment of sexual child abuse resulting from pheromone receptor protein deficiency in tears or blood.
Pheromonal aversion can involve nausea, probably involving the Peyer’s patches full of microvillar cells in the alimentary tract. Disgust sensitivity predicts intuitive disapproval of sexually perverted people,5 so pheromone aversion being the cause of homophobia seems reasonable. Feelings of disgust can be encouraged or overcome with training. Pica, eating dirt, is too common. Japanese people consume raw fish. French people eat snails. Koreans quite happily dine on dogs. Some Africans consume living grubs. Americans eat fast food. Taste is a malleable thing.
PHEROMONE DEFICIENCY ILLNESSES: DIVORCE
Perhaps divorce can be eradicated by the medical profession. Women now get divorces and elect to leave their loving husbands (and vice versa) about half the time. Occupations that keep the family away from heavy air pollution, such as those of fishermen and park or forrest rangers, maintain pair bonds most reliably. Recently air pollution has been associated with depression, anxiety, and learning disabilities.6 It should please rejected husbands to know that the divorcing wife likely has hypoactive sexual desire syndrome, and her promiscuous antics following separation are not entirely her own decision. Divorced wives felt isolated from their husbands, as if he were a stranger. This is a product of desperation, born of sexual frustration from which husbands rarely suffer. Since pheromone reception (and pheromone deficiency) are non-conscious, there is no conscious fault and no blame should be laid.
The cure for divorce, just one annecdote currently, is to alleviate the paternal pheromone deficiency of the divorcing wife or husband with 150-250 mg of healthy adult male facial skin surface lipid p.o. Follow-up to treat hypoactive sexual desire
240

syndrome (if the father-emitted pheromone proves ineffective in bringing back sexual behavior rewards) with ten to twenty milligrams of human (husband) genital and peri-genital sebaceous and pubic scent gland secretions p.o.--or have the husband permit vigorous fellatio prior to consensual intercourse to improve libido. Forcing the wife to intercourse is illegal in civilized jurisdictions and counter-productive, since anger is prophylactic to pheromone reception. Anger in either sex engorges erectile tissue in the nasal passages to close off chemoreceptive middle and upper meati, recall.
In the case of husband and wife, the pheromones of addiction are individually specific. The husband loves only his wife and the wife loves only her husband in mate mate bonding, assured by their unique chemical signatures. If any divorce persists after this book’s publication, perhaps divorce courts will one day order professional pheromone collection and delivery to ex-spouses as part of divorce settlements to short circuit all of the drama and criminality of sexual passion.
Treating either husband or wife with pheromones can disturb the marriage. Any intervention should be limited and any treatments provided should be at least token shared by both to avoid jealousy or suspicion. A 1 milligram ‘taste’ should be supplied to the spouse to allow them to identify and begrudgingly tolerate ‘the intruder’ as beneficial experimental exocrinology. Otherwise an osculation quarantine should be imposed temporarily. A small amount of wine after pheromone oral administration may reduce jealousy as well.
EXOCRINOLOGY
241
1 Sokolov, Vladimir Evgen'evich 1982; Mammal Skin. Berkeley: University of California Press. 695 pages, p 21.

242
2 Bartelsman M, Eckhardt PP. Mental illness in the former Dutch Indies--four psychiatric syndromes: amok, latah, koro and neurasthenia. Ned Tijdschr Geneeskd. 2007 Dec 22;151(51):2845-2851.
3 Stewart, Mary Ellen, and Donald T. Downing 1991; Unusual cholesterol esters in the sebum of young children. Journal of Investigative Dermatology 95(5):603-606.
4 Stewart, Mary Ellen 1992; Sebaceous Stewart, gland lipids. Seminars in Dermatology Jun; 11(2):100-105.
5 Inbar Y, Pizarro DA, Knobe J, Bloom P. 2009; Disgust sensitivity predicts intuitive disapproval of gays. Emotion. 2009 Jun;9(3):435-9.
6 Fonken LK, Xu X, Weil ZM, Chen G, Sun Q, Rajagopalan S, Nelson RJ. Air pollution impairs cognition, provokes depressive-like behaviors and alters hippocampal cytokine expression and morphology. Mol Psychiatry. 2011 Jul 5;. [Epub ahead of print]

Chapter 10: Pheromone Reception & Disease
! Throughout the course of medical history, distinct syndromes of unknown etiology eventually find causation from a single root. Although many different things seem to cause the same disease, when worked out, many different things never cause anything. One disease, one cause. Thus malaria is caused by plasmodia vectored by several different mosquitos. (It is misnamed, instead of “bad” swamp “air” disease, it should be plasmodiasis.) From this long history of ever one disease: one cause, it is intensely irritating that so many psychologists and their sympathizers among the medical profession today adamantly and incomprehensibly attribute multiple causes for virtually every idiopathic syndrome. Such incompetence is an embarrassment to medicine and to science. Our model provides an economic single justification for pheromonal sociopathy and disease. The moderation of pathological intensity from nuisance to mortal consequences enhances the model and explains experimental sociopathological findings. Now, encouraging partial case reports give us hope to control autoimmune, other heretofore idiopathic ills, and cancer. With biochemical tracing of neurological pathways in the brain, the ‘surprising’ abundance of human chemosensory pathways allows the present chapter’s investigation. (If we cannot follow pheromones themselves, we can at least follow the neurological pathways of neurons that respond to pheromones. Happily, these pathways have been traced with good resolution.) For pheromones to control these ills, the nerves that respond to pheromones must be found in disease associated neuroendocrine tissues thought to control these ills. And where pheromones fail, as in alcoholism, there must be a dearth of such pheromonal central nervous system innervation involved in alcoholic reward centers of the brain. This was proved to be true.
EXOCRINOLOGY
243

Humans die appropriate to population needs to maintain the hobbled population equilibrium that got humanity this far. That is also what economics demands if you take things to the Panglossian extreme as we have. Without the feedback devices (pheromones) killing humans to maintain our numbers to available resources, now it will be up to us. Human individual demise selection in an epidemic is accomplished via human pheromones. This is just like pheromones regulating population fertility of bacteria in an enclosed vial. Criminal behavior, susceptibility to addictive drugs, perversion, autoimmune disease, and cancer: all can kill, prevent or limit reproduction, or enfeeble offspring and all are essentially pheromonal. Indeed, the evolution of autoimmune illness and sociopathy in humans and domesticated animals may have resulted from human care-giving.
Physicians must utilize objective criteria to monitor patient welfare or well-being. Evidence-based patient management requires that the physical exam cover behavioral and cognitive pheromone challenges and responses systematically. Human pheromones, the most potent of any drug class by orders of magnitude, must be utilized to attempt to threat these diseases. Some inadvertent successes and fruits of economic calculation augur well for near term success.
Olfaction nerve CNS pathways run with pheromone recognition pathways some of the time as these afferent nerves coarse toward and through the brain. The parallel arrangement allows communication between inputs which may include gates or shoves/encouragements of capacitance via dendritic capacitance ‘analog switching’. Please see Chapter 11. Future examination of the semiochemical recognition areas of the brain may bring notice to these areas being damaged more than their olfactory running mates in autoimmune disease. They might even compare them to normal ones and find a soluble pheromone receptor protein/pheromone etiology.
244

Some sort of poisoning via the retrograde neural transport system from olfactory mucosa to the CNS was suggested as the cause of Alzheimer's disease years ago.1,2 The olfactory tract's neural transport system could accommodate pheromone receptor-proteins as well as pheromones and anti-pheromones and these possibilities must be explored systematically.
The invariable finding of severe and even maximal involvement of the olfactory regions in Alzheimer’s disease is in striking contrast to the minimal pathology in the visual and sensorimotor areas of the neocortex and cannot be without significance. In the olfactory system, the sites that are affected—the anterior olfactory nucleus, the uncus, and the medial group of amygdaloid nuclei—all receive fibres directly from the olfactory bulb. These observations at least raise the possibility that the olfactory pathway is the site of initial involvement of the disease. [A] large majority of patients with Parkinsonism has olfactory dysfunction, as measured by tests of odor identification and detection. This disorder seems to be (1) stable over time, (2) present relatively early in the disease process, (3) general in nature, (4) independent of motor and cognitive aspects of the disease, and (5) indistinguishable, at present, from the olfactory disorder observed in Alzheimer’s disease.3 People with Parkinson's, and other neurological disorders secrete sebaceous secretion copiously.4 A sebaceous, kiss-transferred, addictive pheromone5 would thus provide an increased incentive for patient maintenance by the family and predicts more family accommodation for them.
Fruit flies see a lone pheromone receptor protein homologue in their hemolymph in response to viral infection, with receptor proteins themselves strongly expressed during metamorphosis, suggesting that they participate as chemosensory molecules in tissue remodeling.6 Thus improves the case that pheromone receptor proteins must surely be active during embyological development of the human being, and susceptible to pheromonal influence to account for the currently unknown etiologies of birth defects and still-birth.
Indeed, resistance or immunity of pathogenic bacteria to antibiotics is pheromone based,7 as is pathogenicity.8 Evolving from bacteria, we should be able to respond in kind, would you not think so? After all, individual cells of the body have not lost the sense of smell, so necessary to life as colonial bacteria. Indeed, assembly into an organism should require greater cellular chemosensory acuity, and greater specialization.
EXOCRINOLOGY
245

ANGER or AGGRESSION PHEROMONE IN AUTOIMMUNE
DISEASE:
Anger pheromones9,10 may logically be suspected of both neurotoxic and autoimmune pathogenicity. Incurring wrath might indicate diminished usefulness for group survival and may be directly poisonous by pheromone. A righteous wrath, as when the victim agrees with the emitter/perpetrator and tears-up at the rebuke (enhancing receptivity), might be more poisonous still. Because superior and middle meati of both nasal passages close off by anger-stimulated erectile tissue engorgement,11 therefore anger must itself be prophylactic against auto-reception. Thus, self-righteous anger is also tonic against the unjust wrath of others. The hardy survival of angry and mean patients on the wards is well known to every physician and illustrative.
The evolution of this device becomes warranted when one considers the following. A hunter-gathering group rid of slackers would be expected to outperform those groups lacking an efficient mechanism to dispose of their unprofitable hangers-on. In the absence of effective predators, healthy group members inconvenient to the group could suffer timely demise semiochemically via any number of means, depending upon population needs.12 Violent criminal behavior would be conspecific predation under this model and was examined elsewhere in the previous chapter. (Similarly, the presence of ‘pet-food’ isles in grocery stores is solid evidence for semiochemical parasitism.)
Candidates for anger pheromones, antipheromones, and pheromone-binding proteins might be found in human saliva. Anger pheromones appear in rodent saliva.13,5 Watching angry medical school professors screaming with rage, they do seem to spit tiny hate packets of saliva, sometimes it lands on their beards or even their eyebrows. (No wonder judges and priests sit up high and wear robes with all the self-righteous sputtering?) If one must communicate with an angry fellow human being, doing so by telephone or other electronic means may be the safer strategy. Howard Hughes thought so.
246

Perhaps some anger pheromones may be found among the chemically reactive conjugated dienes of the human male scalp,14 since "blowing one's top" may indicate increased emissions from scalp scent glands. Given the susceptibility of sebaceous secretions to heat,15 "hot heads" may be semiochemically dangerous to health. Systematic testing of various chemical species against psychogenic lachrymal fluid in the presence of sensitive electrodes and under UV examination for conformational change would be a logical place to begin.
AUTOIMMUNE NEURODEGENERATIVE DISEASES:
ALZHEIMER’S DISEASE
Alzheimer’s disease affects the olfactory nerve with a rhyno-central progression suggesting initial insult via the olfactory nerve.16
Aberrant expression of 14-3-3 proteins is associated with both Alzheimer’s and Parkinson’s diseases. The insect homolog is found widely, but includes antenna and pheromone glandular locations, suggesting pheromone receptor protein function, along with its association with Heat Shock Protein 60 (Hsp60)17 Hsp10 is also a partner with Hsp60 to provide the two conformations needed for functionality as dissolved pheromone receptor proteins (PBPs or Pheromone Binding Proteins in other discussions in this book). Hsp10 interacts with “deoxyribonucleic acid checkpoint inactivation, termination of M-phase, messenger ribonucleic acid export, import of nuclear proteins, nucleocytoplasmic transport, and pheromone signaling pathways.” “Recent data suggest that Hsp10 may be not only a component of the folding machine but also an active player of the cell signaling network, influencing cell cycle, nucleo-cytoplasmic transport, and metabolism, with putative roles in the lack of cell differentiation and in the inhibition of apoptosis.”18
Idiopathic inflammation is thought to be the cellular basis for Alzheimer’s disease. Inflammation is essentially an alarm pheromone response of the body’s individual cells and soldier cells called by those individual cells by scent emissions, just like ants and termites.
EXOCRINOLOGY
247

NSAIDs affect alarm pheromone reception responses of the body. So does a mist of aspirin calm bees as well as smoke?
AGE RELATED MACULAR DEGENERATION
Since a diet high in omega-3 fatty acids such as those from fish oil improves macular degeneration outcomes in women,19 it seems reasonable to expect that the human pheromone that these two chemicals resemble might prevent this unfortunate outcome. Testing will tell.
SUBORDINATE PHEROMONE in SUDDEN INFANT DEATH
SYNDROME (SIDS), BLIGHTED OVUM, BIRTH DEFECTS,
ANOREXIA NERVOSA, BULIMIA NERVOSA, OBESITY, and
INFERTILITY
Prolonged stress warps reproduction to adjust population fertility to stressful circumstances, reducing gametogenesis & libido, elevating still-birth rates, and inflicting poor prenatal and maternal care of children. Pheromones emitted during stress may provide positive evidence to stimulate those difficulties, but lack of pheromone reception can accomplish the same ends. Conflicting inputs suggest that pheromones can work in opposition to blunt or counteract each other, similar to the way hormones can be seen to work in opposition. Of course, that has been the observation from anecdotes so far. Economically, pheromones must be involved with all reproductive behavior/libido, from behavioral fertility, gametogenesis, conception, gestation, growth and development to maturity and old age. The delivery of stress information to the individual human being cannot be entirely hormonal, and once the
248

pheromone has been received, an acknowledgement signal pheromone may be needed. Poor wound healing, enhanced disease rates, shortened lifespans, female acyclicity, high infant mortality and cardiovascular disease depend upon stress, too, and provide opportunities to utilize oppositional pheromones (like ‘kissing’ pheromones) to relieve them. It is likely that the pheromone responsible for Anorexia Nervosa is the axillary stress pheromone responsible for most of the effects of stress on reproduction and on populations generally.
Why? Pheromones limit species fertility to available resources.20, 21 Pheromones are why Malthus was all wrong. Subordinate pheromones are typical of stressed, grouped fertile females and emitted by puberty-delayed adolescents22 and human anorexia nervosa sufferers and their poisonous dyads.23 On the same continuum as delaying puberty are behavioral infertility, obesity, physical infertility, blighted ovum, spontaneous abortion, birth defects, sudden infant death syndrome, physical child abuse, sexual child abuse, juvenile delinquency, running away, criminal behavior, homosexuality, sadomasochism, homosexuality, fetishes and other sexual perversions. All diminish effective population fertility and are, or must surely be, controlled by human pheromones. Liberal administration of healthy adult (over 30 for methylation) male facial skin surface lipids p.o. in doses around 150-250 mg should diminish the burden considerably.
Pheromones kill fetuses and cause their resorption in laboratory animals,24, 25 yet before this author’s landmark paper in 2001, the possibility had never been raised that pheromones could explain the nearly identical phenomenon in human beings. (Given the ubiquity and credence for such laboratory animal testing as medical research to be applied to medical treatment of human beings, this thinking deficiency is difficult to explain.) Unexplained spontaneous abortion and blighted ovum must have pheromonal cause in humans.
Subordinate female-emitted pheromone is even more suspect in sudden infant death syndrome (SIDS) and an axillary semiochemical phero-pathogen appears most likely.26 Coleman-Phox Odouli and Li found that the use of a fan during sleep reduced the risk of sudden infant death syndrome,27 suggesting an airborne pathogen. Airborne pheromones travel in plumes of sufficient concentration to be
EXOCRINOLOGY
249

recognizable.28 Surely then, pheromone plumes are being disrupted by the fans used to cause the observed protection from SIDS, preventing or decreasing the reception of poisonous human pheromone. The effect was more pronounced if the nose was less well exposed and with higher temperature (higher axillary emissions from the poisoner). The protection diminished with the use of a pacifier, which increases nasal breathing past chemoreceptive mucosae. Everything fits.
SIDS is common in infants only after they attain six weeks of age, and the protection during the neonatal period baffles other thoughtless investigators.29 In a striking early observation, Darwin noted psychogenic lachrymation only after the first six weeks of age.30 This is the neonatal protected period, and recognizing that tears function in pheromone reception, then an exocrinological SIDS etiology is demonstrated. Subordinate maternal axillary emissions due to stress and low hierarchical status (relieved somewhat by increased status) are the most likely culprits for sudden infant death syndrome, and covering the mother's axillae with petroleum jelly whilst she is in an apneaic infant's breathed atmosphere must be taken as a prudent and easy precaution.
As precaution against stillbirth and blighted ovum, withholding other male ‘kissing’ pheromones from pregnant, lactating, or potentially pregnant females is a reasonable precaution to avoid any human Bruce effect killing of fetuses. Wives with husbands might also be omitted to save the ladies semiochemical discomfort from jealousy.
The anger pheromone alluded to above is also a candidate with either parent the potential source. In which case, admonishing parents to never dispense anger pheromone over helpless infants may suffice to diminish our population’s burden of SIDS and failure to thrive victims. Needless to say, pheromone-barrier shower caps may replace wigs and high rostrums in courts of law, if courts and laws are even necessary once pheromones are under control.
250

EPILEPSY
Right temporal lobe epileptics complicated by depression have hippocampal atrophy that cannot be explained by epilepsy alone.31 Similarly petite mal epilepsy has been treated effectively with the paternal pheromone in just one case. Alarm pheromones cause convulsions in rats treated with the tri-cyclic anti-depressant, imipramine.32 Epileptogenic tissues of the central nervous system are all pheromone reception pathways and association areas.
AUTISM
An autistic child pays little attention to parents, or anyone. He conserves his energy. Autism must be pheromonal. Genetically identical twins can express autism in a single sibling. Families in which one monozygotic twin is diagnosed with autism spectrum disorder, the second twin is unlikely to receive the same diagnosis after elapse of twelve months.33 This suggests economic titration.
Experimental pheromone deficits give mice symptoms of autism.34 In humans, autism causes significant reductions in grey matter volume of medial temporal, fusiform, and cerebellar regions.35 Cerebellar atrophy must be consequential damage, since the cerebellum is unassociated with either pheromone reception or olfaction. However, the right orbitofrontal cortex, right fusiform cortex, and right hypothalamus respond to airborne natural human sexual sweat.36 Young children with autism spectrum disorders show more olfactory sensitivities than children with other developmental delays.37 The olfactory and pheromone recognition pathways (both chemoreceptive, after all) run parallel for much of their runs, recall. Pheromone recognition and reward systems for socialization can account for all symptoms of autism.
Recall that twelve cases of sexual perversion have been remedied with human adult male facial skin surface lipid by mouth, demonstrating that pheromone reception is, itself, subject to pheromonal regulation. These observations suggest a
EXOCRINOLOGY
251

poisonous pheromone reception as the cause of autism. Early onset (after the six week pre-lacrimation hiatus) suggests a parental source.
Predictions, anyone? Of course some central CNS damage must be pheromonally inflicted, but there are simpler explanations for symptoms. The autistic child would have chemically strange emotional tears, perhaps with fewer pheromone receptor proteins, or with inactivated/poisoned pheromone receptor proteins, or even aberrant lachrymal ionic concentrations.
Autism, like physical child abuse, may be semiochemically treatable. Parental facial skin surface lipids provided to the autistic child by mouth should provide full recovery when treated early to avoid the consequential cell death cascade. Differential parental solicitude in identical twins would thus account for differential pheromone reception histories predisposing for the illness. Measuring that may prove more difficult than just trying various doses of parental love pheromones for those already autistic. Perhaps in concert with intravenous epigenetic therapy for resistant cases, maternal and/or paternal facial or other (areolar?) protective skin lipids can be therapeutic?
The possibility of helpful omega-3 fatty acid treatment in autism38 is suggestive that human skin surface lipid pheromone may have important therapeutic effects. Omega-3 fatty acids are, after all, only poor substitutes for the stereochemically similar human skin surface lipids.
POST TRAUMATIC STRESS DISORDER RESOLVED BY
PHEROMONES
PTSD patients display smaller amygdalae and reduced anterior cingulate gray matter density on MRI scans of the brain.39 Reductions have also been found hippocampus and rostral anterior cingulate cortex in PTSD. 40 Thus, PTSD is impaired extinction of fear.41 So why would we need biological cowards? Now that question can be answered.
252

The two amygdalae are pheromone association areas of the brain. You see, animals need to back off the established territories of their conspecifics to avoid chaos in divisions of resources for feeding and breeding, they need innately respect the property of others and without law and order, they need to be biologically afraid. This is why Konrad Lorenz’s trespassing rats climb trees in fear of smaller rats who had previously and quite carefully marked their territory.
Omega-3 fatty acids may provide relief from PTSD.42 Fish oil seems to be a poor substitute for kisses. The similar human kissing pheromone mixture, synergistic and species-specific, must be even more beneficial against PTSD.
The treatment for PTSD is located at the end of this volume. That protocol essentially increases one’s hierarchical status, biologically increasing the rank of the patient. Post Traumatic Stress Disorder now becomes a trivial affliction, easily remedied, with serial administrations of 150 to 250 mg of healthy over 30 year old adult male facial skin surface lipid by mouth on a chewing gum vehicle. The MRI should note improvements and guide titration of the dose, first observing then allowing for regeneration time. Over doses of the facial skin surface lipid would reward cowardly privates with artificially enhanced self-confidence of major generals, and thus these should be avoided. Officers in the field can stimulate resolve and increase morale by providing soldiers with the paternal facial pheromone, as well as the pheromones of loved ones left behind.
Ancient war fighters leave no reports of PTSD in returning veterans. Pheromones are why. We scrub our soldiers clean to protect them from disease but that destroys the microflora that give them band loyalty. The same microflora are transferred by handshaking and backslapping. In addition, our soldiers suffer from pheromone deficiencies that cause difficulties during their service as well as upon their return.
EXOCRINOLOGY
253

ALARM PHEROMONE IN INFECTIOUS DISEASE
SUSCEPTIBILITY AND CANCER
At this author's suggestion, alarm pheromones were tested to demonstrate their ability to affect immunity, suppressing cell-mediated immunity and enhancing humoral in laboratory mice.43 Alarm and anxiety in human beings diminish disease resistance44 and predisposes to cancer.45 Pheromones of alarm in humans diminish resistance to the cancer cells that are thought to arise spontaneously, giving rise to cancers. As the author has had no interest in oncology and only the slightest training in medical school, this area will be left to others. The first place to begin will be to observe for beneficial effects of skin surface and other pheromones in cancer patients of all types and stages, then onto anti-stress pheromone manipulations. Stressful pheromone emitters in waiting rooms on cancer wards likely increase cancer risk.
AUTOIMMUNE DISEASE: DIABETES
Type I Diabetes (known as juvenile onset or insulin dependent diabetes mellitus) should respond well to oral administration of adult male facial skin surface lipids of about 150-250 mg doses as needed. This human male facial skin surface lipid pheromone therapy has never been tried, but given its innocuousness (the equivalent of kissing Daddy a lot), it should be. The author predicts that progression of the disease will be halted, perhaps with full and immediate recovery. Full recovery even after the loss of all beta pancreatic cells may be had with pheromone therapy alone, but almost certainly with epigenetic therapy.
Why does the author have such high hopes for curing this illness? After all, nothing else has ever worked.
The disease provides human population ecological benefit in a reliably regular syndrome: it kills people quickly. Expedient jettison of underperforming troupe members would be a hallmark of Type I Diabetes. Secondly, diabetes is described as
254

an acceleration of aging, and pheromones can affect the insulin signaling pathway to enhance experimental longevity.46, 47
In small open trials, the healthy adult male facial skin surface lipid pheromone seems to reliably halt the progression of Alzheimer’s disease, an autoimmune disease. The eating disorders have characteristics of autoimmune disease, too, and they have autoimmune characteristics. The prospect for pheromone and epigenetic therapies gives hope for all autoimmune disease sufferers everywhere.
Given that modernity has seen dramatic increases in the prevalence of Type II, while Type I is no more prevalent than in olden times, air pollution is unlikely to be involved in the etiology for Type I but it might be for Type II. That would put Type II and Alzheimer’s disease in the same category of increasing prevalence.
The seasonal onset of Type I in winter and early spring suggest closer proximity to the pathological pheromone emitter from cold huddling. Critical periods of susceptibility between ages 5-7 and 11-13 suggest sinus development specific to those ages increases risk of developing Diabetes Type I. Thus a larger, heavier, specifically targeted chemical would first insult a person to diabetes, perhaps a toxic pheromone receptor protein. The episodes of pheromone reception (tears in the patient) and emission (anger/disappointment in the pheromone poison source person) associated with onset episodes are also indicative.
The Human Lucocyte Antigen (HLA) can be discerned via pheromonal human chemosensory analysis,48 while HLA antigens, particularly DR3 and DR4, have strong association with Type I Diabetes. Pheromone receptor protein (murine major urinary protein-1) abundance in the liver is affected by the insulin-sensitizing drug rosiglitazone,49 which is highly suggestive, while the same pheromone receptor protein regulates glucose and lipid metabolism,1, 4 both of which are disjointed in diabetes.
Diabetes diminishes olfaction and may even cause complete anosmia, while insulin can improve olfaction in mice.50 Diabetic and non-diabetics have different protein sets in their olfactory bulbs.51
EXOCRINOLOGY
255

The sex ratio of 3:1 females to males preponderance likely puts Type II into the volatile pheromone group (along with most autoimmune diseases), since sex ratios of pheromone induced puberty delay (AN) are similarly skewed male.
The resolution of diabetes gravitas with delivery suggests that feto-receptor proteins also clearing the system with delivery, may be involved, too.
Moreover, a biochemical process for the cause of diabetes has recently presented itself. The reader will remember that medium and especially long chain free fatty acids of unusual chain length, unsaturation position, methylation, etc. are proposed to be human pheromones (see Chapter 3). Free fatty acid receptors, GPR120 and GPR40 whose endogenous ligands are medium- and long-chain free fatty acids, are thought to play an important physiological role in insulin release.52 If these puppies are pheromone receptors (which they certainly are), then their stimulation with the pheromone of the human face should benefit normal release of insulin, opposing Diabetes Type I and II, curing or at least halting the progression of diabetes.
AUTOIMMUNE DISEASE: JURJANI’S OR GRAVES DISEASE
The sex ratio of 5-10:1 females to males preponderance likely puts Graves disease into the volatile pheromone group (along with most autoimmune diseases), since sex ratios of pheromone induced puberty delay (anorexia nervosa) are similarly skewed male. Frequent adolescent onset also is suggestive for the same reason. Waxing and waning of symptoms suggests pheromone etiology, too.
Excessive lacrimation is a symptom of Graves‘ ophthalmopathy, and given that pheromones have been discovered in emotional tears recently at this author’s suggestion53, lacrimation in excess may signal the pheromone receptivity necessary to overcome the illness.
The improvement of some clinical symptoms with beta-blocker therapy has suggested disruption of adrenergic pathways between the locus ceruleus and the
256

frontal lobe.54 Both brain structures maintain pheromonal neural pathways.55 56 57 Thus pheromone therapy alone or combined with epigenetic chemotherapy should be curative.
!
AUTOIMMUNE DISEASE: LUPUS
Inflammation is a pheromone-mediated response, generally. Systemic lupus erythematosus (SLE) waxes and wanes as a pheromone-mediated condition might. This suggests episodic activity. Lupus’ association with stress suggests the sebaceous anti-stress pheromones should diminish attacks.
Lupus responds to NSAIDs, which are anti-pheromonal. Response to NSAIDs is a red flag for pheromone-mediated ills. The body’s defenses like inflammation, can be stimulated by alarm pheromones.58 Non steroidal anti-inflammatory drugs block the breakdown of long chain free fatty acids.
Lupus also responds to anti-malarials. Quinine, which, like aspirin, derives from tree bark and serves as anti-herbivory allomones. (Quinine and indeed, the caffein in tea and coffee, are both insect repellants/antifeedants and the same fruit fly taste receptors for them also detect the male-avoidance cuticular pheromone.59) Qinidine is a diastereometic stereoisomer of quinine (roughly same chemicals, same bonds, but “different handedness”). Quinine helps lupus, quinidine can cause a form of lupus, both are telling.
Quinidine, procainamide, hydralazine, isoniazid, and phenytoin may cause appearance of anti-nuclear antibodies that signal for lupus in the blood and reversible, drug-induced lupus symptoms. Qinidine and procainamide are both local anesthetics that diminish cardiac arrhythmias. Qinidine is, as mentioned, the diastereomer of the anti-malarial quinine and found with it in the cinchona tree bark. Like many pheromones that depend upon stereochemical shapes, the effects of qinidine and quinine differ dramatically in their medicinal effects on humans. Procainamide helps some arrhythmia patients that fail to respond to quinidine, and marked variation in bioavailability of an active metabolite seems to be genetically (or perhaps
EXOCRINOLOGY
257

epigenetically) dependent. Procainamide is a derivative of procaine anesthetic which renders animals temporarily anosmic, unable to smell.
Hydralazine lowers blood pressure, another likely pheromone-mediated illness since blood pressure responds to stress. Hydralazine also seems to improve murine multiple sclerosis,60 another likely pheromone-mediated illness (in humans). (Interestingly, CNS myelin--white matter, degraded in MS, is promoted by maternal pheromone of the lactating rat.61)
Isoniazid is, of course, the anti-TB antibiotic that will turn the urine orange. Anti-biotics as a class are allomones or artificial allomones. Isoniazid also has anti-depressant activity and is among the most common causes of drug-induced seizures in the US.62
Phenytoin is the Dilantin used to diminish epileptic seizures. Epileptic seizures seem to respond to human pheromone anecdotally. Phynytoin, like procainamide and quinidine, acts as a cardiac anti-arrhythmia drug.
Abnormal protein levels in the urine associated with lupus indicate kidney involvement. The kidneys in many species emit protein binding proteins as pheromones in exactly the same way as lupus affected kidneys. A lipocalin (a protein family that includes pheromone receptor proteins) is a biomarker for lupus nephritis.63, 64
Brain deficits in severe SLE can be localized to the cerebellum’s atrophy. The cerebellum in humans: fine motor skills. The cerebellum’s neuron generation is affected by pheromones in goldfish.65
Being first to suggest that SLE is a human pheromone response means that all these secondary associations are the only ones available. Hopefully, epigenetic and pheromone therapy will alleviate this illness or at least ameliorate its course. Once the pheromonal poison responsible for systemic lupus erythematosus is discovered, semiochemical aseptic techniques and interventions may prove to be of therapeutic benefit.
258

AUTOIMMUNE DISEASE: ARTHRITIS
Pheromones must control the incidence of juvenile rheumatoid arthritis. HLA-B27 histocompatibility antigen is associated with risk of evolving spondyloarthropathy in older boys with pauciarticular JRA. HLA-B27 heavy-chain homodimer formation can be induced by dendritic cell activation.66 Dendrites, the reader will recall, are the nerve cell ‘roots’ that respond to pheromone receptor protein activation as explained in Chapters 6 &7 of this volume. So, pheromone hits pheromone receptor protein, which activates the dendritic cell, which induces HLA-B27 to poison the joints in arthritis some way we do not yet understand.
Weaker HLA associations exist for other subtypes (HLA-DR5, HLA-DR8, HLA-DR4). Of course the HLA Human Leucocyte Antigen or Major Histocompatibility Complex is just a dense set of genes on chromosome #6. The MHC (the name of this homologous area for all animals except humans) controls immunity and chemorecognition, which includes pheromone recognition. Of course, pheromone recognition is, essentially, the immune system’s ability to detect self from non-self. MHC class I (not olfaction located in the class III area) generated proteins respond to pheromones to block pregnancy in the mouse.67 And the MHC class I genes influence the amounts of testosterone-mediated pheromones.68 The skin surface lipid human pheromone (see appendix) may halt the progression of rheumatoid arthritis.
Flooding pheromone reception pathways with ‘good father’ pheromones via the protocol in the appendix should alleviate arthritis and return early onset patients to good health. Concomitant epigenetic therapy with DNA hypomethylation drug 5-azacytidine or equivalent may be needed to ‘reset’ cell nuclei for the influence of the new pheromone, particularly when remodeling of tissues to normal is promoted. Alternatively, epigenetic therapy may not be required, since DNA hypomethylation has been observed in rheumatoid arthritis synovial fibroblasts69 implicating a pheromonal or pheromone deficient causative hypomethylation. We should try the pheromone alone first, and then go from there.
EXOCRINOLOGY
259

AUTOIMMUNE DISEASES: DEPRESSION
General anesthetics like halothane, enflurane, and ether inhibit GABA, gamma-aminobutyric acid, catabolism in the brain. That leaves more GABA, for more neural inhibition, for more anesthesia to avoid the pain and memory of surgery, maybe. Basically, general anesthetics are just harmless solvents that loosen up the oligodendritic (connected) and Schwann cell (unconnected) capacitors that run up and down most nerve axons, increasing the dielectric in the gaps between capacitance holding conductive ‘foil’ and keeping signals from accomplishing saltatory conduction. See Chapter 11.
Metabotropic gamma-aminobutyric acid(B) (GABAB) receptors for the major inhibitory transmitter GABA, together with metabotropic glutamate (mGLuRs) receptors, the extracellular calcium-sensing receptors (CaSRs), some V2R70 pheromone receptors and T1R taste receptors, belong to the family of 3 G-protein-coupled receptors (GPCRs). GABAB receptors are known to control neuronal excitability and modulate synaptic neurotransmission, playing an important role in many physiological activities. These receptors are widely expressed and distributed in the nervous system and have been implicated in a variety of neurodegenerative and pathophysiological disorders including epilepsy, spasticity, chronic pain, depression, schizophrenia and drug addiction.71 Drug addiction’s cure will soon be a fait accompli and its cure should eventually drag the others into the light as well.
To form a functional receptor entity, GABAB receptors must exist as a heterodimer consisting of GABAB1 and GABAB2 receptor subtypes with two 7-transmembrane proteins, and these subunits arise from distinct genes. The GABAB1 subunit binds the endogenous ligand within its extracellular N-terminus, whilst the GABAB2 subunit is not only essential for the correct trafficking of the GABAB1 subunit to the cell surface, but is also responsible for the interaction of the receptor with its cognate G-protein.10
260

Sound familiar? The GABAB1 subunit seems to be a pheromone receptor protein and probably does absolutely no ferrying or ‘trafficking’ of any kind. The reason it is responsible for the interaction of the receptor is because it is changing conformation (the reason for the sulfide bridges, etc.) to release or display ions to alter nearby dendritic potential difference: voltage. Voltage, depolarization from chemical ion presence just outside the dendrite is perceived centrally via EPSP.
Post-partum depression is induced usually on the third day after birth just before fourth day neonatal emissions onto kissable skin surfaces of sebaceous pheromone. PPD stimulates lacrimation with bond-enhancing pheromone receptor proteins in it. (See chapters 6 & 7.) The cure for this post-partum depression might be to transfer sebaceous secretions of the forehead and face of the neonate or infant during the first year of life into the mouth of the post-partum depression patient. That is what the pheromone receptivity is there for, after all, to bond the mother to the infant. The extreme receptivity of post-partum depression may be compensating for some other difficulty in the pheromone recognition system as it is seen less in experienced mothers, so this is probably not autoimmune in nature at all, except perhaps in the general sense.
Depressed people shed slightly more emotional tears than people not suffering from depression and there is a sexual dimorphism of tears and of depression with female preponderance. Perhaps depression is just a state of extreme pheromone receptivity that feeds back on itself, perhaps due to pheromone poisoning by some ‘hot head.’ Oral administration of 150 to 250 mg of healthy adult male facial skin surface lipid taken by mouth on chewing gum vehicle should alleviate the condition instantaneously with or more probably without coordinated epigenetic therapy.
Fluoxetine, an antidepressant in humans, disrupts reproductive behavior in goldfish72 and rats.73 Exposure to female pheromones during pregnancy causes postpartum anxiety in mice.74 All are suggestive even if you have to think about it a little.
EXOCRINOLOGY
261

AUTOIMMUNE DISEASE: ANKYLOSING SPONDYLITIS
Ankylosing spondylitis is strongly associated with the major histocompatibility complex (MHC) class I antigen HLA-B27. Multiple diverse infections and allergies were seen in these patients.75 HLA-B27 is observed on dendrites after their stimulation.33 Recall that pheromones interact with pheromone receptor proteins released from dendrites into intercellular spaces to stimulate dendrites in the model of chemosensory transduction explained in Chapter 7 of this tome.
Here is a good time to point out that this broad spectrum psychopatholytic and anti-autoimmune human pheromone is so obviously innocuous (people never despair of kissing, do they?) and so readily available everywhere (male researchers need literally look no further than the ends of their own noses), that clinical trials of the pheromone should be brought rapidly against every disease, illness, or condition. Physicians remember how many years they tolerated bacterial stomach ulcers while literally holding the tools to defeat it in our idle hands. A sample protocol is appended with clear instructions to patients, physicians, pheromone donors, and technicians to begin clinical trials to test the benefit of healthy adult male facial skin surface lipid p.o. instantly.
AUTOIMMUNE DISEASES: ASTHMA
Vasopressin exerts a local anti-inflammatory effect on the lung through the V2R in a murine model of sepsis.76 It would make sense that pheromone receptors (V2R) of the lung would be involved in asthmatic attacks. Lung inflammation may be influenced by perception of pathogenic pheromones (kairomones to us) emitted by pneumonia’s various septic causes as well as detrimental human pheromones setting them off. V2R and V1R pheromone receptors may be among the huge contingent of
262

microvillar pheromone sensory cells that cover so much of the human upper respiratory system.
Among the first allergens in animal dander to be categorized is a lipocalin,77 an obvious pheromone reception protein in this book’s model of pheromone reception. Dander is generally pheromonal to the species from which it comes, it erupts from the skin and has species-specific value.
In addition to pheromone-laden dander, pollens being gametes emit pheromones and even allomones,78 yes, as do mold spores (more gametes), house dust (mites),79 and feather pillows (the reader will remember the uropygial sebaceous glands of waterfowl from Chapter 3 on pheromone chemistry) are also common allergens. Various low molecular weight chemicals can cause occupational asthma80 among these are isocyanates which have pheromonal effects in a number of species,81,82 aldehydes which are also commonly pheromonal,83,84 as are anhydrides85 colophony,86 sulfates,87 amines,88,89 acrylates,90 dyes are used to trace pheromones and pheromone reception pathways,91 and, of course, metallic salts especially odd metallic salts, take key roles in pheromone reception. Oh my! Everything that triggers asthma is or is potentially of a pheromonal nature. Perhaps that is because asthma is itself, a disease of pheromonal etiology or perhaps it is mere coincidence. Testing can find out.
As asthma waxes and wanes, and since many, nearly half of juveniles ‘grow out’ of it reaching adulthood, asthma should respond favorably to oral administration of 150-250 mg of human adult male facial skin surface lipid on a chewing gum vehicle. Please see appendix for protocol, collection method, and so on.
EXOCRINOLOGY
263

DISEASES OF ERSATS HUMAN PHEROMONES: CRIME AND
OBSESSIVE COMPULSIVE DISORDER
Criminal behavior and obsessive compulsive behavior can be attenuated by brain surgery, commonly called lobotomy, to the human pheromone reception, association, and processing pathways. The surgery was developed empirically from beneficial CNS injuries with outcomes training the surgeon and guiding the knife (or icepick) to the orbitofrontal area of the brain through the ocular orbit by lifting up the eyelid.
Criminal behavior and OCD share dysfunction of the prefrontal-striatal-thalamic circuit. The orbitofrontal area unconsciously processes pheromones92,93,94,95,96 with the majority of reports in humans. Orbitofrontal lesions may produce abnormal social conduct.97
The amygdala processes pheromones, too, another area for lobotomizing surgeons bent on curing violent compulsive psychopaths. Gray matter density in the left superior temporal lobe and the left orbitofrontal cortex diminishes in suicidal schizophrenics.98 The two amygdalae and the orbitofrontal cortex limbic network underpin the neurobiological basis of psychopathy (attempted murder, multiple rape kidnappings).99,100 Schizophrenic psychopaths have blunted amygdala responses to fearful faces.101 Interestingly, left amygdala volume diminishes and right amygdala volume increases with severe early institutional deprivation.102 These disturbances of pheromone processing will abate with paternal facial skin surface pheromone therapy and MRI scanning can provide documentary evidence of cure.
Human pheromone deficiency of both the parents typifies criminal behavior (stealing property, gambling, robbing, burglary, truancy, delinquency). Recall a sufficiency or excess of maternal facial skin surface lipid coupled with a deficiency of paternal facial skin surface lipid seems to result in homosexuality. Criminal gang loyalty might result from paternal deficiency and/or maternal deficiency temporarily sated by gang comfort pheromones and the paternal pheromone deficiency amelioration of criminal thrills.
264

Criminal behavior declines in high winds,103 plausibly because high winds dissipate autoemissions or victim emissions during crimes which attenuate the paternal pheromone deficiency. Criminal behavior increases with ground-level ozone concentration104 perhaps also because ozone irritates chemoreceptive mucosae and because ozone attacks pheromones via ozonolysis.
Scattered anecdotal reports from newspapers suggest that serial appetitive rapists may prefer unintoxicated victims. Such rapists can attack elderly women who can still produce alarm pheromone. Since rapists frequently fail to inseminate victims, pheromonal emissions from the victim may be the addictive pheromone at play. Appetites for torture and rape may evolve from pubertal sexual horseplay and scary movie dates. An addiction to alarm pheromone may plausibly be the device behind human brutality and sexual assault for sexual predators, rapists, and torturers. If so, identifiable human pheromone receptor proteins may divulge the propensity preemptively.
Juvenile OCD patients share disfunction of the prefrontal-striatal-thalamic circuit with involvement of other basal ganglia structures (putamen globus pallidus) and the thalamus in contrast to adults which report mainly involvement of the caudate nucleus and orbitofrontal cortex.105 This ripening of pheromone deficiency responses may reflect changing bonding pheromone preferences of ontogeny, perhaps. The transition suggests useful clinical observations during experimental administration of paternal facial skin surface lipid human pheromone. One might expect a return to the juvenile phase of brain activity during recovery.
Another group using different methodology found consistent activation in cortical and subcortical regions of the orbitofrontal and anterior cingulate loops. The left dorsal frontoparietal network, including the dorsolateral prefrontal cortex and precuneus, and the left superior temporal gyrus were likely OCD activated.
The orbitofrontal area unconsciously processes pheromones106,107,108,109,110
with the majority of reports in humans. The lateral prefrontal cortex and the superior temporal cortex are both activated by the airborne pheromone androstadienone in humans.111 Recall that pheromone and olfactory pathways run parallel part of the
EXOCRINOLOGY
265

way into the CNS. It should come as no surprise that there is impairment of olfactory identification in obsessive-compulsive disorder.112,113
OCD and depression can benefit from electroshock therapy. Perhaps the mechanism renews pheromone receptor proteins much the way blood bubbles in an electric field. OK, this seems strange. You see, hemoglobin loses its load of carbon dioxide and 2,3 bisphosphoglycerate in the right ventricle by the QRS transient electric field’s tugging at unlike charges. Understand that when water or any polar liquid is subjected to electric fields, as in the heart, all the dipoles align--then the released dipoles swing out of alignment and then back again into alignment. This is how the T-Wave, J-Wave, and U-wave show up on an EKG. All the swinging and tugging gets the ions and partial dipoles separated, hemoglobin turns somewhat buff colored immediately before being pumped into the lung for oxygenation and adoption of the familiar red conformation. 2,3 BPG’s higher affinity for hemoglobin now goest to work to deliver oxygen from the blood to the tissues.
Something of the same sort probably takes place in the brain under electroshock therapy. The pheromone receptor proteins throughout the pheromone reception system and the brain are ‘cleaned’ of charged sticky sludge and become ready to receive normal social pheromonal receptions again, roughly. It is cool to watch this on a microscope slide with the naked eye, blood turns yellow and effervesces in a small electric field. Shock-caused conformational changes of olfactory receptor proteins change surface reflectance, and alter their twist under UV irradiation. This should not prove difficult to more formally observe.
OCD has been linked to abnormalities with the neurotransmitter serotonin. Animal pheromone reception is modulated by serotonin.114 Indeed, animal pathological grooming disorders respond to serotonin enhancing drugs.115 Please recall animals groom and allogroom for sebaceous pheromones.
OCD patients will respond favorably to treatment with healthy adult male facial skin surface lipids taken by mouth, as should patients with trichotillomania, body dysmorphic disorder, olfactory reference syndrome, compulsive skin picking,
266

compulsive wound picking, and onychophagia. Please see the appendix at the end of the book.
DISEASES OF ERSATS HUMAN PHEROMONES: PHOBIAS
Phobias replace pheromone with terror. One unreportable case of severe agoraphobia complicated by mitral valve prolapse has been cured by the author. These fears will mostly succumb to the paternal/parental protocol of the appendix.
DISEASES OF ERSATS HUMAN PHEROMONES: GILES DE LA
TOURETTE’S SYNDROME
Tourette’s syndrome is similar to Latah, criminal behavior, obsessive compulsive disorder and compulsive gambling in that an appetite builds for the tic behavior. The syndrome is relieved by rest and exaggerated by stress. Tourette’s waxes and wanes, tics come and go, and up to 50% of patients show marked improvement after a good rest and progressively during the teenage years. Tics kept past 15 years tend to remain chronically.
The author suggested the use of mecamylamine,116 the most central acting nicotinic anti-hypertensive drug, in place of raw nicotine being used to treat severe Tourettes at a local college, because of the devastating peripheral side effects that the nicotine being used had on the children. Of course, the equivalent of having Tourettes patients smoke cigarettes was not truly therapeutic, and the trials failed.117 Human pheromone therapy for one Tourette’s syndrome patient improved his condition and seems to make up for the author’s earlier transgression. Tourette’s syndrome patients may benefit from paternal facial skin surface lipids alone, however
EXOCRINOLOGY
267

optimal therapy may include maternal facial skin surface lipids as well. The logic for paternal (and perhaps maternal) facial skin surface therapy is that stress exaggerates the condition. Axillary stress/alarm/fear pheromones may work in opposition to pheromone reception of facial skin surface lipids. Pheromones working in opposition can overcome previously pheromone-stimulated behavior in mice.118 Paternal facial skin surface lipid pheromone from an over 30 year old male donor, should overcome and cure Tourettes’ syndrome, perhaps even resolving tics in adults if the pheromone is provided in concert with epigenetic therapy and/or artificial pheromone receptivity enhancement by psychogenic lacrimation (watching sad movies).
DISEASES OF ERSATS HUMAN PHEROMONES: GAMBLING
Gambling addiction, is very similar to divorce, delinquency, and running away, as a criminal behavior. The act of gambling obtains hormonal stimulation rewards and probably autoemission and autoreception of pheromones from risky behavior, just like other criminal and delinquent behavior. Gambling addiction, like them, should be amenable to oral administration of healthy human adult male facial skin surface lipid ‘kissing daddy pheromone’ 150 mg or more in one single dose. See appendix.
AUTOIMMUNE DISEASES: ALS
Amyotrophic lateral sclerosis progresses at various rates, probably dependent upon semiochemical exposures which vary from individual to individual. Happy people last longer. Uncontrollable periods of laughing or crying seen in many ALS patients suggest pheromone receptivity is being stimulated to contest against the disease, just as uncontrolled crying of the 3rd day blues contests against bonding failure in new mothers. This indicates that the disease should be treatable with human
268

pheromone to provide a miracle cure for the disease. Thus after oral administration of healthy adult male facial skin surface lipid 150-250 mg, the progression of ALS should stop, followed by complete remission of the disease, the disappearance of all symptoms and complete and rapid asymptomatic recovery.
Schwann cells can depend upon pheromones for normal development in laboratory animals, thus autoimmune diseases that affect myelination may be simple pheromone deficiencies. Given their life-threatening nature, they are more likely to be caused by unfortunate sympathetic reception of poisonous human pheromones. The nearly even sex ratio likely dismisses axillary contribution, but this is only an informed guess.
GABA is diminished in the CSF with increased noradrenalin in both blood and CSF (it passes the bbb, blood-brain large molecule barrier).
Flexible pheromone commerce needs feedback. There are curious physical skin changes119 in ALS and weird immunity from bedsores, even at the terminal stage.120 A similar weird immunity to respiratory illness is seen in Anorexia Nervosa patients.121 ALS patients are more likely to have high densities of Interleukin-6 (IL-6) in the epidermis. IL-6 family proteins act as pheromone receptor proteins.122 Increased amyloid beta protein has been found in the skin of patients with ALS.123 Increased expression of insulin-like growth factor I has been found in ALS.124 Insulin-like growth factors respond to pheromones125 and are probably pheromone receptor proteins. ALS patients have impaired glucose tolerance.126
Eccrine perspiration was elevated (hyperhydrosis) early in ALS, while perspiration declined by 40% over a six month interval.127 This suggests an attempted biological self-remediation of poisonous pheromone reception.
AUTOIMMUNE DISEASE DUE TO BIRTH DEFECTS:
EXOCRINOLOGY
269

There is one report of mandibuloacral dysplasia improvement after chewing the pheromone chewing gum. Premature aging progeriod disorders may respond to paternal facial skin surface lipid pheromone treatment, too.
One of the problem genetic loci known for MAD is for the zinc metalloproteinase (ZMPSTE24),128 which may cause MAD by affecting prelamin A processing.129 ZMPSTE24 was originally discovered in yeast as an enzyme required for maturation or processing of the mating pheromone a-factor.130 These diseases are plainly genetic, but the author has strangely benefitted one case with 150-250 mg healthy over-30 adult male facial skin surface lipid provided per os and he is at a loss as to why. Perhaps this is an epigenetic effect?
CYSTIC FIBROSIS:
Cystic fibrosis and other inheritance-based genetic abnormalities may benefit from pheromone treatment as a palliative, but hope for a cure is unforeseen. (Of course, we thought the same thing about drug addiction.) With epigenetic therapy, there may be hope of some relief in some patients.
On a brighter note, the author noticed that the cystic fibrosis protein defect in transmembrane ion transporter molecules effected loss of magnetic containment to disable transport. Ion transporter proteins generally contain particle confinement fields that allow ions to pass membranes without waters of hydration. (Artificially adjoining these will potentially provide biological ‘plasma conduits’ consisting of long proteins that need no external magnetic containment field, no vacuum pumping, and no metal conductors.) My point here is to propose a novel radiological treatment for easing symptoms of cystic fibrosis. Opening those microscopic containment fields to the passage of ions may be attempted with exogenous magnetic fields. Electromagnets (and motors if necessary) are relatively inexpensive. Since magnetic fields are imperceptible, applying various electromagnetic stimulation to the most affected areas during rest periods should allow increased longevity and improved quality of life in cystic fibrosis patients.
270

You see, CFTR, the cystic fibrosis transmembrane conductance regulator, has significant amino acid homology and length conservation with alpha-factor pheromone export systems proteins, hemolysin B, the MHC-linked peptide transporter, HAM1.131 The CFTR lacks a pairing of juxtaposed magnetic (Hückel’s rule) amino acids right at the bottom of the tube that lets ions pass through without waters of hydration. The tube is so tiny that even water is excluded, so no vacuum pumping is needed and maintaining the magnetic fields requires no refrigeration, obviously, since it works at body temperature. Anyway, the highly suspicious homology (except for the magnetic defect that actually causes cystic fibrosis by preventing transmembrane transport of chloride ion) suggests a pheromonal association, so there actually is hope of a significant improvement with pheromone therapy. If this works, it is a gift from God. If it does not work, then it is an amazing coincidence.
PHEROMONAL ECLAMPSIA, PHEROMONAL PRE-ECLAMPSIA
AND PHEROMONAL PREMATURE LABOR
The control of phenotype from among a database of choices (oocytes and presented spermatozoa) is influenced by pheromones. More human fetuses are lost than delivered because choices are being made with pheromonal information having significant, even fatal influence.
Fetal growth is compromised in animal models with high cortisol availability and cortisol can be stimulated by 11beta-HSD2.132 The author believes that 11beta-HSD2 may be a human placental pheromone or receptor, for a number of reasons. First, 11beta-HSD2 shares the trouble caused by a pollutant with pheromone ‘transport’ proteins,133 suggesting it may have chemosensory receptor function. Second, cortisol can be stimulated in women by human pheromone from the male axilla.134
EXOCRINOLOGY
271

Cortisol is the stress hormone, increasing blood pressure, increasing blood sugar, and reducing immune responses. Pre-eclampsia and eclampsia (pre-eclampsia with seizures) are associated with diabetes mellitus of pregnancy, a risk factor. Thus the male may emit the airborne pheromone which causes the pregnant woman to secrete cortisol, which stimulates some aspects of pre-eclampsia: hypertension of pregnancy, gestational edema, and diabetes mellitus of pregnancy. Since only 5% of pregnant women suffer from pre-eclampsia and a tiny percentage of them reach eclampsia, and since pregnancy is strongly associated with males, some other device must be involved. That device may be a simple pheromone deficiency, perhaps a deficiency of healthy adult male facial or other skin surface lipid? Reduced pre-eclampsia is associated with increased fellatio and ingestion of semen.135 Koelman, Coumans, Nijman, Doxiadis, Dekker, and Claas suggested that soluble HLA found in semen transferred paternal immunity. Their findings suggest that soluble HLA may be a pheromone receptor protein that conveys pheromone recognition. Epidemiological studies attribute the cause of preeclampsia to genetic effects of the mother, both mother and father, a couple effect, and unmeasured factors such as the effect of the length of the sexual relationship and even a ‘dangerous’ father effect.136 All suggest pheromonal etiology and pheromone treatment.
Cortisol levels are higher in constitutional delay of puberty in girls.137 Male airborne pheromone increases cortisol levels in women, but males generally stimulate acceleration of puberty! Honestly, the likelihood of elucidating every pheromone response from browsing the medical and scientific literature is not very high. Pheromone effects on human physiology and disease are, happily, easy to test.
This author discovered that Chronic Fatigue Syndrome and obesity-related spacial perception changes responded to the application of thick gobs of petroleum jelly under the arms. The jelly prevented the auto-emitted axillary pheromones from reaching the nose. Similar axillary petroleum jelly therapy might control hypertension, diabetes, Alzheimer’s disease, anorexia nervosa, and bulimia nervosa. Alternatively, anti-septic bathing may also be beneficial for these pathological conditions, as it destroys the skin surface microflora that
272

is thought responsible for processing axillary apocrine emissions into human pheromones. This is just an idea, but it might just work.
Recently, skin surface microflora have been examined across and within species of amphibians utilizing genetic sequencing techniques.138 This group recognized stable communities of microflora that differed from place to place, even anatomically, just as this author found with his own nose many years ago.
PHEROMONAL CAUSATION OF PSYCHIATRIC DISORDERS:
The development of the tics of Tourette's syndrome, the obsessions of OCD, the binge eating behaviors of bulimia, and the self-starvation of anorexia share ‘abnormal development of [pheromonal-BN] frontostriatal circuits.’139 Epilepsy, spasticity, chronic pain, depression, schizophrenia, and drug addiction should respond favorably to human adult male facial skin surface lipids taken by mouth. Drug addiction has been treated with complete success in three cases so far (2011) and epilepsy, one. Epileptic lesions and surgical interventions affect the pheromone reception and processing pathways in the brain. Meanwhile, alarm pheromones have contributed to clonic epileptic convulsions in rats.140
Schizophrenics and other psychiatric patients suffer from mild alterations in immune functions and their illnesses have autoimmune features. Autoimmune sufferers succumb to more than their fair share of psychiatric disorders, too, frequently with olfactory impairment. A large cluster of olfactory receptor genes has been mapped in proximity to the HLA/MHC locus on chromosome 6. The HLA/MHC and linked olfactory receptor genes on chromosome 6 are clustered in haplotypes and are highly polymorphic.141
Impulsive dysfunctional schizophrenics maintain lower orbitofrontal and hippocampal volumes. Orbitofrontal cortex is dysmorphic in criminals while both are pheromone processing and memory areas.
EXOCRINOLOGY
273

Schizophrenia does not plague us until after puberty. It is similar to Multiple Sclerosis (MS), Anorexia Nervosa (AN), and Bulimia Nervosa in this regard. Might peri-pubertal onset point to development of secondary sex characteristics as playing some role? And dwelling with one’s family reduces schizophrenic symptoms and increases symptom-free episodes. Family therapy is effective in treating schizophrenics while friend therapy fails. What differs? Kissing differentiates the two. That certainly suggests that susceptibility to schizophrenia follows from some family pheromone deficiency. It might be the father’s emissions, or the mothers, or even baby’s, but it would be less likely to be pheromone emissions from sibling children because their levels are low, and we don’t kiss our siblings that much anyway.
Sad to say, exaggerated emission of a short-chain lipid, trans 3-methyl-2-hexenoic acid normally found in human perspiration and produced by specialized skin surface and axillary scent gland microflora, is closely associated with schizophrenia. Indeed, the odor is characteristic142 although that is disputed. For some inexplicable reason, no one has ever observed if schizophrenia improves when the 3-methyl-2-hexenoic acid is prevented from reaching the nose. Bathing schizophrenics in a bathtub of water with a quarter cup of ordinary household bleach and a quarter cup of vinegar should reset the microflora by antisepsis. Healthy microflora could then be transferred to the aseptic skin.
Incidentally, (E)-3-methyl-2-hexenoic acid is reputed to be a pheromone for Anopheles gambiae. which carries malaria.
Given the improved grades seen in delinquents and runaways after being supplied with the paternal facial skin surface lipid pheromone, given schizophrenia’s frequent onset after some life disturbance (as with diabetes I), schizophrenics should benefit from oral paternal pheromone therapy. Why not try it? Masculine facial sebum may be robust enough. Receptivity could be enhanced by examining Bowman’s secretion and inoculating with pheromone receptor proteins, transferring pheromonal microflora, or by merely providing some stimulus to psychogenic lacrimation (such as letting pheromone recipients view the Hollywood film, All Mine To Give), a good cry.
274

EXOCRINOLOGY
275
1 Ferreyra-Moyano H, Barragan E. The olfactory system and Alzheimer's disease. Int J Neurosci 1989 Dec;49(3-4):157-9.
2 Kurtz P, Schuurman T, Prinz H. Loss of smell leads to dementia in mice: is Alzheimer's disease a degenerative disorder of the olfactory system? J Protein Chem. 1989 Jun;8(3):448-451.
3 Doty RL, Snow JB. Age-related alterations in olfactory structure and function. In: Margolis FL, Getchell TV, editors. Molecular Neurobiology of the Olfactory System. New York: Plenum Press. 1988:355-374.
4 Pochi PE, Strauss JS, Mescon H. Sebum production and fractional 17-ketosteroid excretion in Parkinsonism. J. Invest. Dermatol 1962;38:45.
5 Nicholson B. Does kissing aid human bonding by semiochemical addiction? Br J Dermatol 1984 Nov;111(5):623-627.
6 Sabatier L, Jouanguy E, Dostert C, Zachary D, Dimarcq JL, Bulet P, Imler JL. Pherokine-2 and pherokine-3. Eur J Biochem. 2003 Aug;270(16):3398-407.
7 Brurberg MB, Nes IF, Eijsink VG. Brurberg MB, Nes IF, Eijsink VG. Pheromone-induced production of antimicrobial peptides in Lactobacillus Mol Microbiol. 1997 Oct;26(2):347-60.
8 Schlievert PM, Gahr PJ, Assimacopoulos AP, Dinges MM, Stoehr JA, Harmala JW, Hirt H, Dunny GM. Aggregation and binding substances enhance pathogenicity in rabbit models of Enterococcus faecalis endocarditis. Infect Immun. 1998 Jan;66(1):218-223.
9 Sandnabba N. Differences between aggressive and non-aggressive mice in odour signals and marking behaviour. In: Chemical Signals in Vertebrates. MacDonald DW, Müller-Schwarze D, Natynczuky SE, editors. Oxford: Oxford University Press. 1990:125-38.
10 Chamero P, Marton TF, Logan DW, Flanagan K, Cruz JR, Saghatelian A, Cravatt BF, Stowers L. Identification of protein pheromones that promote aggressive behaviour. Nature. 2007 Dec 6;450(7171):899-902.
11 Holmes, Thomas H., et al. (1950). The Nose; an experimental study of reactions within the nose in human subjects during varying life experiences with a foreword by Warfield T. Lonscope. Springfield, Ill: Thomas.

276
12 Wynne-Edwards VC. Animal Dispersion in Relation to Social Behaviour. Edinburgh: Oliver and Boyd. 1962.
13 Albone ES. Mammalian Semiochemistry The Investigation of Chemical Signals Between Mammals. Chichester, England: J Wiley & Sons Limited, 1984.
14 Weitkamp AW, Smiljanic A, Rothman S. The free fatty acids of human hair fat. J Am Chem Soc 1947 Aug;69:1936-9.
15 106. Cunliffe WJ, Burton JL, Shuster S. The effect of local temperature variations on the sebum excretion rate. Br J Dermatol 1970;83:650-654.
16 Hardy JA, Mann DM, Wester P, Winblad B. An integrative hypothesis concerning the pathogenesis and progression of Alzheimer's disease. Neurobiol Aging. 1986 Nov-Dec;7(6):489-502.
17 Tabunoki H, Shimada T, Banno Y, Sato R, Kajiwara H, Mita K, Satoh J. Identification of Bombyx mori 14-3-3 orthologs and the interactor Hsp60. Neurosci Res. 2008 Jul;61(3):271-80. Epub 2008 Apr 3.
18 Czarnecka AM, Campanella C, Zummo G, Cappello F. Heat shock protein 10 and signal transduction: a "capsula eburnea" of carcinogenesis? Cell Stress Chaperones. 2006 Winter;11(4):287-94
19 Christen WG, Schaumberg DA, Glynn RJ, Buring JE. Dietary {omega}-3 Fatty Acid and Fish Intake and Incident Age-Related Macular Degeneration in Women. Arch Ophthalmol. 2011 Mar 14; [Epub ahead of print]
20 Nicholson B. A semiochemical addiction model for human bonding. Paper presented to the Animal Behaviour Society and the International Society for Human Ethology, Bucknell University Lewisburg, PA. June 12, 1983.
21 Drickamer LC, Mikesic DG. Urinary chemosignals, reproduction, and population for house mice (Mus domesticus) living in field enclosures. J Chem Ecol 1990;16(10):2955-2968.
22 Drickamer LC. Puberty-influencing chemosignals in house mice: ecological and evolutionary considerations. In: Duvall D, Müller-Schwarze D, Silverstein RM, editors. Chemical Signals in Vertebrates 4. New York: Plenum Press 1986:441-456.
23 Nicholson B Pheromones cause disease: the exocrinology of anorexia nervosa Med Hypotheses 2000; 54(3): 438-443.
24 Bruce HM. An exteroceptive block to pregnancy in the mouse. Nature 1959;184:105.
25 Keverne EB, Rosser AE. The evolutionary significance of the olfactory block to pregnancy. In: Chemical Signals in Vertebrates 4. Duvall D, Müller-Schwarze D, Silverstein RM, editors. New York: Plenum Press. 1986:433-439.
26 Nicholson B. Human pheromones in the pathogenesis of autoimmune and behavioural illness. Paper presented, informally, to the International Society of Chemical Ecology, University of Hull, England, July 17, 1987.
27 Coleman-Phox K, Odouli R, Li DK. Use of a fan during sleep and the risk of sudden infant death syndrome. Arch Pediatr Adolesc Med. 2008 Oct;162(10):963-968.

EXOCRINOLOGY
277
28 Willis MA, Avondet JL, Finnell AS. Effects of altering flow and odor information on plume tracking behavior in walking cockroaches, Periplaneta americana (L.). J Exp Biol. 2008 Jul;211(Pt 14):2317-2326.
29 Valdes-Dapena MA. Sudden infant death syndrome: a review of the medical literature 1974-1979. Pediatrics 1980;66(4):597-614.
30 Darwin, Charles. The Expression of the Emotions in Man and Animals. London: Murray. 1872:162.
31 Shamim S, Hasler G, Liew C, Sato S, Theodore WH. Temporal lobe epilepsy, depression, and hippocampal volume. Epilepsia. 2009 May;50(5):1067-1071. Epub 2008 Nov 19.
32 Abel EL, Bilitzke PJ, Cotton DB. Alarm substance induces convulsions in imipramine-treated rats. Pharmacol Biochem Behav. 1992 Mar;41(3):599-601.
33 Rosenberg RE, Law JK, Yenokyan G, McGready J, Kaufmann WE, Law PA. Characteristics and concordance of autism spectrum disorders among 277 twin pairs. Arch Pediatr Adolesc Med. 2009 Oct;163(10):907-914.
34 Nicot A, Otto T, Brabet P, Dicicco-Bloom EM. Altered social behavior in pituitary adenylate cyclase-activating polypeptide type I receptor-deficient mice. J Neurosci. 2004 Oct 6;24(40):8786-8795.
35 Toal F, Daly EM, Page L, Deeley Q, Hallahan B, Bloemen O, Cutter WJ, Brammer MJ, Curran S, Robertson D, Murphy C, Murphy KC, Murphy DG. Clinical and anatomical heterogeneity in autistic spectrum disorder: a structural MRI study. Psychol Med. 2009 Nov 6;. [Epub ahead of print]
36 Zhou W, Chen D. Encoding human sexual chemosensory cues in the orbitofrontal and fusiform cortices. J Neurosci. 2008 Dec 31;28(53):14416-14421.
37 Wiggins LD, Robins DL, Bakeman R, Adamson LB. Brief report: sensory abnormalities as distinguishing symptoms of autism spectrum disorders in young children. J Autism Dev Disord. 2009 Jul;39(7):1087-1091. Epub 2009 Mar 13.
38 Meiri G, Bichovsky Y, Belmaker RH. Omega 3 fatty acid treatment in autism. J Child Adolesc Psychopharmacol. 2009 Aug;19(4):449-51.
39 Rogers MA, Yamasue H, Abe O, Yamada H, Ohtani T, Iwanami A, Aoki S, Kato N, Kasai K. Smaller amygdala volume and reduced anterior cingulate gray matter density associated with history of post-traumatic stress disorder. Psychiatry Res. 2009 Dec 30;174(3):210-216. Epub 2009 Nov 13.
40 Felmingham K, Williams LM, Whitford TJ, Falconer E, Kemp AH, Peduto A, Bryant RA. Duration of posttraumatic stress disorder predicts hippocampal grey matter loss. Neuroreport. 2009 Oct 28;20(16):1402-1406.
41 Milad MR, Pitman RK, Ellis CB, Gold AL, Shin LM, Lasko NB, Zeidan MA, Handwerger K, Orr SP, Rauch SL. Neurobiological basis of failure to recall extinction memory in posttraumatic stress disorder. Biol Psychiatry. 2009 Dec 15;66(12):1075-1082. Epub 2009 Sep 12.
42 Matsuoka, Yutaka. Clearance of fear memory from the hippocampus through neurogenesis by omega-3 fatty acids: A novel preventive strategy for posttraumatic stress disorder?BioPsychoSocial Medicine 2011, 5:3

278
43 Cocke R, Thiessen D. Alarm chemosignals suppress the immune system. In: Chemical Signals in Vertebrates V. MacDonald DW, Müller-Schwarze D, Natynczuky SE, editors. Oxford: Oxford University Press. 1990:125-138.
44 Salo M. Effects of anaesthesia and surgery on the immune response. In: Trauman, Stress, and Immunity in Anaesthesia and Surgery. New York: Butterworth Scientific. 1982:211-53.
45 Fan RL, Zheng SH, Wu ZS. [Study on the relationship between lung cancer at preclinical stage and psycho-social factor. A case-control study]. Chung Hua Lin Hsing Ping Hsueh Tsa Chih 1997 Oct;18(5):289-92.
46 Ogg S, Paradis S, Gottlieb S, Patterson GI, Lee L, Tissenbaum HA, Ruvkun G. The Fork head transcription factor DAF-16 transduces insulin-like metabolic and longevity signals in C. elegans. Nature. 1997 Oct 30;389(6654):994-999.
47 Hahm JH, Kim S, Paik YK. Endogenous cGMP regulates adult longevity via the insulin signaling pathway in Caenorhabditis elegans. Aging Cell. 2009 Aug;8(4):473-483. Epub 2009 May 31.
48 Pause BM, Krauel K, Schrader C, Sojka B, Westphal E, Müller-Ruchholtz W, Ferstl R. The human brain is a detector of chemosensorily transmitted HLA-class I-similarity in same- and opposite-sex relations. Proc Biol Sci. 2006 Feb 22;273(1585):471-478.
49 Hui X, Zhu W, Wang Y, Lam KS, Zhang J, Wu D, Kraegen EW, Li Y, Xu A. Major urinary protein-1 increases energy expenditure and improves glucose intolerance through enhancing mitochondrial function in skeletal muscle of diabetic mice. J Biol Chem. 2009 May 22;284(21):14050-7. Epub 2009 Mar 31.
50 Marks DR, Tucker K, Cavallin MA, Mast TG, Fadool DA. Awake intranasal insulin delivery modifies protein complexes and alters memory, anxiety, and olfactory behaviors. J Neurosci. 2009 May 20;29(20):6734-6751.
51 Dennis JC, Coleman ES, Swyers SE, Moody SW, Wright JC, Judd R, Zhong Q, Morrison EE. Changes in mitotic rate and GFAP expression in the primary olfactory axis of streptozotocin-induced diabetic rats. J Neurocytol. 2005 Mar;34(1-2):3-10.
52 Hara T, Hirasawa A, Sun Q, Sadakane K, Itsubo C, Iga T, Adachi T, Koshimizu TA, Hashimoto T, Asakawa Y, Tsujimoto G. Novel selective ligands for free fatty acid receptors GPR120 and GPR40. Naunyn Schmiedebergs Arch Pharmacol. 2009 Sep;380(3):247-55. Epub 2009 May 27.
53 Gelstein S, Yeshurun Y, Rozenkrantz L, Shushan S, Frumin I, Roth Y, Sobel N. Human tears contain a chemosignal. Science. 2011 Jan 14;331(6014):226-30. Epub 2011 Jan 6.
54 Ljunggren JG (August 1983). "[Who was the man behind the syndrome: Ismail al-Jurjani, Testa, Flajina, Parry, Graves or Basedow? Use the term hyperthyreosis instead]". Lakartidningen 80 (32–33): 2902. PMID 6355710.
55 Winberg J, Porter RH. Olfaction and human neonatal behaviour: clinical implications. Acta Paediatr. 1998 Jan;87(1):6-10.
56 Batterton MN, Robarts D, Woodley SK, Baum MJ. Comparison of odor and mating-induced glomerular activation in the main olfactory bulb of estrous female ferrets. Neurosci Lett. 2006 Jun 12;400(3):224-9. Epub 2006 Mar 10.

EXOCRINOLOGY
279
57 Pause BM, Lübke K, Laudien JH, Ferstl R. Intensified neuronal investment in the processing of chemosensory anxiety signals in non-socially anxious and socially anxious individuals. PLoS One. 2010 Apr 23;5(4):e10342.
58 Arakawa H, Arakawa K, Blandino P Jr, Deak T. The role of neuroinflammation in the release of aversive odor cues from footshock-stressed rats: Implications for the neural mechanism of alarm pheromone. Psychoneuroendocrinology. 2011 May;36(4):557-568.
59 Lacaille F, Everaerts C, Ferveur JF. Feminization and alteration of Drosophila taste neurons induce reciprocal effects on male avoidance behavior. Behav Genet. 2009 Sep;39(5):554-563.
60 Leung G, Sun W, Zheng L, Brookes S, Tully M, Shi R. Anti-acrolein treatment improves behavioral outcome and alleviates myelin damage in experimental autoimmune encephalomyelitis mouse. Neuroscience. 2011 Jan 26;173:150-5. Epub 2010 Nov 26.
61 Schumacher SK, Moltz H. Prolonged responsiveness to the maternal pheromone in the postweanling rat. Physiol Behav. 1985 Mar;34(3):471-473.
62 Minns AB, Ghafouri N, Clark RF. Isoniazid-induced status epilepticus in a pediatric patient after inadequate pyridoxine therapy. Pediatr Emerg Care. 2010 May;26(5):380-381.
63 Rubinstein T, Pitashny M, Levine B, Schwartz N, Schwartzman J, Weinstein E, Pego-Reigosa JM, Lu TY, Isenberg D, Rahman A, Putterman C. Urinary neutrophil gelatinase-associated lipocalin as a novel biomarker for disease activity in lupus nephritis. Rheumatology (Oxford). 2010 May;49(5):960-971. Epub 2010 Feb 9.
64 Brunner HI, Mueller M, Rutherford C, Passo MH, Witte D, Grom A, Mishra J, Devarajan P. Urinary neutrophil gelatinase-associated lipocalin as a biomarker of nephritis in childhood-onset systemic lupus erythematosus. Arthritis Rheum. 2006 Aug;54(8):2577-2584.
65 Chung-Davidson YW, Rees CB, Bryan MB, Li W. Neurogenic and neuroendocrine effects of goldfish pheromones. J Neurosci. 2008 Dec 31;28(53):14492-14499.
66 Santos SG, Lynch S, Campbell EC, Antoniou AN, Powis SJ. Induction of HLA-B27 heavy chain homodimer formation after activation in dendritic cells. Arthritis Res Ther. 2008;10(4):R100. Epub 2008 Aug 29.
67 Thompson RN, McMillon R, Napier A, Wekesa KS. Pregnancy block by MHC class I peptides is mediated via the production of inositol 1,4,5-trisphosphate in the mouse vomeronasal organ. J Exp Biol. 2007 Apr;210(Pt 8):1406-1412.
68 Novotny MV, Soini HA, Koyama S, Wiesler D, Bruce KE, Penn DJ. Chemical identification of MHC-influenced volatile compounds in mouse urine. I: Quantitative Proportions of Major Chemosignals. J Chem Ecol. 2007 Feb;33(2):417-34. Epub 2006 Dec 27.
69 Karouzakis E, Gay RE, Michel BA, Gay S, Neidhart M. DNA hypomethylation in rheumatoid arthritis synovial fibroblasts. Arthritis Rheum. 2009 Nov 30;60(12):3613-3622. [Epub ahead of print]
70 Ryba NJ, Tirindelli R. A new multigene family of putative pheromone receptors. Neuron. 1997 Aug;19(2):371-379.
71 Ong J, Kerr DI. Clinical potential of GABAB receptor modulators. CNS Drug Rev. 2005 Autumn;11(3):317-34.

280
72 Mennigen JA, Lado WE, Zamora JM, Duarte-Guterman P, Langlois VS, Metcalfe CD, Chang JP, Moon TW, Trudeau VL. Waterborne fluoxetine disrupts the reproductive axis in sexually mature male goldfish, Carassius auratus. Aquat Toxicol. 2010 Nov 15;100(4):354-64. Epub 2010 Sep 22.
73 Bataineh HN, Daradka T. Effects of long-term use of fluoxetine on fertility parameters in adult male rats. Neuro Endocrinol Lett. 2007 Jun;28(3):321-325.
74 Larsen CM, Grattan DR. Exposure to female pheromones during pregnancy causes postpartum anxiety in mice. Vitam Horm. 2010;83:137-49.
75 Zochling J, Bohl-Bühler MH, Baraliakos X, Feldtkeller E, Braun J. The high prevalence of infections and allergic symptoms in patients with ankylosing spondylitis is associated with clinical symptoms. Clin Rheumatol. 2006 Sep;25(5):648-658. Epub 2005 Dec 23.
76 Boyd JH, Holmes CL, Wang Y, Roberts H, Walley KR. Vasopressin decreases sepsis-induced pulmonary inflammation through the V2R. Resuscitation. 2008 Nov;79(2):325-31.
77 Mäntyjärvi R, Parkkinen S, Rytkönen M, Pentikäinen J, Pelkonen J, Rautiainen J, Zeiler T, Virtanen T. Complementary DNA cloning of the predominant allergen of bovine dander: a new member in the lipocalin family. J Allergy Clin Immunol. 1996 Jun;97(6):1297-1303.
78 Brodmann J, Twele R, Francke W, Yi-bo L, Xi-qiang S, Ayasse M. Orchid mimics honey bee alarm pheromone in order to attract hornets for pollination. Curr Biol. 2009 Aug 25;19(16):1368-1372. Epub 2009 Aug 6.
79 Shimizu N, Miwa K, Noge K, Yakumaru R, Mori N, Kuwahara Y. Stereochemistry of female-specific normonoterpenes, sex pheromone candidates from the acarid mite, Tyreophagus sp. (Astigmata: Acaridae). Biosci Biotechnol Biochem. 2009 Oct;73(10):2332-4. Epub 2009 Oct 7.
80 Di Stefano F, Verna N, Di Giampaolo L, Schiavone C, Di Gioacchino G, Balatsinou L, Burge PS, Boscolo P, Di Gioacchino M. Occupational asthma due to low molecular weight agents. Int J Immunopathol Pharmacol. 2004 May-Aug;17(2 Suppl):77-82.
81 Tótha M, Csonka E, Bakcsa F, Benedek P, Szarukán I, Gomboc S, Toshova T, Subchev M, Ujváry I. Species spectrum of flea beetles (Phyllotreta spp., Coleoptera, Chrysomelidae) attracted to allyl isothiocyanate-baited traps. Z Naturforsch C. 2007 Sep-Oct;62(9-10):772-778.
82 Tóth M, Csonka E, Bartelt RJ, Cossé AA, Zilkowski BW, Muto SE, Mori K. Pheromonal activity of compounds identified from male Phyllotreta cruciferae: field tests of racemic mixtures, pure enantiomers, and combinations with allyl isothiocyanate. J Chem Ecol. 2005 Nov;31(11):2705-20. Epub 2005 Oct 25.
83 Krückert K, Flachsbarth B, Schulz S, Hentschel U, Weldon PJ. Ethyl-branched aldehydes, ketones, and diketones from caimans (Caiman and Paleosuchus; Crocodylia, Reptilia). J Nat Prod. 2006 Jun;69(6):863-870.
84 Wood WF. Volatile components in metatarsal glands of sika deer, Cervus nippon. J Chem Ecol. 2003 Dec;29(12):2729-2733.
85 Villet RH. Involvement of amino and sulphydryl groups in olfactory transduction in silk moths. Nature. 1974 Apr 19;248(450):707-709.

EXOCRINOLOGY
281
86 Trapp S, Croteau R. Defensive resin biosynthesis in conifers. Annu Rev Plant Physiol Plant Mol Biol. 2001 Jun;52:689-724.
87 Bhatt JP, Sajwan MS. Ovarian steroid sulphate functions as priming pheromone in male Barilius bendelisis (Ham.). J Biosci. 2001 Jun;26(2):253-263.
88 Liberles SD. Trace amine-associated receptors are olfactory receptors in vertebrates. Ann N Y Acad Sci. 2009 Jul;1170:168-172.
89 Jarriault D, Barrozo RB, de Carvalho Pinto CJ, Greiner B, Dufour MC, Masante-Roca I, Gramsbergen JB, Anton S, Gadenne C. Age-dependent plasticity of sex pheromone response in the moth, Agrotis ipsilon: combined effects of octopamine and juvenile hormone. Horm Behav. 2009 Jun;56(1):185-91. Epub 2009 May 3.
90 Schildnecht H, Winkler H, Maschwitz U. On arthropod defense substances. XXXI. Comparative chemical studies of substances contained in the pygidial defense bullae of carabids. Z Naturforsch B. 1968 May;23(5):637-644.
91 Ai H, Kanzaki R. Modular organization of the silkmoth antennal lobe macroglomerular complex revealed by voltage-sensitive dye imaging. J Exp Biol. 2004 Feb;207(Pt 4):633-644.
92 Zhou W, Chen D. Encoding human sexual chemosensory cues in the orbitofrontal and fusiform cortices. J Neurosci. 2008 Dec 31;28(53):14416-14421.
93 Treyer V, Koch H, Briner HR, Jones NS, Buck A, Simmen DB. Male subjects who could not perceive the pheromone 5a-androst-16-en-3-one, produced similar orbitofrontal changes on PET compared with perceptible phenylethyl alcohol (rose). Rhinology. 2006 Dec;44(4):278-282.
94 Gelez H, Fabre-Nys C. Neural pathways involved in the endocrine response of anestrous ewes to the male or its odor. Neuroscience. 2006 Jul 7;140(3):791-800. Epub 2006 May 2.
95 Savic I, Berglund H, Lindström P. Brain response to putative pheromones in homosexual men. Proc Natl Acad Sci U S A. 2005 May 17;102(20):7356-61. Epub 2005 May 9.
96 Moncho-Bogani J, Martinez-Garcia F, Novejarque A, Lanuza E. Attraction to sexual pheromones and associated odorants in female mice involves activation of the reward system and basolateral amygdala. Eur J Neurosci. 2005 Apr;21(8):2186-2198.
97 Jurado MA, Junqué C. Criminal behavior after orbitofrontal lesion. Actas Esp Psiquiatr. 2000 Sep-Oct;28(5):337-341.
98 Aguilar EJ, García-Martí G, Martí-Bonmatí L, Lull JJ, Moratal D, Escartí MJ, Robles M, González JC, Guillamón MI, Sanjuán J. Left orbitofrontal and superior temporal gyrus structural changes associated to suicidal behavior in patients with schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry. 2008 Oct 1;32(7):1673-1676. Epub 2008 Jul 11.
99 Craig MC, Catani M, Deeley Q, Latham R, Daly E, Kanaan R, Picchioni M, McGuire PK, Fahy T, Murphy DG. Altered connections on the road to psychopathy. Mol Psychiatry. 2009 Oct;14(10):946-953, 907. Epub 2009 Jun 9.
100 Shamay-Tsoory SG, Harari H, Aharon-Peretz J, Levkovitz Y. The role of the orbitofrontal cortex in affective theory of mind deficits in criminal offenders with psychopathic tendencies. Cortex. 2009 May 18; [Epub ahead of print]

282
101 Dolan MC, Fullam RS. Psychopathy and functional magnetic resonance imaging blood oxygenation level-dependent responses to emotional faces in violent patients with schizophrenia. Biol Psychiatry. 2009 Sep 15;66(6):570-577. Epub 2009 May 15.
102 Mehta MA, Golembo NI, Nosarti C, Colvert E, Mota A, Williams SC, Rutter M, Sonuga-Barke EJ. Amygdala, hippocampal and corpus callosum size following severe early institutional deprivation: the English and Romanian Adoptees study pilot. J Child Psychol Psychiatry. 2009 Aug;50(8):943-951. Epub 2009 Apr 17.
103 Rotton J, Frey J. Air pollution, weather, and violent crimes: concomitant time-series analysis of archival data. J Pers Soc Psychol. 1985 Nov;49(5):1207-1220.
104 Rotton J, Frey J. Air pollution, weather, and violent crimes: concomitant time-series analysis of archival data. J Pers Soc Psychol. 1985 Nov;49(5):1207-1220.
105 Huyser C, Veltman DJ, de Haan E, Boer F. Paediatric obsessive-compulsive disorder, a neurodevelopmental disorder? Evidence from neuroimaging. Neurosci Biobehav Rev. 2009 Jun;33(6):818-30. Epub 2009 Jan 21.
106 Zhou W, Chen D. Encoding human sexual chemosensory cues in the orbitofrontal and fusiform cortices. J Neurosci. 2008 Dec 31;28(53):14416-14421.
107 Treyer V, Koch H, Briner HR, Jones NS, Buck A, Simmen DB. Male subjects who could not perceive the pheromone 5a-androst-16-en-3-one, produced similar orbitofrontal changes on PET compared with perceptible phenylethyl alcohol (rose). Rhinology. 2006 Dec;44(4):278-282.
108 Gelez H, Fabre-Nys C. Neural pathways involved in the endocrine response of anestrous ewes to the male or its odor. Neuroscience. 2006 Jul 7;140(3):791-800. Epub 2006 May 2.
109 Savic I, Berglund H, Lindström P. Brain response to putative pheromones in homosexual men. Proc Natl Acad Sci U S A. 2005 May 17;102(20):7356-61. Epub 2005 May 9.
110 Moncho-Bogani J, Martinez-Garcia F, Novejarque A, Lanuza E. Attraction to sexual pheromones and associated odorants in female mice involves activation of the reward system and basolateral amygdala. Eur J Neurosci. 2005 Apr;21(8):2186-2198.
111 Gulyás B, Kéri S, O'Sullivan BT, Decety J, Roland PE. The putative pheromone androstadienone activates cortical fields in the human brain related to social cognition. Neurochem Int. 2004 Jun;44(8):595-600.
112 Barnett R, Maruff P, Purcell R, Wainwright K, Kyrios M, Brewer W, Pantelis C. Impairment of olfactory identification in obsessive-compulsive disorder. Psychol Med. 1999 Sep;29(5):1227-1233.
113 Hermesh H, Zohar J, Weizman A, Voet H, Gross-Isseroff R. Orbitofrontal cortex dysfunction in obsessive-compulsive disorder? II. Olfactory quality discrimination in obsessive-compulsive disorder. Eur Neuropsychopharmacol. 1999 Sep;9(5):415-420.
114 Roelofs WL. Chemistry of sex attraction. Proc Natl Acad Sci U S A. 1995 Jan 3;92(1):44-49.
115 Feusner JD, Hembacher E, Phillips KA. The mouse who couldn't stop washing: pathologic grooming in animals and humans. CNS Spectr. 2009 Sep;14(9):503-513.

EXOCRINOLOGY
283
116 Sanberg PR, Shytle RD, Silver AA. Treatment of Tourette's syndrome with mecamylamine. Lancet. 1998 Aug 29;352(9129):705-706.
117 Silver AA, Shytle RD, Sheehan KH, Sheehan DV, Ramos A, Sanberg PR. Multicenter, double-blind, placebo-controlled study of mecamylamine monotherapy for Tourette's disorder. J Am Acad Child Adolesc Psychiatry. 2001 Sep;40(9):1103-1110.
118 Drickamer LC. Acceleration and delay of sexual maturation in female house mice (Mus domesticus) by urinary chemosignals: mixing urine sources in unequal proportions. Journal of Comparative Psychology 1988;102(3):215-21.
119 Ono S, Hu J, Shimizu N, Imai T, Nakagawa H. Increased interleukin-6 of skin and serum in amyotrophic lateral sclerosis. J Neurol Sci. 2001 Jun 15;187(1-2):27-34.
120 Ono S. Skin changes in amyotrophic lateral sclerosis. Brain Nerve. 2007 Oct;59(10):1099-1107.
121 Mustafa A, Ward A, Treasure J, Peakman M. T lymphocyte subpopulations in anorexia nervosa and refeeding. Clin Immunol Immunopathol. 1997 Mar;82(3):282-289.
122 Rollmann SM, Houck LD, Feldhoff RC. Proteinaceous pheromone affecting female receptivity in a terrestrial salamander. Science. 1999 Sep 17;285(5435):1907-1909.
123 Tamaoka A, Matsuno S, Ono S, Shimizu N, Shoji S. Increased amyloid beta protein in the skin of patients with amyotrophic lateral sclerosis. J Neurol. 2000 Aug;247(8):633-635.
124 Ono S, Hu J, Imai T, Shimizu N, Tsumura M, Nakagawa H. Increased expression of insulin-like growth factor I in skin in amyotrophic lateral sclerosis. J Neurol Neurosurg Psychiatry. 2000 Aug;69(2):199-2003.
125 Li W, Kennedy SG, Ruvkun G. daf-28 encodes a C. elegans insulin superfamily member that is regulated by environmental cues and acts in the DAF-2 signaling pathway. Genes Dev. 2003 Apr 1;17(7):844-58. Epub 2003 Mar 21.
126 Pradat PF, Bruneteau G, Gordon PH, Dupuis L, Bonnefont-Rousselot D, Simon D, Salachas F, Corcia P, Frochot V, Lacorte JM, Jardel C, Coussieu C, Forestier NL, Lacomblez L, Loeffler JP, Meininger V. Impaired glucose tolerance in patients with amyotrophic lateral sclerosis. Amyotroph Lateral Scler. 2009 Mar 20;. [Epub ahead of print].
127 Beck M, Giess R, Magnus T, Puls I, Reiners K, Toyka KV, Naumann M. Progressive sudomotor dysfunction in amyotrophic lateral sclerosis. J Neurol Neurosurg Psychiatry. 2002 Jul;73(1):68-70.
128 Agarwal AK, Zhou XJ, Hall RK, Nicholls K, Bankier A, Van Esch H, Fryns JP, Garg A. Focal segmental glomerulosclerosis in patients with mandibuloacral dysplasia owing to ZMPSTE24 deficiency. J Investig Med. 2006 May;54(4):208-213.
129 Agarwal AK, Fryns JP, Auchus RJ, Garg A. Zinc metalloproteinase, ZMPSTE24, is mutated in mandibuloacral dysplasia. Hum Mol Genet. 2003 Aug 15;12(16):1995-2001.
130 Barrowman J, Michaelis S. ZMPSTE24, an integral membrane zinc metalloprotease with a connection to progeroid disorders. Biol Chem. 2009 Aug;390(8):761-773.

284
131 Manavalan P, Smith AE, McPherson JM. Sequence and structural homology among membrane-associated domains of CFTR and certain transporter proteins. J Protein Chem. 1993 Jun;12(3):279-290.
132 Aufdenblatten M, Baumann M, Raio L, Dick B, Frey BM, Schneider H, Surbek D, Hocher B, Mohaupt MG. Prematurity is related to high placental cortisol in preeclampsia. Pediatr Res. 2009 Feb;65(2):198-202.
133 Wong JS, Gill SS. Gene expression changes induced in mouse liver by di(2-ethylhexyl) phthalate. Toxicol Appl Pharmacol. 2002 Dec 15;185(3):180-196.
134 Wyart C, Webster WW, Chen JH, Wilson SR, McClary A, Khan RM, Sobel N. Smelling a single component of male sweat alters levels of cortisol in women. J Neurosci. 2007 Feb 7;27(6):1261-1265.
135 Koelman CA, Coumans AB, Nijman HW, Doxiadis II, Dekker GA, Claas FH. Correlation between oral sex and a low incidence of preeclampsia: a role for soluble HLA in seminal fluid? J Reprod Immunol. 2000 Mar;46(2):155-166.
136 Dekker G, Robillard PY, Roberts C. The etiology of preeclampsia: the role of the father. J Reprod Immunol. 2011 Apr 27; [Epub ahead of print]
137 Błogowska A, Rzepka-Górska I, Krzyzanowska-Swiniarska B. Is neuropeptide Y responsible for constitutional delay of puberty in girls? A preliminary report. Gynecol Endocrinol. 2004 Jul;19(1):22-25.
138 McKenzie VJ, Bowers RM, Fierer N, Knight R, Lauber CL. Co-habiting amphibian species harbor unique skin bacterial communities in wild populations. ISME J. 2011 Sep 29. doi: 10.1038/ismej.2011.129. [Epub ahead of print]
139 Marsh R, Maia TV, Peterson BS. Functional disturbances within frontostriatal circuits across multiple childhood psychopathologies. Am J Psychiatry. 2009 Jun;166(6):664-674. Epub 2009 May 15.
140 Abel EL, Bilitzke PJ, Cotton DB. Alarm substance induces convulsions in imipramine-treated rats. Pharmacol Biochem Behav. 1992 Mar;41(3):599-601.
141 Ortega-Hernandez OD, Kivity S, Shoenfeld Y. Olfaction, psychiatric disorders and autoimmunity: is there a common genetic association? Autoimmunity. 2009 Jan;42(1):80-88.
142 Brewer WJ, Wood SJ, Pantelis C, Berger GE, Copolov DL, McGorry PD. Olfactory sensitivity through the course of psychosis: Relationships to olfactory identification, symptomatology and the schizophrenia odor. Psychiatry Res. 2007 Jan 15;149(1-3):97-104. Epub 2006 Dec 5.

Chapter 11: Atherosclerosis When the author was fiddling around in his laboratory (investigating the effects of low cardiac voltage electric pulses on human blood), atherosclerotic plaques formed. Wow! Such plaques had to be electroplated onto vessel walls! The idea explained too much not to be true. It accounted for the plaque build-up in areas of blood turbulence, spared small diameter blood vessels, spared veinous vessels, even atherosclerotic deposits appearing on synthetic vascular prostheses explanted from humans. The atherosclerogenic effects of diabetic blood sugar levels, the nicotine of cigarette smoking, and body habitus all gained a unified explanation in the equation describing electrodeposition. Reviewing the literature, the phenomenon had already been observed and proposed as a model, even though the mechanism by which the plaques formed went unrecognized, amazingly.
Atherosclerotic plaques electrodeposite when blood is used as electrolyte between cathode and anode, and accounts for the resemblance of the electrostimulation model of atherosclerosis1,2,3 to clinical findings. The kicker was that electroplating explained both the mixed tightly spaced crystalline deposits typical of atherosclerotic plaques (which could never be explained by any other mechanism physical or biological) and the blister formations seen in brittle plaques (another previously inexplicable phenomenon), plus it explained the influence of oxygen radicals on thromboembolic risk.
This is a book about human pheromones, right? Atherosclerosis as electrodeposition initiated by human pheromone accounts for all the beneficial effects of dietary fishoil, trans fatty acids and saturated fats, as well as their synergy and species specificity. Moreover, plant allomones like aspirin, cinnamon, quinidine, and dozens of others have similar beneficial cardiac effects that must be based, at least in
EXOCRINOLOGY
285

part, upon their natural anti-herbivory semiochemical function. (Indeed, common antibiotics are naturally allomones, it is often how they were discovered in the first place.) Other hypotheses4 on the etiology of atherosclerosis were primitive by comparison, unable to account for every single risk factor for the disease as this one does.
Artificial atherosclerotic plaques can be formed by low voltage electrodeposition both in animal models and as nodal accumulations from blood in pulsitile, low voltage electric fields: that much we knew. The oddities of atherosclerosis are explained mathematically as elements of electrodeposition, i.e. variables in the electroplater’s equation with just a few perfectly legitimate mathematical substitutions. Atherosclerotic plaques are essentially crystaline with many different crystals (with vastly different temperatures of crystalization) in juxtaposition that form at constant human body temperature: only electrodeposition can account for these phenomena. The phenomenon of electrodeposition mathematically explains the perverse effect of body habitus: why short thick people are so much more susceptible to atherosclerosis than tall skinny ones. The new explanation also takes care of why only conductive innervated arteries are affected leaving non-conductive, non-innervated veins and cut and resected arteries completely free of any plaques whatsoever. Electrodeposition provides the reason for allograft immunity as being due to the broken electrical ciruit. Electrodeposition explains why electrogenic organ proximity is associated with atherosclerosis (heart, gut, brain, & kidneys). Electroplater experience also shows why heavier deposits of plaque are observed under blood turbulence. That same turbulence, produced by shaking articles being plated in the electrolyte bath, is used in the electroplating process to speed deposition! Blood turbulence effects on atherosclerosis had baffled feable minds for generations. Electrodeposits are easily effected on plastics and atherosclerotic plaques have been recorded on artificial plastic vessel prostheses.
Explaining how plaques form on the interiors of blood vessels is a triumph of modern medicine, but the why remained open for a nanosecond. Electrodeposition stimulated centrally by a human pheromone trigger explains human behavior associated with atherosclerosis, and why central lesions on pheromone reception pathways can stimulate systemic atherosclerosis in animals. High status increases the
286

size of some glands, while stress increases alarm pheromone emissions. (The why behind goosepimples and throwing ones hands in the air in panic.) Species-specific beneficial effects of fish oils, health effects of emotion and the anti-sudden infant syndrome (SIDS) effect of fans in nurseries were also elucidated in the same context. That was a summary, so pay close attention to the science part, following.
LOW VOLTAGE ELECTRODEPOSITION OF ATHERSCLEROTIC
PLAQUE
The first four phases of atherosclerotic plaque accumulation are mainly by lipid accumulation.5 When the author passed a small current controlled by a metronome intermittently through whole human blood, electrodeposited atherosclerotic-like plaques formed on the aluminum anode. The experimentally electrodeposited plaque was identical in adherence, thickness, color, consistency, and texture to atherosclerotic plaques observable in human anatomical dissection. The characteristic creepy crackling sound on manipulation was also identical. Early plaque adherence to large arteries also forms patterns closely resembling unfinished electrodeposition of metals on substrates.
Review of the literature showed that low voltage, intermittent pulsatile DC electrodeposition was applied in an animal model of atherosclerosis. Betz and
associates1-3 discovered atheroma and plaque formation between two gold electrodes electrically stimulating rabbit carotid arteries in vivo. Their model was an accurate representation of all stages of plaque development. An atheroma appeared beneath the anode only with cholesterol diet and subendothelial fibromuscular proliferates without,2 and deposits even varied with electric field strength,3 but the notion of electrochemical deposition eluded them. Their finding of gold particles in the arterial wall2 demonstrated electro-deposition as the operative process, but they missed it entirely. The anodal accumulation should also have shed light on the mechanism, but
EXOCRINOLOGY
287

no. They left it a mystery. What is worse, all the learned colleagues reading the article over the years failed to think of electrodeposition, too. Just where did they believe that the gold in their deposits of plaque had come? It had to have come from their own gold electrode.
The literature also revealed a small indirect relationship between hematocrit and atherosclerosis6 or none at all.7 The equation for electrodeposition in ohmic materials8 with hematocrit in denominator explains this phenomenon and others.
weight of electro deposit = (I * t * A)/(z * F)
or by substitution
where I = V/R, from basic physics
and R = Δ * L/a, again from basic physics
and Δ , the resistivity of blood, which depends on the hematocrit as
Δ = 53.2 * e0.022H Function per Geddes and Sadler, 1973.9
Thus: " #Atherosclerotic plaque mass = f $ ___VtAa___________ % $ % $ 53.2 e0.022H LzF % & ' V = voltage in volts I = current in amps
288

t = time in seconds A = atomic weight of species deposited z = valence of species deposited F = Faraday's constant of 96,500 coulombs (amp-sec.) H = blood hematocrit L = length of conductor (basically aorta length, explaining body habitus effects) a = cross sectional area of conductor > 6 mm (complex function), (again, primarily the aorta here.) Aortas are wider in wide people and thinner in skinny people.
Is that cool or what!
Another powerful prediction made by the model is that factors affecting the cross-sectional area and length of the aorta would affect atherosclerosis, predicting that obesity and short stature would carry additional risk. Obese people of short stature do carry increased risk of atherosclerosis and heart disease!10, 11
Concentration of the electrolyte, agitation, temperature, addition agents and the nature of the electrolyte also contribute to electrodeposition. Addition agents employed in metallurgical electrodeposition include sugars, camphor, coal-tar derivatives, colloidal gelatin, agar, glue, rubber or casein. Organic agents such as sugar, coal tars, and casein improve adhesion but do not increase deposits per se. 8 Agitation also speeds electrodeposition8 but oddly, like concentration & temperature, does not appear in theoretical calculations. Both are species-specific near-constants anyway.
Not all atherosclerotic plaque accumulation is electrodeposition. Cholesterol dissolves preferentially in lipid bilayers, for instance, and would accumulate where lipid tails accumulate. But even cholesterol forms liquid crystals in the plaques!11, 12 Nothing but electrodeposition can explain that.
Why small diameter arteries and capillaries remain clear of atherosclerotic plaques has been a baffling problem,13 after all, if atherosclerosis resulted from some blood born factor, presumably that factor was also present in arterioles, capillaries, and veins, right? Electrodeposition in atherosclerosis explains why small diameter arteries remain plaque-free. Diameters of 6 mm and smaller are avoided by
EXOCRINOLOGY
289

electroplaters as impractical. 8 No other model even comes close to explaining this characteristic of atherosclerosis. Now it is no longer a mystery.
Another unusual characteristic of atherosclerotic plaques is that they tend to form in areas of high blood turbulence. Areas of high turbulence within the arterial system are unusually prone to plaque buildup.14 Again, this phenomenon is explained by principles of electrodeposition. Although omitted from their basic equation, it is a truism among electroplaters that the greater the agitation, the greater the electrodeposition!8 Platers use special agitation jigs routinely.
Mantovani et al14 found lipid deposits in synthetic vascular prostheses explanted from humans.15 Electrodeposits onto porous plastic tubing explains their finding. Lipid uptake in synthetic vascular prostheses depended on the duration of implantation of the prosthesis and on the sex of the patient.15 Lipid uptake also depended upon the internal diameter of the prosthesis.15 Electrodeposition is a function of time, and diameter of the tube8 since resistance varies with diameter and current inversely with resistance. Along with hematocrit,15 arterial diameters and lengths depend upon age and sex. Without controlling for these anatomical differences, men accumulate plaque faster than women do.16
The appearance of crystals in atherosclerotic plaques as amorphous and crystalline phases has puzzled observers.17 Electrodeposition explains amorphous and crystalline layers of variably sized crystals that are closely matched!18 Cholesterol and calcium are closely associated in crystal agglomerates within atherosclerotic lesions.19 Again, mixed crystal deposition is typical of alloy electroplating of alloyed metals,19 too! Organic and inorganic constituents of atherosclerotic lesions accumulating together, as observed by Lee et al.,18 are consistent with electrodeposition. Fine crystal grains form near the substrate with much coarser grains at greater distances from the substrate in electrodeposition,19 following the same pattern as observed in early atherosclerotic plaques! Crystallinity varies as a physical property 20 and virtually all chemical species capable of crystallizing from blood are represented in atherosclerotic plaques,18 even cholesterol as a liquid crystal.21 Over-voltage deposition results in larger unstable crystals being electrodeposited.19 Depending upon voltage, electrodeposits can form different
290

structures: amorphous layers (blisters), pyramids, spirals, whiskers and dendrites. In electrodeposition, increasing ion concentration, higher temperature, and increasing turbulence result in increased crystal size, while organic agents (such as sugar and coal tars), increasing current density, and polarization result in finer grain deposits (smaller crystal size).19
Autografted arteries occlude less frequently after bypass surgery.14 Autografts are not as electrically active as the original coronary arteries they replace, due to surgical denervation and scar tissue’s electrical discontinuity. Mammary arteries similarly situated to occlusive arteries, seldom harbor atherosclerotic lesions,14 but the mammary arteries are not within the heart’s electro-stimulation circuit. Scar tissue is much less conductive than healthy coronary pathways as observable on EKG. Do you see? Another conundrum falls.
Electrodeposition explains the accumulation of plaques within arteries supplying electrogenic organs. The heart and mesentery are highly electrogenic. Like the heart, the brain and kidney both possess their own electrical pacemakers.22
As when an artery is autografted, in internally irradiated arteries restenoses form less frequently.23 Radiation scars. Radiation scarring compromises the electrical conductivity of the artery or innervation.
Atherosclerotic plaques are also discontinuous and focal.14 Discontinuous deposits may suggest electrodeposition due to focal anodal charge. Electrodepositions persist after removal of an electrical stimulus, so atherosclerotic electrodeposition may be episodic. Normal lab values and the current of the anodal QRS complex of the AV node can be inserted into the above equation and would be sufficient to deposit atherosclerotic plaque mass. Try it!
An inward rectifier current is present in arterial smooth muscle cells24 while veins lack it. There is evidence that steady direct currents are generated in living bone by metabolic activity.25
Yet another unusual characteristic of atherosclerosis is the unusual propensity of diabetics to develop hardening of the arteries.26 Sugar is among the organic agents
EXOCRINOLOGY
291

used by electroplaters found empirically to improve electro-deposit consistency and adherence to substrates but does not increase mass of deposition per se.8
Tar and nicotine may enhance atherosclerosis in tobacco users. Coal tars, like sugar, are used to enhance adhesion in the electroplating process.8 Cigarette tar may behave similarly in the blood.
A class of nicotinic sites consists of chemoreceptors on sensory nerve endings -- especially certain chemo-sensitive nerve endings in the coronary arteries, the carotid and aortic bodies.27 These are among the commonest sites of atherosclerotic plaque electrodeposition! Pharmacological activation of these sites by nicotine from inhaled tobacco smoke would depolarize them, slightly enhancing anodal electrodeposition of anionic lipids from the blood. This explains why tobacco smokers are more susceptible to atherosclerosis along with other factors such as wall permiability to electronegative species.28
Reactive oxygen species have been implicated in the pathogenesis of atherosclerosis and hypertension, in part by promoting vascular smooth muscle cell growth.29 Oxidation as utilized in fuel cells, generates DC currents. Bioelectric sources are mathematically capable of electrodepositing the heaviest atherosclerotic plaques. Small voltages between electrodes around the carotid artery produce vascular smooth muscle growth1 with electrodeposition functioning.
THE PHEROMONE ETIOLOGY OF ATHEROSCLEROSIS
Free fatty acids are carried in the blood by albumin and other carrier proteins.30 Carrier proteins have been implicated as pheromone receptor proteins in pheromone transduction.31 CD36 is a scavenger receptor important for lipoprotein binding and uptake of cholesterol and lipids in vertebrates. In humans, loss of CD36 is linked to a wide range of disorders including insulin resistance, dyslipidemia, and
atherosclerosis.32 CD36 should be a pheromone receptor protein.33,34 Albumin changes conformation35 on binding to pheromone,36,37 releasing ionic species38 that
292

alter the potential difference of nearby dendritic membranes to transduce chemoreception.31
Heat shock proteins are a group of twenty-five highly conserved proteins up or down regulated in response to stress that show extensive homology at the DNA and protein level among bacterial and mammalian species. Stress releases alarm pheromone in many species, including man. The heat shock proteins play an active role in the development of autoimmune diseases in animals, and have been implicated in human autoimmune diseases (i.e. rheumatoid arthritis, multiple sclerosis). An induced immune response against heat shock protein 65 (HSP-65) results in atherosclerotic lesions in normocholesterolemic rabbits39,40,41 while HSP 60 antibody is associated with atherosclerosis.42 Acquired thermotolerance of cells is unrelated to the rapid synthesis of the characteristic spectrum of heat shock proteins.43 HSP 90, HSP 70 HSP 65, and HSP 28 all stain one way in normal-appearing regions of aorta, but develop a heterogeneous pattern in the presence of atherosclerosis.44 Given that heat shock proteins are emitted under circumstances of alarm, they may serve as extracellular pheromone receptor proteins. Changes of conformation upon pheromone recognition could account for Johnson et al.’s41 inhomogeneity (but so might simple precipitation). Heat shock proteins are necessary for pheromone reception in yeast and other species45,46,47 to account for its electrical effect upon pheromone exposure33-35 albumin must release charged particles like pheromone binding proteins.31 Release of free fatty acids on pheromone reception would allow their accumulation out of solution as fatty streaks by electrolytic attraction to artery walls.
Pheromones probably are responsible for most epigenetic effects observed in atherosclerosis. Pheromones immediately release behavioral and physiological changes, but they also alter behavior and physiology throughout ontogeny and even to follow-on generations. Permanet changes in behavior and physiology suggest epigenetic changes on DNA imprinted by pheromones. Phylogenic earliness is evidenced by the pheromone release of epigenetic changes (fruiting bodies) in colonial fungi.48 Pheromones do change the quartenary structure of DNA (e.g. the pheromone stimulated localization of genes to the nuclear periphery), leading to an abacus-like cellular memory effect.49 Pathological epigenetic changes induced by
EXOCRINOLOGY
293

pheromones should be discoverable and should be reversible with epigentic and pheromone therapy.
Dissatisfaction with one’s social circumstances is associated with susceptibility to atherosclerosis and sudden cardiac death.50 Lin et al.51 associated population density with atherosclerosis experimentally in female mice. Increasing population density leads to increased puberty delay pheromone emissions52 which in this population53 is the likely cause of increased atherosclerosis. Given that pheromones affect the immune system,54 and murine pheromone chemistry is already fathomed,55 testing the pheromone hypothesis will be straightforward.
The sebaceous kissing pheromone of human beings56 has a large triglyceride component with uniquely diverse stereochemistry57 typical of pheromones of most other species. A diversity of chain lengths can be found in the walls of atherosclerotic plaque covered arteries,58 the unusual variety of chain lengths, simultaneous appearance of odd and even chain lengths53 indicate that these lipids may be similar to unique sebaceous lipids and or even sebaceous in origin (taken into the body by kissing).51 Although Claire et al.53 failed to specify unsaturation position, it is plausibly the same unique diversity of fatty acids, esters and triglycerides associated with skin surface sebaceous film.52 The utility of these inhomogeneous components in atherosclerotic lesioned aorta is suggested by the effects of similar lipids. Unusual lipids in large, dietary quantities especially the omega-3 long chain fatty acids have a protective effect against atherosclerosis59,60,61 among many other physiological and behavioral effects.62 All such effects have been observed or would be expected of human pheromones. For instance, Dietary supplementation with either eicosapentaenoic63 or docosahexaenoic64 acids decreases natural killer cell activity. The same phenomenon is seen for murine stress pheromones in much smaller doses.65 Similarly, a diet high in fruits and raw vegetables alters gene function to improve heart disease outcomes.66 Ecological perception determining health & disease includes perception of diet as well as social perception via pheromones.
Men’s regularly kissing a female spouse is anecdotally associated with an increased life span by five years.67 Presumably frequency of kissing is inversely proportional to social dissatisfaction. Atherosclerosis being the most frequent cause
294

of death, the possible longevity benefit suggests that the kissing transfer of human sebum, containing a diverse mixture of human typical free fatty acids, triglycerides, esters and cholesterol esters, may produce some part of this benefit.
The lipids exchanged in kisses are hundreds of oddly substituted and straight chain, saturated and mono-enoic, cis free fatty acids and triglycerides and their esters, alcohols and metabolites. Unlike dietary lipids, these unique skin surface lipids vary widely in both odd and even chain length from 7 carbons to 28 in a chain.52 While kissing does not provide dietary quantities of these unusual lipids, dietary quantities are necessary for pheromonal effect only from near-pheromonal species68 such as the aforementioned n-3 (omega 3) long chain fish oils.
Omega-3 fatty acids may provide relief from PTSD,69 benefit infant growth and development, reduce rheumatoid arthritis pain and reduce overall and cardiovascular mortality, myocardial infarction, and sudden cardiac death.70 Fish oil seems to be only a partial substitute for kisses. The similar human kissing pheromone mixture, synergistic and species-specific, must be even more beneficial against PTSD, heart disease, and infant mortality.
Synergy has been observed in these and other beneficial health effects of unusual dietary cis-monoenoic free fatty acids.71 Synergy of action is another characteristic typical of pheromones,72 and all are cis shaped in humans.
Pheromones are the most potent of all medicines often effective at the ultra-trace level.73 500 fold smaller quantities of pheromonal sebaceous skin lipids have been found to elicit comparable activity when unsaturation position is displaced one carbon on the chain.62
The beneficial health effects of some dietary fatty acids are species specific.63 Pheromones are species specific within ecological niches.65 Rats74 and mice75 have pheromonal lipids in their furs somewhat similar to human sebum, and may help to account for their unfortunate habitat choice in proximity to us.
Various trans-fatty acids have beneficial effects for particular species in large dietary quantities.76 These may resemble species-specific pheromonal lipids
EXOCRINOLOGY
295

transferred in maternal and sexual contacts & via allogrooming, or they may have anti-pheromonal effects.
Some dietary trans-mono- and polyenoic lipids increase coronary artery disease while others, do not.77 Constitutional isomers of human-specific pheromonal sebaleic acid were found in both groups. Sebaleic acid, the free fatty acid C18:2) 5,8 and found only on human skin surfaces is exclusively human.52 Sebaleic acid may be potently beneficial in the treatment of autoimmune illnesses, if presented with synergistic human sebaceous lipids in appropriate quantities. Anecdotal evidence in my laboratory suggests that this same healthy adult male facial skin surface lipid taken p.o. can be a broad-spectrum sociopatholytic with 150 mg in a single dose curing criminal behavior, sexual perversions, and non-alcoholic drug addiction.
Because hostile social environments and low social support are associated with increased carotid atherosclerosis78 anger or aggression pheromones79 become prime suspects. There has been no human anger pheromone yet identified, but masculine testosterone diminishes upon exposure to female tears.80 Polyene lipids, even conjugated trienoics, are found in the hair fat of men81 and would be candidates for "hot-headed" emission. Indeed trienoic free fatty acids, perhaps the C18:3) 6,8,9 triene of human male hair lipid,75 are seen in atherosclerotic aorta walls as both free fatty acid and triglyceride while absent from healthy aortic tissues.53 Its presence as an irritant to vascular innervation might contribute to atherosclerosis electrodeposition.
Because anger causes erection of nasal tumescence tissues to block the chemosensory superior and middle meati,82 anger is prophylactic against autoreception and alloreception of air-born human pheromones. So which patient survives best on the wards, the weepy one or the angry one? Angry, aggressive patients survive because they avoid pheromonal insults that lacrimating patients receive. Such insults include air-born pheromones that probably cause sudden infant death syndrome (SIDS), since electric fans have been associated with improved prognosis83 and electric fans dissipate the pheromone plumes holding clinically relevant concentrations of pheromones. Calm wind conditions are required for pheromone communication in many species. Windy days see reduced crime, while
296

calm winds encourage criminal behavior in humans,84,85 another likely insult escaped by angry patients.
So why do some people get plaque accumulation when others with the same risk factors do not? A strike of substrate or special cleaning procedures may be needed to improve cohesion and initiate deposition on metals, glass, and plastic.8,86,87,88 Presumably, something akin takes place intravascularly. Pheromone receptions are also individually unique and account for accumulated epigenetic differentiation between identical twins.
CLINICAL IMPLICATIONS
A formula derived empirically from electroplating experience explains the biological mechanism of atherosclerotic electrodeposition. The formula shows how currents of injury, advanced age, and large body size with short stature and other factors may contribute to risk of atherosclerosis. Avoiding chronic anemia (raising hematocrit) or anything increasing the resistivity of blood should decrease atherosclerotic electrodeposition.
Given that the first four phases of atherosclerosis progress mainly by lipid accumulation,5 these might be reversed by vascular insertion of pulsatile low voltage DC electrodes for plaque electroacquisition and removal via the femoral artery. Ultrasound might smooth such electronic stripping of plaques as it does in electrodeposit removal.75
One beneficial and two detrimental pheromone sets are suggested in the pathogenesis of atherosclerosis deriving from kissing, stress, and anger, respectively. Lacrimation increases with stress and may indicate pheromone receptivity32 instead of a specific pheromone set. Human pheromones have not yet been tested in the pathology or treatment of atherosclerosis. If pathological autoimmune and cancerous epigenetic changes are indeed induced by pathogenic human pheromones, then
EXOCRINOLOGY
297

human pheromone remedies from among the human skin surface lipids or other semiochemical (naturally derived non-steroidal anti-inflamatory drugs, statins, taxol, and quinine are all allomones) interventions may prove beneficial or perhaps even curative.
Epigenetic pheromone therapy may be promising. For instance, low dose decitabine combined with healthy adult male facial skin surface secretion (paternal kiss) pheromone and perhaps non-steroidal anti-inflamatory drugs for their pheromone prophillaxis, may prove particularly useful in the treatment of atherosclerosis and heart disease, as well.
298
1 Apfel H, Friedmann B, Betz. Experimentelle arteriosklerose: Räumliche Proliferatausbildung als Folge ortsabhängig unterschiedlicher elektrischer Feldstärke? In: Heinle H, Schulte H, Schaeffer H, editors. Vieweg Verlag Braunschweig 1992;306-12.
2 Betz E, Schlote W. Responses of vessel walls to chronically applied electrical stimuli. Basic Res Cardiol Jan/Feb;74(1):10-20.
3 Apfel H, Friedmann B, Betz E, Meyer-Waarden K. [Distribution of the electric field in producing arteriosclerotic changes in the vascular wall by electrostimulation]. Biomed Tech (Berl) 1990;35 Suppl 3:310-311.
4 Tegos TJ, Kalodiki E, Sabetai MM, Nicolaides AN. The genesis of atherosclerosis and risk factors: a review. Angiology 2001 Feb;52(2):89-98.
5 Schoen FJ, Cotran RS. Blood Vessels. In: Cotran RS, Kumar V, Collins T, editors. Robbins pathologic basis of disease 6th ed. Philadelphia: WB Saunders Company; 1999. p.493-541.
6 Hartley CJ, Reddy AK, Madala S, Martin-McNulty B, Vergona R, Sullivan ME, Halks-Miller M, Taffet GE, Michael LH, Entman ML, Wang YX. Hemodynamic changes in apolipoprotein E-knockout mice. Am J Physiol Heart Circ Physiol 2000 Nov;279(5):H2326-34.
7 Jeng JS, Chung MY, Yip PK, Hwang BS, Chang YC. Extracranial carotid atherosclerosis and vascular risk factors in different types of ischemic stroke in Taiwan. Stroke 1994 Oct;25(10):1989-93.
8 Glasstone S. The Fundamentals of Electrochemistry and Electrodeposition. American Electroplaters' Society Palisade, New Jersey: Franklin Publishing Company, Inc; 1943.
9 Geddes LA, Sadler C. The specific resistance of blood at body temperature. Medical and Biological Engineering 1973 May: 336-9.
10 Kortelainen ML, Sarkioja T. Coronary atherosclerosis associated with body structure and obesity in 599 women aged between 15 and 50 years. Int J Obes Relat Metab Disord 1999 Aug;23(8):838-44.

EXOCRINOLOGY
299
11 Scolari F, Tardanico R, Zani R, Pola A, Viola BF, Movilli E, Maiorca R. Cholesterol crystal embolism: A recognizable cause of renal disease. Am J Kidney Dis 2000 Dec;36(6):1089-109.
12 Hata Y, Insull W Jr. Significance of cholesterol esters as liquid crystal in human atherosclerosis. Jpn Circ J 1973 Mar;37(3):269-75.
13 Libby P. Atherosclerosis. In: Fauci AS, Martin JB, Braunwald E, Kasper DL, Isselbacher KJ, Hauser SL, et al., editors. Harrison's principles of internal medicine. 14th ed. New York: McGraw-Hill Health Professions; 1998. p.1345-1352. 14 Mantovani D, Vermette P, Guidoin R, Laroche G. Lipid uptake in synthetic vascular prostheses explanted from humans. Biomaterials 1999 Jun;20(11):1023-32.
15 Hillman RS. Anemia. In: Fauci AS, Braunwald E, Isselbacher KJ, Wilson JD, Martin JB, Kasper DL, et al., editors. Harrison's principles of internal medicine. 14th ed. New York: McGraw-Hill Health Professions Division; 1998. p. 334-339.
16 Di Tullio MR, Sacco RL, Savoia MT, Sciacca RR, Homma S. Gender differences in the risk of ischemic stroke associated with aortic atheromas. Stroke 2000 Nov;31(11):26237.
17 Lee YS, Chen HC, Lai SM. High resolution electron microscopic investigation of atomic structural images in human atherosclerotic plaque. Proc Natl Sci Counc Repub China B 1992 Oct; 16(4):169-176.
18 Paunovic M, Schlesinger M. Fundamentals of Electrochemical deposition. Wiley-Interscience John Wiley & sons, Inc. New York; 1998.
19 Hirsch D, Azoury R, Sarig S, Kruth HS. Colocalization of cholesterol and hydroxyapatite in human atherosclerotic lesions. Calcif Tissue Int 1993 Feb;52(2):94-8.
20 Morrison RT, Boyd RN. Organic chemistry. 5th ed. Boston: Allyn and Bacon, Inc.;1987.
21 Juhasz J. [Embolism caused by cholesterol crystal] Orv Hetil 1961 Sep 10;102:1729-34.
22 With enzymatic processes yielding opposite polarity of proximal & distal convoluted tubule lumens (Brenner BM, Mackenzie HS, 1998 Harrison’s) and medullary salt bridges, kidney structure combines elements of both fuel cell and battery. The juxtaposition of proximal and distal convoluted tubules and the potential difference of their lumens demonstrates catagorically that the kidney electrically scrubs blood of charged particles for waste removal much as similar devices remove polluting charged particles from industrial pollution.
23 Waksman R, Robinson KA, Crocker IR, Gravanis MB, Cipolla GD, Seung KB, et al. Intracoronary radiation decreases the second phase of intimal hyperplasia in a repeat balloon angioplasty model of restenosis. Int J Radiat Oncol Biol Phys 1997 Sep 1;39(2):475-80.
24 Quayle JM, McCarron JG, Brayden JE, Nelson MT. Inward rectifier K+ currents in smooth muscle cells from rat resistance-sized cerebral arteries. Am J Physiol 1993 Nov;265(5 Pt 1):C1363-70.
25 Otter MW, McLeod KJ, Rubin CT. Effects of electromagnetic fields in experimental fracture repair. Clin Orthop 1998 Oct;(355 Suppl):S90-104.

300
26 Alexander CM, Landsman PB, Teutsch SM. Diabetes mellitus, impaired fasting glucose, atherosclerotic risk factors, and prevalence of coronary heart disease. Am J Cardiol 2000 Nov 1;86(9):897-902.
27 Pappano AJ. Wantanabe AM. Cholinoceptor-activating & cholinesterase-inhibiting drugs. In: Katzung BG, editor. Basic & clinical pharmacology. Norwalk, Connecticut: Appleton & Lange Division of Prentice Hall; 1992. p. 88-102.
28 Allen DR, Browse NL, Rutt DL, Butler L, Fletcher C. The effect of cigarette smoke, nicotine, and carbon monoxide on the permeability of the arterial wall. J Vasc Surg 1988 Jan;7(1):139-52.
29 Liao DF, Jin ZG, Baas AS, Daum G, Gygi SP, Aebersold R, Berk BC. Purification and identification of secreted oxidative stress-induced factors from vascular smooth muscle cells. J Biol Chem 2000 Jan 7;275(1):189-96.
30 Malloy MJ, Kane JP. Agents used in hyperlipidemia. In: Katzung BG, editor. Basic & clinical pharmacology. 6th ed. Norwalk, CT: Apple & Lange; 1995. p. 522-35.
31 Nicholson B. Pheromones cause disease: pheromone/odourant transduction. Med Hypotheses 2001 57(3):361-377.
32 Jin X, Ha TS, Smith DP. SNMP is a signaling component required for pheromone sensitivity in Drosophila. Proc Natl Acad Sci U S A. 2008 Aug 5;105(31):10996-11001. Epub 2008 Jul 24.
33 Rogers ME, Sun M, Lerner MR, Vogt RG. Snmp-1, a novel membrane protein of olfactory neurons of the silk moth Antheraea polyphemus with homology to the CD36 family of membrane proteins. J Biol Chem. 1997 Jun 6;272(23):14792-14799.
34 Martin C, Chevrot M, Poirier H, Passilly-Degrace P, Niot I, Besnard P. CD36 as a lipid sensor. Physiol Behav. 2011 Feb 24; [Epub ahead of print]
35 Barnes C, Lewis TJ. Shear-wave detection of structural effects in aqueous solutions of bovine serum albumin and hemoglobin. J Acoust Soc Am 1991 Sep;90(3):1287-1297.
36 Vogt RG. The molecular basis of pheromone reception: its influence on behaviour. In: Prestwich GD, Blomquist GJ, editors. Pheromone biochemistry. Orlando: Academic Press. 1987:385-431.
37 Van den Berg MJ, Ziegelberger G. On the function of the pheromone-binding protein in the olfactory hairs of Antheraea polyphemus. J Insect Physiol 1991;37:79-85.
38 Vogt RG, Prestwich GD, Lerner MR. Odourant-binding-protein subfamilies associate with distinct classes of olfactory receptor neurones in insects. J Neurobiol 1991;22(1):74-84.
39 Gruber R, Lederer S, Bechtel U, Lob S, Riethmuller G, Feucht HE. Increased antibody titers against mycobacterial heat-shock protein 65 in patients with vasculitis and arteriosclerosis. Int Arch Allergy Immunol 1996 May;110(1):95-98.
40 Afek A, George J, Gilburd B, Rauova L, Goldberg I, Kopolovic J, Harats D, Shoenfeld Y.Immunization of low-density lipoprotein receptor deficient (LDL-RD) mice with heat shock protein 65 (HSP-65) promotes early atherosclerosis. J Autoimmun 2000 Mar;14(2):115-121.

EXOCRINOLOGY
301
41 Mukherjee M, De Benedictis C, Jewitt D, Kakkar VV. Association of antibodies to heat-shock protein-65 with percutaneous transluminal coronary angioplasty and subsequent restenosis. Thromb Haemost 1996 Feb;75(2):258-260.
42 Zhu J, Quyyumi AA, Rott D, Csako G, Wu H, Halcox J, Epstein SE. Antibodies to human heat-shock protein 60 are associated with the presence and severity of coronary artery disease: evidence for an autoimmune component of atherogenesis. Circulation 2001 Feb 27;103(8):1071-1075.
43 Barnes CA, Johnston GC, Singer RA. Thermotolerance is independent of induction of the full spectrum of heat shock proteins and of cell cycle blockage in the yeast Saccharomyces cerevisiae.J Bacteriol 1990 Aug;172(8):4352-4358.
44 Johnson AD, Berberian PA, Tytell M, Bond MG.Atherosclerosis alters the localization of HSP70 in human and macaque aortas. Exp Mol Pathol 1993 Jun;58(3):155-68.
45 Louvion JF, Abbas-Terki T, Picard D. Hsp90 is required for pheromone signaling in yeast. Mol Biol Cell 1998 Nov;9(11):3071-3083.
46 Louvion JF, Warth R, Picard D. Two eukaryote-specific regions of Hsp82 are dispensable for its viability and signal transduction functions in yeast. Proc Natl Acad Sci U S A 1996 Nov 26;93(24):13937-42.
47 Savarit F, Sureau G, Cobb M, Ferveur JF. Genetic elimination of known pheromones reveals the fundamental chemical bases of mating and isolation in Drosophila. Proc Natl Acad Sci U S A 1999 Aug 3;96(16):9015-20.
48 Mayrhofer S, Jan M. Weber JM, Pöggeler S. Pheromones and Pheromone Receptors Are Required for Proper Sexual Development in the Homothallic Ascomycete Sordaria macrospora. Genetics, 2006 March; 172:1521-1533.
49 Brickner DG, Cajigas I, Fondufe-Mittendorf Y, Ahmed S, Lee P-C, Widom J, Brickner JH. H2A.Z-mediated localization of genes at the nuclear periphery confers epigenetic memory of previous transcriptional state. PLoS Biol, 2007 April;5(4):e81.
50 Krishnan KR. Depression as a contributing factor in cerebrovascular disease. Am Heart J 2000 Oct;140(4 Suppl):70-76.
51 Lin AH, Castle CK, Melchior GW, Marotti KR. The effect of population density on the development of experimental atherosclerosis in female mice. Atherosclerosis 1995 May;115(1):85-88.
52 Drickamer LC. Puberty-influencing chemosignals in house mice: ecological and evolutionary considerations. In: Duvall D, Müller-Schwarze D, Silverstein RM, editors. Chemical signals in vertebrates 4. New York: Plenum Press; 1986. p.441-456.
53 Drickamer LC, Mikesic DG. Urinary chemosignals, reproduction, and population for house mice (Mus domesticus) living in field enclosures. J Chem Ecol 1990;16(10):2955-2968.
54 Cocke R, Thiessen D. Alarm chemosignals suppress the immune system. In: MacDonald DW, MüllerSchwarze D, Natynczuky SE, editors. Chemical signals in vertebrates 5. Oxford: Oxford University Press, 1990:125-138.

302
55 Novotny M, Jemiolo B, Harvey S. Chemistry of rodent pheromones: molecular insights into chemical signaling in mammals. In: Macdonald DW, Müller-Schwarze D, Natynczuk SE, editors. Chemical signals in vertebrates 5. Oxford: Oxford University Press; 1990. p.1-23.
56 Nicholson B. Does kissing aid human bonding by semiochemical addiction? [Comment] Brit J Dermatol 1984 Nov;111:623-627.
57 Nicolaides N. Skin lipids: their biochemical uniqueness, unlike internal organs, the skin biosynthesizes and excretes unusual fat soluble substances. Science 1974 Oct 4;186:19-26.
58 Claire M, Jacotot B, Robert L. Characterization of lipids associated with macromolecules of the intercellular matrix of human aorta. Connect Tissue Res 1976;4(2):61-71.
59 Yamada T, Strong JP, Ishii T, Ueno T, Koyama M, Wagayama H, Shimizu A, Sakai T, Malcom GT, Guzman MA Atherosclerosis and omega-3 fatty acids in the populations of a fishing village and a farming village in Japan. Atherosclerosis 2000 Dec;153(2):469-481.
60 Angerer P, von Schacky C. n-3 polyunsaturated fatty acids and the cardiovascular system. Curr Opin Lipidol 2000 Feb;11(1):57-63.
61 Das UN. Beneficial effect(s) of n-3 fatty acids in cardiovascular diseases: but, why and how? Prostaglandins Leukot Essent Fatty Acids 2000 Dec;63(6):351-362.
62 Connor WE. Importance of n-3 fatty acids in health and disease. Am J Clin Nutr 2000 Jan;71(1 Suppl):171S-175S.
63 Thies F, Nebe-von-Caron G, Powell JR, Yaqoob P, Newsholme EA, Calder PC. Dietary supplementation with eicosapentaenoic acid, but not with other long-chain n-3 or n-6 polyunsaturated fatty acids, decreases natural killer cell activity in healthy subjects aged >55 y. Am J Clin Nutr 2001 Mar;73(3):539-48.
64 Kelley DS, Taylor PC, Nelson GJ, Schmidt PC, Ferretti A, Erickson KL, Yu R, Chandra RK, Mackey BE. Docosahexaenoic acid ingestion inhibits natural killer cell activity and production of inflammatory mediators in young healthy men. Lipids 1999 Apr;34(4):317-324.
65 Cocke R, Thiessen D, Nicholson B. Alarm chemosignals suppress the immune system. In: MacDonald DW, Müller-Schwarze D, Natynczuk SE, editors. Chemical signals in vertebrates 5. Oxford: Oxford University Press; 1990. p.125-131.
66 Ron Do, Changchun Xie, Xiaohe Zhang, Satu Mannisto, Kennet Harald, Shofiqul Islam, Swneke D. Bailey, Sumathy Rangarajan, Matthew J. McQueen, Rafael Diaz, Liu Lisheng, Xingyu Wang, Kaisa Silander, Leena Peltonen, Salim Yusuf, Veikko Salomaa, James C. Engert, and Sonia S. Anand The Effect of Chromosome 9p21 Variants on Cardiovascular Disease May Be Modified by Dietary Intake: Evidence from a Case/Control and a Prospective Study PLoS Med 9(10): e1001106. doi:10.1371/journal.pmed.1001106
67 Coleman V. The story of medicine. London: R. Hale;1985. P 1.
68 Silverstein RM. Chemistry of insect communication. In: Lewis T, editor. Insect communication. London: Academic Press; 1984. p. 105-21.

EXOCRINOLOGY
303
69 Matsuoka, Yutaka. Clearance of fear memory from the hippocampus through neurogenesis by omega-3 fatty acids: A novel preventive strategy for posttraumatic stress disorder?BioPsychoSocial Medicine 2011, 5:3
70 Oh R. Practical applications of fish oil (Omega-3 fatty acids) in primary care. J Am Board Fam Pract. 2005 Jan-Feb;18(1):28-36.
71 Pakala R, Radcliffe JD, Benedict CR. Serotonin-induced endothelial cell proliferation is blocked by omega-3 fatty acids. Prostaglandins Leukot Essent Fatty Acids 1999 Feb;60(2):115-123.
72 Silverstein RM, JO. Rodin, and D.L. Wood Sex attractants in frass produced by male Ips confusus in ponderosa pine. Science 1966;154:509-510.
73 Wilson EO. Pheromones. Scientific American 1963;208:100-106.
74 Nicolaides N, Ansari MNA. Fatty acids of unusual double-bond positions and chain lengths found in rat skin surface lipids. Lipids 1968;3(5):403-410.
75 Wilkinson DI, Karasek MA. Skin lipids of a normal and a mutant (asebic) mouse strain. J Invest Dermatol 1966;47(5):449-455.
76 Kritchevsky D. Antimutagenic and some other effects of conjugated linoleic acid. Br J Nutr. 2000 May;83(5):459-465.
77 Hodgson JM, Wahlqvist ML, Boxall JA, Balazs ND. Platelet trans fatty acids in relation to angiographically assessed coronary artery disease. Atherosclerosis. 1996 Feb;120(1-2):147-154.
78 Knox SS, Adelman A, Ellison RC, Arnett DK, Siegmund K, Weidner G, Province MA. Hostility, social support, and carotid artery atherosclerosis in the National Heart, Lung, and Blood Institute Family Heart Study. Am J Cardiol 2000 Nov 15;86(10):1086-1089.
79 Sandnabba K. Differences between aggressive and non-aggressive mice in odour signals and marking behaviour. In: MacDonald DW, Müller-Schwarze D, Natynczuk SE, editors. Chemical signals in vertebrates 5. Oxford: Oxford University Press; 1990. p.459-464.
80 Shani Gelstein, Yaara Yeshurun, Liron Rozenkrantz, Sagit Shushan, Idan Frumin, Yehudah Roth, and Noam Sobel Human Tears Contain a Chemosignal. Science 1198331Published online 6 January 2011 [DOI:10.1126/science.1198331]
81 Weitkamp AW, Smiljanic AM, Rothman S. The free fatty acids of human hair fat. J Am Chem Soc 1947;69:1936-1939.
82 Holmes TH, et al. The Nose; an experimental study of reactions within the nose in human subjects during varying life experiences with a forward by Warfield T. Lonscope. Springfield, Ill: Thomas; 1950.
83 Coleman-Phox K, Odouli R, and Li D. Use of a fan during sleep and the risk of sudden infant death syndrome. Arch Pediatr Adolesc Med. 2008;162(10):963-968.
84 Rotton J, Frey J. Air pollution, weather, and violent crimes: concomitant time-series analysis of archival data. J Pers Soc Psychol. 1985 Nov;49(5):1207-1220.

304
85 Indeed in open, anecdotal trials, my laboratory has effected cures of criminal behavior, addiction, and sexual perversion with a home remedy of healthy adult male facial skin surface lipid pheromone delivered p.o. in a single 150mg dose on chewing gum vehicle. We now have a double-blind, cross-over trial under way.
86 Val'syunene YI, Rachinskas VS. Electrodeposition of magnetic coatings on glass. In: Slutzkin D. editor. Electrodeposition of metals; proceedings of the 10th Lithuanian conference of electrochemists, Dec. 12-13 1968. Jerusalem: Israel Program for Scientific Translations; Springfield, Va.: U.S. Dept. of Commerce, Clearinghouse for Federal Scientific and Technical Information; 1970.
87 Kochergin SM, Vyaseleva GY. Electrodeposition of metals in ultrasonic fields. New York: Consultant's Bureau; 1966.
88 Saubestre EB, Typical processing and operating sequences b. plastics. In: Graham AK, editor. Electroplating Engineering Handbook. 3rd ed. New York: Van Nostrand Reinhold Company; 1971. p. 212-213.

Chapter 12: The Function of Myelin
The Schwann/oligodendritic cells/axon device propagates transients electrically, utilizing serial discharges of electric and magnetic fields. With current at the node of Ranvier, a triphasic signal results, inductive and capacitative discharges leading and lagging, respectively.
The peculiar structure of myelinated axons persuades us both the error of previous models and the correctness of this one. The Schwann cell wraps numerous times around the axon. Unnecessary for insulation function, wraps thinned to an extreme constrain cell maintenance unnecessarily. A simple fat cell would do a better insulating job than a Schwann cell.
Electrically conductive "dense lines" found in Schwanns serve no insulative purpose, but must be present to induce an opposite charge for this proposed capacitative function. Maintaining that conductivity comes at an economic cost, as well. The dense line conductivity exceeds that of normal somatic cytoplasm. Specialized rigid proteins for the purpose also obstruct cell maintenance.
As expected by economic theory, Schwann cell membranes are both the best and the thinnest dielectrics available to the body. Both characteristics enhance capacitive properties as all cell membranes except the outer one intervene between planes hypothesized to bear opposite charges.
Unwound, each Schwann cell shows an enormous surface area. Again this is unnecessary for insulation. However, capacitors improve directly with increased surface area. If Schwann cells function as capacitors, their enormous surface areas perform an important service.
EXOCRINOLOGY
305

All Schwann cells along a particular peripheral nervous system nerve unwind to equal surface areas irrespective of the body tissue traversed by that nerve. Yet, different tissues possess differing insulating properties. Accordingly, maintaining uniform insulating cells allocates resources sub-optimally.
Capacitors in series add capacitance as inverses yielding one capacitance smaller than any. Given that capacitance is expensive and logarithmically more expensive as surface area and capacitance increases, and given identical biological material constrains, circuits of capacitors in series are most economical when every capacitor is of equal capacitance.
From basic physics, equilibrium would oppose unequal contributions. At the same time, equal capacitances for sequentially discharging capacitors optimizes functionally as well as economically. Serial discharges of capacitances propagating signals like falling dominos would require equal capacitances, much like dominos require equal potential energies. Equal surface areas in similarly constructed capacitors yield equal capacitances.
Summation of capacitance in series.
306

Electrical diagram of capacitors in series.
Distributed energy storage in the magnetic fields of inductors formed by lapping Schwann wraps and enhanced by the axon core (like a solenoid) might be expected, too. The reactances, inductive and capacitative, would reach equilibrium at equal values. Current across a node of Ranvier would then reflect local capacitance and inductance in a biphasic pulse with inductance leading, capacitance following. Such bi-modality is seen and (hypothesized as erroneously) attributed to respective membrane permeabilities of Na+ and K+.
Capacitance of a parallel plate condenser, the area of whose plates is A and the
distance between them d, is given as
C = (KA)/(4(pi)d)
where K is the dielectric constant of the medium.
EXOCRINOLOGY
307

Capacitors in cross section. The one at lower left looks much like a Schwann cell or an oligodendrite wrapping an axon.
308

EXOCRINOLOGY
309
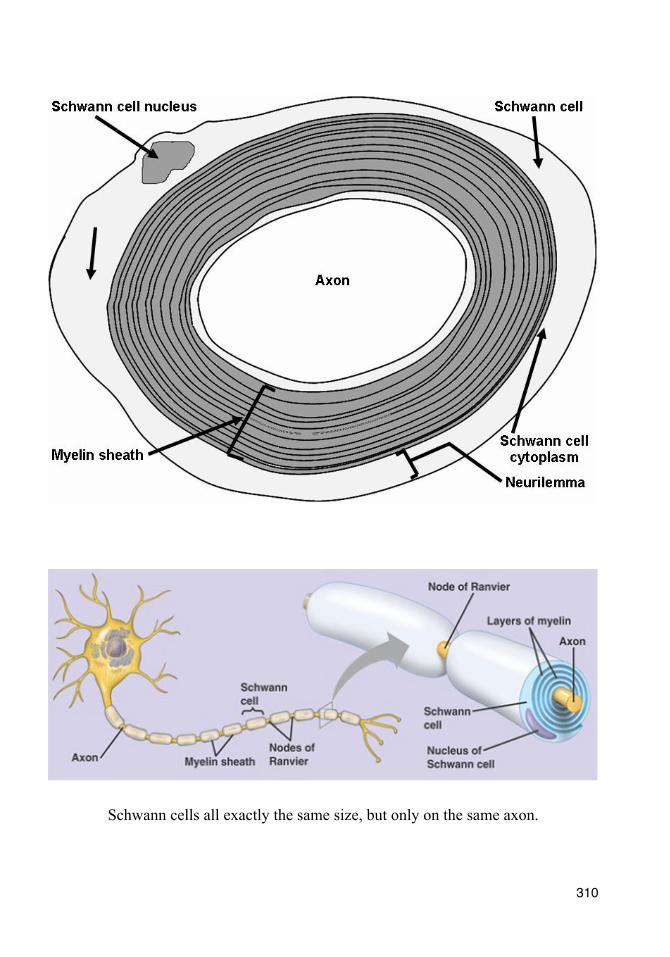
Schwann cells all exactly the same size, but only on the same axon.
310
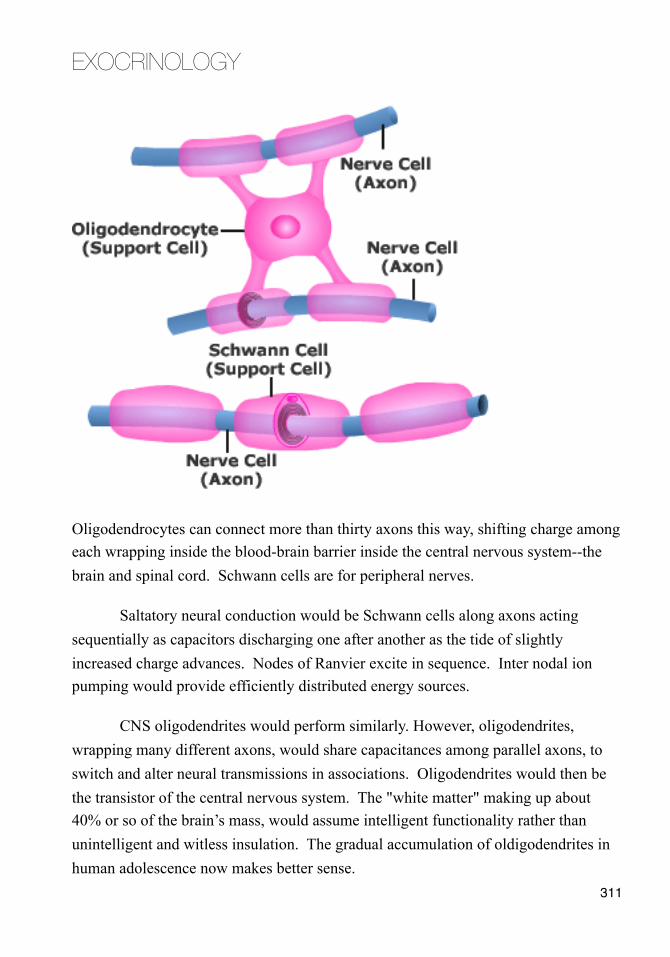
Oligodendrocytes can connect more than thirty axons this way, shifting charge among each wrapping inside the blood-brain barrier inside the central nervous system--the brain and spinal cord. Schwann cells are for peripheral nerves.
Saltatory neural conduction would be Schwann cells along axons acting sequentially as capacitors discharging one after another as the tide of slightly increased charge advances. Nodes of Ranvier excite in sequence. Inter nodal ion pumping would provide efficiently distributed energy sources.
CNS oligodendrites would perform similarly. However, oligodendrites, wrapping many different axons, would share capacitances among parallel axons, to switch and alter neural transmissions in associations. Oligodendrites would then be the transistor of the central nervous system. The "white matter" making up about 40% or so of the brain’s mass, would assume intelligent functionality rather than unintelligent and witless insulation. The gradual accumulation of oldigodendrites in human adolescence now makes better sense.
EXOCRINOLOGY
311

Synesthetes, people who associate colors with letters for instance, mix perceptions localized to adjacent areas of the brain. If oligodendrites of the white matter are shared capacitance switches, now we know how: parasitic capacitance.
THE TRIGGER ZONE
Summation describes the strange behavior of the trigger zone of the axon hillock. The axon hillock is that part of the axon extending from a nerve cell body to the first Schwann cell. This axon hillock has no special structures, and looks to all the world like a simple cell membrane. However, magical powers are attributed to it.
The axon hillock's so called "trigger zone" appears to control the phenomena of "summation," propagation, excitation, and inhibition, all while exhibiting the keenest electro-sensitivity of any cell area. That is quite a feat for a simple membrane. Summation describes the trigger zone's mysterious ability to count up additions and subtractions of electrical stimulation over a short period. When the energy inserted into the trigger zone is high enough over a short, nerve specific period the axon fires, or propagates an excitation (excitatory postsynaptic potential, or EPSP) down the axon.
The trigger zone's magic summation behavior could be duplicated simply by filling and emptying charged ions (capacitance) into the wound Schwann cell right at the edge of the trigger zone. It becomes a circuit describable by physics instead of bio-mumbo-jumbo. Depleting charge from the Schwann would inhibit firing. Several circuit devices could short and restore capacitors across subsequent nodes of Ranvier in series depending upon voltages applied. A voltage sensitive switch, on and off, must be found within the Schwann/node interface.
No structure in trigger zones is capable of EPSP and IPSP summation. A voltage sensitive switch shorted capacitance (and parallel resistance / inductance representing heat and conduction losses and delayed inductor contribution) in series could account for summation, accommodation, and habituation in an electrical model of neural function.
312

HEAT OF CAPACITORS GENERATED WITHOUT O2
Myelinated neurons put off a lot of heat. They are 50 X's hotter than unmyelinated C-fibers of the same size. Yet oxygen consumption is 6% for white matter vs. 94% for grey matter! Only a functioning capacitor would heat with a low O2 consumption.
In-cord coordinated movements (decerebrate cat on a treadmill) and the need for neural decussation follow from oligodendritic shared capacitance switching and limited capacitance transmission. Thus oligodendrites would "connect" crossing nerves and severing the cord below decussation would eliminate all but neural bilateral coordination.
Schwann and especially oligodendritic capacitors are variable. How do you vary that capacitance? Just shrink or swell the cell, the plates move closer or further apart, affecting capacitance. Thus the same old neurotransmitter/hormone receptor mediated channels on the cell membrane which normally affect cell turgidity, also alter neural function. Given shared capacitance switching, a neurohormone can switch firing programs, and alter behavior.
The mechanism for gross chemical neurological influence, hormones that affect the nervous system, would involve influencing membrane bound Na+ ports on the Schwann cell surface. H+ ATPase regulates turgidity in the cell by controlling ionic concentrations. Swelling or shrinking of cells is managed thereby. The same mechanism is proposed for neurohormones. They merely behave exactly like they have been shown to behave. They affect the salt balance inside the Schwann cell by binding and triggering membrane bound proteins which loosen or tighten the ion channel. With a higher interior salt concentration, water moves into the Schwann cell. Water moving into the Schwann cell causes "dense lines" and "thin lines" to move apart slightly by cellular swelling. Moving apart conductive surfaces in a capacitor (the ‘d’ in our capacitance formula) diminishes capacitance, changing neural function.
EXOCRINOLOGY
313

LINEAR ACCELERATORS IN ION TRANSPORT PROTEINS
This last model better explains the kinetics and characteristics of transmembrane ion transmission. Physical and economic efficiencies of cation and anion propulsion argue for use of a simple machine, the linear particle accelerator, in transmembrane ion displacement. Particle confinement devices identical to those of linear accelerators are found in trans-membranous alpha helixes surrounding rigidly juxtaposed Hückel’s rule magnets. Other oddities of kinetics, structure and function find explanations within the model as well.
Phenyl groups set up strong magnetic fields detectable by nuclear magnetic resonance techniques.1 By Lentz's law, current flows in a direction setting up magnetic fields to oppose movement. Thus, the circulation of electrons of two phenyl groups (or other similar magnetic field producing molecular structure) would flow to oppose a mutual approach, and to withhold from a mutual withdrawal. Synchronized approaches and withdrawals of serial phenyl pairs would open and close a linear accelerator. Such approaches and withdrawals characterize vibration. Given the length of the Alpha protein tube with phenyl groups in regular fixed opposition, the natural frequency for the effect can be calculated. Failure of these tiny linear accelerators, where Hückel’s rule amino acids fail to juxtapose to form the containment field, is behind cystic fibrosis. Illuminating these broken cystic fibrosis channel containing tissues with magnetic fields should have an observable and perhaps beneficial effect.
By lining up these natural linear accelerators which transport ions across membranes and joining the parts together in longer than natural rows, we can make room temperature superconductor plasma conduits to carry protons or perhaps microscopic ion propulsion devices. Biologists can now be rocket scientists!
In addition to Scotty’s ion power, a compact phasor design might be produced which requires neither vacuum nor cryo-magnetic devices. The tiny diameter of the
314

tube creates a perfect vacuum without pumps. Many such tubes may be laid down in a fascist bundle.
Those interested must size the tube in proton ATP-ase, since it appears to be the best candidate at first inspection. Calculate or determine resonant frequencies. Count the juxtaposed Hückel’s rule amino acid side chains (at nodes?). Determine if frequencies match observed conformational change frequencies. You may need to ‘peg’ the chain straight to some crystal to get started.
The old model of ion membrane transport--say in the nephron--calls for mechanical binding and releasing of cations across membranes with all sorts of baroque biochemical nonsense. The membranes of the proximal and distal tubules of the glomeruli require a charge gradient between proximal and distal tubules to have a potential difference sufficient to move those charged particles out of circulation. This means that JG cells must function electrically, in similar means. Yet another medical school physiology mystery solved.
Such movements of charged particles under the crazy-quilt of old models all require co-transport of waters of hydration, which, from kinetic studies, would be impossibly slow. The new model would transport only the cation (or anion) through a virtually frictionless mechanism.
If you were God, which would you choose? It is funny that the invention of the linear accelerator was hailed as the first invention divorced from natural precursors (e.g. the electric motor has a naturally flagellar precursor). Oops! Each cell in our body has hundreds of thousands of working linear accelerators, proton ATP-ase and others.
Rapid and regular conformational movements of proximal protein picking up and releasing, say, a couple of magnesium cations in each iteration could act as the pumping mechanism. This heretofore unexplained movement has been observed and recorded. The charge on two magnesium cations are close enough and large enough to have a significant repulsive charge effect on a proton's acceleration.
The design utilizes similar principles as a linear accelerator. The design consists of a proteinaceous alpha helix. Within the lipophilic interior of the helix,
EXOCRINOLOGY
315

tryptophan phenyl groups repeatedly oppose each other in pairs. Oppositions across the interior of the tube are phenyl end to phenyl center, thereby bringing opposite magnetic fields into close, but just wide enough juxtaposition. Magnetic fields on a number of molecular structures affects their NMR spectrometry, and is an effect utilized in their detection. A representation of the toroidal magnetic field of a phenyl group can be seen in the figure. These form the containment field guideway for charged particles.
Additionally, particle acceleration is currently accomplished by a rapid conformational change in another (or another part of the) protein nearby. A pair of large cations are picked up near the proximal end of the alpha helix's tube. The conformational change, ATP driven, moves the two cations distally beside the tube, accelerating a proton or other cation through the tube.
Note that waters of hydration are left behind and only the particle moves down the tube. Of course, the tube's girth being so small and fixed, contamination risks diminish so that no vacuum is required. The event is friction free. Particles move where they are aimed by the opposing opposite magnetic fields (North beside South).
316

The design is sensitive to amino acid substitution into the "gun barrel" of the helix. Any substitution that results in bending renders the accelerator nonfunctional. However, other than linear shapes might be feasible.
The Cl channels2 (Li, M., et al., 1988) may function similarly. The lack of tryptophan's phenyl group at the interior base of the alphahelix closes ion exchange in cystic fibrosis. In addition, an unexplained sensitivity to electric fields has been reported (Li, M., et al., 1988).
[L]arge depolarizations increase the likelihood that Cl channels will open for the first time (activate) in excised patches. Once a Cl channel was activated by depolarization, it remained activated, even at hyperpolarized voltages, .... The way in which depolarization modifies Cl channels is unknown but a voltage of +120 mV is about 160-180 mV away from the normal membrane voltage, creating a very strong electric field.
Multilayer stacked accelerators may have been described in chloroplasts, with the significance of the structures not elucidated.
We have the ability to synthesize proton ATPase, or a portion of the helix repeatedly. A tube such as that described above of any length can be manufactured, however some substrate would be required to maintain linearity to which side chains might attache. A crystal, perhaps?
The author is tempted to suggest a "dilithium crystal."
A MODEL OF MUSCLE FUNCTION
Current thinking has it that muscle contraction is stimulated electrically, fueled chemically, and carried out mechanically.3 The thing between actin and myosine appears more likely to be a levatation process, somewhat like a maglev train.
EXOCRINOLOGY
317

Thus only an amplification would be needed rather than two (2!) energy transductions, plus even the energy of the activating motor neuron is utilized. The friction question becomes manageable, since levitated surfaces never touch. This model's correctness is obvious enough so as to require no further discussion. I will leave it to experimentalists to work out the details.
318
1 Silverstein Robert M., Bassler, G. Clayton, and Morrill, Terrence C. Spectrometric Identification of Organic Compounds. 4th edition. New York: John Wiley & Sons.
2 Li M, McCann JD, Liedtke CM, Nairn AC, Greengard P, Welsh MJ. Cyclic AMP-dependent protein kinase opens chloride channels in normal but not cystic fibrosis airway epithelium. Nature. 1988 Jan 28;331(6154):358-360.
3 Stryer, Lubert 1988; Biochemistry, Third Edition. New York: W.H. Freeman and Company. 1089p.

Chapter 13: The Future This book, for better or worse, is the birth of the medical specialty of exocrinology. Perhaps the most obvious change for the future by exocrinology will be the absence of criminals. Soon, we will need no prisons, no police, no locks, no fences, no military, and we will have no practice of medicine as we know it. It will be like the abandonment of tuberculosis sanitariums closed by inoculations and antibiotics.
The cure for criminal behavior will see changes reminiscent of the changes observed with the finding of the cure for tuberculosis, too. Institutions to deal with criminals, with sexual perversion, with drug addiction, with unwed mothers, and many more social ills will close and society overall will become more efficient. The pace of the change is already gaining speed. The crime rate in the author’s new home town of Tampa keeps dropping and dropping, but pheromone therapy contributes but a small part of the decrease. Clearing ozone from local power plants probably helps the most. Every successful criminal pheromone therapy (all have been successful) takes off the street another criminal to improve our statistics. Soon, the need for hiring policemen, the need for training policemen, these will decline and people who would have endured that occupation will become available for other work. Similarly, cured criminals will be available to become model employees, eager to learn, eager to excel.
Sexual perverts will diminish in danger and number as the cure for their ailment reaches their worried parents. The “cure” for homosexuality is just too easy and universally available (on the end of every healthy father’s nose). Given the fatherly desire for grandchildren, the “malady” will not “fester” much longer in children. Exocrinologists will detect insufficient pheromone receptor proteins and
EXOCRINOLOGY
319

remedy them. On the plus side (for perverted people that is), receptivity for homosexual pheromones may be manipulable as well.
Population rebelliousness may subside, too. Jihadists will be treated with the pheromone, doubtless decried as forced sacraments. (Of course, providing pheromone on chewing gum to captured enemy soldiers will be Christ’s sacrament of communion. Christ must have been the first exocrinologist. That last supper was so very much a meal of pheromones! And that intense feeling eating what remained of Christ’s medicine chest three days after crucifixion, so yes they felt the presence of Jesus Christ three days after his death.) The pheromone exposure history of Moslem people, essentially smelling the feet in front of them, has enabled the jihad phenomenon, which would be juvenile delinquency and criminality in western cultures that lack forced daily foot-sniffing. The holy exposures to each other’s feet by having all face Mecca, the fast of Ramadan, these have saved Muslims from many Christian ills, so the great prophet Mohammed also was an exocrinologist. Human pheromones, discovered and used by Jesus, will drown feelings of jihad entirely. The air pollution that drives the jihad phenomenon, particularly high ozone concentrations, must also drop to effectively end this rebellion against science, learning, and wisdom.
320

Here is a NASA map1 of fine particulate matter in the air, roughly air pollution. The correlation with social turbulence and rebellion is suggestive.
Nosocomial and community acquired infections will respond to human pheromone therapy. Receiving one’s family pheromones will become standard compassionate care for our death beds. Methicillin Resistant Staphylococcus aureus, MRSA patients with weakened immune systems and others with weakened immunity, all should benefit from pheromone therapy immediately. Even DIC, disseminated intravascular coagulation, should respond to conventional antibiotic therapy and 150 mg of adult male facial skin surface lipid p.o. Why? Infection is allowed by the human semiochemical commerce as a pheromonal mechanism to shed population in the face of limited resources. Look at the siege of Mafeking. They kept great records, and those that suffered most were lowest in hierarchical status. Look at the death camps of the nazis, where low status, even the simple pheromone receptivity of lacrimation, tears, could be counted on to reduce chances for survival. Pheromones limit populations to available supplies of food and shelter, even behind barb wire. Human crime and autoimmune pathology provide the lacking predation. Generally more successful species (like humans and livestock) see increased parasitism. The mechanism of reduced immunity is just a tuning of the pheromone vs allomone recognition system that we call the immune system.
EXOCRINOLOGY
321

Animals caring for each other evolved in colonial bacteria, and before, because a gene is a gene is a gene. The free-rider problem of economics and the problem of altruism are both solved when one recognizes that improving the prospects for one’s own genes is accomplished when those same genes reside in others, too. Indeed, with the similarity of genomes, the wonder is that greater care is not taken to preserve each other and nature everywhere.
In our time, autoimmune disease and cancers will cease to exist, as pheromone therapy will be curative. Autoimmune disease and cancer probably evolved to augment natural population pruning immediately after the invention of medicine back at the dawn of time. Just as having antibiotics for soldiers gave the Allied armies advantages against the Axis, caring for the sick ever more effectively advanced societies. Exocrinology will bias the balance of life furthering that human advantage.
Our schools will become centers of brilliance and creativity when all students are free from their spiritual demons. Imagine a high school with only dedicated students, none ever dropping out, with all seeking excellence to the very best of their abilities. Indeed, we may see the need to establish special Orwellian reservations to maintain the sparks to creativity that perversion and mental illness both provide.
The author is no stranger to educational innovation. In the 1950s, educational achievement in Georgia, the author’s home state in America, averaged only to the fourth grade. Now two years of college training is the norm because this author proposed the HOPE Scholarship Program for free college education of deserving students. The program was subsequently emulated in 23 US states.
The parasitism that is pet ownership will lessen to more manageable levels. Allomones of dogs, cats, and horses that reduce our population fertility and diminish the semiochemical commerce will be seen as the dangers they are. With wide availability of healthy adult human male facial skin surface lipids, animal rights activists will recover their sanity again, perhaps enough to contribute themselves to scientific advancement. Similarly, those scientists and physicians who engage in
322

human, animal, and plant torture to assuage their bloody appetites will find peace, happiness, and an end to their unbearable suffering.
Oddly, match-makers can use artificial pheromone exchanges to increase chances of bonding success. The approximate ratio of acceptable to unacceptable skin surface lipids from the opposite sex seems to be about four out of ten. The ratio of highly desirable samples might be about one out of ten. That means that true love is not really that far away. Mutual delight and devotion at one chance in one hundred (the product of the probabilities of delightful matches for each spouse) suggests more work is needed, but it also suggests that such services may be profitable.
In addition to better life matches (and thereby perhaps more blissful marriages), divorce rates should sink to more manageable levels. This is because divorce is treatable with human pheromone therapy.
Not all will be peace and light. This book will eventually cause great tumult at least academically. Psychology will have to become something else since its central dogma will be all hollowed out and gone. The idea that bad thinking causes disease will give up the ghost, taking with it, much of the up to now reasonable argument for political correctness. Criminology will itself become academically criminal given the complete barn door misses of its savagely devoted adherents. Sociology and anthropology will move over and add chemists. College students will begin a much more arduous course of study, too, “Oh happy day!”. Having several Ph.D.’s in different disciplines will become commonplace, just like Star Trek.
That is not to say that observers of family life will all suddenly be looking for new jobs. We will still need surveillance of treated families, because relieving pressure in one place will probably cause problems to bubble up in another. Pheromone therapy will be reserved for serious cases, but the simmering difficulties that caused that serious case may find vent elsewhere. We must be vigilant and professional, and kind.
This book has taken controversial stands well beyond the work of some of the greatest scientists who ever lived: Vero Copner Wynn Edwards,2 Iränus Eibl-
EXOCRINOLOGY
323

Eibesfeldt,3 Edward O. Wilson.4 Hopefully my facile analyses will not embarrass them too badly.
Humanity is a herd being managed by nature, optimization algorithms with feedback loops provide a population homeostasis, we just do not know much about it. Not to worry, if humanity fails, nature has a fall back position, and if that fails, well, there are lots of bare rocks that are earth’s size floating about in space.
Given the awkwardness of humanity’s situation, manipulating our own pheromones overmuch will increase our numbers out of all proportion driving our populations to crash eventually, leaving that cinder behind. Malthus misjudged the rate of increasing agricultural productivity. Thus blanket applications of human paternal pheromones to high government officials, corporate CEOs or to all military personnel to assure ethical war-making or to end all suicidal ideation will backfire. We must reserve pheromone therapy for those of us with problems or illnesses, or else appropriate levels of risk-taking in the society will diminish. We can’t all be president.
Without governance, a reasonable expectation of perhaps another millennium’s survival might be expected. Perhaps the Christian thing to do would be for me to immediately suffer crucifixion, withhold this book from publication, and let these advances slumber another two thousand years.
Instead let us take faith in our hands, time delivers us all to the new future ahead and we must geoengineer it. We will adjust, overcome, and survive. No longer will we humans lie, cheat, steal or kill like Scarlet O’Hara in Margaret Mitchell’s Gone with the Wind. We will need greater food resources for the coming population boom. We must build great air ships to circle our globe and stand between ocean and desert to stir the atmosphere bringing moisture and fertility to badlands. (The umbra cast on the ground causes a Coriolis circular flow of air around it, pulling the moisture ashore.) The future holds shaded glaciers and cities. We will clean our atmosphere. We will drain the sea into the low places of the earth to lower sea level and save our coasts. Our marginal lands will bloom. We will build space ships, and we will go out among the stars of our new destiny. Nothing will hold us back. Amen.
324

EXOCRINOLOGY
325
1 http://www.nasa.gov/topics/earth/features/health-sapping.html
2 Wynn-Edwards, V.C. (1962). Animal Dispersion in Relation to Social Behavior. Oliver & Boyd, London.
3 Eibl-Eibesfeldt, Irenäus. (1970). Ethology the Biology of Behavior, translated by Erich Klinghammer New York: Holt, Rinehart and Winston 530 p.
4 Wilson, Edward O. 1975; Sociobiology The New Synthesis. Cambridge, Mass.: Belknap Press of Harvard University Press. 697p.


Appendix: Administering and collecting healthy adult male facial skin surface lipid
Pheromone Gum Treatment forAddiction and Criminal BehaviorPrepared by: B. Nicholson, Senior Scientist in Charge for Nicholson Science, Tampa, FloridaNicholson Science [email protected]
Executive SummaryObjectiveTest Nicholson Science, Inc. hypothesis that criminal behavior, drug-seeking or addictive behavior, and sexual perversions are due to deficiency of healthy human adult male facial skin surface lipid pheromone, or ‘unconditional paternal love and affection’.
GoalsTo examine the health benefits of our ‘family therapy provided on chewing gum vehicle” including but not limited to criminal behavior as confirmed by arrest records, drug-seeking or addictive behavior as confirmed by toxicology screen, and/or sexual perversion of homosexuality as confirmed by marriage records. In addition, autoimmune diseases of various kinds should benefit dramatically from pheromone therapy.
SolutionA double-blind, cross-over experimental protocol exposing seventy participants in equal proportion to Nicholson Science home remedy of harmless chewing gum and harmless healthy human adult male facial skin surface lipid pheromone (face grease) or canola oil placebo is proposed.Materials and MethodsHuman healthy adult male facial skin surface lipid pheromone is a common environmental substance in constant contact with human skin and ingested by human populations by millions of kilograms on an everyday basis (during and after affectionate behavior). No allergies to face grease have ever been reported in the medical literature. To guard against transmission of infection, the pheromone donors in obvious good health were provided with a physician administered history and physical examination and tested for HIV, Hep B, and Hep C.All participants will receive information about the experiment and their express, written consent obtained and kept on file. However, anyone may leave the trial at any time without penalty. Thirty-five placebos and thirty-five home remedies will be supplied to seventy physician-selected diagnosed drug addicts with empirical evidence of addictive drug exposure, and/or criminals with verifiable records of recent arrest. (These criteria may be narrowed or expanded by your doctor to match available patient population.) Results will be monitored for a convenient period of time for all seventy in the trial. Then, upon completion of the time period, the key will be broken open and those in the placebo group will be provided the pheromone gum, and those results monitored. The key will remain in the possession of your doctor, to be broken only in the event of emergency or when your doctor sees fit. Because chronic criminality and drug addiction may cause brain damage, younger participants will be preferred, i.e. those most prone to recidivism.
EXOCRINOLOGY
327

Expected ResultsOpen, anecdotal trials of identical doses of healthy human adult male facial skin surface lipid on chewing gum vehicle taken by mouth have resulted in behavioral improvement in 73 of 73 cases observed by non-physician Nicholson. Full and complete recoveries, with resumption of family life, increased ambition, higher grades, and promotion have been observed anecdotally and is expected. Pheromone-stimulated recoveries from autoimmune diseases have also been reported, but may not be common enough to warrant reporting. Recoveries from long term autoimmune diseases like diabetes type 1 and Alzheimer’s disease may take years or they may not be possible if all tissue is destroyed.Side EffectsA following effect has been observed in children and young teens. It is an unusual curiosity about the trials and about pheromone donors. It lasts for only a few days. Jealousy can be caused artificially by the pheromone gum, not in the proband, but in the proband’s osculation partner. Apparently, intuition is a chemosense. Scope or Listerine mouthwash before osculation is suggested, or just quarantine by no kissing for 6 to 8 days. Ambition increases. Grades improve. Goal setting takes place and goals are attained in every case so far. Spontaneous recovery from autoimmune diseases (epilepsy, hypothyroidism, Graves aut, diabetes, arthritis, Alzheimer’s disease).• No complaints have been lodged.• Pheromone is odorless, colorless, tasteless, and imperceptible in action after initial relief.• Several recipients have made glowing video taped self-reports, available for viewing on DVD, ask your doctor.• Complete recoveries from criminal and misbehavior (petty delinquency, minor assaults, repeated running away, juvenile burglary, recalcitrance with poor scholastic performance) n=70; drug addiction (1 methadone, 1 crack cocaine), n=2; and homosexual ideation and behavior (1 juvenile male, 7 female) n=8. Homosexuals have not complained and none wished to revert to homosexual behavior, but three confessed something shocking. Had they known that the pheromone would work, they would never have consented. For that reason, consenting homosexual adults must be excluded from these trials initially.Alcoholics (n=4) have not recovered. (Ethanol is processed in the liver and uses a different pathway from direct-acting addictive drugs). Alcoholics must be excluded from these trials initially. Pheromone Gum Treatment for Addiction and Criminal Behavior
Informed Consent FormThank-you for offering to test human paternal skin surface lipid pheromone on chewing gum. You must be in goodgeneral health, and able to consent on your own to participate. Homosexual people must be excluded for technicalreasons. Alcoholics must also be excluded. You must agree not to take aspirin, addictive drugs, or antibiotics for threedays prior to chewing the gum unless your doctor says that you must. You will also need to avoid deep kissing yourspouse or significant other on the mouth for a week or two. You must allow us to inspect your medical and legalrecords.We promise never to disclose your name or identifying information to anyone, ever, except government officials who needto know.First of all, just because you are reading this and said you may want to participate, you are under no obligation to doanything. You can stop right now or at any time. You do not have to report any results to Nicholson Science or toanyone else, either. Your contact information can be erased and no record will be kept if you withdraw, just tell us. Afteryou chew the gum, we must keep a record, however, for the government to see if they ever decide to look. Your name
328

will never be published in any medical journal or news articles that we write, ever.Double-blind Cross-over ExperimentThis is a double-blind cross-over experiment. Double-blind means the person giving you the gum will not know if it isplacebo gum or pheromone gum. We will use the numbers engraved on the can tops to tell whether you got thepheromone gum or the placebo gum after distribution and the first evaluation is over. Cross-over means that people whodon’t get the pheromone gum do get it from us later. The experiment means that we must give participants such asyourself, fake placebos about half the time to keep things scientific. Besides your doctor in an emergency, nobodylocally knows which is which, pheromone gum or placebo gum. Each looks exactly alike, tastes alike, and is packagedalike. You will definitely get the pheromone gum we are testing, but you may get it right away, or you may get it after aperiod of time. No matter what, you will get it, so be sure we have your contact information on file where you receive thegum.Free of ChargeThis pheromone gum for delinquency, criminal behavior, and/or drug addiction is provided free of charge to you for you tochew to hopefully cure yourself of criminal behavior, runaway behavior, or addictive drug-seeking. You may not sell it orgive it away to anyone else as that would be unethical and inappropriate.This is an unpaid enrollment in a clinical trial being run by Nicholson Science, a natural products health care company.Enrollees are maintained in anonymity, but information obtained from you may be provided to government agencies asrequired by law.Home Remedy TestThis is not an investigational new drug, this is a home remedy that is not subject to FDA oversight. It combines ordinarychewing gum and about 0.01% adult male facial skin surface lipid that we believe is a human pheromone.We do not process the putative pheromone in any way, but we are careful to prevent contamination or spoilage. That iswhat the can is for. Other human pheromones are passed through the air. Even human pheromones produced from petroleum, are completely unregulated everywhere in the world. Pheromones often have very specific stereochemistry, and enantiomers or mirror images of pheromones have poisoned pheromone receptors in animals, so the racemic mixtures produced from petroleum-based products might theoretically be dangerous. FDA doesn't worry. Dozens of human pheromone flavors similar to this one are for sale on Ebay. No adverse consequences have ever been reported anywhere to our knowledge, but there is always a first time. Our pheromone gum is of non-racemic stereochemistry and a natural, unprocessed flavor that you cannot see, taste or smell. Even though human pheromones are the most potent drugs ever discovered, their power and potential have until now remained obscure. We do know that pheromone reception presents as “intuition”, and is both cryptic and insidious. We believe that you will get better, and may even feel better initially if you have a severe deficiency, but in a few days after chewing all the gum, you will not notice any changes in the way you feel. In our previous experience, your chief complaint of bad behavior will stop, but the change will not be noticeable to you. For instance, if you are in school, you will only notice that your grades improve if you are making bad grades because of your pheromone deficiency.Source of the Pheromone Gum Home Remedy and Placebo GumYou are getting human paternal pheromone gum obtained from the skin of an adult male donor in excellent health,
EXOCRINOLOGY
329

without skin blemishes. All donors have been screened and cleared by physicians after physical examination and testingfor blood born viruses. Volunteer donors are men of good character, with no criminal background, and living in familiesas fathers and uncles. They are not paid for their services.The Placebo is also ordinary Wrigley’s chewing gum, in the same flavor. Each piece is unwrapped and about 10milligrams of a tasteless oil (canola oil) is applied, then re-wrapped and packaged exactly like the Pheromone Gum, sothat no one without the key can tell which is which. Keeping the key secret is very important in scientific experiments toavoid unintentional errors.DiscussionHuman pheromones are passed in kissing between people. Our best guess is, when we fail to get enough pheromonefrom our families during quality time spent with parents, we contract a range of aberrant behavioral conditions. This teambelieves that each of these conditions may be amenable to pheromone treatment. We propose that the conditioncaused by surplus of maternal pheromone and deficiency of paternal pheromone is homosexual behavior. Similarly wethink that there is a condition which is caused by deficiency of human pheromone of both maternal and paternalpheromone and it is juvenile delinquency and/or criminal behavior, borderline personality disorder, and perhaps, someforms of hysteria probably including obsessive-compulsive disorder. The more extreme deficiencies caused by lack ofparental closeness cause violent behavior or addictive drug seeking, we think.The root cause of the paternal pheromone deficiency is under investigation. Parental withdrawal and lack of receptivitytoward parents in youngsters seems to be due mostly to air pollution, but we do not know for sure. Ozone is the mostclosely correlated pollutant, which makes sense because ozone irritates pheromone receptive areas of the olfactory andrespiratory tract. Ozone also attacks pheromones chemically (ozonolysis). We believe that these super sized naturaldoses of pheromone seem to cure these detrimental effects of ozone pollution.We expect that delinquency and criminal behavior can be stopped completely with a single 15 piece dose of this humanpheromone. One dose (or fake placebo) is supplied on fifteen pieces of fresh, new, dry sugarless chewing gum obtainedfrom a store. We use sugarless chewing gum because it keeps longer and because it maintains the pheromone nearreceptors in the nose, mouth, and upper respiratory system. (The upper respiratory system is covered with pheromone-receptive microvillar or brush border cells.)Previous Experience with Pheromone Gum Home RemedyIn previous open trials, just anecdotes really, illegal drug-seeking behavior has been cured twice out of two tries by thisdosage of paternal skin surface lipid chewing gum. Both cases resulted in complete remission of all symptoms.Recipients ceased inappropriate sexual promiscuity and resumed normal age-appropriate sexual behavior.We must exclude homosexual people from this trial for technical reasons. Homosexual behavior in both young men and
330

women also seems to be treatable by approximately 150 mg of the human pheromone on chewing gum vehicle,however relapses occur and booster doses may be necessary to sustain heterosexuality. We have had eight successesout of eight tries, so far. However, homosexuals are to be excluded from this trial because they have told us that loss ofhomosexual ideation and behavior is a detrimental side effect.
Side Effects Observed in Previous Open TrialsSide effects can include jealousy. It is why you need not to kiss anyone on the mouth for a few days after you take thepheromone gum. The jealousy that is created artificially by a tiny dose of human pheromone is real to the jealous person,so avoid it by not kissing anyone on the mouth. Jealousy, like anger, is a dangerous human emotion, but since we knowthe conditions that cause it, we can avoid any problems.Another side effect is "following effect". The dosed and now former delinquent or problem child feels a slight euphoriaand a deep sense of gratitude which passes within a few days. The euphoria’s intensity depends upon the extent of thepheromone deficiency. No criminal, no addict, nor any homosexual has expressed any interest in returning to formerlifestyles, but Nicholson Science cannot change your lifestyle. You have the free will to do that yourself, as we believeyou will discover once “the monkey is off your back.”Ambition and improved job or school performance has been observed, too. Goals are set and achieved.Ten people who suffered from autoimmune diseases suddenly began to recover and this surprised them enough thatthey sought medical advice on an emergency basis. Nicholson Science will not reimburse you or indemnify you for yourhealth care. We wish we could.RisksThe pheromones are odorless, colorless, tasteless, and imperceptible in action. We believe that they are inherently safe,but we could be wrong. So far as we know, no human being has ever kissed another human being and gotten anallergic reaction. We use good laboratory practices and good sanitary hygiene in collecting human pheromone. We donot use universal precautions (like in surgical operating rooms) in collection of pheromone because people can havedangerous allergies to the latex of gloves. The pheromone on the chewing gum vehicle is exposed briefly to ultravioletlight at antibiotic wavelengths just like many fruits and vegetables and is kept desiccated in a nobel gas atmosphere (nooxygen to cause spoilage) in a sealed and light proof aluminum can. The chewing gum as it came from the storecontains USDA approved preservatives, which also deter spoilage.There is minimal risk of infection for transmissible infectious diseases. However, it is less than the ordinary risk of kissingthe donor on the cheek because we UV sterilize, desiccate the gum, keep it sealed in a can, and use a preservative-protected chewing gum vehicle, Wrigley’s Rain Spearmint gum.We believe that addictive substances are ersatz pheromones. (Indeed most addictive substances, many NSAIDS, and
EXOCRINOLOGY
331

antibiotics and are also semiochemicals that plants and microbes use to influence behavior of animals, but processing isgenerally required to recover them from natural sources.) Avoid the use of addictive substances, aspirin, and antibiotics ifallowed by your physician during the administration period. It usually takes a few hours to chew all 15 pieces of the gumserially. Human paternal pheromone chewing gum has been effective in every case where addicted loved ones sufferedfrom criminal behavior. Every time, the addictive substance seeking was eliminated and criminality ceased.Yours in service,B. Nicholson, Senior Scientist in Charge.Call long distance (1-813 830-1432) to report any problems or concerns by leaving a message.Mechanism of action.Methamphetamine attacks the nucleus accumbens, a pheromone reception center. By providing a small amount of thepheromone, 150 mg or less than half an average aspirin tablet, methamphetamine is no longer sought.
Collection of healthy adult male facial skin surface lipid
Donors must be family men of good character, heterosexual, non-perverted, over 30 years of age, in good health, pass a physical examination, be tested free of blood born viruses, and without skin lesions or clinical skin infections. Donors must agree to avoid cosmetics (except morning use of pure soap), to avoid any medications, to avoid any intoxicating beverages or substances for three days prior to the test. Donors must be free of allergies to chewing gum.The facial skin surface sebaceous skin surface lipid pheromone should be collected by a technician over three sessions over no fewer than three days (each session at least twelve hours apart or until 15 to 25 pieces of pheromone gum are collected.Using a clean dinner plate, open 5 pieces of Wrigley’s Rain Chewing gum (or equivalent), saving the wrappers. Artificially sweetened gum is used. Tare the weight of the fresh, dry, new gum. Rub the gum on the face of the donor anywhere a child would kiss his/her father.Reweigh the gum. Collect until 10 milligrams have been collected. Use both sides if necessary.Irradiate gum with biocidal UV lamp for 30 seconds per side.Re-wrap the gum.Label each piece of the gum: “Doner #, date collected, HOME REMEDY Experimental use only. Not for resale Fresh until (get date from retail package)”, and any other pertinent information at the discretion of the local supervising IRB, Institutional Review Board for human subjects.
NICHOLSON SCIENCE PROVIDES THIS PROTOCOL TO THE GENERAL PUBLIC AS A PUBLIC SERVICE. DONATIONS, WHICH ARE NOT TAX DEDUCTIBLE, SHOULD BE SENT VIA PAYPAL TO
Suggested donation is $25.00 US per person cured.
332





























