Embed Size (px)
Citation preview

ORIGINAL PAPER
Enzymatic and genetic profiles in environmental strainsgrown on polycyclic aromatic hydrocarbons
Lucia Cavalca Æ Nicoletta Guerrieri ÆMilena Colombo Æ Silvia Pagani ÆVincenza Andreoni
Received: 27 June 2006 / Accepted: 11 September 2006� Springer Science+Business Media B.V. 2006
Abstract The possible generation of oxidative
stress induced by aromatic hydrocarbon degra-
dation suggests that ancillary enzyme activities
could facilitate the utilization of polycyclic
aromatic hydrocarbons as sole carbon source.
To investigate the metabolic profiles of low
molecular weight polycyclic aromatic hydrocar-
bon-degrading strains of Sphingobium chlor-
ophenolicum, Rhodococcus aetherovorans,
Rhodococcus opacus and Mycobacterium
smegmatis, the determination of the activity of
putative detoxifying enzymes (rhodanese-like
and glutathione S-transferase proteins) was
combined with genetic analyses. All the studied
strains were able to utilize phenanthrene or
naphthalene. Glutathione S-transferase activity
was found in S. chlorophenolicum strains grown
on phenanthrene and it was related to the
presence of the bphK gene, since modulation of
glutathione S-transferase activity by phenan-
threne paralleled the induction of glutathione
S-transferase transcript in the S. chlorophenoli-
cum strains. No glutathione S-transferase activ-
ity was detectable in R. aetherovorans,
R. opacus and in M. smegmatis strains. All
strains showed 3-mercaptopyruvate:cyanide sul-
furtransferase activity. A rhodanese-like SseA
protein was immunodetected in R. aetherovo-
rans, R. opacus and in M. smegmatis strains,
where increase of 3-mercaptopyruvate:cyanide
sulfurtransferase activity was significantly in-
duced by growth on phenanthrene.
Keywords Phenanthrene � PAH � Glutathione
S-transferase � Rhodanese-like enzymes �Catabolic genes � bphK gene expression
AbbreviationsbphK gene for glutathione S-transferase
CDNB 1,dichloro-2,4-nitrobenzene
GST glutathione S-transferase
MST 3-mercaptopyruvate:cyanide
sulfurtransferase
PAHs polycyclic aromatic hydrocarbons
phnA1 gene for 3,4-phenanthrene dioxygenase
ROS reactive oxygen species
TST thiosulfate:cyanide sulfurtransferase
xylE 2,3-catechol dioxygenase
xylX ring hydroxylating dioxygenase
narAa naphthalene dioxygenase
Sequence accession numbers: narAa of 1BN was underthe GeneBank acc. num.: AJ401612.
L. Cavalca � M. Colombo � V. AndreoniDipartimento di Science e Tecnologie, Alimentari eMicrobiologiche, Universita degli Studi di Milano,Via Celoria 2, 20133 Milano, Italy
N. Guerrieri � S. Pagani (&)Dipartimento di Scienze Molecolari Agroalimentari,Universita degli Studi di Milano, Via Celoria 2, 20133Milano, Italye-mail: [email protected]
123
Antonie van Leeuwenhoek
DOI 10.1007/s10482-006-9119-1

Introduction
Polycyclic aromatic hydrocarbons (PAHs) are
ubiquitous environmental pollutants, that are
considered priority pollutants on the basis of
their toxicity, carcinogenicity and recalcitrance in
the environment (Kanaly and Harayama 2000).
The biodegradation of low-molecular-weight
PAHs has been studied with Gram-negative and
Gram-positive species and attention has also been
tuned to genes involved in their catabolism (Kim
and Zylstra 1999; Pinyakong et al. 2003a; Saito
et al. 1999). Genes coding for putative glutathione
S-transferases (GSTs) have been found within
degradative operons either of monoaromatic
hydrocarbons or PAHs (Bartels et al. 1999;
Favaloro et al. 2000; Xia et al. 2005).
The picture emerging from bacterial genome
sequences is that many bacteria contain large sets
of GST genes of widely divergent sequences.
Although their function, structure and regulation
are still not completely understood (Vuilleumier
and Pagni 2002), their involvement in the bio-
degradation of xenobiotics and/or in detoxifica-
tion from reactive oxygen species (ROS) was
claimed (Nagata et al. 1999). It was reported that
some GSTs show peroxidase activity and partic-
ipate in epoxide hydrolysis and dehalogenation
(Van Hylckama et al. 2000). Chavez et al. (2004)
proved that the growth of bacteria on polychlo-
rinated biphenyls led to a generation of oxidative
stress. ROS and other radicals were also found to
be produced in Ochrobactrum anthropi grown on
aromatic compounds, as a consequence of activ-
ities of either monoxygenases or dioxygenases
(Tamburro et al. 2004). In this respect, enzymes
with putative scavenger activities could help the
cell to overcome xenobiotic-induced stress. In
addition to GSTs, enzymes that could likely be
involved in detoxification processes should be
members of the rhodanese protein homology
superfamily (Accession number: PF00581; http://
www.sanger.ac.uk/Software/Pfam). They are
ubiquitous enzymes displaying sequence homol-
ogy with bovine rhodanese (thiosulfate:cyanide
sulfurtransferase; Ploegman et al. 1978) which
catalyze the in vitro transfer of a sulfur atom from
a suitable sulfur donor (thiosulfate for rhodanes-
es, and 3-mercaptopyruvate for 3-mercaptopyru-
vate:sulfurtransferases) to cyanide, with
concomitant formation of thiocyanate (Westley
1977). Rhodanese modules have been found in
arsenate resistance (Bordo and Bork 2002) and in
stress-related proteins (Adams et al. 2002; Ray
et al. 2000). Up-regulation of genes coding for
rhodanese-like proteins in response to phenol-
induced stress in Pseudomonas putida (Santos
et al. 2004) and in Mycobacterium sp. strain 6PY1
grown on pyrene (Krivobok et al. 2003) was
recently found by proteomic analysis.
In the present study, an investigation aimed to
define enzymatic profiles, as well as the presence
of catabolic genes, was carried out by using
environmental PAH-degrading strains (S. chlor-
ophenolicum, R. aetherovorans, R. opacus and M.
smegmatis). Determination of GST and 3-merca-
ptopyruvate:cyanide sulfurtransferase (MST)
activities after growth on PAHs, as compared to
the activities measured when glucose was the
carbon source, allowed the identification of
strains where these activities were induced selec-
tively by the presence of PAHs. Genetic analyses
revealed that the GST activity found in
S. chlorophenolicum strains was related to the
presence of the bphK gene.
Materials and methods
Chemicals
Benzene, toluene, benzoate (purity >99%) were
purchased from Merck (Darmstadt, Germany);
HPLC grade phenanthrene and naphthalene
(purity >96%), n-hexadecane (purity >99%),
o-phthalate (purity >99.5%), 3-mercaptopyruvate
(purity >90%), 1-dichloro-2,4-nitrobenzene (CD-
NB) (purity >99%) and reduced glutathione
(GSH) (purity >99%) from Sigma-Aldrich Che-
mie (Steinheim, Germany); salicylate (pur-
ity >99.5%) from Fluka Chemie (Buch,
Switzerland).
Antonie van Leeuwenhoek
123

Bacterial strains and culture conditions
Strains were isolated from PAH-contaminated
soils by successive enrichment cultures with
phenanthrene as the only carbon source (Andre-
oni et al. 2004). The growth characteristics of the
isolates on hydrocarbons (Table 1) were studied
in 100 ml flasks containing 20 ml of M9 mineral
medium (Kunz and Chapmann 1981) at 30�C on a
rotary shaker at 150 rpm. Phenanthrene and
naphthalene were dispensed as acetone solutions
sterilized by filtration with a 0.2 lm-pore-size-
membrane filter, at the final concentration of
400 mg l–1. Salicylate, benzoate and o-phthalate
were supplied as aqueous solutions at the final
concentrations of 200 mg l–1. n-Hexadecane, tol-
uene and benzene were supplied as pure com-
pounds at the final concentrations of 200 mg l–1,
the flasks were incubated in separate sealed jars.
Growth was evaluated by determining the tur-
bidity at 600 nm of cultures on a Beckman model
DU 640 spectrophotometer.
Strains were identified on the basis of sequence
analysis of the complete 16S rRNA gene by using
eubacterial universal primers P27f and P1495r
(Table 2) referred to Escherichia coli nucleotide
sequence of 16S rDNA gene, according to the
protocol previously reported (Cavalca et al.
2002). Strains C3R, 1.7, 1.8, 5.1 were identified
as S. chlorophenolicum (ex Sphingomonas flava)
(98% identity to GeneBank acc. num. X87164),
strain F was identified as M. smegmatis (99%
identity to GeneBank acc. num. AY457078),
strains 1.9 and 6.22 were identified as R. aethe-
rovorans (100% identity GeneBank acc. num.
AF447392). R. opacus 1BN was previously iden-
tified (Andreoni et al. 2000).
Preparation of cell extracts
Strains 1.7 and C3R were separately grown in M9
liquid cultures containing either glucose (1 g l–1)
or phenanthrene (400 mg l–1). Cells were har-
vested by centrifugation (15 min at 10,000 · g,
5�C) at fixed incubation times, washed twice with
50 mM Tris–HCl buffer pH 7.5, containing 0.3 M
NaCl. The washed cells were frozen and stored at
–20�C.
Cell-free extracts for immunological detection
and enzymatic assays were prepared from washed
frozen cells (1 g) by treatment with lysozyme
(1 mg) in 50 mM Tris–HCl buffer pH 8.0, con-
taining 0.3 M NaCl followed by pulse sonication
(3 min) and centrifugations (1 h at 10,000 · g,
and 40 min at 19,600 · g).
Enzyme assays
The discontinuous method that quantitates the
product thiocyanate, based on the absorption of
the ferric-thiocyanate complex at 460 nm, was
used to determine either thiosulfate:cyanide sul-
furtransferase (TST) activity (Sorbo 1953), or
MST activity (Jaraback and Westley 1980), with
thiosulfate and 3-mercaptopyruvate the sulfur
donor, respectively. One unit of enzyme activity
Table 1 Growth characteristics of the strains analysed in the study
Strains Growth substrates
Phen Naph Salicylate o-Phthalate Benzoate Benzene Toluene n-Hexa
S. chlorophenolicum 1.7 + – + – + – + –S. chlorophenolicum 1.8 + – + – + – – –S. chlorophenolicum C3R + + + – + – – –S. chlorophenolicum 5.1 + + + – + – + –R. aetherovorans 1.9 + – + – + – – +R. aetherovorans 6.22 +/– + + – + – + +R. opacus 1BN – + + – + + + +M. smegmatis F + – + – + – – +
phenanthrene (Phen); naphthalene (Naph); n-hexadecane (n-Hexa); growth (+); absence of growth (–); weak growth(+/–)
Results were recorded after growth of two subsequent cultures
Antonie van Leeuwenhoek
123
(U) is defined as the amount of enzyme that
produces 1 lmol thiocyanate min–1 at 37�C.
GST activity was determined as described in
Asaoka et al. (1977) in the presence of CDNB
and GSH, by measuring the absorbance increase
at 340 nm due to the formation of S-chloronitro-
phenyl glutathione (e340 = 9.600 M–1 cm–1). One
unit of enzyme activity (U) is defined as the
amount of enzyme that produces the conjugation
of 1 lmol of substrate (CDNB) min–1 at 37�C. All
enzymatic assays were carried out in a spectro-
photometer (Lambda 2, Perkin-Elmer) equipped
with a thermostatic cell holder.
The protein concentration of cell-free extracts
was determined by the method of Bradford
(1976).
Western Blot
Immunological detection of either RhdA-like or
SseA-like proteins blotted onto nitrocellulose
membranes from SDS-PAGE gels of the extracts,
was performed as previously described (Cereda
et al. 2003). Polyclonal anti-RhdA antibodies
raised against purified Azotobacter vinelandii
RhdA after overexpression in E. coli (Bordo
et al. 2000), and polyclonal anti-SseA antibodies
raised against purified E. coli SseA (Colnaghi
et al. 2001) were used.
Phenanthrene degradation experiments
Two strains, named C3R and 1.7, which exhibited
both GST and MST activities after growth on
phenanthrene, were chosen. Phenanthrene degra-
dation experiments were carried out in triplicate
in 100 ml vials. To each vial, 400 mg l–1 phenan-
threne dissolved in acetone was added and, after
evaporation of the solvent, the vials were filled
with 19 ml of M9 medium. The systems were
separately inoculated with 1 ml (OD600 = 0.5) of
cell suspension of S. chlorophenolicum C3R and
1.7 strains. Vials were closed with butyl rubber
stoppers and incubated at 30�C on a rotary shaker
at 150 rpm and they were sacrificed at intervals to
determine growth curves and phenanthrene con-
tent. The viable plate count method on 0.1·Tryptic Soy Agar was used to determine microbial
growth. Phenanthrene was quantified by HPLC
analyses carried out in a HPLC chromatograph
(mod. LG 980/02, Jasco Ternary Gradient Unit)
equipped with an octyl silane column (Merck
Lichrosorb RP-8, length 25 cm). A mixture of
methanol–water (75:25, vol/vol) acidified by the
addition of 0.1% phosphoric acid was the mobile
phase. The flow rate of eluent was 0.8 ml min–1.
The compound was detected by measuring the UV
absorbance at 254 nm in an UV-975 Intelligent
Jasco detector. The concentration of
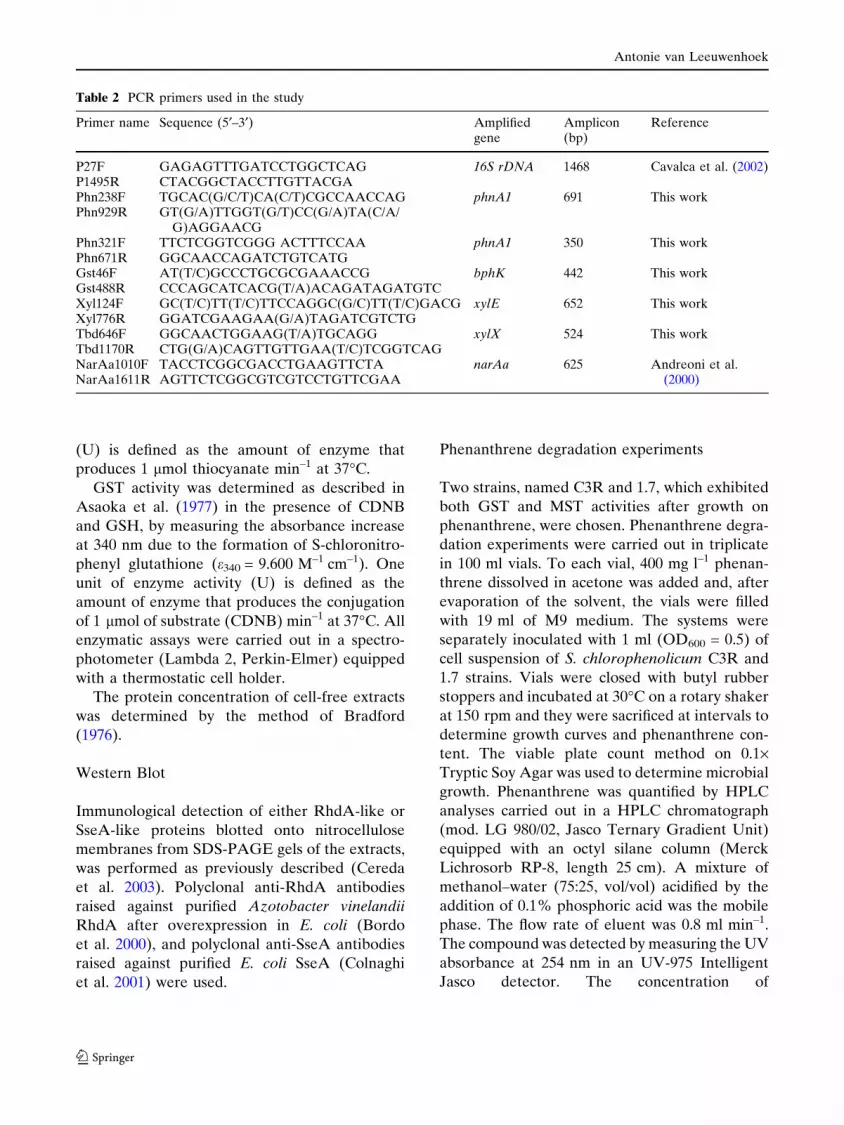
Table 2 PCR primers used in the study
Primer name Sequence (5¢–3¢) Amplifiedgene
Amplicon(bp)
Reference
P27F GAGAGTTTGATCCTGGCTCAG 16S rDNA 1468 Cavalca et al. (2002)P1495R CTACGGCTACCTTGTTACGAPhn238F TGCAC(G/C/T)CA(C/T)CGCCAACCAG phnA1 691 This workPhn929R GT(G/A)TTGGT(G/T)CC(G/A)TA(C/A/
G)AGGAACGPhn321F TTCTCGGTCGGG ACTTTCCAA phnA1 350 This workPhn671R GGCAACCAGATCTGTCATGGst46F AT(T/C)GCCCTGCGCGAAACCG bphK 442 This workGst488R CCCAGCATCACG(T/A)ACAGATAGATGTCXyl124F GC(T/C)TT(T/C)TTCCAGGC(G/C)TT(T/C)GACG xylE 652 This workXyl776R GGATCGAAGAA(G/A)TAGATCGTCTGTbd646F GGCAACTGGAAG(T/A)TGCAGG xylX 524 This workTbd1170R CTG(G/A)CAGTTGTTGAA(T/C)TCGGTCAGNarAa1010F TACCTCGGCGACCTGAAGTTCTA narAa 625 Andreoni et al.
(2000)NarAa1611R AGTTCTCGGCGTCGTCCTGTTCGAA
Antonie van Leeuwenhoek
123

phenanthrene in the samples was calculated by
using standards of known concentrations.
DNA extraction
Cell suspensions (100 ll OD600 = 2.0) were cen-
trifuged at 13,000 · g for 7 min and suspended in
100 ll of sterile MilliQ water, 100 ll of 10 mM
Tris–HCl buffer pH 8.0 and 13 ll of Proteinase K
(1 mg ml–1). The mix was incubated for 2 h at
55�C then boiled for 10 min and centrifuged at
13,000 · g for 5 min. The DNA-containing super-
natant was withdrawn and put in sterile
microtubes.
Polymerase chain reaction (PCR)
amplifications
Amplification of genes for 3,4-phenantrene diox-
ygenase large subunit (phnA1), for GST (bphK),
for toluate/benzoate dioxygenase large subunit
(xylX) and for 2,3-extradiol dioxygenase (xylE),
were performed in 50 ll final volume containing:
10X buffer (BIO101 Qbiogene), 1.75 mM MgCl2,
200 lM nucleotides (dNTPs) (GE Healthcare,
Sweden), 0.2 lM forward and reverse primers
each (Invitrogen, UK), 1 U Taq polymerase
(BIO101 Qbiogene), and 1 ll of template DNA.
On the basis of database available sequences
degenerated primers were designed (Table 2) for
PCR amplification of homologous catabolic genes
in soil isolates using the following strategies.
Primer set Phn238F/Phn929R was placed at
positions 238 and 929 of the reference nucleotide
sequence of phnA1 gene for 3,4-phenanthrene
dioxygenase large subunit of strain Sphingomonas
sp. CHY1 (GeneBank acc. num. AJ633551).
Primer set Gst46F/Gst488R was placed at posi-
tions 46 and 488 of the reference nucleotide
sequence of gene bphK for GST of strain
Sphingomonas aromaticivorans F199 (GeneBank
acc. num. AF079317). Primer set Xyl124F/
Xyl776R was placed at positions 124 and 776 of
the reference nucleotide sequence of xylE gene
for 2,3-extradiol dioxygenase of strain S. aromat-
icivorans F199 (GeneBank acc. num. AF079317).
Primer set Tbd646F/Tbd1170R was placed at
positions 646 and 1170 of the reference nucleotide
of xylX gene for toluate/benzoate dioxygenase
large subunit of strain S. aromaticivorans F199
(GeneBank acc. no. AF079317). The thermal
profile was: 94�C for 2 min, followed by 35 cycles
(94�C for 40 s, 60�C for 50 s, 72�C for 1 min) and
72�C for 7 min. Amplifications of narAa gene for
1,2-naphtalene dioxygenase large subunit was
carried out as previously described (Andreoni
et al. 2000).
All sequences were obtained by using the Taq
Dye-Deoxy Terminator Cycle Sequencing kit
(Applied Biosystems, USA) and automatic
DNA sequencer (373A, Applied Biosystems,
USA) according to the manufacturer’s instruc-
tions. Primers used in the PCR reaction of
sequencing products were the same as in normal
PCR reactions.
RNA extraction and RT-PCR analysis
Total RNA was extracted from S. chlorophenol-
icum cultures (1.7 and C3R strains) after 4 day
growth in mineral medium (M9) supplemented
with 400 mg l–1 phenanthrene or 1 g l–1 glucose.
Samples (1010 cells) were spun down (8000 · g for
15 min), the cell pellets were suspended in
3.25 ml lysing buffer (80 mM Tris–HCl buffer
pH 7.6 containing 800 mM NaCl and 8 mM
EDTA) and digested for 1 h at 50 �C with 1 mg
proteinase K. RNA was further extracted with
phenol, phenol:chlorophorm:isoamil alcohol
(25:24:1, vol/vol/vol) and chlorophorm:isoamil
alcohol (24:1, vol/vol). RNA was recovered after
precipitation with ethanol and treated with
RNase-free DNase I (GE Healthcare, Sweden).
For semi-quantitative RT-PCR, DNAse treated
total RNAs were retrotranscribed to first-strand
cDNA using Stratascript Reverse Transcriptase
(Stratagene, USA) according to the manufac-
turer’s instructions. cDNA aliquots from the RT
reactions were used for the amplification of the
different genes with Taq DNA Polymerase (Invi-
trogen, UK). The PCR primers used were the
followings: Phn321F/Phn671R primer set, specif-
ically designed on the nucleotide sequences
obtained from phnA1 3,4-phenanthrene dioxy-
genase gene of strains 1.7 and C3R (Table 2); the
Gst46F/Gst488R primer set; P27F/P1495R primer
set, used to amplify the 16S rRNA gene as an
amplification control of housekeeping gene. The
Antonie van Leeuwenhoek
123

PCR products were separated on 2% agarose gels
along with O’gene Ruler 1 kb ladder (Fermentas,
Canada). As positive controls, genomic DNA
from the same strains was used as template. As
negative controls, not retro transcribed DNase-
treated RNA was used as template in order to
avoid false positive results.
Results
Growth profiles and genetic traits of the
studied strains
The ability of the strains to utilize PAHs as sole
sources of carbon and energy is reported in
Table 1. The strains were differently able to
utilize phenanthrene and/or naphthalene. All
the strains grew on benzoate and on salicylate,
but not on o-phthalate. R. aetherovorans strains
1.9 and 6.22, R. opacus 1BN and M. smegmatis F
grew also on n-hexadecane. Genetic analyses
were performed to identify in the strains some
genetic traits for catabolic features, and the
presence of GSTs (Table 3). Amplification of
the genes (phnA1 and xylE) for phenanthrene
degradation present, respectively, in the upper
and in the lower degradative operons, for monoa-
romatic ring hydroxylating dioxygenase (xylX)
and for GST (bphK) were obtained in all the
S. chlorophenolicum strains. Genes for 1,2-naph-
thalene dioxygenase (narAa) were amplified in
R. aetherovorans strains 1.9 and 6.22, R. opacus
1BN and M. smegmatis F.
Enzymatic activity profiles
RhdA from A. vinelandii and SseA from E. coli,
are prototypes of rhodanese-like proteins with
TST and MST activity, respectively (Bordo et al.
2001, 2002; Pagani et al. 2000; Colnaghi et al.
2001). Taking advantage of the finding that the
antibody raised against RhdA did not recognize
SseA, and vice versa, a preliminary screening for
the presence of RhdA-like and or SseA-like
proteins in the strains here studied was carried
out by Western blot analysis. RhdA-like proteins
were never detected, whereas SseA-like proteins
were detected in R. aetherovorans strains 1.9 and
6.22, R. opacus 1BN and M. smegmatis F, but not
in S. chlorophenolicum strains. In agreement with
the absence of RhdA-like proteins, TST activity
was very low in all strains (0.30–0.50 U mg–1),
while MST activity was always ‡1.0 U mg–1.
To deepen the possible correlation between
detoxifying activities and phenanthrene utiliza-
tion, the modulation of MST and GST activities as
Table 3 Functional genes found in the studied strains
Strains Enzyme-coding genes Reference organisms GeneBankAcc. Num.
Homology(%)
S. chlorophenolicum1.7, 1.8, C3R, 5.1
phnA1 (3,4-phenanthrenedioxygenase)
S. aromaticivorans F199 AF079317 97.8Sphingomonas sp. CHY1 AJ633551 79.2
xylX (ring hydroxylatingdioxygenase)
S. aromaticivorans F199 AF079317 97Sphingomonas sp. P2 AB091692 87
xylE (2,3-catechol dioxygenase) S. aromaticivorans F199 AF079317 95S. chungbukense DJ77 U83882 95
bphK (glutathione S-transferase) S. chungbukense DJ77 AF001103 96.5S. aromaticivorans F199 AF079317 84.5
R. aetherovorans1.9, 6.22
narAa (naphthalene dioxygenase) Rhodococcus sp. P400 AY392423 98R. opacus 1BN AJ401612 90
R. opacus1BN
narAa (naphthalene dioxygenase) Rhodococcus sp. NCIMB12038
AF082663 99
Rhodococcus sp. I24 AF121905 90
M. smegmatisF
narAa (naphthalene dioxygenase) Mycobacterium sp. 6PY1 AJ494745 86M. vanbaalenii PYR-1 AF249301 83
Antonie van Leeuwenhoek
123

a function of the carbon source (glucose or
phenanthrene) was analysed (Table 4). High level
of MST activity, independent of the presence of
immunodetectable SseA-like proteins, was found
in all strains. A significant increase of MST activity
in the presence of phenanthrene was found in the
strains where GST activity was absent, and where
a SseA-like protein was revealed by immunode-
tection. GST activity correlated well with the
presence of the bphK gene in S. cholorophenol-
icum, in which a very low level of activity
was found in the presence of glucose, whereas a
high activity was induced by the growth on
phenanthrene (Table 4).
Phenanthrene degradation and enzyme
activities
S. chlorophenolicum 1.7 and C3R strains were
chosen for further investigating whether phenan-
threne degradation could be related to the ‘‘acti-
vation’’ of detoxifying enzymes. As shown in
Fig. 1A, the profiles of cell growth and time-
course of phenanthrene disappearance were
essentially identical for both strains. Both strains
exhibited a comparable degradation rate, and
more than 80% of the hydrocarbon was degraded
in the exponential phase. The figures of GST and
MST activities, measured either in the presence of
glucose or phenanthrene at different time growth
(Fig. 1B), indicated different enzymatic modula-
tion behaviours for these S. chlorophenolicum
isolates. Slight time-dependent increase and
induction by phenanthrene of GST activity was
found in S. chlorophenolicum C3R. In S. chlor-
ophenolicum 1.7, GST activity was significantly
induced by phenanthrene after 4 days growth and
doubled after 7 days growth. Similar trend was
found in S. chlorophenolicum 1.7 for MST activ-
ity. Noticeably, the figures of both GST and MST
activities did not change from 4 to 7 days growth
when glucose was the carbon source.
The increase of GST activity found in extracts
from phenanthrene grown cells, as compared to
glucose grown cells, suggested a transcriptional
regulation of the expression of this enzyme.
Glutathione S-transferase and
3,4-phenanthrene dioxygenase gene trancripts
Gene expression was, therefore, measured
by determining levels of both the GST and
3,4-phenanthrene dioxygenase gene transcripts.
To determine whether these genes were actually
transcribed in response to phenanthrene, RT-
PCR experiments were performed with total
RNA isolated from induced (i.e. growth in the
presence of phenanthrene) and uninduced (i.e.
growth in the presence of glucose) cells of
S. chlorophenolicum 1.7 and C3R (Fig. 2). With
Table 4 MST and GST activities of the strains grown on glucose and on PAHs
Strain Growth on MST (U mg-1) GST (U mg–1)
S. chlorophenolicum 1.7 glucose 1.16 ± 0.07 0.05 ± 0.04phenanthrene 1.43 ± 0.23 1.49 ± 0.13
S. chlorophenolicum 1.8 glucose 1.33 ± 0.23 0.00 ± 0.00phenanthrene 1.28 ± 0.12 0.39 ± 0.05
S. chlorophenolicum 5.1 glucose 1.19 ± 0.19 0.09 ± 0.01phenanthrene 1.66 ± 0.04 0.61 ± 0.11
S. chlorophenolicum C3R glucose 1.64 ± 0.39 0.14 ± 0.02phenanthrene 1.58 ± 0.18 0.97 ± 0.11
R. aetherovorans 1.9 glucose 1.74 ± 0.09 0.00 ± 0.00phenanthrene 2.72 ± 0.33 0.00 ± 0.00
R. aetherovorans 6.22 glucose 1.43 ± 0.24 0.00 ± 0.00naphthalene 1.68 ± 0.35 0.00 ± 0.00
R. opacus 1BN glucose 1.00 ± 0.11 0.00 ± 0.00naphthalene 1.41 ± 0.17 0.00 ± 0.00
M. smegmatis F glucose 1.03 ± 0.18 0.00 ± 0.00phenanthrene 2.37 ± 0.44 0.00 ± 0.00
Each value is a mean of four replicates determined after 4 day growth
Antonie van Leeuwenhoek
123

0
50
100
150
200
250
300
350
400
450
0 1 2 3 4 5 6 7
Time (days)
7
7.5
8
8.5
9
9.5
10
Ph
enan
thre
ne
(mg
ml-1
)
Bac
teri
al c
ell g
row
th (
Lo
g C
FU
ml-1
)
MST (U mg-1) GST (U mg-1)Strain
Time of growing (days)
4 7 4 7
S. chlorophenolicum 1.7
Glucose Phenanthrene
1.16±0.02 1.43±0.23
1.27±0.14 2.27±0.28
0.05±0.02 1.49±0.13
0.03±0.01 2.60±0.33
S. chlorophenolicum C3R
GlucosePhenanthrene
1.61±0.21 1.58±0.18
1.66±0.22 1.86±0.33
0.14±0.02 0.97±0.11
0.13±0.02 1.12±0.05
A
B
Fig. 1 (A) S. chlorophenolicum strains 1.7 (circles) andC3R (squares) were grown on 400 mg l–1 phenanthrene.Cell growth (empty symbols) and phenanthrene degrada-tion (full symbols) are shown. Each value is a mean of
three replicates, and standard deviations are reported. (B)MST and GST activities measured at 4 day and 7 dayincubation. The activity figures from control experimentsof cells grown on glucose are also shown
bphK phnA11.7 C3R
G Phen G Phen G Phen G Phen
1.7 C3R
MM M
442 bp
350 bp
Fig. 2 Transcriptional analysis of glutathione S-transfer-ase (bphK) and 3,4-phenanthrene dioxygenase (phnA1)encoding genes of S. chlorophenolicum strains 1.7 and C3R
using RT-PCR. Total RNA from cells grown on glucose(G) or on phenanthrene (Phen) was used as template foramplification. M: O’Gene Ruler 1 kb ladder
Antonie van Leeuwenhoek
123

the use of primer set Gst46F and Gst488R, an
expected fragment of 442 bp corresponding to
GST was amplified from the total RNA extracted
from cells grown with phenanthrene, while no
fragment was amplified from cells grown on
glucose. With the use of primer set Phn321F
and Phn671R, an expected fragment of 350 bp
corresponding to 3,4-phenanthrene dioxygenase
was amplified from the total RNA extracted from
cells grown on phenanthrene, while the product
was barely amplified from cells grown on glucose.
The 1500 bp fragment corresponding to the 16S
rRNA, used as a control, gave the expected
fragment in all the experiments (data not shown).
These results indicated that the growth of
S. chlorophenolicum 1.7 and C3R strains in the
presence of phenanthrene led to increasing the
expression of GST and 3,4-phenanthrene dioxy-
genase genes.
Discussion
S. chlorophenolicum, R. aetherovorans, R. opacus
and M. smegmatis strains were able to utilize both
monocyclic and polycyclic aromatic hydrocarbons
as carbon and energy sources for growth, thus
confirming the common traits of the aromatic
hydrocarbon-degrading strains of these genera.
The ability grown on salicylate and not on
o-phthalate, and the presence of the catechol
2,3-dioxygenase (xylE) gene indicated that these
strains utilized phenanthrene through the salicy-
late meta pathway, as reported for Sphingomonads
(Pyniakong et al. 2003a; Demaneche et al. 2004).
Sequence analysis revealed that the xylX gene
for ring hydroxylating dioxygenase and the
gene bphK for GST were present along with
genes involved in PAH degradation. This gene
organization seems to be highly conserved in
strains exhibiting diverse origins and might be
typical for PAH-degrading Sphingomonas strains
(Pyniakong et al. 2003b; Romine et al. 1999;
Lloyd-Jones and Lau 1997; Xia et al. 2005).
Considering that the reaction of oxygenases
involved in the degradation of xenobiotics might
produce oxidative damage in the cells (Favaloro
et al. 2000), we focussed our studies on GST gene
products for which, in addition to a catalytic role
in the metabolism of endogenous or exogenous
toxic electrophilic chemicals, a detoxification role
could be figured (Vuillemieur and Pagni 2002),
and to members of the rhodanese homology
superfamily whose detoxifying activity may not
be limited to cyanide scavenging.
The availability of selected antibodies that did
not show cross-reactions, allowed the screening
for the presence of rhodanese-like protein exam-
ples (RhdA-like and SseA-like) belonging to the
two different subdomains of the widespread
rhodanese-homology proteins. The active-site
loop of E. coli SseA, shared by a number of
rhodanese-like proteins listed in databases, rep-
resents a signature for MST activity, while the
active-site loop of A. vinelandii RhdA, found in a
limited number of rhodanese-like proteins is
discriminant for TST activity (Bordo et al. 2000;
Pagani et al. 2000; Colnaghi et al. 2001; Bordo
et al. 2001). No presence of RhdA-like proteins,
nor significant level of TST activity were found in
any of the analysed strains. The prominent
presence of MST activity and the modulation of
this activity by phenanthrene in these strains, is in
agreement with a recent proteomic investigation
that revealed up-regulation of a putative SseA in
Pseudomonas putida in phenol-induced stress
conditions (Santos et al. 2004). The presence of
MST activity in strains where a SseA-like protein
was undetectable indicated that other sulfurtrans-
ferase enzymes were present, according to the
revealed presence of paralogs of these proteins in
sequenced genomes. Interestingly, besides the
widespread presence of high level of MST activ-
ity, significant modulation of this activity, inferred
by phenanthrene as carbon source, was found in
the strains where GST activity was absent. GST
activity was modulated by the presence of phen-
anthrene in the strains where molecular analyses
revealed the presence of bphK gene. Genetic
analyses indicated that the GST activity was
related to a bphK gene product, and that the
modulation of GST activity by phenanthrene
paralleled the induction of GST transcript. These
results suggest that the role of GST in S. chlor-
ophenolicum could be related to catabolic func-
tions. In the present study a search for
glutathione-conjugated intermediates was how-
ever not done. Since genes for putative GSTs
Antonie van Leeuwenhoek
123

have been found in the gene clusters responsible
for aromatic degradation in several Sphingomon-
as strains of widely diverse origins (Lloyd-Jones
and Lau 1997; Bae et al. 2003), they should have
been recruited by the strains for specific meta-
bolic functions. The phenanthrene-induced GST
activity showed a time-dependent increase, and
the highest level was measured when cells were in
stationary growth phase. The induction of GST
transcript was determined at 4 day growth, at a
time in which in the presence of glucose the
transcript was undetectable. The low level of the
GST activity found in the presence of glucose
suggests that in strains C3R and 1.7 GST tran-
script can be constitutively expressed. Growth
dependent levels of GST induction on m-toluate
have been related to the presence of multiple
copies of bphK gene in Sphingomonas yan-
oikuyae strain B1 (Bae et al. 2003).
Both C3R and 1.7 strains degraded phenan-
threne with comparable degradation rates (Fig. 1)
and transcriptional analysis of phenanthrene
3,4-dioxygenase gene phnA1 confirmed the pres-
ence of this phenanthrene-activated dioxygenase.
Glucose did not suppress the transcription of
3,4-phenanthrene dioxygenase gene, in agreement
with recent data about the activity of degradative
enzymes on glucose (Basu et al. 2006).
The ability of R. aetherovorans, R. opacus and
M. smegmatis strains to grow on phenanthrene
was not hampered by the absence of GST. In
these examples, the ability to grow by utilizing
phenanthrene as sole carbon source should be
related to the presence of different catabolic
operon and facilitated by the detoxification activ-
ity of MST enzymes. In Mycobacterium sp. strain
6PY1, in addition to dioxygenases, an up-regula-
tion of a rhodanese-like protein induced by
pyrene was found (Krivobok et al. 2003).
The multiple experimental approaches used in
this study allowed the definition of metabolic
traits and enzymatic profiles that differently
characterize PAH-degrading strains of environ-
mental relevance.
Acknowledgements This work was supported by ItalianNational Research Program PRIN 2003 ‘‘Selezione dicolture batteriche per la degradazione di POP e loromonitoraggio in processi di bioarricchimento dei suoli’’.
References
Adams H, Teertstra W, Koster M, Tommassen J (2002)PspE (phage-shock protein E) of Escherichia coli is arhodanese. FEBS Lett 518:173–176
Andreoni V, Bernasconi S, Colombo M, Beilen JB,Cavalca L (2000) Detection of genes for alkane andnaphthalene catabolism in Rhodococcus sp. strain1BN. Environ Microbiol 2:572–577
Andreoni V, Cavalca L, Rao MA, Nocerino G, BernasconiS, Dell’Amico E, Colombo M, Gianfreda L (2004)Bacterial communities and enzyme activities of PAHspolluted soils. Chemosphere 57:401–412
Asaoka K, Ito H, Takahashi K (1977) Monkey glutathioneS-aryltransferases. J Biochem 82:973–981
Bae M, Sul WJ, Koh SC, Lee JH, Zylstra GJ, Kim YM,Kim E (2003) Implication of two glutathione S-transferases in the optimal metabolism of m-toluateby Sphingomonas yanoikuyae B1. Ant van Leeuw84:25–30
Bartels F, Backhaus S, Moore ERB, Timmis KN, Hofer B(1999) Occurrence and expression of glutathione S-transferase-encoding bphK genes in Burkholderia sp.strain LB400 and other biphenyl-utilizing bacteria.Microbiology 145:2821–2834
Basu A, Apte SK, Phale PS (2006) Preferential utilizationof aromatic compounds over glucose by Pseudomonasputida CSV86. Appl Environ Microbiol 72:2226–2230
Bordo D, Forlani F, Spallarossa A, Colnaghi R, Carpen A,Bolognesi M, Pagani S (2001) A persulfurated cyste-ine promotes active-site reactivity in Azotobactervinelandii rhodanese. Biol Chem 382:1245–1252
Bordo D, Bork P (2002) The rhodanese/Cdc25 phospha-tase superfamily. Sequence-structure-function rela-tions. EMBO Rep 8:741–746
Bordo D, Deriu D, Colnaghi R, Carpen A, Pagani S,Bolognesi M (2000) The crystal structure of asulfurtransferase from Azotobacter vinelandii high-lights the evolutionary relationship between rhoda-nese and phosphatase enzymes family. J Mol Biol298:691–704
Bradford MM (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein uti-lizing the principle of protein-dye binding. AnalBiochem 72:248–254
Cavalca L, Colombo M, Larcher S, Gigliotti C, Collina E,Andreoni V (2002) Survival and naphthalene-degrad-ing activity of Rhodococcus sp. strain 1BN in soilmicrocosms. J Appl Microbiol 92:1058–1065
Cereda A, Forlani F, Iametti S, Bernhardt R, Ferranti P,Picariello G, Pagani S, Bonomi F (2003) Molecularrecognition between Azotobacter vinelandii rhoda-nese and a sulfur acceptor protein. Biol Chem384:1473–1481
Chavez FP, Lunsdorf H, Jerez CA (2004) Growth ofpolychlorinated-biphenyl-degrading bacteria in thepresence of biphenyl and chlorobiphenyls generatesoxidative stress and massive accumulation of inor-ganic polyphosphate. Appl Environ Microbiol70:3064–3072
Antonie van Leeuwenhoek
123

Colnaghi R, Cassinelli G, Drummond M, Forlani F, PaganiS (2001) Properties of the Escherichia coli rhodanese-like protein SseA: contribution of the active-siteresidue Ser240 to sulfur donor recognition. FEBSLett 500:153–156
Demaneche S, Meyer C, Micoud J, Louwagie M, WillisonJC, Jounanneau Y (2004) Identification and func-tional analysis of two aromatic ring hydroxylatingdioxygenases from a Sphingomonas strain that de-grades various polycyclic aromatic hydrocarbons.Appl Environ Microbiol 70:6714–6725
Favaloro B, Tamburro A, Trofino MA, Bologna L, RotilioD, Heipieper HJ (2000) Modulation of the glutathi-one S-transferase in Ochrobactrum anthropi: functionof xenobiotic substrates and other forms of stress.Biochem J 346:553–559
Kim E, Zylstra GJ (1999) Functional analysis of genesinvolved in biphenyl, naphthalene, phenanthrene, andm-xylene degradation by Sphingomonas yanoikuyaeB1. J Ind Microbiol Biotechnol 23:294–302
Jarabak R, Westley J (1980) 3-Mercaptopyruvate sulfo-transferase rapid equilibrium-ordered mechanismwith cyanide as the acceptor substrate. Biochemistry19:900–904
Kanaly RA, Harayama S (2000) Biodegradation of high-molecular-weight polycyclic aromatic hydrocarbonsby bacteria. J Bacteriol 182:2059–2067
Krivobok S, Kuony S, Meyer C, Louwagie M, Willison JC,Juoanneau Y (2003) Identification of pyrene-inducedproteins in Mycobacterium sp. strain 6PY1:evidencefor two ring-hydroxylating dioxygenases. J Bacteriol185:3828–3841
Kunz DA, Chapman PJ (1981) Catabolism of pseudocum-ene and 3-ethyltoluene by Pseudomonas putida (ar-villa) mt–2: evidence for new function of the TOL(pWW0) plasmid. J Bacteriol 146:179–191
Lloyd-Jones G, Lau PCK (1997) Glutathione S-transferaseencoding gene as a potential probe for environmentalbacterial isolates capable of degrading polycyclicaromatic hydrocarbons. Appl Environ Microbiol63:3286–3290
Nagata Y, Miyauchi K, Takagi M (1999) Completeanalysis of genes and enzymes for hexachlorocyclo-hexane degradation in Sphingomonas paucimobilisUT26. J Industr Microbiol Biotechnol 23:380–390
Pagani S, Forlani F, Carpen A, Bordo D, Colnaghi R(2000) Mutagenic analysis of Thr232 in rhodanesefrom Azotobacter vinelandii highlighted thedifferences of this prokaryotic enzyme from theknown sulfurtransferases. FEBS Lett 472:307–311
Pinyakong O, Habe H, Omori T (2003a) The uniquearomatic catabolic gene in sphingomonads degrading
polycyclic aromatic hydrocarbons (PAHs). J Gen AppMicrobiol 49:1–19
Pinyakong O, Habe H, Yoshida T, Nojiri H, Omori T(2003b) Identification of three novel salicylate1-hydroxylases involved in the phenanthrene degra-dation of Sphingobium sp. strain P2. Biochem Bio-phys Res Commun 301:350–357
Ploegman JH, Drent G, Kalk KH, Hol WG, HeinriksonRL, Keim P, Weng L, Russell J (1978) The covalentand tertiary structure of bovine liver rhodanese.Nature 273:124–129
Ray WK, Zeng G, Potters MB, Mansuri AM, Larson TJ(2000) Characterization of a 12-Kilodalton rhodaneseencoded by glpE of Escherichia coli and its interactionwith thioredoxin. J Bacteriol 182:2277–2284
Romine MF, Stillwell LC, Wong KK, Thurston SJ, SiskEC, Sensen C, Gaasterland T, Fredrickson JK, SafferJD (1999) Complete sequence of a 184-KilobaseCatabolic Plasmid from Sphingomonas aromaticivo-rans F199. J Bacteriol 181:1585–1602
Saito A, Iwabuchi T, Harayama S (1999) Characterizationof genes for enzymes involved in the phenanthrenedegradation in Nocardioides sp. KP7. Chemosphere38:1331–1337
Santos PM, Benndorf D, Sa-Correla I (2004) Insights intoPseudomonas putida KT2440 response to phenol-induced stress by quantitative proteomics. Proteomics4:2640–2652
Sorbo B (1953) Rhodanese. Acta Chem Scand 7:1137–1145Tamburro A, Robuffo I, Heipieper HJ, Allocati N, Rotilio
D, Di Ilio C, Favaloro B (2004) Expression ofglutathione S-transferase and peptide methioninesulfohoxide reductase in Ochrobactrum anthropi iscorrelated to the production of reactive oxygenspecies caused by aromatic substrates. FEMS Micro-biol Lett 241:151–156
Van Hylckama JET, Leemhuis H, Spelberg JHL, JanssenDB (2000) Characterization of the gene clusterinvolved in isoprene metabolism in Rhodococcus sp.strain AD45. J Bacteriol 182:1956–1963
Vuilleumier S, Pagni M (2002) The elusive roles ofbacterial glutathione S-transferase: new lessons fromgenome. Appl Microbiol Biotechnol 58:138–146
Westley J (1977) Sulfane-transfer catalysis by enzymes. In:Van Tamelen EE (eds) Bio-organic chemistry, vol. 1.Academic Press, Orlando, FL, pp 371–390
Xia Y, Min H, Rao G, Lv Z, Liu J, Ye Y, Duan X (2005)Isolation and characterization of phenanthrene-degrading Sphingomonas paucimobilis strain ZX4.Biodegradation 16:393–402
Antonie van Leeuwenhoek
123





























