Embed Size (px)
Citation preview

Research Article
Epidermal Lipids: Thermotropic Behavior and Role inTranscutaneous Permeation of Levodopa
Kumar Babita,1 Vikas Rana,2 and Ashok Kumar Tiwary1n
1Department of Pharmaceutical Sciences and Drug Research, Punjabi University, Punjab, India2Department of Pharmaceutical Sciences, Govt. Polytechnic for Women, Punjab, India
Strategy, Management and Health Policy
Enabling
Technology,
Genomics,
Proteomics
Preclinical
Research
Preclinical Development
Toxicology, Formulation
Drug Delivery,
Pharmacokinetics
Clinical Development
Phases I-III
Regulatory, Quality,
Manufacturing
Postmarketing
Phase IV
ABSTRACT Skin lipids play a pivotal role in maintaining the barrier properties of skin. The presentinvestigation aimed at studying the effect of skin lipid synthesis inhibitors on the transcutaneous delivery oflevodopa (LD). Skin perturbation with ethanol was found to reduce the content of cholesterol, fatty acid,and sphingosine. Subsequent treatment with various doses of their respective synthesis inhibitorsatorvastatin, cerulenin, and b-chloroalanine sustained the reduced content of these lipids in viable rat skin.A maximum synthesis inhibition of approximately 80, 60, and 55% for cholesterol, triglycerides (anindicator of fatty acid), and sphingosine (a precursor of ceramide) was observed at 2, 2, and 8 h,respectively, after topical application of optimized dose of the inhibitors. Differential scanningcalorimetric (DSC) analysis of treated rat epidermis revealed approximately 70, 52, and 50% reductionin enthalpy (DH) of the lipid endothermic transition peak by the corresponding doses of inhibitors. Theeffective plasma concentration (Ceff) of LD in rats was achieved by treatment with atorvastatin, cerulenin,and b-chloroalanine within 2, 4, and 6 h, respectively. After application of a combination of theseinhibitors, the lag time was reduced to 1 h and Ceff was maintained over 48 h. The inhibition of lipidsynthesis in epidermis and concomitant decrease in enthalpy of lipid endothermic transition were directlycorrelated with enhanced in vitro and in vivo permeation of LD. These findings suggest that lipid synthesisinhibition can be exploited to enhance transcutaneous delivery of a polar drug, levodopa. Drug Dev Res63:190–199, 2004. �c 2004 Wiley-Liss, Inc.
Key words: epidermal cholesterol; triglycerides; sphingosine; differential scanning calorimetry; levodopa
INTRODUCTION
The transdermal route is the most preferred non-invasive route for drug delivery. However, the formid-able barrier properties of stratum corneum (SC) pose asevere limitation. The barrier properties and integrityof SC are attributed to the presence of intercellularlipid multilayers that are enriched in ceramide,cholesterol, and fatty acid [Elias and Menon, 1991].The rate-limiting step for the synthesis of sphingosine(a precursor for ceramide) is the condensationof palmitoyl CoA with L-serine in the presence ofserine palmitoyltransferase (SPT) enzyme. This rate-limiting enzyme has been reported to be inhibited by
b-chloroalanine (b-CA), resulting in decreased cera-mide content in the skin [Medlock and Merrill, 1988].This is accompanied by the appearance of abnormallamellar bodies and extracellular membrane structure
DDR
Grant sponsor: Council of Scientific and Industrial Research(CSIR), New Delhi, India; Grant number: 01(1682)/00/EMR-II.
nCorrespondence to: Ashok Kumar Tiwary, Department ofPharmaceutical Sciences and Drug Research, Punjabi University,Patiala-147 002, Punjab, India. E-mail: [email protected]
Received 9 August 2004; Accepted 2 December 2004
Published online in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/ddr.10413
DRUG DEVELOPMENT RESEARCH 63:190–199 (2004)
�c 2004 Wiley-Liss, Inc.

in the ultrastructure of skin leading to impaired barrierfunction [Behne et al., 2000]. Similarly, cholesterol issynthesized by the conversion of 3-hydroxy-3-methyl-glutaryl coenzyme A (HMG-CoA) to mevalonate in theviable epidermis and atorvastatin (AVN) is a selectiveinhibitor of this enzyme [Phillips, 1997]. There are twokey enzymes, acetyl-CoA carboxylase (ACC) and fattyacid synthase (FAS) enzyme system, that are known tobe involved in the biosynthesis of fatty acids [Otteyet al., 1995]. ACC carboxylates acetyl-CoA to producemalonyl-CoA, which is converted to free palmitate byFAS [Thupari et al., 2001]. The synthesis of fatty acidshas been reported to be inhibited by 5-(tetradecyloxy)-2-furancarboxylic acid (TOFA) or cerulenin (CN). It isfound that the estrification of TOFA with CoAproduces TOFAyl-CoA, which profoundly inhibitedACC [Halvorson and McCune, 1984] whereas cerule-nin non-competitively inhibited the FAS system [Vanceet al., 1972].
Levodopa (LD) is the drug of first choice for thetreatment of Parkinson’s Disease. It has a short half-lifeof 1.4 h and undergoes extensive first-pass metabolism[Robertson et al., 1989], necessitating oral administra-tion 3–4 times a day. The need to maintain non-fluctuating plasma LD concentrations in order to avoidakinesia and dyskinesia [Harder et al., 1995] makestransdermal delivery an attractive alternative. However,the percutaneous permeation of LD is very low due toits hydrophilicity (log Ko/w¼�4.7). Therefore, over-coming the barrier properties of skin seems to beessential for transcutaneous delivery of LD.
Topical application of solvents is known toremove the lipids from SC. This leads to increasedsynthesis of ceramide [Babita et al., 2002; Gupta et al.,2003], cholesterol, and fatty acids in the epidermis[Tsai et al., 1996] in an attempt to restore the barrierstatus of SC. Hence, it can be hypothesized thattreatment of lipid synthesis inhibitors to solventperturbed viable rat skin shall play a vital role inenhancing the percutaneous delivery of drugs.
This study was performed to investigate theinfluence of altered lipid content on in vitro and invivo permeation of LD across rat epidermis. Further,differential scanning calorimetric analysis of rat epi-dermis was carried out to investigate the differences inthermotropic behavior of epidermal lipids due to theaction of lipid synthesis inhibitors.
MATERIALS AND METHODS
Materials
Levodopa and carbidopa were generous giftsamples from Sun Pharma, Baroda, India. D-sphingo-sine, b-chloroalanine, and cerulenin were purchased
from Sigma Chemicals (St. Louis, MO). Atorvastatinwas a gift sample from Ranbaxy Research Laboratories,Gurgaon, India. Cholesterol and triglycerides estima-tion kits were purchased from Span Diagnostics andRanbaxy Laboratories, India, respectively. All otherchemicals were of AR grade.
Albino Wistar rats of either sex (190–210 g),maintained on a standard laboratory diet and tap waterad libitum were used in this study. Various treatmentswere applied to the dorsal skin portion of rats aftershaving with an electric razor. Rats were sacrificed afterdifferent time periods of skin treatment by excessinhalation of anesthetic ether and treated skin portionswere excised for further experiments. The protocol wasapproved by Institutional Animal Ethical Committee.
Methods
Preparation of Epidermal SkinEpidermal sheets were prepared from freshly
excised whole skin portions by soaking it in water at601C for 45 s. The underlying dermis was removed bygentle scraping with a spatula in order to obtainepidermal sheets [Cornwell et al., 1996].
Differential Scanning Calorimetric (DSC) AnalysisDSC analysis (251 to 1101C, 11C/min) was carried
out on both untreated epidermal sheets and thoseobtained from excised viable skin after treatment withAVN, b-CA or CN (Mettler Toledo Star System, 821E,Switzerland). Various doses of AVN (500, 750, or 1,000mg/7cm2), CN (10, 20, or 30 mg/7cm2) or b-CA (400,600, or 1,200 mg/7cm2) were applied to patchesprepared on dorsal skin surface of rats. The epidermiswas prepared from the whole skin excised at differentintervals of treatment. Samples of dry epidermis werehydrated in a closed chamber containing a saturatedsolution of sodium chloride (75% RH at 251C) prior toDSC analysis. All DSC curves were evaluated espe-cially with regard to the phase-transition enthalpies(peak areas) and peak maximum temperatures (Tm) oflipid-phase transitions at near 701C. The percentagereduction in enthalpy (DH) of this endotherm (J/g) wascalculated by using the formula: 100�[(DH of lipidendotherm due to treatmentCDH of lipid endothermin untreated epidermis)� 100]. All experiments werecarried out in triplicate.
Dose Dependent Influence of AVN, CN, or b-CA onCholesterol, Fatty Acid, (as Triglycerides), andSphingosine Content, Respectively, in Viable RatSkin
Three patches were prepared on dorsal skinsurface of rats by shaving with an electric razor. Onepatch was left unperturbed (control). A second patch
THERMOTROPY OF EPIDERMAL LIPIDS: LD PERMEATION 191

was perturbed by ethanol treatment (0.5 ml). The thirdpatch was perturbed by ethanol followed by immediateapplication of AVN (500, 750, or 1,000 mg) or CN (10,20, or 30 mg) or b-CA (400, 600, or 1,200 mg) solutionprepared in 0.25 ml of propylene glycol:ethanol (7:3)mixture (PG:EtOH). The animals were sacrificed after2, 4, 6, 8, 12, 24, 36, or 48 h and epidermis wasseparated from treated skin patches. These epidermalsheets were dried to constant weight and total lipidswere extracted by Folch method. Cholesterol ortriglycerides content in these extracts was determinedby using the respective diagnostic kits. Sphingosinecontent was determined spectrofluorometrically utiliz-ing an excitation wavelength of 340 nm and emissionwavelength of 455 nm according to the proceduredescribed earlier [Babita et al., 2002]. The percentagelipid synthesis inhibited was calculated by using theformula: [1�(lipid content remaining in skin afterinhibitor treatmentClipid content in normalskin)� 100]. Three experiments were carried out foreach study.
In Vitro Permeation Studies of LD Across Ethanol-Perturbed AVN, b-CA, or CN-Treated Excised RatEpidermal Skin
Two patches (7 cm2), one on either side of thespinal cord, were prepared by shaving with an electricrazor. One patch was left unperturbed and served ascontrol. The other patch received ethanol treatmentfollowed by immediate application of AVN (500, 750, or1,000 mg), CN (10, 20, or 30 mg), or b-CA (400, 600, or1,200 mg) dissolved in PG:EtOH (0.25 ml). The animalswere sacrificed after different intervals. The epidermalsheets obtained from these excised patches were usedfor studying the in vitro permeation of LD afterstabilizing them for 4 h in vertical Franz glass diffusioncells. The donor fluid contained LD (64 mg) andsodium sulfite (0.25% w/v) dispersed in PG:EtOHmixture. Phosphate buffer (pH 7.4) containing sodiumazide (0.05% w/v), sodium sulfite (0.25% w/v), andPEG 400 (10% v/v) was used as receptor fluid. Sampleswere withdrawn at various time intervals through 48 hand immediately analyzed spectrophotometrically forthe amount of LD permeated at 280 nm [Gamez et al.,1976]. A standard plot of LD (5–50 mg/ml) wasprepared in phosphate buffer (pH 7.4) containingsodium sulfite (0.25% w/v) at 280 nm. Cartesian plotsof the cumulative amount of drug permeated intoreceptor compartment vs. time were plotted for threeexperiments and flux (mg/cm2/h) was calculated fromthe slope of the steady-state portion of these plots.
Pharmacokinetic Studies
An adhesive transdermal patch was prepared byusing adhesive surgical tape, a plastic ring, and apolyethylene backing membrane. LD (64 mg/ml)dispersed in a PG:EtOH mixture was loaded into thiscavity. The treatment given to various groups, eachconsisting of five rats, was: Group I: normal skin þ LDþ carbidopa (16 mg); Group II: ethanol perturbation(0.5 ml.) þ LD þ carbidopa; Group III: ethanolperturbation þ AVN (750 mg) þ LD þ carbidopa;Group IV: ethanol perturbation þ CN (20 mg) þ LD þcarbidopa; Group V: ethanol perturbation þ b-CA (600mg) þ LD þ carbidopa; Group VI: ethanol perturba-tion þ AVN (750 mg) þ b-CA (600 mg) þ CN (20 mg) þLD þ carbidopa.
Blood samples (0.25 ml) were withdrawn afterspecified time intervals and centrifuged at 2,500 rpmfor 10 min at 41C. The plasma was analyzed for intactLD spectrofluorometrically utilizing excitation andemission wavelengths of 440 and 470 nm, respectively,after coupling it with naphthoresorcinol [Babita et al.,2002].
Statistics
Values were expressed as mean 7 SD. The datawas analyzed using the Student’s t-test. Values(Po0.05) were considered statistically significant.
RESULTS
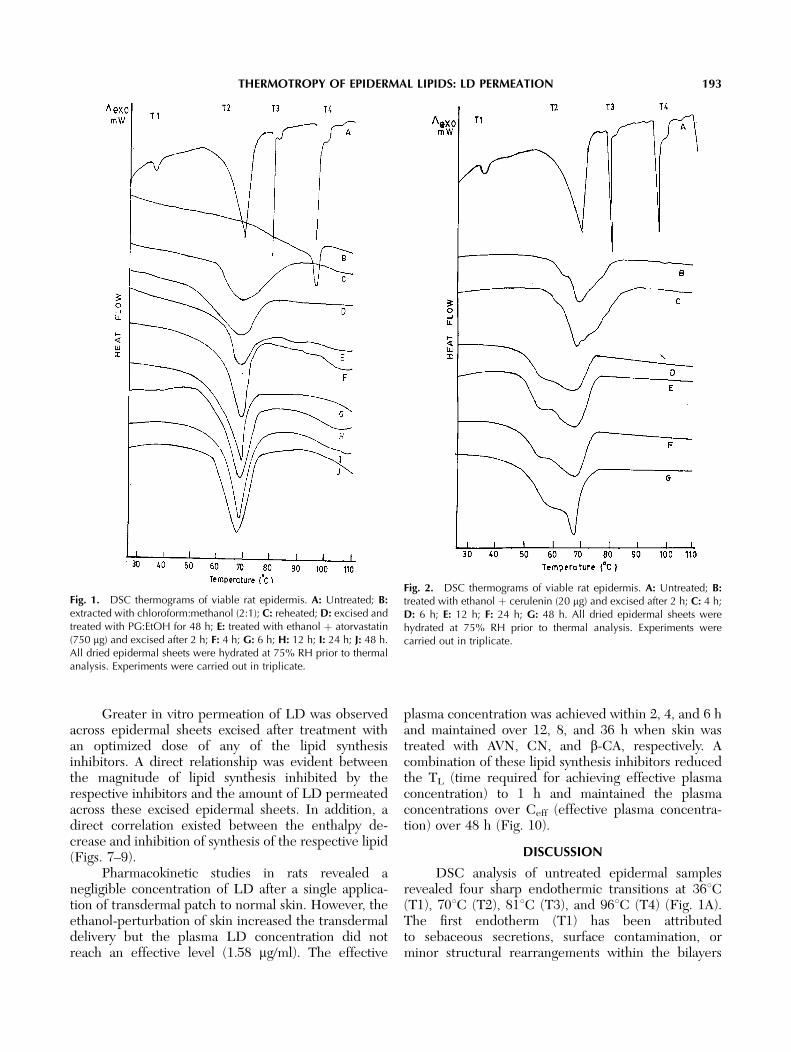
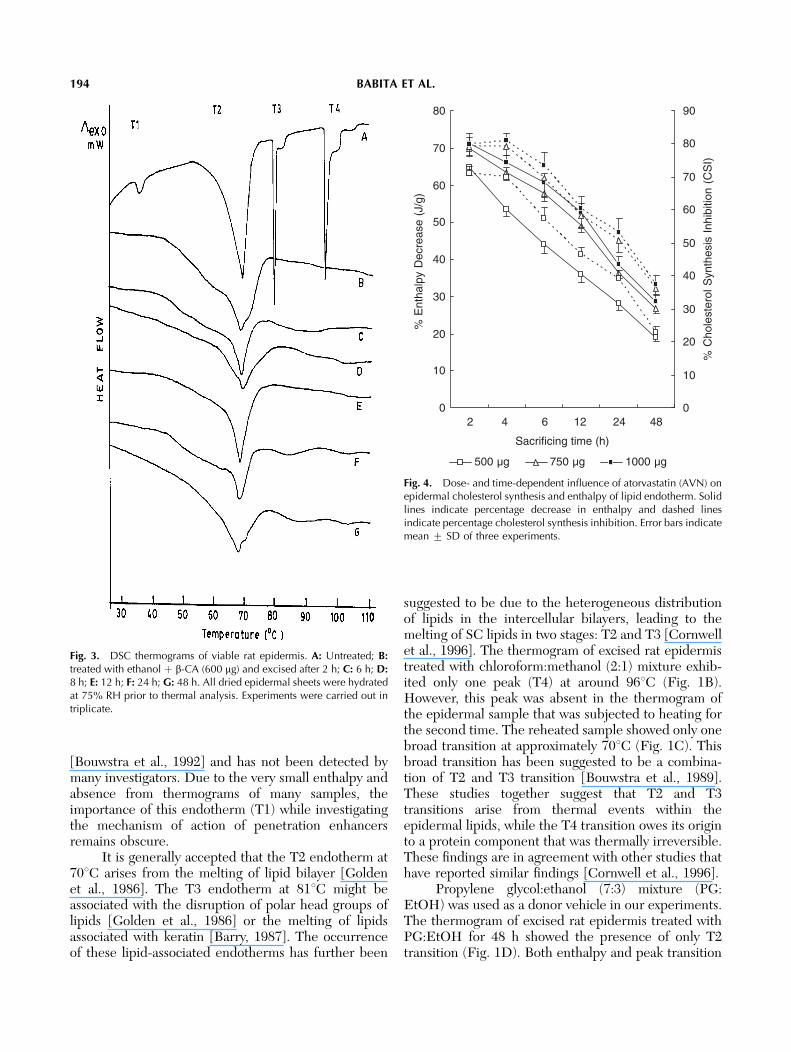
Thermograms obtained by differential scanningcalorimetric analysis of epidermal sheets excised aftervarious periods of treatment with optimized dose of therespective lipid synthesis inhibitors showed a decreasein the intensity (peak area) without any significanteffect on peak temperature of the lipid transitionendotherm. The lowest DH of lipid endothermic peakwas observed at 2 h after treatment with AVN and CNand at 8 h after treatment with b-CA (Figs. 1–3).
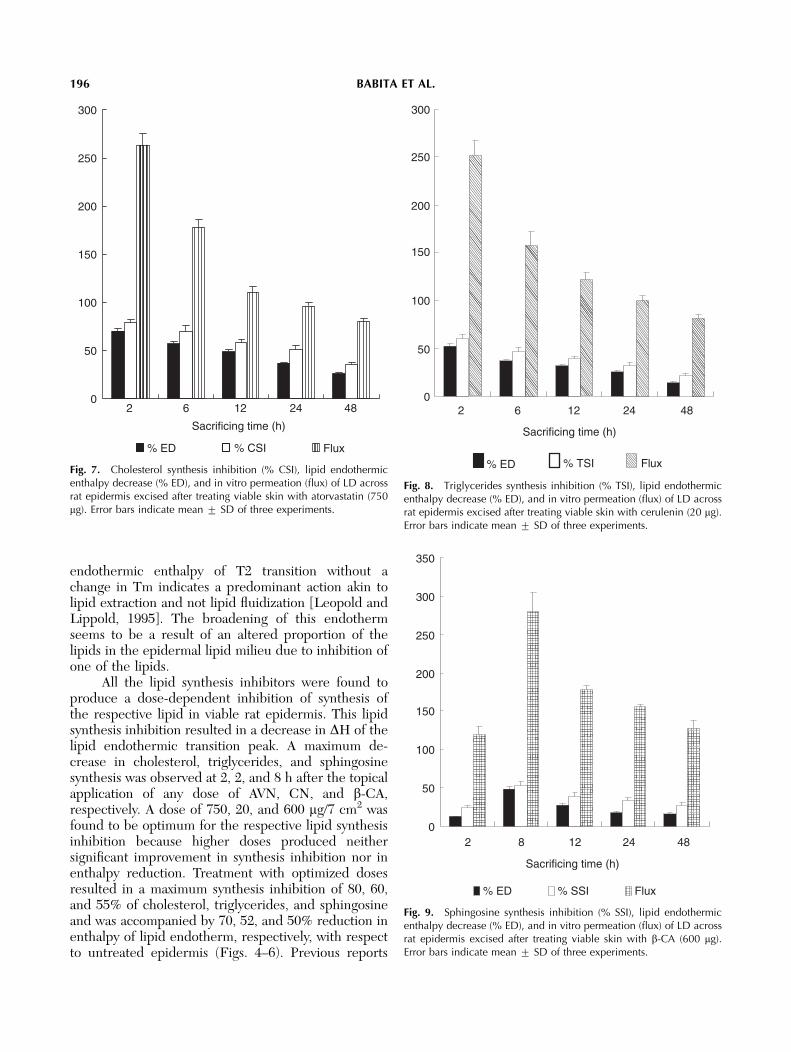
Topical application of different doses of AVN(500, 750, and 1,000 mg), CN (10, 20, and 30 mg) or b-CA (400, 600, and 1,200 mg) to ethanol–perturbedviable rat skin produced a maximum synthesis inhibi-tion of cholesterol, triglycerides (an indicator of fattyacid), and sphingosine (a precursor of ceramide) at 2, 2,and 8 h, respectively. In addition, synthesis inhibitionof these lipids was accompanied with decreasedenthalpies (DH) of lipid endothermic transition peaks.A dose of 750, 20, and 600 mg/7 cm2 of AVN, CN, andb-CA, respectively, was found to be optimum becausehigher doses of these inhibitors produced neithersignificantly higher (Po0.05) synthesis inhibition noran enthalpy decrease (Figs. 4–6).
192 BABITA ET AL.
Greater in vitro permeation of LD was observedacross epidermal sheets excised after treatment withan optimized dose of any of the lipid synthesisinhibitors. A direct relationship was evident betweenthe magnitude of lipid synthesis inhibited by therespective inhibitors and the amount of LD permeatedacross these excised epidermal sheets. In addition, adirect correlation existed between the enthalpy de-crease and inhibition of synthesis of the respective lipid(Figs. 7–9).
Pharmacokinetic studies in rats revealed anegligible concentration of LD after a single applica-tion of transdermal patch to normal skin. However, theethanol-perturbation of skin increased the transdermaldelivery but the plasma LD concentration did notreach an effective level (1.58 mg/ml). The effective
plasma concentration was achieved within 2, 4, and 6 hand maintained over 12, 8, and 36 h when skin wastreated with AVN, CN, and b-CA, respectively. Acombination of these lipid synthesis inhibitors reducedthe TL (time required for achieving effective plasmaconcentration) to 1 h and maintained the plasmaconcentrations over Ceff (effective plasma concentra-tion) over 48 h (Fig. 10).
DISCUSSION
DSC analysis of untreated epidermal samplesrevealed four sharp endothermic transitions at 361C(T1), 701C (T2), 811C (T3), and 961C (T4) (Fig. 1A).The first endotherm (T1) has been attributedto sebaceous secretions, surface contamination, orminor structural rearrangements within the bilayers
Fig. 1. DSC thermograms of viable rat epidermis. A: Untreated; B:extracted with chloroform:methanol (2:1); C: reheated; D: excised andtreated with PG:EtOH for 48 h; E: treated with ethanol þ atorvastatin(750 mg) and excised after 2 h; F: 4 h; G: 6 h; H: 12 h; I: 24 h; J: 48 h.All dried epidermal sheets were hydrated at 75% RH prior to thermalanalysis. Experiments were carried out in triplicate.
Fig. 2. DSC thermograms of viable rat epidermis. A: Untreated; B:treated with ethanol þ cerulenin (20 mg) and excised after 2 h; C: 4 h;D: 6 h; E: 12 h; F: 24 h; G: 48 h. All dried epidermal sheets werehydrated at 75% RH prior to thermal analysis. Experiments werecarried out in triplicate.
THERMOTROPY OF EPIDERMAL LIPIDS: LD PERMEATION 193
[Bouwstra et al., 1992] and has not been detected bymany investigators. Due to the very small enthalpy andabsence from thermograms of many samples, theimportance of this endotherm (T1) while investigatingthe mechanism of action of penetration enhancersremains obscure.
It is generally accepted that the T2 endotherm at701C arises from the melting of lipid bilayer [Goldenet al., 1986]. The T3 endotherm at 811C might beassociated with the disruption of polar head groups oflipids [Golden et al., 1986] or the melting of lipidsassociated with keratin [Barry, 1987]. The occurrenceof these lipid-associated endotherms has further been
suggested to be due to the heterogeneous distributionof lipids in the intercellular bilayers, leading to themelting of SC lipids in two stages: T2 and T3 [Cornwellet al., 1996]. The thermogram of excised rat epidermistreated with chloroform:methanol (2:1) mixture exhib-ited only one peak (T4) at around 961C (Fig. 1B).However, this peak was absent in the thermogram ofthe epidermal sample that was subjected to heating forthe second time. The reheated sample showed only onebroad transition at approximately 701C (Fig. 1C). Thisbroad transition has been suggested to be a combina-tion of T2 and T3 transition [Bouwstra et al., 1989].These studies together suggest that T2 and T3transitions arise from thermal events within theepidermal lipids, while the T4 transition owes its originto a protein component that was thermally irreversible.These findings are in agreement with other studies thathave reported similar findings [Cornwell et al., 1996].
Propylene glycol:ethanol (7:3) mixture (PG:EtOH) was used as a donor vehicle in our experiments.The thermogram of excised rat epidermis treated withPG:EtOH for 48 h showed the presence of only T2transition (Fig. 1D). Both enthalpy and peak transition
Fig. 3. DSC thermograms of viable rat epidermis. A: Untreated; B:treated with ethanol þ b-CA (600 mg) and excised after 2 h; C: 6 h; D:8 h; E: 12 h; F: 24 h; G: 48 h. All dried epidermal sheets were hydratedat 75% RH prior to thermal analysis. Experiments were carried out intriplicate.
0
10
20
30
40
50
60
70
80
2 4 6 12 24 48
Sacrificing time (h)
% E
ntha
lpy
Dec
reas
e (J
/g)
0
10
20
30
40
50
60
70
80
90
% C
hole
ster
ol S
ynth
esis
Inh
ibiti
on (
CS
I)
500 µg 750 µg 1000 µg
Fig. 4. Dose- and time-dependent influence of atorvastatin (AVN) onepidermal cholesterol synthesis and enthalpy of lipid endotherm. Solidlines indicate percentage decrease in enthalpy and dashed linesindicate percentage cholesterol synthesis inhibition. Error bars indicatemean 7 SD of three experiments.
194 BABITA ET AL.
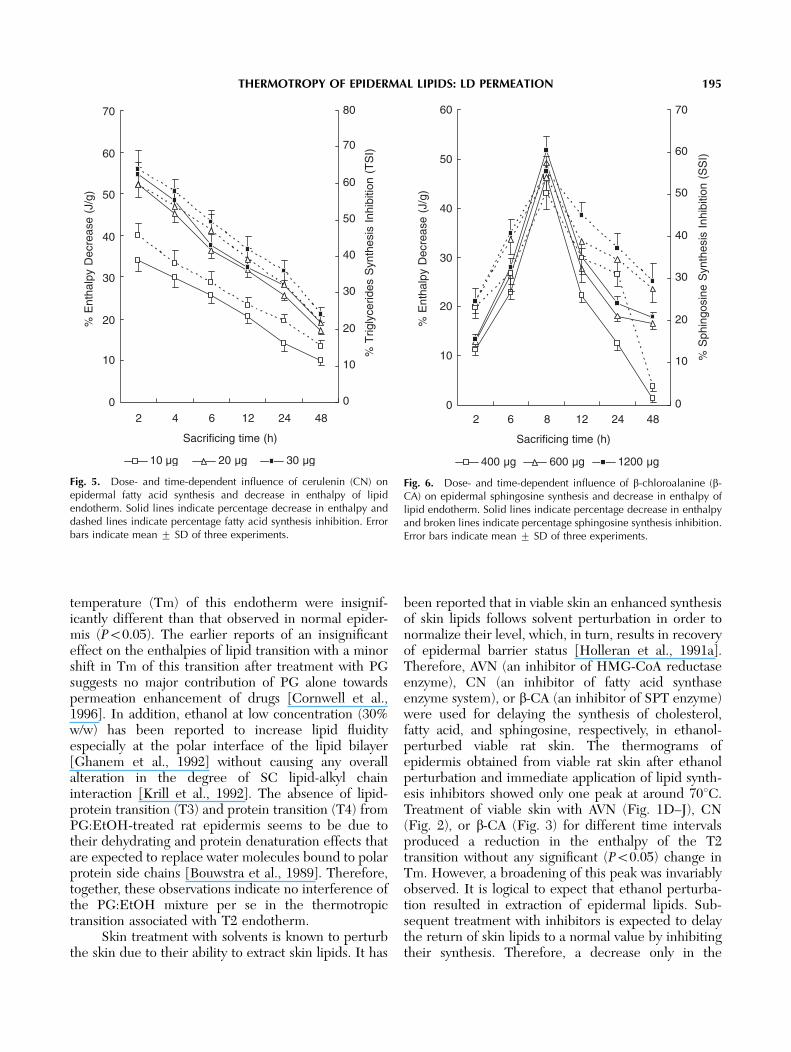
temperature (Tm) of this endotherm were insignif-icantly different than that observed in normal epider-mis (Po0.05). The earlier reports of an insignificanteffect on the enthalpies of lipid transition with a minorshift in Tm of this transition after treatment with PGsuggests no major contribution of PG alone towardspermeation enhancement of drugs [Cornwell et al.,1996]. In addition, ethanol at low concentration (30%w/w) has been reported to increase lipid fluidityespecially at the polar interface of the lipid bilayer[Ghanem et al., 1992] without causing any overallalteration in the degree of SC lipid-alkyl chaininteraction [Krill et al., 1992]. The absence of lipid-protein transition (T3) and protein transition (T4) fromPG:EtOH-treated rat epidermis seems to be due totheir dehydrating and protein denaturation effects thatare expected to replace water molecules bound to polarprotein side chains [Bouwstra et al., 1989]. Therefore,together, these observations indicate no interference ofthe PG:EtOH mixture per se in the thermotropictransition associated with T2 endotherm.
Skin treatment with solvents is known to perturbthe skin due to their ability to extract skin lipids. It has
been reported that in viable skin an enhanced synthesisof skin lipids follows solvent perturbation in order tonormalize their level, which, in turn, results in recoveryof epidermal barrier status [Holleran et al., 1991a].Therefore, AVN (an inhibitor of HMG-CoA reductaseenzyme), CN (an inhibitor of fatty acid synthaseenzyme system), or b-CA (an inhibitor of SPT enzyme)were used for delaying the synthesis of cholesterol,fatty acid, and sphingosine, respectively, in ethanol-perturbed viable rat skin. The thermograms ofepidermis obtained from viable rat skin after ethanolperturbation and immediate application of lipid synth-esis inhibitors showed only one peak at around 701C.Treatment of viable skin with AVN (Fig. 1D–J), CN(Fig. 2), or b-CA (Fig. 3) for different time intervalsproduced a reduction in the enthalpy of the T2transition without any significant (Po0.05) change inTm. However, a broadening of this peak was invariablyobserved. It is logical to expect that ethanol perturba-tion resulted in extraction of epidermal lipids. Sub-sequent treatment with inhibitors is expected to delaythe return of skin lipids to a normal value by inhibitingtheir synthesis. Therefore, a decrease only in the
0
10
20
30
40
50
60
70
2 4 6 12 24 48
Sacrificing time (h)
% E
ntha
lpy
Dec
reas
e (J
/g)
0
10
20
30
40
50
60
70
80
% T
rigly
cerid
es S
ynth
esis
Inh
ibiti
on (
TS
I)
10 µg 20 µg 30 µg
Fig. 5. Dose- and time-dependent influence of cerulenin (CN) onepidermal fatty acid synthesis and decrease in enthalpy of lipidendotherm. Solid lines indicate percentage decrease in enthalpy anddashed lines indicate percentage fatty acid synthesis inhibition. Errorbars indicate mean 7 SD of three experiments.
0
10
20
30
40
50
60
2 6 8 12 24 48
Sacrificing time (h)
% E
ntha
lpy
Dec
reas
e (J
/g)
0
10
20
30
40
50
60
70
% S
phin
gosi
ne S
ynth
esis
Inh
ibiti
on (
SS
I)
400 µg 600 µg 1200 µg
Fig. 6. Dose- and time-dependent influence of b-chloroalanine (b-CA) on epidermal sphingosine synthesis and decrease in enthalpy oflipid endotherm. Solid lines indicate percentage decrease in enthalpyand broken lines indicate percentage sphingosine synthesis inhibition.Error bars indicate mean 7 SD of three experiments.
THERMOTROPY OF EPIDERMAL LIPIDS: LD PERMEATION 195
endothermic enthalpy of T2 transition without achange in Tm indicates a predominant action akin tolipid extraction and not lipid fluidization [Leopold andLippold, 1995]. The broadening of this endothermseems to be a result of an altered proportion of thelipids in the epidermal lipid milieu due to inhibition ofone of the lipids.
All the lipid synthesis inhibitors were found toproduce a dose-dependent inhibition of synthesis ofthe respective lipid in viable rat epidermis. This lipidsynthesis inhibition resulted in a decrease in DH of thelipid endothermic transition peak. A maximum de-crease in cholesterol, triglycerides, and sphingosinesynthesis was observed at 2, 2, and 8 h after the topicalapplication of any dose of AVN, CN, and b-CA,respectively. A dose of 750, 20, and 600 mg/7 cm2 wasfound to be optimum for the respective lipid synthesisinhibition because higher doses produced neithersignificant improvement in synthesis inhibition nor inenthalpy reduction. Treatment with optimized dosesresulted in a maximum synthesis inhibition of 80, 60,and 55% of cholesterol, triglycerides, and sphingosineand was accompanied by 70, 52, and 50% reduction inenthalpy of lipid endotherm, respectively, with respectto untreated epidermis (Figs. 4–6). Previous reports
0
50
100
150
200
250
300
2 6 12 24 48
Sacrificing time (h)
% ED % CSI Flux
Fig. 7. Cholesterol synthesis inhibition (% CSI), lipid endothermicenthalpy decrease (% ED), and in vitro permeation (flux) of LD acrossrat epidermis excised after treating viable skin with atorvastatin (750mg). Error bars indicate mean 7 SD of three experiments.
0
50
100
150
200
250
300
2 6 12 24 48
Sacrificing time (h)
% ED % TSI Flux
Fig. 8. Triglycerides synthesis inhibition (% TSI), lipid endothermicenthalpy decrease (% ED), and in vitro permeation (flux) of LD acrossrat epidermis excised after treating viable skin with cerulenin (20 mg).Error bars indicate mean 7 SD of three experiments.
0
50
100
150
200
250
300
350
2 8 12 24 48
Sacrificing time (h)
% ED % SSI Flux
Fig. 9. Sphingosine synthesis inhibition (% SSI), lipid endothermicenthalpy decrease (% ED), and in vitro permeation (flux) of LD acrossrat epidermis excised after treating viable skin with b-CA (600 mg).Error bars indicate mean 7 SD of three experiments.
196 BABITA ET AL.
have indicated that both lovastatin [Feingold et al.,1990] and CN [Thupari et al., 2001] produced amaximum inhibition of cholesterol and fatty acidsynthesis at 2 h. However, despite a marked decreaseof sphingolipid synthesis within 3 h, b-CA was found todelay the barrier recovery only at later (4 6 h) timepoints [Holleran et al., 1991b]. Therefore, our findingson lipid synthesis inhibition are in consonance with theearlier reports. None of the synthesis inhibitors wascapable of totally arresting the synthesis of therespective lipid. Moreover, the lipid content was foundto increase with time after maximum inhibition,indicating the de novo synthesis of these lipids as anatural process in viable skin. However, at the end of48-h study, approximately 35% of cholesterol, 22% oftriglycerides, and 27% of sphingosine synthesis werefound to be still inhibited by the respective optimizeddose of lipid synthesis inhibitor and resulted in 27%(Fig. 1), 17% (Fig. 2), and 17% (Fig. 3) decrease inenthalpy of T2 endotherm with respect to normalepidermis.
The in vitro flux of LD across epidermis obtainedfrom normal viable rat skin was found to be 74 mg/cm2/h.The flux increased to 263, 252, and 280 mg/cm2/h acrossviable epidermis excised after treatment with opti-mized dose of AVN, CN, or b-CA for 2, 2, and 8 h,respectively. The data indicate an approximately 4-foldenhancement for in vitro permeation of LD due toinhibition of lipid synthesis in viable epidermis. It isnoteworthy that the time of excision of skin throughwhich maximum in vitro permeation of LD wasobserved coincided with the time point at whichmaximum synthesis inhibition of the respective lipidoccurred. In addition, lipid synthesis inhibition pro-duced a maximum decrease in the enthalpy of T2endotherm at the same time point (Figs. 4–6). Thissuggests a direct correlation between the magnitude ofepidermal lipid synthesis inhibited, the reducedenthalpy of lipid endotherm, and the enhancedpermeation of LD. The effect of all the lipid synthesisinhibitors on the in vitro permeation of LD, lipidsynthesis inhibition, and reduction of lipid endothermicenthalpy gradually decreased with the lapse of timeafter a single application of the respective inhibitors.The in vitro permeation of LD across epidermis excisedafter 48 h of single application of b-CA was approxi-mately 2-fold as compared to that across the normalepidermis (Fig. 9) whereas permeation across AVN- orCN-treated epidermis declined to normal level (Figs. 7and 8). This indicates a relatively greater role of thepolar lipid, sphingosine, in influencing permeation of apolar drug, LD, as compared to other lipids.
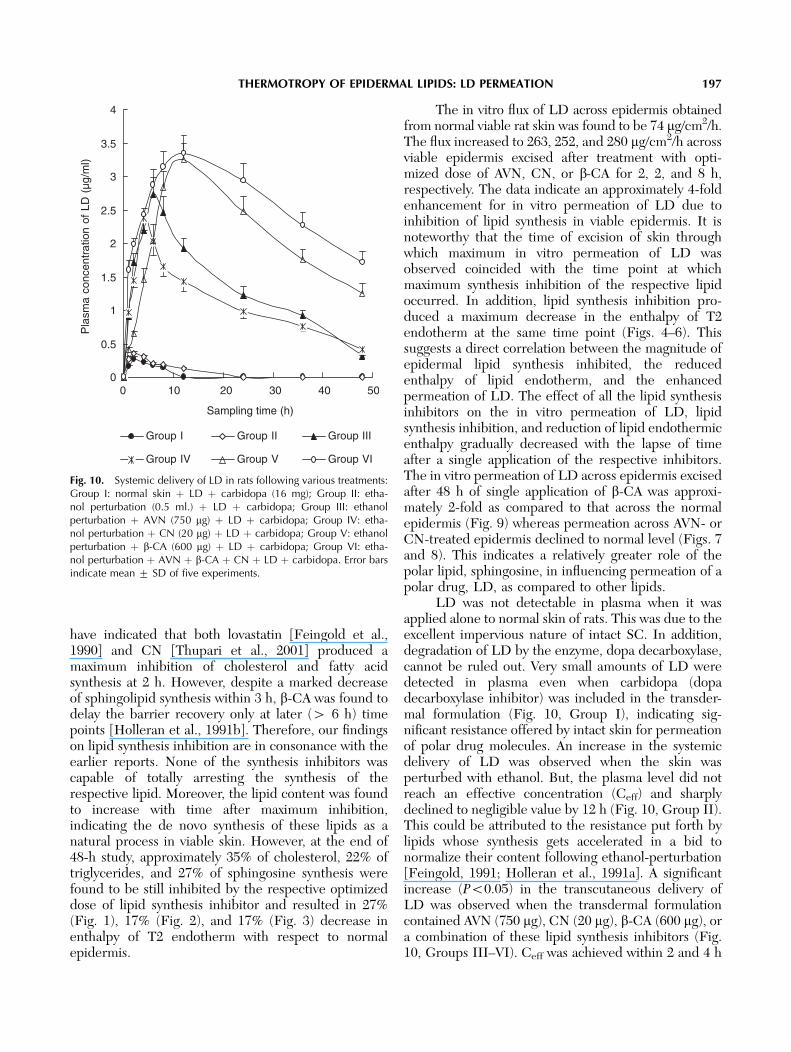
LD was not detectable in plasma when it wasapplied alone to normal skin of rats. This was due to theexcellent impervious nature of intact SC. In addition,degradation of LD by the enzyme, dopa decarboxylase,cannot be ruled out. Very small amounts of LD weredetected in plasma even when carbidopa (dopadecarboxylase inhibitor) was included in the transder-mal formulation (Fig. 10, Group I), indicating sig-nificant resistance offered by intact skin for permeationof polar drug molecules. An increase in the systemicdelivery of LD was observed when the skin wasperturbed with ethanol. But, the plasma level did notreach an effective concentration (Ceff) and sharplydeclined to negligible value by 12 h (Fig. 10, Group II).This could be attributed to the resistance put forth bylipids whose synthesis gets accelerated in a bid tonormalize their content following ethanol-perturbation[Feingold, 1991; Holleran et al., 1991a]. A significantincrease (Po0.05) in the transcutaneous delivery ofLD was observed when the transdermal formulationcontained AVN (750 mg), CN (20 mg), b-CA (600 mg), ora combination of these lipid synthesis inhibitors (Fig.10, Groups III–VI). Ceff was achieved within 2 and 4 h
0
0.5
1
1.5
2
2.5
3
3.5
4
0 10 20 30 40 50
Sampling time (h)
Pla
sma
conc
entr
atio
n of
LD
(µg
/ml)
Group I Group II Group III
Group IV Group V Group VI
Fig. 10. Systemic delivery of LD in rats following various treatments:Group I: normal skin þ LD þ carbidopa (16 mg); Group II: etha-nol perturbation (0.5 ml.) þ LD þ carbidopa; Group III: ethanolperturbation þ AVN (750 mg) þ LD þ carbidopa; Group IV: etha-nol perturbation þ CN (20 mg) þ LD þ carbidopa; Group V: ethanolperturbation þ b-CA (600 mg) þ LD þ carbidopa; Group VI: etha-nol perturbation þ AVN þ b-CA þ CN þ LD þ carbidopa. Error barsindicate mean 7 SD of five experiments.
THERMOTROPY OF EPIDERMAL LIPIDS: LD PERMEATION 197

by application of AVN and CN, respectively. Althoughthe time required to achieve Ceff by application of b-CA was 6 h, the plasma concentration was maintainedabove Ceff for 36 h as against 12 and 8 h after treatmentof skin with AVN and CN, respectively.
The slope of the pre-elimination phase aftertreating the skin with different lipid synthesis inhibitorswas calculated from the systemic delivery profile of LD(Fig. 10). These slopes followed the order: CN4AVN¼b-CA4combination of inhibitors (Po0.05). It is im-portant to note that CN treatment could maintaineffective plasma concentration for only 10 h despiteexhibiting the highest transcutaneous delivery rate ofLD. On the other hand, although, b-CA treatmentexhibited a lower transcutaneous delivery rate, theeffective plasma concentration was maintained over 36h. Further, the combination of these lipid synthesisinhibitors maintained Ceff over 48 h. Hence, together theresults suggest the dominating role of sphingosinesynthesis inhibition in the transcutaneous delivery of LD.
The literature does not reveal much work on fattyacid synthesis inhibition by CN. CN has been reportedto produce more than 50% inhibition of fatty acidsynthesis in MCF-7 human breast cancer cells within 2 h[Thupari et al., 2001]. Another inhibitor of fatty acidsynthesis, 5-(tetradecyloxy)-2-furancarboxylic acid(TOFA), has been reported to exhibit its maximuminhibitory effect within 1–2 h [Halvorson and McCune,1984]. Therefore, fatty acid synthesis has beenimplicated in the initial phase of skin recovery.Similarly, cholesterol synthesis has been reported toinfluence the initial phase of epidermal barrier repair[Feingold et al., 1990]. However, sphingosine synthesishas been reported to be important in the later phase ofepidermal barrier recovery. The inhibition of SPTenzyme activity after skin perturbation has beenreported to start at 30 min and complete normalizationof sphingosine content occurs only by 18 h by a dose of400mg/7 cm2 of b-CA [Holleran et al., 1991b]. There-fore, maintenance of plasma concentration above Ceff
for longer duration after application of b-CA seems tobe due to its overwhelming influence on sphingosinesynthesis inhibition and does not seem to depend onthe transcutaneous delivery rate of a polar molecule,LD. Results of the present investigation revealed adirect correlation between synthesis inhibition ofrespective lipids, lipid endothermic enthalpy decrease,and in vitro permeation of LD. Application of acombination of optimized dose of all the three lipidsynthesis inhibitors reduced the TL to 1 h andmaintained the plasma drug concentration above Ceff
over 48 h. Hence, single transdermal patch containingall three inhibitors seems to offer a non-invasiveapproach for transcutaneous delivery of levodopa.
REFERENCES
Babita, Gupta S, Tiwary AK. 2002. Role of sphingosine synthesisinhibition in transcutaneous delivery of levodopa. Int J Pharm238:43–50.
Barry BW. 1987. Mode of action of penetration enhancers in humanskin. J Control Release 6:85–97.
Behne M, Uchida Y, Seki T, Ortiz de Montellano P, Elias PM,Holleran WM. 2000. Omega-hydroxyceramide are required forcorneocyte lipid envelope formation and normal epidermalpermeability barrier function. J Invest Dermatol 114:185–192.
Bouwstra JA, Peschier L.JC, Brussee J, Bodde HE. 1989. Effect ofn-alkyl azocycloheptan-2-ones including azones on the thermalbehaviour of human stratum corneum. Int J Pharm 2:47–54.
Bouwstra JA, Gooris GS, Salomons-de Vries MA, van der Spek JA,Bras W. 1992. Structure of human stratum corneum as a functionof temperature and hydration: a wide angle X-ray diffractionstudy. Int J Pharm 84:205–216.
Cornwell PA, Barry BW, Bouwstra JA, Gooris GS. 1996. Mode ofaction of terpene penetration enhancers in human skin;differential scanning calorimetry, small X-ray diffraction andenhancer uptake studies. Int J Pharm 127:9–26.
Elias PM, Menon GK. 1991. Structural and lipid biochemicalcorrelates of the epidermal permeability barrier. Adv Lipid Res24:1–26.
Feingold KR. 1991. The regulation and role of lipid synthesis. AdvLipid Res 24:57–82.
Feingold KR, Mao-Qiang M, Menon GK, Cho SS, Brown BE, EliasPM. 1990. Cholesterol synthesis is required for cutaneous barrierfunction in mice. J Clin Invest 86:1738–1745.
Gamez R, Hagel RB, MacMullan EA. 1976. Analytical profile oflevodopa. Analytical Profile of Drug Substance 5:189–223.
Ghanem AH, Mahmoud H, Higuchi WI,Liu P, Good WR. 1992. Theeffects of ethanol transport of lipophilic and polar permeantsacross hairless mouse skin: methods/validation of a novelapproach. Int J Pharm 78:137–156.
Golden GM, Guzek DB, Harris RR, McKie JE, Potts RO. 1986.Lipid thermotropic transitions in human stratum corneum. J ClinInvest 86: 255–259
Gupta M, Mahajan A, Babita, Gupta S, Tiwary AK. 2003. Inhibitionof skin sphingosine synthesis: enhanced percutaneous permeationof 5-fluorouracil. Pharmazie 59:212–216.
Halvorson DL, McCune SA. 1984. Inhibition of fatty acid synthesisin isolated adipocytes by 5-(tetradecyloxy)-2-furoic acid. Lipids19:851–856.
Harder S, Bass H, Rietbrock S. 1995. Concentration effectrelationship of levodopa in patients with Parkinson’s disease. ClinPharmacokinet 29:243–256.
Holleran WM, Feingold KR, Mao-Quing M, Gao WN, Lee JM,Elias PM. 1991a. Regulation of epidermal sphingolipidsynthesis by permeability barrier function. J Lipid Res 32:1151–1158.
Holleran WM, Mao-Quing M, Gao WN, Menon GK, Elias PM.1991b. Sphingolipid are required for mammalian epidermalbarrier function: Inhibition of sphingolipid synthesis delaysbarrier recovery after acute perturbation. J Clin Invest 88:1338–1345.
Krill SL, Knutson K, Higuchi WI. 1992. Ethanol effects on thestratum corneum lipid phase behaviour. Biochim Biophys Acta112:273–280.
198 BABITA ET AL.

Leopold SC, Lippold CB. 1995. An attempt to clarify themechanism of penetration enhancing effect of lipophilic vehicleswith differential scanning calorimetry (DSC). J Pharm Pharmacol47:276–281.
Medlock KA, Merrill AH. 1988. Inhibition of serine palmitoyl-transferase in vitro and long chain base biosynthesis in intactchinese hamster ovary cell by beta chloroalanine. Biochem27:7079–7084.
Ottey KA, Wood LC, Grunfeld C, Elias PM, Feingold KR. 1995.Cutaneous permeability barrier disruption increases fatty acidsynthesis enzyme activity in the epidermis of hairless mice.J Invest Dermatol 104:401–404.
Phillips BB. 1997. Atorvastatin: a review of its pharmacology andplace in therapy. Am J Cardiol 80:39–44.
Robertson DRC, Wood ND, Everest H, Monks K, Waller DG,Renwick AG, George CF. 1989. The effects of age on thepharmacokinetics of levodopa administered alone and in thepresence of carbidopa. Br J Clin Pharmacol 28:61–69.
Thupari JN, Pinn ML, Kuhajda FP. 2001. Fatty acid synthaseinhibition in human breast cancer cells leads to Malonyl CoA-induced inhibition of fatty acid oxidation and cytotoxicity.Biochem Biophys Res Commun 285:217–223.
Tsai JC, Guy RH, Thornfeldt CR, Gao WN, Feingold KR, Elias PM.1996. Metabolic approaches to enhance transdermal drug delivery.1. Effect of lipid synthesis inhibitors. J Pharm Sci 85:643–648.
Vance D, Goldberg I, Mitsuhashi O, Bloch K. 1972. Inhibition offatty acid synthetases by the antibiotic cerulenin. BiochemBiophys Res Commun 48:649–656.
THERMOTROPY OF EPIDERMAL LIPIDS: LD PERMEATION 199





























