Embed Size (px)
Citation preview

Samantha M. Hens& Lyle W.KonigsbergDepartment of Anthropology,252 South Stadium Hall,University of Tennessee,Knoxville, TN 37996-0720.E-mail: [email protected];[email protected]
William L. JungersAnatomical Sciences School ofMedicine, SUNY at StonyBrook, Stony Brook, NY11794-8081. E-mail:[email protected]
Received 17 June 1997Revision received17 November 1997 andaccepted 24 November1997
Keywords: African ape, bodysize, stature estimation,regression techniques.
Estimation of African ape body lengthfrom femur length
The estimation from long bone lengths of stature in humans or bodysize in apes has a deep history in physical anthropology. To date, wecan enumerate at least five different statistical methods for makingsuch estimations. These methods are: (1) the regression of bodylength on long bone length (inverse calibration), (2) regression oflong bone length on body length followed by solving for body length(classical calibration), (3) major axis regression of body length onlong bone length, (4) reduced major axis regression of body length onlong bone length, and (5) use of a long bone/body length ratio. Weexamine some of the statistical properties of these estimators using alarge sample of humans (n=2053) to derive the estimators, andapplying them to smaller samples of Pan troglodytes (n=42), Panpaniscus (n=8), and Gorilla gorilla (n=35).Based on the root mean-squared error (RMSE), the reduced major
axis is the preferred estimator for body length in the combined Pansample. However, inverse calibration is the best estimator for bodylength in gorillas based on the RMSE. Many estimators grosslyunderestimate body length in the apes. Differences in allometriesbetween humans and great apes are obvious, but it is important toshow the assumptions necessary in estimating body size from fossilremains, especially when isolated long bones are recovered and theglobal allometry is consequently unknown.
? 1998 Academic Press Limited
Journal of Human Evolution (1998) 34, 401–411
Introduction
The fields of biology and anthropologyrecognize body size as one of the mostsignificant parameters affecting behavioraland ecological adaptations in mammals(Schmidt-Nielson, 1977, 1984). As ameasurable characteristic, even in fragmen-tary fossil remains, body size estimates areused to infer diet, locomotion, adaptivezones, and social structure in the primatefossil record (Fleagle, 1978; Martin, 1990).Most workers utilize the body massmeasure, which is considered practical anduniversally applicable for comparing animalsto each other and to their environments(Smith, 1993). Another common measure
0047–2484/98/040401+11 $25.00/0/hu970209
of body size is stature in humans or bodylength in non-human primates. As pointedout by Feldesman & Lundy (1988) theseestimates are also important components ofbody size. Stature estimates provide animpression of size for an individual or apopulation. Therefore, it is essential thatpredictions of stature/body length fromskeletal measurements be as accurate aspossible.The determination of living stature
from skeletal remains traces back to thenineteenth century works of Manouvrier(1893), Dwight (1894), and Pearson(1899). However, it was not until the 1950swith the work of Trotter & Gleser (1952,1958) that stature estimation was fullyexplored. Their study of the repatriatedskeletal remains of World War II andKorean War dead led to the development of
? 1998 Academic Press Limited
Address correspondence to: Samantha Hens,Department of Anthropology, 252 South Stadium Hall,University of Tennessee, Knoxville, TN 37996-0720.Tel.: (423) 974-4408; E-mail: [email protected]

402 . . ET AL.
numerous stature formulae. Despite Trotter& Gleser’s warnings about populationspecificity for the regression equations,researchers have applied these equations tonumerous human populations and fossilremains, igniting a debate as to theappropriateness of the equations in certainpopulations. Regression formulae have beenapplied to numerous human groups andfossil hominids, notably australopithecines(Lovejoy & Heiple, 1970; McHenry,1974; Reed & Falk, 1977; Robinson, 1972;White, 1980; Wolpoff, 1973), Homo erectus(Leakey & Walker, 1985; Ruff, 1988), andNeandertals (Trinkaus, 1981, 1983).It was McHenry’s (1974) stature
reconstructions on the remains of 18 Plio-Pleistocene hominids that influenced laterresearch. He used the Trotter & Gleserformulae to gain rough approximations ofbody size on gracile and robust hominidsfrom South Africa. Olivier (1976) chal-lenged McHenry’s conclusions by faultinghis modern reference sample and his choiceof least squares regression techniques.Instead, Olivier suggested using eitherModel II regression techniques or Model Iregressions based on contemporary AfricanPygmies as the reference population. Thechoice of stature estimator was alsocriticized by Geissmann (1986) in his reporton the ability of 45 different regressionequations to correctly determine stature onthe Australopithecus afarensis specimen, A.L.288-1. Using Schmid’s (1986) stature esti-mate based on anatomical reconstruction ofthe entire skeleton, Geissmann concludedthat between inconsistent results andoverestimation, none of the regression for-mulae he tested provided a reliablemethod to predict stature in the gracileaustralopithecines.Reports of non-human limb proportions
for the A.L. 288-1 (Lucy) skeleton(McHenry, 1974; Johanson & Edey, 1981;Jungers, 1982; Jungers & Stern, 1983) ledJungers (1988) to suggest that various
reference populations should be considered(i.e., apes, as well as modern African pygmygroups). Using Model II techniques drawnon a small sample of human pygmies(n=15), Jungers (1988) estimated Lucy’sstature at 105·5 cm, which is remarkablyclose to Schmid’s (1986) reconstruction of105 cm. Feldesman & Lundy (1988) alsorecognized the methodological difficultiesinherent in previous studies of statureestimation. They estimated the stature ofthe Lucy specimen at 106 cm using anadjusted major axis calculation based onequations derived from a South Africanblack sample. Their estimates were superiorto those produced by Trotter and Gleserformulae.An added benefit of the Feldesman &
Lundy (1988) study was the discovery thatLucy’s femur had the same proportion to(reconstructed) stature as in modernhuman groups. On the basis of this finding,Feldesman and coworkers (1989, 1990)developed a femur/stature ratio to predictstature in both modern and fossil hominids.They suggested that the proportion of femurlength to stature is nearly constant bothbetween males and females and acrossmodern human groups. They further sug-gested that applications of the ratio methodto fossil hominids provided more satisfac-tory results than did population specificformulae.The purpose of the present analysis is to
examine five different methods for esti-mating body length that can be found in theliterature. The first is the least squaresregression of body length on long bonelength, which we will refer to as inversecalibration. We call this ‘‘inverse’’ because itis the regression of body size (or ‘‘x’’) onorgan size (or ‘‘y’’), and so it is invertedfrom how one would usually think aboutallometry. The second method uses theregression of long bone length on bodylength, followed by solving for body length.We refer to this as ‘‘classical calibration,’’ if

403
for no other reason then it represents theclassical view of allometry, where local sizeis dependent on global size. Classical cali-bration is the maximum likelihood estimatorwhen all error is presumed to be in longbone length, and none is in body length.Konigsberg & Frankenberg’s (1994) workon age estimators can be adapted to showthat in this situation, the posterior prob-ability for a body size variable is proportionalto the product of the likelihood and anuninformative prior. The likelihood is pro-portional to the probability (from the refer-ence sample) that an individual of a specificbody size will have a long bone measure-ment identical to that observed for the indi-vidual specimen. A potential drawback ofboth of these least squares (Model I) typeregressions is in the dependence and inde-pendence of the variables. Body size mayexplain a long bone measurement, but thelong bone measurement may also explainbody size (Sjøvold, 1990). It is sometimesdifficult to argue direct dependence of y on xor x on y in allometric studies. Thus, manyresearchers seek other methods to accountfor the bilaterality and confusion in errorterms that this bilaterality causes. Webelieve that if body size estimation is the solegoal of a study, then it will generally be mostreasonable to assume that long bone lengthis dependent on body size. As we realize thatthis assumption will meet with some resist-ance from other workers in the field, we alsoconsider symmetrical models where neithervariable is an ‘‘independent’’ variable.Common alternatives to standard least
squares regression are the more symmetricalModel II regressions and comprise the thirdand fourth methods examined in this study:the major axis and reduced major axisregressions of body length on long bonelength. The major axis is defined by thefirst principal component of the variance-covariance matrix. The reduced majoraxis is calculated as the ratio of the standarddeviations, with the sign of the slope indi-
cated by the correlation coefficient(Kermack & Haldane, 1950); equivalently,the reduced major axis slope is simply theordinary least squares slope divided bythe correlation coefficient. These modelsassume that error exists in both the depen-dent and independent variables (Sokal &Rohlf, 1981). With these models, one vari-able just as easily estimates a second as thesecond estimates the first (Sjøvold, 1990).The major axis and reduced major axistechniques are generally preferred whendetermining the central tendency of asample and estimating functional relation-ships (Martin & Barbour, 1989; Rayner,1985).Finally, the fifth method uses a ratio of
body length to long bone length to estimatebody length; this is the femur/stature ratiodeveloped by Feldesman et al. (1989, 1990).The stature to femur ratio can be treated asa regression parameter. This parameter ismultiplied by an actual femur length toobtain an estimate of stature. In this formu-lation, the femur/stature ratio acts like aregression line that passes through the originand the bivariate mean, and it becomes aleast squares line when the femur andstature are isometric. Konigsberg (1997)shows that with this special case of femur/stature isometry, the classical calibrationestimator defaults to the ratio estimator.Consequently, classical calibration and thefemur/stature ratio work equally well incases of isometry, but classical calibrationoutperforms the ratio estimator in othernon-isometric situations. The proof of thisfact is straightforward. If x is stature and y isfemur length, then isometry occurs when thePearson correlation between the two vari-ables is equal to the ratio of the coefficientof variation for stature to the coefficient ofvariation for the femur. Symbolically, wehave:

404 . . ET AL.
Now the regression of y on x can be writtenas:
and substituting equation 1 into equation 2gives:
which shows that the expected femorallength is proportional to stature (as it mustbe in isometry). If âyx is the regressioncoefficient for y on x (femur length onstature), then the classical calibrationestimator can be written as:
x=x+â"1yx (y"y). (4)
In isometry, where the Pearson correlation isequal to the ratio of coefficients of variation(equation 1), â"1
yx is equal to x/y, and as aconsequence equation 4 can be rewritten as:
The estimated stature is then equal to theobserved femur length times the ratio ofaverage stature to average femur length (asin Feldesman and coworkers’ (1989, 1990)femur/stature ratio). This shows that thefemur/stature ratio (equation 5) is thespecial case of classical calibration (equation4) that assumes isometry.We investigate the performance of the five
estimators on a sample of great apes. Morespecifically, we use a large sample of human
femoral and stature measurements todevelop the estimators, and then apply theseto the ape data. This paper consequentlyevaluates the estimators in one of their worstpossible lights—the case where there isextrapolation beyond the characteristic bodysize for the reference sample and whereallometries differ between the reference andthe test material. The comparison in the bestpossible light (i.e., a comparison of esti-mators derived from a reference sample andthen applied back to the same or a verysimilar reference sample) is well understoodand extensively described in the calibrationliterature. In the case where the referencesample forms an appropriate prior for thetarget sample, inverse calibration givesthe minimum variance unbiased estimator.In extrapolation, classical calibration willoutperform inverse calibration as theextrapolation becomes more extreme. Whenallometries differ we will see that there isno a priori basis for choosing an optimalestimator.
Materials and methods
The modern human sample encompasses atotal of 2053 individuals. This sample iscomposed of 159 individuals from theForensic Data Bank maintained at theUniversity of Tennessee, 852 individualsfrom the Terry collection, and 1042 indi-viduals from Trotter’s World War II data.The Terry and Trotter long bone measure-ments were taken by Trotter and areextensively documented in Jantz (1996).The great ape sample comprises mature,wild-shot individuals curated at thePowell-Cotton Museum (England) and theTervuren Museum (Belgium). It isrepresented by 35 Gorilla gorilla, 42 Pantroglodytes, and 8 Pan paniscus, totalling 85specimens. Measurements of body lengthfor the ape sample came from field reports,while the third author took the skeletalmeasurements. All femoral measurements
405
for both the human and non-human primatesamples were maximum length. Stature inthe Terry collection is cadaver stature,where the cadaver is measured in a verticalposition with feet flat on a board and weretaken by Terry (Terry, 1940). The WorldWar II sample statures were taken at thetime of individuals’ induction into thearmed services under standard militaryguidelines (Trotter & Gleser, 1952).Forensic Data Bank statures are listed as‘‘forensic stature’’ and come from varioussources (Moore-Jansen et al., 1994). Boththe Terry and Forensic Data Bank statureswere corrected for by subtracting 2·5 cm tocompensate for age related stature loss andcadaver stature as described in Trotter andGleser (1952). Cousins (1972) illustratesthe method used for the measurement ofbody length on Pan troglodytes and Gorillaspecimens in the field. The apes weremeasured from the crown to the heel withthe knees and hips clearly extended. Thedata on external body length for the pygmychimpanzees come from Coolidge & Shea(1982) where the animals were measuredfrom top of head to heel.The estimators derived from the human
sample are applied to the Gorilla and com-bined Pan samples to predict body length inthese specimens. A general multivariateprogram to fit the five models (inversecalibration, classical calibration, RMA,MA, and ratio) can be obtained fromhttp://konig.la.utk.edu/5reg.html.We use two measures to determine which
of the five equations provides the best esti-mator: the root mean-squared error (RMSE)and bias. The RMSE is the square root ofthe average squared term of the predictedvalues around their true values. A lowRMSE value indicates a better estimator.Bias is calculated as the predicted mean forbody length minus the actual mean for bodylength. The bias provides an idea as to howwell the predictor performs, i.e., how closethe estimates came on average to the actual
value. The RMSE is a measure of efficiencyfor an unbiased estimator. For biased esti-mators the RMSE is less informativebecause it includes terms for both bias andprediction uncertainty.
Results
Table 1 shows the simple statistics forboth the ape and human samples. As thecorrelation, standard deviations, and meansare sufficient statistics for the bivariatenormal distribution, any further parameters(e.g., slopes or their standard errors) areobtainable from Table 1. Table 2 providesthe slope and y-intercept values derivedfrom the human sample for all five regres-sion techniques. Table 3 shows the valuesfor the RMSE and the bias on the combinedPan sample and on the Gorilla sample. Forthe Pan sample, the reduced major axismodel has the lowest RMSE indicating that
Table 1 Means, standard deviations, and corre-lations for the Human sample (n=2053), Gorilla(n=35), P. troglodytes (n=42) and P. paniscussamples (n=8) for femur length and actualstature. Data are given in millimeters
Stature Femur rfem,stat
Mean S.D. Mean S.D.Human 1725·3 85·3 466·4 28·3 0·8684Gorilla 1466·9 138·1 345·5 30·3 0·8873P. trog. 1214·0 63·8 291·7 13·8 0·6742P. paniscus 1153·3 71·2 293·5 14·6 0·8093
Table 2 Regression parameters for inverse cali-bration, classical calibration, major axis (MA),reduced major axis (RMA) and the femur/statureratio models derived from the human sample(n=2053)
y-intercept Slope
Inverse 504·51 2·6175Classical 106·47 3·4709RMA 319·51 3·0141MA 146·84 3·3843Ratio 0·0 3·6992
406 . . ET AL.
the reduced major axis is the optimal indi-cator of body length from femur length inthe chimpanzee sample. The bias for thismodel is minimal. The reduced major axison average underestimates chimpanzee bodylength by only 5 mm. However, the majoraxis, classical, and ratio estimators under-estimate body length by 70 to 124 mm. Theinverse calibration method was the only oneof the five estimators that systematicallyoverestimated the actual values, in this caseby an average of 64 mm.In the Gorilla sample, the inverse cali-
bration of body length on femur length bystandard least squares regression providesthe lowest RMSE. The bias shows that allfive estimators underestimate gorilla bodylength by 58 to 189 mm.The examination of RMSE and bias for
these five estimators in the two genera doesnot provide a clear preference for a bodylength estimator. Least squares regressionby inverse calibration is preferred for thegorilla sample. Reduced major axis isfavored in the Pan sample. The body length/femur length ratio performs poorly in bothsamples due to the scaling differencesbetween the limb proportions of humansand great apes.To visually clarify some of these difficul-
ties, we can inspect graphical representa-
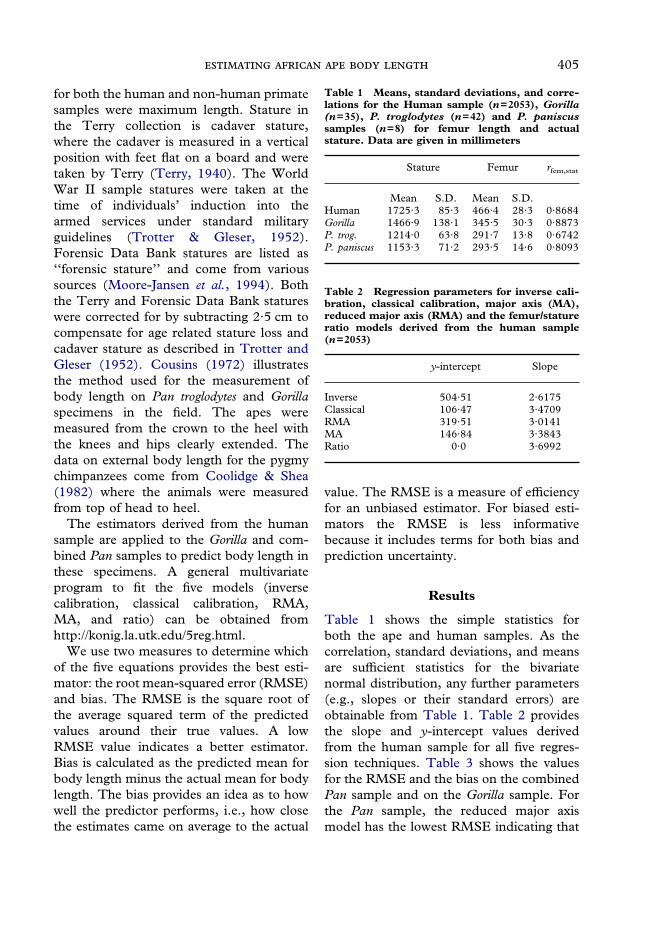
tions of the data. Figure 1 shows the majoraxis (MA), classical, and femur/statureregression lines for humans, along with the95% confidence ellipse (for cases). We haveopted against showing the individual datapoints because the regression lines would bealmost completely obscured. Figure 1 showsthat neither the major axis regression linenor the femur/stature ratio line is muchdifferent from the classical calibration line.For all further graphs we dispatch with themajor axis regression and the femur/statureratio. This leaves the three estimators: clas-sical, reduced major axis (or RMA), andinverse. The relationships of these lines arewell-known (McArdle, 1988; Ricker, 1973;Seim & Sæther, 1983; Sjøvold, 1990),where the classical and inverse calibrationlines cut through tangent points on therectangle enclosing an isodensity, and RMAis on the diagonal of the rectangle. Forspecimens below the human average staturethere will be an ordering of the estimatorswith inverse calibration giving the higheststature estimate from the femur, classical thelowest estimate, and RMA an intermediateestimate.In Figure 2 we make the untenable
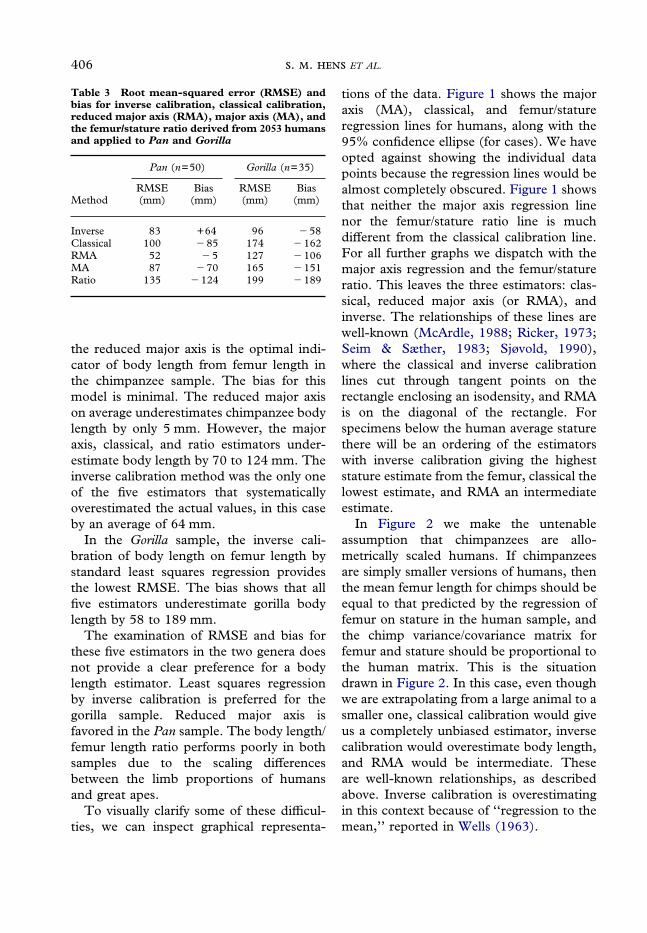
assumption that chimpanzees are allo-metrically scaled humans. If chimpanzeesare simply smaller versions of humans, thenthe mean femur length for chimps should beequal to that predicted by the regression offemur on stature in the human sample, andthe chimp variance/covariance matrix forfemur and stature should be proportional tothe human matrix. This is the situationdrawn in Figure 2. In this case, even thoughwe are extrapolating from a large animal to asmaller one, classical calibration would giveus a completely unbiased estimator, inversecalibration would overestimate body length,and RMA would be intermediate. Theseare well-known relationships, as describedabove. Inverse calibration is overestimatingin this context because of ‘‘regression to themean,’’ reported in Wells (1963).
Table 3 Root mean-squared error (RMSE) andbias for inverse calibration, classical calibration,reduced major axis (RMA), major axis (MA), andthe femur/stature ratio derived from 2053 humansand applied to Pan and Gorilla
Method
Pan (n=50) Gorilla (n=35)
RMSE(mm)
Bias(mm)
RMSE(mm)
Bias(mm)
Inverse 83 +64 96 "58Classical 100 "85 174 "162RMA 52 "5 127 "106MA 87 "70 165 "151Ratio 135 "124 199 "189
407
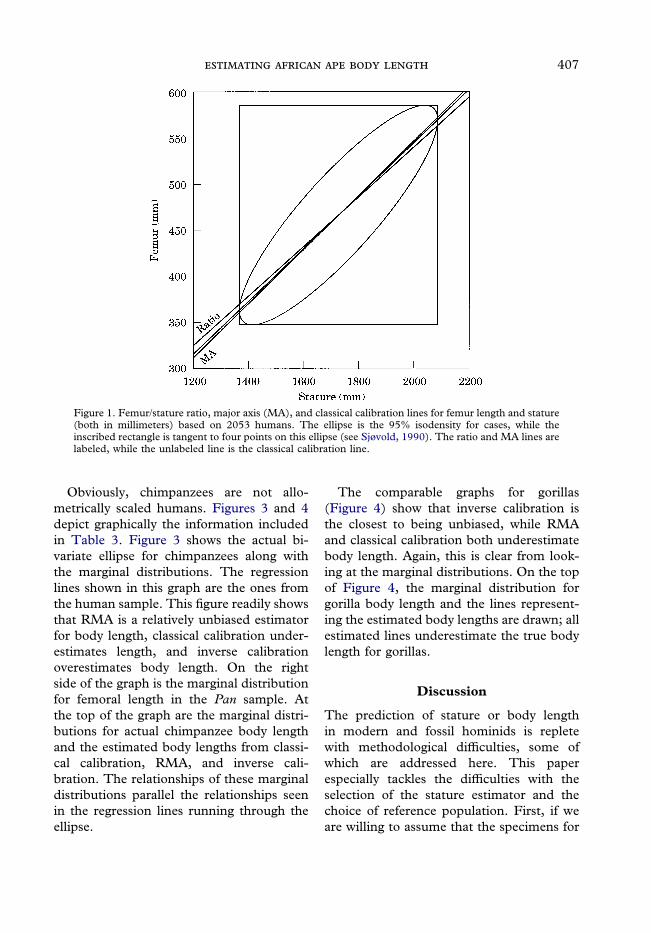
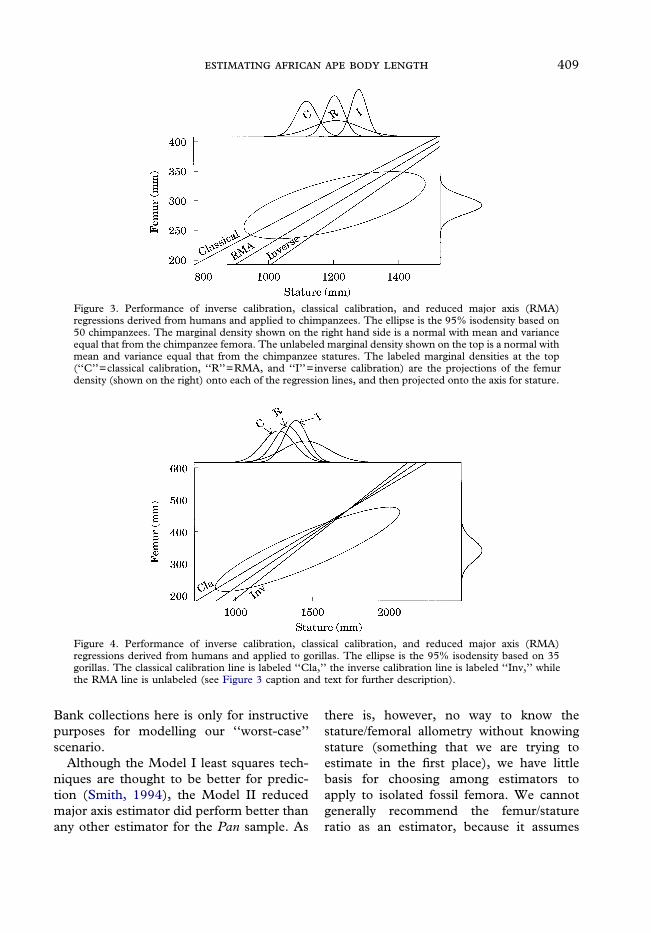
Obviously, chimpanzees are not allo-metrically scaled humans. Figures 3 and 4depict graphically the information includedin Table 3. Figure 3 shows the actual bi-variate ellipse for chimpanzees along withthe marginal distributions. The regressionlines shown in this graph are the ones fromthe human sample. This figure readily showsthat RMA is a relatively unbiased estimatorfor body length, classical calibration under-estimates length, and inverse calibrationoverestimates body length. On the rightside of the graph is the marginal distributionfor femoral length in the Pan sample. Atthe top of the graph are the marginal distri-butions for actual chimpanzee body lengthand the estimated body lengths from classi-cal calibration, RMA, and inverse cali-bration. The relationships of these marginaldistributions parallel the relationships seenin the regression lines running through theellipse.
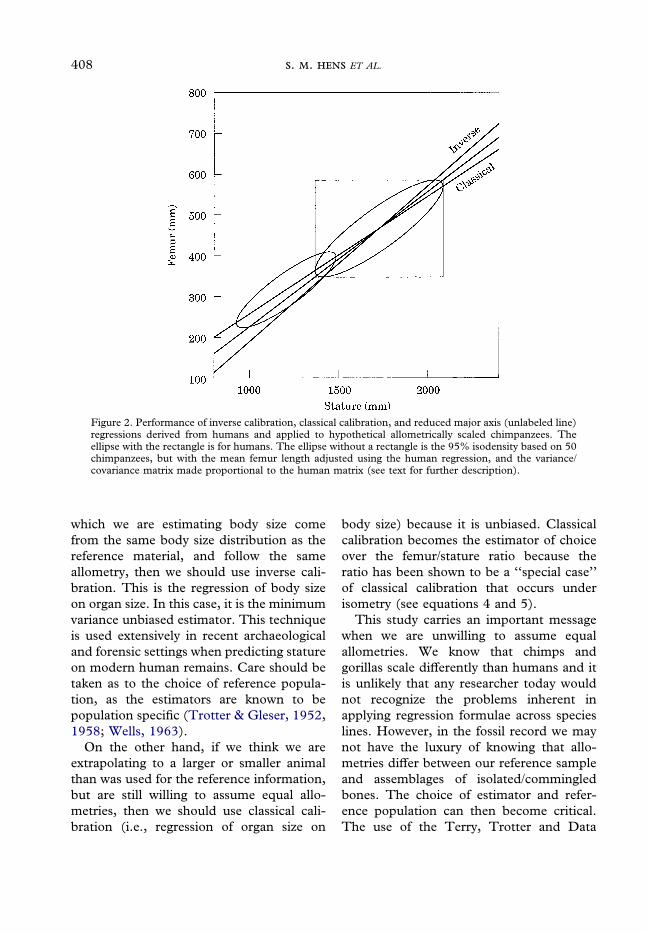
The comparable graphs for gorillas(Figure 4) show that inverse calibration isthe closest to being unbiased, while RMAand classical calibration both underestimatebody length. Again, this is clear from look-ing at the marginal distributions. On the topof Figure 4, the marginal distribution forgorilla body length and the lines represent-ing the estimated body lengths are drawn; allestimated lines underestimate the true bodylength for gorillas.
Discussion
The prediction of stature or body lengthin modern and fossil hominids is repletewith methodological difficulties, some ofwhich are addressed here. This paperespecially tackles the difficulties with theselection of the stature estimator and thechoice of reference population. First, if weare willing to assume that the specimens for
Figure 1. Femur/stature ratio, major axis (MA), and classical calibration lines for femur length and stature(both in millimeters) based on 2053 humans. The ellipse is the 95% isodensity for cases, while theinscribed rectangle is tangent to four points on this ellipse (see Sjøvold, 1990). The ratio and MA lines arelabeled, while the unlabeled line is the classical calibration line.
408 . . ET AL.
which we are estimating body size comefrom the same body size distribution as thereference material, and follow the sameallometry, then we should use inverse cali-bration. This is the regression of body sizeon organ size. In this case, it is the minimumvariance unbiased estimator. This techniqueis used extensively in recent archaeologicaland forensic settings when predicting statureon modern human remains. Care should betaken as to the choice of reference popula-tion, as the estimators are known to bepopulation specific (Trotter & Gleser, 1952,1958; Wells, 1963).On the other hand, if we think we are
extrapolating to a larger or smaller animalthan was used for the reference information,but are still willing to assume equal allo-metries, then we should use classical cali-bration (i.e., regression of organ size on
body size) because it is unbiased. Classicalcalibration becomes the estimator of choiceover the femur/stature ratio because theratio has been shown to be a ‘‘special case’’of classical calibration that occurs underisometry (see equations 4 and 5).This study carries an important message
when we are unwilling to assume equalallometries. We know that chimps andgorillas scale differently than humans and itis unlikely that any researcher today wouldnot recognize the problems inherent inapplying regression formulae across specieslines. However, in the fossil record we maynot have the luxury of knowing that allo-metries differ between our reference sampleand assemblages of isolated/commingledbones. The choice of estimator and refer-ence population can then become critical.The use of the Terry, Trotter and Data
Figure 2. Performance of inverse calibration, classical calibration, and reduced major axis (unlabeled line)regressions derived from humans and applied to hypothetical allometrically scaled chimpanzees. Theellipse with the rectangle is for humans. The ellipse without a rectangle is the 95% isodensity based on 50chimpanzees, but with the mean femur length adjusted using the human regression, and the variance/covariance matrix made proportional to the human matrix (see text for further description).
409
Bank collections here is only for instructivepurposes for modelling our ‘‘worst-case’’scenario.Although the Model I least squares tech-
niques are thought to be better for predic-tion (Smith, 1994), the Model II reducedmajor axis estimator did perform better thanany other estimator for the Pan sample. As
there is, however, no way to know thestature/femoral allometry without knowingstature (something that we are trying toestimate in the first place), we have littlebasis for choosing among estimators toapply to isolated fossil femora. We cannotgenerally recommend the femur/statureratio as an estimator, because it assumes
Figure 3. Performance of inverse calibration, classical calibration, and reduced major axis (RMA)regressions derived from humans and applied to chimpanzees. The ellipse is the 95% isodensity based on50 chimpanzees. The marginal density shown on the right hand side is a normal with mean and varianceequal that from the chimpanzee femora. The unlabeled marginal density shown on the top is a normal withmean and variance equal that from the chimpanzee statures. The labeled marginal densities at the top(‘‘C’’=classical calibration, ‘‘R’’=RMA, and ‘‘I’’=inverse calibration) are the projections of the femurdensity (shown on the right) onto each of the regression lines, and then projected onto the axis for stature.
Figure 4. Performance of inverse calibration, classical calibration, and reduced major axis (RMA)regressions derived from humans and applied to gorillas. The ellipse is the 95% isodensity based on 35gorillas. The classical calibration line is labeled ‘‘Cla,’’ the inverse calibration line is labeled ‘‘Inv,’’ whilethe RMA line is unlabeled (see Figure 3 caption and text for further description).

410 . . ET AL.
isometry both within the reference sampleand for the isolated femur. This is astrong assumption, and one that we suspecthas relatively little empirical support,particularly across primate taxa.
Acknowledgements
We would like to thank Dr Lee Meadows-Jantz for accessing the Trotter World War IIand Korean War data from punch cards andputting it into machine readable format. Wealso wish to express appreciation to theanonymous reviewers for their comments onan earlier draft of this paper.
References
Coolidge, H. J. & Shea, B. T. (1982). External bodydimensions of Pan paniscus and Pan troglodyteschimpanzees. Primates 23, 245–251.
Cousins, D. (1972). Body measurements and weightsof wild and captive gorillas, Gorilla gorilla. Zool.Garten N.F. Leipzig 41, 261–277.
Dwight, T. (1894). Methods of estimating the heightfrom parts of the skeleton. Med. Rec. 46, 293–296.
Feldesman, M. R., Kleckner, J. G. & Lundy, J. K.(1990). Femur/stature ratio and estimates of staturein mid- and late-Pleistocene fossil hominids. Am. J.phys. Anthrop. 83, 359–372.
Feldesman, M. R. & Lundy, J. K. (1988). Statureestimates for some African Plio-Pleistocene fossilhominids. J. hum. Evol. 17, 583–596.
Feldesman, M. R., Lundy, J. K. & Kleckner, J. G.(1989). The femur-stature ratio and estimates ofstature in mid- to late-Pleistocene fossil hominids.Am. J. phys. Anthrop. 78, 219–220.
Fleagle, J. G. (1978). Size distribution of living andfossil primate faunas. Paleobiol. 4, 67–76.
Geissmann, T. (1986). Estimation of australopithecinestature from long bones: A.L. 288-1 as a test case.Folia Primatol. 47, 119–127.
Jantz, L. M. (1996). Secular Change and Allometry inthe Long Limb Bones of Americans from the Mid1700’s Through the 1900’s. Ph.D. Dissertation,University of Tennessee, Knoxville.
Johanson, D. C. & Edey, M. A. (1981). Lucy: TheBeginnings of Humankind. New York: Simon &Schuster.
Jungers, W. L. (1982). Lucy’s limbs: skeletal allometryand locomotion in Australopithecus afarensis. Nature297, 676–678.
Jungers, W. L. (1988). Lucy’s length: staturereconstruction in Australopithecus afarensis (A.L.288-1) with implications for other small-bodiedhominids. Am. J. phys. Anthrop. 76, 227–231.
Jungers, W. L. & Stern, J. T., Jr (1983). Body propor-tions, skeletal allometry and locomotion in the Hadarhominids: A reply to Wolpoff. J. hum. Evol. 12,673–684.
Kermack, K. A. & Haldane, J. B. S. (1950). Organiccorrelation and allometry. Biometrika 37, 30–41.
Konigsberg, L. W. (1997). Statistical inadmissibility ofratios for body size estimation. Am. J. phys.Anthropol. Suppl. 24, 146.
Konigsberg, L. W. & Frankenberg, S. R. (1994). Paleo-demography: ‘‘not quite dead’’. Evol. Anthropol. 3,92–105.
Leakey, R. E. F. & Walker, A. C. (1985). Homo erectusunearthed. Nat. Geogr. 168, 624–629.
Lovejoy, C. O. & Heiple, K. G. (1970). A reconstruc-tion of the femur of Australopithecus africanus. Am. J.phys. Anthrop. 32, 33–40.
Manouvrier, L. (1893). La determination de la statured’apres les grands os des membres. Mem. Soc.Anthropol. Paris 4, 347–402.
Martin, R. A. (1990). Estimating body mass and cor-related variables in extinct mammals: travels in thefourth dimension. In (J. Damuth & B. J. MacFadden,Eds) Body Size in Mammalian Paleobiology: Estimationand Biological Implications, pp. 49–68. Cambridge:Cambridge University Press.
Martin, R. D. & Barbour, A. D. (1989). Aspects ofline-fitting in bivariate allometric analyses. FoliaPrimatol. 53, 65–81.
McArdle, B. H. (1988). The structural relationship:regression in biology. Can. J. Zool. 66, 2329–2339.
McHenry, H. M. (1974). How large were the australo-pithecines? Am. J. phys. Anthrop. 40, 329–340.
Moore-Jansen, P. H., Ousley, S. D. & Jantz, R. L.(1994). Data collection procedures for forensic skeletalmaterial, Third Edition. Knoxville, Tennessee:Department of Anthropology, University ofTennessee, Knoxville, Report of Investigations,no. 48.
Olivier, G. (1976). The stature of australopithecines.J. hum. Evol. 5, 529–534.
Pearson, K. (1899). On the reconstruction of statureof prehistoric races. Mathematical contribution tothe theory of evolution. Trans. Roy. Soc. A 192,169–244.
Rayner, J. M. V. (1985). Linear relations in bio-mechanics: the statistics of scaling functions. J. Zool.Lond. 206, 415–439.
Reed, C. A. & Falk, D. (1977). The stature and weightof Sterkfontein 14, a gracile australopithecine fromthe Transvaal, as determined from the innominatebone. Fieldiana (Geol.) 33, 423–440.
Ricker, W. E. (1973). Linear regressions in fisheryresearch. J. Fish. Res. Board Can. 30, 409–434.
Robinson, J. T. (1972). Early Hominid Posture andLocomotion. Chicago: University of Chicago Press.
Ruff, C. (1988). Hindlimb articular surface allometryin Hominoidea and Macaca, with comparisons todiaphyseal scaling. J. hum. Evol. 17, 687–714.
Schmid, P. (1986). Mehr Funde–mehr Wissen? ZurFundgesschichte der menschlichen Stammesge-schichte. Vjschr. naturf. Ges. Zurich 131, 145–160.

411
Schmidt-Nielsen, K. (1977). Problems of scaling: loco-motion and physiological correlates. In (T. J. Redley,Ed.) Scale Effects in Animal Locomotion, pp. 1–21.New York: Academic Press.
Schmidt-Nielsen, K. (1984). Scaling: Why is AnimalSize so Important? Cambridge: Cambridge UniversityPress.
Seim, E. & Sæther, B. E. (1983). On rethinkingallometry: which regression model to use? J. theor.Biol. 104, 161–168.
Sjøvold, T. (1990). Estimation of stature from longbones utilizing the line of organic correlation. Hum.Evol. 5, 431–447.
Smith, R. J. (1993). Categories of allometry: body sizeversus biomechanics. J. hum. Evol. 24, 173–182.
Smith, R. J. (1994). Regression models for predictionequations. J. hum. Evol. 26, 239–244.
Sokal, R. R. & Rohlf, F. J. (1981). Biometry. New York:Freeman.
Terry, R. J. (1940). On measuring and photographingthe cadaver. Am. J. phys. Anthrop. 26, 433–447.
Trinkaus, E. (1981). Neanderthal limb proportions andcold adaptations. In (C. B. Stringer, Ed.) Aspects ofHuman Evolution, pp. 187–224. London: Taylor &Francis.
Trinkaus, E. (1983). The Shanidar Neanderthals. NewYork: Academic Press.
Trotter, M. & Gleser, G. C. (1952). Estimation ofstature from long bones of American whites andnegroes. Am. J. phys. Anthrop. 10, 463–514.
Trotter, M. & Gleser, G. C. (1958). A re-evaluation ofestimation of stature based on measurements ofstature taken during life and long bones after death.Am. J. phys. Anthrop. 16, 79–124.
Wells, L. H. (1963). Stature in earlier races inmankind. In (D. Brothwell & E. Higgs, Eds) Sciencein Archaeology, pp. 365–378. New York: Basic.
White, T. D. (1980). Evolutionary implications ofPliocene hominid footprints. Science 208, 175–176.
Wolpoff, M. H. (1973). Posterior tooth size, body size,and diet in South African gracile australopithecines.Am. J. phys. Anthrop. 39, 375–394.





























