Embed Size (px)
Citation preview

Estrogen Receptor-b in Quail: Cloning, TissueExpression and Neuroanatomical Distribution
Agnes Foidart,1 Bernard Lakaye,1 Thierry Grisar,1 Gregory F. Ball,2
Jacques Balthazart1
1 Laboratory of Biochemistry, University of Liege, 17 place Delcour, Bat. L1, B-4020 Liege, Belgium
2 Department of Psychology; Behavioral Neuroendocrinology Group, Johns Hopkins University,Baltimore, Maryland 21218
Received 10 December 1998; accepted 3 March 1999
ABSTRACT: A partial estrogen receptor-b (ERb)cDNA had been previously cloned and sequenced inJapanese quail. The 3*- and 5*–rapid amplification ofcDNA ends techniques were used here to identify acDNA sequence of the quail ERb that contains a com-plete open reading frame. For the first time in an avianspecies, this cDNA sequence and the correspondingamino acid sequence are described. They are comparedwith the known ERb sequences previously described inmammals and with the ERa sequences identified in aselection of mammalian and avian species. The analysisby Northern blotting of the ER b mRNA expression inthe brain and kidneys revealed the presence of severaltranscripts. The presence of ERb identified by reversetranscriptase–polymerase chain reaction demonstrateda widespread distribution quite different from the dis-tribution of ER a. The complete neuroanatomical distri-
bution of ERb mRNA as determined by in situ hybrid-ization with 35S- and 33P-labeled oligoprobes is alsopresented. Transcripts are present in many nuclei im-plicated in the control of reproduction such as the me-dial preoptic nucleus, the nucleus striae terminalis, andthe nucleus taeniae, the avian homologue of the amyg-dala. These data demonstrate the presence of ERb in anonmammalian species and indicate that the (neuro)-anatomical distribution of this receptor type has beenconserved in these two classes of vertebrates. The role ofthis receptor in the control of reproduction and otherphysiological processes should now be investigated.© 1999 John Wiley & Sons, Inc. J Neurobiol 40: 327–342, 1999
Keywords: estrogen receptor-b; ERb mRNA; in situhybridization; northern blot; RT-PCR; hypothalamus;neuroanatomical distribution; Japanese quail;Coturnixjaponica
Estrogens play a key role in the control of female andmale reproductive physiology, of sexual behavior aswell as other physiological process such as bone cal-
cification, memory, or cognition (McEwen et al.,1997; Cooke et al., 1998). However, until recently,there has been some confusion about the mechanismsof estrogen action given that estrogens have biologicaleffects in tissues such as the telencephalon or bladderthat appear to be devoid of the intracellular receptorsneeded for physiological responses to estrogen. Al-though estrogens can have rapid nongenomic effectsin the central nervous system (Ramirez et al., 1996;Mermelstein et al., 1996; Joe¨ls, 1997), most of theiractions during development and in adulthood arethought to be mediated by a ligand-activated nucleartranscription factor, the estrogen receptor (ER), that
Correspondence to:J. BalthazartContract grant sponsor: NIMH; contract grant number: R01
MH50388Contract grant sponsor: Belgian FRFC; contract grant number:
9.4565.96FContract grant sponsor: University of Lie`geContract grant sponsor: Belgian FNRS; contract grant number:
1.5.081.98Contract grant sponsor: NATO; contract grant number: CRG
973000© 1999 John Wiley & Sons, Inc. CCC 0022-3034/99/030327-16
327

regulates the expression of specific genes through itsbinding to specific response elements on the DNA.The ER has been cloned (Green et al., 1986; Korke etal., 1987), and subsequentin situhybridization studieshave detected ER mRNA in a large number of ratbrain regions including the preoptic area, nucleusstriae terminalis, and ventromedial hypothalamus(e.g., Simerly et al., 1990; Shughrue et al., 1992).These observations are generally in good agreementwith previous autoradiographic, immunocytochemi-cal, and biochemical studies. However, more recently,a nuclear uptake and retention of radiolabeled estro-gen has been observed in cells of the preoptic area andhypothalamus in ER knockout mice (Lubahn et al.,1993), and it has been shown that estrogens are stillcapable of modulating the expression of specificgenes in the brain of these mice (Shughrue et al.,1997c; Moffat et al., 1998; Shughrue, 1998). Thesedata therefore suggested that another type of ER, notaffected in the ER knockout mice, may also bepresent.
A second form of estrogen receptor has now beencloned in the rat (Kuiper et al., 1996) . The discoveryof this new receptor, referred to as estrogen recep-tor-b (ERb), as opposed to the previously character-ized ER, now called ERa, has generated a consider-able amount of interest because it raises a number ofquestions regarding the physiological roles, sites, andmodes of action of the two receptors in a variety oftarget tissues (Kuiper et al., 1998). To our knowledgeERb has been detected so far in only a few species ofmammals (rat: Kuiper et al., 1996; mouse: Tremblayet al., 1997; human: Mosselman et al., 1996), with theexception of an ER sequence cloned in a fish species,the eel (Todo et al., 1996), and suggested later to behomologue to ERb (Enmark et al., 1997). No infor-mation is available concerning the presence of ERb inother vertebrate classes, including birds.
In avian species as in mammals, estrogens play acritical role in the sexual differentiation and in theactivation in adulthood of reproductive behavior. InJapanese quail specifically, estrogens produced byaromatization of testosterone in the brain activatemale copulatory behavior and regulate the concentra-tion of aromatase and of its messenger RNA (Baltha-zart and Foidart, 1993; Balthazart, 1997; Balthazartand Ball, 1998a). It has usually been assumed thatestrogens produced by aromatization of testosteroneactivate male sexual behavior by acting in the cellswhere they are produced (intracrine action). It wassimilarly postulated that the aromatase synthesis iscontrolled by estrogens acting on ER localized in thecell where estrogens are synthesized (see Balthazart,1997; Balthazart and Ball, 1998b, for further discus-
sion). The distribution of neurons expressing ERa andaromatase has therefore been studied by double-labelimmunohistochemistry in the quail central nervoussystem; surprisingly, it was found that the estrogen-synthesizing enzyme and the ERa are usually notcolocalized (Balthazart et al., 1991) in the areas thatplay a key role in the activation of reproductive be-havior such as the medial preoptic nucleus (POM) andthe nucleus striae terminalis (BST) (Balthazart andFoidart, 1993; Balthazart et al., 1998; Riters et al.,1998). This led us to consider other modes of estrogenaction including nongenomic effects of the steroid onthe activation of sexual behavior and transsynapticcontrols of aromatase synthesis by estrogen-depen-dent catecholaminergic inputs (Balthazart and Ball,1998b). The recent discovery of ERb indicates that areevaluation of these questions might be useful be-cause this new estrogen receptor could be located inaromatase-containing neurons and mediate the actionsof the steroid in these neurons via classical genomicmechanisms.
Our first attempts to localize the ERb in quail brainby immunocytochemistry with commercially avail-able antibodies raised against mammalian ERb se-quences were unsuccessful; we therefore decided tostudy the distribution of the corresponding messengerRNA (mRNA) by in situhybridization. A partial quailERb cDNA from testicular tissue was cloned andsequenced and its distribution was analyzed in therostral forebrain byin situ hybridization with 35S-labeled oligoprobes (Lakaye et al., 1998). This studyidentified a high ERb expression in three limbic nu-clei—the POM, BST, and nucleus taeniae—that haveall been implicated in the control of reproduction(Balthazart and Ball, 1997; Thompson et al., 1998;Balthazart et al., 1998) and contain high levels ofaromatase activity (Schumacher and Balthazart,1987).
However, these studies left unanswered a numberof questions concerning the full sequence of ERb inquail, its tissue distribution, and detailed neuroana-tomical localization. In the present study, we used the39– and 59–rapid amplification of cDNA ends (RACE)technique to identify the cDNA sequence of the quailERb that contains a complete open reading frame. Wedescribe here, for the first time in an avian species,this cDNA sequence, the corresponding amino acidsequence, and its comparison with the known mam-malian sequences. We analyze the expression of theERb mRNA in representative tissues by Northernblotting, we describe its tissue distribution by reversetranscriptase–polymerase chain reaction (RT-PCR),and finally, we present its complete neuroanatomical
328 Foidart et al.

distribution as determined byin situ hybridizationwith 35S- and33P-labeled oligoprobes.
MATERIALS AND METHODS
Construction of Full-Length ERb cDNA
In our previous study, a partial cDNA sequence of the quailERb was isolated and sequenced by using four degenerateprimers based on parts of the rat ERb molecule, as de-scribed by Kuiper and collaborators (1996). These primershad been selected in regions of the molecule that are pre-sumably conserved (DNA binding domain; DBD1 andDBDb1 primers) or have been shown to discriminate be-tween ERb and ERa in rat (ligand binding domain; bER1and bER3 primers). Using nested PCR, we amplified afragment of the expected size (482 bp) whose sequence wasfound to be highly homologous to a region of the rat ERbmRNA located between the DNA- and ligand-binding do-mains (Lakaye et al., 1998).
The 39 and 59 ends of the ERb cDNA were isolated byRACE (Frohman and Marin, 1989), using primers synthe-sized on the basis of the previously cloned quail testis ERbcDNA fragment (nt 400-881 relative to ATG) (Lakaye etal., 1998). Total RNA was prepared from quail testis ac-cording to the acid guanidinium thiocyanate-phenol-chloro-form single-step method and poly (A)1 RNA was isolatedusing oligotex (Qiagen, Hilden, Germany).
The 59-translated region was cloned by 59-RACE usingthe 59/39-RACE kit (Boehringer Mannheim, Mannheim,Germany). Primer qbERSP1 (59-GAGACTGAGATCAAT-GAAGCCAGG-39; nt 831-808 relative to ATG) was usedfor reverse transcription using avian myeloblastosis virus(AMV) reverse transcriptase. After polyA tailing, the oli-godT-anchor primer of the kit and the qbERSP2 primer(59-ACTCACCAACACGTTGGGTGGCTC-39; nt 708-685relative to ATG) were used for PCR amplification (40cycles at 94°C for 20 s, 65°C for 30 s, and 72°C for 90 s).Amplification products were run on a 1% agarose gel,transferred onto a charged nylon membrane (BoehringerMannheim), and hybridized with a dig-labeled oligonucle-otide HISbERq1 (59-GTTTTGCCCATGCAATCTTC-CGAATTACGATGGCGGCGCAGGATTCGATA-39; nt593-544 relative to ATG). Two bands that hybridized to thisoligonucleotide were eluted with the QIAquick gel extrac-tion kit (Qiagen) and reamplified (25 cycles at 94°C for 20 s,57°C for 40 s, and 72°C for 40 s) using primer qbERSP3(59-GGTTTTGCCCATGCAATCTTCCGA-39; nt 594-571relative to ATG) and the anchor primer of the kit.
The 39-translated region was isolated by 39-RACE usingthe 59/39-RACE and the Expand Long Template kits(Boehringer Mannheim). After reverse transcription usingthe oligodT-anchor primer and the M-MLV (Gibco BRL),the anchor and qYRI (59-TATCGAATCCTGCGCCGC-CATCGT-39; nt 544-567 relative to ATG) primers wereused for PCR amplification with the Expand Long templatekit (10 cycles at 94°C for 10 s, 65°C for 30 s, and 68°C for
3 min, followed by 20 cycles at 94°C for 10 s, 65°C for 30 s,and 68°C for 3 min1 20 s each cycle). After electrophore-sis on a 0.8% agarose gel, the gel portion located between1.5 and 3 kb was isolated and the fragments were elutedwith the QIAquick gel extraction kit (Qiagen). A secondPCR using the anchor and the qSED (59-TCGGAAGATT-GCATGGGCAAAACC-39; nt 571-594 relative to ATG)primers was done using the Expand Long template (sameconditions as above except that the annealing temperaturewas 62°C).
All PCR products were subcloned in pCRII by the TAcloning method (Invitrogen, Leek, The Netherlands) andsequenced in both directions by the Sanger dideoxynucle-otide chain termination method using T7 DNA polymerase(Pharmacia, Uppsala, Sweden). The experiment was re-peated with different primers when a discrepancy occurredin the sequences obtained in the two directions. All oligo-nucleotides not included in the kits were from Eurogentec(Seraing, Belgium). Sequences were aligned with the Ge-neworks program on a Macintosh computer.
After reconstruction of the entire transcript based on thesequence of overlapping fragments, an additional PCR wascarried out using primers located at the two ends of theentire molecule (nt219 to 3 and 1410 to 1430 relative toATG). This identified a fragment of the expected size. Inaddition, we sequenced the two ends of this cDNA andshowed that the sequence obtained matched the sequencepresented in the report.
Northern Blot Analysis
Five micrograms of brain and kidney poly (A)1 RNA werefractionated on a 0.8% agarose–0.4M formaldehyde gel(Tsang et al., 1993). RNA was transferred to a Hybond N1membrane (Amersham, Buckinghamshire, UK) by capillarytransfer in 103 SSC. After ultraviolet (UV) crosslinking,hybridization and wash were performed in an hybridizationoven. The membrane was prehybridized for 3 h at68°C in53 SSC, 50% formamide, 53 Denhardt’s, 1% sodiumdodecyl sulfate (SDS), and 100mg/mL denatured salmonsperm DNA. Hybridization was carried out at 68°C for 20 hin the above solution containing 10 ng/mL of32P-labeledriboprobe (specific activity: 108 cpm/mg). The radiolabeledriboprobe, encompassing nucleotides283 to1315 relativeto ATG, was generated by runoff transcription using SP6RNA polymerase (Boehringer Mannheim) and [a32P]-CTP(800 Ci/mmol; ICN). This probe corresponds to the 59 endof the ERb mRNA, and it shows almost no homology withthe corresponding sequence in the ERa molecule (see Re-sults). The membrane was washed in 23 SSC–0.1% SDS atroom temperature for 23 5 min, in 0.23 SSC–0.1% SDSat room temperature for 23 5 min, and 0.13 SSC–0.1%SDS at 68°C for 23 15 min, sealed in a plastic bag, andexposed to Hyperfilm MP (Amersham) for 1 week at270°C in the presence of an intensifying screen.
Estrogen Receptor-b in Quail 329

Tissue Expression
Adult male and female quail were decapitated and differenttissues were collected, frozen on dry ice, and stored at270°C until used. Tissue samples studied were specificbrains areas such as the brain stem, optic lobes, cerebellum,telencephalon excluding nucleus taeniae (known to containhigh levels of ERa) (Balthazart et al., 1989), and dienceph-alon (including preoptic area and hypothalamus), as well asperipheral tissues, including the pituitary gland, spinal cord,kidney, adrenal gland, liver, testis, epididymis, ovary, ovi-duct, heart, lung, and pectoral and gastrocnemius muscles.They were processed for total RNA isolation, extracts weretreated with DNase I, and cDNA synthesis was performedas described previously (Lakaye et al., 1998).
A 513-bp fragment of the quail glyceraldehyde-3-phos-phate dehydrogenase (GAPDH; Accession no. Z19086) wasamplified for 20 cycles by incubation at 94°C for 30 s, 59°Cfor 30 s, and 72°C for 40 s using forward (59-AACTGTCT-GGCACCATTGGC-39) and reverse (59-CACAACACG-GTTGCTGTATCC-39) primers. For ERb, the same RTvolumes as used for GAPDH were submitted to 35 cycles(94°C for 30 s, 70°C for 30 s, and 72°C for 30 s) usingspecific forward (qYRI; see above) and reverse (59-GCTT-TCCAACAGCCTGACTTGGTC-39; nt 859-835 relative toATG) primers. After gel agarose electrophoresis, PCR prod-ucts were stained with ethidium bromide and visualizedunder UV transillumination.
In Situ Hybridization
Three oligonucleotides derived from the quail ERb cDNA,q1 (59-GTTTTGCCCA TGCAATCTTC CGAATTACGATGGCGGCGCA GGATTCGATA-39 nt 593-544 relative toATG), q2 (59-GCAGACCATA ACAGGCTTGG GCTC-GATGTT TGTCTCACAG ATGCACTCTC-39 nt 110-61relative to ATG), and q3 (59-GTGGTGCAGC TTCCTGT-TGC TTTCGGGTTC TTCTGCTGAC AAGGG-39 nt1137-1093 relative to ATG) (Eurogentec), were used asprobes forin situ hybridization based on their low homol-ogy to chicken (Krust et al., 1989) and zebra finches (Jacobset al., 1996) ERa. They were labeled at the 39 end with35S-dATP or 33P-dATP using terminal deoxynucleotidyltransferase (Boerhinger). A series of preliminary studiesrevealed that the three probes produced the same pattern ofanatomical distribution in the brain and that a cocktail oftwo probes produced the densest hybridization signal.Therefore, a cocktail of the q2 and q3 probes was used tomap the distribution of ERb mRNA in the quail brain.
The brains of eight adult male Japanese quail weredissected out of the skull and immediately frozen on dry ice.Twenty-micrometer sections were cut in coronal plane on acryostat, collected on poly-L-lysine–coated slides, air-dried,and then stored at275°C. At the time of processing, slideswere warmed at room temperature, postfixed in formalde-hyde, and rinsed in phosphate-buffered saline 0.1M. Afterdehydration and delipidation, slides were incubated in hu-mid chamber for 2 h at 37°C in prehybridization cocktail
containing 50% formamide, 43 SSC, 13 Denhardt’s solu-tion, 1% sarcosyl, 20 mM phosphate buffer (pH 7.0), 100mg/mL dextran sulfate, 100mg/mL salmon sperm DNA,250 mg/mL tRNA, 100 mg/mL polyadenylic acid, and,eventually, 60 mM dithiothreitol (DTT). The hybridizationproceeded overnight at 42°C in prehybridization buffer con-taining33P- or35S-labeled probes to a final concentration of900,000 cpm/slide, and a total volume of 300mL wasapplied on each slide. After hybridization, slides wererinsed four times for 15 min in 13 SSC at 55°C, dehy-drated, air-dried, and exposed to Amersham Betamax Hy-perfilm for 2–4 weeks. Control sections were incubated for1 h at 37°C with RNAase A (100mg/mL) before hybrid-ization or with the oligoprobe with or without 100 timesexcess of the corresponding nonlabeled probe.
RESULTS
Quail ERb cDNA
We previously published a partial cDNA sequenceencoding part of the DNA binding domain (DBD),hinge region, and the 59 end of the ligand-bindingdomain (LBD) of the quail testes ERb. The 59 and 39ends of the cDNA were obtained here by 59- and39-RACE reactions (Fig. 1).
The 59-RACE procedure generated a 692-bp prod-uct whose analysis identified an ATG initiating codonat position 84. This ATG is flanked by nucleotidesthat fit the Kozak consensus sequence (Kozak, 1986).Alignment of the RACE sequences with the previouspartial cDNA identified an open reading frame of1416 bp encoding a protein of 472 amino acids witha calculated molecular weight of 53.3 kD.
The quail ERb sequence contains several potentialphosphorylation sites for protein kinase A, proteinkinase C, and protein kinase CK2. A single consensusMAP kinase phosphorylation site (Ser60) is present inthe amino-terminal region. This site is equivalent tothe MAP kinase phosphorylation site found in themouse ERb whose phosphorylation by the Ras-Raf1-MAPK kinase-MAPK pathway enhances ER tran-scriptional activity (Tremblay et al., 1997). A con-served tyrosine residue (Tyr439), important for ERdimerization and binding to estrogen response ele-ments (ERE), is present just before the AF2 domain.
Analysis of the predicted protein sequence showsthat it is more related to the estrogen receptor than toother steroid receptors. Based on pairwise proteinalignments, this sequence shares a higher overallidentity with the rat ERb (80%) and human ERb(77%) than with the eel ER (62%) or with the ERaidentified in mammals or birds (52–55%).
As shown in Table 1, the DNA-binding domain is
330 Foidart et al.

highly conserved between ERa and ERb from mam-mals to fish. On the other hand, in the LBD and theamino-terminal domain (NHD), the new quail se-quence is more related to the rat and human ERb, oreven the eel ER, than to the chicken or zebra finchERa. These results support the notion that this se-quence corresponds to the quail ERb.
The quail ERb is slightly smaller than the ERbsequenced in mammals. Most of the deletions areobserved in the carboxy-terminal domain and hingeregion, the two regions showing the lowest percentageof homology between species (Fig. 2).
More surprisingly, two amino acids are also de-leted in the ligand-binding domain of the quail ERbcompared to mammals. These deletions are, however,observed in a region of low identity between the ERisoforms (a and b), and more precisely, near a sur-face-exposed, protease-accessible region (Seielstad etal., 1995). This deletion is therefore of relativelyminor importance for the functional properties of thereceptor. In general, the ERb are markedly shorterthan the ERa (deletions at the C and even moreprominently at the N terminal). This is true for allsequences identified so far in mammals and birds.
Northern Blot
A Northern blot using 5mg of quail poly(A)1 RNAfrom brain and kidney was probed with a6400-bpradioactive riboprobe encompassing the amino-termi-nal domain and part of the 59UTR. As shown inFigure 3, at least three predominant transcripts ofapproximately 3.5, 6.5, and 8.0 kb are expressed inthe kidney. In the brain, only the two larger transcriptscould be detected. These mRNA are approximately ofthe same length as those encoding the mouse or hu-man ERb (Tremblay et al., 1997; Kuiper and Gustafs-son, 1997). The overall expression level is higher inthe kidney than in the brain; in addition, for eachtissue, the relative expression of the various tran-scripts is different. In the kidney, the 3.5- and 6.5-kbtranscripts are expressed at the same but much higherlevel than the 8.0-kb transcript. In contrast, the 8.0-kbtranscript is the most prominent in the brain.
Expression of ERb mRNAin Quail Tissues
Given the low level of ERb mRNA expression in thequail brain, we used RT-PCR with specific primers todetermine the relative distribution of ERb transcriptsin several quail tissues and in specific brain areas. Therelative amount of ERb mRNA was estimated by asemiquantitative procedure with two sets of primers
that allowed the amplification of ERb mRNA andGAPDH transcripts used as internal standard. In alltissues studied, this analysis identified a single band atthe expected length and the transcript distributionobserved in quail was in agreement with data previ-ously reported for the rat ERb (Fig. 4). The highestexpression of ERb mRNA was found in the liver,kidney, ovary, and oviduct. ERb also appeared to bedensely expressed in the telencephalon, diencephalon,optic lobes, brain stem, spinal cord, pituitary, andepididymis. The testes, adrenals, and lung showed alighter but definitely positive expression, whereas cer-ebellum, heart, and pectoral and gastrocnemius mus-cles only displayed a very low, barely detectableexpression of ERb mRNA.
Neuroanatomical Distributionof ERb mRNA
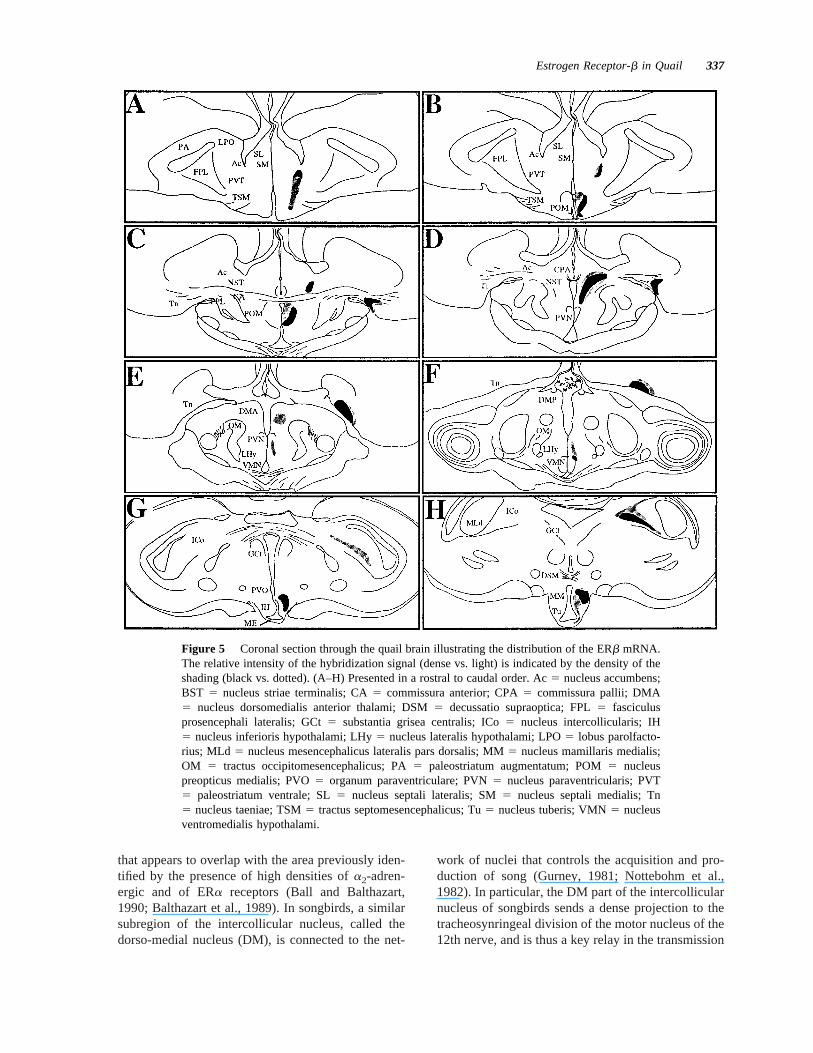
Three antisense oligonucleotide probes correspondingto different domains of the ERb cDNA were used forin situ hybridization studies. q1 is a fragment in thehinge domain, q2 is located in the NHD, and q3 is apart of the LBD. These probes correspond to parts ofquail ERb sequence that have a minimal homologywith the corresponding fragment of the ERa sequencein chicken (Figs. 1 and 2) or with other known se-quences in Genbank. Probes were labeled with either35S-dATP or 33P-dATP. Sections obtained throughthe rostral to caudal extent of the brain were processedby in situ hybridization either with each of these threeprobes separately or with several combinations of twodifferent probes (q11q2, q11q3, and q21q3). In allcases, a similar pattern of ERb mRNA distributionwas observed. Therefore, we present here only theresults obtained with the combination of probes q2and q3 labeled with33P, the condition in which thedensest autoradiographic signal was obtained.
An extensive distribution of the ERb mRNA wasdetected throughout the rostral-caudal extent of thehypothalamus, in the mesencephalic nucleus intercol-licularis, and in the telencephalic nucleus taeniae.This distribution is shown in Figure 5; representativeautoradiograms are presented in Figure 6.
At the level of the most rostral part of the preopticarea, a dense vertical band of hybridization signal wasobserved running from the ventral tip of the lateralventricle to the level of the tractus septomesencephali-cus (TSM) [Figs. 5(A) and 6(A)]. In slightly morecaudal sections, an intense hybridization signal wasfound in the medial preoptic nucleus (POM). TheERb mRNA labeling observed in the POM definedthe exact boundaries of the nucleus throughout itsrostral to caudal extent, although the density of the
Estrogen Receptor-b in Quail 331

Figure 1
332 Foidart et al.
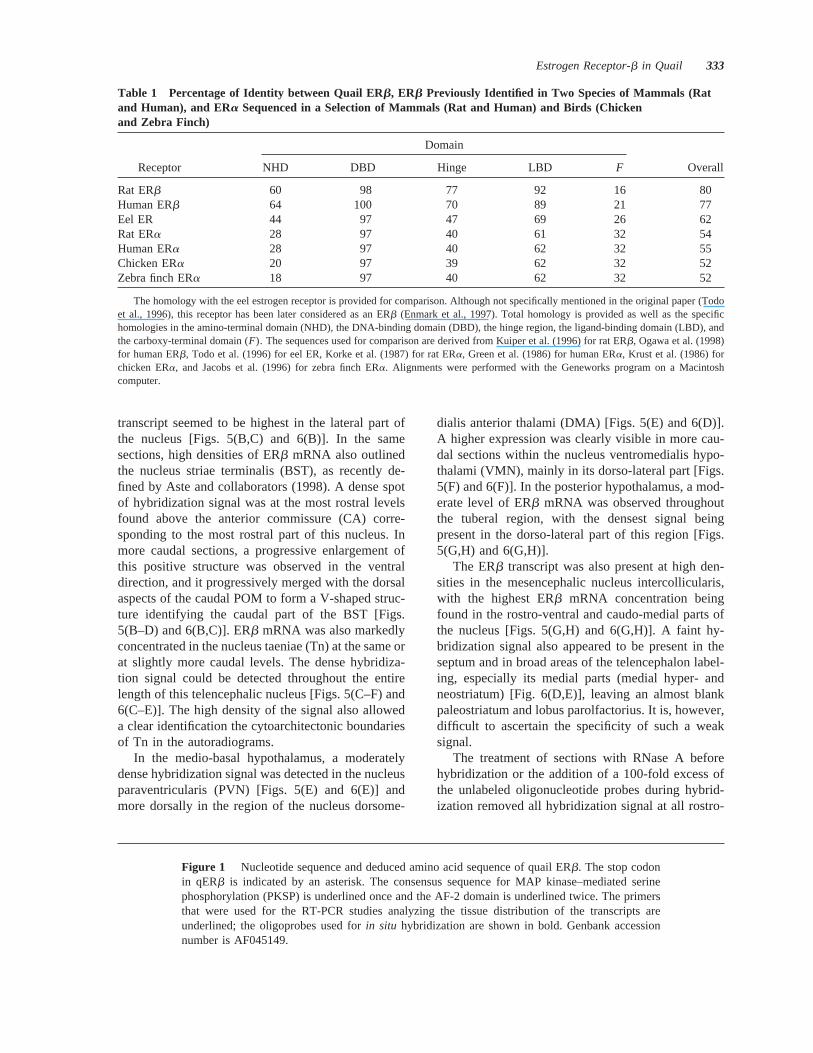
transcript seemed to be highest in the lateral part ofthe nucleus [Figs. 5(B,C) and 6(B)]. In the samesections, high densities of ERb mRNA also outlinedthe nucleus striae terminalis (BST), as recently de-fined by Aste and collaborators (1998). A dense spotof hybridization signal was at the most rostral levelsfound above the anterior commissure (CA) corre-sponding to the most rostral part of this nucleus. Inmore caudal sections, a progressive enlargement ofthis positive structure was observed in the ventraldirection, and it progressively merged with the dorsalaspects of the caudal POM to form a V-shaped struc-ture identifying the caudal part of the BST [Figs.5(B–D) and 6(B,C)]. ERb mRNA was also markedlyconcentrated in the nucleus taeniae (Tn) at the same orat slightly more caudal levels. The dense hybridiza-tion signal could be detected throughout the entirelength of this telencephalic nucleus [Figs. 5(C–F) and6(C–E)]. The high density of the signal also alloweda clear identification the cytoarchitectonic boundariesof Tn in the autoradiograms.
In the medio-basal hypothalamus, a moderatelydense hybridization signal was detected in the nucleusparaventricularis (PVN) [Figs. 5(E) and 6(E)] andmore dorsally in the region of the nucleus dorsome-
dialis anterior thalami (DMA) [Figs. 5(E) and 6(D)].A higher expression was clearly visible in more cau-dal sections within the nucleus ventromedialis hypo-thalami (VMN), mainly in its dorso-lateral part [Figs.5(F) and 6(F)]. In the posterior hypothalamus, a mod-erate level of ERb mRNA was observed throughoutthe tuberal region, with the densest signal beingpresent in the dorso-lateral part of this region [Figs.5(G,H) and 6(G,H)].
The ERb transcript was also present at high den-sities in the mesencephalic nucleus intercollicularis,with the highest ERb mRNA concentration beingfound in the rostro-ventral and caudo-medial parts ofthe nucleus [Figs. 5(G,H) and 6(G,H)]. A faint hy-bridization signal also appeared to be present in theseptum and in broad areas of the telencephalon label-ing, especially its medial parts (medial hyper- andneostriatum) [Fig. 6(D,E)], leaving an almost blankpaleostriatum and lobus parolfactorius. It is, however,difficult to ascertain the specificity of such a weaksignal.
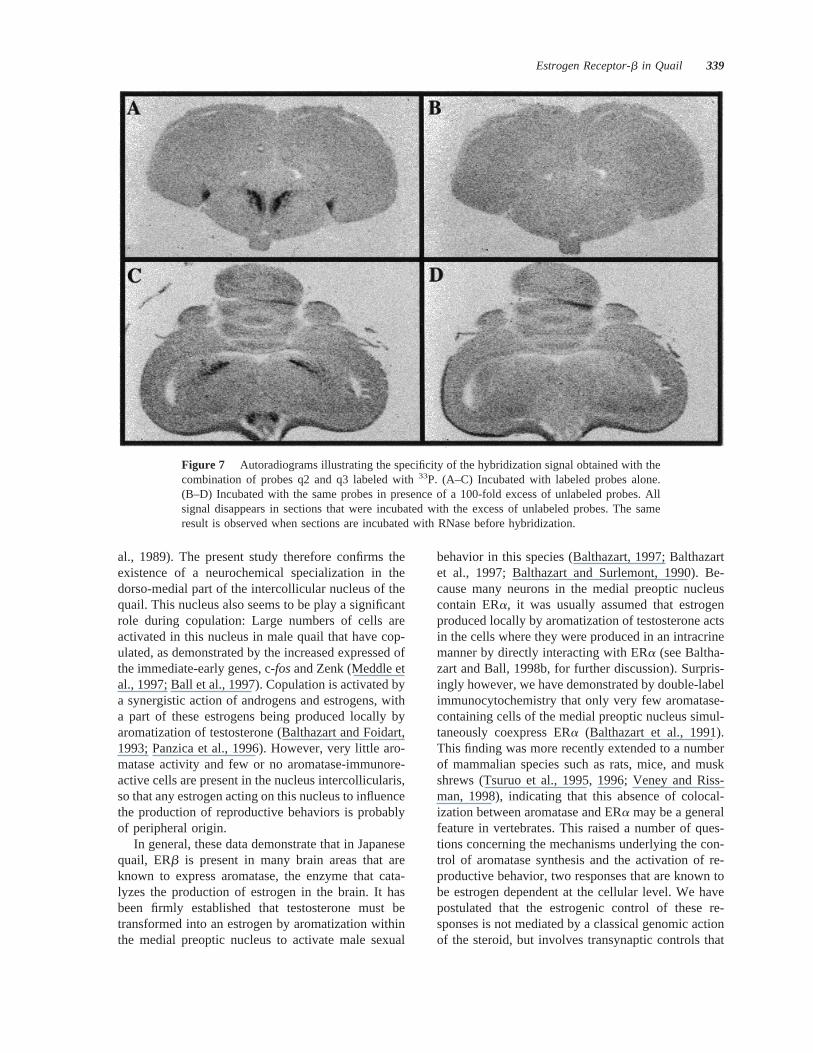
The treatment of sections with RNase A beforehybridization or the addition of a 100-fold excess ofthe unlabeled oligonucleotide probes during hybrid-ization removed all hybridization signal at all rostro-
Table 1 Percentage of Identity between Quail ERb, ERb Previously Identified in Two Species of Mammals (Ratand Human), and ERa Sequenced in a Selection of Mammals (Rat and Human) and Birds (Chickenand Zebra Finch)
Receptor
Domain
OverallNHD DBD Hinge LBD F
Rat ERb 60 98 77 92 16 80Human ERb 64 100 70 89 21 77Eel ER 44 97 47 69 26 62Rat ERa 28 97 40 61 32 54Human ERa 28 97 40 62 32 55Chicken ERa 20 97 39 62 32 52Zebra finch ERa 18 97 40 62 32 52
The homology with the eel estrogen receptor is provided for comparison. Although not specifically mentioned in the original paper (Todoet al., 1996), this receptor has been later considered as an ERb (Enmark et al., 1997). Total homology is provided as well as the specifichomologies in the amino-terminal domain (NHD), the DNA-binding domain (DBD), the hinge region, the ligand-binding domain (LBD), andthe carboxy-terminal domain (F). The sequences used for comparison are derived from Kuiper et al. (1996) for rat ERb, Ogawa et al. (1998)for human ERb, Todo et al. (1996) for eel ER, Korke et al. (1987) for rat ERa, Green et al. (1986) for human ERa, Krust et al. (1986) forchicken ERa, and Jacobs et al. (1996) for zebra finch ERa. Alignments were performed with the Geneworks program on a Macintoshcomputer.
Figure 1 Nucleotide sequence and deduced amino acid sequence of quail ERb. The stop codonin qERb is indicated by an asterisk. The consensus sequence for MAP kinase–mediated serinephosphorylation (PKSP) is underlined once and the AF-2 domain is underlined twice. The primersthat were used for the RT-PCR studies analyzing the tissue distribution of the transcripts areunderlined; the oligoprobes used forin situ hybridization are shown in bold. Genbank accessionnumber is AF045149.
Estrogen Receptor-b in Quail 333

Figure 2
334 Foidart et al.

caudal brain levels that were studied (Fig. 7). Thisstrongly suggests that the dense clusters of hybridiza-tion signal that are described above specifically cor-respond to ERb transcripts. This idea is independentlysupported by the fact that oligoprobes targeting threedifferent parts of the ERb mRNA provided the samevery discrete pattern of neuroanatomical distribution.
DISCUSSION
In the present study, we have identified for the firsttime in a bird species the sequence of the ERb cDNAthat contains a complete open reading frame. The59-RACE study revealed at the 59 end of the moleculethe presence an ATG codon, flanked by nucleotidesthat fit the Kozak consensus sequence. This ATGcorresponds to the beginning of the sequence (firstmethionine residue) that has been reported for rats,mice, and humans (Kuiper et al., 1996; Tremblay etal., 1997; Mosselman et al., 1996; Enmark et al.,1997) and therefore determines a protein of 472amino acids similar to the 485 residues of the rat ERb.However, a more recent report has extended thelength of the ERb protein at the N-terminus to 530residues in human (Ogawa et al., 1998). It is thereforepossible that the quail ERb sequence described hereextends further in the N terminal direction.
The deduced amino acid sequence of the quailERb exhibits a higher homology with mammalianERb, and with the presumptive ERb from eel thanwith mammalian and avian ERa. This homology withmammalian ERb is particularly high in the DNA-binding domain and in the ligand-binding domain, thetwo parts of the molecule that are presumably subjectto the most stringent selection pressure to maintaintheir structure. However, as shown previously for therat and human ERb, the DNA-binding domain of thequail ERb displays a high degree of homology withthe corresponding ERa. Therefore, it appears that ingeneral, this part of the molecule is very well con-served between the two forms of this receptor, but thatthe ERb and ERa differ markedly in the ligand-binding domain and in the other parts of the molecule.These characteristics, together with several features ofthe tissue and neuroanatomical distribution discussed
below, clearly indicate that the molecule we havecloned is an ERb.
Northern blot analysis of mRNAs extracted fromquail brain and kidney revealed the presence of sev-eral transcripts in these tissues. At least two tran-scripts are apparently expressed in the brain, whilethree transcripts could be clearly observed in thekidney that was studied here, because such a veryhigh expression of ERb was observed in our first PCRstudy (Lakaye et al., 1998) . Interestingly, three tran-scripts of similar size were also found in the mouseovary (Tremblay et al., 1997) and in different humantissues (Mosselman et al., 1996), indicating that eitheralternate splicing occurs in the maturation of the RNA(Petersen et al., 1998) or the ERb gene is character-ized by multiple promoters as shown for the ERagene (Grandien et al., 1997).
The tissue distribution of the ERb transcript iden-tified here by RT-PCR is reminiscent of the distribu-tion previously reported in rat by Kuiper et al. (1997)
Figure 3 Northern blot analysis of quail ERb transcriptsin the quail brain (left) and kidney (right). Molecular sizemarkers are shown on the left.
Figure 2 Comparison of the amino acid sequences of the quail ERb (qERb), rat ERb (rERb),human ERb (hERb), eel estrogen receptor (eER), and chicken ERa (cERa). Dots indicate identicalamino acid residues; hyphens indicate gaps or missing residues. The boxes indicate the DNA-binding (DNB) and ligand-binding (LBD) domains (according to Enmark et al., 1997).
Estrogen Receptor-b in Quail 335

using the same technique, and by other authors de-tecting ERb mRNA by in situ hybridization (Enmarket al., 1997) or the corresponding protein by immu-nocytochemistry (Saunders et al., 1997). In the brain,the high expression found in the telencephalon, ex-cluding the nucleus taeniae (known to contain ERaand therefore excluded from the dissection) and thepresence of the ERb at a weaker level in the cerebel-lum confirm that the new sequence do not correspondto an ERa because this type of receptor is not presentin these areas, as demonstrated by our previous im-munocytochemical studies in quail (Balthazart et al.,1989) and by similar studies in a range of otherspecies (Gahr et al., 1993; Gahr and Metzdorf, 1997).
In situhybridization experiments identified a denseexpression of ERb mRNA in a large number of hy-pothalamic and limbic nuclei, including the medialpreoptic nucleus, nucleus striae terminalis, nucleustaeniae, and ventromedial and tuberal part of the hy-pothalamus. All these brain areas are known to playimportant roles in the control of reproduction, includ-ing the activation of appetitive and consummatorycomponents of male sexual behavior as well as thefeedback regulation of gonadotropin hormone–releas-ing hormone (Balthazart and Ball, 1997; Thompson etal., 1998; Balthazart et al., 1998; Davies, 1980; Da-vies and Follett, 1975). This general pattern of neu-roanatomical distribution of the ERb mRNA observedhere in quail closely corresponds to results of otherinsitu hybridization studies carried out in the rat(Shughrue et al., 1996, 1997b) and ERa knockoutmouse brain (Shughrue et al., 1997a). It also largelyoverlaps with the distribution of the ERb protein asidentified by immunocytochemical studies (Li et al.,1997; Saunders et al., 1997, 1998). The presence of anintense hybridization signal in the quail nucleustaeniae, a part of the avian archistriatum, providesfurther neurochemical criterion supporting the homol-ogy of this structure with parts of the mammalian
amygdala (Thompson et al., 1998). The presence ofERb has been reported in the medial and corticalnuclei of the amygdala of rats (Shughrue et al.,1997b). The dense hybridization signal for ERb ob-served throughout the rostral to caudal extent of thenucleus preopticus medialis (Panzica et al., 1991) andof the nucleus striae terminalis, as recently redefinedby Aste et al. (1998), supports the contention thatthese structures are homologous with the rat medialpreoptic nucleus and nucleus striae terminalis, respec-tively.
A relatively high density of ERb mRNA is alsoobserved in the paraventricular nucleus of the hypo-thalamus (PVN) that is known to be devoid of ERa.This nucleus contains neuropeptide systems that areknown to be regulated by estrogen, in particular,vasotocin and oxytocin. These ERb receptors maytherefore mediate the control of the synthesis of theseneuropeptides by estrogens. In rats, recent studieshave demonstrated that the ERb mRNA and ERb-likeimmunoreactivity are colocalized with oxytocinand/or vasopressin (mammalian homologue of theavian vasotocin) in the PVN, suggesting that estro-gens can regulate these two systems through a directgenomic ERb-mediated mechanism (Simonian andHerbison, 1997; Harbovszky et al., 1998; Alves et al.,1998).
A high density of ERb transcripts is present inparts of the nucleus intercollicularis, a brain area thathas been implicated in the control of vocal behavior insongbirds (Nottebohm et al., 1976, 1982) as well as ingallinaceous species including quail (Potash, 1970;Armitage and Seller, 1981; Seller, 1981; Yasaki et al.,1993). The density of this hybridization signal isespecially high in two parts of the nucleus: a rostro-ventral diagonal band that seems to correspond to asubregion that contains high density of muscariniccholinergic receptors (Ball et al., 1990; Ball and Bal-thazart, 1990) and a more caudal dorso-medial area
Figure 4 Tissue distribution of quail ERb mRNA and of glyceraldehyde-3-phosphate dehydro-genase (GAPDH) mRNA (a housekeeping gene) in male quail as determined by RT-PCR on totalRNA extracts. Tel5 telencephalon; Cb5 cerebellum; Di5 diencephalon; OL5 optic lobes; BS5 brain stem; SpC5 spinal cord; Pit5 pituitary gland; Liv5 liver; Pec5 pectoral muscle; Gas5 gastrocnemius muscle; Kdn5 kidney; Ad 5 adrenal gland; Hrt5 heart; Lng5 lung; Tst5 testis; Epi5 epididymis; Ov5 ovary; Ovd5 oviduct.
336 Foidart et al.
that appears to overlap with the area previously iden-tified by the presence of high densities ofa2-adren-ergic and of ERa receptors (Ball and Balthazart,1990; Balthazart et al., 1989). In songbirds, a similarsubregion of the intercollicular nucleus, called thedorso-medial nucleus (DM), is connected to the net-
work of nuclei that controls the acquisition and pro-duction of song (Gurney, 1981; Nottebohm et al.,1982). In particular, the DM part of the intercollicularnucleus of songbirds sends a dense projection to thetracheosynringeal division of the motor nucleus of the12th nerve, and is thus a key relay in the transmission
Figure 5 Coronal section through the quail brain illustrating the distribution of the ERb mRNA.The relative intensity of the hybridization signal (dense vs. light) is indicated by the density of theshading (black vs. dotted). (A–H) Presented in a rostral to caudal order. Ac5 nucleus accumbens;BST 5 nucleus striae terminalis; CA5 commissura anterior; CPA5 commissura pallii; DMA5 nucleus dorsomedialis anterior thalami; DSM5 decussatio supraoptica; FPL5 fasciculusprosencephali lateralis; GCt5 substantia grisea centralis; ICo5 nucleus intercollicularis; IH5 nucleus inferioris hypothalami; LHy5 nucleus lateralis hypothalami; LPO5 lobus parolfacto-rius; MLd 5 nucleus mesencephalicus lateralis pars dorsalis; MM5 nucleus mamillaris medialis;OM 5 tractus occipitomesencephalicus; PA5 paleostriatum augmentatum; POM5 nucleuspreopticus medialis; PVO5 organum paraventriculare; PVN5 nucleus paraventricularis; PVT5 paleostriatum ventrale; SL5 nucleus septali lateralis; SM5 nucleus septali medialis; Tn5 nucleus taeniae; TSM5 tractus septomesencephalicus; Tu5 nucleus tuberis; VMN5 nucleusventromedialis hypothalami.
Estrogen Receptor-b in Quail 337
of motor outputs from the telencephalon to the syrinx.In quail, less is known about the connectivity of theintercollicular nucleus, but, as mentioned above, stud-
ies have shown that ERa anda2-adrenergic receptorscoexist in a dorsomedial subregion of the ICo thatcould be equivalent to the DM of songbirds (Ball et
Figure 6 Autoradiograms of coronal sections in the quail brain illustrating the distribution of theERb mRNA visualized byin situ hybridization with the combination of probes q2 and q3 labeledwith 33P (see text). (A–H) Presented in a rostral to caudal order.
338 Foidart et al.
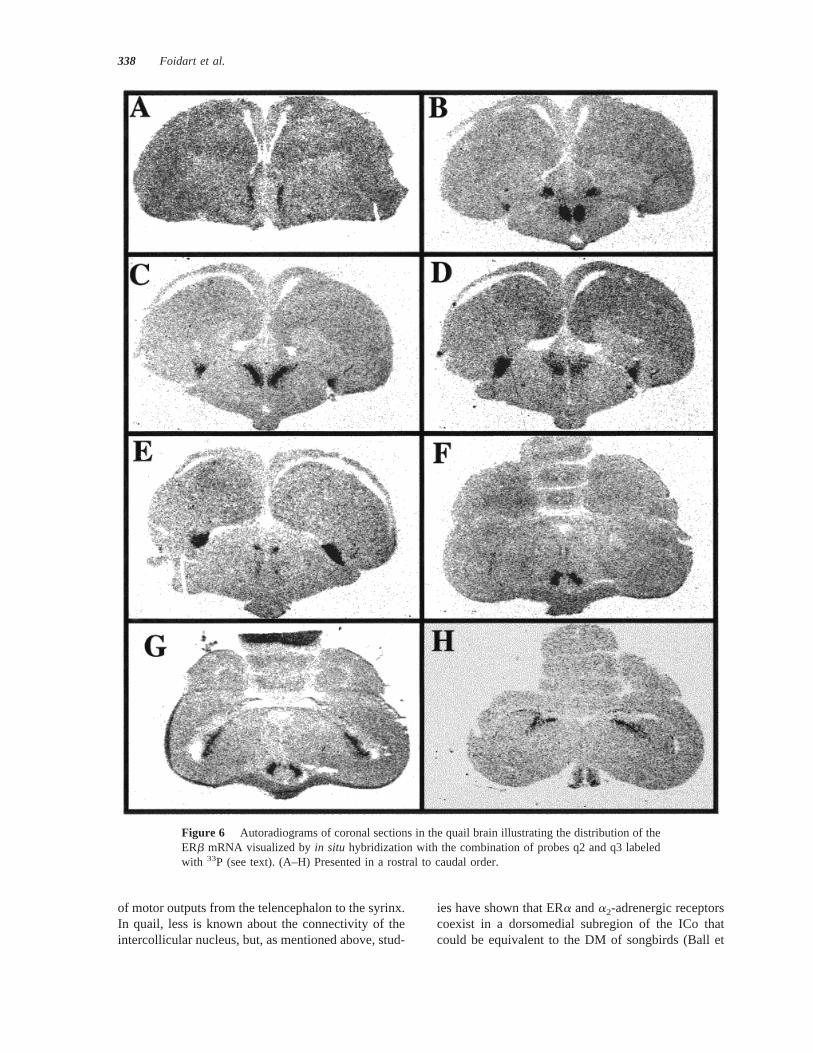
al., 1989). The present study therefore confirms theexistence of a neurochemical specialization in thedorso-medial part of the intercollicular nucleus of thequail. This nucleus also seems to be play a significantrole during copulation: Large numbers of cells areactivated in this nucleus in male quail that have cop-ulated, as demonstrated by the increased expressed ofthe immediate-early genes, c-fosand Zenk (Meddle etal., 1997; Ball et al., 1997). Copulation is activated bya synergistic action of androgens and estrogens, witha part of these estrogens being produced locally byaromatization of testosterone (Balthazart and Foidart,1993; Panzica et al., 1996). However, very little aro-matase activity and few or no aromatase-immunore-active cells are present in the nucleus intercollicularis,so that any estrogen acting on this nucleus to influencethe production of reproductive behaviors is probablyof peripheral origin.
In general, these data demonstrate that in Japanesequail, ERb is present in many brain areas that areknown to express aromatase, the enzyme that cata-lyzes the production of estrogen in the brain. It hasbeen firmly established that testosterone must betransformed into an estrogen by aromatization withinthe medial preoptic nucleus to activate male sexual
behavior in this species (Balthazart, 1997; Balthazartet al., 1997; Balthazart and Surlemont, 1990). Be-cause many neurons in the medial preoptic nucleuscontain ERa, it was usually assumed that estrogenproduced locally by aromatization of testosterone actsin the cells where they were produced in an intracrinemanner by directly interacting with ERa (see Baltha-zart and Ball, 1998b, for further discussion). Surpris-ingly however, we have demonstrated by double-labelimmunocytochemistry that only very few aromatase-containing cells of the medial preoptic nucleus simul-taneously coexpress ERa (Balthazart et al., 1991).This finding was more recently extended to a numberof mammalian species such as rats, mice, and muskshrews (Tsuruo et al., 1995, 1996; Veney and Riss-man, 1998), indicating that this absence of colocal-ization between aromatase and ERa may be a generalfeature in vertebrates. This raised a number of ques-tions concerning the mechanisms underlying the con-trol of aromatase synthesis and the activation of re-productive behavior, two responses that are known tobe estrogen dependent at the cellular level. We havepostulated that the estrogenic control of these re-sponses is not mediated by a classical genomic actionof the steroid, but involves transynaptic controls that
Figure 7 Autoradiograms illustrating the specificity of the hybridization signal obtained with thecombination of probes q2 and q3 labeled with33P. (A–C) Incubated with labeled probes alone.(B–D) Incubated with the same probes in presence of a 100-fold excess of unlabeled probes. Allsignal disappears in sections that were incubated with the excess of unlabeled probes. The sameresult is observed when sections are incubated with RNase before hybridization.
Estrogen Receptor-b in Quail 339

would be originating in another estrogen-dependentneuron (Balthazart and Ball, 1998b). The presentstudy demonstrating the presence of ERb mRNA inbrain areas that contain aromatase cells not colocal-ized with ERa such as the medial preoptic nucleusand the nucleus striae terminalis obviously reopensthis question. Recent studies have revealed that theERb mRNA is coexpressed with ERa in many brainareas, and that the two receptors are often but notalways colocalized in the same cells (Shughrue andScrimo, 1998). The possibility therefore exists thatERb mRNA may be present in cells that expressaromatase but do not contain ERa. Providing thatthese mRNA are translated into functional proteins,the aromatase synthesis and sexual behavior could beregulated by estrogens produced in the same neuronvia a classical intracellular estrogen receptor acting astranscription factor after binding with its ligand. Aro-matase may be selectively colocalized with ERb re-ceptors rather than the ERa type. The functional sig-nificance of such a pattern of colocalization isintriguing. This scenario needs to be tested experi-mentally, and a first step in this process would be todetermine by doublein situ hybridization or by com-bining in situ hybridization for ERb with immunocy-tochemistry for aromatase whether ERb is expressedin aromatase-containing neurons.
The authors thank Dr. Daniel Bernard for useful discus-sions about the cloning strategies employed in this study.This work was supported by a grant from the NIMH (R01MH50388) to GFB and JB, by grants from the BelgianFRFC (No. 9.4565.96F) and the University of Lie`ge (Cred-its speciaux) to JB, and by grants from the Belgian FNRS(1.5.081.98) and University of Lie`ge (Credits speciaux) toTG. The collaboration between JB and GFB was supportedby a NATO collaborative research grant (CRG 973000). AFis a Research Associate of the Belgian FNRS
REFERENCES
Alves SE, Lopez V, McEwen BS, Weiland NG. 1998.Differential colocalization of estrogen receptorb (ERb)with oxytocin and vasopressin in the paraventricular andsupraoptic nuclei of the female rat brain: An immunocy-tochemical study. Proc Natl Acad Sci USA 95:3281–3286.
Armitage SE, Seller TJ. 1981. Midbrain regions involved incall production of Japanese quail. Experientia 37:847–848.
Aste N, Balthazart J, Absil P, Grossmann R, Mulhbauer E,Viglietti-Panzica C, Panzica GC. 1998. Anatomical andneurochemical definition of the nucleus of the stria ter-
minalis in Japanese quail (Coturnix japonica). J CompNeurol 396:141–157.
Ball GF, Balthazart J. 1990. Steroid modulation of musca-rinic cholinergic anda2-adrenergic receptor density inthe nucleus intercollicularis of the Japanese quail. EurJ Neurosci 2:828–835.
Ball GF, Foidart A, Balthazart J. 1989. A dorsomedialsubdivision within the nucleus intercollicularis identifiedin the Japanese quail (Coturnix coturnix japonica) bymeans ofa2-adrenergic receptor autoradiography andestrogen receptor immunohistochemistry. Cell TissueRes 257:123–128.
Ball GF, Nock B, Wingfield JC, McEwen BS, Balthazart J.1990. Muscarinic cholinergic receptors in the songbirdand quail brain: a quantitative autoradiographic study.J Comp Neurol 298:431–442.
Ball GF, Tlemcani O, Balthazart J. 1997. Induction of theZenk protein after sexual interactions in male Japanesequail. Neuroreport 8: 2965–2970.
Balthazart J. 1997. Steroid control and sexual differentiationof brain aromatase. J Steroid Biochem Mol Biol 61:323–339.
Balthazart J, Absil P, Gerard M, Appeltants D, Ball GF.1998. Appetitive and consummatory male sexual behav-ior in Japanese quail are differentially regulated by sub-regions of the preoptic medial nucleus. J Neurosci 18:6512–6527.
Balthazart J, Ball GF. 1993. Neurochemical differences intwo steroid-sensitive areas mediating reproductive behav-iors. Adv Comp Environ Physiol 15:133–161.
Balthazart J, Ball GF. 1997. Neuroendocrine regulation ofappetitive and consummatory aspects of male sexual be-havior in Japanese quail. In: Etches R, Harvey S, editors.Perspectives in avian endocrinology. Bristol, UK: Societyfor Endocrinology. p 241–255.
Balthazart J, Ball GF. 1998a. The Japanese quail as a modelsystem for the investigation of steroid-catecholamine in-teractions mediating appetitive and consummatory as-pects of male sexual behavior. Ann Rev Sex Res. 9:96–176.
Balthazart J, Ball GF. 1998b. New insights into the regula-tion and function of brain estrogen synthase (aromatase).Trends Neurosci 21:243–249.
Balthazart J, Castagna C, Ball GF. 1997. Aromatase inhi-bition blocks the activation and sexual differentiation ofappetitive male sexual behavior in Japanese quail. BehavNeurosci 111:381–397.
Balthazart J, Foidart A. 1993. Brain aromatase and thecontrol of male sexual behavior. J Steroid Biochem MolBiol 44:521–540.
Bathazart J, Foidart A, Surlemont C, Harada N. 1991.Neuroanatomical specificity in the co-localization of aro-matase and estrogen receptors. J Neurobiol 22:143–157.
Balthazart J, Gahr M, Surlemont C. 1989. Distribution ofestrogen receptors in the brain of the Japanese quail: animmunocytochemical study. Brain Res 501:205–214.
Balthazart J, Surlemont C. 1990. Androgen and estrogen
340 Foidart et al.

action in the preoptic area and activation of copulatorybehavior in quail. Physiol Behav 48:599–609.
Cooke PS, Buchanan DL, Lubahn DB, Cunha GR. 1998Mechanism of estrogen action: lessons from the estrogenreceptor-a knockout mouse. Biol Reprod 59:470–475.
Davies DT. 1980. The neuroendocrine control of gonado-trophin release in the Japanese quail. III. The role of thetuberal and anterior hypothalamus in the control of ovar-ian development and ovulation. Proc R Soc Lond B206:424–437.
Davies DT, Follett BK. 1975. The neuroendocrine controlof gonadotrophin release in the Japanese quail. I. The roleof the tuberal hypothalamus. Proc R Soc Lond B 191:285–301.
Enmark E, Pelto-Huikko M, Grandien K, Lagercrantz S,Lagercrantz J, Fried G, Nordenskjold M, Gustafsson J-Å.1997. Human estrogen receptorb-gene structure, chro-mosomal localization, and expression pattern. J Clin En-docrinol Metab 82:4258–4265.
Frohman MA, Marin GR. 1989. Rapid amplification ofcDNA ends using nested primers. Technique1:37–72.
Gahr M, Guttinger H-R, Kroodsma DE. 1993. Estrogenreceptors in the avian brain: survey reveals general dis-tribution and forebrain areas unique to songbirds. J CompNeurol 327:112–122.
Gahr M, Metzdorf R. 1997. Distribution and dynamics inthe expression of androgen and estrogen receptors invocal control systems of songbirds. Brain Res Bull 44:509–517.
Grandien KAJ, Berkenstam A, Gustafsson J-A. 1997. Theestrogen receptor gene: promoter organization and ex-pression. Int J Biochem Cell Biol 29:1343–1396.
Green S, Walter P, Kumar V, Krust A, Bornert J-M, ArgosP, Chambon P. 1986. Human estrogen receptor cDNA:sequence expression and homology to v-erbA. Nature320:134–139.
Gurney ME. 1981. Hormonal control of cell form andnumber in the zebra finch song system. J Neurosci1:658–673.
Harbovszky E, Kallo I, Hajsan T, Shughrue PJ, Merch-enthaler I, Liposits Z. 1998. Expression of estrogen re-ceptor-b messenger ribonucleic acid in oxytocin and va-sotocin neurons of the rat supraoptic and paraventricularnuclei. Endocrinology 139:2600–2604.
Jacobs EC, Arnold AP, Campagnoni AT. 1996. Zebra finchestrogen receptor cDNA: cloning and mRNA expression.J Steroid Biochem Mol Biol 59:135–145.
Joels M. 1997. Steroid hormones and excitability in themammalian brain. Front Neuroendocrinol 18:2–48.
Korke S, Sakai M, Muramatsu M. 1987. Molecular cloningand characterization of rat estrogen receptor cDNA. Nu-cleic Acids Res 15:2499–2513.
Kozak M. 1986. Point mutations define a sequence flankingthe AUG initiator codon that modulates translation byeukaryotic ribosomes. Cell 44:283–292.
Krust A, Green S, Argos P, Kumar V, Walter P, Bornert JM,Chambon P. 1986. The chicken oestrogen receptor se-
quence: homology with v-erbA and the human oestrogenand glucocorticoid receptors. EMBO J 5:891–897.
Kuiper GGJM, Carlsson B, Grandien K, Enmark E, Hagg-blad J, Nilsson S, Gustafsson JÅ. 1997. Comparison ofthe ligand binding specificity and transcript tissue distri-bution of estrogen receptorsa and b. Endocrinology138:863–870.
Kuiper GGJM, Enmark E, Pelto-Huikko M, Nilsson S,Gustafsson JÅ. 1996. Cloning of a novel estrogen recep-tor expressed in rat prostate and ovary. Proc Natl AcadSci USA 93:5925–5930.
Kuiper GGJM, Gustafsson J-Å. 1997. The novel estrogenreceptor-b subtype: potential role in the cell- and promot-er-specific actions of estrogens and anti-estrogens. FEBSLett 410:87–90.
Kuiper GGJM, Shughrue PJ, Merchenthaler I, GustafssonJ-Å. 1998. The estrogen receptorb subtype: a novelmediator of estrogen action in neuroendocrine systems.Front Neuroendocrinol 19:253–286.
Lakaye B, Foidart A, Grisar T, Balthazart J. 1998. Partialcloning and distribution of estrogen receptor beta in theavian brain. Neuroreport 9:2743–2748.
Li X, Schwartz PE, Rissman EF. 1997. Distribution ofestrogen receptor-b-like immunoreactivity in rat fore-brain. Neuroendocrinology 66:63–67.
Lubahn DB, Moyer JS, Golding TS, Couse JF, Korach KS,Smithies O. 1993. Alteration of reproductive function butnot prenatal sexual development after insertional disrup-tion of the mouse estrogen receptor gene. Proc Natl AcadSci USA 90:11162–11166.
McEwen BS, Alves SE, Bulloch K, Weiland NG. 1997.Ovarian steroids and the brain. Neurology 48:S8–S15.
Meddle SL, King VM, Follett BK, Wingfield JC, Ramenof-sky M, Foidart A, Balthazart J. 1997. Copulation acti-vates Fos-like immunoreactivity in the male quail fore-brain. Behav Brain Res 85:143–159.
Mermelstein PG, Becker JB, Surmeier DJ. 1996. Estradiolreduces calcium currents in rat neostriatal neurons via amembrane receptor. J Neurosci 16:595–604.
Moffat CA, Rissman EF, Shupnik MA, Blaustein JD. 1998.Induction of progestin receptors by estradiol in the fore-brain of estrogen receptor-alpha gene-disrupted mice.J Neurosci 18:9556–9563.
Mosselman S, Polman J, Dijkema R. 1996. ER beta: iden-tification and characterization of a novel human estrogenreceptor. FEBS Lett 392:49–53.
Nottebohm F, Kelley DB, Paton JA. 1982. Connections ofvocal control nuclei in the canary telencephalon. J CompNeurol 207:344–357.
Nottebohm F, Stokes TM, Leonard CM. 1976. Centralcontrol of song in the canary,Serinus canarius. J CompNeurol 165:457–486.
Ogawa S, Inoue S, Watanabe T, Hirol H, Orimo A, Hosol T,Ouchi Y, Muramatsu M. 1998. The complete primarystructure of human estrogen receptor beta (hERbeta) andits heterodimerization with ER alpha in vivo and in vitro.Biochem Biophys Res Commun 243:122–126.
Panzica GC, Viglietti-Panzica C, Balthazart J. 1996. The
Estrogen Receptor-b in Quail 341

sexually dimorphic medial preoptic nucleus of quail: akey brain area mediating steroid action on male sexualbehavior. Front Neuroendocrinol 17:51–125.
Panzica GC, Viglietti-Panzica C, Sanchez F, Sante P, Bal-thazart J. 1991. Effects of testosterone on a selectedneuronal population within the preoptic sexually dimor-phic nucleus of the Japanese quail. J Comp Neurol 303:443–456.
Petersen DN, Tkalcevic GT, Koza-Taylor PH, Turi TG,Brown TA. 1998. Identification of estrogen receptorb2,a functional variant of estrogen receptorb in normal rattissues. Endocrinology 139:1082–1092.
Potash LM. 1970. Vocalizations elicited by electrical brainstimulation inCoturnix coturnix japonica. Behaviour 31:149–167.
Ramirez VD, Zheng JB, Siddique KM. 1996. Membranereceptors for estrogen, progesterone, and testosterone inthe rat brain: fantasy or reality. Cell Mol Neurobiol16:175–198.
Riters LV, Absil P, Balthazart J. 1998. Effects of braintestosterone implants on appetitive and consummatorycomponents of male sexual behavior in Japanese quail.Brain Res Bull 47:69–79.
Saunders PTK, Fisher JS, Sharpe RM, Millar MR. 1998.Expression of oestrogen receptor beta (ERb) occurs inmultiple cell types, including some germ cells, in the rattestis. J Endocrinol 156:R13–R17.
Saunders PTK, Maguire SM, Gaughan J, Millar MR. 1997.Expression of oestrogen receptor beta (ERb) in multiplerat tissues visualised by immunohistochemistry. J Endo-crinol 154:R13–R16.
Schumacher M, Balthazart J. 1987. Neuroanatomical distri-bution of testosterone metabolizing enzymes in the Jap-anese quail. Brain Res 422:137–148.
Seielstad DA, Carlson KE, Kushner PJ, Greene GL, Kat-zenellenbogen JA. 1995. Analysis of the structural coreof the human estrogen receptor ligand binding domain byselective proteolysis/mass spectrometric analysis. Bio-chemistry 34:12605–12615.
Seller TJ. 1981. Midbrain vocalization centers in birds.Trends Neurosci 4:301–303.
Shughrue P, Scrimo P, Lane M, Askew R, Merchenthaler I.1997a. The distribution of estrogen receptor-b mRNA inforebrain regions of the estrogen receptor-a knockoutmouse. Endocrinology 138:5649–5652.
Shughrue PJ. 1998. Estrogen action in the estrogen receptoralpha-knockout mouse: is this due to ER-beta? Mol Psy-chiatry 3:299–302.
Shughrue PJ, Bushnell CD, Dorsa DM. 1992. Estrogen recep-tor messenger ribonucleic acid in female rat brain during theestrous cycle: a comparison with ovariectomized femalesand intact males. Endocrinology 131:381–388.
Shughrue PJ, Komm B, Merchenthaler I. 1996. The distri-bution of estrogen receptor-b mRNA in the rat hypothal-amus. Steroids 61:678–681.
Shughrue PJ, Lane MV, Merchanthaler I. 1997b. Compar-ative distribution of estrogen receptor-a and -b mRNA inthe rat central nervous system. J Comp Neurol 388:507–525.
Shughrue PJ, Lubahn D, Negro-Vilar A, Korach KS, Mer-chenthaler I. 1997c. Responses in the brain of estrogenreceptor a-disrupted mice. Proc Natl Acad Sci USA94:11008–11012.
Shughrue PJ, Scrimo PJ. 1998. Evidence for the colocaliza-tion of estrogen receptor-beta and estrogen receptor-alphaimmunoreactivity in neurons of the rat forebrain. Endo-crinology 139:5267–5270.
Simerly RB, Chang C, Muramatsu M, Swanson LW. 1990.Distribution of androgen and estrogen receptor mRNA-containing cells in the rat brain: an in situ hybridizationstudy. J Comp Neurol 294:76–95.
Simonian SX, Herbison AE. 1997. Differential expressionof estrogen receptora and b immunoreactivity by oxy-tocin neurons of rat paraventricular nucleus. J Neuroen-docrinol 9:803–806.
Thompson RR, Goodson JL, Ruscio MG, Adkins-Regan E.1998. Role of the archistriatal nucleus taeniae in thesexual behavior of male Japanese quail (Coturnix ja-ponica): a comparison of function with the medial nu-cleus of the amygdala in mammals. Brain Behav Evol51:215–229.
Todo T, Adachi S, Yamauchi K. 1996. Molecular cloningand characterization of Japanese eel estrogen receptorcDNA. Mol Cell Endocrinol 119:37–45.
Tremblay GB, Tremblay A, Copeland NG, Gilbert DJ,Jenkins NA, Labrie F, Giguere V. 1997. Cloning, chro-mosomal localization, and functional analysis of the mu-rine estrogen receptor beta. Mol Endocrinol 11:353–365.
Tsang SS, Yin X, Guzzo-Arkuran C, Jones VS, Davison AJ.1993. Loss of resolution in gel electrophoresis of RNA: aproblem associated with the presence of formaldehydegradients. Biotechniques 14:380–381.
Tsuruo Y, Ishimura K, Hayashi S, Osawa Y. 1996. Immu-nohistochemical localization of estrogen receptors withinaromatase-immunoreactive neurons in the fetal and neo-natal rat brain. Anat Embryol Berl 193:115–121.
Tsuruo Y, Ishimura K, Osawa Y. 1995. Presence of estro-gen receptors in aromatase-immunoreactive neurons inthe mouse brain. Neurosci Lett 195:49–52.
Veney SL, Rissman EF. 1998. Co-localization of estrogenreceptor and aromatase enzyme immunoreactivities inadult musk shrew brain. Horm Behav 33:151–162.
Yasaki Y, Matsushima T, Aoki K. 1995. The role of themesencephalic call region in the Japanese quail chick. In:Burrows M, Matheson T, Newland PL, Schuppe H, edi-tors. Nervous systems and behaviour: Proceedings of the4th International Congress of Neuroethology. Stuttgart:Georg Thieme Verlag. p 319.
342 Foidart et al.





























