Embed Size (px)
Citation preview

Resources, Conservation and Recycling, 1 (1988) 185-195 185 Elsevier Science Publishers B.V./Pergamon Press plc - - Printed in The Netherlands
Enzymatic Degradat ion of Low-Rank Coals by a Cell-free Enzymatic System from Coriolus Versicolor
JOHN W. PYNE*, DOROTHY L. STEWART, JOHN C. LINEHAN, ROGER M. BEAN, MATHEW A. P0WELL, RICHARD B. LUCKE, BERTA L. THOMAS, JAMES A. CAMPBELL and BARY W. WILSON
Battelle Pacific Northwest Laboratories, Richland, WA 99352 (U.S.A.)**
(Received October 26, 1987; accepted in revised form May 9, 1988)
ABSTRACT
An extracellular fraction from the lignin degrading white-rot fungus, Coriolus versicolor, can biosolubilize leonardite and pretreated lignites in vitro. Leonardite is often found as an overburden to lignite deposits, and is related to humic acid, and is more oxidized than lignites. Extracts from certain fungi such as C. versicolor solubilize leonardite. The biosolubilization activity is at least in part enzymatic in nature. A protein which is at least partly responsible for the solubilization effect has been substantially purified and concentrated by ultrafiltration and ion-exchange chro- matography from the broth of an agitated submerged culture of C. versicolor grown in a defined medium. An in vitro coal biosolubilization assay was used to establish the effects of various phys- ical and chemical factors affecting enzyme activity, including temperature and the salt and buffer composition of the reaction mixture. Some initial studies on the mechanism of the enzymatic leonardite biosolubilization reaction are described.
INTRODUCTION
Cohen and Gabriele [ 1 ] found tha t leonardite particles placed on a mat of white-rot fungi grown on an agar medium were converted to black droplets. Leonardite somewhat resembles lignite and may resemble lignin as much as, or more than, lignites do. Leonardite is therefore a useful substrate for screen- ing microorganisms for their ability to degrade low-rank coals since it appears to be between lignin and low-rank coals in oxidation level. The carbonaceous product formed from leonardite by the action of C. versicolor has been partially characterized by Wilson et al. [2].
A clue to how white-rot fungi degrade leonardite can be gleaned from a con- sideration of the context in which white-rot fungi operate. White-rot fungi are
*To whom correspondence should be addressed. Present address is Cyanotech, 18748 142nd street N.E., Woodinville, WA 98072 (U.S.A.). Tel. (206) 481-2173. **Research funded by Electric Power Research Institute, Palo Alto, CA.
0921-3449/88/$03.50 © 1988 Elsevier Science Publishers B.V./Pergamon Press plc

186
prominent degraders of lignin which is thought to be the precursor of low-rank coals. Lignin is a rigid three-dimensional polymer produced by the dehydro- genative polymerization of the three cinnamyl alcohols: coniferyl, sinapyl, or coumaryl alcohols. Depending on the plant lignin being formed, one, two, or all three of these monomers are used to form lignin. The polymerized product grows by a free radical mechanism that produces a variety of bonds between monomeric units in which the fl-arylether is particularly important [3 ]. When lignin is degraded by white-rot fungi, a number of degradation products can be detected. Examples include guaiacylglycerol-fl-coniferyl ether, p-coumarate, p-hydroxybenzoate, ferulate, vanillate, guaiacylglycerol, syringate, vanillin, 3- methoxy-4-hydroxyphenylpyruvate, 3-o-methylgallate, gallate, and protoca- techuate. The general direction of the reaction proceeds from lignin to mono- meric products which have been demethoxylated so that the aromatic rings have free hydroxyl group substituents. The cells can further metabolize di- phenols by oxidative ring cleavage and fragmentation to yield compounds that can be used by the cells for growth and energy metabolism. During the break- down of lignin by white-rot fungi, a quinone type structure is formed. The quinones will repolymerize unless they are reduced. Carbohydrates are used as a source of reducing potential to convert quinones to diphenols [4].
Lignin breakdown is catalyzed by enzymes that convert the polymer lignin structure into the small molecular weight material that can be used more di- rectly by the cells. Evidence exists showing that a heme containing peroxidase from Phanerochaete chrysosporium can degrade lignin in vitro [5]. This en- zyme requires hydrogen peroxide as an electron acceptor for oxidizing the pro- tein. A model substate that the lignin peroxidase from P. chryosporium can oxidize is veratryl alcohol. Veratryl alcohol is oxidized to water and the corre- sponding aldehyde. A second ligninolytic enzyme system is that of laccase com- bined with glucose oxidase [6]. Laccase is thought to be a polyphenol oxidase, which means that the substrate must have free phenol groups for reactivity. Laccase is a copper-containing enzyme that uses oxygen as the final electron acceptor. A model substrate used to assay for laccases is syringaldazine. The products of laccase oxidation are generally quinones that have a tendency to polymerize. Syringaldazine is designed so that the product quinone has a re- duced tendency to polymerize and is thus useful for enzyme activities. The substrates syringaldazine and leonardite have been used to isolate an enzyme from white-rot fungi that will degrade low-rank coals.
Some properties of the enzyme that solubilize low-rank coals will be pre- sented. This paper will be divided into the following sections: (1) characteri- zation of an extracellular leonardite solubilizing activity, (2) product characterization, and (3) model compounds studies. Hopefully, the informa- tion presented will provide the beginnings of a picture of what Coriolus versi- color does to low-rank coals to increase their water solubility.

187
MATERIALS AND METHODS
Preparation of the enzyme [7]. The fungus was grown in a Chemap CF-20 fermentor at 25°C, and was supplied with 4 L of filtered air per minute at atmospheric pressure to 15 L of growth medium. A two stage Rushton turbine (6 blade) agitator system was used at a speed of 400 rpm. The inoculum used for the fermentation was three 50 mL cultures of Coriolus versicolor grown on Sabourad-maltose broth for 7 to 10 days at 25 ° C with no agitation. The fungi grown under these conditions form a mycelial mat. The mats were transferred to a stoppered vessel containing 250 mL distilled water and 50 mL of 3 mm glass beads. This vessel was shaken vigorously and the mycelial fragments were transferred to the fermentor. After three days growth in the fermentor, 0.3 ml of 2,5-xylidine (Aldrich Chemical Col, Milwaukee, WI ) was added to increase levels of extracellular laccase.
The extracellular fluid formed during the growth of C. versicolor was sepa- rated from the cell mass by filtration through several layers of cheesecloth. This fluid is termed the cell-free filtrate. The resultant fluid was then filtered through an ultrafiltration membrane with a molecular weight cutoff of 100,000 (Amicon H5P10-43). The higher molecular weight material in aqueous fluid was concentrated using an ultrafiltration membrane with a molecular weight cutoff of 10,000 (Amicon H5P10-43). The resultant fluid further washed (di- afiltered) with three 300 mL portions of water while being continually passed through the H5P10-43 membrane to maintain a constant volume of 1 L. The concentration and diafiltration steps caused precipitate formation which was removed by either filtration or centrifugation. The concentrated extracellular fluid was then chromatographed on a 2 cm X 30 cm DEAE-cellulose (Amicon cellufine DEAE-AM). The column had previously been equilibrated to 0.01 M sodium phosphate, pH 7.0 buffer). The final material is the cell-free enzyme.
Model compound preparation. Stock solutions of model compounds were pre- pared at 1 mg/mL in methanol. Benzylbenzoate and methoxybenzophenone were introduced (100 ]~L of the stock solution) to a 50:50 mixture of the cell- free enzyme and a buffer solution of pH 5.2. After equilibration with the en- zyme and buffer for one day, the solution was extracted three times with equal volumes of chloroform. The extracts were combined and then concentrated to 1 mL for analysis by gas chromatography (GC) and gas chromatography/mass spectrometry (GC/MS). Controls were made by adding 100 pL of the stock solution of the model compound to the buffer solution and extracting after one day. The controls were also analyzed by GC.
Extraction efficiencies were determined by GC analysis of a solution of the model compound in the buffer with no cell-free enzyme present. The peak areas obtained were then compared with the 100 ppm standard to determine the efficiency of extraction and possible reaction of the model compound with

188
the medium. The results indicated a 90-95% extraction efficiency and no deg- radation of the model compound by the buffer solution.
Analytical Instrumentation. A Hewlett-Packard 5880 gas chromatograph with a flame ionization detector was used to determine the amount of degra- dation for each model compound. The GC was equipped with a DB-5 30 m column and appropriate flow rates were utilized as recommended by the man- ufacturer. The temperature program used was as follows: 50 °C for 1 minute, 50-250°C at 8 °/min, and 250°C for 10 minutes. The amount of degradation was determined by comparing the peak area of the degraded model compound and the peak area of the standard. The temperature of the injection port was set at 275 ° C. A HP 5985 GC/MS/DS operated in the electron impact ioniza- tion mode was used to identify specific degradation products. The same column and temperature program were used as discussed above.
RESULTS
Characterization of the cell-free enzyme activity
Screening studies have shown that P. chrysosporium has a marginal ability to solubilize low-rank coals. C. versicolor, on the other hand, consistently sol- ubilizes leonardite. C. versicolor culture filtrates solubilize leonardite in vitro. These culture filtrates contain a laccase-like activity, which means that syr- ingaldazine is oxidized without added hydrogen peroxide, and furthermore, the addition of hydrogen peroxide does not enhance the syringaldazine activity. The lignin peroxidase from P. chrysosporium can oxidize veratryl alcohol to the aldehyde [5 ]. It has not been possible to detect veratryl alcohol peroxidase activity in cell-free extracts from C. versicolor.
The cell-free activity from C. versicolor has been further characterized. Pyne et al. [ 7 ] have established several lines of evidence suggesting that a protein is at least in part responsible for the cell-free activity. These workers found that the solubilizing activity of syringaldazine oxidase and the leonardite were sim- ilar in gel permeation and DEAE-cellulose ion-exchange column chromato- graphy experiments. The characterization of the leonardite solubilizing activity has been continued. The enzyme fraction described previously [7] appeared to be homogeneous by DEAE-cellulose chromatography because the ratio of syringaldazine oxidase activity to the absorbance at 280 nm was constant through the protein peak.
When this fraction was subjected to SDS polyacrylamide gel electrophoresis, it appeared homogeneous at normal loading levels. However, when the gel was overloaded, a minor band could be observed. The two proteins have nearly identical mobilities and were difficult to resolve. It was found that syringal- dazine oxidase could be resolved by chromatography on a DEAE-cellulose col- umn using an ammonium bicarbonate concentration gradient. The two peaks

189
(IIA and IIB) were pooled, but the overlapping fractions were not included in either of these two pooled fractions. The two pools were then lyophilized to remove ammonium bicarbonate and concentrate the proteins. After this chro- matographic step, protein-containing fractions that were apparently homo- geneous by polyacrylamide gel electrophoresis were obtained. After redissolving the separately lyophilized fractions in a small volume of 0.01 M sodium phos- phate buffer, pH 5.2, the two fractions were assayed for syringaldazine oxidase and leonardite solubilizing activities. Only the second syringaldazine oxidase peak (IIB ) had much leonardite solubilizing activity. The ratio of leonardite solubilization to syringaldazine oxidase was 50 times smaller for peak IIA as compared to peak IIB. Another syringaldazine oxidase activity (but with little leonardite solubilizing activity) that was separated in an earlier preparatory step in the purification scheme (peak I ) had a ratio of leonardite solubilization to syringaldazine oxidase activity about 100 times smaller than peak IIB. Al- though the purified fraction still retained syringaldazine oxidase activity, it was yellow instead of the blue color that would be expected for a copper con- taining laccase. The protein solution used for chromatography is blue colored, and preliminary work suggests that the syringaldazine oxidase recovered from the ammonium bicarbonate column does contain copper. The possibilities that some copper atoms that are not essential to activity are removed during the isolation procedure, or that the isolation procedure changes the oxidation state of the resting enzyme, are being examined.
Characterization of the bioproduct
The molecular weight of the black, water soluble product formed from the action of C. versicolor on leonardite has been measured by ultrafiltration and osmometry. The average molecular weight is about 50,000 by ultrafiltration and about 340 by osmometry (2). Because of the disparity between these val- ues, another method of size estimation was tried. The method chosen was gel permeation chromatography with two/l-Styragel columns (10 nm and 50 nm in series) using tetrahydrofuran as the eluting solvent. Two methods of detec- tion were used; one was absorbance at 254 nm, and the other a mass detector. Figure 1 shows the elution profile of the base soluble material from leonardite. Leonardites have various amounts of base soluble material that is presumably humic acid. The leonardite sample used in these experiments is about 60% soluble in 10% NaOH. The molecular weight (MW) range of the humic acid fraction is between 17,000 and 200 daltons with a weighted average MW of 1,500 daltons (Fig. 1 ). Figure 2 shows the gel permeation profile of the product formed from leonardite by the action of C. versicolor. The elution profile of this material is biphasic. The large peak at around 300 daltons appears in samples that contain phosphate. It is not clear whether this peak is an artifact, phos- phate, or a phosphate-containing compound. The average molecular weight of
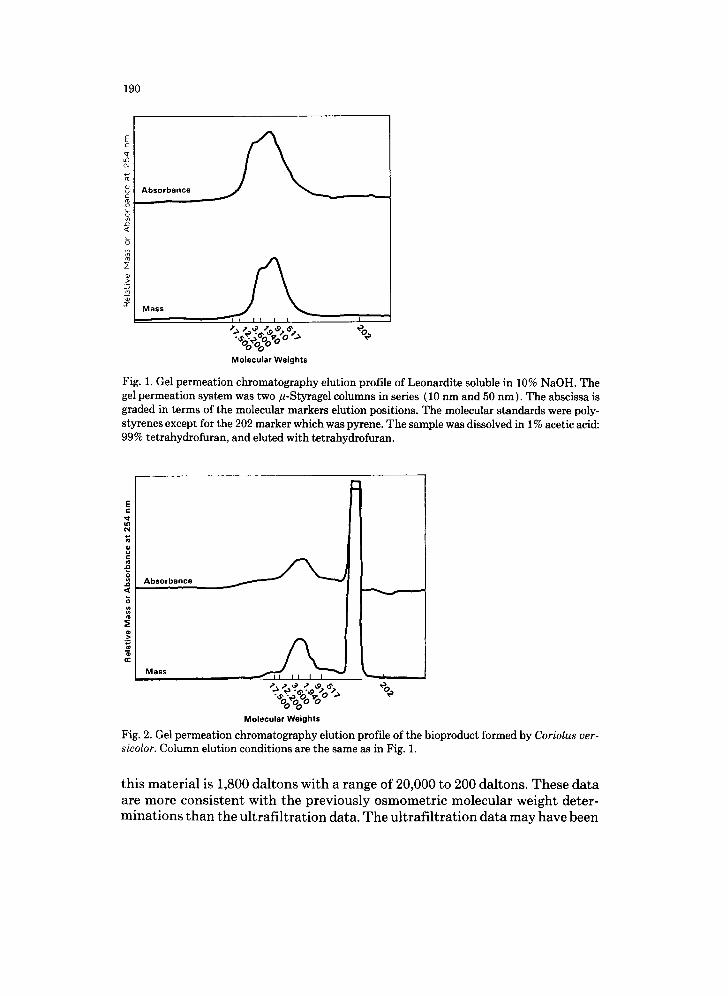
190
E / L c
¢q
Absorbance c
,<
5 M a s s
I I I I I I I
~ ~ ~, o:.<;o "., % %0o0 Molecular Weights
Fig. 1. Gel permeation chromatography elution profile of Leonardite soluble in 10% NaOH. The gel permeation system was two #-Styragel columns in series (10 nm and 50 nm). The abscissa is graded in terms of the molecular markers elution positions. The molecular standards were poly- styrenes except for the 202 marker which was pyrene. The sample was dissolved in 1% acetic acid: 99% tetrahydrofuran, and eluted with tetrahydrofuran.
E c
N
Absorbance
M a s s ~ , ii
Molecular Weights
Fig. 2. Gel permeation chromatography elution profile of the bioproduct formed by Coriolus ver- sicolor. Column elution conditions are the same as in Fig. 1.
this material is 1,800 daltons with a range of 20,000 to 200 daltons. These data are more consistent with the previously osmometric molecular weight deter- minations than the ultrafi l tration data. The ultrafil tration data may have been
191
artifactual because of the tendency of the bioproduct to stick to surfaces. One characteristic of the bioproduct is its tendency to be precipitated by
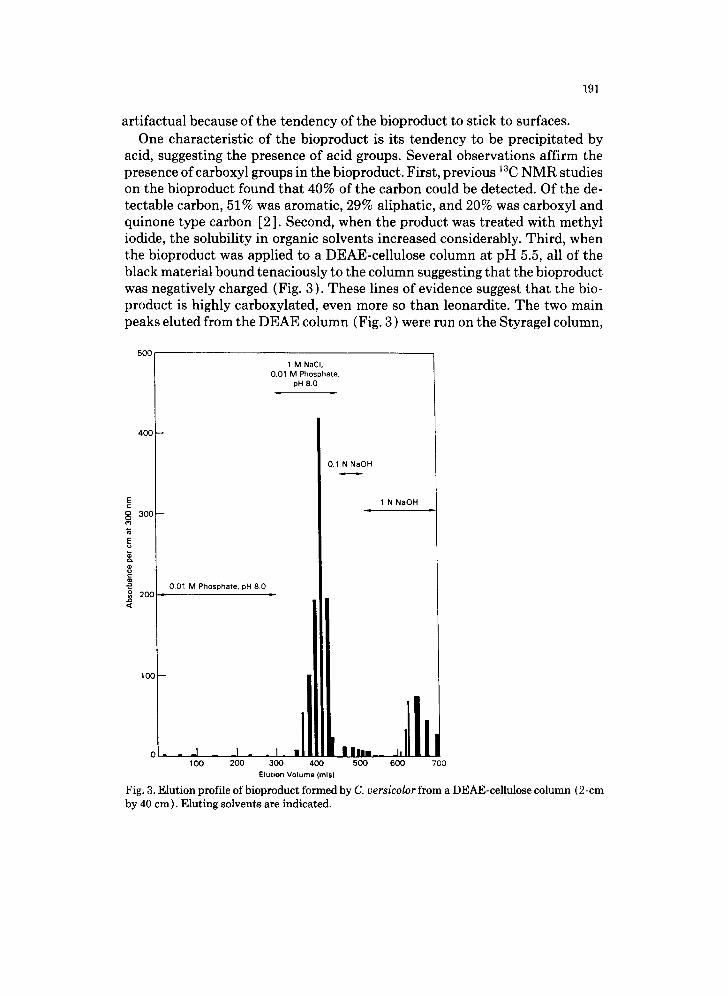
acid, suggesting the presence of acid groups. Several observations affirm the presence of carboxyl groups in the bioproduct. First, previous 13C NMR studies on the bioproduct found that 40% of the carbon could be detected. Of the de- tectable carbon, 51% was aromatic, 29% aliphatic, and 20% was carboxyl and quinone type carbon [2]. Second, when the product was treated with methyl iodide, the solubility in organic solvents increased considerably. Third, when the bioproduct was applied to a DEAE-cellulose column at pH 5.5, all of the black material bound tenaciously to the column suggesting that the bioproduct was negatively charged (Fig. 3). These lines of evidence suggest that the bio- product is highly carboxylated, even more so than leonardite. The two main peaks eluted from the DEAE column (Fig. 3 ) were run on the Styragel column,
5OO
1 M NaCI, 0.01 M Phosphate.
pH 8.0
4 0 0 - -
0.1 N NaOH
E 1 N NaOH
3 0 0 -
001 M Phosphate, pH 8.0 o 20C i
nit
,llll 11 I 0 I -- -- --I I --I -- . I. I Ini,i__ J, •
Elulion Volume (mls)
Fig. 3. Elution profile of bioproduct formed by C. versicolor from a DEAE-cellulose column (2-cm by 40 cm). Eluting solvents are indicated.
192
E ==L
c
L
"~ Abso rbance
I[
- / Mass
i i I I I
OoOoO o Molecu la r We ights
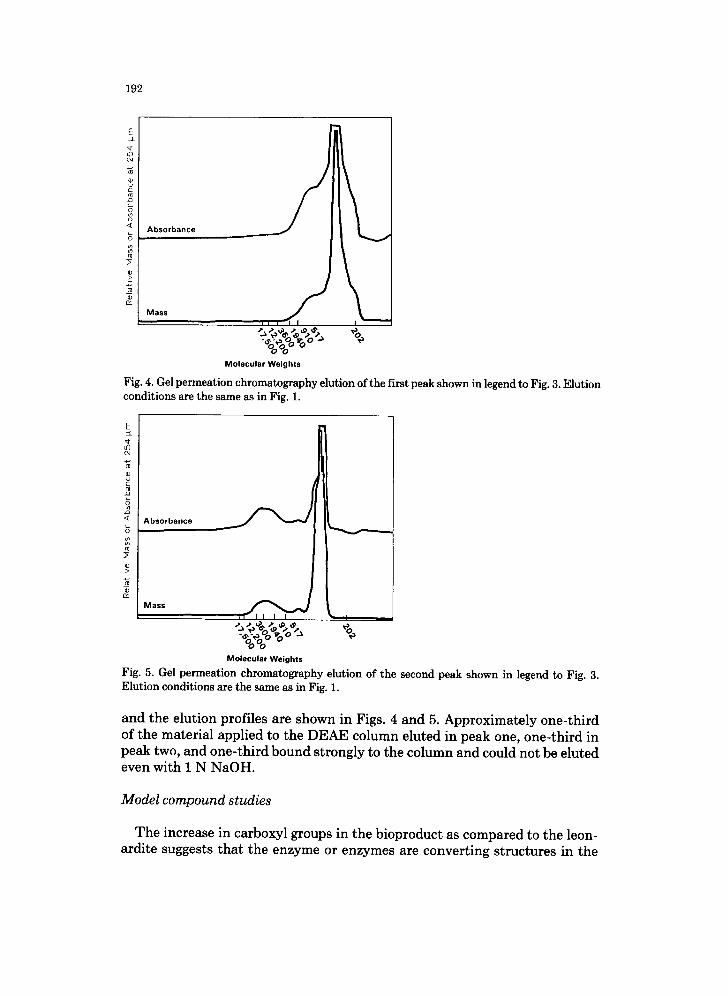
Fig. 4. Gel permeation chromatography elution of the first peak shown in legend to Fig. 3. Elution conditions are the same as in Fig. 1.
E =:k *¢
o
<~ A b s o r b a n c e
M a s s
~l I I I I I
- ;?~ °o . - O ~ % ooo ° o
Molecular Weights
Fig. 5. Gel permeation chromatography elution of the second peak shown in legend to Fig. 3. E l u t i o n c o n d i t i o n s are t he s a m e as in Fig. 1.
and the elution profiles are shown in Figs. 4 and 5. Approximately one-third of the material applied to the DEAE column eluted in peak one, one-third in peak two, and one-third bound strongly to the column and could not be eluted even with 1 N NaOH.
Model compound studies
The increase in carboxyl groups in the bioproduct as compared to the leon- ardite suggests tha t the enzyme or enzymes are converting structures in the

193
(a) 0=[~/_ @ 26.72 (b) 26.71
Y OCHa A = 1700
A = 1800
Fig. 6. Gas chromatogram of: (a) 100 ppm standard of methoxybenzophenone, and (b) after degradation by the cell-free enzyme preparation from C. versicolor.
(a) 0 ( , , . , ,
A = 1400
A
Fig. 7. Gas chromatogram of: (a) 100 ppm standard of benzylbenzoate, and (b) after degradation by a cell-free enzyme preparation from C. versicolor.
leonardite to carboxyl moieties. Chung has suggested that lignite consists of aromatic rings hung together by oxygenated linkages such as esters, carbonyl, and ether groups [8]. Two model compounds were chosen, methoxybenzo- phenone and benzylbenzoate, to test the hypothesis that the purified enzyme is responsible for the conversion of leonardite to a more carboxylated product. Figure 6 shows the GC profile of methoxybenzophenone before and after con- tact with the purified enzyme. There was almost a 5% degradation of the meth- oxybenzophenone after a 20-hour incubation period at room temperature. One

194
of the products appears to be an organic acid, although its exact identity is not known at this time. Figure 7 shows the GC profile of the other model com- pound, benzyl benzoate. In the product profile, the area of the starting material that is labeled "A" is reduced to about half of the area of the unreacted mate- rial, meaning that half has reacted. The large peak in the elution profile labeled "B" is benzyl alcohol. Several small miscellaneous peaks labeled "C" appear to be organic acids but have not as yet been identified. The suggested cleavage site in the case of both model compounds is between the carbon and oxygen bond. This is next to the carbonyl group and apparently produces a free alcohol or phenol and an organic acid. The results with both model compounds suggest tha t the enzyme can at tack a substrate tha t does not contain a free phenol group.
SUMMARY
It has been possible to purify an enzymatic fraction from a cell-free extract of C. versicolor that can solubilize leonardite in vitro. This enzyme is one of three syringaldazine oxidase activities detectable in cell-free extracts. The en- zyme is yellow colored and does not have a laccase-like UV-Vis spectrum. The bioproduct produced from leonardite by C. versicolor is a highly carboxylated material. The action of the purified enzyme on model compounds suggests tha t it may be responsible for the increase in the amount of carboxyl groups seen in the product. The action of the enzyme on the model also suggests that it is not strictly a polyphenol oxidase.
The enzyme from Coriolus has advantages over the ligninases from Phaner- ochaete. One advantage is that the Phanerochaete ligninase is a peroxidase requiring added hydrogen peroxide, whereas the C. versicolor enzyme requires oxygen which is much cheaper to supply in engineering quantities. The C. ver- sicolor enzyme can be easily produced in the submerged culture of the orga- nism. Also, since a single gene product is apparently required for activity and no expensive cofactors are required, then the protein could potentially be pro- duced by genetic engineering techniques in large quantities to reduce costs further.
REFERENCES
1 Cohen, M.S. and Grabiele, P.D., 1982. Degradation of coal by the fungi Polyporus versicolor and Poria monticola. Appl. Environ. Microbiol., 44: 23.
2 Wilson, B.W., Bean, R.M., Franz, J.A., Thomas, B., Cohen, M.S., Aronson, H. and Gray, E.T., 1987. Microbial conversion of low-rank coal: characterization of biodegraded product. J. En- ergy Fuels, 1: 80.
3 Higuchi, T., 1980. Lignin Biodegradation: Microbiology, Chemistry and Potential Applica- tions. Vol. 1, Chap. 1. T. Kirk, T. Higuchi and H. Chang (Eds.)., CRC Press, Boca Raton, FL.

195
4 Westermark, U. and Eriksson, K.E., 1974. Carbohydrate-dependent enzymic quinone reduc- tion during lignin degradation. Acta Chem. Scan., B208: 209.
5 Tien, M. and Kirk, T.K., 1984. Lignin-degrading enzyme from Phanerochaete chrysosporium: purification, characterization and catalytic properties of a unique H202-requiring oxygenase. Proc. Nat. Acad. Sci. (U.S.A.), 81: 2280.
6 Greene, T.R., 1977. Significance of glucose oxidase in lignin degradation. Nature, 268: 78. 7 Pyne, J.W., Stewart, D.L., Fredrickson, J. and Wilson, B.W., 1987. Solubilization of Leonar-
dite by a cell-free extract of Coriolus versicolor. Appl. Environ. Microbiol., (in press). 8 Chung, K.E. and Goldberg, I.B., 1985. Chemical structure of low-rank coals. Presented at DOE/
FE Direct Liquefaction Contractors' Review Meeting, Pittsburgh, PA.



























