Embed Size (px)
Citation preview


ENCYCLOPEDIA OF Biodiversity
xviii+598_Biodiversity_F.indd 1 11/17/11 1:12 PM

xviii+598_Biodiversity_F.indd 2 11/17/11 1:12 PM

ENCYCLOPEDIA OF
Biodiversity
Stanley a. Rice, Ph.D.
xviii+598_Biodiversity_F.indd 3 11/17/11 1:12 PM

ENCYCLOPEDIA OF BIODIVERSITY
Copyright © 2012 by Stanley A. Rice, Ph.D.
All rights reserved. No part of this book may be reproduced or utilized in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage or retrieval systems, without permission in writing from the
publisher. For information contact:
Facts On File, Inc.An imprint of Infobase Learning
132 West 31st StreetNew York NY 10001
Library of Congress Cataloging-in-Publication Data
Rice, Stanley A., 1957– Encyclopedia of biodiversity / author, Stanley A Rice.
p. cm.—(Science encyclopedia) Includes bibliographical references and index.
ISBN 978-0-8160-7726-7 (hardcover) ISBN 978-1-4381-3779-7 (e-book)1. Biodiversity—Encyclopedias—Juvenile literature. I. Title.
QH541.15.B56R53 2012 578.703—dc22 2010050557
Facts On File books are available at special discounts when purchased in bulk quantities for businesses, associations, institutions, or sales promotions. Please call our Special
Sales Department in New York at (212) 967-8800 or (800) 322-8755.
You can find Facts On File on the World Wide Web at http://www.infobaselearning.com
Excerpts included herewith have been reprinted by permission of the copyright holders; the author has made every effort to contact copyright holders. The publishers will be
glad to rectify, in future editions, any errors or omissions brought to their notice.
Text design by Annie O’DonnellComposition by Hermitage Publishing Services
Photo research by Elizabeth H. Oakes and the authorCover printed by Bang Printing, Brainerd, Minn.
Book printed and bound by Bang Printing, Brainerd, Minn.Date printed: December 2011
Printed in the United States of America
10 9 8 7 6 5 4 3 2 1
This book is printed on acid-free paper.

This encyclopedia is dedicated to Edward Osborne Wilson, who has spent his life studying the biodiversity that most people overlook, particularly ants and all the things they do that help to keep Earth alive. He has scientifically described many ant
species and made major discoveries in ant biology. For decades, he has championed the public understanding and appreciation of biodiversity and has recently been focusing his attention on an
online database of the species of the world (encyclopedia of life). Wilson has also made major contributions to evolutionary biology.
Though one of the most famous scientists in the world, he is not too proud to get down on his knees to look at ants.
xviii+598_Biodiversity_F.indd 5 11/17/11 1:12 PM

xviii+598_Biodiversity_F.indd 6 11/17/11 1:12 PM

acknowledgments ix
introduction x
the national Science education Standards xiii
entries a–Z 1
Essays:“Why are there So Many Species?” 50
“Ghosts of Biodiversity Past” 118
“ecosystem Services: Public or Private costs?” 214
“life of earth: Portrait of a Beautiful, Middle-aged, Stressed-Out Planet” 268
“if Humans Vanished . . .” 501
Appendixes:i. chronology 548
ii. Glossary 550
iii. Further Resources 556
iV. classifi cation 564
index 569
Contents
xviii+598_Biodiversity_F.indd 7 11/17/11 1:12 PM

xviii+598_Biodiversity_F.indd 8 11/17/11 1:12 PM

ix
the author acknowledges the energetic encouragement of Facts On File’s executive editor Frank Darmstadt, who was instantly available to guide and assist the author at all stages of manuscript preparation, from start to fi nish. the author also thanks agent Jodie Rhodes for zealous encouragement and for making this book possible. the author also thanks Michael axon, editorial assistant, for his meticulous prepara-tion of the manuscript materials; the production staff
of Facts On File for preparing fi gures and facilitat-ing electronic fi le transfer; the copyediting depart-ment; and elizabeth Oakes, for locating outstanding photographs. the author also greatly benefi ted from Southeastern Oklahoma State University’s providing internet resources and funds from Organized Research for the author to participate in scientifi c conferences. the author thanks his wife, a. lisette Rice, for mak-ing internet publicity of the author’s works possible.
Acknowledgments
xviii+598_Biodiversity_F.indd 9 11/17/11 1:12 PM

x
the loss of global biodiversity is one of the major crises in the world today. Plus, unlike other crises, it is irreparable: Once a species has become extinct, it is gone forever. Driving species into extinction is, as eminent evolutionary biologist edward O. Wil-son states, the act for which future generations are least likely to forgive the present one. Political and economic problems can be solved, and even the most persistent pollutants will eventually decompose, but new species will not evolve to replace those that have been driven into extinction on any time scale mean-ingful to humans.
the extinction of species is a crisis that can be easily dismissed. We may seldom notice the biodi-versity around us. We may be accustomed to lawns that consist of just one species of grass, but natural grasslands contain many dozens of plant species, as did even the lawns around people’s houses in earlier centuries (see fi gure on page xi). to many people, nonhuman species (other than those upon which we directly depend for food) are not criti-cally important. Some people believe that the only effect of global warming is the extinction of polar bears and that the only way in which polar bears are important is that wealthy ecotourists or watch-ers of cable television like to observe them. they are wrong in both ways. First, humans will suf-fer from global warming perhaps even more than the polar bears. Second, polar bears, as well as all other species, provide benefi ts that are essential for the human economy, particularly to agriculture and medicine. Most people do not realize how much our health care system depends on medicinal plants and animals and the native peoples who know about them. (that is right—the physiology of bears may help scientists understand how to treat some human diseases.) Most people do not realize how our food production systems depend on wild pollinators and the genes from wild species. they do not realize that intact ecosystems, with all of their component spe-cies, provide trillions of dollars of free services to the human economy. that is, if we do not protect wild
species and the ecological communities in which they live, we are putting the world economy at risk. Someone needs to explain this to them. this encyclo-pedia does so.
the extinction crisis is also easy to overlook. to someone who has not studied them, the different species of oak trees or of frogs all look pretty much alike, and if one of them becomes extinct, there are plenty of others. However, each species has its own unique set of genes, and no other species can com-pletely substitute for it.
Most extinctions today are caused by human activity. the most detrimental human activity is hab-itat destruction. Humans cut down forests to provide wood and create pastures and pave over grasslands with suburban expansion. Some of the most rapid destruction of natural habitats is in the parts of the world with the most species, such as california and the western amazon basin. extinctions caused by habitat destruction have skyrocketed in the last cen-tury. the rate of extinction occurring today is at least as great as during most of the fi ve mass extinction events in earth history, and human activity is now causing the sixth mass extinction. nevertheless, an even greater extinction crisis looms as global warm-ing begins to disrupt natural habitats and all the species that live in them. in the 19th century, some people tried to “shoot the last buffalo,” but in the 21st century, humans can disrupt the whole prairie ecosystem in which buffalo used to live.
People need to learn to notice and appreci-ate biodiversity, and they have to begin doing so while they are young. as kids, they need to spend some time outdoors in direct contact with trees and grasses and spiders and birds. By the time they are in high school or have entered college, they need to learn a great deal about biodiversity, its importance to humans, and the nature of the threats against it. Encyclopedia of Biodiversity enables high school and college students to fi nd information about all of these things. the 153 cross-referenced entries and fi ve essays average at least 2,000 words each,
Introduction
xviii+598_Biodiversity_F.indd 10 11/17/11 1:12 PM

xi
long enough to provide a good understanding but small enough to not discourage reading. High school teachers can use these entries to meet many of the science education goals established by the national academy of Sciences, as outlined in the section on the national Science education Standards.
if people do not discover biodiversity when they are young, they may never do so. i once had a stu-dent who barely passed a college plant diversity course. However, he told me that he had previously driven his truck down the highway not noticing the trees; now, as he drove, he recognized how many dif-ferent species of trees were there and how the ones that lived near the rivers were different from those that lived on the hillsides (even if he forgot what they were). We had remediated his biodiversity ignorance just a little, and just in time.
educators and scientists realize that citizens need to have “science literacy” in order to make informed decisions as consumers and voters. Despite the abundance of information available on the inter-net and massive expenditures on education, most citizens are unable to draw a valid conclusion from scientific information, whether it concerns their own health or the health of the environment. the national academy of Sciences even referred to the danger that the lack of science literacy poses to the future of the United States as “the Gathering Storm.” One of the goals of this encyclopedia is to make information about groups of organisms, from bacteria to mammals, and about ecological concepts and processes, such as biogeography and ecological succession, clearly and readily available to students and the general public.
Fortunately, biodiversity is one of those topics for which it is easy to generate interest. the world is full of millions of colorful and fascinating species. the entries and colorful illustrations of this encyclo-pedia will invite readers to explore worlds that they already know will interest them, but such material must be presented in an interesting way. that is what i have tried to do. i am a trained professional bota-nist, but most of my work for the past two decades has been to make botany and other biodiversity and ecological topics interesting to students and to read-ers. i have little interest in writing things that will be read only by other scientists. i have traveled all over the United States (and some other parts of the world) in search of ecology and biodiversity stories. i have seen and been excited by many of the things i write about in this encyclopedia and hope to pass the enthusiasm on to my readers.
all stories about biodiversity and ecology are founded in evolutionary science. it is not possible to understand why some species of organisms live
in one part of the world and others in a different part of the world without understanding how they evolved. in this encyclopedia, i consistently provide the evolutionary framework in which biodiversity must be understood.
the figures and tables are designed to be useful. Several of the tables contain lists of species. Such a list would be meaningless in itself, but these lists compare the species in one part of the world with those in other parts, allowing global and evolution-ary patterns of biodiversity to be seen. to my knowl-edge, such lists are available in no other publication.
Humans live in a world of species, and we depend upon them not only for our physical survival but our psychological health. We need to save these other species and the ecological communities that they form. We will save only what we love, and we will love only what we closely observe and learn about. i intend this encyclopedia to help students and general readers learn about and love the world of biodiversity.
See also biodiversity, resources for agri-culture; biodiversity, resources for medicine;
Introduction
Albrecht Dürer painted The Great Piece of Turf in 1503. The lawn depicted in this painting contains spe-cies such as dandelions and plantains, which most people today consider to be weeds and which are seldom tolerated in modern lawns. (Graphische Sam-mlung Albertina, Vienna)
xviii+598_Biodiversity_F.indd 11 11/17/11 1:12 PM

first entryxiixii Introduction
ecosystem services; environmental psychol-ogy; mass extinctions.
Further readingcarson, Rachel. Silent Spring. new york: Houghton Mif-
flin, 1962/2002.Darwin, charles. On the Origin of Species by Means of
Natural Selection, 1st ed. london: John Murray, 1859. available online. URl: http://darwin-online.org.uk/content/frameset?itemiD=F373&viewtype=side&p ageseq=1. accessed July 7, 2011.
louv, Richard. Last Child in the Woods: Saving Our Chil-dren from Nature Deficit Disorder. chapel Hill, n.c.: algonquin Books, 2008.
McKibben, Bill. The End of Nature. new york: Random House, 1990/2006.
Millennium ecosystem assessment. “Millennium ecosys-tem assessment.” available online. URl: http://www.maweb.org/en/index.aspx. accessed December 4, 2010.
Rands, Michael R. W., et al. “Biodiversity conserva-tion: challenges beyond 2010.” Science 329 (2010): 1,298–1,303.
Wilson, edward O. The Diversity of Life. cambridge, Mass.: Harvard University Press, 1992.
———. The Creation: An Appeal to Save Life on Earth. new york: norton, 2006.
xviii+598_Biodiversity_F.indd 12 11/17/11 1:12 PM

xiii
the national Research council has identifi ed life sciences content standards for grades 9–12. entries in this encyclopedia are relevant to four of the con-tent standards (c, D, F, and G) established by the national Research council.Standard c includes:
● Biological evolution ● interdependence of organisms ● Behavior of organisms
the encyclopedia entries that provide information for this standard, which includes the diversity of organisms, are the following:
adaptationadaptive radiationamphibiansanimalsarchaeaarthropodsbacteriabiodiversity, concept ofbiodiversity, patterns ofbiodiversity, quantifi cation ofbiogeographybiological classifi cationbirdsbogs, life inboreal and subalpine forestscambrian explosioncambrian period, biodiversity incamoufl age and mimicrycarboniferous period, biodiversity incarnivorous plantscladisticscloud forestscoevolutioncommensalismcommunities, ecologicalcompetition, ecologicalconifers
continental shelves, biodiversity onconvergencecoral reefscretaceous extinctioncretaceous period, biodiversity incrustaceansDarwin’s fi nchesdecomposersdeep oceans, biodiversity indeserts, cooldeserts, polardeserts, warmDevonian period, biodiversity indicotsdinosaursdisjunct speciesdisturbances, ecologicalDna studies of biodiversityecologyecosystemsediacaran organismsendangered speciesendemic specieseukaryotesextinctionfi shesfl owering plants (angiosperms)fossils and fossilizationfounder effect and genetic driftfungiGalápagos islandsgrasslands and savannasherbivoryhybridizationice agesinsectsisland biogeographyJurassic period, biodiversity inkeystone specieslichenslife history, diversity ofliving fossils
The National Science Education Standards
xviii+598_Biodiversity_F.indd 13 11/17/11 1:12 PM

first entryxivxiv The National Science Education Standards
mammalsmarshes and swampsmass extinctionsmonocotsmutualismnatural selectionneogene period, biodiversity inOrdovician period, biodiversity inPaleogene period, biodiversity inparasitismPermian extinctionPermian period, biodiversity inPleistocene extinctionpollinationpredationprimatesprotistsQuaternary period, biodiversity inreptilesrivers, streams, ponds, and lakesseed dispersalseedless plantsseed plantssexual selectionshrublandsSilurian period, biodiversity inspeciationspiderssuccession, ecologicalsymbiogenesistemperate coniferous foreststemperate deciduous foreststemperate dry foreststemperate rain foreststriassic period, biodiversity intropical rain forests and deforestationtropical seasonal foreststundra
Standard D includes:
● energy in the earth system ● Geochemical cycles ● Origin and evolution of the earth system
the encyclopedia entries that provide information for this standard are the following:
biogeochemical cyclescarbon cycleecosystemsGaiagreenhouse effectgreenhouse effect and biodiversityhydrological cycle
ice agesnitrogen cycle
Standard F includes:
● Personal and community health ● Population growth ● natural resources ● environmental quality ● natural and human-induced hazards ● Science and technology in local, national,
and global challenges
the encyclopedia entries that provide information for this standard are the following:
acid rain and fogagriculturebiodiversity, resources for agriculturebiodiversity, resources for medicinebiomagnificationbiomimicrybioremediationconservation biologycultural diversityecosystem servicesendangered speciesenvironmental ethicsfederal government role in conservation (United
States)greenhouse effectgreenhouse effect and biodiversityhabitat fragmentationintegrated pest management (iPM)invasive speciesnatural systems agriculturepollutionpopulation biologypopulation geneticsresistance, evolution ofrestoration ecologytropical rain forests and deforestation
Standard G includes:
● Historical perspectives
the encyclopedia entries that provide information for this standard are the following:
Bates, Henry Waltercarson, RachelDarwin, charlesGrant, Peter and RosemaryHumboldt, alexander von
xviii+598_Biodiversity_F.indd 14 11/17/11 1:12 PM

xvThe National Science Education Standards
Jackson, WesJanzen, Daniel H.leopold, aldolinnaeus, carolusMaathai, WangariMargulis, lynnMuir, JohnPinchot, Giffordthoreau, Henry DavidWallace, alfred RusselWilson, edward O.
Further readingnational Research council. National Science Education
Standards. Washington, D.c.: national academies Press, 1996. available online. URl: http://www.nap.edu/openbook.php?isbn=0309053269. accessed Janu-ary 6, 2011.
xviii+598_Biodiversity_F.indd 15 11/17/11 1:12 PM

xviii+598_Biodiversity_F.indd 16 11/17/11 1:12 PM

Entries A–Z
xviii+598_Biodiversity_F.indd 17 11/17/11 1:12 PM

xviii+598_Biodiversity_F.indd 18 11/17/11 1:12 PM

1
A
acid rain and fog acid rain and fog, also called acid precipitation, occur when air pollution acidifi es the water droplets in clouds and the resulting acid comes in contact with objects and organisms. the major air pollutants that cause acid precipitation are nitric acid (HnO3) and sulfuric acid (H2SO4), which are the dissolved forms of nitrous and nitric oxide and sulfur dioxide gases, although natural sources of pollutants include wetlands and volcanoes. Sul-fur dioxide is particularly common in the smoke from power plants that burn high-sulfur coal. Many power plants now have “scrubbers” that remove sul-fur dioxide from the effl uent before it is released into the atmosphere. in some cases, rain downwind from industrial areas has been as acidic as orange juice or vinegar. the record pH is 2.4. (Pure water is pH 7, and changes in pH represent orders of magnitude of difference in acidity.)
acid rain and fog can be considered the unin-tended consequence of the control of particulate air pollution (soot) in the second half of the 20th century. Soot helps to neutralize some of the acid. By producing less soot, power plants caused more acid-producing compounds. Moreover, by using taller smokestacks, power plants displaced air pollution away from cities and into layers of air that brought acid rain and fog into the mountains.
acid dissolves limestone. Before the limitation of air pollution in cities, acid rain was steadily dis-solving limestone statues and architecture. acid rain has also been reported to damage the fi nish on auto-mobiles. the major impact of acid precipitation has been on biodiversity, as in these examples.
● acid rain has the potential to cause direct damage to plants by harming the waxy
cuticles of leaves and penetrating into the photosynthetic tissues. this kind of damage has not occurred very extensively, however, because only the fi rst few minutes of a rain event are signifi cantly acidic; thereafter, rain that is less acidic rinses much of the pre-viously deposited acid away before it can damage plants.
● acid rain has the potential to remove nutri-ents from soils. the negatively charged sur-faces of soil clay particles attract and hold positively charged ions such as potassium and magnesium, which are plant nutrients. Hydrogen ions (which are the cause of acid-ity) displace the plant nutrients, thus reducing the fertility of the soil. Moreover, some toxic ions such as lead and aluminum, normally scarce in soil water, may become common if the water is acidifi ed. this does not occur, however, in soils derived from limestone, as the limestone will neutralize the acidic rain.
● acid rain has the potential to accumulate in lakes, thereby reducing the growth of algae and making less food available for the entire food chain. acidity can directly affect some kinds of fi sh, particularly their eggs. trout are especially sensitive to acidi-fi ed water. this does not occur in lakes fed by groundwater that fi lters through limestone.
● acid precipitation has been most harmful in the form of acid fog. acidifi ed droplets in clouds settle on the leaves of plants, espe-cially coniferous trees of mountains, and remain for long periods of time. Often, these trees die. extensive areas of forest in
xviii+598_Biodiversity_F.indd 1 11/17/11 1:12 PM

first entry2 adaptation 2
europe (such as the forests of Switzerland and the Black Forest of Germany) have suf-fered from acid fog. in many cases, acid fog is an indirect cause of death. Much of the subalpine fir forest near Mount Mitchell, at the summit of the appalachians, has died from disease introduced by woolly adelgid bugs (Adelges tsugae). the firs normally resist the bugs—unless they are weakened by acid fog.
acid precipitation was identified as a major prob-lem during the 1970s. congress passed the acid Deposition act in 1980, which established a 10-year study. the Reagan administration convened the acid Rain Peer Review Panel of the national academy of Sciences. they then insisted that a scientist notorious for his outspoken antienvironmental views, S. Fred Singer, be included on the panel. Singer’s fame arose from his work on nuclear weapons three decades earlier, and he did not have recognized expertise in the study of acid pollution. He claimed, with-out adequate study, that controlling the acid rain problem would devastate the american economy and cause utility prices to increase immensely. the panel, with the exception of Singer, concluded that acid rain was a serious problem and that utilities and industry should reduce sulfur emissions. the Reagan admin-istration’s Office of Science and technology Policy, however, altered the executive summary of the report and focused on Singer’s viewpoint as the major con-clusion of the panel. as a result, meaningful action on acid rain was delayed until the administration of George H. W. Bush. congress passed amendments to the clean air act in 1990, establishing a system of “cap and trade” to encourage industrial polluters to limit their emissions. corporations could comply with the law by reducing emissions or by buying pol-lution credits from other corporations that produced fewer emissions than those permitted by law. the economic devastation predicted by Singer did not occur, and the price of electricity even decreased slightly. Singer continues his attacks on environmen-tal issues, presently focusing on global warming.
although sulfur dioxide emissions have decreased by 40 percent since the law took effect and acid rain has decreased 65 percent since 1976, it remains a serious problem. the ecosystem ecologist Gene likens indicated in 2009 that the forests of northeastern north america, damaged by acid rain, have not yet begun to grow back. the cap and trade system apparently worked, but not well enough to reverse the problem.
acid precipitation is also an international prob-lem, as the air pollution of one country (such as the United States or Germany) can cause the death of
forests in another (such as in canada or Switzerland). the control of air pollution, particularly of sulfur dioxide, has reduced the impact of acid precipitation since the 1980s. Due to pressure from industry, the U.S. government has been reluctant to call attention to acid precipitation. For example, national Park Service displays in Shenandoah national Park attrib-uted the death of subalpine fir trees to the adelgid bugs, not mentioning the effects of acid fog.
See also boreal and subalpine forests; greenhouse effect; pollution.
Further readingGalloway, J. n., et al. 1987. “acid rain: a comparison of
china, United States and a remote area.” Science 236 (1987): 1,559–1,562.
Kizlinski, M. l., et al. “Direct and indirect ecosystem con-sequences of an invasive pest on forests dominated by eastern hemlock.” Journal of Biogeography 29 (2002): 1,489–1,503.
likens, G. e., and F. H. Bormann. “acid rain: a serious regional environmental problem.” Science 184 (1974): 1,176–1,179.
———, et al. “long-term effects of acid rain: Response and recovery of a forest ecosystem.” Science 272 (1996): 244–246.
———, and Jerry F. Franklin. “ecosystem thinking in the northern forests—and beyond.” BioScience 59 (2009): 511–513.
Menon, Manoj, et al. “effects of heavy metal soil pollution and acid rain on growth and water use efficiency of a young model forest ecosystem.” Plant and Soil 297 (2007): 171–183.
Mphepya, J. n., et al. “Precipitation chemistry and wet deposition in Kruger national Park, South africa.” Journal of Atmospheric Chemistry 53 (2006): 169–183.
Oreskes, naomi, and erik M. conway. “Sowing the seeds of doubt: acid rain.” chapter 3 in Merchants of Doubt: How a Handful of Scientists Obscured the Truth on Issues from Tobacco Smoke to Global Warming. new york: Bloomsbury, 2010.
adaptation adaptation is the fit of an organism to its environment, which allows successful survival and reproduction. the environment of an organism includes other organisms of the same and different species. For any physical or biological environment, many different adaptations are possible, and this has resulted in exuberant biodiversity on earth.
CharaCTErISTICS ThaT arE NoT GENETICally BaSEd
Physiologists often use the word adaptation to describe environmentally induced characteristics
xviii+598_Biodiversity_F.indd 2 11/17/11 1:12 PM

adaptation 3
rather than genetically based ones. these physiologi-cal adjustments can occur on the following three different levels:
● Physiological response occurs when an organism makes immediate physiological or behavioral changes to its environment. On a hot day, the stomata (pores) of a leaf may open. this allows water to evaporate from the leaf, making it cooler through the process of transpiration. the evaporation of water may also cool a mammal on a hot day (perspiration). the mammal may also respond behaviorally by seeking shade and by exposing its skin surface areas, allowing heat to diffuse into the air. in contrast, on a cold night, the mammal curls into a ball, reducing its heat loss. these responses typi-cally do not involve changes in gene expres-sion. instead, the molecules and structures that are already present behave differently under the changed conditions.
● Acclimatization occurs when an individual organism adjusts its physical characteristics. consider again the example of a leaf on a hot day. the leaf cells accumulate or manufac-ture molecules, which cause water to diffuse into them. the influx of water makes the leaf more resistant to wilting. Or consider humans at high elevations. When humans live at high elevations for a week or more, their bone marrow begins to manufacture more red blood cells, which compensates for the lower availability of oxygen at high ele-vations. these are not immediate responses; they require the genetically controlled manu-facture of new materials. in animals, such changes can occur also as a result of behav-ioral changes (e.g., muscle building).
● Phenotypic plasticity occurs when indi-vidual organisms adjust to their environ-ments as they develop. the leaves of plants that grow in cool, shady, moist conditions are often larger than those of plants that grow in warm, bright, dry conditions, even if all the plants are genetically identical, and the plants that grow in warm, bright, dry conditions have relatively larger root systems. this is illustrated by an experiment with the weed Abutilon theophrasti (see fig-ure on page 4). the characteristics of both kinds of leaf result from the same genes, but the genes are expressed differently in each group of plants. Or consider humans who live from childhood at high elevations. these people may develop larger lungs than
they would have had they lived at sea level; larger lungs compensate for the lower oxy-gen availability at high elevations.
Plasticity is rarely reversible and rarely changes after development of an organ or organism is com-plete. although leaves can change the amount of dis-solved substances in their cells, they cannot change their size once their development is complete. that is, acclimatization is reversible and plasticity, in this case, is not. the red blood cell count can change, but lung size cannot after childhood. in this case also, acclimatization is reversible and plasticity is not. as a result of plasticity, various cells or tissues of an organism can respond differently to the same envi-ronmental conditions. Plasticity and acclimatization are responsible for much of the phenotypic variabil-ity observed in natural environments.
not all environmentally induced differences among individuals in a population are plasticity; in some cases, they may result simply from patterns of growth. as herbaceous plants grow, they accumulate stems and roots but shed their old leaves. a bigger plant will therefore have a relatively lesser amount of leaf material. Plants that grow in the shade are smaller and have relatively more leaf material than plants that grow in bright sunlight. the greater amount of leaf material is not necessarily plasticity; it may be due solely to the smaller size of the shade plants.
Plasticity can be passed down from one genera-tion to another. For example, a large, healthy plant may produce not only more seeds but bigger seeds that contain more nutrients. these seeds may grow into plants that have different characteristics than those that grow from the seeds of smaller plants, even if they are genetically identical. Research by the botanists laura Galloway and Julie etterson showed that in populations of a forest floor wildflower, plants that had grown in deeper shade produced seeds that grew into plants that had superior growth in deeper shade compared to seeds from plants that had grown in patches of sunlight. Such “intergen-erational plasticity” may result from epigenetics, in which the expression of the gene, rather than the gene itself, changes, and this change is passed on to the next generation.
CharaCTErISTICS ThaT arE GENETICally BaSEdevolutionary and conservation biologists restrict the use of the word adaptation to the genetic results of natural selection and sexual selection. therefore, not all genetic variation is adaptation. there are several sources of genetically based variation that are not produced by selection. Some of the genetic differences among individuals, populations, and spe-cies may have resulted from historical accidents that
xviii+598_Biodiversity_F.indd 3 11/17/11 1:12 PM

fi rst entry4 adaptation 4
affected the course of evolution. the most common genes may have been the ones that got lucky, rather than being selected. Genetic drift and the founder effect are examples of historical accidents. the genes that are most common in a population or a spe-cies may simply be those that were most common when the species almost became extinct or when it was fi rst founded by a small number of individu-als. in both cases, the species experienced a genetic bottleneck in which, by chance, much of the genetic
variation was lost. consider a population of fl ower-ing plants on an island. all the fl owers might be red not because natural selection favored red fl owers on the island but because the only seeds that reached the island in the original population were those of red-fl owered individuals. Populations of many endangered species have genetic characteristics that resulted from genetic bottlenecks.
exaptations are genetically based traits that may have originally evolved because they performed one
Plants that develop in the shade grow taller and produce smaller root systems and more leaf area than plants that develop in full sunlight. Plants that develop in dry conditions grow shorter and produce larger root systems and less leaf area than plants that develop in moist conditions. This is an example of phenotypic plasticity, which is sometimes called adaptation, but which does not involve genetic changes. The numbers come from an experiment by the author using the weed Abutilon theophrasti. The three treatments are compared at the overall average weight of 854 mg.
xviii+598_Biodiversity_F.indd 4 11/17/11 1:12 PM

adaptation 5
function but then were available for another func-tion. this means that selection may not have acted directly on the trait being considered; the trait may have been a side effect of selection. a frequently cited example of an exaptation is feathers. the ear-liest birds apparently used feathers as insulation. after feathers had evolved as insulation, natural selection acted upon them as flight structures. after that, sexual selection acted upon them as colorful adornments. Feathers, therefore, can be considered adaptations for insulation and exaptations for flight and sexual adornment.
it can be difficult to distinguish adaptations from exaptations. the classic example of an adapta-tion that has appeared in numerous popular science books and textbooks for more than a century is the neck of the giraffe (see below). to most observers, it would appear obvious that long necks are an adapta-tion that allows giraffes to feed at the tops of trees, but if this is so, why do female giraffes have shorter necks than male giraffes, and why are giraffes so frequently observed actually bending down to eat leaves? Observations of giraffes in the wild have shown that males with longer necks prevail in male-to-male competition, and females choose the males with longer necks. now that giraffes have long necks, they can use them for eating leaves from tree-tops, but that may not have been the original reason that long necks evolved in these animals. the long neck of the giraffe appears to be an adaptation to combat and an exaptation for reaching leaves at the tops of trees.
Some characteristics of organisms are structur-ally inevitable. large plants must have relatively thick trunks, and large animals must have relatively
thick legs. Structurally inevitable features are called allometric characteristics and are not considered to be adaptations.
in most cases, an adaptation does not have just a single function. the glands within an animal’s epidermis have several functions, each of which may have provided a separate evolutionary advantage. Some of the glands produce sweat, which allows the animal to become cooler as the sweat evaporates. Sweat also contains dissolved molecules. the body of the animal uses sweat as one of its methods of disposing of excess or toxic materials. Molecules in sweat can also serve as chemical communication between animals. Mammary glands are modified sweat glands. Sweat glands, therefore, are an adapta-tion with at least four different functions.
adaptations are never perfect. natural and sex-ual selection produce adaptations that are only as good as they need to be. One example is the digestive system of the panda. the immediate ancestors of pandas were carnivores, but pandas are herbivores, living exclusively on leaves. Pandas have intestines that are better suited to a carnivore. in well-adapted herbivores, such as sheep, the intestines are up to 35 times as long as the body, while in well-adapted car-nivores, the intestines are much shorter, only four to seven times as long as the body. With the help of bac-teria, longer intestines allow herbivores more time to digest coarse plant materials, such as cellulose. Meat, in contrast, requires less digestive breakdown. Pandas have intestines that are in the carnivore, not the herbivore, range. the panda’s digestive system is, therefore, not well adapted to its function. Modern evolutionary scientists attribute this to the recent evolutionary shift from meat to leaves in the diet of the panda’s ancestors: there has not been time for better adaptation in this case.
adapTaTIoN aNd BIodIvErSITyadaptation is the reason there is such an astonish-ingly large number of species. if there were only one perfect adaptation for every situation, such as a dry climate, then there would be very few species. all plants in a dry climate would look more or less the same, and one species would dominate that climate. However, this is far from the case. there are many possible plant adaptations to a dry climate. Some plants, such as mesquite shrubs, grow very deep roots to draw water up from the water table. they also have small leaves, which diffuse heat into the air more quickly than do large leaves. in this way, the small-leaved shrubs can keep their leaves cool without much transpiration. Some plants do both of these things. Others, such as cactuses, have shallow roots, but they store water in their succulent tissues. Still others, such as the spring ephemeral wildflowers
Giraffes can drink only with the greatest difficulty. Their long necks appear to be as much a hindrance as an adaptation to their daily work of eating and drinking and seem to have evolved by sexual selec-tion, not natural selection.
xviii+598_Biodiversity_F.indd 5 11/17/11 1:12 PM

first entry6 adaptive radiation 6
of the desert, have neither of these adaptations. instead, they germinate and grow rapidly during the brief rainy season, surviving the long drought in the form of seeds. a similar diversity of adaptation can be seen in desert animals, some of which, such as kangaroo mice, function with very little water, while others, such as desert frogs, remain dormant in the soil between periods of rain. throughout evo-lutionary history, various evolutionary lineages have responded to environmental circumstances in many different and equally successful ways.
coevolution also produces many different adap-tations and promotes biodiversity. coevolution occurs when the evolution of one type of organism influences the evolution of another; they adapt to one another. When flowering plants evolved near the beginning of the cretaceous period, they began to depend upon many different kinds of pollination. Some depended upon the wind, but many engaged the services of animals such as insects to carry their pollen. this began an explosive diversification of flowering plant species, each evolving the ability to attract a different insect species. Meanwhile, the insects were also evolving numerous different and equally successful adaptations that allowed them to obtain food from the flowers they pollinated. evolu-tionary diversification in flowering plants and pol-linators reinforced each other. the adaptations that have resulted from coevolution have been the major source of biodiversity on earth.
See also endangered species; extinction; founder effect and genetic drift; natural selection; sexual selection.
Further readingDawkins, Richard. The Blind Watchmaker: Why the
Evidence of Evolution Reveals a Universe Without Design. new york: norton, 1996.
Galloway, laura F., and Julie R. etterson. “transgenera-tional plasticity is adaptive in the wild.” Science 318 (2007): 1,134–1,136.
Gould, Stephen Jay, and elisabeth Vrba. “exaptation—a missing term in the science of form. Paleobiology 8 (1982): 4–15.
Rice, Stanley a., and Fakhri a. Bazzaz. “Plasticity to light conditions in Abutilon theophrasti: comparing phe-notypes at a common weight.” Oecologia 78 (1989): 502–507.
adaptive radiation adaptive radiation is the evolution of species from a single, ancestral popula-tion. throughout the history of life, many species have become extinct either by natural selection or simply bad luck, while others have produced many new species by adaptive radiation from the surviving
species. adaptive radiation can result either from the dispersal of a species to a new location, after which the founding population undergoes radiation, or the survival of remnant populations of a once wide-spread species, after which the remnants undergo radiation. the net result has been a steady increase in biodiversity through evolutionary time. adaptive radiation occurs because a single ancestral popula-tion separates into distinct populations that do not interbreed, thus allowing separate directions of evo-lution to occur in each.
the world is full of numerous examples of adap-tive radiation. nearly every genus that contains more than one species or any family that contains more than one genus can be considered an example. two examples follow.
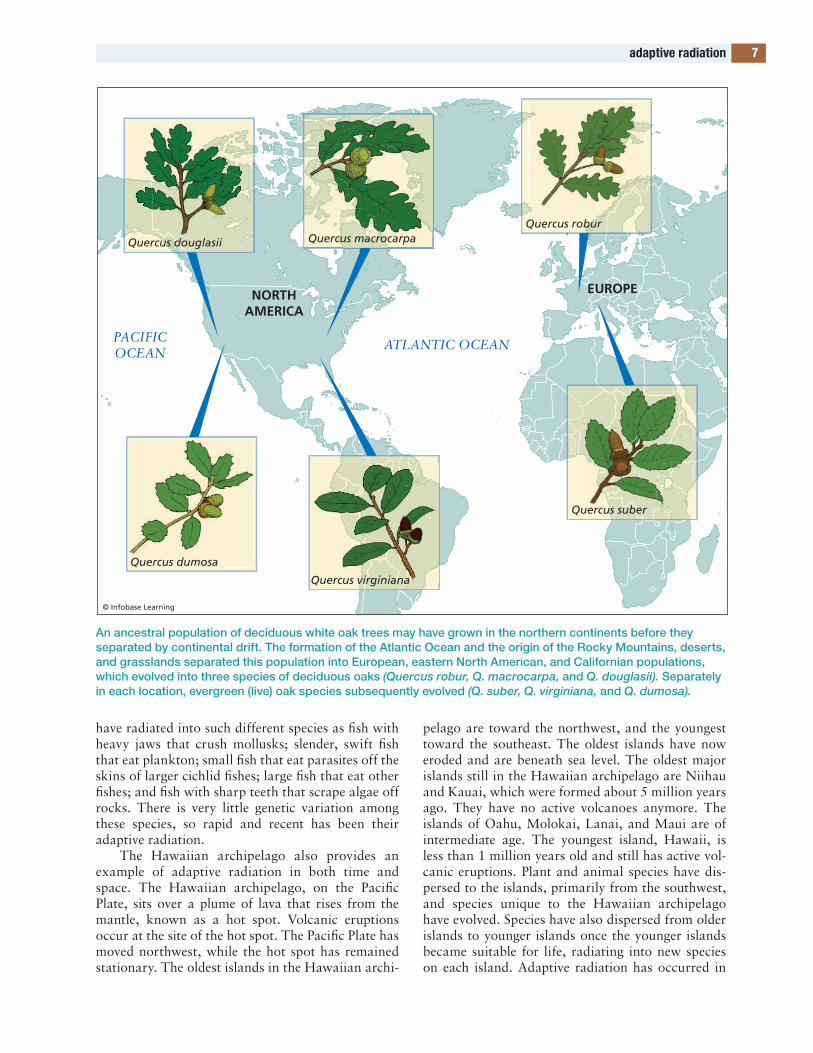
First, consider the white oaks, part of the genus and subgenus Quercus. at one time, probably before 80 million years ago, there was a single species of white oaks that may have lived in what is now east asia. in the late Mesozoic and cenozoic eras, warm conditions were widespread on earth. Oaks spread throughout the northern Hemisphere. three of the locations in which they are now found are europe, eastern north america, and california.
north america then separated from eurasia, new mountain ranges such as the Rocky Mountains arose, and conditions became cooler and drier dur-ing the latter part of the cenozoic era. these events separated the white oaks of europe, eastern north america, and california. Separate species evolved in each of these locations, including Quercus robur (english oak) in europe, Quercus macrocarpa (bur oak) in eastern north america, and Quercus doug-lasii (blue oak) in california (see figure on page 7). Furthermore, in each of these locations, cooler con-ditions allowed the evolution of the deciduous pat-tern of leaf production in some areas, while warmer conditions encouraged the evolution of the evergreen pattern in other areas. the previously mentioned species are all deciduous. Quercus suber (cork oak) is an evergreen white oak of europe; Quercus vir-giniana (live oak) is the evergreen white oak of eastern north america; and Quercus dumosa (scrub oak) is an evergreen white oak of california. in all three areas, both deciduous and evergreen (live) oaks evolved (with some exceptions). these oak species and many others constitute an example of adaptive radiation within the white oak subgenus.
Second, while the adaptive radiation of oaks occurred over the course of about 80 million years, the adaptive radiation of the approximately 500 spe-cies of cichlid fishes of lakes Malawi, tanganyika, and Victoria in africa has occurred in just the last few thousand years. From what was probably a sin-gle ancestral species, the cichlids in lake Victoria
xviii+598_Biodiversity_F.indd 6 11/17/11 1:12 PM
adaptive radiation 7
have radiated into such different species as fi sh with heavy jaws that crush mollusks; slender, swift fi sh that eat plankton; small fi sh that eat parasites off the skins of larger cichlid fi shes; large fi sh that eat other fi shes; and fi sh with sharp teeth that scrape algae off rocks. there is very little genetic variation among these species, so rapid and recent has been their adaptive radiation.
the Hawaiian archipelago also provides an example of adaptive radiation in both time and space. the Hawaiian archipelago, on the Pacifi c Plate, sits over a plume of lava that rises from the mantle, known as a hot spot. Volcanic eruptions occur at the site of the hot spot. the Pacifi c Plate has moved northwest, while the hot spot has remained stationary. the oldest islands in the Hawaiian archi-
pelago are toward the northwest, and the youngest toward the southeast. the oldest islands have now eroded and are beneath sea level. the oldest major islands still in the Hawaiian archipelago are niihau and Kauai, which were formed about 5 million years ago. they have no active volcanoes anymore. the islands of Oahu, Molokai, lanai, and Maui are of intermediate age. the youngest island, Hawaii, is less than 1 million years old and still has active vol-canic eruptions. Plant and animal species have dis-persed to the islands, primarily from the southwest, and species unique to the Hawaiian archipelago have evolved. Species have also dispersed from older islands to younger islands once the younger islands became suitable for life, radiating into new species on each island. adaptive radiation has occurred in
An ancestral population of deciduous white oak trees may have grown in the northern continents before they separated by continental drift. The formation of the Atlantic Ocean and the origin of the Rocky Mountains, deserts, and grasslands separated this population into European, eastern North American, and Californian populations, which evolved into three species of deciduous oaks (Quercus robur, Q. macrocarpa, and Q. douglasii). Separately in each location, evergreen (live) oak species subsequently evolved (Q. suber, Q. virginiana, and Q. dumosa).
xviii+598_Biodiversity_F.indd 7 11/17/11 1:12 PM

first entry8 agriculture 8
space (unique species on each island) and in time (from older to younger islands) in the Hawaiian archipelago.
Perhaps the most famous adaptive radiation has been the evolution of Darwin’s finches in the Galá-pagos islands. a single ancestral species, which may have come from the South american mainland, has radiated out into 13 species in four genera. Finches in three of the genera live in trees. Some of them have some amazing adaptations, such as the spe-cies that uses sticks to pry insects out of holes in branches. the largest genus, with six species, con-sists of finches that hop on the ground and eat mostly seeds.
Sometimes, speciation occurs as one species coevolves in the presence of another. However, the presence of other species can preclude some direc-tions of speciation. On the Galápagos islands, pred-ators are largely absent, and some of the finches have figured out a way to drink blood from the legs of boobies (large marine birds) by pecking them. also, woodpeckers are absent, and some of the finches, as mentioned above, use sticks to get insects out of holes. in a habitat already filled with efficient preda-tors and woodpeckers, the amateurish adaptations displayed by the blood-drinking and insect-prying finches would probably not have succeeded. there-fore, adaptive radiation can occur rapidly when a population invades a new habitat—if, that is, the population is adapted to the conditions of the new habitat and if the new habitat does not have poten-tial competitors.
Release from competition is the reason adaptive radiation has occurred most spectacularly in the his-tory of earth after large disasters. During the Meso-zoic era, mammals were few in number and low in diversity. their constant body temperatures made them successful nocturnal animals. the dinosaurs, meanwhile, ruled the daytime. after the cretaceous extinction, which cleared away the dinosaurs, mam-mals underwent an explosive adaptive radiation. large mammals did not exist before the cenozoic era; any mammal species that began to evolve a large size would have been unsuccessful in competi-tion with large dinosaurs, but many huge mammals evolved in the early cenozoic. along with the big mammals, many of the modern categories of mam-mals, including whales, primates, and bats, evolved. Mammals have continued to have adaptive radia-tions, but none quite as spectacular as the adaptive radiation that occurred early in the cenozoic era.
Within species, the adaptive radiation of subspe-cies and varieties continues. these represent pos-sible future species. the preservation of biodiversity requires not just the protection of species but of subspecies and varieties as well.
See also biogeography; coevolution; Dar-win’s finches; extinction; mass extinctions; speciation.
Further readingGillespie, Rosemary G. “the ecology and evolution of
Hawaiian spider communities.” American Scientist 93 (2005): 122–131.
Grant, Peter R., and B. Rosemary Grant. How and Why Species Multiply: The Radiation of Darwin’s Finches. Princeton, n.J.: Princeton University Press, 2007.
Schilthuizen, Menno. Frogs, Flies, and Dandelions: The Making of Species. new york: Oxford University Press, 2001.
Schluter, Dolph. The Ecology of Adaptive Radiation. new york: Oxford University Press, 2000.
agriculture agriculture is the deliberate cultiva-tion of plants or fungi by animals. cultivation here refers to planting, care, and harvest. agricultural plants and fungi are usually very different from wild plants and fungi; they have characteristics that allow them to be adapted to the agricultural system in which they live, useful to the animals that cultivate them, and poorly adapted to living in the wild. agri-culture is an example of mutualism, which confers benefits on both the agricultural plant or fungus and the animal that cultivates it. in agriculture, crops and the animals that cultivate them are mutually depen-dent. agriculture has been, perhaps, the most signifi-cant component of human cultural evolution. More broadly defined, agriculture may include the care and use of livestock animals by other animals. agriculture interacts with biodiversity in two ways: First, it has an impact upon biodiversity, as explained below, and second, it depends upon wild biodiversity as a source of new crops or genes, as explained in another entry.
EvoluTIoN of aGrICulTurESeveral species of ants carry out activities that bear striking parallels to human agriculture. leaf-cutter ants (genera Atta and Acromyrmex) cultivate gar-dens of fungus (family lepiotaceae). Massive forag-ing parties of leaf-cutter ants gather pieces of leaves from many species of tropical plants and carry them back to their nests. leaf-cutter ants are the single most important source of herbivory in rain forest areas where they are abundant. the ants do not eat the leaves, which contain many toxins. instead, they chew them up into compost, on which a kind of fun-gus grows and breaks down the toxins. the fungus grows nowhere else except in the mounds of leaf-cutter ants; when the ants disperse, they take fungus tissue with them. the ants eat fungus tissue and almost nothing else. Beneficial bacteria that grow on
xviii+598_Biodiversity_F.indd 8 11/17/11 1:12 PM

agriculture 9
the bodies of the ants produce chemicals that inhibit the growth of other bacteria in the compost. For this reason, some biologists consider these and other ants to be a promising source of new antibiotics. Because the ants deliberately prepare compost for the fungus and because of the mutual dependence of ants and fungus upon each other, the ant-fungus relationship can be considered an example of agriculture.
Several species of ants in the seasonal tropics of central america live on and in trees of the genus Acacia. the ants do not eat the leaves of the acacia. instead, they consume nectar that is produced by glands on the stems (not in the flowers) of the trees, and they eat globules of protein and fat, called Beltian bodies, that grow on the tips of immature leaves. the ants chew out the insides of the acacias’ unusually large thorns and live inside the thorns. in some cases, experimental manipulation has shown the ants to be dependent upon the acacias for survival. the ants attack and kill other insects and drive away larger animals that attempt to feed on the acacias. When vines or other plants begin to grow in the immedi-ate vicinity of the acacias, the ants chew them down. in most cases, the acacias are completely dependent upon the ants; when the ants are experimentally removed, vines overgrow the acacias, and animals browse the leaves heavily. the acacias remain green during the dry season, when most of the other trees lose their leaves, but these green targets go undis-turbed by herbivores because of their protective ant army. Because the ants weed out other plants from the vicinity of the acacias and defend their crops and because of the mutual dependence of ant and acacia, the ant-acacia relationship can be considered an example of agriculture. ant partnerships with other kinds of trees in the rain forests of africa have evolved independently of the ant-acacia partnership.
as natural selection favored the characteristics that allowed the partnerships of ants with fungi and ants with acacias, hybridization between these species and others closely related to them was unsuc-cessful. the result was the evolution of new species of fungi, acacias, and ants. in this way, agriculture has increased biodiversity.
as the above examples illustrate, when agri-culture evolved in the human species about 10,000 years ago, it was not the first time that agriculture had evolved on the planet. Hereafter, agriculture will refer to human agriculture.
Some scholars once believed that agriculture was invented by a brilliant man in a tribal society of hunter-gatherers. Other scholars pointed out that since women gathered most of the plant materials, agriculture was probably invented by a woman. Both the brilliant man theory and the brilliant woman theory are incorrect, however, because agriculture
could not have been invented in a single step by anyone. it had to evolve. Unmodified wild plants are unsuitable for agriculture. the following are four reasons for this.
● the seeds of most wild plant species are dor-mant when they are mature; that is, when planted, the seeds will not germinate. Many require a period of exposure to cool, moist conditions before the inhibitors within the plant embryo break down and the seed germinates. if a brilliant man or woman planted the first agricultural seed from a wild plant, it would not have grown, and the person would have rightly concluded that agriculture was not a good idea.
● the seeds of many wild grains shatter or fall off the stem as soon as they are mature. Since natural selection generally favors seeds that disperse to new locations, shattering is beneficial to the plant, but it is extremely inconvenient for a human harvester.
● the seeds of many wild plant species con-tain toxins.
● the seeds of many wild grains are small.
Furthermore, the advantages of primitive agri-culture would not have been immediately apparent to intelligent hunter-gatherers. agriculture requires intense labor. Modern hunter-gatherers often barely eke out an existence in marginal habitats such as the Kalahari Desert of africa or the Great Outback of australia, but these are the habitats to which tribes and nations with more advanced tools have driven them. Before agriculture, many tribes lived in rich habitats in which hunting and gathering in many cases provided a comfortable level of existence. For these reasons also, agriculture had to evolve gradually.
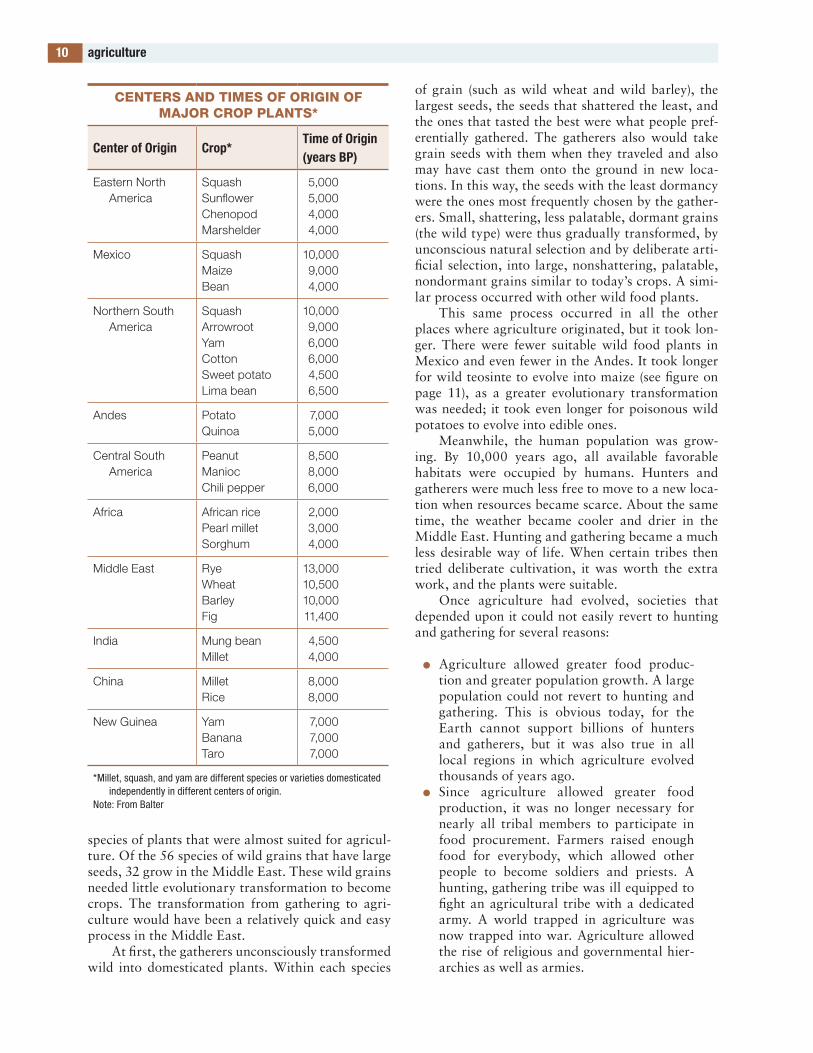
agriculture originated separately in several parts of the world: at least twice in the new World (cen-tral america and South america), in Mesopotamia, and in china. agriculture may have had several sepa-rate origins in each of these areas, as well as in new Guinea. the only inhabited part of the world in which agriculture did not evolve was europe; euro-pean agriculture was imported from the Middle east. Scientists and historians know that agriculture origi-nated separately in these locations because the staple crops were different in each (see table on page 10). the domesticated sumpweeds and sunflowers grown by inhabitants of north america were replaced by Mexican maize and beans in pre-columbian times.
agriculture began earliest (about 10,000 years ago) in the Middle east, especially in the tigris-euphrates floodplain of Mesopotamia. it began there first apparently because there were many wild
xviii+598_Biodiversity_F.indd 9 11/17/11 1:12 PM
first entry10 agriculture 10
species of plants that were almost suited for agricul-ture. Of the 56 species of wild grains that have large seeds, 32 grow in the Middle east. these wild grains needed little evolutionary transformation to become crops. the transformation from gathering to agri-culture would have been a relatively quick and easy process in the Middle east.
at first, the gatherers unconsciously transformed wild into domesticated plants. Within each species
of grain (such as wild wheat and wild barley), the largest seeds, the seeds that shattered the least, and the ones that tasted the best were what people pref-erentially gathered. the gatherers also would take grain seeds with them when they traveled and also may have cast them onto the ground in new loca-tions. in this way, the seeds with the least dormancy were the ones most frequently chosen by the gather-ers. Small, shattering, less palatable, dormant grains (the wild type) were thus gradually transformed, by unconscious natural selection and by deliberate arti-ficial selection, into large, nonshattering, palatable, nondormant grains similar to today’s crops. a simi-lar process occurred with other wild food plants.
this same process occurred in all the other places where agriculture originated, but it took lon-ger. there were fewer suitable wild food plants in Mexico and even fewer in the andes. it took longer for wild teosinte to evolve into maize (see figure on page 11), as a greater evolutionary transformation was needed; it took even longer for poisonous wild potatoes to evolve into edible ones.
Meanwhile, the human population was grow-ing. By 10,000 years ago, all available favorable habitats were occupied by humans. Hunters and gatherers were much less free to move to a new loca-tion when resources became scarce. about the same time, the weather became cooler and drier in the Middle east. Hunting and gathering became a much less desirable way of life. When certain tribes then tried deliberate cultivation, it was worth the extra work, and the plants were suitable.
Once agriculture had evolved, societies that depended upon it could not easily revert to hunting and gathering for several reasons:
● agriculture allowed greater food produc-tion and greater population growth. a large population could not revert to hunting and gathering. this is obvious today, for the earth cannot support billions of hunters and gatherers, but it was also true in all local regions in which agriculture evolved thousands of years ago.
● Since agriculture allowed greater food production, it was no longer necessary for nearly all tribal members to participate in food procurement. Farmers raised enough food for everybody, which allowed other people to become soldiers and priests. a hunting, gathering tribe was ill equipped to fight an agricultural tribe with a dedicated army. a world trapped in agriculture was now trapped into war. agriculture allowed the rise of religious and governmental hier-archies as well as armies.
Centers and times of origin of major Crop plants*
Center of origin Crop*Time of origin (years Bp)
Eastern North America
SquashSunflowerChenopodMarshelder
5,0005,0004,0004,000
Mexico SquashMaizeBean
10,0009,0004,000
Northern South America
SquashArrowrootYamCottonSweet potatoLima bean
10,0009,0006,0006,0004,5006,500
Andes PotatoQuinoa
7,0005,000
Central South America
PeanutManiocChili pepper
8,5008,0006,000
Africa African ricePearl milletSorghum
2,0003,0004,000
Middle East RyeWheatBarleyFig
13,00010,50010,00011,400
India Mung beanMillet
4,5004,000
China MilletRice
8,0008,000
New Guinea YamBananaTaro
7,0007,0007,000
*Millet, squash, and yam are different species or varieties domesticated independently in different centers of origin.
Note: From Balter
xviii+598_Biodiversity_F.indd 10 11/17/11 1:12 PM

agriculture 11
● With the evolution of agriculture, produc-tive farmland became valuable. People settled into cities because they had to stay in one place at least long enough for one harvest. agriculture promoted the rise of civilization. civilization rose earlier in Mes-opotamia than in other places because agri-culture began earlier there. With civilization came advanced technology. Because they needed to defend particular tracts of terri-tory, the armies now had a lot more to fight about. the cultural groups that developed agriculture first were the first to be civi-lized and to have advanced technology that allowed them to conquer the cultural groups in which this process had not progressed as far. the biologist Jared Diamond explains that this is why europeans conquered amer-ica and drove natives onto reservations, rather than native americans conquering europe and driving europeans into remote corners of the alps and Pyrenees.
the evolution of agriculture is a perfect illustra-tion that evolution does not operate for the good of the species. agriculture evolved in the human spe-
cies because it provided an advantage to some indi-viduals within human societies. agriculture did not improve the average health of human beings. in fact, the average life span in early agricultural societies was shorter than that in contemporaneous hunter-gatherer societies. this occurred for two reasons:
● Diseases spread more rapidly in cities, in which people were trapped with one anoth-er’s wastes, garbage, and germs.
● agriculture actually decreased the quality of human nutrition by making people depen-dent upon a few crop plants rather than a diversity of wild foods. in particular, the human body evolved under conditions in which ascorbic acid (vitamin c) was readily available from wild fruits. When entire pop-ulations became dependent upon crops with little vitamin c, scurvy became a way of life.
Herding began when people started to manage herds of wild animal species that were most amenable to survival and reproduction under captivity. Uncon-scious natural selection and then conscious artificial selection resulted in the evolution of livestock species such as the cow, which evolved in western eurasia
Native American farmers developed modern maize from wild teosinte grass by selecting for larger ears (which are clusters of female flowers). (Nicole Rager Fuller, National Science Foundation)
xviii+598_Biodiversity_F.indd 11 11/17/11 1:12 PM

first entry12 agriculture 12
and Southeast asia from two distinct wild species. Once again, the earlier development of herding in the Middle east than in other areas occurred because many wild animal species of the Middle east such as goats and sheep were amenable to herding, whereas wild animal species such as deer in north america were not. livestock animals provided high-quality food (meat and milk), often by consuming wild foods that humans could not eat. this was especially true of goats. Pigs, on the other hand, eat many of the same foods as humans. this competition between pigs and humans for food may be one reason that pigs are considered undesirable (“unclean”) by some cultures in the arid regions of the Middle east. as with agriculture, herding caused a narrowing of the food diversity base, from many wild animal species to a few livestock species.
EffECTS of aGrICulTurE oN BIodIvErSITy of NaTural SySTEmS
agriculture has had several effects upon biodiversity, as indicated by the following examples.
Escape of agricultural species into natural eco-systems. the result of agriculture has been the evolu-tion of many new species of plants and animals that are adapted to human care and often cannot survive in the wild. in this sense, agriculture has created new biodiversity, even though crop species do not grow well in the wild. For example, since wheat (Triticum aestivum) and maize (Zea mays) do not have seed dormancy and do not shatter their seeds, they can usually survive only where they have been planted. However, crop species have contributed, in a largely negative way, to natural biodiversity because wild relatives of cultivated plants are frequent invaders of natural ecosystems. examples include Johnson grass (Sorghum halepense), a relative of cultivated sor-ghum (S. bicolor), and wild oats (Avena fatua and A. barbata), relatives of cultivated oats (A. sativa). Hybridization between crops and wild relatives can produce new, aggressive strains of weeds. the ecolo-gists norm ellstrand and christina Schierenbeck have listed 28 examples. the new weeds are particu-larly hard to control since they so closely resemble the crops they infest. this same process may endan-ger some of the wild relatives also. Some varieties of wild maize in Mexico have been essentially hybrid-ized out of existence.
the use of GMO (genetically modified organism) crops became widespread in the 1990s. For some crops, such as soybean and cotton, most of the acre-age consists of GMO breeds. Many GMO crops have wild relatives with which they can interbreed. For example, cultivated mustards such as canola (domesti-cated Brassica napus and B. campestris) can exchange pollen with wild mustard species. this raises the pos-
sibility that modern agriculture may introduce genetic changes, such as herbicide resistance or the produc-tion of compounds that are toxic to insect herbivores, into some wild populations. Wild, weedy mustards with leaves that are toxic to insects might undergo population explosions as a result of release from her-bivore damage, and the herbivores might suffer a pop-ulation crash. Such “genetic pollution” has occurred, but not yet on a large scale.
numerous domesticated animals have escaped into the wild. livestock animals, though they have also been artificially selected to live under artifi-cial conditions, can thrive under wild conditions. though cows (Bos taurus) are not so well adapted to survival in the wild as were their wild ancestors, they may form extensive wild herds in grasslands from which natural predators such as wolves have been exterminated. the same is true of feral horses (Equus caballus) and donkeys (Equus asinus). Most animals that have been bred as pets are incapable of surviving in the wild, though dogs (Canis familiaris) and especially cats (Felis catus) have formed large and successful feral populations. Feral cats kill hun-dreds of millions of songbirds in the United States each year. these feral populations often lose some of their domesticated characteristics and quickly revert to the characteristics of their wild ancestors.
escaped livestock have had a particularly strong impact upon wild ecological communities. Most notorious are wild pigs and goats. Feral pigs (Sus scrofa), such as the razorbacks of the forests of the southeastern United States, trample and tear up woodlands and wetlands through which they wander in search of food. Wild pigs have caused extensive damage on islands with moist climate, such as the Hawaiian islands, in which the plants have evolved no protection against grazing animals. escaped goats (Capra aegagrus) have had an even more severe impact upon islands with dry climate, such as the channel islands of california and the Galápagos islands of ecuador, though they also thrive in moist climates. a population explosion of sheep (Ovis aries), since their introduction in about 1820, has transformed tasmania, where plants that cannot grow back after grazing have been replaced by grasses that can in many areas.
Destruction of natural habitats for agriculture. large areas—perhaps a third of the dry land of earth—have been transformed into farmland and pasture. as the human population grows, so will the demand for food. expansion of agricultural land is not a good option to meet this demand, as most of the natural habitat that is suitable for agriculture has already been used. Most of what remains is too cold or too dry or has soil that is too poor for good agricultural production. Moreover, most of the pro-
xviii+598_Biodiversity_F.indd 12 11/17/11 1:12 PM

agriculture 13
duction of the principal crops in the United States is used to feed livestock. More people could be fed if farmland were used to raise food that people could eat rather than to raise livestock feed.
the transformation of natural habitats into agricultural land has caused the extinction of many tropical plant and animal species that formerly had very limited ranges; several species of wild plants and animals can be driven to extinction by the develop-ment of a single oil palm plantation. temperate zone plant and animal species generally have greater geo-graphical ranges, and extinctions as a result of agri-culture in this zone are less common. nevertheless, agriculture, like urban sprawl, has brought about the fragmentation of natural ecosystems and a reduction in the size and genetic diversity of wild populations.
agriculture need not have such a negative impact on biodiversity. Hedgerows of shrubs and trees around small agricultural fields provide habitat for predatory birds, bats, spiders, and insects that prey upon insect pests. these predators may not venture out into the middle of large fields because other predators may detect and eat them, or they may simply be killed by insecticides. an agricultural landscape of small fields surrounded by hedgerows will have a much greater biodiversity (and require fewer pesticides) than will large fields devoted to one species of crop. Organic agriculture uses hedgerows and natural pest control. Because they use natural rather than chemical fertilizers, organic farms also have richer soil, which enhances the diversity of soil organisms, including insects and earthworms.
Impact of agriculture on natural ecosystems. agriculture is not simply the replacement of one type of habitat by another. agricultural fields are usually a monoculture of a single crop, perhaps just a single breed of crop. the animals that live in these mono-cultures are also less diverse and consist primarily of insect pests of the crops. Monocultures encour-age the spread of pests and diseases. this has two effects. First, some of these pests, such as grasshop-pers and locusts, can grow into swarms that result in the defoliation of nearby natural ecosystems. Second, monocultures require heavy applications of chemicals to control the pests. all the plants in the field use the same nutrients from the same soil depth, which requires increased dependence on chemical fertilizers. Monoculture also encourages erosion of the soil by wind and water. Because most crops are annuals, large fields are left barren during the non-productive season.
Mainstream agriculture is considered a “high-input” system because a lot of energy and chemi-cals are necessary. Many of these chemicals pollute surrounding ecosystems. Fertilizer from farms, par-ticularly phosphate, is a major contributor to the
explosive growth of cyanobacteria in streams and lakes, resulting in eutrophication, which greatly alters the food chain of such wetland ecosystems. the decomposition of dead cyanobacteria con-tributes to oxygen depletion in wetlands and on ocean bottoms. the huge feedlots used for livestock produce as much sewage as do human cities, and much of this waste also contributes to pollution and eutrophication. the writer Wendell Berry compared preindustrial agriculture, in which animal wastes fertilized the fields, with industrial agriculture, in which crops are raised with artificial fertilizers, while animal wastes become a source of pollution. industrial agriculture, he said, has taken a solution and divided it into two problems.
Many of the environmental problems of modern agriculture can be avoided by increasing the diver-sity of crops and by the use of perennial rather than annual crops. the fact that leguminous crops, such as beans and alfalfa, benefit soil fertility has been known since the time of the Roman writer Virgil. leguminous crops have nitrogen-fixing bacteria in their roots and can actually enrich the soil with nitrogen fertilizer rather than depleting it. terraces, used for millennia in east asia for rice production, not only reduce soil erosion but also allow organic matter in the soil to accumulate, thereby enriching it.
crop rotations, or rotations involving both crops and livestock, also avoid many environmen-tal problems. Fred Kirschenmann, director of the aldo leopold center at iowa State University, has shown that crop rotations in organic agriculture can be successful on a large scale. On a large farm in north Dakota, Kirschenmann grows durum wheat, rye, flax, sunflower, buckwheat, and oats. He uses alfalfa and two species of sweet clover as cover crops and has organic pasture. He alternates warm with cool season crops and cereals with broadleaf crops, thus avoiding buildup of pests. By rotating shallow-rooted with deep-rooted crops, he avoids depleting the soil layers. crop rotation often means that the farmer must give up a marketable crop yield for one year in four while cover crops grow, but lower chemical and energy costs compensate for this loss. the writer Michael Pollan describes the example of argentina, where the agricultural industry raises cattle on grassland for five years, followed by a three-year production of grains or beans. the grass-fed cattle produce high-quality beef, and the cattle wastes provide fertilizer for the crops. insect pests are a relatively small problem, since the insects that attack the crops cannot survive in pastures.
Many GMO crops have been bred to resist herbi-cides. in the past, a farmer could not spray herbicide over a whole field because it would kill the crops as well as the weeds, but now an extensive application
xviii+598_Biodiversity_F.indd 13 11/17/11 1:12 PM

first entry14 agriculture 14
of herbicide will kill only the weeds. the result has been a vast increase in the use of herbicides, which can spread into surrounding natural ecosystems.
Effects of agriculture on global climate. agricul-ture is one of the major sources of greenhouse gases. a tremendous amount of fossil fuel is used in modern systems of food production, from the production of fertilizers and pesticides, to the use of farm equip-ment, to the processing of food. When soil is left bare following the harvest of annual crops, decom-position can release carbon dioxide into the atmo-sphere that was previously sequestered in the humus. agriculture may also be the major source of atmo-spheric methane, another greenhouse gas. Flooded soils, such as rice paddies, and the digestive systems of cows produce a lot of methane. the tremendous agricultural production of greenhouse gases need not occur. Plants can be considered a way of removing carbon dioxide from the air and storing it in the soil as leaves, stems, and roots die and decompose into humus. agriculture based on perennials can store carbon in the soil rather than releasing it.
Modern agriculture might appear to be very efficient (see figure). However, it is efficient only in terms of the amount of food produced per farmer. in every other way, it is wasteful. By some calculations, only 4 percent of americans are farmers or farm-workers, but, as the writer Wendell Berry explains
. . . the four percent of our population that is left on the farm does not, by any stretch of imagination, feed the rest. that four percent is only a small part, the worst-paid part, of a food production network that includes purchas-
ers, wholesalers, retailers, processors, packag-ers, transporters, and the manufacturers and salesmen of machines, building materials, feeds, pesticides, herbicides, fertilizers, medicines, and fuels. . . . [and they] are all dependent on the petroleum industry.
For this reason, agriculture puts a lot of carbon dioxide into the atmosphere.
in addition to being environmentally destruc-tive, modern agriculture is also precarious, because it is dependent on the uninterrupted availability of inexpensive fossil fuels. For numerous environmen-tal and economic reasons, humans need to change the way they raise their food.
See also biodiversity, resources for agri-culture; biodiversity, resources for medicine; greenhouse effect; habitat fragmentation; invasive species.
Further readingBaker, Herbert G. Plants and Civilization. Belmont, calif.:
Wadsworth, 1970.Balter, Michael. “Seeking agriculture’s ancient roots.” Sci-
ence 316 (2007): 1,830–1,835.Berry, Wendell. Bringing It to the Table: On Farming and
Food. Berkeley, calif.: counterpoint, 2009.Diamond, Jared. Guns, Germs, and Steel: The Fates of
Human Societies. new york: norton, 1997.Dutton, a., et al. “Prey-mediated effects of Bacillus thuring-
iensis spray on the predator Chrysoperia carnea in maize.” Biological Control 26 (2003): 209–215.
ellstrand, norman. Dangerous Liaisons? When Cultivated Plants Mate with Their Wild Relatives. Baltimore, Md.: Johns Hopkins University Press, 2003.
———, et al. “Gene flow and introgression from domestic plants into their wild relatives.” Annual Review of Ecology and Systematics 30 (1999): 539–563.
———, and Kristina a. Schierenbeck. “Hybridization as a stimulus for the evolution of invasiveness in plants?” Proceedings of the National Academy of Sciences USA 97 (2000): 7,043–7,050.
Foster, nelson, and linda S. cordell, eds. Chilies to Choco-late: Food the Americas Gave to the World. tucson: University of arizona Press, 1992.
Glover, Jerry D., cynthia M. cox, and John P. Reganold. “Future farming: a return to roots?” Scientific Ameri-can 297 (2007): 82–89.
———. “Harvested perennial grasslands: ecological mod-els for farming’s perennial future.” Agriculture, Eco-systems and Environment 137 (2010): 1–2.
———, and John P. Reganold. “Perennial grains: Food security for the future.” Issues in Science and Technol-ogy, Winter 2010, 41–47.
Harlan, Jack R. The Living Fields: Our Agricultural Heri-tage. new york: cambridge University Press, 1995.
A sign in Kansas boasts of the efficiency of modern agriculture. However, modern agriculture, being utterly dependent on fossil fuels, is very inefficient in its use of energy. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 14 11/17/11 1:12 PM

amphibians 15
Heiser, charles B. Seed to Civilization: The Story of Food. cambridge, Mass.: Harvard University Press, 1990.
Hobhouse, Henry. Seeds of Change: Five Plants that Transformed Mankind. new york: Harper and Row, 1986.
international Society of Organic agriculture Research. “international Society of Organic agriculture Research.” available online. URl: http://www.isofar.org. accessed august 8, 2011.
Jordan, n., et al. “Sustainable development of the agricul-tural bio-economy.” Science 316 (2007): 1,570–1,571.
land institute. “the land institute.” available online. URl: http://www.landinstitute.org. accessed May 1, 2010.
leopold center for Sustainable agriculture, iowa State University. “Welcome to the leopold center.” avail-able online. URl: http://www.leopold.iastate.edu. accessed august 8, 2011.
local Harvest. “local Harvest.” available online. URl: http://www.localharvest.org. accessed august 8, 2011.
lu, yanhi, et al. “Mirid bug outbreaks in multiple crops correlated with wide-scale adoption of Bt cotton in china.” Science 329 (2010): 1,151–1,154.
Marvier, M., and R. Van acker. “can crop transgenes be kept on a leash?” Frontiers in Ecology and the Envi-ronment 3 (2005): 93–100.
natural Resource conservation Service, United States Department of agriculture. “Plants database.” avail-able online. URl: http://www.plants.usda.gov. accessed May 1, 2010.
Pilson, D., and H. Prendeville. “ecological effects of trans-genic crops and the escape of transgenes into wild populations.” Annual Review of Ecology, Evolution, and Systematics 35 (2004): 149–174.
Pollan, Michael. The Omnivore’s Dilemma: A Natural His-tory of Four Meals. new york: Penguin, 2006.
———. “Farmer in chief.” New York Times Magazine, October 12, 2008.
Rice, Stanley a. “How agriculture changed the world.” chapter 10 in Green Planet: How Plants Keep the Earth Alive. new Brunswick, n.J.: Rutgers University Press, 2009.
Rindos, David. The Origins of Agriculture: An Evolution-ary Perspective. San Diego: academic Press, 1984.
Salaman, Redcliffe. The History and Social Influence of the Potato. new york: cambridge University Press, 1949 (reissued 1985).
Simpson, Beryl Brintnall, and Molly conner Ogorzaly. Economic Botany: Plants in Our World. new york: McGraw-Hill, 1995.
Snow, allison a., et al. “a Bt transgene reduces herbivory and enhances fecundity in wild sunflowers.” Ecologi-cal Applications 13 (2003): 279–286.
Stotzky, G. “Persistence and biological activity in soil of the insecticidal proteins from Bacillus thuringiensis, especially from transgenic plants.” Plant and Soil 266 (2004): 77–89.
amphibians amphibians are vertebrates that live in aquatic environments during their juvenile phase and usually on land during their adult phase. Amphi is Greek for “both” and bio is Greek for “life.” Frogs undergo the most dramatic metamor-phosis among the vertebrates, from herbivorous tadpoles with tails and gills to carnivorous adults with lungs and legs. they also undergo changes in physiological adaptation. in adult frogs, as in most vertebrates, hemoglobin releases oxygen into tissues under acid conditions that result from muscu-lar exertion, but many tadpoles live in acidic water. they have a type of hemoglobin that absorbs, rather than releases, oxygen under acidic conditions, thus allowing them to absorb oxygen from the acidic waters in which they live.
EvoluTIoN of amphIBIaNSamphibians are one of the lineages that evolved from the fleshy-finned (sarcopterygian) fishes. Dur-ing the middle Paleozoic era, there were many ani-mals that were intermediate between sarcopterygian fishes and modern amphibians. Most of these “miss-ing link” fish-amphibians were members of lineages that became extinct. all modern amphibians evolved from one amphibian lineage, for which fossils may or may not have been found. the fossils demonstrate, however, that there were many examples of the tran-sition from fish to amphibian during the Paleozoic.
numerous transitional forms have been discov-ered from the Devonian and carboniferous periods, including the following.
Eusthenopteron foordi was a sarcopterygian fish that lived in the late Devonian period. it had all the same fins that modern sarcopterygian fishes still pos-sess, rather than hands or feet. at the bases of the fins, it had bones analogous to the arm and leg bones of terrestrial vertebrates. Eusthenopteron had no bones that corresponded to the digits of modern tetrapods. even though it did not have lungs and did not breathe air, it had nostrils, which perhaps allowed water to flow over olfactory cells and may have enhanced the sense of taste. this organism looked very much like an ordinary sarcopterygian fish, and it probably spent nearly all of its time in shallow water.
Panderichthys rhombolepis was another sarcop-terygian fish that lived during the late Devonian period. it lacked some of the fish fins and had thicker ribs than fishes possess. the thicker ribs were impor-tant in supporting the weight of the body when on land and away from the buoyancy of water. How-ever, lacking legs, this species must have spent nearly all of its time in water.
Tiktaalik roseae created a sensation in the world of science when the evolutionary biologist neil Shu-bin announced its discovery in 2004. He called it a
xviii+598_Biodiversity_F.indd 15 11/17/11 1:12 PM

fi rst entry16 amphibians 16
“fi shopod” because it was almost directly interme-diate between fi shes (such as Eusthenopteron and Panderichthys) and tetrapods (such as Ichthyostegaand Acanthostega, see below). its name derives from a word in the language of the nunavat, the native peoples who live on ellesmere island, where the fos-sil was discovered. Tiktaalik’s fi shlike characteristics included (see fi gure above).
● it had gills. ● it had scales. ● it had no fi ngers.
Tiktaalik’s intermediate characteristics included:
● its shoulder partially resembled that of later amphibians, permitting it a limited ability to move its legs in a walking fashion.
● it had partly amphibian wrist and elbow bones, which allowed it a limited ability to support its weight with its fi ns.
Tiktaalik’s amphibian characteristics included:
● it had amphibian ribs, necessary for sup-porting its body weight outside of water.
● it could move its neck, which fi sh cannot do. ● it had lungs that were like those of amphib-
ians, not lungfi shes.
Ichthyostega stensioei and Acanthostega gun-nari had more adaptations for life on land than did Tiktaalik. they had hands and feet with fi n-gers. their ribs were thicker. their anatomy did not
allow rapid movement on land, and they probably spent most of their time in shallow water, as do many amphibians and reptiles today. in fi shes, the hyomandibu-lar bone helps to support the gills; this bone corresponds to the sta-pes, the ear bone of tetrapods (the innermost ear bone of mammals). the stapes of Acanthostega has been found, and it resembles a fi sh hyomandibular bone. it was not free to vibrate and therefore could not have functioned in hearing on land. later amphibians, in the carboniferous period, had stapes that functioned in hearing.
By the early carboniferous period, there were many differ-ent amphibian lineages. all have become extinct except for two: the branch that led to modern
amphibians, and the branch that led to modern reptiles.
there has been debate regarding the reasons for the evolution of legs in the transitional forms between fi shes and amphibians. Suggestions include the following.
● One early proposal was that fi shes had to walk on land to get from one pond to another if their home pond began to dry up. there are fi shes today that crawl on land and even climb trees entirely without legs.
● Modern salamanders have legs, yet many of them live underwater. they use their legs for walking underwater on rock surfaces against the current. it is possible that the fi rst legged amphibians evolved in rushing water.
● Recent analyses suggest that legs were more useful for lifting the earliest amphibians above the water to watch for predators than for effi cient walking.
● the evolutionary biologist Robert a. Mar-tin suggests that legs assisted in clasping during sexual reproduction, a function they still possess in many modern amphibians.
it is probable that legs proved useful for several different functions over a long period of time. What-ever combination of advantages may have selected for the evolution of legs, it had to be something that worked in a primitive condition. the earliest amphibians with legs could scarcely lift themselves at all.
Tiktaalik roseae, discovered in 2006 by the biologist Neil Shubin and associates, was an early amphibian called a “fi shapod” that had nearly an equal mix of fi sh and amphibian characteristics.
xviii+598_Biodiversity_F.indd 16 11/17/11 1:12 PM

amphibians 17
the evolution of limbs would not require the acquisition of many new genes. Hox genes control the pattern of body part development in most ani-mals. Some of these hox genes (numbers 9 to 13) control limb development in mice, from shoulder to feet. analogs of these genes are found in all verte-brates. activation of specific hox genes can produce limbs; evolution would then work out the structural details of these limbs, rather than produce them from scratch.
While the arm and leg bones have analogs in some fishes, digits were an amphibian invention. the earliest tetrapods had more than five digits (Ichthyo-stega had seven and Acanthostega had eight). all surviving tetrapods have a five-digit fundamental pattern, even though some, such as horses, have lost the ability to produce all five of them.
in addition to the “missing links” between fishes and amphibians, amphibians (as their name implies) are themselves intermediate between fishes and fully terrestrial animals.
dIvErSITy of amphIBIaNSthere are more than 6,000 species of amphibians (see table). amphibians include caecilians, sala-manders, and frogs. Caecilians are legless tropical amphibians that burrow through soil. Both larval and adult salamanders have four legs, all about the same size, and tails. adult frogs have large jumping hind legs and external eardrums but do not have tails.
amphibians, like other groups of vertebrates, vary greatly in size. the smallest salamander (in the genus Thorius) is only an inch (2.5 cm) long, while the largest are asiatic salamanders (genus Andrias), which are up to 5.5 feet (1.7 m) long and can weigh 88 pounds (40 kg). Frogs are generally small. the largest is the goliath frog (Conraua goliath), which can reach 13 inches (33 cm) in length and weigh eight pounds (3 kg).
amphibians have some amazing adaptations to their environments. Frogs can leap for great dis-tances. Some frogs can jump almost 30 feet (9 m).
Frogs keep their tongues folded inside their mouths, with the tip pointing backwards. their tongues can extend almost as long as their bodies to catch flying insects. their sticky tongues capture the insects, which they swallow whole; the small teeth, present only in the upper jaw, serve little function. caeci-lians have a unique sensory structure, called a ten-tacle, which develops from the eyes and nose.
amphibians are important components of aquatic ecosystems. Both as juveniles and adults, they eat a lot of invertebrates. their main pro-tection comes from either camouflage or from its opposite, a bright display of color that indicates poi-sonous skin. Many colorful salamanders produce large amounts of tetrodotoxin in their skins and are among the most toxic animals known. “Poison dart” frogs of many species also have colorful skin in which poison glands produce venom (see figure on page 18). examples include batrachotoxin, epi-batidine, histrionicotoxin, and pumiliotoxin. Most frogs are nocturnal, but poison dart frogs are most visible, and thus safest, during the day. in some frogs, such as the colorado River toad, Bufo alvar-ius, the poison is not deadly and is used as a recre-ational hallucinogen by some people. the poison compounds in amphibian skins have already proven to be of medical importance. Researchers at abbott labs used a poison in the skin of a tree frog from the ecuadorean amazon rain forest, Epipedobates tricolor, as a model for the development of a new painkiller. these rain forests are among the most threatened habitats on earth, and amphibians are among the most threatened species that live in them. captive poison dart frogs do not generate poison because they do not synthesize it themselves; they concentrate it from their prey animals, such as cen-tipedes and ants.
Juvenile amphibians (such as the tadpoles of frogs) obtain oxygen from water and release car-bon dioxide into water through their gills. in con-trast, most adult amphibians breathe with lungs and are therefore at least partly adapted to life on dry land. Salamander larvae have external gills. adult frogs and most adult salamanders lose their gills and develop lungs, but they still rely heavily on oxygen uptake through their soft skins. Most amphibians, even as adults, must live in moist environments. Most have skins that cannot resist desiccation. the skins of some toads resist water loss. the adults of some australian and north american frogs can survive for years in the desert, dormant and underground.
amphibians neither generate very much body heat nor regulate their body temperatures. there-fore, they are both ectothermic (from the Greek for “heat from outside”) and heterothermic (signifying variable body heat).
estimated number of amphibian speCies
Group Number of Species
Anurans (frogs) 5,892
Caecilians 186
Salamanders 586
Total 6,664
*Note: From Amphibiaweb.org
xviii+598_Biodiversity_F.indd 17 11/17/11 1:12 PM

first entry18 amphibians 18
Juvenile amphibians release ammonia as a waste product. ammonia is toxic, but the water washes the ammonia away. in contrast, many land-dwelling adult amphibians produce urea, which is less toxic than ammonia. Wastes from kidneys and intestines are voided together with gametes through a cloaca.
Most adult amphibians return to the water to lay and fertilize eggs. amphibian eggs dry out quickly unless kept in moist conditions. though most amphibians keep their eggs moist by depositing them underwater, some species brood their eggs in a pouch or inside their bodies. even desert amphibians require an aquatic environment for reproduction. they emerge from underground dormancy only dur-ing a rainy season, mate, and lay eggs. their juve-niles grow rapidly and reach adulthood before the pools of water dry up. Some species of amphibians have external fertilization, while in others the males clasp the females while eggs and sperm are released. in some, the males leave sperm on a structure that the females take into their cloacae. in others, the males and females press their cloacae together to achieve internal fertilization.
although most amphibians have an aquatic juvenile and terrestrial adult phase, there is some variation in this pattern. all frogs go through a larval stage, but some of them, such as the Puerto Rican tree frog Eleutherodactylus coqui, do so before the eggs hatch, with the result that adult frogs emerge from the mother. Plethodont salaman-ders also do this. Some salamander species, such as the mudpuppy Necturus maculosus and the axolotl Ambystoma mexicanum, are famous for becoming
sexually mature while they are still larvae. injection of thyroid hormones can cause an axolotl to develop into an adult form that has never been encountered in the wild. this experiment allowed the evolution-ary biologist Julian Huxley to describe how neoteny (retention of juvenile characteristics during sexual maturity) can lead to the evolution of new species.
ExTINCTIoNS of amphIBIaNSall around the world, populations of many amphib-ian species have declined precipitously. the fol-lowing are factors that are directly threatening the survival of amphibians, especially adult frogs.
● the direct cause of most population declines has been funguslike protists that live in the skins and produce toxins. the chytrid pro-tist Batrachochytrium dendrobatidis grows in the skins of tree frogs. By upsetting cel-lular salt balance, toxins from the pathogen cause cardiac arrest in the frogs. Saproleg-nia ferax is another chytrid that kills frogs. Other parasites, such as the trematode worm Ribeiroia ondatrae, cause deformities in frogs. chytrid parasites may have been introduced into susceptible populations by toads that escaped from experimental laboratories, because the common labora-tory toad Xenopus laevis is a symptomless carrier.
● in some cases, amphibians are threatened by invasive fishes that eat their larvae.
● Global warming can cause changes in cli-mate that lead to amphibian extinction. the golden toad (Bufo periglenes) may have become extinct because climate change has caused the cloud forests in which it lived to not be wet enough.
● Frogs also have very permeable skin, which makes them vulnerable to pollutants such as glyphosate and atrazine. these herbicides are very toxic to amphibians at far below the recommended level of agricultural use. atrazine causes frogs to develop hermaph-roditism at a concentration a thousand times less than is commonly found in farm runoff water and 400 times less than is found in some rainwater in agricultural areas. their thin skin also makes frogs vulnerable to environmental threats such as acid rain and ultraviolet radiation.
What has become apparent is that these threat factors interact. Pollutants, ultraviolet light, and acid rain may make the amphibians more suscep-tible to protist and other parasite infections. it is
Many tropical frogs, such as this blue poison dart frog (Dendrobates azureus), have brightly colored skin that functions as a warning that it contains toxins. (Adrian Pingstone)
xviii+598_Biodiversity_F.indd 18 11/17/11 1:12 PM

animals 19
the interaction of pollution and parasites that is threatening so many species of amphibians. Global warming is also contributing to this interaction. For example, the spread of Batrachochytrium has been enhanced by warmer temperatures. amphibians today are facing a rate of extinction greater than they endured during the two largest mass extinc-tions in the past: the Permian extinction and the cretaceous extinction.
See also biodiversity, resources for medi-cine; camouflage and mimicry; carbonifer-ous period, biodiversity in; Devonian period, biodiversity in; fishes; mass extinctions; para-sitism; pollution; reptiles.
Further readingamphibiaweb. “amphibiaweb.” available online. URl:
http://www.amphibiaweb.org. accessed august 8, 2011.Bank, M. S., et al. “Mercury bioaccumulation in northern
two-lined salamanders from streams in the northeast-ern United States.” Ecotoxicology 14 (2005): 181–191.
Beebee, t. J. c. “amphibian breeding and climate.” Nature 374 (1995): 219–220.
Berger, l., et al. “chytridiomycosis causes amphibian mor-tality associated with population declines in the rain forests of australia and central america.” Proceedings of the National Academy of Sciences USA 95 (1998): 9,031–9,036.
Blauenstein, a. R., et al. “UV repair and resistance to solar UV-B in amphibian eggs: a link to population declines.” Proceedings of the National Academy of Sci-ences USA 91 (1994): 1,791–1,795.
———, et al. “Ultraviolet radiation, toxic chemicals, and amphibian population declines.” Diversity and Distri-butions 9 (2003): 123–140.
callery, elizabeth M. “Frogs without polliwogs: evolution of anuran direct development.” BioEssays 23 (2001): 233–241.
chivian, eric, and aaron Bernstein. “threatened groups of organisms valuable to medicine: amphibians.” in Sustaining Life: How Human Health Depends on Bio-diversity, edited by eric chivian and aaron Bernstein, 204–223. new york: Oxford University Press, 2008.
clack, Jennifer. Gaining Ground: The Origin and Evolu-tion of Tetrapods. Bloomington, ind.: indiana Univer-sity Press, 2002.
———. “From fins to limbs.” Natural History (July 2006): 36–41.
coates, Michael i., et al. “ever since Owen: changing per-spectives on the early evolution of tetrapods.” Annual Reviews of Ecology, Evolution, and Systematics 39 (2008): 571–592.
Daeschler, edward B., neil H. Shubin, and Farish a. Jen-kins, Jr. “a Devonian tetrapod-like fish and the evolu-tion of the tetrapod body plan.” Nature 440 (2006): 757–763.
Davic, Robert D., and Hartwell H. Welsh, Jr. “On the eco-logical roles of salamanders.” Annual Review of Ecol-ogy, Evolution, and Systematics 35 (2004): 405–434.
Dupuis, l. a., et al. “Relation of terrestrial-breeding amphibian abundance to tree-stand age.” Conserva-tion Biology 9 (1995): 645–653.
Feder, M. e., and W. W. Burggren, eds. Environmental Physiology of the Amphibians. chicago: University of chicago Press, 1992.
Hall, Brian K., ed. Fins into Limbs: Evolution, Devel-opment, and Transformation. chicago: University of chicago Press, 2007.
Hayes, t. B., et al. “Hermaphroditic, demasculinized frogs after exposure to the herbicide atrazine at low eco-logically relevant doses.” Proceedings of the National Academy of Sciences USA 99 (2002): 5,476–5,480.
laMarca, e., et al. “catastrophic population declines and extinctions in neotropical harlequin frogs (Bufonidae: Atelopus).” Biotropica 37 (2005): 190–201.
lips, K. R., et al. “emerging infectious disease and the loss of biodiversity in a neotropical amphibian com-munity.” Proceedings of the National Academy of Sci-ences USA 103 (2006): 3,165–3,170.
Myers, c. W., et al. “a dangerously toxic new frog (Phyl-lobates) used by emberá indians of western colombia, with discussion of blowgun fabrication and dart poi-soning.” Bulletin of the American Museum of Natural History 161 (1978): 307–366.
Petranka, J. W., et al. “effects of timber harvesting on southern appalachian salamanders.” Conservation Biology 7 (1993): 363–377.
Pierce, B. “the effects of acid precipitation on amphib-ians.” Ecotoxicology 2 (1993): 65–77.
Pounds, J. a., et al. “Widespread amphibian extinctions from epidemic disease driven by global warming.” Nature 439 (2006): 161–167.
Stuart, S. n., et al. “Status and trends of amphibian declines and extinctions worldwide.” Science 306 (2004): 1,783–1,786.
Voyles, Jamie, et al. “Pathogenesis of chytridiomycosis, a cause of catastrophic amphibian declines.” Science 326 (2009): 582–585.
Vredenburg, V. t. “Reversing introduced species effects: experimental removal of introduced fish leads to rapid recovery of a declining frog.” Proceedings of the National Academy of Sciences USA 101 (2004): 7,646–7,650.
animals animals are multicellular eukaryotes that consume other organisms or molecules pro-duced by them and that are usually motile at some stage of their lives. animals (kingdom animalia) are one of four kingdoms within the domain eucarya (eukaryotes). animals differ from the other eukary-otic kingdoms in ways such as the following:
xviii+598_Biodiversity_F.indd 19 11/17/11 1:12 PM

fi rst entry20 animals 20
● animals, unlike animal-like protists, are not only multicellular but organized into tissues and organs.
● animals, unlike animal-like protists, develop from a single fertilized egg.
● animals, unlike plants, never use photosyn-thesis or have cellulose cell walls.
● animals, unlike fungi, never have dikary-otic cells or chitin cell walls.
animals also have a sexual life cycle that con-trasts with almost all plants and fungi. in plants and fungi, meiosis produces spores, which grow into multicellular haploid structures (the mycelia of fungi and the gametophytes of plants). in animals, meio-sis directly produces gametes, which are eggs and sperm. an animal develops from a single fertilized egg. in some cases, development from a fertilized egg is almost the only evidence that an organism is an animal rather than a protist.
the protists that are most closely related to animals are the choanofl agellates, which form small clusters of cells that resemble the choanocytes of sponges. the earliest animals may have been choano-fl agellatelike cells that formed clusters that resem-bled the blastula stage of embryonic development.
From the human viewpoint, animals have been classifi ed as invertebrates and vertebrates based on the absence or presence of bony vertebrae sur-rounding the main nerve cord along the dorsal (back) surface of the body. all the phyla of animals except the chordates are inver-tebrates; even the chordate phy-lum contains some invertebrates. “invertebrate,” therefore, is not a taxonomic category.
the animal kingdom is tradi-tionally divided into phyla. there are 35 phyla currently recognized in the animal kingdom. it appears that all animal phyla evolved more than 540 million years ago, at which time all organisms still lived in the sea; no new animal phyla have arisen since.
Body formS aNd adapTaTIoNSScientists distinguish among ani-mals in numerous ways, including the following:
Habitat. Many animals live only in the oceans or in freshwa-ter. in order to survive and repro-duce on land, animals require protection from water loss and support structures such as skel-
etons. Buoyancy supports the weight of aquatic ani-mals, but terrestrial animals need skeletons to hold them up. arthropods have external skeletons, and vertebrates have internal skeletons. Most animal phyla are fully aquatic, have little or no protection against drying out, and are unable to tolerate terres-trial conditions unless they live in moist soil or inside the bodies of other animals.
Body structure. animals can be distinguished by their types of symmetry. animals in four of the phyla are either too simple to have any recognizable symmetry or are asymmetrical. animals in some phyla have radial symmetry, while in others they have bilateral symmetry. the organs of radially symmetric animals (such as jellyfi sh) are arranged outward from a central point; there is no front or rear, left or right, though there may be a top and bot-tom (see fi gure). the organs of bilaterally symmetric animals, such as humans, are paired left and right along an axis from front to rear. they have dorsal(back) and ventral (belly) surfaces. Many bilaterally symmetrical animals, such as earthworms, have seg-mentation, repeating units from front to rear, some-times with the same organs in each segment. Some animals have a tissue-lined body cavity (coelom),which contains the internal organs, and some do not (acoelomate).
Circulation and respiration. Many animals are so small and simple that they do not have circula-tory or respiratory systems. the ratio of external surface to internal volume is so great that they can
Animals with radial symmetry (left) have a front and back side but radiate equally in all directions. Animals with bilateral (“two-sided”) symmetry (right) have a front and back, top and bottom, and left and right sides.
xviii+598_Biodiversity_F.indd 20 11/17/11 1:12 PM
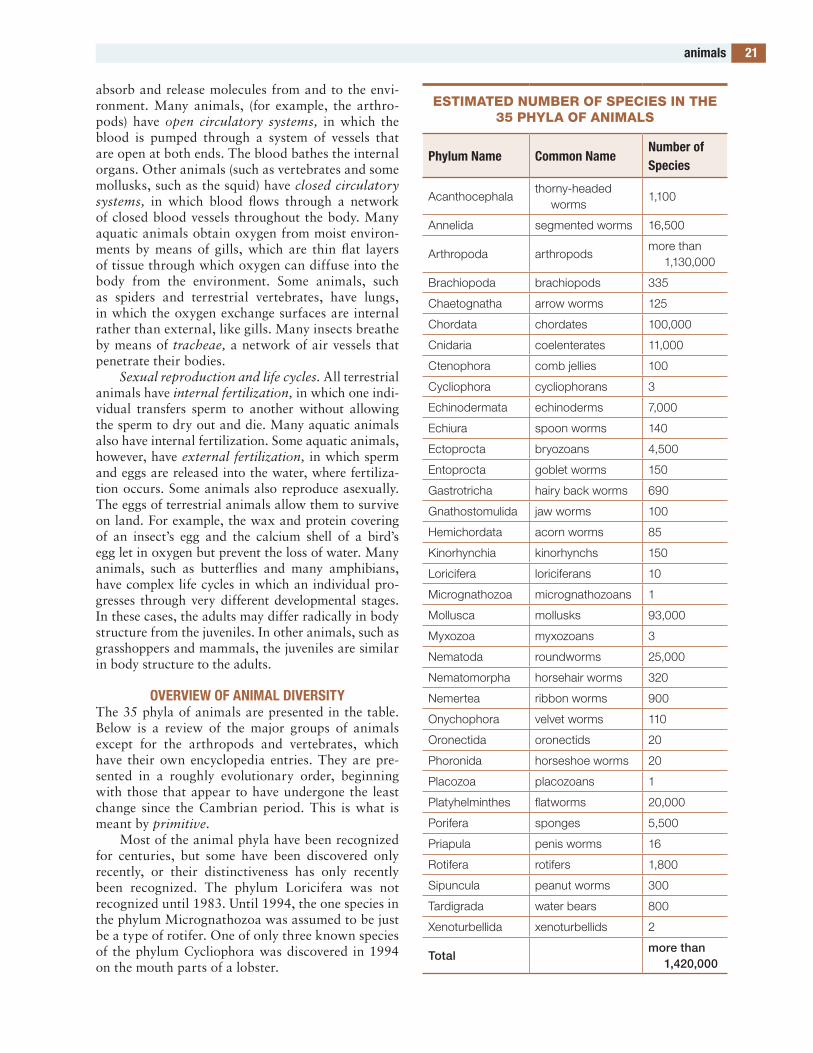
animals 21
absorb and release molecules from and to the envi-ronment. Many animals, (for example, the arthro-pods) have open circulatory systems, in which the blood is pumped through a system of vessels that are open at both ends. the blood bathes the internal organs. Other animals (such as vertebrates and some mollusks, such as the squid) have closed circulatory systems, in which blood flows through a network of closed blood vessels throughout the body. Many aquatic animals obtain oxygen from moist environ-ments by means of gills, which are thin flat layers of tissue through which oxygen can diffuse into the body from the environment. Some animals, such as spiders and terrestrial vertebrates, have lungs, in which the oxygen exchange surfaces are internal rather than external, like gills. Many insects breathe by means of tracheae, a network of air vessels that penetrate their bodies.
Sexual reproduction and life cycles. all terrestrial animals have internal fertilization, in which one indi-vidual transfers sperm to another without allowing the sperm to dry out and die. Many aquatic animals also have internal fertilization. Some aquatic animals, however, have external fertilization, in which sperm and eggs are released into the water, where fertiliza-tion occurs. Some animals also reproduce asexually. the eggs of terrestrial animals allow them to survive on land. For example, the wax and protein covering of an insect’s egg and the calcium shell of a bird’s egg let in oxygen but prevent the loss of water. Many animals, such as butterflies and many amphibians, have complex life cycles in which an individual pro-gresses through very different developmental stages. in these cases, the adults may differ radically in body structure from the juveniles. in other animals, such as grasshoppers and mammals, the juveniles are similar in body structure to the adults.
ovErvIEw of aNImal dIvErSITythe 35 phyla of animals are presented in the table. Below is a review of the major groups of animals except for the arthropods and vertebrates, which have their own encyclopedia entries. they are pre-sented in a roughly evolutionary order, beginning with those that appear to have undergone the least change since the cambrian period. this is what is meant by primitive.
Most of the animal phyla have been recognized for centuries, but some have been discovered only recently, or their distinctiveness has only recently been recognized. the phylum loricifera was not recognized until 1983. Until 1994, the one species in the phylum Micrognathozoa was assumed to be just be a type of rotifer. One of only three known species of the phylum cycliophora was discovered in 1994 on the mouth parts of a lobster.
estimated number of speCies in the 35 phyla of animals
phylum Name Common NameNumber of Species
Acanthocephalathorny-headed
worms1,100
Annelida segmented worms 16,500
Arthropoda arthropodsmore than
1,130,000
Brachiopoda brachiopods 335
Chaetognatha arrow worms 125
Chordata chordates 100,000
Cnidaria coelenterates 11,000
Ctenophora comb jellies 100
Cycliophora cycliophorans 3
Echinodermata echinoderms 7,000
Echiura spoon worms 140
Ectoprocta bryozoans 4,500
Entoprocta goblet worms 150
Gastrotricha hairy back worms 690
Gnathostomulida jaw worms 100
Hemichordata acorn worms 85
Kinorhynchia kinorhynchs 150
Loricifera loriciferans 10
Micrognathozoa micrognathozoans 1
Mollusca mollusks 93,000
Myxozoa myxozoans 3
Nematoda roundworms 25,000
Nematomorpha horsehair worms 320
Nemertea ribbon worms 900
Onychophora velvet worms 110
Oronectida oronectids 20
Phoronida horseshoe worms 20
Placozoa placozoans 1
Platyhelminthes flatworms 20,000
Porifera sponges 5,500
Priapula penis worms 16
Rotifera rotifers 1,800
Sipuncula peanut worms 300
Tardigrada water bears 800
Xenoturbellida xenoturbellids 2
Totalmore than
1,420,000
xviii+598_Biodiversity_F.indd 21 11/17/11 1:12 PM
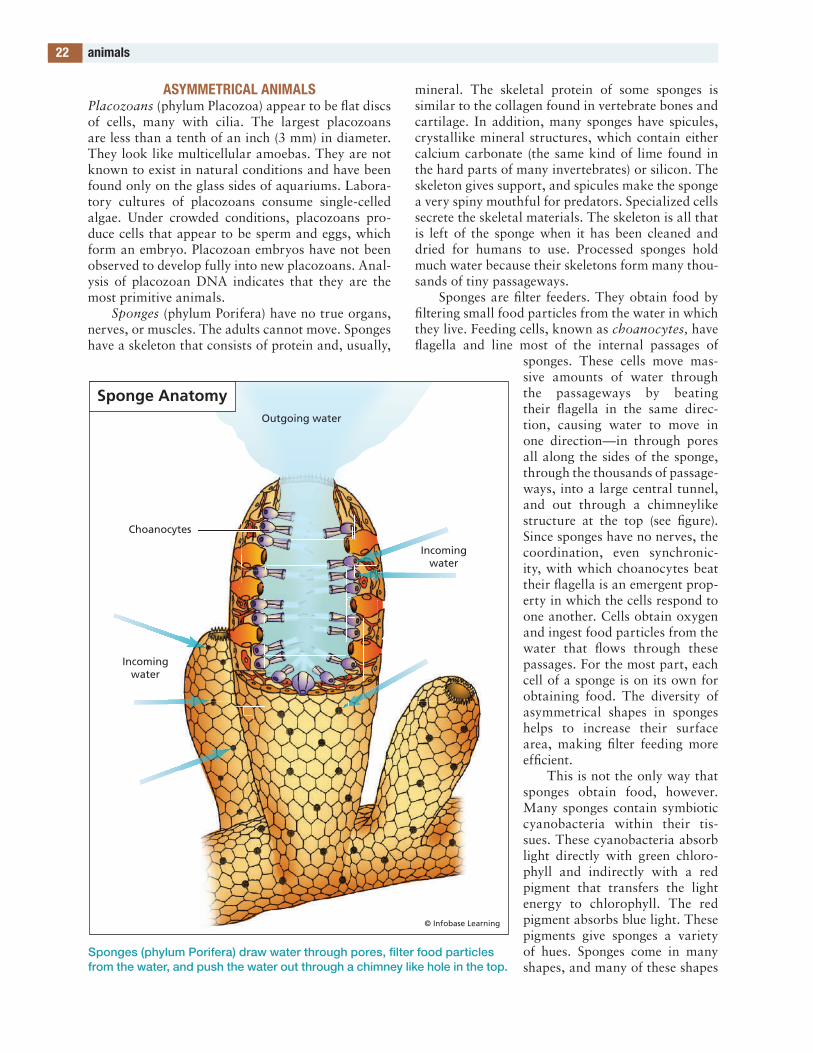
fi rst entry22 animals 22
aSymmETrICal aNImalSPlacozoans (phylum Placozoa) appear to be fl at discs of cells, many with cilia. the largest placozoans are less than a tenth of an inch (3 mm) in diameter. they look like multicellular amoebas. they are not known to exist in natural conditions and have been found only on the glass sides of aquariums. labora-tory cultures of placozoans consume single-celled algae. Under crowded conditions, placozoans pro-duce cells that appear to be sperm and eggs, which form an embryo. Placozoan embryos have not been observed to develop fully into new placozoans. anal-ysis of placozoan Dna indicates that they are the most primitive animals.
Sponges (phylum Porifera) have no true organs, nerves, or muscles. the adults cannot move. Sponges have a skeleton that consists of protein and, usually,
mineral. the skeletal protein of some sponges is similar to the collagen found in vertebrate bones and cartilage. in addition, many sponges have spicules, crystallike mineral structures, which contain either calcium carbonate (the same kind of lime found in the hard parts of many invertebrates) or silicon. the skeleton gives support, and spicules make the sponge a very spiny mouthful for predators. Specialized cells secrete the skeletal materials. the skeleton is all that is left of the sponge when it has been cleaned and dried for humans to use. Processed sponges hold much water because their skeletons form many thou-sands of tiny passageways.
Sponges are fi lter feeders. they obtain food by fi ltering small food particles from the water in which they live. Feeding cells, known as choanocytes, have fl agella and line most of the internal passages of
sponges. these cells move mas-sive amounts of water through the passageways by beating their fl agella in the same direc-tion, causing water to move in one direction—in through pores all along the sides of the sponge, through the thousands of passage-ways, into a large central tunnel, and out through a chimneylike structure at the top (see fi gure). Since sponges have no nerves, the coordination, even synchronic-ity, with which choanocytes beat their fl agella is an emergent prop-erty in which the cells respond to one another. cells obtain oxygen and ingest food particles from the water that fl ows through these passages. For the most part, each cell of a sponge is on its own for obtaining food. the diversity of asymmetrical shapes in sponges helps to increase their surface area, making fi lter feeding more effi cient.
this is not the only way that sponges obtain food, however. Many sponges contain symbiotic cyanobacteria within their tis-sues. these cyanobacteria absorb light directly with green chloro-phyll and indirectly with a red pigment that transfers the light energy to chlorophyll. the red pigment absorbs blue light. these pigments give sponges a variety of hues. Sponges come in many shapes, and many of these shapes
Sponges (phylum Porifera) draw water through pores, fi lter food particles from the water, and push the water out through a chimney like hole in the top.
xviii+598_Biodiversity_F.indd 22 11/17/11 1:12 PM

animals 23
increase the surface area for light absorption. Since blue light penetrates more deeply into the water (which is why underwater scenes appear blue), the red pigment of the cyanobacterial symbionts allows many sponges to survive at depths where only a modest amount of light is available.
Sponges can produce sperm, which sometimes emerge from the chimney in large numbers. the sperm may encounter and be drawn inside other sponges of the same species, where they fertilize eggs. the fertilized egg develops into a larva, which is a cluster of cells that swims by its flagella.
the other very simple phyla of animals, consist-ing of very few cells, are internal parasites: myxozo-ans (phylum Myxozoa) and orthonectidans (phylum Orthonectida).
radIally SymmETrICal aNImalSCnidarians (phylum cnidaria; also called coelenter-ates) include jellyfish, sea anemones, hydras, and corals (see figure). cnidarians can have a major influence on their aquatic environments. For exam-ple, corals secrete lime skeletons, which build up into massive reefs and whole islands. cnidarians can thus be important to many other species, as whole coral reef ecological communities are built upon their activities. explosions of jellyfish populations sometimes occur when predatory fishes become rare.
cnidarians have a very delicate structure that consists of two tissue layers. the inner tissue layer lines a central digestive cavity into which they secrete digestive enzymes. cnidarians have a single opening on one surface for food to enter and wastes to exit. Between the two cell layers is a gelatinous material, which in jellyfish can constitute most of the weight. the gelatinous mass is slightly lighter than water, which gives the animals some buoyancy. a network of nerves stimulates the contraction of cells; cnidar-ians have no specialized muscular tissue. Both the inner and outer tissue layers are in direct contact with the water. each cell exchanges oxygen and car-bon dioxide directly with the water. cnidarians have no circulatory system.
cnidarians produce stinging tentacles around the opening of the central cavity. the stinging cells (nematocysts) on these tentacles inject tiny barbs into the prey. a toxin paralyzes the prey, and the tentacles draw the prey into the central cavity, where the prey is digested. Most cnidarians are harmless to humans, though the stings of some jellyfish can cause pain to and even kill swimmers.
algae cells live in the skin tissue of many cni-darians. this is mutually beneficial both to the photosynthetic cells, which use the animal’s car-bon dioxide for photosynthesis and the animal’s wastes as fertilizer, and the cnidarian, for which
the photosynthetic cells are a built-in food factory. When these photosynthetic guests die, the cnidarian host may die. coral bleaching occurs when corals lose their photosynthetic symbionts and then starve. extensive reefs of corals have been bleached, per-haps due to increases in water temperature caused by global warming or increases in salinity caused by reduced water flow from rivers. the jellyfish Masti-gias papua, found in a lagoon on the Pacific island of Palau, swim from one end of the lagoon to the other and back each day, keeping themselves in the sun and thus promoting the photosynthesis of their symbiotic algae.
Many cnidarians have complex life cycles (see figure on page 24). the cylindrical polyp form is sessile, which means it remains attached to a surface such as a rock. With its tentacles reaching up, it grabs food that happens to come by. new polyps can branch off of, then break free from, old polyps (an example of asexual reproduction). in many cases, however, polyps produce another body form, the medusa, which is umbrella or bell shaped, such as the jellyfish. Medusae break free and swim, trailing their tentacles behind them. the medusae produce eggs and/or sperm for external fertilization. Some species have no medusa stage, some have no polyp stage, and some have both stages. in most species, however, either the polyp or the medusa phase of the life cycle predominates. Some cnidarians form colo-nies that consist of many individuals. For example, the Portuguese man-of-war (Physalia physalis) is a cluster of cnidarians.
ctenophores, or comb jellies (phylum cte-nophora), superficially resemble jellyfish. Unlike jellyfish, ctenophores propel themselves with cilia
This purple-striped jellyfish (Pelagia panopyra) is very large and has very potent stingers. (NOAA/Depart-ment of Commerce)
xviii+598_Biodiversity_F.indd 23 11/17/11 1:12 PM

fi rst entry24 animals 24
found in rows along the outside of the body. also, ctenophores have a mouth on the forward end and an anus on the back end, while jellyfi sh have a single opening for eating and evacuating. ctenophores do not produce their own stinging cells, though in some cases they can incorporate the stinging cells of jel-lyfi sh that they have eaten into their own tentacles.
BIlaTErally SymmETrICal aNImalS: proToSTomES
Protostomes are animals whose embryos undergo cell division (cleavage) in a spiral pattern. as a result, the structures into which the cells develop are deter-mined at a very early stage. the major groups of protostomes are the fl atworms, mollusks, annelids, animals with lophophores, and animals with ecdysis.
Flatworms (phylum Platyhelminthes) are free-living or parasitic worms. Many fl atworms, such as the planarians, live in aquatic habitats. Other fl at-worms, such as the oval-shaped fl ukes (trematodes) and the very long tapeworms (cestodes) live as para-sites in the moist environments inside larger animals.
Planarians have a head, eyespots, and a ner-vous system. Planarians move about to fi nd food and ingest it through a tube in the center of the ventral (lower) surfaces of their bodies. the planar-ian’s internal digestive tract, like that of cnidarians, has a single opening for both the intake of food and
the expulsion of wastes. Flukes and tapeworms, in contrast, absorb nutrients from and release wastes into the tissues or intestinal fl uid of their hosts. therefore, fl ukes and tapeworms, unlike planarians, possess neither digestive tracts nor organs to elimi-nate wastes. the microscopic convolutions of their skin increase the surface area through which they can absorb food. Flukes and tapeworms live under anaerobic conditions, in which cellular respiration is impossible. they are continually bathed in the host’s intestinal fl uid, which is full of digestive enzymes, but are protected by their outer layer of cells. in order to keep from being evacuated from the intes-tine, fl ukes and tapeworms attach themselves to the intestinal lining with suckers or hooks.
Flatworms do not have circulatory systems. Because the digestive tract is highly branched, digested food diffuses from the gut to every part of the body of the planarian. Because planarians are very fl at, oxygen has a very short distance to diffuse from the water into all the internal tissues.
in most fl atworms, each individual has both male and female reproductive organs. Many pla-narians reproduce by simply dividing in two, while others copulate; some switch seasonally between the two modes of reproduction. in most fl ukes and tape-worms, male and female reproductive organs form a large portion of each individual’s body. these organs
Coelenterates (Phylum Cnidaria) alternate between the free-swimming, sexual medusa (“jellyfi sh”) form and the nonsexual polyp form that attaches to hard surfaces.
xviii+598_Biodiversity_F.indd 24 11/17/11 1:12 PM
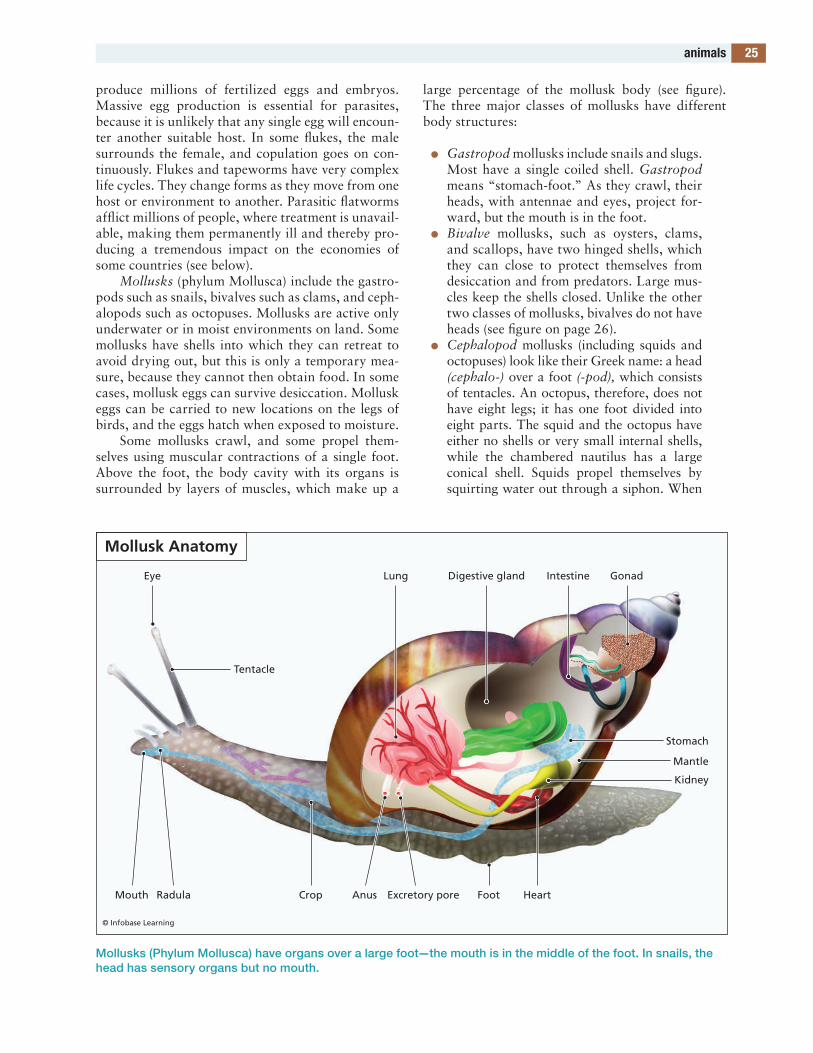
animals 25
produce millions of fertilized eggs and embryos. Massive egg production is essential for parasites, because it is unlikely that any single egg will encoun-ter another suitable host. in some fl ukes, the male surrounds the female, and copulation goes on con-tinuously. Flukes and tapeworms have very complex life cycles. they change forms as they move from one host or environment to another. Parasitic fl atworms affl ict millions of people, where treatment is unavail-able, making them permanently ill and thereby pro-ducing a tremendous impact on the economies of some countries (see below).
Mollusks (phylum Mollusca) include the gastro-pods such as snails, bivalves such as clams, and ceph-alopods such as octopuses. Mollusks are active only underwater or in moist environments on land. Some mollusks have shells into which they can retreat to avoid drying out, but this is only a temporary mea-sure, because they cannot then obtain food. in some cases, mollusk eggs can survive desiccation. Mollusk eggs can be carried to new locations on the legs of birds, and the eggs hatch when exposed to moisture.
Some mollusks crawl, and some propel them-selves using muscular contractions of a single foot. above the foot, the body cavity with its organs is surrounded by layers of muscles, which make up a
large percentage of the mollusk body (see fi gure). the three major classes of mollusks have different body structures:
● Gastropod mollusks include snails and slugs. Most have a single coiled shell. Gastropodmeans “stomach-foot.” as they crawl, their heads, with antennae and eyes, project for-ward, but the mouth is in the foot.
● Bivalve mollusks, such as oysters, clams, and scallops, have two hinged shells, which they can close to protect themselves from desiccation and from predators. large mus-cles keep the shells closed. Unlike the other two classes of mollusks, bivalves do not have heads (see fi gure on page 26).
● Cephalopod mollusks (including squids and octopuses) look like their Greek name: a head (cephalo-) over a foot (-pod), which consists of tentacles. an octopus, therefore, does not have eight legs; it has one foot divided into eight parts. the squid and the octopus have either no shells or very small internal shells, while the chambered nautilus has a large conical shell. Squids propel themselves by squirting water out through a siphon. When
Mollusks (Phylum Mollusca) have organs over a large foot—the mouth is in the middle of the foot. In snails, the head has sensory organs but no mouth.
xviii+598_Biodiversity_F.indd 25 11/17/11 1:12 PM

first entry26 animals 26
a predator pursues them, they may release ink to confuse the pursuer. Jet propulsion imparts great speed to some cephalopods.
Mollusks have diverse ways of getting food. Many gastropods crawl over plant, soil, or rock surfaces on a layer of slime that they secrete, scrap-ing up food (such as algae) with a radula, which is a rasplike structure covered with teeth, as they go along. Other gastropods, such as cone snails, have a poisonous harpoon near the mouth, which captures prey. in contrast, bivalves are filter-feeders. they use the cilia near their mouths to catch microscopic food particles from water that flows through them. Zebra mussels (Dreissena polymorpha) are euro-pean bivalves that were accidentally introduced into american lakes, where they are undergoing a popu-lation explosion. they filter plankton from the water so effectively that they can deplete the food base of whole aquatic food chains. Suckers on the underside of the tentacles allow cephalopods to grasp prey and cling to surfaces. the mouth, with a powerful beak, is located in the middle of the tentacles.
not being small or flat enough to exchange materials directly with their environments, mollusks have circulatory systems. in most mollusks, circula-tion is open and blood fills the whole body, although cephalopods have closed circulation. Mollusks have blood cells, but these cells do not carry oxygen. Oxygen is carried by hemocyanin, a copper-based molecule dissolved in the blood plasma. aquatic mollusks absorb oxygen from a stream of water that is pushed through the body by the movements of internal cilia. Some terrestrial snails have lungs.
Reproductive patterns are varied in mollusks. Gastropods and cephalopods have internal fertil-
ization. Gastropods typically have both male and female organs. a male cephalopod inserts a packet of sperm into a female with one of his tentacles. Bivalves, in contrast, have external fertilization. For this reason, as well as because they are filter feed-ers, bivalves cannot survive on land. Most mollusks, even those in which the adult phase is spent glued to a rock, have aquatic larvae that can swim.
cephalopods have evolved a level of intelli-gence higher than that of all other animals except vertebrates, and they evolved this intelligence inde-pendently of vertebrates. an octopus can learn by watching another. they can open jars with their tentacles. they can escape from predators by chang-ing color and imitating their surroundings. the octopus has a complex eye that, due to its separate evolutionary origin, has a different structure from that of a vertebrate eye: the nerves are behind the retina in the octopus but in front of the retina in the vertebrate.
Annelids (phylum annelida) are segmented worms. Annelid means “little ring.” each segment is a unit in which muscles, nerves, and some organs are repeated nearly throughout the worm’s length. the worm’s muscles exert pressure against water-tight segments, allowing the worm to move. Because water is incompressible, it remains firm when mus-cles press against it. the water therefore acts as a skeleton, called a hydrostatic skeleton (hydrostatic is Greek for “water remaining in place”). the main nerve cord in annelids is ventral (along the belly), unlike humans and other chordates, in which the nerve cord is dorsal (along the back). earthworms are the most familiar annelids.
Many marine annelids grab and eat small par-ticles of food. earthworms ingest leaf litter or soil containing decomposed organic matter, which they further break down. leeches are annelids, many of which suck blood from larger animals. the blood meal may allow a leech to increase greatly in size. it may live off of one meal for up to 200 days.
not being small or flat enough to exchange materials directly with their environments, annelids have circulatory systems. in some annelids, a hemo-globinlike substance that contains iron, which may or may not be in cells, carries oxygen and makes their blood red; in others, hemocyanin carries oxy-gen. the circulatory system is often open to allow the blood to bathe the internal organs.
Marine annelids have external reproduction. in some cases, whole populations release reproduc-tive segments synchronously. the palolo worms of Samoa release millions of reproductive segments in October at the Moon’s last quarter. the fertil-ized eggs develop into larvae, which develop into adults. earthworms, unlike most marine annelids,
This scallop has numerous simple eyes—the eyes allow the scallop to detect moving shadows that may represent danger. (Bill Capman)
xviii+598_Biodiversity_F.indd 26 11/17/11 1:12 PM

animals 27
reproduce by internal fertilization. the easily vis-ible clitellum produces a cocoon from which larval worms hatch. annelids can regenerate readily from fragments, which is fortunate for worms that live in heavily gardened soil.
Because they draw leaf litter down into the soil and fill the soil with burrows, earthworms aerate the soil and enhance its organic matter content. in both ways, earthworms exert an extremely ben-eficial effect on soil fertility, an economic benefit beyond calculation. However, the introduction of earthworms into habitats that previously did not have them can be severely disrupting.
Animals with lophophores. a lophophore is a ring of hollow, ciliated hairs that create a cur-rent and help to capture food items. animals with lophophores are therefore filter feeders. the lopho-phore surrounds the mouth. the digestive tract curls around in a horseshoe shape and usually emerges outside the lophophore.
● Brachiopods, or lampshells (phylum Bra-chiopoda), closely resemble bivalve mollusks, except for the lophophore. Brachiopods and mollusks lived in roughly equal abundance before the Permian extinction, from which the mollusks recovered more vigorously. Brachiopods of the genus Lingula live in sediments and are largely unchanged from the early Paleozoic era.
● Bryozoans, or moss animals (phylum Bryo-zoa, or ectoprocta), are small animals that form extensive colonies on rocks and other marine surfaces. they have an outer layer of tissue that secretes a hard coat, usually calcium carbonate, with an opening on the top through which the animal sticks out its lophophore. individuals within the (sometimes very large) colonies have dif-ferent structures and specialize in different activities.
● Phoronids, or horseshoe worms (phylum Phoronida), look like small roundworms but have an external lophophore around the mouth.
● Goblet worms (phylum entoprocta) are filter feeders with lophophores. Unlike the others, goblet worms have digestive tracts in which both ends are inside the ring of hairs.
Animals with ecdysis. Ecdysis is the molting of a hard outer covering. the major phyla of animals with ecdysis are the roundworms, the arthropods, and their relatives.
Roundworms (phylum nematoda) can, like flat-worms, be free living or parasitic. Most roundworms
are nematodes that live in water and soil. an acre of farmland can contain 100 billion nematodes; a single rotting apple can contain 90,000. Species diversity of nematodes has been explored only barely, and new species are found as new soil samples are examined. Most nematodes are decomposers or predators, but many infect plant roots and cause much crop loss. Some roundworms are parasites inside other animals.
Roundworms are more or less cylindrical rather than flat. a thick covering of collagen (the same protein found in many animal skeletons) protects their bodies. Roundworms move as their muscles exert force against the water contained within their bodies, giving them a hydrostatic skeleton like that of annelids.
Roundworms have a digestive tract with sep-arate openings for ingesting food and expelling wastes. Food can diffuse from the digestive tract as well as oxygen from the environment (unless it is anaerobic, as in the intestines of hosts) to all parts of the roundworm’s body. therefore, roundworms do not have circulatory or respiratory systems.
Roundworms usually live as separate male and female individuals and usually copulate. Parasitic roundworms may have complex life cycles. For example, one species of nematode, Fergusobia cur-riei, spends one part of its life cycle in a fly and another part in eucalyptus trees.
the simple roundworm Caenorhabditis elegans consists, at maturity, of exactly 959 body cells with a variable number of reproductive cells. its Dna and development have been intensively studied. it is one of the most important laboratory animals.
the largest phylum of animals with ecdysis is the arthropods (phylum arthropoda). the most familiar arthropods are the centipedes, millipedes, spiders, and insects. Based on the number of species or the number of individuals, arthropods have been the evolutionary success story of the animal king-dom. these animals are so diverse and important that they are described in their own entries.
two phyla of animals with ecdysis resemble arthropods in structure and on the basis of Dna comparisons, as follows:
● the tardigrades, or water bears (phylum tardigrada), are microscopic aquatic seg-mented animals that walk slowly on eight legs. they are most famous for surviving extreme environmental conditions: they can survive at temperatures close to absolute zero and as high as 300° F (150° c), can absorb a thousand times as much radiation as any other known animal, and can revive after being dehydrated for years. they can even survive in outer space conditions.
xviii+598_Biodiversity_F.indd 27 11/17/11 1:12 PM

first entry28 animals 28
● the velvet worms (phylum Onychophora) look like annelids with stubby legs. they walk around in tropical vegetation look-ing for prey, which they trap in slime. like many arthropods, velvet worms breathe through tracheae, which are air tubes that branch through the body. Some of them produce live offspring, which is very rare outside the vertebrates.
in addition to the roundworms, water bears, velvet worms, and arthropods, the following phyla have ecdysis:
● Kinorhynchians (phylum Kinorhynchia) are animals that look like worms with seg-mented outer plates, and they pull them-selves through sediments with a ring of bristles around their retractable heads.
● Horsehair worms (phylum nematomorpha) are parasitic worms that resemble long, thin roundworms.
● Penis worms (phylum Priapulida) are filter feeders that burrow in sediments and extend a spiny proboscis out into the water.
● Loriciferans (phylum loricifera) are filter feeders that embed themselves in marine sediments. they can retract their heads into the protective outer coverings of their bodies.
the other phyla of protostomes are the following:
● Rotifers (phylum Rotifera) are microscopic animals that live in freshwater or the thin layers of water on soil particles. their heads are surrounded by rapidly moving cilia that scoop food particles into their stomachs. Many bdelloid rotifers are totally asexual. evolutionary scientists generally understand that sexual reproduction creates genetic diversity, which allows animals to evolve faster than their parasites. Bdelloid rotifers, however, can desiccate and blow away in the wind, leaving their fungal parasites behind.
● Ribbon worms (phylum nemertea) are long, thin worms that live mostly on the ocean floor. a ribbon worm can eject a proboscis out of its head to capture prey. Some ribbon worms are almost 100 feet (more than 30 m) long.
● Spiny-headed worms (phylum acantho-cephala) look like simple parasitic round-worms, except that they have a spiny proboscis they can distend from the body and imbed in the intestinal wall of a host.
● Only three species of cycliophore (phylum cycliophora) are known. Symbion pandora lives on the mouth bristles of some lob-sters but is not a parasite. though it has a horseshoe-shaped gut and a ring of cilia around its mouth, it does not have a true lophophore. Because its anatomy matches no other known animal and because of its complex life cycle, which matches the molt-ing phases of the lobster, it is assigned to its own phylum.
● Arrow worms (phylum chaetognatha) are small predatory worms with grasping spines, tiny compound eyes, and tiny teeth.
● Gastrotrichs (phylum Gastrotricha) are tiny worms that live in freshwater or marine sed-iments. they live only about three days.
● there is only one species of micrognatho-zoan (phylum Micrognathozoa). it was mistaken for a rotifer until close observa-tion showed it had complex jaws, which are absent in rotifers.
● Peanut worms (phylum Sipuncula) have a retractable, tentacled proboscis and live in burrows in shallow water.
● Spoon worms (phylum echiura) are genetically similar to annelids but are not segmented.
● Jaw worms (phylum Gnathostomulida) are tiny worms with relatively big jaws that they use to scrape food off sediment particles.
BIlaTErally SymmETrICal aNImalS: dEuTEroSTomES
Deuterostomes are animals whose embryos undergo cell division (cleavage) in a radial pattern. as a result, the structures into which the cells develop are not determined until a later stage than that of protostome embryos. the major groups of deutero-stomes are the echinoderms, the hemichordates, and the chordates.
the simplest deuterostomes are the simple, marine, mollusklike xenoturbellidans (phylum Xenoturbellida), recognized as deuterostomes only by Dna analysis.
echinoderms (phylum echinodermata) include starfish, sea urchins, and sand dollars as well as other forms less frequently seen. adult echino-derms appear radially symmetrical, but their larval forms and their internal organs have some bilaterial symmetry.
echinoderms have an external skeleton con-taining calcium or magnesium, often with knobs or spines (echinoderm is Greek for “spiny skin”). the skeleton offers protection from many predators, but not from desiccation. echinoderms are entirely
xviii+598_Biodiversity_F.indd 28 11/17/11 1:12 PM

animals 29
marine. the five “legs” of a starfish are not really legs, they are portions of the body. Starfish move by means of hundreds of tiny tube feet on the underside of their bodies. Water inside the tube feet is pressur-ized by structures at the bases of the feet. the tube feet create suction against the surfaces on which they walk.
Starfish and many other echinoderms have a mouth in the center of the underside of the body. a starfish uses its tube feet to pull open tightly closed shells of bivalves, then disgorges its stomach out through its mouth and into the bivalve. the bivalve shells need to be pulled apart as little as one 200th of an inch (a 10th of a millimeter) to allow the starfish to insert its stomach. Once it has partly digested the bivalve’s tissues, it absorbs the digested material. Starfish eat animals such as corals by placing their stomachs directly on the corals and digesting them.
no other phylum has the water vascular system found in echinoderms. Seawater circulates and car-ries oxygen throughout the body in this system of canals. the tube feet are connected to this system.
echinoderms release eggs and sperm into the water, and fertilization occurs externally. a bilat-erally symmetrical larva, which swims with cilia, develops from the fertilized egg.
Populations of some starfish have exploded and have eaten many coral polyps, causing extensive damage to some Pacific reefs. Populations of some sea urchins have exploded in the Pacific Ocean. the urchins have eaten up entire seaweed forests, causing disaster for many other species.
Many hemichordates (phylum Hemichordata), the acorn worms, look like simple worms with a pro-boscis, collar, and trunk. they burrow in sediments. Other hemichordates are filter feeders that form extensive colonial networks. as the name implies, they have a number of features that resemble those of chordates: the adults may have gill slits and a nerve cord along the dorsal (back) surface rather than the ventral (front) surface, as in most other phyla. Dna studies have identified them as close relatives of echinoderms and chordates.
the chordates (phylum chordata) include all of the vertebrates, but some invertebrates as well. all chordates have, at some stage of their life cycles, these characteristics:
● a dorsal nerve cord (along the back) ● slits in the neck region called gill slits ● a notochord
the notochord is a flexible rod that provides reinforcement for muscles. Some of the simplest chordates retain these features throughout their
lives, while others have them only in their juvenile stages. Most vertebrates have gill slits and notochord only during embryonic development. at birth, most vertebrates have a backbone with vertebrae instead of a notochord. Because of these similarities, the invertebrate chordates are probably similar to the ancestors of vertebrates.
Lancelets are fish-shaped invertebrate chordates that retain all three chordate features throughout their lives. they lie half buried on the bottoms of shallow seas, with their tails in the sand and their jawless mouths filtering small particles of food from the water. Tunicates have some chordate features during their fishlike larval stage. as adults, however, tunicates do not resemble fish. the larvae settle onto rock surfaces and change into adults. adult tunicates draw water in through an opening on top, filter food particles from the water, and eject the water through a pore along their sides. this man-ner of life has earned adult tunicates the name “sea squirts.” a layer of muscle surrounds their internal organs, including a heart, intestines, and reproduc-tive organs.
the vertebrates are the fishes, amphibians, rep-tiles, birds, and mammals. like the arthropod exo-skeleton, the vertebrate structure has allowed an explosion of diversity and adaptation.
paraSITIC INvErTEBraTESParasitic flatworms and roundworms cause an immense amount of human suffering, mostly in developing tropical countries. Usually, parasitic worms do not kill the human in which they live. the worms may, however, cause much pain and weak-ness in muscles and other organs and if they live in the intestines may consume a large share of the food that the person has eaten, food the victim probably has not gotten enough of anyway. the weakened victim becomes vulnerable to microbial diseases and is a less productive member of society.
adult liver flukes, Opisthorchis sinensis, may reach several centimeters in length inside a human body. they live in the duct that conducts fluids from the liver to the intestines. the flukes mate and can produce millions of eggs, which drain into the intes-tines, ending up in sewage (see figure on page 30). the eggs, if ingested by a snail, hatch into larvae, which form cysts (protective casts) in the snail’s tissues. inside the cysts, these larvae develop into another type of larva. each of these larvae can undergo sex-ual reproduction to form up to 50 larvae of yet a third developmental stage. these larvae emerge from the snail’s skin into the water. When they encounter a fish, they burrow into the fish’s muscle, chang-ing form yet again to become another kind of larva inside cysts. When the fish is caught by a human and
xviii+598_Biodiversity_F.indd 29 11/17/11 1:12 PM
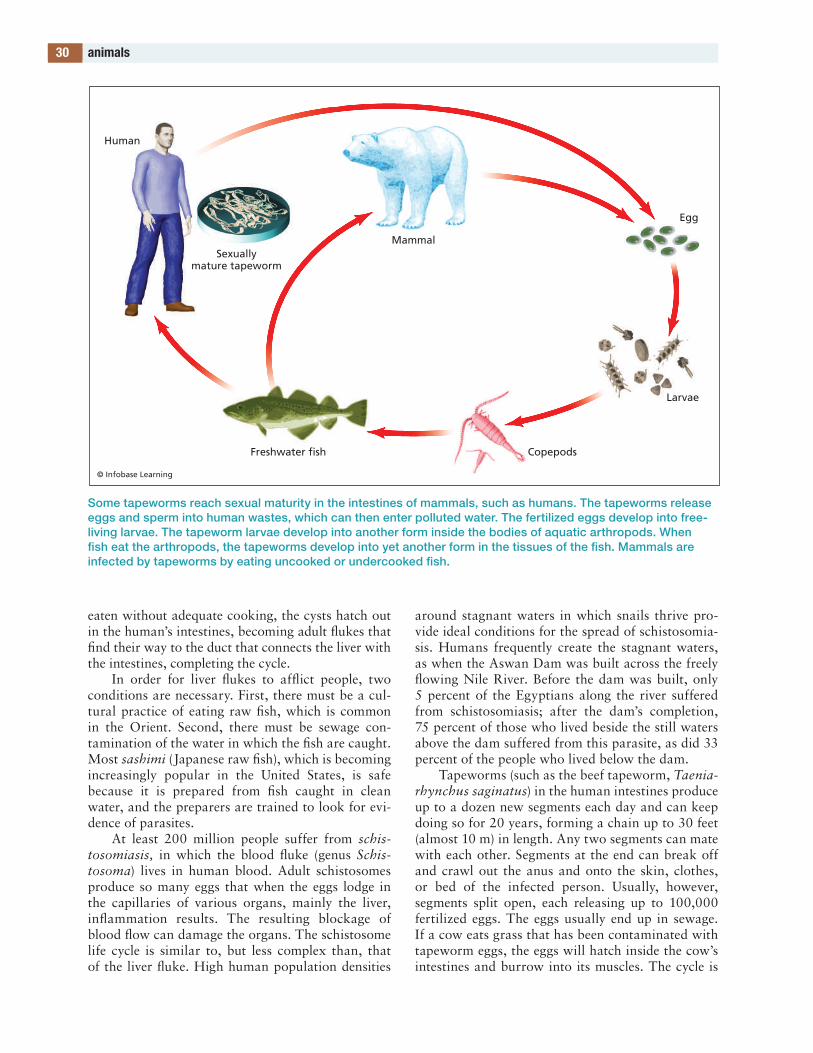
fi rst entry30 animals 30
eaten without adequate cooking, the cysts hatch out in the human’s intestines, becoming adult fl ukes that fi nd their way to the duct that connects the liver with the intestines, completing the cycle.
in order for liver fl ukes to affl ict people, two conditions are necessary. First, there must be a cul-tural practice of eating raw fi sh, which is common in the Orient. Second, there must be sewage con-tamination of the water in which the fi sh are caught. Most sashimi (Japanese raw fi sh), which is becoming increasingly popular in the United States, is safe because it is prepared from fi sh caught in clean water, and the preparers are trained to look for evi-dence of parasites.
at least 200 million people suffer from schis-tosomiasis, in which the blood fl uke (genus Schis-tosoma) lives in human blood. adult schistosomes produce so many eggs that when the eggs lodge in the capillaries of various organs, mainly the liver, infl ammation results. the resulting blockage of blood fl ow can damage the organs. the schistosome life cycle is similar to, but less complex than, that of the liver fl uke. High human population densities
around stagnant waters in which snails thrive pro-vide ideal conditions for the spread of schistosomia-sis. Humans frequently create the stagnant waters, as when the aswan Dam was built across the freely fl owing nile River. Before the dam was built, only 5 percent of the egyptians along the river suffered from schistosomiasis; after the dam’s completion, 75 percent of those who lived beside the still waters above the dam suffered from this parasite, as did 33 percent of the people who lived below the dam.
tapeworms (such as the beef tapeworm, Taenia-rhynchus saginatus) in the human intestines produce up to a dozen new segments each day and can keep doing so for 20 years, forming a chain up to 30 feet (almost 10 m) in length. any two segments can mate with each other. Segments at the end can break off and crawl out the anus and onto the skin, clothes, or bed of the infected person. Usually, however, segments split open, each releasing up to 100,000 fertilized eggs. the eggs usually end up in sewage. if a cow eats grass that has been contaminated with tapeworm eggs, the eggs will hatch inside the cow’s intestines and burrow into its muscles. the cycle is
Some tapeworms reach sexual maturity in the intestines of mammals, such as humans. The tapeworms release eggs and sperm into human wastes, which can then enter polluted water. The fertilized eggs develop into free-living larvae. The tapeworm larvae develop into another form inside the bodies of aquatic arthropods. When fi sh eat the arthropods, the tapeworms develop into yet another form in the tissues of the fi sh. Mammals are infected by tapeworms by eating uncooked or undercooked fi sh.
xviii+598_Biodiversity_F.indd 30 11/17/11 1:12 PM

animals 31
completed when a human eats undercooked beef. the pork tapeworm, Taenia solium, has a similar life cycle. Most beef and pork is not infected, but meat inspections are incomplete. eating rare meat therefore carries significant risk of infection by tape-worms. Trichinella spiralis roundworms have a life cycle similar to tapeworms and also spread through the consumption of undercooked meat.
adults of the dog tapeworm, Dipylidium cani-num, grow and produce eggs in the intestines of dogs. eggs can get into the dog’s blood, also, and from there get into the guts of fleas that feed on the dog’s blood. the fleas can then spread the parasite to another dog. Dogs may get tapeworm eggs in their mouths from licking surfaces contaminated by dog feces. Humans can be accidentally infected by the dog tapeworm, either from the bites of infected fleas or from direct contact with dogs. the tapeworm lar-vae can form cysts in various human organs, includ-ing the brain.
eight kinds of filarial worms cause disease in 400 million people. adult worms feed and mate within the lymph system of humans, blocking the vessels and causing the swelling and disfigurement often called elephantiasis. after mating, the female worms release larvae into the lymph. the larvae can spread to other humans by mosquito bites. Other filarial worms cause river blindness. adult worms live in the outer cell layer of the cornea, causing severe itching and eventually blindness. the bites of black flies transmit the worms to humans. in some african and South american villages, nearly every elderly person is blind from this disease.
acanthocephalan worms usually live in the intestines of crustaceans. When shorebirds eat the crustaceans, the acanthocephalans can infest the birds’ intestines as well. in at least one case, the acanthocephalan produces massive amounts of sero-tonin, a hormone that influences the crustacean’s behavior. crustaceans that normally hide will, as a result of the serotonin, stick their bodies out where ducks can easily find and eat them.
Parasitic worms have a major impact on a bil-lion people in the world. it is not necessary to wait for a medical breakthrough in order to treat these illnesses. Medicines, though sometimes expensive, are available. it is more effective, however, to control the disease ecologically by preventing the spread of the worms, in most cases by sewage treatment. However, many governments are unable or unwill-ing to undertake the building of sewage treatment facilities. Until they do, these diseases will persist. the unsanitary conditions of poverty encourage the spread of these worms, but the illnesses caused by these worms, even when mild, make it difficult for the victims to raise themselves out of poverty.
See also amphibians; arthropods; birds; continental shelves, biodiversity on; crus-taceans; deep oceans, biodiversity in; eukaryotes; fishes; insects; invasive species; mammals; mutualism; parasitism; reptiles.
Further readingarizono, naoki, et al. “Diphyllobothriasis associated with
eating raw Pacific salmon.” Emerging Infectious Dis-eases 15 (2009): 866–870.
Dellaporta, Stephen l., et al. “Mitochondrial genome of Trichoplax adhaerens supports Placozoa as the basal lower metazoan phylum.” Proceedings of the National Academy of Sciences USA 103 (2006): 8,751–8,756.
Donoghue, P. c. J., and M. a. Purnell. “Genome duplica-tion, extinction, and vertebrate evolution.” Trends in Ecology and Evolution 20 (2005): 312–319.
emlen, Douglas J. “the evolution of animal weapons.” Annual Reviews of Ecology, Evolution, and Systemat-ics 39 (2008): 387–413.
Fenwick, alan. “Waterborne infectious diseases—could they be consigned to history?” Science 313 (2006): 1,077–1,081.
Glenner, Henrik, et al. “the origin of insects.” Science 314 (2006): 1,883–1,884.
Jönsson, K. ingemar, et al. “tardigrades survive exposure to space in low earth orbit.” Current Biology 18 (2008): R729–731.
lafferty, K. D. “Fishing for lobsters indirectly increases epidemics in sea urchins.” Ecological Applications 14 (2004): 1,566–1,573.
Mather, Jennifer a. “cephalopod consciousness: Behav-ioral evidence.” Consciousness and Cognition 17 (2008): 37–48.
Pearse, J. S. “ecological role of purple sea urchins.” Science 314 (2006): 940–941.
Peigné, J., et al. “earthworm populations under different tillage systems in organic farming.” Soil and Tillage Research 104 (2009): 207–214.
Power, Brian, and carolyn Goossens. “Hallucinations aris-ing in the context of torn attachment, traumatic child-hood, and tapeworms.” Australasian Psychiatry 17 (2009): 240–242.
Skelly, Patrick. “Fighting killer worms.” Scientific Ameri-can, May 2008, 94–99.
Wilson, christopher G., and Paul W. Sherman. “anciently asexual bdelloid rotifers escape lethal fungal parasites by drying up and blowing away.” Science 327 (2010): 574–576.
Zimmer, carl. Parasite Rex: Inside the Bizarre World of Nature’s Most Dangerous Creatures. new york: Free Press, 2001.
———. “How smart is the octopus? Bright enough to do the moving-rock trick.” Slate, June 23, 2008. avail-able online. URl: http://www.slate.com/id/2192211. accessed May 11, 2011.
xviii+598_Biodiversity_F.indd 31 11/17/11 1:12 PM

first entry32 archaea 32
archaea Formerly known as archaebacteria, the archaea, or archeans, are single-celled bacteria-like organisms, most of which live in extreme condi-tions of temperature, salt, or acidity. Most of them have been discovered only recently, because they live in conditions in which scientists did not expect to find any organisms. their superficial resemblance to bacteria also contributed to their uniqueness being overlooked for many decades. among their similari-ties to bacteria are the following:
● like bacteria, archaeans have simple shapes, usually spheres, rods, or spirals.
● like most bacteria, most archaeans have cell walls. in both bacteria and archaeans, there are some types that lack cell walls (Mycoplasma in bacteria, Thermoplasma in archaeans) and have irregular shapes.
● like bacteria, archaeans have Dna but no nucleus; both are prokaryotes.
● like many bacteria, many archaeans can form colonies.
● Both bacteria and archaeans are very small, often 10 times smaller than eukaryotic cells.
● Bacteria can be aerobic (grow in the pres-ence of oxygen gas) or anaerobic. Most archaeans are anaerobic, although some Sulfolobus archaeans can tolerate oxygen.
However, archaeans are distinct from bacteria in some fundamental ways, such as the following:
● Bacterial cell walls consist largely of pepti-doglycan; archaean cell walls do not.
● cell membranes of all cells consist largely of phospholipids, but these are different in bacteria and archaeans. in bacteria, the phospholipids have ester linkages, while archaean cell membranes have phospholip-ids with ether linkages instead. the fatty chain of the bacterial phospholipid is a fatty acid, while in archaeans it is an iso-prene. eukaryotic cell membrane phospho-lipids are similar to those of bacteria, not archaeans.
● cell membranes in bacteria (and eukaryotes) are double layered, while in some archaeans they form a single layer.
● Some archaeans have shapes not found in bacteria, such as flat, square forms and extremely narrow, needlelike forms.
● archaean Dna is associated with histone proteins, like the chromosomes of eukary-otic cells; the Dna of bacteria is not.
● archaeans have enzymes associated with the transcription of Rna and translation of
proteins that more closely resemble those of eukaryotic cells than of bacteria.
● Both bacteria and archaeans often have fla-gella, whiplike structures that allow them to swim, but the flagella of bacteria and archaeans appear to have evolved separately, the former from secretory proteins, the lat-ter from hairlike extensions called pili.
● Both bacteria and archaeans have many of the same ways of obtaining metabolic energy (see table on page 33). For example, some species in both groups obtain energy from sunlight or obtain carbon from inorganic sources such as carbon dioxide. Some bacteria (such as cyanobacteria) can do both, but there are no archaeans that can do both. Some types of metabolism, such as the production of meth-ane gas, are unique to archaeans.
these differences, plus the fact that archaeans have very different Dna sequences from those of bacteria (up to 15 percent of archaean genes encode proteins not known in bacteria), justify the classifi-cation of the archaea as one of the three principal domains of life, equally distinct from bacteria and eucarya (eukaryotes).
Many of the organisms that live in extreme con-ditions are archaeans. Most of them live in conditions of extreme acidity, temperature, salinity, or pres-sure that biologists at one time considered impos-sible for organisms to endure. Pyrolobus fumarii, for example, lives in deep ocean vents where the water reaches 235°F (113°c). Water boils at 212°F (100°c) at the air pressure of sea level, but the pressure at the bottom of the ocean allows superheated water to exist, in which this archaean lives. However, the most primitive lineages of bacteria, represented by Aquifex and Thermotoga, thrive under extreme con-ditions as well. the earliest bacteria, like the earli-est archaeans, must therefore have evolved under extreme conditions on the early earth.
the extreme conditions in which most archae-ans live are not necessarily distant from the places where most eukaryotes reside. Many archaeans live in animal intestines (under extreme anoxic, basic, or acidic conditions), where they digest cellulose and other molecules. Some intestinal archaeans are commensalistic, neither beneficial nor harmful. an example is Methanobrevibacter smithii, a methane-producing archaean that lives in human intestines. not all archaeans, however, live in extreme condi-tions. For example, archaeans are a major compo-nent of marine plankton. no archaeans are known to cause diseases in eukaryotes.
the evolutionary scientist carl Woese began making nucleic acid comparisons among many dif-
xviii+598_Biodiversity_F.indd 32 11/17/11 1:12 PM
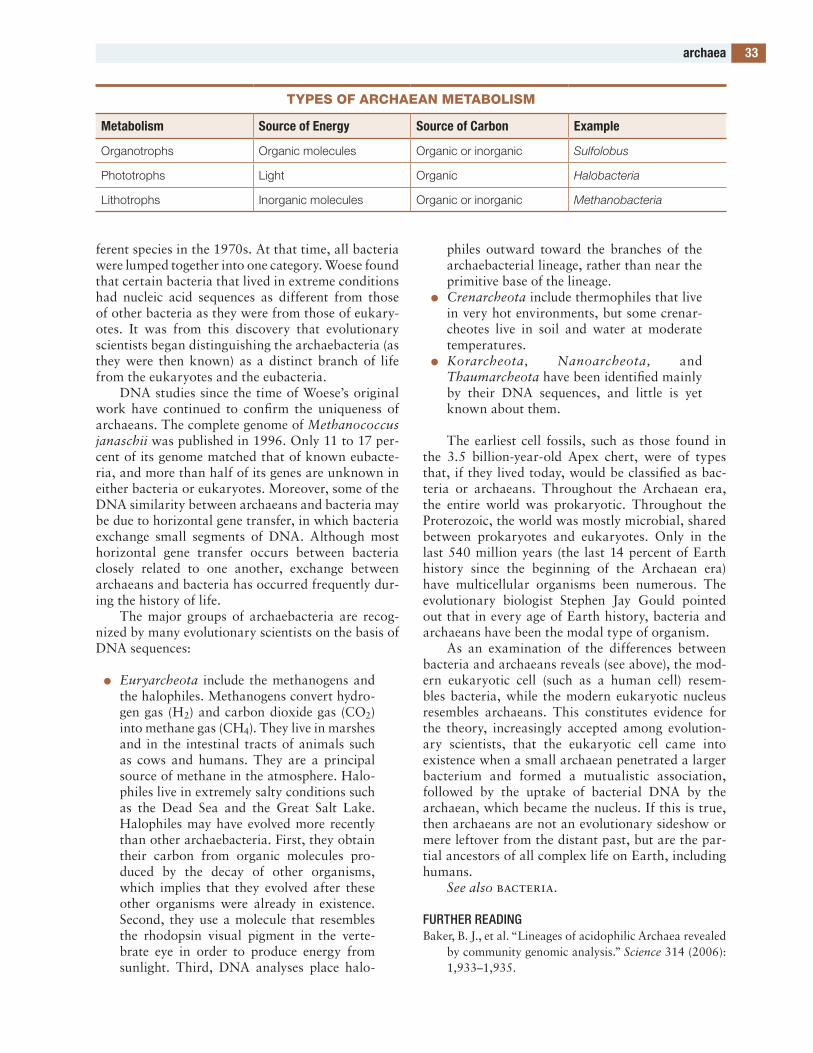
archaea 33
ferent species in the 1970s. at that time, all bacteria were lumped together into one category. Woese found that certain bacteria that lived in extreme conditions had nucleic acid sequences as different from those of other bacteria as they were from those of eukary-otes. it was from this discovery that evolutionary scientists began distinguishing the archaebacteria (as they were then known) as a distinct branch of life from the eukaryotes and the eubacteria.
Dna studies since the time of Woese’s original work have continued to confirm the uniqueness of archaeans. the complete genome of Methanococcus janaschii was published in 1996. Only 11 to 17 per-cent of its genome matched that of known eubacte-ria, and more than half of its genes are unknown in either bacteria or eukaryotes. Moreover, some of the Dna similarity between archaeans and bacteria may be due to horizontal gene transfer, in which bacteria exchange small segments of Dna. although most horizontal gene transfer occurs between bacteria closely related to one another, exchange between archaeans and bacteria has occurred frequently dur-ing the history of life.
the major groups of archaebacteria are recog-nized by many evolutionary scientists on the basis of Dna sequences:
● Euryarcheota include the methanogens and the halophiles. Methanogens convert hydro-gen gas (H2) and carbon dioxide gas (cO2) into methane gas (cH4). they live in marshes and in the intestinal tracts of animals such as cows and humans. they are a principal source of methane in the atmosphere. Halo-philes live in extremely salty conditions such as the Dead Sea and the Great Salt lake. Halophiles may have evolved more recently than other archaebacteria. First, they obtain their carbon from organic molecules pro-duced by the decay of other organisms, which implies that they evolved after these other organisms were already in existence. Second, they use a molecule that resembles the rhodopsin visual pigment in the verte-brate eye in order to produce energy from sunlight. third, Dna analyses place halo-
philes outward toward the branches of the archaebacterial lineage, rather than near the primitive base of the lineage.
● Crenarcheota include thermophiles that live in very hot environments, but some crenar-cheotes live in soil and water at moderate temperatures.
● Korarcheota, Nanoarcheota, and Thaumarcheota have been identified mainly by their Dna sequences, and little is yet known about them.
the earliest cell fossils, such as those found in the 3.5 billion-year-old apex chert, were of types that, if they lived today, would be classified as bac-teria or archaeans. throughout the archaean era, the entire world was prokaryotic. throughout the Proterozoic, the world was mostly microbial, shared between prokaryotes and eukaryotes. Only in the last 540 million years (the last 14 percent of earth history since the beginning of the archaean era) have multicellular organisms been numerous. the evolutionary biologist Stephen Jay Gould pointed out that in every age of earth history, bacteria and archaeans have been the modal type of organism.
as an examination of the differences between bacteria and archaeans reveals (see above), the mod-ern eukaryotic cell (such as a human cell) resem-bles bacteria, while the modern eukaryotic nucleus resembles archaeans. this constitutes evidence for the theory, increasingly accepted among evolution-ary scientists, that the eukaryotic cell came into existence when a small archaean penetrated a larger bacterium and formed a mutualistic association, followed by the uptake of bacterial Dna by the archaean, which became the nucleus. if this is true, then archaeans are not an evolutionary sideshow or mere leftover from the distant past, but are the par-tial ancestors of all complex life on earth, including humans.
See also bacteria.
Further readingBaker, B. J., et al. “lineages of acidophilic archaea revealed
by community genomic analysis.” Science 314 (2006): 1,933–1,935.
types of arChaean metabolism
metabolism Source of Energy Source of Carbon Example
Organotrophs Organic molecules Organic or inorganic Sulfolobus
Phototrophs Light Organic Halobacteria
Lithotrophs Inorganic molecules Organic or inorganic Methanobacteria
xviii+598_Biodiversity_F.indd 33 11/17/11 1:12 PM
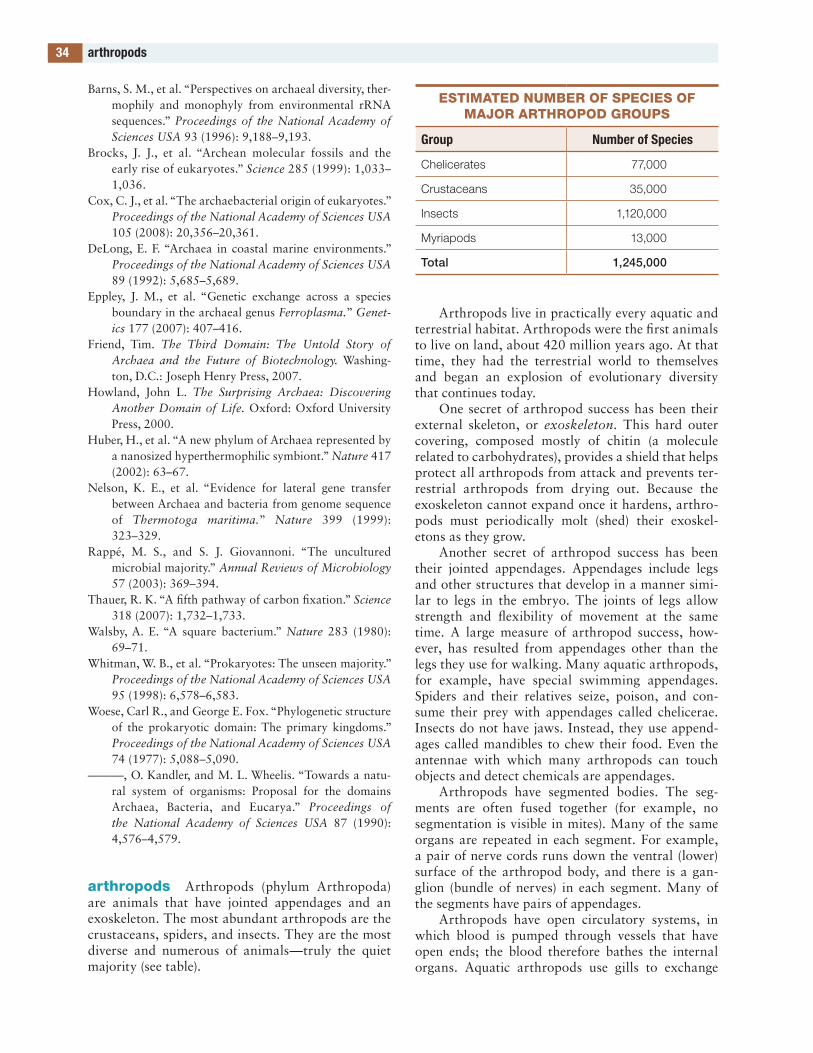
first entry34 arthropods 34
Barns, S. M., et al. “Perspectives on archaeal diversity, ther-mophily and monophyly from environmental rRna sequences.” Proceedings of the National Academy of Sciences USA 93 (1996): 9,188–9,193.
Brocks, J. J., et al. “archean molecular fossils and the early rise of eukaryotes.” Science 285 (1999): 1,033–1,036.
cox, c. J., et al. “the archaebacterial origin of eukaryotes.” Proceedings of the National Academy of Sciences USA 105 (2008): 20,356–20,361.
Delong, e. F. “archaea in coastal marine environments.” Proceedings of the National Academy of Sciences USA 89 (1992): 5,685–5,689.
eppley, J. M., et al. “Genetic exchange across a species boundary in the archaeal genus Ferroplasma.” Genet-ics 177 (2007): 407–416.
Friend, tim. The Third Domain: The Untold Story of Archaea and the Future of Biotechnology. Washing-ton, D.c.: Joseph Henry Press, 2007.
Howland, John l. The Surprising Archaea: Discovering Another Domain of Life. Oxford: Oxford University Press, 2000.
Huber, H., et al. “a new phylum of archaea represented by a nanosized hyperthermophilic symbiont.” Nature 417 (2002): 63–67.
nelson, K. e., et al. “evidence for lateral gene transfer between archaea and bacteria from genome sequence of Thermotoga maritima.” Nature 399 (1999): 323–329.
Rappé, M. S., and S. J. Giovannoni. “the uncultured microbial majority.” Annual Reviews of Microbiology 57 (2003): 369–394.
thauer, R. K. “a fifth pathway of carbon fixation.” Science 318 (2007): 1,732–1,733.
Walsby, a. e. “a square bacterium.” Nature 283 (1980): 69–71.
Whitman, W. B., et al. “Prokaryotes: the unseen majority.” Proceedings of the National Academy of Sciences USA 95 (1998): 6,578–6,583.
Woese, carl R., and George e. Fox. “Phylogenetic structure of the prokaryotic domain: the primary kingdoms.” Proceedings of the National Academy of Sciences USA 74 (1977): 5,088–5,090.
———, O. Kandler, and M. l. Wheelis. “towards a natu-ral system of organisms: Proposal for the domains archaea, Bacteria, and eucarya.” Proceedings of the National Academy of Sciences USA 87 (1990): 4,576–4,579.
arthropods arthropods (phylum arthropoda) are animals that have jointed appendages and an exoskeleton. the most abundant arthropods are the crustaceans, spiders, and insects. they are the most diverse and numerous of animals—truly the quiet majority (see table).
arthropods live in practically every aquatic and terrestrial habitat. arthropods were the first animals to live on land, about 420 million years ago. at that time, they had the terrestrial world to themselves and began an explosion of evolutionary diversity that continues today.
One secret of arthropod success has been their external skeleton, or exoskeleton. this hard outer covering, composed mostly of chitin (a molecule related to carbohydrates), provides a shield that helps protect all arthropods from attack and prevents ter-restrial arthropods from drying out. Because the exoskeleton cannot expand once it hardens, arthro-pods must periodically molt (shed) their exoskel-etons as they grow.
another secret of arthropod success has been their jointed appendages. appendages include legs and other structures that develop in a manner simi-lar to legs in the embryo. the joints of legs allow strength and flexibility of movement at the same time. a large measure of arthropod success, how-ever, has resulted from appendages other than the legs they use for walking. Many aquatic arthropods, for example, have special swimming appendages. Spiders and their relatives seize, poison, and con-sume their prey with appendages called chelicerae. insects do not have jaws. instead, they use append-ages called mandibles to chew their food. even the antennae with which many arthropods can touch objects and detect chemicals are appendages.
arthropods have segmented bodies. the seg-ments are often fused together (for example, no segmentation is visible in mites). Many of the same organs are repeated in each segment. For example, a pair of nerve cords runs down the ventral (lower) surface of the arthropod body, and there is a gan-glion (bundle of nerves) in each segment. Many of the segments have pairs of appendages.
arthropods have open circulatory systems, in which blood is pumped through vessels that have open ends; the blood therefore bathes the internal organs. aquatic arthropods use gills to exchange
estimated number of speCies of major arthropod groups
Group Number of Species
Chelicerates 77,000
Crustaceans 35,000
Insects 1,120,000
Myriapods 13,000
Total 1,245,000
xviii+598_Biodiversity_F.indd 34 11/17/11 1:12 PM

arthropods 35
oxygen with their environments. insects breathe by means of tracheae, breathing tubes that pen-etrate throughout their bodies. this is a very effi-cient system for a small animal. tracheae would not, however, provide enough surface area to pro-vide sufficient oxygen to the tissues of a very large insect. therefore, one of the reasons that insects are mostly small is that huge, “monster,” insects would be unable to breathe. Spiders have simple lungs.
arthropods have two kinds of eyes. the ocelli are usually little more than pits with visual pigments and allow the animal to detect the direction from which danger may be approaching. the ocelli of spiders, however, can form images. the compound eyes of arthropods form simple images because each facet is a pixel of light, darkness, and, in many cases, color.
all terrestrial and many aquatic arthropods have internal fertilization. this is true even of adult barnacles, which cannot move. the male barnacle extends a lengthy penis to fertilize a nearby female. Many arthropods have appendages specialized for sexual reproduction. Some arthropods can repro-duce asexually. aphids (insects that consume sap from plants) may have many generations during a single summer in which new aphids hatch from unfertilized eggs laid by female parents that have not mated. in many arthropods, the larval phase (such as a caterpillar) has a structure and function very different from the adult phase (such as a butterfly).
myrIapodSMyriapods are the centipedes, millipedes, and related groups. their name refers to their large num-ber of legs: Most have a pair of legs on nearly every one of their many segments. Most live in moist forests. they have simple eyes. they breathe with tracheae; myriapod tracheae evolved separately from those of insects. Myriapods were among the earliest land animals, beginning in the late Silurian period. Myriapods include the following.
● Centipedes (chilopoda) have venom claws on their heads and are predators. “centi-pede” means “hundred legs” in latin, but none have quite this many.
● Millipedes (Diplopoda) consume decaying plant material. they appear to have two pairs of legs per segment rather than one due to the fusion of segment pairs during development. they defend themselves by curling into a coil.
● Pauropods (Pauropoda) and symphylans (Symphyla) resemble tiny, pale millipedes. they live primarily in the pore spaces of soil.
CruSTaCEaNSCrustaceans, nearly all aquatic, include crayfish, lobsters, crabs, shrimp, barnacles, and many micro-scopic plankton. they are very abundant and impor-tant members of aquatic ecosystems around the world and have their own entry in this encyclopedia. Some isopods, such as pill bugs, live on land in moist soil mostly underneath rocks. the largest group of crustaceans, the decapods (from the latin for “ten legs”), have five pairs of walking legs on the thorax. they manipulate food with the front appendages of the thorax, which are sometimes modified into big pincers. the appendages of their abdomens are often modified for swimming.
ChElICEraTESChelicerates include spiders, scorpions, ticks, mites, and their relatives. they often have two body parts: the cephalothorax (head and thorax fused), with appendages, and the abdomen, without appendages. the first pair of appendages on the cephalothorax is the chelicerae, which are paired mouthparts used to seize and pierce prey. in spiders, the chelicerae insert venom into the prey. Behind the chelicerae are the pedipalps. if the chelicerae are large, the pedipalps may be used primarily for walking or as sensory structures. Otherwise, they grasp and manipulate the prey.
the largest group of chelicerates is the arachnids. they lack antennae and have four pairs of walking legs. Sexual reproduction in arachnids such as spiders is a touchy and dangerous operation. after the final molt, the adult male stops eating. the male spider puts sperm on one of his front appendages and goes searching for a female. He uses special approach signals so that he is not mistaken for prey. the male inserts the appendage with sperm into a pore in the female’s abdomen. Mating in scorpions is even more risky than in spiders and involves complex dances and the transfer of sperm into the female’s abdomen by one of the male’s legs. in some species, the female spider or scorpion eats the male after mating.
the largest group of arachnids is the spiders, described in a separate entry. among the groups of arachnids that resemble spiders are the following.
● Harvestmen (“daddy longlegs”) resemble spiders, but their body segments are largely fused into a single structure. they have extremely long legs. Unlike spiders but like insects, they breathe through tracheae. they are not known to produce venom or silk. also unlike spiders, they can eat, not just suck. they are harmless to humans.
● Sunscorpions (solifugids), unlike spiders, have a segmented abdomen. Because their
xviii+598_Biodiversity_F.indd 35 11/17/11 1:12 PM

first entry36 arthropods 36
front legs are small and sensory, they walk primarily on six legs. like harvestmen and insects, they breathe through tracheae. their pedipalps produce an adhesive that captures prey. they produce no venom. the chelicerae cut up the prey. enzymes liquefy the internal organs of the prey. Sunscor-pions must suck because they cannot eat. they also differ from true scorpions (see below).
● Hooded tickspiders are small and blind and have a hood with which they can cover their heads.
Scorpions are nocturnal predators with big clawlike chelicerae and a stinging tail that they hold over their bodies (see figure). Unlike other arthro-pods, they bear live young. like spiders, they cannot eat food items but must liquefy the internal organs of their victims with enzymes. they are associated with deserts, although scorpions live in a wide range of terrestrial environments. Many people greatly fear scorpions not because scorpions seek out and attack people, but because an accidental encounter with a scorpion hiding in a shoe or a corner can result in a painful and possibly deadly sting. Only about 20 of the 2,000 species are lethal to humans, but these cause 10 times as many human deaths as do snake-bites, mostly in the Middle east and north africa. the only deadly species in the United States is the arizona bark scorpion (Centruroides exilicauda). its sting, if untreated, kills fewer than 1 percent of adults but up to a quarter of children. adult scor-pions are fluorescent in ultraviolet light; blacklights are used to locate scorpions at night.
among the groups that resemble scorpions are the following.
● Pseudoscorpions are not scorpions. although they have large pincers with venom, they do not have stinging tails.
● Whip scorpions are also not true scorpions. their tails may appear whiplike. there are several groups of whip scorpions. the tail-less whip scorpions have front sensory legs that can be very large relative to their bod-ies, up to 10 inches (25 cm) long. Microwhip scorpions use their pedipalps for walking and their front legs as sensory structures. the vinegaroons also use their front legs as sensory structures and have long tails with-out stingers. the split-middle whipscorpi-ons appear to have three body parts because the thorax has two plates.
the other major group of arachnids is the mites and ticks. Mites and ticks are small and have little visible segmentation. they often have eight legs, but some, such as the follicle mite, Demodex folliculo-rum, and the larva of the deer tick, Ixodes dammin-nii, have fewer. ticks and mites have mouthparts that can bite, sting, saw, or suck.
Herbivorous mites can be major agricultural pests. Follicle mites are harmless and live in the fol-licles of human eyelashes, although under some con-ditions they can cause an immune response. Mites can also be predators or parasites. chiggers (genus Trombicula) are immature mites that dig into human skin by mistake, where they die, as they cannot com-plete their development on a human host—small comfort to the host. Some mites live in carpet dust or pillows, where they consume dead cells. they are harmless except when they cause allergic reactions in humans.
Most ticks are small external parasites that feed on the blood of vertebrates. the first immature stage hatches in the spring and waits on branches and leaves to encounter a host. after feeding until enor-mously swollen, the immature tick falls off, molts into a second immature stage, and finds another host. after it feeds this second time, it falls off, molts into adulthood, and finds its final host. ticks and mites can sometimes be harmful to humans, mainly because they transmit microbial diseases such as lyme disease.
two of the lesser-known groups of nonarachnid chelicerates are the following.
● Sea spiders (Pycnogonida) are usually small, and their muscles often consist of just one muscle cell. Some antarctic species can be An emperor scorpion (Reptile Apartment)
xviii+598_Biodiversity_F.indd 36 11/17/11 1:12 PM

arthropods 37
almost three feet (90 cm) in diameter, which includes the legs. Most have eight legs, though the number can vary.
● Only four species of horseshoe crabs (Xyphosura) survive from a group that was abundant in the Paleozoic era. largely unchanged in anatomy since that time, they have been considered one of the best examples of “living fossils.” they are not true crabs. atlantic horseshoe crabs (Limu-lus polyphemus), about a foot (less than half a meter) in length, can be very abun-dant when they come ashore to breed during spring high tides of full and new moons. the female can lay 20,000 eggs. a large carapace covers the cephalothorax. they have five pairs of walking legs, the first four of which are also used to process food. as in many arthropods, the blood of horseshoe crabs contains blue, copper-based hemocya-nin rather than red, iron-based hemoglobin.
INSECTS aNd ThEIr rElaTIvESinsects and their relatives are by far the most numer-ous and diverse arthropods. the world could not function without more than 1 million species of insects. at any given moment, there are a billion bil-lion (1018) insects on the planet. as herbivores, car-nivores, pollinators, and decomposers, they play a greater role in the flow of energy and matter through earth’s ecosystem than any other group. about 800 species of insects have become pests of agricultural crops. On the other hand, three-quarters of all com-mercial crops depend on bees for pollination. Flight has allowed insects to evolve many different pol-lination relationships with plants; the evolution of pollination relationships may have been the most important factor that contributed to the evolution of insect diversity. Because of their ecological and evolutionary importance, the insects have their own encyclopedia entry.
insects have three body parts and six legs. the abdomen contains digestive and reproductive organs. their six legs emerge from the thorax. the head has many appendages, including antennae and many mouthparts. instead of jaws and tongues, insects have appendages that are modified into struc-tures that fulfill the same roles. Many adult insects have a proboscis mouthpart with which they can penetrate the tissues of plants or animals. Some tiny arthropods that live in soil have six legs but do not have three body parts. therefore, most scientists classify them separately from the insects.
the evolution of wings, unique among arthro-pods, is one of the insects’ most important terrestrial adaptations. Wings allow some insects to travel at
great speeds. the sphinx moth can fly at 1,000 body lengths per second. a jet flying at three times the speed of sound travels only 100 body lengths per second. in addition to speed, wings allow insects tremendous maneuverability.
Most modern insects are either amphibious (liv-ing first in water, then on land) or entirely terrestrial. the first insects lived partly or wholly on land; a few modern insects have returned to an entirely aquatic existence.
insects develop from juvenile to adult by pass-ing through a series of molts, or instars. Some insects, such as grasshoppers, have relatively sim-ple life cycles, in which the juvenile insect that hatches from the egg closely resembles the adult. Wings may develop only on the final instar. Other insects undergo metamorphosis. they have a larval stage that looks almost completely different from the adult, as “grubs” bear little resemblance to adult beetles or bees, nor caterpillars to butterflies. the larvae are almost always adapted as “eating machines,” while the adults, which devote them-selves to reproduction, eat much less, if anything at all. in amphibious insects, such as the dragonfly, the juvenile stage lives underwater, but the adult lives on land and in the air.
examples of ecologically and economically important groups of insects are the following.
Coleopterans. there are more species of beetles (order coleoptera, sheath-winged) than of any other group of organisms in the world—at least 300,000 species, which is a quarter of all the species of organ-isms that are known in the world! With such diver-sity, beetles fill almost every ecological role. Some, such as the potato beetle and the boll weevil, are crop pests, while others, such as the ladybird beetle, eat crop pests. “lightning bugs” are actually beetles. their grubs pupate before developing into adults.
Lepidopterans. adult butterflies and moths have four usually colorful scaly wings. Moths often have brushy antennae, dusky wings, and nocturnal habits. the brushy antennae allow the male moths to detect even a few molecules of female sex attractant, which guide the male many kilometers to a mate. Butter-flies are often diurnal and have more wirelike anten-nae and colorful wings. the larvae of this order are caterpillars, with three pairs of jointed legs in front, followed by numerous pairs of prolegs, which are stubby projections with bristles. like beetle grubs, caterpillars are specialized to do mostly one thing: eat. the larva pupates as a cocoon (in moths) or a chrysalis (in butterflies), allowing a striking meta-morphosis to occur, including the development of wings and a change of mouthparts from chewing mandibles to hollow sucking mouthparts. caterpil-lars can devastate large areas of cropland or forest.
xviii+598_Biodiversity_F.indd 37 11/17/11 1:12 PM

first entry38 arthropods 38
Many adults, however, are important pollinators of wild and domesticated plants.
Hymenopterans. ants, bees, and wasps often have complex social systems. Many species live in colonies of individuals that hatch from eggs all laid by the same large, fertile female, the queen. the fer-tilized eggs develop only into females, while unfertil-ized eggs develop into male drones. Female honeybee larvae that receive regular food develop into work-ers, which perform many different jobs. Worker honeybees do not lay eggs, as their egg duct is modi-fied into a stinger. Some hymenopterans, especially wasps, are important to agriculture because they either hunt or parasitize the caterpillars that cause much crop damage and loss.
Dipterans. adult flies and mosquitoes can be pollinators but are often parasites that spread dis-ease. an adult mosquito eats little, if anything, but the female requires blood for the production of eggs. She seeks warm-blooded vertebrates by flying toward heat and carbon dioxide. Mosquitoes spread several diseases, such as malaria and various viral fevers. eradication of certain species of mosquitoes has been a major focus of public health efforts. Fly larvae can be parasitic (for example, the bot flies in cattle), but most maggots are decomposers.
Orthopterans. Since ancient times, grasshoppers and locusts have been major insect pests. even today, plagues of locusts can occur on the same level as those described in ancient biblical records.
Isopterans. termites are also social insects. Members of their colonies all hatch from eggs fer-tilized by a male king and laid by a large, fertile queen. Some of the eggs develop into males, some into females. termites eat wood and other plant materials. Many termites, however, cannot digest wood. they rely on microbes in their guts to digest cellulose. although many termites damage human structures, they are very important decomposers, especially in semiarid regions, and make a major contribution to the carbon cycle.
Homopterans. insects in the order Homop-tera, especially the aphids, also damage crop plants. aphids have piercing mouthparts that insert into the plant cells that carry sap. Because plant sap is pressurized, the aphids do not even need to drink; they let the plant pump the food through them faster than they can digest it. as a result, aphid excrement is mostly sap, which can drip on many things, such as a car that stands beneath an aphid-infested tree. ants may tend aphids as cowboys tend cattle, eating some of the sap that the aphids have funneled out. a homopteran, besides sucking sap, can spread micro-bial diseases from one plant to another by its probos-cis. aphid infestations can be controlled by ladybird beetle larvae or adults or by spraying them with soap.
Siphonapterans. Fleas spread several diseases, most notably bubonic plague. they are flattened ver-tically, which allows them to cling closely to the skin and hair of hosts. the human flea, Pulex irritans, does not spread plague; plague bacteria are not spread from one human to another by flea bites. the rat flea, Xenopsylla cheopsis, prefers to bite rats. When the rats die from plague or the fleas’ throats are blocked by the plague bacteria, fleas may bite humans, thereby spreading plague from rats to the humans.
Anoplurans. lice can be distinguished from fleas by their horizontal, rather than vertical, flat-tening. the flat bodies of lice, like those of fleas, are not easily scratched off. lice spread such diseases as typhus from one human to another under unsanitary conditions.
Many insects become pests when their popula-tions get out of control. Massive use of pesticides has not only failed to control them but has resulted in the evolution of superpests. Modern agricultural researchers now realize that the best and only per-manent control of insect pests is to release diseases, predators, or parasites to kill the pests or release sterilized adults of the pest species that will mix with the pest population and reduce its reproductive success. in addition, if humans try to spray all insect pests, they will also kill the many insects that are not only beneficial but also essential, especially the pollinators.
insects may provide great medical benefits. Medicines may be ineffective against some infected wounds, but doctors can sometimes use maggots—yes, the larvae of flies—for debridement, which is cleaning out decaying flesh from a wound! Scientists have wondered why some insects, such as ants, can live in dense populations without having outbreaks of disease. it appears to be because these insects pro-duce antibiotics that protect them. in the continual search for new antibiotics, researchers may have to turn to insects.
See also crustaceans; insects; living fos-sils; spiders.
Further readingBeccaloni, Jan. Arachnids. Berkeley: University of califor-
nia Press, 2009.Berenbaum, May. Bugs in the System: Insects and Their
Impact on Human Affairs. Upper Saddle River, n.J.: addison-Wesley, 1996.
———. Buzzwords: A Scientist Muses on Sex, Bugs, and Rock ‘n’ Roll. Washington, D.c.: national academies Press, 2000.
eisner, thomas. For Love of Insects. cambridge, Mass.: Harvard University Press, 2003.
———, and Jerrold Meinwald. “Defensive secretions of arthropods.” Science 153 (1966): 1,341–1,350.
xviii+598_Biodiversity_F.indd 38 11/17/11 1:12 PM

arthropods 39
Farinós, Gema P., et al. “Diversity and seasonal phenol-ogy of aboveground arthropods in conventional and transgenic maize crops in central Spain.” Biological Control 44 (2008): 362–371.
Farmer, edward e., and lucie Dubugnon. “Detritivorous crustaceans become herbivores on jasmonate-deficient plants.” Proceedings of the National Academy of Sci-ences USA 106 (2009): 935–940.
Fountain, Henry. “competing for food, beetle becomes a killer.” available online. New York Times, January 27, 2009. URl: http://www.nytimes.com/2009/01/27/science/27obdung.html. accessed May 12, 2011.
Franks, n. R. “army ants: a collective intelligence.” Ameri-can Scientist 77 (1989): 139–145.
Hillyard, Paul. The Private Life of Spiders. Princeton, n.J.: Princeton University Press, 2008.
Hölldobler, Bert, and edward O. Wilson. The Ants. cam-bridge, Mass.: Harvard University Press, 1990.
Richmond, Sonya, and David c. lasenby. “the behav-ioural response of mayfly nymphs (Stenonema sp.) to chemical cues from crayfish (Orconectes rusticus). Hydrobiologia 560 (2006): 335–343.
Sargent, William. Crab Wars: A Tale of Horseshoe Crabs, Bioterrorism, and Human Health. lebanon, n.H.: University Press of new england, 2006.
Sharley, David J., et al. “the effects of soil tillage on ben-eficial invertebrates within the vineyard.” Agricultural and Forest Entomology 10 (2008): 233–243.
Shuster, carl n., et al., eds. The American Horseshoe Crab. cambridge, Mass.: Harvard University Press, 2004.
Snyder, Bruce a., and Paul F. Hendrix. “current and poten-tial roles of soil macroinvertebrates (earthworms, millipedes, and isopods) in ecological restoration.” Restoration Ecology 16 (2008): 629–636.
———, et al. “competition between invasive earthworms (Amynthas corticis, Megascolecidae) and native north american millipedes (Pseudopolydesmus era-sus, Polydesmidae): effects on carbon cycling and soil structure.” Soil Biology and Biochemistry 41 (2009): 1,442–1,449.
thomas, Richard H. “Wingless insects and plucked chick-ens.” Science 299 (2003): 1,854–1,855.
University of Delaware Sea Grant Program. “Horseshoe crab.” available online. URl: http://www.ceoe.udel.edu/horseshoecrab. accessed July 16, 2009.
Weeks, andrew R., et al. “a mite species that consists entirely of haploid females.” Science 292 (2001): 2,479–2,482.
Willemart, Rodrigo H., et al. “Sexually dimorphic legs in a neotropical harvestman (arachnida, Opiliones): Orna-ment or weapon?” Behavioural Processes 80 (2009): 51–59.
Wu, yanling, et al. “anti-atherogenic effects of centipede acidic protein in rats fed an atherogenic diet.” Journal of Ethnopharmacology 12 (2009): 509–516.
xviii+598_Biodiversity_F.indd 39 11/17/11 1:12 PM

40
bacteria Of all the organisms in the world, bac-teria are among the (1) smallest, (2) simplest, (3) most numerous, (4) most metabolically diverse, and (5) oldest. the bacteria share these distinctions with the morphologically similar archaea, which were once considered to be bacteria.
Smallest. the typical bacterial cell is about 10 times smaller than the typical cell of complex organisms in each dimension. therefore, the typi-cal bacterium has a volume 1,000 times smaller than that of a typical complex cell, although there are exceptions. the largest known bacterium is Thiomargarita namibiensis, which is 1/25th of an inch (1 mm) in diameter. this is larger than many eukaryotic cells.
Simplest. Bacteria are prokaryotic, which means they do not have a nucleus (Greek, karyo). Human, plant, fungus, and protist cells are eukaryoticbecause they have a nucleus. Most bacteria have:
● a cell wall, which consists mostly of a com-bination of protein and polysaccharide called peptidoglycan. Because Gram’s stain indicates the presence of peptidoglycan, the gram-positive bacteria are those with a thick cell wall, and the gram-negative bac-teria have thinner cell walls. Some bacteria have a capsule covering outside the cell wall.
● a cell membrane, which regulates the mol-ecules that can enter and leave the cell. all cells have cell membranes, although the chemical composition of archaea mem-branes is different from that of bacteria and of all other cells.
● cytoplasm, the liquid portion in which the molecules are dissolved.
● ribosomes, which make proteins. Bacte-ria have smaller ribosomes than those of eukaryotic cells.
● a circular molecule of Dna. Bacterial Dna, in contrast to the Dna in eukaryotic chro-mosomes, is very effi ciently used for genetic storage. that is, there is very little noncod-ing Dna. in some cases, bacterial genes are so compressed that they overlap. Bacteria also have small fragments of Dna called plasmids that can move from one bacterium to another, even of a different species. Bacte-rial Dna has sometimes been incorporated into eukaryotic chromosomes, a process called horizontal gene transfer. Few bac-teria produce cholesterol; the ones that do may have obtained the gene from eukaryotic cells. the Dna can have as few as 160,000 base pairs (as in some degenerate bacteria that live inside eukaryotic cells) to as many as 12 million base pairs. Some bacteria, such as Borrelia burgdorferi, the germ that causes lyme disease, have a linear rather than circular Dna molecule.
Many bacteria also have fl agella (singular fl agel-lum), a whiplike structure that can move and propel the bacteria through a liquid medium. it is propelled by a protein-based rotor mechanism. the bacterial fl agellum is different from the fl agella and cilia of eukaryotic cells.
the shapes of bacteria are relatively simple. Small, spherical bacteria are cocci (singular coccus), rod-shaped bacteria are bacilli (singular bacillus), and spiral bacteria are spirilla (singular spirillum). Spirochetes are tightly coiled around a central axis.
B
xviii+598_Biodiversity_F.indd 40 11/17/11 1:12 PM
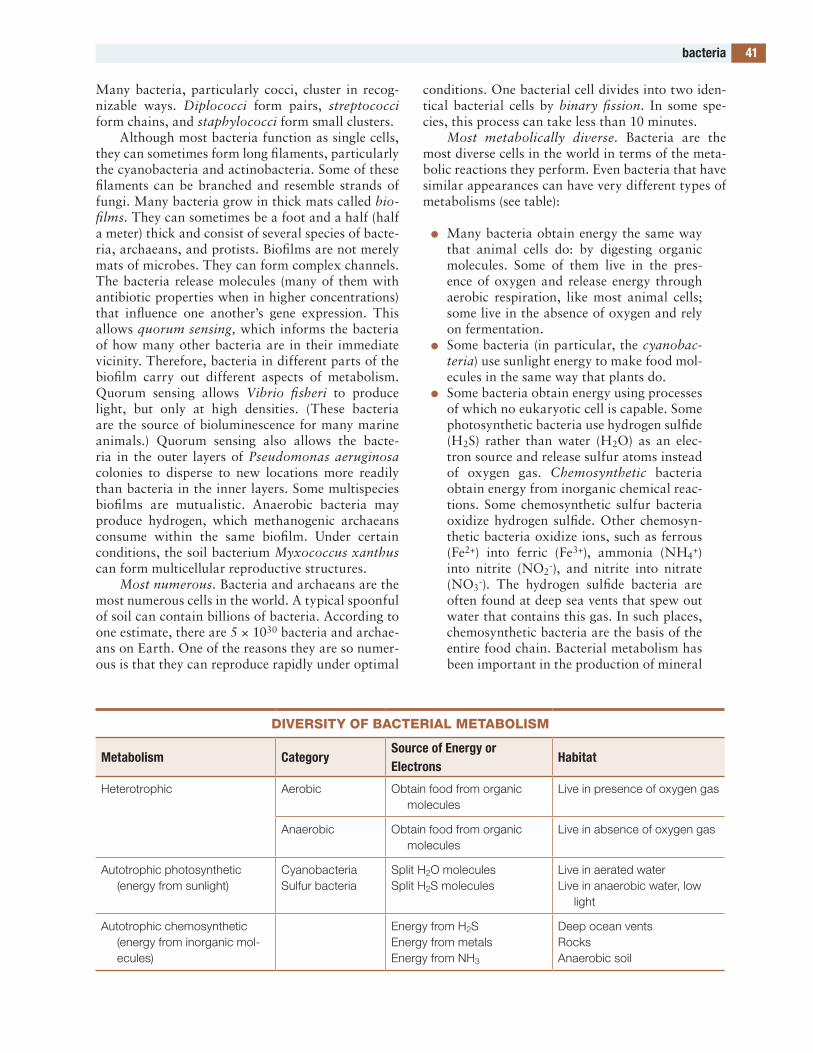
bacteria 41
Many bacteria, particularly cocci, cluster in recog-nizable ways. Diplococci form pairs, streptococci form chains, and staphylococci form small clusters.
although most bacteria function as single cells, they can sometimes form long filaments, particularly the cyanobacteria and actinobacteria. Some of these filaments can be branched and resemble strands of fungi. Many bacteria grow in thick mats called bio-films. they can sometimes be a foot and a half (half a meter) thick and consist of several species of bacte-ria, archaeans, and protists. Biofilms are not merely mats of microbes. they can form complex channels. the bacteria release molecules (many of them with antibiotic properties when in higher concentrations) that influence one another’s gene expression. this allows quorum sensing, which informs the bacteria of how many other bacteria are in their immediate vicinity. therefore, bacteria in different parts of the biofilm carry out different aspects of metabolism. Quorum sensing allows Vibrio fisheri to produce light, but only at high densities. (these bacteria are the source of bioluminescence for many marine animals.) Quorum sensing also allows the bacte-ria in the outer layers of Pseudomonas aeruginosa colonies to disperse to new locations more readily than bacteria in the inner layers. Some multispecies biofilms are mutualistic. anaerobic bacteria may produce hydrogen, which methanogenic archaeans consume within the same biofilm. Under certain conditions, the soil bacterium Myxococcus xanthus can form multicellular reproductive structures.
Most numerous. Bacteria and archaeans are the most numerous cells in the world. a typical spoonful of soil can contain billions of bacteria. according to one estimate, there are 5 × 1030 bacteria and archae-ans on earth. One of the reasons they are so numer-ous is that they can reproduce rapidly under optimal
conditions. One bacterial cell divides into two iden-tical bacterial cells by binary fission. in some spe-cies, this process can take less than 10 minutes.
Most metabolically diverse. Bacteria are the most diverse cells in the world in terms of the meta-bolic reactions they perform. even bacteria that have similar appearances can have very different types of metabolisms (see table):
● Many bacteria obtain energy the same way that animal cells do: by digesting organic molecules. Some of them live in the pres-ence of oxygen and release energy through aerobic respiration, like most animal cells; some live in the absence of oxygen and rely on fermentation.
● Some bacteria (in particular, the cyanobac-teria) use sunlight energy to make food mol-ecules in the same way that plants do.
● Some bacteria obtain energy using processes of which no eukaryotic cell is capable. Some photosynthetic bacteria use hydrogen sulfide (H2S) rather than water (H2O) as an elec-tron source and release sulfur atoms instead of oxygen gas. Chemosynthetic bacteria obtain energy from inorganic chemical reac-tions. Some chemosynthetic sulfur bacteria oxidize hydrogen sulfide. Other chemosyn-thetic bacteria oxidize ions, such as ferrous (Fe2+) into ferric (Fe3+), ammonia (nH4
+) into nitrite (nO2
-), and nitrite into nitrate (nO3
-). the hydrogen sulfide bacteria are often found at deep sea vents that spew out water that contains this gas. in such places, chemosynthetic bacteria are the basis of the entire food chain. Bacterial metabolism has been important in the production of mineral
diversity of baCterial metabolism
metabolism CategorySource of Energy or Electrons
habitat
Heterotrophic Aerobic Obtain food from organic molecules
Live in presence of oxygen gas
Anaerobic Obtain food from organic molecules
Live in absence of oxygen gas
Autotrophic photosynthetic (energy from sunlight)
CyanobacteriaSulfur bacteria
Split H2O moleculesSplit H2S molecules
Live in aerated waterLive in anaerobic water, low
light
Autotrophic chemosynthetic (energy from inorganic mol-ecules)
Energy from H2SEnergy from metalsEnergy from NH3
Deep ocean ventsRocksAnaerobic soil
xviii+598_Biodiversity_F.indd 41 11/17/11 1:12 PM

first entry42 bacteria 42
deposits. although bacteria do not metabo-lize gold, they filter it from seawater and produce gold deposits.
Bacteria can carry out such a wide range of chemical activities that they are used for many eco-nomically and environmentally important processes. Many bacteria play essential roles in food process-ing, especially of dairy products such as butter, cheese, and yogurt. Bacteria, especially those that have been genetically engineered, produce important industrial chemicals. Many pharmaceutical chemi-cals are most effective if only one mirror-image form of the molecule is produced. it is difficult for inor-ganic industrial processes to produce just one mirror image, but bacterial enzymes do it easily and consis-tently. there are bacteria that can break down crude oil; these bacteria have already proven useful in helping to clean up oil spills. there are bacteria that can break down almost any toxic compound, and it is just a matter of finding them. Searching for tech-nologically useful bacteria is an important part of bioprospecting, which is the search for wild species of organisms that do or produce something useful in the human economy. it is especially in bioprospect-ing that the importance of bacterial biodiversity can be seen.
Oldest. Bacteria and archaeans were the first life forms on earth. they probably existed 3.5 billion years ago, and it was nearly 2 billion years later that the first eukaryotic cells evolved. the earliest cell fossils, such as those found in the 3.5 billion-year-old apex chert, are of types that, if they lived today, would be classified as bacteria. throughout the archaean era, the entire world was bacterial. throughout the Proterozoic, the world was mostly microbial, shared between prokaryotes and eukaryotes. Only in the last 540 million years (the last 14 percent of earth history since the begin-ning of the archaean era) have multicellular organ-isms been numerous. the evolutionary biologist Stephen Jay Gould points out that in every age of earth’s history, bacteria have been the modal type of organism.
Bacteria have a bad reputation. Many people look upon them as agents of disease. there are hun-dreds of examples of diseases caused by bacteria, particularly in animals, but for every kind of bac-teria that causes a disease, there are many that are harmless or beneficial to humans.
Bacteria are essential to the survival of all organisms and to the function of all ecosystems on earth. according to the ecologist Paul Falkowski, bacteria and other microbes are the foundation of all metabolism on earth. all aerobic respiration depends on mitochondria, which are the degener-
ate evolutionary descendants of proteobacteria, and photosynthesis occurs in chloroplasts, which are the descendants of cyanobacteria. the bacterial ances-tors of chloroplasts and bacteria took up residence inside ancestral eukaryotic cells, a process called symbiogenesis, but even ignoring the symbiogenetic descendants of bacteria in eukaryotic cells, bacteria perform essential functions, such as the following.
● Bacteria, along with fungi, are the principal decomposers. Without them, carbon would not become available for photosynthesis and the food chain. actinobacteria are decom-posers that produce the “smell of rain,” the refreshing scent that comes from dry soil when it first becomes wet.
● Bacteria perform some unique chemical reactions that are important and perhaps necessary for the ecosystems of earth. in particular, some bacteria carry out nitro-gen fixation, by which they transform atmospheric nitrogen gas (n2) into ammo-nia (nH3), which can then be oxidized into other forms, such as nitrite (nO2
-) and nitrate (nO3
-). ammonia, nitrite, and nitrate are essential soil nutrients upon which the entire food chain depends. Many nitrogen-fixing bacteria live in the root nod-ules of plants, such as the Rhizobium bac-teria in leguminous plants and the Frankia bacteria in the roots of alder trees. they obtain n2 from the air that penetrates into the soil. another unique chemical reaction carried out by some bacteria is denitrifica-tion, which transforms nitrogen-containing molecules back into n2.
● there are many independently living photo-synthetic bacteria. Bacteria exhibit several different forms of photosynthesis. cyano-bacteria (previously known as blue-green algae) have the same kind of photosynthesis as the chloroplasts that evolved from them. a large amount of energy that enters aquatic food chains does so through cyanobacterial photosynthesis. Prochlorococcus, a genus of cyanobacteria discovered in 1988, is found in oceans throughout the world and may be the most abundant kind of cell on earth.
● Bacteria live on and in the bodies of every organism, including humans. a human body consists of about 70 trillion human cells and about 700 trillion bacterial cells. that is, only one 10th of a person’s cells are human. Bacteria live all over the skin and consume dead skin cells and oils. they are particularly abundant in warm, moist areas.
xviii+598_Biodiversity_F.indd 42 11/17/11 1:12 PM

bacteria 43
they also live in the mouth and digestive tract, particularly the intestines, which have about 15 billion bacteria per cubic inch (1 billion per cubic centimeter). Perhaps the best known is Escherichia coli, or E. coli, which is one of the most common bacteria of research and teaching laboratories. Many of them are harmless and cause no more inconvenience than odors or gas. Other bacteria of the human body are mutual-istic. each part of the human body has a unique community of bacteria. the bacte-ria that live on the inside of the elbow, for example, are different from those that live on the outside. Harmless bacteria on the skin and in the intestines can depress the growth of dangerous bacteria that might otherwise grow there. Some bacteria that live in human intestines produce vitamins. Bacteria of the genus Lactobacillus are particularly beneficial in human intestines. Digestive bacteria in ruminant mammals break down otherwise indigestible cellulose, which is an abundant component of plant material, allowing these mammals to obtain much more food value from the plant mate-rial they have eaten. Bacteria, therefore, are major components not only of the external environments but also the internal environ-ments of organisms.
the total number of bacterial species is impossi-ble to calculate. Some studies suggest that a spoonful of soil contains thousands of kinds of bacteria, while a different spoonful of soil from a different loca-tion contains thousands of other kinds of bacteria. Because they have simple structures, they can be dis-tinguished mainly by their metabolisms, but scien-tists have to know what that metabolism is in order to grow them and study them. there are many kinds of bacteria whose presence researchers overlook because they cannot grow them properly. Recently, biotechnology has provided new ways of identifying bacteria on the basis of their Dna without having to grow them in a laboratory. these methods have revealed that a quart (liter) of seawater can have 12,000 species of bacteria. Many bacteria have been identified only, and scientists do not understand how they live or what role they may play in ecosystems. For example, Nitrospira moscoviensis was discov-ered in 1995 in a corroded heating pipe in Moscow; it is genetically distinctive enough to be classified into its own division, but there is little else to distin-guish this division from others. Bacteria have been classified, on the basis of Dna and metabolism, into 17 divisions (see table on page 44).
Many scientists speculate that there is more life under earth than on top of it and that this life con-sists of bacteria and archaeans. they obtain energy from minerals and live in the wet cracks in rocks up to two miles (almost 4 km) below the surface, metabolizing very slowly (dividing perhaps once a century). Some scientists estimate that the bacte-ria of these “subsurface lithoautotrophic microbial ecosystems” (abbreviated SliMe) would cover the surface of the earth to a depth of five feet (almost 2 m) if they were concentrated.
Many kinds of bacteria can tolerate extreme environmental conditions. Some bacilli, such as Bacillus and Clostridium, form endospores in which the Dna and a small amount of cytoplasm are pro-tected inside an impervious coat. endospores have no detectable metabolism and can survive ultravi-olet and gamma irradiation, chemicals, pressure, heat, and desiccation. Boiling water is not enough to destroy endospores; it is principally to destroy the endospores of toxic bacteria such as botulism (Clostridium botulinum) that high-pressure canning methods must be used to store moist food. Some bacteria can not only survive but also grow under extreme conditions: high temperatures, strong acid, or concentrated salt, all conditions that would kill eukaryotic cells. For example, Thiobacillus concre-tivorans grows in sulfuric acid at concentrations that would dissolve metal. Acidithiobacillus ferooxidans makes food by using energy from iron and sul-fur while living in sulfuric acid. Other bacteria can tolerate extremely basic conditions. Deinococcus radiodurans can live in the water tanks of nuclear reactors. it resists nuclear radiation because it has a unique ability to reform its Dna after damage by radiation. Streptococcus mitis was found inside the lens of a camera that was left on the Moon for two years. Bacteria live even in deep ocean vents so hot that the temperature probes start to melt. Thermus aquaticus (shortened to taq), which lives in hot springs, is the source of an enzyme that copies Dna at high temperatures and is therefore very use-ful in genetic engineering. Some bacteria have been reported to survive in suspended animation for up to 250 million years.
the ability of bacteria to exchange fragments of Dna confers upon them the ability to evolve rapidly. Rather than having to wait for a new muta-tion to arise, a bacterium might be lucky enough to obtain a new gene from another bacterium, even of a different species. this is the principal reason that pathogenic bacteria can evolve resistance to antibiot-ics so rapidly. For example, the green sulfur bacteria (which use hydrogen sulfide as an electron source in photosynthesis) have only one set of reactions for absorbing and using light energy. the cyanobacteria,
xviii+598_Biodiversity_F.indd 43 11/17/11 1:12 PM
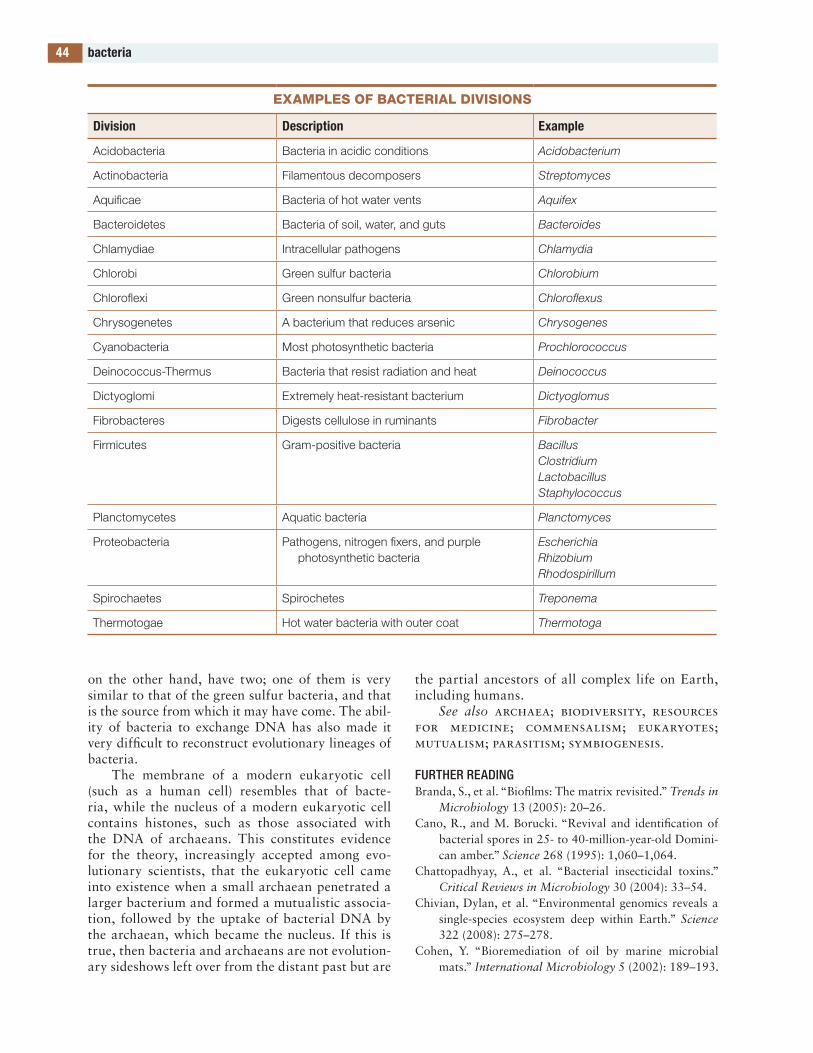
first entry44 bacteria 44
on the other hand, have two; one of them is very similar to that of the green sulfur bacteria, and that is the source from which it may have come. the abil-ity of bacteria to exchange Dna has also made it very difficult to reconstruct evolutionary lineages of bacteria.
the membrane of a modern eukaryotic cell (such as a human cell) resembles that of bacte-ria, while the nucleus of a modern eukaryotic cell contains histones, such as those associated with the Dna of archaeans. this constitutes evidence for the theory, increasingly accepted among evo-lutionary scientists, that the eukaryotic cell came into existence when a small archaean penetrated a larger bacterium and formed a mutualistic associa-tion, followed by the uptake of bacterial Dna by the archaean, which became the nucleus. if this is true, then bacteria and archaeans are not evolution-ary sideshows left over from the distant past but are
the partial ancestors of all complex life on earth, including humans.
See also archaea; biodiversity, resources for medicine; commensalism; eukaryotes; mutualism; parasitism; symbiogenesis.
Further readingBranda, S., et al. “Biofilms: the matrix revisited.” Trends in
Microbiology 13 (2005): 20–26.cano, R., and M. Borucki. “Revival and identification of
bacterial spores in 25- to 40-million-year-old Domini-can amber.” Science 268 (1995): 1,060–1,064.
chattopadhyay, a., et al. “Bacterial insecticidal toxins.” Critical Reviews in Microbiology 30 (2004): 33–54.
chivian, Dylan, et al. “environmental genomics reveals a single-species ecosystem deep within earth.” Science 322 (2008): 275–278.
cohen, y. “Bioremediation of oil by marine microbial mats.” International Microbiology 5 (2002): 189–193.
examples of baCterial divisions
division description Example
Acidobacteria Bacteria in acidic conditions Acidobacterium
Actinobacteria Filamentous decomposers Streptomyces
Aquificae Bacteria of hot water vents Aquifex
Bacteroidetes Bacteria of soil, water, and guts Bacteroides
Chlamydiae Intracellular pathogens Chlamydia
Chlorobi Green sulfur bacteria Chlorobium
Chloroflexi Green nonsulfur bacteria Chloroflexus
Chrysogenetes A bacterium that reduces arsenic Chrysogenes
Cyanobacteria Most photosynthetic bacteria Prochlorococcus
Deinococcus-Thermus Bacteria that resist radiation and heat Deinococcus
Dictyoglomi Extremely heat-resistant bacterium Dictyoglomus
Fibrobacteres Digests cellulose in ruminants Fibrobacter
Firmicutes Gram-positive bacteria BacillusClostridiumLactobacillusStaphylococcus
Planctomycetes Aquatic bacteria Planctomyces
Proteobacteria Pathogens, nitrogen fixers, and purple photosynthetic bacteria
EscherichiaRhizobiumRhodospirillum
Spirochaetes Spirochetes Treponema
Thermotogae Hot water bacteria with outer coat Thermotoga
xviii+598_Biodiversity_F.indd 44 11/17/11 1:12 PM

Bates, henry walter 45
curtis, t., et al. “estimating prokaryotic diversity and its limits.” Proceedings of the National Academy of Sci-ences USA 99 (2002): 10,494–10,499.
DasSarma, Shiladitya. “extreme microbes.” American Sci-entist 95 (2007): 224–231.
Delong, e., and n. Pace. “environmental diversity of bacteria and archaea.” Systematic Biology 50 (2001): 470–478.
Dexter-Dyer, Betsey. A Field Guide to Bacteria. ithaca, n.y.: cornell University Press, 2003.
DiGiulio, M. “the universal ancestor and the ancestor of bacteria were hyperthermophiles.” Journal of Molecu-lar Evolution 57 (2003): 721–730.
eagon, R. “Pseudomonas natriegens, a marine bacterium with a generation time of less than 10 minutes.” Jour-nal of Bacteriology 83 (1962): 736–737.
Falkowski, Paul G., et al. “the microbial engines that drive earth’s biogeochemical cycles.” Science 320 (2008): 1,034–1,039.
Fraser, christophe, et al. “the bacterial species challenge: Making sense of genetic and ecological diversity.” Sci-ence 323 (2009): 741–746.
Gorbach, S. “lactic acid bacteria and human health.” Annals of Medicine 22 (1990): 37–41.
Guerrero, R., et al. “Predatory prokaryotes: Predation and primary consumption evolved in bacteria.” Proceed-ings of the National Academy of Sciences USA 83 (1986): 2,138–2,142.
Hinnebusch, J., and K. tilly. “linear plasmids and chro-mosomes in bacteria.” Molecular Microbiology 10 (1993): 917–922.
Horner-Devine, M. c., et al. “an ecological perspective on bacterial biodiversity.” Proceedings of the Royal Society of London B (Biological Sciences) 271 (2004): 113–122.
Johnson, Zackary i., et al. “niche partitioning among Prochlorococcus ecotypes along ocean-scale environ-mental gradients.” Science 311 (2006): 1,737–1,740. Summary by Pennisi, elizabeth, “How a marine bac-terium adapts to multiple environments.” Science 311 (2006): 1,697.
ley, R., et al. “Obesity alters gut microbial ecology.” Pro-ceedings of the National Academy of Sciences USA 102 (2005): 11,070–11,075.
Mehta, Mausmi P., and John a. Baross. “nitrogen fixation at 92 c by a hydrothermal vent archaeon.” Science 314 (2006): 1,783–1,786.
nakabachi, a., et al. “the 160-kilobase genome of the bac-terial endosymbiont Carsonella.” Science 314 (2006): 267.
nicholson, W., et al. “Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environ-ments.” Microbiology and Molecular Biology Review 64 (2000): 548–572.
———, et al. “the solar UV environment and bacterial spore UV resistance: considerations for earth-to-Mars
transport by natural processes and human spaceflight.” Mutation and Resistance 571 (2005): 249–264.
Reith, Frank, et al. “Biomineralization of gold: Biofilms on bacterioform gold.” Science 313 (2006): 233–236.
Schulz, H., and B. Jorgensen. “Big bacteria.” Annual Review of Microbiology 55 (2001): 105–137.
Stevens t. O., and J. P. McKinley. “lithoautotrophic micro-bial ecosystems in deep basalt aquifers.” Science 270 (1995): 450–454.
Velicer, G. J., and K. l. Stredwick. “experimental social evolution with Myxococcus xanthus.” Antonie Van Leeuwenhoek 81 (2002): 155–164.
Vreeland, R., et al. “isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal.” Nature 407 (2000): 897–900.
Whitman, W., et al. “Prokaryotes: the unseen majority.” Proceedings of the National Academy of Sciences USA 95 (1998): 6,578–6,583.
Zimmer, carl. Microcosm: E. coli and the New Science of Life. new york: Pantheon, 2009.
Zoetendal, e., et al. “a microbial world within us.” Molec-ular Microbiology 59 (2006): 1,639–1,650.
bates, henry Walter (1825–1892) British Naturalist Born on February 8, 1825, Henry Wal-ter Bates was a British naturalist who specialized in collecting and studying insects. His insights were very important to the modern understanding of bio-diversity and coevolution.
Bates befriended another British naturalist, alfred Russel Wallace. in 1848, Bates and Wal-lace traveled together to the amazon rain forest of South america, where both of them extensively col-lected insects and other animals. Wallace returned to england and then traveled to indonesia, but Bates remained in the amazon rain forest for 14 years.
While studying amazonian insects, Bates dis-covered that certain nonpoisonous species of heli-conid butterflies had wing coloration patterns that resembled those of poisonous heliconid butterfly species. Bates considered this to be mimicry, which afforded protection to the nonpoisonous butterflies without the cost of producing the poison. Since Bates’s initial discovery, numerous other examples of mimicry have been found. this type of mim-icry is now called Batesian mimicry, in contrast to Mullerian mimicry, in which numerous prey species converge upon a common set of warning coloration patterns. Bates’s discovery of mimicry contributed significantly to charles Darwin’s work on evolution. Darwin cited Bates’s discovery in later editions of The Origin of Species. Bates and Darwin both main-tained that such mimicry could not have resulted from the use and disuse of the structures involved and must have resulted from natural selection.
xviii+598_Biodiversity_F.indd 45 11/17/11 1:12 PM

first entry46 Berenbaum, may46
Bates returned to england in 1862. He pub-lished an account of his work in the rain forest, The Naturalist on the River Amazons, in 1863. Bates continued scientific research and was elected a fel-low of the Royal Society in 1881. He died on Febru-ary 16, 1892.
See also biodiversity, concept of; cam-ouflage and mimicry; coevolution; Darwin, charles; natural selection; Wallace, alfred Russel.
Further readingBeddall, Barbara G. Wallace and Bates in the Tropics: An
Introduction to the Theory of Natural Selection, Based on the Writings of Alfred Russel Wallace and Henry Walter Bates. london: Macmillan, 1969.
berenbaum, may See insects.
biodiversity, concept of Biodiversity is the diversity of life. it is frequently defined as the num-ber of species in a defined location (a forest, a coun-try, or the earth), but the concept of biodiversity incorporates other levels of diversity as well, includ-ing the following.
Genetic diversity within populations. the pres-ervation of biodiversity cannot be considered suc-cessful if a species is merely saved from extinction, for example as a single pair in a zoo. natural selec-tion works upon genetic diversity; if a species is to persist, it must be able to evolve, and a lack of genetic diversity prevents evolutionary adjustments. Moreover, harmful mutations show up more often in small populations than in larger ones. a species usually does not begin to lose genetic diversity until it has been driven close to extinction, but from that point forward, the loss of genetic diversity pushes the species ever closer to extinction, despite the best efforts of conservationists.
Genetic diversity among populations. Different populations of a species have unique sets of alleles, either because natural selection has operated differ-ently in the separate populations or due to genetic drift. if one population becomes extinct, this is con-sidered a loss of biodiversity, even if the species survives. all wolves are in the species Canis lupus. However, there are different populations of wolves (different enough to be considered subspecies). the visible differences in coat color among these subspe-cies is only the most visible characteristic that makes them distinct.
Diversity of ecological functions. it is mislead-ing to think that the loss of one species from an eco-logical community with a hundred species is merely
a 1 percent loss of biodiversity in any meaningful sense, because different species can rarely substi-tute for one another. For example, if one member of a symbiotic relationship becomes extinct, the other may likewise become extinct. Of particular importance are keystone species, the disappearance of which may produce profound changes in an eco-logical community. One recently discovered example is that large herbivorous mammals were keystone species in the tundra and boreal forests that covered much of what is now the United States during the most recent ice age. the largest of these mammals became extinct at the end of that ice age, an event known as the Pleistocene extinction. Various reasons have been proposed for this extinction event, includ-ing changes in climate that altered the vegetation upon which these animals depended. a more careful study of the sequence of events during the Pleis-tocene extinction revealed that the populations of large mammals began to decline rapidly before there were changes in vegetation. that is, the vegetation changed because the mammals became rare, not the other way around. the huge amount of grazing for which these animals were responsible had kept for-ests from spreading and encouraged the growth of grasslands. With the extinction of these animals, the forests spread, and the accumulation of dead wood enhanced the frequency of fires. the extinction of mammoths and mastodons from a Pleistocene for-est edge community is small when calculated on the basis of the number of species, but its effect was considerable.
Diversity of higher taxonomic levels. Most ecol-ogists consider a grassland plant community that consists of 100 species, all of which are grasses, to be less diverse than a grassland plant commu-nity that consists of 100 species, some of which are grasses and some of which are dicot wildflowers. the greater the number of genera, families, orders, classes, phyla, kingdoms, and domains, the more diverse an ecological community is considered to be. the atlantic horseshoe crab (Limulus polyphemus) is but one species of many in the atlantic Ocean, but its loss would be noticed more than the loss of just one of the many species of dinoflagellates in the same waters because it is (along with Tachypleus gigas and Carcinoscorpus rotundicaudatus) one of the last survivors of its evolutionary lineage, the xiphosuran arthropods.
nobody knows how many species there are. it is certain that many species remain unidentified, but it is impossible even to make a good estimate of the percentage of species that have not yet been identified. nobody even knows how many species have been identified, as the accounts have been pub-lished in thousands of different scientific journals;
xviii+598_Biodiversity_F.indd 46 11/17/11 1:12 PM

biodiversity, patterns of 47
nobody looks at all of them. all the scientists who study biodiversity are specialists in some relatively narrow group of organisms. it has proven difficult even to establish an internet database of known spe-cies. One such ongoing effort is the encyclopedia of life. the numbers in the table, taken from a recent book, are minimum estimates and not accepted by all experts.
Most species on earth are microbes. there may be 5 × 1030 individual microbes on earth—more than the number of stars in the universe. nobody knows how many species are represented in this total. there are at least 700 species of bacteria and archaeans in the human mouth. Scientists most read-ily identify species on the basis of their visible char-acteristics, of which microbes have few. the table does not even attempt to list the number of bacterial and archaean species. the few prokaryotic species that can be grown in a laboratory cannot readily be distinguished by physical or metabolic characteris-tics. analysis of Dna is necessary for the identifi-cation of microbe species and is increasingly being used to identify plants and animals as well.
See also biodiversity, quantification of; communities, ecological; Dna studies of biodiversity; extinction; founder effect and genetic drift; keystone species; mutualism; natural selection; Pleistocene extinction; population genetics.
Further readingchivian, eric, and aaron Bernstein. Sustaining Life: How
Human Health Depends on Biodiversity. new york: Oxford University Press, 2008.
encyclopedia of life. “encyclopedia of life.” available online. URl: http://www.eol.org. accessed June 25, 2010.
Gill, Jacquelyn l., et al. “Pleistocene megafaunal collapse, novel plant communities, and enhanced fire regimes in north america.” Science 326 (2009): 1,100–1,103. Summarized by Johnson, christopher. “Megafaunal decline and fall.” Science 326 (2009): 1,072–1,073.
Wilson, edward O. The Diversity of Life. cambridge, Mass.: Harvard University Press, 1992.
biodiversity, patterns of Some parts of earth have more species than do others. in par-ticular, tropical latitudes (and to a certain extent low elevations at all latitudes) are home to more species than are temperate or polar latitudes. this pattern holds true when all species are considered and also within ecological or evolutionary groups of species, such as trees, mammals, or insects. there are a few exceptions: Parasites, aquatic plants, and aquatic mammals, such as seals, have no greater diversity in the tropics than away from them.
ever since alexander von Humboldt in the early 19th century, scientists have attempted to explain why some parts of earth have more species than do others—and have yet to answer this deceptively sim-ple question. Von Humboldt suggested that tropical forests had more plant species because the only plant or animal species that can live outside the tropics are those that can avoid or resist damage from cold temperatures. a related observation is that deserts have fewer plant or animal species because they must have adaptations that allow them to avoid drought or to resist it by means of enhanced water absorption or reduced water loss. For many species, life appears to be easier in tropical climates.
nevertheless, why have more species not evolved adaptations to resist cold or dry conditions? While the adaptations that allow organisms to live in cold or dry conditions may be complex and costly, there is a benefit also: the species that are adapted to cold or dry conditions escape from competition with organisms not so adapted. these costly adaptations confer a survival advantage. it is generally true that, for most species, the edge of the distribution range toward higher resource concentrations (e.g., the equatorial edge) is determined by competition with other species, while the edge of the range toward lower resource concentrations (e.g., the poleward edge) is determined by tolerance of the climatic con-ditions. that is, most temperate animals and plants
minimum estimates of speCies diversity in eukaryotes
Taxonomic GroupEstimated Number of Species
Protists (including algae) 80,000
Plants 270,000
Fungi 70,000
Nematodes 15,000
Mollusks 70,000
Vertebrates 45,000
Arthropods: 855,000
Crustaceans 40,000
Arachnids 75,000
Insects 720,000
Other animals 95,000
Total About 1.5 million
Note: From Chivian and Bernstein
xviii+598_Biodiversity_F.indd 47 11/17/11 1:12 PM

first entry48 biodiversity, patterns of 48
could survive in the tropics if they were sheltered from competition. even so, why should it be easier for a species to compete with, or be eaten or parasit-ized by, other species in a jungle than to live in harsh conditions? it is even difficult to define what “harsh conditions” are—archaebacteria live in extremely hot, salty, or acidic conditions and cannot even sur-vive in what most species would regard as benign conditions.
More than 30 hypotheses have been proposed to explain global biodiversity patterns. Some of them are here examined, with a focus on terrestrial species.
arE BIodIvErSITy paTTErNS aN IlluSIoN?Null models describe biodiversity patterns that would result even if species were randomly dis-tributed on earth’s dry land surface. One of them is called the mid-domain effect. it claims that if the ranges of species are distributed randomly, the ranges of most of them would overlap in the middle of a continent or island. the middle of a continent (the mid-domain) would therefore have the greatest species richness. One reason that the tropics have so many species may be that they are the mid-domain of large continents.
there are several other reasons why one might expect more species to evolve in the middle of a con-tinent than near the edge, regardless of the climatic conditions on the continent. large areas allow more opportunities for evolution in ways such as the fol-lowing. they allow larger populations, thus enhanc-ing the chances for evolution as well as reducing the chance of extinction; they contain more microhabi-tat diversity; and they may have fewer restrictions to gene flow between populations. in almost every case, a larger area contains more species. this is known as the species-area relationship. However, the relation-ship is not constant: Doubling a small area adds more species than doubling a large area. this occurs because both within-habitat and between-habitat diversity contribute to species richness in small areas, while only between-habitat diversity contrib-utes to species richness in large areas.
at first glance, the mid-domain effect cannot be true. all one has to do is look at the vast expanses of the northern continents, where relatively few spe-cies live in the tundra and northern forests. in north america, the tropical zone has relatively little land-mass and yet has the most species.
there is an important sense, however, in which the mid-domain effect may help explain tropical diversity. all the tropical regions within a conti-nental region are in one place—the equator. the temperate regions, however, are divided between the northern and Southern Hemispheres. accord-
ing to this null model, temperate forests might have as many species as do tropical forests if all of them were in one place, but this explanation also does not go very far. Most of the temperate land surface of earth is already in one place: the northern Hemi-sphere. the Southern Hemisphere is mostly ocean.
large-scale measurements of biodiversity may disguise underlying biodiversity patterns. this, how-ever, does not nullify the reality of the patterns. the greater number of mammal species in the tropics is due mainly to the greater number of tropical bat spe-cies; other mammalian orders (such as dogs, cats, and ungulates) may not have a greater species richness in the tropics. the greater diversity of the bats may have resulted from evolutionary events specific to them rather than from a general process that applies to all mammals. this explanation is not convincing, because there is a huge number of examples of evo-lutionary lineages that have diversified in the tropics and far fewer that have diversified in the temperate or polar regions; bats are not an isolated example.
the null models indicate that, in the absence of other factors, there would still be more species in some parts of the world than in others, but they do not appear to explain the patterns that actually exist.
proCESSES ThaT may ExplaIN BIodIvErSITy paTTErNS
Since the null models are not entirely adequate, what are some processes that could explain why some parts of earth, particularly the tropics, have more species than do other parts?
Climate. On a worldwide scale, tropical rain forests have greater species diversity than any other ecological community. Some researchers attribute this to the great amount of energy (in the forms of temperature and of light) and water in rain forests. Bright sunlight and abundant moisture allow rain forests to have the world’s highest levels of produc-tivity (growth of organisms), starting with photo-synthesis and continuing up the food chain. if there is more energy flowing through the food chain, there can be more trophic levels and thus more species. the ecologist Bradford Hawkins and his associates analyzed 85 data sets of species; climate was the strongest predictor of species richness in 83 of the 85 data sets.
neither energy nor water can be considered alone; ecologists usually combine them into a vari-able known as potential evapotranspiration, which is the amount of water that could be removed from an area of land by evaporation and by transpira-tion of plants. Both low temperatures and low rain-fall contribute to low evapotranspiration; both high temperatures and high rainfall contribute to high
xviii+598_Biodiversity_F.indd 48 11/17/11 1:12 PM

biodiversity, patterns of 49
evapotranspiration. Populations can grow faster, and fewer individuals die, in warmer, wetter condi-tions than in colder or drier conditions. Warmth and moisture enhance the growth of plants and animals, which promotes species diversity.
On a smaller scale, however, the highest pro-ductivity is not always associated with the greatest number of species, especially if the high productivity results from abundant soil nutrients. Within tropical and temperate forests and grasslands, species rich-ness is highest at intermediate levels of productiv-ity. When productivity is very high, competition is strong, and rare species may be driven to extinction. in grasslands, for example, adding nutrients to the soil causes the dominant species to grow and cast shade on the less common species, often eliminating them. the ecologist yann Hautier and his associates counteracted this process by experimentally provid-ing supplemental light underneath the grass canopy along with extra nutrients in the soil.
Similar patterns are found in aquatic habi-tats. On a worldwide scale, the most productive and diverse marine habitats are those that receive the most sunlight—that is, shallow tropical seas and coral reefs. continental shelves also have more nutrients than do the open oceans, and this further stimulates the productivity of the food web. How-ever, as on land, an overabundance of nutrients (for example, with eutrophication) results in a few spe-cies dominating the community.
Rate of evolution. evolution itself may run faster and produce more species under warm, wet conditions than under cool, dry conditions. One reason is that plants and cold-blooded animals have more generations per year in tropical forests, not only due to warmer temperatures but because there is no winter in these forests. another reason is that mutations can occur more often under conditions of higher productivity. Both of these processes may allow natural selection to occur more rapidly in the tropics than in cooler or drier regions. this is appar-ently true not only of cold-blooded species but of warm-blooded species as well. the ecologist Shane Wright and his associates demonstrated that tropical mammals have 50 percent faster mutations than do related temperate mammals.
Once biodiversity has begun to increase in a region, it feeds upon itself. as the ecologist andrew Forbes wrote, “a major cause for biodiversity may be biodiversity itself. as new species form, they may create . . . a chain reaction of speciation events across trophic levels.” this process can continue all year long in a tropical forest, while in a temperate decidu-ous forest, it slows down or stops during the winter.
Time for evolution. another reason the tropics contain so many species is that they have persisted
for a long time. Particularly since the beginning of the Pleistocene epoch, the temperate zones in northern continents have experienced glaciations that have disrupted entire ecological communities. Glaciations also indirectly affected tropical forests as well: Periods of glaciation in the north resulted in drier conditions in the tropical zone. During these dry periods, however, the tropical forests contracted but were not totally displaced. intense evolution has been going on without interruption in some tropical areas for hundreds of millions of years. the west-ern amazon has greater species richness than does the central amazon, and this may be because the western amazon remained forested throughout the Pleistocene droughts that turned central amazonia into a savanna.
even within temperate zones, the regions most affected by glaciation in the past have fewer spe-cies today. For example, compare europe, north america, and china. the temperate forests of china were little disturbed by the ice ages. north american deciduous forests recovered from the most recent ice age relatively quickly, as the north-south mountain ranges did not block the migration of species. euro-pean forests did not recover from the most recent ice age so well, because east-west mountain ranges and the Mediterranean Sea blocked the migration of some species. this may explain why chinese forests have more species than do north american forests, which have more species than do european forests.
Range of tolerance. Species that live in temper-ate, boreal, or arctic zones must tolerate a large (sometimes very large) annual range of temperature conditions. Without going anywhere, members of a temperate species experience conditions that corre-spond to a wide geographical range. in many cases, temperate species have wider geographical ranges, since they are already adapted to a wide range of tem-perature and moisture conditions. tropical species experience relatively constant conditions and may specialize on a small geographical range. Despite the intuitive appeal of this hypothesis, there are many exceptions to it. tree species in the new World trop-ics, for example, may be exceedingly rare in any one location but have a wide geographical range.
Many other factors contribute to species rich-ness, such as a moderate level of disturbance. How-ever, these factors do not differ consistently from one part of the world to another.
why arE ThErE So maNy TropICal SpECIES?any combination of the foregoing processes may explain why tropical zones have more species than do temperate or polar zones, but they seem insuf-ficient to explain the breathtaking species richness of
(continues on page 52)
xviii+598_Biodiversity_F.indd 49 11/17/11 1:12 PM

first entry50 biodiversity, patterns of 50
As explained in many entries in this ency-clopedia, the number of species on Earth is literally incalculable. Scientists have described more than a million species, and many other species have recently or will soon become extinct as a result of human activity before they could or can be documented. There will probably never be a complete answer to the ques-tion of why there are so many species.
Many pre-Darwinian scholars believed that God’s creation consisted of many lev-els, from rock to worm to plant to animal to man to angel, and that God had created something to fill in each of the gaps in the great fabric of being. This idea, known as the scala naturae, or great chain of being, was their explanation of why all levels of complexity existed in the natural world. It also led these scholars to believe in intermediate forms, such as half-human half-apes, even though they did not have any concept of evolution. To answer the question of why there are so many spe-cies by simply saying that God made the scala naturae was not satisfying even to theologians, who wanted to believe in something more than the Creator’s whim.
Some pre-Darwinian scholars at-tempted to explain why some parts of Earth had more species than others, even though they could not explain why there were so many species or where they had come from. Alexander von Humboldt extensively explored the Tropics and ob-served their tremendous species diversity. He speculated that the warm, moist condi-tions of the Tropics allowed more species to survive than did the colder and drier regions of Earth. There were more species in the places where life was easier.
Once Charles Darwin had presented evidence for evolution and explained how it could happen, scientists could inves-tigate the origin of Earth’s astounding biodiversity. After life had begun, new species would evolve as they progres-sively adapted to new climates. Even before Darwin, geologists had known that the earliest life forms all lived in the oceans and that life on land existed only in
more recent geological ages, but this did not seem adequate to explain the sheer number of species, since there were not that many different climatic or geographi-cal habitats to which organisms could adapt. Why were there so many species of beetles in the Tropics, a fact revealed by Darwin’s friends Henry Walter Bates and Alfred Russel Wallace, when they all appeared to be doing more or less the same things in the same places? Why would not the “struggle for existence” wipe many of them out, leaving only the species best adapted?
Darwin expanded his concept of nat-ural selection to include species evolv-ing in response to one another (which scientists today call coevolution), and he explained the process of sexual selection. Coevolution and sexual selection could explain what natural selection in response to climate could not: New species could evolve in response to previous species by parasitizing them, eating them, or help-ing them, and even newer species could then evolve in the same fashion. New spe-cies could evolve that recognized different mating signals or decorations, of which there is a potentially limitless variety. Spe-cies diversity prompted the evolution of greater species diversity. Once biodiver-sity has begun to increase, it feeds upon itself. As ecologist Andrew Forbes wrote, “A major cause for biodiversity may be biodiversity itself. As new species form, they may create . . . a chain reaction of speciation events across trophic levels.”
Furthermore, biodiversity could be propagated up the food chain. If there are more species of plants in one area than another, there can be more species of leaf-eating insects—a pattern confirmed by the ecologist Vojtek Novotny and his associates in the tropical forests.
Still, scientists have not been happy with simply saying there are so many species because they evolved, any more than they would have been happy with the idea of God’s whim. Hence, rather than simply document each new species and speculate about how it might have
come into existence, each species an iso-lated story of evolution, they looked for general patterns.
A 1959 article by the ecologist G. Evelyn Hutchinson (based on his presi-dential address to the American Society of Naturalists) claimed to address directly the question of why there were so many species of animals. Actually, Hutchin-son turned the question around and laid the foundation for the modern scientific understanding of why there are not more species in the world than there are. Among his suggestions were the following.
• Food chains. Hutchinson noted that, at each level of the food chain, the consumers were larger and rarer than the organisms they consumed. Many ecologists today refer to the “ten percent law,” indicating that each level of the food chain contains only about 10 percent as much energy and matter as the level below it. How-ever, 10 percent is just an average figure; Hutchinson used 20 percent and calculated that if the oceans were full of single-celled algae and there were 50 links in the food chain, there would not be enough room in all the oceans to feed even one of those top predators. Natural selection, which operates within physical limits, can-not keep producing predators that eat predators that eat predators.
• Limiting similarity. Hutchinson also noted that two species that are too similar to each other cannot both sur-vive in the same place. He noted that food-gathering structures of related animals living in the same location tended to differ by a factor of 1.3—that is, two species of similar animals could exist in the same place if one of them was larger by a factor of approx-imately 1.3. It was and is far from certain that this pattern and others like it are not simply random.
Hutchinson and many ecologists after him have pointed out ecological pro-
“Why Are There So MAny SpecieS?”
xviii+598_Biodiversity_F.indd 50 11/17/11 1:12 PM

biodiversity, patterns of 51
cesses that could allow species to avoid driving one another to extinction. They include the following:
• Predator switching. Predators seldom specialize entirely on one prey spe-cies. As a result, there is no predator species that will drive its single spe-cies of prey into extinction and then suffer extinction itself.
• Disturbance. Species differ in their life histories—that is, in how long they live and how much they reproduce. Some species grow slowly, live a long time, and produce few offspring; oth-ers grow quickly, have short lives, and produce many offspring. Species form a continuum between these two extremes. The short-lived, fecund species often live in areas recently disturbed by, for example, a forest fire. The fire clears away the dominant plants and makes resources such as light and nutrients available. The quickly growing plant species require the abundant resources made avail-able by the fire. They grow quickly in the disturbed areas, which soon become thick with plants. The quickly growing species can no longer sur-vive, nor can their seedlings grow, in the habitat that they have helped to make crowded. With animals as with plants, species with different life his-tories can live in the same place at the same time by living differently.
• Microhabitat specialization. A habi-tat may appear uniform to a human observer but actually consists of a great variety of small-scale condi-tions. To small organisms, the over-all habitat is actually a collection of microhabitats. While this is undeni-ably true, it has only limited useful-ness in explaining biodiversity. The ecologist Stephen Hubbell found that the trees of Barro Colorado Island in Panama appeared to be randomly dis-tributed with respect to small distur-bances and differences in slope and the direction the slope faced.
• Foraging specialization. The ecologist Robert MacArthur studied the foraging of warblers in trees. He found that dif-ferent species of warblers specialized
on different portions of the trees—some near the top, some lower, some inside the branches, some on the out-side. To birds, a tree is no more of a single habitat than is a whole forest.
These and other processes allow large numbers of species to persist, once evo-lution has produced them—and evolution can produce a lot of species.
One of the most obvious patterns of biodiversity is that it has increased over evolutionary time. Although more than 99 percent of all species that have ever lived are now extinct, more species have evolved than have become extinct. Eco-logical communities are therefore not just nurseries of new species but museums of old ones. It is not uncommon for newly discovered species to be “living fossils” that were presumed to have become extinct millions of years ago. This was the case with the dawn redwood (Metase-quoia glyptostroboides) and the Wollemi pine (Wollemia nobilis).
Part of the evolutionary explanation for why there are so many species may be historical contingency or evolution-ary accident. Today there are a quarter of a million species of flowering plants (angiosperms). Since they came into existence about 120 million years ago, they have coevolved with insect pollina-tors as well as with other animals that disperse their seeds. Since insect diver-sity began to explode about the same time as the origin of flowering plants, it is likely that insects and flowering plants contributed to each other’s diversity. Coevolution is no accident, but the origin of the first flowering plants may have been. For about 290 million years after the first terrestrial life forms had evolved, the oceans had greater biodiversity than did the land. This changed quickly after the origin of the flowering plants, and now terrestrial species outnumber marine species 10 to one.
Still, none of these explanations, sin-gly or in combination, seems to do justice to the prodigious number of species in the world, particularly in tropical forests, where two and a half acres (a hectare) can have 250 species of trees, compared to 20 species in a North American decid-
uous forest of the same size. About 0.2 square miles (half a square kilometer) of tropical rain forest can contain more tree species than all of the temperate forests of Europe, North America, and Asia com-bined. In a Venezuelan rain forest, 978 species of beetles were found on just six trees. The mystery of why there are so many species is not simply that tropical rain forests are warm and wet or that coevolution produces incredible species diversity, but that the biodiversity of the Tropics compared to that of temperate zones seems wildly disproportionate.
See also Bates, Henry Walter; Bio-diversity, concept of; Biodiversity, pat-terns of Biodiversity, quantification of; BiogeograpHy; coevolution; darWin, cHarles; disturBances, ecological; eco-systems; life History, diversity of; living fossils; natural selection; sexual selec-tion; Wallace, alfred russel.
Further readingForbes, Andrew A., et al. “Sequential sym-
patric speciation across trophic levels.” Science 323 (2009): 776–779.
Gilbert, Benjamin, and Martin J. Lechowicz. “Neutrality, niches, and dispersal in a temperate forest understory.” Proceed-ings of the National Academy of Sciences USA 101 (2004): 7,651–7,656.
Hubbell, Stephen P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton, N.J.: Princeton University Press, 2001.
Hutchinson, G. Evelyn. “Homage to Santa Rosalia or why are there so many kinds of animals?” American Naturalist 93 (1959): 145–159.
Jablonski, David, et al. “Out of the tropics: Evolutionary dynamics of the latitudinal diversity gradient.” Science 314 (2006): 102–106.
MacArthur, Robert H. “Population ecol-ogy of some warblers of northeastern coniferous forests.” Ecology 39 (1958): 599–619.
Novotny, Vojtek, et al. “Why are there so many species of herbivorous insects in tropical rain forests?” Science 313 (2006): 1,115–1,118. Summary by Kitch-ing, R. L. “Crafting the pieces of the diversity jigsaw puzzle.” Science 313 (2006): 1,055–1,057.
xviii+598_Biodiversity_F.indd 51 11/17/11 1:12 PM

first entry52 biodiversity, patterns of 52
tropical rain forests. two and a half acres (a hectare) of tropical forest can have 250 species of trees, com-pared to 20 species in a north american deciduous forest. about 0.2 square miles (half a square kilome-ter) of tropical rain forest can contain more tree spe-cies than all the temperate forests of europe, north america, and asia combined. astonishing diversity also occurs within evolutionary groups in the trop-ics: One forest on Sarawak in indonesia contains 27 species of hemiparasitic figs, and a forest in ecuador has 34 species of palms.
One reason (aside from reasons presented previ-ously) that tropical rain forests have such a dispro-portionately large number of plant species may be that each one is attacked by specialized herbivores and parasites, as pointed out by the ecologist Dan Janzen. in temperate forests, a young acorn can grow into a mature oak tree in close proximity to its parent, though not in the parent’s shade. in tropi-cal forests, a young tree that grows near its parent would quickly be killed by the herbivores and pests on the parent tree. this may be the reason that the individual trees of each species are often so widely separated from one another. Since no single tree spe-cies can usually dominate a canopy, there is literally room for many tree species to grow. this process occurs during the seedling stage. the ecologist liza comita and her colleagues, working with the ecolo-gist Stephen Hubbell, found that seedlings of rare tropical forest tree species (which is to say, most of them) grow poorly near other members of the same species, while seedlings of common tree species grow well near other members of their species. along with tree species diversity goes the diversity of small ani-mals that specialize upon them. trees defend their leaves with poisonous chemicals, each species using its own special toxic mix, and each tree species has herbivores that can detoxify that species’ particular mix of poisons. in contrast, tropical aphids do not have as much diversity as the tropical insects that eat leaves, since aphids drink nonpoisonous sap.
another reason may be that tropical forests have entire ways of living that are not readily available in other regions. in particular, much of the diversity of plant and animal species in tropical forests occurs in the epiphytic plants that live in the upper branches of the trees. Bromeliad plants capture pools of water, and orchids absorb water through aerial roots. Both create habitats for whole communities of animals. in temperate forests, it does not rain enough for many plant and animal species to live up in the branches, totally disconnected from the soil.
if these explanations are the most important ones, it would follow that tropical forest species do not specialize upon particular microclimates. that
is, a small plot of tropical rain forest does not con-tain 250 tree species because it contains 250 dis-tinguishable microclimates. the ecologist Stephen Hubbell found that the trees of Barro colorado island in Panama appeared to be randomly distrib-uted with respect to small disturbances and differ-ences in slope and the direction the slope faced.
these large-scale ecological patterns may not apply to the more limited scale of biodiversity hot-spots. even though the tropics contain the greatest number of species, most biodiversity hotspots are not tropical.
why arE moST SpECIES TErrESTrIal?about 90 percent of nonmicrobial species are terres-trial. in 1994, Robert M. May estimated 85 percent, and in 2009, Michael Benton estimated 95 percent, of species to be terrestrial. this pattern is true even of biodiversity hotspots: less than half an acre (a hectare) of rain forest may have 475 tree species and 25,000 insect species, but the same area of coral reef (often called “rain forests of the oceans”) may have only 300 coral species, 600 fish species, and 200 algae species.
One explanation proposed for lower marine bio-diversity is the tremendous contribution of flowering plants to the evolution of terrestrial biodiversity. Sessile organisms (in which the adults do not move) have to disperse their eggs, sperm, seeds, or spores to new locations. in the oceans, water provides enough buoyant force to fulfill this role very well. air is not as buoyant a medium for dispersal. On land, there-fore, sessile organisms (mostly plants) usually rely on animals for dispersal. a tremendous burst of coevo-lution occurred when the flowering plants evolved on land about 120 million years ago. Moreover, the ecologists Rick Grosberg and Geerat Vermeij indicate that flowering plants have three to 10 times as much water supply per unit leaf surface area as do conifers. this simply allows them to grow more. therefore, the greater biodiversity of terrestrial than of marine regions may be the direct and indirect result of the evolution of flowering plants. For about 290 mil-lion years, until flowering plants evolved, there were more species in the oceans than on land. the oceans, however, remain very poorly explored—not just the seafloors but also the intermediate depths—com-pared to land. it seems unlikely that a large enough trove of newly discovered marine species will be found to alter the assessment that most species are terrestrial. Of course, who knows for sure?
although shallow tropical seas may generally have the highest marine species diversity, there are exceptions. according to the ecologist andrew Bar-ton, phytoplankton hotspots do not form a tropical band but are found off the coasts of Japan, Baja
(continued from page 49)
xviii+598_Biodiversity_F.indd 52 11/17/11 1:12 PM

biodiversity, patterns of 53
california, newfoundland, and argentina and are due mostly to the upwelling of deep ocean nutrients by marine currents.
Summarytropical areas on land and in the oceans usu-ally have more species than other regions because warmth and moisture on land and direct sunlight in water strongly promote the growth of plants and animals and evolution within their popula-tions. However, in habitats with very high nutrient availability, a few dominant species may take over, which results in lower biodiversity. in addition, tropical regions have high biodiversity for histori-cal reasons, such as less disruption during the ice ages. the main reason there are so many species of tropical trees may be that the trees cannot survive near their parents; hundreds of tree species are often spread out over the landscape. there are also more known species of organisms on land than in the oceans perhaps because of the explosion of coevolution that began 120 million years ago with the origin of the flowering plants. none of these reasons, individually or together, seems satisfactory to explain the breathtaking diversity of tropical forests.
See also essay: “Why are there So Many Spe-cies?”; communities, ecological; deep oceans, biodiversity in; disturbances, ecological; Janzen, Daniel H.; population genetics; tropi-cal rain forests and deforestation.
Further readingadams, J. M., and F. ian Woodward. “Patterns in tree spe-
cies richness as a test of the glacial extinction hypoth-esis.” Nature 339 (1989): 699–701.
allen, a. P., et al. “Global biodiversity, biochemical kinet-ics, and the energetic-equivalence rule.” Science 297 (2002): 1,545–1,548.
Barton, andrew D., et al. “Patterns of diversity in marine phytoplankton.” Science 327 (2010): 1,509–1,511.
Brown, J. H. “two decades of homage to Santa Rosalia: toward a general theory of diversity.” American Zool-ogist 21 (1981): 877–888.
clark, James S. “individuals and the variation needed for high species diversity in forest trees.” Science 327 (2010): 1,129–1,132.
colwell, Rita K., and D. c. lees. “the mid-domain effect: Geometric constraints on the geography of species richness.” Trends in Ecology and Evolution 15 (2000): 70–76.
comita, liza S., et al. “asymmetric density dependence shapes species abundances in a tropical tree commu-nity.” Science 329 (2010): 330–332.
connell, Joseph H. “Diversity in tropical rain forests and coral reefs.” Science 199 (1978): 1,302–1,310.
———, and eduardo Orias. “the ecological regulation of species diversity.” American Naturalist 98 (1964): 399–414.
crutsinger, Gregory M., et al. “Plant genotypic diversity predicts community structure and governs an ecosys-tem process.” Science 313 (2006): 966–968.
currie, D. J. “energy and large-scale patterns of animal- and plant-species richness.” American Naturalist 137 (1991): 27–49.
———, and V. Paquin. “large-scale biogeographical pat-terns of species richness of trees.” Nature 329 (1987): 326–327.
Forbes, andrew a., et al. “Sequential sympatric speciation across trophic levels.” Science 323 (2009): 776–779.
Gentry, alwyn H. “Patterns of neotropical plant-species diversity.” Evolutionary Biology 15 (1982): 1–85.
———. “tree species richness of upper amazon forests.” Proceedings of the National Academy of Sciences USA 85 (1988): 156–159.
Givnish, thomas J. “On the causes of gradients in trop-ical tree diversity.” Journal of Ecology 87 (1999): 193–210.
Grassle, J. F., and n. J. Maciolek. “Deep-sea species rich-ness: Regional and local diversity estimates from quantitative bottom samples.” American Naturalist 139 (1992): 313–341.
Hautier, yann, et al. “competition for light causes plant biodiversity loss after eutrophication.” Science 324 (2009): 636–638.
Hawkins, Bradford a., et al. “energy, water, and broad-scale geographic patterns of species richness.” Ecology 84 (2003): 3,105–3,117.
Hodgson, J. G. “Why do so few plant species exploit pro-ductive habitats? an investigation into cytology, plant strategies and abundance within a local flora.” Func-tional Ecology 1 (1987): 243–250.
Hubbell, Stephen P. The Unified Neutral Theory of Biodi-versity and Biogeography. Princeton, n.J.: Princeton University Press, 2001.
Huston, M. “a general hypothesis of species diversity.” American Naturalist 113 (1979): 81–101.
Hutchinson, G. evelyn. “Homage to Santa Rosalia or why are there so many kinds of animals?” American Natu-ralist 93 (1959): 145–159.
Janzen, Daniel H. “Why mountain passes are higher in the tropics.” American Naturalist 101 (1967): 233–249.
———. “Herbivores and the number of tree species in tropical forests.” American Naturalist 104 (1970): 501–528.
latham, R. e., and Robert e. Ricklefs. “Global patterns of tree species richness in moist forests: energy-diversity theory does not account for variation in species rich-ness.” Oikos 67 (1993): 325–333.
leigh, egbert Giles, Jr., et al. “Why do some tropical forests have so many species of trees?” Biotropica 36 (2004): 447–473.
xviii+598_Biodiversity_F.indd 53 11/17/11 1:12 PM

first entry54 biodiversity, quantification of 54
McKenna, Duane D., and Brian D. Farrell. “tropical for-ests are both evolutionary cradles and museums of leaf beetle diversity.” Proceedings of the National Academy of Sciences USA 103 (2006): 10,947–10,951.
Mittelbach, Gary G., et al. “What is the observed relation-ship between species richness and productivity?” Ecol-ogy 82 (2001): 2,381–2,396.
Pärtel, Meelis, et al. “contrasting plant productivity-diver-sity relationships across latitude: the role of evolu-tionary history.” Ecology 88 (2007): 1,091–1,097.
Pennisi, elizabeth. “On rarity and richness: two scientists take a stab at explaining why oceans have far fewer species than terrestrial habitats.” Science 327 (2010): 1,318–1,319.
Prance, Ghillean t., ed. Biological Diversification in the Tropics. new york: columbia University Press, 1982.
Ritchie, M. e., and H. Olff. “Spatial scaling laws yield a synthetic theory of biodiversity.” Nature 400 (1999): 557–560.
Rohde, K. “latitudinal gradients in species diversity: the search for the primary cause.” Oikos 65 (1992): 514–527.
Rosenzweig, M. l. Species Diversity in Space and Time. cambridge: cambridge University Press, 1995.
Snelgrove, P. V. R. “Getting to the bottom of marine biodi-versity: Sedimentary habitats. BioScience 49 (1999): 129–138.
Waide, R. B., et al. “the relationship between productivity and species richness.” Annual Review of Ecology and Systematics 30 (1999): 257–300.
Willig, M. R., et al. “latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis.” Annual Review of Ecology and Systematics 34 (2003): 273–309.
Willis, K. J., and R. J. Whittaker. “Species diversity: Scale matters.” Science 295 (2002): 1,245–1,248.
Wright, Shane, et al. “the road from Santa Rosalia: a faster tempo of evolution in tropical climates.” Pro-ceedings of the National Academy of Sciences USA 103 (2006): 7,718–7,722.
biodiversity, quantification of Biodiversity is usually considered to be the total number of spe-cies that exist in a defined area, although the concept also includes genetic diversity within species and the diversity of habitats and ecological communities. the quantification of biodiversity involves three prob-lems: counting the species that exist in a defined area, calculating indices that reflect the relative impor-tance of each species to the others, and determining how many of those species are “new to science,” that is, recognized as a distinct species by scientists.
CouNTING SpECIESit would seem to be easy to determine how many species exist in a particular location: Go out, find
all the species, and write down their names. this was the approach used by early european scientific explorers, such as Joseph Banks, who accompanied captain James cook on his Polynesian expeditions, and charles Darwin, who accompanied captain Robert FitzRoy on his expedition to South america. the number of species in an area is known as species richness. in reality, it can be very difficult to do this. For example, the insect species in a tropical rain for-est can be so numerous and so well hidden that they can be only indirectly counted. the ecologist terry irwin estimated insect species richness in a rain for-est by choosing trees of different species, gassing them with insecticide, and catching and identifying the insects that fell from the trees. not only did he find that a single tropical tree could contain as many insect species as some european countries, but that each species of tree had many of its own unique species of insects. Vast expertise is required to dis-tinguish one species from another. Scientists typi-cally work in teams, each scientist being an expert on a different group of organisms. For microbes, the look-and-count system is practically impossible. Scientists use Dna methods to quantify the species richness of microbes.
BIodIvErSITy INdICESthe use of species richness as the only measurement of biodiversity conceals a great deal of information about the ecological community. in ecological and evolutionary terms, scientists distinguish between a community that contains, say, 50 equally abundant species and one that contains 50 species, 49 of which are rare. the first of these communities would be considered more diverse than the second, which is overwhelmingly dominated by a single species. ecol-ogists have, accordingly, devised different diversity indices (symbolized by H), rather than to rely simply on species richness.
in order to calculate a diversity index, it is first necessary to determine the importance of the species in the community. this must be a numerical calcu-lation rather than a value judgment by the investi-gator. Different investigators use different methods to calculate the relative importance of each species within a community. (Usually, these investigators focus on one group of organisms within an eco-logical community, such as a plant community or an insect community.) ecologists such as John t. curtis distinguished three factors that contributed to the importance of a species in a community, concepts that he developed for deciduous forest trees:
● Relative frequency. How many samples within the sampling area contain the species of interest? a common tree species may be
xviii+598_Biodiversity_F.indd 54 11/17/11 1:12 PM

biodiversity, quantification of 55
found in all of them, a rare species in only one.
● Relative density. How many individuals of the species of interest are found in the sam-pling area? a common tree species will be represented by more individuals than will a rare species.
● Relative dominance. How much area does each tree species use within the sampling area? an important tree species, for exam-ple, will cover more area of ground than will a rare tree species. Sometimes, the relative dominance of a species of tree is calculated from its crown area, the area of ground shaded directly by its branches, and some-times by its basal area, the cross-sectional area of its trunk.
curtis and others calculated an importance value (iV) by adding all three (expressed as percentages) into a total, in which all species summed to 300. importance values can be converted to proportions that add up to 1. importance values can, in this way, be measured quickly in the field. ecologists such as Robert Whittaker preferred the more laborious but much more informative measurement of net produc-tion, which is the net growth of each species. in the case of a plant species, this would be its total photo-synthetic production minus the biomass lost through cellular respiration. By this measure, the most impor-tant species is the one that grows the most.
in each of these indices, there are s species, and the proportional importance of the ith species is represented by pi. if one simply adds up all of these proportions, the result is 1. the indices are calcu-lated by adding up mathematical derivatives of the proportions. One of these indices, Simpson’s diver-sity index, emphasizes the dominant species because it adds up the squares of the proportions:
H pii
s
==∑ 2
1
the other, the Shannon-Wiener diversity index, which is based upon information theory developed by the mathematicians claude Shannon and norbert Wiener, emphasizes the rare species because it multi-plies each proportion by its natural logarithm:
H p pi ii
s
= −=∑ ln
1
Besides diversity, scientists can also quantify equitability. in a community of species with high equitability, the species are all about equally impor-tant; in a community with low equitability, there are
a few dominant species, and the rest are rare. equi-tability (e) is the diversity divided by the natural logarithm of the number of species:
e = H/ln s
Within the United States, appalachian cove for-ests not only have many plant species, but these spe-cies are equitable. in contrast, a boreal forest has few species and low equitability. this can be illustrated by dominance-diversity curves. in these curves, the relative importance of each species (as a percent) is graphed against the species sequence (ranging from the most important to the least important species). the relative importances (on the vertical axis) are usually displayed on a logarithmic scale.
While an infinite number of patterns is possible in a dominance-diversity curve, three general pat-terns are usually recognized:
● the geometric series assumes that the com-munity is divided among its component spe-cies by a constant proportion. For example, if the most important species has an impor-tance of 50 percent, the second-most-impor-tant will have 25 percent importance, the next will have 12.5 percent importance, and so on. the resulting logarithmic dominance-diversity curve is steep and straight.
● the MacArthur distribution assumes that the community is divided among its com-ponent species in a random fashion. as a result, there will be a few species that are very important, a few that are very rare, but most will be of intermediate importance. the dominance-diversity curve will display a gentle downward slope. it was proposed by the ecologist Robert Macarthur in the middle of the 20th century.
● the log-normal distribution assumes that the importance values have a normal, or bell-shaped, distribution. this, too, pro-duces a gently sloping dominance-diver-sity curve, with few rare and few common species.
in general, communities that live in recently dis-turbed or extreme environments and have few spe-cies display a geometric series of importance values. an example is the vascular plants of the subalpine forest of the Great Smoky Mountains in tennessee (see figure on page 56), based upon net production of each plant species. in communities that have more species, such as the vascular plants of a cove forest of the Great Smoky Mountains, the species divide up the resources more equably (see figure).
xviii+598_Biodiversity_F.indd 55 11/17/11 1:12 PM
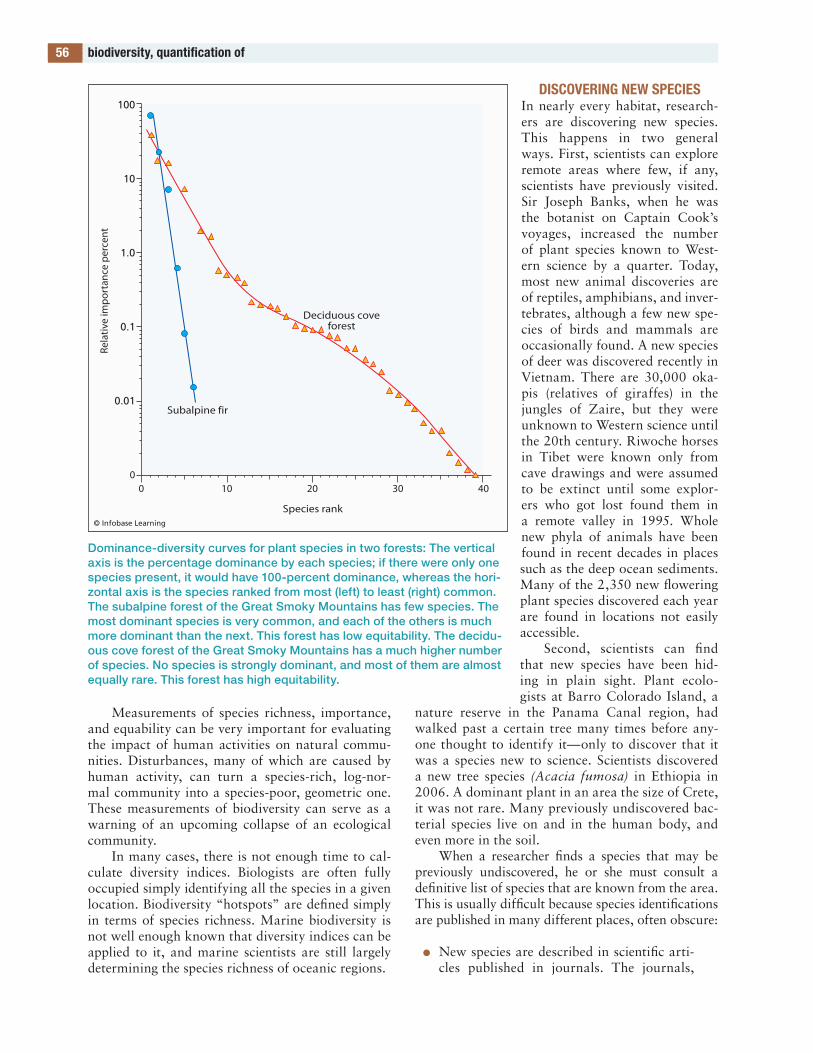
fi rst entry56 biodiversity, quantifi cation of 56
Measurements of species richness, importance, and equability can be very important for evaluating the impact of human activities on natural commu-nities. Disturbances, many of which are caused by human activity, can turn a species-rich, log-nor-mal community into a species-poor, geometric one. these measurements of biodiversity can serve as a warning of an upcoming collapse of an ecological community.
in many cases, there is not enough time to cal-culate diversity indices. Biologists are often fully occupied simply identifying all the species in a given location. Biodiversity “hotspots” are defi ned simply in terms of species richness. Marine biodiversity is not well enough known that diversity indices can be applied to it, and marine scientists are still largely determining the species richness of oceanic regions.
dISCovErING NEw SpECIESin nearly every habitat, research-ers are discovering new species. this happens in two general ways. First, scientists can explore remote areas where few, if any, scientists have previously visited. Sir Joseph Banks, when he was the botanist on captain cook’s voyages, increased the number of plant species known to West-ern science by a quarter. today, most new animal discoveries are of reptiles, amphibians, and inver-tebrates, although a few new spe-cies of birds and mammals are occasionally found. a new species of deer was discovered recently in Vietnam. there are 30,000 oka-pis (relatives of giraffes) in the jungles of Zaire, but they were unknown to Western science until the 20th century. Riwoche horses in tibet were known only from cave drawings and were assumed to be extinct until some explor-ers who got lost found them in a remote valley in 1995. Whole new phyla of animals have been found in recent decades in places such as the deep ocean sediments. Many of the 2,350 new fl owering plant species discovered each year are found in locations not easily accessible.
Second, scientists can fi nd that new species have been hid-ing in plain sight. Plant ecolo-gists at Barro colorado island, a
nature reserve in the Panama canal region, had walked past a certain tree many times before any-one thought to identify it—only to discover that it was a species new to science. Scientists discovered a new tree species (Acacia fumosa) in ethiopia in 2006. a dominant plant in an area the size of crete, it was not rare. Many previously undiscovered bac-terial species live on and in the human body, and even more in the soil.
When a researcher fi nds a species that may be previously undiscovered, he or she must consult a defi nitive list of species that are known from the area. this is usually diffi cult because species identifi cations are published in many different places, often obscure:
● new species are described in scientifi c arti-cles published in journals. the journals,
Dominance-diversity curves for plant species in two forests: The vertical axis is the percentage dominance by each species; if there were only one species present, it would have 100-percent dominance, whereas the hori-zontal axis is the species ranked from most (left) to least (right) common. The subalpine forest of the Great Smoky Mountains has few species. The most dominant species is very common, and each of the others is much more dominant than the next. This forest has low equitability. The decidu-ous cove forest of the Great Smoky Mountains has a much higher number of species. No species is strongly dominant, and most of them are almost equally rare. This forest has high equitability.
xviii+598_Biodiversity_F.indd 56 11/17/11 1:12 PM

biodiversity, resources for agriculture 57
particularly older issues or journals that have ceased publication, may be very dif-ficult to locate.
● lists of all the species (for example, a flora is a compilation of all the plant species of a defined location) are usually organized into keys that allow identification. they may be equally difficult to locate.
Keeping track of even the known species is a dif-ficult task. Sometimes, new species turn out to have been already described, although this occurs only about 20 percent of the time. Some scientists esti-mate that there may be 100 million species on earth. at current rates, it would take biologists 14,000 years to catalog them.
the modern answer to this difficulty is com-puter databases. the evolutionary biologist edward O. Wilson said, “imagine an electronic page for each species of organism on earth . . .” the result was the encyclopedia of life. it is a database that may eventually include all species and to which people can submit information. although it circumvents the ultimately necessary but slow process of sci-entific review, information is accepted only from people who have expertise in the relevant group of organisms. each species has its own Web page. all the information (except genetic sequences) that someone may need in order to identify a specimen can be downloaded in the field and used on the spot. the encyclopedia of life or similar databases will not identify a specimen for a researcher, but just allow the researcher to tentatively confirm an iden-tification already made or to confirm that the spe-cies has not been previously discovered. the online information is available by satellite in remote and little-explored regions. Similar online repositories of species information include the catalogue of life and, for north america, the integrated taxonomic information System, although these databases focus mainly on the list of species names that are accepted by experts.
even with advanced databases, it may not be possible to recognize a new species. in some cases, populations of birds that had been considered a single species turn out to be more than one. accord-ing to the biological species concept, populations are of the same species if they can potentially interbreed in the wild. in some cases, birds of one population do not recognize the mating call of another and may therefore qualify as two different species. in some cases, species might be recognized only on the basis of Dna dissimilarity.
Because of the difficulties just cited, there is not and probably never will be a definitive list of the spe-cies that live on earth.
See also communities, ecological; Dna studies of biodiversity; speciation.
Further readingBiodiversity Heritage library. “BHlwiki.” available online.
URl: http://biodivlib.wikispaces.com/about. accessed June 21, 2010.
Encyclopedia of Life. “encyclopedia of life.” available online. URl: http://www.eol.org. accessed June 21, 2010.
ertter, Barbara. “Floristic surprises in north america north of Mexico.” Annals of the Missouri Botanical Gar-den 87 (2000): 81–109. available online. URl: http://ucjeps.berkeley.edu/floristic_surprises.html. accessed June 22, 2010.
integrated taxonomic information System. “integrated taxonomic information System.” available online. URl: http://www.itis.gov. accessed July 2, 2010.
———. “Species 2000.” available online. URl: http://www.catalogueoflife.org. accessed July 2, 2010.
Mabberley, David J. “exploring terra incognita.” Science 324 (2009): 472.
Rodrigues, ana S. l., and thomas M. Brooks. “Shortcuts for biodiversity conservation planning: the effective-ness of surrogates.” Annual Review of Ecology, Evolu-tion, and Systematics 38 (2007): 713–737.
Scholes, R. J., et al. “toward a global biodiversity observ-ing system.” Science 321 (2008): 104–105.
Simons, Marlise. “a stone-age horse still roams a tibetan plateau.” available online. New York Times, november 12, 1995. URl: http://www.nytimes.com/1995/11/12/world/a-stone-age-horse-still-roams-a-tibetan-plateau. html ?pagewanted=1. accessed May 12, 2011.
Whittaker, Robert H. Communities and Ecosystems, 2nd ed. new york: Macmillan, 1975.
biodiversity, resources for agriculture every species of organism plays an important, if not essential, role in local and global ecosystems. How-ever, the diversity of species, and genetic diversity within species, has provided and will continue to provide direct and important benefits to human wel-fare and the world economy. these benefits are often the ones that most easily convince people that spe-cies—as many as possible—need to be rescued from extinction. this entry explains the essential role of wild species in maintaining agriculture. Other entries provide examples of wild species that have made dramatic differences in human health and of new ideas for inventions and processes that have come from the study of wild species.
wIld plaNT SpECIES for food aNd aGrICulTurEWhen humans lived as hunters and gatherers, which was the case for more than 90 percent of the time
xviii+598_Biodiversity_F.indd 57 11/17/11 1:12 PM

first entry58 biodiversity, resources for agriculture 58
that Homo sapiens has existed, they ate a great diversity of foods, including wild seeds and fruits. the physiology of human bodies evolved a depen-dence on this diversity of food sources. For example, some mammals can synthesize ascorbic acid, a mol-ecule that is necessary in the formation of certain tissues in their bodies, but the ancestors of modern humans lost this ability. this was not a problem, however, as ascorbic acid (also known as vitamin c) was abundant in the wild fruits that primitive humans ate. a diversity of plants has always been important in the human diet.
Vitamin c deficiency became a problem after the advent of civilization as many of the poorest humans had to survive on a bland diet of grains. they suffered the symptoms of scurvy as a result. Fortunately, agriculture supplies humans not with just the basic staples, such as wheat, but with a large number of other foods, such as apples and oranges, that supply nutrients such as vitamin c that the staples cannot provide. agriculture can meet the full range of human nutritional needs, not just the need for calories. the database of the United nations Food and agriculture Organization (FaO) lists 152 different major crops produced in the world. even though only a dozen or so crops constitute the lion’s share of agricultural production, human health and economy would suffer substantially without the oth-ers. Human health continues to depend on plant diversity—the diversity of crop species.
the new science of evolutionary medicine has revealed that human health may be best served by a diet that most closely resembles what primitive hunter-gatherers ate. in particular, disorders such as diabetes appear to be stimulated by a reliance on modern, processed foods. if everyone were to go back to a traditional, diverse diet, it would seem that 152 kinds of crops would provide sufficient variety without the need to gather wild foods or domesti-cate new kinds of crops. However, as the botanist Gary Paul nabhan has explained, there is no single primitive diet to which all humans have adapted. He notes that certain genetic lineages of people, such as native americans, native Hawaiians, and native australians, suffer the most from diseases such as diabetes when they depart from traditional cuisines and adopt modern culinary uniformity. He proposed that, during the course of the past few thousand years, cultural groups have biologically adapted to consuming local foods. the wild succulent plants consumed by desert peoples, he points out, not only have less of the quickly metabolized starches that contribute to diabetes, but also contain more of the molecules that help them resist diabetes. if a close coevolution has occurred between local peoples and their local crops and wild plants, then 152 kinds of
plant-based foods may not be enough for the opti-mal health of all humans.
However, the story of diversity does not stop with plant species diversity. Humans need not only a great diversity of crop plant species, but also a great genetic diversity of breeds or varieties within each crop species. there are scores or hundreds of breeds of each species of crop plant.
Plant breeders have traditionally produced new breeds of crop plants by cross-breeding existing varieties of crops to produce new combinations of characteristics. More recently, the techniques of bio-technology have allowed plant breeders to move the Dna from one breed of plant to another. Because the Dna of all species is chemically the same, bio-technologists can move genes from one species of plant to another or even insert a viral, bacterial, or animal gene into plant Dna. thousands of acres are now devoted to crops that contain viral or bacterial genes.
the importance of maintaining a diversity of crop breeds is not immediately apparent to most people. although it is appealing to have different breeds of apples from which to choose, just how important is it? after all, these breeds do not differ greatly in their nutritional values to humans.
the importance of having a great diversity of crop breeds is not so much for everyone’s pleasure in eating as it is for farmers’ success in growing them. it is necessary to preserve a great diversity of crop breeds because they each have adaptations to dif-ferent climates and to different biological environ-ments. Some breeds of wheat tolerate cold winters or droughts better than others, and some breeds of corn can complete their life cycle in a shorter period of time than others, which allows them to grow farther north, where the growing season is shorter. these are examples of adaptations to different cli-mates. adaptations to various biological environ-ments, which consist of the other plants, the viruses, the fungi, and the insects that the crop encounters, are even more important. Some crop breeds are bet-ter than others at growing in the crowded condi-tions of modern agricultural fields. Some can resist viruses, fungi, or insect pests that others cannot. Plant breeders have to keep producing new breeds, because once an established breed has been grown for a long time, viruses or bacteria or fungi will evolve the ability to infect it. it is an evolutionary race between the natural evolution of the pests and the artificial evolution carried out by plant breeders. Breeders keep reshuffling the alleles of different crop varieties to produce new adaptations to changing climate and to new pests.
neither traditional plant breeders nor modern biotechnologists create new genetic variation—they
xviii+598_Biodiversity_F.indd 58 11/17/11 1:12 PM

biodiversity, resources for agriculture 59
just move it around. they have to obtain the alleles from existing populations of organisms, and if those organisms become extinct, the alleles they contain vanish and are no longer available for the production of new breeds of crops.
Wild species have also proven to be good sources of natural pesticides. the Bt toxin, from the bacterium Bacillus thuringiensis, kills insects by disrupting the lining of their digestive tracts. Pyre-thrin insecticides (now modified into the safer and more stable pyrethroids) were discovered in a species of chrysanthemum. Four compounds that prevent insects from feeding as well as their ability to reach maturity have been found in the leaves of the neem tree (Azadirachta indica). the bacterium Strepto-myces avermitilis produces avermectins, which kill roaches and fire ants. a new group of antifungal compounds, the strobilurins, were discovered in Strobilurus tenacellus, a mushroom.
ThrEE ThrEaTS To plaNT dIvErSITy IN aGrICulTurE
there are three kinds of extinction that threaten the future of agriculture as well as the development of new commercial products: extinction of traditional varieties, extinction of wild relatives of existing crops, and extinction of wild species that may serve as future crops.
Genetic extinction of traditional crop varieties. traditional crop varieties are often called “heirloom” varieties. around the world, subsistence farmers and gardeners have developed numerous breeds of crop that are well suited to their local conditions and the preferences of their cultures. they pass the seeds of these varieties from one generation to another. this is why the world contains hundreds of varieties of major crops such as wheat and minor ones such as chili peppers and eggplants. nevertheless, often these heirloom varieties have inferior yields in com-parison to the varieties sold by the large seed com-panies of europe and north america. Subsistence farmers may, if they can afford to do so, choose to eat their old seeds rather than save them for planting the next year and buy the varieties that they perceive are superior. the old varieties and the benefit that they may offer to the future of plant breeding may be lost as a result. By consuming the last seeds of the old varieties, they are literally eating the future.
Plant breeders do not want these traditional varieties of crops to become extinct, and some gov-ernments are trying to rescue them before they van-ish. the U.S. Department of agriculture (USDa), for example, has cold-temperature storage facilities for major crops, such as the one for potato varieties in Sturgeon Bay, Wisconsin. Such facilities are expen-sive, and they are used only to save the varieties of
the crops that earn the most money for large-scale agriculture. Because cold-temperature storage in warm countries is expensive, five northern countries (led by norway) established a seed vault in the arctic Svalbard islands in 2006. the facility will eventu-ally have 3 million seeds and store more varieties of more crops than any previous project. But these measures by themselves are not enough. Seeds will die after being stored for a few decades, so they must be planted and grown, to produce new seeds, which are then stored again.
there is, however, a dark side to the Svalbard seed vault. although the actual seeds in the vault are not available to them, large agricultural corpora-tions have free access to the genetic varieties repre-sented by those seeds. these corporations can use genetic engineering to modify the varieties and then patent them. this would give the corporations own-ership over the crop varieties, and no one can then use these varieties without paying the corporations. Families, communities, and tribes have for millennia planted their traditional varieties of garden plants, passing the seeds down from one generation to the next. Since many of these varieties are threatened with extinction, nongovernmental organizations, such as Seed Savers exchange, have rescued thou-sands of crop varieties from oblivion by storing and growing them. Such organizations have aspired to help tribes and communities, which freely donated their traditional and often sacred seeds, to keep their food plant genetic variation alive. large corpora-tions, however, may now be able to patent these very same varieties and charge farmers for the privilege of planting their own traditional seeds. Many conser-vationists consider this to be corporate profit based on a freely donated public resource and consider the Svalbard seed vault, however lofty its purpose may appear to be, to constitute corporate acquisition of genetic resources for which they have not paid. Seed Savers exchange is one of the organizations that has agreed to these conditions, thus making them a conduit of traditional varieties from families, tribes, and communities to the control of large agricultural corporations.
Often, people save varieties not just for their usefulness but for pleasure. in addition to the regu-lar kind of basil with which most aficionados of italian cuisine are familiar, there are many other varieties, which have flavors similar to everything from cinnamon to lemon. there is also a garden variety of “chocolate mint” whose raw leaves live up to their name. Diversity within major and minor crop species is important for both practical reasons and for sheer enjoyment.
Genetic extinction of wild relatives of crop plants. Wild breeds or species of plants that can
xviii+598_Biodiversity_F.indd 59 11/17/11 1:12 PM

first entry60 biodiversity, resources for agriculture 60
cross-breed with crop plants can provide even more genetic variety than the heirloom crop varieties. For example, there are at least 212 species of cultivated plants from the tropics alone that have wild popula-tions closely related to them. chocolate, the health benefits of which are now being revealed, is Theo-broma cacao. the genus Theobroma has 21 wild species that can be useful in producing new breeds of cocoa. there is at least one famous example of a wild species that rescued a cultivated one from danger. When phylloxera bug (Daktulosphaira viti-foliae) infestations threatened european grapevines (Vitis vinifera) and the wine industry in the late 19th century by infecting vines with a mildew, the botanist t. V. Munson found that wild grapes (Vitis rotundifolia) from Oklahoma and texas could resist this insect. He shipped thousands of wild grape rootstocks to France, and French grapevines were grafted to these roots. this action is reputed to have saved the european wine industry.
However, like the traditional garden varieties, the wild relatives of crops are vanishing, mostly because the natural habitats in which they live are being destroyed. the wild ancestors of wheat, for instance, persist in the hills of the Middle east, where war and land degradation endanger them. Some species of teosinte, the wild ancestor of maize, barely survive in the foothills of Mexico. in some cases, traditional garden varieties have avoided extinction by escaping into the wild. For example, wild coffee trees live in the forests of ethiopia adja-cent to the fields of cultivated coffee, but these for-ests, like most others, are shrinking under human influence. For a few crops, such as mangoes and papayas, the wild relatives thrive in disturbed areas and are therefore in no danger of extinction. Such examples, however, are in the minority, and for even these species, at least some of the wild relatives are threatened with oblivion.
Extinction of possible new crops. there are many wild species of plants that offer the promise of future development as crops. Given that there are already so many kinds of crops (even more than the 152 major categories listed by the FaO), why would it be necessary to develop new ones? this is a good question, especially because the development of a new crop is a protracted and difficult process, but there are definite economic and ecological benefits to the development of new crops from wild species. Wild plants are not only important for individual survival in an emergency—the U.S. Army Survival Manual describes 74 of them—but for the future of agriculture.
new crops may offer significant market pos-sibilities if their taste or usefulness is unique. For example, most persimmons sold in the United States
are Japanese persimmons (Diospyros kaki), but the wild american persimmon (Diospyros virginiana) provides a fruit whose wonderful flavor differs not only from all others in the supermarket but from the Japanese species as well. likewise, there is no other fruit with a flavor quite like that of the mul-berry, Morus alba. a significant amount of work would be necessary to prepare these two wild species for orchard cultivation. the american persimmon fruit is small and full of large seeds, and mulberries typically lose their flavor if frozen. Wild fruits usu-ally ripen over a prolonged period that would make simultaneous harvest impractical. Still, someday we may find that there is a market niche for them. Other wild plants, such as dandelions and violets and even (after cooking) nettles, are eminently edible and nutritious.
Perhaps more important, new crops may offer significant ecological advantages. Some might, for example, be able to grow exceptionally well in degraded or dry soils otherwise unsuitable for agri-culture. the spread of soil degradation through ero-sion and salinization creates an incentive to develop new crops that can live in degraded soil. there are three ecological circumstances for which new crops might offer a partial solution.
● New crops for saline soils. around the world, irrigated land in warm, dry regions is the most productive. irrigation, however, causes salt to build up in the soil. according to the United nations Food and agricul-ture Organization (FaO), of the more than 500 million acres (230 million hectares) of agricultural land that are irrigated, almost 20 percent have been salinized. Relatively few crop plants can grow in salty soil (beets and barley are among the few exceptions), and even these cannot tolerate very much salt. However, some wild plants can tol-erate high levels of salt. Wildwheat (Dis-tichlis palmeri), a wild grass native to the southwestern United States, produces an edible grain, and the oil-rich seeds of the pickleweed Salicornia can be added to ani-mal feed. S. bigelovii can produce 0.8 tons per acre (2.0 metric tons per hectare) of seed. in contrast, the average sunflower oil crop produces only 60 percent as much. in theory, genetic engineering could be used to produce salt-tolerant crops, but salt toler-ance turns out to be a complex set of traits that have not yet been successfully intro-duced into existing crops. if humans are to raise crops in salinized soil, finding wild plants may be the only option.
xviii+598_Biodiversity_F.indd 60 11/17/11 1:12 PM

biodiversity, resources for agriculture 61
● New crops for dry soils. Some wild plants that thrive under desert conditions might prove useful for agriculture in the future. the jojoba (Simmondsia chinensis), a bush from the north american southwestern desert, produces a liquid wax that can be useful in industrial processes because it is stable under high temperature and pres-sure. Many acres of jojoba have been raised in irrigated lands in the desert Southwest. although the bush does not need irrigation, water supplementation boosts its growth and yields. Unfortunately, this bush became a victim of its own success. irrigation raised the land values so much that jojoba planta-tions, which yield only a moderate profit, have been converted to lucrative subdivi-sions. Other arid land plants, however, grow well without irrigation. Prickly pear cac-tus (Opuntia ficus-indica) produces edible fruits (tuna in Spanish) and edible pads or stems (nopalitos in Spanish). Buffalo gourds (Cucurbita foetidissima) grow well on road-sides of the deserts and high plains. as its latin name suggests, it is inedible. However, its massive root can yield starch for etha-nol production. its seeds can produce more oil than some cultivated oil crops, with no irrigation.
● New crops for polluted areas. Wild cattails can grow in water contaminated by sewage. the bacteria that grow around their stems and roots break down the sewage, and the cattail plants absorb the released nutrients. arcata, california, uses cattail marshes for its municipal sewage treatment. the under-ground stems of the plants produce a great deal of starch, which can be harvested for the production of fuel ethanol.
Plant breeders can also produce new crops that cause less environmental degradation in the first place. Perhaps the best example of this is natural systems agriculture (nSa), which is the focus of research at the land institute in Salina, Kansas. nSa uses perennial crops rather than annual crops and therefore requires less energy and less fertilizer, as well as minimizing soil erosion. Only a few of today’s main agricultural crops, among them pota-toes and alfalfa, are perennials. in order to promote an agriculture based on perennials, land institute scientists have to develop new crops, and to do so, they draw from the resources of wild plant species. For example, wild wheatgrass (Thinopyrum inter-medium) is a perennial relative of wheat that could be used as a source of grains. Maximilian sunflower
(Helianthus maximiliani) and rosinweed (Silphium integrifolium) are perennials that could be used as a source of oilseeds. Bundleflower (Desmanthus illi-noensis) is a perennial legume that produces seeds similar to those of soybean, only smaller.
the main work of plant breeders at the land institute is to cross wild perennial plants with annual crop species. this is a quicker and more promising route to the development of perennial replacements for annual crops than is the direct breeding of wild perennials. the researchers cross perennial wheatgrass with annual wheat, Maximil-ian sunflower with annual sunflower, and peren-nial Johnson grass (Sorghum halepense) with annual sorghum (S. bicolor). they then cross these hybrids with the annual parents (see figure on below), thus producing plants that more closely resemble the annual crops, except that they may be perennials.
Plant breeders at the Land Institute in Salina, Kansas, are cross-breeding domesticated annual sunflowers and wild perennial sunflowers, hoping to obtain a perennial breed of sunflower that has commercially valuable properties, such as high seed-oil content. The outer (ray) florets, which do not produce pollen, have been removed from each of the sunflowers. Breeding new crops is one of the economic benefits of wild plant species. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 61 11/17/11 1:12 PM

first entry62 biodiversity, resources for agriculture 62
they plant thousands of cross-bred specimens and then see which ones grow for more than a year. From among these survivors, they choose the plants with the best characteristics. the best characteristics are usually the same ones that the earliest farmers selected: breeds that have large seeds, do not shat-ter the mature seeds, have no seed dormancy, have a high harvest index, and do not collapse under the weight of their own seeds.
if successful, perennial crops will allow farmers to produce grains, oilseeds, and legumes that have characteristics desirable to humans, that minimize soil erosion, and that promote fertility of the soil and its ability to hold water.
Saving old crops can be as useful as finding new ones. native americans of Mexico and South amer-ica grew large amounts of amaranth and quinoa, the seeds of which can be used as grains even though they are not grasses. the conquistadores tried to stamp out the cultivation of these crops, ostensibly because of their association with paganism. Fortu-nately, they were not entirely successful. although they are not major crops, amaranth and quinoa are still grown and eaten in the places where they once flourished and available for purchase in some ameri-can supermarkets. Many of the traditional grains of africa, such as an african species of rice, finger and pearl millets, fonio, sorghum, and tef, are still grown in rural areas.
plaNT aNd aNImal dIvErSITy dowN oN ThE farm
the importance of plant diversity is perhaps most obvious not in what is grown on farms but in the way it is grown. Modern agriculture is overwhelm-ingly based on monoculture, in which farmers plant large expanses of just a single breed of a single species of crop plant. Monoculture invites many kinds of problems. For example, pests and diseases can spread rapidly through a monoculture of crops because their preferred food plant is always just a few inches away. if a whole field consists of one species or even one variety of crop, all of them are competing for the same nutrients in the same soil level and the same sunlight at the same height above the ground. Despite these problems, farmers in industrialized nations usually grow their crops in monocultures, because a huge, genetically uniform field is easier to manage by means of machinery and chemicals. if a field contains nothing but corn and all the ears of corn are exactly the same height above the ground, a mechanical harvester can get all of the corn by just driving back and forth across the field.
One of the worst examples of monoculture is the modern banana. thousands of square miles of land in latin america are a monoculture of just
one kind of banana: the cavendish. Because domes-ticated bananas do not produce seeds, they must be propagated by cuttings, all of which are geneti-cally identical. a genetically uniform monoculture is an invitation for the spread of disease. this is pre-cisely what has happened: Panama disease, caused by a fungus, Fusarium oxysporum, has devastated banana plantations in Southeast asia, and it is only a matter of time before it spreads to latin amer-ica. Fruit companies first marketed the cavendish banana because the previously popular variety, the Gros Michel, had been devastated by a different strain of Panama disease.
Planting more than one kind of crop in the same field, a practice called polyculture, can greatly reduce these problems. Because each of the crop species is at a lower density in a polyculture than it would be in a monoculture, pests spread much more slowly through fields that consist of more than one crop species. Some tomato fields in cali-fornia are interrupted with rows of sunflowers. the sunflowers can impede the movement of sphinx moths through tomato fields, causing them to lay fewer eggs, which hatch into voracious tomato hornworms. even planting different breeds of the same crop together in a large field can confer some polyculture benefits. interplanting disease-resistant with disease-susceptible crops can help protect the susceptible ones. Planting rice varieties that grow to different heights together in the same field creates pockets of temperature variation that reduce the spread of fungus disease.
Polyculture is an essential component of natural systems agriculture. By using perennial plants in polyculture, nSa is following the lead of the geneti-cist Wes Jackson, founder of the land institute, who takes the tallgrass prairie as his inspiration and model. a mixture of perennial species has made the prairie a successful natural habitat; the same system should work for agricultural lands. His vision is to make agriculture incorporate natural processes that have been perfected by millions of years of evolution.
if the crops in the mixture are of different sizes, they might be able to use the sunlight and the dif-ferent soil depths more efficiently than could a monoculture. the gardens of tribal peoples almost always contained mixtures of species growing side by side. native american gardens were centered on the “three sisters”—maize, beans, and squash (see figure on page 63). the beans and squash grew near the ground and used the tall corn as a trellis. Other examples include growing rows of papaya trees in pineapple fields in the Philippines and grow-ing orange trees beneath date palms in southeast-ern california. the papaya trees do not shade the
xviii+598_Biodiversity_F.indd 62 11/17/11 1:12 PM

biodiversity, resources for agriculture 63
pineapples nor do the date palms shade the oranges enough to impede their growth. in fact, some crops such as coffee bushes grow better in moderate shade.
another way in which polyculture can enhance the efficiency of agricultural production is that a farmer can benefit from raising more than one prod-uct. although this may not particularly matter in the market economies of industrialized countries, it can be very important in rural areas of the developing world. For example, farmers who plant the fast-growing tropical tree leucaena (Leucaena leuco-cephala) along with their primary crop can harvest the crop as well as the leucaena. Because the trees are leguminous, they have nitrogen-fixing bacteria in their roots. this allows their pods and leaves to be nutritious food for livestock and even means that they can enrich the soil with nitrogen fertilizer. in addition, the trees can help prevent soil erosion.
in crop rotation, a farmer plants different crops in a field each year. Pests of the first crop may build
up, only to die the next year when a different kind of crop is planted. a leguminous crop one year can enrich the soil with nitrogen that the next year’s crop can use. crop rotation can be considered a tempo-ral polyculture that alternates in time rather than in space. Farmers have known about crop rotation for many centuries, though they did not understand how it worked. the Roman poet Virgil wrote about legume-grain crop rotation:
Sow in the golden grain where previously you raised a crop of beans that gaily shook Within their pods, or a tiny brood of vetch . . . thus will the land find rest in its change of crop, and earth left unploughed show you gratitude.
there is no reason that polyculture cannot be integrated into modern mechanized agriculture. instead of having a square mile of corn and another square mile of soybeans, a farmer can plant alternat-ing swaths of corn and soybeans that are still wide enough to allow farm equipment to maneuver. Such fields used to be a rare sight but are increasingly common in the midwestern United States.
is it possible that all the wild plants that could be domesticated have already been brought under cultivation? this is probably not the case. Once agri-culture evolved, it was easier for farmers who moved to new locations to bring crops with them than to develop crops from wild plants in the new loca-tion. the success of the early crop plants therefore preempted the development of equally promising or even superior new crop plants. the great diversity of wild plant species therefore still represents a cornu-copia of possible new resources for agriculture.
there is a parallel decline in the diversity of livestock breeds. in the last 150 years, 190 breeds of livestock have become extinct, 60 of them in just the last five years. While the remaining breeds are very productive, the extinction of “inferior” breeds has reduced the ability to breed for disease resistance, just as is the case with crops. Holsteins are 90 per-cent of dairy cattle, and leghorn chickens produce almost all eggs.
See also biodiversity, resources for medi-cine; biomimicry; Jackson, Wes; natural sys-tems agriculture.
Further readingchivian, eric, and aaron Bernstein, eds. Sustaining Life:
How Human Health Depends on Biodiversity. new york: Oxford University Press, 2008.
collins, W. W., and c. O. Quaslet, eds. Biodiversity in Agroecosystems. new york: lewis, 1998.
cox, t. S., et al. “Breeding perennial grain crops.” Critical Reviews in Plant Sciences 21 (2002): 59–91.
Native American agriculture (for example, as at New Echota, a reconstructed Cherokee town in Georgia) used polycultures, in which several crop species grow together. Polyculture reduces the density and therefore the pest load of each crop species. Hollow gourds allow swallows, which eat insect pests, a place to nest. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 63 11/17/11 1:12 PM

first entry64 biodiversity, resources for medicine 64
Donia, Mohamed S., et al. “a global assembly line for cyanobactins.” Nature Chemical Biology 4 (2008): 341–343.
elias, thomas, and Peter Dykeman. Edible Wild Plants: A North American Field Guide. new york: Sterling, 1990.
Farnham, timothy J. Saving Nature’s Legacy: Origins of the Idea of Biological Diversity. new Haven, conn.: yale University Press, 2007.
Fischbach, Michael a., and christopher t. Walsh. “anti-biotics for emerging pathogens.” Science 325 (2009): 1,089–1,093.
Glover, Jerry D., cindy M. cox, and John P. Reganold. “Future farming: a return to roots?” Scientific Ameri-can, august 2007, 82–89.
Hinman, c. Wiley, and Jack W. Hinman. The Plight and Promise of Arid Land Agriculture. new york: colum-bia University Press, 1992.
Jackson, Wes. New Roots for Agriculture. lincoln, neb.: University of nebraska Press, 1980.
Kalka, Margareta B., et al. “Bats limit arthropods and her-bivory in a tropical forest.” Science 320 (2008): 71.
Koeppel, Dan. Banana: The Fate of the Fruit That Changed the World. new york: Hudson Street Press, 2007.
Maxwell, nicole. Witch-Doctor’s Apprentice: Hunting for Medicinal Plants in the Amazon. new york: citadel, 1961.
nabhan, Gary Paul. Why Some Like It Hot: Food, Genes, and Cultural Diversity. Washington, D.c.: island Press, 2004.
national Research council, Board on Science and tech-nology for international Development. Lost Crops of Africa. Vol. 1: Grains. Washington, D.c.: national academy Press, 1996.
Partida-Martinez, laila P., and christian Hertweck. “Pathogenic fungus harbours endosymbiotic bacteria for toxin production.” Nature 437 (2005): 884–888.
Piel, Jörn. “Metabolites from endosymbiotic bacteria.” Natural Product Reports 26 (2009): 338–362.
Plotkin, Mark. Tales of a Shaman’s Apprentice: An Ethno-botanist Searches for New Medicines in the Amazon Rain Forest. new york: Viking, 1993.
Rice, Stanley a. Green Planet: How Plants Keep the Earth Alive. new Brunswick, n.J.: Rutgers University Press, 2009.
Rosenthal, elizabeth. “near arctic, seed vault is a Fort Knox of food.” available online. New York Times, February 29, 2008. URl: http://www.nytimes.com/2008/02/29/world/europe/29seeds.html?ei=5124&en=006cb7 68718869ca&ex=1362027600&exprod=permalink&pagewanted=print. accessed May 12, 2011.
Rozema, Jelte, and timothy Flowers. “crops for a salinized world.” Science 322 (2008): 1,478–1,481.
Schiebinger, londa. Plants and Empire: Colonial Bio-prospecting in the Atlantic World. cambridge, Mass.: Harvard University Press, 2004.
Smith, nigel J. H., et al. Tropical Forests and Their Crops. ithaca, n.y.: cornell University Press, 1992.
Southern exposure Seed exchange. “Southern exposure Seed exchange.” available online. URl: http://www.southernexposure.com. accessed July 2, 2010.
United nations Food and agriculture Organization. “Biodiversity for a World Without Hunger.” avail-able online. URl: http://www.fao.org/biodiversity/en/. accessed august 8, 2011.
Williams-Guillén, Kimberly, ivette Perfecto, and John Van-dermeer. “Bats limit insects in a neotropical agrofor-estry system.” Science 320 (2008): 70.
Wilson, edward O. The Future of Life. new york: Vintage, 2003.
Wolfe, M. S. “crop strength through diversity.” Nature 405 (2000): 681–682.
Zhu, y. y., et al. “Genetic diversity and disease control in rice.” Nature 406 (2000): 18–722.
biodiversity, resources for medicine the diversity of species and genetic diversity within spe-cies has provided and will continue to provide direct and important benefits to human welfare and the world economy. these benefits are often the ones that most easily convince people that species—as many as possible—need to be rescued from extinc-tion. Biodiversity has been and will continue to be crucial to the improvement of human health in two general ways. First, many wild species produce molecules that have medicinal value to humans. Second, some wild species have provided valu-able insights in medical research. Other entries in this encyclopedia are devoted to the importance of biodiversity to agriculture and to technological progress.
wIld SpECIES aS SourCES of mEdICINal CompouNdS
Humans already owe a tremendous debt of grati-tude to wild species, which have provided nearly all medicinal compounds. new compounds continue to be found. nearly always, the medicinal compound is discovered in a species that seemed to have little importance to human activities and whose extinc-tion would not have been noticed by many people. the following is an overview of examples.
Medicinal compounds from microorganisms. the most famous medicinal compounds are those produced by fungi and bacteria (see table on page 65). they produce antibiotic compounds that are, in effect, chemical warfare against other microorgan-isms with which they compete for food supplies. Sometimes the compounds serve other functions, such as signaling between microbial cells. Because of their short generation times, microbes can evolve
xviii+598_Biodiversity_F.indd 64 11/17/11 1:12 PM
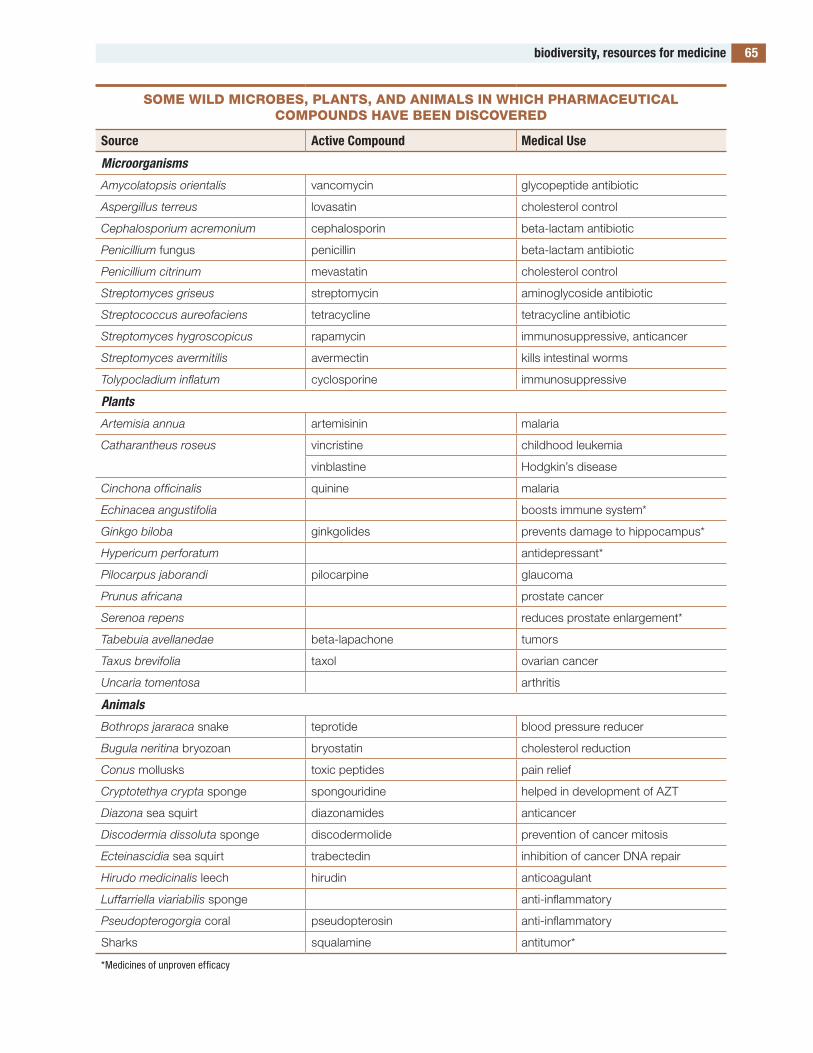
biodiversity, resources for medicine 65
some Wild miCrobes, plants, and animals in WhiCh pharmaCeutiCal Compounds have been disCovered
Source active Compound medical use
Microorganisms
Amycolatopsis orientalis vancomycin glycopeptide antibiotic
Aspergillus terreus lovasatin cholesterol control
Cephalosporium acremonium cephalosporin beta-lactam antibiotic
Penicillium fungus penicillin beta-lactam antibiotic
Penicillium citrinum mevastatin cholesterol control
Streptomyces griseus streptomycin aminoglycoside antibiotic
Streptococcus aureofaciens tetracycline tetracycline antibiotic
Streptomyces hygroscopicus rapamycin immunosuppressive, anticancer
Streptomyces avermitilis avermectin kills intestinal worms
Tolypocladium inflatum cyclosporine immunosuppressive
Plants
Artemisia annua artemisinin malaria
Catharantheus roseus vincristine childhood leukemia
vinblastine Hodgkin’s disease
Cinchona officinalis quinine malaria
Echinacea angustifolia boosts immune system*
Ginkgo biloba ginkgolides prevents damage to hippocampus*
Hypericum perforatum antidepressant*
Pilocarpus jaborandi pilocarpine glaucoma
Prunus africana prostate cancer
Serenoa repens reduces prostate enlargement*
Tabebuia avellanedae beta-lapachone tumors
Taxus brevifolia taxol ovarian cancer
Uncaria tomentosa arthritis
Animals
Bothrops jararaca snake teprotide blood pressure reducer
Bugula neritina bryozoan bryostatin cholesterol reduction
Conus mollusks toxic peptides pain relief
Cryptotethya crypta sponge spongouridine helped in development of AZT
Diazona sea squirt diazonamides anticancer
Discodermia dissoluta sponge discodermolide prevention of cancer mitosis
Ecteinascidia sea squirt trabectedin inhibition of cancer DNA repair
Hirudo medicinalis leech hirudin anticoagulant
Luffarriella viariabilis sponge anti-inflammatory
Pseudopterogorgia coral pseudopterosin anti-inflammatory
Sharks squalamine antitumor*
*Medicines of unproven efficacy
xviii+598_Biodiversity_F.indd 65 11/17/11 1:12 PM

first entry66 biodiversity, resources for medicine 66
quickly, and pathogenic bacteria can evolve resis-tance against antibiotics within a very short period. there are scores of antibiotics, but for each of them, there is a population of bacteria somewhere in the world that is resistant to it. a random sample of soil microbes contains bacteria that can resist anti-biotics. Bacteria can exchange Dna, even between species, with the result that antibiotic resistance can spread from one species of bacterium to another within a habitat, such as a human body. Some fungi produce compounds that have anticancer and anti-viral effects. the fungus Tolypocladium inflatum produces cyclosporine, a compound that happens to suppress the human immune system and is therefore extremely valuable in controlling the rejection of transplanted organs. Many microbial compounds are produced by species that are not known except from the analysis of their Dna.
Medicinal compounds from plants. Wild plants have been and will continue to be important sources of medicines (see table on page 65). the leaves of wild plants produce toxins that deter fungi and her-bivorous animals from damaging them. Many of these toxins, when used in small quantities, have medicinal effects on humans. indeed, most medi-cines began as natural plant extracts. For example, aspirin, though today synthesized artificially as ace-tylsalicylic acid, was once extracted from willow bark and received its name because it was first dis-covered in a wild spiraea bush (Spiraea ulmaria). the plants of the tropical forest are an especially rich source of medicines because their leaves must be very toxic to protect themselves from year-round attack by insects and other animals. Wild plant com-munities, particularly tropical forests, are rich labo-ratories for the production of medicinal compounds, as millions of plants in thousands of species produce new genetic variations of chemicals. Pharmaceuti-cal research corporations often base their research on plant extracts. Because each kind of plant has a unique combination of protective chemicals, we need to save all of them. the extinction of any wild plant species might mean the loss of a yet-undiscovered medicinal compound. Moreover, these compounds are not randomly produced molecules but molecules whose biological potency has been confirmed by natural selection: the plants that produce them have resisted the attacks of organisms.
Medicinal compounds from animals. Many ani-mals also produce compounds that have proven to be of medicinal value. Some amphibians, for example, have poisonous skin. the poison has its effect by relaxing the muscles or reducing the heart rate of the predator. in small quantities, such a compound can be used as a muscle relaxant. Recently, sponges have been found to produce medicinal compounds.
Sponges live in close mutualism with bacteria, and these compounds may help them prevent the bacteria from growing and killing them. the anticoagulant lepirudin was originally known from leeches, which also produce a related compound, hirudin. lepiru-din is now produced commercially, but the leeches themselves are still sometimes used in medical treat-ments, for example, to drain fluid from reattached body parts while the veins are still growing back. cone snails (genus Conus) have up to 140,000 dif-ferent toxic peptides. Some of these molecules may function as pain relievers and anticonvulsants by blocking membrane channels. each kind of peptide blocks a specific kind of channel. Squalamine from sharks, after initial hopes that it would function as an effective antibiotic, may instead be developed into an antitumor drug and an appetite suppressant that does not lower metabolism, as most appetite sup-pressants do.
there is no way to predict, beforehand, which wild species might be “worth saving” for medicine or for some other useful product or to what use they might be put. Who would ever have guessed that a compound that could suppress the human immune system and control the growth of tumors would be found in a soil bacterium from easter island? even so, that is where rapamycin came from (named after Rapa nui, the Polynesian name of the island). Who could ever have guessed that certain forms of leuke-mia could be treated with vincristine and vinblas-tine found in the leaves of Catharanthus roseus, the rosy periwinkle, a small pink wildflower that lives in the vanishing rain forests of Madagascar (see figure on page 67)? Or that prostate cancer could be treated with a compound from the african cherry (Prunus africana)? Or that ovarian cancer could be treated by taxol from the bark of the Pacific yew, Taxus brevifolia? Who would have guessed that the bile of bears contained ursodeoxycholic acid, which may be useful in treating gall bladder conditions in humans? Who would have expected to find a new kind of glue in a frog? the frog Notaden bennetti lies dormant in desert mud for nine months each year encased in a kind of glue that is strong and yet, unlike other strong adhesives, allows molecules to diffuse through it, which is how they breathe.
One promising way of seeking natural pharma-ceuticals is to listen to the lore of tribal peoples who have had millennia of experience with medicinal plants. they can, at least, point the way toward find-ing plant species that have potent properties wor-thy of investigation. they can also indicate which species of frogs have poison skin, which the native people use for hunting darts. Ethnobotanists are sci-entists who visit tribal peoples and ask them about their native traditions concerning medicinal plants.
xviii+598_Biodiversity_F.indd 66 11/17/11 1:12 PM

biodiversity, resources for medicine 67
Occasionally, naturally occurring compounds may reveal new modes of therapy. epibatidine, an alkaloid from the ecuadorean frog Epipedobates tricolor, is a painkiller 200 times as potent as mor-phine, and the human body does not build up tol-erance to it as it does to morphine. not just new antimicrobials but new chemical families of antimi-crobials have been discovered in amphibian skins. this is not unexpected, since amphibians have soft skin that is vulnerable to infection. amphibian anti-microbials include some that are useful against the fungi that infect aiDS victims and against myco-plasmas, which are bacteria that are difficult to control. the sponge Cryptotethya crypta produces compounds such as spongouridine, which at the time of its discovery was a mere curiosity. the compound used the sugar arabinose in its nucleotides instead of the usual ribose (of Rna) or deoxyribose (of Dna). this discovery eventually allowed the devel-opment of therapies that used alternative nucleotides to inhibit the growth of retroviruses, such as HiV. the atlantic horseshoe crab (Limulus polyphemus) has blue blood, which contains antimicrobial and antiviral compounds. lysates from horseshoe crabs are used as extremely sensitive detectors of the pres-ence of gram-negative bacterial toxins.
it may turn out that hibernating bears have something to teach medical researchers. (Bears do not actually hibernate; their winter sleep is more properly called denning.) they are inactive for five months, yet they do not suffer bone loss during that time. they do not urinate during that time; their bodies recycle urea into amino acids. they do not develop diabetes, even though they have high blood levels of fat and sugar and low levels of insulin. Solv-
ing the mysteries of how they do this may lead to major medical breakthroughs for humans.
the medicinal compounds are sometimes the result of an ecological relationship between species. Wild dendrobatid frogs make medicinally useful alkaloids in their skins, but they do not do so in captivity. the toxin may actually come from insects that they eat. Sometimes frogs will make the chemi-cals in their food more toxic by concentrating it. an anticancer compound that was found in a marine mollusk turned out to have been produced by sea-weeds eaten by the mollusks—or perhaps even by symbiotic bacteria associated with the seaweeds.
Gathering wild plants, especially rare ones, for pharmaceutical extraction can easily deplete wild populations. three responses to this problem are possible.
● First, researchers may grow the wild plants in plantations or greenhouses, which is now being done on a small scale for the rosy periwinkle, for Pilocarpus jaborandi (source of pilocarpine, a medicine that reduces glau-coma pressure), for Artemisia annua (source of the antimalarial drug artemisinin), and for the african cherry.
● Second, researchers may find a close relative of a plant that is not rare and that may con-tain the same or a related active compound (a european yew now substitutes for the Pacific yew as a source of taxol).
● third, researchers may artificially synthe-size an active compound. artificial synthesis is now the main source of vincristine and vinblastine. Salicylic acid, found in many wild plants, is modified into acetylsalicylic acid as the basis for aspirin.
the same problem arises with animals that pro-duce pharmaceutical compounds. the soft coral Pseudopterogorgia elisabethae (or a symbiont that lives inside it) produces an anti-inflammatory com-pound. With careful management, this coral can rapidly regrow after harvesting to produce a sustain-able source of this compound.
the search for medically potent natural com-pounds is particularly important for the discovery of new antibiotics. according to the medical biol-ogists Michael Fischbach and christopher Walsh, most new antibiotics developed between about 1962 and 2000 have been modifications of old classes of antibiotics (penicillins, cephalosporins, quinolones, macrolids, and tetracyclines). Bacterial populations evolved resistance to older penicillins but were not resistant to modified penicillins. However, the bacte-ria quickly evolve resistance to the new ones because
The rosy periwinkle (Catharanthus roseus) is a wild-flower that lives in Madagascar. The leaves of this plant just happened to contain compounds, such as vincristine and vinblastine, that help control leukemia. (Wikimedia)
xviii+598_Biodiversity_F.indd 67 11/17/11 1:12 PM

first entry68 biodiversity, resources for medicine 68
they are structurally similar to the old ones. the authors call for the development of whole new chem-ical categories of antibiotics, something that was not done during the “innovation gap” of 1962 to 2000. One of the ways that the authors suggested as a source of whole new categories of antibiotics was previously unexplored artificial and natural prod-ucts, including products that may appear to have a function unrelated to killing bacteria. they mention examples such as:
● the first sulfa drug, Prontosil, was origi-nally developed as a dye by Bayer.
● the first quinolone was nalidixic acid, an intermediate in the production of chloro-quine (synthetic quinine).
● the antibacterial action of oxazolidinones was discovered by DuPont, but they were originally developed to treat plant diseases.
For a while, the development of new antibiot-ics from natural sources seemed to be an outmoded approach. as the entire genomes of bacteria were deciphered, researchers believed they could use this information to identify proteins that the bacteria needed to survive and design compounds that would block them. However, noted Fischbach and Walsh, this approach has not yet yielded any new anti-biotics. Furthermore, most existing antibiotics are broad-spectrum, that is, they kill many kinds of bac-teria. this would appear to be a good thing, since the physician may not know what kind of bacterium is causing an infection, but broad-spectrum antibiot-ics kill so many kinds of bacteria that they can leave a patient open to opportunistic infections by oth-erwise harmless bacteria. this is the most frequent cause of Clostridium dificile infections. it appears that the door remains wide open to the discovery of new antibiotics, including narrow-spectrum anti-biotics, from wild organisms—even (as in the oxa-zolidinone example) from plants.
Some medicinally useful compounds have been found in organisms just before they were about to become extinct. epibatidine is a relaxant that has fewer side effects than opiates. Scientists discovered it in the skins of a species of poison frog in ecua-dor. this species survived in only one forest; the surrounding forests had all been made into banana plantations. calanolide a is a powerful inhibitor of reverse transcriptase, an enzyme unique to ret-roviruses such as HiV and therefore a promising anti-aiDS drug. it is known only from one variety of Calophyllum lanigerum (var. austrocoriaceum); other subspecies are not known to produce the com-pound. the last wild member of this subspecies was cut down in Borneo before researchers could return
to it. luckily, living specimens were alive in a tropi-cal botanical garden.
Meanwhile, as tribal peoples are incorporated into modern civilization, they begin to depend more and more on the products of modern agriculture and industry. tribal children in rural areas can iden-tify hundreds of wild plants and know what they are useful for; tribal children in urban areas, like most children in highly industrial countries, know very few plants. tribal knowledge is vanishing much more rapidly than are tribes themselves.
wIld SpECIES ThaT yIEld mEdICal INSIGhTSadaptations of wild species have proven to be valu-able in medical research aside from their production of medicinal compounds. Many have done so by serving as experimental organisms.
the use of laboratory nematodes, fruit flies, mice, rats, and primates has proven indispensible in medical research to benefit humans. Mice and rats are mammals that have most of the same physiologi-cal responses to pharmaceutical compounds that humans do and have been inbred to produce genetic uniformity. Scientists can also produce “knockout mice” in which one version of a gene can knock out another version of the same gene to allow the physiological effects of just that one gene to be stud-ied. nematodes (Caenorhabditis elegans) and fruit flies (Drosophila melanogaster) are much simpler than mammals, but they have many of the same physiological processes. For example, humans have a much more complex nervous system than do fruit flies, but the nerve cells are largely the same. there are mutants of Drosophila that correspond closely to human mutations that contribute to problems with circadian rhythms, aggression, addiction, prema-ture aging (Werner syndrome), Parkinson’s disease, alzheimer’s disease, and Huntington’s disease, to name a few. the genetic basis of these diseases can be studied more easily in fruit flies (which are sim-ple, have rapid generation times, and present no ethi-cal problems for experimentation) than in humans. Some politicians do not realize this. in 2009, the for-mer alaska governor and vice presidential candidate Sarah Palin made a statement in which she ridiculed the use of government funds for fruit fly research (“i kid you not,” she said).
Who can say what new experimental species may be in the wild, its usefulness not yet discovered? One example of an animal that might prove useful in experimentation is the gastric brooding frogs (Rheo-batrachus silus and R. vitellinus). the tadpoles of this species develop while they are protected inside the stomachs of the parents and emerge as small adult frogs. in order to do this, the parents must keep from digesting the offspring, which means that the produc-
xviii+598_Biodiversity_F.indd 68 11/17/11 1:12 PM

biogeochemical cycles 69
tion of pepsin and acid is blocked while the young are brooding. if scientists discover how the frogs do this, they may be able to develop better treatments for some stomach diseases in humans. these frogs were unknown until the 1980s because they are rare and, therefore, in danger of extinction. Hagfishes and lam-preys are useful in the study of the vertebrate immune system, since they are the only vertebrates that have only innate immunity. the ability of nerve cells to regenerate (neurogenesis) was discovered in chicka-dees (Parus atricapillus) when scientists studied the enlargement of their brains that occurred when the breeding season began and the birds began to sing.
Biotechnology research has been revolutionized by the discovery of useful genes in wild species. the GFP (green fluorescent protein) gene, linked to another gene, produces a pigment whenever the gene to which it is linked is activated. a researcher can determine which tissues have the activated gene by shining ultraviolet light on the organism and look-ing for a green glow. this gene was found in the sponge Aequorea victoria. Biotechnology has been revolutionized by the use of a special kind of Dna polymerase that can resist high temperatures. this taq polymerase was first discovered in Thermus aquatilis, a bacterium that lives in hot springs.
Most of the newly discovered wild species that produce medicinal compounds live in tropical rain forests. Many tropical countries are poor. the United nations convention on Biological Diversity recognizes the economic interest these countries have in the development of medicinal compounds from the plants within their borders. Pharmaceutical prospectors and the companies for which they work now operate within strict guidelines that guarantee profit-sharing with the country in which the com-pound was discovered and the native peoples who called attention to it.
See also biodiversity, resources for agri-culture; biomimicry; resistance, evolution of.
Further readingchivian, eric, and aaron Bernstein. Sustaining Life: How
Human Health Depends on Biodiversity. new york: Oxford University Press, 2008.
cragg, G. M., and D. J. newman. “Biodiversity: a continu-ing source of novel drug leads.” Pure and Applied Chemistry 77 (2005): 7–24.
Donia, Mohamed S., et al. “a global assembly line for cyanobactins.” Nature Chemical Biology 4 (2008): 341–343.
Dumbacher, J. P., et al. “Melyrid beetles (choresine): a putative source for the batrachotoxin alkaloids found in poison-dart frogs and toxic passerine birds.” Pro-ceedings of the National Academy of Sciences USA 101 (2004): 15,857–15,860.
engel, S., et al. “antimicrobial activities of extracts from atlantic marine plants against marine pathogens and saprophytes.” Marine Biology 49 (2006): 991–1,002.
Fenical, W., et al. “new anticancer drugs from cultured and collected marine organisms.” Pharmaceutical Biology 41 (2003): 6–14.
Fischbach, Michael a., and christopher t. Walsh. “anti-biotics for emerging pathogens.” Science 325 (2009): 1,089–1,093.
Maxwell, nicole. Witch-Doctor’s Apprentice: Hunting for Medicinal Plants in the Amazon. new york: citadel, 1961.
Moore, K. S., et al. “Squalamine—an aminosterol anti-biotic from the shark.” Proceedings of the National Academy of Sciences USA 90 (1993): 1,354–1,358.
Partida-Martinez, laila P., and christian Hertweck. “Pathogenic fungus harbours endosymbiotic bacteria for toxin production.” Nature 437 (2005): 884–888.
Piel, Jörn. “Bacterial symbionts: Prospects for the sustain-able production of invertebrate-derived pharmaceuti-cals.” Current Medicinal Chemistry 13 (2006): 39–50.
———. “Metabolites from endosymbiotic bacteria.” Natu-ral Product Reports 26 (2009): 338–362.
Plotkin, Mark. Tales of a Shaman’s Apprentice: An Ethno-botanist Searches for New Medicines in the Amazon Rain Forest. new york: Viking, 1993.
Saporito, R. a., et al. “Formicine ants: an arthropod source for the pumiliotoxin alkaloids of dendrobatid poison frogs.” Proceedings of the National Academy of Sci-ences USA 101 (2004): 8,045–8,050.
Schaeffer, D. J., and V. S. Krylov. “anti-HiV activity of extracts and compounds from algae and cyanobac-teria.” Ecotoxicology and Environmental Safety 45 (2000): 208–227.
Stewart, K. M. “the african cherry (Prunus africana): can lessons be learned from an overexploited medicinal tree?” Journal of Ethnopharmacology 89 (2003): 3–13.
Wilson, e. O. The Future of Life. new york: Vintage, 2003.
biogeochemical cycles atoms and mol-ecules circulate endlessly through the ecosystems of earth. these cycles are called biogeochemical because the driving forces are biological processes such as photosynthesis (and, ultimately, evolution), geological processes such as erosion (and, ultimately, continental drift), and chemical processes such as oxidation. the entire concept of biogeochemical cycles is based on the insight that organisms live and populations evolve not as actors on an environmen-tal stage but as participants in the “nonliving” earth itself. Organisms have altered earth into a form very different from what it would have been if life had not evolved or had evolved differently. For example, nearly all the oxygen gas in the atmosphere has come
xviii+598_Biodiversity_F.indd 69 11/17/11 1:12 PM

first entry70 biogeochemical cycles 70
from photosynthesis, and many carbonate deposits are biological in origin. For this reason, some sci-entists prefer to view earth as a single living entity, figuratively referred to as Gaia, rather than as living and nonliving components.
Some of these cycles are complex enough that they have their own entries in this encyclopedia:
● Water molecules change state but remain intact as they circulate through the hydro-logical cycle. Water evaporates, primarily from the oceans, condenses in clouds, and precipitates onto land and ocean. a small amount of the water remains on land in lakes and aquifers. Organisms, primarily plants, play a crucial role in preventing run-off of water from snow and rain, allowing water to percolate into aquifers, and also release much water vapor back into the air during transpiration.
● carbon atoms in atmospheric carbon diox-ide gas enter the food web by photosynthesis, then return to the atmosphere by respiration and decomposition, mostly over the course of tens to hundreds of years. Organisms use carbon atoms to build virtually all their molecules. carbon atoms also enter and are removed from rocks by chemical reactions, mostly over the course of thousands of years (see figure on page 71). carbon dioxide in the atmosphere is an important cause of the greenhouse effect.
● Oxygen atoms circulate through ecosys-tems in two ways. Some remain associated with carbon atoms throughout the carbon cycle. Others enter the atmosphere as oxy-gen molecules when photosynthesis splits water molecules and return to the atmo-sphere when respiration puts those oxygen atoms back into water molecules. Oxygen also reacts with minerals in rocks.
● nitrogen atoms in the form of largely inert nitrogen gas in the atmosphere are made into compounds such as ammonia by some bacteria. the ammonia and other mole-cules derived from it remain in the soil until plant roots remove them, and the nitrogen atoms enter the food web. Organ-isms use nitrogen atoms primarily in their proteins and nucleic acids. Some bacteria release nitrogen gas back into the atmo-sphere from decomposing organic material (see figure on page 71).
Sulfur atoms also participate in biogeochemical cycles with both soil and atmospheric components.
Plant roots absorb sulfate ions (SO42-) from the soil,
and from there the sulfur atoms pass through the food chain, primarily as a component of proteins. Decomposers release sulfate back into the soil. Under anaerobic conditions, decomposers may produce hydrogen sulfide gas (H2S) instead, which enters the atmosphere. Hydrogen sulfide reacts with oxygen to form sulfate, which returns to the ground as sulfuric acid dissolved in rain.
all the previously mentioned cycles have both gaseous and atmospheric components. in contrast, most minerals cycle through ecosystems in solid or dissolved form (see figure on page 71).
Plant roots absorb minerals that are dissolved in soil water. Plant cells use these minerals, which then travel through the food web, where they are used in animal cells and released back into the soil by decomposers. this is a short-term mineral cycle, in which a mineral atom or molecule may complete the cycle in as short a time as a few weeks. Most of the minerals that function in organisms circulate in short-term cycles. the most common of these miner-als are as follows:
● Potassium (K+) ions function primarily in the salt balance of cells, which do not syn-thesize them into larger molecules.
● calcium (ca2+) ions function primarily in assisting the operation of enzymes and are major components of bones (calcium phos-phate), shells (usually calcium carbonate), and plant cell walls.
● Phosphorus atoms are usually found in phosphate ions such as HPO4
2- in the soil. Organisms use phosphorus as a component of nucleic acids as well as in the calcium phosphate of bones.
● Magnesium (Mg2+) ions are found in the center of every chlorophyll molecule.
● iron atoms primarily exist as ferrous (Fe2+) and ferric (Fe3+) ions in the soil. every hemo-globin molecule contains an iron atom in its center, as do proteins that conduct electrons in photosynthesis and respiration.
Minerals may also undergo a long-term mineral cycle, in which a mineral atom or molecule may accumulate in sediment at the bottom of a shallow sea. the sediment is transformed into rock, which, when uplifted above sea level, can erode back into sediments. all the minerals found in short-term cycles can also participate in long-term cycles. cal-cium accumulates along with carbonate in lime-stone deposits. Under anaerobic conditions, sulfur and iron atoms can form pyrite (FeS2) molecules, which accumulate in crystals. When exposed to
xviii+598_Biodiversity_F.indd 70 11/17/11 1:12 PM

biogeochemical cycles 71
oxygen gas, pyrite degrades into iron and sulfate ions (sulfuric acid). Sulfate can be ejected into the atmosphere by volcanic explosions. in both of these ways, sulfur atoms can reenter short-term cycles. Organisms play important roles in long-term min-eral cycles. For example, Acidothiobacillus bacteria considerably speed up the oxidation of pyrite into sulfuric acid.
Minerals that do not play a signifi cant role in most biological systems generally travel through long-term mineral cycles. Silicon atoms exist pri-marily in the form of quartz (SiO2). although some organisms use SiO2 (for example, in the cell walls of grasses), for the most part, silicon remains in the soil until, eventually, that soil is transformed into rock.
in addition to the minerals mentioned, many relatively rare minerals circulate through short- and long-term cycles. Many of these, such as aluminum, zinc, manganese, molybdenum, nickel, and copper, are required in very small quantities to assist some enzyme reactions in cells. (iron also functions in this role.) copper, like iron, is also a common component of proteins that conduct electrons in photosynthesis and respiration. animals require a small quantity of iodine in order for their thyroid glands to work prop-erly. Because they are required in such small quanti-ties, they are called micronutrients. in most cases, soils contain enough micronutrients, although this is not always the case. Humans can get enough iodine by eating seafood or even by breathing sea spray. cer-tain inland locations, however, are defi cient in iodine.
All of the biogeochemical cycles operate through the food chain. The carbon cycle is shown in solid lines. Plants absorb carbon dioxide and produce organic molecules, which pass through the food chain; decomposers release carbon dioxide back into the atmosphere. The nitrogen cycle is shown in dashed lines. Nitrogen-fi xing bacteria, most of which live in the roots of leguminous plants, transform nitrogen gas into fertilizers, which the leguminous plants use to make their proteins. Decomposers release the fertilizers from dead leguminous plants. Other plants absorb nitrogen fertilizers from the soil amd make their own proteins. Animals make their proteins from the proteins in the plants that they eat. The cycle of minerals (such as calcium or potassium) is shown in dotted lines. Plants obtain minerals from the soil, animals obtain minerals by eating plants, and decomposers release minerals from dead plants and animals into the soil.
xviii+598_Biodiversity_F.indd 71 11/17/11 1:12 PM

first entry72 biogeography 72
Micronutrients are generally poisonous at high concentrations. in addition, there are some miner-als, such as arsenic and mercury, that are poisonous at low concentration and have no positive biologi-cal function. Some soils, generally of very limited geographical extent, contain high concentrations of these minerals. an example is serpentine soils (named after the mineral from which they form), which cover about 1 percent of california and much less of the appalachians. these soils have a low ratio of calcium to magnesium. this is important because the same proteins allow plant roots to absorb both calcium and magnesium. as a result, in serpentine soils, the plants absorb too much magnesium and too little calcium. Serpentine soils are also deficient in phosphorus and contain high levels of heavy met-als such as nickel and chromium. as a result, the plants that live in serpentine soils grow slowly and are sometimes of species or varieties (such as the oak Quercus durata and the wildflower Phlox hirsuta) that are specifically adapted to these conditions.
in some cases, biological processes can bring toxic minerals into the food chain and enhance their biogeochemical cycling. Mercury atoms enter the air and water from the erosion of mineral deposits or the burning of coal. Most of the mercury contamina-tion in the food chain, however, occurs in the form of methyl mercury, a biological derivative of ele-mental mercury that is produced by some bacteria. Plant roots also absorb strontium-90, a dangerous product of nuclear fission, because the proteins that absorb calcium cannot distinguish between it and strontium. this is how strontium-90, once it enters the food web, accumulates in the bones of animals. Strontium-90 entered ecosystems as a result of atmo-spheric testing of nuclear weapons.
intact ecosystems with high levels of biodiver-sity are essential to biogeochemical cycles. in some cases, only certain species can transform molecules from one form to another. in particular, only certain bacteria carry out nitrogen fixation, which brings nitrogen atoms into the food chain. as mentioned, only certain plant species can survive in serpen-tine soils. Without these species, serpentine deposits would experience high rates of erosion, thus intro-ducing contaminants into nearby ecosystems. as the ecologist Peter Vitousek and associates (including Jane lubchenco, head of the national Oceanic and atmospheric administration) explain, human activi-ties have greatly altered biogeochemical cycling. this includes not only human impacts on soil ero-sion and global warming but also on the spread of invasive species.
See also carbon cycle; ecosystems; Gaia; greenhouse effect; hydrological cycle; inva-sive species; nitrogen cycle.
Further readingasner, Gregory P., and Peter M. Vitousek. “Remote analysis
of biological invasion and biogeochemical change.” Proceedings of the National Academy of Sciences USA 102 (2005): 4,383–4,386.
Barrett, J. e., et al. “co-variation in soil biodiversity and biogeochemistry in northern and southern Victo-ria land, antarctica.” Antarctic Science 18 (2006): 535–548.
Beare, M. H., et al. “a hierarchical approach to evaluating the significance of soil biodiversity to biogeochemical cycling.” Plant and Soil 170 (1995): 5–22.
DeGraaf, Maaike c. c., et al. “Biodiversity, vegetation gradients and key biogeochemical processes in the heathland landscape.” Biological Conservation 142 (2009): 2,191–2,201.
Gadd, G. M., ed. Fungi in Biogeochemical Cycles. new york: cambridge University Press, 2006.
turner, t. l., et al. “Population resequencing reveals local adaptation of Arabidopsis lyrata to serpentine soils.” Nature Genetics 42 (2010): 260–263.
Vitousek, Peter M., Harold a. Mooney, Jane lubchenco, and Jerry M. Melillo. “Human domination of earth’s ecosystems.” Science 277 (1997): 494–499.
biogeography Biogeography is the study of where organisms live and how they got there. it is an essential part of understanding biodiversity. Biogeography is also inseparable from evolutionary science. in fact, biogeography allowed some initial insights into the fact that evolution had occurred. in The Origin of Species, Darwin noted that major groups of animals lived on certain continents and not on others that had similar climates because they had evolved on those continents. this insight from biogeography was also valuable to Darwin’s friend alfred Russel Wallace, who also discovered natural selection. Wallace noticed that the (primarily placen-tal) mammals of asia were very different from the (primarily marsupial) mammals of new Guinea and australia. this is still one of the best examples of evolutionary biogeography.
Probably the biggest process that has affected bio-geography over the history of life on earth has been continental drift. continents have moved, coalesced into supercontinents, then split apart in different clusters. continental coalescence brought different species into contact for the first time and allowed new species to evolve in response to one another. Subsequent splitting isolated them and allowed them to follow separate evolutionary directions. conti-nental movements are the major explanation for the biogeographic realms, which have largely separate sets of species. Using the classification system of the biogeographer e. c. Pielou, these realms are:
xviii+598_Biodiversity_F.indd 72 11/17/11 1:12 PM

biogeography 73
● nearctic (north america) ● neotropical (central and South america) ● Palaearctic (europe and western asia
including the Mediterranean region) ● ethiopian (africa south of the Mediterra-
nean region) ● Oriental (eastern asia) ● australasian (australia and nearby islands) ● Oceanian (Pacific islands) ● antarctic (antarctica)
not only have the continents moved, but moun-tains have arisen and subsequently eroded away. climatic changes have also created barriers such as deserts between populations that were once in con-tact with one another. Mountains and deserts can separate a population into two or more populations as surely as can an ocean. When populations are separated, one species can become many, a process known as adaptive radiation, and different species can evolve similar adaptations independently of one another in each isolated region, a process known as convergence.
GloBal paTTErNS of vICarIaNCE aNd dISpErSalWhen geographical barriers split a previously uni-fied population or species, allowing speciation, the resulting geographical pattern is due to vicariance. alternatively, individuals of a species can migrate to a new location. the resulting geographical pat-tern is due to dispersal. Since each dispersal event is independent, it is unlikely that broad-scale biogeo-graphic patterns are due to dispersal.
Often, biogeography on a higher taxonomic level (such as class or family) may need to invoke more ancient continental movements and climatic changes, while biogeography on a lower taxonomic level (such as genus or species) may be attributed to more recent continental movements and climatic changes. consider the example of the oak family, Fagaceae:
● the oak family, Fagaceae, apparently evolved soon after the breakup of the super-continent Pangaea at the end of the Meso-zoic era. this family may have been found throughout the supercontinent remnants. Since the northern and southern continents diverged, they have remained largely sepa-rate, except for the connection of africa to eurasia, the collision of the indian subconti-nent into eurasia, and the recent formation of the isthmus of Panama. Because the fam-ily Fagaceae was already in existence, both hemispheres contain members of this plant family. Recently, some scientists have split
the southern members of Fagaceae into their own family, the nothofagaceae.
● the modern genera within the Fagaceae family evolved after the separation of north-ern from southern continents. therefore, oaks (genus Quercus), chestnuts (genus Castanea), and beeches (genus Fagus) are restricted to the northern continents, where they evolved (see table on page 74), while South america and australia have the genus Nothofagus, where it evolved.
● Some 50 million years ago, north american and european forests were continuous with those of northern asia. thereafter, climates began to change, and the northern conti-nents drifted apart. asian, north american, and european oaks were separated from one another. Within north america, the development of the Rocky Mountains and the southwestern desert separated the west-ern oaks (especially in california) from the oaks of the eastern deciduous forest. europe and north america had apparently already separated by the time the red oak subgenus (section Lobatae) and cork oak subgenus (section Cerris) had evolved; red oaks are found only in america and cork oaks only in eurasia. Separate species within the white oak subgenus (section Quercus) are found in america, europe, and eastern asia (see table on page 74).
● Further speciation occurred after the sepa-ration of north america and europe. cali-fornia, separated by mountains and deserts, has a completely separate set of oak species from eastern north america.
Many plant families are largely tropical, and they have remained in the tropical areas of the world as the continents have moved. Some of them have a pantropical distribution, in tropical forests all over the world, while others are found only in the Old World or the new World.
there are numerous examples of families, gen-era, or subgenera of plants that are found only in the northern Hemisphere, for example the oaks, beeches, and chestnuts; elms (genus Ulmus); birches (genus Bet-ula); cottonwoods (genus Populus, section Aegiros); aspens (genus Populus, section Populus); and maples (genus Acer) (see table on page 74). there are fewer examples of families, genera, or subgenera restricted to the temperate regions of the Southern Hemisphere in the manner of Nothofagus. this may be because the cold climates of the Southern Hemisphere are not as cold as those of the northern Hemisphere. there is almost no land surface in the Southern Hemisphere
xviii+598_Biodiversity_F.indd 73 11/17/11 1:12 PM
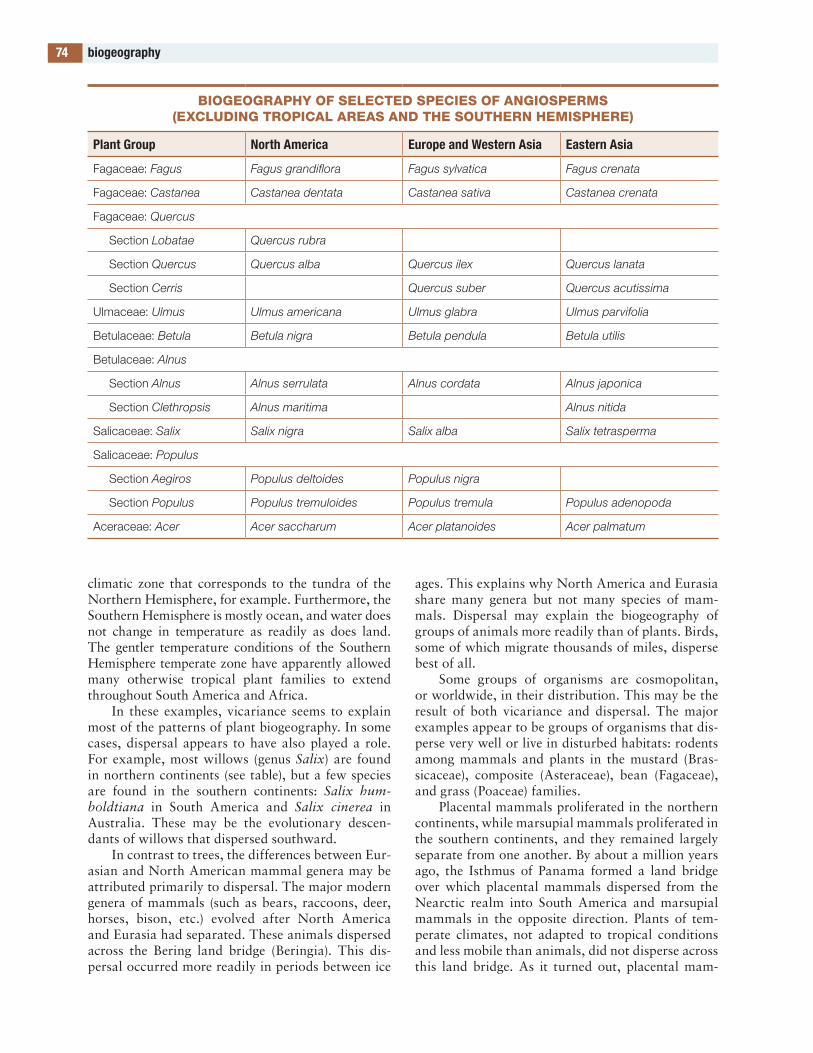
first entry74 biogeography 74
climatic zone that corresponds to the tundra of the northern Hemisphere, for example. Furthermore, the Southern Hemisphere is mostly ocean, and water does not change in temperature as readily as does land. the gentler temperature conditions of the Southern Hemisphere temperate zone have apparently allowed many otherwise tropical plant families to extend throughout South america and africa.
in these examples, vicariance seems to explain most of the patterns of plant biogeography. in some cases, dispersal appears to have also played a role. For example, most willows (genus Salix) are found in northern continents (see table), but a few species are found in the southern continents: Salix hum-boldtiana in South america and Salix cinerea in australia. these may be the evolutionary descen-dants of willows that dispersed southward.
in contrast to trees, the differences between eur-asian and north american mammal genera may be attributed primarily to dispersal. the major modern genera of mammals (such as bears, raccoons, deer, horses, bison, etc.) evolved after north america and eurasia had separated. these animals dispersed across the Bering land bridge (Beringia). this dis-persal occurred more readily in periods between ice
ages. this explains why north america and eurasia share many genera but not many species of mam-mals. Dispersal may explain the biogeography of groups of animals more readily than of plants. Birds, some of which migrate thousands of miles, disperse best of all.
Some groups of organisms are cosmopolitan, or worldwide, in their distribution. this may be the result of both vicariance and dispersal. the major examples appear to be groups of organisms that dis-perse very well or live in disturbed habitats: rodents among mammals and plants in the mustard (Bras-sicaceae), composite (asteraceae), bean (Fagaceae), and grass (Poaceae) families.
Placental mammals proliferated in the northern continents, while marsupial mammals proliferated in the southern continents, and they remained largely separate from one another. By about a million years ago, the isthmus of Panama formed a land bridge over which placental mammals dispersed from the nearctic realm into South america and marsupial mammals in the opposite direction. Plants of tem-perate climates, not adapted to tropical conditions and less mobile than animals, did not disperse across this land bridge. as it turned out, placental mam-
biogeography of seleCted speCies of angiosperms (exCluding tropiCal areas and the southern hemisphere)
plant Group North america Europe and western asia Eastern asia
Fagaceae: Fagus Fagus grandiflora Fagus sylvatica Fagus crenata
Fagaceae: Castanea Castanea dentata Castanea sativa Castanea crenata
Fagaceae: Quercus
Section Lobatae Quercus rubra
Section Quercus Quercus alba Quercus ilex Quercus lanata
Section Cerris Quercus suber Quercus acutissima
Ulmaceae: Ulmus Ulmus americana Ulmus glabra Ulmus parvifolia
Betulaceae: Betula Betula nigra Betula pendula Betula utilis
Betulaceae: Alnus
Section Alnus Alnus serrulata Alnus cordata Alnus japonica
Section Clethropsis Alnus maritima Alnus nitida
Salicaceae: Salix Salix nigra Salix alba Salix tetrasperma
Salicaceae: Populus
Section Aegiros Populus deltoides Populus nigra
Section Populus Populus tremuloides Populus tremula Populus adenopoda
Aceraceae: Acer Acer saccharum Acer platanoides Acer palmatum
xviii+598_Biodiversity_F.indd 74 11/17/11 1:12 PM

biogeography 75
mals frequently drove marsupial mammals toward extinction by competition. the placental mammals of South america, now dominant, are the descen-dants of invaders from north america. Most of the marsupials that moved north into north america became extinct except for the opossum. Meanwhile, australia and new Guinea remained separate and the territory of marsupials. Few placental mammals made the journey into the australasian realm. One exception was humans. Homo erectus ventured into the islands of indonesia and became Java Man, but there is no clear evidence that they traveled to aus-tralia. Some may have become isolated on the island of Flores, evolving into the diminutive species Homo floresiensis. Modern humans dispersed to australia at least 50,000 years ago and brought dogs with them. For thousands of years, humans and dogs were the main placental mammals of australia. aus-tralian marsupials included many species that were similar to placental species but that had evolved separately. europeans brought placental mammals, such as rodents and livestock, with them. Once again, the placental mammals proved to be the supe-rior competitors. Most of the australian marsupials, except for various species of kangaroo, have become rare or extinct, largely as a result of competition with the placental invaders.
When a continental fragment separates, it may sometimes retain a collection of species that closely resemble their ancestors because they are not exposed to many of the newly evolved continental species. this appears to be what happened with the island of new caledonia. the forests consist largely of conifers more similar to those that domi-nated Mesozoic forests than to those that dominate modern forests. this is why film crews who want a realistic backdrop for dinosaur movies choose new caledonia for their film site. the species of flower-ing plant that most closely resembles the presumed ancestor of all flowering plants, Amborella trichop-oda, is found only on new caledonia. new caledo-nia is one of the biodiversity hot spots and contains many more unique plant species per unit area than any of the other hot spots.
Some distribution patterns remain mysterious. One example is the plant family empetraceae (crow-berries), which are found in the cool regions of north america and eurasia but also at the cool tip of South america and in the warm climate of Florida. this is difficult to explain in terms of con-tinental movements or dispersal to another habitat with similar climatic conditions.
ISlaNdSislands, starting with Darwin’s experience in the Galápagos islands, have proven to be evolutionary
showcases. When a new island forms, it receives only a genetic subset of organisms from the nearby main-land. in particular:
● in any species some of whose members dis-perse to a new island, only a small por-tion of the mainland genetic variability may be represented on the island. this founder effect may be followed by random genetic changes in the population (genetic drift) if the island population remains small.
● the newly arrived species may have fewer parasites and perhaps no predators or com-petitors. this may, if the species happens to be well suited to the physical environment of the island, allow a population explo-sion, leading to evolutionary change and diversification.
● Many small mammals remain small partly because their small size allows rapid popula-tion growth, giving them an advantage in competition, but on an island with fewer or no competitors, the small mammals may evolve into larger forms. the same thing appears to happen in plants. Many small, weedy plants have an advantage due to rapid growth and early reproduction. in this way, they avoid competition with large trees. When the plants disperse to an island where there are no trees, the weedy plants may evolve into a tree form. the plant family asteraceae consists mostly of weedy herba-ceous plants, except on some islands (such as the channel islands of california and St. Helena in the atlantic), where they have evolved into small trees. the nettle family Urticaceae consists mostly of weedy herba-ceous plants, but on Hawaii there is a bush nettle.
● Many large mammals remain large partly because their large size allows them to resist the attacks of predators, but on an island with no predators, the large mammals may evolve into smaller forms. Mammoths on Wrangel island, northeast of Siberia, and on Santa Rosa island, off the coast of cali-fornia, evolved into smaller forms. another example is the previously mentioned Flores island people, Homo floresiensis. Some large mammals evolved into smaller forms on this island at the same time that the Flores island people lived there; the predator exclusion explanation, however, is unlikely to be true for the small size of these people. the explanation for the small size of the Flores island people remains a mystery.
xviii+598_Biodiversity_F.indd 75 11/17/11 1:12 PM

first entry76 biogeography 76
● Many island species lose their defenses. Since defenses (spines or chemicals in plants and behaviors in animals) can be expen-sive, natural selection favors their loss in conditions where they are not useful. Many island plants have lost their defenses. For example, not only is the Hawaiian nettle unusual in its family for being a bush but also for its lack of stinging hairs. numerous island animal species (reptiles, birds, and mammals) are notorious for their lack of fear of humans, with whom they have had little contact until recently.
● in order to get to an island in the first place, plants and animals need to have superior dispersal abilities. Once on the island, many lose their dispersal abilities. One extreme example is the flightless cormorant of the Galápagos islands. On some islands, plant families with small seeds have produced species with seeds larger than are normal for the family.
ICE aGESGenera and even species of plants are shared between the arctic and alpine tundras, enough to allow these two regions to both be called tundras. arctic tundra is found around the arctic Ocean, while alpine tundra is on mountaintops. this bio-geographical pattern is explained mostly by the ice ages. When glaciers were at their maximum extent, the arctic tundra formed a band across what is now the northern United States and extended as far south as chicago. the Rocky Mountains, Sierra nevada, and cascades received tundra species from contact with this band of tundra, and some moun-tains that were not in direct contact with the tundra (e.g., the San Francisco Peaks of arizona) were close enough to receive tundra species by dispersal. When the glaciers retreated northward, the tundra plants retreated into the remaining zones of cold climate, either northward, where they are found today in the arctic tundra, or up the mountains, where they are today found on alpine peaks. the alpine tundra is now stranded, sometimes in very small patches (as in the San Francisco Peaks). this is not the entire explanation for tundra plant spe-cies. the tundra of the Sierra nevada contains not only species affiliated with the arctic but also the evolutionary descendants of desert species, such as the buckwheat Eriogonum. the tundra of the Sierra nevada not only has drier soil than other tundras but also is close to the desert. adaptations to cold and to drought are often similar, and this similar-ity allowed some desert plants to adapt to tundra conditions in california. this pattern is also evident
in some animals: the caribou of the cold climates of north america is the same species as the reindeer of the cold climates of eurasia.
Most of the north american continent south of the tundra was boreal (mainly spruce) and pine forests. nearly the entire area that is now the Great Plains was covered with white spruce (Picea glauca). there was a small area of grassland in what is now texas, but there was no hot desert. it would have been an alien world to modern eyes. there were many modern mammals but also many mammals such as mammoths and mastodons that have become extinct. Most of the trees and other plants would have been familiar to modern observers, but they were in combinations that no longer exist. For exam-ple, in some places there were spruce trees scattered in grassland, an arrangement uncommon today.
as the ice sheets retreated, each surviving spe-cies moved to new locations independently, and some species dispersed more rapidly than others. the glacial retreats and the movements of species were not uniform; glaciers sometimes temporar-ily advanced again. the forests are still moving, though too slowly for human observers to notice without consulting long-term records. the tallgrass prairie did not exist before or during the ice ages. the tall, deep-rooted, warm-weather grasses of the prairie were established during the period of maximum warmth (the hypsithermal period, about 7,000 years ago). the prairies persisted even when conditions became cooler and wetter until ameri-can agriculture and civilization destroyed most of them. the prairies persisted because periodic fires killed any forest trees that began to encroach. in north america, where most mountain ranges run in a north-south orientation, there were few barri-ers to the primarily northward movement of plant and animal species. in europe, however, the Medi-terranean Sea and mountain ranges such as the alps were barriers to northward movement. this helps explain why British forests have fewer spe-cies of trees and spring wildflowers than do many american forests. the explanations for current pat-terns of distribution and diversity are rooted in the past rather than in modern climatic conditions. the greenhouse effect may be altering the patterns of species movement that have been occurring for the past few millennia.
Humans have had a tremendous effect on bio-geographical patterns. First, humans have carried species of plants and animals from one place to another. in some cases, these plants and animals have become invasive, displaced native species, and greatly altered the original environment. Humans have allowed species to disperse, especially by ship and airplane, more effectively than almost any spe-
xviii+598_Biodiversity_F.indd 76 11/17/11 1:12 PM

biological classification 77
cies could previously have dispersed. Second, humans have created a great deal of disturbed habitat (such as farming, road building, and construction). Spe-cies that specialize in disturbed areas (especially weeds) have found a worldwide network of suitable habitats, courtesy of humankind. Some of the most widespread species, such as dandelions and barn swallows, live in areas altered by human activity. Finally, humans are causing a greenhouse effect that greatly exceeds any of the periods of global warming that have occurred between the ice ages.
See also adaptive radiation; boreal and subalpine forests; coevolution; convergence; Darwin, charles; founder effect and genetic drift; invasive species; island biogeography; mammals; Pleistocene extinction; speciation; tundra; Wallace, alfred Russel.
Further readingBonnicksen, thomas M. America’s Ancient Forests: From
the Ice Age to the Age of Discovery. new york: John Wiley and Sons, 2000.
cox, c. Barry, and Peter D. Moore. Biogeography: An Eco-logical and Evolutionary Approach. Hoboken, n.J.: John Wiley, 2010.
Gillespie, Rosemary G. “the ecology and evolution of Hawaiian spider communities.” American Scientist 93 (2005): 122–131.
Grehan, John R., and Jeffrey H. Schwartz. “evolution of the second orangutan: Phylogeny and biogeography of hominid origins.” Journal of Biogeography 36 (2009): 1,823–1,844.
international Biogeography Society. “the international Biogeography Society.” available online. URl: http://www.biogeography.org. accessed July 4, 2010.
MacDonald, Glen M. Biogeography: Space, Time, and Life. new york: John Wiley and Sons, 2003.
Mccarthy, Dennis. Here Be Dragons: How the Study of Animal and Plant Distributions Revolutionized Our Views of Life and Earth. new york: Oxford Univer-sity Press, 2009.
Médail, Frédéric, and Katia Diadema. “Glacial refugia influence plant diversity patterns in the Mediter-ranean Basin.” Journal of Biogeography 36 (2009): 1,333–1,345.
Pielou, e. c. Biogeography. Malabar, Fla.: Krieger, 1992.Quammen, David. Song of the Dodo: Island Biogeogra-
phy in an Age of Extinctions. new york: Scribner, 1996.
Whittaker, Robert J. Island Biogeography: Ecology, Evolu-tion, and Conservation. new york: Oxford University Press, 1998.
Woodruff, David S., and leslie M. turner. “the indochi-nese-Sundaic zoogeographic transition: a description and analysis of terrestrial mammal species distribu-tions.” Journal of Biogeography 36 (2009): 803–821.
biological classification the system used by most biologists to classify species of organisms into nested categories was invented by the Swedish botanist carl von linné, also known as carolus lin-naeus. Taxonomy is the study of classification.
Before linnaeus, the classification of organisms was in disarray. First, each species was usually iden-tified by long latin descriptions. For example, the common ground cherry was called Physalis amno ramosissime ramis angulosis glabirs foliis dentoser-ratis, which means, in part, that it had angled stems and that the leaves were hairless and had toothed margins. linnaeus shortened these long names to two-word names (in this case, Physalis angulata, a name it still carries). thus, linnaeus established the practice of binomial nomenclature (“naming system that uses two names”).
Second, the same species could be given differ-ent descriptive names by different botanists. lin-naeus standardized the names. Was Rosa sylvestris alba cum rubore, folio glabro, the same as Rosa syl-vestris inodora seu canina? linnaeus standardized it to Rosa canina. Mistakes still happen, in which the same species is accidentally given two different names, but this error is uncommon.
these innovations were accepted by the bio-logical community with a sense of relief, but this was not the most important innovation of the linnaean system. Before linnaeus, there was no recognizable basis for classification. Some zoologists classified animals on the basis of terrestrial versus aquatic, others on the basis of big versus small. the French paleontologist Georges Buffon organized animals according to their usefulness in the human economy.
linnaeus brought order into this chaos. in classi-fying plants, linnaeus recognized that reproductive characteristics were less variable than the charac-teristics of leaves and stems. He therefore classified plants on the basis of the numbers and condition of the stamens (male parts) and pistils (female parts) of their flowers. consistent with the male bias of science and society, linnaeus’s major axis of clas-sification was the number of stamens: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 12, and many individual stamens; the grouping of the stamens (tetradidymous, dididy-mous, monadelphous, diadelphous, polyadelphous); the fusion of stamens to one another or to pistils; and plants with separate male and female flowers on the same tree (monoecious) or different trees (dioe-cious) or a mixture of arrangements. His last class, the cryptogams (crypto- means hidden), consisted of plants that had no obvious flowers. Within the male axis, he divided plants on the basis of their female parts. He applied his system in a rigid fashion that resulted in classifications that are surprising to modern botanists. For example, he placed oaks,
xviii+598_Biodiversity_F.indd 77 11/17/11 1:12 PM

first entry78 biological classification 78
beeches, hazelnuts, and cattails into the same cat-egory; modern botanists recognize cattails as very different from the other three.
the central feature of the linnaean system is that it is a nested hierarchy. the specific epithet (the second of the two words in the name) referred to the species, and the genus name (the first word) was given to a set of closely related species. as the system was modified by later scientists, closely related gen-era were clustered into families, closely related fami-lies into orders, closely related orders into classes, closely related classes into phyla, and phyla into the major kingdoms, of which there were originally only two: plants and animals. linnaeus inspired the sci-entific world because he brought, or revealed, order within the diversity of organisms on earth, which was just then being thoroughly explored.
the linnaean system seemed to correspond closely to the concept of the Great chain of Being, also known as the scala naturae. all things, from rocks to plants to animals to humans to angels to God, were believed to be connected like a tapes-try of fibers. Scientists were not surprised to find intermediate entities—for example, a sponge as intermediate between animal and rock, or beings halfway between humans and apes—because they were all part of the created tapestry. linnaeus had no thought of the evolutionary origins of organisms. this is revealed by the fact that he applied his sys-tem to rocks as well as to organisms. linnaeus tried to make the system as natural as possible by using characteristics that were a realistic reflection of the chain of Being.
While linnaeus gave no thought to evolution, the fact that the natural world could be so read-ily classified into nested categories struck charles Darwin as being a major piece of evidence favor-ing evolution. Despite naturalists’ belief in the scala naturae, there actually were not many intermedi-ates between modern entities. Sponges are not really half-rock, half-animal. Dogs, wolves, coyotes, and foxes form a group of similar canines; cats, lions, tigers, leopards, and cheetahs form a group of simi-lar felines; and there are no cat-dogs. Organisms, as linnaeus classified them, display a pattern like branches of an evolutionary tree.
today, biologists attempt to classify organ-isms using the linnaean system but try to make the system as natural as possible by defining genera, families, orders, classes, and phyla on the basis of evolutionary divergence. the species themselves are recognized on the basis of reproductive isolation. When a group of species share many features with one another, scientists conclude that they evolved from a common ancestor more recently than species that share few features.
the latin (or Greek) generic and specific names may describe the characteristics that make the genus or species different from other species or genera; they may describe visible features of the organism or the location in which it lives; they may be the ancient Greek or latin names for the organisms; or they may be a tribute to a scientist who studied the organ-isms. For example, dogs are Canis familiaris—genus Canis, species familiaris. Canis is a latin word for “dog.” Other members of the genus Canis include Canis lupus, the wolf (lupus is latin for “wolf”), and Canis latrans, the coyote (latrans comes from latin for “barking”).
this system of groupings also explains the higher levels of the linnaean system (see figure on page 79):
● Genera that evolved from a common ances-tor most recently and are therefore most similar to one another are grouped together into the same family. the canid family, can-idae, contains all the doglike animals, such as foxes, along with the true dogs.
● Families that evolved most recently from a common ancestor and are therefore most similar to one another are members of the same order. canids, along with cats, bears, raccoons, weasels, badgers, skunks, and otters, are members of the order carnivora, the carnivores. the families in the carnivore order share common features such as the kinds of teeth they possess.
● Orders that evolved most recently from a common ancestor and are therefore most similar to one another are members of the same class. carnivores, along with all other mammals, are members of the class Mam-malia. the orders within the mammal class share common features such as the produc-tion of hair and milk.
● classes of animals and protists that evolved most recently from a common ancestor and are therefore most similar to one another are members of the same phylum (plural, phyla). in plants, fungi, and bacteria, this category has traditionally been called a divi-sion. Mammals, along with all other verte-brates (most of which possess a backbone), and those other animals that have a flexible rod instead of a backbone are members of the phylum chordata.
● Phyla or classes of organisms that evolved most recently from a common ancestor and are therefore most similar to one another are members of the same kingdom. Most biologists recognize five kingdoms of organisms.
xviii+598_Biodiversity_F.indd 78 11/17/11 1:12 PM

biological classification 79
in recent years, biologists have classified king-doms into three domains. two of the domains, often placed into the kingdom Monera, consist of single-celled organisms commonly called bacteria:
● the archaebacteria (domain archaea) live in environments such as deep ocean vol-canic vents and salt slush that, while they seem extremely inhospitable to humans, are similar to the conditions found on the early earth.
● the eubacteria are mostly the bacteria that live in soils and water and on or in larger organisms on earth’s surface. Some of them are green and make food energy from sun-light by photosynthesis.
the other three kingdoms consist of organisms that either are, or are constructed from, a more com-plex form of cell called eukaryotic, and make up the domain eucarya:
● the protists are either single-celled or, if multicellular, are not organized into fully integrated bodies.
● the fungi secrete digestive enzymes, then absorb the digested food molecules individ-ually from the environment.
● Plants make food from small molecules in the environment using energy from the Sun.
● Animals consume food as particles or chunks rather than as individual molecules.
the protists do not all have a common ances-tor that differs from the common ancestor of fungi, plants, and animals. therefore, it is not a mono-phyletic group. Many biologists no longer recognize the kingdom Protista as a formal category. instead, they divide all of life into three domains; the fungi, plants, and animals are kingdoms within one of these domains.
early in the 20th century, some biologists applied the linnaean system at least roughly to com-munities, particularly of the plants that dominated the landscape. therefore, to ecologists such as Fred-erick clements, a forest of pines (genus Pinus) was quite literally a Pinetum, and an oak forest (genus Quercus) was a Quercetum. By the middle of the 20th century, however, biologists realized that each species of organism had its own geographical range and set of habitat preferences. a pine forest was not a coherent entity but rather a collection of interact-ing species that happened to be in the same place.
Scientists have described more than 1 million species. they have had to extensively modify the linnaean system in order to accommodate the aston-ishing diversity of the natural world. For example, some large families must be broken into subfamilies and tribes, and orders into infraorders; groups of orders are superorders. that is, the linnaean system still works only because of extensive fixes. Some evolutionary scientists have decided the linnaean system is not worth all the bother. For example, the tree of life project Web site classifies many of the known species of organisms within nested groups (each connected to the next by hyperlinks), but the
The chordates and the arthropods are two of the phyla within the animal kingdom. The mammals are one of the classes within the chordate phylum. Carnivores and primates are two of the orders within the mammalian class. The canids (dogs, wolves, and coyotes) and the felids (lions, cheetahs, and housecats) are two families within the carnivore order.
xviii+598_Biodiversity_F.indd 79 11/17/11 1:12 PM

first entry80 biological species concept 80
levels do not correspond rigidly to the linnaean system. in its classification of the flowering plants, for example, it separates Amborella trichopoda from a branch (or clade) containing all other flowering plants. Within this other branch, one branchlet con-tains the families chloranthaceae, ceratophyllaceae, and the monocot, magnoliid, and eudicot groups—a mixture of families and clusters of orders. it is pos-sible to fit this into a linnaean system, but only by placing Amborella by itself in its own order and family. evolutionary biologists may assign a name to each clade, regardless of whether it is a species, genus, or higher level of classification. they refer to this system as “rank-free” classification. the inter-national Society for Phylogenetic nomenclature is working on an evolution-based system, called Phy-locode, to replace the linnaean system. Some have even suggested that the species concept should be replaced with “least-inclusive taxonomic unit.” the Phylocode approach is not without its critics, one of whom wrote an article entitled “critique of pure folly.” Some Phylocode critics, such as the biodiver-sity expert edward O. Wilson, see it as too much of a disruption right at the time that scientists are try-ing to identify as many species as possible.
See also cladistics; Darwin, charles; eukaryotes; speciation.
Further readingBenton, Michael J. “Stems, nodes, crown clades, and rank-
free lists: is linnaeus dead?” Biological Reviews 75 (2000): 633–648.
cantino, P. D., et al. “Species names in phylogenetic nomen-clature.” Systematic Biology 48 (1999): 790–807.
Dayrat, B., et al. “Suggestions for a new species nomencla-ture.” Taxon 53 (2005): 485–591.
DeQueiroz, Kevin, and J. Gauthier. “Phylogenetic taxon-omy.” Annual Review of Ecology and Systematics 23 (1992): 449–480.
Dubois, a. “naming taxa from cladograms: a caution-ary tale.” Molecular Phylogenetics and Evolution 42 (2007): 317–330.
Fisher, K. “Rank-free monography: a practical example from the moss clade Leucophanella (Calymperaceae).” Systematic Botany 31 (2006): 13–30.
international Society for Phylogenetic nomenclature. “international Society for Phylogenetic nomencla-ture.” available online. URl: http://phylonames.org. accessed august 8, 2011.
Kastner, Joseph. “an adam and his apostle.” chap. 2 in A Species of Eternity. new york: Dutton, 1977.
nixon, Kevin c., et al. “the Phylocode is fatally flawed, and the ‘linnaean’ system can easily be fixed.” Botani-cal Review 69 (2003): 111–120.
Pleijel, F., and G. W. Rouse. “least-inclusive taxonomic unit: a new taxonomic concept for biology.” Proceed-
ings of the Royal Society of London B (Biology) 267 (2000): 627–630.
Polaszek, a., and edward O. Wilson. “Sense and stability in animal names.” Trends in Ecology and Evolution 20 (2005): 421–422.
Schuh, R. t. “the linnaean system and its 250-year persis-tence.” Botanical Review 69 (2003): 59–78.
tree of life Web Project. “explore the tree of life.” avail-able online. URl: http://www.tolweb.org. accessed august 8, 2011.
biological species concept See speciation.
biomagnification Biomagnification is the increasing concentration of toxic compounds in the tissues of organisms higher on the food chain. these compounds do not quickly break down upon expo-sure to environmental conditions such as sunlight and oxygen, nor are they readily broken down by enzymes in the tissues of organisms. in fact, the reduced ability of enzymes to break them down is what makes these compounds toxic. Some of these pollutants are elemental contaminants such as mer-cury, and others are organic molecules known as persistent organic pollutants (POPs). Biomagnifica-tion differs from bioaccumulation, which is the stor-age of toxins in the tissues of an organism.
the pollutants may be present in the environ-ment at concentrations sufficiently low that direct human contact with them is harmless. Organisms at the base of the food chain (for example, microscopic aquatic plants) may accumulate these toxins to a higher concentration because they metabolize the nontoxic compounds from the environment but store the toxins in their tissues. at each level of the food chain, an organism consumes its food, metabolizes the nontoxic compounds, and may accumulate the toxins. this causes the toxins to reach a higher con-centration in the tissues of organisms with each level in the food chain. One of the functions of the liver is to break down toxic materials that are absorbed into the bloodstream, particularly toxic materials that are part of the food absorbed by the intestines and delivered to the liver by the hepatic portal vein. toxins may accumulate in the liver rather than being eliminated by the kidneys.
One famous example of biomagnification and one of the earliest to be discovered involved the pesticide DDt, which was sprayed extensively to control mosquitoes (see figure on page 81). DDt lev-els were particularly high in wetlands, but still not high enough to cause harm to humans upon direct exposure. Ospreys, which are birds of prey, eat large fish from wetlands. in the 1960s, biologists noticed
xviii+598_Biodiversity_F.indd 80 11/17/11 1:12 PM
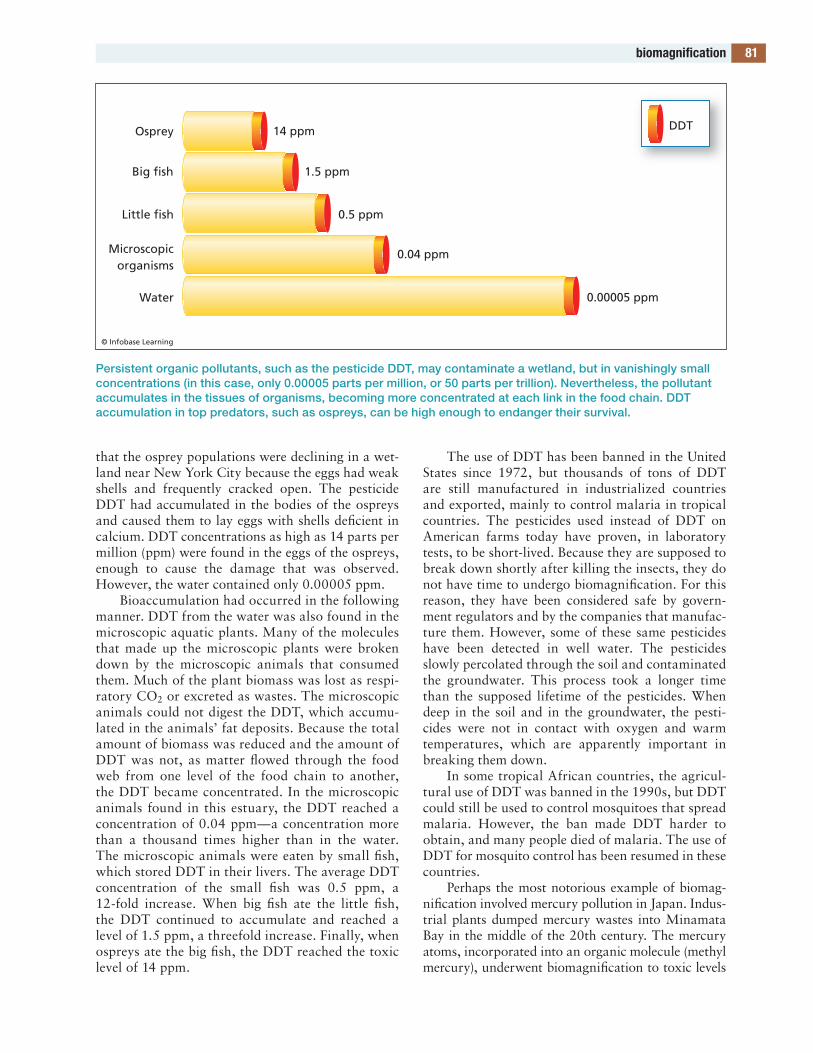
biomagnifi cation 81
that the osprey populations were declining in a wet-land near new york city because the eggs had weak shells and frequently cracked open. the pesticide DDt had accumulated in the bodies of the ospreys and caused them to lay eggs with shells defi cient in calcium. DDt concentrations as high as 14 parts per million (ppm) were found in the eggs of the ospreys, enough to cause the damage that was observed. However, the water contained only 0.00005 ppm.
Bioaccumulation had occurred in the following manner. DDt from the water was also found in the microscopic aquatic plants. Many of the molecules that made up the microscopic plants were broken down by the microscopic animals that consumed them. Much of the plant biomass was lost as respi-ratory cO2 or excreted as wastes. the microscopic animals could not digest the DDt, which accumu-lated in the animals’ fat deposits. Because the total amount of biomass was reduced and the amount of DDt was not, as matter fl owed through the food web from one level of the food chain to another, the DDt became concentrated. in the microscopic animals found in this estuary, the DDt reached a concentration of 0.04 ppm—a concentration more than a thousand times higher than in the water. the microscopic animals were eaten by small fi sh, which stored DDt in their livers. the average DDt concentration of the small fi sh was 0.5 ppm, a 12-fold increase. When big fi sh ate the little fi sh, the DDt continued to accumulate and reached a level of 1.5 ppm, a threefold increase. Finally, when ospreys ate the big fi sh, the DDt reached the toxic level of 14 ppm.
the use of DDt has been banned in the United States since 1972, but thousands of tons of DDt are still manufactured in industrialized countries and exported, mainly to control malaria in tropical countries. the pesticides used instead of DDt on american farms today have proven, in laboratory tests, to be short-lived. Because they are supposed to break down shortly after killing the insects, they do not have time to undergo biomagnifi cation. For this reason, they have been considered safe by govern-ment regulators and by the companies that manufac-ture them. However, some of these same pesticides have been detected in well water. the pesticides slowly percolated through the soil and contaminated the groundwater. this process took a longer time than the supposed lifetime of the pesticides. When deep in the soil and in the groundwater, the pesti-cides were not in contact with oxygen and warm temperatures, which are apparently important in breaking them down.
in some tropical african countries, the agricul-tural use of DDt was banned in the 1990s, but DDt could still be used to control mosquitoes that spread malaria. However, the ban made DDt harder to obtain, and many people died of malaria. the use of DDt for mosquito control has been resumed in these countries.
Perhaps the most notorious example of biomag-nifi cation involved mercury pollution in Japan. indus-trial plants dumped mercury wastes into Minamata Bay in the middle of the 20th century. the mercury atoms, incorporated into an organic molecule (methyl mercury), underwent biomagnifi cation to toxic levels
Persistent organic pollutants, such as the pesticide DDT, may contaminate a wetland, but in vanishingly small concentrations (in this case, only 0.00005 parts per million, or 50 parts per trillion). Nevertheless, the pollutant accumulates in the tissues of organisms, becoming more concentrated at each link in the food chain. DDT accumulation in top predators, such as ospreys, can be high enough to endanger their survival.
xviii+598_Biodiversity_F.indd 81 11/17/11 1:12 PM

first entry82 biome 82
in the local seafood. Many people experienced per-manent nerve damage from eating the seafood in the 1960s.
Bioaccumulation of radioactive wastes occurred in reindeer that ate plants contami-nated with the fallout from the nuclear plant that exploded at chernobyl, in the Ukraine, in 1986. Unlike Minamata in the 1960s, the Ukraine in the 1980s avoided extensive human illness from bioac-cumulation because the concept had become well established and health officials were able to prevent the consumption of contaminated food following the release of radioactive wastes from the cher-nobyl nuclear disaster.
the biologist Rachel carson brought intense and rapid public attention to biomagnification, as well as other problems associated with the use of pesticides, in her 1962 book Silent Spring. a decade later, the United States banned the use of DDt. Most nations now recognize that biomagnifications and other pollution problems cross national boundar-ies and that international agreements are necessary in order to control the problem. the United States was one of 90 signatories to the Stockholm conven-
tion of 2001, which limited the production, use, and release of the 12 most abundant POPs, often called “the dirty dozen” (see table). environmental activists continue to oppose the construction of new coal-fired power plants because the combustion of coal releases a small amount of mercury—a small amount that can be magnified by the food chain. Organic chemicals that are not on the “dirty dozen” list, such as endosulfan, can persist long enough in the environment to penetrate into ecosystems far from their point of release.
See also biogeochemical cycles; carson, Rachel.
Further readingenvironmental Protection agency. “Persistent organic pol-
lutants: a global issue, a global response.” available online. URl: http://www.epa.gov/oia/toxics/pop.html. accessed august 8, 2011.
Hu, Guo-cheng, et al. “Bioaccumulation behavior of poly-brominated diphenyl ethers (PBDes) in the freshwater food chain of Baiyangdian lake, north china.” Envi-ronment International 36 (2010): 309–315.
Kidd, Karen a., et al. “Biomagnification of DDt through the benthic and pelagic food webs of lake Malawi, east africa: importance of trophic level and carbon source.” Environmental Science and Technology 35 (2001): 14–20.
Molina, carlos israel, et al. “transfer of mercury and methylmercury along macroinvertebrate food chains in a floodplain lake of the Beni River, Bolivian ama-zonia.” Science of the Total Environment 408 (2010): 3,382–3,391.
Morel, François M. M., et al. “the chemical cycle and bio-accumulation of mercury.” Annual Review of Ecology and Systematics 29 (1998): 543–566.
Weber, Jan, et al. “endosulfan, a global pesticide: a review of its fate in the environment and occurrence in the arctic.” Science of the Total Environment 408 (2010): 2,966–2,984.
biome See communities, ecological.
biomimicry Biomimicry is the human imitation of the adaptations of wild species for commercially useful purposes. the invention of new processes and products for medicine, industry, and agricul-ture is limited by the knowledge and imagination of researchers. in contrast, natural selection has been operating for billions of years in millions of species; relative to human knowledge, natural selection has tried out almost everything that can be done and eliminated the things that did not work well within the range of conditions in the biosphere.
the tWelve persistent organiC pollutants Controlled by the 2001
stoCkholm Convention
pollutant use/Source
Aldrin insecticide
Chlordane insecticide
Dichlorodiphenyltrichloro-ethane (DDT)
insecticide
Dieldrin insecticide
Endrin insecticide
Heptachlor insecticide
Hexachlorobenzene fungicide, industrial chemical, combustion
Mirex insecticide
Toxaphene insecticide
Polychlorinated biphenyls (PCBs)
industrial chemical, combustion
Polychlorinated dibenzo-p-dioxins (dioxins)
combustion
Polychlorinated dibenzo-furans (furans)
combustion
Note: From U.S. Environmental Protection Agency
xviii+598_Biodiversity_F.indd 82 11/17/11 1:12 PM

bioremediation 83
the Swiss engineer George de Mestral closely observed the barbs surrounding cocklebur seeds in the 1940s, which appeared perfectly adapted to entangle mammalian fur. He realized that the sys-tem of microscopic hooks and hairs could be imi-tated industrially. the result was Velcro, marketed in 1953. Most of the possibilities of biomimicry remain untapped. Below are some products of bio-logical evolution that have had or may soon have great importance to the human economy.
● computers use binary digital code, while Dna in cells uses a digital code of four kinds of molecules, which vastly increases the potential to store and process information. the development of Dna computers, which would operate in a test tube (in vitro) rather than on a chip (in silico), currently has major hurdles, as it is difficult to control the move-ment and interaction of the Dna molecules.
● chloroplasts use photosynthesis to convert solar energy to chemical energy with higher efficiency than any industrial process. chlo-roplasts produce sugar, while solar energy technology needs to produce electricity. Researchers may be able to design artificial chloroplasts that produce electricity.
● Geckos (a kind of lizard) can walk on walls and ceilings, but they do so without chemi-cal adhesives. Geckos form very short-lived bonds when electrons in the molecules of their feet displace electrons in the molecules of a wall or ceiling. temporary adhesion that does not leave chemical residues may have very important industrial applications.
● the surfaces of the eyes of some moths admit light very efficiently, without reflec-tion. Understanding the physical and chemi-cal basis of moth eye surfaces may allow the production of much more efficient photo-graphic imaging.
● Spider silk is one of the strongest materials known.
● a beetle in Brazil, Lamprocyphus augustus, has crystals in its iridescent green scales that focus light more precisely than can industri-ally produced photonic crystals. Studying this beetle may allow the development of photonic crystals that will allow the produc-tion of ultrafast optical computers.
Scientists can even gain insights from the study of ecosystems that may allow the more efficient management of commercial systems. the agricul-tural ecologist Wes Jackson has demonstrated that it is possible to produce a much more efficient system
of agriculture (which he calls natural systems agri-culture) by imitating the ecological processes of a tallgrass prairie.
the direct application of biological adaptations to human use may be difficult or impossible in some cases. Many biological processes occur on a micro-scopic scale and will not work the same way on the human scale. Spider silk is stronger than steel for mak-ing very small fibers, but not necessarily for making larger structures. Partly for this reason, the promise of strong yet pliant spider silk body armor has thus far remained elusive. Similarly, it is unlikely that organ-isms much larger than geckos could walk on walls and ceilings using only the forces of electron displacement.
natural adaptations have been successful in the global environment. in contrast, many human indus-trial processes are destructive to the environment in the long term. Biomimicry may allow the develop-ment of processes that bring the human economy into better compliance with the ecological limita-tions of the biosphere.
along with the benefits to agriculture and medi-cine, biodiversity offers the promise of biomimicry. this is yet another reason to save as many species as possible from extinction. While some adaptations, such as Dna, are found in all organisms, many oth-ers may have evolved in only one obscure species. Who would have guessed that the key to improved photonic crystals might have been found in a Brazil-ian beetle? in addition, just as spider silk is stronger than silkworm silk, a breakthrough for the indus-trial production of yet stronger fibers might be hid-den in the silk produced by some unknown species of rain forest spider.
See also biodiversity, resources for agri-culture; biodiversity, resources for medi-cine; natural systems agriculture.
Further readingayoub, nadia a., et al. “Blueprint for a high-performance
biomaterial: Full-length spider dragline silk genes.” Public Library of Science ONE 2 (2007): e514. avail-able online. URl: http://www.plosone.org/article/info:doi/10.1371/journal.pone.0000514. accessed July 3, 2010.
Benyus, Janine M. Biomimicry: Innovation Inspired by Nature. new york: William Morrow, 1997.
Mueller, tom. “Biomimetics: Design by nature.” National Geographic, (april 2008): 68–91.
biophilia See environmental psychology.
bioremediation Bioremediation is the use of organisms to break down or remove pollution from
xviii+598_Biodiversity_F.indd 83 11/17/11 1:12 PM

first entry8484 birds
the environment, such as organic pollutants or heavy metals such as lead, chromium, cadmium, nickel, zinc, copper, selenium, arsenic, and even radioactive metals such as cesium and uranium. chemists gen-erally restrict the use of the word to the biological breakdown, rather than accumulation, of pollutants.
Some natural bacteria, such as Alcanivorax bor-kumensis, live in sediments that contain natural petroleum seepage, and they have evolved the ability to break down oil. Other bacteria can break down compounds that are not found in nature. they do this because they have enzymes that break down chemically similar natural compounds. examples include bacteria that break down vinyl chloride and pesticides.
Some natural species are very good at accumu-lating toxic ions. examples include the following:
● indian mustard (Brassica juncea) ● alpine pennycress (Thlaspi caerulescens) ● Sunflower (Helianthus annuus) ● lichen (Trapelia involucrata) ● Fungi (Phanerochaete chrysosporium and
Gloeophyllum sp.) ● Brakefern (Pteris ensiformis) ● Water hyacinth (Eichhornia crassipes)
Brakefern and water hyacinth absorb arsenic, and the Trapelia lichen absorbs uranium. Other wild species have a significant, though lesser, ability to accumulate toxic ions: alfalfa, cordgrass, tomato, pumpkin, bamboo, willow, poplar, and kudzu vines.
Some wild plants absorb heavy metal ions pre-cisely because they are toxic. they store the ions in vacuoles, where they will not damage the tissues, but if a herbivore eats the leaf, the vacuoles burst open and make the food poisonous. locoweeds of the genera Oxytropis and Astragalus accumulate selenium in leaf vacuoles, a toxin against herbivores.
Without bioremediation, it is nearly impossible to detoxify soil that has been polluted with indus-trial chemicals without destroying the structure of the soil and turning it into a big adobe brick on which most plants cannot grow. Plants that absorb toxins, however, penetrate the soil with their roots and leave it undamaged. the stems and leaves of the plants, which contain most of the toxins, can then be harvested and disposed of safely.
Some biologists have produced genetically modi-fied strains of plants that have an enhanced ability to absorb toxins. in nature, the plants absorb only as much toxin as they need for defense, but biologists can manipulate the plant’s genes such that they over-express the genes involved in absorption.
Success stories include the following. chrys-ler corporation used mustard plants to reduce the
amount of lead from contaminated soil at its industrial complex in Detroit by half, bringing it into compliance with pollution laws. Sunflowers have been used to absorb radioactive metals from areas around cher-nobyl, the site of the 1986 Ukrainian nuclear disaster.
See also biogeochemical cycles; bio-magnification.
Further readingal agely, a., et al. “Mycorrhizae increase arsenic uptake by
the hyperaccumulator chinese brake fern (Pteris vit-tata l.).” Journal of Environmental Quality 34 (2005): 2,181–2,186.
arthur, e. l., et al. “Phytoremediation—an overview.” Crit-ical Reviews in Plant Sciences 24 (2005): 109–122.
connell, S., and S. al-Hamdani. “Selected physiological responses of kudzu to different chromium concentra-tions.” Canadian Journal of Plant Science 81 (2001): 53–58.
Mitra, J., et al. “Bioremediation of DDt in soil by geneti-cally improved strains of soil fungus Fusarium solani.” Biodegradation 12 (2001): 235–245.
Ralebitso, t. K., et al. “Microbial aspects of atrazine deg-radation in natural environments.” Biodegradation 13 (2002): 11–19.
Revkin, andrew. “new pollution tool: toxic avengers with leaves.” New York Times, March 6, 2001. available online. URl: http://www.nytimes.com/2001/03/06/science/ new-pollution-tool-toxic-avengers-with-leaves.html. accessed august 4, 2011.
Singh, O. V., et al. “Phytoremediation—an overview of metallic ion decontamination from soil.” Applied Microbiology and Biotechnology 61 (2003): 405–412.
Soudek, P., et al. “laboratory analyses of cs-137 uptake by sunflower, reed, and poplar.” Chemosphere 55 (2004): 1,081–1,087.
Vidali, M. “Bioremediation: an overview.” Pure and Applied Chemistry 73 (2001): 1,163–1,172.
birds Birds are vertebrates that have feath-ers. Feathers not only allow birds to fly but keep them warm. Feathers can have spectacular colors that function in sexual reproduction (see figure on page 85). Feathers are the defining characteristic of birds. One of the earliest birds, Archaeopteryx, which lived about 150 million years ago during the Jurassic period, had a skeleton indistinguishable from those of other theropod dinosaurs. it would have been classified as an ordinary dinosaur had it not been for impressions of feathers that were found in the rocks along with its fossilized bones. indeed, some Archaeopteryx specimens without the feather impressions had been misclassified in this way. Since birds are one evolutionary lineage among the dinosaurs, many biologists consider
xviii+598_Biodiversity_F.indd 84 11/17/11 1:12 PM

birds 85
birds to actually be dinosaurs. Once feathers had evolved, bird evolution continued rapidly. another bird fossil from about this same period had a large breastbone, short tail, and perching feet, all of them features found in birds today. there were many bird lineages during the Jurassic and cre-taceous periods, all of which (including the one with Archaeopteryx) became extinct except one, the ancestral lineage of all modern birds.
aNaTomICal fEaTurES ThaT allow BIrdS To fly
Birds possess many features that enable them to fly. these features provide a strong surface for push-ing against the air, enhance birds’ metabolism, and reduce birds’ weight.
Feathers. Feathers emerge from the skin of all birds and form most of birds’ outer covering. they apparently evolved from reptile scales, as the early stages of development in both feathers and scales are the same. Both are made of keratin, the same protein that predominates in the skin tissue of mammals. Feathers have numerous functions:
● they provide a very strong and light-weight surface that allows flight.
● they provide insulation by trapping air pockets. this is important because birds are warm blooded and have a high metabolic rate. Most birds are small and would lose significant warmth through their body sur-faces were it not for feathers.
● they repel water, which would add to a bird’s weight during flight. the outer feath-ers are layered like shingles on a roof to shed water. During preening, many birds transfer oil from an oil gland to the feathers, making them even more water repellent.
Contour feathers cover the outside of an adult bird’s body. they have a central shaft. Many of the shafts grow out of the bones, leaving small indenta-tions that can be identified in fossils. these indenta-tions are sometimes the only evidence that a dinosaur had feathers. On either side of the shaft, vanes con-sist of hundreds of barbs, which branch into thou-sands of barblets, which are connected together by
Many birds have colorful feathers, such as this peacock. (Pavo cristatus) (Duisberg Zoo)
xviii+598_Biodiversity_F.indd 85 11/17/11 1:12 PM

fi rst entry86 birds 86
thousands of tiny hooks. the most important fl ight feathers are called primaries and grow on the ends of the wings. Bird wings provide lift because the air moves faster and therefore has a lower pressure on the upper side. the feathers come together on the downstroke to force the bird upward but move apart on the upstroke to avoid pushing the bird downward. Flight feathers have a central quill with barbs projecting from the quill; barbules with hooks interlock the barbs (see fi gure). During preening, birds often zip disconnected barbules back together. the vanes are asymmetrical: the outer (leading) vane is smaller than the inner vane, which further contributes to air movements that provide lift. Some contour feathers are modifi ed into bristles.
Down feathers line the trunk of a bird’s body. Very weak, their function is insulation. the fi rst feath-ers to evolve may have resembled down feathers and functioned as insulation rather than as fl ight feathers.
Respiration. Birds breathe very effi ciently. their lungs are structurally different from those of all other living vertebrates. Most vertebrates breathe in and then breathe out, but in birds the air fl ows in only one direction, exposing the lung tissues to a continuous supply of fresh air. Birds do not have a diaphragm muscle; the entire body acts like bel-lows, pushing the air through the respiratory sys-tem. alligators also have lungs in which air fl ows in one direction. this implies that unidirectional lungs were an adaptation shared by the common ancestors of dinosaurs, pterosaurs, and alligators.
in addition to unidirectional air fl ow, birds have rapid heartbeats (up to 1,200 beats per minute in hummingbirds) that allow the blood to carry oxygen rapidly to all parts of the body.
Digestion. Because they have no teeth, birds must swal-low their food whole, sometimes regurgitating indigestible por-tions. they have crops in which to store food and gizzards, which are muscular stomachs that may use small stones and other hard objects to help grind food. the stones are called gastroliths;large, smooth gastroliths have also been found associated with the skeletons of large herbivorous dinosaurs. in birds, as in these dinosaurs, the use of gastroliths reduces the need for heavy teeth to grind food and thus reduces weight. (Of course, gastroliths add to the bird’s weight.)
Water balance. Most birds drink by dipping their beaks in water and raising their heads, allowing the water to fl ow down their throats. Most doves, in contrast, can drink water by swallowing, as mam-mals do. Water is heavy, and birds do not carry a lot of it around with them. Birds eliminate nitrogenous wastes in the form of solid uric acid rather than urea dissolved in water. the white nitrogenous wastes mix with dark intestinal wastes and leave the bird’s body through a single opening, called a cloaca.
Bones. the very large sternum (breastbone) allows the attachment of large fl ight muscles. Birds have partially hollow bones, which greatly reduces the weight relative to the solid bones of other verte-brates. Modern birds have no teeth, though ancient birds did. their beaks, similar to hardened lips, allow exploitation of different types of food: some for cracking large seeds, some for gathering small seeds or insects. Hummingbirds have long beaks and tongues that allow them to obtain nectar from tubu-lar fl owers. Modern birds do not have tail vertebrae, although early birds did. Modern birds have tail feathers rather than tails.
Muscles. Birds have more muscle, relative to body weight, than almost any other group of ani-mals. about 40 percent of the body weight of a dove consists of wing muscle.
Temperature regulation. Birds produce internal body heat (are endothermic) and regulate their body temperatures (are homeothermic). their body tem-peratures are higher than those of humans and most
Flight feathers have a central shaft and barbs that are held together by hooks. Feathers are extremely strong relative to their light weight.
xviii+598_Biodiversity_F.indd 86 11/17/11 1:12 PM

birds 87
other mammals. this is an important feature in pro-viding the high levels of energy needed for flight. at rest, a bird’s metabolic rate is 10 times higher than the metabolic rate of a mammal of the same size. Small birds must produce more body heat per unit body weight than do large birds in order to main-tain a constant warm body temperature. Small birds must eat 30 percent and large birds 5 percent of their body weight each day. For comparison, a 150-pound (70-kg) human would have to eat up to 45 pounds (20 kg) of food per day in order to “eat like a bird”! Some birds eat three times their body weight per day during cold seasons. Bird metabolism is so high and so expensive in terms of food energy that very small birds cannot maintain a constant body tempera-ture all the time. Some hummingbirds, for example, allow their body temperatures to drop to about 60°F (14°c) on cool nights.
Sexual reproduction and life cycle. Sexual reproduction in birds is well adapted to life in the air and on dry land.
● the reproductive organs of birds remain reduced in size except during the breeding season, and even then in many species the females have only a left ovary. this allows a great reduction in weight during flight. Most birds do not have external reproductive organs. Some male birds, such as waterfowl, turkeys, and ostriches, have penises that remain hidden except during copulation.
● Birds typically achieve internal fertilization by pressing their cloacae together. Bird eggs have calcium carbonate shells with small pores that allow air to diffuse in but prevent the rapid evaporation of water. the yolk is the egg cell itself and contains many nutri-ents. the albumin, or “egg white,” consists mostly of protein for nourishment of the embryo. as a result, the gametes and the embryo are protected from drying.
Some humans have attempted to fly under their own power by building winglike structures. these have proven unsuccessful largely because birds, unlike humans, are very lightweight and have an enormous amount of muscle tissue. Still, a combination of light-weight materials and pedal-powered propellers has allowed some very muscular humans to fly significant distances. the distance record is 74 miles (115 km), and the speed record is about 20 miles (about 30 km) per hour for about 1 mile (about 1.5 km).
Early BIrdSBirds, like lizards, snakes, crocodiles, tuataras, and dinosaurs, are in the diapsid lineage, which means
that their skulls have two openings on each side of the skull, in contrast to the anapsids (turtles), which have no such openings, and the synapsids (mammals and their ancestors), the earliest members of which lineage had a single opening. in the late Paleozoic era, the diapsids split into two evolutionary lineages: the lepidosaurs (lizards, snakes, tuataras, and others) and the archosaurs (crocodiles, dinosaurs, and others). there were many important lineages of archosaurs during the triassic period, but only the crocodile and dinosaur lineages have survived. Dinosaurs evolved into many forms during the Jurassic and cretaceous periods. Some, the theropods, were predators. Many (except for the tyrannosaurs) were small and ran rap-idly on the ground, occasionally jumping into the air or gliding from tree to tree. these were the ancestors of birds. Since birds arose from a lineage of dinosaurs, modern taxonomists consider birds to be dinosaurs. the common ancestor of all bird lineages had feathers.
Several lineages of primitive birds, such as Archaeopteryx, Confuciusornis, Ichthyornis, and Hesperornis, eventually became extinct. the surviv-ing lineage diverged into the paleognath and neog-nath lineages (see table on page 88).
modErN BIrdSthe following description focuses on the major lin-eages of birds and is not a complete list of orders. all the orders and the number of species in each are included in the table on page 88. the groupings fol-low the 2008 publication by Shannon J. Hackett (see Further Reading source on page 90).
Paleognaths. the paleognaths are represented today by ostriches in africa, rheas in South amer-ica, cassowaries in Papua new Guinea, emus in australia, kiwis in new Zealand, and tinamous in South america. two other paleognath lineages, the elephant birds of Madagascar and the moas of new Zealand, became extinct recently when humans first took up residence on the islands they inhabited. Paleognath means “old jaw,” which refers to a palate structure more similar to that of reptile lineages than are the palates of other birds.
all paleognaths are or were flightless except the tinamous, which fly weakly and slowly. if the common ancestor of paleognaths was flightless, the tinamous must have regained the ability to fly. it is more likely that all these paleognath lineages inde-pendently lost the ability to fly. Flightless paleog-naths still have wings but use them for things such as mating displays. Kiwi wings are very small.
all paleognaths live in the Southern Hemi-sphere on continents that were once connected. this suggests that they evolved before the formation of Pangaea preceding the Mesozoic era, although fos-sils that old have not been found.
xviii+598_Biodiversity_F.indd 87 11/17/11 1:12 PM
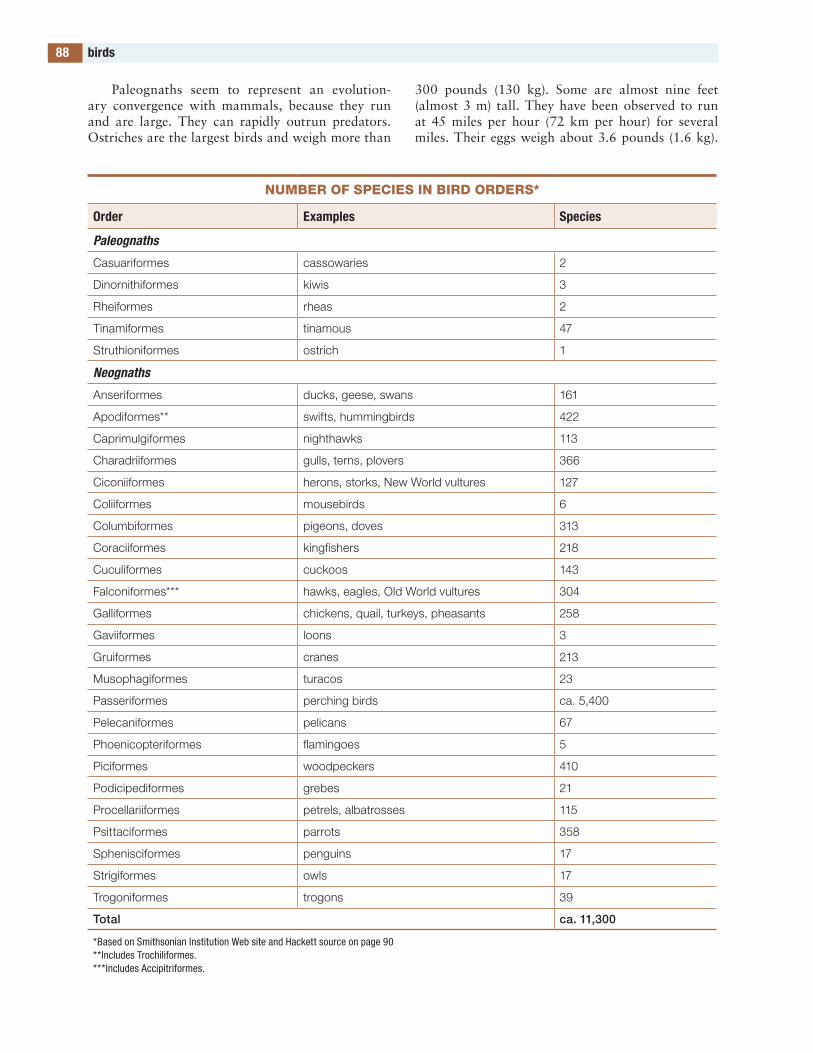
first entry88 birds 88
Paleognaths seem to represent an evolution-ary convergence with mammals, because they run and are large. they can rapidly outrun predators. Ostriches are the largest birds and weigh more than
300 pounds (130 kg). Some are almost nine feet (almost 3 m) tall. they have been observed to run at 45 miles per hour (72 km per hour) for several miles. their eggs weigh about 3.6 pounds (1.6 kg).
number of speCies in bird orders*
order Examples Species
Paleognaths
Casuariformes cassowaries 2
Dinornithiformes kiwis 3
Rheiformes rheas 2
Tinamiformes tinamous 47
Struthioniformes ostrich 1
Neognaths
Anseriformes ducks, geese, swans 161
Apodiformes** swifts, hummingbirds 422
Caprimulgiformes nighthawks 113
Charadriiformes gulls, terns, plovers 366
Ciconiiformes herons, storks, New World vultures 127
Coliiformes mousebirds 6
Columbiformes pigeons, doves 313
Coraciiformes kingfishers 218
Cuculiformes cuckoos 143
Falconiformes*** hawks, eagles, Old World vultures 304
Galliformes chickens, quail, turkeys, pheasants 258
Gaviiformes loons 3
Gruiformes cranes 213
Musophagiformes turacos 23
Passeriformes perching birds ca. 5,400
Pelecaniformes pelicans 67
Phoenicopteriformes flamingoes 5
Piciformes woodpeckers 410
Podicipediformes grebes 21
Procellariiformes petrels, albatrosses 115
Psittaciformes parrots 358
Sphenisciformes penguins 17
Strigiformes owls 17
Trogoniformes trogons 39
Total ca. 11,300
*Based on Smithsonian Institution Web site and Hackett source on page 90**Includes Trochiliformes.***Includes Accipitriformes.
xviii+598_Biodiversity_F.indd 88 11/17/11 1:12 PM

birds 89
the kiwis resemble mammals exceptionally well. new Zealand has no native mammals except bats, and kiwis evolved into a mammalian mode of liv-ing. Kiwis, unlike most birds, are nocturnal and use scent to locate prey. even their feathers resemble mammal fur.
Neognaths. neognaths include the following lineages.
One group of neognaths consists of order anser-iformes (ducks, geese, swans, etc.) and the order Gal-liformes (fowl, quail, turkeys, pheasants, etc.). the ducks, geese, and swans are large birds with long necks and spatulalike bills and are among the most abundant and noticeable members of freshwater eco-systems. Most of them swim with webbed feet and forage by sticking their necks underwater. they are typically migratory. Some swans mate for life. Geese are important in some tundra ecosystems because they consume so much plant material and produce fertilizer. Some geese have been seen migrating over the Himalayas, the highest mountains in the world. Members of order Galliformes (fowl, quail, turkeys, and pheasants) fly slowly and near the ground, pre-ferring to walk as they forage on seeds and insects. the most well-known fowl is the red jungle fowl Gallus gallus, which has been domesticated into the chicken. the chicken is perhaps the most energy-efficient producer of animal protein, in the form of meat and eggs, and as such is extremely important to billions of poor people around the world. While 54 calories of fossil fuel is required to produce a calorie of beef, only 26 are required for a calorie of eggs, and only four for a calorie of chicken meat.
another group contains the order apodiformes (hummingbirds and swifts). Hummingbirds live pri-marily off nectar, and they beat their wings up to 70 times per second to hover in front of tubular flowers into which they insert their long bills. Because they are so small, they are often unable to maintain their body temperatures at night, and they enter a brief version of hibernation. Swifts are small birds with short bodies and narrow wings that extend beyond the tail. they eat insects with their short bills and are best known for nesting in chimneys.
another group contains the order columbi-formes (doves and pigeons). Pigeons and the more slender doves have small legs, heads, and beaks. they are strong fliers. they reproduce rapidly and adapt readily to human habitats. Urban pigeons come from the wild european rock dove, Columba livia. Doves and pigeons are almost the only birds that can drink water by suction. Perhaps because of their gentle “cooing,” doves have been symbols of peace for millennia.
another group contains the orders Procellari-iformes (petrels and albatrosses), Pelecaniformes
(pelicans), ciconiiformes (herons, storks, new World vultures), Spheniciformes (penguins), and Gaviiformes (loons). Most are water birds that eat fish, which they may store (as pelicans do) in their sac-like bills. Penguins cannot fly; their wings are modified as flippers. they are famous for survival in the antarctic even during the winter. new World vultures evolved separately from the Old World vultures, which are related to hawks and eagles (see below). like them, the new World vultures have featherless heads, which prevents an excessive buildup of decaying flesh when they stick their heads into carcasses.
another group contains the order charadri-iformes (sandpipers, plovers, and gulls). Sandpipers and plovers feed on the seashore by inserting their bills, of various lengths, into the sand. Gulls and terns live mostly on seashores and catch fish or crabs or eat carrion. they seldom travel over the open ocean. Seagulls have followed rivers and canals (especially those with slightly saline water) hundreds of miles inland. the arctic tern (Sterna paradisaea) migrates each year between the arctic and antarctic regions of the world, an annual journey of 22,000 miles (about 36,000 km).
the group that has evolved furthest from the neognath common ancestor contains the following orders:
Order Strigiformes (owls) are nocturnal preda-tors. they have large wings and short tails. their forward-facing eyes allow binocular vision. Serra-tions on the leading edges of their wing feathers minimize the sound they make while flying. Some owls have begun moving into suburban areas, where their rodent-controlling activity is welcome.
Order Falconiformes (falcons) usually includes the accipitriformes (hawks, eagles, and Old World vultures). they are hovering and swiftly flying pred-ators with eyesight far more acute than that of any human. they catch their prey with extremely sharp talons. Peregrine falcons can fly at almost 200 miles per hour (320 km per hour). Old World vultures evolved separately from the new World vultures. like them, the Old World vultures have feather-less heads, which prevents an excessive buildup of decaying flesh when they stick their heads into car-casses. Most recent evidence separates the falcons, which are close relatives of perching birds, from the accipitriformes.
Order Piciformes (woodpeckers) have numerous adaptations for forming holes in the wood of dead branches: very sharp beaks, the ability to pound their beaks rapidly, and feet and stiff tail feathers that prop them firmly against the branch. they have extremely long tongues, which may be as long as their bodies. if, while pecking a hole, a woodpecker
xviii+598_Biodiversity_F.indd 89 11/17/11 1:12 PM

first entry90 bogs, life in 90
encounters an insect burrow, it can insert its barbed tongue several inches and skewer the insect. they also return to old holes to seek insects that may have colonized them. the acorn woodpecker of califor-nia (Melanerpes formicivorus) pecks holes in oak trees, each of which is precisely the right size for storing a particular acorn.
Order Passeriformes (the perching birds) is the largest and most recently evolved order of neog-nath birds. almost half of bird species (by some estimates, 5,400 of 11,240) are passerines. not only do passerines represent the greatest number of species, they often have the largest populations. the number of insects consumed by the billions of passerines in the United States is nearly incalcu-lable, and they keep many insect populations under control that would otherwise cause massive pest outbreaks. they are small birds and specialize on many different food sources, most of them small, such as seeds and insects. Because they can produce a great diversity of vocalizations and colors, it is likely that sexual selection has played an important role in their diversification. examples include the following families.
● corvidae (jays, crows) ● Fringillidae (finches) ● Hirundinidae (swallows) ● icteridae (blackbirds, meadowlarks) ● Mimidae (mockingbirds, thrashers) ● Paradisaeidae (birds of paradise) ● Passeridae (sparrows) ● troglodytidae (wrens) ● turdidae (thrushes, robins) ● tyrannidae (flycatchers)
Birds have sex chromosomes. Male birds have two Z chromosomes, while female birds have a Z and a W chromosome. in paleognaths, considered to be more primitive, the Z and W chromosomes are about the same size, while in neognaths the W chro-mosome is smaller.
See also convergence; dinosaurs; reptiles; sexual selection.
Further readingBenton, Michael. “Dinosaur summer.” chap. 4 in Gould,
Stephen Jay, ed., The Book of Life: An Illustrated History of the Evolution of Life on Earth. new york: norton, 1993.
chiappe, luis M. “Downsized dinosaurs: the evolutionary transition to modern birds.” Evolution Education and Outreach 2 (2009): 248–256.
cornell lab of Ornithology. “all about birds.” available online. URl: http://www.allaboutbirds.org. accessed august 8, 2011.
Dalsgaard, Bo, et al. “Plant-hummingbird interactions in the West indies: Floral specialisation gradients associ-ated with environment and hummingbird size.” Oeco-logia 159 (2009): 757–766.
Dingus, l., and t. Rowe. The Mistaken Extinction: Dino-saur Evolution and the Origin of Birds. new york: Freeman, 1997.
Dyrcz, andrzej, and Konrad Halupka. “Why does the fre-quency of nest parasitism by the cuckoo differ consid-erably between two populations of warblers living in the same habitat?” Ethology 113 (2007): 209–213.
Farmer, c. G., and Kent Sanders. “Unidirectional airflow in the lungs of alligators.” Science 327 (2010): 338–340.
Hackett, Shannon J., et al. “a phylogenomic study of birds reveals their evolutionary history.” Science 320 (2008): 1,763–1,768.
Haroldson, Kurt J., et al. “association of ring-necked pheasant, gray partridge, and meadowlark abundance to conservation Reserve Program grasslands.” Journal of Wildlife Management 70 (2006): 1,276–1,284.
Houston, David. Condors and Vultures. Osceola, Wis.: Voyageur Press, 2001.
Mathiessen, Peter. Birds of Heaven: Travels with Cranes. new york: Farrar Straus and Giroux, 2001.
Ornithological council, national Museum of natural His-tory, Smithsonian institution. “Birdnet: the Ornitho-logical information Source.” available online. URl: http://www.nmnh.si.edu/BiRDnet/. accessed august 8, 2011.
Padian, Kevin and l. M. chiappe. “the origin of birds and their flight.” Scientific American 278 (1998): 38–47.
Paul, Gregory S. Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore, Md.: Johns Hopkins University Press, 2002.
Reichard, Dustin G., and J. Jordan Price. “Species recogni-tion in a vocal mimic: Repetition pattern not the only cue used by northern mockingbirds in discriminating songs of conspecifics and brown thrashers.” Wilson Journal of Ornithology 120 (2008): 717–724.
Short, lester l. The Lives of Birds: Birds of the World and Their Behavior. new york: Henry Holt, 1993.
Snyder, noel, and Helen Snyder. Raptors of North Amer-ica: Natural History and Conservation. Osceola, Wis.: Voyageur Press, 2006.
Sodhi, navjot S., et al. “avian extinctions from tropical and subtropical forests.” Annual Review of Ecology, Evo-lution, and Systematics 35 (2004): 323–346.
tilford, tony. Complete Book of Hummingbirds. Holt, Mich.: thunder Bay Press, 2009.
bogs, life in Bogs are among the most special-ized and unusual of aquatic habitats. Most bogs formed from fragments of glacier left behind at the end of the most recent ice age and are most com-mon in regions that were recently glaciated. they
xviii+598_Biodiversity_F.indd 90 11/17/11 1:12 PM

boreal and subalpine forests 91
are clay-lined bowls of glacial meltwater. Because of the dense clay lining, bogs retain water from rain. Furthermore, they are ombrotrophic because they receive no mineral nutrients from rivers or streams. the water is therefore very low in nutrients.
One kind of plant that thrives in these condi-tions is sphagnum moss, which forms rafts that float on and may entirely conceal the surface of the water. not only is the bog nutrient-poor, the moss releases acid into it, inhibiting the growth of larger plants even further. Moreover, the mat of moss keeps the water beneath it cool. Sphagnum is also known as peat moss.
the only plants that grow in bogs are those that can tolerate cool acidic water with few nutrients. Some plant species specialize on bogs and similar habitats. For example, in the Black Hills, the bog birch (Betula glandulosa) inhabits bogs, while the mountain birch (B. occidentalis) inhabits nearby streamsides. Many bog plants, such as the larch (Larix decidua) and the cotton sedge (Eriophorum vaginatum), are adapted to nutrient-depleted, wet, cool conditions and can be found in the boreal and subalpine forests or the tundra as well as in bogs. indeed, the bog could be considered a southerly remnant of the tundra as vegetation zones shifted northward after the most recent ice age. Other bog plants include heath bushes of the plant fam-ily ericaceae. examples include cranberries and blueberries. around the world, heath bushes (for example, the ones that dominate the heather of Scotland) tolerate acidic soils. Bog bushes may be evergreen, which is an adaptation that prevents the plants from losing the scarce mineral nutrients in their leaves.
the most famous plants of the bog are the carnivorous plants, which trap and digest insects. Pitcher plants (such as Sarracenia purpurea) trap insects in pools of digestive fluid; sundews (such as Drosera rotundifolia) trap insects on sticky hairs. Why do carnivorous plants eat insects? the plants are green and make their own sugar through photo-synthesis. therefore, they do not eat insects in order to obtain food energy. instead, they use the insects as a source of fertilizer, such as nitrates and phos-phates, which are scarce in the acidic moss or soil.
animals are less abundant in bogs than in reg-ular ponds. the acidic conditions make calcium, which is necessary for bones, unavailable. Obstruc-tion of the surface also inhibits the diffusion of oxy-gen from the air into the water. Because of the acidic and low-oxygen conditions, fishes are rare in bogs. amphibians and insects, which have access to the air, may be abundant.
a fen is similar to a bog but contains open water and may receive some minerals from groundwater.
ecological succession may in time fill in the fen and turn it into a bog.
Decomposition is very slow in the cold, acidic waters of a bog. it may therefore hide many ancient mysteries. layers of bog sediments preserve ancient pollen and chemicals that allow the reconstruction of past environments. Human corpses, some thou-sands of years old, have been found well preserved in bogs, although the hair has been bleached and the skin literally tanned like leather. analysis of the corpses of “bog people,” particularly in europe, has helped to reveal facts about the daily lives of people in prehistoric cultures and, in some cases, methods of execution. therefore, bogs, besides being interest-ing microclimates, act as time capsules that preserve bits of the past. Bogs are also disproportionately important to the global atmosphere because their bacterial decomposition may emit large amounts of methane (and other molecules) into the atmosphere.
See also boreal and subalpine forests; car-nivorous plants; ice ages; tundra.
Further readingcadillo-Quiroz, Hinsby, et al. “Diversity and community
structure of archaea inhabiting the rhizoplane of two contrasting plants from an acidic bog.” Microbial Ecology 59 (2010): 757–767.
Krachler, Regina, et al. “Relevance of peat-draining riv-ers for the riverine input of dissolved iron into the ocean.” Science of the Total Environment 408 (2010): 2,402–2,408.
Simmonds, P. G., et al. “natural chloroform emissions from the blanket peat bogs in the vicinity of Mace Head, ireland over a 14-year period.” Atmospheric Environment 44 (2010): 1,284–1,291.
tsujino, Riyō, et al. “Restoration of floating mat bog veg-etation after eutrophication damages by improving water quality in a small pond.” Limnology 11 (2010): 289–297.
Wright, Margaret e., and tom a. Ranker. “Dispersal and habitat fidelity of bog and forest growth forms of Hawaiian Metrosideros (Myrtaceae).” Botanical Jour-nal of the Linnaean Society 162 (2010): 558–571.
bony fishes See fishes.
boreal and subalpine forests Boreal for-ests, also called taiga, cover one-third of the forested area of earth. Boreal comes from Greek for “north-ern.” these forests occur between 50° and 60° north latitudes, which is about the extent of the arctic air mass in winter and summer, respectively. Boreal forests form a broad belt of forest across north america, Scandinavia, and Siberia.
xviii+598_Biodiversity_F.indd 91 11/17/11 1:12 PM

first entry92 boreal and subalpine forests 92
Boreal forests have short, moderately warm sum-mers and long, cold, snowy winters. the growing season is about three months long—only 50 to 100 days are frost free. this is the principal reason that needle-leaved evergreen trees dominate the boreal forests. the summers are too short to allow trees to grow a new set of leaves each year. therefore, none of the dominant tree species is deciduous. Further-more, the leaves must be adapted to surviving long, snowy winters. their needlelike shape allows them to prevent the buildup of snow, the weight of which could damage tree branches. in both of these ways, boreal forests contrast with broad-leaved decidu-ous forests. the trees in boreal forests also have a conical shape that allows them to shed snow in the winter. Summers can, however, be much warmer. Verkoyansk, Siberia, has recorded extremes of tem-perature from -90° to 90°F (about -68° to 35°c). the precipitation does not much exceed that of an arid climate, but there is so little evaporation that the region is humid.
the soils in a boreal forest tend to be thin, nutri-ent poor, and acidic, both because the low tempera-tures do not allow extensive decomposition to form humus and because needle leaves tend to be acidic. the acid leaches minerals from the shallow soils, leaving them gray in color.
the diversity of tree species in boreal forests is much lower than that of temperate forests; spruce (genus Picea) and fir (genus Abies) trees are abundant over large geographical areas (see table on page 93). a few deciduous trees, such as birches (genus Betula), alders (genus Alnus), and aspens (Populus tremuloi-des), grow in disturbed and streamside habitats of the boreal forest. Fires are common in the boreal and subalpine forests. Burned areas are quickly filled by jack pine (Pinus banksiana) and aspens.
Boreal forests were found in southern europe and southern north america during the most recent ice age. When the glaciers retreated, starting about 12,000 years ago, the trees of the boreal forest migrated (through seed dispersal) northward to their present locations and also to the tops of moun-tains, where they exist today as subalpine forests (see figure). this is why many subalpine forests of the Rocky Mountain region are dominated by trees very similar to those of the boreal forest. How-ever, pines dominate some subalpine forests of the Rocky Mountains, especially of the Sierra nevada of california (see table on page 93). these are not the same pine species that occur in lower-eleva-tion montane or other temperate coniferous forests. examples include the limber pine (Pinus garryana) and the bristlecone pines (P. longeava in the Sierras, P. aristida in the Rockies). the bristlecone pines of california grow extremely slowly in the cold, dry
environment and are the oldest known trees in the world, some exceeding 4,700 years of age.
Some subalpine forests also exist in the Southern Hemisphere. One example is the subalpine forests of australia, where the snow gum (Eucalyptus pauci-flora) is a dominant tree species.
Boreal forests intergrade with tundra. a tran-sitional zone that can be 60 miles (almost 100 km) in width connects the arctic tundra and the boreal forest in canada. the boreal forest dominates the warm, south-facing, well-drained slopes in this zone. the numerous bogs in some parts of the taiga (form-ing a mixture known as muskeg) are tundra inclu-sions. the transitional zone between alpine tundra and subalpine forest is characterized by stunted subalpine trees (called krummholz) that are often short enough to be protected by snow in the winter and that may have branches only on the leeward side where winter winds have not killed them. the krummholz trees form a vaguely defined timberline. Boreal forests also intergrade with temperate forests, for example, in northern Minnesota.
to a certain extent, the ecological border between tundra and boreal forest is regulated by the boreal forest itself. Boreal forest trees are dark and become warm by absorbing sunlight. this hastens
Coniferous trees grow in subalpine forests, such as these red firs (Abies magnifica) in the Sierra Nevada of California. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 92 11/17/11 1:12 PM
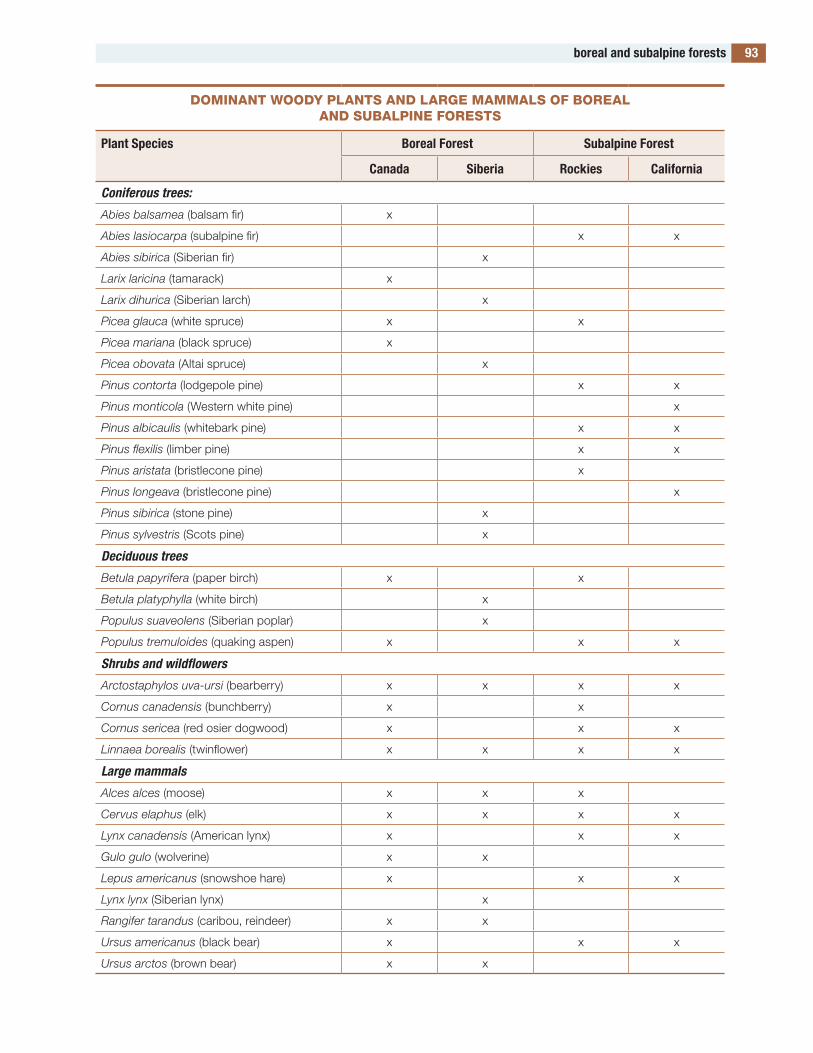
boreal and subalpine forests 93
dominant Woody plants and large mammals of boreal and subalpine forests
plant Species Boreal forest Subalpine forest
Canada Siberia rockies California
Coniferous trees:
Abies balsamea (balsam fir) x
Abies lasiocarpa (subalpine fir) x x
Abies sibirica (Siberian fir) x
Larix laricina (tamarack) x
Larix dihurica (Siberian larch) x
Picea glauca (white spruce) x x
Picea mariana (black spruce) x
Picea obovata (Altai spruce) x
Pinus contorta (lodgepole pine) x x
Pinus monticola (Western white pine) x
Pinus albicaulis (whitebark pine) x x
Pinus flexilis (limber pine) x x
Pinus aristata (bristlecone pine) x
Pinus longeava (bristlecone pine) x
Pinus sibirica (stone pine) x
Pinus sylvestris (Scots pine) x
Deciduous trees
Betula papyrifera (paper birch) x x
Betula platyphylla (white birch) x
Populus suaveolens (Siberian poplar) x
Populus tremuloides (quaking aspen) x x x
Shrubs and wildflowers
Arctostaphylos uva-ursi (bearberry) x x x x
Cornus canadensis (bunchberry) x x
Cornus sericea (red osier dogwood) x x x
Linnaea borealis (twinflower) x x x x
Large mammals
Alces alces (moose) x x x
Cervus elaphus (elk) x x x x
Lynx canadensis (American lynx) x x x
Gulo gulo (wolverine) x x
Lepus americanus (snowshoe hare) x x x
Lynx lynx (Siberian lynx) x
Rangifer tarandus (caribou, reindeer) x x
Ursus americanus (black bear) x x x
Ursus arctos (brown bear) x x
xviii+598_Biodiversity_F.indd 93 11/17/11 1:12 PM

first entry9494 brown algae
the melting of snow in the spring and creates warmer conditions in which the boreal species can grow.
Boreal and subalpine forests are experiencing threats from human activity. logging is very com-mon in boreal forests because of the abundant soft-wood (coniferous) trees with little undergrowth. logging in boreal forests has become a controversial issue, as many ecologists believe these forests are subject to destruction if well-planned management is not practiced. Global warming also threatens boreal and subalpine forests. Since boreal forests have low species diversity, they are subject to outbreaks of insect and other pests. Such outbreaks have been rare because of the short growing season. Global warming has produced longer, warmer summers that have allowed insect outbreaks to kill trees in large areas of alaska. Subalpine forests can survive only on cold mountaintops; if mountaintops become even slightly warmer as a consequence of global warming, montane and deciduous forests may grow all the way to the summits, resulting in the elimina-tion of the subalpine forests. acid rain has weakened the remnant subalpine forest of Fraser firs (Abies fra-seri) in the highest appalachians and allowed them to fall victim to the woolly adelgid bug.
Further readingallison, Steven D., et al. “nitrogen alters carbon dynamics
during early succession in boreal forest.” Soil Biology and Biochemistry 42 (2010): 1,157–1,164.
Grundale, Michael J., et al. “Vascular plant removal effects on biological n fixation vary across a boreal forest island gradient.” Ecology 91 (2010): 1,704–1,714.
Hoffman, ary a., et al. “Phenological changes in six aus-tralian subalpine plants in response to experimental warming and year-to-year variation.” Journal of Ecol-ogy 98 (2010): 927–937.
Josefsson, torbjörn, et al. “linking forest history and con-servation efforts: long-term impact of low-intensity timber harvest on forest structure and wood-inhabit-ing fungi in northern Sweden.” Biological Conserva-tion 143 (2010): 1,803–1,811.
lynch, Wayne. Life in the Boreal Forest. Markham, Ont.: Fitzhenry & Whiteside, 2006.
Rice, Stanley a. “Plants create habitats.” ch. 8 in Green Planet: How Plants Keep the Earth Alive. new Bruns-wick, n.J.: Rutgers University Press, 2009.
Runesson, Ulf t. “Boreal forests of the world.” available online. URl: http://www.borealforest.org/world.htm. accessed May 4, 2009.
tremblay, Junior a., et al. “Foraging ecology of black-backed woodpeckers (Picoides arcticus) in unburned eastern boreal forest stands.” Canadian Journal of For-est Research 40 (2010): 991–999.
brown algae (Phaeophyta) See protists.
burgess shale See cambrian explosion.
xviii+598_Biodiversity_F.indd 94 11/17/11 1:12 PM

95
Cambrian explosion the cambrian explo-sion was the fi rst major eruption of biodiversity in earth history. the cambrian period began about 540 million years ago. life had existed for almost 3 billion years before it. the fundamental components of life had evolved before the cambrian period: metabolism, prokaryotic cells, eukaryotic cells, and multicellularity. Protists, including seaweeds, existed before the cambrian. Fossilized animal embryos and burrows indicate the existence of at least simple animal life before the cambrian, although recent evidence suggests that the burrows may have been produced by a giant protist similar to the modern genus Gromia. immediately before the cambrian period, simple organisms called ediacarans domi-nate fossil deposits worldwide.
However, at the beginning of the cambrian period, a rapid diversifi cation of complex animals, particularly animals with hard skeletons, occurred in the oceans of the world. Because much of this evolution occurred right at the beginning of the cambrian period, it has been called the cambrian explosion. the paleontologist andrew Knoll wrote that the cambrian explosion looked like a “brief moment . . . when all things were possible in animal evolution.” the appearance of new animal forms was rapid, but not sudden: it occurred over a period of about 5 million years.
Some scientists explain the cambrian explosion as the appearance of animals that had previously existed but had not been fossilized. according to this view, complex animals without hard parts had previously existed, although complex animals with hard skeletons had not. Hard skeletons are much more readily preserved in the fossil record. How-ever, organisms without hard parts have often been
preserved in the fossil record, including the Pre-cambrian ediacaran organisms and animal embryos mentioned previously. the cambrian explosion, then, must be explained as the rapid evolution of new animals, not just the evolution of hard skeletons. Some observers have classifi ed ediacaran organisms as relatives of modern animal groups such as cnidar-ians, but even if this is true, the cambrian explosion represents spectacular diversifi cation within those invertebrate groups as well as the origin of new ani-mal groups such as the arthropods.
a number of worldwide environmental changes occurred in the late Precambrian that were necessary for the cambrian explosion, as follows.
● Oxygen gas accumulated in the atmosphere from the photosynthesis of bacteria and eukaryotes. Metabolism that uses oxygen is essential for complex animals. therefore, the explosive evolution of animals could not have occurred before the atmosphere and oceans contained abundant oxygen. Oxy-gen gas accumulated in the atmosphere and then in the oceans long before the cambrian period. For example, the Great Oxidation event of 2.4 billion years ago may have produced enough oxygen to fi lter out most of the ultraviolet radiation from the Sun. nevertheless, modern levels of atmospheric oxygen, suffi cient for metabolism of large animals, may not have been reached until just before the beginning of the cambrian period.
● at least three major worldwide glaciations occurred during the Precambrian. the most recent of them may not have ended until
C
xviii+598_Biodiversity_F.indd 95 11/17/11 1:12 PM

first entry96 Cambrian period, biodiversity in 96
about 600 million years ago. an explosive evolution of animals was unlikely to occur under such cold conditions. the melting of the ice also made more mineral nutrients available to the seaweeds and microscopic protists that were the basis of the marine food chain. However, there was a 60-mil-lion-year delay before the cambrian explo-sion. late Precambrian conditions were certainly suitable for the ediacarans; why not complex animals?
the leading explanation for the cambrian explosion takes its cue from the rapid appearance of animals with hard parts. this explanation sug-gests that the cambrian explosion began with the evolution of predation. the first predators were animals with a feeding apparatus that allowed them to chew their food. Previously, large organisms such as the ediacarans were defenseless but safe. now, any soft-bodied animal was helpless before the predators. the cambrian explosion occurred when animals began to evolve different kinds of defenses as predators or against predators, such as the following.
● natural selection favored many differ-ent structures that permitted predators to search for, catch, and chew their prey.
● Some evolved hard external skeletons. in many cases, this required the evolution of new body configurations. For example, the bivalve body form (as in mussels and oys-ters) could not function without hard shells. Hard shell defenses also evolved in a wide variety of forms. even single-celled photo-synthetic eukaryotes evolved hard and com-plex external coverings.
● Some prey, then as now, had soft bodies but hid from predators by burrowing into the mud. the complexity and depth of animal burrows increased rapidly during the cam-brian explosion.
as a result of these processes, the cambrian world was filled with diverse and complex animals, as exemplified by fossil deposits such as the Burgess Shale in canada and others. animals with internal skeletons did not evolve until the latter part of the cambrian period.
See also cambrian period, biodiversity in.
Further readingBudd, Graham e. and Sören Jensen. “a critical reappraisal
of the fossil record of the bilaterian phyla.” Biological Reviews 75 (2000): 253–295.
conway Morris, Simon. The Crucible of Creation: The Burgess Shale and the Rise of Animals. new york: Oxford University Press, 1998.
Knoll, andrew H. Life on a Young Planet: The First Three Billion Years of Evolution on Earth. Princeton, n.J.: Princeton University Press, 2003.
lane, nick. Oxygen: The Molecule that Made the World. new york: Oxford University Press, 2002.
Rokas, antonis, Dirk Krüger, and Sean B. carroll. “animal evolution and the molecular signature of radiation compressed in time.” Science 310 (2005): 1,933–1,938.
Cambrian period, biodiversity in the cambrian period (540 to 510 million years ago) was the first period of the Phanerozoic eon and the Paleozoic era. During the previous eons, major evolutionary innovations had occurred, such as the origins of prokaryotic cells, of eukaryotic cells, and of multicellular organisms. at the beginning of the cambrian period, complex animal forms rapidly evolved, a burst of biodiversity commonly known as the cambrian explosion. this is why the cambrian period is considered the beginning of the Phanero-zoic eon, or the eon of “visible life.” an average of about 300 families of marine organisms existed during the cambrian, compared to 1,000 marine families (and 2,200 total families) today.
Climate. all cambrian life lived underwater, with the possible exception of lichens on continental rocks. temperatures are much more stable under-water than on land. atmospheric oxygen levels were much lower and carbon dioxide levels much higher than today. Global temperatures were also higher than today (see table on page 97).
Continents. Because there were no continents located near the poles, ocean currents were able to circulate freely. as a result, there was no significant formation of ice sheets. Sea level was more than 180 feet (about 60 m) higher than today.
Marine life. Most and perhaps all of the major animal phyla evolved before or during the cambrian period (see figure on page 97). Fossil deposits such as the Burgess Shale in canada, chengjiang in china, and Sirius Passet in Greenland preserve fossils of many animal phyla, including those that had no hard parts. these deposits reveal a somewhat uniform worldwide assemblage of animals that persisted for about 30 million years. the animals within these phyla were different from the ones within those phyla today. For example, the arthropods were represented by trilobites and by animals such as Anomalocaris, a large predator with a round mouth, both of which have long been extinct. the first vertebrates, jawless fishes, evolved during the cambrian period. the 530-million-year-old chengjiang deposit in china
xviii+598_Biodiversity_F.indd 96 11/17/11 1:12 PM

Cambrian period, biodiversity in 97
contains two jawless vertebrate fossils, Myllokun-mingia and Haikouichthys, considered to be the fi rst fi shes. conodont animals existed as far back as 540 million years ago. these fi shlike animals had long bodies, large eyes, and conelike teeth but no jaws. Single-celled and multicellular photosynthetic pro-tists (such as seaweeds) were the basis of the marine food chain.
Continental life. there is no clear evidence that either animals or plants existed on land or in freshwater during the cambrian period. the con-tinents consisted of bare rock, sand, silt, and clay. Moist rocks may have supported green algae and lichens.
Extinctions. although the cambrian period is not considered a time of mass extinction, the edia-
caran organisms and other Precambrian life forms apparently became extinct during this period. there were far fewer species in the cambrian period than in the following Ordovician period, but, on a percentage basis, cambrian extinctions were considerable.
See also arthropods; cambrian explosion; fishes.
Further readingBenton, Michael. “Diversifi cation and extinction in the his-
tory of life.” Science 268 (1995): 52–58.Briggs, Derek e. G., et al. The Fossils of the Burgess Shale.
Washington, D.c.: Smithsonian Books, 1994.conway Morris, Simon. The Crucible of Creation: The
Burgess Shale and the Rise of Animals. Oxford: Oxford University Press, 2000.
Gould, Stephen Jay. Wonderful Life: The Burgess Shale and the Nature of History. new york: norton, 1989.
Hou, Xian-Guang, et al. The Cambrian Fossils of Chengji-ang, China: The Flowering of Early Animal Life. new york: Wiley-Blackwell, 2004.
multiCellular biodiversity in the Cambrian period
Conditions
Atmospheric oxygen 63% of modern level
Atmospheric carbon dioxide 13 times modern level
Global temperature 9°F (7°C) higher than today
Sea level 190 feet (60 m) higher than today
Approximate Number of Families*
Continental 0
Marine 100
Total 300
Examples of Marine Animals
Anomalocaris canadensis arthropod
Diagonella hinderi chordate
Fallotapsis longa trilobite arthropod
Hallucigenia sparsa onychophoran
Myllokunmingia fengjiaoa jawless “fi sh”
Nectocaris pteryx mollusk?
Opabinia regalis lobopod?
Palaecmaea typica cnidarian
Phragmochaeta canicularis annelid
Pikaia graciliens chordate
Wiwaxia corrugata mollusk?**
*Based on Benton**Identity uncertain
Three of the most unusual animals of the Cambrian period were (a) Anomalocaris, a predator with a round mouth and grasping appendages; (b) Halluci-genia, protected by a double row of spikes; and (c) Opabinia, with fi ve eyes and an extended grasping mouthpart.
xviii+598_Biodiversity_F.indd 97 11/17/11 1:12 PM

first entry98 camouflage and mimicry 98
camouflage and mimicry Mimicry occurs when natural selection favors characteristics that make organisms (the mimics) resemble other organ-isms or objects (the models). By resembling mod-els, the mimics are often able to avoid being eaten by predators or are able to obtain benefits at the expense of other species.
Mimicry does not have to be perfect in order to be highly effective. all that is necessary is that other species be unable to distinguish the mimic from the model at least part of the time. Most popula-tions have a range of characteristics that affect their appearance. if some members of the population even slightly resemble a model, they may benefit from this resemblance. For example, an insect may not resemble a leaf very closely, but closely enough that a predator with imperfect eyesight or in the semidark-ness of shade or dusk may overlook it. Male animals may rush to mate with artificial objects that (to human eyes) hardly resemble the female of the spe-cies at all. this likewise demonstrates that imperfect mimicry, in this case of a mimic that resembles a female animal, can be favored by natural selection. in examples of nest parasitism, the mimics may look very different from the hosts whose nests they para-sitize, but the hosts are not using vision as their way of identifying which animals should be allowed into the nest. in many cases, generation by generation, the incipient mimics are selected to more and more closely resemble the model. after a long period of evolution, the resemblance can sometimes be very striking. Since mimicry involves the evolution of characteristics in the mimic and the evolution of the ability of other animal species to recognize them, it is an example of coevolution.
types of mimicry include the following.Camouflage mimicry. camouflage is widespread
among animals. Many mammals, for example, have dappled coats that help them blend in with vegetation and shade, and many lizards are colored like rocks. Many insects have a striking resemblance to sticks or leaves. the mantid Acanthodis curvidens closely resembles a stick, while the mantid Phyllium gigan-teum and the katydid Mimetica viridifolia closely resemble leaves. the scientific names define these insects by their ability to mimic leaves. the tropi-cal leaf litter frog Rhinella margaritifera comes in several color patterns, all of which resemble decay-ing leaves. Most animals are countershaded, which means that their backs are darker than their bellies. Some caterpillars have a shape and coloration that resembles bird droppings. Some examples of camou-flage, such as zebra stripes, appear quite imperfect in the middle of the day but make the zebras blend well into the grass at dusk, when their predators are hunting them.
Some animals mimic the appearance of their predators to escape notice. One example is the nymph of the texas bow-legged bug Hyalymenus tarsatus, which closely resembles ants and feeds alongside them on plants.
camouflage does not always involve the mim-ic’s own coloration. Some caterpillars eat leaves by removing tissue from the outer edges while main-taining the overall shape of the leaf. Predators may seek caterpillars by looking for patches of missing leaf material and may be less likely to notice leaves that are smaller but otherwise almost normal in appearance.
camouflage mimicry is less common but not unknown among plants. living stones, succulent plants of the genus Lithops in the deserts of southern africa, resemble green rocks and thus avoid being eaten. the young leaves of many plant species, while still tender and not yet toxic, are often reddish or brownish in color, which may make herbivores mis-take them for dead leaves.
even fungi can use camouflage mimicry, such as a fungus that lives in termite mounds. normally, termites remove fungal hyphae and reproductive structures, but this fungus has reproductive struc-tures that resemble termite eggs in size, shape, and texture.
Distraction mimicry. the bug Anisocelis fla-volineata has a large thorax, which a predator can easily see, but it also has large red flaps on two of its legs, which it holds away from its body. When the bug sits on a flower, the three red zones resemble flower parts. if the predator recognizes the prey, it may still grab at one of the leg flaps and eat just the leg, leaving behind an injured but surviving bug. another example is the sea krait Laticauda colubrine (a type of sea snake), in which the tail is colored like the head. When the snake curls around, a predator may think that there are two venomous snakes rather than just one. Some insects have markings on them that look like large eyes, which may startle predators (see figure on page 99).
Sexual and prey mimicry. there are numer-ous examples of organisms that mimic the sexual characteristics of another species in order to take advantage of them. consider the example of the blis-ter beetle Meloe fransiscanus, which parasitizes the nests of Habropoda pallida solitary bees. the beetle larvae gather together into clusters on the stems of plants; these clusters not only look a bit like female Habropoda bees, but the larvae produce a chemical that resembles the bee sex pheromone. a male bee attempts to mate with the cluster of beetle larvae, and they cling to him. He flies away with the clus-ter, and when he finds a female bee and attempts to
xviii+598_Biodiversity_F.indd 98 11/17/11 1:12 PM

camouflage and mimicry 99
mate again, the beetle larvae jump onto the female. eventually, the female bee carries the beetle larvae to the nest.
certain flowers resemble, to some wasps and flies, either their food or the female of their species. Male wasps pollinate some species of orchids by attempting to mate with the flowers. the appear-ance of the orchids is close enough to that of female wasps that the males are tricked, and the orchids produce a scent that, to the male wasps, is indis-tinguishable from that of a female wasp. the green spider orchid Chiloglottis trapeziformis produces a chemical, 2-ethyl-5-propylcyclohexan-1,3-dione, that is the same as the sex hormone produced by the female Neozeleboria cyroptoides wasp. in many other cases, female flies lay eggs in flowers (espe-cially arums and carrion flowers) that look and smell like rotting meat (see table on page 100). even more numerous are examples of plant species (once again, many of them orchids) whose flowers do not produce their own nectar but that closely resemble the flowers of plant species that do produce nectar. the pollinator visits the nectarless flower to pol-
linate it, discovering only too late that no nectar is available.
angler fishes are so called because they have extensions from their heads that resemble worms; they eat other fishes that approach to investigate the worm. in a similar fashion, the tongue of the alliga-tor snapping turtle resembles a moving worm, and the turtle eats fishes attracted by it. Some freshwater mussels attract fishes with colored tissues that look like prey; when the fishes approach, the mussels spurt out larvae, which adhere to the fishes. the mollusks use the fishes as vehicles to transport their eggs back upstream.
Some predators imitate their prey in order to attract them. the bolas spider produces sex hor-mones of female moths, thus attracting male moths. One kind of firefly beetle produces the light signals of the females of another kind of firefly beetle, which is its prey. a kind of katydid imitates the wing-flick sound of female cicadas, thus attracting male cicadas.
Mullerian mimicry. named after the 19th-cen-tury German zoologist Fritz Müller, this form of mimicry occurs when poisonous prey species evolve to resemble one another. the entire set of species is both mimic and model. they converge upon a set of characteristics such as warning coloration that allows the predator to recognize them as poisonous. Black alternating with white, red, yellow, and/or orange are widespread warning colorations among animals, such as bees, wasps, and snakes.
Batesian mimicry. named after the 19th-cen-tury British naturalist Henry Walter Bates, who first observed this phenomenon in the amazon rain forest, this form of mimicry occurs when non-poisonous prey (mimics) evolve a resemblance to poisonous animals (models). the mimic therefore benefits from the protection afforded by the poison-ous models without themselves having to expend resources to make themselves poisonous. examples include stingless flies that resemble bees and wasps and the viceroy butterfly that resembles the poison-ous monarch butterfly. if the mimic becomes too common relative to the model, the system becomes unstable because when the predators occasionally eat the mimic, they do not suffer the consequences that originally selected for the ability to recognize the model (see table on page 100).
Social mimicry. in this kind of mimicry, also called social parasitism, the parasites mimic the social signals of the hosts. in turn, the hosts take care of them. cowbirds and cuckoos are notorious for laying their eggs in the nests of other bird species. Baby cowbirds hatch sooner and grow faster and larger than the host species’ own babies and begin pushing the host species’ babies out of the nest, but
This click beetle (family Elateridae) has large spots on its thorax, which may deceive predators into thinking it is a large animal with large eyes. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 99 11/17/11 1:12 PM

first entry100 camouflage and mimicry 100
the parents do not seem to notice. the parents use the location of the baby bird as the sole indicator of its identity. the common cuckoo, as a species, parasitizes the nests of many nesting bird species, but each female specializes on just one species of host, the eggs of which most closely resemble those she lays.
there are more than 10,000 species of insects that live in ant nests. not only do the ants not kill them, they also feed them, often mistaking them for a queen of their own species. Usually, the parasites (mimics) produce a chemical that is indistinguish-able from that produced by ant queens. in one
case, the parasitic caterpillar (and later the pupa) produces a sound that the worker ants cannot dis-tinguish from the sound produced by their queens (see table).
in some cases, a parasite causes one of its hosts to undergo changes that make it resemble the prey of another of its hosts. For example, some parasitic worms live in snails and cause the snails to develop brightly colored structures and to crawl onto the tip of a branch or a leaf. this makes the snail obvious to predatory birds—the next of the worms’ hosts. there are other parasites that cause ant abdomens to swell and become red to imitate berries. Once again, birds, which eat the berrylike abdomens, are the next of the worms’ hosts.
Mimicry provides numerous excellent examples of natural selection and coevolution. Batesian mimicry formed an important piece of evidence that charles Darwin cited in his original presentation of evolution-ary theory. Mimicry, like all other forms of coevolu-tion, has stimulated the evolution of many new species.
See also coevolution; natural selection; parasitism.
Further readingangier, natalie, and Jennifer S. Holland. “the art of
deception.” National Geographic, august 2009, 70–87.
Barbero, Francesca, et al. “Queen ants make distinctive sounds that are mimicked by a butterfly social para-site.” Science 323 (2009): 782–785.
Kaiser, Roman. “Plants and fungi use scents to mimic each other.” Science 311 (2006): 806–807.
Kronfrost, M. R., and lawrence e. Gilbert. “the popula-tion genetics of mimetic diversity in Heliconius but-terflies.” Proceedings of the Royal Society of London B 275 (2008): 493–500.
Mallet, J. “causes and consequences of a lack of coevolu-tion in Müllerian mimicry.” Evolutionary Ecology 13 (2001): 777–806.
———, and M. Joron. “evolution of diversity in warning color and mimicry: Polymorphisms, shifting balance, and speciation.” Annual Review of Ecology and Sys-tematics 30 (1999): 201–233.
Moksnes, a., and e. Roskraft. “egg-morphs and host pref-erence in the common cuckoo (Cuculus canorus): an analysis of cuckoo and host eggs from european museums and collections. Journal of Zoology 236 (1995): 625–648.
Peakall, Rod. “Pollination by sexual deception in austra-lian terrestrial orchids.” available online. URl: http://www.anu.edu.au/BoZo/orchid_pollination/. accessed May 11, 2011.
Randall, J. e. “a review of mimicry in marine fishes.” Zoo-logical Studies 44 (2005): 299–328.
examples of mimiCry
mimic model
Pollination mimicry
Bee orchid (Ophrys apifera)
Female Eucera bee
Hammer orchid (Drakaea glyptodon)
Female Zapilothynnus trilo-batus wasp
Orchid (Cryptostylis ovata)
Female Lissopimpla excelsa wasp
Spider orchid (Chiloglottis trapezi-formis)
Female Neozeleboria cryp-toides wasp
Orchid (Cyanicula gemmata)
Other orchids, which provide nectar
Predator mimicry
Bolas spiders Moths
Firefly (Photurus) Photinus firefly
Katydid (Chlorobalius) Cicadas
Batesian mimicry
Kingsnake (Lampropeltis triangulum)
Micrurus fulvius coral snake
Bombyliid bee flies Apid bees
Viceroy butterfly (Lime-nitis archippus)
Danaus plexippus monarch butterfly
Social mimicry
Cowbird (Molothrus ater) Many nesting bird species
Cuckoo (Cuculis canoris) Many nesting bird species
Caterpillar (Maculinea rebeli)
Myrmica schencki ant
xviii+598_Biodiversity_F.indd 100 11/17/11 1:12 PM

carbon cycle 101
Ruxton, Graeme D., thomas n. Sherratt, and Michael P. Speed. Avoiding Attack: The Evolutionary Ecology of Crypsis, Warning Signals and Mimicry. new york: Oxford University Press, 2005.
Saul-Gershenz, leslie S., and Jocelyn G. Millar. “Pho-retic nest parasites use sexual deception to obtain transport to their host’s nest.” Proceedings of the National Academy of Sciences USA 103 (2006): 14,039–14,044.
Schiestl, Florian P., et al. “the chemistry of sexual decep-tion in an orchid-wasp pollination system.” Science 302 (2003): 437–438.
Soler, M., et al. “Magpie host manipulation by great spot-ted cuckoos: evidence for an avian mafia?” Evolution 49 (1995): 770–775.
carbon cycle the carbon cycle is the move-ment of carbon atoms from the atmosphere, through the crust of earth and the food web, and back into the atmosphere. all the large molecules of which organisms consist are built from chains of carbon atoms. nearly all of these molecules contain oxygen atoms, and many contain nitrogen atoms as well. When photosynthesis removes carbon dioxide (cO2) from the atmosphere, its oxygen atoms as well as its carbon atoms end up in carbohydrates and enter the food web. carbon cycle concepts therefore apply to oxygen as well. the nitrogen cycle, described in a separate entry, differs from the carbon cycle at some points.
the carbon cycle is of immense interest today not only because life is based on it but because the greenhouse effect has caused certain parts of the carbon cycle to change in a way that is not beneficial to humans and to the earth as humans have come to depend upon it. Human activities, such as trans-portation and industry, have released tremendous amounts of cO2 into the atmosphere. Other human activities, such as deforestation, have reduced the ability of natural ecosystems to remove cO2 from the atmosphere. agriculture has done both of these things by replacing forests and grasslands, which remove much cO2 from the air, with farmland, which does not remove as much. indeed, farmland may release more cO2 into the atmosphere than it removes, because exposure of the soil to warmer conditions has stimulated the decomposition of humus that was previously stored in the soil, and soil erosion has removed much of the carbon-rich topsoil.
as with all biogeochemical cycles, carbon fol-lows long-term and short-term paths. the long-term paths occur on the scale of millions of years, while the short-term paths occur on the scale of tens to
thousands of years. therefore, only the short-term cycles are likely to be of any use in helping humans bring the greenhouse effect under control.
ShorT-TErm CarBoN CyClEthe short-term carbon cycle is powered by photo-synthesis. Plants and other photosynthetic organ-isms (some protists and bacteria) have chlorophyll and associated molecules, which absorb sunlight energy and deliver it into energy and electron carrier molecules. Photosynthetic organisms absorb cO2 from the atmosphere and convert it into carbohy-drates such as sugar, using the energy and electrons from the carrier molecules. in this way, photosyn-thesis creates food molecules and removes cO2 from the air.
not all of this living matter enters the food chain right away. at any given time, about 4 per-cent of the carbon that is removed from the air by photosynthesis is stored up—about a third of this in vegetation and about two-thirds in the soil.
Most of the living matter travels through the food web via herbivores and carnivores. Plants and animals respire. Respiration puts cO2 back into the atmosphere. When plants and animals die, decom-posers consume their food molecules; the respiration of decomposers also puts cO2 back into the atmo-sphere. Other biological processes also put carbon back into the atmosphere. anaerobic decomposers in wetlands release methane (cH4) into the atmo-sphere, where it quickly reacts with oxygen and becomes cO2. Before this happens, methane is an even more potent greenhouse gas than cO2. in addi-tion, many plant leaves release isoprene compounds into the air. these compounds help to stabilize their photosynthesis under hot, dry conditions. these volatile molecules react with hydroxyl radicals in the atmosphere and form haze, which is eventually rinsed from the atmosphere by rain.
Most ocean absorption of cO2 occurs as a result of photosynthesis, but some occurs because the water itself absorbs cO2, converting it to carbonic acid. the excess carbonic acid has caused a measur-able worldwide increase in ocean acidity.
loNG-TErm CarBoN CyClEBoth on land and in the oceans, some of the carbon from the food web enters long-term storage, in ways such as the following.
● Soil decomposers do not return all of the carbon to the atmosphere; some accumu-lates as humus in soil.
● in the oceans, bodies of many dead organisms accumulate on the sea floor. their calcium
xviii+598_Biodiversity_F.indd 101 11/17/11 1:12 PM

first entry102 carbon cycle 102
carbonate (limestone) shells may accumulate over millions of years to form limestone.
● in addition, at certain times in the past, the bodies of dead organisms have accumulated. Organic molecules from forests and to a certain extent peat bogs have accumulated and been trapped in geological deposits. after being subjected to high temperature and pressure by the rocks above and below them, these deposits have become coal. Shallow sea deposits, primarily of microbes, have been similarly transformed into oil.
● Much ancient organic matter has also been stored as natural gas, which consists mostly of methane. in many continental shelves, methane molecules are frozen with water in the form of methane hydrates or clathrates. the methane does not become a gas because of the pressure of ocean waters above the continental shelves. Occasionally, how-ever, methane “burps” have occurred and released methane into the atmosphere. no large methane releases are known to have occurred during human history.
in these ways, ecosystems provide “biological carbon pumps” that transfer carbon from short-term to long-term organic storage.
in addition, the silicate rocks of earth’s crust remove cO2 from the air. cO2 reacts with calcium silicate (caSiO3) to produce limestone (cacO3) and quartz (SiO2). this reaction occurs more rapidly under conditions of high soil erosion. in the past, during times of severe greenhouse effect on earth (such as the Permian extinction), there was much soil erosion, which allowed this reaction to remove some of the cO2 from the air. limestone formation is also stimulated by the acids produced by decompos-ers. therefore, vegetation cover enhances the rate of geological cO2 removal from the atmosphere. lime-stone deposits can hence have either a biological or a geological origin, and both of these processes remove cO2 from the atmosphere.
Over both long and short terms, biological and geological processes have favored slightly more cO2 removal from the atmosphere (carbon sinks) than cO2 release into the atmosphere (carbon sources). natural processes in the land and oceans remove about 151.5 gigatons of carbon from the atmosphere each year (see table). natural processes in the land and oceans release about 146 gigatons of carbon per year, with the result that natural processes remove about 5.5 gigatons of carbon per year. (a gigaton is a billion metric tons, which are roughly the same as english tons. in order to allow comparisons between cO2 and organic car-bon, these calculations are done in carbon dioxide equivalents, rather than using just the weight of the carbon atom.)
Occasional major perturbations of the long-term carbon cycle have occurred during the history of earth, for example the following.
● the Permian extinction, mentioned previ-ously, brought the Paleozoic era to a close about 251 million years ago. Volcanic erup-tions released much cO2 into the air, and acid rain killed much of the vegetation, the decomposition of which released even more cO2 into the air.
● an asteroid hit earth during the cretaceous extinction, about 65 million years ago, bringing the Mesozoic era to a close. Mas-sive decomposition of vegetation released much cO2 into the atmosphere at that time.
● about 55 million years ago, between the Paleocene and eocene epochs of the ceno-zoic era, a period of global warming (the “Paleocene-eocene thermal Maximum”) lasted about 100,000 years. it may have been caused by a massive emergence of methane from continental shelf clathrates (even though the Republican congressman
sinks and sourCes of Carbon on earth (gigatons of Co2
equivalent per year)
Sinks (removing CO2 from atmosphere)
Land processes 59.3
Ocean processes 92.2
Total 151.5
Sources (putting CO2 into atmosphere)
Land processes 55.5
Ocean processes 90.5
Fossil fuels* 6.4
Changes in vegetation 1.2
Total 153.6
Net Release of CO2 into Atmosphere 2.1
*Includes all industrial processesNote: Adapted from Normile
xviii+598_Biodiversity_F.indd 102 11/17/11 1:12 PM

Carboniferous period, biodiversity in 103
Dana Rohrabacher speculated in 2007 that it may have been due to dinosaur flatu-lence). Methane releases may have occurred in the Permian and cretaceous extinctions as well.
humaN EffECTS oN ThE CarBoN CyClEthe most important human effect on the carbon cycle is the release of cO2 from the burning of fos-sil fuels for industry and transportation. this cO2, together with cO2 produced by other industrial pro-cesses, releases about 6.4 gigatons of cO2 into the atmosphere per year. in addition, the destruction of natural habitats causes the release of another 1.2 gigatons of cO2 per year. the total, 7.6 gigatons of cO2, released by human activity exceeds the 5.5 gigatons per year that natural processes absorb by 2.1 gigatons. this additional 2.1 gigatons of cO2 per year is what has caused atmospheric cO2 levels to increase by about one-third since the beginning of the industrial Revolution (as well as widespread deforestation and land degradation) and is causing atmospheric cO2 to accumulate by just less than 2 parts per million (ppm) each year. the resulting greenhouse effect, if left unabated, will not create so much heat that humans cannot tolerate it, but it will greatly disrupt human economic and social activities.
tree plantations cannot remove cO2 from the air as effectively as natural forests, nor can crop-lands do so as effectively as can natural grasslands. natural forests and grasslands have many plant spe-cies, each of which removes cO2 from the air in a different way (up in the canopy or down in the shade, for example) and at different times of year, while plantations and croplands each have only one way of removing cO2 from the atmosphere. loss of plant biodiversity can even slow down the decompo-sition of leaf litter. therefore, one of the most impor-tant reasons humans need to protect biodiversity is that it is essential to the proper functioning of the carbon cycle.
See also biogeochemical cycles; creta-ceous extinction; greenhouse effect; nitro-gen cycle; Permian extinction.
Further readingBonan, Gordon B. “Forests and climate change: Forcings,
feedbacks, and the climate benefits of forests.” Science 320 (2008): 1,444–1,449.
Field, christopher B., et al. “Primary production of the biosphere: integrating terrestrial and oceanic compo-nents.” Science 281 (1998): 237–240.
Hättenschwiler, Stephan, et al. “Biodiversity and litter decomposition in terrestrial ecosystems.” Annual
Review of Ecology, Evolution, and Systematics 36 (2005): 191–218.
imhoff, M. l., et al. “Global patterns in human consump-tion of net primary production.” Nature 429 (2004): 870–873.
normile, Dennis. “Round and round: a guide to the car-bon cycle.” Science 325 (2009): 1,642–1,643.
Vitousek, Peter M., et al. “Human appropriation of the products of photosynthesis.” BioScience 36 (1986): 368–373.
Carboniferous period, biodiversity in the carboniferous period (360 to 290 million years ago) was the fifth period of the Paleozoic era, fol-lowing the Devonian period. in north america, the earlier Mississippian time, with extensive marine limestone deposits, is distinguished from the later Pennsylvanian time, with extensive coal deposits derived from terrestrial forests, within the carbon-iferous period.
Climate. in land that is now in the northern continents, climatic conditions were very warm and wet with shallow lakes and seas. these conditions were perfect for the growth of extensive swamps, from which coal deposits formed. this is why most of the coal and oil deposits are found today in the northern Hemisphere. Much of the land that is now in the southern continents had cold polar conditions. Glaciations on the large southern continent caused intermittent reductions in sea level during the car-boniferous period.
Some researchers have suggested that the atmo-sphere contained up to 35 percent oxygen gas, in contrast to today’s 21 percent (see table on page 104). this would have resulted from the extensive growth of plants, which produced oxygen and which accumu-lated into thick deposits of peat (later to become coal) rather than decomposing. the high concentration of oxygen would explain how some animals, particularly flying insects, could grow so large during this period: the abundant oxygen would have penetrated their breathing tubes, and the denser atmosphere would have supported their flight. Such high levels of oxygen would also have promoted spontaneous combustion, especially under dry conditions. During the car-boniferous period, almost all organisms lived in wet conditions. Despite this, many of the plants showed evidence of fire damage.
Continents. the land that is today the north-ern continents (such as europe and north amer-ica) formed the continent of laurasia, which was near the equator, which created warm climates. the land that is today the southern continents (such as africa and South america) formed the continent of
xviii+598_Biodiversity_F.indd 103 11/17/11 1:12 PM
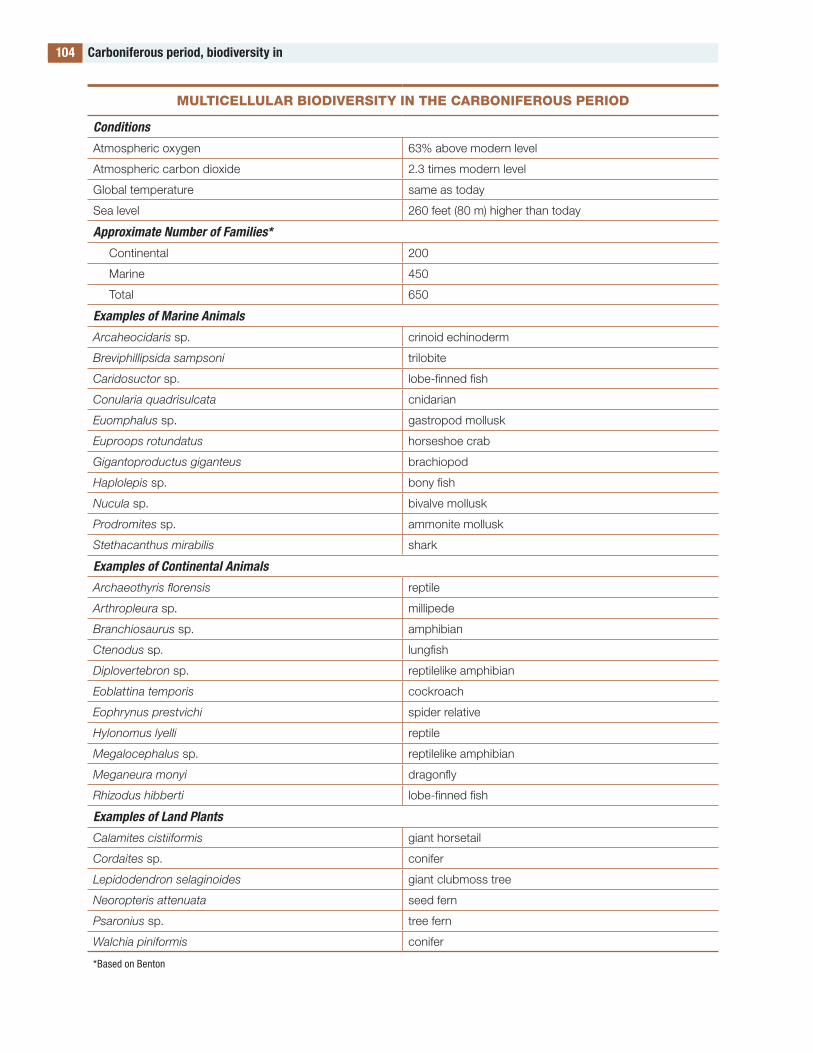
first entry104 Carboniferous period, biodiversity in 104
multiCellular biodiversity in the Carboniferous period
Conditions
Atmospheric oxygen 63% above modern level
Atmospheric carbon dioxide 2.3 times modern level
Global temperature same as today
Sea level 260 feet (80 m) higher than today
Approximate Number of Families*
Continental 200
Marine 450
Total 650
Examples of Marine Animals
Arcaheocidaris sp. crinoid echinoderm
Breviphillipsida sampsoni trilobite
Caridosuctor sp. lobe-finned fish
Conularia quadrisulcata cnidarian
Euomphalus sp. gastropod mollusk
Euproops rotundatus horseshoe crab
Gigantoproductus giganteus brachiopod
Haplolepis sp. bony fish
Nucula sp. bivalve mollusk
Prodromites sp. ammonite mollusk
Stethacanthus mirabilis shark
Examples of Continental Animals
Archaeothyris florensis reptile
Arthropleura sp. millipede
Branchiosaurus sp. amphibian
Ctenodus sp. lungfish
Diplovertebron sp. reptilelike amphibian
Eoblattina temporis cockroach
Eophrynus prestvichi spider relative
Hylonomus lyelli reptile
Megalocephalus sp. reptilelike amphibian
Meganeura monyi dragonfly
Rhizodus hibberti lobe-finned fish
Examples of Land Plants
Calamites cistiiformis giant horsetail
Cordaites sp. conifer
Lepidodendron selaginoides giant clubmoss tree
Neoropteris attenuata seed fern
Psaronius sp. tree fern
Walchia piniformis conifer
*Based on Benton
xviii+598_Biodiversity_F.indd 104 11/17/11 1:12 PM

Carboniferous period, biodiversity in 105
Gondwanaland, much of which was over the South Pole. During the later part of the carboniferous, laurasia collided with Godwanaland. the resulting geological forces produced mountain ranges even in the middle of tectonic plates, such as the appala-chian Mountains of north america. the land that is now Siberia collided with eastern europe to create what are now the Ural Mountains. the worldwide ocean (Panthalassic Ocean) was not divided by con-tinents, as it is today.
Marine life. all modern groups of marine organ-isms existed during the carboniferous except aquatic mammals (see fi gure).
● Invertebrates. Several modern lineages of crustaceans are fi rst known from the car-boniferous period. they had not yet evolved the pincer claws that are so common in modern crustaceans. Brachiopods remained more common than the superfi cially similar bivalves. the largest brachiopods (though only four inches [10 cm] in diameter) lived at this time. Shelled cephalopod mollusks (relatives of today’s chambered nautilus) remained common. trilobites persisted, but there were fewer species of them than during their Ordovician peak of diversity.
● Vertebrates. Perhaps as a result of the extinction of the placoderm (armored fi shes) at the end of the Silurian, sharks proliferated in diversity during the carboniferous. the shark Stethacanthus mirabilis had an anvil-shaped dorsal fi n, the func-tion of which is unknown. Bony fi shes continued to pro-liferate in the seas.
Continental life. extensive swamps fi lled the shallow seas and lakes of the carboniferous period in the tropical areas.
● Plants. large trees had been rare in the Devonian period, but trees up to 130 feet (40 m) in height became abundant in the carboniferous. Most of these were tree-sized versions of modern clubmosses (such as Lepidodendron), ferns (such as Psaronius, which had fronds almost fi ve feet [1.5 m] long), and horsetails (such as Calamites). these trees had systems of water transport and sexual reproduction that
would not work well for tree-sized plants on earth today but functioned well in the very wet conditions of the carboniferous. the fi rst seed plants evolved, possibly in drier upland regions, during this time. Before the carboniferous period, land plants grew only in wetlands, but now some grew on moist hillsides. the seed plants became more com-mon, and the seedless trees began their slide toward extinction during a period of drier weather. the seed ferns (now extinct) were not ferns, but seed plants with fern-shaped leaves. By the end of the carboniferous, the fi rst true conifers had evolved, fully adapted to growing on dry hillsides. there were no fl owering plants and were no plants such as grasses or shrubs that could grow in very dry conditions.
● Invertebrates. insects fi rst proliferated dur-ing the carboniferous. examples include Meganeura, a dragonfl y with a two-and-a-half-foot (75-cm) wingspan (see fi gure below). the fi rst cockroaches evolved dur-ing this period. Other arthropods included relatives of spiders; some millipedes, such as Arthropleura, were six feet (almost two m) long.
● Vertebrates. lungfi shes and lobe-fi nned fi shes (such as the 24-foot-[7-m-]) long Rhizodus hibberti) became less common as amphibians became more numerous and diverse. One lineage of amphibians
One of the largest arthropods of the Carboniferous period was the drag-onfl y Meganeura, with a wingspan of almost two feet (more than 60 cm). The higher oxygen content of the denser air in the Carboniferous period allowed large fl ying insects to evolve.
xviii+598_Biodiversity_F.indd 105 11/17/11 1:12 PM

first entry106 carnivorous plants 106
developed reptilelike characteristics. By the end of the carboniferous period, the first reptiles, with hard-shelled eggs that could be laid on dry land, evolved. Hylonomus may have been the ancestor of lizards, while Archaeothyris may have been the ancestor of the mammal-like reptiles. there were not yet any dinosaurs, mammals, or birds.
Extinctions. the carboniferous period is not recognized to contain one of the major extinction events. a worldwide period of warmer, drier climate near the end of the carboniferous created conditions that favored the spread and diversification of seed plants and reptiles.
See also conifers; fishes; insects; seed plants.
Further readingBenton, Michael J. “Diversification and extinction in the
history of life.” Science 268 (1995): 52–58.———. “Four feet on the ground.” in: Gould, Stephen
J., ed. The Book of Life: An Illustrated History of the Evolution of Life on Earth. new york: norton, 2001.
Béthoux, Olivier, and Frank Wieland. “evidence for car-boniferous origin of the order Mantodea (insecta: Dictyoptera) gained from forewing morphology.” Zoological Journal of the Linnaean Society 156 (2009): 79–113.
clarkson, e. n. K. “carboniferous crustaceans.” Geology Today 1 (2008): 11–15.
Dudley, Robert. “atmospheric oxygen, giant Paleozoic insects and the evolution of aerial locomotor perfor-mance.” Journal of Experimental Biology 201 (1998): 1,043–1,050.
Gastaldo, Robert a., et al. “community heterogeneity of early Pennsylvania peat mires.” Geology 32 (2004): 693–696.
Gugliotta, Guy. “Forest primeval.” Smithsonian, July 2009, 14–16.
Smithson, t. R. “the earliest known reptile.” Nature 342 (1989): 676–678.
carnivorous plants carnivorous plants are flowering plants that trap and digest animals, pri-marily insects. Since these plants are photosynthetic, they do not use the animals as sources of food energy but rather as sources of minerals. carnivorous plants live in wet conditions in which the water is usually acidic as well as nutrient poor. these conditions include bogs, tropical rain forests, and pine barrens.
carnivorous plants trap insects in several differ-ent ways.
● the most famous is the Venus flytrap (Dio-naea muscipula), in which the insect trips over a fiber that causes the leaf to fold up and trap it. the upper surface of the folded leaf secretes digestive enzymes, then absorbs molecules from the digested insect.
● Many carnivorous plants such as sundews (Drosera rotundifolia) and butterworts (Pinguicula vulgaris) (see figure) have sticky hairs on the leaf or stem surfaces. the sur-face cells secrete enzymes that digest the entangled insects.
● Many carnivorous plants such as pitcher plants (Sarracenia purpurea) and cobra lil-ies (Darlingtonia californica) have leaves that form a vertical cylinder. Hairs on the inside of the cylinder point downward to a pool of digestive juices. insects fall in and are digested, and their molecules are absorbed.
● Many carnivorous plants such as blad-derworts (Utricularia vulgaris) have small underwater bladders with a door that
The leaf of the carnivorous plant Drosera capen-sis entangles an insect in sap. The leaf produces digestive enzymes and absorbs molecules from the insects. (Noah Elhardt; Wikipedia)
xviii+598_Biodiversity_F.indd 106 11/17/11 1:12 PM
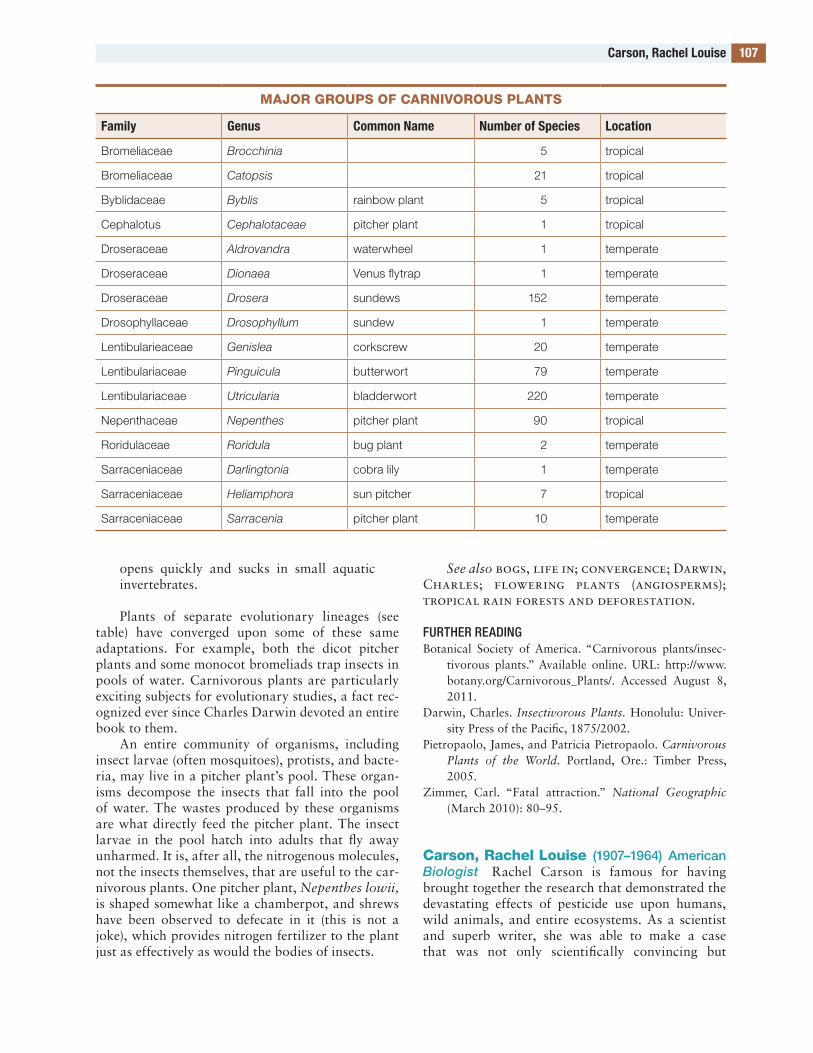
Carson, rachel louise 107
opens quickly and sucks in small aquatic invertebrates.
Plants of separate evolutionary lineages (see table) have converged upon some of these same adaptations. For example, both the dicot pitcher plants and some monocot bromeliads trap insects in pools of water. carnivorous plants are particularly exciting subjects for evolutionary studies, a fact rec-ognized ever since charles Darwin devoted an entire book to them.
an entire community of organisms, including insect larvae (often mosquitoes), protists, and bacte-ria, may live in a pitcher plant’s pool. these organ-isms decompose the insects that fall into the pool of water. the wastes produced by these organisms are what directly feed the pitcher plant. the insect larvae in the pool hatch into adults that fly away unharmed. it is, after all, the nitrogenous molecules, not the insects themselves, that are useful to the car-nivorous plants. One pitcher plant, Nepenthes lowii, is shaped somewhat like a chamberpot, and shrews have been observed to defecate in it (this is not a joke), which provides nitrogen fertilizer to the plant just as effectively as would the bodies of insects.
See also bogs, life in; convergence; Darwin, charles; flowering plants (angiosperms); tropical rain forests and deforestation.
Further readingBotanical Society of america. “carnivorous plants/insec-
tivorous plants.” available online. URl: http://www. botany.org/carnivorous_Plants/. accessed august 8, 2011.
Darwin, charles. Insectivorous Plants. Honolulu: Univer-sity Press of the Pacific, 1875/2002.
Pietropaolo, James, and Patricia Pietropaolo. Carnivorous Plants of the World. Portland, Ore.: timber Press, 2005.
Zimmer, carl. “Fatal attraction.” National Geographic (March 2010): 80–95.
Carson, rachel louise (1907–1964) American Biologist Rachel carson is famous for having brought together the research that demonstrated the devastating effects of pesticide use upon humans, wild animals, and entire ecosystems. as a scientist and superb writer, she was able to make a case that was not only scientifically convincing but
major groups of Carnivorous plants
family Genus Common Name Number of Species location
Bromeliaceae Brocchinia 5 tropical
Bromeliaceae Catopsis 21 tropical
Byblidaceae Byblis rainbow plant 5 tropical
Cephalotus Cephalotaceae pitcher plant 1 tropical
Droseraceae Aldrovandra waterwheel 1 temperate
Droseraceae Dionaea Venus flytrap 1 temperate
Droseraceae Drosera sundews 152 temperate
Drosophyllaceae Drosophyllum sundew 1 temperate
Lentibularieaceae Genislea corkscrew 20 temperate
Lentibulariaceae Pinguicula butterwort 79 temperate
Lentibulariaceae Utricularia bladderwort 220 temperate
Nepenthaceae Nepenthes pitcher plant 90 tropical
Roridulaceae Roridula bug plant 2 temperate
Sarraceniaceae Darlingtonia cobra lily 1 temperate
Sarraceniaceae Heliamphora sun pitcher 7 tropical
Sarraceniaceae Sarracenia pitcher plant 10 temperate
xviii+598_Biodiversity_F.indd 107 11/17/11 1:12 PM

first entry108 Carson, rachel louise108
emotionally compelling in her 1962 book Silent Spring, which remains one of the classics of popular science and nature literature.
Born on May 27, 1907, carson grew up with an interest in nature. She earned a Ph.D. in marine zoology in 1932 (see figure). She wrote radio scripts for the U.S. Fish and Wildlife Service during the depression era and after 1936 was in charge of all Fish and Wildlife Service publications. She wrote pamphlets about conservation, which had become a major concern of the U.S. federal government. On her own time, she wrote popular books about nature for a general readership (such as The Sea Around Us) and books for children. She resigned her government position to become a full-time writer in 1952.
carson became increasingly concerned about the effects of pesticides on the human and natural environments. She amassed numerous references from scientific literature to use in Silent Spring. She not only demonstrated that pesticides were danger-ous to the humans who used them or happened to
get in the way of the spray but that they also had calamitous effects on populations of wild animals such as birds (hence the title of her book). not only that, but even when pesticides were used in low concentrations, the food web could concentrate them until the tissues of top predators contained high levels of pesticide, a process called biomagni-fication. this was particularly true of DDt, which was a long-lasting pesticide. DDt interfered with calcium metabolism and caused birds at the top of the food chain, such as pelicans and eagles, to lay eggs that cracked open. largely as a result of the use of DDt, birds such as the bald eagle were threatened with extinction. Furthermore, the prof-ligate use of pesticides encouraged pest populations to evolve resistance to them, requiring the use of larger and larger amounts of pesticide to achieve less and less effect.
Silent Spring was vigorously attacked by the corporations that manufactured pesticides. carson was, however, able to maintain her case during tes-
Rachel Carson conducts marine biology research with her colleague Bob Hines in 1952. (United States Fish and Wildlife Service and NCTC Archives/Museum)
xviii+598_Biodiversity_F.indd 108 11/17/11 1:12 PM

cladistics 109
timony to the U.S. congress in 1963. carson died of cancer in 1964 but lived to see the beginning of legislation that restricted the use of pesticides to safer levels and situations. She would have been very pleased to know that the use of DDt was banned in the United States in 1972 and that the bald eagle made a considerable recovery in the three decades following this ban. She would not be pleased, how-ever, to see that pesticides are still used heavily in the United States, especially where migrant farm-workers are exposed to them, and that DDt, while not used in the United States, is manufactured in the United States, and used in other countries. as the book by angus Wright explains, the heavy use of pesticides in regions of Mexico that supply winter vegetables to american supermarkets has a devas-tating effect on the health of the peasants who raise the vegetables.
in 2007, a resolution of respect for Rachel car-son was introduced into the U.S. Senate. Such reso-lutions require a unanimous vote. there was one dissenting vote for this resolution (tom coburn, Republican of Oklahoma). He claimed that car-son’s attack on DDt resulted in the deaths of mil-lions of african children. in african countries, DDt had been banned in the 1990s for agricultural use, but not for the control of mosquitoes that spread malaria. in Silent Spring, carson had not called for a complete ban on pesticides but for their limited and focused use. it appears that today Rachel car-son still faces a campaign of misinformation.
See also biomagnification; pollution; resis-tance, evolution of.
Further readingcarson, Rachel. The Sea Around Us. new york: Oxford
University Press, 1951/2003.———. Silent Spring. new york: Houghton Mifflin,
1962/2002.lear, linda. Rachel Carson: Witness for Nature. new
york: Henry Holt, 1997.lytle, Mark Hamilton. The Gentle Subversive: Rachel Car-
son, Silent Spring, and the Rise of the Environmental Movement. new york: Oxford University Press, 2007.
Matthiessen, Peter, ed. Courage for the Earth: Writers, Sci-entists, and Activists Celebrate the Life and Writing of Rachel Carson. new york: Mariner, 2007.
Wright, angus. The Death of Ramón González: The Mod-ern Agricultural Dilemma, Rev. ed. austin: University of texas Press, 2005.
cartilaginous fishes See fishes.
chaparral See shrublands.
chimpanzees See primates.
chordates See animals.
cladistics cladistics is the modern, evolu-tion-based method of classifying biodiversity. it is a technique that classifies organisms on the basis of shared derived characters, which are similari-ties that they have inherited from a common ances-tor. this method assumes that two species with a greater number of shared derived characters are more closely related than two species that have fewer shared derived characters. the common ancestor of two species represents a branch point (Greek clados means “branch”) in the evolutionary histories of these species. cladistic analysis can be applied to any set of evolutionary lineages, not just to species. cladistic analysis is also called phylogenetic analy-sis. this method was developed in 1950 by the Ger-man entomologist Willi Hennig. a similar method, developed by the american botanist Warren Wag-ner, contributed methodologies that are now part of cladistic analysis.
cladistic analysis generates clusters of branches entirely on the basis of the characters of species. the diagram that represents the branching pattern is called a cladogram. the older systems of clas-sification, in contrast, attempted to reconstruct the evolutionary history of the organisms in question. By making no a priori evolutionary assumptions, cladistic analysis generates a set of possible evolu-tionary relationships and allows the investigator to choose the one that most closely matches the data of modern organisms or, if known, the data from the fossil record.
Grouping the species objectively on the basis of similarities has two advantages over older systems of classification:
● all the species are placed at the tips of the branches rather than at branch points. Mod-ern green algae and modern flowering plants are descendants of a common ancestor, but the lineage that led to green algae has under-gone less evolutionary modification than the lineage that led to flowering plants. Green algae and flowering plants are at separate branch tips of the cladogram. the ances-tral population of organisms from which both green algae and flowering plants are descended no longer exists.
● cladistics allows a more objective approach to classification than the traditional sys-tems. the cladogram can be generated by
xviii+598_Biodiversity_F.indd 109 11/17/11 1:12 PM

first entry110 cladistics 110
mathematical rules. this is often done by a computer. the investigator’s preferences do not influence the results.
as a result of this approach, cladists will not say that one modern group evolved from another mod-ern group. consider the example of green algae and flowering plants. if one could actually have looked at their common ancestor, it would have looked like green algae. cladists will not generally say that flow-ering plants “evolved from” green algae. they will say that green algae and flowering plants evolved from a common ancestor.
Choosing the traits. the investigator must choose which traits are to be used in the analysis. Some traits are ancestral (plesiomorphies), and some are derived (apomorphies). there are four categories of traits:
1. Shared ancestral traits (symplesiomorphy). ancestral traits are shared by almost all of the spe-cies in the analysis; ancestral traits cannot be used to distinguish among the species. in a cladistic analysis of flowering plants, for example, chlorophyll would not be a useful trait, since almost all flowering plants have chlorophyll in their leaves, a trait they inherited from their common ancestor. Most human character-istics are symplesiomorphies shared with other apes.
2. Unique traits (autapomorphy). characteris-tics that are totally unique to one species also cannot be used to distinguish among the species. consider a cladistic analysis of flowering plants in which one of the species is a parasitic plant, lacking chlorophyll in its leaves. the lack of chlorophyll is not a useful trait for this analysis, since only one of the species has this trait. an example from human evolution would be the unique skull characteristics, such as the projecting face and strong bite, of the neanderthals.
3. Convergences (homoplasy). in many cases, a trait may evolve more than once. this could hap-pen in either of two ways. First, the trait may have evolved into a more advanced form, then reverted back to the primitive form, during the course of evo-lution. this would cause a species to resemble a dis-tantly related species that had retained the ancestral form all along. Second, two species with separate origins may have evolved the same trait. the evo-lutionary biologist caro-Beth Stewart calls homo-plasy the “ultimate trickster” of cladistics. consider examples of homoplasy from plants, from nonhu-man animals, and from humans:
● a cladistic analysis of flowering plants may include two parasitic plant species that lack chlorophyll, one of them a parasitic relative of heath plants (such as genus Pterospora), the other a parasitic relative of morning glo-
ries (genus Cuscuta). the common ancestor of these two plants had chlorophyll. the chlorophyll was lost in two separate evolu-tionary events. it would be incorrect to clas-sify Pterospora and Cuscuta together into one group, with the other heath and morn-ing glory plants into another group.
● a cladistic analysis of vertebrates would include both mammals and reptiles. all mammals except monotremes have live birth. Most reptiles lay eggs, but some snakes have live birth. no biologist would classify live-bearing snakes together with the mammals, and the monotremes together with the reptiles. live birth evolved more than once: in the ancestors of certain snakes and in the ancestors of most mammals.
● increased brain size evolved independently in Homo erectus, neanderthals, and the lineage that led to modern humans.
it is generally easier for convergence to occur in the loss of an old adaptation (e.g., the loss of chlo-rophyll) than in the acquisition of a new one. traits that result from convergence are called analogous to each other. in contrast, traits that are inherited from a common ancestor are considered to be homolo-gous to each other. convergence represents a prob-lem for cladistics, just as it does for older methods of evolutionary classification.
4. Shared derived traits (synapomorphy). these are the traits that evolutionary scientists try to use in cladistic analysis. a cladistic analysis requires the choice of traits that distinguish different lineages: the species within each lineage shares the traits with one another, but not with species in other lineages. examples of synapomorphies include hair (which all mammals, and only mammals, possess) and feathers (which all birds, and only birds, pos-sess). it is generally easy to eliminate ancestral and unique traits from a phylogenetic analysis. the rec-ognition and elimination of homoplasy can some-times be difficult.
evolutionary scientists use more than one trait in a cladistic analysis. not only must the traits be shared derived traits, they must be independent of one another. to consider leaf length and leaf width as two traits would be incorrect, since longer leaves are usually wider as well. Rather than two traits, these may be just two ways of measuring leaf size.
the above examples used easily visible char-acteristics to classify organisms into groups. the characteristics need not be visible. in fact, many cladistic analyses use Dna sequence information. Suppose a scientist has segments of Dna from three species. if the segment of Dna is 100 nucleo-
xviii+598_Biodiversity_F.indd 110 11/17/11 1:12 PM

cladistics 111
tides in length, this allows 100 characters to be used to test each cladogram. nearly every phylo-genetic study includes at least one analysis that uses Dna nucleotides as traits. it is no easier to use Dna than to use visible characters in cladistic analysis. the reason is that one cannot compare just any old piece of Dna of one species with any old piece of Dna in another, any more than one can make comparisons of just any visible trait. Just as visible traits must be homologous, so the Dna segments must be homologous. the investigator must compare the same gene in all the species in the analysis. Further complications arise when one con-siders that some mutations exchange one nucleotide for another, while other mutations add or delete nucleotides, or reverse segments, or even move seg-ments to different places. Usually, researchers use certain well-studied Dna sequences rather than trying to fi gure out a new one for themselves. in comparisons among plants, matK (the gene for an enzyme that acts upon the amino acid tyrosine) and
rbcl (the gene for the large subunit of the enzyme rubisco) are frequently used.
Some analyses use noncoding Dna for phy-logenetic analysis. Researchers may use transpos-able elements because homoplasy is very unlikely to occur with them. it is very unlikely that the same transposable element would evolve twice: it would neither insert itself in the same location in the chro-mosomes of two different species nor transfer to a new location without gaining or losing a few nucleo-tides in the process. Other analyses use short or long interspersed sequences in the noncoding Dna. the fi gure uses the presence or absence of 20 segments of noncoding Dna to construct a cladogram of mam-mals. cows and deer are clustered together because they share all the segments for which information is available. Whales and hippos are clustered together because they share 15 of the 17 segments for which information is available. Whales and cows are not clustered closely together because they share only nine of the 17 segments for which information is
This cladogram is based on 20 noncoding sequences (“locations”) in the genomes of six mammal species. The “1” refers to the presence, and the “0” to the absence, of the sequence. The camel (the outgroup) has none of the 20 sequences and is less related to any of the others than they are to one another. Pigs form a clade sepa-rate from the remaining four species; they have sequences 19 and 20, while the others do not. The others all have sequences 10 and 12. Cattle and deer share four sequences that none of the others have, whereas whales and hippos share four sequences that none of the others have. This cladogram is meant to show patterns, not degrees, of relationship. Therefore, the horizontal line lengths are arbitrary. (Based on data from Nikaido, et al.)
xviii+598_Biodiversity_F.indd 111 11/17/11 1:12 PM

first entry112 cladistics 112
available. camels are not closely related to cows, as they have none of the 11 Dna segments that cows possess.
Choosing the best cladogram. a computer can generate many possible cladograms for any set of spe-cies that are being compared. Which of these clado-grams is the correct one? the correct one is the one that most closely matches what actually happened during evolutionary history. nobody can know exactly what happened during evolutionary his-tory; cladists must make an assumption about which cladogram is best. they often make the assumption of parsimony, that is, they choose the simplest expla-nation. to do this, they choose the cladogram that requires the fewest number of character changes. consider this simplified example in the classification of dogs, cats, and lions. Since each group has two branches, one could classify them in three ways:
1. [Dogs] [cats, lions]2. [Dogs, cats] [lions]3. [Dogs, lions] [cats]
cats and lions both have retractable claws; dogs do not. cladograms 2 and 3 require that retract-able claws should have evolved twice, separately in cats and lions. therefore, cladogram 1 is the most parsimonious. it would not be safe to draw a con-clusion based upon just one characteristic (in this case, retractable claws). cladograms based on any of a number of other characteristics would lead to the same conclusion: cats and lions form part of the feline group, while dogs are part of a separate (canine) group. From this, one would conclude that cats and lions shared a common ancestor that lived much more recently than the common ancestor of canines and felines.
it is easy enough to compare the three clado-grams that are possible with three species. However, as more species are compared, the analysis becomes exponentially more difficult. When comparing even a slightly larger number of species, the calculations become so complex that they can be performed only by a computer. as the number of species in the anal-ysis increases, even a computer cannot sort through all the possibilities. For 10 species, there are more than 2 million possible cladograms; for 50 species, there are 3 × 1076 possible cladograms. to provide some idea of how large a number this is, consider that the universe has existed for less than 1018 sec-onds! the computer must randomly generate a sub-set of several hundred possible cladograms and make comparisons among them.
even when a cladistic analysis compares only a few hundred of the possible cladograms, there may
be a large number of almost equally parsimonious ones. the computer must then generate a consensus tree that preserves the patterns that the best clado-grams share with one another.
Applying cladistics to classification. a cladistic approach to classification would group organisms together on the basis of the branch points in the cladogram. a clade (whether it corresponds to a genus, a family, an order, a class, a phylum, or a kingdom) must consist of all species that share a common ancestor, and only those species. Such a clade is called monophyletic (see figure on page 113). cladists will not accept a genus, family, order, class, phylum, or kingdom that includes only some of the species that have evolved from a common ancestor. Such a group is called paraphyletic. cladists most vigorously reject taxonomic groups that both omit some species that share a common ancestor and include some species that do not, a group referred to as polyphyletic.
a cladistic approach can lead to a different sys-tem of classification than what is traditionally recog-nized, as in these examples.
● “Monkey” is a paraphyletic category. the new World monkeys diverged from the Old World lineage, which includes Old World monkeys and all of the apes. to a cladist, either a human is a monkey, or else there is no such thing as a monkey.
● “Reptile” is a paraphyletic category. terres-trial vertebrates are traditionally classified into reptiles, birds, and mammals. Reptiles are cold-blooded and have scales, birds are warm-blooded and have feathers, and mam-mals are warm-blooded and have hair and milk. this sounds like a straightforward classification based upon characteristics: classes Reptilia, aves, and Mammalia. cla-dists point out that several lineages of reptiles branched off from one another to produce separate lineages such as turtles, lizards, tua-taras, more than one branch of dinosaurs, and therapsids. Mammals evolved from the-rapsids, and birds from dinosaurs. Reptiles, birds, and mammals are not separate clades, or branches. “Reptile” is therefore a paraphy-letic group that contains some, but not all, of the species descended from the group’s com-mon ancestor. Strict cladists insist that either the term reptile should not be used at all, or else the term reptile should include birds and mammals. to a cladist, the term dinosaur includes birds, and birds are dinosaurs. One can always tell a cladist because he or she
xviii+598_Biodiversity_F.indd 112 11/17/11 1:12 PM

cladistics 113
will refer to the big lumbering beasts of the Mesozoic era as “nonavian dinosaurs” rather than simply as dinosaurs. to a cladist, either a human is a reptile, or else there is no such thing as a reptile.
● “Fish” is a paraphyletic cat-egory. Vertebrates include fi shes, amphibians, reptiles, mammals, and birds. the reptile clade (which includes birds and mammals) and the amphibian clade share a common ancestor, but the amphibian-reptile clade branched off from the lobe-fi nned fi shes, which is just one branch of several branches that are usually called “fi shes”; the oth-ers are the ray-fi nned fi shes, cartilaginous fi shes, hagfi shes, and lampreys. “Fishes” is therefore also a paraphyletic group. cladis-tically speaking, then, there is no such thing as a fi sh, unless by fi sh one includes amphib-ians and reptiles (which includes birds and mammals). to a cladist, either a human is a fi sh, or else there is no such thing as a fi sh.
Some cladists, in fact, insist that scientists should not even bother with the linnaean sys-tem. instead of kingdoms, phyla, classes, orders, families, and genera, scientists should refer only to clades. this approach is called the PhyloCode.Most people, from common experience and also from the biology classes they took in school, tend to think of the old-fashioned classifi cations such as fi sh, amphibian, reptile, bird, and mammal. the new classifi cations are unfamiliar and confusing to most lay people. What many science teachers do is accept the evolutionary insights of cladistic analysis but do not attempt cladistic consistency in terminology.
Cladistics confi rms evolution. the grouping patterns produced by one cladistic analysis often closely resemble the grouping patterns produced by another. For example, a cladistic analysis that uses visible characteristics usually comes out almost the same as a cladistic analysis that uses Dna. this is highly unlikely to happen by chance and almost certainly means that the resulting classifi cation represents what really happened in the evolution-ary history of the organisms.
Moreover, the grouping patterns produced by cladistic analyses usually bear a close resemblance
to the grouping patterns that had been proposed previous to the development of cladistics. One example is within the walnuts (genus Juglans). For at least a century, botanists have recognized four major groups of trees in this genus, based upon leaf and fruit characteristics, as well as upon the geogra-phy of where the trees grew:
● the black walnuts of north and South america, section Rhysocaryon (16 species)
● the asian butternuts, section Cardiocaryon(three species)
● the butternut of eastern north america, sec-tion Trachycaryon (one species)
● the eruasian walnut, section Dioscaryon(one species)
the botanist alice Stanford and colleagues per-formed cladistic analyses upon the walnuts using three different regions of Dna: the matK gene, an itS (internal transcribed spacer, a type of noncod-ing Dna), and RFlP (restriction fragment length polymorphism Dna). they did not base their results on just one region of Dna, but on a consen-sus of three regions. the result was the same four groupings of walnut species. cladistics, therefore, often confi rms that the evolutionary classifi cations that had been previously produced were not the mere product of evolutionists’ imaginations. com-puters, which have no imagination, produce the same groupings.
this result is similar to what happened when charles Darwin published On the Origin of Spe-cies. Before Darwin’s theory, in fact before the acceptance of any form of evolution, scientists had been classifying organisms by the linnaean
The fl owering plants (angiosperms) are a monophyletic group that consists of all the species descended from a common ancestor; within the angio-sperms, the monocots are a monophyletic group.
xviii+598_Biodiversity_F.indd 113 11/17/11 1:12 PM

fi rst entry114 cladistics 114
system, based on what seemed to them to be the most “natural” groupings. Darwin said that these “natural” groupings were, in fact, the product of common descent and that scientists, to their amazement, had been studying evolution all along without knowing it.
cladists pay close attention to the occasional differences between cladistic and traditional clas-sifi cation systems. One signifi cant example of these differences is the reclassifi cation of the fl ow-ering plants. traditionally, all fl owering plants were classifi ed as either monocots or dicots. More recent analysis indicates that the monocots are a monophyletic group, but the dicots are not. the monocots are one branch from within the dicots. therefore, dicot is a paraphyletic group (see fi gure). cladistics has shown the monocot-dicot dichot-omy that every college biology student learns is an oversimplifi cation.
Perhaps the main advantage of cladistics in evolutionary science is that it allows investigations to proceed without having detailed knowledge of which ancient species, represented today by fossils, were or were not ancestral to which modern spe-cies. For example, the existence of Homo habilisabout 2 million years ago is well known. However, scientists cannot know if H. habilis represented the actual ancestral population from which the modern human species evolved or whether modern humans evolved from another population similar to H. habi-lis. With cladistics, research can continue despite this uncertainty.
the cladistic method has received some experi-mental confi rmation. the evolutionary microbiolo-gist Daniel Hillis grew cultures of viruses in his laboratory, allowing them to undergo mutations and evolve into different lineages. He knew precisely
what the pattern of evolutionary branching was for these viruses. He then performed cladistic anal-yses using Dna base sequences. the resulting cladogram closely matched the evolutionary pattern that had actually occurred.
cladistics has also proven its worth in the study of evolution-ary medicine. cladograms regu-larly appear in articles published in such journals as the Journal of Emerging Infectious Diseases,published by the national insti-tutes of Health. epidemiologists routinely perform cladistic analy-ses on strains of diseases in order to determine which strains are most closely related to which oth-
ers in the hope of deducing where new strains of diseases originated. this knowledge helps in the production of vaccines, to control the spread of the disease, and to predict the emergence of new diseases. it was cladistic analysis that allowed epi-demiologists to identify the strain of anthrax that was used in bioterrorist attacks in the United States in 2001. cladistics is also important to epidemiolo-gists who want to know from which animal species a human disease may have evolved. cladistics was the method that allowed researchers to determine that HiV came from chimpanzees. cladistics has proven not only a revolutionary technique within evolutionary science but has allowed evolutionary science to produce practical benefi ts to humankind.
See also biological classification; convergence.
Further readingcracraft, Joel, and Michael J. Donoghue. Assembling the
Tree of Life. new york: Oxford University Press, 2004.
Hillis, Daniel M., et al. “experimental phylogenies: Gen-eration of a known phylogeny.” Science 255 (1992): 589–592.
nikaido, M., a. P. Rooney, and n. Okada. “Phylogenetic relationships among cetartiodactyls based on inser-tions of short and long interspersed elements: Hippo-potamuses are the closest extant relatives of whales.” Proceedings of the National Academy of Sciences USA96 (1999): 10,261–10,266.
Stanford, alice M., Rachel Harden, and clifford R. Parks. “Phylogeny and biogeography of Juglans (Juglanda-ceae) based on matK and itS sequence data.” Ameri-can Journal of Botany 87 (2000): 872–882.
Stewart, caro-Beth. “the powers and pitfalls of parsi-mony.” Nature 361 (1993): 603–607.
The dicots are a paraphyletic group of fl owering plants because the mono-cot lineage is excluded from it. The term dicot is used descriptively but is not a phylogenetic classifi cation.
xviii+598_Biodiversity_F.indd 114 11/17/11 1:12 PM
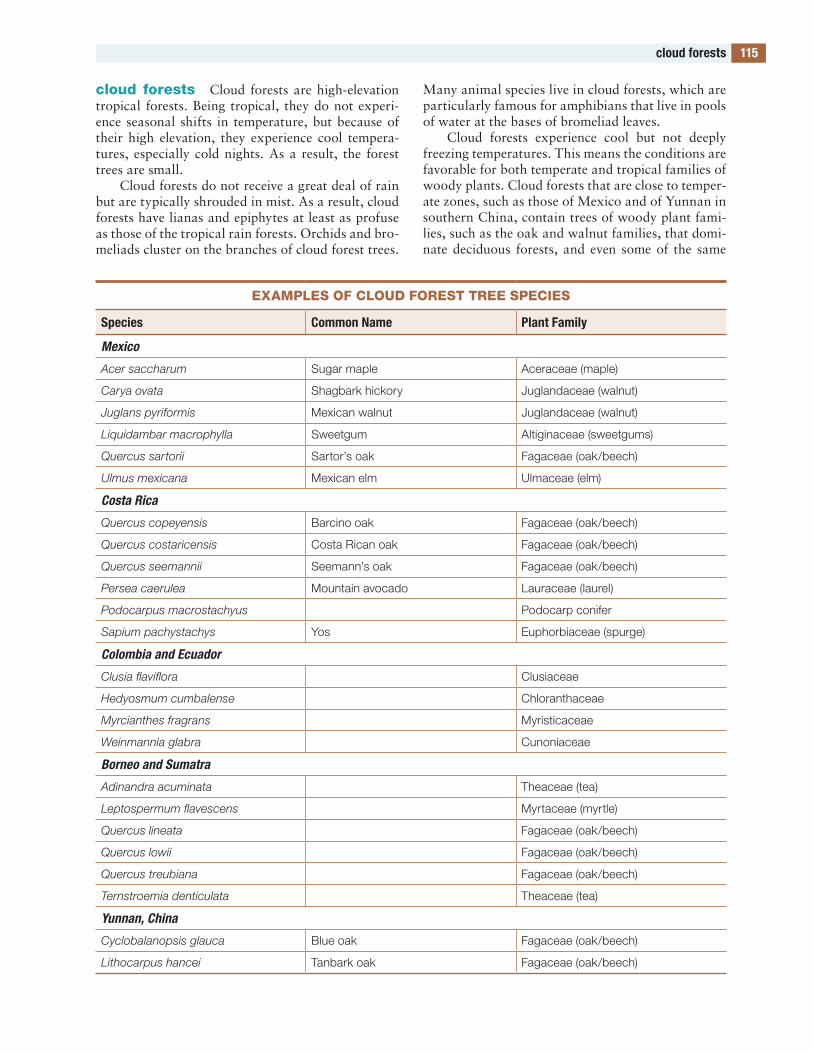
cloud forests 115
cloud forests cloud forests are high-elevation tropical forests. Being tropical, they do not experi-ence seasonal shifts in temperature, but because of their high elevation, they experience cool tempera-tures, especially cold nights. as a result, the forest trees are small.
cloud forests do not receive a great deal of rain but are typically shrouded in mist. as a result, cloud forests have lianas and epiphytes at least as profuse as those of the tropical rain forests. Orchids and bro-meliads cluster on the branches of cloud forest trees.
Many animal species live in cloud forests, which are particularly famous for amphibians that live in pools of water at the bases of bromeliad leaves.
cloud forests experience cool but not deeply freezing temperatures. this means the conditions are favorable for both temperate and tropical families of woody plants. cloud forests that are close to temper-ate zones, such as those of Mexico and of yunnan in southern china, contain trees of woody plant fami-lies, such as the oak and walnut families, that domi-nate deciduous forests, and even some of the same
examples of Cloud forest tree speCies
Species Common Name plant family
Mexico
Acer saccharum Sugar maple Aceraceae (maple)
Carya ovata Shagbark hickory Juglandaceae (walnut)
Juglans pyriformis Mexican walnut Juglandaceae (walnut)
Liquidambar macrophylla Sweetgum Altiginaceae (sweetgums)
Quercus sartorii Sartor’s oak Fagaceae (oak/beech)
Ulmus mexicana Mexican elm Ulmaceae (elm)
Costa Rica
Quercus copeyensis Barcino oak Fagaceae (oak/beech)
Quercus costaricensis Costa Rican oak Fagaceae (oak/beech)
Quercus seemannii Seemann’s oak Fagaceae (oak/beech)
Persea caerulea Mountain avocado Lauraceae (laurel)
Podocarpus macrostachyus Podocarp conifer
Sapium pachystachys Yos Euphorbiaceae (spurge)
Colombia and Ecuador
Clusia flaviflora Clusiaceae
Hedyosmum cumbalense Chloranthaceae
Myrcianthes fragrans Myristicaceae
Weinmannia glabra Cunoniaceae
Borneo and Sumatra
Adinandra acuminata Theaceae (tea)
Leptospermum flavescens Myrtaceae (myrtle)
Quercus lineata Fagaceae (oak/beech)
Quercus lowii Fagaceae (oak/beech)
Quercus treubiana Fagaceae (oak/beech)
Ternstroemia denticulata Theaceae (tea)
Yunnan, China
Cyclobalanopsis glauca Blue oak Fagaceae (oak/beech)
Lithocarpus hancei Tanbark oak Fagaceae (oak/beech)
xviii+598_Biodiversity_F.indd 115 11/17/11 1:12 PM

first entry116 coevolution 116
species, such as shagbark hickory and sugar maple (see table on page 115). cloud forests near the equa-tor, such as those in ecuador, have many trees in families of tropical origin, although oaks live in the cloud forests of Borneo and Sumatra (see table). in some areas of tropical mountains, conditions such as a fire cycle maintain grasslands, such as the andean páramo, rather than forest.
cloud forests are particularly vulnerable to the greenhouse effect. even a slight increase in tem-perature or change in weather patterns can cause the cloud layer to form at a greater altitude, too high to provide the constant drip of water upon which the plants and animals depend. the golden toad (Bufo periglenes) of central america appears to have become extinct because the cloud forests are no longer cloudy enough.
See also disturbances, ecological; green-house effect and biodiversity; tropical rain forests and deforestation.
Further readingchurchill, S. P., et al. Biodiversity and Conservation of
Neotropical Montane Forests. Bronx, n.y.: new york Botanical Garden, 1995.
Gomes, laurens G. l., et al. “tolerance of frugivorous birds to habitat disturbance in a tropical cloud forest.” Biological Conservation 141 (2008): 860–871.
Hamilton, lawrence S., et al. Tropical Montane Cloud For-ests. new york: Springer, 1995.
Pounds, J. a., et al. “Biological response to climate change on a tropical mountain.” Nature 398 (1999): 611–615.
Shi, J., and H. Zhu. “tree species composition and diversity of tropical mountain cloud forest in the yunnan, south-western china.” Ecological Research 24 (2009): 83–92.
tennesen, Michael. “Fog forests.” Discover, april 2009, 60–65.
coevolution coevolution occurs when the evo-lution of one species influences and is influenced by the evolution of another species. every species has experienced coevolution, since every species exists in a web of ecological relationships. in the general sense, every species coevolves with every other spe-cies. in the specific sense, the term coevolution is usually restricted to important and specific relation-ships between species rather than the diffuse back-ground of ecological relationships. For example, many plants have evolved the ability to compensate for the loss of tissue to herbivores, and herbivores have evolved the ability to digest plants, but this interaction is usually considered too diffuse to qual-ify as coevolution. Plants also evolved the ability to produce toxins, and herbivores evolved the ability to detoxify them. this can be considered coevolution
because the toxins and the ability to detoxify them are both often specific to certain groups of plants and herbivores.
in coevolution, natural selection favors those indi-viduals within a population that most effectively resist or cooperate beneficially with individuals of another species. in many cases, coevolution has produced symbiotic relationships between species (symbiosis), in which the life processes of individuals within one species are closely linked to those of individuals of another species (see table). the following are exam-ples, rather than a thorough analysis, of coevolution.
CoEvoluTIoN of plaNTS aNd hErBIvorESDuring the 1950s, ecologists such as Gottfried Fraenkel began to ask, why is the world green? insects are capable of massive population growth, and by now they should have eaten every bit of green leaf on the planet. Part of the reason this has not happened is that many insects die during the winter, and their populations must grow again the following spring. Still, even the winterless tropical forests are green. Why? the answer lies with coevolution.
Plants produce a great variety of secondary compounds in their tissues, especially their leaves. Secondary compounds have no primary metabolic role in organisms; they usually serve a defensive role instead. Many secondary plant compounds are famous for the uses to which humans put them. the active ingredients of spices and of many pharma-ceuticals as well as many miscellaneous compounds such as caffeine are all secondary compounds of plants. Some of the chemicals, such as tannins, are digestion inhibitors, while many others are mildly to strongly poisonous. in some cases, plant secondary compounds can work in ways other than as toxins. compounds in the needles of some conifers function as inhibitors of insect growth, so that insects that eat them cannot reach maturity. Some plants produce compounds that aphids mistake for predator alarm signals; this scares the aphids away. Other plants release chemicals into the air when herbivorous insects chew on them, and these chemicals attract wasps that then lay their eggs on the herbivores. Most plants have evolved chemical defenses against
types of symbiotiC interaCtions
Type of Interaction
Effect on host
Effect on other Species
Parasitism Harmful Beneficial
Commensalism Unaffected Beneficial
Mutualism Beneficial Beneficial
xviii+598_Biodiversity_F.indd 116 11/17/11 1:12 PM

coevolution 117
herbivores. Because of coevolution, the world is not a big salad bowl.
Plants usually produce the greatest amounts of secondary compounds in their leaves, as the stems are tougher (and often defended by thorns or spines), while roots are underground, where herbivorous ani-mals are fewer. leaves need to be relatively free of defensive spines and to be somewhat soft because they have to release water and oxygen into, and absorb carbon dioxide and light from, their environment.
Some plants have relatively little toxic defense against herbivores. these plants are usually the ones that can grow back quickly after being eaten. Grasses, for example, have few toxic compounds in their leaves, but they grow back quickly from under-ground structures after being grazed. Grass leaves have high levels of silicon dioxide, which makes them tough. Only specialized grazing animals can chew and digest them. Other plants get by without second-ary compounds because they live for only a short time and vanish from the scene after reproducing.
chemical defenses are costly. Some plants may produce fewer chemical defenses in years of abun-dant rainfall, when they can more easily afford to grow new leaves to replace those lost to herbivores. in some cases, plants wait until the herbivores begin to eat them, then undergo a rapid response and pro-duce toxins. in some cases, plants respond to drought by producing chemicals that make them more toxic. they may respond to drought and to herbivory by some of the same reactions. in all these and other ways, herbivores have influenced plant evolution.
Herbivores have evolved ways of eating plant tissue despite these defenses. Some animals consume massive quantities of herbage, get what little nutri-tion they can from it, and evacuate the rest. these animals (such as caterpillars, koalas, and sloths), deriving little energy from their food, are generally lethargic. Other animals can tolerate the toxins of specific plants and even specialize upon them. the caterpillar of the monarch butterfly (Danaus plexip-pus) eats milkweeds (genus Asclepias), which have chemicals that interfere with heartbeats, but they sequester the poisons in special sacs. they even use the chemical for their own defense: Having stored up the milkweed poison, they are now themselves poi-sonous to their predators. caterpillars of Heliconius butterflies similarly use toxic compounds from the passionflower vines (genus Passiflora) upon which they feed. Humans eat tissues of plants as spices. the fact that spices contain potent chemicals pre-vents humans from eating very much plant tissue. the human attraction to spices may have evolved because some spices have antimicrobial activity. in all these and other ways, plants have influenced her-bivore evolution.
Plants and herbivores are participating in what has been called an arms race, in which increased plant defense selects for increased ability of the her-bivore to tolerate or to avoid the plant defense and increased herbivore success selects for increased plant defense. Since plant-herbivore coevolution occurs over a long time period, the only thing similar to experimental confirmation is that on some islands plants do not have strong defenses. For example, a stingless nettle has evolved in Hawaii, which has no native browsing mammals.
evolution of defense against herbivores has occurred separately in every evolutionary lineage of plants. therefore, hardly any two plant defenses are the same. So vast is the diversity of plant chemical defenses that these chemicals are sometimes used to assist in the evolutionary classification of plants.
CoEvoluTIoN of prEdaTorS aNd prEyPredators are animals that kill entire organisms for food. Usually, their prey is other animals, but ani-mals that eat seeds are usually called seed predators because a seed contains an entire plant.
the evolution of predators has been overwhelm-ingly influenced by their search for prey. they tend to have greater intelligence, sensory acuteness, and agility than prey. Some predators have evolved pack behavior. the evolution of the prey has been just as strongly influenced by the need to avoid or resist predators. in some cases, this involves sensory acute-ness, agility, and herd behavior, just as it does with predators. Some prey form large herds in which the members can watch out for one another; in some flocks of birds and schools of fish, the coordinated and sudden movements of the members can disorient and confuse the predators. in some cases, the prey animals have evolved to simply become too big for most predators to be able to handle. the evolution of predators constantly influences the evolution of prey, and vice versa, because individuals that are slower, less intelligent, or less able to function as part of the group are more likely to be eaten or to not eat.
Seed predators have evolved the ability to find and sometimes store and retrieve seeds. the fact that seed predators, such as squirrels, fail to retrieve all the seeds they store is what makes them effective dispersers of seeds to new locations. as such, they often constitute a net benefit to the plant, some of whose seeds they eat. Seeds are a rich source of food for predators as well as for the embryonic plants inside of them. Seed plants have evolved, in turn, to resist seed predators, at least the ones such as weevils that offer them no benefit. Many seeds are mildly or strongly toxic. For example, while the apple fruit is delicious and nutritious, the seeds contain cyanide. this encourages the evolution of animals that will eat
xviii+598_Biodiversity_F.indd 117 11/17/11 1:12 PM

first entry118 coevolution 118
the apple but pass the seeds unharmed through their digestive tracts, obtaining nutrition from the fruit but not from the seeds. Many fruits have a laxative effect on animals, which encourages the animals to evacu-ate the seeds before they die in the intestines.
Perhaps the most interesting way in which plants have evolved in response to seed predators is mast seeding. Many trees produce large seed crops every few years and few seeds at other times. in this way,
the seed predator populations do not build up; when the trees produce a huge (mast) crop, the predator populations cannot consume them all.
not only has coevolution occurred between plants and the animals that both eat and disperse their seeds, it has produced a radiation of species within each. Many pines have seeds with wings and that blow in the wind, but some have wingless seeds that are stored and eaten by crows and jays. a prolif-
Species evolve in response to one another, a process known as coevo-
lution. This has been one of the most important ways that the astonishing bio-diversity of Earth has come into exis-tence. Many adaptations of one species make sense only in relation to another. But what happens if one of the coevo-lutionary partners becomes extinct? The surviving species may continue to manifest its adaptations, perhaps for thousands of years, even though the adaptations are now meaningless. The adaptations of the surviving species now become puzzling, “ghosts of biodiversity past.” In order for coevolutionary adapta-tions to continue, even when the other partner is a ghost, the adaptations must not cost so much that natural selection would get rid of them quickly. Also, the ghost must have become extinct recently, otherwise natural selection, operating over long periods of time, would have eliminated the coevolutionary adapta-tions even if they were not very costly.
Several species of trees in North America appear to participate in a coevo-lutionary relationship in which the partner has become extinct. First, consider some examples of trees that produce fruits that no wild animal appears to eat.
Bois d’arcs. These small trees are noted for their very strong wood and have been planted widely across North Amer-ica as fencerow windbreaks. Like other trees in the mulberry family, Maclura pomifera has separate male and female trees, and the females produce compos-ite fruits that consist of a cluster of ber-
ries. While mulberries are small, sweet, and edible by many species of birds and mammals (including humans), the fruits of the bois-d’arc are large (up to seven inches [or 15 cm] in diameter), green, hard, and sticky. (These trees are also called Osage oranges, because the fruits look a bit like oranges and grow wild in the tribal lands of the Osage tribe, and hedge-apples, because the fruits look a bit like apples and the trees were planted as hedges.) While not highly poisonous, the fruits are definitely repulsive and dif-ficult to digest. Squirrels consume the seeds, but this is not dispersal; they kill the seeds rather than dispersing them. The fruits commonly fall to the ground and rot; several of the fruit’s dozens of seeds grow, resulting in a clump of trunks that grow together. In some cases, the trunks fuse to form what appears to be a single trunk. It is rare to see a sin-gle-trunked bois d’arc tree in the wild. (Single-trunked bois d’arc trees are com-mon, but these were planted in fields that were subsequently abandoned and are not really wild trees.) Another con-sequence of the absence of a disperser is that wild bois d’arcs are found only in river valleys in Oklahoma, Texas, Arkan-sas, and nearby areas. Without dispers-ers, the only direction for such large fruits to roll is downhill, into river valleys. Had they not been planted widely across east-ern North America, they might eventu-ally have become extinct as the last fruit rolled into the sea.
Avocados. Wild avocados (Persea americana) are not nearly as large as the
ones in supermarkets, but they are still large enough that no extant species of animal swallows them whole. Many ani-mals can nibble at the nutritious fruit, but the seeds are simply left near the parent in most cases. Which large animal might have eaten these fruits in the past?
Honey locusts. Gleditsia triacanthos, a tree in the bean family, produces very long, spiral seed pods. No native species of mammal regularly eats these pods, even though they are juicy and sweet when young. Once cattle were introduced to North America, many of them ate these pods when available. Honey locusts have been widely planted as urban trees and have escaped again into the wild. There appears to be a similar lack of dispersal in the mesquite, a bush in the bean family that also produces sweet pods, though much smaller than those of the honey locust.
In each of these examples, it is prob-able that the true disperser would have been an animal large enough to swal-low the fruit whole, leaving the seeds intact—the way birds do with mulber-ries, for example. While there are no native mammals large enough to swal-low these fruits today, there were many such mammals about 13,000 years ago, at the end of the Pleistocene epoch. The mammals became extinct for reasons that are not entirely clear. The trees that produce these fruits have not become extinct—a few of their seeds germinate even without adequate dispersal—but their populations have suffered from the extinction of the large Pleistocene
“GhoSTS of BiodiverSiTy pAST”
xviii+598_Biodiversity_F.indd 118 11/17/11 1:12 PM

coevolution 119
eration of both pine and jay species occurred in the Miocene epoch, which some evolutionary scientists attribute to coevolution between them.
CoEvoluTIoN of flowErING plaNTS aNd pollINaTorS
Pollination is necessary for sexual reproduction in seed plants. the pollen must be carried from the male organs (male cones of conifers, or stamens of
flowers) to the female organs (female cones of coni-fers, or the pistils of flowers). the pollen lands upon the surface of the female cone or the stigma of the flower’s pistil and grows a tube down to the imma-ture seed.
in the ancestors of flowering plants, pollen was carried from one plant to another primarily by the wind. Modern conifers continue to rely on wind pollination. the earliest flowering plants relied on
mammals. Mammals that the Europeans brought to North America sometimes eat the fruits. For example, horses eat bois-d’arc fruits, which is why they are also called horse apples. Other examples of fruits that no longer have dispersers may be papayas, mangos, melons, gourds, and watermelons.
There are numerous fruits in Africa and Asia that are consumed and the seeds dispersed by animals (such as ele-phants) similar to the ones that became extinct in the Americas. Some of the fruits are inedible to most animals, but the now-extinct specialist dispersers may have had, as African and Asian fruit dis-persers do today, intestinal bacteria that break down otherwise toxic products. Elephants eat clay, which adsorbs toxins from the digestive system. Might mam-moths and mastodons have had similar adaptations that allowed them to con-sume fruits such as bois-d’arc?
There are also some flowers whose pollinators appear to be extinct. Pawpaws (Asimina triloba), a small wild fruit tree of the eastern deciduous forest of North America, persist primarily by the spread and resprouting of roots and underground stems. No native pollinator appears to be reliable.
Recently, native pollinator populations have declined precipitously, which threat-ens many native plant species with ulti-mate extinction. This may especially be true of large cacti in the deserts of south-western North America, which rely on bats to pollinate their flowers. Thus, very soon these cacti may become ghosts of biodiversity past.
There are also stems whose char-acteristics make sense only as adapta-tions to animals that no longer exist. The
bois d’arc also has large thorns, widely spaced on the branches, which appear totally ineffective at defending the leaves against the animals that eat them today, such as deer. Perhaps the thorns as well as the fruits are the leftover responses to the now-extinct herbivores. Wild honey locust trees have three-pointed thorns (hence the name tri-acanthos) bristling from their trunks. Hawthorns (genus Crataegus) have thorns even longer and more widely spaced than those of bois d’arc. Evolutionary biologists estimate that 10 percent of the woody plant spe-cies in New Zealand have a particularly tangled method of branching that pro-tected them at one time from browsing by moas, giant birds that were driven into extinction by the first Maori inhabitants.
The Pleistocene extinction occurred, as mentioned earlier, about 13,000 years ago. This is not long enough for trees to evolve some other mechanism of disper-sal. One would expect that, eventually, the unnecessarily large fruits would no longer be produced, as mutant forms of the trees produced fruits that could be easily dispersed by wind or water. This may have happened in the case of the honey locust. An apparently new spe-cies of locust, the swamp locust Gledit-sia aquatica, has evolved in southern North America, perhaps from a Gleditsia triacanthos ancestor. It produces much smaller pods that float easily in water.
There are also some animal exam-ples of the ghosts of biodiversity past. Pronghorns in the western grassland run much faster than necessary to escape modern predators, but prior to the Pleis-tocene extinction, they were pursued by the American cheetah, which was probably as fast as its barely-surviving
African counterpart, the fastest animal in the world.
As the ecologist Paul Martin wrote, “We live on a continent of ghosts, their prehistoric presence hinted at by sweet-tasting pods of mesquite [and] honey locust.” As the destruction of wild habi-tats by humans continues at a rapid pace, humans may be inaugurating the sixth of the mass extinctions in the his-tory of the world. Sometimes, when two species are partners, human activities cause the extinction of one partner but not the other. Starting in the 19th cen-tury, scientists began to understand that the present is the key to under-standing the past. To understand what happened in the past, look at what is happening today in the natural world. However, it is just as true that the past is the key to understanding the present. This is just one of the many meanings that emerge from the brilliant statement made by Theodosius Dobzhansky, one of the founders of modern evolutionary sci-ence: “Nothing in biology makes sense except in the light of evolution.”
See also coevolution; HerBivory; mass extinctions; pleistocene extinction; pol-lination; seed dispersal.
Further readingBarlow, Connie. The Ghosts of Evolution: Non-
sensical Fruit, Missing Partners, and Other Ecological Anachronisms. New York: Basic Books, 2000.
Buchmann, Stephen L., and Gary Paul Nab-han. The Forgotten Pollinators. Washing-ton, D.C.: Island Press, 1996.
Janzen, Daniel H., and Paul S. Martin. “Neo-tropical anachronisms: The fruits the gomphotheres ate.” Science 215 (1982): 19–27.
xviii+598_Biodiversity_F.indd 119 11/17/11 1:12 PM

first entry120 coevolution 120
animals (usually insects) to carry pollen from one plant to another. Very soon after the evolution of these early insect-pollinated flowers, some flowering plants reverted to wind pollination, and many flow-ering plants today rely on wind pollination. the evo-lution of wind pollination is not coevolution, since the wind is not an organism. Flowers that rely on wind pollination cannot control where their pollen goes and have characteristics such as the following.
● their petals are reduced or absent, as petals would only get in the way of the transporta-tion of pollen by the wind from stamens of one flower to pistils of other flowers.
● they produce massive amounts of pollen, since most of the pollen misses its target.
● the stigmatic surfaces are large, to increase the chance of the pollen hitting its target.
● the flowers of many wind-pollinated trees, such as oaks, open in the early spring, before the leaves of deciduous plants emerge. Oth-erwise, leaves would slow down the wind, and pollen might stick, uselessly, to the leaves.
in contrast, flowers that are pollinated by ani-mals have characteristics such as the following.
● their petals and nectar attract the animals. ● they produce less pollen, since animals can
carry the pollen directly to another flower. ● the stigmatic surfaces are not usually large. ● their flowers may open at a later time dur-
ing the growing season.
Flowers often attract pollinators with a variety of rewards: nectar is high in calories; pollen is a high-protein food; in some cases, flowers in cold cli-mates offer a warm spot (by concentrating sunlight and blocking wind) for the pollinators to get in from the cold. in some cases, flowers produce extra pollen that is sterile and intended specifically to feed the pollinators.
in many cases, flower characteristics attract and offer rewards to specific kinds of pollinators. Red tubular flowers, such as those of trumpet creep-ers, Campsis radicans, attract hummingbirds, whose long beaks and tongues fit into the tube, while they exclude animals that are too large to crawl into the tube and do not have long tongues (for example, bumblebees). Hummingbirds may have evolved a preference for red because it helped them find flow-ers, and the flowers may have evolved red pigment production because it brought pollinators to them. in contrast, red flowers do not strongly attract bees, which cannot see red. Moths and butterflies have
long tongues and often visit tubular flowers. Some flowers have closed petals and a landing platform; only large bees can push their way into the flower. in these and many other examples, coevolution has produced characteristics that benefit both species: the pollinators have evolved the ability to find and exploit these flowers as food sources, and the flowers benefit from being pollinated.
in other cases, flowers attract pollinators by deception. Many species of flowers have dark red-dish petals and produce the putrid scent of rotten meat, which attracts flies as pollinators. a few spe-cies of orchids attract male wasps by imitating the appearance and chemical characteristics of female wasps. the male wasps attempt to mate with the flower but succeed only in either getting pollen stuck to them from stamens or rubbing pollen off onto the stigmas. in these examples, the flowers benefit, but the insects do not. it is questionable whether these adaptations were produced by coevo-lution. natural selection favored flowers that most closely imitated dead meat or female wasps. How-ever, the evolution of the flies and wasps was pri-marily influenced by the search for food and mates, not by the flowers.
in some cases, insects obtain nectar from flow-ers without pollinating them. Some flowers have tubes that are too small for large bees to enter, but the large bees chew a hole from the outside and drink the nectar. Such nectar thieves are quite common.
Whether or not the pollinator benefits from the relationships, the pollination relationships just men-tioned are specific. Only certain wasp species will respond to the orchid that fakes the female wasp. While the specificity is not always absolute (hum-mingbirds sometimes drink nectar from flowers that are not red, and bees sometimes visit red flowers), it does assure that, in most cases, the animal carries pollen from one flower to another flower of the same species. an indiscriminate animal carrying pollen to a flower of a different species would not benefit either of the plants. in fact, it would not even be a pollinator. coevolution that leads to specific pollina-tion relationships rather than indiscriminate ones helps the plants transfer their pollen more efficiently and may help the pollinators find a more reliable food source.
in other cases, flowers are open to a wider vari-ety of pollinators. Many flowers are shallow, their stamens and pistils open to any insect that can fly or crawl in. in these cases, the plant population may be locally dense, so that pollen is seldom carried to the wrong species or the pollinators are relatively faith-ful to one species even when not forced to be by the flower structure.
xviii+598_Biodiversity_F.indd 120 11/17/11 1:12 PM

coevolution 121
Once coevolution had begun between flower-ing plants and insects 120 million years ago, at the beginning of the cretaceous period, it resulted in an astonishing diversification of flowering plants. a floral innovation that allowed even a slight change in which insects pollinated it would cause an almost immediate isolation of the new form from the rest of the population, starting it on the road to becoming a new species of flowering plant. this is probably the reason that each flowering plant family usually contains a considerable variety of growth forms and environmental adaptations but all of its species have similar flowers: the family itself began its evolution as a partner in a new coevolutionary pollination relationship. today there are nearly 250,000 spe-cies of flowering plants. Hymenopterans (bees and wasps) and lepidopterans (butterflies and moths) had existed since the Paleozoic era, and their diversifica-tion was not stimulated so much by flowers as the diversification of flowers was stimulated by them.
CoEvoluTIoN lEadING from paraSITISm To CommENSalISm
the preceding discussion focused how coevolution influenced general relationships among groups of species. coevolution has also produced symbiotic species relationships, in which one species depends upon another.
Parasitism occurs when individuals of one spe-cies (the parasite) obtain their living from individu-als of another species (the host). Some parasites are occasional visitors, such as mosquitoes; some, such as lice, may live on the host a long time; some, such as intestinal worms, live inside the host; and some microbial parasites live among and even in the host cells. Parasites have a negative impact on the host, either by consuming directly from it, by consuming some of its food (as when intestinal worms harm the host’s nutrition), by producing toxic waste products (as do many bacterial and viral parasites, such as cholera and bubonic plague), or by causing the host’s own immune system to have harmful effects on the host. the evolution of parasites has been strongly influenced by the hosts. For example, the life cycles of fleas that live on rabbits are coordinated with hormone levels in the rabbits’ blood, which ensures that the young fleas hatch at the same time as the baby rabbits are born. the evolution of the hosts has been influenced by parasites; the very existence of immune systems may be considered proof of this.
in contrast, a commensalistic relationship is one in which the commensal obtains its living from the host, but the host is indifferent to its presence. (this is why, according to some biologists, commensalism is not true symbiosis, but represents an intermediate condition between parasitism and mutualism.) the
human body contains trillions of commensalistic microorganisms, primarily on the skin and in the digestive tract. in fact, there are 10 times as many bacteria as there are human cells in the human body. the commensals consume materials that the body does not need. the worst thing they do is produce disagreeable odors. they are commensalistic not out of goodwill toward humans but because the body’s defenses keep them in check. People with compro-mised immune systems (such as from HiV) may be infected and killed by microbes that would other-wise have been commensalistic.
coevolution may select parasites that are more effective and hosts that have adaptations that will limit, resist, or eliminate the parasite. if the host species’ evolution is successful in causing the para-site to become harmless to it, the relationship is no longer parasitism, but commensalism. Similarly, in many but not all cases, the parasites actually benefit from not harming the host; if the parasite kills the host, it has killed its habitat and must find another. natural selection often favors the evolu-tion of milder and milder parasites. this does not happen with parasites, such as cholera bacteria, that disperse to new hosts rapidly or impersonally (in the case of cholera, through sewage). However, when a parasite must disperse to a new host by personal contact among hosts (as with smallpox), the parasite benefits when the host remains well enough to walk around and infect other people. the ebola virus is one of the deadliest diseases known to humankind, so deadly that it has never had a chance to spread through more than a small group of people at any one time.
coevolution, therefore, can cause host species to become more resistant and parasite populations to become milder. Many diseases, such as smallpox, were evolving toward milder forms even before the introduction of modern medical practices. Some dis-eases have, in fact, disappeared, perhaps because the parasitic relationship evolved into a completely com-mensalistic one. this process, also called balanced pathogenicity, has been directly measured during the course of a disease outbreak in rabbits. com-mensalism can be produced by coevolution, but once it has been attained, it does not involve coevolution any longer. natural selection may prevent the rela-tionship from slipping back into parasitism.
CoEvoluTIoN lEadING To muTualISmanother kind of symbiosis is mutualism, in which both species benefit. in some cases, the activities of commensals can evolve into mutualism. Most human intestinal bacteria are commensals. Some of them are mutualists because they provide a benefit to humans. the presence of some kinds of intestinal
xviii+598_Biodiversity_F.indd 121 11/17/11 1:12 PM

first entry122 coevolution 122
bacteria may prevent the growth of parasitic bacte-ria. Some of the more than 400 species of bacteria that live in the human intestine are mutualistic: they digest some foods that the intestine by itself cannot, making as much as 20 percent more calories avail-able for absorption. Some of these bacteria produce vitamin B, which is to them a waste product. Bac-teria on the skin and in body orifices may prevent infection by parasitic bacteria and fungi. nonhuman examples of beneficial internal microbes abound. cows cannot digest grass; it is the bacteria in their stomachs that digest it (and get food for themselves in the process). Many termites cannot digest wood; it is the microbes in their intestines or their nests that digest it. the microbes in cow and termite guts may well have begun as parasites or commensals and evolved a mutualistic function.
insectivorous birds may benefit from associating with grazing animals in a pasture; the large animal stirs up insects that the birds eat. the bird com-mensals benefit, and the large animal hosts are unaf-fected. Some birds have evolved an extra step. they eat parasites from the skins of the large host ani-mals, which turns the relationship into mutualism, but even this mutualism can be mixed with parasit-ism. yellow-billed oxpeckers (Buphagus africanus) eat ticks from mammals in the savannas of africa, but they also drink blood from wounds, including some that they cause.
Scientists may have observed an example of coevolution in the process of turning parasitism into mutualism. Fungi infect and reduce the growth and reproduction of grasses such as Festuca arundinacea and Lolium perenne. this sounds like parasitism. the infected grass smells bad to grazing animals such as cows, which avoid it, thus benefiting the grass. the cows that fail to avoid this grass suffer numerous physical ailments and may become lame from limb damage. What began as a parasitic infec-tion of the grass may be evolving into a mutualistic protection for the grass. Over the long term, how-ever, the grasses infected by fungi do not appear to grow any more than do the grasses without fungi, for both the herbivores and the fungi reduce the growth of the grasses.
Mutualism need not evolve through the stages of parasitism and commensalism. it may evolve from antagonistic or casual associations, as in the follow-ing examples.
● Big fish usually eat little fish. However, little fish can eat parasites from the big fish. if the big fish evolve behavior patterns in which they allow the little fish to come close to them, even to search around inside their mouths, and if the little fish evolve behavior
patterns to trust them, a mutualism may result: the big fish are relieved of their para-site load, and the little fish get food. Such cleaning mutualisms are common, especially in coral reef ecosystems.
● ants crawl on many kinds of plants looking for prey. this may have the effect of helping the plant by killing its herbivores. the plant may, in turn, benefit by rewarding the ants, encouraging them to stay. acacia trees of tropical dry forests are protected by ants, which kill herbivores and even vines that may threaten the acacias. the acacias, in turn, feed the ants with nectar and special morsels of protein and fat. in some cases, the mutual-ism has evolved so far that the ants and trees depend upon one another for survival.
● leaf cutter ants in tropical forests can-not eat leaves, but they can eat fungi that decompose the leaves. What may once have been a casual association, in which the ants preferred molded leaves, has evolved into an intricate mutualism in which the ants gather leaves, chew them up, and raise underground fungus gardens on the compost. as with the acacias and ants, the leaf-cutter ants and fungi have evolved a mutual dependence: neither can survive without the other.
● Human agriculture may have begun as a casual association between gatherers and wild plants and evolved into the biggest mutualism in the world.
Some mutualists are so intimate that they have begun the process of melding into single organ-isms. Zooxanthellae are single-celled algae that live in the skins of aquatic invertebrates. nitrogen-fix-ing bacteria and mycorrhizal fungi live in the roots of plants, most notably many leguminous plants (beans, peas, clover, vetch, alfalfa, locust), as well as alders, cycads, and casuarinas. Riftia pachyptila, a vestimentiferan worm that lives in deep sea volcanic vents, is more than three feet (1 m) in length, but it does not eat: it has no mouth, gut, or anus. it has specialized compartments in which symbiotic sulfur bacteria live, which live off of the hydrogen sulfide from the vents and then become food for the worm. Some mutualists, such as the bacteria that obtain nitrogen in sponges, are passed from one generation of the host to another when it reproduces.
Some organisms have gone all the way, melding mutualistically into unified creatures. the cells of all photosynthetic eukaryotes contain chloroplasts, and almost all eukaryotic cells contain mitochondria, the evolutionary descendants of mutualistic bacteria. in fact, the evolution of mutualism may have made the
xviii+598_Biodiversity_F.indd 122 11/17/11 1:12 PM

commensalism 123
evolutionary advancement of life beyond the bacte-rial stage possible. the evolutionary biologist lynn Margulis points out that the formation of mutual-isms has been a major process in the evolution of biodiversity.
the evolution of adaptations to the physical environment might have produced only a few spe-cies. Just how many ways are there for a plant or an animal to adapt to desert conditions? the greatest number of evolutionary innovations, from the huge array of plant chemicals to the astonishing diversity of flowers and even the symbioses that keep human cells alive, have resulted from coevolution of organ-isms in response to one another.
See also essay: “Ghosts of Biodiversity Past;” biodiversity, resources for medicine; herbiv-ory; pollination; predation; seed dispersal; symbiogenesis.
Further readingBäckhed, Fredrik, et al. “Host-bacterial mutualism in the
human intestine.” Science 307 (2005): 1,915–1,920.Barbosa, Pedro, and ignacio castellanos, eds. Ecology of
Predator-Prey Interactions. new york: Oxford Uni-versity Press, 2005.
Birkle, l. M., et al. “Microbial genotype and insect fitness in an aphid-bacterial symbiosis.” Functional Ecology 18 (2004): 598–604.
currie, cameron R., et al. “coevolved crypts and exocrine glands support mutualistic bacteria in fungus-growing ants.” Science 311 (2006): 81–83.
Dawkins, Richard, and J. R. Krebs. “arms races between and within species.” Proceedings of the Royal Society of London B (Biology) 205 (1979): 489–511.
Fisher, Jeffrey a. The Plague Makers: How We Are Cre-ating Catastrophic New Epidemics—And What We Must Do to Avert Them. new york: Simon & Schus-ter, 1994.
Fraenckel, Gottfried S., et al. “effect of soybean inhibitors on growth of Tribolium confusum.” Agriculture and Food Chemistry 2 (1954): 410–411.
Gill, Steven R., et al. “Metagenomic analysis of the human distal gut microbiome.” Science 312 (2006): 1,355–1,359.
Janzen, Daniel H. “co-evolution of mutualism between ants and acacias in central america.” Evolution 20 (1966): 249–275.
———. “Why fruits rot, seeds mold, and meat spoils.” American Naturalist 111 (1977): 691–713.
Kiers, e. toby, and R. Ford Denison. “Sanctions, coopera-tion, and the stability of the plant-rhizosphere mutu-alisms.” Annual Reviews of Ecology, Evolution, and Systematics 39 (2008): 215–236.
Koenig, Walter D., and Johannes M. H. Knops. “the mys-tery of masting in trees.” American Scientist 93 (2005): 340–347.
Pichersky, eran. “Plant scents.” American Scientist 92 (2004): 514–521.
Proctor, Michael, Peter yeo, and andrew lack. The Nat-ural History of Pollination. Portland, Ore.: timber Press, 1996.
Ruby, edward, Brian Henderson, and Margaret McFall-ngai. “We get by with a little help from our (little) friends.” Science 303 (2004): 1,305–1,307.
Sachs, Jessica Snyder. Good Germs, Bad Germs: Health and Survival in a Bacterial World. new york: Hill & Wang, 2007.
Saikkonen, K., et al. “Fungal endophytes: a continuum of interactions with host plants.” Annual Review of Ecol-ogy, Evolution, and Systematics 29 (1998): 319–343.
Scarborough, claire l., Julia Ferrari, and H. c. J. Godfray. “aphid protected from pathogen by endosymbiont.” Science 310 (2005): 1,781.
Schoonoven, louis M., Joop J. a. Van loon, and Marcel Dicke. Insect-Plant Biology, 2nd edition. new york: Oxford University Press, 2006.
Sessions, laura a., and Steven D. Johnson. “the flower and the fly.” Natural History (March 2005): 58–63.
Sherman, Paul W., and J. Billing. “Darwinian gastronomy: Why we use spices.” BioScience 49 (1999): 453–463.
thomas, Frédéric, François Renaud, and Jean-François Guégan, eds. Parasitism and Ecosystems. new york: Oxford University Press, 2005.
thompson, John n. The Coevolutionary Process. chicago: University of chicago Press, 1994.
Vega, Fernando e., and Meredith Blackwell, eds. Insect-Fungal Associations. new york: Oxford University Press, 2005.
Wills, christopher. Yellow Fever, Black Goddess: The Coevolution of People and Plagues. Reading, Mass.: addison-Wesley, 1996.
Wilson, Michael. Microbial Inhabitants of Humans: Their Ecology and Role in Health and Disease. new york: cambridge University Press, 2005.
Wu, licheng, et al. “Recognition of host immune activation by Pseudomonas aeruginosa.” Science 309 (2005): 774–777.
commensalism commensalism is a symbiotic relationship between two species in which the com-mensal derives part or all of its livelihood from the host, but without harming the host.
the most abundant commensals are the microbes and small organisms that live on or in the bodies of larger multicellular organisms. ani-mal bodies, including human bodies, have millions of bacteria on the skin, millions of bacteria in the mouth and throat, and trillions of bacteria in the intestines. commensalistic bacteria associated with the human body outnumber human cells 10 to one. Mites are found in the hair follicles of nearly all
xviii+598_Biodiversity_F.indd 123 11/17/11 1:12 PM

first entry124 communities, ecological 124
human beings, no matter how thoroughly they wash. they consume oils from the follicles. Some commen-sals even remain in suspended animation until the host dies. the nematode Pristionchus pacificus does not become active until the beetle on which it lives has died, at which time it eats the bacteria and fungi that decompose the dead beetle.
Humans are also surrounded by commensals. a mattress can have 2 million microscopic mites eat-ing the oils and skin flakes produced by their human hosts. they are so small they can go right through pillowcases and sheets. One-tenth of the weight of a six-year-old pillow may consist of skin flakes, living mites, dead mites, and mite dung.
Different portions of a multicellular body may support different ecological communities of com-mensals. Skin bacteria are almost entirely different from intestinal bacteria. even the bacterial communi-ties on the inside and the outside of the elbow differ from each other. a study of the skin of six healthy people found 182 bacterial species of 91 genera on the forearms. Only four of the species were found on all six people, and the original species were no longer present 10 months later. Some species of commensals are transient. their populations build up quickly and are washed away. Other species are permanent and live deeply in even well-washed skin. the most com-mon skin bacteria are in the genus Pseudomonas, which are also common in soil and water.
close ecological relationships of larger organ-isms can also be considered commensalistic. When vines grow on the branches of trees, their leaves may intercept the sunlight that the tree’s own leaves have not used or may grow on a dead branch; the addi-tional weight of the vines may be negligible. the vine benefits because it does not produce much of its own woody tissue. Many bird species (most famously cattle egrets, Bubulcus ibis) follow grazing animals and eat the insects stirred up by the movement of the grazers. the birds benefit because they do not have to expend their own energy stirring up the insects. in a similar fashion, commensalistic birds may follow army ants and eat the insects stirred up by the ants.
in north america, the black vulture (Coragyps atratus) is a commensalistic partner of the turkey vulture (Cathartes aura). turkey vultures, with well-developed scent recognition and good eyesight, are very good at finding carrion. Black vultures do not have well-developed scent recognition, and they usu-ally follow turkey vultures. the black vultures wait until the turkey vultures have finished eating, then clean up the leftovers.
the border between commensalism and other ecological relationships is unclear and shifting. Most microbes on and in the human body are potentially parasitic; they are commensals only because the
human body defends itself from them. if the human immune system is weakened by illness, the erstwhile commensals become parasites. if the human body dies, they become decomposers. the commensalistic vines described earlier can quickly become com-petitors of the tree on which they grow if their leaves cast shade on the tree’s leaves, if they become too heavy, or if their roots compete with those of the tree. the dust mites described previously, living in pillows or the carpet, might be considered decom-posers. they can be harmful to those people who have allergic reactions to their waste products. con-sider the example of barnacles growing on whales. if the barnacles are sufficiently uncommon, the whale may not notice them, and the barnacles get a free ride to areas with new food supplies. if the barnacle populations build up on the whales, they may irri-tate the whale, slow it down, or even allow infec-tions to occur.
See also coevolution; mutualism; parasitism.
Further readingFierer, noah, et al. “the influence of sex, handedness, and
washing on the diversity of hand surface bacteria.” Proceedings of the National Academy of Sciences USA 105 (2008): 17,994–17,999.
———, et al. “Forensic identification using skin bacterial communities.” Proceedings of the National Academy of Sciences USA 107 (2010): 6,477–6,481.
Gao, Z., et al. “Molecular analysis of human forearm superficial skin bacterial biota.” Proceedings of the National Academy of Sciences USA 104 (2007): 2,927–2,932.
Pennisi, elizabeth. “Bacteria are picky about their homes on human skin.” Science 320 (2008): 1,001.
communities, ecological an ecological com-munity is all of the interacting organisms in the same place at the same time. Because the web of interact-ing organisms extends nearly unbroken across the entire earth, the only well-defined community is that of the entire biosphere of earth. (even this has been called into question by those who claim that bacteria, called “space plankton,” can survive inside rocks thrown into outer space by asteroid impacts.) However, organisms may form recognizable clumps of interactions in which there are more interactions within the clump than between them. these clumps are ecological communities.
whaT arE EColoGICal CommuNITIES?More than a century ago, ecologists such as Fred-eric clements maintained that ecological communi-ties were distinct entities (superorganisms) with their own life cycles. these communities even received
xviii+598_Biodiversity_F.indd 124 11/17/11 1:12 PM

communities, ecological 125
latin names: a forest of Quercus oaks was a Querce-tum, and a woodland of Pinus pines was a Pinetum. ecologists such as Henry Gleason and Robert Whit-taker demonstrated that each species of plant had its own distributional range. the distinction between one ecological community and another is, according to this view, gradual, rather than being discrete like the body of an organism. Strictly speaking, there is no such thing as a forest. “Forest” is the name humans give to a set of species of plants, animals, and microorganisms that grow together and interact. animals, in particular, can be members of more than one ecological community. if a disturbance, such as a fire, wipes away a community, other species of plants, animals, and microorganisms live in the new environment created by the fire. Because of ecological changes that occur during the subsequent decades, the original set of species may, in fact, return, but it was the individual plants and animals within each species that grew back—not the forest.
More recently, some ecologists have begun to reexamine the extent to which ecological com-munities are associations of organisms that live in the same habitat conditions. While none of them have gone back to the clementsian view, they have emphasized the species interactions that occur much more strongly within than between communities. it is true that animals readily move from one ecologi-cal community to another, but most of them spend most of their time in one type of ecological commu-nity. the web of community interactions can even help to define the border of a community, indistinct as it is. One example is the transition from tundra to boreal or subalpine forest. the forest trees live in areas where the temperature is warm enough that their branches will not usually be killed by the win-ter wind and the permafrost is not near the surface of the soil. However, the trees themselves influence the environmental conditions. their dark needles absorb sunlight and increase the temperature where they live. in this way, the boreal or subalpine com-munity itself can increase its range northward or up a mountain. Perhaps the most realistic view of an ecological community is that it is a clump of frequent ecological interactions among species imperfectly but significantly separated from other such clumps.
adjacent ecological communities overlap in areas known as ecotones. an example is the tran-sition from tundra to boreal or subalpine forest already mentioned. each ecotone consists of loosely defined zones. Between a forest and grassland, for example, there is an ecotonal area dominated by saplings of shade-intolerant trees, an area influenced by the shade of the forest and litter from the trees in a way the center of the grassland is not. a terrestrial community, such as a forest, may intergrade into
an aquatic community, such as a lake. Some plants grow in the mud at the lake margin. Others, such as cattails, sedges, and rushes, specialize in the littoral zone, in which emergent plants are rooted under-water but have their leaves and flowers primarily above the water’s surface. each aquatic plant species specializes on a different water depth—water lilies grow farther out in the water than do cattails. Some plants live underwater or float on top of it. the plant species Hottonia inflata, for example, has finely dis-sected leaves that absorb dissolved carbon dioxide and bicarbonate from the water, but it floats because the peduncle (which bears the flowers) encloses a bubble of air. ecological succession typically occurs in these ecotonal zones, such as when the wetland fills with soil and decaying plant material.
Since interactions can take place on all scales, from a few inches to many miles, it is impossible to define the spatial extent of an ecological community. to a mosquito larva, a tree hole is the world, while wolves can range over a wide area, moving between forests and grasslands. For ease of discussion, this encyclopedia uses the following rough scale of com-munity identity to define its entries: a community is large and distinct enough to be separately identified on ecological maps (see figures on pages 126 and 127). the following ecological communities have entries in this encyclopedia:
● bogs ● boreal and subalpine forests ● cloud forests ● continental shelves ● coral reefs ● deep oceans ● deserts, cool ● deserts, polar ● deserts, warm ● grasslands and savannas ● marshes and swamps ● rivers, streams, ponds, and lakes ● shrublands ● temperate coniferous forests ● temperate deciduous forests ● temperate dry forests ● temperate rain forests ● tropical rain forests ● tropical seasonal forests ● tundra
lEvElS of dIvErSITy wIThIN CommuNITIESBiodiversity within ecological communities can be considered at three scales:
● Alpha diversity results from the species that live and interact in a single location.
xviii+598_Biodiversity_F.indd 125 11/17/11 1:12 PM

fi rst entry126 communities, ecological 126
● Beta diversity results from the change of species that occurs over the course of a few feet or meters.
● Gamma diversity results from the spe-cies differences between locations that are around a mile or kilometer apart.
as these defi nitions suggest, there is no clear way to distinguish the three levels of community diversity.
alpha diversity consists of the species that are in a single location. these species coexist and interact because they specialize on different ways of making a living within the community. Within a decidu-ous forest, for example, some trees, such as maples, dominate the canopy, while others, such as dog-woods, specialize in shaded conditions and fl ourish in the understory. Smaller shrubs, such as spicebush, and herbs, such as snakeroot, grow underneath them. Shrubs may specialize on using the light that reaches the forest fl oor in the early spring, before the forest canopy casts shade. in a fi eld of weeds or a desert, the plant species may all receive full sunlight but dominate different layers of soil or fl ourish at different times of the year. in the colorado Desert of california, shrubs have deep roots, while cactuses have shallow but widespread roots. tiny wildfl owers grow only during the brief spring rains and have very small root systems. the herbivorous animals may specialize on different species of plants or on eating
the plants in different ways: Some chew entire leaves, some eat photosynthetic cells inside the leaves, some tap the pressurized fl uid within the phloem, and some specialize on the roots of plants. the predators may also specialize. Birds may specialize on seeds or on insects. Parasites specialize on hosts to differ-ent degrees: Some parasitic wasps may search for a single species of caterpillar or leaf bud on which to lay their eggs, while mistletoe plants may grow on many different species of trees. Some animals pol-linate a wide variety of plants, and others are limited to a narrow range of choices. Other animals spe-cialize on eating fruits without harming the seeds, which they disperse to a new location. Detritivores eat dead organic matter, mostly the leaves and stems of plants. Within each general category of organ-ism (plant, herbivore, predator, parasite, pollinator, detritivore), specialization allows individuals of a species to fl ourish by avoiding direct competition with other species that have a similar way of making a living. each location within each general type of ecological community has its own unique set of spe-cies interactions, which is constantly changing over the seasons and over the years.
another way in which species coexistence can enhance alpha diversity is through mutualism. Some species create habitats for other species with which they have a mutually benefi cial ecological relation-ship. although mites are frequently found on plants, some leaves produce acarodomatia (mite-houses)
This world map shows the approximate locations of the major ecological communities.
xviii+598_Biodiversity_F.indd 126 11/17/11 1:12 PM

communities, ecological 127
that encourage the presence of a resident population of mites, which may help to protect the leaves from herbivore damage.
Beta diversity consists of species that inhabit dif-ferent, adjacent microhabitats. every habitat, such as a forest or grassland, contains microhabitats, which usually are home to some species that are not found in other parts of the habitat. these microhabitats are considered part of the ecological community, since individuals of many species move in and out of them freely.
One or more microhabitats can be created by something as simple as a large rock. a single rock can create a strikingly different set of environmen-tal conditions upon which some plant and animal species may depend. a large rock in a forest, for example, is likely to discourage the growth of large trees, and the result is a sunny, warm, dry microhab-itat, which lizards prefer to the surrounding forest.
On the other hand, crevices in rocks may be cooler and wetter than the surrounding forest. in the Black Hills of South Dakota, for example, ponderosa pines cast moderate shade on the relatively dry forest floor. Prickly pear cactus lives in the sunny spots on these rocks, right alongside ferns that specialize in the shaded crevices. neither the cacti nor the ferns could thrive in the surrounding soil. Rocks also allow very little soil accumulation, and small plant spe-cies with shallow roots (or mosses and lichens that have no roots at all) can grow on rock surfaces and in crevices without interference from the roots of larger plants. a large rock in dry grassland may have the opposite effect on environmental conditions by creating shaded, moist conditions otherwise absent from the grassland.
consider granite boulders in the foothills of the Sierra nevada of california. the grassland is dominated by grasses and forbs that flourish briefly
This map of the United States shows the approximate locations of the major ecological communities.
xviii+598_Biodiversity_F.indd 127 11/17/11 1:12 PM

first entry128 communities, ecological 128
before the onset of the long summer drought, but some grass and forb species are found only near or on the rocks. the tiny succulent plant Crassula erecta grows only in the shallow pockets of soil created by mosses, while the tiny grass Lamarckia aurea grows in slightly deeper pockets of soil adja-cent to them. the wildflower Mimulus dudleyi grows only in the shallow crevices on these rock surfaces. the only shade-tolerant wildflower species in these grasslands, Pholistoma membranaceum, grows in the shade between boulders. another wild-flower, Phacelia cicutaria, grows almost nowhere else except at the bases of boulders, where water running off the rock surfaces creates a zone of increased moisture. in these same foothills, blue oaks (Quercus douglasii) grow only on hillsides where the boulders are common. Researchers have shown that grasses use water from the soil and can thereby cause oak seedlings to die. Perhaps the oaks grow around boulders because grasses do not grow well there, and the boulders allow water and tree roots to penetrate deeper into the soil. Graz-ing mammals eat oak leaves and damage seedlings. Perhaps the animals do not graze in boulder fields as much as on smooth, grassy hillsides.
Edaphic conditions refer to the material in which a plant grows, usually but not always soil. it can also refer to rocks on which plants grow or the floating mat of moss that covers a bog. normal, rich soil is a complex structure of many components. edaphic conditions, such as unusual soils, can cause an almost entirely different ecological community to dominate over a small area. Rocky or sandy soils are both drier and poorer in nutrients than normal soils. in Michigan, areas of rich loam soil alternate with poor sandy soils left by glaciers when they melted. Beech-maple forests dominate the rich soils, while jack pines dominate the sandy soils. Jack pines are more commonly found in the poorer soils of the boreal forest. Hemlocks (Tsuga canadensis) are often found on poor, rocky soils in the eastern decid-uous forest. in parts of illinois, normal soils also alternate with sand deposits. Black oaks dominate the normal soils, but the sand deposits are so nutri-ent-poor and dry that no trees at all grow on them. instead, grasses that are more typical of the western prairies and even beavertail cacti grow there.
along the coast of northern california, tall coni-fers such as redwoods dominate the normal soils. in some small areas, the soils are very shallow and acidic. in these places, rather than tall redwoods, there are “pygmy forests” of short bishop pines, tiny cypresses, and plants such as rhododendron bushes that are typical of acidic soils. the trees resemble bonsai trees because, just as bonsai trees do, they grow in very poor soils.
Some soils contain toxic minerals that inhibit the growth of most plants. For example, soils derived from serpentine rock contain high levels of chromium and nickel. these rocks resemble those of earth’s mantle rather than the continental crust, and they have been lifted up by the movement of the continental plates. therefore, serpentine deposits are common near california’s San andreas Fault. Some species of plants, or varieties of plants within spe-cies, are able to grow in these toxic soils. there is a cost to this adaptation, however. these plants grow more slowly in normal soils than do normal plants. this is what restricts such plants to the patches of toxic soil.
another unusual soil is diatomaceous earth, an almost perfectly white, powdery soil formed from rock that is made of the shells of billions of dia-toms (single-celled protists) that accumulated on the bottoms of ancient oceans. in the Purísima Hills north of lompoc, california, large deposits of this white dirt support a forest of bishop pines (Pinus muricata). although this tree species is common (maintained by a fire cycle) farther north from this location, it is found nowhere else in the immediate vicinity. the surrounding community is chaparral, which also depends on a fire cycle. the soil factors that allow this unusual plant community have not been identified.
Some microhabitats are created by very small-scale disturbances. in the shortgrass prairies of west-ern north america, grass roots densely pack the soil, and other plants seldom have a chance to grow among the grasses. When bison or prairie dogs dis-turb the grasses, other species of plants can grow there, such as Sphaeralcea coccinea. in the badlands of the South Dakota grasslands, soil erosion may also reduce competition from grasses enough that wildflowers such as Oenothera caespitosa can grow.
even a hole in a tree can create a microhabitat. Many animal species depend on tree holes for parts of their lives. Birds may require these holes for nest-ing, while insect larvae may be unable to live any-where else.
While animals (due to their mobility) specialize on microhabitats less than do plants, the diversity of animal species may depend heavily on the presence of microhabitats that interrupt the dominant species and create openings where they can locate a greater range of food plants and where prey may be easier to capture.
Gamma diversity in a community results from smaller, adjacent ecological communities. they are mixed together enough that it is not useful to distinguish them as separate communities. these differences usually result from major features of the landscape, such as mountains. in the north-
xviii+598_Biodiversity_F.indd 128 11/17/11 1:12 PM

communities, ecological 129
ern Hemisphere, the northeast slopes of mountains receive less direct sunlight and are therefore cooler and wetter than the southwest slopes. in many cases, a forest may grow on the northeast slope, while grassland or desert dominates the southwest slope of the same mountain (see figure). Where two water-sheds come together, wetter conditions may allow the growth of plant species that could not toler-ate drier conditions. in the chaparral of the coastal mountains of california, live oaks (such as Quer-cus agrifolia) and the california bay (Umbellularia californica) are found primarily in such conjoined watersheds, both because of the greater availability of soil moisture and the fact that fires, which are essential for the maintenance of the chaparral, are less likely to burn in these slightly wetter areas.
Gamma diversity intergrades seamlessly into the differences between ecological communities. Because of the striking environmental differences between water and land, aquatic habitats are usually considered separate communities, even when they are very small. a single mountain produces cooler, wetter conditions on the northeast side than the southwest side, but it usually takes a whole range of mountains to produce a rain shadow (see figure on page 130), in which most of the precipitation falls on the windward slopes of the mountains, or to create temperature differences large enough to allow the persistence of subalpine forests and alpine tundra.
communities may differ strikingly from one another not only in the number of species but in their relative abundances. typically, communi-ties that have few species (occupying disturbed or extreme habitats) are dominated by one or a few spe-cies, while the species within diverse communities are much more equal in their relative abundances.
individuals within each species are often located at random with respect to one another. Departures from random distribution reveal some underlying community process. Most often, indi-viduals within a species may be clumped together—that is, they tend to be found close to one another more often than they are distant from one another. this pattern could result from limited dispersal of offspring, such as seeds that do not travel far from the parent plant, or from specialization on patchy resources in the habitat, such as tree seedlings clumping in a patch of sunlight on the forest floor. Occasionally, individuals within a species are more evenly dispersed through the community than one would expect from a random pattern. One example of this is the apparently regular spacing of desert shrubs. this pattern may result from competition between the root system of each shrub and that of the next shrub for access to scarce water and nutri-ent resources.
Within ecological communities, one species may influence the distribution patterns of another. there is disagreement regarding how important competi-tion is in determining the species composition of eco-logical communities. the ecologist Steven Hubbell studied the distribution of tree species on Barro col-orado island in the Panama canal area and found that they were randomly distributed with respect to one another; dispersal, speciation, and extinction were sufficient to explain which kind of tree grew where. in other cases, however, competition plays a vital role in determining which species are pres-ent in a community. During the process of ecologi-cal succession, long-lived, shade-tolerant plants that produce fewer, larger offspring (seeds) replace short-lived, shade-intolerant plants that produce more, smaller offspring due to the competitive superiority of the former. the vital role of competition in the successional development of ecological communi-ties, however, applies more to the general character-istics of the plants rather than to the species of the plant. the ecologist tadami Fukashi and colleagues experimentally manipulated the species composition of grasslands, then allowed secondary succession to occur. the result was a convergence upon simi-lar types of woodlands, but each woodland had its own set of species. competition, therefore, often influences the set of traits but not the actual species composition of the species that make up an ecologi-cal community.
EColoGICal INTEraCTIoNS IN CommuNITIESecological interactions (such as herbivory and pre-dation) not only influence where many of the spe-cies live within an ecological community, but the populations of one species can influence those of
Oak trees (Quercus douglasii) grow on the northeast-facing slopes of the foothills of the Sierra Nevada in California, while grasses, which can tolerate drier con-ditions, grow on the southwest-facing slopes. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 129 11/17/11 1:12 PM

first entry130 communities, ecological 130
another species. the result may be a trophic cascade. Predators may reduce the populations of herbivores on which they prey; in turn, the plants that would otherwise be eaten by the herbivores can grow more. Predators can therefore increase the growth of plants. in general, a trophic cascade occurs when an organism at one level of the food chain promotes the growth of an organism two levels below it.
the result is that species within an ecologi-cal community appear to keep one another under control. this concept is often called the “balance of nature.” the balance of nature, though real, is very approximate, and populations of wild species (especially predators and prey) experience large fluctuations.
the web of relationships (trophic, competi-tive, symbiotic) in an ecological community can be astoundingly complex. in the Origin of Species, charles Darwin provided an example of the complex web of relationships found in even the “simplest” ecological community, a pasture in england. Red clover depends upon bumblebees for pollination, and therefore the production of seeds. Field mice
destroy bumblebee nests, and cats eat mice. Darwin concluded,
Hence it is quite credible that the presence of [cats] in large numbers in a district might deter-mine, through the intervention first of mice and then of bees, the frequency of certain flowers in that district!
in this manner, Darwin described the ecological complexity within which all evolution occurs.
See also biodiversity, patterns of; bogs, life in; boreal and subalpine forests; cloud forests; competition, ecological; continen-tal shelves, biodiversity on; coral reefs; deep oceans, biodiversity in; deserts, cool; deserts, polar; deserts, warm; grasslands and savan-nas; herbivory; marshes and swamps; mutual-ism; parasitism; pollination; rivers, streams, ponds, and lakes; shrublands; succession, ecological; temperate coniferous forests; temperate deciduous forests; temperate dry forests; temperate rain forests; tropical rain
The Sierra Nevada create a rain shadow in California. Most of the rain and snow falls on the Pacific slope, creat-ing lush meadows and forests of giant sequoias (right). After the wind has reached the eastern side of the moun-tains, it is dry, and desert conditions prevail, as seen in this eastward view from the top of Mount Whitney (left). Both the meadow and the desert are at about the same elevation. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 130 11/17/11 1:12 PM

competition, ecological 131
forests and deforestation; tropical seasonal forests; tundra.
Further readingamarasekare, Priyanga. “Spatial dynamics of foodwebs.”
Annual Reviews of Ecology, Evolution, and Systemat-ics 39 (2008): 479–500.
Begon, Michael, colin R. townsend, and John l. Harper. “the nature of the community: Patterns in space and time.” ch. 16 in Ecology: From Individuals to Ecosys-tems, 4th ed. Oxford: Blackwell Publishing, 2006.
Black, R. alan, and Richard n. Mack. “Tsuga canadensis in Ohio: Synecological and phytogeographical rela-tionships.” Plant Ecology 32 (1976): 11–19.
Bukovinszky, tibor, et al. “Direct and indirect effects of resource quality on food web structure.” Science 319 (2008): 804–807.
clarke, K. R., and R. M. Warwick. “Quantifying structural redundancy in ecological communities.” Oecologia 113 (1998): 278–289.
côté, Steeve D., et al. “ecological impacts of deer over-abundance.” Annual Review of Ecology, Evolution, and Systematics 35 (2004): 113–148.
Dunne, Jennifer a., et al. “compilation and network analy-ses of cambrian food webs.” Public Library of Science Biology 6 (2008): e102. available online. URl: http://www.plosbiology.org/article/info:doi/10.1371/journal.pbio.0060102. accessed august 4, 2011.
Farber, Paul lawrence. Finding Order in Nature: The Nat-uralist Tradition from Linnaeus to E. O. Wilson. Balti-more: Johns Hopkins University Press, 2000.
Finke, Deborah l., and William e. Snyder. “niche par-titioning increases resource exploitation by diverse communities.” Science 321 (2008): 1,488–1,490.
Fukashi, tadami, et al. “Species divergence and trait con-vergence in experimental plant community assembly.” Ecology Letters 8 (2005): 1,283–1,290.
George, l. O., and F. a. Bazzaz. “the fern understory as an ecological filter: emergence and establishment of canopy tree seedlings.” Ecology 80 (1999): 833–845.
Gilbert, B., and M. J. lechowicz. “neutrality, niches, and dispersal in a temperate forest understory.” Proceed-ings of the National Academy of Sciences USA 101 (2004): 7,651–7,656.
Gordon, Doria R., and Kevin J. Rice. “competitive sup-pression of Quercus douglasii (Fagaceae) seedling emergence and growth.” American Journal of Botany 87 (2000): 986–994.
Hairston, nelson G., Frederick e. Smith, and lawrence B. Slobodkin. “community structure, population con-trol and competition.” American Naturalist 94 (1960): 421–425.
Hubbell, Steven P. The Unified Neutral Theory of Biodi-versity and Biogeography. Princeton, n.J.: Princeton University Press, 2001.
Kraft, nathan J. B., Renato Valencia, and David D. ack-erly. “Functional traits and niche-based tree commu-nity assembly in an amazonian forest.” Science 322 (2008): 580–582.
Kuchler, a. W. Potential Natural Vegetation of the Conter-minous United States. american Geographical Society, Special Publication no. 36, 1964.
lankau, Richard a., and Sharon y. Strauss. “Mutual feed-backs maintain both genetic and species diversity in a plant community.” Science 317 (2007): 1,561– 1,563.
Petraitis, Peter S., et al. “experimental confirmation of multiple community states in a marine ecosystem.” Oecologia 161 (2009): 139–148.
Slobodkin, lawrence B., Frederick e. Smith, and nelson G. Hairston. “Regulation in terrestrial ecosystems, and the implied balance of nature.” American Naturalist 101 (1967): 109–124.
Stokstad, erik. “On the origin of ecological structure.” Sci-ence 326 (2009): 33–35.
Strauss, Sharon y., and Rebecca e. irwin. “ecological and evolutionary consequences of multispecies plant-ani-mal interactions.” Annual Review of Ecology, Evolu-tion, and Systematics 35 (2004): 435–466.
thébault, elisa, and colin Fontaine. “Stability of ecological communities and the architecture of mutualistic and trophic networks.” Science 329 (2010): 853–856.
Volkov, i., et al. “neutral theory and relative species abun-dance in ecology.” Nature 424 (2003): 1,017–1,028.
Wardle, D. a. “ecological linkages between above-ground and belowground biota.” Science 304 (2004): 1,629–1,633.
Whittaker, Robert H. Communities and Ecosystems, 2nd ed. new york: Macmillan, 1975.
competition, ecological ecological competi-tion occurs when organisms use similar resources and when these resources are in limited supply. competition is an essential component of natural selection. charles Darwin called it “struggle for existence,” a phrase now seldom used because com-petition need not be a visible struggle. competi-tion may result in some individuals growing larger and leaving more offspring than others, and it may result in the deaths of some individuals. Darwin explained how competition results in natural selec-tion, in which the population evolves to resemble the successful competitors.
ecologists generally distinguish between two mechanisms of competition. When a flush of new resources becomes available, populations may grow quickly, and the winners of this kind of competition (scramble competition) are those that can preempt the resources most quickly. When a flush of resources
xviii+598_Biodiversity_F.indd 131 11/17/11 1:12 PM

first entry132 competition, ecological 132
is not available, some individuals can do something that inhibits the growth of other individuals (inter-ference competition). examples of interference com-petition include the production of poisons by plants that enter the soil (allelopathy) and aggression of one animal against another.
competition can occur between individuals within populations of the same species (intraspecific competition). Food can be in short supply in animal populations, as can access to light and soil in plant populations. the shortage of resources contributes to the deaths of most offspring in most populations of plants and animals. Particularly in plant popula-tions, the individuals that (by good luck or superior characteristics) obtain more resources can grow much larger than individuals that do not. intraspe-cific competition can also occur for access to mates. Since plants have more plasticity of growth than do animals, dense plant populations frequently con-tain a large number of small individuals, while in animal populations these individuals would prob-ably die.
While very high densities can have negative effects on individuals within populations, there can also be beneficial effects of moderate crowding. While a large population of prey might be more easily spotted by a predator, it might also be able to defend itself more effectively than an isolated individual. For example, a herd of animals can carry out defensive maneuvers to ward off predators and protect the young. among plants, an isolated individual in a bare field might suffer the consequences of soil erosion and hot dry winds, from which a moderate density of other plants might be able to shield it. the benefits of moderate population density relative to low population density were first pointed out by the ecologist Warder c. allee and are called the Allee effect.
intraspecific competition is often more severe than competition between members of different spe-cies, which is called interspecific competition. this is because individuals within the same species may use almost exactly the same resources, while mem-bers of different species use different resources. Dif-ferent species can coexist and avoid competition not only by using different resources but by using the same resources differently. this idea was concep-tualized by the ecologist G. evelyn Hutchinson in the 20th century as an “n-dimensional hyperspace.” each individual uses numerous resources, and each of these resources can be represented as an axis in a multidimensional hyperspace. Such a concep-tual hyperspace is most commonly called a niche. coexistence is possible if different species occupy niches that do not overlap much. the application of this concept has been limited, however, because no species and no individual occupies an unchang-
ing niche of resources. Phenotypic plasticity allows organisms to adjust to the availability or the short-age of resources, within their genetic limits. Plants that grow best in bright light can adjust their chloro-phyll production and leaf area in a way that allows them to survive in shade. Predators may specialize on a certain type of prey but can switch to another if the preferred prey population becomes critically small. even broad categories such as herbivore and carnivore have a certain amount of flexibility. Most carnivores are actually omnivores that can substitute plant sources for scarce meat to a certain extent. it would surprise most people to learn that herbivores can become carnivores if the opportunity presents itself. People who raise chickens know that they have to use double layers of wire to keep deer from eating the chicks. all of these examples of flexibility indi-cate that hyperspaces and niches are diffuse concepts and therefore of limited use in explaining interspe-cific competition.
among the ways that species that use the same resource may reduce the degree of competition among them are:
● in a dense mixture of several plant species, individuals of some species (for instance, of larger plants) may grow faster than and cast shade over the smaller plants of other species. evolutionary adaptations, how-ever, can allow some of the smaller plants to either avoid or tolerate competition. For example, some of the smaller plant species may have evolved specialized characteristics of shade tolerance. they can therefore sur-vive and reproduce in the shade of the larger plants, rather than competing with them for the light. alternatively, the smaller plants may become active earlier in the spring and complete most of their growth and repro-duction before the larger plants have grown up and cast shade over them.
● consider a certain mixture of plant species in which all the species depend on the same pollinators. if these plant species bloomed simultaneously, not only might there not be enough pollinators to visit all of them, but the pollinators might transfer pollen among rather than within species. if each plant spe-cies blooms at a different time, however, both of these competition-related problems are avoided.
● lions, hyenas, and vultures all eat wilde-beest meat. they avoid competition, how-ever, because the lion kills the wildebeest and eats first, the hyenas clean up the leftovers, and vultures consume the rotting meat the
xviii+598_Biodiversity_F.indd 132 11/17/11 1:12 PM

competition, ecological 133
hyenas do not want. Some biologists specu-late that early species of humans, such as Homo ergaster, avoided competition with lions and hyenas because they used stone tools to break open bones and consume bone marrow from animals killed by lions, after the hyenas had finished with them.
a species that competes with another for resources may benefit from making the resources unfit for use by the competing species. For instance:
● Many species of plants (such as walnut trees and california sage bushes) release toxic chemicals into their immediate environ-ment. in some cases, these toxic chemicals (allelochemicals) may inhibit the growth of other plants.
● Bacteria and fungi may release toxic chemi-cals as they putrefy foods, thereby prevent-ing animals from consuming the food items upon which they are growing. this is the principal reason that meats and fruits smell bad when they rot: the bacteria and fungi are protecting themselves by preventing ani-mals from eating the food in which they are already living.
Sometimes, competition can displace individu-als of a species from their optimal habitat into one for which they are not as well suited. this occurs because competition is usually most intense where resources are most abundant, not where they are most scarce. three examples follow.
● in general, competition is stronger among species in tropical rain forests than in temperate forests and in temperate forests than in boreal forests. in many cases in the northern Hemisphere, northern spe-cies will survive in southern climates if they are transplanted and if they are protected from competition; their southern distribu-tion is limited by competition. in contrast, the northern distribution of most northern Hemisphere species is limited by the ability to survive cold winter conditions.
● Barnacles that live on rocks in the intertidal zone of the seashore must close their shells and cannot obtain food while exposed to low tide. the lower portion of the intertidal zone is inundated by ocean water more of the time than the upper portion; for the bar-nacle species living on rocks, the lower por-tion of the zone represents higher resources. Research by the ecologist Joseph connell
showed that Balanus barnacles dominate the lower intertidal; they cannot survive well in the upper intertidal zone, but they are the superior competitors in the lower inter-tidal zone. Chthamalus barnacles dominate the upper intertidal, even though they sur-vive well when transplanted into the lower zone and are protected from competition. For both kinds of barnacles, dry conditions determine the upper limit of growth, and for Chthamalus, competition determines the lower limit of growth.
● the knotweed Polygonum pensylvanicum grows in recently disturbed areas in the mid-western United States. living in a disturbed area is itself a way of escaping competition. the knotweed, however, escapes competi-tion from other weeds by growing in wet areas that other weeds cannot tolerate as well. When grown in isolation, knotweeds prefer the same moderately moist conditions that the other weeds prefer, but it cannot compete with them.
interference competition may perpetuate the effects of a chance event. in the shade of boulders or of live oaks in the arid hills of Southern califor-nia, populations of the herb Pholistoma auritum may become established. Once established, this herb produces allelochemicals that inhibit the growth of grasses that commonly grow in the shade of oak trees. the initial establishment of the population under some oaks and not under others may have been a chance event.
competition can also cause a species to use its resources in a way that is not optimal. Many fruits entice animals to eat them and to disperse their seeds to new locations. they do this by making the fruits soft, sweet, colorful, and flavorful. the fruits have these characteristics only after the seed is mature. Plants discourage animals from eating unripe fruits by making these fruits hard, green, and distasteful. How-ever, frugivorous animals that consume the fruits first will get more food resources than an animal of the same or different species that waits until the fruits are fully ripe. Frugivorous animals therefore frequently consume fruits that are not fully ripe. in some cases, as with parrots, they need to eat dirt, which helps to inactivate toxins from the unripe fruits.
Recurring disturbances can maintain conditions in which some species remain free from competition for long periods of time. One common example is an overgrazed pasture. in the south central United States, weeds such as milkweed (Asclepias viridis), spurge (Euphorbia marginata), and bull nettle (Cni-doscolus texanus) would normally be smothered by
xviii+598_Biodiversity_F.indd 133 11/17/11 1:12 PM

first entry134 competition, ecological 134
competition from grasses. cattle will not eat these plants, which are either poisonous or have irritat-ing hairs. the cattle eat the grasses, which liberates resources that the weeds use.
One of the resources for which there is the most competition is access to mates. this competi-tion leads to sexual selection, a process also first described by Darwin. Many species of birds have evolved astonishing coloration and courtship behav-iors, and primate species have evolved intricate social interactions, all as mechanisms that reward individuals most successful in the competition for mates. even plants compete for mates. in many plant species that have separate male and female individu-als (such as cottonwoods and hollies), the male flow-ers open earlier than the female flowers. the first male flowers to open after the female flowers may be able to pollinate a large number of them. the risk of being the first male flower to open is that the flower may open too early.
competition has contributed to the evolution of biodiversity also by allowing the evolution of species that tolerate harsh conditions. there are many thousands of species that tolerate cold, dry, salty, or toxic conditions, where they encounter less competition. examples range from desert kangaroo rats, which obtain nearly all their moisture from the dew that condenses on the plants that they eat, to Tidestromia oblongifolia, a plant that lives nearly alone in the bottom of Death Valley, to fishes that live in water so cold they are supercooled and run the risk of freezing solid if they touch a rock. the champions are the extremophile microbes. Some algae live in salt slush, while many bacteria live in superheated water or extreme acidity or toler-ate high levels of nuclear radiation. although they appear simple, extremophiles have amazing bio-chemical adaptations. One reason they do this is that they have persisted in those places from the archaean era, when the entire earth had those con-ditions, but they also persist because it allows them an escape from competition.
While competition, or the avoidance of it, has stimulated the evolution of many species, it can also prevent evolution. in a forest with woodpeckers, any other kind of bird that attempts to make a liv-ing by extracting insects from inside tree bark may have a difficult time competing with the woodpeck-ers. On the Galápagos islands, however, there are no woodpeckers. One of the species of Darwin’s finches, the woodpecker finch, uses cactus spines to extract insects from bark. it has evolved into a new species, with its own “niche,” because the woodpecker way of making a living was an unfilled occupation in the natural economy of the islands. the Galápagos islands are a laboratory of evolution
precisely because the immigrant species have been free from competition long enough to evolve new adaptations.
Because organisms specialize on different sets of resources at different times, the resources of the bio-logical world are used more efficiently than would be the case if all species competed for exactly the same resources. Some species live on the resources that other species have left. the avoidance of com-petition allows the world to have a greater weight, number, and diversity of organisms than it would otherwise have. However, each organism specializes on its peculiar set of resources not in order to help the world but to maximize its own reproductive suc-cess or fitness.
CompETITIoN aNd EColoGICal CommuNITIESDisagreement continues on the extent to which com-petition influences the species composition of entire ecological communities. can competition actually keep some species out of an ecological community? the evolutionary scientist Jared Diamond studied birds on islands in Southeast asia and determined that the black honey-eater (Myzomela pammelaena) did not coexist with the black sunbird (Nectarinia sericea) on the same islands. He claimed that there are “assembly rules” that determine which spe-cies are compatible and which are not. if two spe-cies (such as these two birds) are too similar in the way they obtain resources, one will out-compete the other. in contrast, the ecologist Daniel Simber-loff said that the distribution of most bird species on these islands was random with respect to one another. Diamond’s response was that although the initial colonization of each island by any bird species may have been a random event, subsequent competi-tion eliminated inferior competitors as well as newly arrived species on an island that already contained a species too similar to it.
One ecological process that would seem to be largely dependent on competition is ecological suc-cession. the species that dominate ecological com-munities late in the process of succession are mostly the superior competitors, yet there is also a ran-dom element in this process. the ecologist tadami Fukashi and colleagues manipulated experimental grasslands and allowed them to undergo secondary succession. all the fields showed the same pattern of convergence upon forests (dominance by large, long-lived, shade-tolerant plants), but the actual species composition of the resulting forest depended strongly upon the initial composition of the grassland. they concluded that there were competition-based assem-bly rules for plant traits but not for plant species.
in many cases, the species that first colonizes a new habitat—for example, a newly created volcanic
xviii+598_Biodiversity_F.indd 134 11/17/11 1:12 PM

conifers 135
island—is the winner in competition against spe-cies that arrive later, even if these species are not as similar to one another as black honey-eaters and black sunbirds. the ecologist Peter Petraitis and col-leagues have shown that rocky seashores in Maine can be dominated either by seaweeds or by mussels, depending on which of them were able to colonize newly exposed rock surfaces more quickly. the win-ner in ecological competition is, therefore, frequently the species that arrives first.
See also communities, ecological; natural selection.
Further readingBais, H. P., et al. “allelopathy and exotic plant invasion:
From molecules and genes to species interactions.” Sci-ence 301 (2003): 1,377–1,380.
connell, Joseph H. “the influence of interspecific com-petition and other factors on the distribution of the barnacle Chthamalus stellatus.” Ecology 42 (1961): 710–723.
———. “Diversity and the coevolution of competitors, or the ghost of competition past. Oikos 35 (1980): 131–138.
———. “On the prevalence and relative importance of interspecific competition: evidence from field experi-ments. American Naturalist 122 (1983): 661–696.
———. “apparent versus ‘real’ competition in plants.” in Perspectives on Plant Competition, edited by J. B. Grace and David tilman, pp. 9–26. new york: aca-demic Press, 1990.
Diamond, Jared M. “assembly of species communities.” in Ecology and Evolution of Communities, edited by Martin l. cody and Jared M. Diamond, pp. 342–444. cambridge, Mass.: Harvard University Press, 1975.
ebert, e., et al. “Dose effects and density-dependent regula-tion in two microparasites of Daphnia magna.” Oeco-logia 122 (2000): 200–209.
Fukashi, tadami, et al. “Species divergence and trait con-vergence in experimental plant community assembly.” Ecology Letters 8 (2005): 1,283–1,290.
Gause, G. F. The Struggle for Existence. Baltimore: Wil-liams and Wilkins, 1934.
Hutchinson, G. evelyn. “concluding remarks.” Cold Spring Harbor Symposium on Quantitative Biology 22 (1957): 415–427.
Janzen, Daniel H. “Why fruits rot, seeds mold, and meat spoils.” American Naturalist 111 (1977): 691–713.
Muller, cornelius H. “the role of chemical inhibition (alle-lopathy) in vegetational composition.” Bulletin of the Torrey Botanical Club 93 (1966): 332–351.
Parker, V. thomas, and cornelius H. Muller. “allelo-pathic dominance by a tree-associated herb in a california annual grassland.” Oecologia 37 (1979): 315–320.
Parrish, Judy a. D., and Fakhri a. Bazzaz. “Underground niche separation in successional plants.” Ecology 57 (1976): 1,281–1,288.
Petraitis, Peter S., et al. “experimental confirmation of multiple community states in a marine ecosystem.” Oecologia 161 (2009): 139–148.
Reeve, J. D., et al. “Scramble competition in the southern pine beetle, Dendroctonus frontalis.” Ecological Ento-mology 23 (1998): 433–443.
Stoll, P., and D. Prati. “intraspecific aggregation alters inter-specific interactions in experimental plant communi-ties.” Ecology 82 (2001): 319–327.
Stowe, l. G. “allelopathy and its influence on the distribu-tion of plants in an illinois old-field.” Journal of Ecol-ogy 67 (1979): 1,065–1,085.
Watkinson, a. R., and J. l. Harper. “the demography of a sand dune annual: Vulpia fasciculata. i. the natu-ral regulation of populations.” Journal of Ecology 66 (1978): 15–33.
conifers conifers are seed plants that produce pollen in staminate (male) cones and seeds in pistil-late (female) cones. a cone is a compact branch that produces short, thick leaves called bracts, under-neath which pollen or seeds develop (see figure on page 136). the terms male and female strictly refer to the pollen and to the unfertilized seeds. coni-fers are trees or shrubs, not vines or herbaceous plants. conifers are one of the evolutionary lin-eages, along with cycads, ginkgoes, and Gnetales (and some extinct lineages) often considered to be gymnosperms (Greek for naked seed), so called because the seeds are not completely surrounded by parental tissue while they develop. conifers usually produce female cones near the tips of branches and near the top of the tree and male cones farther back on the branch or nearer to the base of the tree. this arrangement maximizes the chances for the wind to carry pollen from the staminate cones of one tree to the pistillate cones of another. if the staminate cones were above the pistillate cones, most of the pollen would fall on the pistillate cones of the same tree. conifers are pollinated by wind.
Most conifers have needlelike or scalelike leaves. they evolved during the Mesozoic era in relatively cold and dry regions, and they still live in such places, though they are not found in deserts. their small, thick leaves expose relatively little surface area to desiccation. this is an especially important adapta-tion during the long, cold winters that most conifers experience, for example in the boreal and subalpine forests. needlelike leaves also absorb light from all directions, which may also be useful for trees that grow in mountainous areas with many exposed rock surfaces. these leaves have a photosynthetic rate
xviii+598_Biodiversity_F.indd 135 11/17/11 1:12 PM

first entry136 conifers 136
about half that of the thin, broad leaves found on deciduous trees. Over a leaf’s lifetime, however, an evergreen leaf with a low photosynthetic rate may produce as much food as a deciduous leaf with a high photosynthetic rate.
conifers reproduce slowly. When the staminate cones release pollen, wind blows some of it to small, tender, purplish pistillate cones of another tree. the pollen grains germinate, grow tubes into the pistillate cone tissue, penetrate the ovules (immature seeds) inside, and deposit a sperm nucleus that fuses with the egg. the growth of the tubes takes about a year, and the eggs are not fertilized until the pistillate cone has grown to be much larger and green. Only in the third year do the pistillate cones become hard and brown, at which time the bracts open and release mature seeds into the wind. Some conifers have undergone coevolution with birds such as nutcrackers that pry open the cones, remove the seeds, and then store them in caches in surrounding rocks. the stored seeds that the birds misplace grow into new trees. the cones of these trees, such as the whitebark pine (Pinus albicau-lis; see figure on page 137), produce sticky resin that prevents other animals from getting the seeds.
even though they reproduce slowly, some coni-fers can grow rapidly. Some pines that are adapted to a fire cycle, such as the bishop pine (Pinus muricata) of coastal california, can produce cones in just a few years after the trees germinate. in many regions that are dominated by temperate deciduous forest, pines grow rapidly in recently disturbed areas. the pines cannot germinate and grow in shade; shade-tolerant deciduous trees grow up underneath them and replace them.
conifers such as podocarps and auracarians that most closely resemble the primitive ones that evolved in the Mesozoic era live in the Southern Hemisphere.
the evolutionary lineages that have undergone more change and diversification are very common in the northern Hemisphere. Some important families of conifers are:
Podocarpaceae. Podocarps live in continents of the Southern Hemisphere. Some of the kauri trees (Agathis australis) of new Zealand have trunks that are 15 feet (5 m) thick at 150 feet (30 m) above the ground.
Araucariaceae. araucarians also live in the Southern Hemisphere. they include the monkey puzzle trees (Araucaria araucana) of chile, whose relatively wide, flat needles are very sharp and stiff; the norfolk island Pine (Araucaria heterophylla) of the australian region; and the Wollemi pine (Wol-lemia nobilis), which was known only from the fossil record until its recent discovery in a national park in australia.
Taxaceae. the yews (genus Taxus) are often understory trees or shrubs of moist forests in the northern Hemisphere. their soft needles are rela-tively flat. the female seeds do not develop in cones but exposed on branches. the seed coat often has a red swelling called an aril, which attracts animals that eat and disperse them.
Taxodiaceae. the sequoia family contains the largest trees in the world. the most massive trees in the world are the giant sequoia trees (Sequoiaden-dron giganteum) of the Sierra nevadas of califor-nia. the General Sherman tree in Sequoia national Park has a trunk that is 36 feet (12 m) thick at the base and still 14 feet (more than 4 m) thick at a height of 180 feet (60 m). the coast redwoods (Sequoia sempervirens) of southern Oregon and northern california are the tallest trees in the world; 26 of them are known to exceed 360 feet (120 m) in height. the dawn redwood (Metasequoia glyp-
Pines, such as these ponderosa pines in the Black Hills of South Dakota (Pinus ponderosa), produce pollen in male cones (left) and seeds in female cones (right, here shown at their pollen-receptive stage). (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 136 11/17/11 1:12 PM

conifers 137
tostroboides) was thought to be extinct until it was found growing in the mountains of china during the 20th century. Bald cypresses (Taxodium distichum), so called because they are deciduous, are important swamp trees in the southeastern United States.
Cupressaceae. Most of the cypresses, cedars, and junipers have short, waxy needles or scales that press against the branches. Species such as the incense cedar (Calocedrus decurrens) of the Sierra nevadas are relatives of conifers that dominated ancient temperate coniferous forests. the Jomon Sugi tree of yakushima island (Cryptomeria japon-ica) is one of the largest trees in Japan. Juniper shrubs (genus Juniperus) are the most drought-toler-ant conifers in north america. the pistillate cones of junipers remain fleshy when mature and look like berries. Birds eat these cones and disperse the seeds.
Pinaceae. trees of this family dominate thou-sands of square miles. it is one of two plant families
(the other is the grass family) visible by humans from outer space. important genera in the pine family include the following:
● Spruces (genus Picea) have pistillate cones that hang downward and needles inserted individually on the stems. Spruces dominate the boreal and most subalpine forests.
● Firs (genus Abies) have pistillate cones that point upward, and each of their individually inserted needles has a woody peg.
● Hundreds of species of pines (genus Pinus) have relatively long needles in clusters of two to five, wrapped with a membrane at the base. (the exception is Pinus monophylla of the Great Basin mountains of north amer-ica, which has just a single needle in each bundle.) Pines, firs, and Douglas firs (Pseu-dotsuga menziesii) dominate the montane forests of the american West. Pines such as the foxtail pine (Pinus flexilis) dominate dry subalpine forests of the Rocky Mountains and the Sierra nevadas.
● larches (genus Larix) are deciduous and live in moist areas of the boreal forests.
● the hemlocks (genus Tsuga) are often small trees associated with coniferous or hard-wood forests in eurasia and north america.
the oldest trees in the world are the bristlecone pines (Pinus longeava) of the White Mountains of california. they live not only at high elevation but in the rain shadow of the Sierra nevadas. they tol-erate cold, dry conditions by growing very slowly. each new layer of wood may be only a third of a millimeter (a few hundredths of an inch) thick, and the needles may survive for several decades (see figure on page 138). their very long record of tree rings makes them very useful in the study of past climate. Many bristlecone pines exceed 4,000 years of age.
Many conifers are called “softwoods” because they produce light wood and are the mainstay of the timber industry. timber companies manage many thousands of square miles of pine plantations, some of which have been bred for rapid and uniform growth.
Some conifer species are very rare. this includes the dawn redwood of china and Torreya taxifolia, a tree relative of the yew that lives near the border of Florida and Georgia. their populations consist of only some 2,000 trees. Several species of pines that live only in certain coastal forests of california are also rare. their cones will not open unless a fire burns the forest. the torrey pine (Pinus torreyi) is found only in a few patches near San Diego, and the
The seeds of whitebark pines (Pinus albicaulis) are dispersed to new locations by Clark’s nutcrackers (Nucifraga columbiana) in the subalpine forests of California. The birds plant the seeds and eat some of them later, but some of the seeds grow into new trees. The pines (like several other subalpine pine species) produce thick sap that prevents squirrels from eating the seeds; only nutcracker birds can open them. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 137 11/17/11 1:12 PM

first entry138 Connell, Joseph138
Monterey pine (Pinus radiata) only in a few patches near the Monterey Peninsula. though rare in its native habitat, the Monterey pine has become an aggressively invasive species in new Zealand, where it was planted for timber production.
See also boreal and subalpine forests; disturbances, ecological; seed plants; suc-cession, ecological; temperate coniferous forests; temperate deciduous forests.
Further readingamerican conifer Society. “conifer Society Home Page.”
available online. URl: http://www.conifersociety.org. accessed July 9, 2010.
Dickie, ian a., et al. “co-invasion by Pinus and its mycor-rhizal fungi.” New Phytologist 187 (2010): 475–484.
Farjos, aljon. A Natural History of Conifers. Portland, Oreg.: timber Press, 2008.
Gymnosperm Database. “Gymnosperm Database.” avail-able online. URl: http://www.conifers.org. accessed July 9, 2010.
lanner, Ronald M. The Bristlecone Book: A Natural His-tory of the World’s Oldest Trees. Missoula, Mont.: Mountain Press, 2007.
———. The Piñon Pine: A Natural and Cultural History. Reno: University of nevada Press, 1981.
llorens, laura, et al. “Water-use responses of living fossil conifers to cO2 enrichment in a simulated cretaceous polar environment.” Annals of Botany 104 (2009): 179.
Schwartz, Mark W., et al. “Population persistence in Flor-ida Torreya: comparing modeled projections of a declining coniferous tree.” Conservation Biology 14 (2000): 1,023–1,033.
Connell, joseph See competition, ecological.
conservation biology conservation biology is the interdisciplinary study of all levels of biodi-versity, from populations to large-scale ecological communities, with the aim to rescue them from
Bristlecone pines (Pinus longeava) in the White Mountains of California grow very slowly for thousands of years. The trunk of this tree, which is probably more than 4,000 years old, is more than a meter in diameter. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 138 11/17/11 1:12 PM

conservation biology 139
human destruction and to do so, if possible, without harming human economic activities. conservation biology therefore not only includes many biological disciplines (from genetics to ecology) but also human sciences such as economics, political science, and history. it is a very broad and urgent area of scien-tific study. For many of its component sciences, the main question is “How can people understand the way the world works?” But for conservation biol-ogy, the main question is “What can people do to save the natural world?” in order to accomplish this, conservation biologists must bring together infor-mation and concepts from all of these fields. More information about conservation biology is available in numerous entries in this encyclopedia. Most of the biographical entries in this encyclopedia are on people who have had or continue to have a major impact on conservation.
Population genetics and reproduction. conser-vation biologists do not just save species but must save populations of species that contain enough genetic variability. Genetic variation allows the population to avoid the negative effects of inbreed-ing and provides the basis for further evolutionary adjustments to environmental changes. it is also nec-essary to determine how successfully the popula-tions are reproducing. For example, the Oklahoma subspecies of the seaside alder (Alnus maritima ssp. oklahomensis) consists of a few thousand large shrubs, each of which may produce hundreds of seeds in a typical year. However, few of these seeds germinate, and almost none of them grow into new shrubs. therefore, to preserve population genetic variation, nearly the entire population of shrubs must be protected.
Habitat conservation. it is seldom possible to protect the entire range of any species. it is there-fore necessary for conservation biologists to identify habitats within the species’ range that are relatively undisturbed, that are least likely to be threatened by human activity in the near future, and in which the species have sufficient genetic diversity. One of the major sources of habitat disruption is invasive species, which can greatly alter the habitat as they spread unchecked, even within protected habitats. the habitats that have highest conservation priority are the biodiversity hot spots, which contain a large number of endemic species (species found nowhere else). Because elements of the human landscape, such as cities, farms, and roads, break the species range into small fragments, a protected habitat should be as large as possible or connected by natural cor-ridors that allow populations in one fragment to mingle with those in another. Sometimes a habitat might be split in such a way that amphibians need to go from one fragment to the other in order to
complete their life cycle; in such cases corridors are particularly important. examples of corridors that connect protected areas include:
● the yellowstone to yukon conservation initiative, which connects habitats along a 2,000 mile (3,200 km) line
● the Mesoamerican Biological corridor, which covers 80,000 square miles (208,000 square km), or 27 percent of central america
Just setting aside a protected area, even if it is not a fragment, may not be enough to save the populations that live in it. if the protected area is long and narrow, it may be too exposed to human-altered habitats. Many species can survive only deep within an undisturbed habitat, such as a forest, and cannot survive near the edge. this is known as the edge effect and has often resulted in “empty forest syndrome,” in which the common species survive but the rare species vanish.
in some cases, economic interests so greatly threaten a habitat that protection is nearly impos-sible. in these cases, however, government agencies can require a developer to protect or even to attempt to re-create a similar habitat elsewhere. this process is known as mitigation banking and is usually used in the conservation of wetland areas.
Extinction. One of the highest priorities of con-servation biology is to prevent extinction. this is particularly important for keystone species, which influence the structure and function of entire ecolog-ical communities. extinction of species may be pre-vented by saving their habitats, but this will prove ineffective if global warming causes the conditions of those habitats to change such that they are no lon-ger suitable for the survival of the species they were intended to protect. Scientists expect global warm-ing to cause the extinction of many species despite intense efforts at habitat conservation. the “sixth mass extinction” is currently under way, mainly as a result of habitat destruction, and global warming will only intensify it.
Economic benefits and social justice. an emphasis on the economic benefits of wild habitats will help generate support for saving them and for reducing global warming as well. the benefits are not only for large corporations, such as pharmaceu-tical companies that can discover new drugs from plant leaves or frog skins in rain forests, but for the livelihood of native peoples that live in or near the natural habitats. issues of large-scale social justice may also arise from the lack of habitat conservation. if a private corporation cuts down a hillside forest, is it liable for the damage to downslope villages if the deforestation allows floods and mudslides to occur?
xviii+598_Biodiversity_F.indd 139 11/17/11 1:12 PM

first entry140 conservation biology 140
according to some political leaders, destruction of habitats are not only “crimes against nature” but against humans as well.
Education. conservation of biodiversity is pos-sible only if people know why biodiversity is impor-tant, but even this is not enough. Students may learn in a classroom all about plants and animals yet not support or participate in the measures nec-essary to protect them. it is much more effective—as well as more enjoyable—for students to go out into natural habitats, explore their biodiversity, and think about the ecological processes occurring in them. even the moderate discomfort of ticks and spiders may instill in them a greater sense of respect for nature.
it is also important for issues of biodiversity conservation to be integrated into an entire educa-tional curriculum, including topics such as history and literature, rather than to be isolated into a single class that a student might not take or later forget. education is the source of future conservation biolo-gists, but even more importantly, of a citizenry that understands the importance of conservation.
Diplomacy. Biodiversity and other conservation issues do not respect national borders any more than do the animals that migrate or the seeds that blow over them. Meaningful conservation requires inter-national cooperation. conservation issues are, at one and the same time, the issues most easily brushed aside by diplomats who work on international issues and the issues that most seldom cause disagreement. Most nations agree that species must be saved, and most nations give these species little attention. the good news is that international conservation agree-ments between nations that have problems getting along may pave the way for agreements on the issues that cause the most friction between them. How-ever unlikely a prospect this may be, conservation biologists are professionally optimistic that such agreements must at least be attempted. international agreements with narrowly defined goals, such as the convention on international trade in endangered Species (citeS), have had more success than agree-ments that are broader in scope or have greater economic impact. For example, meaningful carbon emissions agreements remain elusive. the United nations has established the convention on Biologi-cal Diversity (cBD), an attempt to facilitate interna-tional agreements to protect endangered species. the cBD declared 2010 to be the international year of Biodiversity.
See also biodiversity, resources for agri-culture; biodiversity, resources for medi-cine; ecosystem services; endangered species; extinction; federal government role in con-servation (United States); greenhouse effect
and biodiversity; habitat fragmentation; integrated pest management (ipm); invasive species; mass extinctions; population genet-ics; restoration ecology.
Further readingBecker, carlos Guilherme, et al. “Habitat split and the
global decline of amphibians.” Science 318 (2007): 1,775–1,777.
Bowen, B. W. “Preserving genes, species, or ecosystems? Healing the fractured foundations of conservation policy.” Molecular Ecology 8 (1999): S5–S10.
Brooks, t. M., et al. “Global biodiversity conservation pri-orities.” Science 313 (2006): 58.
Budd, ann F., and John M. Pandolfi. “evolutionary novelty is concentrated at the edge of coral species distribu-tions.” Science 328 (2010): 1,558–1,561.
chan, K. M. “Value and advocacy in conservation biology: crisis discipline or discipline in crisis?” Conservation Biology 22 (2008): 1–3.
costanza, Robert, et al. “the value of the world’s ecosys-tem services and natural capital.” Nature 387 (1997): 253–260.
crowder, larry B., and elliott a. norse. Marine Conser-vation Biology: The Science of Maintaining the Sea’s Biodiversity. Washington, D.c.: island Press, 2005.
Gaston, Kevin J., et al. “the ecological performance of pro-tected areas.” Annual Reviews of Ecology, Evolution, and Systematics 39 (2008): 93–113.
Gibson, J. Phil, Stanley a. Rice, and clare M. Stucke. “comparison of population genetic diversity between a rare, narrowly distributed species and a common, widespread species of Alnus (Betulaceae). American Journal of Botany 95 (2008): 588–596.
Kareiva, Peter, and M. Marvier. “conserving biodiversity coldspots.” American Scientist 91 (2003): 344–351.
Kennedy, Robert F., Jr. Crimes Against Nature. new york: Harpercollins, 2004.
Koh, l. P., et al. “Species coextinctions and the biodiversity crisis.” Science 305 (2004): 1,632–1,634.
leroux, S. J., and F. K. Schmiegelow. “Biodiversity concor-dance and the importance of endemism.” Conserva-tion Biology 21 (2007): 266–268.
luck, Gary W., Gretchen c. Daily, and Paul R. ehrlich. “Population diversity and ecosystem services.” Trends in Ecology and Evolution 18 (2003): 331–336.
Margules, c. R., and R. l. Pressey. “Systematic conserva-tion planning.” Nature 405 (2000): 243–253.
May, Robert l., and John lawton. Extinction Rates. Oxford: Oxford University Press, 1995.
Mccallum, M. l. “amphibian decline or extinction? cur-rent declines dwarf background extinction rate.” Jour-nal of Herpetology 41 (2008): 483–491.
Mikkelson, G. M., et al. “economic inequality pre-dicts biodiversity loss.” Public Library of Science ONE 2 (2007): e444. available online. URl: http://
xviii+598_Biodiversity_F.indd 140 11/17/11 1:12 PM

continental shelves, biodiversity on 141
www.plosone.org/article/info:doi/10.1371/journal.pone.0000444. accessed august 4, 2011.
Molnar, J., et al. “the sum is greater than the parts.” Con-servation Biology 18 (2004): 1,670–1,671.
Myers, norman, et al. “Biodiversity hotspots for conserva-tion priorities.” Nature 403 (2000): 853–858.
naidoo, R., et al. “Global mapping of ecosystem ser-vices and conservation priorities.” Proceedings of the National Academy of Sciences USA 105 (2008): 9,495–9,500.
Orr, David W. Ecological Literacy: Education and the Tran-sition to a Postmodern World. albany: State Univer-sity of new york Press, 1992.
Primack, Richard B. Essentials of Conservation Biology. Sunderland, Mass.: Sinauer associates, 2006.
Rice, Stanley a., and J. Phil Gibson. “is seedling establish-ment very rare in the Oklahoma seaside alder, Alnus maritima ssp. oklahomensis?” Oklahoma Native Plant Record 9 (2009): 59–63.
Sachs, Jeffrey D., et al. “Biodiversity conservation and the millennium development goals.” Science 325 (2009): 1,502–1,503.
Society for conservation Biology. “Society for conservation Biology.” available online. URl: http://www.conbio. org. accessed July 11, 2010.
terborgh, John. Requiem for Nature. Washington, D.c.: island Press, 1999.
Underwood, e. c., et al. “Protecting biodiversity when money matters: maximizing return on investment.” Public Library of Science ONE 3 (2008): e1515. available online. URl: http://www.plosone.org/arti-cle/info:doi/10.1371/journal.pone.0001515. accessed august 4, 2011.
United nations. “convention on Biological Diversity.” available online. URl: http://www.cbd.int. accessed July 11, 2010.
Wake, D. B., and V. t. Vredenburg. “are we in the midst of the sixth mass extinction? a view from the world of amphibians.” Proceedings of the National Academy of Sciences USA 105 (2008): 11,466–11,473.
Wilson, edward O. “On the future of conservation biol-ogy.” Conservation Biology 14 (2000): 1–3.
Wood, c. c., and M. R. Gross. “elemental conservation units: communicating extinction risk without dictat-ing targets for protection.” Conservation Biology 22 (2008): 36–47.
Woods institute for the environment. “aldo leopold lead-ership Program.” available online. URl: http://leopold leadership.stanford.edu/. accessed July 11, 2010.
continental shelves, biodiversity on con-tinental shelves are the undersea edges of continents. they are covered by shallow water and have at times of lower sea level in the past (for example, during the ice ages) been exposed as dry land. therefore, con-
tinental shelves often contain many fossils and even some archaeological deposits. Some terrestrial areas that were once continental shelves now exist in the middle of each continent. the edges of the continental shelves drop precipitously to the deep ocean floor. large areas of continental shelf are found between alaska and Siberia, off the coasts of Florida and yuca-tán in the Gulf of Mexico, off the coast of argentina, throughout the north Sea, in Hudson Bay, in much of Southeast asia, and off the north coast of australia.
continental shelves have abundant marine life because sunlight can penetrate the shallow water and because of mineral nutrients that erode from the continents. excessive nutrient load from the conti-nents, however, can cause eutrophication and reduce biodiversity. the most productive fishing banks are continental shelves. the shallow water supports not only visits from open ocean animals such as fishes and whales but also animals such as sea otters (Enhydra lutris) that do not venture far into the sea.
among the most diverse of the continental shelf habitats are:
Kelp forests. Dense stands of kelp (brown algae) grow on many temperate continental shelves, espe-cially near areas where nutrient-rich water rises from the deep oceans. Many of these seaweeds are anchored to the bottom and extend perhaps hun-dreds of feet up to the surface, held upright by bladders that contain gas (see figure on page 142). Species of Laminaria dominate kelp forests on both sides of the atlantic Ocean and the coasts of china and Japan; Ecklonia dominates the kelp forests of australia, new Zealand, and South africa; Macro-cystis primarily dominates the Pacific kelp forests. the northeastern Pacific Rim, from california to the aleutian islands, has the kelp forests with the great-est diversity of seaweed species. Kelp forests, like ter-restrial forests, have microhabitats, such as canopy and shaded understory, with their own species of seaweeds and animals.
Kelp forests provide examples of keystone spe-cies that operate up or down the food web. Kelp is a keystone species, not only forming (along with photo-synthetic plankton) the base of the food web but also creating surfaces on which many animals (and other algae) live. However, top predators are also keystone species. Overfishing can remove predatory fishes that normally control populations of herbivorous animals. When predators such as sea otters become rare, popu-lations of their prey (mostly sea urchins) explode and eat many of the kelp strands, particularly the young ones. in some cases, the result is “urchin barrens.”
Coral reefs. coral reefs have tremendous bio-diversity not only because of the penetration of sunlight into warm waters, but because the coral animals themselves build the reefs. Seaweeds are
xviii+598_Biodiversity_F.indd 141 11/17/11 1:12 PM

first entry142 continental shelves, biodiversity on 142
rare in coral reefs because the corals themselves and other animals consume young seaweeds. Dense populations of small seaweeds can cover coral reefs when eutrophication occurs. if disturbance kills the corals, populations of starfish, which had formerly been controlled by the corals, may explode.
Intertidal zone. the intertidal zone is the shore-line that is exposed to the air at low tide and covered by the ocean at high tide. Most permanent resi-dents of the intertidal zone are invertebrates, such as barnacles and bivalves, that can close their shells when they are exposed to air. competition is stron-gest in the lower intertidal zone, where the animals can obtain food from the ocean except during brief periods. animals that cannot withstand temporary dry conditions, such as sea anemones, crabs, and echinoderms, may be abundant in tidepools, where seawater is trapped.
continental shelves need to be protected from pollution and from overharvesting of organisms such as fishes and kelp. Most preserved natural areas are on land, but in recent decades governments around the world have begun to establish marine biodiversity preserves in continental shelf areas as well as around deep sea volcanic islands. the larg-est marine preserve in the United States is off the coast of Hawaii. Several national parks exist pri-
marily to protect continental shelf and other marine areas. Protection of continental shelf areas has been more successful than the protection of deep sea areas because continental shelves are usually within a sin-gle nation’s jurisdiction.
CoNTINENTal ShElvES: daNGErouSly aBuNdaNT SourCES of oIl
Many continental shelves contain vast supplies of oil. this is primarily because some of them have had shallow marine water for millions of years and are the repository of dead oil-rich phytoplankton that have accumulated during that time. examples include the Gulf of Mexico and the north Sea. to many people, particularly those who reject all the scientific evidence related to global warming, these oil supplies represent an irresistible source of fuel for the indefinite future. in the United States, “Drill, baby, drill!” was a common conservative slogan. as industrial civilizations have burned more and more oil, drilling operations have moved into deeper water, where there is a greater risk of accidents.
One such accident occurred on april 19, 2010. the Deepwater Horizon drilling rig, operated by a contractor for BP (British Petroleum), exploded. this allowed pressurized oil to gush into Gulf of Mexico waters unchecked for 87 days, until July 15. the flow of oil may have been 4.2 million gallons (16,000 cubic m) per day.
BP was apparently unprepared to respond to any such disaster. its response to the spill, coordinated by the U.S. coast Guard, was primarily to use dis-persants that allowed the components of the oil to no longer be visible in the hope that observers would ignore it. they also pumped some of the oil onto ships and burned some of it on the ocean surface. a great deal of natural gas as well as oil emerged from the leaking well. long before the leak was capped, it was widely considered to be the worst environmental disaster in the history of the United States. environ-mentalists countered the conservatives with their own slogan: “Spill, baby, spill!” tarballs on beaches and birds coated with oil were the most famous yet probably the least significant ecological conse-quences of the spill. Millions of people felt helpless to respond and even attempted some symbolic but useless tactics, such as sending some of their hair to the Gulf coast to absorb the oil.
as this encyclopedia goes to press, the U.S. Justice Department continues its investigation into whether criminal negligence was involved in the BP oil spill. it appears that the federal government (the Minerals Management Service of the Department of the interior) deliberately overlooked dangerous problems with the drilling rig before its explosion in return for favors from BP and associated companies.
Kelp forests support abundant marine life, including many species of fishes. (Earthgauge and Department of Commerce/NOAA)
xviii+598_Biodiversity_F.indd 142 11/17/11 1:12 PM

convergence 143
the liability that the federal government may have along with BP, including the deaths of 11 rig workers, has not been determined.
the long-term consequences will not be known for many years but include toxicity from the dis-persed, invisible oil components (and from the chem-icals used to disperse them) and the methane from the natural gas. Besides the direct environmental consequences, the Gulf Oil Spill of 2010 will con-tribute to the greenhouse effect directly (through the release of methane) and indirectly (as a result of con-tinued economic reliance on continental shelf oil).
See also competition, ecological; coral reefs; deep oceans, biodiversity in; federal government role in conservation (United States); greenhouse effect; keystone species.
Further readingandrew, n. l. “Spatial heterogeneity, sea urchin grazing,
and habitat structure on reefs in temperate australia.” Ecology 74 (1993): 292–302.
Begley, Sharon. “What the spill will kill.” Newsweek, June 14, 2010, 24–28.
carpenter, K. e., et al. “One-third of reef-building corals face elevated extinction risk from climate change and local impacts.” Science 321 (2008): 560–566.
carr, M. H. “effects of macroalgal dynamics on recruit-ment of a temperate reef fish.” Ecology 75 (1994): 1,320–1,333.
Dayton, P. K. “ecology of kelp communities.” Annual Review of Ecology and Systematics 16 (1985): 215–245.
Gell, F. R., and c. M. Roberts. “Benefits beyond boundar-ies: the fishery effects of marine reserves.” Trends in Ecology and Evolution 18 (2003): 448–455.
Gold, Russell, and Ben casselman. “alarm was disabled before BP blast.” Wall Street Journal, July 24, 2010. available online. URl: http://online.wsj.com/article/SB10001424052748703294904575385160342490350.html?mod=WSJ_hps_MiDDleSecondnews. accessed May 12, 2011.
Graham, M. H., et al. “Deep-water kelp refugia as poten-tial hotspots of tropical marine diversity and pro-ductivity.” Proceedings of the National Academy of Sciences USA 104 (2007): 16,576–16,580.
Gulf Oil Spill. “new BP oil spill task force is headed by criminal Division of Justice Department.” available online. URl: http://www.gulfspilloil.com/new-bp-oil-spill-task-force-is-headed-by-criminal-division-of- justice-department. accessed august 4, 2011.
Halpern, B. S., et al. “Strong top-down control in South-ern california kelp forest ecosystems.” Science 312 (2006): 1,230–1,232.
Harrold, c., and D. c. Reed. “Food availability, sea urchin grazing and kelp forest community structure.” Ecology 66 (1985): 1,160–1,169.
Hughes, t. P., et al. “climate change, human impacts, and the resilience of coral reefs.” Science 301 (2003): 929–933.
Jackson, J. B. c., et al. “Historical overfishing and the recent collapse of coastal ecosystems.” Science 293 (2002): 629–638.
lafferty, K. D. “Fishing for lobsters indirectly increases epidemics in sea urchins.” Ecological Applications 14 (2004): 1,566–1,573.
Paddack, M. J., and J. a. estes. “Kelp forest fish popula-tions in marine reserves and adjacent exploited areas of central california.” Ecological Applications 10 (2000): 855–870.
Pearse, J. S., and a. H. Hines. “expansion of a central cali-fornia kelp forest following the mass mortality of sea urchins.” Marine Biology 51 (1987): 83–91.
Sala, e., and M. H. Graham. “community-wide distri-bution of predator-prey interaction strength in kelp forests.” Proceedings of the National Academy of Sci-ences USA 99 (2002): 3,678–3,683.
Santelices, B. “the discovery of kelp forests in deep-water habitats of tropical regions.” Proceedings of the National Academy of Sciences USA 104 (2007): 19,163–19,164.
Sotka, e. e., and Mark e. Hay. “effects of herbivores, nutrient enrichment, and their interactions on mac-roalgal proliferation and coral growth.” Coral Reefs 28 (2009): 555–568.
Steneck, R. S., et al. “Kelp forest ecosystems: Biodiversity, stability, resilience and future.” Environmental Con-servation 29 (2002): 436–459.
convergence convergence occurs when similar adaptations evolve in separate lineages of organisms. convergent adaptations evolve separately (by con-vergent evolution), rather than being inherited from a common ancestor. convergent evolution is respon-sible for a great deal of the biodiversity on earth.
convergence can often be recognized because the organisms evolve the adaptation in different ways. this has been experimentally demonstrated in bacte-ria. Scientists grew some populations of the gut bac-terium Escherichia coli in the absence of the sugar maltose, and they lost the genes that would allow them to use maltose as a food source. then they were exposed to maltose again. the populations produced bacteria that had mutations that allowed them to metabolize the maltose. each of these populations evolved a different set of mutations. the reinvention of the ability to use maltose was a convergence in the separate bacterial populations, but each population achieved this result in a different way.
the following are just a few of the many thou-sands of possible examples from plants and from animals.
xviii+598_Biodiversity_F.indd 143 11/17/11 1:12 PM

first entry144 convergence 144
plaNT CoNvErGENCESPlant adaptations to climatic conditions. Plants have evolved convergent adaptations to stressful environ-mental conditions, including these examples.
● Bushes that live in dry climates have small leaves. Small leaves dissipate heat into the air more efficiently than do large leaves, which is important in hot, dry climates, where plants must restrict the amount of water they use in evaporative cooling. Small-leaved bushes have evolved within several differ-ent plant families, all of which have species with larger leaves. Small leaves are therefore a new adaptation in several different plant families. in some environments, these bushes are also adapted to sprouting or seed germi-nation after periodic fires. Habitats that are dominated by bushes with small leaves and that experience periodic fires have evolved separately in california, chile, the Mediter-ranean Basin, and South africa.
● the succulent growth form, which allows water storage and caM photosynthesis, has evolved in several plant families, for exam-ple the cactus and lily families in north american deserts, and the spurge family in South african deserts.
● the c4 form of photosynthesis may have evolved more than 45 different times.
● Several lineages of plants have evolved the ability to grow in toxic serpentine soil.
Plant adaptations to pollination methods. Plants have evolved convergent adaptations to the same pol-lination mechanisms, as in the following examples.
● Flowers that attract hummingbirds as pol-linators are usually tubular and red. the tubular red form of flower has evolved in many different plant families. in one instance, a single mutation changed yellow, bee-pollinated flowers into red, humming-bird-pollinated flowers and resulted in an immediate reproductive isolation of two populations, resulting in the evolution of two separate species.
● Different lineages of plants have evolved adaptations for wind pollination, in which the petals are reduced in size or lost, pollen production is increased, and the pollen-cap-turing surfaces of the flowers increase in size.
Plant defenses. Plants have evolved convergent adaptations as defenses against herbivores, as in these examples.
● Spines and thorns have evolved many times in separate lineages of plants. Some plants have seeds with sharp spines that entangle in animal fur, causing the animal to dis-perse the seeds to a new location. Strikingly similar structures for animal fur dispersal have evolved in Tribulus terrestris, a dicot, and genus Cenchrus, a grass (see figure on page 145). Grasses and dicots evolved from a common ancestor, without spiny seeds, that lived more than 100 million years ago.
● the use of latex (a sticky, milky sap) as a defense against herbivores and as a way of sealing damaged areas has also evolved sepa-rately in several plant families. as a result, rubber latex is produced by a spurge (Hevea brasiliensis), a fig (Ficus elastica), and the dita bark tree, Alstonia, in the dogbane family.
● chemical defense in plants has evolved thousands of times. Usually, each lineage of plant uses a different set of chemicals, which is why the plant kingdom produces such an astounding variety of pharmaceuti-cal and spice substances. Some plant chemi-cals, such as limonene, reappear over and over, producing the lemon scent separately in lemons (a citrus), lemon basil and lemon balm (mints), lemon grass, and the conifer Thuja standishii. Onion scent has evolved separately in onions (Allium cepa), which are monocots, and in the salmwood tree, a dicot (Cordia alliadora).
Other examples of convergent evolution in plants include the following.
● Parasitism has evolved separately in several lineages of flowering plants. these plants have lost their chlorophyll and obtain food from other plants. examples include plants in the heath family (genera Pterospora and Monotropa), relatives of snapdragons (genus Epifagus), and plants in the orchid family (genus Corallorhiza).
● Within the genus Calochortus, the same flower shapes appear to have evolved in sev-eral different lineages: the cat’s ear form, which is hairy and has open petals; the mariposa form, which is cuplike; the fairy lantern form, which hangs down, with the petals together; and the star tulip form, with smooth spreading petals.
● at least three lineages of plants have sepa-rately evolved mutualisms with bacteria in their roots that convert nitrogen gas into
xviii+598_Biodiversity_F.indd 144 11/17/11 1:12 PM

convergence 145
nitrogen fertilizer: the legume family Faba-ceae, alders (in the birch family Betulaceae), and Vitex keniensis (in the verbena family Verbenaceae). the bacteria involved in these mutualisms are different: legumes have coevolved with bacteria such as Rhizobium, while alders have coevolved with Frankia bacteria.
● Strikingly similar leaf forms, so similar as to be almost indistinguishable, have evolved in Jacaranda mimosifolia (family Bignonia-ceae) and the mimosa (Albizzia julibrissin of the Mimosoideae).
aNImal CoNvErGENCESEyes. eyes have evolved several times in the animal kingdom, as in these examples.
● among the earliest eyes were the crystalline eyes of trilobites, which evolved separately from the compound eyes found in other arthropods and the calcitic eyes of certain echinoderms.
● Most arthropods have compound eyes, but some spiders (the jumping spiders that rely on acute vision) have a lens eye similar to that of vertebrates. Some shrimp have both compound and lens-type eyes.
● eyes have evolved in three lineages of gas-tropod mollusks (snails and their relatives) separately from the eyes of cephalopods such as squid and octopus.
● Some marine worms have also evolved eyes. ● One group of coelenterates has evolved eyes. ● Vertebrates have also evolved eyes.
each of these groups has eyes that receive visual information, but in different ways. For example, the eyes of the squid are nearly as complex as vertebrate eyes. in vertebrates, the nerve cells are in front of the retina, while in the squid they are behind the retina. Humans, other apes, and Old World monkeys have three-color vision; this type of vision has evolved independently in howler monkeys of the new World. all the above examples are true eyes, with lenses and retinas, as opposed to simple eyespots that detect the presence of light.
Animal societies. Social evolution has converged in different lineages of animals, as in the following examples.
● the “army ant” lifestyle may have evolved separately in africa and South america, including the behavior of forming a living nest out of the ants’ own bodies that has a limited ability to regulate the internal tem-perature. in turn, seven different lineages of staphylinid beetles have evolved into army ant mimics, which can live among the ants without being recognized as intruders.
● the reproductive system in which one dominant female reproduces and the other females do not is well known in insects (ants, bees, and wasps). three different lin-eages of coral reef shrimp have evolved a similar system. it is also found in one group of mammals, the naked mole rat. the insect and mammal lineages achieved this sys-tem differently, however. in ants, bees, and wasps, the males have only half the chro-mosomes of the females, and the difference between queens and workers results from being fed different foods as they develop. in mole rats, the males have the same number
Both the puncture vine (left, Tribulus terrestris) and the sandbur (right, Cenchrus spinifex) produce seeds that entangle in the fur of mammals. The seeds look remarkably similar to one another, even though punc-ture vines (a dicot) and sandburs (a grass) have had separate evolutionary lineages for about 100 million years. They have independently evolved seeds with spines. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 145 11/17/11 1:12 PM
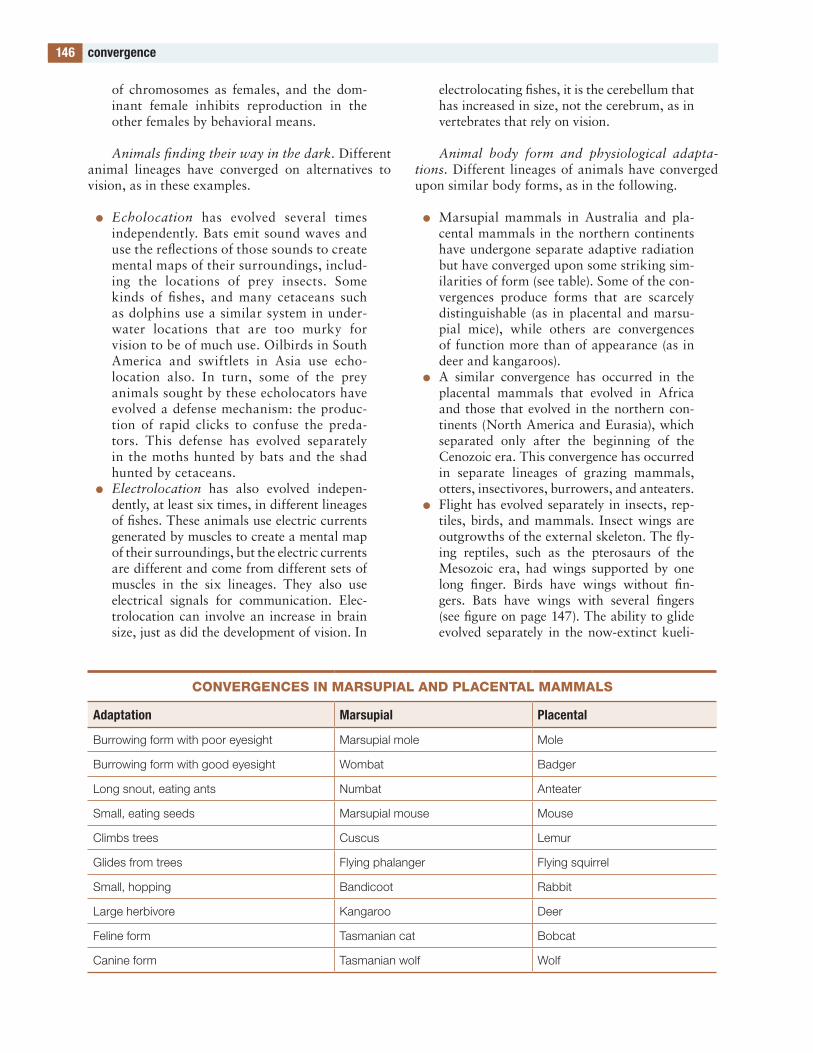
first entry146 convergence 146
of chromosomes as females, and the dom-inant female inhibits reproduction in the other females by behavioral means.
Animals finding their way in the dark. Different animal lineages have converged on alternatives to vision, as in these examples.
● Echolocation has evolved several times independently. Bats emit sound waves and use the reflections of those sounds to create mental maps of their surroundings, includ-ing the locations of prey insects. Some kinds of fishes, and many cetaceans such as dolphins use a similar system in under-water locations that are too murky for vision to be of much use. Oilbirds in South america and swiftlets in asia use echo-location also. in turn, some of the prey animals sought by these echolocators have evolved a defense mechanism: the produc-tion of rapid clicks to confuse the preda-tors. this defense has evolved separately in the moths hunted by bats and the shad hunted by cetaceans.
● Electrolocation has also evolved indepen-dently, at least six times, in different lineages of fishes. these animals use electric currents generated by muscles to create a mental map of their surroundings, but the electric currents are different and come from different sets of muscles in the six lineages. they also use electrical signals for communication. elec-trolocation can involve an increase in brain size, just as did the development of vision. in
electrolocating fishes, it is the cerebellum that has increased in size, not the cerebrum, as in vertebrates that rely on vision.
Animal body form and physiological adapta-tions. Different lineages of animals have converged upon similar body forms, as in the following.
● Marsupial mammals in australia and pla-cental mammals in the northern continents have undergone separate adaptive radiation but have converged upon some striking sim-ilarities of form (see table). Some of the con-vergences produce forms that are scarcely distinguishable (as in placental and marsu-pial mice), while others are convergences of function more than of appearance (as in deer and kangaroos).
● a similar convergence has occurred in the placental mammals that evolved in africa and those that evolved in the northern con-tinents (north america and eurasia), which separated only after the beginning of the cenozoic era. this convergence has occurred in separate lineages of grazing mammals, otters, insectivores, burrowers, and anteaters.
● Flight has evolved separately in insects, rep-tiles, birds, and mammals. insect wings are outgrowths of the external skeleton. the fly-ing reptiles, such as the pterosaurs of the Mesozoic era, had wings supported by one long finger. Birds have wings without fin-gers. Bats have wings with several fingers (see figure on page 147). the ability to glide evolved separately in the now-extinct kueli-
ConvergenCes in marsupial and plaCental mammals
adaptation marsupial placental
Burrowing form with poor eyesight Marsupial mole Mole
Burrowing form with good eyesight Wombat Badger
Long snout, eating ants Numbat Anteater
Small, eating seeds Marsupial mouse Mouse
Climbs trees Cuscus Lemur
Glides from trees Flying phalanger Flying squirrel
Small, hopping Bandicoot Rabbit
Large herbivore Kangaroo Deer
Feline form Tasmanian cat Bobcat
Canine form Tasmanian wolf Wolf
xviii+598_Biodiversity_F.indd 146 11/17/11 1:12 PM

convergence 147
neosaurs, in which extended ribs produced a gliding surface; fl ying squirrels, in which fl aps of skin between the legs produce a glid-ing surface; and even fl ying snakes, in which the entire fl attened body produces a gliding surface. there is even a species of gliding ant.
● the fi shlike body form has evolved several times: in fi shes, in aquatic reptiles of the Mesozoic era, and in dolphins and other cetaceans.
● Several different families of birds, on differ-ent continents and islands, have evolved the hummingbird body form and way of living: small bodies, long bills, and rapid fl utter-ing of wings. this includes the humming-birds of north and South america (family trochilidae), australian honeyeaters (fam-ily Meliphagidae), Hawaiian honeycreepers (family Drepanididae), and african sunbirds (family nectariniidae). this adaptation has also evolved in sphinx moths, which many people mistake for hummingbirds when they fi rst see them.
● Mammals in 11 families in three different orders, including some marsupials, have evolved the molelike lifestyle of digging through the soil, with big claws and reduced eyes. this adaptation has evolved eight times in rodents.
● earthworms (which are invertebrates) and caecilians (which are vertebrate amphibians) have separately evolved long, fl exible bodies without limbs and the use of a hydrostatic skeleton (muscles pressing against water compartments) to move through the soil.
● One type of barnacle produces a shell made of calcium phosphate and protein, like the
bones of vertebrates, although most bar-nacles produce shells of calcium carbonate.
● insect species with a grasping foreleg (the praying mantis body form) have evolved in three separate insect orders.
● Both bombardier beetles and certain kinds of mites spray caustic liquids against ani-mals that threaten them. However, it is not the same chemical in the two groups.
● Both dipterans and strepsipteran fl ies as well as african beetles have separately evolved halteres. Halteres are modifi ed wings that function as balancing devices. in dipterans (such as the house fl y), it is the hind wings that become halteres; in strepsipterans, it is the forewings; in some african beetles, it is the wing cover.
● cephalopod molluscs such as the octopus have evolved a closed circulatory system, in which the blood cells remain inside the ves-sels, independently of vertebrates.
● live birth has evolved separately in mam-mals, some reptiles, some bony fi shes, and some sharks.
● Warm-bloodedness has evolved separately in birds, mammals, and tuna.
● arctic and antarctic fi shes have separately evolved a very similar set of antifreeze proteins.
● Vultures, which are birds that have featherless heads and eat carrion, have evolved separately in the new World (the family cathartidae is related to storks) and the Old World (the fam-ily accipitridae is a type of hawk).
Physiological convergence has occurred in dif-ferent lineages of humans. the ability of adults to
Both bats and birds have wings, but wings evolved separately in bird and mammal lineages. Their separate evolutionary origin is revealed by their internal structure: Bat wings have fi nger bones, and bird wings do not.
xviii+598_Biodiversity_F.indd 147 11/17/11 1:12 PM

first entry148 convergence 148
digest lactose has evolved separately in europeans and in some african tribes that live by herding cattle.
Coevolution between animal species and other species. Similar types of coevolution have occurred in different sets of organisms, as in the following examples.
● at least five different species of ants have evolved the ability to grow underground gardens of fungus. these ant gardens may represent an evolution of agriculture sepa-rate from humans.
● in 70 different lineages, insects and spiders have evolved characteristics that allow them to parasitize ant nests without being recog-nized as intruders.
● Bioluminescence, in which living tissues produce light, has evolved separately in insects and in vertebrates. in vertebrates, the light is usually produced by mutualistic bacteria that live in the vertebrate tissues. Bioluminescence may have evolved 30 sepa-rate times.
● in some vertebrates that eat plant material, bacteria in the digestive system digest the cellulose. the vertebrate then uses a mutant form of lysozyme to digest the bacteria. this adaptation has evolved separately in cows, langur monkeys, and hoatzin birds.
in many cases, convergent evolution does not require a total reinvention of an adaptation, as in these examples.
● limonene has evolved separately in sev-eral lineages of plants (see above), but most flowering plants have terpene synthases, the enzymes that produce the immediate chemi-cal precursors of limonene.
● the ornithologist n. i. Mundy explains that the evolution of similar color patterns in separate bird lineages has resulted from the modification of an underlying genetic pat-tern that they inherited from their common ancestor.
● eyes have evolved perhaps 40 separate times in the evolutionary history of animals (see above). all these animals share an organizer gene, called pax, that stimulates the devel-opment of eyes. in each of these lineages, the actual structure of the eye has evolved separately. there is a marine worm that has compound eyes, like those of other inver-tebrates, but also has brain lobes that have photosensitive chemicals similar to those of vertebrate eyes. apparently, vertebrate eyes
evolved from invertebrate brain lobes rather than from invertebrate eyes.
in modern organisms, scientists can recognize convergence because, despite the external similari-ties, the organisms are internally different and have achieved the convergence in different ways. in look-ing at fossils, scientists cannot always tell whether organisms shared a characteristic because they inherited it from a common ancestor or because they evolved it separately by convergence. this sometimes makes reconstructing fossil history difficult. Some-times in the fossil record, a species appears to have become extinct, then reappears. is it because the species was rare for a long time, then reappeared in greater abundance (what the paleontologist David Jablonski called Lazarus taxa, because it appeared to rise from the dead), or because another lineage evolved similar features by convergence and it just looked like a reappearance (what the paleontologist Douglas erwin called Elvis taxa)?
every evolutionary scientist understands that there are many examples of convergent evolution. they differ in how much importance they give con-vergence in the overall pattern of the evolution of life. Some evolutionary scientists claim that evolution has so many different possible directions that the actual course taken by evolution is a matter of historical contingency, a viewpoint expressed by the evolution-ary biologist Stephen Jay Gould. Gould said that evolution produced a bush of different forms, not a ladder leading upward to human perfection. these scientists consider convergence to be details that have occurred at the tips of the branches of the bush of life. Others, such as Simon conway Morris, insist that convergence is so frequent that it constitutes one of the major features of evolution. they point out that even though there may be an almost infinite number of possible biological adaptations, there are only a limited number of strategies that work, and evolution keeps finding these strategies over and over. conway Morris agrees with Gould that evolu-tion is a bush, not a ladder, but in this bush, he says, many of the branches are parallel. conway Morris cites the parallel evolution of large brains in ceta-ceans (such as dolphins) and humans. in the human lineage, there was a separate increase in brain size in the lineage that led to modern humans, the lin-eage that led to neanderthals, and perhaps the lin-eages that led to asian Homo erectus and the robust australopithecines. agriculture and civilization in humans evolved separately in east asia, the Middle east, and america. intelligence does not always offer an evolutionary advantage, but when it does, one would expect that the large processing center for the nerves would be near the front end of the animal,
xviii+598_Biodiversity_F.indd 148 11/17/11 1:12 PM

coral reefs 149
where most of the sensory organs are located (a big brain in a head with eyes and nose). conway Morris says that the evolution of intelligence is a rare event, but that if humans encounter an intelligent life form somewhere else in the universe, it would probably be something that they would recognize as a person, perhaps even as a human. this is why he refers to “inevitable humans in a lonely universe.”
in convergent evolution, the same or similar solutions to environmental and social challenges evolve over and over again, contributing to the pro-liferation of species throughout evolutionary history.
See also adaptive radiation; coevolution; mutualism; parasitism; speciation.
Further readingconway Morris, Simon. Life’s Solution: Inevitable Humans
in a Lonely Universe. cambridge: cambridge Univer-sity Press, 2003.
Mundy, n. i., et al. “conserved genetic basis of a quantita-tive plumage trait involved in mate choice.” Science 303 (2004): 1,870–1,873. Summarized by Hoekstra, Hopi e. and trevor Price. “Parallel evolution is in the genes.” Science 303 (2004): 1,779–1,781.
Patterson, thomas B., and thomas J. Givnish. “Geographic cohesion, chromosomal evolution, parallel adaptive radiations, and consequent floral adaptations in Calo-chortus (calochortaceae): evidence from a cpDna phylogeny.” New Phytologist 161 (2003): 253–264.
Pichersky, eran. “Plant scents.” American Scientist 92 (2004): 514–521.
Shadwick, Robert e. “How tunas and lamnid sharks swim: an evolutionary convergence.” American Scientist 93 (2005): 524–531.
Sheppard, Samuel K., et al. “convergence of Campylo-bacter species: implications for bacterial evolution.” Science 320 (2008): 237–239.
Weinrech, Daniel M., et al. “Darwinian evolution can fol-low only very few mutational paths to fitter proteins.” Science 312 (2006): 111–114.
coral reefs coral reefs are reefs, or ridges, con-structed from the accumulated limestone bodies of corals. new corals attach to the limestone shells of dead corals, further building the reefs. Some coral-line algae also have limestone in their cell walls and contribute to reef formation. Some animals bore into the reefs, and parrotfish chew and spit out coral. the resulting coral sediments settle into reef fissures and strengthen the reefs. in ancient oceans, corals were not the only organisms that built reefs. Bryozoans and brachiopods also formed reefs, as did kinds of coral that are no longer common.
Fringing reefs are attached to shoreline rocks and form shallow lagoons. Barrier reefs are sepa-
rated from shoreline rocks and form deep lagoons. if a reef forms a complete circle, it is a coral atoll. these reefs, full of crevices, provide a habitat for numerous other species. Because of their immense biodiversity, they have been called the “rain forests of the sea,” even though their diversity is less than that of rain forests.
coral reefs are most abundant in shallow tropi-cal waters. the main reason is that corals grow best in water with bright sunlight. although cor-als (which are animals in the phylum cnidaria) can grab food with their stinging tentacles, most of their food comes from mutualistic single-celled algae (zooxanthellae) that live in their tissues and produce food by photosynthesis. Zooxanthellae create the many bright colors of corals, and the shapes of corals appear to be adaptations to expos-ing their zooxanthellae to light (see figure on page 150). corals do not grow well if ocean water is diluted by freshwater, as occurs near the mouths of the amazon and Ganges Rivers.
corals thrive in intermediate conditions of tem-perature and nutrients. if temperatures are too high, the corals lose their zooxanthellae, which causes them to become white. this is known as coral bleaching. the coral animals slowly starve. if nutrient levels are too high, algae overgrow the coral reefs. Most of the nutrient cycling occurs very close to the reef itself; nutrient levels in the water are low. the zooxanthellae use the carbon dioxide and the nitrogenous wastes of the corals inside the tissues of the corals themselves.
Some corals, such as Oculina varicosa and Lophelia pertusa, live in deep water (more than 6,000 feet [2,000 m]) or on continental shelves as far north as norway. However, they do not have zooxanthellae, and they grow more slowly. they therefore do not form large reefs.
Because of the abundant coral growth, a com-plex ecological community, including as many as 4,000 fish species (about one-quarter of the total fish species in the sea), depends on the corals. among the top resident predators in reefs are moray eels, in the fish family Muraenidae. even larger predators such as sharks pass through, feeding on the abundant fishes. Because the zooxanthellae create many col-ors, colorful fish are well camouflaged in coral reefs.
the coral reef community exists in a delicate balance of disturbance. echinoderm larvae cannot settle and develop on intact reefs because the corals eat them. However, in disturbed reefs, echinoderm larvae can develop, and once they are large enough, they eat the corals. During World War ii, many coral reefs were damaged by bombing. this tipped the bal-ance in favor of echinoderms, especially Acanthaster planci, the crown-of-thorns starfish. large numbers of starfish began to devastate the coral reefs.
xviii+598_Biodiversity_F.indd 149 11/17/11 1:12 PM

first entry150 coral reefs 150
coral reefs are facing numerous environmental threats. they include:
● eutrophication occurs when wind erosion brings in dust from land surfaces or when coastal communities discharge sewage.
● Some fishermen use explosives or cyanide to kill large numbers of fish.
● Many tropical fishes for aquaria come from coral reefs, particularly those that are unable to reproduce in captivity.
● Global warming is causing the water to become warm enough to bleach the corals.
● carbon dioxide in the atmosphere is acidify-ing the water.
coral bleaching and water acidification may be serious enough problems that coral reefs may be unable to survive by the end of the 21st century, except as small patches. the ocean biologist nancy Knowlton predicts that coral reefs will no longer exist by the year 2100, and perhaps by 2050.
there are attempts to restore coral reefs that have been damaged. these include the construction of metallic frames on which coral larvae can settle and begin to form artificial reefs. the process may be encouraged by electric currents, which precipi-tate limestone onto the frames. While some of these efforts have been successful, they cannot prevail against the higher water temperatures and acidifica-tion associated with global climate change.
See also animals; continental shelves, bio-diversity in.
Further readingBellwood, D. R., et al. “Functional versatility supports
coral reef biodiversity.” Proceedings of the Royal Soci-ety of London B (Biology) 273 (2006): 101–107.
Burt, J., et al. “are artificial reefs surrogates of natural habitats for corals and fish in Dubai, United arab emirates?” Coral Reefs 28 (2009): 663–675.
carpenter, Kent e., et al. “One-third of reef-building corals face elevated extinction risk from climate change and local impacts.” Science 321 (2008): 560–566.
This coral reef, near Port Ghalib, Egypt, has abundant corals and colorful fishes. (J. Hutsch)
xviii+598_Biodiversity_F.indd 150 11/17/11 1:12 PM

Cretaceous extinction 151
cheng, y.-R., and c.-F. Dai. “endosymbiotic copepods may feed on zooxanthellae from their coral host, Pocillo-pora darnicornis.” Coral Reefs 29 (2010): 13–18.
coral Reef alliance. “coral Reef alliance.” available online. URl: http://www.coral.org. accessed July 8, 2010.
Hobbs, Jean-Paul a., et al. “Rarity and extinction risk in coral reef angelfishes on isolated islands: interrelation-ships among abundance, geographic range size, and specialization. Coral Reefs 29 (2010): 1–11.
Hughes, t. P., et al. “climate change, human impacts, and the resilience of coral reefs.” Science 301 (2003): 929–933.
Knowlton, nancy. “coral reef biodiversity—habitat size matters.” Science 292 (2001): 1,493–1,495.
Maragos, J. e., et al. “coral reefs and biodiversity: a crit-ical and threatened relationship.” Oceanography 9 (1996): 83–99.
Mumby, Peter J. “Herbivory versus corallivory: are par-rotfish good or bad for caribbean coral reefs?” Coral Reefs 29 (2009): 683–690.
national Oceanic and atmospheric administration. “nOaa coral Reef conservation Program.” available online. URl: http://coralreef.noaa.gov. accessed July 8, 2010.
Orr, J. c., et al. “anthropogenic ocean acidification over the 21st century and its impact on calcifying organ-isms.” Nature 437 (2005): 681–686.
Prada, c., et al. “Octocoral bleaching during unusual ther-mal stress.” Coral Reefs 29 (2010): 41–45.
Selkoe, K. a., et al. “a map of human impacts to a ‘pris-tine’ coral reef ecosystem, the Papahānaumokuākea Marine national Monument.” Coral Reefs 28 (2009): 635–650.
Sotka, e. e., and Mark e. Hay. “effects of herbivores, nutrient enrichment, and their interactions on mac-roalgal proliferation and coral growth.” Coral Reefs 28 (2009): 555–568.
Cowles, henry Chandler See succession, ecological.
Cretaceous extinction Some 65 million years ago, at the end of the cretaceous period, one of the five mass extinctions of earth history occurred. it was not the greatest of the mass extinctions (the Permian extinction resulted in the disappearance of more species) but has long been the subject of intense speculation and research because it was the most recent of the mass extinctions, and it involved the disappearance of the dinosaurs, the prehistoric ani-mals that have attracted perhaps the most attention from scientists and the general public.
Part of the problem in explaining the cretaceous extinction, as with the other mass extinctions, was that not all groups of organisms suffered the same rate of disappearance. the largest land animals were dinosaurs, all of which suffered extinction, and the large marine reptiles also became extinct. How-ever, not all dinosaurs were large; even the small ones became extinct. Most mammals, amphibians, crocodiles, turtles, insects, and land plants sur-vived the cretaceous extinction. among the marine mollusks, the ammonoids became extinct, but the nautiloids, which appeared to have had very similar ecological roles, did not. Birds survived, but only a few lineages of them. Some plankton, the floating microscopic organisms of the oceans, experienced a dramatic set of extinctions (for example, the fora-miniferans), while others, such as diatoms, did not. extinction rates were much higher on land (about 90 percent of species) than in freshwater (about 10 percent of species).
there was also the question of whether the cre-taceous extinction was a single catastrophic event or the outcome of gradual declines toward extinc-tion. Several lineages of dinosaurs were already in decline during the cretaceous period; others were not. although land plants survived the extinc-tion, their populations were apparently set back by some catastrophe. at the end of the cretaceous, ferns and fern spores were very numerous. Ferns often lived in areas from which the larger vegeta-tion had been removed by some disturbance. the cretaceous extinction appears to have been a cata-strophic event.
Some investigators noticed that a catastrophic col-lapse of the food chain could have resulted in the pat-tern of extinctions. the large reptiles (which may have had relatively rapid metabolisms) would need con-tinual food, often in vast quantities. in contrast, many amphibians, reptiles with slow metabolisms such as crocodiles and turtles, and insects can hibernate until food resources again became available. the creta-ceous mammals were small and, despite their high metabolisms, might also have been able to hibernate, as many do today. Omnivores and carrion feeders usu-ally survived the cretaceous extinction. Plants could have survived as rootstocks or seeds. Some researchers speculated that plankton may have been less capable of surviving an interruption of conditions suitable for photosynthesis, although most plankton have resting phases that can resist very cold and dry conditions.
there was, however, no agreement on what might have interrupted the food supply. Volcanic eruptions can have a massive impact on organisms around the world. the indonesian volcano tam-bora eruption in 1815 was followed by what has
xviii+598_Biodiversity_F.indd 151 11/17/11 1:12 PM

first entry152 Cretaceous extinction 152
been called the “year without a summer” because the volcanic dust, propelled into the stratosphere where it could not be cleared by rain, blocked sun-light. it was the worst agricultural year on record; the resulting famine killed 65,000 people in ireland alone. all this happened, even though the global temperature decreased by only about 2°F (1½°c). a “supervolcano” eruption occurred 74,000 years ago (toba, in Sumatra). evidence from Greenland ice cores, which preserve a record of global climate, suggests that this volcanic eruption was followed by six cold years. clearly, massive volcanic eruptions could have interrupted the food chain at the end of the cretaceous. there were massive volcanic erup-tions (producing the Deccan traps) at the end of the cretaceous period. However, these eruptions do not appear to have been large enough to have, by them-selves, caused the cretaceous extinction.
in a 1980 paper, the physicist luis alvarez and his son, the geologist Walter alvarez, and their associates made a dramatic and risky proposal: that the cretaceous extinction was caused by an extra-terrestrial impact. Such an impact would, in fact, explain global extinctions. the impact itself would project dust high into the atmosphere and ignite fires that would add great clouds of smoke, both of which would darken the earth, plunging it into win-try conditions disastrous for the food chain. Mas-sive acid rain events might also have occurred as a result of the fires, especially if the rock underlying the impact contained sulfur. the shock to the crust could have set off volcanic eruptions and tsunamis (tidal waves), which would have contributed further to local and worldwide extinctions. Some research-ers estimate that such an impact would have influ-enced climate around the world for up to 10,000 years afterward.
Because the prevalence of uniformitarianism in geological science made paleontologists extremely hesitant to invoke processes that are not normally occurring on the earth as explanations for events in the history of the earth, the alvarez suggestion was resisted at first. nevertheless, their initial evidence was good, and more evidence later reinforced his proposal.
the alvarez team had not begun their research with the intention of explaining the cretaceous extinction. they were more interested in determin-ing accurate dates for the strata that spanned the cretaceous-tertiary interval. they expected that the rare element iridium might be useful in studying these strata. they found an astonishing “iridium anomaly” in rocks over a wide area right at the cre-taceous-tertiary boundary: a transient and very high concentration of this element. iridium is much more common in interplanetary particles and objects than
on earth. the iridium spike in the sedimentary lay-ers at the end of the cretaceous period strongly suggested an asteroid impact, but scientists sought corroboration. they found it.
● not only was iridium common in the last cretaceous layers, but also microtektites, which are small glass spheres characteristic of the impact of an asteroid with terrestrial rocks that contain silicon. Shocked quartz grains, characteristic of powerful impacts, were also found.
● Sootlike carbon particles were also unusu-ally common in the last cretaceous deposits.
● Sediments in the caribbean from the end of the cretaceous suggest that a very large tsunami had struck coastal areas.
● Perhaps most important, scientists wanted to find the crater caused by the asteroid. if the impact had occurred in an area with abundant rainfall and erosion or in the ocean, it might have eroded away but still left some evidence. Just such evidence was found on the yucatán Peninsula. the chicx-ulub crater has mostly filled in, but evi-dence of sedimentary layers shows that there was once a massive impact crater.
the cretaceous extinction changed the face of biodiversity on earth. after the large dinosaurs were gone, many large mammals evolved as well as many small ones. even though fishes continue to be the most diverse group of vertebrates and there are still more reptile species than mammals, the cenozoic era has become known as the “age of mammals.” Before the cretaceous extinction, conifers dominated many forests, with flowering plants playing a minor role; afterward, their roles were reversed. Since the cre-taceous extinction, the only extensive coniferous for-ests have been in the cold regions of earth.
See also cretaceous period, biodiversity in; Permian extinction.
Further readingalvarez, luis W., Walter alvarez, Frank asaro, and Helen
V. Michel. “extraterrestrial cause for the cretaceous-tertiary extinction.” Science 208 (1980): 1,095–1,107.
alvarez, Walter. T. Rex and the Crater of Doom. Princeton, n.J.: Princeton University Press, 1997.
Duncan, R. a., and D. G. Pyle. “Rapid eruption of the Dec-can flood basalts at the cretaceous/tertiary bound-ary.” Nature 333 (1988): 841–843.
Fawcett, a., et al. “Plants with double genomes might have had a better chance to survive the cretaceous-tertiary extinction event.” Proceedings of the National Acad-emy of Sciences USA 106 (2009): 5,737–5,742.
xviii+598_Biodiversity_F.indd 152 11/17/11 1:12 PM

Cretaceous period, biodiversity in 153
Frankel, charles. The End of the Dinosaurs: Chicxulub Crater and Mass Extinctions. cambridge: cambridge University Press, 1999.
labandeira, conrad c., et al. “impact of the terminal cretaceous event on plant-insect associations.” Pro-ceedings of the National Academy of Sciences USA 99 (2002): 2,061–2,066.
liangquan, l., and G. Keller. “abrupt deep-sea warming at the end of the cretaceous.” Geology 26 (1998): 995–998.
Marshall, c. R., and Peter D. Ward. “Sudden and grad-ual molluscan extinctions in the latest cretaceous of western european tethys.” Science 274 (1996): 1,360–1,363.
Morgan, J., et al. “analyses of shocked quartz at the global K-P boundary indicate an origin from a single, high-angle, oblique impact at chicxulub.” Earth and Plan-etary Science Letters 251 (2006): 264–279.
Poinar, George, Jr., and Roberta Poinar. What Bugged the Dinosaurs? Insects, Disease, and Death in the Cre-taceous. Princeton, n.J.: Princeton University Press, 2008.
Robertson, D. S., et al. “Survival in the first hours of the cenozoic.” Bulletin of the Geological Society of Amer-ica 116 (2004): 760–768.
Sheehan, Peter M., and thor a. Hansen. “Detritus feeding as a buffer to extinction at the end of the cretaceous.” Geology 14 (1986): 868–870.
Springer, M. S., et al. “Placental mammal diversification and the cretaceous-tertiary boundary.” Proceedings of the National Academy of Sciences USA 100 (2003): 1,056–1,061.
Vajda, Vivi, et al. “indication of global deforestation at the cretaceous-tertiary boundary by new Zealand fern spike.” Science 294 (2001): 1,700–1,702.
———, and S. Mcloughlin. “Fungal proliferation at the cretaceous-tertiary boundary.” Science 303 (2004): 1,489–1,490.
Wilf, Peter, and K. R. Johnson. “land plant extinction at the end of the cretaceous: a quantitative analysis of the north Dakota megafloral record.” Paleobiology 30 (2004): 347–368.
———, et al. “Decoupled plant and insect diversity after the end-cretaceous extinction.” Science 313 (2006): 1,112–1,115.
Cretaceous period, biodiversity in the cretaceous period (140 to 65 million years ago) was the third and final period of the Mesozoic era, following the Jurassic period. the Mesozoic era is also known as the age of dinosaurs, because dino-saurs were the largest land animals during that time. Dinosaurs became extinct during the cretaceous extinction, which is one of the two most impor-tant mass extinctions in the history of earth. the
multiCellular biodiversity in the CretaCeous period
Conditions
Atmospheric oxygen 50% above modern level
Atmospheric carbon dioxide
4.9 times modern level
Global temperature 7°F (4°C) above modern level
Sea level 560 feet (170 m) higher than today
Approximate Number of Families*
Continental 500
Marine 600
Total 1100
Examples of Marine Animals
Aristonectes parvidens plesiosaur reptile
Haccourtaster sp. starfish
Hoploscaphites pumilis ammonite mollusk
Keuppia levante octopus (mollusk)
Maiaspondylus lindoei ichthyosaur reptile
Micropycnodon kansasensis
bony fish
Mosasaurus missouriensis mosasaur reptile
Pentaditrupa subtorquata annelid
Prehepatus werneri crab
Squalicorax falcatus shark
Examples of Continental Animals
Alphadon marshi marsupial-like mammal
Anatotitan copei hadrosaur dinosaur
Ankylosaurus magniventris herbivorous dinosaur
Aurorachelys gaffneyi turtle
Beelzebufo ampinga frog
Confuciusornis sanctus toothed bird
Conodella suciensis snail
Costasaurus sp. lizard
Crephanogaster rara wasp
Deinosuchus sp. crocodile
(continues)
xviii+598_Biodiversity_F.indd 153 11/17/11 1:12 PM

first entry154 Cretaceous period, biodiversity in 154
cretaceous period was named for the deposits of chalk (latin creta) that were abundant in european strata. in north america, the cretaceous is divided into marine Mississippian deposits and coal-rich Pennsylvanian deposits.
Climate. the mild climate of the Jurassic period continued into the cretaceous period. Swamps and shallow seas covered widespread areas. as con-tinents continued to drift apart, the climate over much of the land became cooler and drier during the cretaceous. Occasional glaciations in the antarctic
regions caused sea levels to fall, then rise again when the glaciers melted (see table on pages 153–154).
Continents. the massive continent of Pangaea continued to break apart, mainly into what would become the northern continents and the southern continents of today. africa and South america split apart, allowing the southern part of the atlantic Ocean to form. the alps formed in europe. india broke free from Gondwanaland and became a sepa-rate continent. the continents began to assume their modern shapes and locations. the geographical iso-lation of plants and animals on the newly separated continents allowed the evolution of new species in the separate areas.
Marine life. Overall marine species diversity increased greatly over that of the preceding Jurassic period (see figure on page 155).
● Invertebrates. the invertebrate fauna of the oceans began to assume a modern appear-ance. among the mollusks, species of octo-pus (without external shells) became more common than ammonites and other shelled cephalopods, and starfish became more com-mon than stalked crinoids among the echi-noderms. the crustaceans (such as crabs) that are most common today were also the dominant crustaceans of the cretaceous.
● Vertebrates. Bony fishes and sharks contin-ued to diversify, while jawless fishes contin-ued to diminish. Very large aquatic reptiles, such as the mosasaurs and the elasmosaurs (plesiosaurs with very long necks), lived in the oceans. ichthyosaurs, which were rep-tiles that looked like porpoises, became less common, perhaps because of competi-tion from modern forms of fishes that were diversifying during the cretaceous.
Continental life. terrestrial forests were trans-formed by the appearance of the angiosperms, or flowering plants, at or just before the beginning of the cretaceous. the replacement of gymnosperm forests by angiosperm trees strongly affected the dominant animals.
● Plants. Gymnosperms had dominated all previous Mesozoic forests. Flowering trees evolved in tropical regions. Once they had evolved, they had an explosive increase in diversity as a result of coevolution with insects. During the cretaceous period, these trees displaced most of the gymno-sperms toward the polar regions. the for-ests of tropical and temperate regions began to look like they do today. By the end of
multiCellular biodiversity in the CretaCeous period (continued)
Eomaia scansoria placental-like mammal
Gallornis straeleni modern bird relative
Hesperornis regalis flightless bird
Ichthyornis dispar bird
Lapparentophis defrenni snake
Pachycephalosaurus sp. dinosaur
Purgatorius unio primatelike mammal
Quetzalcoatlus northropi pterosaur
Shuvuuia deserti feathered dinosaur
Sphecomyrma freyi ant
Triceratops horridus herbivorous dinosaur
Tyrannosaurus rex predatory dinosaur
Velociraptor mongoliensis feathered dinosaur
Examples of Land plants
Alnus sp. alder
Diospyros primaeva ebony
Ginkgo apodes ginkgo
Ettingshausenia cuneifolia sycamore
Ephedrites sinensis ephedra
Ficus ceratops fig
Juglans sp. walnut
Laurus plutonia laurel
Magnolia sp. magnolia
Picea sp. spruce
Pinus belgica pine
*Based on Benton
xviii+598_Biodiversity_F.indd 154 11/17/11 1:12 PM

Cretaceous period, biodiversity in 155
the cretaceous period, tem-perate forests were probably dominated by angiosperms such as maples, beeches, birches, magnolias, and wal-nuts, although it is diffi cult to assign cretaceous fossils to modern genera. the gym-nosperm forests consisted mostly of conifers such as pines and spruces in moun-tainous and northern regions, where they are found today.
● Invertebrates. all modern groups of insects appear to have evolved by the Jurassic period, although the evidence of Mesozoic butterfl ies and moths is unclear. What is clear is that insects exploded in diversity during the cre-taceous, especially the fl ies, ants and wasps, and probably the butterfl ies and moths, as a result of coevolution with fl owering plants. these many new species of insects were mostly pollinators or con-sumers of fl owering plants.
● Vertebrates. Freshwater and land vertebrates continued to diversify. Many lineages produced large animals. For example, the frog Beelzebufo (whose name means “devil frog”) was 16 inches (40 cm) long. Snakes are fi rst known from the cretaceous period.
Dinosaurs such as hadro-saurs, which may have special-ized in eating gymnosperms, were abundant worldwide at the beginning of the cretaceous. as angiosperm forests spread, new kinds of dinosaurs, such as cera-topsians, evolved that specialized on them. the gymnosperms and the dinosaurs that ate them were displaced into the polar regions, which were cold but not nearly as cold as today. Primitive birds may have driven fl ying reptiles, such as pterosaurs, toward extinction during the cretaceous, although some of the largest pterosaurs,
Some animals of the Cretaceous period: (A) Ankylosaurus was a heavily armored dinosaur. (B) Oviraptor was a dinosaur that laid eggs in clutches and protected them. The fi rst scientists to discover Oviraptor fossils believed that it had stolen the eggs from another dinosaur. (Oviraptor means “egg thief.”) Oviraptor, and many other dinosaurs, protected their eggs just as birds do. (C) Triceratops dinosaurs. (D) In the oceans, ichthyosaur reptiles resembled modern dolphins.
xviii+598_Biodiversity_F.indd 155 11/17/11 1:12 PM

first entry156 crustaceans 156
such as Quetzalcoatlus, lived at this time. Many of the small predatory dinosaurs were feathered, as in the Jurassic period, but these fossils provided chemi-cal evidence of feathers. a Shuvuuia had traces of the kind of keratin found in feathers, and Velocirap-tor had quill knobs on its bones.
there were about 100 genera of cretaceous dino-saurs that looked like modern birds. they had greater specialization to specific ways of living than did those of the Jurassic. For example, Hesperornis was a flight-less, nearly wingless diving bird. Confuciusornis, like modern birds, had no teeth, but it was not ancestral to modern birds; the ancestors of modern birds, similar to Hesperornis and Ichthyornis, still had teeth. Ich-thyornis could probably be mistaken for a gull, except for the teeth. Gallornis was similar to the common ancestor of all surviving birds.
about 90 genera of mammals lived during the cretaceous. they were small and represented by few species compared to the dinosaurs. One spe-cies, Repenomamus robustus, was large enough to eat small dinosaurs; one of them died with a baby dinosaur in its stomach. Many of the mammals were in lineages, such as the Gondwanatheria and the Multituberculata, that are extinct today, but the modern mammal groups apparently evolved during the cretaceous. Alphadon may have been a primitive marsupial, and Eomaia may have been one of the earliest placental mammals. Purgatorius may have been related to the earliest primates.
Extinctions. no major extinction event sep-arated the Jurassic from the cretaceous periods. Many dinosaur lineages went into decline during the cretaceous, while new groups of dinosaurs evolved. Some, but not all, dinosaurs would probably have become extinct even if the cretaceous extinction had not occurred. this extinction event was caused largely by an asteroid that hit earth 65 million years ago. the large marine and flying reptiles also became extinct at the end of the cretaceous. the extinction of the dinosaurs allowed mammals to diversify into the forms that dominated the cenozoic era.
See also cretaceous extinction; flowering plants (angiosperms).
Further readingBenton, Michael J. “Diversification and extinction in the
history of life.” Science 268 (1995): 52–58.coiffard, c., et al. “early cretaceous angiosperm invasion
of western europe and major environmental changes.” Annals of Botany 100 (2007): 545–553.
Falcon-lang, H. J., and D. J. cantrill. “leaf phenology of some mid-cretaceous polar forests, alexander island, antarctica.” Geological Magazine 138 (2001): 39–52.
Field, t. S., and n. c. arens. “Form, function, and envi-ronments of the early angiosperms: Merging extant
phylogeny and ecophysiology with fossils.” New Phy-tologist 166 (2005): 383–408.
Kaer, Benjamin P., et al. “Unusual gut contents in a creta-ceous ichthyosaur.” Proceedings of the Royal Society of London B (Biology) 270 (2003): S206–208.
lehman, thomas M., and elisabeth a. Wheeler. “a fossil dicotyledonous woodland forest from the Upper cre-taceous of Big Bend national Park, texas.” Palaios 16 (2001): 102–108.
Morell, Virginia. “When sea monsters ruled the deep.” National Geographic, December 2005, 58–87.
Schweitzer, Mary Higby, et al. “Beta-keratin specific immu-nological reactivity in feather-like structures of the cretaceous alvarezsaurid, Shuvuuia deserti.” Jour-nal of Experimental Zoology B (Mol Dev Evol) 285 (1999): 146–157.
Skelton, Peter, et al. The Cretaceous World. new york: cambridge University Press, 2003.
turner, alan H., et al. “Feather quill knobs in the dinosaur Velociraptor.” Science 317 (2007): 1,721.
———, et al. “a basal dromaeosaurid and size evolu-tion preceding avian flight.” Science 317 (2007): 1,378–1,381.
Wilson, edward O., and Bert Hölldobler. “the rise of the ants: a phylogenetic and ecological explanation.” Pro-ceedings of the National Academy of Sciences USA 102 (2005): 7,411–7,414.
Xu, X., et al. “Basal tyrannosauroids from china and evi-dence for protofeathers in tyrannosauroids.” Nature 431 (2004): 680–684.
crustaceans crustaceans are a major group of mostly aquatic arthropods. crayfish, lobsters, crabs, shrimp, barnacles, and many components of micro-scopic plankton are crustaceans. Most crustaceans have exoskeletons that cover the entire body, although the more than 500 species of hermit crabs have unpro-tected abdomens and must find shells of dead mol-lusks in which to live. typical crustaceans have two pairs of feeding appendages, unlike other arthropods.
crustaceans exhibit an astonishing diversity of ways of making a living. Most are fully aquatic, but some live on land. Some isopods, such as pill bugs, live in the moist soil beneath rocks. Some crabs live on land, though usually on islands not far from the sea. christmas island, in the Pacific Ocean, has 14 species of land crab. One of these species, the red crab Gecarcoidea natalis, has a population of more than a hundred million on this island (see figure on page 157). land crabs breed in the ocean and breathe with gills. the exoskeleton covering the gills is inflated and contains blood vessels that can extract oxygen from air. Some tropical hermit crabs are also terrestrial. One of them, Coenobita clypeatus, lives in trees. Snapping shrimp use their
xviii+598_Biodiversity_F.indd 156 11/17/11 1:12 PM

crustaceans 157
claws to produce a shock wave that is strong enough to actually kill small prey. Pentastomes look just like parasitic worms that live in vertebrates, with few external features to identify them as crustaceans. Previous to the study of Dna evidence, they were classified in their own phylum.
Some crustaceans have complex behavior. Several dozen individuals of a species of atlantic spiny lobster, Panulirus argus, form long lines and maintain touch contact with their antennae. Migrating in long lines reduces the drag from water currents and gives them protection from predators—not unlike a V of migrat-ing geese. Some species of snapping shrimp form colo-nies, with a reproductive queen, inside sponges.
evolutionary diversification of crustaceans began very early in the history of multicellular life. By the early cambrian period, several crustacean lineages had already differentiated. this makes it difficult to reconstruct the evolutionary history of this group of arthropods.
the most diverse and well-known group of crus-taceans are the Malacostraca. their bodies have these characteristics.
● the cephalothorax consists of the head and thorax and is usually covered by a protective
carapace. the head has six segments, with appendages that include mouthparts and large and small antennae. the thorax has eight segments, each with a pair of legs, that can be pincers, walking legs, or small legs that manipulate food. the thorax is covered by a shell or carapace.
● the abdomen has six segments, with small legs, some of which are modified for swimming.
the largest group in the Malacostraca is the decapods, which include crabs, crayfish, lobsters, prawns, and shrimp. Decapods (from latin for “ten legs”) have five pairs of walking legs on the thorax. they manipulate food with the appendages of the head, which are sometimes modified into large pin-cers. the appendages of the abdomen are often mod-ified for swimming. the females of most decapods carry the fertilized eggs beneath their abdomens until they hatch. important groups of decapods are:
● True shrimp are primarily marine filter feeders with large populations. they are able to swim backward rapidly. a female shrimp can release up to a million eggs, which hatch into tiny nauplius larvae that live on yolk reserves. they develop through a series of larval phases until they resem-ble small adults. Many shrimp migrate into estuaries at this stage. the preservation of shrimp populations, on which many marine fish depend, is one reason estuaries must be protected. Prawns differ from true shrimp primarily in having branched gills.
● Lobsters are mostly solitary marine preda-tors. their front legs are modified into large pincers. they can live for up to a century, and some have reached 44 pounds (20 kg).
● Crayfish (or crawdads) are omnivorous freshwater relatives of lobsters. they occa-sionally live in mud burrows.
Other decapods include the following. in steno-pods (from Greek for “narrow feet”), it is the third pair of walking legs that is modified into large pin-cers. in the blind polychelids (from Greek for “many claws”), most or all of the walking legs have claws. in achelates (from Greek for “no claws”), often called spiny lobsters, none of the legs have claws, and the antennae are very large.
Other groups of Malacostraca include the following.
● Krill (euphausids) look like shrimp but are usually smaller, and the head and thorax are
Massive populations of red crabs (Gecarcoidea nata-lis) periodically migrate across Christmas Island in the Indian Ocean. (Attorney General’s Department, Australian Government)
xviii+598_Biodiversity_F.indd 157 11/17/11 1:12 PM

first entry158 cultural diversity 158
fused together. they are very abundant filter feeders and the basis of many marine food chains. Baleen whales eat large quantities of them.
● Isopods are flattened vertically and have seven pairs of legs. Most are aquatic, but some have diversified into terrestrial habi-tats. Pillbugs (genus Armadillidium), also known as woodlice, roly-polies, or crawly-ballies, live in moist recesses under rocks and logs, where they scavenge dead plant material.
● Amphipods look like shrimp but are flat-tened from side to side. Some of the append-ages of the thorax are enlarged into pincers.
● Mantis shrimp (stomatopods) are not shrimp. they are predators with powerful pincers.
● Some groups of malacostracans are simple organisms of caves or hot springs.
Other groups of crustaceans, aside from mala-costracans, include the following.
● the maxillopods are crustaceans with reduced abdomens and include the copepods and the barnacles. adult barnacles cement themselves to solid surfaces (including the skins of whales) in the ocean and secrete a limestone shell. their legs have modified into comblike structures with which they sweep in small bits of food. the arrangement of organs within the adult barnacle gives little hint to its arthropod identity, and barnacles were once considered mollusks until the lar-val form of the barnacle, with recognizable arthropod features, was identified. Many barnacles attach directly to rock and other surfaces, while some, such as gooseneck bar-nacles, grow a stalk. Some, such as genus Sacculina, are parasites of other crustaceans.
● Branchiopods are small, shrimplike crusta-ceans that live as plankton in fresh, brine, salt, and hypersaline conditions. they have flattened abdominal appendages that have gills, and they have hairs that help trap small particles of food. they have two pairs of antennae, the second of which is used for propulsion. Because they breed abundantly, brine shrimp (genus Artemia, also known as sea monkeys) and Daphnia are used as fish food and as experimental organisms.
● Ostracods are small, flattened, planktonic crustaceans.
● Remipedia and cephalocarida are groups of blind, elongated marine crustaceans.
Many of the 35,000 species of crustaceans are abundant and important parts of aquatic food chains.
See also arthropods.
Further readingBayly, ian a. e., and Geoffrey a. Boxshall. “an all-conquer-
ing ecological journey: From the sea, calanoid cope-pods mastered brackish, fresh, and athalassic saline waters.” Hydrobiologia 630 (2009): 39–47.
Browder, Joan a., and Michael B. Robblee. “Pink shrimp as an indicator for restoration of everglades ecosys-tems.” Ecological Indicators 9 (2009) (supplement): S17–S28.
cumberlidge, neil, et al. “Freshwater crabs and the bio-diversity crisis: importance, threats, status, and con-servation challenges.” Biological Conservation 142 (2009): 1,665–1,673.
Duffy, J. e. “eusociality in a coral reef shrimp.” Nature 381 (1996): 512–514.
MacDonald, c. D., et al. “Queuing behavior of the Hawai-ian spiny lobster Panulirus marginatus.” Bulletin of Marine Science 35 (1984): 111–114.
Schram, F. Crustacea. new york: Oxford University Press, 1986.
taylor, Jennifer R. a., and William M. Kier. “Switching skeletons: Hydrostatic support in molting crabs.” Sci-ence 301 (2003): 209–210.
Versluis, Michel, et al. “How snapping shrimp snap: through cavitating bubbles.” Science 289 (2000): 2,114–2,117.
cultural diversity Genetic diversity is essential for the continued evolution and survival of wild populations of organisms; it is an essential part of biodiversity. in animal populations, mimetic, or cul-tural, diversity is also very important to survival and continued evolution. this fact has tremendous impli-cations for the preservation of animal biodiversity on this planet.
the evolution of animal adaptations has fre-quently occurred by gene-culture coevolution. ani-mal populations invent new behavioral patterns (often done by one or a few brilliant individuals within the population) that are an important component of their success. these behavioral patterns are not encoded in the genes. However, they create a situation in which natural selection favors genetic traits that facilitate the behavior patterns. Over time, the behavior may even become genetically encoded. in many cases, however, the traits are useless unless the animals have learned the behavior that makes use of them.
the behavioral patterns themselves are trans-mitted through learning. Passive learning occurs when young animals watch adults handling prey and working out social interactions. in a few cases,
xviii+598_Biodiversity_F.indd 158 11/17/11 1:12 PM

cultural diversity 159
nonhuman animals actively teach their young. adult meerkats, for example, partially disable their prey (scorpions) so that the young meerkats can easily kill and eat them, and the adults disable the prey less for older juveniles than they do for younger juveniles (see figure).
if the chain of transmission is broken, the cul-tural component of adaptation can be lost forever. this has occurred several times when wild animal populations have been driven into extinction and conservationists have reintroduced zoo animals into the habitats. animals that have been fed by zookeep-ers do not know how to hunt. Sometimes, there are even more basic behavioral lapses suffered by zoo animals reintroduced into the wild. Bears have been extinct in the forests of northern France for centu-ries, and when zoo bears were released there in the early 21st century, some of them fell off of cliffs and died. the zoo bears had not learned what cliffs were. Bears with mental problems, which would probably have died in the wild, can survive in captivity. it was one of these “psychotic” bears that wreaked havoc in France when it was released. the basic structure of bird songs is encoded in their brains, but young birds learn their specific songs from listening to adults. Occasionally, birds learn the songs of the wrong parental species. the repertoire of bird song is lost when hatchlings raised by humans are the founders of new populations.
in zoos, at least a limited amount of cultural transmission occurs from one generation to another. if an extinct or nearly extinct species is produced by cloning, even this limited amount of cultural information is lost. the guar is a nearly extinct Southeast asian wild cow. Researchers at advanced cell technologies cloned a guar inside the egg and with surrogate mother of the domesticated cow in late 2000. even though it apparently had no genetic defects, the young cloned guar died shortly after birth from a bacterial infection. even if it had sur-vived, it would have been helpless if introduced into the wild. Some researchers want to clone mammoths and mastodons inside the eggs and with surrogate mothers of elephants in order to reintroduce them into habitats in which they once lived. Such animals may, in time, invent new cultural adaptations. the genetic rescue of endangered species is better than extinction, but the preservation of wild populations is a far superior process, not least because the lines of cultural transmission are intact.
europeans and white americans once consid-ered human cultural diversity to be debris that the advance of white civilization would clear away. the eradication of tribal cultures was official policy, even when the lives of the people were spared. the indian schools of the United States were a notorious
example, in which children from many tribes were boarded, taught english and the white american lifestyle, and forbidden to speak in their native lan-guages. in the 20th century, european and white american cultures began to realize that native cul-tures were valuable in their own right, and both they and the tribes tried to rescue the remains of sometimes shattered cultures. today, for example, there may be no member of the cherokee tribe who cannot speak english. the cherokee language is therefore unnecessary for tribal survival. neverthe-less, the tribe puts much effort into maintaining the language, which includes a unique written syl-labary invented by the cherokee scholar Sequoyah in the early 19th century. the tribe not only prints many articles in both english and cherokee in the tribal newspaper, but has begun immersion schools in which the students must speak cherokee. the preservation of tribal languages and cultures is not merely an attempt to counteract the unfair practices of the past. each language represents a uniquely different way of understanding the world and orga-nizing mental concepts. the huge corpus of human knowledge, far exceeding the capacity of any group of people to memorize and transmit, is perhaps the most important human adaptation. Human cultural
Many vertebrate species, such as these African meer-kats, have complex social behavior, which is an adap-tation culturally transmitted from one generation to the next. (Vivapets)
xviii+598_Biodiversity_F.indd 159 11/17/11 1:12 PM

first entry160 cyanobacteria 160
diversity is therefore an essential part of this adap-tation. Scholars have used language analysis along with genetic analysis to reconstruct the history of human migrations. therefore, the preservation of cultural diversity is important to understanding human history as well as to promoting the continued success of the most important human adaptation.
Further readingBBc news. “endangered animal clone dies.” available
online. URl: http://news.bbc.co.uk/1/hi/sci/tech/ 1113719.stm. accessed March 21, 2009.
campbell, Matthew. “ ‘Psycho’ bear tears apart eco-idyll.” Sunday Times (london), (July 29, 2007). available online. URl: http://www.timesonline.co.uk/tol/news/world/europe/article2159387.ece. accessed May 12, 2011.
thornton, alex, and Katherine Mcauliffe. “teaching in wild meerkats.” Science 313 (2006): 227–229.
cyanobacteria See bacteria.
xviii+598_Biodiversity_F.indd 160 11/17/11 1:12 PM

161
darwin, Charles (1809–1882) British Evo-lutionary scientist charles Darwin (see fi gure) was the scientist whose perseverance and wisdom changed the way professional scientists, amateurs, and almost everyone else viewed the world. He explained where all biodiversity comes from. Dar-win presented a convincing case that all life forms had evolved from a common ancestor and for a mechanism, natural selection, by which evolution occurs. earlier scholars had proposed evolutionary theories, but, until charles Darwin, no proposals of evolution were credible. Perhaps just as important is the fact that he explained that all the diversity of life sprang from a common ancestor.
charles Robert Darwin was born February 12, 1809, in Shrewsbury, england, the same day that abraham lincoln was born in the state of Kentucky. Both men, in different ways, led the human mind to consider new possibilities and freedoms without which future progress would not have been possible. Darwin, like many other aristocratic englishmen, was born into a rich and inbred family. His father, Robert Darwin, was a prominent physician, and his mother, Susannah Wedgwood Darwin, was an heir of the Wedgwood pottery fortune. Robert Darwin and Susannah Wedgwood were, in addition, cous-ins. Unlike other rich, inbred englishmen, charles Darwin was to put his inherited wealth to better use than perhaps anyone ever has.
the Darwin family was respectably but not pas-sionately religious. Robert Darwin was secretly a freethinker but remained associated with the angli-can church. Susannah Darwin continued the Wedg-wood family tradition of Unitarianism. the family had a history of creative thinking that went beyond the bounds of common opinion. Robert Darwin’s
father, erasmus, also a physician, doubled as a poet and wrote books in which he openly speculated on evolution—even to the extent of suggesting that life began in a warm little pond from nonliving chemi-cals. charles Darwin grew up, however, with a nega-tive view of evolution. the only major evolutionary theory that had been considered by scientists was that of Jean Baptiste de lamarck, which to the Brit-ish elite was associated with the chaos of the French Revolution. While young Darwin was not repressed into religious narrowness, he explored the path of evolutionary theory hesitantly and on his own.
though not passionate about religious doctrine, the Darwin and Wedgwood families were passion-ate about the religious argument against slavery. Both families contributed considerable money to the antislavery movement in england and put much per-sonal effort into publicizing the abuses of slavery and insisting that england prohibit slavery in all of its colonies. the Wedgwood fortune came from the production of ceramics. charles Darwin’s uncle, Josiah Wedgwood, produced the famous plaque that depicted a slave in chains, with the words, “am i not a man and a brother?” and distributed thousands of them free of charge. charles Darwin was raised to hate slavery.
Meanwhile, there was plenty of opportunity for charles Darwin to develop scientifi c interests. His family kept a dovecote, and he collected insects and rocks and performed chemical experiments (earning the nickname “Gas” among his friends). Darwin got none of this important training from the schools he attended. He learned adequately in school, but noth-ing except direct contact with the world of nature inspired him. Being a naturalist, however, was not a career. When it came time for him to go to college,
D
xviii+598_Biodiversity_F.indd 161 11/17/11 1:12 PM

first entry162 darwin, Charles162
he followed his older brother to the University of edinburgh to study medicine.
Darwin stayed in edinburgh about a year. First, he could not keep his mind on the study of medicine. He spent more time attending lectures in geology and zoology and was more interested in the reproduction of marine invertebrates than in surgery. During his brief stay, he was exposed to some of the radical anticlerical views of fellow students and even of one professor, the zoologist Robert Grant, who openly defended lamarckian evolution. edinburgh Univer-sity, being free from anglican theology, was a haven for scientific and political debate. While Darwin experienced the bracing atmosphere of free inquiry, he also saw what could happen if one went too far. One student presented a paper at a local scientific society claiming that the human mind consisted only of the brain, without any soul; his remarks were stricken from the record. During his year and a half in edinburgh working with Grant, Darwin learned how to conduct scientific research, and he presented his first paper there in 1827 on a discovery he had made about the life cycle of the cnidarian Flustra. Second, Darwin could not stand the sight of blood. When he had to witness operations administered without anesthesia, he decided to abandon plans for a medical career.
another experience that Darwin had in edin-burgh was formative of his entire future. He hunted animals, primarily birds, and wanted to learn to pre-serve them. He learned taxidermy from a freedman named John edmondstone, who had been a slave in South america. Darwin spent a lot of time talking to him and discovered three things. First, he wanted to travel the world and see some of the amazing diversity of plants and animals that John described. Second, he heard eyewitness accounts (as opposed to the second-hand stories his family used in their tracts) of the brutality experienced by black slaves in South america. third, he learned that black people were no less intelligent and had the same capacities for emotion and morality as the British whites—which put him in conflict with many leading British intellectuals.
after leaving edinburgh, Darwin chose to attend cambridge and study theology. He planned to become a country parson. at the time, when the church of england was securely supported by the government, a country parson needed only to per-form his round of duties, then he would have plenty of time for studying birds and flowers and rocks. Some of the best natural history studies, such as the Natural History of Selborne, written by parson-naturalist Gilbert White, were the work of coun-try ministers. even at cambridge, Darwin spent far more time attending lectures in science classes in
which he was not actually enrolled than in studying theology. He befriended the botany professor John Stevens Henslow and frequently went on field trips with the botany students. He was particularly pas-sionate about collecting beetles. Robert Darwin was openly skeptical about the prospects for success of his son, who, he said, cared “for nothing but shoot-ing, dogs, and rat-catching,” adding, “you will be a disgrace to yourself and all of your family.” at the last minute, before his comprehensive examinations, Darwin studied enough to earn a respectable grade in theology.
there was one aspect of his theological studies that charles was eager to pursue. He was required to read Reverend William Paley’s book Natural The-ology, which maintained that the evidence of geol-ogy and biology strongly indicated that there was a creator. at the time, the only alternative to natu-ral theology was some unspecified and mysterious process of self-formation, of which most scientists were understandably skeptical. little did Darwin, who nearly memorized the book, suspect that he would be the one to present a credible, nonmiracu-lous mechanism by which nature could design itself.
Darwin’s future seemed clear to him: He would get married, assume a parsonage, and settle down to live in the country, where he could collect beetles forever. it would not pay much, but charles was in line to inherit at least part of a small fortune. then, at the last minute, an unexpected opportunity arose. the captain of the HMS Beagle, Robert FitzRoy, needed a companion for a voyage around the world. it was a scientific expedition dedicated mainly to surveying the major ports of the world (into which the British assumed they had unlimited access) but also to every other aspect of scientific exploration. the ship already had a naturalist, the ship’s sur-geon, but the captain wanted a fellow aristocrat with whom to share meals and conversation. FitzRoy con-tacted Henslow, who told him the perfect person for the job would be charles Darwin. it was hardly a job—Darwin had to pay his own way—but it was an opportunity to see the wonders of creation around the world in a way that, as it turned out, Darwin would never have again. FitzRoy interviewed Dar-win and offered him the post. this voyage changed Darwin and changed the world. Robert Darwin at first refused to pay for his son’s trip, but Josiah Wedgwood convinced Robert to support charles.
During the 1831–36 voyage of HMS Beagle, Darwin was constantly putting his knowledge of natural history to work. the Beagle went down the east coast of South america to the west coast, across to australia and the indian Ocean, around africa, and back to england. there were long periods in which the ship remained in harbor for surveying
xviii+598_Biodiversity_F.indd 162 11/17/11 1:12 PM

darwin, Charles 163
work, during which Darwin was able to go ashore and travel far inland. this was fortunate for him, for he suffered constantly from seasickness. Darwin studied the geology of islands and coastlines; he col-lected fossils as well as plants and animals. He took careful notes on everything and periodically sent his specimens back to england. He visited tropical rain forests and the heights of the andes Mountains. the rain forest was, for him, a particularly spiritual experience; one could not look at the beauties of such a forest, he said, without believing that there was more to man than merely the breath of his body.
even so, the voyage had more than spiritual inspiration for him. He read the first volume of the geologist Sir charles lyell’s famous and controver-sial Principles of Geology. While on the voyage, he received the other volumes and read them as well. He was convinced that lyell was correct about the long ages of earth history and the gradual changes that had occurred during them—in contrast to an earth that had been recently created, or at least recently re-created, by a God whose main purpose was humankind. in addition to this, many of the things that Darwin saw made him question some aspects of his belief in creation. He collected fossils and found that animal fossils most nearly resembled the animals that were currently alive in the region. For example, South america is where llamas, capy-baras, sloths, and armadillos now live; this is also where the fossils of extinct species of what appeared to be llamas, capybaras, sloths, and armadillos were found. australia was the place to find most of the marsupials, and most of the marsupial fossils, of the world. it suggested strongly to him that the fossils represented ancestors of the animals now present, and when he saw the animals of the Galápagos islands, he began to wonder if at least some species had come into existence on the islands where they were now found.
Darwin also had experiences with people on this voyage that would influence his later thoughts about evolution and biodiversity. in particular, one of the objectives of the Beagle voyage was to return three Fuegian native americans to their tribes as missionaries. two men and one woman had been captured from a Fuegian tribe and taught christian-ity and english life. now they were going back to enlighten their tribe. instead, they reconverted to their tribal ways. this made captain FitzRoy upset but made Darwin think about why the people whom europeans called primitive had levels of intelligence nearly or completely equal to theirs. He also saw slaves in Brazil (he did not visit the United States) and developed a deep hatred of slavery and of violent and brutal european colonialism. in particular, he was bothered by the way slave owners used Bibli-
cal passages to justify slavery. Perhaps these experi-ences laid the groundwork for his later rejection of christianity.
Darwin’s firsthand observations in South amer-ica not only filled him with a lifelong hatred of slavery but also had an important, and perhaps the decisive, effect on his views of evolution and biodi-versity. He believed passionately that all humans had a common ancestor, that the darker races were not separate creations. He jotted his thoughts in secret notebooks, which scholars have now analyzed. in one passage, he clearly links the idea that all spe-cies have a common ancestor to his belief that all humans have a common ancestor.
Darwin returned to england as he had left, still believing in some form of creationism, but his mind was prepared for rapid changes of viewpoint. Upon his return, he found that he had become a bit famous among the scientific community because of his cor-respondence and his specimens. lyell, instead of just being a hero who wrote a book, became a friend. Darwin spent a couple of years in london catalog-ing his specimens and writing his travel notes, which became a popular book, The Voyage of HMS Beagle.
Darwin married his cousin emma Wedgwood. emma was very firmly religious. even before mar-riage, Darwin had already begun to entertain doubts about the religious view of the world. in later years, as Darwin developed his evolutionary theories, he
Charles Darwin in old age
xviii+598_Biodiversity_F.indd 163 11/17/11 1:12 PM

first entry164 darwin, Charles164
had an ever more difficult time reconciling them with christianity, even though some of his friends (espe-cially the Harvard botanist asa Gray) managed to retain conventional christian faith while embracing evolutionary science. By the end of his life, charles Darwin was an agnostic. Historians have considered that the death of Darwin’s beloved daughter annie at age 10 may have been the precipitating factor in making him abandon belief in, at least, a providen-tial God. all this time, Darwin was in great pain that his loss of faith was distressing to emma, which indeed it was. When they wrote notes to each other about it, she remained his loyal partner even though she feared he was losing his soul, and he responded, “you do not know how many times i have wept over this.” Darwin’s religious agnosticism, therefore, was not easy or arrogant but came at the expense of the pain that he caused his dearest friend.
charles and emma bought a country house in Down, just far enough from london to be able to visit it but not be bothered by it. as it turned out, it would not have mattered how far away from lon-don he lived, because Darwin became ill at this time, remained so for the rest of his life, and hardly ever traveled. He suffered recurring symptoms of headaches, heart flutters, indigestion, boils, muscle spasms, and eczema, on which he kept scientific notes but for which neither he nor any physician had a solution. Several possibilities have been proposed for the cause of Darwin’s illness. Some scholars have suggested that he had contracted chagas disease, caused by a parasite from a bug bite, while in South america. this is unlikely, because Darwin was developing some of his symptoms shortly before the voyage began. Other scholars have suggested that his illness was genetic in origin and resulted from the inbreeding of the Darwin and Wedgwood fami-lies. emma’s brother Josiah and his wife, who were also first cousins, lost a sickly baby. it has also been suggested that recessive mutations from inbreeding caused the death of Darwin’s daughter annie. the most commonly accepted explanation is that Dar-win was nervous about announcing his controversial theory to the world.
emma was the perfect companion and nurse. emma’s care and friendship enabled Darwin to con-tinue his research and writing despite his illness and despite the criticisms he received throughout his life. She would play the piano for him, and they would read together in the evenings. the conclusion seems inescapable that charles Darwin could not have accomplished what he did had it not been for emma.
Darwin’s illness kept him home, and his wealth meant that he did not have to work. He was, accord-ingly, able to devote full time to the study of sci-ence—that is, the few hours of the day in which he
was not suffering the symptoms of his malady. He read every scientific book and article he could find and corresponded widely with fellow scientists all over the world. the solitude, especially during his strolls on the Sandwalk around his property, allowed him to think. He put the time to good use, not only proposing the modern theory of evolution, but also almost singlehandedly inventing the sciences of ecol-ogy, pollination biology, and plant physiology. Until the very end of his life, he noticed things that hurried and shallow people did not: His last book was about the slow but cumulative effects of earthworms on transforming the landscape and creating the soil.
it was not long after the Darwins moved to Down House that charles had to deal with a crisis of thought. By 1837, he had worked out the theory of natural selection. He thought of natural selection, first, because he was very familiar with individual differences within populations, whether of pigeons or of people, and second, because he realized that populations could grow rapidly. He got the latter idea from reading Malthus’s Essay on Populations. Malthus said that because populations grow by dou-bling but that resources increase, if at all, only in a linear fashion, then people were condemned to an eternal cycle of overpopulation and famine. though Malthus, a clergyman, used this argument against the concept of Divine Providence, he nevertheless concluded that humankind made progress because of this suffering. Darwin, his christian faith already slipping, undoubtedly rejected Divine Providence along with Malthus, but he also realized that the whole world of plants and animals was similarly condemned to eternal overpopulation. Out of this crisis came an insight that changed the world. Dar-win realized that overpopulation in nature could be a creative force as well as a destructive one, just as Malthus had seen it to be for the human species. the superior individuals in a population would be the ones to survive, and this would cause the population to evolve.
One of the consequences of Malthus’s doctrine was the political opinion that any attempts to help the poor through public works and welfare would only cause their populations to grow larger and to increase the total sum of their misery. throughout Darwin’s adult life, there were riots and political movements in which the poor and their populist leaders threatened the entrenched powers of lords, priests, and the leaders of industry. a new set of Poor laws in england specifically invoked Malthus as the justification for forcing the poor to leave england, work for lower wages, or be content with poverty. the political theory of the Malthusians was that ruthless competition among companies guar-anteed the progress of england and of humankind.
xviii+598_Biodiversity_F.indd 164 11/17/11 1:12 PM

darwin, Charles 165
it was exactly the political viewpoint that philoso-pher Herbert Spencer would later defend, inventing the phrase “survival of the fittest.” Darwin feared that by incorporating Malthus into his theory of natural selection, he might be seen to side with those who oppressed the poor, an association he did not desire. even today, Darwinian natural selection is often associated with ruthless competition. the nov-elist Kurt Vonnegut, for example, said that Darwin “taught that those who die are meant to die, that corpses are improvements.” During the 20th cen-tury, evolutionary scientists proposed ways in which natural selection could lead to cooperation rather than just to competition.
Darwin began to keep a notebook on the subject of natural selection but told almost nobody about it for many years. He had seen what would happen if someone came up with a half-baked theory of evolu-tion and rushed it into print: an anonymous author (now known to be Robert chambers) wrote such a book in 1844. Darwin did not want his theory to suffer such deserved ridicule. He was afraid to admit that he accepted evolution (called at that time trans-mutation) partly because it was the reigning doc-trine of political revolutionaries. He told his friend the botanist Sir Joseph Hooker that admitting that he believed in transmutation was “like confessing a murder.” Darwin wanted to make sure that he had all the scientific facts in order, to certify every aspect of his theory, before he published it. this was a tall order. Darwin had to demonstrate that populations of plants and animals did have a tremendous amount of variability, that plants and animals did experience overpopulation and other natural stresses, that selec-tion can produce significant changes in populations, and that an evolutionary explanation fit the facts of geology and of the distribution patterns of plants and animals. He gathered information for 21 years, showing that pigeon breeders could use artificial selection to produce astonishing new varieties start-ing only with the variation present in wild pigeon populations, that plants and animals did suffer the same population stresses that humans do, and that an evolutionary pattern would explain the living and extinct species of barnacles. Darwin spent eight years studying every kind of barnacle in the world, living or extinct. it became quite a burden to him; no man, he said, hated a barnacle quite as much as he did. His son George grew up seeing him study barnacles, and he assumed that is what grownups did. George asked one of his friends, “Where does your father study his barnacles?” Darwin’s evidence piled up, nicely matching his theory of natural selection, but after 21 years, he had hardly begun. He might have worked until he died without completing the “big book” in which he planned to explain everything.
in 1858, Darwin received a letter and manu-script from alfred Russel Wallace, a young english naturalist in indonesia. Wallace had figured out all the same aspects of natural selection that Darwin had, only he was ready to rush into print with it. Fortunately, Darwin had written a brief summary of his theory in 1842 and had shown it to lyell, Hooker, and emma. lyell could therefore vouch that Darwin had thought of natural selection first. natural selection was presented as a joint theory of Darwin and Wallace in 1858. Some scholars, such as the science historian loren eiseley, have said that evolution by means of natural selection was an idea that was in the air already, and Darwin happened to be the first person to write about it. the science historian Peter Bowler has pointed out, however, that Darwin’s version of evolution differed strikingly from all the others, even from that of Wallace, who had not thought of the importance of plant and ani-mal breeding as evidence for the heritable variation within natural populations.
For the next year, Darwin worked on what he considered to be only a brief abstract of his ideas, which became a 400-page book, On the Origin of Species by Means of Natural Selection. the editor of the Quarterly Review read an advance copy of the book. He thought it was good but that nobody would be interested in it. He suggested that Darwin write a book about pigeons instead; he pointed out that everybody likes pigeons. However, he was wrong about the popular reception of Darwin’s book. the first printing, 1,250 copies, of The Origin of Species sold out the first day. Perhaps the buyers of Origin of Species were expecting to find something about human evolution in this book, but Darwin stayed away from that subject, saying only that “light will be thrown on the origin of man and his history,” a statement that remained unchanged in later editions.
in the Origin of Species, Darwin argued con-vincingly that all life had a common evolutionary origin and history and that evolution was caused by natural selection. When Darwin’s friend thomas Henry Huxley read it, he is reported to have said, “How stupid not to have thought of this before!” it was primarily to the fact of evolution rather than to the mechanism of natural selection that Huxley referred. natural selection remained a theory seldom embraced until the 20th century. the codiscoverer of natural selection, alfred Russel Wallace, defended both evolution and natural selection but argued from it (quite differently from Darwin) that the human mind was a product of miraculous creation. Dar-winian evolution (soon called Darwinism) survived numerous attacks, for example from the biologist St. George Jackson Mivart. Some of the criticisms, such as those of the engineer Fleeming Jenkin, stemmed
xviii+598_Biodiversity_F.indd 165 11/17/11 1:12 PM

first entry166 darwin, Charles166
not so much from problems with evolution as from the fact that biologists did not as yet understand how genetic traits were inherited. One of his fierc-est critics was a man who had once been his friend, the anatomist Richard Owen, who accepted some aspects of transmutation but vehemently rejected Darwin’s nontheistic mechanism of it. after this brief flurry of controversy, within a decade most scientists had accepted Darwin’s proposal of the fact of evolution. (creationism is primarily a product of the 20th century.)
Darwin, being ill, did not defend his views in public. He left this primarily to his “bulldog” friend Huxley. Meanwhile, in america, it was the botanist (and traditional christian) asa Gray who publicly defended Darwinian evolution.
the world did not have to wait long for Darwin to make very clear statements about human evolu-tion. in 1871, he published a book, The Descent of Man, stating that humans evolved from african apes. Since the size of the human brain and the diversity of human racial characteristics seems to be almost outlandish in proportion to what survival would require and natural selection would produce, Darwin proposed another mechanism of evolution in this same book: sexual selection, by which charac-teristics evolved in one sex because they were pleas-ing to the other sex, allowing the individuals that had these characteristics to produce more offspring. in case anyone thought Darwin meant that only the human body had evolved and not the human spirit, they could read another of his books, The Expres-sion of Emotions in Men and Animals.
Darwin made yet other very important contri-butions to the study of biodiversity. three of them were about pollination. He showed how intricate the process of coevolution could be in his 1862 book about the pollination of orchids. For one flower, the pollinator of which was unknown, Darwin pre-dicted that the pollinator would be a long-tongued moth. Decades after Darwin’s death, this pollinator was discovered, exactly as Darwin had predicted. in an 1876 book, he compared cross-pollination with self-pollination. in 1877, he wrote a book about dif-ferent forms of flowers on the same plant. He wrote other books about plants, including two books (1875 and 1880) in which he described the ways in which plants move. He also studied and wrote a book about carnivorous plants in 1875. His last book, in 1881, was about the role of earthworms in the formation of soil. all these books were based upon his own greenhouse or field observations and measurements.
although charles Darwin remains a popular target for antievolutionists and for evolutionists who have their own new theories of evolution, the care-
fulness of Darwin’s work and breadth of his imagi-nation have assured that his theory has survived far longer than he might have imagined. His personal integrity and humility have also made him one of the most admired scientific figures in history.
Darwin not only made the study of evolution, but of biodiversity, possible. as the evolutionary biologist Olivia Judson wrote,
Before the “Origin,” similarities and differences between species were mere curiosities; questions as to why a certain plant is succulent like a cac-tus or deciduous like a maple could be answered only, “Because.” Biology itself was nothing more than a vast exercise in catalog and description. after the “Origin,” all organisms became con-nected, part of the same, profoundly ancient, family tree. Similarities and differences became comprehensible and explicable. in short, Darwin gave us a framework for asking questions about the natural world, and about ourselves.
charles Darwin died on april 19, 1882, while emma was cradling his head in her arms. Darwin’s friends (primarily Huxley) successfully arranged to have him buried in Westminster abbey, right next to Sir isaac newton.
See also carnivorous plants; Darwin’s finches; natural selection; pollination; sex-ual selection; Wallace, alfred Russel.
Further readingadler, Jerry. “charles Darwin: evolution of a Scientist.”
Newsweek, november 28, 2005: 50–58.american Museum of natural History. “Darwin Digital
library of evolution.” available online. URl: http://darwinlibrary.amnh.org. accessed august 7, 2011.
armstrong, Patrick. Darwin’s Other Islands. london: con-tinuum, 2004.
Boulter, Michael. Darwin’s Garden: Down House and the Origin of Species. Berkeley, calif.: counterpoint, 2009.
Bowler, Peter J. “Darwin’s originality.” Science 323 (2009): 223–226.
Browne, Janet. Charles Darwin: Voyaging. Princeton, n.J.: Princeton University Press, 1995.
———. Charles Darwin: The Power of Place. new york: Knopf, 2002.
Darwin, charles. Autobiography of Charles Darwin 1809–1882. edited by nora Barlow. new york: norton, 1993.
Desmond, adrian, and James Moore. Darwin: The Life of a Tormented Evolutionist. new york: Warner, 1991.
——— and ———. Darwin’s Sacred Cause: How a Hatred of Slavery Shaped Darwin’s Views on Human Evolu-tion. Boston: Houghton Mifflin, 2009.
xviii+598_Biodiversity_F.indd 166 11/17/11 1:12 PM

darwin’s finches 167
———, ———, and Janet Browne. Charles Darwin. Oxford: Oxford University Press, 2007.
eiseley, loren. Darwin’s Century: Evolution and the Men Who Discovered It. new york: Doubleday, 1961.
eldredge, niles. Darwin: Discovering the Tree of Life. new york: norton, 2005.
Healey, edna. Emma Darwin: The Inspirational Wife of a Genius. london: Headline, 2001.
Judson, Olivia. “the origin of Darwin.” New York Times (February 12, 2009). available online. URl: http://www.nytimes.com/2009/02/12/opinion/12judson.html. accessed May 13, 2011.
Keynes, Richard Darwin. Fossils, Finches, and Fuegians: Darwin’s Adventures and Discoveries on the Beagle. new york: Oxford University Press, 2003.
lamoureux, Denis O. “theological insights from charles Darwin.” Perspectives on Science and Christian Faith 56 (2004): 2–12.
leff, David. “aboutDarwin.com: Dedicated to the life and times of charles Darwin.” available online. URl: http://www.aboutdarwin.com. accessed august 7, 2011.
loy, James D., and Kent M. loy. Emma Darwin: A Victorian Life. Gainesville: University Press of Florida, 2010.
Moore, James. The Darwin Legend. Grand Rapids, Mich.: Baker Book House, 1994.
Quammen, David. The Reluctant Mr. Darwin: An Intimate Portrait of Charles Darwin and the Making of His Theory of Evolution. new york: norton, 2006.
Van Wyhe, John. “the complete works of charles Darwin online.” available online. URl: http://darwin-online.org.uk/Freeman_intro.html. accessed July 7, 2011.
darwin’s finches Darwin’s finches are a group of 14 species of finches that inhabit the Galápa-gos islands off the coast of ecuador. the finches and the islands are famous because charles Darwin visited them in September 1835 during an around-the-world trip on HMS Beagle. Because evolution is occurring rapidly in these finches, they represent an almost unique opportunity for studying evolution in action and seeing how new biodiversity devel-ops. they have become perhaps the most famous examples of how biodiversity originates. Because the species are still differentiating, some experts classify the finches into a slightly different set of species than do other experts.
as had every previous visitor, Darwin noticed the giant tortoises that are unique to the islands. He also observed that the islands seemed to have similar geography and climate yet were inhabited by slightly different groups of animals. He collected specimens of many of the small birds he found. Because their beaks were all so different from one another, Dar-win did not recognize that the birds were all closely related finches. When Darwin returned to london,
the ornithologist John Gould identified them as dif-ferent species of finches. Darwin realized that each island might have had its own species of finch, but he had not taken careful notes regarding from which island he had obtained which finch specimen. Fortu-nately, other visitors to the islands, including Robert FitzRoy, the captain of the Beagle, had kept more careful notes. When Darwin published the account of his voyage on HMS Beagle, he had not yet begun to fully develop an evolutionary theory. a famous passage indicates that he had already begun thinking about evolution: “Seeing this gradation and diversity of structure in one small, intimately related group of birds, one might really fancy that from an original paucity of birds in this archipelago, one species had been taken and modified for different ends.” these finches, then, could be considered the spark that started the fire of evolutionary theory.
Between 5 and 10 million years ago, a group of finches from the South american mainland migrated (or perhaps were blown by a storm) to the Galápagos islands. From this original founding group, the 14 species of finches present today evolved (see table on page 168). Most of the finch species live on more than one of the islands, but none live on all the islands. no island in the Galápagos archipelago has its own unique species of finch. Some species live on or near the ground and eat seeds; others live on or in cactuses and eat seeds, pollen, and nectar; the others live in trees. the tree finches eat fruits, leaves, seeds, and insects, and one of them strips the outer bark and eats the rich inner bark. One group of finches, though it primarily consumes insects, occasionally drinks the blood or eats the eggs of seabirds such as blue-footed boobies. Some finches occasionally perch on iguanas or tortoises and eat the parasites from them. the finches have undergone adaptive radiation in two ways: first, by evolving on different islands, and sec-ond, by specializing on different food resources.
One of the earliest extensive studies of these finches, which brought them to the attention of the world as an excellent example of evolution in action, was the 1947 study by David lack. He visited for only four months, not long enough to get extensive data regarding the ongoing processes of evolution in these finch populations. He built an aviary, in which he was able to confirm that the different kinds of finches did not readily interbreed and were, there-fore, different species. He showed that evolution had occurred, but his evidence did not show that evolu-tion was still occurring.
in order to obtain convincing evidence about natural selection, it is important to closely monitor entire isolated populations of organisms. if this is not done, the researcher cannot determine whether the disappearance of individuals is due to natural
xviii+598_Biodiversity_F.indd 167 11/17/11 1:12 PM

first entry168 darwin’s finches 168
selection (the individuals die or fail to reproduce) or due to migration (the individuals leave). this was one of the problems with what had once been considered a classic study of natural selection, the zoologist H. B. D. Kettlewell’s studies on the pep-pered moths of Britain. Darwin’s finches provided a perhaps unique opportunity to study natural selec-tion and other evolutionary processes, since they are isolated on different islands, to and from which they seldom migrate. a prodigious amount of work is required to keep track of each individual finch, how many offspring it produces, how it lives, and its physical characteristics, even with relatively small populations on small islands. nevertheless, that is precisely what Peter and Rosemary Grant, evolu-tionary biologists at Princeton University, and their associates have been doing on some of the islands since the 1970s. Having monitored the lives of more than 20,000 individual finches they have been able to present convincing evidence of natural selection and other evolutionary processes at work. they can recognize the individual finches, and know the gene-alogies of each, the kinds and sizes of seeds that these individuals eat, and their individual songs as well as the kinds of seeds available on the islands and in what amounts.
the Grants studied the beak sizes of Geospiza fortis ground finches and the relationship between beak size and the kinds of seeds that these finches ate. the ground finches eat seeds. Some plants (for example grasses) produce small seeds that are relatively easy to crack. these plants grow abun-dantly during rainy seasons. During dry seasons, large seeds (such as those of the puncture vine) are the most abundant. the large seeds also have very hard coats. Finches with larger beaks can crack larger seeds. the finches with smaller beaks can more easily handle the smaller seeds. a difference of less than a millimeter in beak size can make a tremendous difference in the food resources avail-able to a finch. a finch with a 0.44-inch (11-mm) beak can crack open a puncture vine seed, while an individual with a 0.42-inch (10.5-mm) beak may not be able to do so. the individuals with smaller beaks may starve, or they may eat seeds from the Chamaesyce bush, which produces a sticky, milky sap. the sap gets on the heads of the birds. the birds use their heads to push gravel around as they search for seeds. the birds with sticky heads, when they do this, rub the feathers off the tops of their heads, leaving the skin exposed, which can lead to overheating of the brain, followed by death. as Darwin wrote, “the smallest grain in the balance, in the long run, must tell on which death shall fall, and which shall survive.”
the Grants have gathered data about natural selection, character displacement, and incipient spe-ciation among Darwin’s finches.
Natural selection in finch populations. in order to show that natural selection is occurring, the Grants had to establish that the beak size differ-ences among the individual Geospiza fortis birds are genetically based. to do this, the Grants switched eggs among nests and found that individual birds had beak sizes similar to those of their biological, not their adoptive, parents.
next, the Grants had to measure changes that occurred in genetically based characteristics, such as beak size. they measured beak sizes of Geospiza fortis during a long drought, when vegetation was sparse and most of the seeds came from the puncture vine. then, during an el niño Southern Oscillation event, a tremendous amount of rain fell, and the island was overgrown with plants that produced smaller seeds. almost immediately, the average sizes of the birds and the sizes of their beaks began to decrease. When the drought returned and the plants that produced small seeds became progressively less abundant, the average body and beak size began to increase in the G. fortis population. this is an exam-ple of natural selection in action, first one direction, then the other.
darWin’s finChes
Ground Finches
Small ground finch Geospiza fuliginosa
Medium ground finch Geospiza fortis
Large ground finch Geospiza magnirostris
Cactus finch Geospiza scandens
Large cactus finch Geospiza conirostris
Sharp-beaked ground finch
Geospiza difficilis
Tree Finches
Small tree finch Camarhynchus parvulus
Medium tree finch Camarhynchus pauper
Large tree finch Camarhynchus psittacula
Woodpecker finch Cactospiza pallidus
Vegetarian finch Platyspiza crassirostris
Gray warbler finch Certhidea fusca
Olive warbler finch Certhidea olivacea
Cocos finch Pinaroloxias inornata
xviii+598_Biodiversity_F.indd 168 11/17/11 1:12 PM

darwin’s finches 169
Character displacement between finch spe-cies. natural selection occurs not only in response to weather patterns but also in response to con-tact between species of finches. On the island of Genovesa, the large cactus finch Geospiza conirostris and the large ground finch G. magnirostris (which has a larger average beak size) live together. During rainy seasons, when there are plenty of seeds, both species exhibit a wide range of beak sizes, and there is overlap between the beak sizes of the largest G. conirostris and the smallest G. magnirostris. the finches in the zone of overlap would compete but do not because there are plenty of seeds for both. Dur-ing droughts, when seeds become scarce, the two species specialize: natural selection favors the G. magnirostris with the largest beaks but favors the G. conirostris with the smallest beaks. this is an exam-ple of character displacement between two species that minimizes competition between them. character displacement is expected to occur only when two species are in contact, and this is precisely what we see with these finches. On the island of española, the large ground finch G. magnirostris is absent. On this island, without competition from the large ground finch, the cactus finch G. conirostris has a much greater range of beak sizes regardless of the weather conditions. competition between species has not only caused character displacement but may have caused some local extinctions as well. Generally speaking, finches that are very similar to one another in size and the type of food they eat are not found on the same island. the Grants documented in 2006 that natural selection favored larger beaks in the medium ground finch, G. fortis, during a drought in 1977, but dur-ing a 2003 drought, after the large ground finch G. magnirostris had invaded the island, natural selection favored smaller beaks. competition had caused evo-lution to proceed in the opposite direction.
Incipient speciation. the Grants have also observed some possible examples of speciation beginning to occur. During a dry year, they identi-fied two different groups of cactus finch, Geospiza conirostris: One group had shorter beaks than the other group. the differences were only slight: the two groups differed by only 15 percent. this was more than twice as much difference in beak size as there was among individuals within each group. the birds with the longer beaks extracted seeds from cactus fruits, while those with shorter beaks frequently tore open cactus pads and ate the pulp. Moreover, the two groups tended to breed sepa-rately; each group had its own type of mating song. this had the elements—genetic variation and repro-ductive isolation—needed to begin the formation of separate species. When the rains returned, the two groups began to blend back together.
computer simulation can retrace the pathway of evolution for the seed-eating finches of the Galápa-gos. a program designed by the evolutionary biolo-gist Dolph Schluter begins with the ranges of beak sizes within each species, the range of seed sizes available on each island, and the presence or absence of competition and calculates the expected ranges of beak sizes on each of the islands. the result of Schluter’s analysis matches the real pattern of beak sizes very closely.
Limits to adaptive radiation. When the finches first arrived on the Galápagos islands, there were no other similar kinds of birds with which to compete. the islands represented a new world open to them for adaptive radiation. they were able to exploit new ways of making a living without having to com-pete with other bird species that already possessed superior adaptations. the woodpecker finch (Cama-rhynchus pallidus), for example, would not have been able to evolve on an island that already had woodpeckers. Some of the cactus finches eat nectar from the flowers of Waltheria ovata, and smaller finches are able to obtain and use nectar more effi-ciently. (Most birds that consume nectar, such as hummingbirds, are small.) On islands that do not have native bees, some of the finches specialize on nectar, although not exclusively: nectar constitutes 20 percent of their diet, and they are smaller than the finches that do not eat nectar. On islands that already have populations of the native bee Xylocopa darwini, nectar makes up only 5 percent of the finch diet, and the finches do not differ in size from those that do not eat nectar.
Hybridization is rare between species of Galá-pagos finches, but when it does occur, the Grants often make note of it. in more than one case, hybrids have proven fertile for more than several generations during wet seasons, but under dry seasons, when the successful individuals are the ones that special-ize, the hybrids (with intermediate beak sizes) have inferior fitness. it appears that it is the dry seasons in which natural selection keeps the species apart.
in the Pacific Ocean, the island of cocos is the home of one of the same species of finches that is found in the Galápagos, the cocos island finch, Pin-aroloxias inornata. Unlike the Galápagos islands, the island of cocos is not large enough to promote reproductive isolation among populations of finches. Speciation is not occurring in these finches. Rather than specializing on food resources, the cocos finches have remained generalists, exploiting many different food resources and learning how to do so by watching other finches.
See also adaptive radiation; Darwin, charles; hybridization; natural selection; speciation.
xviii+598_Biodiversity_F.indd 169 11/17/11 1:12 PM

first entry170 decomposers 170
Further readingGrant, Peter R. Ecology and Evolution of Darwin’s Finches.
Princeton, n. J.: Princeton University Press, 1999.Grant, Peter R., and B. Rosemary Grant. “evolution of
character displacement in Darwin’s finches.” Science 313 (2006): 224–226. Summary by Pennisi, elizabeth. “competition drives big beaks out of business.” Sci-ence 313 (2006): 156.
Sato, a., et al. “On the origin of Darwin’s finches.” Molecu-lar Biology and Evolution 18 (2001): 299–311.
Weiner, Jonathan. The Beak of the Finch: A Story of Evolu-tion in Our Time. new york: Knopf, 1994.
decomposers Decomposers are organisms that consume dead organisms. Because many of them release carbon dioxide into the air and nutri-ents into the soil, they have an essential role in the biogeochemical cycles. Most decomposers live on or in the soil. However, when an animal dies, its formerly commensalistic intestinal bacteria become decomposers. Dead animals are frequently consumed by scavengers such as vultures, and the decomposers consume the leftovers and wastes of the scavengers.
every kingdom of organisms has decompos-ers. Most bacteria (and perhaps some archaea) are decomposers, as are most fungi and many pro-tists. Some plants are decomposers. the coral-root orchid (Corallorhiza) has no chlorophyll. it obtains all of its organic compounds from decomposing wood and forest litter. the most famous decom-poser animal is the earthworm, which consumes soil and digests the organic matter within it. Many small arthropods that live in the soil, such as col-lembolans, are also decomposers. Some beetles are decomposers also. Burying beetles, such as the rare Nicrophorus americanus, bury small mammals and lay their eggs in the carcasses. Scarab beetles (family Scarabaeidae) bury feces and lay their eggs in it. Maggots of thousands of species of flies con-sume carrion.
Decomposition occurs in stages. Decompos-ers and scavengers transform corpses into complex organic molecules, such as the humus that makes soil dark brown. Other decomposers transform these organic molecules into simple inorganic molecules, such as carbon dioxide, ammonia, and phosphate. Many bacteria transform one kind of inorganic molecule into another. For example, some bacteria change ammonia to nitrite, others change the nitrite to nitrate, and some release nitrogen gas into the atmosphere (denitrification).
See also biogeochemical cycles; carbon cycle; nitrogen cycle.
Further readingHaimi, J. “Decomposer animals and bioremediation of
soils.” Environmental Pollution 107 (2000): 233–238.Harte, John, and ann P. Kinzig. “Mutualism and competi-
tion between plants and decomposers: implications for nutrient allocation in ecosystems.” American Nat-uralist 141 (1993): 829–846.
lodge, D. Jean. “Factors related to diversity of decomposer fungi in tropical forests.” Biodiversity and Conserva-tion 6 (1997): 681–688.
Setälä, H., et al. “Functional diversity of decomposer organisms in relation to primary production.” Applied Soil Ecology 9 (1998): 25–31.
Shahid, naeem, et al. “Producer-decomposer co-depen-dency influences biodiversity effects.” Nature 403 (2000): 762–764.
Wardle, David a., et al. “Plant removals in perennial grassland: Vegetation dynamics, decomposers, soil biodiversity, and ecosystem properties.” Ecological Monographs 69 (1999): 535–568.
deep oceans, biodiversity in the deep oceans, or open oceans, occur away from the conti-nental shelves. Because ocean waters are constantly moving, the life of the deep oceans does not have as much of a geographical pattern as does life on the land. Deep ocean habitats can be roughly divided into pelagic and benthic zones.
SurfaCE pElaGIC waTErSthe pelagic zone is the water of the open oceans above the sea floor. Most of the biologically impor-tant nutrients in the oceans come from the continents. therefore, the water of the pelagic zone of the open oceans is relatively poor in nutrients such as nitrogen and iron. Most of the photosynthetic organisms are single-celled protists and cyanobacteria. Because of the low nutrients, these organisms are not abundant in any one location, which is why the oceans are blue and not green. However, because open oceans cover about 70 percent of earth’s surface, there is a lot of photosynthetic productivity in the pelagic zone.
a complex food web depends on these protists and cyanobacteria. Billions of small arthropods, such as krill, eat the photosynthetic and herbivorous protists. Organisms such as the protists and the krill that are small enough that they float without swimming are called plankton. One of the reasons that ocean waters have low nutrient levels is that the photosynthetic plankton organisms absorb them quickly. Small fish and invertebrates such as shrimp eat the plankton, larger fish eat the smaller fish and shrimp, and top predators such as sharks and por-poises eat the larger fish.
xviii+598_Biodiversity_F.indd 170 11/17/11 1:12 PM

deep oceans, biodiversity in 171
the largest animals in the open oceans are whales, which strain plankton from the water with their baleen plates. Since water can hold more organ-isms than air, terrestrial ecosystems contain no ana-log of the large marine filter feeders. For 100 million years during the cretaceous period, large pachy-ormid fishes fulfilled this role in the oceans. after their extinction, rays (family Mobulidae) and sharks (families cetorhinidae and Rhinocodontidae) acted as large filter feeders. Predatory whales evolved early in the Paleogene period, but filter-feeding whales (Mysticeti) did not evolve until 25 million years ago.
atmospheric temperatures and sunlight cause tropical pelagic waters to be warmer than temperate and polar waters. However, the following two pro-cesses cause the open ocean to have less latitudinal difference in temperature than does the land.
● Ocean currents spread warmth among the open oceans. For example, currents carry the very warm surface waters near South-east asia toward South america, bring-ing rain during el niño periods of the el niño Southern Oscillation (enSO). the Gulf Stream carries warm caribbean waters toward europe and is largely responsible for warm european temperatures. england is much warmer than labrador, which is at the same latitude, largely because of the Gulf Stream. Before about 3 million years ago, a large ocean current passed through what is now the isthmus of Panama, carrying even more warm waters into the north atlantic and much of it into an arctic Ocean that was more open than it is today. as a result, prior to the formation of the isthmus, the arctic Ocean was much warmer than it is today. about 30 million years ago, decidu-ous forests grew to the northern extremes of the continents because of the warmer climate created by this ocean current.
● Water retains more heat than land. this is why antarctic waters, though very cold, are not as cold as the antarctic continent. the antarctic Ocean, rich in nutrients, pro-duces enough krill and fish that penguins and other birds that live on the shores of the antarctic continent can survive on them, and whales visit these waters.
currents also move nutrients among the open oceans. the most productive open oceans are not always tropical but are frequently those in which currents bring nutrients up from the ocean depths. this is what happens along the west coast of South
america during the la niña periods of the enSO. the Gulf Stream and other currents largely bypass the Sargasso Sea, leaving it nutrient poor. However, the Sargasso Sea is filled with Sargassum seaweeds upon which a complex food chain depends.
the large organisms of the surface pelagic waters are reasonably well known. the micro-scopic life, like microscopic life everywhere, remains largely mysterious. the biotechnologist J. craig Ven-ter sailed through the Sargasso Sea and analyzed the Dna in water samples. the Dna was sufficiently diverse that it must have come from thousands of unknown microbes.
dEEp pElaGIC waTErSin contrast to the shallow waters, the deep waters remain mysterious. Water pressure increases by about one atmosphere for each 30 feet (about 10 m) of depth. this does not present as much of a problem for the animals that live there all the time as it does for animals, such as whales and humans, who dive. light is also unable to penetrate very far beneath the surface waters. Moreover, some of the deep-est waters may contain very little dissolved oxygen. Many species of giant cephalopods (squid and octo-pus) live in deep pelagic waters. Some cephalopods, such as Vampyroteuthis infernalis, live in waters partway toward the bottom of the sea, while others, such as Architeuthis dux, live nearly at the bottom. Deep pelagic waters are also the home to preda-tory angler fishes, which use bioluminescent lures to attract prey in the dark waters. these waters remain largely unexplored except by remotely controlled probes because of the difficulty of dealing with the tremendous pressures of the deep waters.
BENThIC waTErSBenthic waters occur at the bottom of the sea. Most benthic habitats depend on dead organisms that fall from the surface waters. Widely scattered animals, including echinoderms and angler fishes, live there. the deepest benthic waters occur in the Marianas trench near Southeast asia; the maximum depth is 35,827 feet (ca. 12,000 m) at the challenger Deep.
two types of unusual benthic habitats have been discovered that are not entirely dependent on food items settling down from above. these are the cold seeps and the hydrothermal vents.
Cold seeps. Hydrogen sulfide, methane, and hydrocarbons seep out of some locations on the ocean floor. Some bacteria and archaea can metabo-lize these molecules. larger animals can either eat these microbes, or the microbes may live symbi-otically within their tissues. these animals include tubeworms and clams. the metabolic reactions are
xviii+598_Biodiversity_F.indd 171 11/17/11 1:12 PM

first entry172 deep oceans, biodiversity in 172
very slow, and the animals may have very long life spans. For example, the tubeworm Lamellibrachia luymesi can probably live as long as 250 years. loca-tions of cold seeps include the Gulf of Mexico, Mon-terey canyon off the coast of california, the Sea of Japan, the Pacific Ocean off the coast of costa Rica, the atlantic Ocean off africa, the ocean floor near alaska, and under an antarctic ice sheet. the deep-est cold seep so far discovered occurs at 24,035 feet (7,326 m) in the Sea of Japan.
Hydrothermal vents. at some locations on the seafloor, volcanic activity heats seawater and fills it with hydrogen sulfide gas to make it very acidic. Superheated water (sometimes more than 800°F or [450°c] and which, because of the tremendous depths, does not boil) emerges from the seafloor. When the hot water comes into contact with the very cold water of the ocean depths, minerals precipitate out of the water. if the minerals contain a lot of sul-fur, they form dark-colored cones known as “black smokers.” if the minerals contain a lot of silicon or calcium, they form “white smokers.” the hottest water is actually a supercritical fluid that is neither gas nor liquid.
Hydrothermal vents resemble cold seeps, only they are hotter and metabolism occurs more rapidly. Bacteria and archaea can live in the hottest water and metabolize hydrogen sulfide to form the basis of a food chain. Most of the animals live in cooler water nearby. Small arthropods graze directly on the microbes, while larger animals have symbiotic microbes in their tissues. the most common animals are gigantic tubeworms, especially the species Tevnia jerichonana and Riftia pachyptila. annelid worms, snails, clams, shrimp, and crabs are also common. in one location, the most common animals are eels.
Hydrothermal vents are wellsprings of biodiver-sity. Most of the animal species are closely related to animals that live in shallower waters. Some have evolved unique adaptations. Pompeii worms (Alvi-nella pompejana) can withstand very hot water, and the scaly-footed gastropod Crysomallon squa-miferum has scales of iron on its foot.
dEEp oCEaN CoNSErvaTIoNtwo of the major threats to the open oceans are pol-lution and overfishing.
Pollution. a huge amount of garbage, much of it toxic, is dumped into the oceans. Most sewage con-tamination affects only the waters of the continental shelves, and most metallic garbage sinks to the bot-tom of the ocean. Plastic, however, floats. Pieces of plastic, some large but most of them small, accu-mulate in gyres of slow water surrounded by ocean currents, where they form “garbage patches.” the Pacific trash Vortex is located between about 135°
and 155° west longitude and 35° to 42° north lati-tude. its most highly concentrated portion is about the size of texas. Fishes, sea turtles, and birds often mistake the plastic for prey items; many dead ani-mals have been found with hundreds of pieces of plastic in their digestive systems. the plastic can also transport toxic molecules.
Overfishing. Modern fish harvest technology provides the ability to scoop up hundreds of thou-sands of fishes at a time. techniques include purse-seine nets, drift nets, trawl bags, and very long lines with hooks on them. aerial surveillance also allows many fishing vessels to converge upon schools of fish. Ocean invertebrate populations such as those of shrimp are also overfished. Overfishing has had two profound effects on ocean animal populations:
● the populations of some fish have precipi-tously declined. an example is the atlantic cod (Gadus morhua). Starting in the 1980s, catches of cod increased over previous lev-els by a factor of about four. in 1992, cod populations plummeted. Other fishes whose populations have plummeted are the blue-fin tuna (Thunnus thynnus) and the orange roughy (Hoplostethus atlanticus).
● animals other than fish can get caught in the nets. this “by-catch” can exceed the total weight of the target fish or shrimp. For each pound of shrimp that is caught on the open oceans, there are two pounds of by-catch. thousands of dolphins and sea turtles have perished in this manner.
Overfishing has other effects as well. Millions of people in africa depend on ocean fish for their protein. Fish populations near africa have declined, largely due to overfishing by european vessels. the result is that many africans compensate for this protein by eating “bushmeat,” the flesh of rain for-est animals that are already endangered, including chimpanzees and gorillas. therefore, sustainable fishing is important not only to maintain ocean fish populations but for global food security. Many con-sumers now insist on eating only fish that are har-vested in a sustainable fashion. More information about which fishes are environmentally responsible can be found at the Monterey Bay aquarium and Blue Ocean institute Web sites.
One alternative to open ocean fishing is aqua-culture, in which fish and shrimp are raised in enclo-sures. these enclosures may be on land, in shallow water, or in the open ocean (where the practice is called mariculture). aquaculture can be sustainable but usually is not. the fish may be fed meat that comes from open ocean fish; in these cases, aquacul-
xviii+598_Biodiversity_F.indd 172 11/17/11 1:12 PM

deep oceans, biodiversity in 173
ture results in even more destruction of open ocean populations. crowded aquaculture populations pro-duce concentrated pollution and may experience disease outbreaks. in response, aquaculture man-agers may use antibiotics. the pollution (including antibiotics), as well as diseased fishes, may escape into the wild. Fishes from aquaculture may not only spread diseases to wild populations but interbreed with them. aquaculture fishes grow faster and breed when they are younger, and they may confer the genes for these life history patterns to the wild fish populations.
the good news is that fish populations can often recover once the pressures on them are relieved. Some international agreements have restricted fish harvests, but these mostly affect fishing on the conti-nental shelves, within territorial waters. consumers are beginning to show a preference for fish that have been harvested sustainably. However, fishing on the open oceans remains largely uncontrolled.
See also continental shelves, biodiversity in; coral reefs; pollution.
Further readingBlue Ocean institute. “Guide to Ocean Friendly Seafood.”
available online. URl: http://www.blueocean.org/sea-food. accessed august 8, 2011.
cherel, y., et al. “Stable isotopes document the trophic structure of a deep-sea cephalopod assemblage includ-ing giant octopod and giant squid.” Biology Letters 5 (2009): 364–367.
cowles, David l., and James J. childress. “aerobic metabolism of the anglerfish Melanocetus johnsoni, a deep-pelagic marine sit-and-wait predator.” Deep Sea Research 42 (1995): 1,631–1,638.
Dekas, anne e., et al. “Deep-sea archaea fix and share nitrogen in methane-consuming microbial consortia.” Science 326 (2009): 422–426. Summarized by Ful-weiler, Robinson W., “Fantastic fixers.” Science 326 (2009): 377–378.
Department of Biology, Pennsylvania State University. “life Without light.” available online. URl: http://homes.bio.psu.edu/cold_seeps. accessed July 9, 2010.
deRoos, a. M., et al. “evolutionary regime shifts in age and size at maturation of exploited fish stocks.” Proceed-ings of the Royal Society of London B (Biological Sci-ences) 273 (2006): 1,873–1,880.
Doney, Scott c. “the growing human footprint on coastal and open-ocean biogeochemistry.” Science 328 (2010): 1,512–1,516.
earle, Sylvia. The World Is Blue: How Our Fate and the Oceans’ Are One. Washington, D.c.: national Geo-graphic, 2009.
Friedman, Matt, et al. “100-million-year dynasty of giant planktivorous bony fishes in the Mesozoic seas.” Sci-ence 327 (2010): 990–993.
Halpern, Benjamin S., et al. “a global map of human impact on marine ecosystems.” Science 319 (2008): 948–952.
Hilborn, R. “Salmon-farming impacts on wild salmon.” Proceedings of the National Academy of Sciences USA 103 (2006): 15,277.
———, et al. “Biocomplexity and fisheries sustainability.” Proceedings of the National Academy of Sciences USA 103 (2006): 6,564–6,568.
Kaiser, Jocelyn. “the dirt on ocean garbage patches.” Sci-ence 328 (2010): 1,506.
levin, lisa a., et al. “environmental influences on regional deep-sea species diversity.” Annual Review of Ecology and Systematics 32 (2001): 51–93.
levinton, Jeffrey S. Marine Biology: Function, Biodiver-sity, Ecology. new york: Oxford University Press, 2008.
longhurst, alan. Mismanagement of Marine Fisheries. new york: Oxford University Press, 2010.
Marx, Felix G., and Mark D. Uhen. “climate, critters, and cetaceans: cenozoic drivers of the evolution of mod-ern whales.” Science 327 (2010): 993–996.
Mcleod, Karen, and Heather leslie, eds. Ecosystem-Based Management for the Oceans. Washington, D.c.: island Press, 2009.
Monterey Bay aquarium. “Seafood watch.” available online. URl: http://www.montereybayaquarium.org/cr/seafoodwatch.aspx. accessed august 8, 2011.
Pauly, D., et al. “towards sustainability in world fisheries.” Nature 418 (2002): 689–695.
Pelagic Shark Research Foundation. “Pelagic Shark Research Foundation.” available online. URl: http://www.pelagic.org. accessed May 13, 2011.
Pennisi, elizabeth. “in the deep blue sea: a survey of corals and their kin shows the ocean’s evolutionary poten-tial.” Science 327 (2010): 519.
Rios, l. M., charles Moore, and P. R. Jones. “Persistent organic pollutants carried by synthetic polymers in the ocean environment.” Marine Pollution Bulletin 54 (2007): 1,230–1,237.
Roberts, c. M., and J. P. Hawkins. “extinction risk in the sea.” Trends in Ecology and Evolution 14 (1999): 241–246.
Smith, Martin D., et al. “Sustainability and global seafood.” Science 327 (2010): 784–786.
Sogin, M. l., et al. “Microbial diversity in the deep sea and the underexplored ‘rare biosphere.’ ” Proceedings of the National Academy of Sciences USA 103 (2006): 12,115–12,120.
Stachowicz, John J., et al. “Understanding the effects of marine biodiversity on communities and ecosystems.” Annual Review of Ecology, Evolution, and Systemat-ics 38 (2007): 739–766.
Venter, J. craig, et al. “environmental genome shotgun sequencing of the Sargasso Sea.” Science 304 (2004): 66–74.
xviii+598_Biodiversity_F.indd 173 11/17/11 1:12 PM

first entry174 deserts, cool 174
deserts, cool cool deserts are dry shrublands, usually with grasses. they are found poleward from the warm deserts. they receive moderate amounts of snow in the cold winters but very little rain in the warm summers. Unlike warm deserts, which often result from subtropical dry air at just less than 30° north and south latitudes, cool deserts occur between 30° and 40° north and south latitudes. the main cause of cool desert conditions is rain shadows, in which mountains block moisture from the oceans. Some deserts are so far from oceans that rain and snow do not reach them, even though there is no pronounced rain shadow effect. in some cases, cool deserts are found near oceans with cold water from which little evaporation occurs. examples include the following.
● the Great Basin Desert of north america, which covers much of nevada, Utah, and the northern parts of arizona and new Mexico, is dry because the Pacific mountain ranges (Sierra nevada, White Mountains, cascades) block moisture from the Pacific Ocean and the Rocky Mountains block moisture from the Gulf of Mexico.
● the Owens Valley Desert of california is dry because it is in the immediate rain shadow of the Sierra nevada. it is cool not merely because of its latitude but also because its average elevation is about 5,000 feet (about 1,500 m) above sea level.
● the Patagonian Desert in southern argen-tina is dry because the andes block mois-ture from the Pacific Ocean, and there is little evaporation from the cold currents of the atlantic Ocean.
● the central asian deserts (turkestan Des-ert, taklamakan Desert, Gobi Desert) are far from any ocean and surrounded by mountains.
as in some warm deserts, the shrubs of cool des-erts have deep and expansive root systems. Because of the cold winters, most cool desert shrubs are decidu-ous, unlike most of those in warm deserts. their leaves are small to allow heat to be conducted into the air without much water loss. Some, such as sage-brush, produce slightly larger leaves in the spring, after snowmelt, than in the dry summer. Many cool desert shrubs are remarkably tolerant of salty soils.
Because deserts have come into existence some-what recently in earth history, the cool deserts of the world are largely isolated from one another and have different dominant shrubs (see table). cool deserts intergrade into shortgrass prairies (north america) steppes (central asia), and warm deserts.
Because cool deserts also have grasses, they can support some of the same large animals as the adjacent shortgrass prairies. in north america, pronghorns (Antilocapra americana) move between shortgrass prairie and sagebrush desert. Gazelles (Procapra gutturosa) and wild donkeys known as onagers (Equus hemionus) of the Mongolian steppe also live at the edges of the Gobi Desert. in South america, camel-like guanacos (Lama guanicoe) and large flightless birds called rheas (Rhea americana) live both in grasslands and in parts of the Patago-nian Desert. central asian cool deserts are home to the Bactrian camel (Camelus bactrianus), the two-humped relative of the single-humped dromedary camel that lives in the warm Middle east and Sahara. Both species of camel can tolerate thirst for long peri-ods and drink tremendous amounts of water at oases. the presence of grasses in the sagebrush desert is the reason it is often called sagebrush steppe.
Wildfires maintain grasslands by destroying woody plants. in the tallgrass prairies, the woody plants are the trees of the temperate deciduous for-est. in the shortgrass prairies, it is often the cool des-ert shrubs that are burned. Both overgrazing and fire suppression can cause grasses to become rare and cool desert shrubs to dominate, and both of these processes are symptomatic of human activity in cool desert regions. Severe overgrazing can destroy the shrubs as well. cool deserts withstand fires better than do dry coniferous forests, however, and fire suppression has encouraged the spread of junipers into the Great Basin sagebrush desert.
See also communities, ecological; deserts, warm; disturbances, ecological; grasslands and savannas; temperate deciduous forests.
Further readinganderson, Jay e., and Karl e. Holte. “Vegetation develop-
ment over 25 years without grazing on sagebrush-dominated rangelands in southeastern idaho.” Journal of Range Management 34 (1981): 25–29.
examples of Cool desert shrubs
Great Basin Sagebrush (Artemisia tridentata)
Owens Valley Greasewood (Sarcobatus vermiculatus)
Shadscale (Atriplex confertifolia)
Patagonia Acantholippia deserticola
Benthamiella patagonica
Central Asian deserts
Camel’s thorn (Alhagi maurorum)Saxaul (Haloxylon ammodendron)
xviii+598_Biodiversity_F.indd 174 11/17/11 1:12 PM

deserts, warm 175
Biondini, Mario e., et al. “Secondary successional patterns in a sagebrush (Artemisia tridentata) community as they relate to soil disturbance and soil biological activ-ity.” Plant Ecology 60 (1985): 25–36.
Miller, Richard F., and Jeffrey a. Rose. “Fire history and western juniper encroachment into sagebrush steppe.” Journal of Range Management 52 (1999): 550–559.
Raedeke, Kenneth J., and Javier a. Simonetti. “Food habits of Lama guanicoe in the atacama Desert of northern chile.” Journal of Mammalogy 69 (1988): 198–201.
Romo, J. t., and M. R. Haferkamp. “Water relations of Artemisia tridentata ssp. wyomingensis and Sarco-batus vermiculatus in the steppe of southeastern Oregon.” American Midland Naturalist 121 (1989): 155–164.
Saltz, David, and Daniel i. Rubenstein. “Population dynamics of a reintroduced asiatic wild ass (Equus hemionus) herd.” Ecological Applications 5 (1995): 327–335.
deserts, polar Polar deserts are regions with either bare rock or thick layers of ice, found in the coldest land surfaces on earth. they are called polar deserts because little snow falls, and the bare rocks and small pockets of soil retain little water. they are colder and drier than arctic tundra. examples include the McMurdo dry valleys of antarctica, which have been ice-free for thousands of years; the coast of antarctica; the northern edges of eur-asia and north america; and arctic islands such as Greenland, ellesmere island, Franz Josef land, and Svalbard, all of which have land surface within 10° of latitude of the north Pole.
Some tundra plants can tolerate dry conditions and live in polar desert regions adjacent to the tun-dra. examples are Saxifraga oppositifolia and the dwarf azaelea Loiseleuria procumbens around the arctic Ocean. the only native flowering plants of antarctica are the pearlwort Colobanthus quitensis and the hairgrass Deschampsia antarctica, which live in sheltered rock recesses near the ocean.
as global warming continues, some of the tun-dra plants that now live in polar deserts may be able to expand their ranges and form tundra vegetation. this will not happen rapidly enough to compensate for the loss of the species-rich arctic tundra, which may be rapidly replaced by boreal forests.
See also boreal and subalpine forests; greenhouse effect and biodiversity; tundra.
Further readingBay, christian. “Floristical and ecological characterization
of the polar desert zone of Greenland.” Journal of Veg-etation Science 8 (1997): 685–696.
Gold, Warren G., and lawrence c. Bliss. “Water limita-tions and plant community development in a polar desert.” Ecology 76 (1995): 1,558–1,568.
deserts, warm Deserts are regions with little rainfall. Some deserts have had very little rainfall throughout recorded history. For example, most of the atacama Desert of Peru has had no noticeable amount of rainfall since observations began in 1571. Furthermore, deserts receive only a little snow, and most of their rainfall comes in powerful storms. all desert plants are therefore adapted to long periods of dryness. Warm deserts also have mild winters and hot summers. cool deserts (treated in a separate entry) may have cold winters.
Warm deserts are found in southwestern north america, on the west coast of South america, in north africa and the Middle east, near the west coast of southern africa, and in australia. the three major regions of warm desert in north america are as follows.
● in the Mojave Desert of california, creosote and bursage bushes are regularly spaced, almost like a plantation or a decorated car-pet, because the area between them is filled with roots that soak up the little rain that falls. the only major species of succulent plant in the Mojave Desert is the tree-sized yucca known as Joshua tree.
● in the colorado (or Sonoran) Desert of ari-zona and northern Mexico, many species of cactus predominate as well as acacia trees, under which the cactus seedlings germinate.
● in the chihuahuan Desert of new Mex-ico, texas, and northern Mexico, creosote and mesquite bushes dominate. Because of its proximity to the Gulf of Mexico, the chihuahuan Desert receives more rain than the other deserts, allowing grasses to grow between the bushes.
the major South american deserts are in Peru, especially the atacama Desert. the major african deserts are the Sahara, which covers most of north africa, and the Kalahari Desert, near the western coast of southern africa, particularly in namibia and Botswana. Desert covers much of the Middle east, including the entire arabian Peninsula, and some other eurasian areas, such as the thar Desert of india and Pakistan. the Great Outback of australia is a desert.
CauSES of dESErT CoNdITIoNSWarm deserts have mild winters, with some rainfall, and hot summers, with occasional thunderstorms.
xviii+598_Biodiversity_F.indd 175 11/17/11 1:12 PM

first entry176 deserts, warm 176
the following are three main reasons that warm deserts are dry.
● Hadley cells. Hadley cells are pockets or bands of air circulation, named after the British meteorologist who discovered them. the warmest places on earth are along the equator, where moist air rises above tropi-cal rain forests. Much of the moisture con-denses above the equator and produces abundant tropical rainfall. the air circu-lates poleward, then descends to the surface of earth again at about 30° n and 30° S latitude. the air becomes even drier as it descends and becomes warmer. the return of the air toward the equator, producing trade winds, completes the cycle of each Hadley cell. the result is a band of warm deserts in the subtropical zone, roughly 30° n and 30° S. the northern continents have the greatest land mass and the largest warm deserts, all of them between roughly the 20th and 30th parallels of north latitude. the deserts of the southern hemisphere are also between roughly the 20th and 30th parallels of south latitude.
● Rain shadows. Mountains force moist oce-anic winds upward, where they become cooler and their moisture condenses as rain or snow on the windward side. the wind, now much drier, descends the leeward side of the mountains and becomes warmer and drier. nearly every warm desert is slightly drier than the Hadley cells would make it. the southwestern desert in north amer-ica is surrounded by low mountains, not enough themselves to cause desert condi-tions. the atacama Desert is between the andes and the chilean coast Range, which block moisture from both east and west.
● Ocean currents. coastal regions near cold ocean currents often have little rainfall. this is yet another reason that the atacama Desert receives almost no rainfall.
the extreme conditions of Death Valley in cali-fornia are due not only to Hadley cells and rain shadows but to its low elevation, which allows hot air to accumulate. its lowest point, Badwater, is 282 feet (86 m) below sea level. Plants that live near the bottom, such as the shrub Tidestromia oblongifolia, have photosynthetic reactions that function best at high temperatures (see figure). as the name would suggest, saline water has accumulated at the bot-tom of Death Valley, where Salicornia pickleweeds are almost the only plants. Death Valley was a lake
when glaciers were melting at the end of the most recent ice age.
Warm deserts have the highest average tem-peratures on earth. the highest temperature ever recorded was 136°F (58°c) in the Sahara of libya in 1922. However, temperatures can drop rapidly at night. in the Sahara, a high temperature of 126°F (52°c) was followed by a nighttime low temperature of 26°F (-5°c). Both of these temperature records were made possible by the dry conditions. Water gains and releases heat more slowly than dry soil, causing humid environments to have cooler days and warmer nights.
dESErT plaNT adapTaTIoNS To drouGhTPlants are small and few in deserts compared to other ecological communities, though more abun-dant than casual observers often recognize. Plants have numerous adaptations to make efficient use of the scarce moisture that warm deserts receive. their
A severe rain shadow keeps most of the rain and snow out of Death Valley, California. Conditions at the bottom of Death Valley (282 feet [86 m], below sea level) are very hot, dry, and salty. Few plants can survive there, but Tidestromia oblongifolia thrives under conditions that would kill most complex plants. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 176 11/17/11 1:12 PM

deserts, warm 177
adaptations are either avoidance (maintaining moist conditions inside their tissues) or tolerance (grow-ing despite dry internal conditions), and include the following.
Reduced leaf area. Desert shrubs generally have small leaves. Succulents may have no leaves at all (as in desert cacti) or leaves only during the rainy season (as in desert spurges). Small leaves allow more of the heat load to diffuse into the air, thereby requiring less transpiration of water vapor to cool them. Succulent stems store the heat in the water inside their tissues.
Storage. Many warm desert plants are succu-lents and store water in their tissues, which are usually protected by spines and thorns from thirsty animals. examples include the cacti of the north and South american deserts and the spurges of the african deserts (see table on page 178 and 179). they often have wide, shallow root systems that quickly absorb rain. their succulence also allows them to absorb carbon dioxide from the air during the cool nights and store it in the form of acid; they can then produce food by photosynthesis during the hot daytime without opening their pores.
Deep roots. Many warm desert shrubs have deep roots that reach the water table. While these roots may have essentially unlimited water, they cannot provide much of it to the aboveground part of the shrub, because such roots, expensive to grow, are not numerous, and they must transport the water a long distance upward. Some mesquites have been found to have roots that are more than 100 feet (30 m) deep. Many shrubs, despite limited access to groundwater, produce leaves and bloom in the spring. the ocotillo bush produces leaves and bright red flowers in the spring, while it looks like dead sticks the rest of the year.
Some shrubs grow more quickly than others. near the bottom of Death Valley, the shrubs exploit soil water sources that do not last for many years. Tidestromia oblongifolia shrubs live only a few years; they produce seeds and die when the water runs out. in contrast, Atriplex hymenelytra shrubs grow slowly when water is available and stop grow-ing when it is not.
Ephemeral plants. Many desert wildflowers ger-minate during the winter rainy season, then bloom in the spring when the rains have ended. the warm des-ert spring may produce a carpet of wildflowers, such as Mexican poppies in the american southwest. the entire plant of Mimulus bigelovii may be only about two inches (a few cm) in height, almost a third of which consists of flowers (see figure). another exam-ple is the paper daisy (Leucochrysum fitzgibbonii) of australia. By summer, the wildflowers have produced seeds and begun to die. ephemeral plants grow very quickly during the brief wet season. Some desert
plants, such as the desert nardoo fern of australia (Marsilea drummondii), grow only when temporary ponds are wet; only its spores survive the dry season.
Condensation of fog. Some coastal deserts, such as in South america and africa, receive little, if any, rainfall but frequently receive fog and mist from the oceans. the fog droplets collect on the stems and leaves of the plants already present and drip into the soil.
Some desert plants grow well in moist micro-habitats, such as the following.
● Some warm desert plants may create moist microhabitats for other plants. Phacelia campanularia is a southwest american des-ert wildflower that often grows beneath the creosote shrubs of the Mojave Desert. Many cactus seedlings will not survive past ger-mination unless they are in the moist shade cast by shrubs such as acacias. the cactus seedlings eventually grow up through the “nurse shrub” (see figure on page 180).
● Dry creek beds (called arroyos in southwest-ern north america and wadi in the Middle east) are also a relatively moist microhabi-tat. after the rainy season, arroyos are the
This ephemeral wildflower (the monkeyflower Mimu-lus bigelovii) grows only a few centimeters tall and completes its entire life cycle within the few weeks of wet weather in the desert spring. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 177 11/17/11 1:12 PM
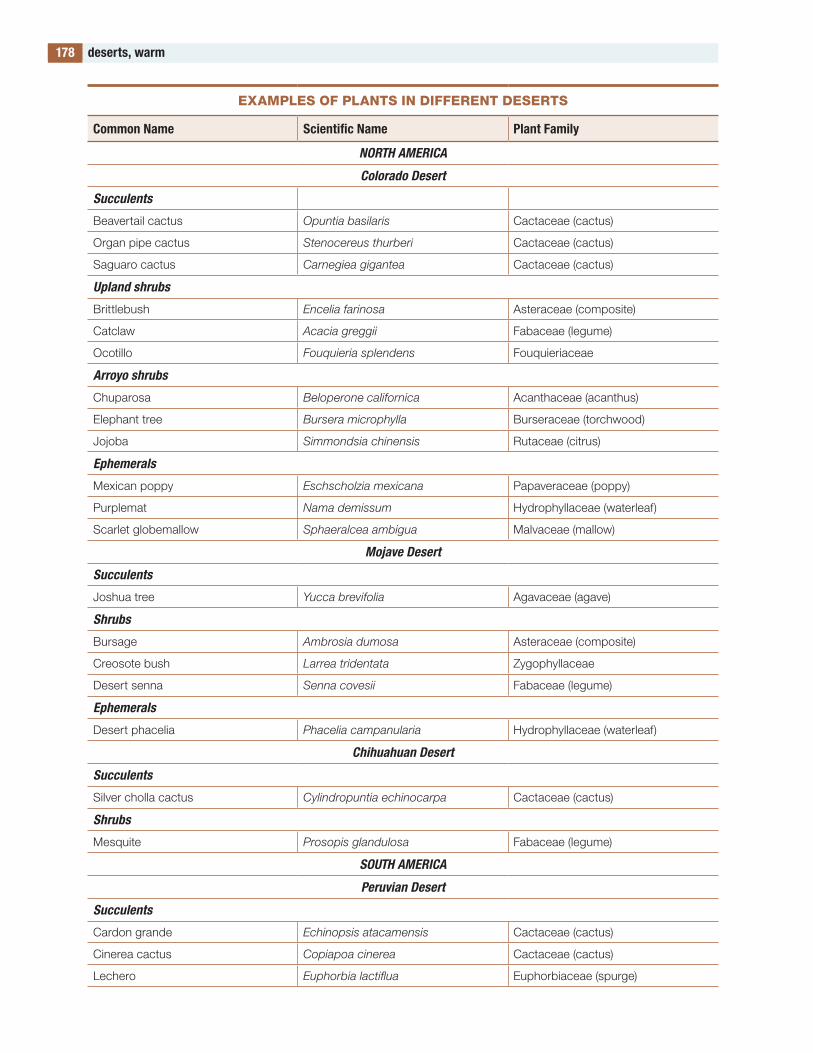
first entry178 deserts, warm 178
examples of plants in different deserts
Common Name Scientific Name plant family
NORTH AMERICA
Colorado Desert
Succulents
Beavertail cactus Opuntia basilaris Cactaceae (cactus)
Organ pipe cactus Stenocereus thurberi Cactaceae (cactus)
Saguaro cactus Carnegiea gigantea Cactaceae (cactus)
Upland shrubs
Brittlebush Encelia farinosa Asteraceae (composite)
Catclaw Acacia greggii Fabaceae (legume)
Ocotillo Fouquieria splendens Fouquieriaceae
Arroyo shrubs
Chuparosa Beloperone californica Acanthaceae (acanthus)
Elephant tree Bursera microphylla Burseraceae (torchwood)
Jojoba Simmondsia chinensis Rutaceae (citrus)
Ephemerals
Mexican poppy Eschscholzia mexicana Papaveraceae (poppy)
Purplemat Nama demissum Hydrophyllaceae (waterleaf)
Scarlet globemallow Sphaeralcea ambigua Malvaceae (mallow)
Mojave Desert
Succulents
Joshua tree Yucca brevifolia Agavaceae (agave)
Shrubs
Bursage Ambrosia dumosa Asteraceae (composite)
Creosote bush Larrea tridentata Zygophyllaceae
Desert senna Senna covesii Fabaceae (legume)
Ephemerals
Desert phacelia Phacelia campanularia Hydrophyllaceae (waterleaf)
Chihuahuan Desert
Succulents
Silver cholla cactus Cylindropuntia echinocarpa Cactaceae (cactus)
Shrubs
Mesquite Prosopis glandulosa Fabaceae (legume)
SOUTH AMERICA
Peruvian Desert
Succulents
Cardon grande Echinopsis atacamensis Cactaceae (cactus)
Cinerea cactus Copiapoa cinerea Cactaceae (cactus)
Lechero Euphorbia lactiflua Euphorbiaceae (spurge)
xviii+598_Biodiversity_F.indd 178 11/17/11 1:12 PM
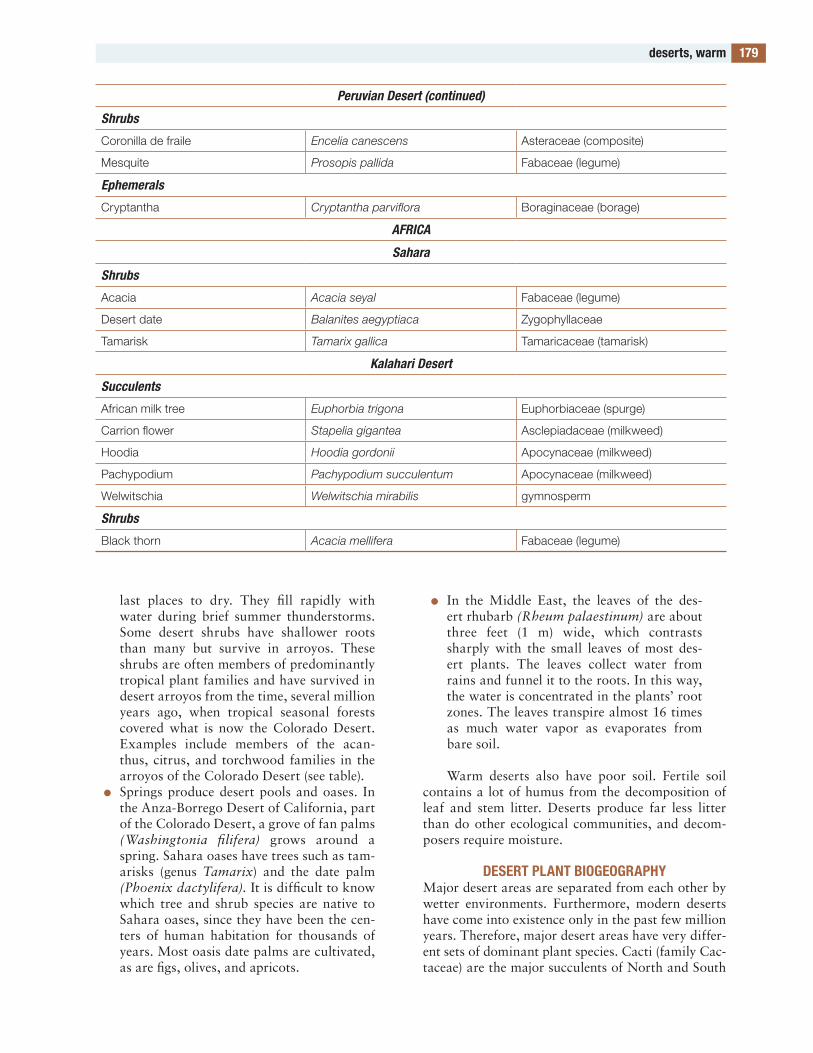
deserts, warm 179
last places to dry. they fill rapidly with water during brief summer thunderstorms. Some desert shrubs have shallower roots than many but survive in arroyos. these shrubs are often members of predominantly tropical plant families and have survived in desert arroyos from the time, several million years ago, when tropical seasonal forests covered what is now the colorado Desert. examples include members of the acan-thus, citrus, and torchwood families in the arroyos of the colorado Desert (see table).
● Springs produce desert pools and oases. in the anza-Borrego Desert of california, part of the colorado Desert, a grove of fan palms (Washingtonia filifera) grows around a spring. Sahara oases have trees such as tam-arisks (genus Tamarix) and the date palm (Phoenix dactylifera). it is difficult to know which tree and shrub species are native to Sahara oases, since they have been the cen-ters of human habitation for thousands of years. Most oasis date palms are cultivated, as are figs, olives, and apricots.
● in the Middle east, the leaves of the des-ert rhubarb (Rheum palaestinum) are about three feet (1 m) wide, which contrasts sharply with the small leaves of most des-ert plants. the leaves collect water from rains and funnel it to the roots. in this way, the water is concentrated in the plants’ root zones. the leaves transpire almost 16 times as much water vapor as evaporates from bare soil.
Warm deserts also have poor soil. Fertile soil contains a lot of humus from the decomposition of leaf and stem litter. Deserts produce far less litter than do other ecological communities, and decom-posers require moisture.
dESErT plaNT BIoGEoGraphyMajor desert areas are separated from each other by wetter environments. Furthermore, modern deserts have come into existence only in the past few million years. therefore, major desert areas have very differ-ent sets of dominant plant species. cacti (family cac-taceae) are the major succulents of north and South
Peruvian Desert (continued)
Shrubs
Coronilla de fraile Encelia canescens Asteraceae (composite)
Mesquite Prosopis pallida Fabaceae (legume)
Ephemerals
Cryptantha Cryptantha parviflora Boraginaceae (borage)
AFRICA
Sahara
Shrubs
Acacia Acacia seyal Fabaceae (legume)
Desert date Balanites aegyptiaca Zygophyllaceae
Tamarisk Tamarix gallica Tamaricaceae (tamarisk)
Kalahari Desert
Succulents
African milk tree Euphorbia trigona Euphorbiaceae (spurge)
Carrion flower Stapelia gigantea Asclepiadaceae (milkweed)
Hoodia Hoodia gordonii Apocynaceae (milkweed)
Pachypodium Pachypodium succulentum Apocynaceae (milkweed)
Welwitschia Welwitschia mirabilis gymnosperm
Shrubs
Black thorn Acacia mellifera Fabaceae (legume)
xviii+598_Biodiversity_F.indd 179 11/17/11 1:12 PM

first entry180 deserts, warm 180
american deserts, while spurges and milkweeds dominate the succulent flora of africa (see table on pages 178 and 179). these plants look enough like cacti that many horticulturalists call them cacti. Some desert shrubs are found nearly worldwide, for example, the acacia and mesquite genera.
the fluctuation between wet and dry conditions is ecologically similar to a disturbance, in which a flush of resources follows a long period without them. this explains the overlap between weed and desert plant families. caltrop (Tribulus terrestris and T. cistoides, also known as puncture vine and devil’s thorn) is a desert plant from South africa. in other parts of the world, such as the warmer regions of north america, it is a weed. it was naturalized in recent centuries on the Galápagos islands. it is in the family Zygophyllaceae, which is also known as desert shrubs in north america, africa, and the Middle east. the composite family (asteraceae) is also famous for its weeds. Many grasses are also weeds, but a few grasses live in (and stabilize) desert
sand dunes. an example is the australian sandhill cane Zygochloa paradoxa, which stores water in its underground stems.
Perhaps the strangest desert plant is Welwitschia mirabilis, which grows in the southern african des-ert. it is the only species within its gymnosperm evo-lutionary lineage. it is the only gymnosperm that has (independently) evolved the kind of photosynthesis that is common in flowering plant succulents. the stem remains underground. long leaves keep grow-ing as their tips are destroyed by desert conditions. During occasional periods of moisture, reproductive structures emerge from between the two leaves.
Warm deserts intergrade into adjacent ecologi-cal communities. the chihuahuan Desert contains grasses; no clear line separates it from arid grass-lands. the Mojave Desert intergrades with cool des-erts to the north. the deserts of southwest africa intergrade into the shrublands of South africa.
dESErT aNImal adapTaTIoNSDesert animals have adaptations that allow them to avoid or tolerate heat and drought.
Avoidance. all desert animals avoid drought to the extent that is possible. Some desert animals are active only during moist seasons or in areas of per-manent moisture. examples include the following.
● the Death Valley desert pupfish (Cyprin-odon salinus) lives in the saline conditions at the bottom of Death Valley. a spring maintains the moisture all year. Because desert springs are isolated, different species of pupfish have evolved in desert pools in other areas of the american Southwest. it avoids drought but must physiologically tol-erate hot water and salt.
● the australian shield shrimp Triops aus-traliensis lives and breeds in desert saline pools.
● adult desert frogs emerge from under-ground during the wet season, mate, and lay eggs. the tadpoles mature, and the adult frogs dig tunnels in the moist soil. Once underground, the australian water-holding frog (Cyclorana platycephalus) sheds a layer of skin that forms a membrane that traps much of the moisture as the soil dries out. it can remain underground for several years if rains fail to come. it thus avoids drought by becoming dormant during the dry season.
Tolerance. Other desert animals tolerate drought. their major way of doing this is to be active primarily at night. Most of them also remain in wet microhabitats whenever possible. this is where most
Seedlings of many cactus species, such as the cholla, organ pipe (Stenocereus thurberi), and saguaro (Car-negeia gigantea), grow in the shade of shrubs such as acacias and continue to grow after the acacias have died. (Organ Pipe Cactus National Monument; Stanley A. Rice)
xviii+598_Biodiversity_F.indd 180 11/17/11 1:12 PM

deserts, warm 181
prey (such as lizards and rodents) and most preda-tors (such as snakes and roadrunners) live. Many desert insects, such as ants and termites, live under-ground. Scorpions, abundant in deserts worldwide, hunt at night. Other examples include the following.
● the australian whistling spider (Selenocos-mia stirlingi) spends the day at the end of a three-foot-long (1 m-long) burrow and at night waits behind the silk door of the bur-row to ambush arthropods that come near.
● australian sand grasshoppers (Urnisa gut-tulosa) bury themselves in sand, with only their eyes and antennae exposed, during the day and emerge to eat plant tissues at night (only during the wet season).
● Pit viper snakes (crotalinae) have special sen-sory organs that detect infrared light, which all warm objects emit. this allows them to “see the heat” of mammalian prey species.
Some desert mammals have adaptations that allow them to dissipate body heat without the loss of water. the ears of american jackrabbits (such as Lepus californicus) have large exposed areas that allow body heat to radiate into the air. all desert animals have ways to avoid water loss. Snakes and lizards are abundant and diverse in the deserts of the world. they all have scales that prevent evaporation, and they excrete a slurry of uric acid rather than watery urine. Many desert animals also have inge-nious ways of obtaining water, as in the following examples.
● Some desert animals get all their water from their food. the ayrean grasswren (Amytor-nis goyderi), a desert bird of australia, gets all its water from the seeds and arthropods that it eats; it also excretes less water than most other birds. in many cases, desert ani-mals that eat seeds gather them just before dawn, when there is the greatest chance that moisture will have condensed on them from the air.
● Kangaroo rats (genus Dipodomys) of the american Southwest and the australian spinifex hopping-mouse (Notomys alexis) not only get all their water from the food that they eat, but rely heavily on metabolic water. this is water that the mitochondria in their cells produce in the process of releasing energy from the metabolism of sugar.
● Some beetles that live in the deserts of southern africa stand on sand dunes when fog moves in from the ocean. they drink the water that condenses on their bodies.
among the most amazing desert animal adap-tations are those of dromedary camels (Camelus dromedarius) of the Sahara and Middle east deserts. they include the following:
● Dromedary camels store most of their body fat in a single hump rather than distrib-uted throughout the body under the skin. this reduces the insulation of the skin and helps body heat dissipate into the air. camel humps do not store water.
● like other desert mammals, camels make use of metabolic water, of which the metab-olism of their fat produces a great deal.
● camel blood cells are oval-shaped, which helps to prevent them from sticking together when the blood thickens due to dehydration. their bodies can withstand up to 25 percent fluid loss from sweating, in contrast to most mammals, in which heart failure begins at 15 percent fluid loss. the cells are remark-ably resilient to changes in blood salinity, which occurs when camels drink a large amount of water (up to 40 gallons [150 l] at a time) after a long thirst.
● camels can tolerate a body temperature up to 106°F (41°c).
● camels have thick coats of hair. this inter-feres with the loss of body heat but also shields camels’ bodies from the intense heat of the desert. their long legs keep their bod-ies far from the hot desert soil.
● camels lose very little water through urine (which can be as thick as syrup) or feces.
Deserts have low biodiversity compared to most other ecological communities, but plants and animals have evolved numerous adaptations inde-pendently in deserts around the world to avoid or tolerate dry conditions. Few deserts are threatened with human development, which requires water. Solar power plants have been constructed in some desert areas, particularly the Mojave Desert in cali-fornia, and many more are planned. there are few endemic species in the Mojave Desert, and it would require the development of many solar power plants before this desert were harmed. Desert oases may have endemic species and require habitat protection. Some deserts, such as the Sonoran Desert, are par-ticularly rich in species and require particular atten-tion, especially since much of the Sonoran Desert is near tucson and Phoenix. conservation is particu-larly important for bats and other pollinators of the Sonoran Desert cacti. although most pharmaceuti-cals come from rain forest plants, a few come from desert plants. an extract of the african succulent
xviii+598_Biodiversity_F.indd 181 11/17/11 1:12 PM

first entry182 deuterostomes 182
Hoodia gordonii has been used for centuries by local tribes to reduce the symptoms of hunger and is now marketed as a natural supplement for weight loss. increasing drought in connection with global warm-ing may cause some desert areas, whose plants and animals are already on the brink of survival, to lose many of their species.
See also biogeography; communities, eco-logical; convergence; deserts, cool; distur-bances, ecological; grasslands and savannas; shrublands.
Further readingatchley, Michael c., et al. “arroyo water storage and soil
nutrients and their effects on gas-exchange of shrub species in the northern chihuahuan Desert.” Journal of Arid Environments 43 (1999): 21–33.
Brown, James H., and c. Robert Feldmeth. “evolution in constant and fluctuating environments: thermal toler-ances of desert pupfish (Cyprinodon).” Evolution 25 (1971): 390–398.
Gulmon, Sally l., and Harold a. Mooney. “Spatial and temporal relationships between two desert shrubs, Atriplex hymenelytra and Tidestromia oblongifolia in Death Valley, california.” Journal of Ecology 65 (1977): 831–838.
Holzapfel, claus, and Bruce e. Mahall. “Bidirectional facilitation and interference between shrubs and annuals in the Mojave Desert.” Ecology 80 (1999): 1,747–1,761.
Kaufman, Donald W., and Glennis a. Kaufman. “effect of moonlight on activity and microhabitat use by Ord’s kangaroo rat (Dipodomys ordii).” Journal of Mam-malogy 63 (1982): 309–312.
Mahall, Bruce e., and Ragan M. callaway. “Root com-munication mechanisms and intracommunity distri-butions of two Mojave Desert shrubs.” Ecology 73 (1992): 2,145–2,151.
Mcauliffe, Joseph R. “Sahuaro-nurse tree associations in the Sonoran Desert: competitive effects of sahuaros.” Oecologia 64 (1984): 319–321.
nagy, K. a. “Seasonal water, energy and food use by free-living, arid-habitat mammals.” Australian Journal of Zoology 42 (1994): 55–63.
Pockman, William t., and John S. Sperry. “Vulnerability to xylem cavitation and the distribution of Sonoran Desert vegetation.” American Journal of Botany 87 (2000): 1,287–1,299.
Polis, Gary a., and Sharon J. Mccormick. “intraguild predation and competition among desert scorpions.” Ecology 68 (1987): 332–343.
Reichman, O. J., and David Oberstein. “Selection of seed distribution types by Dipodomys merriami and Perog-nathus amplus.” Ecology 58 (1977): 636–643.
Rundel, Philip W., James ehleringer, Harold a. Mooney, and Sally l. Gulmon. “Patterns of drought response in
leaf-succulent shrubs of the coastal atacama Desert in northern chile.” Oecologia 46 (1980): 196–200.
tucker, compton J., and Sharon e. nicholson. “Variations in the size of the Sahara Desert from 1980 to 1997.” Ambio 28 (1999): 587–591.
Vasek, Frank c. “creosote bush: long-lived clones in the Mojave Desert.” American Journal of Botany 67 (1980): 246–255.
Ward, David. The Biology of Deserts. Oxford: Oxford Uni-versity Press, 2009.
Winter, Klaus, and Michael J. Schramm. “analysis of stoma-tal and nonstomatal components in the environmental control of cO2 exchange in leaves of Welwitschia mirabilis.” Plant Physiology 82 (1986): 173–178.
deuterostomes See animals.
devonian period, biodiversity in the Devonian period (410 to 360 million years ago) was the fourth period of the Paleozoic era. the first significant advances of life onto land had occurred during the preceding Silurian period. During the Devonian period, plants filled the wet areas of earth, making large areas of earth’s surface green for the first time.
Climate. the climate in the equatorial regions was warm and wet. Portions of the southern conti-nent, over the South Pole, were cold. atmospheric oxygen levels were lower and carbon dioxide levels higher than at present (see table on page 183).
Continents. During the Devonian, there were three major continental areas. What is now north america and europe formed a single landmass near the equator, much of it covered by shallow oceans. a portion of what is now Siberia lay to the north. Gondwanaland, a continent composed of what is now South america, africa, antarctica, india, and australia, formed a large mass in the Southern Hemisphere.
Marine life. Marine life, especially fishes, diver-sified during the Devonian (see figure on page 184).
● Invertebrates. the invertebrate groups that dominated the Silurian period continued to proliferate during the Devonian. Bra-chiopods continued to be more common than superficially similar bivalves. among the cephalopod mollusks, a new group appeared—the ammonites, with coiled shells. trilobites were much less diverse than in previous periods but remained common.
● Vertebrates. Many kinds of fishes evolved during the Devonian period. So great was their diversity that the Devonian period has
xviii+598_Biodiversity_F.indd 182 11/17/11 1:12 PM

devonian period, biodiversity in 183
been called the age of fishes, even though there were also many species of plants and animals on land. armored jawless fishes evolved into some large forms such as the five-foot (1.5-m) Psammolepis, then declined toward extinction, mainly due to compe-tition with fishes that had jaws and that could therefore forage more efficiently. One lineage of jawless fishes without armor sur-vived as the ancestors of modern lampreys and hagfishes. the jawed fishes included the heavily armored placoderms (“plate-skins”), a group now extinct. Some, such as Dunkleosteus, were 33 feet (10 m) in length. the placoderms were the dominant group of fishes in the sea at this time. although fossilized shark scales have been found in Silurian deposits, the first full shark fossils date from the Devonian period, at which time shark species proliferated. the most abundant group of fishes today, the ray-finned fishes, began to proliferate during the Devonian period.
Continental life. at the beginning of the Devo-nian period, there were only small plants (about 3 feet [1 m] in height) and a few arthropods in terres-trial wetlands. By the end of the Devonian period, the first trees and terrestrial vertebrates had evolved.
● Plants. early Devonian Rhynia was not much bigger than the late Silurian Cook-sonia. these plants had xylem cells, which carry water from the ground up to the tips of the stems. Xylem allowed the evolution-ary descendants of these plants to grow to the size of trees. By the end of the Devo-nian, some trees were about 30 feet (10 m) tall. their descendants became much larger trees during the subsequent carboniferous period. Silurian plants evolved into Devo-nian clubmosses, horsetails, and ferns, which reproduced by spores rather than by seeds. Silurian plants had only simple branching stems, but during the Devonian, the first leaves evolved. Some of the branches were united by layers of green tissue, which became the veins and blades of the leaves. Before the end of the Devonian, progymno-sperms such as Archaeopteris had primitive forms of seeds.
● Vertebrates. the lobe-finned fishes, which had fingerlike bony reinforcements in their fins, evolved in continental freshwater dur-ing the Devonian period. One group of lobe-finned fishes was the lungfishes. a few
multiCellular biodiversity in the devonian period
Conditions
Atmospheric oxygen 75 percent of modern level
Atmospheric carbon dioxide 6.3 times modern level
Global temperature 11°F (6°C) higher than today
Sea level 260 feet (80 m) higher than today
Approximate number of families*
Continental 70
Marine 450
Total 520
Examples of marine animals
Angustidontis sp. crustacean arthropod
Cladoselache sp. shark
Dipleura dekayi trilobite
Dunkleosteus sp. armored jawed fish
Hydroblastus hendyi echinoderm
Lepidocoleus hohensteini annelid
Cheirolepis sp. ray-finned fish
Orthonychia protei gastropod mollusk
Psammolepis sp. armored jawless fish
Sporadoceras orbiculare ammonite mollusk
Examples of continental animals
Acanthostega gunneri fishlike amphibian
Attercopus fimbriungus spider relative
Dipnorhynchus sp. lungfish
Eusthenopteron foordi amphibianlike fish
Ichthyostega stensoi fishlike amphibian
Orsadesmus rubecollus millipede
Tiktaalik roseae fishlike amphibian
Examples of land plants
Archaeopteris obtusa seed-bearing tree
Gillespiea randolphensis fernlike plant
Protolepidodendron sp. tree clubmoss
Psilophyton sp. trimerophyte plant
Rhacophyton sp. fern
Zosterophyllum rhenanum zosterophyllophyte plant
*Based on Benton
xviii+598_Biodiversity_F.indd 183 11/17/11 1:12 PM

fi rst entry184 dicots 184
lungfi shes still live in tropical rivers today. their lungs allow them to survive long peri-ods in mud when ponds become seasonally dry. From one of the lineages of lobe-fi nned fi shes, the fi rst tetrapod amphibians evolved during the Devonian. the crucial fi rst steps, from lobe-fi nned fi shes to amphibianlike fi shes (such as Eusthenopteron) to fi sh-like amphibians (such as Ichthyostega) to amphibians occurred during this geological period. the recently discovered Tiktaalik roseae was almost exactly in the middle of the fi sh-to-amphibian transition.
living among the small plants in the early Devo-nian mud were the fi rst land animals, relatives of modern arthropods such as scorpions, mites, and spiders. Shortly thereafter, the ancestors of insects, centipedes, and millipedes were living among the small plants of early Devonian wetlands.
Extinctions. One of the mass extinction events in earth history occurred at the end of the Devonian, at which time an estimated 22 percent of families, representing 83 percent of species, became extinct. the causes of this extinction event are unclear.
See also amphibians; fishes; mass extinctions.
Further readingalgeo, t. J., et al. “effects of the Middle to late Devonian
spread of vascular land plants on weathering regimes, marine biota, and global climate.” in Plants Invade the Land: Evolutionary and Environmental Approaches,edited by Patricia G. Gensel and D. edwards, 213–236. new york: columbia University Press, 2001.
Benton, Michael. “Diversifi cation and extinction in the his-tory of life.” Science 268 (1995): 52–58.
Bond, David, et al. “extent and duration of marine anoxia during the Frasnian-Famennian (late Devonian) mass extinction in Poland, Germany, austria, and France.” Geological Magazine 141 (2004): 173–193.
Boyce, c. Kevin, et al. “chemical evi-dence for cell wall lignifi cation and the evolution of tracheids in early Devo-nian plants.” International Journal of Plant Sciences 164 (2003): 691–703.clack, Jennifer a. “Devonian climate change, breathing, and the origin of the tetrapod stem group.” Integrative and Comparative Biology 47 (2007): 510–523.Murphy, Dennis c. “Devonian times.” available online. URl: http://www.devoniantimes.org/index.html. accessed September 17, 2009.Rode, alycia l., and Bruse S. lieber-man. “integrating evolution and bio-
geography: a case involving Devonian crustaceans.” Journal of Paleontology 79 (2005): 267.
Rolfe, W. D. ian, and Jerzy Dzik. “Angustidontus, a late Devonian pelagic predatory crustacean.” Transactions of the Royal Society of Edinburgh (Earth Sciences) 97 (2006): 75–96.
Roth-nebelsick, anita, and Wilfried Konrad. “assimila-tion and transpiration capabilities of rhyniophytic plants from the lower Devonian and their implica-tions for paleoatmospheric cO2 concentration.” Pal-aeogeography, Palaeoclimatology, and Palaeoecology202 (2003): 123–176.
dicots Dicots are the most abundant fl owering plants. there are at least 170,000 species of dicots, ranging from tiny herbaceous plants to trees. along with the conifers, the fl owering plants have trans-formed the landscape of earth into a living surface of green. in addition, fl owering plants have under-gone coevolution with animals that pollinate their fl owers and disperse their seeds. as a result, the fl owering plants, especially the dicots, have fi lled the world with an astounding diversity of form, color, and scent.
Flowering plants have fl owers and fruits. Flow-ers may contain:
● Sepals: leafl ike protective structures ● Petals: often colorful structures that attract
pollinators ● Stamens: male structures that produce
pollen ● Pistils: female structures that produce seeds
the term dicot refers to the embryo of the plant. When it emerges from the seed, the embryo has two cotyledons, or embryonic leaves. Most dicots also have veins that form cores in the middle of the roots, rings in the young stems, and networks in the
Placoderms were large armored fi shes that lived in the oceans during the Devonian period.
xviii+598_Biodiversity_F.indd 184 11/17/11 1:12 PM

dicots 185
leaves, in contrast to the arrangement of veins in the monocots, the embryos of which have only one cotyledon.
taxonomists (scientists who study evolution-ary relationships among organisms) have tradition-ally divided the flowering plants into two groups: the monocots and the dicots. Recent evolution-ary research, however, has shown that the dicots actually represent several distinct lineages. this makes them a paraphyletic cladistic group. all of these dicot lineages except one are considered to be “primitive dicots” and are discussed in the entry on flowering plants. the remaining dicot lineage, which contains the vast majority of dicots, is called the eudicots, and is the subject of the remainder of this entry.
prImITIvE aNd CorE EudICoTSthere are several relatively primitive lineages within the eudicots. Here are three of the important fami-lies of primitive eudicots that are important in north america and characteristics by which to recognize them.
the poppies (Papaveraceae) are herbaceous plants with milky sap. this sap helps protect them from animals and often contains potent chemicals such as morphine. Poppy flowers have numerous sta-mens (the male parts), all separate from one another.
the buttercups (Ranunculaceae) are also herba-ceous plants whose flowers have numerous separate stamens. Relatives of buttercups include larkspurs and columbines, the petals of which have long spurs. Only specialized pollinators can obtain nectar from inside these spurs.
the sycamores (Platanaceae) are large, fast-growing trees found mostly near water. their large leaves and striking white bark make them easily rec-ognizable. the fruits form fluffy parachutes and are condensed into spheres that hang from the branches during winter. in spring, the wind blows the seeds with their fruit parachutes away. Platanaceae is no longer considered a well-defined family.
Most other eudicots are part of an advanced lineage called the core eudicots. taxonomists have identified several major lineages within the core eudicots. two of these lineages are the rosids and the asterids.
roSIdSin most rosids, the components of the flowers are separate, just as in the primitive dicots. Here are some families of rosids that are important in north america and characteristics by which to recognize them.
the oaks and beeches (Fagaceae) are major for-est trees. they grow slowly and dominate old for-
ests. they produce tiny male flowers on dangling stalks known as catkins. the female flowers pro-duce a single large nut surrounded by tiny leaflike bracts (see figure). in oaks, the nut is an acorn, and the bracts form a cap. the leaves of oaks and beeches consist of a single blade. north american oaks (genus Quercus) consist of two groups. the red and black oaks have leaves with spines, while white oaks have leaves with smooth lobes. eastern and western north america have entirely separate species of oaks. in eastern north america, all the oaks are deciduous except the live oak, Quercus vir-giniana, which is evergreen. in western north amer-ica, most of the oaks are evergreen. Oaks dominate several kinds of temperate deciduous forests. For example, white oaks (Quercus alba) grow in moist upland forests, post oaks (Quercus stellata) grow in dry forests, and water oaks (Quercus nigra) grow near water. europe and asia have a set of oak spe-cies entirely different from those of north america. there are few species of beech, and one major spe-cies of beech (Fagus grandifolia) dominates north
Water oaks (Quercus nigra) produce separate male and female flowers, but the flowers are on the same plant. The male flowers hang in catkins (green arrow), while the female flowers are acorns (red arrow), here shown at the pollen-receptive stage. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 185 11/17/11 1:12 PM

first entry186 dicots 186
american beech forests; in contrast, north america has 90 species of oaks. the american chestnut (Cas-tanea americana) used to dominate large areas of the deciduous forest of eastern north america, but they have (except for resprouted saplings) been destroyed by an accidentally introduced fungus.
the walnuts and hickories (Juglandaceae) are also important forest trees, although they are usu-ally minor components mixed into old forests domi-nated by oaks or beeches. Walnuts (genus Juglans) and hickories (genus Carya) also produce catkins of male flowers. the female flowers produce a single large nut surrounded by a single husk (walnuts) or a husk divided into four parts (hickories). the large leaves consist of several to many small leaflets.
the birches and alders (Betulaceae) are small trees that often grow near water. they produce cat-kins of male flowers and tight clusters of female flowers that sometimes look like cones. the leaves consist of a single blade. By vigorously resprouting, alders (genus Alnus) can recover quickly from flood damage. alders have bacteria in their roots that manufacture nitrogen fertilizer. therefore, although they are far less abundant than oaks (there are only eight species of alders in north america), they play an important ecological role. Birches seldom domi-nate whole forests except in the far north. the sea-side alder, Alnus maritima, is an interesting example of a disjunct species, found in only a few locations very distant from one another.
the rose family (Rosaceae) has flowers with five petals and many stamens that grow from the edge of a central disc. although horticultural varieties of roses (genus Rosa) may have many petals, wild roses have only five. the flowers are open, easily accessible by many kinds of pollinators. While the flowers in this family look very much alike, there is great diversity in their growth patterns (small wild-flowers to large trees) and fruits, including most of the commercially valuable fruits. in peaches, plums, and cherries (Prunus), a single seed develops inside the fruit. in blackberries (Rubus), many small ber-ries cluster together. in apples (Malus) and pears (Pyrus), the stem at the base of the flower grows up and surrounds the seeds. in strawberries (Fragaria), the stem at the base of the flower grows up and pushes the seeds outward. the strawberry is not actually a fruit; the fruits are the black “seeds” on the outside of the expanded stem. although com-mercially valuable, the rose family plays a minor role in natural landscapes compared to oaks: short-lived black cherry trees (Prunus serotina), a few shrubs, and many species of wildflowers.
the mulberry family (Moraceae) includes mul-berries and figs. Male trees produce catkins of male flowers. Female trees produce clusters of tiny fruits.
in mulberries (genus Morus), the tiny fruits are on the outside of the cluster. in figs (Ficus), the tiny fruits are on the inside of the cluster; a fig is an inside-out mulberry. Mulberries and figs are very common in tropical rain forests. the north ameri-can bois d’arc (Maclura pomifera) produces a fruit that was eaten by mammals that are now extinct. Mulberries, like poppies, produce a defensive milky sap. the leaves consist of a single blade.
the legume family (Fabaceae) produces beanlike fruits in which the seeds line up in single file inside a capsule. in most species, the flowers have a boatlike structure, with a banner that resembles a sail and a keel that encloses the stamens and pistil. in some legumes, such as mimosas, the numerous, long, and colorful stamens are the most noticeable feature of the flowers. the leaves often consist of three to several dozen small leaflets. legumes range in size from small herbaceous plants to large trees. like alders, legumes have bacteria in their roots that manufacture nitrogen fertilizer. the bacteria are different from those in alders; fertilizer production in alder and legume roots is therefore an example of evolutionary convergence. north american forests have a few leguminous trees, such as the black locust (Robinia pseudoacacia) and honey locust (Gleditsia triacanthos), and grasslands have many leguminous wildflowers, such as the poi-sonous locoweeds (genus Astragalus). However, most legumes grow in tropical rain forests and tropical sea-sonal forests. leguminous acacias are very important components of tropical savannas. Many acacias have evolved close cooperation with ants.
the willow family (Salicaceae) consists of wil-low, cottonwood, poplar, and aspen trees. Male trees produce catkins of male flowers, and female trees produce catkins of female flowers (see figures on page 187). the leaves consist of a single blade. these trees usually grow fast in areas that have been recently disturbed, for example, by flooding. Because of its fast growth, the black poplar (Populus trichocarpa) was one of the first plant species for which biotechnologists decoded the entire genome.
Members of the mustard family (Brassicaceae) are often called “crucifers” because their flowers have four petals in the shape of a cross. almost all of them are wildflowers, often weedy components of recently disturbed fields. Many important veg-etables, such as radishes, are in the mustard family; cabbage, cauliflower, broccoli, and brussels sprouts were all bred from a single species of wild plant, Brassica oleracea. a species that lives in europe and asia, Arabidopsis thaliana, goes through its entire life cycle in a few weeks and has therefore been of great interest to genetic researchers; it was one of the first plant species for which biotechnologists decoded the entire genome.
xviii+598_Biodiversity_F.indd 186 11/17/11 1:12 PM

dicots 187
the sumac family (Anacardiaceae) consists of bushes such as the north american sumac, tropical trees such as cashew and mango, and vines such as poison ivy, poison oak, and poison sumac (genus Toxicodendron). Poison ivy is not actually poi-sonous to animal metabolism. the urushiol in its leaves provokes a strong allergic reaction in most people—but not in everyone, and not in mammals that eat its leaves (such as deer) and its berries (such as squirrels). Poison ivy and poison oak are easily recognizable because their leaves consist of three leaflets.
the maple family (Sapindaceae, formerly acera-ceae) includes maple trees and soapberry trees. Sugar maples (Acer saccharum) are major components of the temperate deciduous forest, while silver maples (Acer saccharinum) grow vigorously in floodplains. Maple trees and shrubs may be recognized by the fruits, in which a seed is enclosed in a single wing. this allows the fruit to whirl like a helicopter blade in the wind. Sugar maples are famous because their sap can be boiled into syrup and for their brilliant autumn colors in eastern north america.
the grape family (Vitaceae) consists of vines, including several species of wild grape (genus Vitis) and the Virginia creeper (Parthenocissus quinquefo-lia). the leaves consist of a single blade, and clusters of small flowers emerge from the opposite side of the stem from the leaf. Domesticated grapes (Vitis vinifera) come from europe, but rootstocks of wild american grapes from texas and Oklahoma (Vitis rotundifolia) were resistant to fungi that killed euro-pean grapes in the late 19th century. Grafting euro-pean grapes onto american rootstocks helped to save the French wine industry.
the spurge family (Euphorbiaceae) consists of herbs and shrubs that contain poisonous milky sap. Male flowers cluster around a female flower; in the genus Euphorbia, this cluster is reduced into a structure that looks like a single flower. in north america, most spurges are weedy herbaceous plants or small shrubs in dry areas. in african deserts, spurges have evolved a resemblance to the cacti of american deserts, another example of evolution-ary convergence. the most poisonous plant chemi-cal known, ricin, is produced by the castor bean
Cottonwoods (Populus deltoides) grow as separate male and female plants. Male cottonwood trees produce catkins of male flowers that release pollen (left), while female cottonwood trees produce catkins of female flow-ers that produce seeds (right). (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 187 11/17/11 1:12 PM

first entry188 dicots 188
bush Ricinus communis. in tropical forests, some are trees, such as the Brazilian rubber tree (Hevea brasiliensis), in which the milky sap can be dried to form rubber. Manioc (Manihot esculentum) is a small tree whose starchy roots are the tropical coun-terpart of potatoes; in some varieties, people must leach out the poisons before consuming the starch.
aSTErIdSin many asterids, parts of the flowers (particularly the petals) have fused together over evolutionary time, in some cases forming tubular structures that can be pollinated only by specialized animals. Here are some families of asterids that are important in north america and characteristics by which to rec-ognize them.
the nightshade family (Solanaceae) consists of mostly herbaceous plants noted for their poisonous leaves, stems, and fruits. Some members of this fam-ily, however, are important foods, such as potatoes (Solanum tuberosum) and tomatoes (Lycopersicon esculentum).
the mint family (Lamiaceae), most of which are herbaceous, is recognizable because its tubular flowers have a landing platform for pollinators, and its stems are square in cross-section. Many spices come from the leaves of these plants, including mint, oregano, basil, thyme, and sage. Some species are poisonous. in tropical forests, some mints are trees, such as teak (Tectona grandis), which is famous for wood used in shipbuilding.
the carrot family (Apiaceae), also herbaceous, is easily recognizable because its flowers are in umbrella-shaped clusters. carrots and celery are members of this family. Many spices come from the fruits of these plants, including cumin, coriander, dill, fennel, anise, and caraway. the seeds of celery and the leaves of coriander (known as cilantro) are also popular flavors. Some species are poisonous.
the milkweed family (Apocynaceae) consists mostly of herbaceous plants with defensive milky sap. the clustered flowers are easily recognizable because the five coronas, which look like an extra set of petals, form beaks that enclose each of the five stamens.
the composite family (Asteraceae) consists of almost 23,000 species of plants, most of them herba-ceous weeds that grow rapidly in recently disturbed areas or in grasslands. composites have flowers that form highly complex reproductive structures. these structures are called composite because each of them consists of several or many small flowers surrounded by small leaflike bracts. examples are the wild bal-samroot (see figure) and the sunflower. a sunflower appears to be a flower that consists of yellow petals radiating outward from a central disc of small struc-
tures. each of these “petals,” however, is a separate ray flower. each ray flower consists of five petals fused into a single petal surrounding a pistil. the central disc contains several to many disc flowers, each one of which consists of five petals fused into a single petal surrounding stamens and a pistil. a sunflower, therefore, is dozens of flowers united into a single structure. Most species of composites have both ray and disc flowers. Some, such as dandelions, consist only of raylike flowers and have no central disc. Some, such as thistles, consist only of disc flow-ers and have no rays.
oThEr CorE EudICoTSlineages of core eudicots other than rosids and aste-rids also contain important plants. One of these is the cactus family (Cactaceae), whose flowers (usu-ally large) have numerous petals and stamens (see
Composite dicots (family Asteraceae) have numerous small flowers (florets) compressed into a single flow-erlike structure called a head. Disk florets (red arrow) are tubular and erect; they produce both pollen and seeds. Unopened disk florets are in the center of the head. Each structure that looks like a petal is actually a ray floret (green arrow) that radiates from the center of the head. This species is Balsamorhiza sagittata from the Black Hills of South Dakota. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 188 11/17/11 1:12 PM

dicots 189
figure). Primitive cactus trees in central and South america have spines and succulent leaves. leafless cacti, in the deserts of north and South america have green, succulent stems but no leaves.
From simple ancestors that lived near the beginning of the cretaceous period, the flowering plants have diversified into hundreds of thousands of species, produced many specialized reproductive structures, and adapted to every habitat on earth, including coastal antarctica. Of the two flowering plant species that live in antarctica, one is a monocot and one is a dicot. Most flowering plants are dicots. no other group of multicellular organisms, except insects, have evolved such tremendous diversity. explorers continue to discover new species of dicots, usually, but not always, in tropical forests. Many species are driven to extinction, often before they have been studied by scientists, when humans destroy their habitats. this is a tremendous loss to the eco-logical communities of which they are the basis, to scientific research, and to the human economy.
See also essay: “Ghosts of Biodiversity Past;” adaptive radiation; biodiversity, resources for agriculture; biodiversity, resources
for medicine; cladistics; coevolution; cre-taceous period, biodiversity in; flowering plants (angiosperms); monocots; pollination.
Further readingabrams, Marc D. “adaptations and responses to drought
in Quercus species of north america.” Tree Physiol-ogy 7 (1990): 227–238.
anderson, edward F. The Cactus Family. Portland, Ore.: timber Press, 2001.
california Oak Mortality task Force. “california Oak Mortality task Force.” available online. URl: http://www.suddenoakdeath.org. accessed May 13, 2011.
callaway, Ragan M. “effect of shrubs on recruitment of Quercus douglasii and Quercus lobata in california.” Ecology 73 (1992): 2,118–2,128.
———, nalini M. nadkarni, and Bruce e. Mahall. “Facili-tation and interference of Quercus douglasii on under-story productivity in central california.” Ecology 72 (1991): 1,484–1,499.
carney, Shanna e., et al. “invasive argentine ants (Linepi-thema humile) do not replace native ants as seed dispersers of Dendromecon rigida (Papaveraceae) in california, USa.” Oecologia 135 (2003): 576–582.
DeBeaulieu, antoine leHardy. An Illustrated Guide to Maples. Portland, Ore.: timber Press, 2003.
ertter, Barbara. “Floristic surprises in north america north of Mexico.” Annals of the Missouri Botanical Gar-den 87 (2000): 81–109. available online. URl: http://ucjeps.berkeley.edu/floristic_surprises.html. accessed March 23, 2005.
Keator, Glenn. The Life of an Oak: An Intimate Portrait. Berkeley, calif.: Heyday Books, 1998.
logan, William Bryant. Oak: The Frame of Civilization. new york: norton, 1996.
Miller, Howard a., and Samuel lamb. Oaks of North America. Happy camp, calif.: naturegraph Publish-ers, 1984.
Mohan, Jacqueline e., et al. “Biomass and toxicity responses of poison ivy (Toxicodendron radicans) to elevated atmospheric cO2.” Proceedings of the National Acad-emy of Sciences USA 103 (2006): 9,086–9,089.
national Research council (U.S.). Tropical Legumes: Resources for the Future. Washington, D.c.: national Research council, 2002.
Panizzi, l., et al. “composition and antimicrobial proper-ties of essential oils of four Mediterranean lamiaceae.” Journal of Ethnopharmacology 39 (1993): 167–170.
Petrides, George a. A Field Guide to Trees and Shrubs. Boston: Houghton Mifflin, 1972.
Rizzo, D. M., et al. “Phytophthora ramorum as the cause of extensive mortality of Quercus spp. and Lithocarpus densiflorus in california.” Plant Disease 86 (2002): 205–214.
Russell, charles e., and Peter Felker. “the prickly-pears (Opuntia spp., cactaceae): a source of human and
Cactus flowers (such as this Opuntia basilaris from the Anza Borrego Desert of California) have numerous petals and numerous stamens. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 189 11/17/11 1:12 PM

first entry190 dinosaurs 190
animal food in semiarid regions.” Economic Botany 41 (1987): 433–445.
Russell, tony, and catherine cutler. The World Encyclope-dia of Trees. london: anness Publishing, 2003.
Strauss-Debenedetti, Sylvia, and Fakhri a. Bazzaz. “Plastic-ity and acclimation to light in tropical Moraceae of different sucessional positions.” Oecologia 87 (1991): 377–387.
tudge, colin. The Secret Life of Trees: How They Live and Why They Matter. new york: Penguin, 2006.
Wanvestraut, Robert H., et al. “competition for water in a pecan (Carya illinoensis K. Koch)-cotton (Gossy-pium hirsutum l.) alley cropping system in the south-ern United States.” Agroforestry Systems 60 (2004): 167–179.
dinosaurs the term dinosaur usually refers to reptiles, many of them large, that dominated earth during the Mesozoic era. they are probably the most famous examples of extinct biodiversity. the earliest dinosaurs lived during the triassic period. Dinosaurs diversified into numerous forms during the Juras-sic and cretaceous periods, perhaps because one of the mass extinctions, which occurred at the end of the triassic period, removed competition from other vertebrate groups. the dinosaurs themselves became extinct at the end of the cretaceous period.
Modern phylogeny classifies organisms based on evolutionary divergence. Since birds and mammals diverged from the group of vertebrates usually called reptiles, there is no such thing as reptiles unless one includes birds and mammals in the group. Dinosaurs had many characteristics of skin, teeth, and bones that would remind observers of the reptiles with which they are familiar. in fact, since birds evolved from dinosaurs, the cladistic approach would indi-cate that the dinosaurs still exist: they are birds. therefore, the dinosaurs that most people think of are more properly called nonavian dinosaurs. although dinosaurs are truly ancient, the therapsid (mammal-like) reptile lineage had diverged before the dinosaur lineage, and true mammals already existed by the time the first dinosaurs roamed earth.
the first dinosaurs to be studied were large, which is why Sir Richard Owen gave them the name dinosaurs (Greek for “terrible lizards”) in 1842. the first famous dinosaur discoveries came from the northern plains of the United States. in the late 19th century, almost 150 new dinosaur species were revealed by the excavations and reconstructions of the american paleontologists edward Drinker cope and Othniel charles Marsh. they began as collabo-rators but ended up in a three-decade-long intense rivalry. Marsh did not perform much field work. One account claims that Marsh visited a spot where
dinosaur bones were lying about like logs but did not recognize them. Marsh was, however, rich enough to buy fossils and to hire field workers. His uncle, George Peabody, endowed a museum at yale Univer-sity specifically so that Marsh could study dinosaurs. cope, who was also wealthy, spent much productive time in the field. He worked in Montana at the same time that General custer’s army was being annihi-lated nearby. When suspicious native americans checked out his camp, he entertained them by taking out his false teeth until they went away. a school-teacher who found some dinosaur bones genially informed both Marsh and cope about them. cope sent the schoolteacher money and told him to ask Marsh to send all the specimens back to him. the rivalry grew intense enough that the excavators of the Marsh and cope camps threw rocks at one another. cope lost his money, but not his ego, in financial speculation and died poor. He willed his body to a museum, hoping to have his skeleton used as the “type specimen” to represent the human spe-cies. When the skeleton was prepared, it was found to have syphilitic lesions and was unsuitable as a type specimen.
Most of what scientists know about dinosaurs comes from the study of fossilized bones, in the tradition of cope and Marsh. Recently, this has included the microscopic study of the spaces left by blood vessels in the bones. Other kinds of fossil evi-dence have also allowed many insights into the lives of dinosaurs, such as the following.
● Gastroliths. these are “stomach stones” that helped grind coarse plant material in the stomachs of some dinosaurs that ate branches and leaves. Many modern birds swallow stones that help them grind their food.
● Eggs. Fossilized eggs with embryos inside reveal some of the processes of dinosaur development. in addition, the discovery of caches of eggs reveals that dinosaurs cared for nests of eggs in a manner similar to that of most modern birds.
● Soft material. in 2005, the paleontologist Mary Higby Schweitzer discovered soft tis-sue, including what may be blood vessels and cells, in a dinosaur bone. Some of this soft tissue was collagen, the protein that makes up cartilage. analysis of this colla-gen revealed a close relationship with birds rather than with reptiles or mammals.
Since the time of Owen, Marsh, and cope, dinosaurs have entered the popular imagination as emblematic both of huge size and of evolutionary failure. in both ways, the image is unfair. First, many
xviii+598_Biodiversity_F.indd 190 11/17/11 1:12 PM

dinosaurs 191
dinosaurs were small or medium sized. Many of the smaller dinosaurs were sleek and rapid, and some had feathers. Second, there were many kinds of dinosaurs all over the world; they were, for more than 100 mil-lion years, an astounding evolutionary success.
Some dinosaurs were extremely large. Sauro-poseidon proteles, which lived 110 million years ago in what is now Oklahoma, had a body 100 feet (30 m) in length, with a neck more than 30 feet (10 m) long. the size estimate of the neck of this dinosaur is based on the fact that most vertebrates have the same number of neck vertebrae, and the longest ver-tebrae found for this dinosaur were about fi ve feet (152.4 cm) long. the sauropod Supersaurus weighed nearly 110 tons (100 metric tons), the largest animals to ever live on land (see fi gure). they were also the largest four-legged animals that could possibly have lived; if they had grown to 154 tons (140 metric tons), the size of the legs necessary to support the weight would have become so great that the legs would not have been able to move.
Sauropods were herbivorous dinosaurs whose large size and long necks allowed them to eat leaves even in the tallest trees. instead of having large teeth to grind the branches and leaves, they had small sharp teeth that could tear leaves and branches to be swallowed whole. Grinding occurred in the stomach with the aid of gastroliths. if dinosaurs such as Apa-tosaurus (formerly Brontosaurus) held their heads and necks upright, they would have needed seven times the blood pressure of a human, and the huge heart would have required the energy equivalent to about half the food intake of the dinosaur. Some
paleontologists say that huge, long-necked dinosaurs did not raise their heads. Perhaps they lifted their heads just long enough to eat. in contrast, the car-nivorous dinosaurs, even the famous Tyrannosaurus rex, were much smaller (about fi ve tons [4.6 metric tons]) because meat was much less available than vegetation (see fi gure on page 192).
the huge sauropod dinosaurs were probably also stupid. the huge, long-necked dinosaurs had small brains (and small teeth) because the force of leverage that would have been necessary to raise a large head at the end of a long neck would have been too great. large dinosaurs were also stupid because a single, large brain in the head would have been too far away from the tail to allow quick control over muscle movement. the large dinosaurs had a brain but also had a secondary ganglion of nerves near the base of the tail, which aided in control of posterior muscles.
among the oldest known dinosaur fossils are Herrerasaurus and Eoraptor, from the late trias-sic period about 230 million years ago. Because these fossils already had dinosaur specializations, the ancestral dinosaur must have lived even earlier. these earliest dinosaurs were about the size of a human and had narrow heads, long pointed snouts, mouths that extended from ear to ear, many small sharp teeth, and large eyes. Unlike nearly all other reptiles, these early dinosaurs walked on their two hind legs, which allowed their “hands” to evolve the ability to grasp prey. From animals similar to Herrerasaurus, two lineages of reptiles evolved: the bird-hipped dinosaurs (ornithischians) and the liz-ard-hipped dinosaurs (saurischians), which evolved
Grazing dinosaurs, such as Supersaurus, were the largest land animals to ever live—each leg was the size of a human.
xviii+598_Biodiversity_F.indd 191 11/17/11 1:12 PM

fi rst entry192 dinosaurs 192
into numerous forms during the ensuing Jurassic and cretaceous periods.
Ornithischians had a pelvis that reminded early paleontologists of birds. their teeth and jaws allowed for the tearing and grinding of plant mate-rials, and the enlarged ribs and pelvis allowed for the digestion of large quantities of plant material. Despite their name, the ornithischians were not the ancestors of birds; they all met extinction at the end of the cretaceous period. as time went on, ornithis-chians diverged into different lineages, all of which became much larger than their Herrerasaurus ances-tors. they included the following.
● Pachycephalosaurs, or “thick-headed” dino-saurs, had skulls with thick domes that may have served as battering rams during fi ghts. Recent research has failed to fi nd evidence of tiny fractures that would have resulted from such confl icts.
● Ceratopsians, or horned dinosaurs, reverted to walking on all fours. By the late Paleo-zoic, some were quite large and had large horns, such as Triceratops.
● Thyreophores, or “shield-bearing” dino-saurs, evolved into large animals with armored plates and, in the stegosaurs, a paired row of large plates along the back.
● Ornithopods also evolved into large forms but remained primarily bipedal.
Saurischian dinosaurs had long necks and “hands” with long index fi ngers, a basic pattern now found in many birds. they diverged into the sauro-pods and the theropods.
● Sauropods became very large. an example is the Supersaurus mentioned above.
● Theropods had long, fl exible necks, sharp teeth and claws, and “hands” with three curved claws that were good at grasping. their bones were thin, light, and hollow, like those of modern birds. their collar-bones were fused into a wishbone, or fur-cula, a feature found only in birds today. these features made the later development of fl ight possible.
From large theropod ancestors, coelourosauri-ans evolved. Some of them, such as Tyrannosau-rus, were quite large, with big heads but very small arms. Others were much smaller, with larger brains and eyes like modern birds, resembling an ostrich in body form, even to the point of having a tooth-less beak. all but one of these birdlike lineages became extinct. the lineage that survived to eventu-
Predatory dinosaurs, such as Tyrannosaurus rex, were much smaller than the grazing dinosaurs but were the largest terrestrial predators to ever live.
xviii+598_Biodiversity_F.indd 192 11/17/11 1:12 PM

dinosaurs 193
ally become birds was the mani-raptorans, which were dinosaurs with feathers.
the maniraptoran dinosaurs apparently had hollow bones, a wishbone, and feathers before they had the ability to fl y. evolu-tionary biologists conclude from this that the original function of feathers was not for fl ight but to hold in body heat. this may have been the fi rst step in warm-blood-edness, which is important in providing the energy needed for fl ight. Specimens of one of them, Oviraptor, have been found brooding eggs in the fashion of modern birds, another indication of warm-bloodedness.
One evolutionary branch of maniraptorans not only had feathers but could fl y. the earli-est known of these was Archae-opteryx, from 150 million years ago, during the Jurassic period (see fi gure). although skeletally sim-ilar to other maniraptorans, Archaeopteryx resem-bled modern birds in having long arms that formed a wing with fl ight feathers, reduced teeth, and a large brain. Because maniraptorans were runners, evolutionary biologists believe that fl ight originated from rapid running “from the ground up” rather than from gliding “from the trees down.” the evo-lutionary lineage of Archaeopteryx became extinct, but related lineages continued to evolve more bird-like characteristics, such as a larger breastbone that allowed massive fl ight muscles to anchor. By the early cretaceous period, before the extinc-tion of nonavian dinosaurs, there were fi nch-sized dinosaur birds capable of a modern type of fl ight. Other lineages adapted to an aquatic existence and resembled modern waterfowl, some of them even losing the ability to fl y. all but one lineage, the birds, became extinct by the end of the cretaceous period.
Dinosaurs diversifi ed into many forms, from huge consumers of branches and leaves to small, sleek predators, mainly during the Jurassic and cre-taceous periods. When fl owering plants evolved at the beginning of the cretaceous, some dinosaurs evolved the ability to consume their leaves, but there was apparently no coevolution between nonavian dinosaurs and fl owering plants for pollination of fl owers or dispersal of fruits. insects (and some birds) evolved many pollinating forms, and mammals were particularly good dispersers of fruits and seeds. the increasing dominance of fl owering plants in the
landscape may have been one reason that dinosaurs were in decline during the late cretaceous. Massive volcanic eruptions changed the climate and further hastened the decline of dinosaurs. it was appar-ently an asteroid that hit earth 65 million years ago that sent all remaining dinosaurs, except birds, into extinction. Until almost the last moment, there were at least 100 species of dinosaurs. Fossil evidence sug-gests that some dinosaurs survived the cretaceous extinction, but not many and not for long.
See also cretaceous period, biodiversity in; Jurassic period, biodiversity in; triassic period, biodiversity in.
Further readingachenbach, Joel. “Dinosaurs: cracking the mystery of how
they lived.” National Geographic, March 2003, 2–33.asara, John M., et al. “Protein sequences from mastodon
and Tyrannosaurus rex revealed by mass spectros-copy.” Science 316 (2007): 280–285.
Bakker, Robert t. The Dinosaur Heresies. new york: Wil-liam Morrow, 1986.
———. “the bite of the Bronto.” Earth 3 (1994): 26–33.———. “Dinosaur mid-life crisis: the Jurassic-cretaceous
transition in Wyoming and colorado.” in: lucas, S.G., et al., eds. Lower and Middle Cretaceous Terrestrial Ecosystems: New Mexico Museum of Natural History and Science Bulletin 14 (1998): 67–77.
carpenter, K., K. F. Hirsch, and J. R. Horner, eds., Dinosaur Eggs and Babies. cambridge: cambridge University Press, 2007.
carrano, Matthew t., and Patrick M. O’connor. “Bird’s-eye view.” Natural History, May 2005, 42–47.
Archaeopteryx was a feathered dinosaur (a bird) that retained many dino-saur characteristics, such as teeth, claws on its wings, and vertebrae in its tail. It had feathers similar to those of modern birds.
xviii+598_Biodiversity_F.indd 193 11/17/11 1:12 PM

first entry194 disjunct species 194
clark, James M., and Xing Xu. “evolutionary transitions among dinosaurs: examples from the Jurassic of china.” Evolution Education and Outreach 2 (2009): 236–247.
currie, Philip e., and Kevin Padian. Encyclopedia of Dino-saurs. new york: academic Press, 1997.
erickson, Gregory M., et al. “tyrannosaur life tables: an example of nonavian dinosaur population biology.” Science 313 (2006): 213–214.
Fiorillo, anthony R. “the dinosaurs of arctic alaska.” Sci-entific American, December 2004, 84–91.
Fipper, Steve. Tyrannosaurus Sue: The Extraordinary Saga of the Largest, Most Fought Over T. Rex Ever Found. new york: Freeman, 2000.
Foster, J. Jurassic West: The Dinosaurs of the Morrison For-mation and Their World. Bloomington, ind.: indiana University Press, 2007.
Horner, John R., Kevin Padian, and armand de Ricqlès. “How dinosaurs grew so large—and so small.” Scien-tific American (July 2005): 56–63.
irmis, Randall B., et al. “a late triassic dinosauromorph assemblage from new Mexico and the rise of dino-saurs.” Science 317 (2007): 358–361.
levin, eric. “Dinosaur family values.” Discover, June 2003, 34–41.
norell, Mark. Unearthing the Dragon: The Great Feath-ered Dinosaur Discovery. new york: Pi Press, 2005.
Organ, chris l., et al. “Molecular phylogenetics of mast-odon and Tyrannosaurus rex.” Science 320 (2008): 499.
Parker, Steve. Dinosaurus: The Complete Guide to Dino-saurs. Richmond Hill, Ont.: Firefly Books, 2003.
Prum, R., and a. H. Brush. “the evolutionary origin and diversification of feathers.” Quarterly Review of Biol-ogy 77 (2002): 261–295.
Sampson, Scott D. Dinosaur Odyssey. Berkeley: University of california Press, 2009.
Sander, P. Martin, and nicole Klein. “Developmental plas-ticity in the life history of a prosauropod dinosaur.” Science 310 (2005): 1,800–1,802.
Schweitzer, Mary Higby, et al. “Beta-keratin specific immu-nological reactivity in feather-like structures of the cretaceous alvarezsaurid, Shuvuuia deserti.” Jour-nal of Experimental Zoology B (Mol Dev Evol) 285 (1999):146–157
———, et al. “Gender-specific reproductive tissue in rat-ites and Tyrannosaurus rex.” Science 308 (2005): 1,456–1,460.
———, et al. “analyses of soft tissue from Tyrannosau-rus rex suggest the presence of protein.” Science 316 (2007): 277–280.
Stevens, Kent a., and J. Michael Parrish. “neck posture and feeding habits of two Jurassic sauropod dino-saurs.” Science 284 (1999): 798–800.
Stokstad, erik. “Did horny young dinosaurs cause illusion of separate species?” Science 318 (2007): 1,236.
turner, alan H., et al. “a basal dromaeosaurid and size evolution preceding avian flight.” Science 317 (2007): 1,378–1,381.
———, et al. “Feather quill knobs in the dinosaur Veloci-raptor.” Science 317 (2007): 1,721.
Weishampel, David B., Peter Dodson, and Halszka Osmól-ska. The Dinosauria, 2nd ed. Berkeley: University of california Press, 2004.
Xu, X., et al. “Basal tyrannosauroids from china and evi-dence for protofeathers in tyrannosauroids.” Nature 431 (2004): 680–684.
yeoman, Barry. “Schweitzer’s dangerous discovery.” Dis-cover, april 2006, 36–41, 77.
disjunct species Disjunct species contain or consist of populations (disjunct populations) that are isolated from one another. Because of the distances that separate the populations of disjunct species, gene flow rarely occurs between them. Gene flow occurs when pollen or seeds (of plants) or animals travel from one population or part of a population to another. if the populations of a disjunct species are genetically different, the populations can often be described as subspecies. if enough time passes, it is practically inevitable that the disjunct populations will evolve into separate species unless they become extinct.
two ways are generally recognized in which populations within a species could have become dis-junct. the first is that the populations are relictual, that is, they are relicts of a large ancestral popula-tion (see figure on page 195). Populations of the spe-cies that were formerly present between the disjunct populations have disappeared, leaving the relictual populations isolated from one another. if the two relictual populations diverge, they are considered the product of vicariance. the other way that disjunct populations can form is that a few organisms dis-persed (as seeds or as a small flock) from the main population and settled in a distant location, forming a new population, an explanation called dispersal.
How can scientists determine which explana-tion, the relictual versus the dispersal, is correct? the best evidence is if fossil or other evidence of the formerly intervening populations can actually be discovered. in some cases, evidence for popula-tions of that particular species cannot be found, but if it is found for other species with similar dis-junct distributions, then it can be inferred for the species in question as well. For example, there are many species of wildflowers in the alpine tundra of the high elevations of the Sierra nevada, cascade, Olympic, Rocky, and Sangre de cristo Mountains of western north america that are very similar, in some cases nearly identical, to those of the arctic tundra of northern alaska and canada. Hundreds
xviii+598_Biodiversity_F.indd 194 11/17/11 1:12 PM

disjunct species 195
or thousands of miles separate the disjunct arctic and alpine populations. these distributions were, in fact, somewhat difficult to explain before louis agassiz and charles Darwin. agassiz explained the ice ages: at various times, much of north america was, in fact, in the arctic tundra zone. as the glaciers retreated, some tundra plants migrated north, where they are now found as arctic tundra; others migrated up the mountains, where they are now stranded as alpine tundra. Darwin explained the subsequent vicariance of these populations by evolution occur-ring separately in each. this is also the explanation of how disjunct populations of tundra species, such as the sedge Eriophorum vaginatum and boreal spe-cies such as the larch tree, came to be stranded in the bogs of northern states in the United States, such as Michigan.
the next best evidence comes from genetic com-parisons. if the disjunct populations can be shown to have several distinct mutations within their Dna, mutations not shared with one another, then they may be relictual. if one of the populations has Dna that is a subset of another population, then the first population dispersed from the second. the only problem with this approach is that even if one popu-lation dispersed from another, the new population can evolve its own distinct mutations, which would
make it appear as relictual. therefore, genetic stud-ies are most useful to detect recently dispersed dis-junct populations.
numerous examples of disjunct populations have been found. One example is the seaside alder, Alnus maritima. Many alders are large bushes that grow in swamps or next to streams and rivers with rocky beds. the hazel alder, Alnus serrulata, grows in this kind of streamside habitat through-out much of the eastern United States, but unlike the hazel alder, the seaside alder is a disjunct spe-cies and consists entirely of three small disjunct populations. One population lives on the Delmarva Peninsula, east of chesapeake Bay. another lives almost entirely within Johnston county in central Oklahoma. the third lives in a single swamp in Bartow county in northwest Georgia. Why is the seaside alder so rare, despite its apparent similarity to the hazel alder, which grows throughout eastern north america?
First, consider the vicariance hypothesis. alders reproduce in two ways. their seeds, produced in autumn, grow best in sunny locations with moist mineral soil. along many rivers, locations that have moist mineral soil are often shaded and not good for the growth of alder seedlings. in a second reproduc-tive mode, once a bush is established, it can keep resprouting from the same roots even if floods wash many of its branches away or if it is subsequently shaded by the growth of larger trees. the botanists Stanley Rice and J. Phil Gibson have demonstrated that both species of alders prefer sunny over shady conditions, but the hazel alders are found in shade more frequently than seaside alders.
in the past (perhaps after the most recent ice age), both species of alders may have established themselves abundantly in the new landscape of sunny, moist, mineral soil. Subsequent regrowth of forests shaded the alder habitats, and the hazel alders have tolerated these shaded conditions at least a little better than the seaside alders. Hazel alders have therefore persisted throughout eastern north america, whereas the seaside alders have persisted in only three places. the seaside alder sur-vived in these three places by chance, not because the three places were the most suitable habitats. the climatic conditions in Oklahoma, Georgia, and Del-aware are quite different, and without doubt there are many places between these three locations that are better for seaside alder growth but in which the seaside alder by chance became extinct. Researchers have found that separate mutations have occurred in the three populations. these three populations are now classified as three distinct subspecies. there is no direct evidence that earlier populations of the seaside alder lived in places between Oklahoma,
Fan palms (Washingtonia filifera) have been planted widely throughout the western United States. However, they are native only to palm oases in the deserts of the southwestern United States, where their survival is in jeopardy. The wild populations of palms, such as this one in the Anza-Borrego Desert of California, are relicts of larger populations that grew before the formation of deserts in the South-west. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 195 11/17/11 1:12 PM

first entry196 disjunct species 196
Georgia, and Delaware. Fossilized leaves that are identical to modern seaside alder leaves have been found in northwestern north america, which sug-gests that this species once grew across the entire continent. the species of alder that are most closely related to the seaside alder, in fact, grow in eastern asia. this is the vicariance hypothesis for the dis-junct distribution of the seaside alder.
now consider the dispersal hypothesis. the seaside alder might have originated in one loca-tion, perhaps Delmarva, and subsequently dispersed to the other locations. How could this have hap-pened? Something, or someone, might have carried the seeds from Delmarva to the other locations. neither wind nor watershed patterns seem to suggest how this could have occurred. native americans use alder bark as a medicine and may have carried seeds or cuttings with them. the Georgia population is within the former cherokee nation territory, and the cherokees are known to have moved south from the iroquoian lands of what is now the northeastern United States. Subsequently, the cherokee were relo-cated by the United States government to Oklahoma in 1835. could cherokee have carried the seeds or small plants with them from the northeast to Geor-gia to Oklahoma, connecting all three of the existing populations? this explanation is unlikely to be true, because the three populations (subspecies) are differ-ent enough that they must have been separated for a long time, much longer than the cherokee migra-tions of ca. 1000 and 1835 c.e.
in the case of the seaside alder and in many other cases as well, the disjunct populations are threatened with extinction. not only do small populations have a greater risk of extinction, disjunct populations are frequently not in the habitats most suitable for them. Oklahoma seaside alders have been planted and are surviving well in distant locations, while they are barely surviving in Oklahoma, where they can with-stand the summer heat only if the soil is continually wet. this is because the climatic changes that iso-lated the disjunct populations in the first place are now threatening them with extinction. the Georgia and Delmarva populations are similarly threatened.
in other cases, a species may have a large, cen-tral population and several disjunct populations at some distance away from the central population, as in the following examples:
● the least tern (Sternula antillarum) is a shorebird that is abundant in many areas, but the interior subspecies of the least tern (which migrated up the Mississippi and arkansas Rivers into Oklahoma) is rare.
● the white spruce (Picea glauca) is abundant in the subalpine forests of the mountains
of central north america and the boreal forests of canada. the limber pine (Pinus flexilis) is fairly common in the Rocky Mountains and Sierra nevada. in the Black Hills of South Dakota, relictual populations of both the white spruce and the limber pine survive. they became stranded in the Black Hills after the most recent ice age and have declined in abundance since then. the spruce remains abundant in the Black Hills because it grows well in cool, moist, shaded areas of the northern hills, although global warming could rapidly create condi-tions in which they can no longer survive. in contrast, the limber pine has nearly become extinct in the Black Hills because it requires cool, dry, sunny conditions, which are now rare in the Black Hills. the few remaining limber pines are being crowded and shaded out by the extremely abundant ponderosa pines.
● the bishop pine (Pinus muricata) grows near the northern coast of california. there is a small population of bishop pines about 200 miles from the main population in the Purísima Hills near the central california coast. it persists on a rare kind of soil com-posed almost entirely of dead diatoms that accumulated in shallow oceans millions of years ago.
● a genus of heather shrub, Erica, is abundant in europe but also in South africa.
● Olive trees are also disjunct. the familiar olive tree (Olea europaea) grows around the Mediterranean Sea, O. paniculata grows in the forests of Queensland in australia, and the black ironwood O. laurifolia grows in South africa.
● the crowberry family empetraceae has disjunct genera. Corema album grows in the azores, Spain, and Portugal, while C. conradii grows in the northeastern United States and adjacent areas in canada. Species of the genus Empetrum live in cool temper-ate regions of the northern and Southern Hemispheres, but not in between.
● Many freshwater fishes have disjunct popu-lations because their habitats are separated from one another by large expanses of dry land.
in some cases, humans are strongly implicated in disjunct distributions of other species. the pygmy shrew (Sorex minutus) has an irish population dis-junct from the mainland european population. Genetic analysis shows that the irish shrews are all
xviii+598_Biodiversity_F.indd 196 11/17/11 1:12 PM

disturbances, ecological 197
descended from a recent founding event. they may have been stowaways in a human vessel.
Disjunct populations are frequently the failed dead ends of evolution. they can also be wellsprings of new biodiversity. Because they are disjunct, these populations are reproductively isolated from the larger population, which allows them to evolve in a different direction. if they have dispersed from a large, central population, they may also by chance be genetically different from the original population. evolution can occur more rapidly in small popula-tions. if the disjunct population evolves rapidly, it can produce a new species that will appear on the scene in what amounts to a geological instant. Much of the work of saving biodiversity is in saving small disjunct populations of rare species in the face of rapid climate change.
See also biogeography; bogs, life in; extinction; founder effect and genetic drift; ice ages; population genetics; specia-tion; tundra.
Further readingBernardi, G., and J. lape. “tempo and mode of specia-
tion in the Baja california disjunct fish species Aniso-tremus davidsonii.” Molecular Ecology 14 (2005): 4,085–4,096.
coleman, Max, et al. “Repeat intercontinental dispersal and Pleistocene speciation in disjunct Mediterranean and desert Senecio (asteraceae).” American Journal of Botany 90 (2003): 1,446–1,454.
Fok, Koon Wah, et al. “inferring the phylogeny of disjunct populations of the azure-winged magpie Cyanopica cyanus from mitochondrial control region sequences.” Proceedings of the Royal Society of London B (Biol-ogy) 269 (2002): 1,671–1,679.
Gibson, J. Phil, Stanley a. Rice, and clare M. Stucke. “comparison of population genetic diversity between a rare, narrowly distributed species and a common, widespread species of Alnus (Betulaceae).” American Journal of Botany 95 (2008): 588–596.
Habel, Jan christian, and thorsten assmann. Relict Spe-cies: Phylogeography and Conservation Biology. Ber-lin: Springer, 2010.
Koenemann, Stefan, et al. “a new, disjunct species of Spe-leonectes (Remipedia, crustacea) from the canary islands.” Marine Biodiversity 39 (2009): 215–225.
Masheretti, S., et al. “How did pygmy shrews colonize ireland? clues from a phylogenetic analysis of mito-chondrial cytochrome b sequences.” Proceedings of the Royal Society of London B (Biology) 270 (2003): 1,593–1,599.
Penman, t. D., et al. “Bioclimatic analysis of disjunct populations of the giant burrowing frog, Heleiopo-rus australiacus.” Journal of Biogeography 32 (2005): 397–405.
Schrader, James a., William R. Graves, Stanley a. Rice, and J. Phil Gibson. “Differences in shade tolerance help explain varying success in two sympatric Alnus species.” International Journal of Plant Sciences 167 (2006): 979–989.
terry, astrid, et al. “Restricted gene flow and incipient speciation in disjunct Pacific Ocean and Sea of cortez populations of a reef fish species, Girella nigricans.” Evolution 54 (2000): 652–659.
Wisheu, irene c., et al. “Disjunct atlantic coastal plain spe-cies in nova Scotia: Distribution, habitat and conser-vation priorities.” Biological Conservation 68 (1994): 217–224.
disturbances, ecological ecological distur-bances are any events that damage or remove the dominant vegetation in an ecological community. they can be as small as a gopher mound in a grass-land or a tree falling in a forest or as large as a fire or hurricane that damages an extensive area. they usually occur individually, but during the ice ages glaciers caused many thousands of disturbances simultaneously.
the term disturbance carries negative connota-tions, and for the individual plants in the dominant vegetation a disturbance is indeed negative. How-ever, a disturbance can also be considered an abrupt release of resources: Suddenly, light shines down on the forest floor, or nutrients from ashes are released into the soil. this was experimentally demonstrated in the 1960s, when some forests on the Hubbard Brook watershed in new Hampshire were clear-cut. a flush of nutrients came out in the water that drained from the clear-cut slopes. to the organisms that can use this flush of resources, a disturbance is positive. Many of these organisms have rapid life cycles and population growth rates that allow them to capitalize on the briefly available resources. after plants begin to grow in the disturbed area and ani-mals return, the resources are again locked up within the tissues of organisms. as forests grew back on the Hubbard Brook watershed, the flush of nutrients at the bottom of the slope decreased. Within decades, forests had grown back, and the runoff of nutrients was no greater than that of adjacent intact slopes.
Many species of organisms depend on periodic disturbances and cannot survive in ecological com-munities that do not experience them. it is even questionable whether ecological communities could persist without at least small-scale disturbances, since there are no communities that do not experi-ence them. it is clear that disturbances enhance bio-diversity, and the management of disturbances is an important part of the work of people who maintain the healthy function of ecosystems. in the old days
xviii+598_Biodiversity_F.indd 197 11/17/11 1:12 PM

first entry198 disturbances, ecological 198
of Smokey the Bear, foresters put out every fire they could; now they spend as much time conducting “controlled burns” as they do fighting wildfires.
Disturbances can be placed roughly in two cat-egories: those that remove the soil, and those that leave the soil largely intact. When a disturbance, such as a volcanic eruption or a landslide, removes the soil, the plants that move into the disturbed area must build the soil up before the ecosystem can recover. if a disturbance, such as a storm or a fire, leaves the soil in place, many plants can move right in and begin to grow. in fact, many seeds that are already in the soil can simply sprout. the plants that grow after a disturbance do not all grow at the same time or the same rate. Some plants grow faster and dominate the habitat sooner, while others grow more slowly and dominate the habitat later. the result is a series of stages, known as ecological suc-cession, by which the habitat recovers.
even the smallest disturbances can release resources from the dominant plant species and make
them available for other plant species. the grasses of the shortgrass prairie of the western plains of north america (itself dependent on fire; see below) form a thick sod of underground stems and fibrous roots that few seedlings can penetrate. even fires do not disturb this sod, but bison and prairie dogs disrupt the sod. Bison scratch their parasites by roll-ing around in the grass, tearing it up and creating “buffalo wallows.” new seedlings of grass and other plants grow in these roughly circular wallows, called greenups because they produce new leaf area earlier in the spring than do the established grasses. Prairie dogs dig burrows and produce piles of newly turned earth (see figure). the prairie around each burrow may consist almost solely of the grama grasses Bou-teloua curtipendula and B. gracilis, while wild-flowers such as scarlet globemallow (Sphaeralcea coccinea) can grow only on these mounds. eroded areas of the shortgrass prairie may be the only place to find wildflowers such as the evening primrose Oenothera caespitosa (see figure on page 199). Fires and other disturbances can, therefore, greatly enhance biodiversity.
animals may also benefit from disturbances. Generations of children have learned from the Walt Disney movie Bambi that hunters and fires are evil. conservationists have called this the Bambi syndrome. the postfire regrowth, however, can produce a bounty of rich food for grazing and browsing mammals that would find little to eat in the middle of an old forest. Fires and other distur-bances can also enhance animal biodiversity. the extent to which disturbances enhance the diversity of fungi and microbes has not been extensively investigated.
EColoGICal CommuNITIES ThaT dEpENd upoN dISTurBaNCE
it is probable that nearly every terrestrial benefits from occasional disturbances such as fires, which clear away dead wood, release nutrients, and allow sunlight to reach the ground. Some ecological com-munities, however, would not even exist without recurring disturbances (usually fire) to which their species have adapted but which other species cannot tolerate. the disturbances prevent ecological suc-cession and maintain a fire-tolerant set of species. examples include the following.
Grasslands. the grasslands (also known as prai-ries and steppes) of the world could not exist without periodic fires. an ocean of grassland, the tallgrass prairie, once covered major portions of Minnesota, iowa, illinois, the eastern portions of north and South Dakota, nebraska, and Kansas and extended down into Oklahoma and texas. the tallgrass prai-rie received enough rainfall that many species of
Prairie dogs and bison create disturbed areas in grasslands, such as in the badlands of South Dakota. Prairie dogs dig burrows and produce mounds, while bison graze, trample, and roll in the dirt. The disturbed areas may have higher levels of soil nutrients. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 198 11/17/11 1:12 PM

disturbances, ecological 199
forest trees, such as oaks, could have easily grown there. tree seeds continually germinated near the edges of the tallgrass prairie, but frequent fires killed many of the germinating saplings. the saplings that survived, such as those of bur oak (Quercus mac-rocarpa), grew into small shrubs rather than tall trees. the fires did not harm the grasses, which are perennials and have underground buds and whose leaves grow from the bases. after a fire, the grasses would simply resprout. this growth form allowed grasses to flourish not only after fires but in response to grazing animals. the nongrass wildflowers, called forbs, were also perennials and could sprout after fires. Shortgrass (semiarid) prairies, such as the west-ern prairies of colorado, Wyoming, and the western Dakotas, do not have enough moisture for trees to grow. they do, however, have enough moisture to support cool desert shrubs such as sagebrush, yet sagebrush does not dominate these grasslands, largely because of fire. Fire kills sagebrush and other shrubs but not the grasses. the steppes of central asia experience a similar cycle of renewal by fire.
Fire actually benefits the grasses and forbs of the prairies. the plants die back to the ground each
winter, then sprout in the spring. after a few years, dead stems and leaves have built up a thick layer that not only shades the ground but also ties up the nutrients. Fire transforms dead litter, with trapped minerals, into fertilizing ash. the sun penetrates to the soil surface, now dark with ash, and warms it more quickly in the spring. Grasses and forbs grow profusely after a fire if it occurs during early spring or late fall. Fires are unlikely to occur at other times, when the grassland has wet green foliage.
Prairies, especially those with tall grass, have deep, rich soil. nearly all the north american tall-grass prairies have been converted to prime agricul-tural land. Very few of these grasslands now remain, most of them along railroad tracks or cemeteries that were established before agriculture. Preserving these prairies is therefore very important. Because it keeps the trees and shrubs out, fire is a necessary management tool for maintaining prairies. Fall and spring are the best times for controlled burns, as the prairies are not filled with green blades of grass, but these are also the times when the wind can be stron-gest and fires can get out of control.
Chaparral. Found mostly in california, chap-arral habitats consist of fire-adapted shrubs. after fire, the shrubs resprout either from their roots or from seeds. if several decades pass without fire, the chaparral shrubs have more dead than living stems, and the litter layer beneath them is deep and crisp in the hot, dry summers. One can even smell vola-tile combustible chemicals produced by the shrubs themselves. Fire is inevitable. after the fire, not only do the shrubs grow back, but many wildflowers as well, whose seeds have lain dormant in the soil since the previous chaparral fire. Many other shrublands, such as the matorral of chile and the fynbos of South africa, also depend on a fire cycle.
Serotinous pine forests. there are certain pine forests in coastal california mountains that rely on a fire cycle. they are called closed-cone pines because their cones do not open as soon as they become mature. the seeds usually remain sealed inside until a fire bakes them open. their cones are serotinous, that is, hardened. then, after the fire, the seeds fall on the ground and germinate into a thick regrowth of pine seedlings. One example is the forest of bishop pines (Pinus muricata) in the Purísima Hills near lompoc, california. a fire destroyed the forest in 1995. Before the end of the summer, the ground beneath standing black trunks was covered with almost nothing but pine seedlings (see figure on page 200). Beneficial fungi help the pines grow rapidly in soil that was fertilized by the ashes. By 2002, some of the new pine trees were already producing cones. Often, these forests have no other dominant tree species (see table on page 201). Older cones on some
In grasslands, most wildflowers grow in eroded or otherwise disturbed areas, where the thick grass sod has been broken. This is Oenothera caespitosa in the badlands of South Dakota. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 199 11/17/11 1:12 PM

first entry200 disturbances, ecological 200
individuals of these species eventually open and drop their seeds even if a fire does not occur.
Many pine forests may not absolutely require a fire cycle for reproduction, but the fires strongly enhance regeneration. young lodgepole pines (Pinus contorta) in california and in the Rocky Mountains often have cones that open without fire, whereas older trees often have cones that will not open unless they are burned. the populations of pitch pines (P. rigida) of the pine barrens in new Jersey also consist of a mixture of closed-cone and open-cone trees. the cones of ponderosa pines (P. ponderosa) in western north america do not need fire to open, nor do their seeds need to grow on burned soil. How-ever, fires clear out openings in which pine seedlings can grow much more readily than in the shade of trees in an undisturbed forest.
Floodplain and streamside habitats. Fire is the most common, but by no means the only, type of disturbance that determines the species composi-tion of wild habitats. Riparian (riverside) forests consist of tree species, such as sycamores, cotton-
woods, box elders, and willows, whose seedlings begin their growth in sunny, wet conditions. these conditions are created by floods that wash away the overstory of trees in part of the floodplain and open up a sunny space in which the seedlings can grow. these seedlings do not grow well in shade and therefore do not establish well in places that have not flooded.
dISTurBaNCES aNd CoNSErvaTIoNto most people, disturbances (particularly fire) are nothing more than disasters that need to be stopped. When wildfires, fanned by hot, dry Santa ana winds, make the bushes of the california chap-arral explode into flame in a rapidly advancing front, firefighters work around the clock to stop this natural disaster. Humans want to make the natural world “behave itself.” they do not want the fire to burn the houses of people who insist on living in the chaparral in defiance of the natural fire cycle. in such circumstances, total suppression of fires is expected. it is, however, impossible. When fires do not burn in western forests or chaparral, dead tim-ber builds up. When there are no small fires, the risk of large fires increases. When fires begin as a result of lightning or arson, they are much larger and more damaging than they would otherwise have been. Forest managers often clear some areas as an artificial substitute for fires. they also use con-trolled burns, small fires deliberately set, to reduce the danger of larger fires. Rarely, these fires get out of control.
the widespread practice of fire suppression dur-ing the 20th century has dramatically altered the north american landscape. almost all the tallgrass prairies have been destroyed by agriculture and the growth of cities and suburbs. However, even if this had not occurred, fire suppression would have diminished the prairie and allowed forests to spread. the area of the shortgrass prairie is dimin-ishing, and ponderosa pine woodland and sage-brush are spreading. in large areas of the central United States, eastern red cedar (Juniperus virgin-iana), formerly controlled by natural fires, is spread-ing. although it is a native species, it has assumed the role of an invasive. in the arbuckle Mountains of central Oklahoma, native forests of post oaks (Quercus stellata) were cut down, and the land was used for pasture. Fire suppression has subsequently allowed ashe junipers (Juniperus ashei) to cover these mountains. Both species of junipers cast deep shade that prevents the successful regeneration of the oak forests. (Besides all this, the ashe juniper is a major source of allergenic pollen.) Woodlands of oaks, which can partially tolerate fires, have been replaced by forests of trees that thrive in the
Some pines, such as these bishop pines (Pinus muri-cata) in the Purísima Hills of coastal California, open their cones only after a fire has destroyed the adult trees. A thick growth of pine seedlings springs up after the fire. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 200 11/17/11 1:12 PM
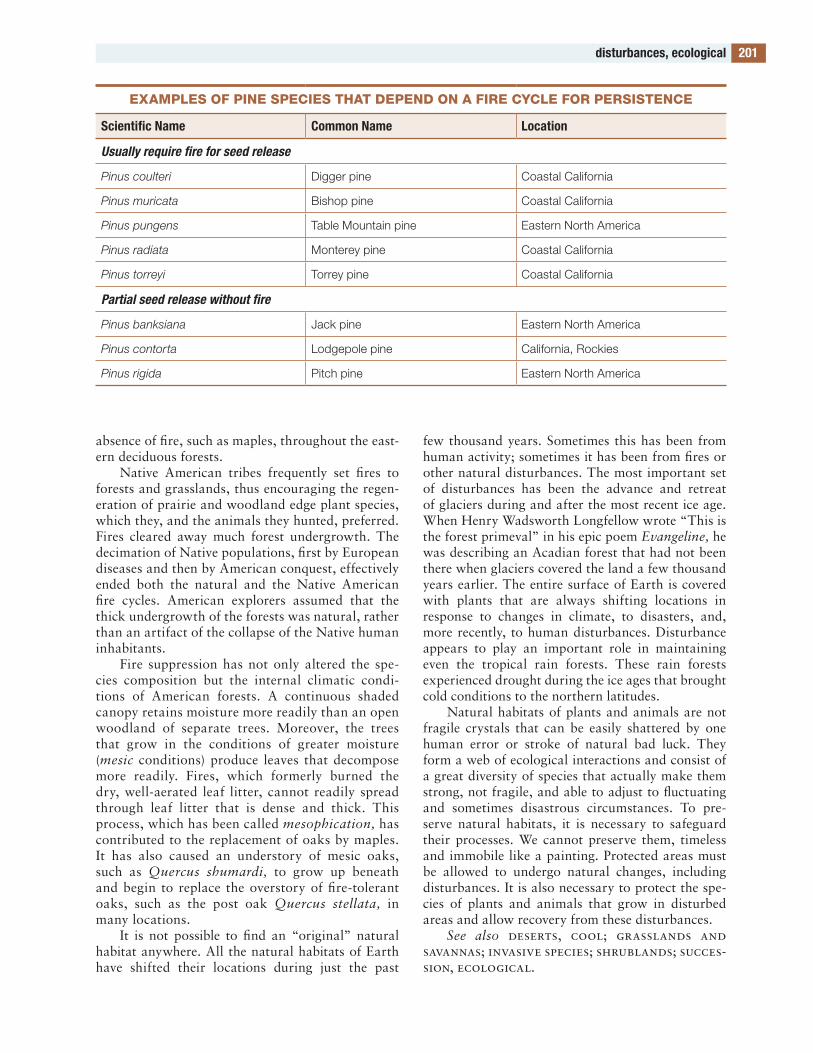
disturbances, ecological 201
absence of fire, such as maples, throughout the east-ern deciduous forests.
native american tribes frequently set fires to forests and grasslands, thus encouraging the regen-eration of prairie and woodland edge plant species, which they, and the animals they hunted, preferred. Fires cleared away much forest undergrowth. the decimation of native populations, first by european diseases and then by american conquest, effectively ended both the natural and the native american fire cycles. american explorers assumed that the thick undergrowth of the forests was natural, rather than an artifact of the collapse of the native human inhabitants.
Fire suppression has not only altered the spe-cies composition but the internal climatic condi-tions of american forests. a continuous shaded canopy retains moisture more readily than an open woodland of separate trees. Moreover, the trees that grow in the conditions of greater moisture (mesic conditions) produce leaves that decompose more readily. Fires, which formerly burned the dry, well-aerated leaf litter, cannot readily spread through leaf litter that is dense and thick. this process, which has been called mesophication, has contributed to the replacement of oaks by maples. it has also caused an understory of mesic oaks, such as Quercus shumardi, to grow up beneath and begin to replace the overstory of fire-tolerant oaks, such as the post oak Quercus stellata, in many locations.
it is not possible to find an “original” natural habitat anywhere. all the natural habitats of earth have shifted their locations during just the past
few thousand years. Sometimes this has been from human activity; sometimes it has been from fires or other natural disturbances. the most important set of disturbances has been the advance and retreat of glaciers during and after the most recent ice age. When Henry Wadsworth longfellow wrote “this is the forest primeval” in his epic poem Evangeline, he was describing an acadian forest that had not been there when glaciers covered the land a few thousand years earlier. the entire surface of earth is covered with plants that are always shifting locations in response to changes in climate, to disasters, and, more recently, to human disturbances. Disturbance appears to play an important role in maintaining even the tropical rain forests. these rain forests experienced drought during the ice ages that brought cold conditions to the northern latitudes.
natural habitats of plants and animals are not fragile crystals that can be easily shattered by one human error or stroke of natural bad luck. they form a web of ecological interactions and consist of a great diversity of species that actually make them strong, not fragile, and able to adjust to fluctuating and sometimes disastrous circumstances. to pre-serve natural habitats, it is necessary to safeguard their processes. We cannot preserve them, timeless and immobile like a painting. Protected areas must be allowed to undergo natural changes, including disturbances. it is also necessary to protect the spe-cies of plants and animals that grow in disturbed areas and allow recovery from these disturbances.
See also deserts, cool; grasslands and savannas; invasive species; shrublands; succes-sion, ecological.
examples of pine speCies that depend on a fire CyCle for persistenCe
Scientific Name Common Name location
Usually require fire for seed release
Pinus coulteri Digger pine Coastal California
Pinus muricata Bishop pine Coastal California
Pinus pungens Table Mountain pine Eastern North America
Pinus radiata Monterey pine Coastal California
Pinus torreyi Torrey pine Coastal California
Partial seed release without fire
Pinus banksiana Jack pine Eastern North America
Pinus contorta Lodgepole pine California, Rockies
Pinus rigida Pitch pine Eastern North America
xviii+598_Biodiversity_F.indd 201 11/17/11 1:12 PM

first entry202 dNa studies of biodiversity 202
Further readingBarbour, Michael G., et al. “Fire.” chapter 16 in Terrestrial
Plant Ecology. Menlo Park, calif.: addison Wesley Benjamin cummings, 1999.
Botkin, Daniel B. Discordant Harmonies: A New Ecology for the Twenty-first Century. new york: Oxford Uni-versity Press, 1990.
cartmill, M. “the Bambi syndrome.” Natural History (June 1993): 6–12.
Grogan, P., et al. “Below-ground ectomycorrhizal commu-nity structure in a recently burned bishop pine forest.” Journal of Ecology 88 (2000): 1,051–1,062.
Heckenberger, Michael J., et al. “Pre-columbian urbanism, anthropogenic landscapes, and the future of the ama-zon.” Science 321 (2008): 1,214–1,217. Summarized by Mann, charles c. “ancient earthmovers of the amazon.” Science 321 (2008): 1,148–1,152.
Keeley, Jon e. “Demographic structure of california chap-arral in the long-term absence of fire.” Journal of Veg-etation Science 3 (1992): 79–90.
———, and c. D. Fotheringam. “Smoke-induced seed germination in californian chaparral.” Science 276 (1997): 1,248–1,250.
likens, Gene e., et al. “effects of forest cutting and her-bicide treatment on nutrient budgets in the Hubbard Brook watershed-ecosystem.” Ecological Monographs 40 (1970): 23–47.
nowacki, Gregory J., and Marc D. abrams. “the demise of fire and the ‘mesophication’ of forests in the eastern United States.” BioScience 58 (2008): 123–138.
Rice, Stanley a. “Plants heal the landscape.” ch. 9 in Green Planet: How Plants Keep the Earth Alive. Piscataway, n.J.: Rutgers University Press, 2009.
thomas, Peter, and Rob Mcalpine. Fire in the Forest. new york: Oxford University Press, 2010.
West, neil e. “intermountain deserts, shrub steppes, and woodlands.” chapter 7 in Barbour, Michael G., and William Dwight Billings, eds. North American Ter-restrial Vegetation. new york: cambridge University Press, 1988.
Wheelwright, Jeff. “Fire in the sky: Why america’s ecologi-cal treasures sometimes just need to burn.” Discover (June 2006): 50–57.
Willis, K. J., and H. J. B. Birks. “What is natural? the need for a long-term perspective in biodiversity conserva-tion.” Science 314 (2006): 1,261–1,265.
dna studies of biodiversity Dna is the molecule, found in every cell, that encodes an organ-ism’s genetic information (genes). Most cells have noncoding Dna in addition to genes. Mutations in the Dna accumulate over time and alter the sequence of bases in the Dna. Usually, more muta-tions accumulate in noncoding Dna than in genes because natural selection may eliminate organisms
that have deleterious mutations in their genes. the result of the accumulation of mutations is that each species has a recognizable set of base sequences. in theory, the species of any organism could be deter-mined by identifying these base sequences.
it is much easier to identify a species of multicel-lular organisms by its physical characteristics than by its genes, because genetic identification involves extraction and analysis of Dna, which takes longer and is expensive. However, if genetic identification of individual organisms could be made quick and inexpensive, it would solve several problems in field work on biodiversity, such as the following.
● it often takes a well-trained expert to dis-tinguish among closely related species. to identify all the species in a habitat, the exper-tise of several scientists is needed, since each specializes on a certain set of organisms. the number of habitats that can be studied is thus limited by the number of available experts.
● Some organisms cannot readily be identified by physical characteristics—most notably, bacteria.
● Organisms often cannot be identified unless they are at a certain stage of their life cycle. Many plant species are difficult to iden-tify when they are not flowering, and fungi are often indistinguishable unless they are undergoing sexual reproduction.
new techniques offer the promise of rapid genetic identification of organisms. it is not neces-sary to analyze the base sequence of all, or even of very many, genes in order to identify a species. For animals, there is enough variation in the cytochrome oxidase i gene (coxi) to allow identification in most cases. For fungi, an internal transcribed spacer (itS), which is part of the noncoding Dna, can be used. For plants, two genes are used: neither the large sub-unit of the ribulose bisphosphate carboxylase gene (rbcl) nor the megakaryocyte-associated tyrosine kinase gene (matK) may be sufficient by themselves to identify a plant species, but when used together, they may work very well. Furthermore, quick and inexpensive ways of identifying the genetic variants of these genes are available and could conceivably be packaged for field use. a researcher would need only to know how to use the kit; the information could be compared with an online database from the field. currently, the equipment is expensive, but once it is in place, the newest equipment can identify a speci-men for as little as $2.
this approach, while imperfect, is quite reliable. in a 2006 study, it had a 98 percent success rate of distinguishing among 521 species of tropical butter-
xviii+598_Biodiversity_F.indd 202 11/17/11 1:12 PM

dNa studies of biodiversity 203
flies and moths. the two plant genes allow the spe-cies of a plant to be identified correctly 72 percent of the time; if the comparison is limited to plant species that are known to occur locally, identification is cor-rect nearly 100 percent of the time for seed plants. For seedless plants, identification is a bit less accu-rate. the rbcl and matK genes are found in chloro-plasts, while the coxi gene is found in mitochondria, both of which are inherited only from the female parent (seeds and eggs). as a result, hybrid organ-isms may not be recognized. Still, these sources of error are much less than what normally happens with mistaken identifications based on appear-ance. there will probably always be a necessity for voucher specimens (a specimen as an example of the species the researcher has claimed to identify) and the supervision of experts, but the experts do not need to do all the work. there is already a huge backlog of unsorted and unidentified field specimens awaiting identification in herbaria and museums around the world; one biologist estimated that, with experts working alone, it would take 50,000 years to identify all of them.
in some cases, genetic techniques have allowed new species to be identified. Organisms that had been assumed to be a single species may be several species, indistinguishable even by experts. this is especially the case with certain insects, in which the insects that have Wolbachia bacteria in their cells cannot interbreed with those that do not, resulting in a speciation event. On the other hand, these tech-niques may identify genetic variation within species as species-level differences, thus inflating the number of species that are identified.
Sometimes, scientists gather Dna specimens from entire ecological communities and then analyze it to determine the number of species the sample rep-resents. this approach, known as metagenomics, has been especially useful in sampling microbial commu-nities in the soil and in the oceans. it has also been used to estimate the diversity of multicellular organ-isms. in 2009, some scientists drove a vehicle for sev-eral hundred miles in the eastern United States and collected Dna from the bumper of the vehicle as an estimate of species diversity along the route. Besides finding bacteria, the researchers also collected insect Dna in this manner. Only 8 percent of the Dna fragments matched those of known sequences, sug-gesting that even in well-studied regions there are many unknown species.
the genetic approach has been called the “bar-code of life,” since the computerized sequence of bases looks like a barcode of colored bars, each color representing the base a, c, t, or G. the consortium for the Barcode of life (cBOl) is a scientific organi-zation that is advancing the development and appli-
cation of these techniques. the name “barcode of life” is also meant to reflect the same kind of ease of species identification as supermarket barcodes allow for item identification, pricing, and inventory. if the experts do not have to do all of the work, more habi-tats can be sampled more frequently, allowing not only more comprehensive biodiversity studies but the monitoring of changes in biodiversity over time. One researcher, the botanist Vincent Savolainen, raised the possibility that genetic barcoding may be “Star trek’s tricorder coming to reality.” this is, at pres-ent, an excessive expectation, but barcoding may prove revolutionary in studies of biodiversity.
See also natural selection.
Further readingchase, Mark W., and Michael F. Fay. “Barcoding of plants
and fungi.” Science 325 (2009): 682–683.———, et al. “land plants and Dna barcodes: Short-term
and long-term goals.” Philosophical Transactions of the Royal Society of London Biology 360 (2005): 1,889–1,895.
consortium for the Barcode of life, Smithsonian institu-tion. “What is cBOl?” available online. URl: http://www.barcoding.si.edu. accessed august 24, 2009.
Hajibabaei, Mehrdad, et al. “Dna barbodes distinguish species of tropical lepidoptera.” Proceedings of the National Academy of Sciences USA 103 (2006): 968–971.
Hebert, Paul D. n., et al. “ten species in one: Dna bar-coding reveals cryptic species in the neotropical skip-per butterfly Astraptes fulgerator.” Proceedings of the National Academy of Sciences USA 101 (2004): 14,812–14,817.
Jones, Matthew B., et al. “the new bioinformatics: inte-grating ecological data from the gene to the bio-sphere.” Annual Review of Ecology, Evolution, and Systematics 37 (2006): 519–544.
Kress, W. John, et al. “Use of Dna barcodes to identify flowering plants.” Proceedings of the National Acad-emy of Sciences USA 102 (2005): 8,369–8,374.
lahaye, Renaud, et al. “Dna barcoding the floras of biodi-versity hot spots.” Proceedings of the National Acad-emy of Sciences USA 105 (2008): 2,923–2,928.
Ratnasingham, Sujeevan, and Paul D. n. Hebert. “Barcod-ing of life Data System.” available online. URl: http://www.barcodinglife.org. accessed august 10, 2011.
Savolainen, Vincent, et al. “towards writing the encyclo-paedia of life: an introduction to Dna barcoding.” Philosophical Transcactions of the Royal Society of London Biology 360 (2005): 1,805–1,811.
Smith, M. alex, et al. “Dna barcodes affirm that 16 spe-cies of apparently generalist tropical parasitoid flies (Diptera, tachinidae) are not all generalists.” Proceed-ings of the National Academy of Sciences USA 104 (2007): 4,967–4,972.
xviii+598_Biodiversity_F.indd 203 11/17/11 1:12 PM

first entry204 dNa studies of biodiversity 204
Song, Hojun, et al. “Many species in one: Dna barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified.” Proceed-ings of the National Academy of Sciences USA 105 (2008): 13,486–13,491.
Spooner, David M. “Dna barcoding will frequently fail in complicated groups: an example in wild
potatoes.” American Journal of Botany 96 (2009): 1,177–1,189.
Venter, J. craig, et al. “environmental genome shotgun sequencing of the Sargasso Sea.” Science 304 (2004): 66–74.
xviii+598_Biodiversity_F.indd 204 11/17/11 1:12 PM

205
ecology ecology is the study of the relationships between organisms and their living and nonliving environments. ecology can be divided broadly into two areas: the relationship between an organism and its nonliving environment (autecology, from Greek auto-, meaning self) and the relationship between an organism and other organisms (synecology, from Greek syn-, meaning with).
auTEColoGyi. Energy balance of the Earth and organisms. every organism must maintain the proper balance of heat and chemical energy. an organism must remain within a relatively narrow range of tempera-ture if it is to survive and must obtain energy for its metabolism.
a. Energy balance of the Earth. the surface of the earth obtains almost all of its energy in the form of photons from the Sun. the clouds, ice, and light-colored rocks of the earth refl ect some of the sunlight, called albedo, back into outer space. albedo does not increase the temperature of the earth. the darker rocks, water, and organisms of the earth absorb sunshine. the sunshine increases the temperature of the earth and all its surface com-ponents, which results from increased movement (kinetic energy) of its molecules. the earth cannot conduct kinetic energy into outer space. the only way in which the earth can cool off is to radiate low-energy infrared photons into outer space. the Sun shines visible photons on one side of the earth; invisible photons radiate outwards in all directions from the earth.
Over time, the input and output of photons to and from the earth is in balance. the average temperature of the earth can temporarily fl uctu-
ate. Some of the infrared photons are absorbed by greenhouse gases such as carbon dioxide, methane, and water vapor, with the result that high concen-trations of these gases can cause the atmosphere to become warmer. this is the source of the greenhouse effect and is the reason that the earth was much warmer in the past than it is today. Methane reacts with oxygen in the atmosphere, producing carbon dioxide. Plants remove carbon dioxide from the air. in this way, photosynthesis prevents an excessive greenhouse effect from occurring. in fact, excessive photosynthesis may have been a contributing factor to worldwide glaciations that occurred during Pre-cambrian time.
B. Chemical energy in organisms. all organ-isms need chemical energy to build the complex molecules that make up their bodies and to run their metabolic reactions, which they use for the uptake of molecules, for growth, for movement, and for reproduction. Plants absorb photons of sunlight and transfer them to the potential energy of chemical bonds in sugar molecules. From these sugar mol-ecules, plants make all of their other molecules. animals obtain chemical energy by eating plants or by eating other animals that have eaten plants. When plants and animals die, decomposers obtain chemical energy from them. the result is the food chain or food web. the evolution of every organ-ism has been strongly infl uenced by its place in the food web. Since only 10 percent of the food energy is transferred from one step in the food chain to the next, the total weight of photosynthetic organisms is usually 10 times as great as the total weight of her-bivorous animals, which is 10 times as great as the total weight of carnivorous animals. Since 100 calo-ries of sugar in plants can support only 1 calorie of
E
xviii+598_Biodiversity_F.indd 205 11/17/11 1:12 PM

first entry206 ecology 206
carnivore, carnivores are often rare and have evolved adaptations that allow them to search for and pursue prey over a wide area.
c. Energy balance of organisms. Organisms obtain some of their warmth from sunshine and the warm air around them and the rest from their food. Organisms must also get rid of energy, thus main-taining an energy balance. the energy that origi-nally entered the organisms as photons or as food can be lost in three ways. First, organisms can radi-ate infrared photons. Warm-blooded animals radiate more infrared energy than do cold-blooded animals, plants, and their surroundings. Second, organisms can conduct kinetic energy into the air or water around them. third, plants and animals on dry land can evaporate water. a great deal of energy is required to evaporate water, therefore evaporation (transpiration from plants, perspiration from ani-mals) is a very efficient cooling mechanism.
the evolution of each species has been influenced by the necessity of maintaining energy balance. For example, bushes in deserts have evolved small leaves that allow them to disperse more of their heat load in the form of kinetic energy rather than the evapo-ration of water, which is rare in their habitats. at the same time, these bushes have often evolved deep roots, which maximize the amount of water they can obtain. Plants and animals from separate evo-lutionary lineages have often evolved similar adap-tations to these climatic conditions. animals need to increase their energy loss in hot environments and restrict it in cold environments. Warm-blooded animals have evolved thick layers of hair or fat to insulate them in cold environments. Hibernation is an evolutionary adaptation that allows animals to avoid excessive energy loss during winter.
the temperature and rainfall patterns of differ-ent parts of the earth are influenced by the move-ments of the earth, as well as the arrangement of the continents. Before the Pleistocene epoch, oce-anic currents carried warm water into north Polar regions, making them much warmer than they are today. the movement of continents subsequently closed off some of this circulation, causing the north Polar region to become permanently cold, and beginning a cycle of ice ages.
ii. Obtaining matter. Organisms not only need energy but also need matter. Photosynthesis removes carbon dioxide gas from the air or water and makes sugar from it. animals and decomposers obtain matter from the food chain at the same time and from the same sources where they obtain energy. all organisms, including plants, release carbon diox-ide gas back into the air or water. Organisms also need nitrogen and minerals such as phosphorus, potassium, and calcium. Plants obtain them mostly
from the soil by absorbing inorganic chemicals with their roots. Plants store some of these minerals and use others in the construction of their large organic molecules. animals and decomposers obtain miner-als from their food.
One corollary to the eternal cycling of mat-ter is that nothing ever goes away. Molecules may change form, as when exposure to oxygen causes an organic toxin to break down into harmless com-ponents. Some toxins are persistent, such as the slowly degrading pesticide DDt or the immortal element mercury. Humans often think that they can “throw things away” and forget about them, but one of the major slogans of ecology is “there is no such place as away.” this way of thinking was very evident during the 2010 Gulf oil spill. BP, with the guidance and permission of the federal government, used tons of dispersant chemicals (themselves of undetermined toxicity) to break the oil globules into invisibly small particles. they correctly assumed that people would not be concerned about petro-leum toxins that they could not see. Just as the laws of nature dictate that one cannot get something for nothing, they also dictate that one cannot get noth-ing from something.
SyNEColoGyi. Interactions within populations. a population is all of the individuals of a species that interact. Populations can grow exponentially because of the reproduction of the individuals that make it up. Spe-cies have evolved different reproductive systems that allow individuals to produce offspring successfully and disperse them into new locations. Populations also contain genetic variability, which is essential for natural selection. the fitness of an individual organism is its reproductive success relative to the other members of the population. individuals com-pete with one another not only for resources such as food and territory but also for opportunities to mate, resulting in sexual selection. Different popula-tions in a species can become new species. Practi-cally every aspect of evolution is influenced by the synecology of populations.
ii. Interactions between species. evolution has produced millions of species, each having com-plex interactions with many others. Species, just like individuals within species, can compete with one another. Plant species have evolved different ways of protecting themselves from herbivores, including thousands of kinds of toxic chemicals, and herbi-vores have evolved ways of getting around the plant defenses. Prey animals have evolved many ways of protecting themselves from predators, and predators have evolved many ways of finding and eating the prey. Many animals obtain their food by pollinat-
xviii+598_Biodiversity_F.indd 206 11/17/11 1:12 PM

ecosystems 207
ing the plants that have made use of those animals to transport their pollen. Many species have evolved very close relationships, called symbioses, in which the other species is the most important environmen-tal factor. Often, one species benefits but the other does not (parasitism), while in others one species benefits while the other is unaffected (commensal-ism). in many cases, symbioses have evolved toward the mutual benefit of both species (mutualism). Sym-bioses result from coevolution. in some cases, one species has evolved such a dependence on another that its individuality has been lost and the two have fused into one (symbiogenesis).
iii. Ecological communities. the total of all the interspecific interactions in a location is the ecological community. this is the context within which all evolutionary change occurs. evolution has produced a great diversity of species within each community (biodiversity). Biodiversity results from a balance between speciation and extinction. the movements of continents not only influence the cli-mate, as explained above, but also separate species and bring species together, allowing new interactions to occur and new biodiversity to evolve. When new islands emerge from the ocean, new species quickly evolve. Disturbances such as fire and storm occur within ecological communities, which are disastrous to some individuals but create openings that can be colonized by others. Species diversity has been much enhanced by the evolution of species that specialize in disturbed areas. individuals within species grow first and rapidly in disturbed areas but are usually later replaced by individuals in species that grow more slowly, a process known as ecological succes-sion. Many human activities create disturbances. the disturbances created by humans, however, may be occurring more rapidly than ecological communi-ties or the evolutionary process can handle, with the result that human activity may now be causing the sixth of the earth’s great mass extinctions.
Because all the species in ecological communi-ties interact, forming a complex network of effects, it is very difficult to predict the effects of a single ecological event. One cannot assume that pesticides will simply kill pests and leave everything else in the ecological community unaltered. the pesticides may have unintended victims, such as predators, with the paradoxical result that the pest populations may actually increase (a process known as resurgence). the toxins may concentrate in the food chain, a pro-cess known as biomagnification. this is the reason for the other major slogan of ecology, “you can’t do just one thing.”
iV. Ecosystems. an ecosystem incorporates the ecological community with the flow of energy and cycling of matter into a single system. ecosys-
tem ecology ties together autecology and synecology by considering both organisms and the nonliving environment as components of an interacting system.
at every level, from the energy balance of an individual organism to the interactions of all the species in the world, ecology is the exterior context within which all interior functions of organisms must occur and is the setting within which biodiver-sity develops.
See also biogeochemical cycles; bioge-ography; biomagnification; carbon cycle; coevolution; communities, ecological; dis-turbances, ecological; ecosystems; green-house effect; herbivory; hydrological cycle; mass extinctions; mutualism; nitrogen cycle; parasitism; pollination; population genetics; predation; speciation; succession, ecological; symbiogenesis.
Further readingBarbour, Michael G. and William Dwight Billings, eds.
North American Terrestrial Vegetation. new york: cambridge University Press, 1988.
Bazzaz, Fakhri a. Plants in Changing Environments: Link-ing Physiological, Population, and Community Ecol-ogy. new york: cambridge University Press, 1996.
Begon, Michael, colin R. townsend, and John l. Harper. Ecology: From Individuals to Ecosystems, 4th ed. Oxford, U.K.: Blackwell, 2006.
Hagen, Joel B. An Entangled Bank: The Origins of Ecosys-tem Ecology. new Brunswick, n. J.: Rutgers Univer-sity Press, 1992.
Harper, John l. “a Darwinian approach to plant ecology.” Journal of Ecology 55 (1967): 247–270.
Mayhew, Peter J. Discovering Evolutionary Ecology: Bringing Together Ecology and Evolution. Oxford, U.K.: Oxford University Press, 2006.
ecosystems ecosystems are the systems by which energy flows from the Sun, through the earth, and into outer space and by which matter cycles on the earth. ecosystems can be defined on a large scale (e.g., the entire earth) or on smaller scales (e.g., the tallgrass prairie ecosystem of north america or the cedar lake Bog ecosystem of Minnesota). Within ecosystems, the amount of energy and of each kind of atom or molecule (matter) within each ecosystem compartment and the transfer of energy and matter from one compartment to another are quantified. a compartment may be organisms with a certain eco-logical function (e.g., producers) or a certain stage of energy flow or cycling of matter (e.g., leaf litter). the concept of ecosystems incorporates ecological communities, which are the living organisms within ecosystems.
xviii+598_Biodiversity_F.indd 207 11/17/11 1:12 PM
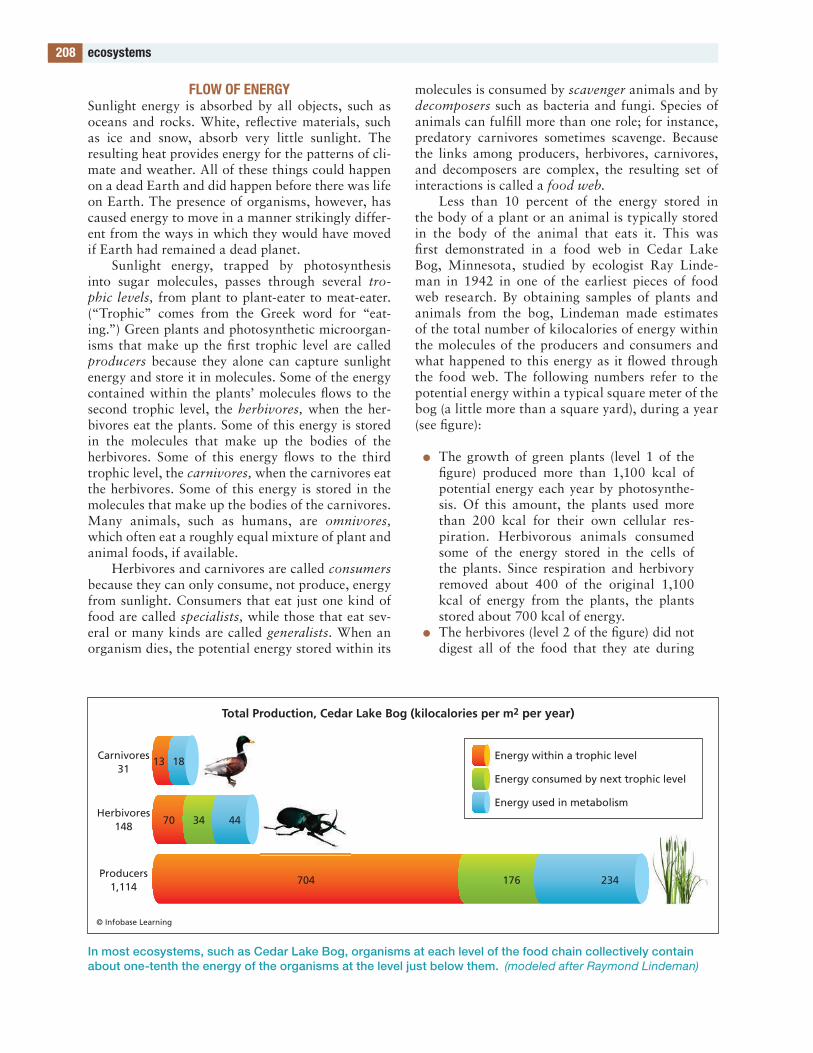
fi rst entry208 ecosystems 208
flow of ENErGySunlight energy is absorbed by all objects, such as oceans and rocks. White, refl ective materials, such as ice and snow, absorb very little sunlight. the resulting heat provides energy for the patterns of cli-mate and weather. all of these things could happen on a dead earth and did happen before there was life on earth. the presence of organisms, however, has caused energy to move in a manner strikingly differ-ent from the ways in which they would have moved if earth had remained a dead planet.
Sunlight energy, trapped by photosynthesis into sugar molecules, passes through several tro-phic levels, from plant to plant-eater to meat-eater. (“trophic” comes from the Greek word for “eat-ing.”) Green plants and photosynthetic microorgan-isms that make up the fi rst trophic level are called producers because they alone can capture sunlight energy and store it in molecules. Some of the energy contained within the plants’ molecules fl ows to the second trophic level, the herbivores, when the her-bivores eat the plants. Some of this energy is stored in the molecules that make up the bodies of the herbivores. Some of this energy fl ows to the third trophic level, the carnivores, when the carnivores eat the herbivores. Some of this energy is stored in the molecules that make up the bodies of the carnivores. Many animals, such as humans, are omnivores,which often eat a roughly equal mixture of plant and animal foods, if available.
Herbivores and carnivores are called consumersbecause they can only consume, not produce, energy from sunlight. consumers that eat just one kind of food are called specialists, while those that eat sev-eral or many kinds are called generalists. When an organism dies, the potential energy stored within its
molecules is consumed by scavenger animals and by decomposers such as bacteria and fungi. Species of animals can fulfi ll more than one role; for instance, predatory carnivores sometimes scavenge. Because the links among producers, herbivores, carnivores, and decomposers are complex, the resulting set of interactions is called a food web.
less than 10 percent of the energy stored in the body of a plant or an animal is typically stored in the body of the animal that eats it. this was fi rst demonstrated in a food web in cedar lake Bog, Minnesota, studied by ecologist Ray linde-man in 1942 in one of the earliest pieces of food web research. By obtaining samples of plants and animals from the bog, lindeman made estimates of the total number of kilocalories of energy within the molecules of the producers and consumers and what happened to this energy as it fl owed through the food web. the following numbers refer to the potential energy within a typical square meter of the bog (a little more than a square yard), during a year (see fi gure):
● the growth of green plants (level 1 of the fi gure) produced more than 1,100 kcal of potential energy each year by photosynthe-sis. Of this amount, the plants used more than 200 kcal for their own cellular res-piration. Herbivorous animals consumed some of the energy stored in the cells of the plants. Since respiration and herbivory removed about 400 of the original 1,100 kcal of energy from the plants, the plants stored about 700 kcal of energy.
● the herbivores (level 2 of the fi gure) did not digest all of the food that they ate during
In most ecosystems, such as Cedar Lake Bog, organisms at each level of the food chain collectively contain about one-tenth the energy of the organisms at the level just below them. (modeled after Raymond Lindeman)
xviii+598_Biodiversity_F.indd 208 11/17/11 1:12 PM

ecosystems 209
the year. they excreted part of it, and their digestive systems absorbed the remainder. the herbivores used some of the energy for their own cellular respiration. carnivores such as fishes consumed some of the herbi-vores and the energy contained within them. Since respiration and predation removed some of the original energy that was con-sumed by the herbivores, the herbivores stored about 70 kcal of potential energy.
● the carnivores (level 3 of the figure) did not digest all of the food that they ate. they excreted part of it, and their digestive sys-tems absorbed the remainder. the carnivores used some of the energy for their own cellu-lar respiration. the remaining 13 kcal were stored in the molecules of the carnivores.
the result has been described as an energy pyra-mid because the energy stored within the molecules of the organisms of one trophic level is usually less than 10 percent as much as the energy stored within the molecules of the organisms in the trophic level on which it feeds. it has also been described as the ten percent law for this reason. in the bog studied by lindeman, the energy in the molecules of the her-bivores is exactly 10 percent of the potential energy in the molecules of the producers, and the energy in the molecules of the carnivores is almost 20 percent of the energy in the molecules of the herbivores. in most food chains, a trophic level stores far less than 10 percent of the energy that is stored in the trophic level below it.
the organic molecules produced by plants in an ecosystem are collectively called primary pro-ductivity. the ecosystems with the greatest primary productivity are those with the greatest temperature and moisture; the ecosystems with the least primary productivity are cold or dry (see figure on page 210). in general, marine ecosystems have less primary productivity than do terrestrial ecosystems, largely because of the lack of nutrients in the waters of the deep oceans (see table).
in general, aquatic and terrestrial ecosystems dif-fer markedly in the flow of energy from one trophic level to another. the primary producers in aquatic systems are primarily phytoplankton, which are a very energy-rich food source. land plants, unlike phytoplankton, produce a lot of tissue such as wood that allows them to grow large. this tissue is not very nutritious to herbivores. this tissue also takes longer to decompose than do the bodies of phyto-plankton organisms. Moreover, some phytoplankton store energy in the form of oil, which allows them to float but which is also very nutritious to herbivores. therefore aquatic systems have a greater biomass of
herbivores and a lower mass of detritus and decom-posers. the rich source of primary production often allows aquatic food chains to have many more levels than terrestrial food chains. a typical aquatic food chain might be phytoplankton to small zooplankton to small arthropods to large arthropods to small fish to large fish to predatory bird, while a typical terres-trial food chain might be grass to rodent to predator.
the ten percent law explains how humans can use farmland more efficiently by eating plants than by eating meat from grain-fed animals. the United States is the world’s leading producer of agricul-tural crops. More than half of the grain raised in the United States and nearly 40 percent of the grain raised in the world are used as livestock feed rather than as human food. David Pimentel, an ecologist at cornell University, estimated in 1997 that the food produced in the United States could feed 800 million people if it were used to feed people directly, rather than to feed livestock. a strictly vegetarian diet could support about 10 times as many people as could a
average primary produCtivity of some eCosystems on earth, in kilograms dry Weight per
square meter per year
Tropical forests 2,000
Swamp and marsh 2,000
Reefs and estuaries 1,800
Temperate forests 1,250
Savanna 900
Boreal forests 800
Dry woods and shrublands 700
Cultivated land 650
Temperate grasslands 600
Continental shelves 360
Lakes and streams 250
Tundra 140
Open oceans 125
Deserts 40
Total continental 773
Total marine 152
World 333
*To convert to pounds per square foot per year, multiply by 0.2.Note: From Whittaker
xviii+598_Biodiversity_F.indd 209 11/17/11 1:12 PM

fi rst entry210 ecosystems 210
strictly carnivorous diet. therefore a larger number of people can receive adequate nutrition if all people ate relatively less meat. americans (who eat more than 90 pounds [40 kg] of beef per capita each year) consume a disproportionate share of scarce food resources. if americans ate less meat, a great deal of food energy might become available to the poor people of the world. it is not necessary to avoid meat totally in order to bring these benefi ts about.
For each 100 kcal of food energy in the live-stock feed, the animal uses between 30 and 60 kcal for its own metabolism; between 20 and 30 kcal of the energy is in the animal’s waste; the remain-der contributes to the growth of the animal. the effi ciency by which feed is converted into livestock growth is higher than the effi ciency of herbivores in the wild because the livestock feed has already been processed: the stems and roots of the crop plants have been removed. also, the animals do not need to expend much energy to locate and consume their food, since it is usually provided for them in a trough. consider, also, that humans do not consume very much of the animal’s body, eating primarily the muscles (meat). therefore, in reality, the effi ciency of energy in the agricultural food chain from farm to beef on the plate is very low.
the effi ciency of energy use is even lower when we consider that sunlight energy is not the only
energy used in producing meat: We also use a huge amount of fossil fuel to plant, fertilize, spray, har-vest, and process the grains and to maintain and process the livestock. Raising beef cattle requires 54 kcal of fossil fuel energy for each calorie of pro-tein that reaches the consumer; eggs require 26, pork requires 17, and turkey requires 13. the most energy-effi cient meat production is chicken, in which only 4 kcal of fossil fuel energy are needed for each kilocalorie of protein that reaches the consumer. these fi gures also do not take into consideration the tremendous expenditure of water needed for raising and processing the grains and livestock and the soil erosion that comes from farms and feedlots. these calculations apply only to livestock fed on grain that is raised on mechanized farms. Pastures are usu-ally located on land that is too poor or where water resources are too scarce for raising crops. cattle raised on pastures provide meat that is not at the expense of other food. also, in some cases, animal feed contains waste material that could not be con-sumed by humans. this practice, while enhancing the energy effi ciency of livestock production, can result in public health disasters. For example, the use of leftover animal parts, such as brains, in animal feed has allowed the spread of the prions that cause bovine spongiform encephalopathy (BSe, otherwise known as mad cow disease).
This map shows the global pattern of gross primary production (GPP, the total amount of photosynthesis) in grams of carbon per square meter per year. Tropical rain forests have the highest plant productivity. (Adapted from Beer, et al.)
xviii+598_Biodiversity_F.indd 210 11/17/11 1:13 PM
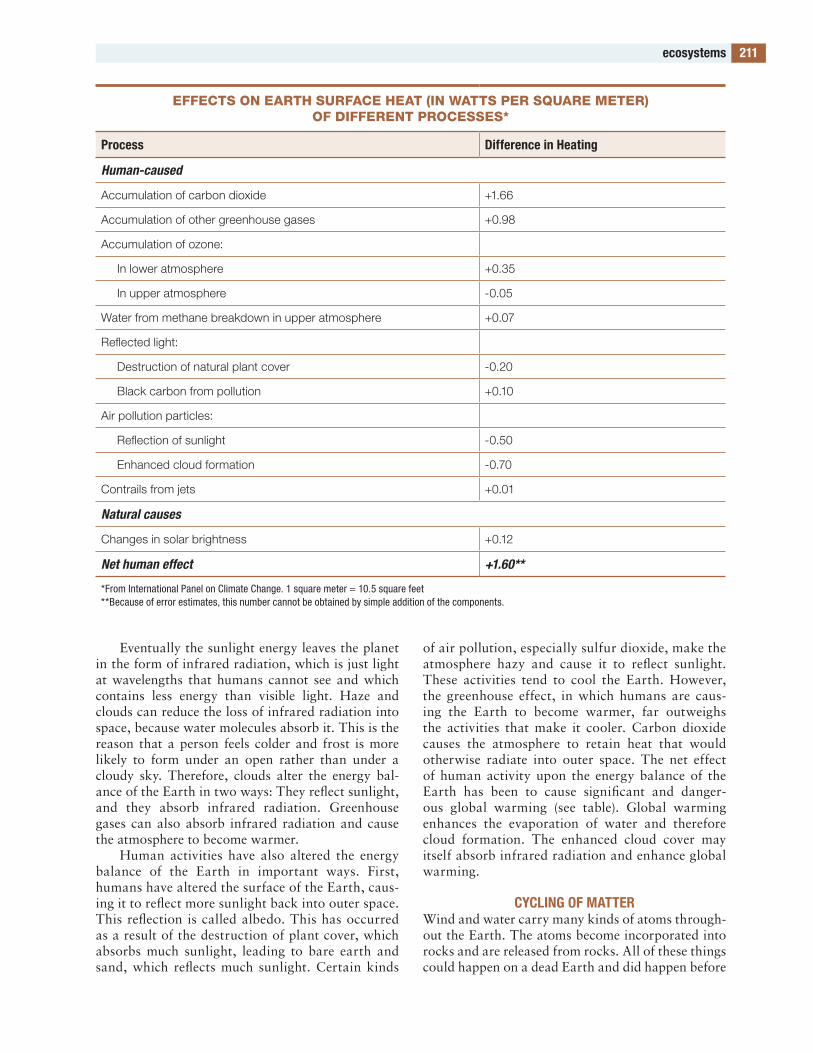
ecosystems 211
eventually the sunlight energy leaves the planet in the form of infrared radiation, which is just light at wavelengths that humans cannot see and which contains less energy than visible light. Haze and clouds can reduce the loss of infrared radiation into space, because water molecules absorb it. this is the reason that a person feels colder and frost is more likely to form under an open rather than under a cloudy sky. therefore, clouds alter the energy bal-ance of the earth in two ways: they reflect sunlight, and they absorb infrared radiation. Greenhouse gases can also absorb infrared radiation and cause the atmosphere to become warmer.
Human activities have also altered the energy balance of the earth in important ways. First, humans have altered the surface of the earth, caus-ing it to reflect more sunlight back into outer space. this reflection is called albedo. this has occurred as a result of the destruction of plant cover, which absorbs much sunlight, leading to bare earth and sand, which reflects much sunlight. certain kinds
of air pollution, especially sulfur dioxide, make the atmosphere hazy and cause it to reflect sunlight. these activities tend to cool the earth. However, the greenhouse effect, in which humans are caus-ing the earth to become warmer, far outweighs the activities that make it cooler. carbon dioxide causes the atmosphere to retain heat that would otherwise radiate into outer space. the net effect of human activity upon the energy balance of the earth has been to cause significant and danger-ous global warming (see table). Global warming enhances the evaporation of water and therefore cloud formation. the enhanced cloud cover may itself absorb infrared radiation and enhance global warming.
CyClING of maTTErWind and water carry many kinds of atoms through-out the earth. the atoms become incorporated into rocks and are released from rocks. all of these things could happen on a dead earth and did happen before
effeCts on earth surfaCe heat (in Watts per square meter) of different proCesses*
process difference in heating
Human-caused
Accumulation of carbon dioxide +1.66
Accumulation of other greenhouse gases +0.98
Accumulation of ozone:
In lower atmosphere +0.35
In upper atmosphere -0.05
Water from methane breakdown in upper atmosphere +0.07
Reflected light:
Destruction of natural plant cover -0.20
Black carbon from pollution +0.10
Air pollution particles:
Reflection of sunlight -0.50
Enhanced cloud formation -0.70
Contrails from jets +0.01
Natural causes
Changes in solar brightness +0.12
Net human effect +1.60**
*From International Panel on Climate Change. 1 square meter = 10.5 square feet**Because of error estimates, this number cannot be obtained by simple addition of the components.
xviii+598_Biodiversity_F.indd 211 11/17/11 1:13 PM

first entry212 ecosystems 212
there was life on earth. the presence of organisms, however, has caused atoms to move in a manner strikingly different from the ways in which they would have moved if earth had remained a dead planet.
carbon, hydrogen, oxygen, nitrogen, and min-eral atoms travel throughout the earth and through the food web. Because the chemical cycles have bio-logical and geological components, they are called biogeochemical cycles. When a green plant carries out photosynthesis or when an animal eats food, they are participating not only in the flow of energy through the earth but also in the cycling of matter on the earth. everything, matter as well as energy, goes somewhere.
Some kinds of atoms have an atmospheric component in their biogeochemical cycles. Plants remove carbon dioxide (cO2) from the air. Nitro-gen-fixing bacteria, some of them in the soil but most of them in the nodules of roots of many kinds of plants, remove nitrogen gas (n2) from the air spaces in the soil. the other kinds of atoms usually do not have a significant atmospheric component. Plants absorb phosphorus, potassium, calcium, iron, and sulfur atoms through their roots directly from the water solution found in soils. Plants obtain most of their nitrogen from soil solution ammo-nium and nitrates, rather than from nitrogen-fixing bacteria. carbon, oxygen, nitrogen, and mineral elements absorbed by plants are incorporated into biological molecules. For example, carbohydrates, proteins, lipids, and nucleic acids all contain car-bon and oxygen atoms; proteins and nucleic acids contain, in addition, nitrogen atoms; nucleic acids also have phosphorus atoms, and proteins have sulfur atoms. Organisms use calcium primarily for structural uses: between plant cell walls and within the bones of animals. Plants use potassium, and animals use both sodium and potassium, for water balance of cells. cells use iron for electron trans-port in metabolism, and animals use iron in hemo-globin. When organisms die, decomposers release the molecular components: cO2 goes back into the atmosphere, while nitrates, phosphates, calcium, potassium, and other minerals enter the soil water solution. Denitrifying bacteria change some of the nitrogenous minerals back into n2.
Water molecules cycle through the ecosystems of the earth (the hydrological cycle). Unlike the atoms and molecules of most of the other cycles, water mol-ecules are not chemically altered in the hydrologic cycle.
carbon, oxygen, nitrogen, and minerals cycling through the ecosystem constitute a short-term cycle. atoms and molecules in short-term cycles transfer from one compartment to another in periods of
time as short as a few days to a few centuries. if these atoms and molecules become trapped in sedi-ments, they may become components of rocks, from which they are released only when the rocks erode. these atoms and molecules therefore participate in a long-term cycle. Humans, like other animals, obtain nutrition from many atoms and molecules in food and energy and economically valuable materi-als from recently produced organic materials such as wood. in these ways, humans participate in short-term cycles. Humans also obtain materials (such as petroleum and natural ores) for all other activities from biogeochemical cycles. in many cases, human activities deplete resources that have accumulated by the action of long-term cycles. Materials that have accumulated by long-term cycles are called nonre-newable resources because their renewal requires millions of years. Serious economic and ecological problems can be avoided if humans adjust their eco-nomic activities to a balanced participation in short-term cycles, for example by use of renewable energy sources and recycling of material resources.
While it is easy enough to weigh organisms and to determine how much carbon or nitrogen is in them and therefore to reconstruct a generalized food web, it may be very difficult to determine the complex pathways by which the carbon or nitrogen moves through the food web. in some cases, investi-gators trace the movement of stable natural isotopes through food webs. One new method is to introduce nanoparticles into a food web and observe where they go.
Most organic matter that moves from one tro-phic level to another is either released as carbon dioxide or used by decomposers, resulting in a ten percent law for matter not very different from the one for energy. Some organic molecules, however, are persistent, which means they are not metabo-lized by organisms. Many of them are pollutants produced by humans. these persistent organic pol-lutants (POPs) therefore undergo biomagnification, in which they are highly concentrated in the tissues of animals near the top of the food chain.
no ecosystem, aside from the entire earth, is discrete from others. lindeman chose to study a bog because bogs are largely distinct from surround-ing ecological communities. Some ecosystems are strongly influenced by others. the deep ocean floors are almost entirely dependent on organic material floating down from the surface. Pond ecosystems may depend strongly on detritus from surrounding forests, and terrestrial animals may depend strongly on prey from the ponds. the boundaries between ecosystems are not, however, arbitrary. each ecosys-tem, such as a forest or a pond, consists mostly of a self-reinforcing network of ecological interactions
xviii+598_Biodiversity_F.indd 212 11/17/11 1:13 PM

ecosystem services 213
among its species. these interactions may help rein-force what might otherwise be an arbitrary bound-ary. Boreal forest trees, for example, do not merely live in places that are a little warmer than the tun-dra; the trees absorb sunlight and melt the snow, a process by which the boreal forest makes itself warmer than the nearby tundra.
See also biogeochemical cycles; biomagni-fication; carbon cycle; communities, ecologi-cal; ecology; greenhouse effect; nitrogen cycle.
Further readingarím, Matias, et al. “Food web structure and body size:
trophic position and resource acquisition.” Oikos 119 (2010): 147–153.
Becker, D. S., and G. n. Bigham. “Distribution of mer-cury in the aquatic food web of Onondaga lake, new york.” Water, Air, and Soil Pollution 80 (1995): 563–571.
Beer, christian, et al. “terrestrial gross carbon dioxide uptake: Global distribution and covariation with cli-mate.” Science 329 (2010): 834–838.
Belgrano, andrea, et al., eds. Aquatic Food Webs: An Eco-system Approach. Oxford, U.K.: Oxford University Press, 2005.
cabana, Gilbert, and Joseph B. Rasmussen. “comparison of aquatic food chains using nitrogen isotopes.” Pro-ceedings of the National Academy of Sciences USA 93 (1996): 10,844–10,847.
Doucett, Richard R., et al. “Measuring terrestrial subsidies to aquatic food webs using stable isotopes of hydro-gen.” Ecology 88 (2007): 1,587–1,592.
Golley, Frank Benjamin. A History of the Ecosystem Con-cept in Ecology: More Than the Sum of the Parts. new Haven, conn.: yale University Press, 1993.
Holbrook, R. David, et al. “trophic transfer of nanopar-ticles in a simplified invertebrate food web.” Nature Nanotechnology 3 (2008): 352–355.
lindeman, Raymond e. “trophic-dynamic aspect of ecol-ogy.” Ecology 23 (1942): 399–418.
Pauly, Daniel, et al. “Fishing down aquatic food chains.” American Scientist 88 (2000): 46–52.
Persson, a., et al. “effects of enrichment on simple aquatic food chains.” American Naturalist 157 (2001): 654–669.
Rice, Stanley a. Green Planet: How Plants Keep the Earth Alive. new Brunswick, n.J.: Rutgers University Press, 2009.
Shurin, Jonathan B., et al. “all wet or dried up? Real dif-ferences between aquatic and terrestrial food chains.” Proceedings of the Royal Society of London B (Biol-ogy) 273 (2006): 1–9.
Solomon, Susan, and the intergovernmental Panel on cli-mate change. Climate Change 2007: The Physical Science Basis. available online. URl: http://www.ipcc.
ch/publications_and_data/ar4/wg1/en/contents.html. accessed July 31, 2010.
Whittaker, Robert H. Communities and Ecosystems, 2nd ed. new york: Macmillan, 1975.
ecosystem services the ecosystems of the earth provide many services to the human economy, free of charge. this is not their purpose. all of the individuals of each species pursue their own fitness. But as trees (for example) photosynthesize, grow roots, and transpire, they end up doing many things upon which human life and the human economy depend.
nearly everyone has at least a vague under-standing that, without the natural world, human life would be impossible, and that humans extract many resources from the living world of nature that have great economic value. the ecosystem services approach differs from this widely accepted view in these major ways:
● First, it replaces a vague awareness of the value of nature with actual monetary values. the approach usually taken is to calculate how much it would cost for an ecosystem service, such as the production of oxygen by plants, to be replaced by an industrial process.
● Second, it does not rely solely on the mon-etary value of extracted resources (such as the value of the timber in a forest) but on all of the services that the ecosystem provides.
as a result of ecosystem services calculations, it becomes obvious that an intact ecosystem is worth much more, even in strictly monetary terms, than the resources that would come from the destruc-tion of the ecosystem. For example, the ecosystem services of a forest exceed the value of the timber by many orders of magnitude. the results are nearly always astonishing. the calculations apply to wild ecosystems or, in urban areas, reconstructed ecosys-tems such as parks.
ecosystem services, like ecosystem processes, are interconnected and difficult to isolate from one another. Most of these services result from the growth of plants. this should not be surprising since animals (and to a lesser extent fungi and bacteria) perform metabolic roles similar to those of humans, which are to that extent competitors rather than beneficiaries. in other words, most wild animals receive the same ecosystem services from plants that humans receive.
Humans, however, are more dependent on eco-system services than is any wild animal species,
xviii+598_Biodiversity_F.indd 213 11/17/11 1:13 PM

first entry214 ecosystem services 214
because humans depend upon a complex economy which, in turn, depends to a greater extent on envi-ronmental stability than do the lives of other animal species. Fires burn the aerial portions of plants and chase away the animals; afterward, the plants resprout and the animals return. the same fire will probably cause millions of dollars of damage to humans. Hunters and gatherers had many of the options, such as migrating to a new location, which wild animals have, but which civilized humans, liv-ing in a thick landscape of land ownership, do not.
ExamplE of ECoSySTEm SErvICESHere are some processes of intact wild ecosystems that happen to provide benefits to humans, which would take billions of dollars to provide artificially:
● Oxygen production. Photosynthetic organ-isms release 17.3 million tons (15.7 million metric tons) of oxygen into the atmosphere each year.
● Carbon dioxide removal. Photosynthetic organisms absorb about 115 million tons (105 million metric tons) of carbon diox-ide from the atmosphere each year. carbon dioxide is the principal long-term cause of the greenhouse effect.
● Shade. trees absorb the heat from sun-light, which they transfer into water vapor. the shade of the trees is therefore cooler than the atmosphere above unshaded land. though the shade of a tree may not make air-conditioning unnecessary in a small shaded building, it will reduce the amount of air conditioning that is needed. Shade saves millions of dollars of air-conditioning costs in the United States each year.
● Filtering pollution. trees reduce the amount of many pollutants in the atmo-sphere of cities, because the pollutants adhere to leaf surfaces and same leaves neu-tralize the pollutants.
Services provided by wild ecosystems provide at least $33 trillion dollars
of benefits to the human economy each year. Examples of such ecosystem ser-vices are plants putting oxygen into and removing carbon dioxide from the atmo-sphere; forests controlling floods and allowing rain to percolate into the soil and recharge aquifers; and wild plants not only holding down but building up the soil. In every case, the services provided by intact natural ecosystems (such as forests) vastly overshadow the immedi-ate profit to be gained by extraction of resources (such as timber) from them.
One problem with ecosystem ser-vices calculations is that they often do not convince narrow corporate interests to treat the natural world as a sustainable rather than as an extractive resource. The numerous benefits of an intact forest have tremendous value to the people who live near the forest but will not increase the profit margin of a timber corporation. To such a corporation, the lumber will always be worth more than the forest. It is gov-ernments, to the extent that they reflect
the welfare of the people, that can make the most use of ecosystem services calcu-lations. This is because it is governments that have to clean up the mess from, for example, mudslides and floods following deforestation, after the timber corporation has cut the trees and moved on. One of the few ways in which the corporations can have an economic interest in ecosystem services is if governments hold them liable for the disruption of these services.
The Gulf oil spill, following the explo-sion of the Deepwater Horizon drilling rig on April 19, 2010, was the greatest envi-ronmental disaster to that point in Ameri-can history. Initially, much of the cost was borne by Gulf Coast residents and the United States government, but the government insisted that BP, the com-pany whose contractors operated the rig, establish a $20 billion fund to pay all of the direct environmental costs. BP com-plied with this demand, and most citizens and politicians of both major political par-ties agreed that it was fair, except for Representative Joe Barton (Republican of Texas) who considered it to be federal
oppression of the corporation. Without government intervention, almost the entire cost of the oil spill would have been borne by the government and citizens, while BP would have lost only the money it spent on the oil rig itself.
Another way in which private corpora-tions may take an interest in ecosystem services is when insurance companies (and the reinsurers that insure them) face the possibility of having to pay for the mess caused by an environmental disaster. It is the insurance companies that are becom-ing concerned with the effects, already beginning to be felt, of global warming. In the short run, they can refuse to issue poli-cies to people along the Gulf Coast, due to hurricanes, oil spills, and rising sea levels. In the long run, as the effects of global warming become more pervasive and worldwide, insurance corporations cannot refuse coverage to very many more people or else they will have very little business.
Moreover, the calculated costs of envi-ronmental damage almost never include indirect costs, such as greenhouse effect. The 2010 BP blowout released huge
“ecoSySTeM ServiceS: puBlic or privATe coSTS?”
xviii+598_Biodiversity_F.indd 214 11/17/11 1:13 PM

ecosystem services 215
● Flood prevention. trees slow down the impact of rainfall that reaches the soil. this has two effects. First, trees help to prevent runoff that would cause flooding and mud-slides. Second, the rain is more likely to percolate into the soil if the leaves of trees, shrubs, and grasses slow it down.
● Groundwater recharge. Much of the water that percolates into the soil recharges the water supplies in aquifers, which is where most people obtain water.
● Erosion prevention. By slowing down the wind and the rain at the soil surface, plants reduce the loss of soil by wind and water erosion. it costs a lot of money to compen-sate for topsoil loss by applying commercial fertilizers and, in severe cases, compensa-tion cannot be made at all.
● Pest control. agricultural pests and the pests that spread human diseases are seldom species that live primarily in the wild. Many
wild animal species help to control pests. examples include birds and bats, which col-lectively consume billions of pest insects, and wasps, which parasitize and kill them.
● Pollination. Honeybees (Apis mellifera) carry out much agricultural pollination, but wild ecosystems also provide many pollina-tors, and honeybees may rely on wild nec-tar sources during times when agricultural sources are not available.
● Sources of agricultural germplasm. Wild relatives of crop plants are important sources of genes that plant breeders can use to produce disease-resistant crop varieties. an entire entry in this encyclopedia explores the value of biodiversity for agriculture.
● Sources of pharmaceuticals. Wild plants and animals are important sources for the discovery of pharmaceutical compounds. an entire entry in this encyclopedia explores the value of biodiversity for medicine.
amounts of methane into the atmosphere, where it is no longer visible and can be ignored by most people. But methane and the carbon dioxide produced from it con-tribute substantially to greenhouse gases. Also, the blowout has put pressure on American society to become even more heavily dependent on fossil fuels. At the very time that the oil spill was gushing unabated into the Gulf, most Americans remained in favor of continued oil drill-ing in the deep water at the edges of the continental shelves. None of these greenhouse-related costs are reflected in the $20 billion escrow account that the government required BP to establish for the cleanup. If the full indirect costs of the greenhouse effect are taken into account for environmental disasters, they would overwhelm all private profits. Such calcu-lations will never happen.
And these are only the costs associ-ated with disasters, rather than with the slow, persistent depletion of the func-tions of wild ecosystems. In a few cases, corporations have made contributions to private conservation organizations. It is safe to say that, aside from such volun-tary gifts, almost none of the $33 trillion is balanced by corporations setting aside money for environmental restoration.
Corporations generally do not consider it worthwhile to invest in natural capital—that is, the monetary value of nature. The people who depend directly on natural ecosystems for their survival—for exam-ple, tribal farmers who have to supple-ment their diet with wild game—have essentially no economic power in their countries. It is the poorest people and the poorest countries that will suffer most from environmental depletion and from the greenhouse effect.
In the valuation of ecosystem ser-vices, as in so many other ways, the environment is a medium through which people transmit altruism, or injustice, to one another. Furthermore, it is routine for politicians to make lofty statements about the kind of world they are leaving for their grandchildren, while assigning a value of essentially zero to the kind of environment in which their grandchildren will live (a process known as future dis-counting). Most people do not care that future generations will resent the present generation for depleting the ecosystem services of the planet, and even fewer are willing to invest money specifically to make a better world for them.
See also ecosystem services; environ-mental etHics.
Further readingChaddock, Gail Russell. “With Joe Barton’s
BP apology, Republicans’ point lost in oblivion.” Christian Science Moni-tor, June 18, 2010. Available online. URL: http://www.csmonitor.com/USA/Politics/The-Vote/2010/0618/With-Joe-Barton-s-BP-apology-Republicans-points-lost-in-oblivion. Accessed August 7, 2011.
Costanza, Robert, et al. “The value of the world’s ecosystem services and natural capital.” Nature 387 (1997): 253–260.
Davidson, Eric. You Can’t Eat GNP: Economics as if Ecology Mattered. New York: Basic Books, 2001.
Hawken, Paul, Amory Lovins, and L. Hunter Lovins. Natural Capitalism: Creating the Next Industrial Revolution. New York: Hachette, 2008.
Rehmeyer, Julie. “ ‘Discounting’ the future cost of climate change.” Science News, May 21, 2010. Available online. URL: http://costingthefuture.org/?p=66. Accessed August 7, 2011.
Rice, Stanley A. Green Planet: How Plants Keep the Earth Alive. New Brunswick, N.J.: Rutgers, 2009.
Roodman, David Malin. The Natural Wealth of Nations: Harnessing the Market for the Environment. New York: Norton, 1998.
xviii+598_Biodiversity_F.indd 215 11/17/11 1:13 PM
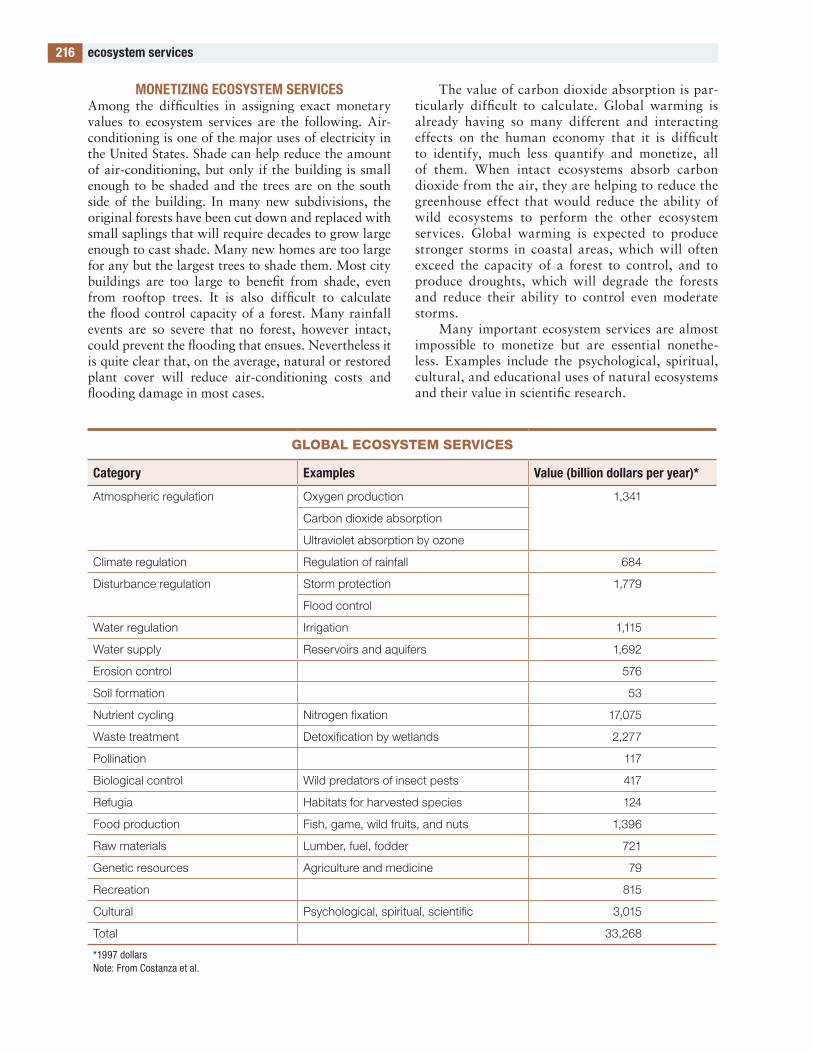
first entry216 ecosystem services 216
moNETIzING ECoSySTEm SErvICESamong the difficulties in assigning exact monetary values to ecosystem services are the following. air-conditioning is one of the major uses of electricity in the United States. Shade can help reduce the amount of air-conditioning, but only if the building is small enough to be shaded and the trees are on the south side of the building. in many new subdivisions, the original forests have been cut down and replaced with small saplings that will require decades to grow large enough to cast shade. Many new homes are too large for any but the largest trees to shade them. Most city buildings are too large to benefit from shade, even from rooftop trees. it is also difficult to calculate the flood control capacity of a forest. Many rainfall events are so severe that no forest, however intact, could prevent the flooding that ensues. nevertheless it is quite clear that, on the average, natural or restored plant cover will reduce air-conditioning costs and flooding damage in most cases.
the value of carbon dioxide absorption is par-ticularly difficult to calculate. Global warming is already having so many different and interacting effects on the human economy that it is difficult to identify, much less quantify and monetize, all of them. When intact ecosystems absorb carbon dioxide from the air, they are helping to reduce the greenhouse effect that would reduce the ability of wild ecosystems to perform the other ecosystem services. Global warming is expected to produce stronger storms in coastal areas, which will often exceed the capacity of a forest to control, and to produce droughts, which will degrade the forests and reduce their ability to control even moderate storms.
Many important ecosystem services are almost impossible to monetize but are essential nonethe-less. examples include the psychological, spiritual, cultural, and educational uses of natural ecosystems and their value in scientific research.
global eCosystem serviCes
Category Examples value (billion dollars per year)*
Atmospheric regulation Oxygen production 1,341
Carbon dioxide absorption
Ultraviolet absorption by ozone
Climate regulation Regulation of rainfall 684
Disturbance regulation Storm protection 1,779
Flood control
Water regulation Irrigation 1,115
Water supply Reservoirs and aquifers 1,692
Erosion control 576
Soil formation 53
Nutrient cycling Nitrogen fixation 17,075
Waste treatment Detoxification by wetlands 2,277
Pollination 117
Biological control Wild predators of insect pests 417
Refugia Habitats for harvested species 124
Food production Fish, game, wild fruits, and nuts 1,396
Raw materials Lumber, fuel, fodder 721
Genetic resources Agriculture and medicine 79
Recreation 815
Cultural Psychological, spiritual, scientific 3,015
Total 33,268
*1997 dollarsNote: From Costanza et al.
xviii+598_Biodiversity_F.indd 216 11/17/11 1:13 PM

Ediacaran organisms 217
the difficulty of making these calculations has not stopped economists from trying. economist Rob-ert costanza and colleagues have estimated that the total global value of 17 ecosystem services is $33 tril-lion (in 1997 U.S. dollars), which is almost double the total global gross domestic product ($18 trillion) (see table on page 216). On a smaller scale, new york city Parks commissioner adrian Benepe, using a computer program developed at the University of california, Davis, estimates that the 592,130 trees in new york public areas generate $5.60 in savings for each $1.00 spent to plant and maintain them.
the value of ecosystem services is calculated only on the basis of renewable production. accu-mulated past production (such as coal and oil) are not counted. However, even oil can be considered an ecosystem service, to a very limited extent. in the Gulf of Mexico, organic material continues to accu-mulate and, after a long time, is compressed into oil.
Of most immediate concern to most people is not so much the value of ecosystem services as the costs that are necessary to mitigate disasters that damage them. it will be a long time before the total costs of the Gulf oil spill of 2010, the biggest envi-ronmental disaster in american history, are known. the $20 billion that the federal government required BP to set aside for the cleanup is almost certainly not going to be enough. and this is only for the imper-fect cleanup of an ecosystem.
See also essay: “ecosystem Services: Pub-lic or Private costs?”; biodiversity, resources for agriculture; biodiversity, resources for medicine; biogeochemical cycles; ecosys-tems; environmental psychology; greenhouse effect.
Further readingcostanza, Robert, et al. “the value of the world’s ecosys-
tem services and natural capital.” Nature 387 (1997): 253–260.
Randall, David K. “Maybe only God can make a tree, but only people can put a price on it.” New York Times (april 18, 2007). available online. URl: http://www.nytimes.com/2007/04/18/nyregion/18trees.html?pagewanted=print. accessed May 13, 2011.
Rice, Stanley a. Green Planet: How Plants Keep the Earth Alive. new Brunswick, n. J.: Rutgers University Press, 2009.
ediacaran organisms the ediacaran organ-isms were the first notable diversification of multicel-lular life on earth. they lived immediately before the cambrian period, which began about 540 mil-lion years ago. the abundance of multicellular forms after the cambrian explosion contrasts sharply with
the relatively few multicellular forms represented by the ediacarans.
the earliest ediacarans lived about 580 million years ago. these marine organisms may have had a worldwide distribution, as their fossils are found in 24 locations on six continents, including australia (especially the ediacara Hills after which they are named), newfoundland, and namibia.
Reginald Sprigg, an australian geologist, was investigating mines in the ediacara Hills in 1946. While eating lunch, he turned over a rock. He was not expecting to find any fossils, since the rocks were Precambrian in age, and at the time no multicellular organisms were known from before the cambrian explosion. But he saw many leaflike impressions in the rock. He reported his findings at meetings, but they were rejected for publication. then in 1957, John Mason, a student, found a fossil in england, also of Precambrian age, that closely resembled Sprigg’s aus-tralian fossil. now that these fossils were being found more than once and in such diverse places, they were taken seriously. Sprigg’s priority as discoverer has been recognized by calling them ediacaran fossils.
nobody is entirely sure what these organisms were. Some of them bear at least a superficial resem-blance to existing kinds of marine invertebrates. For example, Tribrachidium, Arkarua, and Albumares resemble modern echinoderm “sand dollars”; Maw-sonites resembles a modern cnidarian sea anemone; Spriggina and Bomakellia resemble the arthropod trilobites that were abundant in oceans until about 250 million years ago; and Charniodiscus and Pteri-dinium resemble modern sea pens, all of them inver-tebrate animals known from the cambrian period. these resemblances are often pointed out because it is reasonable to expect the immediate ancestors of cambrian animals to have lived during the edia-caran period.
there are numerous puzzling differences between the ediacaran organisms and the inverte-brate animals they resemble. in particular, the edia-carans are not known to have had legs or mouths. While many invertebrate animals lack legs, none lack mouths. if ediacarans were animals, how and what did they eat?
another difficulty in identifying ediacarans as animals related to modern invertebrates is the question of whether the modern phyla of animals were yet in existence. Molecular studies suggest that the diversification of the modern groups of inver-tebrates had already begun. Precambrian fossils of what are clearly animal embryos and what were probably bilaterally symmetrical animals have been found. these observations suggest that some mod-ern groups of invertebrates were in existence during ediacaran times.
xviii+598_Biodiversity_F.indd 217 11/17/11 1:13 PM

first entry218 endangered species 218
Some researchers claim that the ediacarans had to be animals because they moved. in several exam-ples, ediacaran fossils are found in groups of four: three depression fossils and one with positive relief. the interpretation given in this case is that the edia-caran organism flopped around four times, leaving three prints in the bottom of the sea where they lived; the fourth, positive structure is the fossil of the organism itself. However, this does not prove that they moved themselves; they may have been moved by ocean currents.
Other researchers believe that the ediacarans were organisms that are completely unrelated to modern animals; in fact, they may not have been animals at all. consider, for example, the Spriggina fossil. to human observers it looks like a worm or trilobite with a head. But if an observer turns it upside down he or she may see a seaweedlike struc-ture with a holdfast to anchor it to the bottom of the ocean. Researchers generally accept that animals were present at that time. Fossilized burrows and other trace fossils (evidence of things the organisms did, rather than the organisms themselves) have been found, although some of the burrows may have been caused by single-celled protists rolling through the mud. this does not prove that the ediacarans were the animals that left the traces.
although red algae are known from 1.2 billion years ago, long before the ediacaran organisms, the ediacarans possess no structures that identify them as seaweeds, just as they possess no struc-tures clearly identifying them as animals. Research-ers such as paleontologists Dolf Seilacher and Mark McMenamin have suggested that the ediacarans were actually symbiotic combinations of protist cells, some of which were photosynthetic. Many symbi-otic combinations of plantlike and animallike protist cells are known from the modern world, such as Ophrydium, a protist colony that lives underwater. the symbiosis would explain how the ediacarans obtained food without having mouths: the plantlike cells made food from sunlight. it might even explain why many of these fossils were sometimes found to be filled with sand: they contained sand, as ballast, while alive, which would have been inconvenient for an animal, but perfectly workable for symbiotic bags of cells. alternatively, these organisms could have lived by absorbing dissolved organic materials. they may even have been large single-celled protists, similar to the modern oceanic xenophyophores. in 1992, Dolf Seilacher proposed that the ediacaran organisms were a separate kingdom of life, which he called Vendobionta.
it is possible that some of the ediacarans were members of modern invertebrate phyla, while some were not. after all, the resemblance of Charniodis-
cus to the cambrian cnidarian Thaumaptilon and to modern sea pens is very close. Sea pens, like many other cnidarians, form symbiotic associations the way many researchers claim the ediacarans did. therefore, some ediacarans may have been cnidar-ians but also had endosymbiotic algae living in their tissues, just as many coelenterates such as jellyfish and corals do today.
One of the major features of the ediacarans is that they had no hard shells, spines, or other defenses against predators. the implication is that predatory animals had not yet evolved. Once preda-tors evolved, the ediacarans may have been driven to extinction by them; only the hard-shelled animals or chemically defended seaweeds could survive. it has been suggested that the appearance of the first pred-ators was the stimulus for the cambrian explosion.
ediacaran organisms were the first to become prominent after prolonged periods of global cool-ing that have been called Snowball earth. a long period of earth history, in which (aside from some seaweeds) most organisms were colonies of single cells (such as stromatolites), came to an end with the global spread of ice on land and over most of the oceans as well. Organisms survived only underneath a few openings in the ice. it was in the rapid and pos-sibly severe period of global warming that followed Snowball earth that multicellular organisms, first the ediacarans, then the cambrian organisms, evolved.
See also animals; cambrian explosion; cambrian period, biodiversity in.
Further readingconway Morris, Simon. The Crucible of Creation: The
Burgess Shale and the Rise of Animals. new york: Oxford University Press, 1998.
McMenamin, Mark a. S. The Garden of Ediacara: Discov-ering the First Complex Life. new york: columbia University Press, 1998.
endangered species endangered species are species that are at risk of extinction in their natural habitats. Other than the convention on interna-tional trade in endangered Species of Wild Fauna and Flora (citeS), there are few international pro-tections for endangered species or even international standards for what constitutes an endangered spe-cies. the international Union for conservation of nature (iUcn), an environmental network that brings together more than 1,000 governments and nGOs (nongovernmental organizations), maintains a list of species all over the world that are critically endangered, endangered, extinct in the wild, vulner-able, or near-threatened. iUcn is universally recog-nized as the international information source about
xviii+598_Biodiversity_F.indd 218 11/17/11 1:13 PM
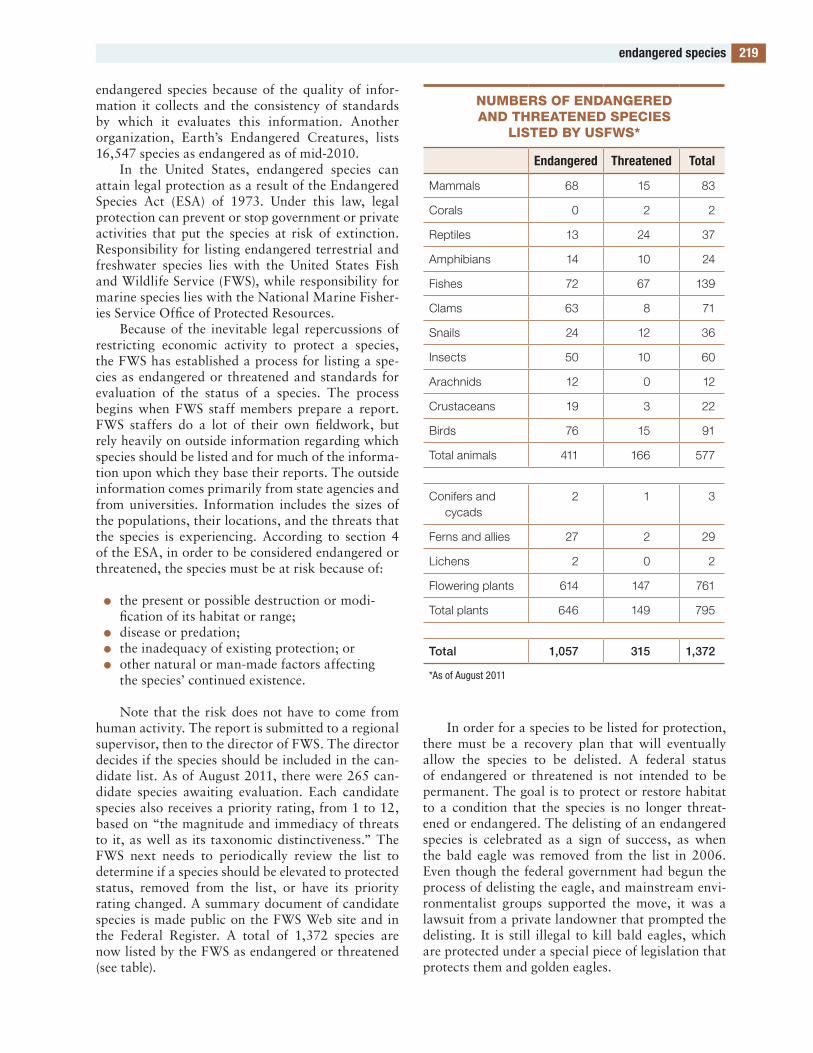
endangered species 219
endangered species because of the quality of infor-mation it collects and the consistency of standards by which it evaluates this information. another organization, earth’s endangered creatures, lists 16,547 species as endangered as of mid-2010.
in the United States, endangered species can attain legal protection as a result of the endangered Species act (eSa) of 1973. Under this law, legal protection can prevent or stop government or private activities that put the species at risk of extinction. Responsibility for listing endangered terrestrial and freshwater species lies with the United States Fish and Wildlife Service (FWS), while responsibility for marine species lies with the national Marine Fisher-ies Service Office of Protected Resources.
Because of the inevitable legal repercussions of restricting economic activity to protect a species, the FWS has established a process for listing a spe-cies as endangered or threatened and standards for evaluation of the status of a species. the process begins when FWS staff members prepare a report. FWS staffers do a lot of their own fieldwork, but rely heavily on outside information regarding which species should be listed and for much of the informa-tion upon which they base their reports. the outside information comes primarily from state agencies and from universities. information includes the sizes of the populations, their locations, and the threats that the species is experiencing. according to section 4 of the eSa, in order to be considered endangered or threatened, the species must be at risk because of:
● the present or possible destruction or modi-fication of its habitat or range;
● disease or predation; ● the inadequacy of existing protection; or ● other natural or man-made factors affecting
the species’ continued existence.
note that the risk does not have to come from human activity. the report is submitted to a regional supervisor, then to the director of FWS. the director decides if the species should be included in the can-didate list. as of august 2011, there were 265 can-didate species awaiting evaluation. each candidate species also receives a priority rating, from 1 to 12, based on “the magnitude and immediacy of threats to it, as well as its taxonomic distinctiveness.” the FWS next needs to periodically review the list to determine if a species should be elevated to protected status, removed from the list, or have its priority rating changed. a summary document of candidate species is made public on the FWS Web site and in the Federal Register. a total of 1,372 species are now listed by the FWS as endangered or threatened (see table).
in order for a species to be listed for protection, there must be a recovery plan that will eventually allow the species to be delisted. a federal status of endangered or threatened is not intended to be permanent. the goal is to protect or restore habitat to a condition that the species is no longer threat-ened or endangered. the delisting of an endangered species is celebrated as a sign of success, as when the bald eagle was removed from the list in 2006. even though the federal government had begun the process of delisting the eagle, and mainstream envi-ronmentalist groups supported the move, it was a lawsuit from a private landowner that prompted the delisting. it is still illegal to kill bald eagles, which are protected under a special piece of legislation that protects them and golden eagles.
numbers of endangered and threatened speCies
listed by usfWs*
Endangered Threatened Total
Mammals 68 15 83
Corals 0 2 2
Reptiles 13 24 37
Amphibians 14 10 24
Fishes 72 67 139
Clams 63 8 71
Snails 24 12 36
Insects 50 10 60
Arachnids 12 0 12
Crustaceans 19 3 22
Birds 76 15 91
Total animals 411 166 577
Conifers and cycads
2 1 3
Ferns and allies 27 2 29
Lichens 2 0 2
Flowering plants 614 147 761
Total plants 646 149 795
Total 1,057 315 1,372
*As of August 2011
xviii+598_Biodiversity_F.indd 219 11/17/11 1:13 PM

first entry220 endangered species 220
the wolf was not as fortunate. listing the wolf as endangered meant that it was illegal for hunters to kill it. the national Park Service reintroduced wolves to yellowstone national Park, where they had been eradicated. Wolf populations returned to levels that were not considered dangerously low. When the wolf was delisted in March 2008, Wyoming officials imme-diately declared nearly the entire state a “free-fire zone.” Some hunters immediately began killing as many wolves as they could. in some cases, they even trespassed on private property (whose landowners did not approve of the hunting) to do so. Within months, nearly one-tenth of the wolves of the Rocky Mountain region had been shot. the hunters claimed that wolves were killing their sheep, which did in fact sometimes happen. But according to research conducted by the United States Department of agriculture (which man-ages the national forests), only 1 percent of sheep deaths were caused by wolves, less than the number caused by bears, cougars, or even wild dogs (see table). More than a quarter of the sheep deaths were caused by coyotes, and almost a quarter of them died due to the weather. then a federal judge put a stop to the hunting of wolves, restoring them to protected status. Just days before leaving office, on January 14, 2009, President George W. Bush once again removed fed-eral protection from wolves. a federal judge restored wolves to the endangered species list in 2010. another federal judge revoked their protected status in 2011.
the administration of George W. Bush slowed the process of protection for endangered species to a historic low. as of 2004, the Bush administra-tion added an average of 9.5 species a year to the endangered list, compared with 65 a year under President Bill clinton and 59 a year under President George H. W. Bush. the period from May 2006 to May 2007 was the first time since 1981 that an entire year had passed without a single species being approved for endangered species listing. even for those that they did list, Bush administration appoin-tees approved only half as much critical habitat as FWS staffers had recommended based upon scien-tific research. Julie MacDonald, a deputy assistant secretary for Fish and Wildlife appointed in 2004, personally instructed FWS staffers to not find any more species to be at risk of extinction. as a result of these activities, she resigned in 2007. Republican lawmakers, in particular california representative Richard Pombo and Utah representative James V. Hansen, were strongly supportive of strangling the endangered Species act. Hansen proposed, in 2002, that all private property, all military bases, and all plant life be excluded from the endangered Spe-cies act. He predicted that, as a result, “we would, in short order, have a more prosperous and secure nation.” after the election of Barack Obama but before his inauguration, President Bush eliminated the requirement for independent scientific review of the potential environmental impact of federal proj-ects (in particular, leases for oil drilling).
the endangered Species act has, since its incep-tion, been the law that conservatives have loved to hate, and it will continue to be so. But it remains the only significant process by which endangered species can receive protection. alternative strategies, such as cloning, have proven to be expensive failures. in 2001, advanced cell technologies, a Massachu-setts biotechnology company, successfully cloned a guar, an endangered relative of the domestic cow, by implanting a genetically modified egg cell in a cow. Full of hope, they named the little guar noah. it quickly died of a bacterial infection. Had the guar survived, what could be done then, if the wild habi-tat has been destroyed? Habitat preservation, made possible by the eSa, remains the only cost-effective way to rescue endangered species.
See also federal government role in con-servation (United States).
Further readingBBc news. “endangered animal clone dies.” available
online. URl: http://news.bbc.co.uk/2/hi/science/nature/1113719.stm. accessed May 13, 2011.
Brooke, M. de l., et al. “Potential for rat predation to cause decline of the globally threatened Henderson
Causes of sheep deaths, aCCording to the united states
department of agriCulture*
Source of death percentage of deaths
Wolves 1
Coyotes 25.3
Weather 22.6
Disease 11
Complications during lambing 9
Other predators 6.5
Old age 5.8
Bear 1.8
Cougar 1.4
Dog 1.1
Other 7
Unknown 7.7
*Data from Chadwick
xviii+598_Biodiversity_F.indd 220 11/17/11 1:13 PM

endemic species 221
petrel Pterodroma atrata: evidence from the field, sta-ble isotopes and population modelling.” Endangered Species Research 11 (2010): 47–59.
cart, Julie. “Predator becomes prey in Wyoming.” Los Angeles Times, September 28, 2008. available online. URl: http://articles.latimes.com/2008/sep/28/nation/na-wolf28/2. accessed May 13, 2011.
———. “endangered Species act is modified.” Los Angeles Times, December 12, 2008. available online. URl: http://articles.latimes.com/2008/dec/31/local/me-spe cies31. accessed May 13, 2011.
chadwick, Douglas H. “Wolf wars.” National Geographic, March 2010, 34–55.
christiansen, F., et al. “effects of tourist boats on the behav-iour of indo-Pacific bottlenose dolphins off the south coast of Zanzibar.” Endangered Species Research 11 (2010): 91–99.
earth’s endangered creatures. “World endangered Species list.” available online. URl: http://www.earthsendan-gered.com/full_list_html.asp. accessed august 8, 2011.
eilperin, Juliet. “endangered Species act’s protections are trimmed.” Washington Post, July 4, 2004: a01.
international Union for the conservation of nature. “iUcn.” available online. URl: http://www.iucn.org. accessed august 8, 2011.
loi, Pasqualino, et al. “Genetic rescue of an endangered mammal by cross-species nuclear transfer using post-mortem somatic cells.” Nature Biotechnology 19 (2001): 962–964.
loomis, John B., and Douglas S. White. “economic benefits of rare and endangered species: Summary and meta-analysis.” Ecological Economics 18 (1996): 197–206.
Sherley, R. B., et al. “Spotting the difference: towards fully-automated population monitoring of african penguins Spheniscus demersus.” Endangered Species Research 11 (2010): 101–111.
Slevin, Peter. “Bald eagle to be taken off endangered list.” Washington Post, December 25, 2006. available online. URl: http://www.washingtonpost.com/wp-dyn/content/article/2006/12/24/aR2006122400666.html. accessed May 13, 2011.
U.S. Fish and Wildlife Service. “endangered Species Pro-gram.” available online. URl: http://www.fws.gov/endangered/. accessed May 13, 2011.
endemic species endemic species are spe-cies whose entire range is a single, limited locality. they may be locally abundant within that locality but are rare on a regional and global level. Biodi-versity hot spots have many endemic species. Many endemic species are endangered because if local populations become extinct there are no others to replace them.
there are two main reasons that may cause a species to be endemic. First, the species may have
evolved only in the one location in which it persists. this is the case with many species that are endemic to geographical islands. Second, the species may have formerly had a wide range but has become extinct everywhere except that one location. this is the case with many species that are endemic to eco-logical islands on continents.
there are many examples in which a disjunct species consists only of endemic subspecies. the seaside alder, Alnus maritima, is a disjunct spe-cies that consists only of three endemic subspecies: ssp. maritima, which lives only on the Delmarva Peninsula east of chesapeake Bay; ssp. georgiensis, which lives in a single swamp in Bartow county, in northwestern Georgia; and ssp. oklahomensis, which lives only in south central Oklahoma, primar-ily in Johnston county. there are also examples of endemic subspecies of a widespread species. Cervus canadensis, the elk, has populations across north america. One subspecies, ssp. nannodes, the tule elk, is endemic to the marshes of central california and has nearly become extinct as these marshes have been drained and transformed into an agricultural and urban landscape. in 1895, only 28 tule elk sur-vived. Habitat preservation has allowed their num-bers to expand to a few thousand.
GEoGraphIC ISlaNd ENdEmICSSome large islands are actually continental fragments that have remained separate from the larger con-tinents for many millions of years. Madagascar, a large island off the coast of africa, has a very large number of endemic species, most notably the 99 spe-cies of lemurs. another continent-island is new cale-donia, which has many endemic species, including about half of the trees in the conifer family araucari-aceae. the flowering plant that most nearly resembles the ancestral flowering plant, Amborella trichopoda, is endemic to new caledonia. taiwan, the world’s fourth largest island, has also been isolated for many millions of years and has many endemic species. twenty percent of its animals (including 64 percent of its mammals, such as the Formosan rock macaque) and 26 percent of its plants are endemic.
young volcanic islands have had much less time for the evolution of endemic species, yet they have many. the oldest islands of the Hawaiian archi-pelago are only about 5 million years old. Despite its youth, the Hawaiian archipelago has (or had, prior to extinction caused by the aboriginal Polynesian humans) many endemic bird species, including 51 species of honeycreepers. the Galápagos islands, off the coast of ecuador, also have many endemic species, most famously the giant tortoises. the croc-odile Crocodylus mindorensis is found only on the Philippine islands.
xviii+598_Biodiversity_F.indd 221 11/17/11 1:13 PM

first entry222 endemic species 222
EColoGICal ISlaNd ENdEmICSecological islands are small habitats surrounded by large expanses of an environment unsuitable for its species. Frequently they are remnants of previously large habitats. examples include the following:
● cloud forests are deciduous forests at the tops of tropical mountains. Some of the spe-cies are the same as those of temperate decid-uous forests, while others are endemics.
● in california, all of the coastal closed-cone pine species and coastal dry temperate oak species are endemic.
● Some oak species, such as Quercus carm-enensis and Q. deliquescens, are endemic to mountaintop dry forests surrounded by desert in northern Mexico and southwestern United States.
● Ginkgoes were once widespread in temper-ate forests, but by modern times were nearly extinct. Just one species, Ginkgo biloba, survived as an endemic species in small patches of forest in china.
● the dawn redwood (Metasequoia glypto-stroboides) once grew widely in the forests of asia but today is endemic to a small region in Sichuan and Hubei Provinces of china.
● the giant redwood (Sequoiadendron gigan-teum) was once widespread in western north america but is now restricted to a few moist coniferous forests in the Sierra nevada.
● the Serbian spruce (Picea omorika), which today grows only in the Drina River Valley, was once more widespread in eurasia.
● the Santa lucia fir (Abies bracteata) now grows only in moist microenvironments of the Santa lucia Mountains of coastal california.
● Torreya taxifolia is a conifer now endemic to the apalachicola River area of northern Florida and southwestern Georgia.
isolated lakes can also have numerous endemic species. the food web of lake Baikal in Siberia is based upon an endemic zooplankton species, Epischura baikalensis. among the endemic fishes in lake Baikal are the Baikal oil fish species Come-phorus baikalensis and C. dybowskii and the Baikal sturgeon subspecies Asipenser baerri baicalensis. lake Baikal has an endemic seal species Phoca sibirica.
Some endemic plants have proven to be popu-lar ornamentals, partly because they can tolerate a greater range of climatic conditions than their restricted natural range would suggest and partly
because of the allure of their rarity. Ginkgoes are now planted worldwide, and dawn redwoods are not uncommon in parks and gardens. a few people have planted seaside alders in their yards, and a volun-teer group of citizen-naturalists has planted Torreya trees at several other locations in the southeastern United States. the umbrella palm (actually a sedge, Cyperus alternifolius) is endemic to Madagascar but is now a popular water garden ornamental. the traveler’s palm (Ravenala madagascariensis, actu-ally a relative of the bird of paradise plant), endemic to Madagascar, is also a popular ornamental in warm regions of the world.
See also biogeography; cloud forests; dis-junct species; endangered species; Galápagos islands; island biogeography.
Further readingBowen, lizabeth, and Dirk van Vuren. “insular endemic
plants lack defenses against herbivores.” Conservation Biology 11 (1997): 1,249–1,254.
Harrison, Susan. “local and regional diversity in a patchy landscape: native, alien, and endemic herbs on serpen-tine.” Ecology 80 (1999): 70–80.
lavergne, Sébastien, et al. “the biology and ecology of nar-row endemic and widespread plants: a comparative study of trait variation in 20 congeneric pairs.” Oikos 107 (2004): 505–518.
Malcolm, Jay R., et al. “Global warming and extinction of endemic species from biodiversity hotspots.” Conser-vation Biology 20 (2006): 538–548.
Médail, Frédéric, and Régine Verlauque. “ecological char-acteristics and rarity of endemic plants from south-east France and corsica: implications for biodiversity conservation.” Biological Conservation 80 (1997): 269–281.
Orme, c. David l., et al. “Global hotspots of species rich-ness are not congruent with endemism or threat.” Nature 436 (2005): 1,016–1,019.
Park, young-Seuk, et al. “conservation for endemic fish species threatened by the three Gorges Dam.” Biologi-cal Conservation 17 (2003): 1,748–1,758.
Philippart, Jean claude. “is captive breeding an effective solution for the preservation of endemic species?” Bio-logical Conservation 72 (1995): 281–295.
Silvertown, Jonathan. “the ghost of competition past in the phylogeny of island endemic plants.” Journal of Ecology 92 (2004): 168–173.
thompson, John D. “Population differentiation in Medi-terranean plants: insights into colonization history and the evolution and conservation of endemic spe-cies.” Heredity 82 (1999): 229–236.
Van der Werff, Henk, and trisha consiglio. “Distribution and conservation significance of endemic species of flowering plants in Peru.” Biodiversity and Conserva-tion 13 (2004): 1,699–1,713.
xviii+598_Biodiversity_F.indd 222 11/17/11 1:13 PM

environmental ethics 223
environmental ethics Members of the human species are unique in having the need to invent ethi-cal systems to determine the actions that they should take. ethical systems evolved first for local then national and then international relationships among human beings. environmental ethics prescribe cor-rect actions of humans toward other species and toward the earth as a whole. it was the last to evolve and is still in rudimentary stages of development. While wide consensus has been reached on inter-national ethical issues such the condemnation of nuclear war and torture, such consensus is a long way off for environmental issues. Many people see environmental ethics as an inevitable extension of the trajectory of the ethics of human interactions. at the time that Odysseus hung all of his disobedi-ent household slaves on one rope, such action was accepted as ethical. today it is considered murder. Many people say it is time to consider the abuse of nonhuman creatures and of the earth just as evil as the abuse of one human or human group by another. a magazine article in 2008 described the slaughter
of gorillas in africa; it was entitled, “Who Murdered the Mountain Gorillas?” in the United States in the 1930s, the massive soil erosion of the dust bowl in the western plains was considered a natural resource disaster, but many observers also considered it an immoral violation of the earth (see illustration).
any group of organisms acting only in its own interests has the capacity of destroying its local environment or the entire earth. While the Gaia hypothesis, first proposed by climate scientist James lovelock, describes the many ways in which organ-isms have promoted their own success through cooperation with others, and the ways in which such cooperation has influenced the entire earth (especially its atmosphere), biologist Peter Ward has proposed an alternative. the Medea hypoth-esis describes the ways in which organisms have devastated their local environments and the entire earth through the pursuit of their own interests. He cites the Permian extinction, caused by bacterial hydrogen sulfide, as an example. it appears that, without environmental ethics, humans metabolizing
Alexandre Hogue painted Erosion No. 2—Mother Earth Laid Bare during the dust bowl in the central United States—it depicts erosion, caused principally by irresponsible farming, as a violation of a sacred, living planet. (Philbrook Museum, Tulsa, Oklahoma)
xviii+598_Biodiversity_F.indd 223 11/17/11 1:13 PM

first entry224 environmental ethics 224
their way through the biosphere cannot avoid envi-ronmental destruction perhaps as disastrous as that of the Permian extinction.
Most if not all of the world’s environmental problems could be solved by pragmatism. Global warming should be curbed because of the devas-tating effects it will have on all aspects of human activity. Heavy metal pollution should be prevented because the metals concentrate in the food chain and can endanger human health. Biodiversity should be protected because many of the species may provide immense economic benefit in the future. ecologist aldo leopold, considered by many to be the founder of environmental ethics, stated that the first law of intelligent tinkering was to not throw out any of the parts. these pragmatic arguments apply to human society as a whole; it is usually profitable for cor-porations to continue releasing carbon dioxide and heavy metals as long as they can get away with it, since the profit is theirs and the damage is shared by everyone. it may not be necessary to go beyond merely pragmatic arguments, since the implementa-tion of these arguments appears to be beyond human capacity at this time: the human species appears incapable of reducing its carbon output enough to prevent disaster. What is the point of going beyond a practical argument?
this question is difficult to answer, other than to point out that many people are simply not any more satisfied with a pragmatic argument for envi-ronmental preservation than for a pragmatic argu-ment against genocide. nearly everyone admits the necessity of moral, rather than just pragmatic, envi-ronmental ethics—some from a desire to conform, others out of passion. Proposed systems of envi-ronmental ethics can be roughly classified on the basis of what a person considers the environment to be: creation or nature. the first considers the environment and all of its living and nonliving com-ponents to be the product of a deity that has made humankind separate from it in some way, while the second considers the environment to be the source of humankind (“nature” comes from the latin for “birth”).
ThE ENvIroNmENT aS CrEaTIoNthe major monotheistic religions of the world (Juda-ism, christianity, and islam) believe that a single, all-powerful deity created the world and made humans the rulers of it. Moreover, these religions base their beliefs upon scriptures written long before people even imagined the questions of environmental eth-ics. Until the last few centuries, humans quite rightly viewed the earth as unconquerable and usually dan-gerous. the earth could take care of itself. Modern adherents of these religions therefore turn to their
pre-ecological scriptures and pick out passages that were not written for the purpose of answering such questions and try to base systems of environmental ethics upon them.
Because of the importance of the Judeo- christian Bible in the history of western culture, historian lynn White, Jr., wrote in 1967 that chris-tianity, guided by the first chapter of Genesis, had been in the forefront of a largely successful attempt to conquer the created world. it is beyond dispute that, according to this chapter, God gives humans the command to conquer and subdue the earth and have dominion over it. the words clearly refer to the kind of conquest that kings exercised over van-quished nations. White did not, however, say that christianity should be rejected, but that christians should use Saint Francis of assisi, a medieval italian monk who considered the Sun, Moon, wind, fire, and all creatures to be his brothers and sisters, as the model of ethical behavior rather than the Bible.
not surprisingly, many christians became defensive and claimed that White had misinterpreted the first chapter of Genesis. Since 1967, christian writers have found many passages in the Bible that paint a very different picture of the creation. Here are a few examples:
● the second chapter of Genesis depicts God as commanding humans to take care of the earth the way one would care for a garden (the Garden of eden).
● the third chapter of Genesis depicts expul-sion from the Garden of eden and a host of environmental problems (“cursed is the ground . . . thorns and thistles it shall bring forth”) as consequences of sin.
● the sixth chapter of Genesis describes noah, whose family was to be the sole sur-vivors of a worldwide flood. God told noah to build an ark big enough for a pair of each species of terrestrial animal, presum-ably even those that noah did not especially care about.
● the ten commandments included a prohi-bition against work on one day of the week, called the sabbath. the result of this com-mandment, according to conservative Jew-ish scholars, is the deliberate interruption of the momentum of acquisition—to desist, once a week, from using other people (and the earth) as a source of profit.
● the Old testament law included some com-mandments that dealt directly with the earth. Most famous is the Sabbath of the Fields, in which agricultural fields were sup-posed to be left fallow every seventh year.
xviii+598_Biodiversity_F.indd 224 11/17/11 1:13 PM

environmental ethics 225
● in the book of Job, God addresses Job from a whirlwind and describes a vast unknow-able and unconquerable world of creatures, from large ones such as Behemoth and leviathan to humble ones such as donkeys and conies. the message is clear that the purposes of these creatures have absolutely nothing to do with humans. the fact that these creatures are untamed was a perfectly acceptable part of God’s order of the world.
● Many passages in the psalms and prophets describe the beauty of creation entirely apart from any benefit it may confer on humans. a passage from the prophet isaiah describes trees reclaiming wasteland and restoring its water resources and claims that this process, which would now be called ecological, was the work of God.
● Jesus frequently used symbolism from the creation, from wildflowers to sparrows, in his parables—frequently things that would be noticed only by people who stopped to carefully observe them.
ecology-minded christians, who focus attention on these passages, claim that God has given humans the responsibility to be stewards, or caretakers, of the earth, rather than its conquerors. this is an important concept, even though the terms steward and caretaker do not appear in the Bible in connec-tion with the earth.
the above examples aside, White was right in his conclusion. One reason is that the Bible is not a book. it is a collection of books, written by many people over many centuries, and it does not have a single coherent message. Most of the biblical books are themselves the product of editorial redaction from different primordial traditions. the first chap-ter of Genesis, correctly interpreted by White, pres-ents a different view of creation from the second chapter, which was originally written by a different person. each part of the Bible potentially contradicts every other part. an adherent of christianity may choose to accept a biblical image of conquering the earth or a biblical image of taking care of it. it all depends on which passage is chosen. adherents of christianity, Judaism, or islam can, however, force these passages into harmony, and in this way they can use the second chapter of Genesis to ameliorate the first.
White was right for another reason. christian theologians have forced all the biblical passages into harmony, and the climax of this harmonized Bible is the book of Revelation, also known as the apoca-lypse. Revelation, the last book of the Bible, is the only example of an entire style of literature, popular
in the Mediterranean world about 100 c.e., which is familiar to most modern readers. in this book, the entire biosphere of the earth is destroyed dur-ing cosmic battles between good and evil, and those who are saved live in Heaven. the Heavenly city there described is the antithesis of biodiversity and contains virtually nothing of the original creation. there is a river and a single tree. everything else is gold and jewels—an utterly artificial environment. if this is, as about half of americans believe, the future of the universe, what is the point of saving species and ecosystems? For this reason also, if christians are to participate in the rescue of the environment, they must look beyond the Bible and perhaps, as White suggests, emulate St. Francis.
the extreme popularity of apocalyptic chris-tianity meant that christians, as a whole, were a feeble source of support for environmental protec-tion. their main passion was to support the politi-cally conservative George W. Bush administration, which rejected the evidence of global warming and paid little attention to environmental protection. in fact, after the end of the Bush administration, the full extent of conservative neglect for even the most basic environmental protection became obvious. the Minerals Management Service of the Depart-ment of the interior was supposed to license fossil fuel extraction on federal lands with due consider-ation of the impact that this extraction would have on natural habitats. instead, they gave licenses to corporations in return for bribes and sexual favors. When President Bush left the 2007 economic sum-mit, he said, “Goodbye from the world’s biggest polluter,” making environmental ethics into what he considered to be a joke.
edward O. Wilson, perhaps the world’s lead-ing defender of biodiversity, wrote a book that was addressed to an unnamed Baptist minister. Wilson said that a conservative christian could find com-mon ground with even a secular humanist like him-self by defending the earth, which Wilson called the creation. Wilson, however, wrote this book only after originally considering the christian environ-mental movement to be a hopeless cause.
With the end of the Bush administration and the disgrace of several leading christian antienvi-ronmentalists, a tide of opinion has turned toward christian environmentalism. Despite this, many conservative christians still consider the earth to be unworthy of serious attention. they focus their attention instead on fighting against evolutionary science. When edward O. Wilson visited Oklahoma in March 2009, a group of creationists disputed him about whether charles Darwin did or did not believe in a creator, and in the process Wilson’s impas-sioned plea for saving the earth’s biodiversity was
xviii+598_Biodiversity_F.indd 225 11/17/11 1:13 PM

first entry226 environmental ethics 226
largely obscured. the view of the earth that many conservative christians have can be summarized in this way: “it’s okay to spit on it, to drive a truck over it, to spill oil on it, to chop it down, or to shoot it, so long as you don’t believe that it evolved!”
all during this time, a significant minority of christians continued to insist that protecting the earth was a christian responsibility because the earth is God’s creation.
ThE ENvIroNmENT aS NaTurEOther religious traditions consider the earth to be nature, from which humans evolved and of which humans are a part, just like every other species. Some of these religions are polytheistic, such as Buddhism and paganism, while others are more akin to spiri-tual philosophies, such as taoism. there is even a little of this viewpoint in the Bible. the 104th psalm describes a world of creatures, of which humans are depicted as merely one component. although these religions have not led their adherents on a deliber-ate conquest of the earth, the homelands of these religions have suffered their share of environmental problems. the natives of north and South amer-ica experienced collapsed civilizations just like the europeans.
the person most responsible for the emergence of environmental ethics based upon ecology was aldo leopold. as mentioned previously, leopold made a pragmatic argument for the preservation of biodiversity. But he also promoted what he called the “land ethic” and championed what he called “thinking like a mountain.” Humans, rather than rulers or even stewards, would be merely “plain member and citizen” of the world of species. Right and wrong were defined as those actions that con-tributed to or detracted from the health of nature. His essays in A Sand County Almanac remain a classic, even though they do not develop a complete ethical system. More recently, writer Wendell Berry has called for an ecological humility as the proper ethical system for humans. Biologist Wes Jackson promoted a very pragmatic view of the steward-ship of natural resources, particularly soil, in New Roots for Agriculture and by founding the land institute in Salina, Kansas. like leopold, Jackson then went beyond pragmatism and calls for humans to recognize their membership in the world of inter-acting species.
attention is now being given to evolution, as well as ecology, as a foundation for environmental ethics. Humans are one of several million species and, like the rest of them, evolved by natural selec-tion; humans therefore have no rights not possessed by the other species. Some thinkers have gone so far as to develop a viewpoint known as Deep ecology,
in which the human species has become a disease upon the earth that needs to be brought into control.
Most environmentalists seek peaceful and orderly solutions to environmental problems—for both ethical and pragmatic reasons. a few organiza-tions, however, use violent tactics to gain attention, even though they have little hope of actually coun-teracting the economic forces behind environmen-tal destruction. these tactics usually take the form of arson on resorts that are closed for the season. although no one has yet died from what conserva-tives call “environmental terrorism,” Senator James inhofe, Republican of Oklahoma, said that these organizations were the greatest threat of domestic terrorism in the United States. He said this even though Oklahoma was the site of the largest act of domestic terrorism that has ever occurred in the United States—the Oklahoma city bombing, which had nothing to do with environmentalism. conser-vatives conveniently ignore acts of violence carried out by extremists on the political right (again, with-out deaths) against government agencies, such as the Bureau of land Management, whose charge is to protect natural areas.
ThE proBlEm of fraGmENTaTIoNPerhaps the major impediment to environmental ethics is academic fragmentation. nearly every-thing that humans do and every area of study have environmental implications and a role to play in the development of environmental ethics. Science, religion, business, psychology, sociology, and the humanities are all necessary components of envi-ronmental ethics. yet all of these fields of study are largely separate from one another. environmental issues are largely left to environmental scientists. Meanwhile, economists often continue to do their work as if the world were understood the same today as in the time of adam Smith. economist Kenneth Boulding said, “anyone who believes that exponential growth can go on forever in a finite world is either a madman or an economist.” Only by bringing environmental science and ethics into all fields of study will humankind have a coor-dinated response to the threat of environmental disaster. edward O. Wilson calls for a “consilience” of all academic disciplines, centered around the rec-ognition of environmental and evolutionary realities of the human species. this idea, necessary as it is, has made little progress.
the most promising direction for success in envi-ronmental ethics is to work toward a convergence. if people become aware of the pragmatic reasons for protecting the environment and biodiversity, they will turn to their own ethical background—whether monotheistic religion, natural religion, ecology, or
xviii+598_Biodiversity_F.indd 226 11/17/11 1:13 PM

environmental ethics 227
evolution—and join with others to implement solu-tions to environmental problems, even people who have reached the same conclusion from a different starting point.
ThE INEvITaBIlITy of dIlEmmaSWhen a species with as much technological potential as Homo sapiens evolves, whatever happens next is going to be wrong. every action taken by such a species is going to have a destructive impact, if only because of enormous population growth. Dilemmas, therefore, are inevitable.
One example of such a dilemma involves the use of plastics. Objects made of plastics, such as con-tainers and sacks, never decompose. if they become litter they end up in the ocean, often in gigantic gyres filled with garbage. Over a dozen such oceanic trash heaps exist, some as large as texas. Marine animals such as turtles and birds have not evolved the ability to distinguish between garbage and their normal food items and often die after eating large numbers of plastic objects. the plastic in aquatic ecosystems may become brittle from exposure to ultraviolet light and break into ever smaller pieces. these pieces then become part of the plankton and are consumed by other aquatic animals. no one knows what will happen when all of the plastic that today consists of large chunks eventually breaks into plankton-sized pieces. even when most people are careful about proper refuse disposal and recy-cling, much plastic garbage ends up in the environ-ment: Wind can rip plastic bags right out of refuse bins when their contents are being transferred to garbage trucks. Some cities and even entire coun-tries (such as ireland) have banned plastic bags, and shoppers increasingly bring their own bags, some made from recycled plastic.
if plastic containers had not been invented, many more containers would have been made of metal or glass. transportation of these heavier containers would have resulted in the burning of more fossil fuels and a greater contribution to the greenhouse effect. Plastics, therefore, have contrib-uted to a major worldwide pollution problem but relieved the earth from some of its greenhouse effect. Should the use of plastics be encouraged or prohibited? the only way around such a dilemma is to find an altogether different solution. For exam-ple, if consumers buy and use less material, there will be less need for containers. Meanwhile, the dilemma remains.
environmental scholar Martin lewis points out another dilemma. Burning coal has contrib-uted immensely to air pollution and the greenhouse effect. yet it was coal, as a new energy source, that spared the english forests from complete destruc-
tion at the beginning of the industrial Revolution. the dilemma—is coal good or bad?—can be solved only by finding an altogether different solution, which is to use neither coal nor wood, an option that was not available in the 19th century. Dilem-mas are unavoidable in environmental ethics and can only be resolved if dedicated people employ creative thinking.
See also essay: “ecosystem Services: Public or Private costs?”; ecosystem services; greenhouse effect; Wilson, edward O.
Further readingBailey, liberty Hyde. The Holy Earth. ithaca, n.y.: cornell
University Press, 1915/1980.Berry, thomas. The Dream of the Earth. San Francisco:
Sierra club, 1988.Berry, Wendell. The Gift of Good Land. San Francisco:
north Point Press, 1981.———. The Unsettling of America. San Francisco: Sierra
club, 1977.Douglas, William O. “Mineral King and the Rights of
Rocks.” Supreme court of the United States, Sierra Club v. Morton, 70–34, april 19, 1972.
Foreman, Dave. Confessions of an Eco-Warrior. new york: Random House, 1993.
Hardin, Garrett. “ecology and the death of providence.” Zygon 15 (1980): 57–68.
Jackson, Wes. Becoming Native to This Place. Berkeley, calif.: counterpoint, 1996.
———. New Roots for Agriculture. lincoln: University of nebraska Press, 1980.
Jenkins, Mark. “Who murdered the Virunga gorillas?” National Geographic, July 2008, 34–65.
leopold, aldo. A Sand County Almanac: With Essays on Conservation from Round River. new york: Sierra club/Ballantine Books, 1974.
lewis, Martin H. Green Delusions: An Environmental-ist Critique of Radical Environmentalism. Durham, n.c.: Duke University Press, 1992.
lovelock, James. The Vanishing Face of Gaia: A Final Warning. new york: Basic Books, 2009.
Manes, christopher. Green Rage: Radical Environmental-ism and the Unmaking of Civilization. Boston, Mass.: little, Brown, 1990.
McKibben, Bill. The Comforting Whirlwind: God, Job, and the Scale of Creation. cambridge, Mass.: cowley, 2005.
Orr, David W. Earth in Mind: On Education, Environment, and the Human Prospect. Washington, D.c.: island Press, 1994.
Shaw, James H. “assessing the progress toward leopold’s land ethic.” Wildlife Society Bulletin 15 (1987): 470–472.
Snyder, Gary. The Practice of the Wild. San Francisco: north Point Press, 1990.
xviii+598_Biodiversity_F.indd 227 11/17/11 1:13 PM

first entry228 environmental psychology 228
Stone, christopher D. Should Trees Have Standing?: Towards Legal Rights for Natural Objects. los altos, calif.: W. Kaufmann, 1981.
Van Dyke, Fred, David c. Mahan, Joseph K. Sheldon, and Raymond H. Brand. Redeeming Creation: The Bibli-cal Basis for Environmental Stewardship. Downers Grove, ill.: inter-Varsity Press, 1996.
Ward, Peter. The Medea Hypothesis: Is Life on Earth Ultimately Self-Destructive? Princeton, n.J.: Princeton University Press, 2009.
White, lynn, Jr. “the historical roots of our ecologic cri-sis.” Science 155 (1967): 1,203–1,207.
Wilson, edward O. Consilience: The Unity of Knowledge. new york: Knopf, 1998.
———. The Creation: An Appeal to Save Life on Earth. new york: norton, 2006.
environmental psychology environmental (or conservation) psychology is the study of the ben-eficial effects of nature on the human mind. this includes a person’s frequent presence within natural settings and direct contact with a diversity of wild organisms.
it is widely recognized that humans have an innate desire for direct contact with the natural world. evolutionary biologist e. O. Wilson coined the term biophilia for the universal human love (-philia) of nature and life (bio-). it may have a genetic component, as well as a learning component that is acquired by children during exposure to the outdoors. it will perhaps be impossible for scientists to rescue the biodiversity of the earth without draw-ing upon the psychological resources of citizens of countries all over the world—to tap into the feelings of biophilia.
Biophilia, contends Wilson, was as much the product of natural selection as any other aspect of human psychology such as the fear of strangers. Biophilia provided a fitness advantage for humans and their evolutionary ancestors. the enjoyment of birds, trees, and mammals encouraged learning about them. By carefully watching the other species, primitive humans could learn to make more effective use of them as natural resources. the people who enjoyed listening to birds would be more likely to recognize a bird alarm call that might indicate the presence of a dangerous predator. While a feeling of biophilia was not necessary to these beneficial actions, it made them easier and more effective. the extensive use of animal motifs in such paintings as those in the caves of lascaux and altamira suggests that biophilia may have been closely connected to religion, which also provided important advantages to primitive human societies. an appreciation of the beauty of the landscape and a love for its living
community of species may have kept some primi-tive humans from giving up in the frequently harsh circumstances of life, and these were the humans who were the ancestors of modern people. Modern humans are, according to this concept, happiest in environments that most closely resemble the savanna in which the human genus first evolved 2 million years ago.
Psychologists and urban planners have indepen-dently arrived at a concept similar to biophilia. Psy-chologists have found that humans are happier in, and choose environments that contain, a balance of trees and open areas. Urban planners have been careful to include trees and landscaping with build-ings and parks between them. Biophilia may also explain the frequent success of horticultural therapy in the recovery of patients from physical and mental diseases. Programs to help guide children away from social problems often include an outdoor compo-nent. even though many people can be as happy in a big city as if it were canyons, most people seek occa-sional escape into the natural world. Biophilia may be essential to the successful preservation of natural areas simply because most people cannot conceive of living in a world in which natural areas have been destroyed.
in modern society, the learning component of biophilia has overlooked a great deal of biodiversity. Many people like trees but do not particularly care which ones. While central american native tribal children can identify many plant species, as could rural american children just a few decades ago, modern children in the United States can identify very few. Moreover, when modern humans visit the outdoors, they usually take a great deal of their manufactured environment with them. Wilson and many others believe that, in order to rescue bio-diversity from what appears to be the sixth of the mass extinctions that is now resulting from human activity, humans must rediscover the love not just of nature but of species. in this regard, evolution-based biological and environmental education may be essential for the protection of the natural world.
it is not just modern scholars who have claimed that exposure to the natural world may be essential to healthy psychological development in children. in one of his last essays, “the Dispersion of Seeds,” 19th-century american naturalist Henry David thoreau wrote:
a man fits out a ship at a great expense and sends it to the West indies with a crew of men and boys, and after six months or a year, it comes back with a load of pine-apples; now, if no more gets accomplished than the speculator commonly aims at, if it simply turns out what is called a
xviii+598_Biodiversity_F.indd 228 11/17/11 1:13 PM

environmental psychology 229
successful venture, i am less interested in this expedition than in some child’s first excursions a-huckleberrying, in which it is introduced into a new world, experiences a new development, though it brings home only a gill of berries in its basket.
Research, such as that conducted by landscape ecologist Frances e. Kuo, has clearly demonstrated that humans receive psychological benefits from contact with natural landscapes. it is well known that children with attention deficit hyperactivity dis-order (aDHD) have reduced symptoms if they have time for recess. Kuo has shown that aDHD symp-toms were reduced more by outdoor playtime than by indoor play, and by outdoor play in the vicinity of trees and other plants than by play on cement or asphalt. She has also shown that girls growing up in a chicago housing project score higher on tests of self-discipline (concentration, control of com-pulsive behavior, delay of gratification) when their apartments have views of trees and green spaces than when they do not. there was a similar effect of green spaces on the ability of inner-city women to deal with major life issues. Green surroundings can even reduce crime. Within a chicago hous-ing project, areas that have trees and grass had 48 percent fewer property crimes and 56 percent fewer violent crimes than areas that lacked vegeta-tion. Reasons may include that green landscaping encourages people to spend more time outdoors, thus increasing their surveillance and preventing crime, and that green landscaping is an indication that residents take better care of the property in which they live. these results run directly coun-ter to the misconception that trees are dangerous because criminals can hide behind them. Residents that live in housing project buildings surrounded by trees and grass report in surveys that they knew their neighbors better, socialized with them more often, and had a stronger sense of community than residents of buildings without green surroundings. these results show that natural settings do more than just make people feel better; they have measur-able effects on reducing crime and on the quality of life in a community.
Other researchers have also shown the medi-cal and social benefits of green environments. in one study, volunteers watched a stressful movie, fol-lowed by pictures of either urban or natural settings. those who watched the natural images recovered from the stress more quickly, as measured by blood pressure, heartbeat rate, and facial muscle tension. Other studies have demonstrated that patients expe-rience less stress prior to surgery if the waiting room has plants, aquaria, or windows with a natural view.
after surgery, patients recover more quickly and require less painkiller if they have a window with a view of trees. Studies with psychiatric patients and prisoners also show that views of nature can reduce symptoms of stress. all of this is in addition to the enhancement of physical health that comes from reg-ular exercise, such as hiking and jogging, in natural settings. nature is not just a place to kill time but an essential investment in physical and mental health. national legislation called no child left inside (a counterpart to the educational legislation no child left Behind) was introduced to congress in 2011.
it would seem to be the easiest thing in the world to enhance the health of both children and adults by preserving and creating urban natural areas. there are problems, however. One is liability. if a child, or anyone else, is injured while hiking or climbing in a natural area, the government or private owner risks a lawsuit. the fear of litigation is one reason that many schools have reduced the amount of recess and limited most of this to highly supervised and very unnatural playgrounds. in addition, green spaces are often not readily accessible because developed resi-dential areas are cut off from them by busy streets and highways.
not all ways of spending time in nature are equally good. in several places around the United States, desert dunes have been set aside on which people can ride all-terrain vehicles; these vehicles are also permitted on some trails on Forest Service land. this sort of recreation does not encourage close observation and appreciation of nature and biodiver-sity, nor does it tap into the psychological benefits of contact with nature. Frequently, users of all-terrain vehicles wander from designated areas and destroy natural and archaeological sites. Just being outdoors is, therefore, not enough.
One problem that has resulted from lack of con-tact with nature is that people do not realize what nature is supposed to look like. When a natural habitat is degraded by human activity, a new genera-tion of children grows up thinking that this is the way it is supposed to look—that invasive species are natural components of a habitat, that eutrophication is natural. as a result, they are not motivated to take action to preserve truly natural environments or to remediate degraded environments.
Most scientists and environmental profession-als agree that the increasing awareness of environ-mental problems is good, because it will motivate people to change the habits that contribute to these problems (e.g., reducing one’s carbon footprint). But this awareness can have a negative effect also. Psy-chologists Jessica Frizte and colleagues have pointed out that the changes associated with the greenhouse effect, particularly disastrous storms and droughts,
xviii+598_Biodiversity_F.indd 229 11/17/11 1:13 PM

first entry230 Eocene epoch 230
may significantly contribute to anxiety and men-tal health problems. By encouraging people to have more direct contact with nature, it is nearly inevita-ble that they will not only begin to care more about biodiversity but will begin to experience nature not as a problem to be solved but as a world to be enjoyed and protected.
See also greenhouse effect; thoreau, Henry David; Wilson, edward O.
Further readingcalifornia Roundtable on Recreation, Parks, and tour-
ism. “california children’s Outdoor Bill of Rights.” available online. URl: http://www.calroundtable.org. accessed august 26, 2009.
chicago Wildnerness. “leave no child inside.” available online. URl: http://www.kidsoutside.info. accessed august 25, 2009.
children and nature network. “children and nature net-work: Building a movement to reconnect children and nature.” available online. URl: http://www.children andnature.org. accessed august 8, 2011.
clayton, Susan, and Gene Myers. Conservation Psychol-ogy: Understanding and Promoting Human Care for Nature. Oxford, U.K.: Wiley-Blackwell, 2009.
connecticut Department of environmental Protection. “no child left inside.” available online. URl: http://www.nochildleftinside.org. accessed May 13, 2011.
Dean, Bradley P., ed. Faith in a Seed: The First Publication of Thoreau’s Last Manuscript. covelo, calif.: island Press, 1993.
Faber taylor, a., et al. “coping with aDD: the surprising connection to green play settings.” Environment and Behavior 33 (2001): 54–77.
———, et al. “Views of nature and self-discipline: evidence from inner city children.” Journal of Environmental Psychology 22 (2002): 49–63.
Fritze, Jessica G., et al. “Hope, despair, and transforma-tion: climate change and the promotion of mental health and wellbeing.” International Journal of Mental Health Systems 2 (2008): 13–22.
Hooked on nature. “tools, resources, and workshops to help people deepen their relationship with the natural world.” available online. URl: http://www.hookedon-nature.org/. accessed May 13, 2011.
Kuo, Frances e., et al. “Fertile ground for community: inner-city neighborhood common spaces.” Ameri-can Journal of Community Psychology 26 (1998): 823–851.
Kuo, Frances e., and a. Faber taylor. “a potential natural treatment for attention-Deficit/Hyperactivity Disor-der: evidence from a national study.” American Jour-nal of Public Health 94 (2004): 1,580–1,586.
Kuo, Frances e., and W. c. Sullivan. “environment and crime in the inner city: Does vegetation reduce crime?” Environment and Behavior 33 (2001): 343–367.
lewis, charles a. Green Nature Human Nature: The Meaning of Plants in Our Lives. Urbana: University of illinois Press, 1996.
louv, Richard. Last Child in the Woods: Saving Our Chil-dren from Nature Deficit Disorder. chapel Hill, n.c.: algonquin Books, 2008.
takano, t., et al. “Urban residential environments and senior citizens’ longevity in megacity areas: the impor-tance of walkable green spaces.” Journal of Epidemiol-ogy and Community Health 56 (2002): 913–918.
Ulrich, R. S., et al. “View through a window may influence recovery from surgery.” Science 224 (1984): 420–421.
———, et al. “effects of environmental simulations and television on blood donor stress.” Journal of Architec-tural and Planning Research 20 (2003): 38–47.
University of illinois at Urbana-champaign. “landscapes and Human Health laboratory.” available online. URl: http://lhhl.illinois.edu. accessed august 8, 2011.
Wilson, edward O. Biophilia. cambridge, Mass.: Harvard University Press, 1986.
eocene epoch See Paleogene period, biodi-versity in.
estuaries See marshes and swamps.
eukaryotes eukaryotes are organisms that con-sist of complex cells. all cells have a cell membrane, which is a thin double layer of lipids that maintains a chemical difference between the inside of a cell and the outside; cytoplasm, which is the liquid con-tents of a cell with its suspended components; ribo-somes, which make proteins; and Dna, which stores genetic information. the simplest cells (though by no means simple) are the prokaryotic cells which have very little more than this. Bacteria and archaea have, in addition, a cell wall and, in some cases, internal membranes.
the cells of protists, fungi, plants, and animals are eukaryotic cells, which are often about 10 times larger than prokaryotic cells and much more com-plex. eukaryotic cells have a nucleus, which contains the Dna in the form of chromosomes inside of a membrane; an internal membrane system involved in making large molecules inside the cell; and organ-elles such as chloroplasts and mitochondria. Eukary-otic refers to the nucleus (Greek for “true kernel”). the Dna molecules in the nucleus are associated with proteins called histones, and the membrane surrounding the nucleus has pores, which are not merely holes but are passageways controlled by pro-teins. these pores allow genetic instructions and
xviii+598_Biodiversity_F.indd 230 11/17/11 1:13 PM

eukaryotes 231
materials, in the form of Rna, to go out into the cytoplasm and allow hormone messages to enter the nucleus. Part of the reason bacteria have rapid metabolism is that as soon as the genetic informa-tion of Dna is transcribed into Rna, the ribosomes start using the information to make proteins. in eukaryotic cells, the Rna must travel out to the cytoplasm first, a process that is slower but allows a greater degree of control over the use of genetic information.
the events involved in the origin of eukaryotes have not been resolved. it is known that chloroplasts and mitochondria are the degenerate evolutionary descendants of prokaryotes that invaded and per-sisted in ancestral eukaryotic cells. the bacterial ancestors of chloroplasts and mitochondria had enough Dna to control their own activities, but chloroplasts and mitochondria do not now have enough genes to survive on their own. Many genes transferred from the primitive chloroplasts and mitochondria to the nucleus of the host eukaryote. in some cases, part of a complex gene transferred to the nucleus and part remained behind in the organelle. an important enzyme in photosynthesis, the rbc gene for the rubisco enzyme, is constructed from genetic instructions in both the chloroplast (the large component, rbcl) and the nucleus (the small component). Mitochondrial genes occasionally act in a manner that is against the best interests of the nucleus (for example, cytoplasmic male sterility fac-tors in plants). Smaller cells moving into and living inside a larger cell is an example of symbiosis, and when it leads to the origin of a new species, it is called symbiogenesis.
the evolution of the nucleus has proven much more difficult to explain. three theories have been proposed, each with provocative evidence. the three theories for the origin of the nucleus are symbio-genesis, the evolution of the nucleus from structures in bacterial ancestors, and that the nucleus evolved from a virus.
The nucleus originated through symbiogenesis. Some evolutionary scientists propose that a primor-dial archaea entered into a larger bacterial host cell and became its nucleus. Subsequently, horizontal gene transfer merged the bacterial and the archaeal Dna. according to this theory, the nucleus was originally archaeal, to which eubacterial genes were added.
it is not clear what kind of bacterium might have been the original host. it may have resembled today’s myxobacteria, which unlike other bacteria use mol-ecules for intercellular communication and can form multicellular structures, just like eukaryotes.
it is also not clear where the membrane that surrounds the nucleus came from. lynn Margulis, who convinced most evolutionary scientists of the
symbiogenetic origin of chloroplasts and mitochon-dria, proposes that the nuclear membrane began as a fusion of the membranes of bacterial and archaeal attachment structures.
The nucleus evolved from structures in bacterial ancestors Most biologists, biology teachers, and biology students assume that eukaryotes have fully formed nuclei, and bacteria have nothing of the sort. it turns out that an obscure group of bacteria called planctomycetes have their Dna almost completely enclosed in a membrane that looks like a nuclear membrane. another group of recently discovered bacteria, which live only inside of sponges, also appear to have structures that look like nuclei. Some researchers even claim that these bacteria have nuclei and that the eukaryotic nucleus may have evolved from this bacterial nucleus. critics point out that, despite appearances, the eukaryotic nuclear mem-brane and the membrane inside these bacteria are structurally very different.
The nucleus evolved from a virus. a few researchers point out that nuclei, like viruses, are genetic material surrounded by a protein coat. they maintain that an ancient virus invaded an archaeal cell, and instead of killing the host, the virus main-tained a multigenerational mild infection inside of it. Gradually, horizontal gene transfer moved bacte-rial genes into the virus, which became the nucleus. their evidence is that nuclei, like viruses, depend on the cell around them for metabolism. and while bacterial Dna molecules are circular, the Dna of viruses is linear, like that of eukaryotic nuclei. Some viruses not only have a protein coat but have an envelope, which they make by stealing a chunk of the host cell membrane, just like the eukaryotic cell uses internal membranes to make the nuclear membrane. Some large viruses even have telomeres, nucleotide sequences at the ends of chromosomes that, in eukaryotes, help keep the process of Dna replication itself from breaking off chunks of essen-tial genes.
these three possibilities are not mere guesses but can be framed as scientific hypotheses. if the first eukaryotic cells were mergers between archaea and bacteria, then some eukaryotic genes should more closely resemble those of archaea, while others should more closely resemble those of bacteria. this appears to be the case. Dna sequences for the genes in eukaryotes that are involved in the use of genetic information resemble archaeal genes. this includes the production of histone proteins, known only from eukaryotic and archaeal cells. Dna sequences for the eukaryotic genes involved in metabolism (energy use and construction of molecules) resemble bacte-rial genes. cell biologists are pursuing the infor-mation necessary to test other hypotheses. if the
xviii+598_Biodiversity_F.indd 231 11/17/11 1:13 PM

first entry232 eukaryotes 232
nucleus began as an extension of a flagellum, which began as an embedded spirochete, as proposed by Margulis and associates, then there should be signif-icant similarities between the proteins that move the chromosomes during cell division, the proteins of eukaryotic flagella, and the proteins of spirochetes. if the first eukaryotic cells evolved from plancto-mycete bacteria, these bacteria should have a closer genetic resemblance to eukaryotes than do other kinds of bacteria.
according to two of these views, the first nucle-ated cell was a chimera, a fusion of two different life-forms. in Greek mythology, a chimera was a combination of two different animals. While it is impossible for two complex multicellular organisms to merge into one, it happens even today on the cellular level and may have been the source of the nucleus. this would mean that the origin of many structures found in the eukaryotic cell occurred through symbiogenesis. if this is the case, the reason that scientists have not found a gradual series of transitional forms in the evolution of eukaryotic cell structures is because they never existed.
the evolutionary patterns constructed from Dna analysis show that the three main branches of life are the archaea, bacteria, and eukaryotes. the branches within the eukaryotes cannot be clearly distinguished from one another. Most of the diver-sity of eukaryotes exists among the primarily single-celled organisms usually called protists (formerly “algae” and “protozoans”). Being usually small, of relatively simple structure, and living in places humans seldom look (such as oceans, ponds, soil, and inside of animal guts), protists have been less intensively studied than plants and animals. not only is less known about protists than about plants and animals, but new groups of protists, including some as small as bacteria, continue to be discovered.
each such discovery has the possibility of changing scientific understanding of eukaryotic evolutionary relationships.
Many eukaryotes are multicellular. Multicellu-larity evolved several times independently (see table): in brown algae, in red algae, in fungi, in plants, and in animals. at least twice, protists have evolved temporary multicellularity: in the cellular and the plasmodial slime molds. each lineage that contains multicellular organisms also contains single-celled organisms.
Some surprises have come from the study of protists. One surprise is that the evolution of chlo-roplasts from cyanobacteria occurred only in the lineage leading to red algae, green algae, and plants. there are other photosynthetic eukaryotes, such as the brown algae and diatoms in one lineage, some euglenas in another lineage, and dinoflagellates in yet another lineage. the chloroplasts in these last three lineages did not evolve directly from cyanobac-teria. instead, they are the evolutionary descendants of eukaryotic algae. this means that eukaryotes such as brown algae and dinoflagellates do not con-sist of cells within cells, but rather cells within cells within cells! another surprise is that fungi and ani-mals are more closely related to each other than to the other eukaryotes.
Within the linnaean system of classification, there are four kingdoms of eukaryotes: protists, fungi, plants, and animals. in this classification, protists are the eukaryotic organisms that do not produce complex tissues. But complex tissues have evolved separately in fungus, plant, and animal lin-eages. the fungi, plants, and animals are therefore branches of the protist kingdom.
the origin of the complex eukaryotic cell allowed an explosion of evolutionary diversification and novelty. although some bacteria can form mul-
major lineages of eukaryotes
lineage Examples
Excavates Single-celled protists without mitochondria (e.g., Giardia)
Discicristates Euglenas, parasitic protists, cellular slime molds
Heterokonts Brown algae, diatoms, water “molds”
Alveolates Ciliates, dinoflagellates, parasitic protists (e.g., Plasmodium)
Cercozoans Shelled amoebae (e.g., foraminiferans)
Green eukaryotes Red algae, green algae, plants
Amoebozoans Plasmodial slime molds, lobed amoebas
Opisthokonts Choanoflagellates, fungi, animals
xviii+598_Biodiversity_F.indd 232 11/17/11 1:13 PM

extinction 233
ticellular filaments or clusters, it appears that only eukaryotes can form complex multicellular organ-isms. the early evolution of eukaryotes occurred during Precambrian time and left few fossils. today scientists attempt to reconstruct these evolutionary events, and to do so they must study the organisms that most closely resemble the earliest eukaryotes: the protists.
See also animals; archaea; bacteria; fungi; Margulis, lynn; protists; seedless plants; seed plants; symbiogenesis
Further readingBaldauf, S. l. “the deep roots of eukaryotes.” Science 300
(2003): 1,703–1,706.Kurland, G. c., l. J. collins, and D. Penny. “Genomics and
the irreducible nature of eukaryote cells.” Science 312 (2006): 1,011–1,014.
lane, nick. Power, Sex, and Suicide: Mitochondria and the Meaning of Life. new york: Oxford University Press, 2005.
Margulis, lynn, Michael F. Dolan, and Ricardo Guerrero. “the chimeric eukaryote: origin of the nucleus from the karyomastigont in amitochondriate protists.” Pro-ceedings of the National Academy of Sciences USA 97 (2000): 6,954–6,959.
———, ———, and Jessica H. Whiteside. “ ‘imperfections and oddities’ in the origin of the nucleus.” Paleobiol-ogy 31 (2005): 175–191.
Morrison, Hilary G., et al. “Genomic minimalism in the early diverging intestinal parasite Giardia lamblia.” Science 317 (2007): 1,921–1,926. Summary by Keel-ing, Patrick J. “Deep questions in the tree of life.” Sci-ence 317 (2007): 1,875–1,876.
Pennisi, elizabeth. “the birth of the nucleus.” Science 305 (2004): 766–768.
Zimmer, carl. “On the origin of eukaryotes.” Science 325 (2009): 666–668.
eutrophication See rivers, streams, ponds, and lakes.
extinction extinction is the disappearance of a species. it usually involves (1) a decrease in popu-lation sizes within a species and (2) a decrease in genetic diversity within a species. these two pro-cesses are related, since small populations usually have less genetic diversity than large populations.
extinction usually occurs for a variety of inter-related reasons and occurs one species at a time. When a large number of species becomes extinct at the same time for the same reason, a mass extinction event is said to have occurred. Five mass extinc-tions have occurred during the history of life and
have been important in the pattern of evolutionary history. Many biologists consider that a sixth mass extinction of species is now occurring, largely due to human disturbance of the environment, and that the mass extinction now under way may well prove to be the biggest in the history of life on earth.
changes in the environment, whether naturally occurring or caused by human activity, may impair the ability of organisms to reproduce; this will cause populations to decline. a species with a small num-ber of populations is in much greater danger of extinction than a species with larger population number and size. For example, a plant species that is limited to a single locality may become extinct if that locality is disturbed by fire, flood, or human activity. the bush Franklinia alatamaha was discovered by the american botanists John and William Bartram in the 18th century. it was growing in just one loca-tion in the mountains of the colony of Georgia, on the plateau adjacent to the altamaha River. the Bar-trams took cuttings from these bushes and planted them in a garden; descendants of these bushes are still alive in gardens throughout the United States and europe. the wild population has disappeared, perhaps when the forest that contained it was cleared for cultivation, perhaps even by a single farmer. the tree Ginkgo biloba was very nearly extinct in its habitat, the forests of north central china. Buddhist priests planted specimens of this tree in their mon-asteries. Ginkgo trees are now abundant throughout the world where they have been planted in cities and more recently in plantations for the production of medicinal compounds. Franklinias and ginkgos survive today largely because a small sample of them was rescued from their very small and vulnerable wild populations.
loss of genetic diversity also makes a species more vulnerable to extinction. a species with fewer genes is less capable of responding through natural selection to changes in the environment. a species with little diversity of genes that confer resistance to specific diseases runs the risk of being devastated by a disease for which it has no resistance. in addition, the loss of genetic diversity can lead to inbreeding. Within most populations, deadly genetic mutations are hidden in the cells of even apparently healthy individuals. in small populations, the chance that two closely related individuals will mate and pro-duce offspring is greater than in large populations. if two individuals that have the same hidden muta-tions mate, these mutations may show up in double dose in the offspring, causing the offspring to die before or shortly after birth. this can be a problem even in a population that has begun to recover from a previous decline. the number of individuals in the population may grow but the number of genes will
xviii+598_Biodiversity_F.indd 233 11/17/11 1:13 PM

first entry234 extinction 234
not, except over a very long period of time. a loss of genetic diversity in the past (a genetic bottleneck) can condemn a population to reduced survival abil-ity even if circumstances should become favorable to its growth.
an example of a species that is near extinc-tion because of a genetic bottleneck is the nene, or Hawaiian goose (Nesochen sandiviensis). Due to the efforts of conservation workers and public cooperation, populations of the nene (the Hawaiian state bird) are growing, following its near extinction in the first half of the 20th century. Dna studies that compare modern birds with birds preserved in museums indicate that modern populations of the nene have much lower genetic diversity than did populations in the 19th century. Many goslings, whether hatched in captivity or in the wild, have birth defects, and many of them die. this is the same reason that small, isolated human populations also exhibit traits that remain hidden in larger popula-tions, for example extra fingers and toes (polydac-tyly) and metabolic disorders.
a third risk factor for extinction is the depen-dence of the species on other species. the fate of a species of butterfly that depends on just one host plant for its caterpillars depends on the fortunes of that host plant.
Rescuing species from extinction may involve a lot more than just making sure the survivors have a place to live. an example is the american chestnut (Castanea dentata). Once a dominant tree in appa-lachian forests, nearly all adult american chest-nut trees were killed by the fungus Cryphonectria (Endothia) parasitica, which was accidentally intro-duced from europe near the beginning of the 20th century. Plenty of american chestnuts remain alive as saplings. they never produce seeds, however, because as soon as the saplings become large enough to have mature bark, the fungus that decimated their original populations kills the sapling. new saplings resprout from the rootstock. a few mature individu-als survive in areas where they were planted far from and before the fungus outbreak occurred. Bring-ing the chestnut back from the brink of extinction involves locating or breeding individuals that are resistant to the fungus.
Rescuing animal species from extinction poses its own set of challenges. Reintroduction of zoo animals into the wild has proven largely unsuccess-ful because animal species that have lived in zoos for several generations have forgotten how to live in the wild and have lost their fear of humans. this is because a lot of animal behavior is passed socially, not genetically, from one generation to the next. On the other hand, some types of natural animal behavior directly hinder the recovery of endangered
species. the magpie robin of the Seychelle islands (Copsychus sechellarum) consisted of only seven breeding pairs in 1988. its population recovered to about 150 but could not exceed this number, since the dominant breeding pairs suppressed the repro-duction of other pairs.
Saving species from extinction can provide a tre-mendous benefit to science and to human well-being. Plant breeders, for example, wish to introduce genes for drought resistance into maize (Zea mays; also called corn) raised in the United States. Drought-resistant genes are available in Tripsacum grass, but Tripsacum cannot crossbreed with maize. a wild species of perennial maize, Zea diploperennis, can crossbreed with both Tripsacum and Zea mays and can thereby act as the bridge for introducing Trip-sacum genes into agricultural maize populations. Zea diploperennis was almost extinct when botanist Hugh iltis discovered it in the 20th century.
Most species have not been identified, and many of these species will become extinct before they are identified. currently, most extinctions occur because of the destruction of habitats, due to urban sprawl, agriculture, soil erosion, and the spread of deserts. nearly half of the terrestrial surface of the earth has been significantly altered by humans. Few wild species can survive in places completely changed by humans, and some wild species will become extinct even in places only slightly altered by humans. Most extinctions occur due to habitat destruction in areas with very high species diversity, known as “biodi-versity hot spots.” in the future, the greatest cause of extinction will be global warming, which has already begun to alter the climatic conditions in nearly all habitats. a severe change in climate can decimate the species in a habitat, even one set aside to preserve those very species, as surely as can a bull-dozer or a tractor. Hunting seldom causes extinc-tion, but there are some exceptions. the Pleistocene extinction may have been caused by overhunting by the first humans to arrive in north america, with similar extinctions in new Zealand and Madagas-car. the dodo (Raphus cucullatus) was hunted to extinction on the indian Ocean island of Mauritius. the passenger pigeon (Ectopistes migratorius), of which billions once darkened the skies and broke the tree boughs of eastern north america, was hunted almost to extinction. Once their populations were small, other processes led to their final disappear-ance. the last one, a female named Martha, died in the cincinnati Zoo on September 1, 1914. White americans with rifles attempted to drive the bison to extinction, partly to undermine the food base of the native americans and make them easier to conquer, but some bison were spared, and today their num-bers are increasing.
xviii+598_Biodiversity_F.indd 234 11/17/11 1:13 PM

extinction 235
On the other hand, it is often difficult to prove that a species has become extinct. the fact that experts cannot find them is pretty good evidence of extinction, but one never knows if a golden toad (Bufo periglenes, one of the few species that many scientists claim have become extinct due directly or indirectly to global warming) might turn up some-where that the experts have not looked. One can be fairly certain that the passenger pigeon is extinct, for surely one would have shown up during the century since Martha died.
Mass extinctions occur when major global events cause the simultaneous extinctions of many species. Five mass extinctions have occurred dur-ing the history of the earth and, according to many scientists, a sixth mass extinction is now under way as a result of human impacts upon the global eco-system. the background rate of extinction, based on the fossil record, is about one species out of 1 mil-lion each year. current extinction rates are at least 100 times as great as this. Many species are almost extinct because their populations are too small. Of the 136 species of native Hawaiian birds, many have become extinct; only 11 of them have populations large enough to ensure survival in the near future. Of the 980 species of native Hawaiian plants, 84 have become extinct, and 133 have populations that consist of fewer than 100 individuals.
While the definition of extinction is fairly clear, the definition of an endangered species is less clear. Government agencies and conservation organiza-tions have adopted guidelines to help identify which species are most in danger of extinction. it is an important matter, as an endangered status may qual-ify the species for protection at some cost to the public and with some restrictions to private land owners. the international Union for conservation of nature (iUcn) publishes the iUcn Red list of threatened Species. a species is considered to be critically threatened if
● more than 80 percent of the population has been lost over the last decade or three gen-erations, whichever is longer; or
● fewer than 250 individuals remain; or ● less than about four square miles (about 10
square kilometers) of habitat remains.
as of 2010, the iUcn red list had 47,677 spe-cies known to be at risk of extinction. this includes 12 percent of bird species; 21 percent of mammal species; 27 percent of reef-building corals; and 30 percent of conifers and cycads.
Some species exist as just a single individual. a single wild tree of Carpinus putoensis remains in the wild, on Mount Froding on Putuo island in china’s
Zhoushan archipelago. a tree hibiscus (Hibiscus liliflorus) of Rodrigues island became extinct after the single surviving individual died. the story of lonely Martha the passenger pigeon continues to be repeated.
in addition to all the other problems faced by endangered species, their novelty and rarity attract collectors. the global black market trade in endan-gered species is worth $10 billion a year, making it second only to drugs and guns. the convention on international trade in endangered Species of Wild Fauna and Flora (citeS) coordinates efforts of dif-ferent countries in an attempt to stop the traffic in endangered species. the rhinoceros is threatened with extinction because of the belief that powder made from its horn has medicinal properties and because the horn is used for handles of traditional arabic jambiya daggers.
Sometimes individuals of endangered species are killed out of desperation. Hunger, caused by among other things a collapse in the availability of fish, has driven many africans to eat bushmeat, which is frequently obtained from endangered species of pri-mates, including gorillas and chimpanzees.
in 1993, the United nations adopted the con-vention on Biological Diversity, which included agreements to protect endangered species. accord-ing to associates of the United nations environment Programme (UneP) World conservation Monitor-ing centre, the rate of biodiversity loss has not decreased in recent years, despite such international efforts and agreements. there have been consistent increases in protection, as indicated by extent of protected areas and sustainable forests. However, the pressures upon biodiversity are increasing (for example, ecological footprint, pollution, invasive species, and overfishing) and the indicators of bio-diversity (such as forest and coral reef extent) are decreasing. Moreover, in most cases, these trends are not very different, per year, since 2000 than since 1970—and in some cases, biodiversity is far-ing even worse (see table on page 236). this indi-cates that the threat of extinction has not declined in the most recent decade, despite great increases in conservation efforts—probably because population and resource pressures are greater than ever and overwhelm conservation efforts.
the extinction of just one species may not seem significant in the context of a million species. But extinctions keep adding up. evolutionary biologist edward O. Wilson says that, through these extinc-tions, nature is dying a “death of a thousand cuts.” conservationists frequently say that “extinction is forever.” this statement is obviously true for a spe-cies—once it is gone it cannot come back—but is also true of the genetic diversity within a species.
xviii+598_Biodiversity_F.indd 235 11/17/11 1:13 PM
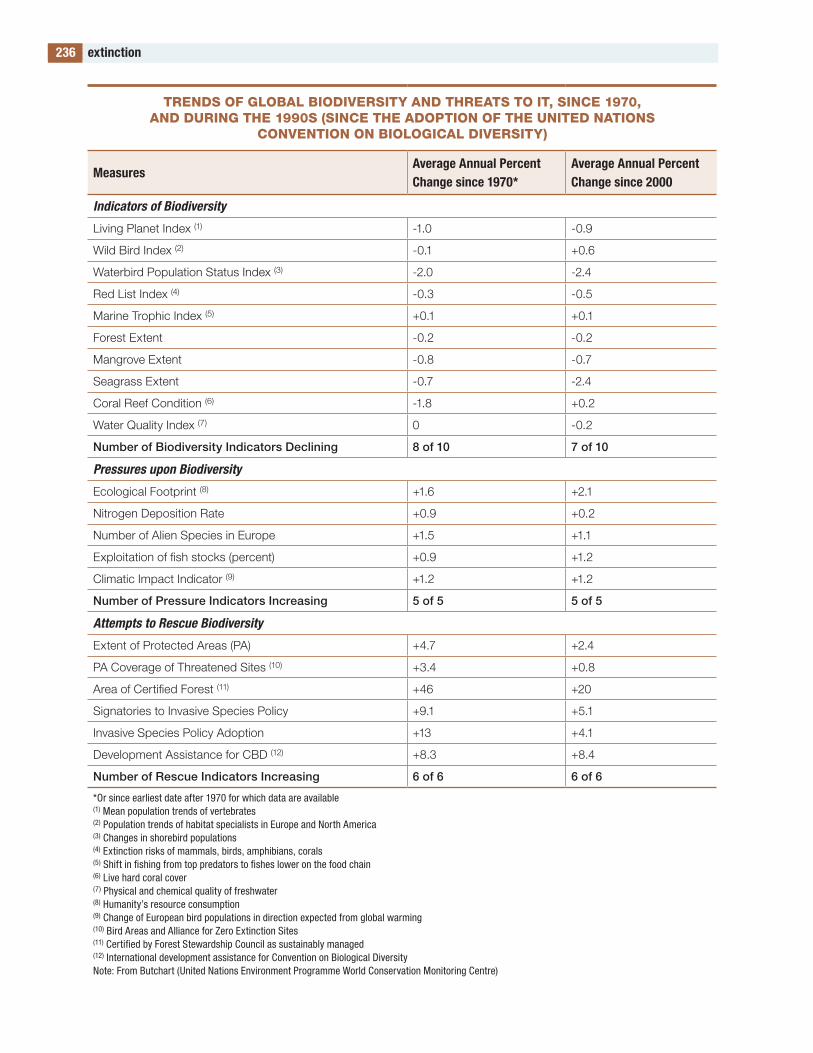
first entry236 extinction 236
trends of global biodiversity and threats to it, sinCe 1970, and during the 1990s (sinCe the adoption of the united nations
Convention on biologiCal diversity)
measuresaverage annual percent Change since 1970*
average annual percent Change since 2000
Indicators of Biodiversity
Living Planet Index (1) -1.0 -0.9
Wild Bird Index (2) -0.1 +0.6
Waterbird Population Status Index (3) -2.0 -2.4
Red List Index (4) -0.3 -0.5
Marine Trophic Index (5) +0.1 +0.1
Forest Extent -0.2 -0.2
Mangrove Extent -0.8 -0.7
Seagrass Extent -0.7 -2.4
Coral Reef Condition (6) -1.8 +0.2
Water Quality Index (7) 0 -0.2
Number of Biodiversity Indicators Declining 8 of 10 7 of 10
Pressures upon Biodiversity
Ecological Footprint (8) +1.6 +2.1
Nitrogen Deposition Rate +0.9 +0.2
Number of Alien Species in Europe +1.5 +1.1
Exploitation of fish stocks (percent) +0.9 +1.2
Climatic Impact Indicator (9) +1.2 +1.2
Number of Pressure Indicators Increasing 5 of 5 5 of 5
Attempts to Rescue Biodiversity
Extent of Protected Areas (PA) +4.7 +2.4
PA Coverage of Threatened Sites (10) +3.4 +0.8
Area of Certified Forest (11) +46 +20
Signatories to Invasive Species Policy +9.1 +5.1
Invasive Species Policy Adoption +13 +4.1
Development Assistance for CBD (12) +8.3 +8.4
Number of Rescue Indicators Increasing 6 of 6 6 of 6
*Or since earliest date after 1970 for which data are available(1) Mean population trends of vertebrates(2) Population trends of habitat specialists in Europe and North America(3) Changes in shorebird populations(4) Extinction risks of mammals, birds, amphibians, corals(5) Shift in fishing from top predators to fishes lower on the food chain(6) Live hard coral cover(7) Physical and chemical quality of freshwater(8) Humanity’s resource consumption(9) Change of European bird populations in direction expected from global warming(10) Bird Areas and Alliance for Zero Extinction Sites(11) Certified by Forest Stewardship Council as sustainably managed(12) International development assistance for Convention on Biological DiversityNote: From Butchart (United Nations Environment Programme World Conservation Monitoring Centre)
xviii+598_Biodiversity_F.indd 236 11/17/11 1:13 PM

extinction 237
See also biodiversity, resources for agri-culture; biodiversity, resources for medi-cine; greenhouse effect; mass extinctions; population genetics.
Further readingamerican chestnut Foundation. “american chestnut
Foundation.” available online. URl: http://www.acf.org. accessed May 13, 2011.
animal Welfare institute. “endangered Species Hand-book.” available online. URl: http://www.endangered specieshandbook.org/dinos.php. accessed May 13, 2011.
Barrett, Meredith a., et al. “citeS designation for endan-gered rosewood in Madagascar.” Science 328 (2010): 1,109–1,110.
Barrow, Mark V., Jr. Nature’s Ghosts: Confronting Extinc-tion from the Age of Jefferson to the Age of Ecology. chicago, ill.: University of chicago Press, 2009.
Brook, B. W., et al. “catastrophic extinctions follow defor-estation in Singapore.” Nature 424 (2003): 420–423.
Bushmeat crisis task Force. “Bushmeat crisis task Force.” available online. URl: http://www.bushmeat.org. accessed May 13, 2011.
Butchart, Stuart H. M., et al. “Global biodiversity: indicators of recent declines.” Science 328 (2010): 1,164–1,168.
cardillo, Marcel, et al. “Multiple causes of high extinction risk in large mammal species.” Science 309 (2005): 1,239–1,241.
collins, James P., and Martha l. crump. Extinction in Our Times: Global Amphibian Decline. new york: Oxford University Press, 2009.
convention on international trade in endangered Spe-cies (citeS). “convention on international trade in endangered Species of Wild Fauna and Flora.” avail-able online. URl: http://www.cites.org. accessed Feb-ruary 25, 2011.
international Union for the conservation of nature. “iUcn Red list of threatened Species.” available online. URl: http://www.iucnredlist.org. accessed May 13, 2011.
Magurran, anne e. “threats to freshwater fish.” Science 325 (2009): 1,215–1,216.
Marton-lefèvre, Julia. “Biodiversity is our life.” Science 327 (2010): 1,179.
McGavin, George c. Endangered: Wildlife on the Brink of Extinction. Richmond Hill, Ont.: Firefly Books, 2006.
Menino, Holly. Darwin’s Fox and My Coyote: Why Sci-ence Alone Can’t Win the Race to Save Wild Animals. charlottesville, Va.: University of Virginia Press, 2008.
Morell, Virginia. “into the wild: Reintroduced animals face daunting odds.” Science 320 (2008): 742–743.
Myers, norman, and andrew Knoll. “the biotic crisis and the future of evolution.” Proceedings of the National Academy of Sciences 98 (2001): 5,389–5,392.
Paddle, Robert. The Last Tasmanian Tiger: The History and Extinction of the Thylacine. new york: cam-bridge University Press, 2002.
Passenger Pigeon Society. “Passenger Pigeon Society.” available online. URl: http://www.passengerpigeon.org. accessed May 13, 2011.
Pimm, Stuart l. “the future of biodiversity.” Science 269 (1995): 347–350.
———, Peter Raven, alan Peterson, Çağan H. Şekercioğlu, and Paul R. ehrlich. “Human impacts on the rates of recent, present, and future bird extinctions.” Proceed-ings of the National Academy of Sciences USA 103 (2006): 10,941–10,946.
Pounds, J. a., et al. “Widespread amphibian extinction from epidemic disease driven by global warming.” Nature 439 (2006): 161–167.
thomas, J. a., et al. “Successful conservation of a threat-ened Maculinea butterfly.” Science 325 (2009): 80–83.
Wright, Karen. “Return of the king of trees.” Discover, May 2004, 58–61.
xviii+598_Biodiversity_F.indd 237 11/17/11 1:13 PM

238
federal government role in conservation (united states) the federal government of the United States has had a larger role in the manage-ment of natural resources and the preservation of wilderness and biodiversity than any other institu-tion in the history of the world. the government acquired this role by historical accident rather than by choice. after the american Revolution, most americans considered it the manifest destiny of the United States to rule the entire north american con-tinent between the borders of canada and Mexico. the american government acquired territories both through purchases from France (the louisiana Pur-chase of 1803) and Mexico (the Gadsden Purchase of 1854) and by war against Mexico and against native american nations. the result was that the govern-ment had millions of square miles of public lands under its control. the General land Offi ce was in the Department of the treasury and the indian affairs Offi ce was part of the Department of War until congress created the Department of the inte-rior to oversee all of the federal land and its native inhabitants in 1849. today, the Department of the interior still oversees about one-fi fth of the land area of the United States. the Department of agriculture oversees the national forests.
european colonists had driven many native american nations to extinction by massacre and dis-ease before the american Revolution, but hundreds of native nations remained in control of their ances-tral lands west of the american frontier. the United States military waged war against many of these nations. after defeating them, the United States government would establish treaties that allowed the nations to keep part of their land—and would then ignore the treaties and take the land. in this
way, native american nations were reduced to poor and generally landless tribes on reservations. in one case, the United States government expelled native americans against the express order of the Supreme court. the cherokee nation sued the United States to prevent its expropriation of their appalachian homelands; the Supreme court, under chief Justice John Marshall, ruled in favor of the cherokees. But President andrew Jackson disregarded the Supreme court order, saying, “let John Marshall enforce it,” and forced the cherokees westward to indian ter-ritory (now Oklahoma) in 1835. During the 20th century, some tribes were partially restored to their ancestral lands; for example, some navahos and apaches returned to arizona from Oklahoma. to this day, native american communities remain gen-erally poor, and in some cases (such as northeastern Oklahoma) their land is severely polluted by indus-trial activities from which the native communities received no profi t.
Pioneers could generally acquire land from which the government had displaced the native residents by moving to it and establishing residency. Proof of residence included cutting down trees and planting farms or orchards. this right was formalized in the 1862 Homestead act, which permitted any family to establish a 160-acre (65 ha) farm. During the second half of the 19th century, the federal government gave thousands of square miles of land to railroad, lumber, and mining corporations at minimal charge. the railroads were built primarily to establish net-works of towns across the continent, connecting the eastern states with california, which had grown to statehood during the gold rush that began in 1849. even when corporations paid the government, it was still practically a giveaway. in the fi rst decade of the
F
xviii+598_Biodiversity_F.indd 238 11/17/11 1:13 PM

federal government role in conservation (united States) 239
20th century, the Weyerhauser corporation paid the government a dime a tree for lumber from the lush forests around Mount St. Helens and Mount Rainier in Washington State.
Wholesale destruction of natural resources was the rule of the day in the late 19th century. entire forests were clear-cut and abandoned. lumberjacks cut down the giant sequoias and coast redwoods of california as fast as they could. People rode trains out into the prairie for the express purpose of shoot-ing entire herds of bison and leaving the carcasses to rot. the federal government supported the bison massacres because they undermined the subsistence economy of the last native american nations to avoid subjugation.
a few pioneers looked up long enough to notice that the land that they were destroying was beauti-ful, often breathtakingly so. there seemed, however, to be no way to preserve any of it. Geologist Ferdi-nand V. Hayden conducted a geological expedition in 1871 to the yellowstone Basin of Wyoming and made a detailed map of its natural wonders, includ-ing geysers and hot springs. His documentation included large photographs by photographer Wil-liam Henry Jackson and huge paintings by american landscape artist thomas Moran. it was the pictures as much as the facts that convinced congress in 1872 to withdraw the land from public auction and make it into the first national park in the United States and in the world. the major purpose of the park was not to preserve natural habitats or species but to create a tourist destination for people riding the newly built railroads.
it was not until the last decade of the 19th cen-tury that the federal government began to preserve a serious amount of public land. Prior to that time the General land Office transferred public land to large railroad, timber, and mining corporations for a min-imal price or no price at all. But a clause in an 1891 act of congress gave the president the power to set aside forestlands. in 1897, just 10 days before leaving office, President Grover cleveland used that power to protect 21 million acres (8.4 million hectares) of forestland in the western United States. congress quickly passed a bill to nullify this protection—a bill that cleveland vetoed as he walked out of the door. the incoming president, William McKinley, trans-ferred the land to the General land Office, which continued the land giveaway.
President McKinley was assassinated in 1901, and his vice president, theodore Roosevelt, took his place. Roosevelt was an avid outdoorsman and nature enthusiast. He formed the United States For-est Service in 1905 with Gifford Pinchot as its first director. With the establishment of the national for-ests, the federal government retained ownership of
the land, established the rules for how it could be used, and collected fees on an ongoing basis for sus-tainable resource extraction. in 1907, a congressio-nal bill forbade the further creation of government forest preserves. in the last days before the bill took effect, Roosevelt and Pinchot set aside 16 million acres (65,000 sq km) of forestland. During the dust bowl era, the federal government acquired much eroded land in the western Great Plains that had been abandoned by farmers, and this land is today the national Grasslands.
puBlIC laNdS IN ThE uNITEd STaTESMost of the public lands (owned by the federal gov-ernment) are managed by the following agencies in the Department of the interior (DOi), Department of Defense (DOD), and Department of agriculture (DOa):
● National Park Service (DOI). the nPS manages national parks, national monu-ments, and other locations whose principal purpose is preservation of natural landscapes and wildlife and public recreation (see table on pages 240–241). yellowstone was the only national park for almost two decades. three more were created before the end of the 19th century: yosemite national Park and Sequoia national Park (california) in 1890, and Mount Rainier national Park (Washington) in 1899. congress passed a na-tional parks bill, formalizing the role of national parks, in 1899. twenty-two more national parks were created between 1900 and 1935. there are now 58 national parks in the United States. the national park Ser-vice was created as an agency within the DOi in 1916 by a bill that required the agency “to conserve the scenery and the nat-ural and historic objects and wildlife therein, and to provide for the enjoyment of the same in such manner and by such means as will leave them unimpaired for the enjoyment of future generations.” national parks are, therefore, intended for the preservation of nature and of historically important sites. national monuments are lands that are not considered quite significant enough to qual-ify for national park status, but in recent de-cades many national monuments (such as Death Valley) have become national parks.
● United States Forest Service (DOA). national parks are intended for preserva-tion, following the vision of american con-servationist John Muir. in contrast, the national forests are intended for managed
xviii+598_Biodiversity_F.indd 239 11/17/11 1:13 PM
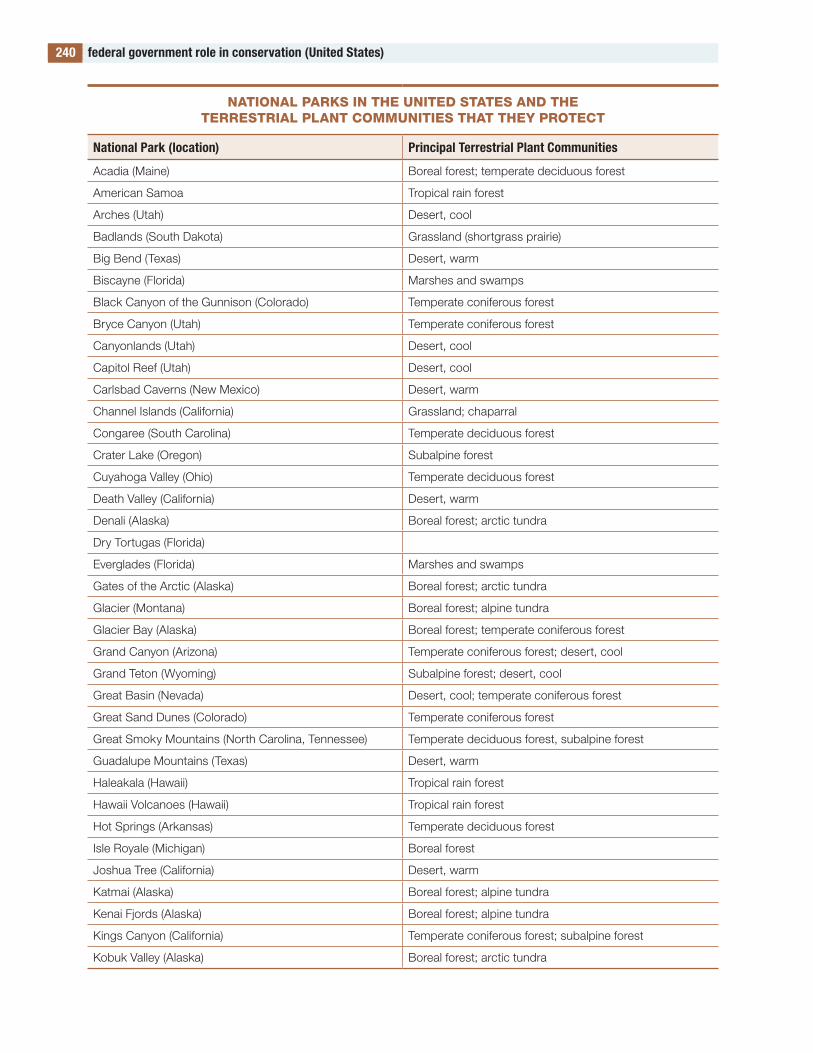
first entry240 federal government role in conservation (united States) 240
national parks in the united states and the terrestrial plant Communities that they proteCt
National park (location) principal Terrestrial plant Communities
Acadia (Maine) Boreal forest; temperate deciduous forest
American Samoa Tropical rain forest
Arches (Utah) Desert, cool
Badlands (South Dakota) Grassland (shortgrass prairie)
Big Bend (Texas) Desert, warm
Biscayne (Florida) Marshes and swamps
Black Canyon of the Gunnison (Colorado) Temperate coniferous forest
Bryce Canyon (Utah) Temperate coniferous forest
Canyonlands (Utah) Desert, cool
Capitol Reef (Utah) Desert, cool
Carlsbad Caverns (New Mexico) Desert, warm
Channel Islands (California) Grassland; chaparral
Congaree (South Carolina) Temperate deciduous forest
Crater Lake (Oregon) Subalpine forest
Cuyahoga Valley (Ohio) Temperate deciduous forest
Death Valley (California) Desert, warm
Denali (Alaska) Boreal forest; arctic tundra
Dry Tortugas (Florida)
Everglades (Florida) Marshes and swamps
Gates of the Arctic (Alaska) Boreal forest; arctic tundra
Glacier (Montana) Boreal forest; alpine tundra
Glacier Bay (Alaska) Boreal forest; temperate coniferous forest
Grand Canyon (Arizona) Temperate coniferous forest; desert, cool
Grand Teton (Wyoming) Subalpine forest; desert, cool
Great Basin (Nevada) Desert, cool; temperate coniferous forest
Great Sand Dunes (Colorado) Temperate coniferous forest
Great Smoky Mountains (North Carolina, Tennessee) Temperate deciduous forest, subalpine forest
Guadalupe Mountains (Texas) Desert, warm
Haleakala (Hawaii) Tropical rain forest
Hawaii Volcanoes (Hawaii) Tropical rain forest
Hot Springs (Arkansas) Temperate deciduous forest
Isle Royale (Michigan) Boreal forest
Joshua Tree (California) Desert, warm
Katmai (Alaska) Boreal forest; alpine tundra
Kenai Fjords (Alaska) Boreal forest; alpine tundra
Kings Canyon (California) Temperate coniferous forest; subalpine forest
Kobuk Valley (Alaska) Boreal forest; arctic tundra
xviii+598_Biodiversity_F.indd 240 11/17/11 1:13 PM

federal government role in conservation (united States) 241
use, following the vision of american for-ester Gifford Pinchot. the national forest slogan is “land of Many Uses,” of which recreation and protection of species and habitats is just one. timber and mineral extraction is permitted from national for-ests, and they contain many private land-holdings. there are 155 national forests. the federal government controls which trees can be removed and how. For example, timber extraction, but not of giant sequoia trees, is permitted in Sequoia national For-est in california.
● Bureau of Land Management (DOI). the BlM manages hundreds of thousands of square miles of woodland, grassland, and desert in the western United States. One example is the national Grasslands. the national Grasslands system began when the federal government bought eroded land from farmers who moved out of the western Great Plains during the dust bowl of the 1930s. today, the government leases most
of these grasslands to ranchers for cattle grazing. the federal government sets con-servation rules for these lands: For example, they control the number of cattle and the conservation of endangered black-footed ferrets in the Buffalo Gap national Grass-land in South Dakota.
● Fish and Wildlife Service (DOI). the FWS oversees the system of national Wildlife Ref-uges. they are small areas set aside as habitat for wildlife. there are 551 national Wildlife Refuges and 37 wetland management dis-tricts in the national Wildlife Refuge sys-tem. the first national Wildlife Refuge was Pelican island, off the coast of Florida. it was created by President theodore Roosevelt in 1903. He asked his advisers if there was any law that prohibited him from establishing this refuge. Hearing no objections, Roosevelt said, “then i do so declare it.”
● Department of Defense. Military bases around the United States contain thousands of square miles of uninhabited and largely
National park (location) principal Terrestrial plant Communities
Lake Clark (Alaska) Boreal forest; alpine tundra
Lassen Volcanic (California) Subalpine forest; alpine tundra
Mammoth Cave (Kentucky) Temperate deciduous forest
Mesa Verde (Colorado) Temperate coniferous forest
Mount Rainier (Washington) Subalpine forest; alpine tundra
North Cascades (Washington) Subalpine forest; alpine tundra
Olympic National Park (Washington) Temperate coniferous forest; subalpine forest
Petrified Forest (Arizona) Desert, warm
Redwood (California) Temperate coniferous forest
Rocky Mountain (Colorado) Subalpine forest, alpine tundra
Saguaro (Arizona) Desert, warm
Sequoia (California) Temperate coniferous forest; subalpine forest; alpine tundra
Shenandoah (Virginia) Temperate deciduous forest
Theodore Roosevelt (North Dakota) Grassland (shortgrass prairie)
Virgin Islands Tropical rain forest
Voyageurs (Minnesota) Boreal forest
Wind Cave (South Dakota) Temperate coniferous forest
Wrangell-St. Elias (Alaska) Boreal forest; subalpine tundra
Yellowstone (Wyoming) Subalpine forest; alpine tundra
Yosemite (California) Temperate coniferous forest; subalpine forest
Zion (Utah) Desert, cool
xviii+598_Biodiversity_F.indd 241 11/17/11 1:13 PM

first entry242 federal government role in conservation (united States) 242
natural land. Since the military uses these lands for military training and as bombing ranges, they are not undisturbed, but there are no trespassers. in recent decades, the Department of Defense has made serious efforts to protect habitats and wildlife in these areas as far as is possible consistent with military uses, and scientific research is often conducted on these lands.
● Army Corps of Engineers (DOD). Begin-ning in the 1930s, the federal government used federal land and acquired private land to create reservoirs for flood con-trol. Because reservoirs get larger as they store floodwater, the army corps has also acquired much of the land around the mar-gins of the reservoirs. Much of this land is leased for limited recreational development or used for ecological research.
● Bureau of Reclamation (DOI). this gov-ernment agency, like the army corps, cre-ated reservoirs, but with a different purpose: the development of irrigation.
in addition, there are 756 Wilderness areas. the Wilderness act of 1964 gave the federal government the power to designate Wilderness areas within fed-eral lands that were already protected by one of the foregoing agencies. For example, Black elk Wilder-ness area was established within the Black Hills national Forest. Wilderness areas have no develop-ment and only primitive trail access.
fEdEral JurISdICTIoN IN ThE CoNTrol of ENvIroNmENTal proBlEmS
During the 20th century, it became apparent that many environmental problems could not be solved by uncoordinated actions of state and local govern-ments or private corporations. Primary examples are soil erosion, water resources, pollution, and endan-gered species.
Soil erosion During the dust bowl of the late 1920s and 1930s, millions of tons of topsoil from the western great plains of the United States blew for thousands of miles in the air, leaving a withered and almost sterile substrate that would grow very little food or forage. in many Midwestern towns, midday became as black as night, and some people even died from the inhalation of dust. this was at the end of a long drought; but the dust storms were not completely the fault of the drought. they were also the result of overgrazing and excessive plowing, which left the soil that could otherwise have survived a drought vulnerable to the relentless wind.
in april 1935, during the height of the dust bowl, soil scientist Hugh Bennett met with the U.S.
Senate Public lands committee. He wanted to con-vince them to create a government agency to combat soil erosion. When the clouds of dust blew over Washington D.c., he invited the senators to look out the window. they saw the nation’s farmlands, upon which both the present and the future of the country depended, blowing out to sea. the Senate established the Soil conservation Service (now the natural Resources conservation Service) of the U.S. Department of agriculture on april 27, 1935. this agency helps farmers to incorporate new techniques that will reduce the loss of topsoil to the forces of wind and rain. although cooperation is voluntary, nearly every county in the United States is also a Soil conservation District.
Water resources. During the 20th century, the federal government also began to assume an impor-tant role in the management of water resources, primarily by the construction of large dams, which not only created reservoirs but also electricity. Dam construction was a major component of federal job creation during the depression. as explained previ-ously, the army corps of engineers constructed dams that were primarily intended for flood control, while the Bureau of Reclamation built dams that fed into irrigation systems. aside from this, however, water resource policy in the United States, and the federal government’s role in it, are often unclear. For exam-ple, the tarrant Regional Water District (Fort Worth, texas) sued the state of Oklahoma in 2007 because, it claimed, Oklahoma’s moratorium on out-of-state water sales violated the commerce clause of the con-stitution. the case continues in court as of 2011.
Pollution and endangered species. environ-mental problems such as pollution went largely unchecked into the 1960s. a government biologist, Rachel carson, wrote a book in 1962 that quickly gained national and international attention. Silent Spring presented extensive documentation that the widespread use of pesticides on the american landscape was producing severe pollution, much of which was concentrated up the food chain and killing unintended targets such as birds (hence the title of her book). chemical corporations attacked her, but congressional hearings substantiated her claims. Scientists verified that birds at the top of the food chain, such as eagles and pelicans, were dying because the pesticide DDt, concentrated in their tissues, interfered with their ability to lay eggs. carson demonstrated, moreover, that insect and other pest populations evolved resistance to those pesticides, making them ineffective. at this same time, awareness of the world population explo-sion was spreading. among the iconic events that spurred environmentalism were the Santa Barbara oil spill and the cuyahoga River in Ohio, which
xviii+598_Biodiversity_F.indd 242 11/17/11 1:13 PM

federal government role in conservation (united States) 243
caught on fire due to petroleum pollution, both in 1969. the environmentalism of the 1960s was a brief moment of intense social and political trans-formation, because it occurred at the same time as protests against the Vietnam War and against racial segregation. Protestors saw corporate profits as a motivating force behind pollution as well as the war. Since that time, much larger environmental disasters have failed to generate nearly as much protest. the Deepwater Horizon (BP) oil spill of 2010 is the larg-est environmental disaster in american history to date, yet did not have the kind of effect on national policy that the comparatively small disasters of the 1960s did.
it is therefore no surprise that, in quick suc-cession, the federal government adopted sweeping changes in environmental protection, including the following:
● National Environmental Policy Act (NEPA). this law, passed by congress in 1969, cre-ated the environmental Protection agency (ePa), which has the authority to enforce environmental legislation. ePa monitors state and local governments and corporations for compliance with federal environmen-tal laws and must approve environmental impact Statements before the federal govern-ment can undertake any significant activity that may have an impact on the environment. the ePa banned the widespread use of the pesticide DDt in 1972, allowing its use to control the spread of disease vector insects.
● Clean Air Act. this law, passed by con-gress in 1970, required that industry, utili-ties, and transportation reduce air pollution. One example of a technology that arose to control air pollution is the catalytic con-verter. incomplete combustion in internal combustion engines produces toxic carbon monoxide; the converter uses a palladium catalyst to make the carbon monoxide into carbon dioxide. the law, still in effect, has been very successful. emissions of lead, primarily from gasoline, have dropped 98 percent. emissions of sulfur dioxide, mostly from power plants, have dropped by 35 percent even though the gross domes-tic product has more than doubled since 1970. emissions of carbon monoxide have dropped by 32 percent even though driving has increased 127 percent. amendments to the act helped to reduce acid rain and fog, and in 2009 the ePa declared carbon diox-ide to be a pollutant because of its role in the greenhouse effect.
● Clean Water Act. this law, passed by congress in 1972, prohibited pollution from point sources, that is, specific locations from which pollution was released, which includes industries and feedlots. nonpoint sources, such as storm runoff and drain-age from agricultural lands, were exempted, although a 1987 amendment required the separation of sewage from storm runoff in municipal drainage systems that had previ-ously combined them.
● Endangered Species Act (ESA). this law, passed by congress in 1973, established a mechanism by which scientific data could be gathered and used to determine which species were threatened by human activities and then to enable protection of those spe-cies. the law, administered by the FWS (on land and in freshwater) and the national Oceanic and atmospheric administration (in the oceans), protects not only those spe-cies but the habitats on which they depend, and for this reason many scientists consider the eSa to be the single most important mechanism for the protection of biodiver-sity in the United States. the purpose of the law is not, however, to identify spe-cies for permanent protection but to restore them to a nonthreatened status. therefore, whenever an agency or a group of citizens proposes a species for federal protection under this act, they must include a recovery plan, by which the species can be brought back to safe population levels. in some high-profile cases, species that were threat-ened with extinction have been delisted. the FWS removed the bald eagle from fed-eral protection in 2007, as its populations had sufficiently recovered, largely because of the disappearance of DDt from the food chain and the protection of nesting site habitats. the bald eagle continues to be protected by the Bald and Golden eagle Protection act of 1940 and the Migra-tory Bird treaty. While this was greeted as evidence of success in conservation, the delisting of wolves has proven controver-sial, because as soon as federal protection for wolves was lifted in 2008, bands of hunters in Wyoming and Montana orga-nized what many conservationists consider pointless slaughters.
all of these laws were enacted during the presi-dency of Richard nixon, who was not a particu-larly passionate environmentalist. He recognized,
xviii+598_Biodiversity_F.indd 243 11/17/11 1:13 PM

first entry244 ferns 244
however, that it was political suicide to resist the environmental movement of the 1960s.
Federal government protection of resources on public lands is imperfect. the most notorious exam-ple is the Minerals Management Service (DOi) [now the Bureau of Ocean energy Management, Regula-tion and enforcement], which regulates the leasing of public lands for coal, oil, and mineral extraction by private corporations and is responsible for col-lecting revenues from those corporations. in 2008, the news broke that up to one-third of the employ-ees in the Denver office of the MMS accepted gifts (which included sex and drugs) from corporations in return for profitable leases and lax collection of fees. in 2010, the news broke of lax enforcement of safety violations by the new Orleans office of the MMS, which may have played a direct role in allowing the Deepwater Horizon oil spill to occur. Despite these scandals, public lands appear to be better protected in the United States than in many other countries. conservationists often refer to national parks in many countries as “paper parks,” meaning that they are protected only by lines drawn on a map, not by enforcement. although 12 percent of the earth’s land surface has official protection, conservationists estimate that only about 0.5 percent is protected by meaningful enforcement.
the United States participates in some interna-tional agreements to protect biodiversity, such as the convention on international trade in endan-gered Species (citeS). Most of these endangered species on the international market come from out-side the United States. However, there is an inter-national market for products from some american species. For example, the eggs of paddlefish, illegally obtained in the United States by the Russian mafia, are marketed as caviar in eastern europe.
Due to limited resources and political pressure, the federal government cannot assume the complete role of protecting biodiversity. Private organizations (most notably the nature conservancy) purchase land or receive it by donation, then manage it to pro-tect the integrity of the ecological communities in it.
although the federal government has only the powers delegated to it by the constitution, it plays an overwhelming part in the protection of biodi-versity, because federal lands are so extensive and because many pollution problems as well as spe-cies cross state lines. the federal government will remain the single most powerful force in biodiversity protection in the United States indefinitely into the future. it also plays an essential role by approv-ing international agreements on biodiversity protec-tion, pollution, and carbon emissions—and by not approving them. the single most important reason that carbon dioxide emissions have not been ade-
quately controlled around the world is probably that the Republican Party has refused to allow the federal government to commit to international agreements that limit them.
See also acid rain and fog; biomagnification; carson, Rachel; endangered species; green-house effect; Muir, John; Pinchot, Gifford.
Further readingBrinkley, Douglas. The Wilderness Warrior: Theodore
Roosevelt and the Crusade for America. new york: Harper Perennial, 2010.
Duncan, Dayton, and Ken Burns. The National Parks: America’s Best Idea. new york: Knopf, 2009.
egan, timothy. The Big Burn: Teddy Roosevelt and the Fire that Saved America. Boston: Houghton Mifflin, 2009.
———. The Worst Hard Time: The Untold Story of Those Who Survived the Great American Dust Bowl. Bos-ton: Houghton Mifflin, 2006.
Flippen, J. Brooks. Nixon and the Environment. albuquer-que: University of new Mexico Press, 2000.
Fretwell, Holly lippke. Who is Minding the Federal Estate?: Political Management of America’s Public Lands. lanham, Md.: lexington Books, 2009.
langholz, Jeff. “Saving nature privately.” Worldwatch, Sep-tember/October 2009, 7–11.
Merchant, carolyn. American Environmental History: An Introduction. new york: columbia University Press, 2007.
natural Resources conservation Service. “Biography of Hugh Hammond Bennett.” available online. URl: http://www.nrcs.usda.gov/about/history/bennett.html. accessed June 24, 2010.
nature conservancy. “the nature conservancy.” available online. URl: http://www.nature.org. accessed august 14, 2010.
Savage, charlie. “Sex, drug use and graft cited in inte-rior Department.” New York Times, Septem-ber 11, 2008. available online. URl: http://www.nytimes.com/2008/09/11/washington/11royalty.html?pagewanted=print. accessed May 13, 2011.
ferns See seedless plants.
fishes Fishes were the first vertebrates. there are more species of fishes than of amphibians, reptiles, birds, or mammals. they are the only group of ver-tebrates that is almost exclusively aquatic, which was the primitive condition of all life. When referring to more than one individual of the same species, “fish” is the correct plural; for more than one species, “fishes” is correct. Since one of the lineages of fishes evolved into the tetrapods (amphibians, reptiles,
xviii+598_Biodiversity_F.indd 244 11/17/11 1:13 PM
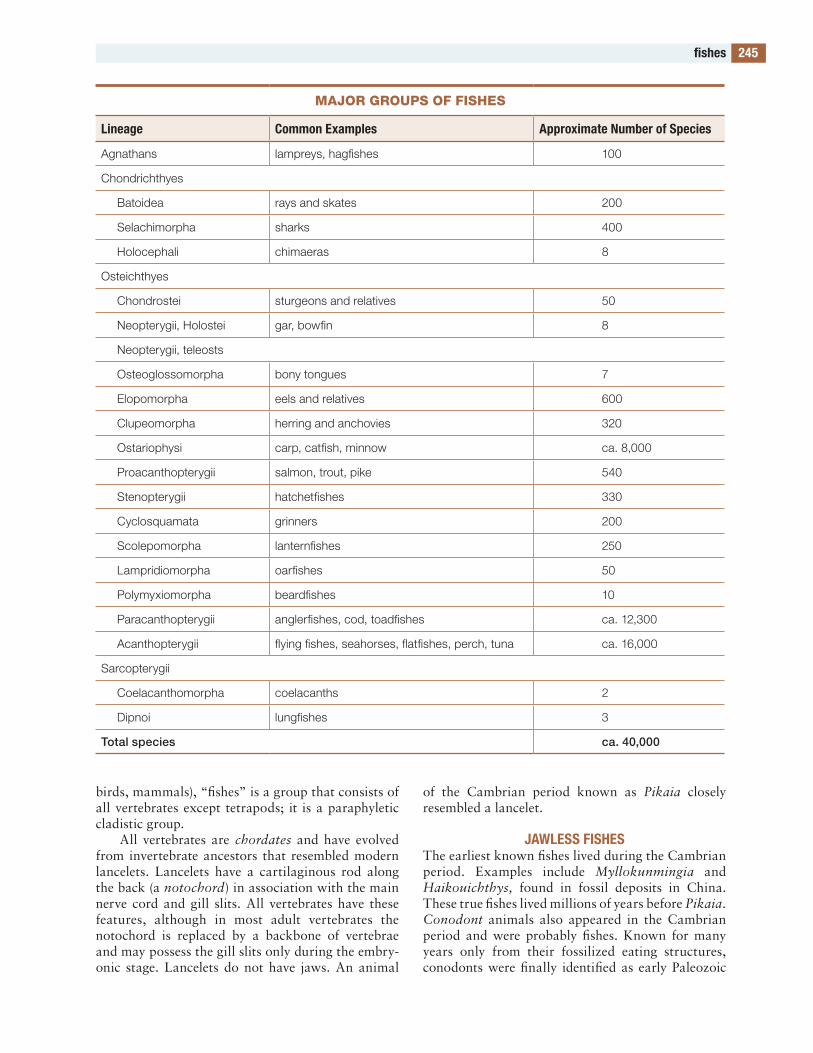
fishes 245
birds, mammals), “fishes” is a group that consists of all vertebrates except tetrapods; it is a paraphyletic cladistic group.
all vertebrates are chordates and have evolved from invertebrate ancestors that resembled modern lancelets. lancelets have a cartilaginous rod along the back (a notochord) in association with the main nerve cord and gill slits. all vertebrates have these features, although in most adult vertebrates the notochord is replaced by a backbone of vertebrae and may possess the gill slits only during the embry-onic stage. lancelets do not have jaws. an animal
of the cambrian period known as Pikaia closely resembled a lancelet.
JawlESS fIShESthe earliest known fishes lived during the cambrian period. examples include Myllokunmingia and Haikouichthys, found in fossil deposits in china. these true fishes lived millions of years before Pikaia. Conodont animals also appeared in the cambrian period and were probably fishes. Known for many years only from their fossilized eating structures, conodonts were finally identified as early Paleozoic
major groups of fishes
lineage Common Examples approximate Number of Species
Agnathans lampreys, hagfishes 100
Chondrichthyes
Batoidea rays and skates 200
Selachimorpha sharks 400
Holocephali chimaeras 8
Osteichthyes
Chondrostei sturgeons and relatives 50
Neopterygii, Holostei gar, bowfin 8
Neopterygii, teleosts
Osteoglossomorpha bony tongues 7
Elopomorpha eels and relatives 600
Clupeomorpha herring and anchovies 320
Ostariophysi carp, catfish, minnow ca. 8,000
Proacanthopterygii salmon, trout, pike 540
Stenopterygii hatchetfishes 330
Cyclosquamata grinners 200
Scolepomorpha lanternfishes 250
Lampridiomorpha oarfishes 50
Polymyxiomorpha beardfishes 10
Paracanthopterygii anglerfishes, cod, toadfishes ca. 12,300
Acanthopterygii flying fishes, seahorses, flatfishes, perch, tuna ca. 16,000
Sarcopterygii
Coelacanthomorpha coelacanths 2
Dipnoi lungfishes 3
Total species ca. 40,000
xviii+598_Biodiversity_F.indd 245 11/17/11 1:13 PM

first entry246 fishes 246
fishes when a fossil was found in which numerous conodont eating structures were imbedded within a shadowy eel-like creature with large eyes at the loca-tions where one would expect teeth to be. (Since the exact function of the “teeth” is unknown, the eating structures are called “conodont elements.”)
neither conodonts nor the fishes of the cam-brian or early Ordovician period had jaws. two classes of modern agnathan (jawless) fishes survive: Lampreys live by sucking blood from larger fishes (see figure), and hagfishes feed on carrion. Hagfishes are classified together with the vertebrates even though they have no vertebrae, ribs, or jaws. agna-thans do not have paired fins; they have fins only along the dorsal (back) and ventral (front or belly) surfaces. they do not have eyes that are homologous to the eyes of most vertebrates; they have a pineal eye that is homologous to the pineal gland in the heads of vertebrates. they do not have scales. lam-preys dig shallow nests in the bottom of the body of water, into which they place eggs and sperm. the fertilized eggs hatch into a larval form that looks more like a lancelet than an adult lamprey.
One lineage of fishes evolved jaws during the Ordovician period. this allowed a major evolu-tionary advancement in the efficiency of predation. Many of the earliest jawed fishes, including the placoderms, were covered with bony armor, as the predatory arms race became severe. Placoderms had bony plates in their mouths that served in place of teeth. Placoderms apparently became extinct with-out descendants. Other lineages of jawed fishes sur-vived to become the chondrichthyes (cartilaginous fishes) and the Osteichthyes (bony fishes). Modern fishes have less armor and are more maneuverable than placoderms were.
CarTIlaGINouS fIShESCartilaginous fishes include the sharks and rays. their skeletons consist only of cartilage. as everyone knows, they have teeth; but teeth are not bones—shark teeth are structurally similar to their scales and are often the only part of the shark to be pre-served as fossils. Sharks have sharp incurved teeth, sometimes several rows of them. Sharks are continu-ally producing new teeth; an individual shark can lose many thousands of teeth in a lifetime. their jaws are not connected to their skulls. Shark bones do not have marrow. their red blood cells are pro-duced in the spleen.
Sharks and rays have the following kinds of fins:
● they have unpaired fins which include the dorsal fin along the back (top); the anal fin along the front (bottom); and the caudal fin (the tail). Rays do not have anal fins.
● they have paired pectoral (“breast”) and pelvic (“hip”) fins, on each side, connected to one another. the inflexible horizontal pectoral fins of sharks make them much less maneuverable than other fishes; once they have begun moving rapidly, they can-not stop or back up. the pectoral fins of rays are much more flexible.
not all sharks are large. the dwarf lantern shark Etmopterus perryi is only seven inches (17 cm) long. the largest shark, and the largest fish, is the whale shark Rhincodon typus, which can be almost 40 feet (12 m) long. Whale sharks, like the baleen whales to which they are quite unrelated, eat only by filter feed-ing, though sometimes they swallow squid and fishes.
Sharks have keen senses. they can detect blood at a concentration of as little as a part per million. they have good senses of sight and hearing but also rely very much on electroreception. all muscle movements produce electric fields, and sharks may have thousands of receptors that are sensitive to these electric fields. this allows sharks to find even prey that are hidden in the sand, as their muscles produce electrical fields. Hammerhead sharks have nostrils and electroreceptors spaced widely apart on the front of the head, allowing a more accurate detection of the locations of prey.
Most sharks have internal fertilization, in which males transfer sperm directly to females. Some shark species have live birth, in which the embryos, con-nected by an umbilical cord, develop inside the mother. live birth in sharks evolved independently from live birth in some reptiles and the live birth that characterizes most mammals. Other sharks release an egg capsule, containing the developing embryo, into the environment.
Mouth of a lamprey, which is a jawless fish that has invaded the Great Lakes. It embeds its teeth into the skins of larger fishes and sucks blood from them. (Great Lakes Image Collection, EPA)
xviii+598_Biodiversity_F.indd 246 11/17/11 1:13 PM

fishes 247
Sharks have tremendous evolutionary diversity. tiger sharks, the “garbage cans of the sea,” will eat almost anything. Biologist edward O. Wilson reports that one giant tiger shark’s stomach con-tained three overcoats, a raincoat, a driver’s license, a cow’s hoof, the antlers of a deer, twelve lobsters, and a chicken coop with feathers and bones still inside. in contrast, the great white shark selectively hunts seals, sometimes catching humans by mis-take. Sharks are fierce predators but not of humans; worldwide, there are about four people killed each year in unprovoked attacks. the small cookie-cutter shark takes shallow bites out of the skins of larger animals, sometimes chewing the plastic coatings of submarine navigation equipment by mistake. Chi-maeras, or ghost sharks, resemble sharks but have no scales and instead of sharp teeth have three pairs of grinding tooth plates in their mouths. the largest shark to have ever lived, Carcharodon megalodon, could reach over 50 feet (16 m) in length. it became extinct only about a million and a half years ago.
Rays are vertically flattened, very broad carti-laginous fishes that swim along the bottom of the sea in search of prey. their gill slits are on the ven-tral surface, behind the mouth, rather than on the side of the head as in sharks. the pectoral fins can be very broad and winglike. Muscles in the “wings” of some ray species generate enough electric current to immobilize prey (these are called “electric rays”). they have tails that can be much longer than the rest of the body, and in one family of rays (the sting-rays), venomous spines in the tail can kill a human-sized animal.
BoNy fIShESBony fishes evolved in freshwater conditions and were restricted to freshwater for the first 160 million years of their existence. evolutionary scientists spec-ulate that their bones were more important as a way of storing calcium, a mineral that could be scarce in freshwater, than as a skeletal reinforcement. today, 41 percent of fish species are freshwater species. this may have resulted because systems of lakes and rivers are largely isolated from one another, while marine fishes can swim over large areas. Bony fishes diverged into two major lineages:
● the ray-finned fishes, which have fins rein-forced with bony rays that do not correspond to the fingers or toes of other vertebrates. the major lineage of ray-finned fishes is the teleosts, which includes most modern fish species.
● the flesh-finned fishes diverged into two lineages: the lungfishes and the crossopter-ygians. Some lungfishes survive today in
shallow tropical ponds. When the ponds have water, the oxygen levels are low and the lungfishes gulp air. When the ponds dry up, the lungfishes continue to breathe while estivating in the mud. the crossopterygian fishes have bones at the bases of their fins that correspond to the one upper and two lower limb bones of tetrapods (four-legged animals). two branches of crossopteryg-ian fishes became the coelacanths and the tetrapods.
Ray-finned fishes live in nearly all aquatic envi-ronments. they have an astonishing variety of body forms and range in size from Paedocypris progenet-ica, in which adults can be as small as a third of an inch (8 mm), to the ocean sunfish Mola mola, which can weigh more than 5,000 pounds (2,300 kg), and the oarfish Regalecus glesne, which can be 36 feet (11 m) long.
the sturgeons and their relatives are among the most primitive bony fishes, having changed little over the last 200 million years. they do not have scales, and their skeletons are primarily cartilaginous. they spawn in freshwater, where some of them remain while others migrate to shallow marine waters. Because they grow slowly and mature after only many years, overfishing is particularly dangerous to the survival of these species. among the relatives of sturgeons are african fishes known as bichirs. they have undergone less evolutionary change since the common ancestor of ray-finned and flesh-finned fishes; they share some characteristics with the flesh-finned fishes, including primitive lungs. the stur-geons, bichirs, and paddlefishes constitute a separate lineage (chondrostei) from all the other ray-finned fishes (neopterygii).
the gars and bowfins are the most primitive members of the neopterygii lineage of ray-finned fishes. Gars live in shallow water and catch prey with their sharp teeth in elongated snouts. their swim bladders function partly as lungs.
all of the neopterygii other than gars and bow-fins are teleosts. teleost fishes can protrude their jaws outward, and (unlike other bony fishes and unlike sharks) their vertebrae do not extend into their tails. the relatively minor anatomical differ-ence between teleosts and other fishes does not explain why teleosts have had explosive evolution-ary success. Major lineages of teleosts are listed in the table on page 245, and major superorders of teleosts are presented below:
Eels (see figure on page 248) are long skinny fishes that have no pelvic fins and most also lack pec-toral fins. their dorsal, anal, and caudal fins are fused into a single ribbon that runs along the entire body.
xviii+598_Biodiversity_F.indd 247 11/17/11 1:13 PM

first entry248 fishes 248
the giant moray eel (Gymnothorax javanicus) can be over 12 feet (almost 4 m) in length. eels are predators.
Herrings and anchovies are small, narrow, sil-very fishes that live in large groups and consume mostly plankton. especially because of their large numbers, they are important to marine food webs.
the ostariophysians are the second largest superorder of fishes and the largest group of fresh-water fishes. examples include the following:
● Carp and minnows carp (Cyprinus carpio) are often farmed inland and are an impor-tant food source. Domesticated asian koi are just colorful carp.
● Goldfishes (Carassius auratus) ● Catfishes are bottom-feeders in freshwater.
their flattened heads, lack of buoyancy, and their sensory barbels (which they use more than their small eyes) allow them to navigate on the bottoms of ponds. the front sac of the air bladder is connected by bone to the inner ear, greatly enhancing their sense of hearing. they have no scales, and many breathe through their skin. catfishes are also farmed inland and an important food source.
● Electric eels (Electrophorus electricus), even though they are long and skinny, are ostari-ophysians, not eels. they live in rivers of the amazon basin. the organs that produce electricity make up four-fifths of their bod-ies. they can produce a 500-volt charge that can kill prey. they also use electric fields for electrolocation of prey and landmarks.
● Piranhas have big teeth and a reputation for group ferocity against prey. However, they are not known for unprovoked attacks. Some piranha species are scavengers or eat seeds.
Salmon and trout are the best-known members of the superorder Protacanthopterygii. they are predators that breed in fresh cool upstream waters. trout remain in these waters throughout their lives, but salmon are anadromous, which means that they live in the ocean but return to the streams in which they hatched to spawn. Upstream migration of salmon (salmon runs) are dramatic, as the fish even jump up waterfalls (see figure on page 249). they have fatty adipose fins behind their dorsal fins.
Most members of the superorder Stenoptery-gii, such as dragonfishes and hatchetfishes, live in deep oceans and have special organs that produce white, yellow, red, or purple bioluminescence. Most of the light in marine bioluminescent animals comes from bacteria. the light-producing structures of these fishes, however, are not produced by bacteria, although the presence of bacterial Dna suggests that symbiotic bacteria were originally present.
Members of the superorder Paracanthopterygii include the following:
● Toadfishes have no scales; they have large eyes on large heads. they can remain out of water for 24 hours as they push themselves across mud flats with their pectoral fins. they are omnivores but can also ambush their prey when under water. they are also able to “sing” with their air bladders and have well-developed parental care. Some have venom glands on their fins.
● Many anglerfishes live in deep water. they have enormous mouths and inwardly curved teeth. they can consume prey up to twice their size. they are famous for the way that they can attract their prey. the front spines of their dorsal fins develop into a filament (a “fishing pole”) from which a fleshy tip pro-trudes. the fleshy tip may be bioluminescent. the pole attracts other fishes which they quickly consume. they can “walk” along the bottom of the sea with modified fins.
the acanthopterygii is the largest superorder of teleosts, containing half of the known species. examples include:
● Flying fishes have large pectoral fins that allow them to glide through the air near the ocean surface. they do not actually fly. they propel themselves from the water, then glide for up to 160 feet (50 m), often for a half a minute. their pectoral fins are shapes somewhat like bird wings.
● Pipefishes and sea horses are long, slender fishes that swim vertically, often camou-
Geometric moray eels in Rocktail Bay, Australia (Clacherty and Associates)
xviii+598_Biodiversity_F.indd 248 11/17/11 1:13 PM

fishes 249
flaged in the vegetation of estuaries and coastal swamps (see figure on page 250). they are among the most famous evolu-tionary novelties because the males have the babies. Females deposit eggs in a pouch in the male’s body, where the eggs are fertilized, develop, and hatch. Sea horses (genus Hippocampus) have plates instead of scales as their outer surface and have no tail fin.
● Flatfishes swim on their sides. in newly hatched flatfishes, one eye is on each side of the head but during development one eye migrates, so that both eyes are on the upper side of the fish as it lies on its side in the mud. the flatfish group contains a large number of economically important fishes, such as flounders, sole, turbot, and halibut.
● the perciform fishes are one of the largest groups. among them are the gobies, tunas, and perches. tuna have red meat because of the very high concentration of myoglo-bin, which supplies oxygen to the muscles. therefore they can swim rapidly (up to 45 miles [70 km] per hour). Some tunas are warm-blooded, also due to muscular activity.
Ray-finned fishes have been the evolutionary success story of the aquatic vertebrates. in contrast, the flesh-finned fishes are far less diverse, but it is from this group that the tetrapods evolved. the sarcopterygians (flesh-finned fishes) were abundant in the middle Paleozoic era but are rare today. their pectoral and pelvic fins have bonelike lobes that are
homologous to the limbs of tetrapods: pectoral lobes to arms or front legs, and pelvic lobes to back legs. two lineages of sarcopterygians evolved during the Paleozoic. the coelacanths evolved and remain in deep ocean waters, while the lungfishes lived and live in shallow freshwater. lungfishes can use their fins to push themselves from one body of shallow water to another. their lungs allow them to breathe air while embedded in mud for long periods during the dry season and to survive in murky water that contains little dissolved oxygen.
coelacanths were thought to have been extinct since the cretaceous extinction 65 million years ago. However, in 1938, captain Hendrick Goosen of the trawler Nerine brought a fish that had been caught deep in the indian Ocean to Marjorie cour-tenay-latimer, curator of a museum in east london, South africa. She identified the fish, now named Latimeria chalumnae, as a crossopterygian. Subse-quent searches revealed that native fishermen of the comoro islands (in the indian Ocean off the coast of africa) reported that they had caught these fish for years and had thrown them back as inedible. in 1998, another species, Latimeria menadoensis, was found several thousand miles away, in indonesia, by a scientist on his honeymoon.
the lineage of fishes that became tetrapods included fishes such as Eusthenopteron, which had skeletal characteristics intermediate between those of earlier fishes and later amphibians. later species in this lineage had legs but still retained many fish skeletal characteristics. as is the case with any so-called missing links, there is no clear line of division between ancestor and descendant.
Most bony fishes release eggs and sperm into the water, where external fertilization occurs. in some bony fishes, such as sea horses and pipefish (see above), fathers may hold fertilized eggs within their bodies while the eggs undergo embryonic develop-ment. Bony fishes display a great variety of repro-ductive habits. in some species, individual males and females form pairs, while in other species they form large reproductive groups. in some species, a fish may begin its life as a female then develop into a male, while other species contain some individuals that are hermaphroditic (both male and female at the same time).
adapTaTIoNS To aquaTIC ENvIroNmENTSMost fishes, except for the tuna and the great white shark, are ectothermic (from the Greek, roughly translating to “outside temperature”), which means their temperatures fluctuate with those of their environments. the tuna and the great white shark generate some internal body heat, which makes them at least partly endothermic (Greek for “inside
Salmon migrate from the ocean upstream to the ponds in which they hatched, often jumping upstream over water-falls to do so. Bears easily catch some of them during this migration. (Sandra vom Stein; iStockPhoto.com)
xviii+598_Biodiversity_F.indd 249 11/17/11 1:13 PM

first entry250 fishes 250
temperature”). there are no fishes that maintain a constant body temperature. therefore, fishes are all heterothermic (Greek for “variable temperature”) rather than homeothermic (Greek for “constant temperature”).
Fishes need to maintain buoyancy so that they do not have to swim continually to keep from sink-ing. in sharks, buoyancy is provided by a large liver filled with fat (which is less dense than water). the liver can counterbalance only about one-third of the shark’s weight, however; sharks therefore do need to keep swimming, if only slowly, to keep from sinking. Most sharks also need to keep swimming in order to provide oxygenated water to their gills, which are inside of slits behind the head. Some sharks, however, can rest in caves or on the bottom of the sea while they pump water past their gills. Bony fishes, however, use air bladders or lungs. lungs are a primitive condition for bony fishes. lungs began as expansions of the upper digestive tract that were open to the pharynx, allowing the fish to gulp air. Gars and bowfins can still do this. in most bony
fishes today, the lung has evolved into the air blad-der, not connected to the pharynx, which the fish uses for buoyancy.
Fishes have a single opening through which both nitrogenous wastes and gametes are released. in some fish, this opening is a cloaca (plural cloacae) through which intestinal wastes are also eliminated. Hagfishes cannot regulate their tissue salt concen-tration, which is therefore the same as that of the water surrounding them. However, some lampreys live in freshwater and can regulate their internal salt concentration. the tissues of cartilaginous fishes contain less salt than the ocean water surround-ing them, but they maintain isotonic balance with the seawater by accumulating urea in their blood. Sharks also release ammonia into the water through their gills. the blood of marine bony fishes contains less salt than the hypertonic seawater, and they must continually drink water to replace the osmotic water loss. Freshwater bony fishes must continually excrete the water that diffuses into their bodies from the hypotonic environment.
CoNSErvaTIoN of fIShESMany fishes are rare and in danger of extinction. But there are many threats to the survival of even the more common fish species. these include:
Overfishing. Many species of commercially important fishes have large populations and fill large expanses of the oceans. they appear to be inex-haustible resources. But commercial fishing fleets use advanced technologies to capture prodigious num-bers of them: nets that are many miles long, for example. as a result, many commercial fish species have experienced steep population declines. Because the open oceans are international, the only controls of ocean fishing are international agreements and voluntary measures by fishing corporations, neither of which have been satisfactory. Significant over-fishing has occurred even in the shallow waters of continental shelves that are under the jurisdiction of specific nations. in some places, such as california, new england, and iceland, sustainable harvest stan-dards were adopted. in 2009, after about a decade of these standards, fish populations began to recover. nevertheless, some fish species such as the north-ern bluefin tuna (Thunnus thynnus) are critically endangered and unlikely to recover soon. their very rarity contributes to their decline: a single bluefin can sell for $100,000 on the Japanese wholesale sushi market.
Pollution. the oceans have been significantly polluted, particularly with garbage. Ocean currents have deposited large accumulations of floating gar-bage in numerous locations. the largest of these, the Great Pacific Garbage Patch, is located in the north
A male pink pygmy sea horse (Hippocampus barg-ibanti), with its pouch full of young offspring, is excel-lently camouflaged against a Muricella sea fan coral. (Dan Schmitt; iStockPhoto.com)
xviii+598_Biodiversity_F.indd 250 11/17/11 1:13 PM

fishes 251
Pacific Ocean and is the size of texas. the garbage is mostly plastic, which decomposers cannot break down. Some of the plastic garbage is toxic or has surfaces upon which persistent organic pollutants can be carried. even though they cover 70 percent of the earth’s surface, the oceans are not in fact too large to pollute.
Protection of spawning grounds. Many spe-cies of fish spawn in habitats different from those in which they live their adult lives. Many marine fishes spawn and hatch in estuaries, which is where freshwater and salt water meet. estuaries are highly productive because of the nutrients from the fresh-water and the sunlight penetrating the shallow depths and are, therefore, rich sources of food for young fish. they are also a focal point for indus-trial pollution and for commercial development. anadromous fishes, such as salmon, return to the streams in which they hatched to lay their eggs. it is a dangerous enough journey as it is, but becomes nearly impossible when the streams are blocked by dams. Many dams have salmon ladders, which allow salmon to bypass the dam. this has proven an imperfect solution, and in some cases dams that are not of great economic importance have been destroyed by the same agencies that built them—not only allowing fish migrations, but the recovery of entire aquatic ecosystems.
Releases from hatcheries and selection by fishing. it would seem strange to consider the replenishment of fish populations by the release of young fish from hatcheries to be a conservation problem. However, when large numbers of fish are released and then many of them are caught by fishermen, natural selection favors the fish that reproduce before they reach the legal size limit for capture. these fish evolve the ability to reproduce at smaller sizes. their genes spread into wild populations may cause a reduction in average fish size in those popula-tions as well. there is special concern about the escape of genetically modified fish, such as salmon, resulting in the introduction of transgenes into wild populations.
See also amphibians; animals; cladistics; deep oceans, biodiversity in.
Further readingallen, thomas B. Shark Almanac: A Fully Illustrated His-
tory of Sharks, Skates, and Rays. Guilford, conn.: lyons Press, 1999, 2003.
Barlow, George. The Cichlid Fishes: Nature’s Grand Exper-iment in Evolution. new york: Perseus, 2002.
Berra, tim M. Freshwater Fish Distribution. San Diego: academic Press, 2001.
Briggs, D. “conodonts: a major extinct group added to the vertebrates.” Science 256 (1992): 1,285–1,286.
chapleau, Francois, et al., eds. Encyclopedia of Fishes. San Diego: academic Press, 1998.
Ford, Michael J. “Selection in captivity during supportive breeding may reduce fitness in the wild.” Conservation Biology 16 (2002): 815–825.
Fountain, Henry. “Red-bellied piranha is really yel-low.” New York Times, May 24, 2005. available online. URl: http://www.nytimes.com/2005/05/24/science/24obse.html. accessed august 7, 2011.
Froese, R., and D. Pauly, eds. “FishBase.” available online. URl: http://www.fishbase.org. accessed august 8, 2011.
Hutchings, J. a., and D. a Fraser. “the nature of fisheries- and farming-induced evolution.” Molecular Ecology 17 (2008): 294–313.
Jackson, J. B. c., et al. “Historical overfishing and the recent collapse of coastal ecosystems.” Science 293 (2002): 629–638.
long, John a. The Rise of Fishes: 500 Million Years of Evolution. Johns Hopkins University Press, 1996.
Martin, R. aidan. “ReefQuest center for Shark Research.” available online. URl: http://www.elasmo-research.org. accessed august 8, 2011.
Monterey Bay aquarium. “Seafood watch.” available online. URl: http://www.montereybayaquarium.org/cr/seafoodwatch.aspx. accessed May 13, 2011.
Moritz, t., and K. e. lisenmair. “the air-breathing behav-iour of Brevimyrus niger (Osteoglossomorpha, Mormyridae).” Journal of Fish Biology 71 (2007): 279–283.
national Oceanic and atmospheric administration. “Our fear of and fascination with sharks.” available online. URl: http://www.magazine.noaa.gov/stories/mag36.htm. accessed May 13, 2011.
nelson, Joseph S. Fishes of the World. Hoboken, n.J.: John Wiley & Sons, 2006.
Quammen, David. “Where the salmon rule.” National Geographic, august 2009, 28–55.
Rios, l. M., charles Moore, and P. R. Jones. “Persistent organic pollutants carried by synthetic polymers in the ocean environment.” Marine Pollution Bulletin 54 (2007): 1,230–1,237.
Shubin, neal. Your Inner Fish: A Journey into the 3.5-bil-lion-year History of the Human Body. new york: Basic Books, 2007.
Sweet, W. c., and Donoghue, P. c. J. (2001), “conodonts: past, present and future.” Journal of Paleontology 75 (2001): 1,174–1,184.
United nations environmental Programme. “Overfish-ing: a threat to marine biodiversity.” available online. URl: http://www.un.org/events/tenstories/06/story.asp?storyiD=800. accessed May 13, 2011.
Xian-guang, H., et al. “new evidence on the anatomy and phylogeny of the earliest vertebrates.” Proceedings of the Royal Society B: Biological Sciences 269 (2002): 1,865–1,869.
xviii+598_Biodiversity_F.indd 251 11/17/11 1:13 PM
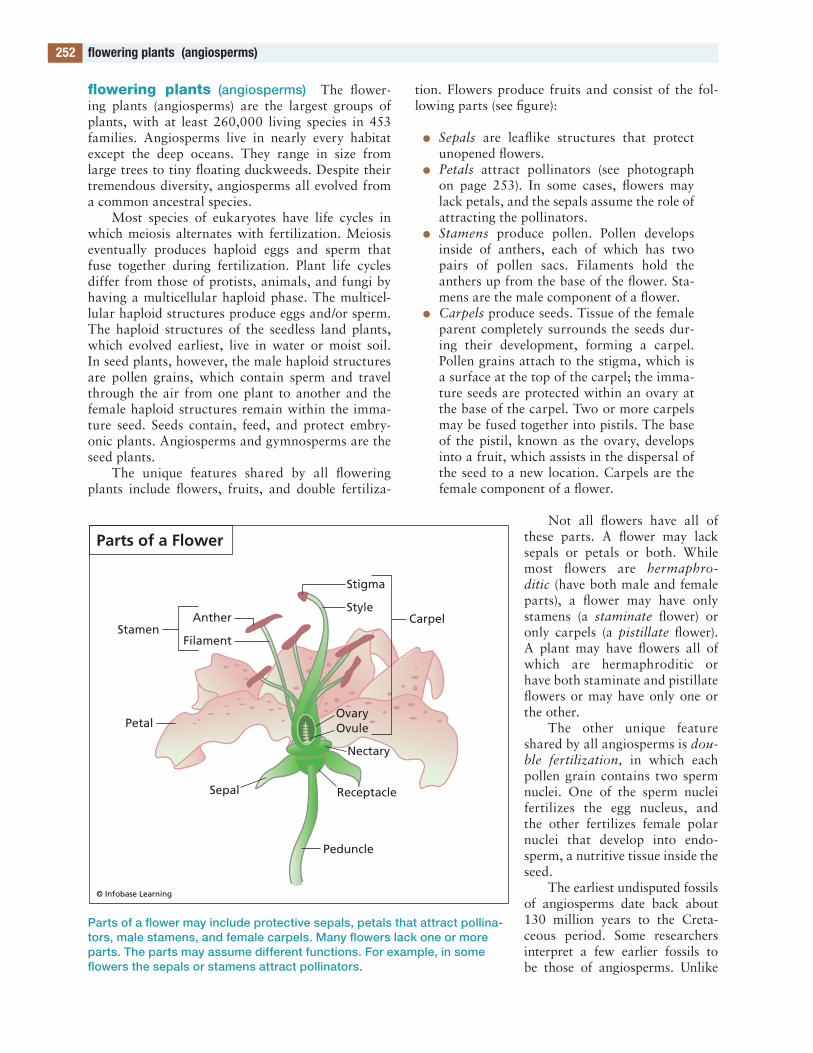
first entry252 flowering plants (angiosperms) 252
flowering plants (angiosperms) the flower-ing plants (angiosperms) are the largest groups of plants, with at least 260,000 living species in 453 families. angiosperms live in nearly every habitat except the deep oceans. they range in size from large trees to tiny floating duckweeds. Despite their tremendous diversity, angiosperms all evolved from a common ancestral species.
Most species of eukaryotes have life cycles in which meiosis alternates with fertilization. Meiosis eventually produces haploid eggs and sperm that fuse together during fertilization. Plant life cycles differ from those of protists, animals, and fungi by having a multicellular haploid phase. the multicel-lular haploid structures produce eggs and/or sperm. the haploid structures of the seedless land plants, which evolved earliest, live in water or moist soil. in seed plants, however, the male haploid structures are pollen grains, which contain sperm and travel through the air from one plant to another and the female haploid structures remain within the imma-ture seed. Seeds contain, feed, and protect embry-onic plants. angiosperms and gymnosperms are the seed plants.
the unique features shared by all flowering plants include flowers, fruits, and double fertiliza-
tion. Flowers produce fruits and consist of the fol-lowing parts (see figure):
● Sepals are leaflike structures that protect unopened flowers.
● Petals attract pollinators (see photograph on page 253). in some cases, flowers may lack petals, and the sepals assume the role of attracting the pollinators.
● Stamens produce pollen. Pollen develops inside of anthers, each of which has two pairs of pollen sacs. Filaments hold the anthers up from the base of the flower. Sta-mens are the male component of a flower.
● Carpels produce seeds. tissue of the female parent completely surrounds the seeds dur-ing their development, forming a carpel. Pollen grains attach to the stigma, which is a surface at the top of the carpel; the imma-ture seeds are protected within an ovary at the base of the carpel. two or more carpels may be fused together into pistils. the base of the pistil, known as the ovary, develops into a fruit, which assists in the dispersal of the seed to a new location. carpels are the female component of a flower.
not all flowers have all of these parts. a flower may lack sepals or petals or both. While most flowers are hermaphro-ditic (have both male and female parts), a flower may have only stamens (a staminate flower) or only carpels (a pistillate flower). a plant may have flowers all of which are hermaphroditic or have both staminate and pistillate flowers or may have only one or the other.
the other unique feature shared by all angiosperms is dou-ble fertilization, in which each pollen grain contains two sperm nuclei. One of the sperm nuclei fertilizes the egg nucleus, and the other fertilizes female polar nuclei that develop into endo-sperm, a nutritive tissue inside the seed.
the earliest undisputed fossils of angiosperms date back about 130 million years to the creta-ceous period. Some researchers interpret a few earlier fossils to be those of angiosperms. Unlike
Parts of a flower may include protective sepals, petals that attract pollina-tors, male stamens, and female carpels. Many flowers lack one or more parts. The parts may assume different functions. For example, in some flowers the sepals or stamens attract pollinators.
xviii+598_Biodiversity_F.indd 252 11/17/11 1:13 PM

flowering plants (angiosperms) 253
the dinosaurs, the angiosperms as a group survived and recuperated from the asteroid impact of the cretaceous extinction, perhaps because their seeds remained dormant and sprouted after the earth recovered from this event.
Botanists understand that the angiosperms evolved from a gymnosperm ancestor. this had to occur, unless seeds evolved more than once, which is unlikely. However, fossil evidence of the ancestor of angiosperms is lacking. the earliest angiosperms in the fossil record are already differentiated into distinct evolutionary lineages. Based just upon fossil evidence, the origin of angiosperms remains today as it was when charles Darwin called it an “abomi-nable mystery.” Fortunately, there is evidence, much of it molecular, that allows botanists to reconstruct the origin of angiosperms.
One major difference between gymnosperms and angiosperms is that the wood of most angio-sperms contains large water-conducting vessels, while most gymnosperms have only small conduct-ing tubes. a few gymnosperms, including the now extinct Bennettitales and the modern group often classified as Gnetales, have wood with large ves-sels, while a few angiosperms have wood without
large vessels. the Bennettitales and Gnetales had or have reproductive structures that are not flowers but resemble them in some ways. the angiosperms that have wood without vessels are those that have also been identified, for other reasons, as the most primi-tive angiosperms.
One major difference between angiosperms and gymnosperms is that most angiosperms have a much faster life cycle than that of gymnosperms. in gym-nosperms, a year may be required for the pollen nucleus to reach the egg and bring about fertiliza-tion. there are therefore no annual gymnosperms (i.e. that live only for a year, completing their repro-duction at the end of the same year in which they ger-minated). in angiosperms, fertilization can happen in a few days or less. this observation has led many researchers to believe that the earliest angiosperms lived in disturbed environments. these disturbed areas may have included places that experienced a forest fire or landslide, seasonal drought, or even damage by dinosaurs. Rapid growth provides an evolutionary advantage in disturbed areas. if the earliest angiosperms lived in such environments, it may not be surprising that no fossils of them have been found, as such environments are very unsuit-able for fossilization.
there is also widespread recognition among botanists that angiosperms have proliferated largely because of coevolution with animals, as in these examples:
● although many lineages of angiosperms retain or have re-evolved wind pollination, most angiosperms have coevolved with the insects and other animals that pollinate their flowers. insects as a whole did not experience an increased burst of diversity during the cretaceous period. But insect lin-eages such as bees and butterflies, in which the adults pollinate flowers, have prolifer-ated, along with the angiosperms, since the cretaceous period.
● although many lineages of angiosperms have re-evolved wind dispersal of seeds, most angiosperms have coevolved with mammals and other animals that disperse their seeds. the fact that many mammals ate fruits, while dinosaurs appeared to rarely do so, paralleled the decline of dinosaurs dur-ing the cretaceous and the proliferation of mammals since the cretaceous.
the earliest angiosperm fossils do not provide a clear indication of what the earliest flowers were like, because by the time of these earliest fossils, angiosperms had already evolved a great diversity
Many flowers, such as this Linanthus montanus from the Sierra Nevada of California, have colorful mark-ings that show pollinators where to find the nectar. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 253 11/17/11 1:13 PM

first entry254 flowering plants (angiosperms) 254
of flowers, from small, simple flowers to large mag-nolia-like flowers. angiosperms radiated into many diverse forms very early in their evolutionary history. Since botanists have no fossils of the ancestral angio-sperm, they have attempted to reconstruct the earli-est flower by cladistics, an evolutionary analysis of existing groups of angiosperms. Most of these analy-ses have clearly identified a small woody plant that today exists only in the cloud forests of new cale-donia, Amborella trichopoda, as the only surviving member of the lineage that most closely resembles the ancestor of all flowering plants. characteristics of Amborella include the following:
● five to eight sepals and petals, which are small, greenish-yellow, and largely separate from one another
● numerous stamens, largely separate from one another
● five to six separate carpels, each of which has an ovary with a single seed. the carpel does not completely close with tissue during growth. it is sealed by a secreted liquid.
if these characteristics represent what the first flowers were like, then the following major changes have occurred during the evolution of the angiosperms:
● in many but not all lineages, the floral parts have fused together. two kinds of floral fusion are connate fusion, in which simi-lar parts have fused, and adnate fusion, in which different parts have fused. in some lineages, petals have fused together to form a tube or to form a landing platform that accommodates pollinators; in many lin-eages, carpels have fused into pistils; in a few, stamens have fused together. all these are examples of connate fusion. in some lineages, stamens have fused to petals, an example of adnate fusion.
● in many but not all lineages, the numbers of floral parts have changed. in many lin-eages, the numerous stamens have been replaced by a small number of stamens, usually the same as the number of petals. in some lineages, the number of parts has increased, as in the lineages in which the flowers have dozens or even hundreds of petals.
● Amborella and other primitive flowering plants are woody. the herbaceous condi-tion, in which extensive woody tissue never develops, has evolved numerous times in separate lineages of flowering plants.
in many lineages of flowers, a mosaic of such changes has occurred. in roses, for example, all of the floral parts are still separate, the stamens are still numerous, but there are only five petals. in gerani-ums, all of the floral parts are still separate, but there are only five stamens and five petals. Mallow flowers have five separate petals but numerous stamens that have fused together.
the traditional classification of angiosperms divides them into monocots and dicots. Monocots have characteristics such as these:
● the embryo has one leaf (mono- for one, -cot for cotyledon).
● the leaves usually have parallel veins. ● the stems usually have scattered bundles of
veins. ● the flowers usually have parts in threes or
multiples of three.
Dicots have characteristics such as these:
● the embryo has two leaves (di- for two, -cot for cotyledon).
● the leaves usually have netlike veins. ● the veins usually form a ring just under-
neath the surface of the young stem. ● the flowers usually have parts that are numer-
ous, or in fours, fives, or multiples thereof.
the arrangement of the stem veins results in important differences between monocots and dicots. each vein contains both xylem (which con-ducts water upward from the roots to the leaves) and phloem (which conducts sugar water primarily downward from the leaves to the stems and roots). the phloem is oriented toward the outside of the stem. Between the xylem and phloem there are cells that produce new xylem and phloem cells. in young dicot stems, since the xylem of the veins forms a discontinuous ring, the production of new xylem cells produces a continuous ring of xylem, or wood. Furthermore, a new layer of wood can be produced each year. Meanwhile, the discontinuous ring of phloem can become a continuous ring of phloem, or inner bark. Many dicots are therefore capable of producing large woody trunks that increase in diam-eter throughout the life of the tree (see photographs on page 255). (the bark does not accumulate, as it is pushed outward by the accumulating wood.) in contrast, the scattered veins in monocot stems do not produce very many new xylem and phloem cells. if they were to do so, the result would be an unstable mixture of wood and bark. Most mono-cots are therefore small, herbaceous plants. Only a few, such as palm trees, grow large. the trunks of
xviii+598_Biodiversity_F.indd 254 11/17/11 1:13 PM

flowering plants (angiosperms) 255
these trees do not increase in diameter as the plant becomes older: the trunk of a palm tree is no thicker at the bottom than it is at the top (see figure). evo-lutionary analysis has allowed the classification of angiosperms into the following lineages:
● Basal angiosperms include several indepen-dent evolutionary lineages, such as Ambo-rella, water lilies, magnolias, and laurels. all of these basal lineages have sepals, petals, stamens, and carpels that are not attached to one another, and their trunks are woody.
● Monocots are a distinct lineage of flowering plants.
● Eudicots, or true dicots, are a lineage that includes all of the rest of the flowering plants and are described in a separate entry.
therefore, although the monocots are a mono-phyletic group, the dicots are a paraphyletic group.
the 260,000 species of flowering plants far sur-pass the species diversity of any other plant group.
See also cladistics; coevolution; creta-ceous period, biodiversity in; dicots; mono-cots; pollination; seedless plants; seed plants.
Further readingcrepet, William l., and Karl J. niklas. “Darwin’s sec-
ond ‘abominable mystery’: Why are there so many angiosperm species?” American Journal of Botany 96 (2009): 366–381.
endress, Peter K., and James a. Doyle. “Reconstructing the ancestral angiosperm flower and its initial specializa-tions.” American Journal of Botany 96 (2009): 22–66.
Friedman, William e. “the meaning of Darwin’s ‘abomina-ble mystery.’ ” American Journal of Botany 96 (2009): 5–21.
Klesius, Michael. “the big bloom: How flowering plants changed the world.” National Geographic, July 2002, 102–121.
Magallon, Susana, and amanda castillo. “angiosperm diversification through time.” American Journal of Botany 96 (2009): 349–365.
Soltis, Douglas e., et al. Phylogeny and Evolution of Angio-sperms. Sunderland, Mass.: Sinauer associates, 2005.
Most trees are dicots, which produce xylem (wood) and phloem (bark) in orderly layers, allowing a strong trunk to develop (left). Palm trees produce xylem and phloem in scattered bundles throughout the trunk, which pro-duces a trunk that is not as strong (right). (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 255 11/17/11 1:13 PM

first entry256 food web 256
Soltis, Pamela S. and Douglas e. Soltis. “the origin and diversification of angiosperms.” American Journal of Botany 91 (2004): 1,614–1,626.
taylor, edith l., and thomas n. taylor. “Seed ferns from the late Paleozoic and Mesozoic: any angiosperm ancestors lurking there?” American Journal of Botany 96 (2009): 237–251.
food web See ecosystems.
fossils and fossilization Fossils are the evi-dence of past biodiversity and therefore the basis upon which scientists can reconstruct the evolu-tionary history of biodiversity. Fossils are the physi-cal evidence of organisms that have been dead for many years, past the normal duration of decompo-sition; fossilization is the set of processes by which they are formed. When volcanic or other rocks erode, water carries sediment to the continental shelves at the edges of the oceans, where it accumu-lates in layers called strata. this process is occur-ring right now. as silt and mud layers are buried, the increased temperature and pressure transform them into sedimentary rocks. the layers in sedi-mentary rocks are visually distinguishable. clay particles become shale, while sand particles become sandstone. Fossils are usually found in sedimentary rocks, since dead plants and animals are often buried in the sediment. intense heat and pressure can transform volcanic or sedimentary rocks into metamorphic rocks, in which the fossils are usu-ally destroyed. Fossils are not always destroyed by metamorphosis. Structures that may be fossilized cyanobacteria can be seen in the 3.5-billion-year-old apex chert of australia.
Fossils have been known for millennia, but until the last three centuries, most people thought fossils were simply peculiar rocks that just happened to look like plants and animals, rather than being the remains of actual plants and animals. leonardo da Vinci was one of the earliest scholars to recognize fossils for what they really were, and he puzzled about why fossils of seashells were found at the tops of mountains. His solution to the problem reflected more of the medieval than the modern mind. He conceived of the earth as an organism, therefore it must have a circulatory system, which means that ocean water must circulate underground to the tops of mountains, where it comes out as creeks and riv-ers—and sometimes brings seashells with it. He was right about the fossils, though wrong about how they got there. another early scholar to recognize fossils as remnants of formerly living organisms was Danish geologist niels Stensen (nicholas Steno).
When an organism dies, it almost always decays. Under some circumstances, decomposition is delayed, such as in the presence of water and the absence of oxygen. as the organism decays slowly, the space inside its body, sometimes even inside of its cells, is filled with mineral deposits from the water (see figure on page 257). the fossil may be completely mineralized; alternatively, permin-eralization occurs when minerals enter the dead organism without completely replacing the original molecules. Under conditions of high temperature and pressure, the sediments surrounding the organ-ism and the organism itself are both transformed into rock. Because of chemical differences between the fossil and its surrounding matrix, fossils usu-ally stand out and may cause a zone of weakness in the rock that contains them. Fossils are there-fore frequently found by geologists who strike the rocks with hammers; the fracture reveals the fos-sils. Mineral deposits inside of a coal seam (coal balls), rather than the coal itself, often contain fos-sils of the plants from which the coal was derived. the chemical differences between the fossil and its matrix can sometimes be of help in extracting the fossil. For example, shells of invertebrates can sometimes be isolated if the matrix is dissolved in concentrated hydrofluoric acid.
Sediments are the major, but not the only, place to find fossils. Many insects, stumbling into coni-fer sap, became entangled and engulfed; the sap became amber, and the insects were preserved in exquisite detail. this happened frequently enough that there is a worldwide market for insects pre-served in amber.
ancient wood other than petrified wood, human bodies from a bog, and mummies preserved in andean caves are not usually considered fossils, nor are bodies preserved by human activity (such as egyptian mummies). Fossils retain the structure of the original organisms, sometimes with consider-able internal detail; therefore coal and oil are not considered fossils, since they have been transformed into uniform organic materials (even though they are called fossil fuels). these remains can be as valuable as fossils in scientific research. leaves that have been preserved for 20 million years in cold, wet, anoxic conditions in deposits near clarkia, idaho, are still green, but oxidize into black films upon exposure to air. the leaves were so chemically intact that scien-tists could extract chloroplast Dna from them.
the hard parts of organisms (such as the cal-cium carbonate shells of invertebrates, and the cal-cium phosphate bones of vertebrates) are most easily preserved and constitute the vast majority of fos-sils. teeth are particularly popular fossils to study, not only because they are hard (and abundantly
xviii+598_Biodiversity_F.indd 256 11/17/11 1:13 PM

fossils and fossilization 257
preserved) but also because they can reveal what kind of diet the animal had—grinding teeth for grazers, sharp teeth for carnivores. the study of fossil bones can also reveal more than just the ana-tomical structure of the animal. Study of tissue lay-ers inside of bones reveals the growth rate of the animal and its age at death. this is how scientists have determined that Tyrannosaurus dinosaurs had a high metabolic rate and grew rapidly, up to 880 pounds (400 kg) by the age of 20 years.
Under conditions of very slow decomposition and very thorough burial, soft parts of organisms, and entire organisms without hard parts, can be preserved. a famous example is the Burgess Shale in canada. impressions of soft-bodied animals, many parts of plants, and even single-celled microbes have frequently been preserved. Since fossilization almost
always requires burial in sediment, most fossils are of the aquatic organisms that already live in or near the sediments. aquatic invertebrates therefore con-stitute the best fossil assemblages. terrestrial organ-isms are fossilized when they fall into or are washed into the sediments, as when primitive birds fell into low-oxygen pools that later became the limestone deposits of Solnhofen limestone in Germany, where ancient Archaeopteryx birds were fossilized. the odds are very much against the preservation of fos-sils from terrestrial organisms: it is estimated that only one bone in a billion is preserved as a fossil. if all 300 million americans, most of whom have 206 bones, were to die, only about 60 bones would be fossilized under normal conditions. the odds are not even good for organisms of shallow oceans, where only 40 percent of the organisms have even the possi-bility of being fossilized. Rarely, massive graveyards of fossils (Lagerstätten) such as the Burgess Shale or the Solnhofen limestone have resulted from condi-tions that were remarkably good for preservation.
an intimate knowledge of biology is necessary in the study of fossils. this allows a great deal of information about the living animal to be recon-structed from the fossils of just its bones. Sites of muscle attachment leave traces on the bones, therefore the musculature of an extinct animal can be reconstructed from bone fossils. Blood vessels inside the living bones may have left traces in the fossilized bones, allowing scientists to determine the extent of blood vessel formation and, hence, whether the animal was warm-blooded. Occasional lucky finds reveal even more about the animal’s life. Gastroliths are smooth stones found associated with dinosaur skeletons. like modern birds, many dinosaurs swallowed stones that helped to grind plant food in their stomachs. the grinding action caused the stones to become smooth. coprolites (fossilized feces) often allow scientists to recon-struct the diet of the animal.
it is usually easy to tell which structures in sedi-mentary rocks are fossils and which are inorganic in origin, but not always. there was some controversy about whether a “shell” that was found in a Russian fossil deposit was actually a shell or a percussion fracture from the drill bit that extracted the sample.
Unfortunately for researchers, fossils are usually found in fragments and have to be put back together. examples include:
● Plants often shed their organs separately or are torn apart during fossilization. thus it is often impossible to tell which leaf grew in connection with which trunk.
● the same thing can happen to animals. the largest animal of the cambrian seas,
Petrified wood is produced when minerals replace biological material during slow decomposition in water. The resulting mineral structure reveals where the tree rings and wood rays used to be (the rays contain red iron in this photograph). The petrified logs at Petrified Forest National Park in Arizona were a forest of coni-fers related to modern Araucaria more than 200 million years ago. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 257 11/17/11 1:13 PM

first entry258 fossils and fossilization 258
Anomalocaris, was formerly described as three separate animal species. the animal’s body had been described as a sea cucumber; its mouth had been described as a jellyfish; and its front legs had been described as head-less shrimp. the true identity of these frag-ments was revealed only when a fossil was found that had these fragments together.
● One of the most abundant kinds of Paleo-zoic fossils was the conodont (cone-tooth). everybody suspected that it was an animal tooth but nobody could find the animal. in 1983, after many decades of research, a conodont was found in association with a soft-bodied vertebrate and then only from a faint minnowlike depression.
● When fossils are found individually, it is often impossible to determine which of them belonged to the same species. Juvenile dino-saurs have sometimes been classified as a sep-arate species from the parents. When whole herds or populations of dinosaurs are found fossilized together, it may be possible to deter-mine which ones were the juvenile forms.
there are a few huge deposits of fossils, such as the hills of diatomaceous earth near lompoc, california, and the white hills of Dover in england, which consist of billions of shells of tiny single-celled organisms. a huge graveyard of dinosaur bones, turned to stone, can be seen at Dinosaur national Monument in colorado. numerous skeletons of rhi-noceroses and horses that died from breathing ash from a volcanic eruption 12 million years ago are preserved, in exquisite detail and still in their origi-nal arrangement, at ashfall Beds State Historic Park in nebraska.
trace fossils are the preserved evidence of the activities of the organisms. this includes worm bur-rows and animal footprints. Dinosaur footprints have allowed the analysis of how large the dino-saurs were and how fast they ran, which has fueled the suggestion that some of them, at least, were warm-blooded. Perhaps the most convincing evi-dence that the human ancestor Australopithecus afarensis walked on two legs was the discovery, by Mary leakey, of a trail of remarkably human footprints produced by this species. Mistakes can be made in the interpretation of trace fossils. What was believed to be worm burrows, before the beginning of the cambrian period, were considered the earliest signs of the rapid evolution of animal life. it is likely, however, that these burrows were created by single-celled protists rolling in the seafloor mud.
Fossils can be used to reconstruct the climate in which the organisms lived.
● Plants with large leaves today grow in moist areas, and plants with smaller leaves in drier areas. Serrated leaf margins allow modern plants to cool more efficiently than plants with smooth margins, without the evapo-ration of as much water. the study of leaf shapes can help reconstruct ancient tem-perature and moisture conditions.
● the pollen of seed plants has an outer cover-ing that is very resistant to decay, and usu-ally each genus of plant has its own visually distinguishable kind of pollen. even when all other parts of a plant have decomposed, the pollen (though dead) may still be rec-ognizable. Pollen from mud deposited dur-ing the most recent ice age, for example, indicates that spruce trees (now found only far north and on mountaintops) grew as far south as Oklahoma and that much of what is now amazonian rain forest was grass-land. the study of ancient pollen is called palynology.
the isotope ratios of fossils may also help determine the climate in which they grew. One example is the isotopes of carbon. carbon dioxide molecules (cO2) in the atmosphere contain mostly carbon-12 (12c) but also a little bit of the heavier isotope carbon-13 (13c). the kind of photosynthe-sis that occurs in most plants discriminates against 13cO2, with the result that organic matter contains relatively less 13c than does the atmosphere. Many grasses that live in warm, dry grasslands have a different kind of photosynthesis, which does not discriminate against 13cO2 and produces organic matter that contains more 13c than is usually the case with organic matter. animals that eat fruits may contain relatively less 13c in their bodies than animals that eat grains. tooth enamel that contains more 13c may therefore reflect a diet of grains in a grassland or savanna setting, while tooth enamel with less 13c may indicate a diet of fruits and nuts in a forest setting. this has proven very important for reconstructing the behavior of human ancestors. the human lineage was assumed to have evolved in the savanna, until the analysis of tooth enamel carbon indicated in 2009 that Ardipithecus ramidus, a human ancestor that lived about a million years after the earliest hominids, lived in forests.
See also birds; dinosaurs; fishes.
Further readingDeSalle, Rob, et al. “Dna sequences from a fossil termite
in Oligo-Miocene amber and their phylogenetic impli-cations.” Science 257 (1992): 1,933–1,936.
xviii+598_Biodiversity_F.indd 258 11/17/11 1:13 PM

founder effect and genetic drift 259
Fortey, Richard. Earth: An Intimate History. new york: Knopf, 2004.
Gould, Stephen Jay. “the lying stones of Marrakech.” chap. 1 in The Lying Stones of Marrakech: Penulti-mate Reflections in Natural History. new york: Har-mony, 2000.
———. “Magnolias from Moscow.” chap. 31 in Dinosaur in a Haystack: Reflections in Natural History. new york: Harmony, 1995.
———. “the upwardly mobile fossils of leonardo’s living earth.” chap 1. in Leonardo’s Mountain of Clams and the Diet of Worms. new york: three Rivers, 1998.
Masterson, Jane. “Stomatal size in fossil plants: evidence of polyploidy in majority of angiosperms.” Science 264 (1994): 421–424.
niklas, Karl J., Robert M. Brown, Jr., and R. Santos. “Ultra-structural states of preservation in clarkia angiosperm leaf tissues: implications on modes of fossilization in late cenozoic history of the Pacific.” in Late Cenozoic History of the Pacific, edited by charles J. Smiley, 143–159. San Francisco: american association for the advancement of Science, 1985.
Olson, Storrs l., and Helen F. James. “Fossil birds from the Hawaiian islands: evidence for wholesale extinction by man before western contact.” Science 217 (1982): 633–635.
Palmer, Douglas. Fossil Revolutions: The Finds That Changed Our View of the Past. new york: collins, 2004.
WoldeGabriel, Giday, et al. “the geological, isotopic, botan-ical, invertebrate, and lower vertebrate surroundings of Ardipithecus ramidus.” Science 326 (2009): 65.
founder effect and genetic drift the founder effect and genetic drift occur when a popu-lation is reduced in size (undergoing a population “bottleneck”), either soon after it is established as an offshoot of a larger population or as a result of bad luck that it experiences afterward. these circum-stances allow random events, rather than processes such as natural selection, to control the genetic makeup of the population.
consider a large population that diminishes in size and becomes a small population. in this small population, just by chance, some of the genes may be lost. consider a population consisting of some plants with red flowers and some plants with white flowers. if the population experiences a crash, the red-flowered individuals may be the only ones to sur-vive, even though their survival had nothing to do with the color of their flowers. By chance, the genes for white flowers have been lost. Such genetic drift can almost permanently reduce the genetic diversity in a population. then, when the population begins to grow, genetic diversity remains low.
consider a new population that is founded (for example, on an island) by a small number of emi-grants that disperse from a larger, central popula-tion. the small number of founders will not carry all of the genetic diversity that was present in the central population. if a central population contains individual plants with red and with white flowers and a few seeds disperse to an island, the seeds may happen to be from red-flowered individuals. From the moment that the island population is founded, it already has lower genetic diversity than the population that produced it. it is unlikely that red flowers confer any advantage on the island population; the fact that the island population has no white-flowered plants is due to chance. this is the founder effect. the founder effect can result in a population in which, even if it grows, genetic diversity remains low.
Both the founder effect and genetic drift reduce genetic diversity in a population: the founder effect, when a new population is formed, and genetic drift, when the old population suffers a bottleneck event. Both the founder effect and genetic drift were inves-tigated by geneticist Sewall Wright.
Genetic drift has been experimentally con-firmed. in one experiment with fruit flies, geneticist Peter Buri investigated a gene with two alleles. at the beginning of each experiment, all of the flies were heterozygous, therefore both of the alleles were equally common. every generation, Buri chose a small sample of males and females, from which he raised the next generation. every generation of flies experienced a genetic bottleneck. By the 19th gen-eration, in over half of the populations, one or the other of the two alleles became more and more com-mon until it was the only allele that was present.
Genetic drift can leave its mark on a population for many generations afterward. One in 20 humans who live on Pingelap atoll in the Pacific Ocean has a mutation that causes color blindness and extreme light sensitivity. the usual population average for this mutation is one in 20,000 people. the reason the mutation is so common on Pingelap atoll is that it was carried by one of the few survivors of a typhoon and famine that struck in 1776.
Genetic drift can affect entire species, which is why it is of immense interest to conservation biolo-gists. the Hawaiian goose, or nene (Branta sand-iviensis), experienced a genetic bottleneck which greatly reduced the genetic variability in its popula-tions and now faces extinction. even when a spe-cies consists of a large number of individuals, the fragmentation of the species into many small popu-lations can cause genetic drift to occur in each of the small populations, resulting in the catastrophic loss of genetic diversity. this is occurring in many
xviii+598_Biodiversity_F.indd 259 11/17/11 1:13 PM

first entry260 fungi 260
natural habitats, which are being set aside as only small, disconnected nature preserves.
Humans provide one of the best examples of a genetic bottleneck. the human species has a remark-ably low genetic diversity. Geneticists estimate that the entire human species experienced a genetic bot-tleneck event about 70,000 years ago, at which time the entire human species consisted of only a few thousand individuals.
See also natural selection; population genetics.
Further readingaguilée, Robin, et al. “allele fixation in a dynamic meta-
population: Founder effects vs. refuge effects.” Theo-retical Population Biology 76 (2009): 105–117.
Buri, Peter. “Gene frequencies in small populations of mutant Drosophila.” Evolution 10 (1956): 367–402.
chung, Mi yoon. “low levels of genetic variation within populations of the four rare orchids Gymnadenia cucullata, Gymnadenia camtschatica, Amitostigma gracile, and Pogonia minor in South Korea: indica-tion of genetic drift and implications for conserva-tion.” Plant Systematics and Evolution 281 (2009): 65–76.
Ramírez-Valiente, J. a., et al. “elucidating the role of genetic drift and natural selection in cork oak dif-ferentiation regarding drought tolerance.” Molecular Ecology 18 (2009): 3,803–3,815.
templeton, alan, et al. “the genetic consequences of habi-tat fragmentation.” Annals of the Missouri Botanical Garden 77 (1990): 13–27.
Wright, Shane D., et al. “Slower tempo of microevolution in island birds: implications for conservation biology.” Evolution 63 (2009): 2,275–2,287.
young, andrew, et al. “the population genetic conse-quences of habitat fragmentation for plants.” Trends in Ecology and Evolution 11 (1996): 413–418.
Zlojutro, Mark, et al. “coalescent simulations of yakut mtDna variation suggest small founding popula-tion.” American Journal of Physical Anthropology 139 (2009): 474–482.
fungi Fungi are eukaryotic multicellular organ-isms with the following distinctive characteristics:
● their cells have cell walls made of chitin. Fungus cell walls represent an example of evolutionary convergence: Bacterial cell walls consist of peptidoglycan, and plant cell walls consist of cellulose (often with other materials) and evolved separately from the cell walls of fungi. chitin is the same molecule that makes up the exoskeleton of arthropods.
● they have external digestion. they release digestive enzymes into their immediate surroundings and absorb the products of digestion.
● they consist of strands (hyphae, making up a mycelium) of haploid cells. During sexual reproduction, some hyphal cells fuse together to form dikaryotic cells, which have two haploid nuclei that do not fuse together in most cells. Fungi do not have diploid tissue. in some dikaryotic cells, the nuclei fuse together; the resulting diploid cell immediately undergoes meiosis and pro-duces haploid spores, which are the tiny cells that blow through the air and disperse to a new location (see figure on page 261).
Because of these unique characteristics, fungi are classified into their own kingdom within the eukary-otic domain.
Most of the time, fungi consist of hyphae that grow through living or dead organic material and consume it. Haploid hyphae often produce asex-ual sporangia, which release spores. the spores are genetically identical to the hyphae. Occasionally, two genetically different hyphae of the same species of fungus may encounter one another. When this occurs, they may fuse together, forming dikaryotic cells that are characteristic of their evolutionary lin-eage. Fungi do not have male and female gender. they carry out sexual reproduction with genetically different fungi of the same species.
Many fungi release their spores directly into the wind. Some fungi, such as puffballs, allow col-lisions to release the spores (for example, raindrops propel the spores of earth stars into the air). Some fungi attract animals to disperse their spores. Stink-horns produce a putrid scent that attracts flies, while truffles are famous for producing a version of the sex
estimated number of fungus speCies
division Common NameNumber of Species
Chytridiomycota chytrids 1,000
Microsporidia microsporidians 1,500
Zygomycota bread molds 1,000
Ascomycota sac fungi 30,000
Basidiomycota club fungi 30,000
Total 33,500
xviii+598_Biodiversity_F.indd 260 11/17/11 1:13 PM

fungi 261
hormone of a female pig. Male pigs dig them up, eat them, and transport the spores to a new location.
Several groups of organisms, such as slime molds and water molds, have traditionally been classifi ed with the fungi but do not share a common ancestor with fungi. these funguslike lineages are classifi ed with the protists.
EvoluTIoNary dIvErSITy of fuNGIthere are several lineages within the kingdom Fungi, of which two (ascomycota and Basidiomycota) are considered the “higher fungi” (see table on page 260). the table presents a rough estimate of the number of identifi ed species, but many mycologists (biologists who study fungi) estimate that there may be a million and a half species. the lineages of fungi are distinguished by their sexual reproductive struc-tures. the category “Deuteromycota” refers to fungi that have never been observed to reproduce sexually. in many cases, their identity has been determined by Dna analysis, and they have now been assigned to fungus lineages.
Division Zygomycota are mostly terrestrial decomposers; a well-known example is bread mold, Rhizopus nigricans. its haploid hyphae spread over moist bread and absorb nutrients from it. During its asexual phase, it produces a stalk with a sphere on top, inside of which haploid spores form. When the sphere breaks open, the spores fl oat away and may grow on another piece of bread. When two genetically different hyphae meet, they may form a spiky sphere known as a zygosporangium, in which sexual recombination produces haploid spores with a mixture of genes from the two parental hyphae.
these spores are resistant to environmental condi-tions and are the form in which zygomycotes often persist in a dormant state. Pilobolus fungi grow on dung. the zygosporangium stalk is sensitive to light, aims the sporangium at the Sun, and explo-sively shoots the spores into the air. the spores may land up to seven feet (two m) away, far away from the dung, on vegetation that a grazing herbivore is likely to eat. the spores pass through the digestive system of the animal and end up on another lump of dung. another zygomycote is Empusa muscae,which grows inside the bodies of fl ies and kills them. the fungus reproductive structures emerge and release spores onto vegetation, where more fl ies will encounter them.
Divisions Chytridiomycota and Blastocladio-mycota are fungi that often live in water and are unique in producing spores that swim with fl agella.
Division Microsporidia consists of single-celled parasites of animals. Because of their simple struc-ture and lack of mitochondria, they were once classi-fi ed as primitive protists. Genetic analyses, however, identify them as degenerate fungi.
Division Neocallimastigomycota consists of anaerobic fungi that live in the stomachs of rumi-nant animals and aid in the digestion of coarse plant material.
Division Ascomycota, when they reproduce sexually, produce an ascus (plural asci), or sac, with eight spores: four produced by meiosis, each of which divides once by mitosis. Many ascomy-cota, however, do not reproduce sexually. there is a tremendous diversity of ascomycote forms. they include the following:
When two fungal hyphae meet, they may form a joint reproductive structure, such as a mushroom. The mushroom consists of cells, each of which has two separate nuclei, one from each hypha. In some of the cells beneath the cap the nuclei fuse and then sexual cell division produces spores, which blow away in the wind.
xviii+598_Biodiversity_F.indd 261 11/17/11 1:13 PM

first entry262 fungi 262
● fungi that produce large reproductive struc-tures on which the asci form, such as morels and truffles
● fungi that decompose food in a way that humans like. the best known is baker’s or brewer’s yeast (Saccharomyces cerevesiae), which exists primarily as single cells and reproduces asexually by budding. yeasts produce the carbon dioxide bubbles in leavened bread and in beer, as well as the alcohol in fermented beverages. in addition, other ascomycotes help in processing foods: Aspergillus oryzae is essential in process-ing soybeans into soy sauce, and species of Penicillium produce the distinctive flavors of several kinds of cheese.
● fungi such as Pneumocystis, Candida, and Tinea that live inside the tissues of animals. Pneumocystis is usually harmless, but in people with severely compromised immune systems (especially due to HiV/aiDS), these fungi can produce fatal pneumonia. Can-dida albicans is responsible for vaginal yeast infections, as well as mouth infections called thrush. Tinea infects the outer, dead layers of the epidermis, forming dark rings as it advances, hence its name ringworm. it is most commonly found on feet (athlete’s foot) or in the genital areas (jock itch). it is annoy-ing, difficult to eliminate, but not dangerous.
● fungi that are highly toxic to humans. Aspergillus flavus grows on peanuts and produces deadly aflatoxin.
● the ergot fungus Claviceps and other plant pathogens such as the Dutch elm disease (genus Ophiostoma), which has killed mil-lions of large elm trees throughout the United States. the ascomycote Cryphonectria para-sitica causes the chestnut blight that has elim-inated almost all native adult chestnut trees in the eastern United States. Penicillium italicum is the green fungus that grows on oranges.
● powdery mildews on leaf surfaces and molds that grow in houses
● Penicillium chrysogenum, the hyphae of which produce penicillin, the first antibiotic to be discovered. Tolypocladium niveum is an ascomycote that produces cyclosporin, a very important immune suppressor used during organ transplants.
● most lichens, which consist primarily of ascomycote hyphae
Division Basidiomycota, when they repro-duce sexually, produce spores on the end of basidia (clubs).
● many fungi with large reproductive struc-tures are basidiomycotes. examples include mushrooms (see figure), which have gills on the surface of which billions of spores are produced; boletes, which look like mushrooms but have pores instead of gills, through which spores are released into the air; puffballs; and earth stars.
● many plant pathogens, such as rusts and smuts
● Cryptococcus is a human pathogen.
EColoGICal rolES of fuNGIFungi are very important in ecosystems as decom-posers of plant material. their hyphae constitute a single individual fungus. Since in most cases each hyphal cell is on its own to obtain nutrition, the hyphae can extend only as far as there is a rich supply of dead leaves. Some fungi, such as honey mushrooms of the genus Armillaria, form rhizo-morphs, which are reinforced cables of parallel hyphae that can transport nutrients. Rhizomorphs can grow through nutrient-poor soil and allow the hyphae to locate new pockets of food and allow the fungus to extend over large areas. clones of geneti-cally identical hyphal cells can be identified by Dna techniques. One Armillaria clone in Michigan was mapped using these techniques and found to cover 37 acres (15 hectares). it quickly gathered media atten-tion as the world’s largest organism, even though it would not be noticeable to a person standing nearby. the researchers estimated that the clone was 1,500 years old and weighed 100 tons (9,700 kg), which is approximately the mass of an adult blue whale. an even larger fungus, Armillaria ostoyae, was discov-
Some fungi, such as this Amanita muscaria, are col-orful and deadly. (Brandon Brooks)
xviii+598_Biodiversity_F.indd 262 11/17/11 1:13 PM

fungi 263
ered in Oregon. it covered 2,384 acres (165 hectares) and was estimated to be 2,400 years old. the soils of the world are filled with hyphal strands of decom-poser fungi.
Many fungi cause diseases. this list of animal diseases caused by fungi is rather short, but fungus diseases of plants are widespread. every wild plant has a fungus that can kill it, and some fungi are specific to certain plants. in many cases, the fungi have only a single host: Gibberella zeae causes only head blight of cereals, although it can infect more than one kind of cereal. in other cases, fungi such as Scleroctinia and Rhizoctonia can infect a wide range of different agricultural host plants such as peanuts and soybeans. as its name might suggest, Phymato-trichum omnivorum infects many trees other than citrus, its principal host. But in several very interest-ing cases, pathogenic fungi have specific alternate hosts: the asexual phase of the fungus life cycle lives in the tissues of one host, while the sexual phase lives in the tissues of another. examples include some astonishing alternations, such as between wheat and barberry bushes, wheat and meadow rue (a wild-flower), and between apples and junipers (see table). Usually the fungus causes very different symptoms in the two hosts and appears to be two different spe-cies of fungi. to control the spread of agriculturally important fungi, it is only necessary to keep the crop populations apart from the wild plants that serve as alternate hosts. Since fungus spores can spread so widely, however, this has proven more difficult than it sounds. During the cold war, the United States military researched the use of Puccinia cereal rusts as biological weapons to use primarily against the Soviet Union, which depended heavily on wheat, and Magnaporthe grisea as a weapon against the Japa-nese rice crop during World War ii.
as important as fungi are as decomposers and pathogens, their role as mutualists is just as impor-tant. Hyphae of fungi known as mycorrhizae grow in the cortex of the roots of many plant species. the hyphae extend out into the soil farther than the roots or root hairs. the hyphae then absorb and
transport minerals, especially phosphate, into the roots, for their own use—but the plant makes use of these minerals also. this is particularly important as phosphate is a mineral that diffuses very slowly through soil and is thus frequently a limiting factor to the growth of plants. evidence of mycorrhizal growth has been found in the fossilized stems of Aglaophyton major that lived 400 million years ago—one of the first land plants. it is therefore pos-sible that, without the assistance of mycorrhizae, plants would not have been able to grow in the poor soils of the previously barren dry land. Ectomy-corrhizae grow between the root cells but do not penetrate them, while endomycorrhizae penetrate the walls of root cells. the fungi feed on food-rich exudates from root cells.
Ectomycorrhizae. about one-tenth of plant families (including the pine, oak, birch, eucalyp-tus, and rose families) have ectomycorrhizae. these fungi may belong to any of the three major divisions (Zygomycota, ascomycota, and Basidiomycota). they form a network of cells inside and outside of the host root. Some ectomycorrhizae chemically attract springtails (small arthropods related to insects) and use the nitrogen in their bodies as a source of fertil-izer for themselves and for their host plants.
Endomycorrhizae. the most common endo-mycorrhizae are the arbuscular mycorrhizae, known only from the division Glomeromycota. they form balloonlike structures between the cell membranes and cell walls of root cortex cells, producing an intri-cate surface area of contact. Most families of plants (and many crop species) are hosts to these fungi. arbuscular mycorrhizae are not known to have sexual reproduction, although there are genetically diverse nuclei in their cells. Plants in the heath and orchid families have other kinds of endomycorrhizae.
in some cases, mycorrhizal fungi can form asso-ciations with the roots of more than one plant at a time. in doing so, they can transfer nutrients from a plant that has enough to another plant, even of a different species, that does not have enough. this is an example of mutualism that connects several
examples of pathogeniC fungi With alternate hosts
fungus Common Name host alternate host
Cronartium ribicola Pine blister rust White pine Currants (Ribes)
Gymnosporangium Apple cedar rust Apple Red cedar (Juniperus)
Puccinia coronata Cereal crown rust Cereals Buckthorn (Rhamnus)
Puccinia graminis Cereal stem rust Cereals Barberries (Berberis)
Puccinia triticina Wheat leaf rust Wheat Meadow rue (Thalictrum)
xviii+598_Biodiversity_F.indd 263 11/17/11 1:13 PM

first entry264 fungi 264
species. Plant ecologist Peter Högberg and associ-ates found that mycorrhizal fungi transferred car-bon compounds from overstory Scots pines (Pinus sylvestris) and spruces (Picea abies) to understory birches (Betula pendula) in a Swedish forest. not only does this keep the conifers and the birches alive but promotes the transformation of a pine to a birch forest. Plant ecologist David Hartnett and associates found that mycorrhizae transferred phos-phorus from grasses such as Sorghastrum nutans to forbs (nongrass herbaceous plants) such as Arte-misia ludoviciana in the tallgrass prairie. according to mycologist Paul Stamets, it is possible to interpret this activity as the fungi cultivating the forest or the grassland, maintaining the plant cover and the diversity of species as if it were an agricultural field producing food for the fungus. it is not in the evolu-tionary interest of any plant to promote the growth of a plant of a different species, but it may be in the interest of the fungus to promote the growth of sev-eral plant species.
another ecologically important group of fungi is the genus Xylaria. leafcutter ants (genus Atta) gather pieces of tropical rain forest leaves, bring them into the nests, and chew them up as mulch for growing Xylaria fungi. the fungi are the only food source for the ants. leafcutter ants can be major “herbivores” in tropical forests even though they do not actually eat the plants.
Perhaps the most extraordinary role that fungi play in the world of biodiversity is that their hyphae (usually of ascomycotes) form the structures of lichens. Single-celled eukaryotic green algae and prokaryotic cyanobacteria live amid the hyphae. the algae and cyanobacteria produce food by photosyn-thesis but cannot tolerate conditions that are both sunny and dry. the fungi protect the algae from excess sunlight during dry conditions. as a result, lichens live in places, such as tree trunks and sur-faces of rocks, where neither fungi nor algae could live by themselves. the algal portion of the lichen (the phycobiont), the fungus portion of the lichen (the mycobiont), and the lichen itself each receives a scientific name.
Some fungi have played important roles in human history. examples include the following:
● “King tut’s curse” is believed to have been Aspergillus fungi. this fungus grew on organic material such as grains that were buried with the pharaohs, and its spores (still alive after millennia) were breathed by the first explorers to open the tombs. the fungus spores then grew in their lungs and killed them by pneumonia.
● the ergot fungus Claviceps purpurea grows in rye plants and produces the hallucino-gen lysergic acid diethylamide (lSD). When humans consumed contaminated rye (in the form of rye bread), they hallucinated. this is apparently the cause of many of the witchcraft and werewolf trials in europe and america several centuries ago. the hal-lucinogen caused some people to think they were werewolves and others to think they had seen werewolves. Research has shown that witchcraft and werewolf trials were most common in regions and years with mild winters and cool summers (conditions excellent for the growth of fungus) and among the poor, who ate most of the rye bread, rather than the rich, who ate wheat bread.
the conservation of fungi is both very important and very difficult. the main difficulty is that scien-tists do not even know what most of them are and therefore have no idea how to save them. Many of them live only in undisturbed forests; the destruction of an old-growth forest could, therefore, lead to the extinction of important species of fungi. Researchers such as Paul Stamets have found new species of fungi that can be of immense importance to humans, such as in the processing of organic wastes. Perhaps even more important than their benefit to humans is the way fungi tie together the dominant plants of an ecosystem into interacting units that scientists are only beginning to understand.
See also decomposers; lichens; parasitism.
Further readingBurdon, J. J. “Fungal pathogens as selective forces in plant
populations and communities.” Austral Ecology 16 (2006): 423–432.
Dighton, John, James Francis White, and Peter Oudemans. The Fungal Community: Its Organization and Role in the Ecosystem. Boca Raton, Fla.: cRc Press, 2005.
Fischer, W. M., and J. D. Palmer. “evidence from small-subunit ribosomal Rna sequences for a fungal origin of Microsporidia.” Molecular Phylogenetics and Evo-lution 36 (2005): 606–622.
Gill, e. e, and n. M. Fast. “assessing the microsporidia-fungi relationship: combined phylogenetic analysis of eight genes.” Gene 375 (2006): 103–109.
Högberg, Peter, et al. “natural 13c abundance reveals trophic status of fungi and host-origin of carbon in mycorrhizal fungi in mixed forests.” Proceedings of the National Academy of Sciences USA 96 (1999): 8,534–8,539.
Klironomos, J. n., and M. M. Hart. “animal nitrogen swap for plant carbon.” Nature 410 (2001): 651–652.
xviii+598_Biodiversity_F.indd 264 11/17/11 1:13 PM

fungi 265
lee, Soo chan, et al. “Microsporidia evolved from ancestral sexual fungi.” Current Biology 18 (2008): 1,675–1,679.
li, H., et al. “arbuscular mycorrhizal fungi contribute to phosphorus uptake by wheat grown in a phosphorus-fixing soil even in the absence of positive growth responses.” New Phytologist 172 (2006): 536–543.
Matossian, Mary Kilbourne. Poisons of the Past: Molds, Epidemics, and History. new Haven, conn.: yale Uni-versity Press, 1991.
Money, nicholas P. The Triumph of the Fungi: A Rotten History. new york: Oxford University Press, 2006.
Simard, Suzanne W., et al. “net transfer of carbon between ectomycorrhizal tree species in the field.” Nature 388 (1997): 579–582.
Simon, l., et al. “Origin and diversification of endomy-corrhizal fungi and coincidence with vascular land plants.” Nature 363 (1993): 67–69.
Smith, Myron, Johann Bruhn, and James anderson. “the fungus Armillaria bulbosa is among the largest and oldest living organisms.” Nature 356 (1992): 428–431.
Stamets, Paul. Mycelium Running: How Fungi Can Help Save the World. Berkeley, calif.: ten Speed Press, 2005.
Stephenson, Steven l. The Kingdom Fungi: The Biology of Mushrooms, Molds, and Lichens. Portland, Ore.: timber Press, 2010.
Wilson, Gail W. t., David c. Hartnett, and c. W. Rice. “Mycorrhizal-mediated phosphorus transfer between tallgrass prairie plants Sorghastrum nutans and Arte-misia ludoviciana.” Functional Ecology 20 (2006): 427–435.
yafetto, l., et al. “the fastest flights in nature: High-speed spore discharge mechanisms among fungi.” PLoS One 3 (2008): e3237. available online. URl: http://www.plosone.org/article/info:doi/10.1371/journal.pone.0003237. accessed May 13, 2011.
xviii+598_Biodiversity_F.indd 265 11/17/11 1:13 PM

266
gaia the Gaia hypothesis claims that organisms have transformed the entire earth and maintain it in a state very different than it would have been without organisms. “Gaia” is the name of an ancient Greek earth goddess and is intended by the scientists who defend the hypothesis as an image, not referring to the earth as a literal organism or person. Biologist lewis thomas proposed the concept that the earth was like a cell in his 1984 book Lives of a Cell.However, thomas’s imagery resulted in no program of scientifi c investigation. the scientifi c predictions based upon the Gaia hypothesis began with the work of atmospheric scientist James lovelock. its principal proponent within the world of biological and evolutionary science is evolutionary biologist lynn Margulis. Many biological scientists defend some version of the Gaia hypothesis.
the Gaia hypothesis is sometimes worded as “the earth is an organism,” but this is not correct. the scientists who defend the Gaia hypothesis claim not that the earth is an organism but that the atmo-sphere and water of the earth are a nonliving part of the earth’s living system of interacting organisms the same way that a shell is a nonliving part of a mollusk. the Gaia hypothesis states that organisms transform the earth in a way that has resulted in a regulation of the temperature and chemical composition of the earth. a cell may contain both living components, such as organelles, and nonliving components, such as starch granules, but when viewed from the outside, the whole cell is alive. Similarly, the earth consists of both living components such as organisms and non-living components such as air molecules and minerals, but when viewed from the outside, the earth is alive.
Biological activities of organisms have caused the earth to be very different from other planets
and maintain it in a chemically unstable condition. thus, even without visiting a distant planet, observ-ers should be able to determine whether life exists on the planet. if all of its atmospheric components (which can be studied by analyzing the light that they refl ect) are in equilibrium, the planet is either lifeless or nearly so. life creates chemical and physi-cal disequilibrium which can be visible from millions of miles away. this was the basis on which James lovelock predicted that no life would be found on Mars, a prediction that is either entirely or very nearly accurate.
as a result of these biological processes, the earth behaves as a system, rather than as simply a stage upon which life-forms act. But the hypoth-esis goes further than just to say that the earth is an ecosystem. Perhaps the clearest statement of the Gaia hypothesis is the amsterdam Declaration of 2001: “the earth system behaves as a single, self-regulating system comprised of physical, chemical, biological, and human components.” ecosystem sci-ence does not include the regulation component. For example:
● everyone knows that nearly all of the energy that fl ows through earth ecosystems arrives as photons of light from the Sun. Most of the absorbed sunlight warms the earth, and organisms use a little bit of this energy. eventually this energy radiates into outer space as infrared photons. But the Gaia hypothesis goes further. it claims that the web of interacting organisms regulates the temperature of the earth within certain lim-its. this, the hypothesis claims, is why the earth does not become too hot or too cold.
G
xviii+598_Biodiversity_F.indd 266 11/17/11 1:13 PM

Gaia 267
● everyone knows that, in the earth ecosys-tem, photosynthesis produces oxygen gas and absorbs carbon dioxide gas, while respiration does the opposite. as a result, carbon and oxygen molecules cycle end-lessly through the earth ecosystem. But the Gaia hypothesis goes further. it claims that the earth regulates the amount of carbon dioxide and oxygen in the air, within cer-tain limits. this, the hypothesis claims, is why carbon dioxide does not become so abundant in the atmosphere as to become poisonous and oxygen does not become so abundant as to cause the spontaneous com-bustion of all organic matter on the earth.
What could cause such regulation to occur? One prevailing idea, called “wasteworld” by ecosystem scientist tyler Volk, is that what is waste to one type of organism is a resource to another. consider, for example, the carbon and oxygen cycles described previously. natural selection favored photosynthesis because it gave cyanobacteria a fitness benefit: it allowed them to manufacture food. to cyanobacte-ria, carbon dioxide gas was a resource and oxygen gas was a waste product. But it was not long before other bacteria evolved an ability to use oxygen—not as food, but as a way of extracting energy from food, and they produced carbon dioxide as a waste. When the atmosphere contains a lot of carbon diox-ide, plants carry out more photosynthesis, which removes carbon dioxide from the air. But when these abundant plants begin to die, decomposers remove oxygen from the air in the process of rotting the plants—and of returning carbon dioxide into the air. Regulation of atmospheric oxygen and carbon is an emergent property of different kinds of metabolism. natural selection favors organisms that can obtain whatever resources are in short supply.
at the beginning of life on earth, resources were not wastes of other organisms but the primordial molecules of the planet. the original atmosphere of the earth had so much carbon dioxide that the aver-age temperature of the planet would have been about 122°F (about 50°c) at present levels of sunlight intensity. (Sunlight, however, was dimmer at that time.) Photosynthesis eventually removed enough carbon dioxide from the atmosphere that the average earth temperature today is about 55°F (13°c). the emergence of the continents from underneath the ocean also helped to remove carbon dioxide from the air, since weathering of minerals consumes it.
Organisms capitalize upon the wastes of other organisms not only on the planetary scale but in local regions as well. if the soil in a certain location does not have very much nitrogen in it (in the form
of ammonia, nitrates, and nitrites), natural selec-tion may favor plants that form mutualistic relation-ships with bacteria, which fix atmospheric nitrogen gas into ammonia within root nodules. When these plants die, decomposers release nitrogen-containing molecules into the soil. Once the soil contains high levels of nitrogen compounds, there is no longer a selective advantage to the nitrogen fixation symbio-sis. in fact, in such circumstances, some bacteria can make a living by turning soil nitrogen compounds back into nitrogen gas. the result is a regulation of soil nitrogen content by organisms, each of which are acting in their own interests.
as a result, ecosystem regulation emerges from processes of negative feedback. this is not, however, a completely satisfying explanation, for two reasons, which follow:
● Organisms do not always evolve in response to negative feedback. Sometimes they evolve in response to positive feedback. that is, their wastes create an environment that is favorable to them, rather than to the other species of the world. One example is plants that pollute their surroundings with poi-sonous chemicals that inhibit the growth of other plants, a process known as alle-lopathy. their wastes allow them to domi-nate their habitats more, not less. another example is plants that are adapted to a fire cycle. they can tolerate fire, and their life cycles allow them to take advantage of fires and even to promote fires. (Some chaparral shrubs produce volatile, flammable chemi-cals in their old wood.) in a bog, sphagnum moss produces acid and grows as a thick clump, which prevents most other kinds of plants from growing with them. in each case, the plants benefit from creating condi-tions that are worse for other organisms, and other organisms have been unable to use these conditions as an opportunity for evolution and growth. therefore, the natu-ral world is as likely to contain examples of positive feedback as of negative feedback.
● negative feedback seems unable to explain how the conditions of life are kept within such narrow limits. there is no guarantee that negative feedback will operate quickly enough to keep earth’s conditions from getting out of control. eventually, negative feedback will prevail, but in the meantime, any number of conditions—temperature, oxygen, carbon dioxide—may fluctuate out-side the limits that life can tolerate. Why has this not occurred?
xviii+598_Biodiversity_F.indd 267 11/17/11 1:13 PM

first entry268 Gaia 268
One answer to these objections is simply luck. Plants that release allelopathic chemicals can, by chance, tolerate only a narrow range of conditions, and their polluting activities are limited to areas that have these conditions. Bogs are limited to small wetlands in which dense clay walls them off from the surrounding groundwater. a fire cycle can be established only in warm, dry regions. By chance, no organisms have produced any positive feedbacks that allow them to change the conditions of the whole world. earth is just lucky that neither oxygen nor car-bon dioxide levels have risen or fallen to deadly lev-els. Other planets may not have been so lucky, and on those planets, there are no people to ask these ques-tions. astrobiologist Peter Ward has pointed out that biological processes have sometimes in the past cre-
ated conditions that worsened the earth as a whole, for example the bacterial production of hydrogen sulfide during the Permian extinction. Ward refers to this as the “Medea hypothesis,” named after a venge-ful character in Greek mythology, as a contrast to Gaia. earth is lucky not only for being just the right distance from the Sun but because negative feedbacks have prevailed over positive ones.
another answer is that earth has not always been as well regulated as it now is. in fact, atmo-spheric temperature, carbon dioxide, and oxygen have reached levels in the past that were almost deadly. two examples follow:
● During Precambrian time, the earth almost completely froze, a condition popularly
Earth is a middle-aged planet. A little less than 5 billion years ago,
the rocks of the Earth’s crust cooled off enough that the oceans could form and, soon thereafter, bacterial life-forms were living in it. A little less than 5 bil-lion years from now, the Sun will have expanded enough that, even if it has not yet engulfed the Earth, life on this planet will have been obliterated, except for some bacteria living deep in the rocks. The universe is about 13.7 billion years old. If the age of the universe were equivalent to the distance across North America, from the Atlantic Ocean to the geographic center of the United States (which is a rock pedestal in Kansas), a human lifetime would be about the width of a finger. All of human civilization could fit into a few feet of space in front of that pedestal.
Earth is a planet that does not just contain biodiversity but is held together by biodiversity. Earth is not itself an organism, but it is a living planet, kept alive by its millions of species, most of which (to the human viewpoint) have evolved just in the most recent 10 per-cent of Earth’s history. Earth’s species have transformed it into a planet almost completely different from what it would otherwise have been or what the other
planets are. Photosynthesis has filled the atmosphere with oxygen and removed most of the carbon dioxide that was origi-nally present. Photosynthetic organisms, and the organisms that eat them, have created vast limestone deposits. Plants transpire a tremendous amount of water vapor into the atmosphere. Decompos-ers, consuming dead plant tissues, have turned sterile dirt into rich dark soil. Over much of the planet’s surface, the physical conditions (as experienced by medium-sized organisms) are not the direct prod-uct of sun and rain but are mediated through the shade cast by trees. All of its species form a living network.
And this network appears to have regulated the physical conditions of the planet. Such regulation was not in any way the intention of the organisms mak-ing up this network. Natural selection has favored organisms that pass their genes into the next generation most effectively. Period. But, perhaps just by chance, these organisms have stabilized the conditions of the Earth in the pro-cess of creating conditions suitable for themselves. Billions of years ago, the Sun was much dimmer, but the high levels of carbon dioxide in the atmosphere cre-ated a greenhouse effect. As the Sun has become brighter, photosynthesis
has removed much of the carbon diox-ide. That is, as the Sun became brighter, the greenhouse effect diminished, result-ing in temperatures that have remained within the general boundaries in which life is possible during that entire period of time. Organisms appear to be keeping the temperature, oxygen, and carbon dioxide levels within habitable limits.
Earth could be said to have had an unstable childhood and adolescence. Until about a half billion years ago, most life on Earth was microscopic. The con-tinents were barren, and there was not as much biomass in the oceans as there is today. Photosynthesis and respiration got out of balance. At least three times, massive ice sheets covered much of the planet, as microbial metabolism proved incapable of maintaining an adequate greenhouse effect. But once Earth entered its young adulthood, life began to proliferate into many, teeming, mul-ticellular forms, first in the oceans and then on the land. Ever since life began to cover the Earth, there have been no massive disruptions of the conditions of life—with a couple of exceptions: the Permian and Cretaceous extinctions. But these were caused by massive volcanic eruptions (in the first) and an asteroid impact (in the second). Neither of these
“life of eArTh: porTrAiT of A BeAuTiful, Middle-AGed, STreSSed-ouT plAneT”
xviii+598_Biodiversity_F.indd 268 11/17/11 1:13 PM

Gaia 269
known as Snowball earth. the reason may have been that microbial photosynthesis was so effective that it removed too much carbon dioxide from the air, endangering earth’s protective greenhouse effect. Had the snow-ball conditions been just a little worse or persisted just a little longer, the earth might never have warmed back up, since most of the sunlight would have been reflected back into outer space. it is possible that the rea-son this occurred in the first place is that there simply was not yet enough of either the diversity or the abundance of life. in this case, Gaia was not yet large enough of a system to prevent fluctuations as great as the one that allowed Snowball earth.
● During the Paleozoic era, small trees lived in shallow wetlands all over the continents. not only were the wetlands extensive, but the small trees were very crowded. these worldwide forests carried out a lot of photo-synthesis and put a lot of oxygen in the air. not only that, but herbivores and decom-posers were apparently unable to consume as wide a variety of plant tissues, living and dead, as they do today. the trees in these worldwide forests accumulated as coal. Decomposition did not occur enough to remove very much of the oxygen that these trees produced. Most scientists accept that the atmosphere contained very high levels of oxygen—about 35 percent, rather than
events tipped the Earth into a spiral of massive heating or cooling. Even during these times of crisis, the atmosphere was never as hot, or as cold, as it had been before and during the primordial greenhouse conditions or worldwide glaciations. Life itself now appeared to be in charge of the planet and of stabiliz-ing its conditions.
Earth is now middle-aged and is astonishingly beautiful—primarily because of the very same network of biodiversity that allowed it to survive and keeps it alive. Earth was not always a pleasant place for every species. Species interactions were as likely to be nega-tive (as with predators and parasites) as mutualistically positive. Nevertheless, Earth is at the peak of its beauty and health.
With the arrival of the human spe-cies, unprecedented levels of intelligence evolved. In an instant of time, relative to Earth’s life, it was as if life had become conscious of itself; and in an even shorter instant, just in the last few centuries, human minds have begun to understand how the Earth and its life-forms evolved. Humans today represent the most extreme examples of many evolutionary processes, such as sexual selection and altruism. And with human intelligence has come the unique ability of a species actually to become a deliberate force of nature. As no other species (except per-haps for the photosynthetic bacteria that
first filled the atmosphere with oxygen), humans have transformed the surface of the Earth, turning over a third of it into managed lands such as farms and cities, and transformed the atmosphere. Global warming has happened many times in the past but never as rapidly as it is occur-ring today as a result of human activity. Thus just at the moment that Earth might have, by means of human intelligence, begun to appreciate its own beauty, it has entered a period of stress. Earth is unlikely to die from this stress, but it may temporarily lose much of the stability that its biodiversity has allowed it to have—biodiversity that is being rapidly depleted by humans.
Conservationists, professional and amateur, who are trying to save the bio-diversity of the Earth are doing so not just for the sake of the individual spe-cies. The efforts to prevent global warm-ing are not just about polar bears, nor is the campaign against tropical deforesta-tion just about beautiful (and sinister) orchids. It is about saving the integrity of the network of life that allows Earth to maintain its life-supporting conditions. Nobody knows how many species can become extinct without that integrity beginning to unravel. Nobody knows how much environmental destruction it might take to turn the life-sustaining opera-tions into life-effacing ones. This is what nearly happened during the Permian extinction, when oxygen-starved oceans
emitted hydrogen sulfide gas into the atmosphere. The network of life saved the planet that time, just barely. And it will almost certainly do so again this time. Nobody is saying that the Sixth Mass Extinction or the greenhouse effect is going to make humans fall over dead. But the remaining biodiversity of Earth may not stabilize the planet until after the conditions of life have fluctuated out-side of the limits in which the network of civilization, upon which most humans depend for survival, have been exceeded. Will Earth enter its old age crippled?
See also BiogeocHemical cycles; cre-taceous extinction; gaia; mass extinc-tions; mutualism; parasitism; permian extinction.
Further readingLovelock, James. The Vanishing Face of Gaia:
A Final Warning. New York: Basic, 2009.Rice, Stanley A. Life of Earth: Portrait of a
Beautiful, Middle-Aged, Stressed-Out World. Amherst, N.Y.: Prometheus Books, 2010.
Ward, Peter. Under a Green Sky: Global Warming, the Mass Extinctions of the Past, and What They Can Tell Us About Our Future. New York: HarperCollins, 2007.
———, and Donald Brownlee. The Life and Death of Planet Earth: How the New Sci-ence of Astrobiology Charts the Ultimate Fate of Our World. New York: Henry Holt, 2003.
xviii+598_Biodiversity_F.indd 269 11/17/11 1:13 PM

first entry270 Gaia 270
the 21 percent that is in the air today. in this case, some important processes within the Gaia system had not yet evolved to their cur-rent levels of efficiency.
the earth today has more species and perhaps a greater mass of organisms than at any time in the past. if there had been as many organisms during the Precambrian as there are today, Snowball earth may not have happened. a green scum earth produced a Snowball earth, something that would not have happened if there had been an abundance of organ-isms that could eat the green scum. today, trees are consumed by animals when they are alive and fungi when they are dead. Photosynthesis would not get a chance to produce a hyperoxygenated atmosphere today as it did during the Paleozoic era. life itself regulates the earth much more effectively today than ever before in the past.
Positive feedback on a worldwide scale, in which organisms amplify rather than dampen the extremes of earth conditions, is rare but significant. the main example is the ice ages. the cool periods of gla-ciation and the warmer periods of glacial retreat are initiated by slight changes in the amount of sunlight that reaches the earth, as a result of the Milanković cycles, in which changes in the shape of earth’s orbit and the angle of earth’s rotation produce an approx-imately 100,000-year cycle of cooling and warm-ing. the changes in solar radiation are very slight, enough to cause less than one-tenth of the amount of temperature fluctuation that characterized the ice ages. Biological processes, however, appear to enhance the Milanković cycle. the slight increase in sunlight and warmth allows more decomposition, which enhances the greenhouse effect and ampli-fies the warming. Photosynthesis removes enough carbon dioxide that the greenhouse effect is damp-ened. then when the slight decrease in sunlight and warmth occurs, the lack of a greenhouse effect amplifies the cooling. in this way, atmospheric car-bon dioxide levels are both a cause and an effect of global warming.
the rarity of such positive feedback mecha-nisms, however, assures that the “butterfly effect” cannot be true. the butterfly effect is a common aphorism that the flapping of a butterfly’s wings in one part of the world can cause a storm to occur in another part. instead, the second law of ther-modynamics insures that the random movement of molecules will drown out small events, such as the movement of a butterfly, rather than amplify them.
the Gaia hypothesis indicates that organisms do not simply live upon the earth; evolution has not simply caused organisms to adapt to the physi-cal conditions of the earth. Organisms have radi-
cally transformed the earth and have partly created the very conditions to which evolution has adapted them. lynn Margulis says that Gaia is just symbiosis seen from outer space. it may therefore be not just certain species interactions but the entire planet that is symbiotic.
Some critics of the Gaia hypothesis claim that it is merely an earth-goddess religion cloaked in scientific terminology. a religious view of earth ecosystems is especially threatening to conservative and religiously motivated critics, who react to Gaia as to a pagan religion used in the service of envi-ronmentalism, which they oppose. But it is clear that conservative antienvironmentalists have their own, thoroughly unscientific, version of Gaia, upon whose altar they are willing to place the survival of the planet and humankind. they believe that, no matter what humans do, no matter how much car-bon is released into the air or how many pesticides into the waters, the earth will clean up the mess and keep everything—including temperature, rain-fall, and soil fertility—in a range that is comfortable to humans. this may be the principal motivation behind conservative, especially religious, opposition to any laws or treaties regarding carbon emissions or pollution. as noted previously, life is much better at regulating the conditions of the earth than it ever was in the past; but this ability is limited. Human activities are pushing the earth beyond the ability of Gaia to regulate it. But to the conservatives in the Republican Party, there is nothing that Gaia cannot do. their opposition to environmentalism is based on a version of earth-religion far more extreme than has been proposed by any apologist for Gaia. the result is likely to be an earth that is much warmer—not warm enough to threaten life in general but warm enough to be disastrous to human activities, one that may be known to future generations as “the Republican climate.”
See also essay: “life of earth: Portrait of a Beau-tiful, Middle-aged, Stressed-out Planet”; carbon cycle; competition, ecological; disturbances, ecological; ecosystems; greenhouse effect; ice ages; Margulis, lynn; nitrogen cycle.
Further readingGribbin, John, and Mary Gribbin. James Lovelock: In
Search of Gaia. Princeton, n.J.: Princeton University Press, 2009.
Hamilton, William D. “ecology in the large: Gaia and Genghis Khan.” Journal of Applied Ecology 32 (1995): 451–453.
———. “Gaia’s benefits.” New Scientist 151 (1996): 62–63.———, and timothy M. lenton. “Spora and Gaia: How
microbes fly with their clouds.” Ecology, Ethology and Evolution 10 (1998): 1–16.
xviii+598_Biodiversity_F.indd 270 11/17/11 1:13 PM

Galápagos Islands 271
Jaenicke, Ruprecht. “abundance of cellular material and proteins in the atmosphere.” Science 308 (2005): 73.
lenton, timothy M. “Gaia and natural selection.” Nature 394 (1998): 439–447.
———. “testing Gaia: the effect of life on earth’s habit-ability and regulation.” Climatic Change 52 (2002): 409–422.
———, and M. van Oijen. “Gaia as a complex adaptive system.” Philosophical Transactions of the Royal Soci-ety of London B (Biology) 357 (2002): 683–695.
lovelock, James. The Vanishing Face of Gaia: A Final Warning. new york: Basic, 2009.
———. Gaia: A New Look At Life On Earth. new york: Oxford University Press, 1987.
———, and lynn M. Margulis. “atmospheric homeostasis by and for the biosphere: the Gaia Hypothesis.” Tellus 26 (1974): 2–10.
———, and M. Whitfield. “life-span of the biosphere.” Nature 296 (1982): 561–563.
Malin, G. and G. O. Kirst. “algal production of dimethyl sulfide and its atmospheric role.” Journal of Phycology 33 (1997): 889–896.
Margulis, lynn, and James e. lovelock. “Biological mod-ulation of earth’s atmosphere.” Icarus 21 (1974): 471–489.
———, and Dorion Sagan. Slanted Truths: Essays on Gaia, Symbiosis, and Evolution. new york: copernicus, 1997.
Rice, Stanley a. Life of Earth: Portrait of a Beautiful, Middle-Aged, Stressed-Out World. amherst, n.y.: Prometheus, 2010.
Schneider, Stephen H., et al., eds. Scientists Debate Gaia. cambridge, Mass.: Mit Press, 2004.
Schwartzman, D. W., and tyler Volk. “Biotic enhancement of weathering and the habitability of earth.” Nature 340 (1989): 457–460.
Swenson, W., David Sloan Wilson, and R. elias. “artifi-cial ecosystem selection.” Proceedings of the National Academy of Sciences USA 97 (2000): 9,110–9,114.
Volk, tyler. “Gaia is life in a wasteworld of by-products.” in Scientists Debate Gaia, edited by Stephen H. Sch-neider et al. cambridge, Mass.: Mit Press, 2004.
———. Gaia’s Body: Toward a Physiology of Earth. new york: copernicus, 1998.
Williams, George c. “Gaia, nature worship, and biocentric fallacies.” Quarterly Review of Biology 76 (2002): 479–486.
galápagos islands the Galápagos islands are volcanic islands that have played and continue to play a very important role in evolutionary sci-ence and are the showcase of how biodiversity can develop. they belong to ecuador and are about 600 miles (1,000 km) off the coast of that country. Much of the volcanic activity that produced them occurred
about 5 million years ago, which is very recent in evolutionary time.
Being directly on the equator, these islands experience almost no variation in sunlight. they are right in the path of oceanic currents that influ-ence temperature and moisture conditions. there is an annual fluctuation of temperature caused by the ocean currents. Because of this, the cactuses can use a temperature cue that allows them to bloom every January. the el niño Southern Oscillation alter-nates between dry conditions, when the ocean water near the South american coast is cool, and wet conditions, when the water is warm. During an el niño year, large amounts of rainfall stimulate plant growth, particularly of grasses, vines, and weeds, but diminish the productivity of the oceans. the food chain of the ocean depends upon the miner-als brought up from cool, deep ocean waters; when the water is warm, these minerals are not available. However, periods of heavy rainfall are uncommon, and the lower slopes of the Galápagos islands are usually hot and dry, with scattered trees and a lot of hot, bare ground. the upland areas of the larger islands are frequently shrouded in mist, which pro-vides enough moisture for forests to grow.
the islands are famous for their unusual plant and animal life. there are no large native graz-ing animals or predators. the native animals have evolved, until recently, in the absence of preda-tors and human contact. From the fur seals to the ground-nesting birds, they show no fear of humans. Many of the species are unique to these islands, or almost so. this is because they evolved on the islands from small founding populations of main-land species. all over the world, islands are home to unique species. the unique species found on islands was one of the observations that led charles Darwin to begin thinking about the process of evolution. During his round-the-world voyage on HMS Beagle, he visited many islands, including the Galápagos. His 1835 visit to the Galápagos turned out to be per-haps the single most important event that sparked Darwin’s understanding that evolution occurred by means of natural selection.
among the plants and animals that have under-gone adaptive radiation on the Galápagos islands are:
● Finches. these birds have evolved from a single founding population into about 14 species that specialize on different ways of life and types of food. ever since David lack studied them in 1947, they have constituted a premier example of adaptive radiation. Since Peter and Rosemary Grant began their studies in the 1970s, the finches have pro-vided some of the most detailed information
xviii+598_Biodiversity_F.indd 271 11/17/11 1:13 PM

first entry272 Galápagos Islands 272
in the world about natural selection, specia-tion, and hybridization.
● Mockingbirds. a single founding popula-tion, probably of the ecuadorian long-tailed mockingbird Mimus longicaudatus, has evolved into four species of mockingbirds on the Galápagos: Nesomimus trifasciatus, N. macdonaldi, N. melanotis, and N. par-vulus. they are distinct from one another in coloration and behavior; they differ enough from their mainland ancestors that they have been placed in a different genus, Nesomi-mus, from the genus Mimus of which the north and South american mainland mock-ingbirds are members.
● Land iguanas, or lava lizards. Seven spe-cies of land iguanas have evolved on the Galápagos islands.
● Tortoises. the giant tortoises after which the Galápagos islands are named are the most famous animals in the islands. Several of the islands have recognizably different kinds of tortoises—a fact that the governor of the islands pointed out to charles Dar-win when he visited. the tortoises move very slowly and live for a long time; a slow metabolism is a major part of their adapta-tion to these islands. Darwin rode on some when he visited. a few of the tortoises alive today may have been alive when Darwin visited, though this is unlikely.
● Daisy trees. the Galápagos islands have 13 species of Scalesia trees, in the composite (daisy) family asteraceae. almost all mem-bers of this huge, worldwide family of plants are herbaceous (like the familiar daisies, dandelions, thistles, and ragweeds)—except on islands, such as the channel islands of california, St. Helena in the atlantic, Hawaii, and the Galápagos. these daisy trees are somewhat flimsy and small as trees go and probably could not compete with trees from plant families with a long evolu-tionary history of producing trees, such as pines and oaks. On these islands, there are relatively few trees, which has allowed natu-ral selection to produce tree-sized daisies.
● Tree cactuses. cactuses in mainland north and South american deserts do not have bark; the six species of Galápagos prickly pear cactuses, however, do. the bark may have evolved as protection against the abun-dant sea animals that eat them.
there are also species that are unique to the Galápagos islands, although they have not under-
gone as much adaptive radiation as can be seen in the animals mentioned above. two hundred of the 700 species of Galápagos plants are unique, including the Galápagos tomato. Marine iguanas are highly unusual in the reptile world because they swim and eat algae. the Galápagos penguin is the only pen-guin that lives in a warm environment. there are Galápagos species of sharks, doves, hawks, rats, bees, butterflies, centipedes, martins, and flycatchers (see table).
Other animals such as frigate birds travel widely in the oceans and have not evolved into unique forms confined to the islands. the isolation of the islands makes them a haven for birds and mammals that travel the oceans.
some noteWorthy speCies of the galápagos islands
Plants
Brachycereus nesioticus Lava cactus (Cactaceae)
Bursera graveolens Palo santo (Burseraceae)
Miconia robinsoniana Cacaotillo (Melastomataceae)
Opuntia megasperma Opuntia (Cactaceae)
Scalesia spp. Daisy trees (Asteraceae)
Scutia pauciflora Espino bush (Rhamnaceae)
Solanum cheesemanii Galápagos tomato (Solanaceae)
Animals
Anblyrhynchus cristatus Marine iguana
Buteo galapagoensis Galápagos hawk
Chelonia mydas agassisi Galápagos green turtle
Conolophus spp. Land iguana
Geochelone elephantopus Galápagos giant tortoise
Grapsus grapsus Sally lightfoot crab
Phalacrocorax harrisi Flightless cormorant
Phoebastria irrorata Waved albatross
Spheniscus mendiculus Galápagos penguin
Sula nebouxii Blue-footed booby
Zalophus wollebaeki Galápagos sea lions
See text for other species.
xviii+598_Biodiversity_F.indd 272 11/17/11 1:13 PM

Grant, peter and rosemary 273
the Galápagos islands offer very little in the way of natural resources. they have been used for centuries as a place where ships could stop, but they were not even very good for this, for they have very little freshwater. the tortoises were actually a popu-lar attraction for sailing ships. Because of their slow metabolism, the tortoises could survive for many weeks in the cargo holds of the ships and could sup-ply fresh meat on long voyages. When Darwin vis-ited, one of the islands was used as a prison colony.
Since none of the native animals (except the tortoises) had economic value to the europeans, they introduced livestock that could survive the harsh island conditions. today, populations of feral animals continue to damage the natural veg-etation. the most abundant feral animal is the goat. Because of grazing by goats, the native cac-tus Opuntia megasperma nearly became extinct; the tortoises, now rare, also dispersed the cactus seeds. Goats have been eradicated from some of the islands. introduced black rats (Rattus rattus) attack young tortoises. Rats prevented tortoises from reproducing on the island of Pinzón for a half century, during which time only adult tortoises lived on the island. cows and donkeys have been introduced also, which eat the vegetation and com-pete with native animals for water. Feral dogs and cats often kill birds.
a considerable part of the Galápagos islands is ecuadorian national parkland. cruise ships and air-plane flights take thousands of tourists each year to see some of what Darwin saw.
in 2007, the United nations educational, Scien-tific and cultural Organization (UneScO) declared the Galápagos islands to be endangered by invasive species, immigration, and the impact of tourism. after the ecuadorian government took measures to control these problems, UneScO removed the Galápagos islands from its list of endangered habi-tats in July 2010.
See also adaptive radiation; biogeography; Darwin, charles; Darwin’s finches; hybrid-ization; invasive species; natural selection; speciation.
Further readingadsersen, Henning. “the rare plants of the Galápagos
islands and their conservation.” Biological Conserva-tion 47 (1989): 49–77.
campbell, Karl, et al. “eradication of feral goats Capra hircus from Pinta island, Galápagos, ecuador.” Oryx 38 (2004): 328–333.
causton, c. e., et al. “alien insects: threats and implica-tions for conservation of Galápagos islands.” Annals of the Entomological Society of America 99 (2006): 121–143.
charles Darwin Foundation. “charles Darwin Founda-tion.” available online. URl: http://www.darwinfoun-dation.org/english/pages/index.php. accessed July 28, 2010.
DeRoy, tui. Galápagos: Preserving Darwin’s Legacy. Ontario: Firefly Books, 2009.
Dowler, Robert c., et al. “Rediscovery of rodents (genus Nesoryzomys) considered extinct in the Galápagos islands.” Oryx 34 (2000): 109–117.
Galápagos conservancy. available online. URl: http://www.galapagos.org/2008/index.php?id=68. accessed august 8, 2011.
Kricher, John. Galápagos: A Natural History. Princeton, n.J.: Princeton University Press, 2006.
Stewart, Paul D. Galápagos: The Islands that Changed the World. new Haven, conn.: yale University Press, 2007.
Whiteman, noah Kerness, et al. “Disease ecology in the Galápagos Hawk (Buteo galapagoensis): host genetic diversity, parasite load and natural antibodies.” Pro-ceedings of the Royal Society of London B (Biology) 273 (2006): 797–804.
genetic drift See founder effect and genetic drift.
grant, peter and rosemary (1936– ) and (1936– ) British Evolutionary biologists Peter Raymond Grant and Barbara Rosemary Grant are among the most famous living evolutionary biolo-gists. their work with Darwin’s finches on the Galápagos islands has provided the most thorough set of data upon which modern scientists base their understanding of how evolution produces biodiver-sity. Since 1973, they have spent half of each year on the islands of Daphne Major and Genovesa. they have numbered, tagged, and measured nearly every finch on Daphne Major and recognize most of them. in this way, they can understand the lives of each of the finches as if they were part of a complex Russian novel. they can determine which finches mate with which others and which finches are their offspring. in this way, they have been able to demonstrate the heritability of traits; natural selection acting upon those traits; and character displacement in those traits caused by competition between finch species.
Peter Grant was born October 26, 1936, in lon-don, and attended cambridge University. Rosemary was born on October 8, 1936, in a small village on the northwest coast of england and attended edin-burgh University. Both grew up with a passion for the natural world. Peter met Rosemary when he was a graduate student and she a lecturer at the University
xviii+598_Biodiversity_F.indd 273 11/17/11 1:13 PM

first entry274 grasslands and savannas 274
of British columbia. they married, and Peter com-pleted his graduate work on the evolution of birds on tropical islands. Peter and Rosemary went to yale University, he as a postdoctoral researcher and she as a lecturer. they had two daughters, both of whom undertook research projects, later published, in the Galápagos. after university positions in Montreal and Michigan, and after Rosemary completed her doctoral degree, both Peter and Rosemary became faculty members at Princeton University. Both have retired but continue their research. Rosemary’s greatest wish is “that we have transmitted, and will continue to transmit, some of our own sense of won-der, joy, and excitement in curiosity-driven research and in life to our students and to others.”
See also Darwin’s finches; Galápagos islands; natural selection.
Further readingGrant, Peter, and Rosemary Grant. “in charles Dar-
win’s footsteps.” available online. URl: http://www.inamori-f.or.jp/laureates/k25_b_peter/img/lct_e.pdf. accessed august 15, 2010.
Weiner, Jonathan. The Beak of the Finch: A Story of Evolu-tion in Our Time. new york: Knopf, 1994.
grasslands and savannas Grasslands are regions dominated by grasses, and savannas are grasslands with individual trees. even though grasses comprise the majority of the biomass of grasslands, most of the species of plants are forbs, which are her-baceous wildflowers. Grasslands are generally found in regions that are drier than boreal forests or tem-perate forests but wetter than deserts. Grasslands are not defined by temperature; some grasslands, such as the prairies of Saskatchewan, can be very cold, while others, such as the prairies of texas, can be very warm, and yet not only be grasslands but contain many of the same grass and forb spe-cies. Buffalo grass (Buchloë dactyloides) is one of the species that grows from texas to Saskatchewan. Grasses, along with sedges, rushes, and cattails, can dominate marshes, such as the everglades of Florida, which are treeless and flooded; these are not called grasslands.
the wettest grasslands in north america are the tallgrass prairies, which once dominated what is now western Minnesota, iowa, eastern north and South Dakota, eastern nebraska, eastern Kansas, and north central Oklahoma (see table on page 275). Many of the grasses and forbs could grow over six feet (two m) tall; a person on horseback could hide in some of these grasslands. they received enough rain and snow that deciduous broad-leaved forests could grow in these regions. However, periodic fires burned the tallgrass prairies, killing any trees
that happened to grow there. the grasses and forbs would resprout from underground buds. the fire would not only remove the trees but also transform dead grass stalks into ashes that acted as fertilizers and allowed sunshine to warm the ground more quickly in the springtime. Fires, by keeping out the trees and promoting the growth of grasses and forbs, were essential to the tallgrass prairie. therefore the grasses themselves help to maintain the boundary between prairie and forest. the tallgrass prairie became established during a period of weather after the most recent of the ice ages that was warmer and drier than the present—when it was too dry for for-ests to grow there. after wetter weather returned, fires kept the deciduous trees from growing there. the tallgrass prairie was one of the last regions to be converted to agriculture; the thick sod, with roots penetrating up to 10 feet (three m) into the soil, could not be plowed until the invention of the steel plow. the topsoil, very rich and deep, proved to be some of the world’s best soil, and nearly the entire tallgrass prairie was converted to farmland. Most of it today is dominated by another tall grass: corn. For the most part, only small parcels of prairie were protected, mostly as pioneer cemeteries and railroad rights-of-way (see photograph). the tallgrass prairie has been more completely destroyed than any other natural habitat on earth.
the driest grasslands are the shortgrass prairies, forming a band across western north and South Dakota, nebraska, Kansas, and Oklahoma, and eastern Montana, Wyoming, colorado, and new Mexico. there is also shortgrass prairie (the Palouse)
A remnant tallgrass prairie grows along a railroad track in Illinois. Most original prairies are found near railroads or cemeteries, the only places that were not transformed into farmland by settlers. The railroad company bulldozed the prairie plants that were along the track in the mistaken impression that the plants were a nuisance. Soon thereafter, the embankment began to erode and was soon filled with ragweeds and poison ivy, which the thick prairie sod had previ-ously kept out. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 274 11/17/11 1:13 PM
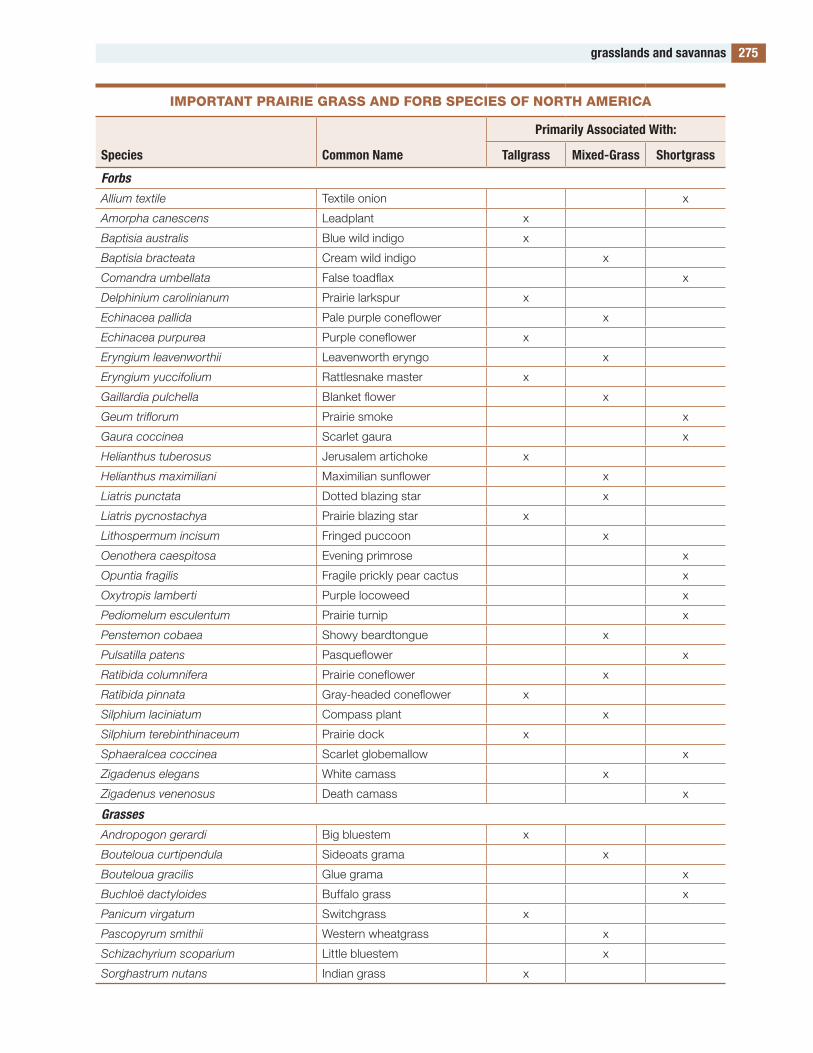
grasslands and savannas 275
important prairie grass and forb speCies of north ameriCa
Species Common Name
primarily associated with:
Tallgrass mixed-Grass Shortgrass
ForbsAllium textile Textile onion x
Amorpha canescens Leadplant x
Baptisia australis Blue wild indigo x
Baptisia bracteata Cream wild indigo x
Comandra umbellata False toadflax x
Delphinium carolinianum Prairie larkspur x
Echinacea pallida Pale purple coneflower x
Echinacea purpurea Purple coneflower x
Eryngium leavenworthii Leavenworth eryngo x
Eryngium yuccifolium Rattlesnake master x
Gaillardia pulchella Blanket flower x
Geum triflorum Prairie smoke x
Gaura coccinea Scarlet gaura x
Helianthus tuberosus Jerusalem artichoke x
Helianthus maximiliani Maximilian sunflower x
Liatris punctata Dotted blazing star x
Liatris pycnostachya Prairie blazing star x
Lithospermum incisum Fringed puccoon x
Oenothera caespitosa Evening primrose x
Opuntia fragilis Fragile prickly pear cactus x
Oxytropis lamberti Purple locoweed x
Pediomelum esculentum Prairie turnip x
Penstemon cobaea Showy beardtongue x
Pulsatilla patens Pasqueflower x
Ratibida columnifera Prairie coneflower x
Ratibida pinnata Gray-headed coneflower x
Silphium laciniatum Compass plant x
Silphium terebinthinaceum Prairie dock x
Sphaeralcea coccinea Scarlet globemallow x
Zigadenus elegans White camass x
Zigadenus venenosus Death camass x
GrassesAndropogon gerardi Big bluestem x
Bouteloua curtipendula Sideoats grama x
Bouteloua gracilis Glue grama x
Buchloë dactyloides Buffalo grass x
Panicum virgatum Switchgrass x
Pascopyrum smithii Western wheatgrass x
Schizachyrium scoparium Little bluestem x
Sorghastrum nutans Indian grass x
xviii+598_Biodiversity_F.indd 275 11/17/11 1:13 PM

first entry276 grasslands and savannas 276
in eastern Oregon and Washington. Shortgrass prai-rie also grew on the foothills of the Sierra nevada in california. the topsoil and plant roots are much shallower in the shortgrass than in the tallgrass prai-rie. the grasses such as the grama grasses (see table on page 275) may be only a foot (⅓ m) tall; some, such as buffalo grass (Buchloë dactyloides) may be only four inches (10 cm) tall. it is the lack of mois-ture that keeps trees from growing in the shortgrass prairie; where water is available, as along rivers, trees (mostly cottonwood, Populus deltoides) can grow. Fire is essential to maintaining the shortgrass prairie also, not by killing tree seedlings but by kill-ing cool desert shrubs such as sagebrush in the Great Basin and warm desert shrubs such as creosote bush in the southern Great Plains.
a band of grasslands intermediate between tall and shortgrass prairies dominate the landscape across central north and South Dakota, nebraska, Kansas, and Oklahoma, making up the mixed-grass prairies, dominated by grasses that grow about three feet (one m) tall (see table on page 275).
the grasses and forbs of the tallgrass, mixed-grass, and shortgrass prairies of north america are often different species of the same genus that have different abilities to tolerate drought. For example,
the prairie dock, Silphium terebintinaceum, has enormous leaves without lobes, while the compass plant (S. laciniatum) has smaller leaves that are lobed. Smaller leaves and lobes allow a leaf to dis-seminate its heat load without transpiring as much water. in addition, the compass plant tends to orient its leaf blades to face east and west (hence its name), thus avoiding direct sunlight at noon. the forbs of the tallgrass prairie, like the grasses, have deep roots. in the shortgrass prairies, however, the root sod is formed mostly by the grasses; the forbs grow mostly in areas disturbed by animals and erosion.
Grasslands around the world contain or once contained large grazing mammals and other mam-mals such as rodents that act as keystone species that determine the structure of the entire ecological community. Grasses and grazing mammals have coevolved: the mammals have complex digestive systems with symbiotic bacteria that allow them to digest coarse grass, while the grasses grow from buds that are near or below the surface, allowing them to grow back after grazing as well as after fire.
Grasslands and savannas are found in califor-nia, australia, South america (especially the pampas of argentina), the steppes of central asia, and the
The savannas of Africa support many large mammals, such as elephants. (Fogstock)
xviii+598_Biodiversity_F.indd 276 11/17/11 1:13 PM
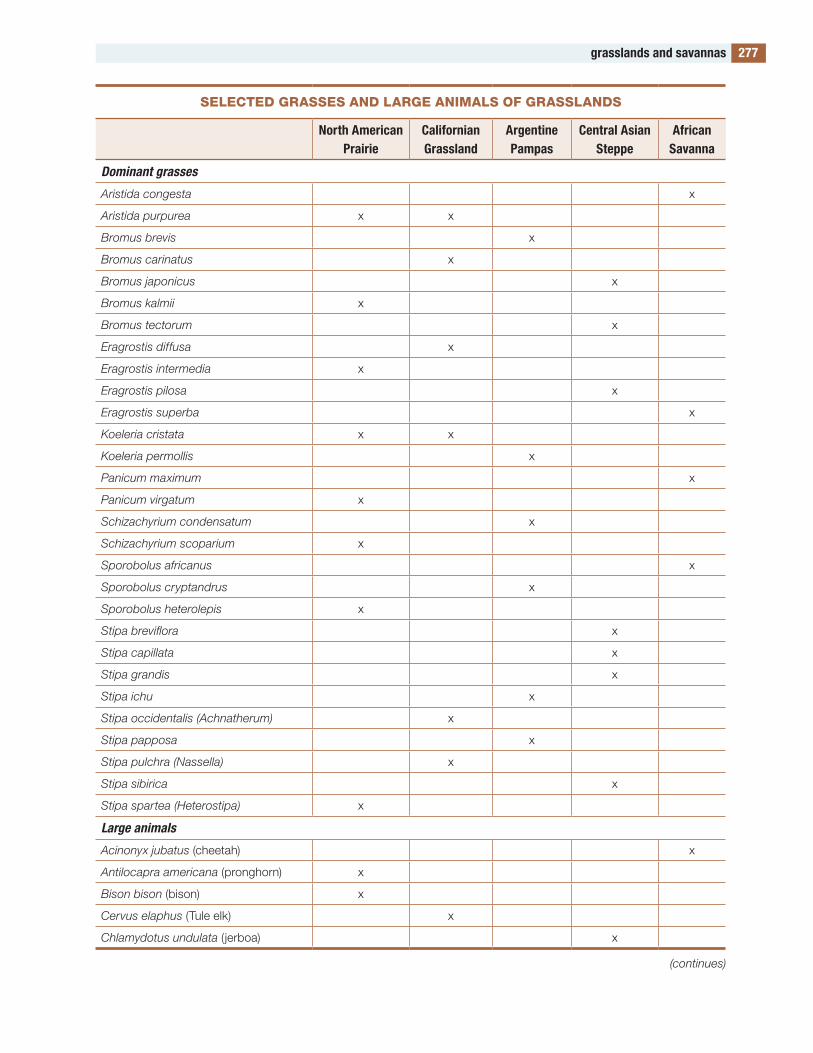
grasslands and savannas 277
seleCted grasses and large animals of grasslands
North american prairie
Californian Grassland
argentine pampas
Central asian Steppe
african Savanna
Dominant grasses
Aristida congesta x
Aristida purpurea x x
Bromus brevis x
Bromus carinatus x
Bromus japonicus x
Bromus kalmii x
Bromus tectorum x
Eragrostis diffusa x
Eragrostis intermedia x
Eragrostis pilosa x
Eragrostis superba x
Koeleria cristata x x
Koeleria permollis x
Panicum maximum x
Panicum virgatum x
Schizachyrium condensatum x
Schizachyrium scoparium x
Sporobolus africanus x
Sporobolus cryptandrus x
Sporobolus heterolepis x
Stipa breviflora x
Stipa capillata x
Stipa grandis x
Stipa ichu x
Stipa occidentalis (Achnatherum) x
Stipa papposa x
Stipa pulchra (Nassella) x
Stipa sibirica x
Stipa spartea (Heterostipa) x
Large animals
Acinonyx jubatus (cheetah) x
Antilocapra americana (pronghorn) x
Bison bison (bison) x
Cervus elaphus (Tule elk) x
Chlamydotus undulata (jerboa) x
(continues)
xviii+598_Biodiversity_F.indd 277 11/17/11 1:13 PM
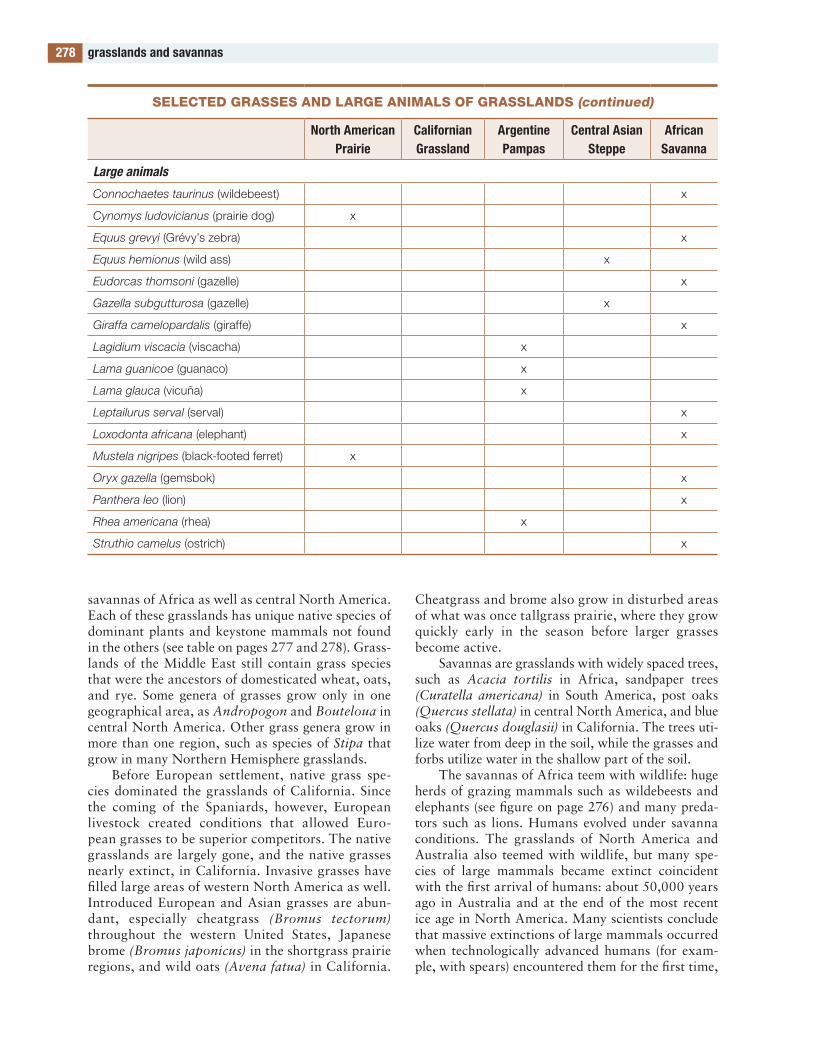
first entry278 grasslands and savannas 278
savannas of africa as well as central north america. each of these grasslands has unique native species of dominant plants and keystone mammals not found in the others (see table on pages 277 and 278). Grass-lands of the Middle east still contain grass species that were the ancestors of domesticated wheat, oats, and rye. Some genera of grasses grow only in one geographical area, as Andropogon and Bouteloua in central north america. Other grass genera grow in more than one region, such as species of Stipa that grow in many northern Hemisphere grasslands.
Before european settlement, native grass spe-cies dominated the grasslands of california. Since the coming of the Spaniards, however, european livestock created conditions that allowed euro-pean grasses to be superior competitors. the native grasslands are largely gone, and the native grasses nearly extinct, in california. invasive grasses have filled large areas of western north america as well. introduced european and asian grasses are abun-dant, especially cheatgrass (Bromus tectorum) throughout the western United States, Japanese brome (Bromus japonicus) in the shortgrass prairie regions, and wild oats (Avena fatua) in california.
cheatgrass and brome also grow in disturbed areas of what was once tallgrass prairie, where they grow quickly early in the season before larger grasses become active.
Savannas are grasslands with widely spaced trees, such as Acacia tortilis in africa, sandpaper trees (Curatella americana) in South america, post oaks (Quercus stellata) in central north america, and blue oaks (Quercus douglasii) in california. the trees uti-lize water from deep in the soil, while the grasses and forbs utilize water in the shallow part of the soil.
the savannas of africa teem with wildlife: huge herds of grazing mammals such as wildebeests and elephants (see figure on page 276) and many preda-tors such as lions. Humans evolved under savanna conditions. the grasslands of north america and australia also teemed with wildlife, but many spe-cies of large mammals became extinct coincident with the first arrival of humans: about 50,000 years ago in australia and at the end of the most recent ice age in north america. Many scientists conclude that massive extinctions of large mammals occurred when technologically advanced humans (for exam-ple, with spears) encountered them for the first time,
seleCted grasses and large animals of grasslands (continued)
North american prairie
Californian Grassland
argentine pampas
Central asian Steppe
african Savanna
Large animals
Connochaetes taurinus (wildebeest) x
Cynomys ludovicianus (prairie dog) x
Equus grevyi (Grévy’s zebra) x
Equus hemionus (wild ass) x
Eudorcas thomsoni (gazelle) x
Gazella subgutturosa (gazelle) x
Giraffa camelopardalis (giraffe) x
Lagidium viscacia (viscacha) x
Lama guanicoe (guanaco) x
Lama glauca (vicuña) x
Leptailurus serval (serval) x
Loxodonta africana (elephant) x
Mustela nigripes (black-footed ferret) x
Oryx gazella (gemsbok) x
Panthera leo (lion) x
Rhea americana (rhea) x
Struthio camelus (ostrich) x
xviii+598_Biodiversity_F.indd 278 11/17/11 1:13 PM

greenhouse effect 279
but such extinctions did not occur in places such as africa in which the large mammals and humans had evolved together for millions of years.
the most famous large mammal of the north american prairie is the bison. When white ameri-cans were traveling and homesteading across the Great Plains, they saw herds of millions of bison, and native americans hunting them. they assumed that this is the way it had always been—then proceeded to attempt the extermination of the bison in order to undermine the economy of the native tribes. Recent scholarship has suggested, however, that hunting by native americans had kept the bison populations under control for thousands of years, and the bison population exploded only after diseases such as smallpox reduced native populations by as much as 90 percent. a similar explanation has been offered for why there were so many billions of passenger pigeons in the forests of eastern north america just prior to white american expansion. according to the earliest explorers, who visited native tribes prior to their decimation by smallpox, native americans extensively transformed the landscape by agriculture and fire. if this is true, then much of the “original” distribution of the north american prairies, imme-diately before white settlement, may have been as much due to native american land management as to natural processes.
See also communities, ecological; des-erts, cool; deserts, warm; disturbances, eco-logical; invasive species; keystone species; marshes and swamps.
Further readingarcher, Steve, et al. “autogenic succession in a subtropi-
cal savanna: conversion of grassland to thorn wood-land.” Ecological Monographs 58 (1988): 111–127.
Brown, lauren. Grasslands. new york: Knopf, 1985.carlsen, tina M., et al. “Reducing competitive suppression
of a rare annual forb by restoring native california perennial grasslands.” Restoration Ecology 8 (2001): 18–29.
Hamilton, J. G., et al. “coexistence and interference between a native perennial grass and nonnative annual grasses in california.” Oecologia 121 (1999): 518–526.
Hayes, Grey F., and Karen D. Holl. “cattle grazing impacts on annual forbs and vegetation composition of mesic grasslands in california.” Conservation Biology 17 (2003): 1,694–1,702.
Hector, a., et al. “Plant diversity and productivity experi-ments in european grasslands.” Science 286 (1999): 1,123–1,127.
Huenneke, laura Foster, et al. “effects of soil resources on plant invasion and community structure in californian serpentine grassland.” Ecology 71 (1990): 478–491.
Kirt, Russell R. Prairie Plants of the Midwest: Identifica-tion and Ecology. champaign, ill: Stipes Publishing, 1998.
Kurtz, carl. A Practical Guide to Prairie Reconstruction. iowa city: University of iowa Press, 2001.
Mcnaughton, S. J. “Serengeti grassland ecology: the role of composite environmental factors and contingency in community organization.” Ecological Monographs 53 (1983): 291–320.
Manning, Richard. Grassland: The History, Biology, Poli-tics, and Promise of the American Prairie. new york: Penguin, 1997.
Menke, John W. “Grazing and fire management for native perennial grass restoration in california grasslands.” Fremontia 20 (1992): 22–25.
Moul, Francis, and Georg Joutras. The National Grass-lands: A Guide to America’s Undiscovered Treasures. lincoln: University of nebraska Press, 2006.
O’connor, t. G., et al. “influence of precipitation and spe-cies composition on phytomass of a semi-arid african grassland.” Journal of Ecology 89 (2002): 850–860.
Seabloom, eric W., et al. “invasion, competitive dominance, and resource use by exotic and native california grass-land species.” Proceedings of the National Academy of Sciences USA 100 (2003): 13,384–13,389.
tilman, David, and John a. Downing. “Biodiversity and stability in grasslands.” Nature 367 (1994): 363–365.
Uys, Roger G., et al. “the effect of different fire regimes on plant diversity in southern african grasslands.” Bio-logical Conservation 118 (2004): 489–499.
greenhouse effect the earth is getting warmer and humans are the principal cause. Humans have increased the amount of carbon dioxide (cO2) gas in the atmosphere, and this gas retains heat in the atmosphere, in a manner suggestive of the way a glass roof holds sunlight heat in a greenhouse. along with global warming have come many other changes in climate. consensus of scientists on these points, and about the importance that they have for the future of natural ecosystems and human society, has grown over recent decades and is now as strong as any scientific consensus can be.
ThE EarTh IS GETTING warmErin order to determine whether global climate is becoming warmer, it is necessary to determine tem-perature patterns over extended periods of time and over the whole earth. climate can be thought of as a long-term average of weather conditions. One hot year does not prove that global climate is becom-ing warmer. However, scientific evidence shows that the climate of the entire earth has gotten abruptly warmer in the last couple of centuries.
xviii+598_Biodiversity_F.indd 279 11/17/11 1:13 PM
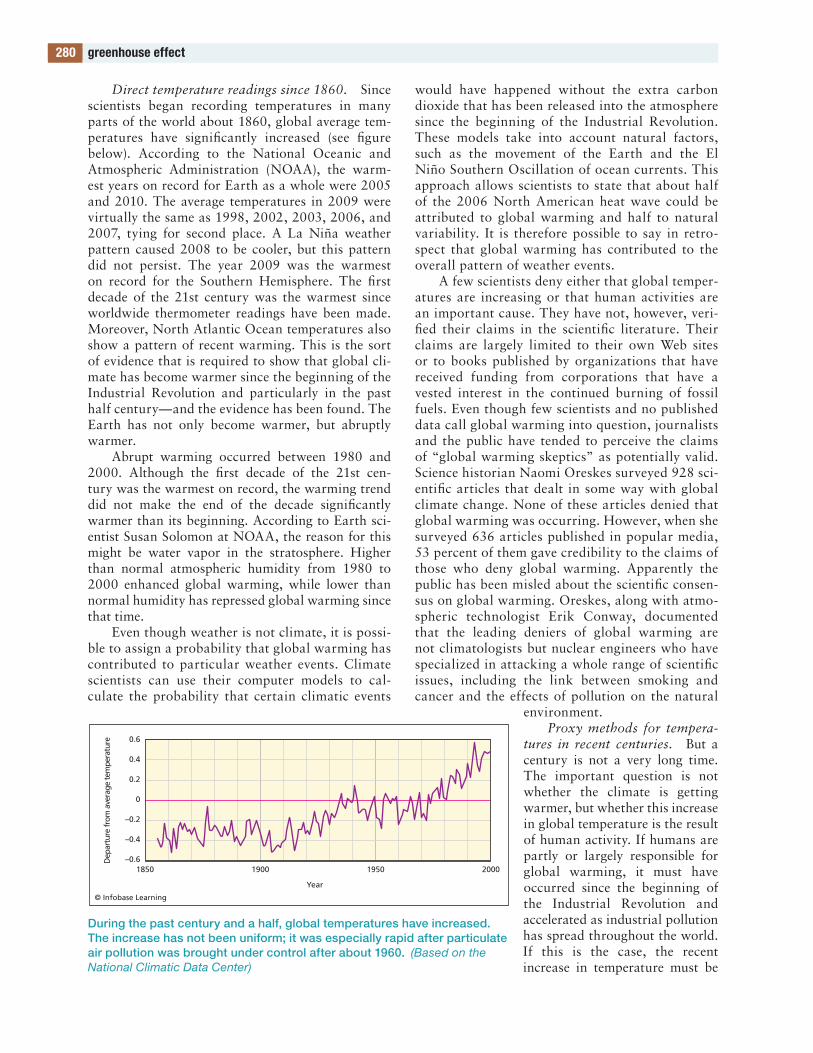
first entry280 greenhouse effect 280
Direct temperature readings since 1860. Since scientists began recording temperatures in many parts of the world about 1860, global average tem-peratures have significantly increased (see figure below). according to the national Oceanic and atmospheric administration (nOaa), the warm-est years on record for earth as a whole were 2005 and 2010. the average temperatures in 2009 were virtually the same as 1998, 2002, 2003, 2006, and 2007, tying for second place. a la niña weather pattern caused 2008 to be cooler, but this pattern did not persist. the year 2009 was the warmest on record for the Southern Hemisphere. the first decade of the 21st century was the warmest since worldwide thermometer readings have been made. Moreover, north atlantic Ocean temperatures also show a pattern of recent warming. this is the sort of evidence that is required to show that global cli-mate has become warmer since the beginning of the industrial Revolution and particularly in the past half century—and the evidence has been found. the earth has not only become warmer, but abruptly warmer.
abrupt warming occurred between 1980 and 2000. although the first decade of the 21st cen-tury was the warmest on record, the warming trend did not make the end of the decade significantly warmer than its beginning. according to earth sci-entist Susan Solomon at nOaa, the reason for this might be water vapor in the stratosphere. Higher than normal atmospheric humidity from 1980 to 2000 enhanced global warming, while lower than normal humidity has repressed global warming since that time.
even though weather is not climate, it is possi-ble to assign a probability that global warming has contributed to particular weather events. climate scientists can use their computer models to cal-culate the probability that certain climatic events
would have happened without the extra carbon dioxide that has been released into the atmosphere since the beginning of the industrial Revolution. these models take into account natural factors, such as the movement of the earth and the el niño Southern Oscillation of ocean currents. this approach allows scientists to state that about half of the 2006 north american heat wave could be attributed to global warming and half to natural variability. it is therefore possible to say in retro-spect that global warming has contributed to the overall pattern of weather events.
a few scientists deny either that global temper-atures are increasing or that human activities are an important cause. they have not, however, veri-fied their claims in the scientific literature. their claims are largely limited to their own Web sites or to books published by organizations that have received funding from corporations that have a vested interest in the continued burning of fossil fuels. even though few scientists and no published data call global warming into question, journalists and the public have tended to perceive the claims of “global warming skeptics” as potentially valid. Science historian naomi Oreskes surveyed 928 sci-entific articles that dealt in some way with global climate change. none of these articles denied that global warming was occurring. However, when she surveyed 636 articles published in popular media, 53 percent of them gave credibility to the claims of those who deny global warming. apparently the public has been misled about the scientific consen-sus on global warming. Oreskes, along with atmo-spheric technologist erik conway, documented that the leading deniers of global warming are not climatologists but nuclear engineers who have specialized in attacking a whole range of scientific issues, including the link between smoking and cancer and the effects of pollution on the natural
environment.Proxy methods for tempera-
tures in recent centuries. But a century is not a very long time. the important question is not whether the climate is getting warmer, but whether this increase in global temperature is the result of human activity. if humans are partly or largely responsible for global warming, it must have occurred since the beginning of the industrial Revolution and accelerated as industrial pollution has spread throughout the world. if this is the case, the recent increase in temperature must be
During the past century and a half, global temperatures have increased. The increase has not been uniform; it was especially rapid after particulate air pollution was brought under control after about 1960. (Based on the National Climatic Data Center)
xviii+598_Biodiversity_F.indd 280 11/17/11 1:13 PM

greenhouse effect 281
greater than any that occurred in the past one or two millennia. therefore scientists have investigated tem-perature patterns farther into the past than is possi-ble with just the “instrumental” (thermometer) data. Scientists must use other methods to estimate tem-peratures before that time. Several of these “proxy” methods have been developed.
One example of a proxy method is the measure-ment of tree rings. each year, trees add another layer of wood just underneath the bark. the layer is visible as a ring because the wood produced in the spring is more porous, therefore lighter in color, than wood produced in late summer. the age of the tree can be accurately determined by counting the rings all the way to the center of the trunk. Dur-ing years with favorable weather conditions, trees produce a thick ring of wood; when the weather is poor, the ring is thinner. Good and bad weather result from temperature and moisture conditions. For trees in the north, good years are warm and bad years are cool; for trees in the south, good years are wet and bad years are dry. the barcode pattern of thick and thin layers of wood represents a record of good and bad years, extending back into the past, as long as the tree has been alive. Some trees, such as the bristlecone pines of california, have been alive and accumulating layers of wood for more than 4,000 years (see figure). Dendrochronologists, scientists who study tree rings, do not limit them-selves to the study of trees that are alive right now. if they find a well-preserved log, they can line up the barcode pattern of thick and thin rings in the log with those in a living tree, allowing them to deter-mine what year the tree died. they can then extend the tree ring record back to the year the dead tree had begun growing. By this method, forest scientist edward Schulman was able to determine that a cer-tain bristlecone pine log at the ancient Bristlecone Pine Reserve in california, now called the Schulman log, began growing about 1530 b.c.e. and died in 1676 c.e. at the age of 3,206 years. tree rings in very long-lived trees are more difficult to interpret than those in trees that grow faster and have shorter life spans. But the climate record from tree rings is fairly reliable for the past thousand years, especially when verified by other proxy methods such as the chemistry of the annual layers of sediment on conti-nental shelves.
the conclusion of these proxy methods is that the global climate today is hotter than it has been at any time in the past thousand years, and the current warming trend is faster than any warming trend dur-ing that period. Several different proxy reconstruc-tions of past temperatures have been published by scientists, and all are in close agreement with this major conclusion. the most famous of these graphs
is climate scientist Michael Mann’s “hockey stick model,” so called because a long period of relatively stable temperature (the hockey handle) has given way to a sudden warming as brief and abrupt as the end of a hockey stick. Other researchers have inde-pendently compiled similar data (see figure on page 282). Since its publication in 1998, Mann’s graph has withstood criticisms, one of which involves the so-called Medieval Warm Period and little ice age. Over the past millennium in europe, the aver-age temperature has fluctuated. temperatures were higher during the Medieval Warm Period in the 14th century and thereafter became cooler, produc-ing what has been named the little ice age. Some observers noted that Mann’s graph does not show these two events. it turns out, however, that these phenomena may have been limited to the northern Hemisphere, rather than global in extent. One rea-son for the fame of this graph is that it so angered a member of congress (Joe Barton, a Republican from texas) that he began an investigation into Mann’s competence, a move that was widely recognized as an attempt to suppress the evidence for global warm-ing. in 2006, a panel of the national academy of Sciences confirmed Mann’s data, although it noted that the estimates of temperatures of the past four centuries are more reliable than the estimates of the older dates.
the intergovernmental Panel on climate change (iPcc) is a group of scientists and government rep-resentatives from more than 100 nations, including the United States, brought together by the United nations environment Programme (UneP) and the World Meteorological Organization. their reports
Bristlecone pine trees (Pinus longeava) grow very slowly for thousands of years and produce very thin layers of wood. Each layer is about a hundredth of an inch (less than a third of a millimeter) wide. Some are thicker than others, however, and scientists use this information to reconstruct the history of climate change. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 281 11/17/11 1:13 PM
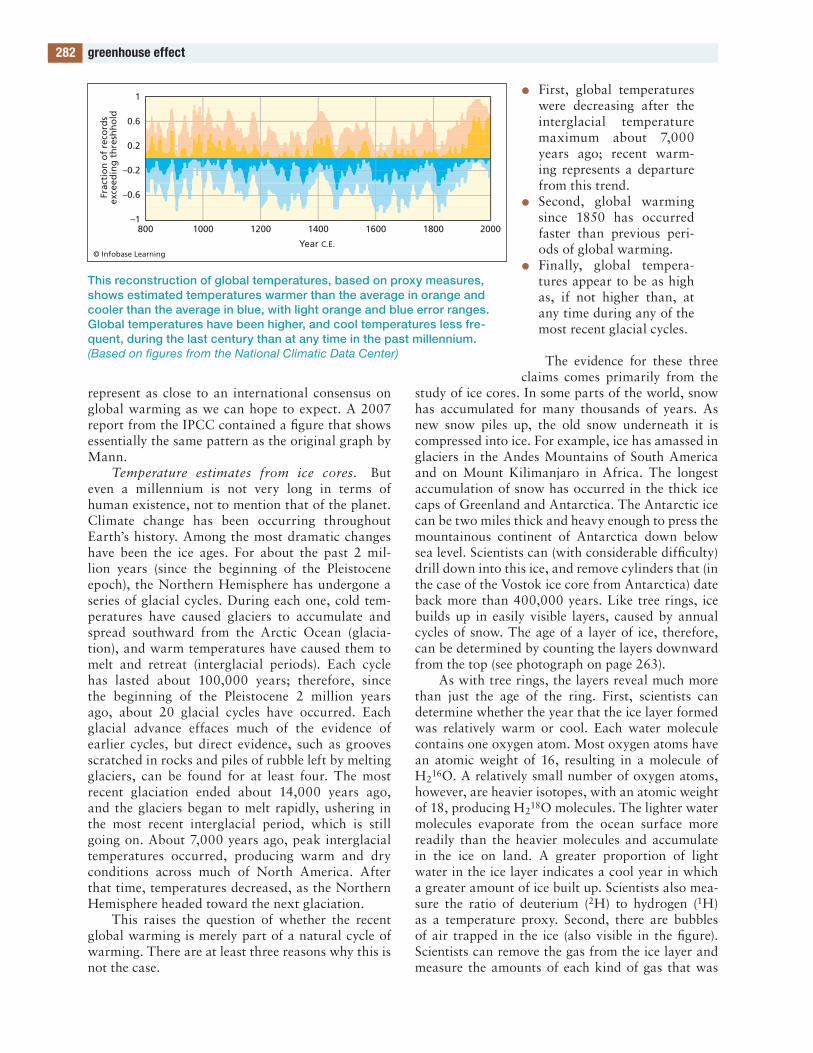
fi rst entry282 greenhouse effect 282
represent as close to an international consensus on global warming as we can hope to expect. a 2007 report from the iPcc contained a fi gure that shows essentially the same pattern as the original graph by Mann.
Temperature estimates from ice cores. But even a millennium is not very long in terms of human existence, not to mention that of the planet. climate change has been occurring throughout earth’s history. among the most dramatic changes have been the ice ages. For about the past 2 mil-lion years (since the beginning of the Pleistocene epoch), the northern Hemisphere has undergone a series of glacial cycles. During each one, cold tem-peratures have caused glaciers to accumulate and spread southward from the arctic Ocean (glacia-tion), and warm temperatures have caused them to melt and retreat (interglacial periods). each cycle has lasted about 100,000 years; therefore, since the beginning of the Pleistocene 2 million years ago, about 20 glacial cycles have occurred. each glacial advance effaces much of the evidence of earlier cycles, but direct evidence, such as grooves scratched in rocks and piles of rubble left by melting glaciers, can be found for at least four. the most recent glaciation ended about 14,000 years ago, and the glaciers began to melt rapidly, ushering in the most recent interglacial period, which is still going on. about 7,000 years ago, peak interglacial temperatures occurred, producing warm and dry conditions across much of north america. after that time, temperatures decreased, as the northern Hemisphere headed toward the next glaciation.
this raises the question of whether the recent global warming is merely part of a natural cycle of warming. there are at least three reasons why this is not the case.
● First, global temperatures were decreasing after the interglacial temperature maximum about 7,000 years ago; recent warm-ing represents a departure from this trend.
● Second, global warming since 1850 has occurred faster than previous peri-ods of global warming.
● Finally, global tempera-tures appear to be as high as, if not higher than, at any time during any of the most recent glacial cycles.
the evidence for these three claims comes primarily from the
study of ice cores. in some parts of the world, snow has accumulated for many thousands of years. as new snow piles up, the old snow underneath it is compressed into ice. For example, ice has amassed in glaciers in the andes Mountains of South america and on Mount Kilimanjaro in africa. the longest accumulation of snow has occurred in the thick ice caps of Greenland and antarctica. the antarctic ice can be two miles thick and heavy enough to press the mountainous continent of antarctica down below sea level. Scientists can (with considerable diffi culty) drill down into this ice, and remove cylinders that (in the case of the Vostok ice core from antarctica) date back more than 400,000 years. like tree rings, ice builds up in easily visible layers, caused by annual cycles of snow. the age of a layer of ice, therefore, can be determined by counting the layers downward from the top (see photograph on page 263).
as with tree rings, the layers reveal much more than just the age of the ring. First, scientists can determine whether the year that the ice layer formed was relatively warm or cool. each water molecule contains one oxygen atom. Most oxygen atoms have an atomic weight of 16, resulting in a molecule of H2
16O. a relatively small number of oxygen atoms, however, are heavier isotopes, with an atomic weight of 18, producing H2
18O molecules. the lighter water molecules evaporate from the ocean surface more readily than the heavier molecules and accumulate in the ice on land. a greater proportion of light water in the ice layer indicates a cool year in which a greater amount of ice built up. Scientists also mea-sure the ratio of deuterium (2H) to hydrogen (1H) as a temperature proxy. Second, there are bubbles of air trapped in the ice (also visible in the fi gure). Scientists can remove the gas from the ice layer and measure the amounts of each kind of gas that was
This reconstruction of global temperatures, based on proxy measures, shows estimated temperatures warmer than the average in orange and cooler than the average in blue, with light orange and blue error ranges. Global temperatures have been higher, and cool temperatures less fre-quent, during the last century than at any time in the past millennium. (Based on fi gures from the National Climatic Data Center)
xviii+598_Biodiversity_F.indd 282 11/17/11 1:13 PM

greenhouse effect 283
in the atmosphere at the time the layer was formed. the importance of this procedure will become obvi-ous a little later in this entry.
Using this information, scientists can recon-struct a record of global average temperatures reach-ing back as far in the past as the ice layers will allow. an analysis of the changes in global temperature derived from the Vostok ice core from antarctica clearly reveals the four most recent glacial cycles, each with glacial and interglacial periods (see fig-ure). Global temperatures have not yet increased beyond those of previous interglacial periods, but, as previous graphs have shown, that increase is now occurring rapidly. Global warming is nothing new in earth’s history; but the current rate of global warm-ing is 10 times as rapid as the warming that occurred at the end of the most recent glaciation.
there is other evidence that global temperatures have been increasing during the recent centuries and millennia. the most famous evidence is that,
all around the world, ice that has accumulated for thousands of years is beginning to melt. in antarc-tica, shelves of sea ice have been breaking off and forming icebergs. Because antarctica is the cold-est place on earth, and because it has so much ice, scientists had expected that global warming would melt antarctic ice slowly and gradually. Much to their surprise, they found that it is melting more rapidly than they could have imagined. the larsen B ice shelf in antarctica, the size of Rhode island, rather than gradually melting, collapsed over the course of two months in 2002, most of the cave-in occurring during two eventful days. a portion of the Wilkins ice shelf collapsed in March 2008. Meanwhile, in Greenland, sheets of land ice are melting and retreating. as in antarctica, Greenland ice is thick, and scientists expected it to melt slowly. they were surprised to discover in 2006 that water from glacial melt leaks down to the bottom of the glacier and lubricates it, allowing local ice masses
Scientists remove cylinders of ice from places such as Greenland and Antarctica, where snow accumulated for hundreds of thousands of years and was then compressed into ice. Each layer represents one year of accumu-lation. The chemistry of each layer indicates the global climate during that year. Air bubbles in the ice are actual air samples from that year. (American Museum of Natural History; Stanley A. Rice)
xviii+598_Biodiversity_F.indd 283 11/17/11 1:13 PM
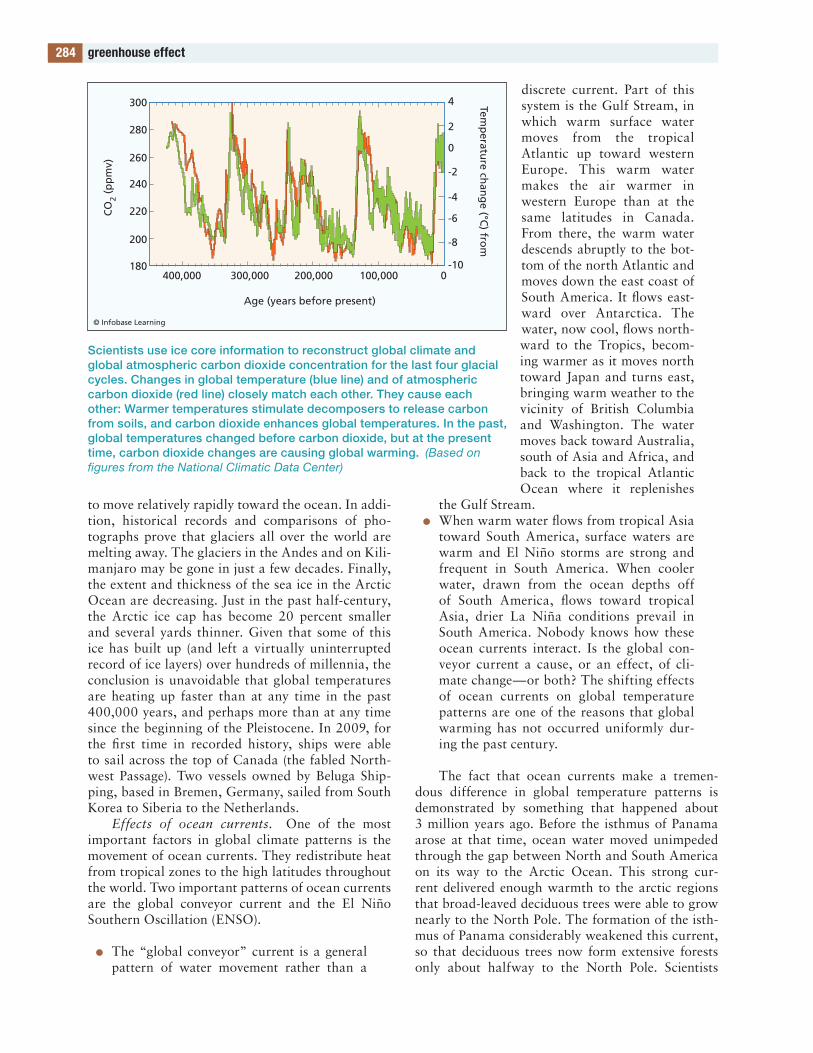
first entry284 greenhouse effect 284
to move relatively rapidly toward the ocean. in addi-tion, historical records and comparisons of pho-tographs prove that glaciers all over the world are melting away. the glaciers in the andes and on Kili-manjaro may be gone in just a few decades. Finally, the extent and thickness of the sea ice in the arctic Ocean are decreasing. Just in the past half-century, the arctic ice cap has become 20 percent smaller and several yards thinner. Given that some of this ice has built up (and left a virtually uninterrupted record of ice layers) over hundreds of millennia, the conclusion is unavoidable that global temperatures are heating up faster than at any time in the past 400,000 years, and perhaps more than at any time since the beginning of the Pleistocene. in 2009, for the first time in recorded history, ships were able to sail across the top of canada (the fabled north-west Passage). two vessels owned by Beluga Ship-ping, based in Bremen, Germany, sailed from South Korea to Siberia to the netherlands.
Effects of ocean currents. One of the most important factors in global climate patterns is the movement of ocean currents. they redistribute heat from tropical zones to the high latitudes throughout the world. two important patterns of ocean currents are the global conveyor current and the el niño Southern Oscillation (enSO).
● the “global conveyor” current is a general pattern of water movement rather than a
discrete current. Part of this system is the Gulf Stream, in which warm surface water moves from the tropical atlantic up toward western europe. this warm water makes the air warmer in western europe than at the same latitudes in canada. From there, the warm water descends abruptly to the bot-tom of the north atlantic and moves down the east coast of South america. it flows east-ward over antarctica. the water, now cool, flows north-ward to the tropics, becom-ing warmer as it moves north toward Japan and turns east, bringing warm weather to the vicinity of British columbia and Washington. the water moves back toward australia, south of asia and africa, and back to the tropical atlantic Ocean where it replenishes
the Gulf Stream. ● When warm water flows from tropical asia
toward South america, surface waters are warm and el niño storms are strong and frequent in South america. When cooler water, drawn from the ocean depths off of South america, flows toward tropical asia, drier la niña conditions prevail in South america. nobody knows how these ocean currents interact. is the global con-veyor current a cause, or an effect, of cli-mate change—or both? the shifting effects of ocean currents on global temperature patterns are one of the reasons that global warming has not occurred uniformly dur-ing the past century.
the fact that ocean currents make a tremen-dous difference in global temperature patterns is demonstrated by something that happened about 3 million years ago. Before the isthmus of Panama arose at that time, ocean water moved unimpeded through the gap between north and South america on its way to the arctic Ocean. this strong cur-rent delivered enough warmth to the arctic regions that broad-leaved deciduous trees were able to grow nearly to the north Pole. the formation of the isth-mus of Panama considerably weakened this current, so that deciduous trees now form extensive forests only about halfway to the north Pole. Scientists
Scientists use ice core information to reconstruct global climate and global atmospheric carbon dioxide concentration for the last four glacial cycles. Changes in global temperature (blue line) and of atmospheric carbon dioxide (red line) closely match each other. They cause each other: Warmer temperatures stimulate decomposers to release carbon from soils, and carbon dioxide enhances global temperatures. In the past, global temperatures changed before carbon dioxide, but at the present time, carbon dioxide changes are causing global warming. (Based on figures from the National Climatic Data Center)
xviii+598_Biodiversity_F.indd 284 11/17/11 1:13 PM

greenhouse effect 285
have concluded that this cannot be the entire reason that the north Pole was much warmer in the past than it is today, but the decrease in ocean currents was probably an important factor.
if anything slowed or stopped the Gulf Stream, the weather in northern europe could become much colder. this apparently happened about 12,800 years ago, right when the glaciers of the most recent ice age were melting and the earth was becoming warmer. Very quickly, the warming trend reversed, and temperatures as cold as those of the glacial period returned. this massive cold snap is called the “younger Dryas.” Dryas is the name of an arctic wildflower, whose range had retreated northward as the glaciers melted, but which quickly returned to europe and north america when the temperatures became frigid again. the cold snap was apparently caused by a flood of freshwater that entered the north atlantic. the collapse of a glacier released a huge amount of cold water through the St. law-rence Seaway into the north atlantic Ocean. the cold freshwater was lighter than the salt water of the ocean and remained near the surface of the north atlantic. then the warm Gulf Stream waters appar-ently sank long before they reached europe. europe, no longer receiving Gulf Stream warmth, experi-enced a rapid chilling. nobody knows what caused this sudden glacial melt.
could the north atlantic again be flooded with cold freshwater? the melting of Greenland ice could cause something like this to occur. therefore the effects of global warming may, paradoxically, include a temporary, sudden cooling in europe. this is, however, one of the predictions that has the least support among climate scientists.
Sunspots also appear to affect global temper-ature. For example, there was very little sunspot activity during the little ice age. as is the case with ocean currents, variations in sunspot activity appear to superimpose global temperature variations upon an overall warming trend.
EffECTS of GloBal warmING oN ThE EarThit is clear that the earth is becoming warmer. How much warmer will it become in the future? in order to predict future temperatures, scientists run complex “general circulation models” on some of the world’s most powerful computers. these models make, and integrate, predictions for each of many grid squares into which the earth’s surface is divided. Different computer models are used at the Goddard institute for Space Studies in new york, lawrence livermore laboratories in california, the Hadley centre at the University of east anglia in england, and the Max Planck institute in Germany. although they are all different models, they yield very similar results. and
there has been at least one opportunity to test the models for reliability. When the volcano Pinatubo erupted in the Philippines in 1991, it spewed par-ticulate matter into the upper atmosphere. Using the Goddard institute program, naSa climatolo-gist James Hansen predicted that global temperature would decrease by about a half a degree F as a result. His prediction turned out to be very accurate.
Global warming is raising the sea level. During the most recent glaciation, the sea level was almost 300 feet (100 m) lower than it is today, because enor-mous amounts of water were locked up in glaciers. then the glaciers melted and the ocean levels rose. Scientific measurements indicate that ocean levels are rising by about a quarter of an inch per year. the sea level appears to be rising at a faster rate than in the past, as a result of global warming. there are two reasons for this. First, when sea ice melts, the additional water does not alter the ocean level. But when global warming causes ice on land to melt, the water drains into the ocean and raises its level. Water from melting glaciers, especially in antarctica and Greenland, is already causing the sea level to rise. if all of the ice melted, which may occur in a few centu-ries, the ocean levels would rise several feet. Second, as global temperature increases, the oceans become warmer, causing the water to expand.
if the Greenland ice cap melted or just frag-mented and fell into the ocean as icebergs, the ocean level would rise by almost 20 feet (about six m). the West antarctic ice sheet, if it slid into the ocean, would cause the oceans to rise by a similar amount. (the east antarctic ice sheet is much larger but is more stable and unlikely to collapse.) this rise would be enough to flood the major coastal cities of the world, such as new york, Boston, Washing-ton, los angeles, San Francisco, Seattle, Shanghai, tokyo, Bangkok, Bombay, london, and Marseilles. the ocean levels would rise slowly enough that peo-ple would have time to move, but the expense of moving and rebuilding—or, alternatively, of build-ing sea walls—is nearly incalculable. the map of the world would be redrawn, including a Pacific Ocean in which several island nations have vanished and a United States with most of Florida missing. the sud-den influx of glacial meltwater into the north atlan-tic at the start of the younger Dryas raised ocean levels by more than 30 feet.
Global warming is causing more rainfall in coastal areas and droughts inland. Warm air can hold more moisture than cool air. as a result, when temperatures rise, more water evaporates from the oceans, most of which falls in coastal areas. the average amount of rainfall in the world has already increased by 20 percent in coastal areas. But warm air can also cause more water to evaporate from
xviii+598_Biodiversity_F.indd 285 11/17/11 1:13 PM

first entry286 greenhouse effect 286
plants and soil. therefore, away from the oceans, in continental areas, rainfall could decrease. these continental areas include most of the agricultural region of the United States, where droughts are becoming more frequent. a study released in 2007 predicts that in the american Southwest droughts will become the norm rather than the exception. this region depends on water from snowmelt, and if snowfall decreases, large areas may experience droughts severe enough that crops cannot be irri-gated. Meanwhile in the middle of africa, droughts may become more frequent and intense. Moreover, the year 2010 was the ninth year of one of austra-lia’s worst droughts. this drought was followed, in 2011, by devastating floods. the drier conditions will undoubtedly cause more frequent, and more severe, wildfires. Fires burned large areas of forest worldwide in the summer of 2005, not only in the tropics, but in alaska as well.
another way in which global warming can con-tribute to continental drought is by causing glaciers to disappear. Millions of people depend on glacial meltwater in mountainous areas of South america and asia. Glaciers are melting in the andes and on Kilimanjaro.
Global warming is causing stronger storms. even though 2005 had more hurricanes in the atlan-tic and more typhoons and cyclones in the Pacific than any previously recorded year, the historical record of storms indicates that the frequency of hur-ricanes is not increasing. every few decades, a period of either more or fewer hurricanes begins. However, recent hurricanes and other storms have been more severe than those in the past. in india, for example, there has been an increase in heavy storms, but not in the total number of storms, over the past half century. the intensity of storms is influenced by the warmth of the surface waters in the north atlantic Ocean; warmer surface waters cause more evapora-tion and stronger wind, both of which contribute to creating intense storms. Hurricanes Katrina (in 2005) and Dean (in 2007) may therefore provide a very realistic glimpse into the kind of damage that will become commonplace in future decades, as global warming continues. in a warmer world, strong storms can also form in places where they have not previously occurred. the first recorded South atlantic hurricane occurred in 2004.
all of these processes can occur simultaneously. the last week of July 2010 saw not only the most severe flooding in Pakistan’s history but also a pro-longed heat wave and drought in Russia that resulted in the worst forest fires in a century.
Inevitable surprises. With more sunlight energy being trapped in the climate systems of the earth, the result will almost certainly not be a uni-
form warming of the planet. instead, scientists expect that climate fluctuations will become more extreme. in effect, global warming is stirring the climate pot more vigorously. the exact results will be impossible to predict. in summer 2007, for example, arctic sea ice melted at a much greater rate than had been pre-dicted. the summer arctic ice minimum had been declining at 8.6 percent per decade until 2005, then it declined almost 20 percent in 2007. this rapid rate of melting has not continued.
One reason that the detailed patterns of climate change are unpredictable is that they occur by means of “tipping points” or “thresholds.” a complex sys-tem may be in a relatively stable state for long peri-ods of time, then suddenly change to another stable state when a threshold is reached. Historical and sociological examples abound, such as revolutions and the Reformation. engineering examples are also well known, in which a building or bridge suddenly collapses. everyone knows about the proverbial “straw that broke the camel’s back.” Many scien-tific processes occur by means of threshold events. Rocks move along geological faults, not gradually, but suddenly: a major earthquake may occur after a buildup of pressure crosses a threshold. chemical reactions may not occur at all until threshold condi-tions are reached.
climate change may occur slowly until some threshold incident catalyzes a cascade of events that feed on one another. Here are some examples:
● the arctic ice cap reflects a great deal of sunlight back into outer space; this reflected light, called albedo, does not cause the earth to become warmer. But as the arctic ice melts, the ocean that the ice formerly cov-ered now absorbs more sunlight and retains the heat. Global warming causes the ice to melt, but the melting of the ice also causes more global warming.
● Decomposition of organic matter in peat bogs and soils around the world, especially in the far north, releases greenhouse gases such as carbon dioxide and methane (see next section) into the atmosphere. Huge areas of Siberia, including a continental shelf that today is under the sea but was exposed land surface during the most recent ice age, are producing huge amounts of methane. Some of these organic deposits are more than 150 feet (about 50 m) thick. Global warming causes more decomposition, and decomposition causes more global warm-ing in the cold peat bogs; some bog lakes in Siberia no longer freeze in the winter because of the warmth created by decom-
xviii+598_Biodiversity_F.indd 286 11/17/11 1:13 PM

greenhouse effect 287
position. in tropical areas such as indone-sia, global warming dries out peat bogs and encourages them to catch fire.
● Global warming also causes drier conditions in the interiors of continents. these drier conditions allow more wildfires to occur—which is what happened in thousands of square miles of tropical forests in 2005. the fires put greenhouse gases into the air and, at least temporarily, destroy the forests that could remove the greenhouse gases from the air. the exposed soils become warmer, which causes them to release more carbon dioxide.
therefore, greenhouse gases cause global warm-ing that causes the release of more greenhouse gases, resulting in more global warming.
Because of the complex interactions in the natu-ral world, the effects of global warming on wild species are difficult to predict. Humans may notice most of all the impact of global warming on the spread of organisms, such as mosquitoes, that carry diseases. in 2007, the World Health Organization announced that global warming threatened the livelihoods and health of millions of people, par-ticularly in those countries that put the least carbon dioxide into the air.
therefore even the most sophisticated computer models of what will happen in the future can be erro-neous. leading climate scientists announced in 2007 that this is already proving to be the case. in an ear-lier 2001 report, the iPcc predicted future increases in atmospheric carbon dioxide, global temperature, and sea level. With each prediction, the iPcc sci-entists specified an error range, which means the increases might be less, or might be greater, than they had predicted. Since 2001, all three factors have been increasing even more rapidly than the iPcc predicted—and the increases have been right at the very top of the range of predicted possibilities. the climate predictions, as frightening as they have been, have already proven to be underestimates.
aTmoSphErIC CarBoN dIoxIdE IS a CauSE of GloBal warmING
the greenhouse effect makes the earth warmer than it would otherwise be. in a greenhouse, sun-light shines in through the glass roof and heats up the plants and benches, which conduct heat to the air. the glass roof holds in a lot of the heat from this warm air and therefore acts as a largely one-way portal for heat to enter the greenhouse. Green-house gases (GHG) perform an analogous service for the earth’s atmosphere. light shines through the atmosphere and warms the earth’s surface, which
then emits infrared radiation. infrared radiation is low-energy wavelengths of light that are invisible to humans. the earth’s atmosphere consists mostly of nitrogen and oxygen gases. in an atmosphere that consisted only of nitrogen and oxygen gases, all of the infrared radiation would be lost into outer space. GHG molecules, however, absorb the infrared radia-tion, which makes them warm. they conduct some of their energy with the other gases in the atmo-sphere, making the whole atmosphere warmer. the more GHG molecules there are in the atmosphere, the warmer that atmosphere and the planet will be.
carbon dioxide (cO2) is the most famous green-house gas. the atmosphere contains only about 380 parts per million (ppm), or 0.038 percent, cO2, by volume, fewer than four molecules out of every 10,000 in the atmosphere. But without this cO2, the earth would be uninhabitably cold. there are other GHG in the atmosphere, including water vapor and methane (the major component of natural gas). Methane is about a thousand times less common than cO2 in the atmosphere but at least 20 times as potent at retaining heat. However, of these three gases, only cO2 can accumulate indefinitely. Water vapor in the lower atmosphere can build up until it condenses as clouds and falls as rain. Methane reacts with oxygen gas, producing cO2. Other GHG, such as nitrogen oxides and ozone, are even more potent and (in the case of ozone) persistent, but are rare compared to cO2. even though methane is rare and persists only about a decade in the atmosphere, scientists estimate that it is responsible for about 40 percent of the global temperature increase. nitrous oxide is also a greenhouse gas. although it is nor-mally rare in the atmosphere, its concentration has been increasing because of china’s overuse of nitro-gen fertilizer.
the greenhouse effect is, to a certain extent, a good thing. Without it, the earth would be about as cold as Mars now is. Mars itself has an atmosphere consisting mostly of cO2, but its greenhouse effect is minor because its atmosphere is so thin. the atmo-sphere of Venus contains huge amounts of cO2, which is the reason that Venus is hotter than Mer-cury even though it is farther from the Sun.
But it is possible to have too much of a good thing. too much cO2 can cause the earth to become too warm for the life-forms that are now living on it. this is a major reason that scientists view the increasing concentration of GHG in earth’s atmo-sphere with some alarm.
Atmospheric carbon dioxide levels are increasing. the longest-running measurement of atmospheric cO2 began in 1958 at the U.S. national Oceanic and atmospheric administration (nOaa) observatory near the top of Mauna loa. the top of a volcanic
xviii+598_Biodiversity_F.indd 287 11/17/11 1:13 PM
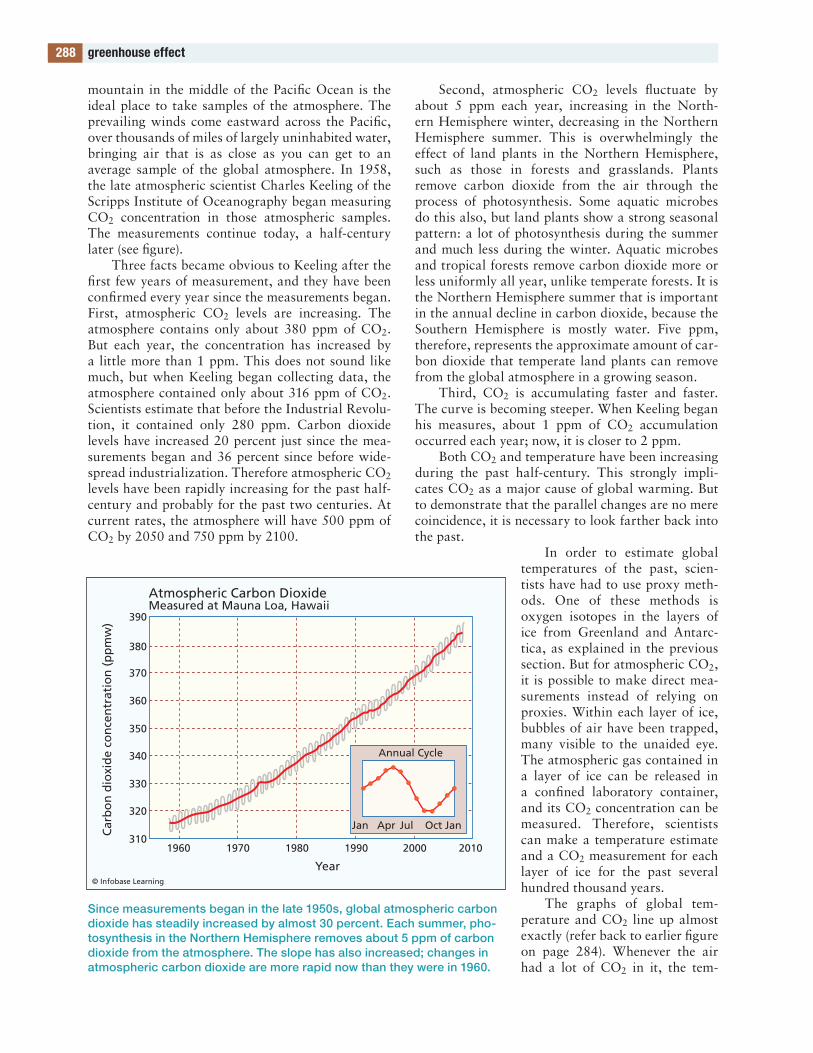
first entry288 greenhouse effect 288
mountain in the middle of the Pacific Ocean is the ideal place to take samples of the atmosphere. the prevailing winds come eastward across the Pacific, over thousands of miles of largely uninhabited water, bringing air that is as close as you can get to an average sample of the global atmosphere. in 1958, the late atmospheric scientist charles Keeling of the Scripps institute of Oceanography began measuring cO2 concentration in those atmospheric samples. the measurements continue today, a half-century later (see figure).
three facts became obvious to Keeling after the first few years of measurement, and they have been confirmed every year since the measurements began. First, atmospheric cO2 levels are increasing. the atmosphere contains only about 380 ppm of cO2. But each year, the concentration has increased by a little more than 1 ppm. this does not sound like much, but when Keeling began collecting data, the atmosphere contained only about 316 ppm of cO2. Scientists estimate that before the industrial Revolu-tion, it contained only 280 ppm. carbon dioxide levels have increased 20 percent just since the mea-surements began and 36 percent since before wide-spread industrialization. therefore atmospheric cO2 levels have been rapidly increasing for the past half-century and probably for the past two centuries. at current rates, the atmosphere will have 500 ppm of cO2 by 2050 and 750 ppm by 2100.
Second, atmospheric cO2 levels fluctuate by about 5 ppm each year, increasing in the north-ern Hemisphere winter, decreasing in the northern Hemisphere summer. this is overwhelmingly the effect of land plants in the northern Hemisphere, such as those in forests and grasslands. Plants remove carbon dioxide from the air through the process of photosynthesis. Some aquatic microbes do this also, but land plants show a strong seasonal pattern: a lot of photosynthesis during the summer and much less during the winter. aquatic microbes and tropical forests remove carbon dioxide more or less uniformly all year, unlike temperate forests. it is the northern Hemisphere summer that is important in the annual decline in carbon dioxide, because the Southern Hemisphere is mostly water. Five ppm, therefore, represents the approximate amount of car-bon dioxide that temperate land plants can remove from the global atmosphere in a growing season.
third, cO2 is accumulating faster and faster. the curve is becoming steeper. When Keeling began his measures, about 1 ppm of cO2 accumulation occurred each year; now, it is closer to 2 ppm.
Both cO2 and temperature have been increasing during the past half-century. this strongly impli-cates cO2 as a major cause of global warming. But to demonstrate that the parallel changes are no mere coincidence, it is necessary to look farther back into the past.
in order to estimate global temperatures of the past, scien-tists have had to use proxy meth-ods. One of these methods is oxygen isotopes in the layers of ice from Greenland and antarc-tica, as explained in the previous section. But for atmospheric cO2, it is possible to make direct mea-surements instead of relying on proxies. Within each layer of ice, bubbles of air have been trapped, many visible to the unaided eye. the atmospheric gas contained in a layer of ice can be released in a confined laboratory container, and its cO2 concentration can be measured. therefore, scientists can make a temperature estimate and a cO2 measurement for each layer of ice for the past several hundred thousand years.
the graphs of global tem-perature and cO2 line up almost exactly (refer back to earlier figure on page 284). Whenever the air had a lot of cO2 in it, the tem-
Since measurements began in the late 1950s, global atmospheric carbon dioxide has steadily increased by almost 30 percent. Each summer, pho-tosynthesis in the Northern Hemisphere removes about 5 ppm of carbon dioxide from the atmosphere. The slope has also increased; changes in atmospheric carbon dioxide are more rapid now than they were in 1960.
xviii+598_Biodiversity_F.indd 288 11/17/11 1:13 PM

greenhouse effect 289
perature was greater than at times when there was less cO2 in the atmosphere. this is a striking and convincing proof of the correlation of carbon diox-ide with global warming.
correlation, however, is not the same as cau-sation. not only could an increase in cO2 cause global warming, but global warming could cause an increase in cO2. Warmer temperatures stimulate decomposition by bacteria and fungi in the soils of the world; decomposition releases cO2 into the air. increases in cO2 and global warming therefore cause each other. this accounts for the correlation. in the past, ice ages came and went, largely because processes such as the rotation and revolution of the earth initiated cycles of warming and cooling. a warming trend would stimulate the release of cO2, which would accentuate the warming trend. Before civilization, global temperatures appear to have increased prior to the increase in cO2. today, however, it is the other way around. atmospheric cO2 has increased to its highest level in almost a half million years, and global temperatures have yet to catch up. For this reason, even if humans stopped putting cO2 into the atmosphere right now, global warming would still occur. the difference between what happened before civilization, in which global warming caused increases in cO2, and what is hap-pening today, in which cO2 is causing global warm-ing, is that billions of humans are producing vast amounts of cO2.
Increased carbon dioxide levels are largely responsible for global warming. cO2 is not the only factor that may contribute to global warming. as mentioned previously, natural climatic variabil-ity can be caused by the movement of the earth and changes in the intensity of sunlight. the advance and retreat of glaciers across north america and europe appear to be caused by changes in the movements of the earth relative to the Sun, which produce an approximately 100,000-year cycle of cold (glaciation) and warm (interglacial) periods. However, the changes in solar intensity are very small. By themselves, they seem unable to explain the ice ages. apparently these variations in solar intensity have stimulated a series of processes that, working together, have produced ice ages during the past 2 million years. although the ice ages have followed the pattern imposed by cycles of solar illu-mination, recent global warming cannot be attrib-uted to it.
Other human activities may counteract global warming but at a cost. the destruction of forests, and their replacement by barren land reflect more sunlight back into outer space but also reduce pho-tosynthesis. Particulate air pollution (such as smoke and sulfur dioxide) reflects sunlight back into outer
space but creates environmental and health prob-lems. in fact, the reduction of particulate air pol-lution in recent decades, allowing more sunlight to reach the surface of the earth, may be the reason that global temperatures have risen sharply in recent decades, even though cO2 has been steadily accu-mulating for a century and a half. none of these other processes has anywhere near the effect that carbon dioxide has had on recent global warming.
Human activities have increased the amount of energy at the earth’s surface by 1.9 watts per square yard (1.6 watts per square meter). this is the equiva-lent of having an additional heat load of 200 30-watt lightbulbs burning continuously on each acre of the earth’s land and ocean surface. this is approxi-mately equal to the amount of heating that is attrib-utable to the buildup of cO2 in the air.
carbon dioxide and other GHG have also been a major influence in the earth’s long-term history, during the past 2 million years, during the climate change that occurred about 30 million years ago, and during the sudden warming that occurred about 55 million years ago. at only one time, about 300 million years ago, has a period of ice ages given way to a prolonged warm period, and this event was apparently caused by an increase in atmospheric carbon dioxide.
whaT IS CauSING ThE INCrEaSE IN CarBoN dIoxIdE?
although in previous millennia the changes in tem-perature and cO2 were related to the ice ages, the present-day increase in cO2 is undoubtedly the result of human activities. the main sources of cO2 emissions are from transportation and manu-facturing. the burning of fuels such as coal, oil, and natural gas for transportation and industry releases huge amounts of cO2 into the air. Since the beginning of the industrial Revolution, humans have burned more and more of these fossil fuels (so-called because they were formed millions of years ago primarily from dead plants and microbes) each year. Of these three fuels, coal (often used in power plants and factories) emits the most cO2 per unit of energy produced, because it already contains so much carbon. Oil, including oil refined into gaso-line, also produces a lot of cO2 from the tailpipes of the billion cars and trucks in the world. another large source of carbon emissions is the burning of firewood. Firewood produces about 30 percent of the cO2 that enters the atmosphere. this is the principal or only source of energy for billions of poor people.
another reason for the prodigious increase in carbon emissions is that humans are destroying plants. the destruction of forests and other plant
xviii+598_Biodiversity_F.indd 289 11/17/11 1:13 PM

first entry290 greenhouse effect 290
habitats puts cO2 into the air in two major ways. First, when the trees are burned or decompose, they release cO2 into the air. Second, dead plants can no longer carry out the photosynthetic process that removes cO2 from the air. in many instances, the natural habitats do not recover. in such cases, the loss of plant cover represents a permanent release of carbon dioxide and reduction in the ability of plants to cleanse the air of excess cO2.
Humans are not only destroying plants, but causing cO2 to be released from dead plant mate-rials. conversion of forests to agricultural lands causes the soil to become warmer, which enhances the decomposition of humus—while at the same time reducing the amount of new humus that is cre-ated. about 2 billion tons (metric tons) of carbon have entered the air as a result of draining and burn-ing peatlands in indonesia for oil palm plantations. (this also caused fruit bats to roost outside of the forest, in closer contact with humans. their feces contain viruses that could infect people.)
the sheer volume of these processes—all of them caused by humans—accounts for almost all of the increase in atmospheric cO2. it is ludicrous to assert that scientists cannot know why its level is rising. One U.S. Republican congressman, Dana Rohrabacher of california, speculated in 2007 that a global warming event 55 million years ago was caused by unknown and unknowable processes—his guess was that it might have been dinosaur flatu-lence. this suggestion is as irrelevant as it is ridicu-lous. even if the causes of global warming in the past were not fairly well understood, the cause today is not in doubt. nearly all scientists agree that human activities are the main cause of the global warming that is now occurring.
the agreement of scientists is perhaps best indicated by the strong conclusions issued by the iPcc. in February 2007, the iPcc stated that global warming from carbon dioxide was a certainty and that the human contribution to this warming was 90 percent certain. in april 2007, the panel released a second report that predicted droughts in some areas, flooding in others, and the spread of disease and insect pests as consequences of global warming. in May 2007, it released a third report that proposed possible solutions. in november 2007, it produced a synthesis report that summarized all of the organi-zation’s conclusions. When this international panel of scientists issues a consensus statement, it is as close to a scientific fact as we can get about climate change. the panel’s statements are reviewed by rep-resentatives of many governments and therefore rep-resent something as close to world consensus as can be reasonably expected. the iPcc is now assem-bling data for their 2013 report.
atmospheric cO2 levels have been much higher in the distant past than they are now. and global warming is nothing new on this planet. Before 25 million years ago, antarctica was fully forested. then cO2 fell below 500 ppm and ice sheets began to expand in antarctica. no competent scientist claims that global warming or the accumulation of atmospheric cO2 will prove directly lethal to life on this planet. in fact, the earth was greener when there was more carbon dioxide in the air millions of years ago. But the consequences of global warming, as outlined above, may be disastrous to the habitats and ecological relationships of plants and animals, as well as to the human economy and to global politics. never before has the earth had to support 7 billion humans. the entire system of agriculture and commerce is based on the assumption that global climate patterns and ocean levels will remain as they are except for occasional droughts or storms. this comfortable belief will almost certainly be shaken. it will not be necessary for global warming to make humans fall over dead in order to wreak havoc on global civilization. Human activity is pushing tem-peratures and cO2 levels outside of the range that they have occupied for the past 2 million to 3 million years, at which time continents and global ocean circulation assumed their present patterns. there is thus no comparable situation in the past to which scientists can look that will allow them to predict what will happen next. the economic consequences, from higher sea levels, from storms, from disease, and from agricultural losses, are unpredictable and will probably be massive.
CaN ThE GrEENhouSE EffECT BE CoNTrollEd?For the future security of the human and natural world, it is essential that the greenhouse effect be brought under control. Some scientists have made wildly speculative proposals to do this by reflecting sunlight back into outer space by launching giant mirrors into orbit or by spewing massive amounts of sulfate particles into the upper atmosphere. Most scientists are convinced that the greenhouse effect can be reduced only by managing the amount of GHG in the atmosphere. the two general ways to do this are to release fewer GHG into the atmosphere and to remove them from the atmosphere. Some scientists have proposed to reduce carbon emis-sions through “carbon sequestration,” or chemical removal of carbon dioxide from effluent gases before they enter the air, for example by removing cO2 from the smokestacks of power plants. the technol-ogy to do this on any meaningful scale is currently experimental and expensive. a few other scientists have championed the chemical removal of carbon dioxide from the air. this method would be even
xviii+598_Biodiversity_F.indd 290 11/17/11 1:13 PM

greenhouse effect 291
more uncertain and expensive. thus most scientists say that humans should produce less cO2 in the first place and remove it from the air by enhancing natu-ral processes such as plant photosynthesis.
Photosynthesis is not the only natural process that removes cO2 from the air. Over a very long period of time, the weathering of some kinds of rocks removes cO2 from the air. carbon dioxide reacts with silicates in the earth’s crust, forming carbonates (limestone) that are washed away and accumulate as sediments on the ocean floor. the cycle is completed when continental drift thrusts the sediments into the crust and the carbon is released as cO2 from volcanoes. this process acts as a regula-tory mechanism, because a greater abundance of atmospheric cO2 produces a warmer climate that stimulates the reaction between cO2 and silicate. this process cycles a much greater amount of carbon through the earth than does photosynthesis. How-ever, it requires millions of years and cannot rescue the earth from an excessive greenhouse effect on any timescale meaningful to humans. Photosynthesis is the only natural process that can quickly remove cO2 from the air. if humans promote the growth of plants, they may not only stimulate photosynthesis but even the formation of limestone: Plants speed up the process of silicate weathering by acidifying the soil with their leaf litter.
at least one “think tank” of american antienvi-ronmentalists, the competitive enterprise institute, has claimed that photosynthesis is the reason that humans do not have to worry about global warming. Humans can pour all the carbon dioxide they want into the air, and plants will clean up the mess. their advertisements asserted: “carbon dioxide—we call it life!” Unfortunately, this argument is lethally sim-plistic. Perhaps the most obvious fact that contra-dicts this hypothesis is that plants are not, right now, absorbing the excess cO2 in the atmosphere. also, at the very time that they are most needed, humans are destroying plants. Furthermore, plants carry out photosynthesis only if they have enough nutrients and water. if large continental areas expe-rience droughts, the trees will not remove very much carbon from the air. Researchers at Duke University performed a free-air carbon enhancement (Face) study in a forested area of north carolina. in Face studies, the air is enriched in cO2, but the plants are not enclosed by any kind of barrier. the researchers increased the cO2 in forest plots by 50 percent over the course of an entire decade. they announced in the summer of 2007 that the extra cO2 had aided tree growth only in those areas that had plenty of water and good soil. Other studies have found that global warming might enhance soil and plant res-piration, which releases carbon dioxide, even more
than it stimulates photosynthesis, which removes it. Finally, there are just not enough plants in the world to counteract all of the carbon that human activities are putting into the air.
the world not only needs more plant cover but plants that remove carbon from the air per-manently. as stated earlier, each northern summer, plants remove about 5 ppm of carbon dioxide from the air. Most of this is returned, however, through decomposition of leaf litter. this is particularly true of short-lived plants such as crops. Researchers at the University of illinois used Face experiments to enhance the growth of soybeans and observed only modest increases, far less than the increases that had been predicted from laboratory experiments. Other recent experiments have shown that extra cO2 in the air may boost the growth of forests, but that these results may be transient—forests do not con-tinue to show year after year of additional growth in the presence of elevated carbon dioxide. if, in contrast, forests are allowed to store carbon in the form of wood, the carbon has been removed from the air for the life of the tree. the trees that remove the most carbon from the air during their lives are also the ones that grow slowly. a huge california redwood may remove 30 tons (about 30 metric tons) of carbon from the air—but may require two millen-nia to do so. Preserving forests and planting more trees is a necessary but not entirely sufficient part of removing excess cO2 from the atmosphere.
another way in which plants may be able to remove carbon dioxide from the atmosphere for a long period is by the growth of roots. When dead roots decompose, much of the carbon goes into humus rather than directly into the atmosphere. increased plant growth will store more carbon in the soil. Humus in soils all around the world contains a massive amount of carbon. the cold, soggy soils of Siberia alone may hold 70 billion tons of carbon. However, global warming is causing soils every-where to release cO2, which will enhance global warming still further.
no matter how many forests and grasslands humans may plant, they will not remove all of the additional carbon that is entering the atmosphere. Plant ecologist christopher Field and colleagues esti-mated that the earth’s natural ecosystems produce 117 billion tons of biomass per year. this biomass either consists of carbohydrates or is derived from carbohydrates, which are produced by photosynthe-sis. Most of the mass of these carbohydrates comes from cO2. the burning of fossil fuels produces about 7 billion tons of carbon, which corresponds to about 25 billion tons of carbon dioxide, each year. (car-bon dioxide consists of one carbon and two oxygen atoms; mass of cO2 is 3.7 times the mass of carbon.)
xviii+598_Biodiversity_F.indd 291 11/17/11 1:13 PM

first entry292 greenhouse effect 292
if plants and aquatic microbes were to absorb the entire 25 billion tons, it would be necessary to increase the growth of plants worldwide by about 21 percent relative to the plant cover that existed before civilization. While this will probably never happen while the human economy is as large as it now is, a 21 percent increase in forest cover is not impossible.
not all plant cover would be equally helpful in reducing the greenhouse effect. Boreal forests (of the far north) absorb much more sunlight than does snow cover. this causes the trees to become warm and release heat into the surrounding air, which would not happen with sun shining on snow. One scientist pointed out that an expansion of boreal forests at the expense of tundra, which is covered by snow except for a couple of months each year, would therefore result in a net warming of the planet. this point is true but may be moot, because with global warming, the snow cover of the tundra will be severely reduced anyway, and scientists predict that the boreal forest will move northward whether trees are planted or not. Moreover, it will do no good to plant trees under conditions in which they cannot survive. Millions of trees have been planted in arid, dusty regions of china, but many of these have died. the restoration of grasslands would have been a more effective way of stabilizing the soil and of removing carbon from the air. Grasses have massive root systems and are particularly good at putting carbon into the soil.
the oceans also absorb cO2. Many aquatic microbes carry out photosynthesis just as land plants do. Many small aquatic organisms, and some
that are not so small, produce shells of calcium car-bonate. When these organisms die, their shells, and the carbon they contain, settle to the bottom of the sea, forming organic limestone. in addition, ocean water itself absorbs cO2, producing carbonic acid. currently, by these processes, the oceans remove about 2 billion tons of carbon from the atmo-sphere—about one-fourth of what is added to the atmosphere—each year. However, absorption of carbon by the oceans may not continue indefinitely at its current rate. carbonic acid in the oceans may reduce the growth of the organisms that now make organic limestone, and warmer oceans will absorb less carbon dioxide. arctic Ocean waters near canada were measurably depleted of calcium carbonate in 2008.
natural processes (photosynthesis on land and in oceans, absorption by ocean water, reaction with silicates) cannot compensate for the amount of cO2 humans are adding to the atmosphere. this simple conclusion is obvious when we consider that they are not now doing so; atmospheric cO2 increases measurably every year. about 45 percent of the cO2 added to the atmosphere stays there.
Because about half of the cO2 that humans put into the atmosphere stays there during any given year, there is no way around imposing limits on carbon emissions and increasing the efficiency of energy use in power plants and vehicles. timed to coincide with the start of the United nations cli-mate Summit in copenhagen on December 7, 2009, the environmental Protection agency announced that it would regulate cO2 as a substance that is dangerous to human health. On that same day, con-gressional Republicans issued a statement criticiz-ing President Barack Obama for generating carbon emissions (which, they claimed, were not respon-sible for global warming anyway) by flying in Air Force One to Oslo to accept the nobel Peace Prize. caps on carbon emissions appear to be both neces-sary and impossible in the United States, to which the rest of the world is looking for leadership.
each person leaves an “ecological footprint” on the world. this refers to the acreage of produc-tive land and water (such as forests and agricultural fields) that would be required to remove the carbon dioxide from the air that a person is responsible for releasing and to provide the raw materials for that person’s activities on a continuing basis. this includes the absorption of the cO2 not only from what each person directly releases, such as from an automobile tailpipe, but the cO2 released by power plants and factories that provide the person with what he or she consumes. the average american has an ecological footprint of about 23 acres (10 hect-
Comparisons of the eCologiCal footprints (per person) of
seleCted Countries*
Country hectares**
United States 10
Canada 8
United Kingdom 6
Japan 4
Germany 2
China 2
World 2
India 1
Kenya 1
*Rounded to the nearest hectare. From Global Footprint Network**1 hectare = 2.5 acres
xviii+598_Biodiversity_F.indd 292 11/17/11 1:13 PM
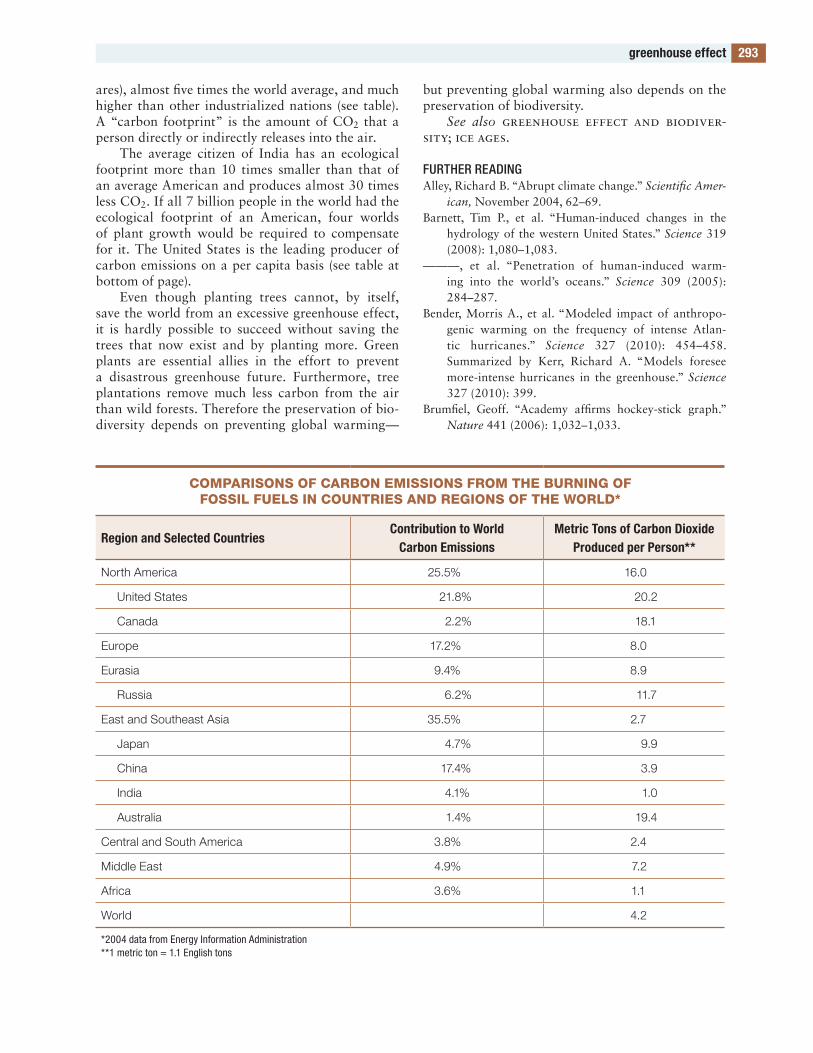
greenhouse effect 293
ares), almost five times the world average, and much higher than other industrialized nations (see table). a “carbon footprint” is the amount of cO2 that a person directly or indirectly releases into the air.
the average citizen of india has an ecological footprint more than 10 times smaller than that of an average american and produces almost 30 times less cO2. if all 7 billion people in the world had the ecological footprint of an american, four worlds of plant growth would be required to compensate for it. the United States is the leading producer of carbon emissions on a per capita basis (see table at bottom of page).
even though planting trees cannot, by itself, save the world from an excessive greenhouse effect, it is hardly possible to succeed without saving the trees that now exist and by planting more. Green plants are essential allies in the effort to prevent a disastrous greenhouse future. Furthermore, tree plantations remove much less carbon from the air than wild forests. therefore the preservation of bio-diversity depends on preventing global warming—
but preventing global warming also depends on the preservation of biodiversity.
See also greenhouse effect and biodiver-sity; ice ages.
Further readingalley, Richard B. “abrupt climate change.” Scientific Amer-
ican, november 2004, 62–69.Barnett, tim P., et al. “Human-induced changes in the
hydrology of the western United States.” Science 319 (2008): 1,080–1,083.
———, et al. “Penetration of human-induced warm-ing into the world’s oceans.” Science 309 (2005): 284–287.
Bender, Morris a., et al. “Modeled impact of anthropo-genic warming on the frequency of intense atlan-tic hurricanes.” Science 327 (2010): 454–458. Summarized by Kerr, Richard a. “Models foresee more-intense hurricanes in the greenhouse.” Science 327 (2010): 399.
Brumfiel, Geoff. “academy affirms hockey-stick graph.” Nature 441 (2006): 1,032–1,033.
Comparisons of Carbon emissions from the burning of fossil fuels in Countries and regions of the World*
region and Selected CountriesContribution to world
Carbon Emissionsmetric Tons of Carbon dioxide
produced per person**
North America 25.5% 16.0
United States 21.8% 20.2
Canada 2.2% 18.1
Europe 17.2% 8.0
Eurasia 9.4% 8.9
Russia 6.2% 11.7
East and Southeast Asia 35.5% 2.7
Japan 4.7% 9.9
China 17.4% 3.9
India 4.1% 1.0
Australia 1.4% 19.4
Central and South America 3.8% 2.4
Middle East 4.9% 7.2
Africa 3.6% 1.1
World 4.2
*2004 data from Energy Information Administration**1 metric ton = 1.1 English tons
xviii+598_Biodiversity_F.indd 293 11/17/11 1:13 PM

first entry294 greenhouse effect 294
chen, J. l., et al. “Satellite gravity measurements confirm accelerated melting of Greenland ice sheet.” Science 313 (2006): 1,958–1,960.
collins, William, et al. “the physical science behind climate change.” Scientific American, august 2007, 64–73.
energy information administration. “environment: energy-related emissions data and environmental analyses.” available online. URl: http://www.eia.doe.gov/environment.html. accessed august 1, 2010.
Field, christopher B., et al. “Primary production of the biosphere: integrating terrestrial and oceanic compo-nents.” Science 281 (1998): 237–240.
Froese, Duane G., et al. “ancient permafrost and a future, warmer arctic.” Science 321 (2008): 1,648.
Fu, Qiang, et al. “enhanced mid-latitude tropospheric warming in satellite measurements.” Science 312 (2006): 1,179.
Global Footprint network. “national footprints.” avail-able online. URl: http://www.footprintnetwork.org/gfn_sub.php?content=national_footprints. accessed august 1, 2010. this Web site compares the ecological footprints of all nations.
Goswami, B. n., et al. “increasing trend of extreme rain events over india in a warming environment.” Science 314 (2006): 1,442–1,445.
Heath, James, et al. “Rising atmospheric cO2 reduces sequestration of root-derived soil carbon.” Science 309 (2005): 1,711–1,713.
Heimann, Martin. “How stable is the methane cycle?” Sci-ence 327 (2010): 1,211–1,212.
Howat, ian M., et al. “Rapid changes in ice discharge from Greenland outlet glaciers.” Science 315 (2007): 1,559–1,561.
Kerr, Richard a. “Record U.S. warmth of 2006 was part natural, part greenhouse.” Science 317 (2007): 182–183.
King, anthony W. “Plant respiration in a warmer world.” Science 312 (2006): 536–537.
Körner, christian, et al. “carbon flux and growth in mature deciduous forest trees exposed to elevated cO2.” Sci-ence 309 (2005): 1,360–1,362.
Kunzig, Robert. “Drying of the West.” National Geo-graphic, (February 2008): 90–113.
long, Stephen P., et al. “Food for thought: lower-than-expected crop yield stimulation with rising cO2 con-centrations.” Science 312 (2006): 1,918–1,921.
luo, yiqi. “terrestrial carbon cycle feedback to climate warming.” Annual Review of Ecology, Evolution, and Systematics 38 (2007): 683–712.
luthcke, S. B., et al. “Recent Greenland ice mass loss by drainage system from satellite gravity observations.” Science 314 (2006): 1,286–1,289.
Mann, Michael e., et al. “Global-scale temperature pat-terns and climate forcing over the past six centuries.” Nature 392 (1998): 779–787.
———. “Global signatures and dynamical origins of the little ice age and the Medieval climate anomaly.” Science 326 (2009): 1,256–1,260.
Meehl, Gerald a., et al. “amplifying the Pacific climate sys-tem response to a small 11-year solar cycle forcing.” Science 325 (2009): 1,114–1,118.
Meier, Mark F., et al. “Glaciers dominate eustatic sea-level rise in the 21st century.” Science 317 (2007): 1,064–1,067.
Mishchenko, Michael i., et al. “long-term satellite record reveals likely recent aerosol trend.” Science 315 (2007): 1,543.
national academies, committee on Stabilization targets for atmospheric Greenhouse Gas concentrations. “climate Stabilization targets: emissions, concen-trations, and impacts over Decades to Millennia.” Released July 16, 2010. available online. URl: http://www.nationalacademies.org/includes/Stabilization targetsFinal.pdf. accessed July 16, 2010.
national Oceanic and atmospheric administration. “nOaa: 2010 tied for warmest year on record.” available online. URl: http://www.noaanews.noaa.gov/stories2011/20110112_globalstats.html. accessed august 8, 2011.
———. “a Paleo Perspective on Global Warming: the instrumental Data of Past Global temperatures.” available online. URl: http://www.ncdc.noaa.gov/paleo/globalwarming/instrumental.html. accessed august 1, 2010.
———. “a Paleo Perspective on Global Warming: Paleocli-matic Data for the last 2000 years.” available online. URl: http://www.ncdc.noaa.gov/paleo/globalwarm ing/paleolast.html. accessed august 1, 2010.
———. “trends in carbon dioxide.” available online. URl: http://www.esrl.noaa.gov/gmd/ccgg/trends/co2_data_mlo.html. accessed august 1, 2010.
Oreskes, naomi. “the scientific consensus on climate change.” Science 306 (2004): 1,686.
———, and erik M. conway. Merchants of Doubt: How a Handful of Scientists Obscured the Truth on Issues from Tobacco Smoke to Global Warming. new york: Bloomsbury, 2010.
Osborn, timothy J., and Keith R. Briffa. “the spatial extent of 20th-century warmth in the context of the past 1200 years.” Science 311 (2006): 841–844.
Overpeck, Jonathan t., et al. “Paleoclimatic evidence for future ice-sheet instability and rapid sea-level rise.” Science 311 (2006): 1,747–1,750.
Pagani, Mark, et al. “an ancient carbon mystery.” Science 314 (2006): 1,556–1,557.
———. “Marked decline in atmospheric carbon dioxide concentrations during the Paleogene.” Science 309 (2005): 600–603.
Pearce, Fred. With Speed and Violence: Why Scientists Fear Tipping Points in Climate Change. Boston: Beacon, 2007.
xviii+598_Biodiversity_F.indd 294 11/17/11 1:13 PM

greenhouse effect and biodiversity 295
Petit, J. R., et al. “climate and atmospheric history of the past 420,000 years from the Vostok ice core, antarc-tica.” Nature 399 (1999): 429–436.
Rahmstorf, Stefan, et al. “Recent climate observations compared to projections.” Science 316 (2007): 709.
Realclimate. “Realclimate: climate science from climate scientists.” available online. URl: http://www.realcli mate.org. accessed august 8, 2011.
Retallack, Gregory J. “a 300-million-year record of atmo-spheric carbon dioxide from fossil cuticles.” Nature 411 (2001): 287–290.
Seager, Richard, et al. “Model projections of an immi-nent transition to a more arid climate in southwestern north america.” Science 316 (2007): 1,181–1,184.
Shakhova, natalia, et al. “extensive methane venting to the atmosphere from sediments of the east Siberian arctic Shelf.” Science 327 (2010): 1,246–1,250.
Shepherd, andrew, and Duncan Wingham. “Recent sea-level contributions of the antarctic and Greenland ice sheets.” Science 315 (2007): 1,529–1,532.
Siegenthaler, Urs, et al. “Stable carbon cycle-climate rela-tionship during the late Pleistocene.” Science 310 (2005): 1,313–1,317.
Solomon, Susan, and the intergovernmental Panel on cli-mate change. “climate change 2007: the Physical Science Basis.” available online. URl: http://www.ipcc.ch/publications_and_data/ar4/wg1/en/contents.html. accessed august 8, 2011.
———, et al. “contributions of stratospheric water vapor to decadal changes in the rate of global warming.” Sci-ence 327 (2010): 1,219–1,223.
Spahni, Renato, et al. “atmospheric methane and nitrous oxide of the late Pleistocene from antarctic ice cores.” Science 310 (2005): 1,317–1,321.
Sutton, Rowan t., and Daniel l. R. Hodson. “atlantic Ocean forcing of north american and european sum-mer climate.” Science 309 (2005): 115–118.
trenberth, Kevin e. “Warmer oceans, stronger storms.” Scientific American, July 2007, 44–47.
tripati, aradhna K., et al. “coupling of cO2 and ice sheet stability over major climate transitions of the last 20 million years.” Science 326 (2009): 1,394–1,397.
Velicogna, isabella, and John Wahr. “Measurements of time-variable gravity show mass loss in antarctica.” Science 311 (2006): 1,754–1,756.
Walter, Katey M., et al. “Methane bubbling from Siberian thaw lakes as a positive feedback to climate warming.” Nature 443 (2006): 71–75.
Weart, Spencer R. The Discovery of Global Warming. cambridge, Mass.: Harvard University Press, 2003.
Webster, P. J., et al. “changes in tropical cyclone number, duration, and intensity in a warming environment.” Science 309 (2005): 1,844–1,846. Summarized by Richard a. Kerr, “is Katrina a harbinger of still more powerful hurricanes?” Science 309 (2005): 1,807.
Westerling, a. l., et al. “Warming and earlier spring increase western U.S. forest wildfire activity.” Science 313 (2006): 940–943. Summary by Running, Steven W. “is global warming causing more, larger wildfires?” Science 313 (2006): 927–928.
yamamoto-Kawai, Michiyo, et al. “aragonite undersatura-tion in the arctic Ocean: effects of ocean acidification and sea ice melt.” Science 326 (2009): 1,098–1,100.
greenhouse effect and biodiversity the climate of the earth is becoming warmer. Global warming will have major impacts on natural ecosys-tems and the species of organisms that inhabit them.
Global warming is nothing new in the history of the earth; in fact it happened once before during the recent history of the modern human species, Homo sapiens. as skeptics of global warming tire-lessly point out, the earth was warmer millions of years ago, such as the middle of the Paleozoic era and of the Mesozoic era. at those times, life flour-ished on the earth. Wetland forests were exten-sive enough during the Paleozoic era that their remains form today’s coal deposits, and the earth had enough plant growth to support large popula-tions of massive dinosaurs during the Mesozoic era. clearly, a warmer earth, per se, is not incompatible with ecosystem productivity and species diversity. a period of global warming also occurred at the end of the most recent ice age, during which Stone age humans lived nearly everywhere on the earth.
there are two main reasons that global warm-ing is a bigger threat to biodiversity than it was in the past:
● The rate of temperature increase. climatic shifts in the past were generally much slower than the shifts occurring today. Despite tem-porary (and catastrophic) changes such as the younger Dryas (a sudden reversal of the warming trend), the global warming that occurred at the end of the most recent ice age was 10 times slower than the global warming that is occurring today.
● The ability of organisms to disperse. in the Holocene, the northern continents were largely free of obstructions to the migration of animals and the dispersal of plants. today, cities and farms and roads block their migra-tion. Species that live on the shore may be trapped there as sea levels rise, and animals and the seeds of plants will find few suitable places to live while they migrate northward. Humans, who did not have national boundar-ies or own property, were also free to migrate.
xviii+598_Biodiversity_F.indd 295 11/17/11 1:13 PM

first entry296 greenhouse effect and biodiversity 296
For these two reasons, global warming (and associated changes such as droughts and wildfires in continental areas) presents a major threat to biodiversity.
GloBal warmING aNd plaNTSGlobal warming is already having measurable effects on plants. Plant species are growing closer to the poles and farther up on mountains as a result of global warming. antarctica has only two native species of higher plants, one of which is the grass Deschampsia antarctica. it used to form only widely spaced tussocks in protected crevices along the coast, but antarctica now has its first grass meadows.
a major indicator of global warming is that buds of deciduous trees in the northern continents are opening sooner now than even a few decades ago—spring is coming earlier. a few isolated instances stand out. Perhaps the most famous example and most noticed by the United States federal govern-ment in Washington, D.c. was when the cherry trees bloomed along the Potomac in January 2007, far sooner than their usual budburst time.
But scientists do not rely on isolated examples. Botanist a. J. Miller-Rushing and associates com-pared pressed plant specimens and photographs from the 19th and early 20th centuries with recent specimens and photographs. their analysis shows that trees are leafing earlier and herbaceous plants emerging earlier in the very same locations—about 11 days earlier per century. Miller-Rushing and colleagues used 285 photographs and numerous preserved specimens. this pattern is confirmed by satellite imagery, which indicates that deciduous forests are becoming green earlier in the spring than in previous decades. another study, published by climatologist Xiaoyang Zhang and associates, shows that lilacs planted all over north america have been flowering earlier in the spring. inter-estingly, some of the southern lilacs have been blooming later rather than earlier, because in those southern locations the winters have not been cold enough to allow the biochemical breakdown of budburst inhibitors.
in addition to changing the seasonal patterns of plant growth, global warming is also causing changes in where the plants live. ecologist camille Parmesan has reported the results of a survey of 1,700 plant species. On average, these plant spe-cies migrated northward about four miles (almost 6 km) per decade and migrated higher up in the mountains by about 24 feet (about 8 m) per decade. She also found that, on average, spring budburst occurred about two days earlier per decade. Daniel W. McKenney of the canadian Forest Service and
associates studied 130 tree species. they have esti-mated that most of the trees will experience severe reductions in geographical range even if they man-age to migrate hundreds of miles northward. in Brit-ain, 385 species of plants flowered an average of four and a half days earlier over the course of a decade. in Spain, high-altitude grasslands are being replaced by lower-altitude shrublands.
One reason that these shifts in range and sea-sonality are important is that they may result in extinctions. the plants that already live at the tops of mountains, such as in the alpine tundra of the Rocky Mountains and Sierra nevada and in the cloud forests of central america, have no place to go if conditions become warmer or drier. the alpine tundra, which consists of small grasses, sedges, and wildflowers, survives only because conditions are too cold for trees to grow over them and shade them. Once the subalpine spruce and pine forests begin to grow to the very tops of the mountains, the tundra plants and animals will face extinction. the cloud forests of central america persist because they are inside of clouds and receive abundant water from mists. Warmer temperatures are already causing the cloud levels to rise, leaving the cloud forests high and dry. One of the richest biodiversity hot spots is the cape region of South africa, which is at the south-ern tip of the african continent. the endemic plants of this area have no place to go when temperature and moisture conditions change.
emerging from dormancy earlier in the spring may even cause direct harm to plants. this could occur if an extended warm period in the winter awakened buds or animals, which may then be killed by the return of winter. Buds of post oak trees (Quercus stellata) in Oklahoma open on warm days in winter and then die when cold weather returns. later, when temperatures become and remain warmer, another set of buds open in place of the damaged buds. as global warming progresses, such events will become more common. Whether this loss of some of the buds may have long-term effects on the growth of the oaks is not known.
coniferous forest trees in the western United States—even in forests that appear to be otherwise healthy—are currently experiencing a higher death rate than they were four decades ago. according to forest scientist Phillip van Mantgem and associates, death rate of trees per year has increased from 0.3 percent to 1.3 percent in the Pacific northwest, from 1.0 percent to 1.7 percent in the mountains of cali-fornia, and from 0.2 percent to 0.6 percent in the interior forests. this pattern held true at all 76 loca-tions and at all sizes, elevations, species, and ages of trees. the researchers attribute the increased death to higher temperatures and less precipitation, both
xviii+598_Biodiversity_F.indd 296 11/17/11 1:13 PM

greenhouse effect and biodiversity 297
of which are associated with global warming that has occurred during that period.
GloBal warmING aNd TErrESTrIal aNImalSanimals are also responding to climatic changes. armadillos have been expanding from their native habitats in the South, as far north as illinois. Several bird species have moved their breeding ranges north-ward. Over the course of 20 years, British birds have shifted their ranges about 12 miles (15 km) north-ward, and British birds are laying their eggs sooner. camille Parmesan studied not only plants but also nonmigratory european butterflies, most of which shifted their range northward between 22 to 148 miles (28 to 185 km) during the 20th century. Over the course of 100 years, mammals that live at lower elevations in mountains have moved to higher eleva-tions, and the mammals that live at high elevations have experienced range restrictions. Global warming has caused changes in migration patterns of some birds. the european blackcap breeds in Germany and has traditionally migrated to Spain and Portugal to overwinter. Recently, some flocks of blackcaps have spent the winter in england instead. the flocks return from england earlier than the flocks from Portugal, with the result that the england-wintering birds mate earlier and leave more offspring. Scien-tists estimate that climate-induced changes in the migration patterns of this species will result in evolu-tionary changes in their migration instinct.
animal species are already facing extinction from the climatic changes in their habitat. the golden toad has already become extinct at least in part because of the drier conditions in the cloud for-ests. this is only the beginning. Ornithologist Çağan Şekercioğlu and associates have predicted the extinc-tion of between 100 and 500 bird species for each 1°F increase in global temperature.
GloBal warmING aNd marINE orGaNISmSas the oceans become warmer, the extinction of some marine creatures appears to be inevitable. in particular, warmer waters and higher salt concen-trations have already caused many coral reefs to experience bleaching. When the corals die, the entire food chain collapses and is replaced by one that is based upon algae. living corals prevent algae from growing on the reefs, but the algae grow abundantly on dead coral structures. the result is a striking decrease in the diversity of species at all levels of the food chain.
Scientists have also given much attention to the effects of ocean acidification. Oceans absorb cO2, too slowly to prevent cO2 buildup in the atmo-sphere, but quickly enough to have already caused a measurable increase in the acidity of the water.
acidic waters dissolve calcium carbonate. this con-tributes to the death of animals that use calcium for their structure, including corals, molluscs, and zooplankton. the death of these organisms has a feedback effect on the ocean’s ability to absorb cO2. Fishes excrete a lot of carbonate and also make a substantial contribution to the removal of carbon from surface waters. Organisms use carbon as well as calcium to make their shells, then when they die they sink to the bottom of the sea, taking the carbon away from the surface and thus from the atmosphere. if there are fewer shelled organisms, the ocean will remove less carbon from the atmosphere. Ocean acidification has already reduced the rate of calcium deposition in reefs.
GloBal warmING aNd EColoGICal INTEraCTIoNS
Global warming can also disrupt ecological relation-ships between species. Warmer temperatures may give insects a longer reproductive season and allow them to spread rapidly on their host plants. (this includes the insect pests of crop plants.) Warmer temperatures are already allowing outbreaks of spruce bark beetles (Dendroctonus rufipennis) and pine bark beetles (D. ponderosae) to destroy mil-lions of spruces in alaska and canada, as well as englemann spruces and lodgepole and piñon pines in the Rocky Mountains. the beetles transmit a fungus (Conartium ribicola).Warmer temperatures allow beetle populations to explode because they can complete their life cycle in one year rather than the normal two. Reduced sap production in the trees, associated with warmer, drier conditions, also encourages the survival of the beetles, since sap blocks the burrows created by the beetle larvae. D. ponderosae is spreading the same fungus among whitebark pines (Pinus albicaulis) in the subalpine forests of california. Because diseases are spread-ing through the boreal and subalpine forests, global warming is causing these forests to turn brown at the same time that it is causing the tundra adjacent to them to turn greener.
Global warming can also have ecological effects via drought. in one example, salt marsh blue crabs (Callinectes sapidus) normally kept periwinkle snail (Littoraria irrorata) populations in check. the snails would damage the salt marsh cordgrass (Spartina alterniflora) by puncturing the grass leaf cuticle, allowing a Fusarium fungus to invade the leaves. During droughts, the cordgrass is stressed by higher concentrations of salt as well as acid and toxic chemi-cals in the water. But in addition, the drought reduces the crab population, allowing an outbreak of snails.
Other disrupted ecological relationships result when global warming causes some organisms to
xviii+598_Biodiversity_F.indd 297 11/17/11 1:13 PM

first entry298 greenhouse effect and biodiversity 298
become active earlier in the spring than do others. One example is the relationship between flower-ing plants and their insect pollinators. Most wild plant species, other than conifers, grasses, and sedges, depend on insects for pollination. if warmer temperatures change the seasonal life cycles of the plants and the pollinators in different ways, the flowers may open when the pollinators are not pres-ent. as a result, the pollinators may find no food, and the flowers will not be pollinated. another example is the disruption of the food chain. Her-bivorous animals may respond more quickly to the warming climate than do the plants that they eat, with the result that the animals may hatch and find little or nothing to eat. Or, if warmer springs cause caterpillars to hatch before the baby birds that eat them, the nestlings may not have enough to eat. Within bird populations, those individuals that could best adjust the timing of their reproduction have been more successful than those that could not alter their reproduction to adjust to the earlier arrival of spring.
an experiment in a california grassland indi-cates that increased precipitation during the nor-mally rainy winter would have little effect on the growth of plants or the dominant species of plants and invertebrate animals, but rainfall that extended the spring rains into summer would have a profound effect. What that effect would be is also unclear. in this experiment, there was an initial burst of growth in some plant species, followed by a decline in both plant growth and number of plant and invertebrate species in the later years of the experiment.
GloBal warmING aNd EvoluTIoNthere is some evidence that global warming may have already begun to induce evolutionary changes. Researchers have observed genetic alteration within species. Genetic changes have occurred in wild pop-ulations of fruit flies similar to those that occur in hot and dry conditions.
Researchers have also observed what might be the beginning of the evolution of one species into two or more as a result of global warming. in the european blackcap example mentioned above, the birds that overwinter in england mate mostly with one another rather than with the birds that over-winter in Spain and Portugal; that is, they are repro-ductively isolated from the birds that migrate from Spain and Portugal. this may eventually allow one bird species to diverge into two.
these evolutionary changes are not necessarily bad. in fact, they should allow species to adjust to the changes associated with global warming—except that the changes are occurring too fast for evolution
to help them. they illustrate the very broad impact of global warming on the natural world.
GloBal warmING, wIld SpECIES, aNd humaNSShifts in the geographical ranges of plants and ani-mals and other biological changes may have direct impact on humans. Studies led by Fakhri Bazzaz suggest that global climate change may enhance the growth and the pollen production of ragweeds. Warm weather causes poison ivy to produce more irritant. and tropical mosquitoes, which spread numerous diseases such as malaria and dengue fever, may spread northward and introduce these diseases, which americans have been privileged to ignore, to the United States.
Government agencies and private landowners have set aside “nature preserves” for wild plants and animals, but as the climate becomes warmer, wild species will be trapped in preserves no longer suitable for them. ecologists are beginning to design preserves that are connected by corridors, which will allow plants and animals to move northward from one preserve to another in response to global warm-ing. But such designs are still far from widespread implementation and will probably prove to be too little too late for the wild species. Some ecologists are even suggesting that humans need to intervene to assist the migration of species to new locations where they can survive.
these are just the effects of global warming that scientists have already begun to observe. if global warming disrupts human civilization (for example, by creating environmental refugees and by causing droughts in agricultural areas), humans may turn in desperation to even more harmful exploitation of natural resources than they already are. at the pres-ent time, africans are eating the meat of wild apes partly because of the collapse of ocean fish popula-tions. Will global warming cause such impacts to become more severe and widespread?
See also greenhouse effect; ice ages.
Further readingaronson, Richard B., et al. “climate change and the inva-
sibility of the antarctic benthos.” Annual Review of Ecology, Evolution, and Systematics 38 (2007): 129–154.
Bearhop, Stuart, et al. “assortative mating as a mechanism for rapid evolution of a migratory divide.” Science 310 (2005): 502–504.
Berteaux, D., et al. “Keeping pace with fast climate change: can arctic life count on evolution?” Integrative and Comparative Biology 44 (2004): 140–151.
Bradshaw, W. e., and c. M. Holzapfel. “Genetic shift in photoperiodic response correlated with global warm-
xviii+598_Biodiversity_F.indd 298 11/17/11 1:13 PM

greenhouse effect and biodiversity 299
ing.” Proceedings of the National Academy of Sciences USA 98 (2001): 14,509–14,511.
charmantier, anne, et al. “adaptive phenotypic plasticity in response to climate change in a wild bird popula-tion.” Science 320 (2008): 800–803.
colwell, Rita R. “Global climate and infectious disease: the cholera paradigm.” Science 274 (1996): 2,025–2,031.
colwell, Robert K., et al. “Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics.” Science 322 (2008): 258–261.
crick, H. Q. P., et al. “UK birds are laying eggs earlier.” Nature 388 (1997): 526.
De’ath, Glenn. “Declining coral calcification on the Great Barrier Reef.” Science 323 (2009): 116–119. Summa-rized by Pennisi, elizabeth. “calcification rates drop in australian reefs.” Science 323 (2009): 27.
Fitter, a. H., and R. S. R. Fitter. “Rapid changes in flow-ering time in British plants.” Science 296 (2002): 1,689–1,691.
Grabherr, G., et al. “climate effects on mountain plants.” Nature 369 (1994): 448.
Hoegh-Guldberg, O., et al. “assisted colonization and rapid climate change.” Science 321 (2008): 345–346.
———, et al. “coral reefs under rapid climate change and ocean acidification.” Science 318 (2007): 1,737–1,742.
lenoir, J., et al. “a significant upward shift in plant species optimum elevation during the 20th century.” Science 320 (2008): 1,768–1,771.
lindgren, e., and R. Gustafson. “tick-borne encephalitis in Sweden and climate change.” Lancet 358 (2001): 16–18.
lovejoy, thomas, and l. Hannah, eds. Climate Change and Biodiversity. new Haven, conn.: yale University Press, 2005.
lyon, Bruce e., et al. “a matter of timing.” Science 321 (2008): 1,051–1,052.
McKenney, Daniel W., et al. “Potential impacts of climate change on the distribution of north american trees.” BioScience 57 (2007): 939–948.
Miller-Rushing, abraham J., et al. “Photographs and her-barium specimens as tools to document phenological changes in response to global warming.” American Journal of Botany 93 (2006): 1,667–1,674.
Moritz, craig, et al. “impact of a century of climate change on small-mammal communities in yosemite national Park, USa.” Science 322 (2008): 261–264.
Myneni, R. B., et al. “increased plant growth in the north-ern high latitudes from 1981 to 1991. Nature 386 (1997): 698–702.
nussey, Daniel H., et al. “Selection on heritable pheno-typic plasticity in a wild bird population.” Science 310 (2005): 304–306.
Parmesan, camille, and Gary yohe. “a globally coherent fingerprint of climate change impacts across natural systems.” Nature 421 (2003): 37–42.
———, et al. “Poleward shifts in geographical ranges of butterfly species associated with regional warming.” Nature 399 (1999): 579–583.
Patz, J. a., et al. “impact of regional climate change on human health.” Nature 438 (2005): 310–317.
Pounds, J. a., et al. “Biological response to climate change on a tropical mountain.” Nature 398 (1999): 611–615.
———, et al. “Widespread amphibian extinctions from epidemic disease driven by global warming.” Nature 439 (2006): 161–167.
Powell, J., and J. logan. “Ghost forests, global warming and the mountain pine beetle.” American Entomolo-gist 47 (2001): 160–172.
Pulido, Francisco, and Peter Berthold. “current selection for lower migratory activity will drive the evolution of residency in a migratory bird population.” Proceed-ings of the National Academy of Sciences 107 (2010): 7,341–7,346.
Rice, Stanley a. “Plants help to keep the earth from over-heating.” chap. 3 in Green Planet: How Plants Keep the Earth Alive. new Brunswick, n.J.: Rutgers Uni-versity Press, 2009.
Rogers, christine a., et al., “interaction of the onset of spring and elevated atmospheric cO2 on ragweed (Ambrosia artemesiifolia l.) pollen production.” Environmental Health Perspectives 114 (2006): 865–869.
Sanz-elorza, Mario, et al. “changes in the high-mountain vegetation of the central iberian Peninsula as a prob-able sign of global warming.” Annals of Botany 92 (2003): 273–280.
Schneider, Steven H., and t. l. Root. “ecological implica-tions of climate change will include surprises.” Biodi-versity and Conservation 5 (1996): 1,109–1,119.
Şekercioğlu, Çağan H., et al. “climate change, elevational range shifts, and bird extinctions.” Conservation Biol-ogy 22 (2008): 149–150.
Silliman, B. R., et al. “Drought, snails, and large-scale die-off of southern US salt marshes.” Science 310 (2005): 1,803–1,806.
Sturm, Matthew. “arctic plants feel the heat.” Scientific American, May 2010, 66–73.
thomas, chris D., and Jack J. lennon. “Birds extend their ranges northward.” Nature 399 (1999): 213.
Umina, P. a., et al. “a rapid shift in a classic clinal pattern Drosophila reflecting climate change.” Science 308 (2005): 691–693.
van Mantgem, Phillip J., et al. “Widespread increase of tree mortality rates in the western United States.” Science 323 (2009): 521–524. Summarized by Pennisi, eliza-beth. “Western U.S. forests suffer death by degrees.” Science 323 (2009): 447.
Visser, Marcel e., and leonard J. M. Holleman. “Warmer springs disrupt the synchrony of oak and winter moth
xviii+598_Biodiversity_F.indd 299 11/17/11 1:13 PM

first entry300 gymnosperms 300
phenology.” Proceedings of the Royal Society of Lon-don B (Biology) 286 (2001): 289–294.
———, ———, and Phillip Gienapp. “Shifts in caterpil-lar biomass phenology due to climate change and its impact on the breeding biology of an insectivorous bird.” Oecologia 147 (2006): 164–172.
Willis, Kathy J., and Shonil a. Bhagwat. “Biodiversity and climate change.” Science 326 (2009): 806–807.
Wilson, R. W., et al. “contribution of fish to the marine inorganic carbon cycle.” Science 323 (2009): 359–362.
Zeebe, Richard e., et al. “carbon emissions and acidifica-tion.” Science 321 (2008): 51–52.
gymnosperms See conifers; seed plants.
xviii+598_Biodiversity_F.indd 300 11/17/11 1:13 PM

301
habitat fragmentation Habitat fragmenta-tion occurs when a large habitat, with large pop-ulations of many of its species, becomes divided into numerous smaller habitats. Fragmentation can occur as a result of natural processes. as the climate became drier over much of north america dur-ing the late neogene period, moist habitats became smaller and isolated from one another. Some species that were once widespread, such as the california fan palm Washingtonia fi lifera, may survive only as endemic species in desert springs. the isolated populations may diversify into different species or subspecies. For example, pupfi shes were common in rivers and streams of what is now the Mojave and Sonora Deserts of southwestern north america. as desert conditions spread, the fi shes were isolated into separate desert springs, where they evolved into separate species. today, Cyprinodon diabolis lives only in Devil’s Hole, and C. nevadensis nevadensislives only in Saratoga Springs, both in Death Valley.
today, human activity is causing more habitat fragmentation than has probably ever occurred dur-ing the history of the earth. Humans have altered most of the surface of the earth. they sometimes set aside wildlife preserves, which are only tiny fragments of the original habitat range, in which they permit wild species to survive. even these few remaining habitats are divided by roads, even the smallest of which may represent a barrier to the movement of some species.
Habitat fragmentation has numerous signifi cant effects on wild species, including the following:
● Edge effect. the edges of a habitat, the cli-matic conditions of which are strongly infl u-
enced by the environment outside, may not be suitable for the survival of many of the habitat’s species. in particular, many large animals stay away from the edges, where they would frequently come in contact with people.
● Breaking larger populations into smaller ones. each species has populations in each fragment that are effectively isolated from the populations in other fragments. each population therefore has a reduced amount of genetic variation upon which natural selection can act. the populations may be small enough that genetic drift may cause random genetic changes.
● Blocked migration. the species in each fragment are unable to migrate to other fragments. if corridors of migration were open, individuals could migrate from one fragment to another in the event that one of the fragments becomes uninhabitable. it is particularly important that organisms have corridors of migration over long distances, which will allow them to migrate (as they did after the most recent ice age) in response to global warming.
the only solution to the edge effect problem would be to establish larger protected areas. this is extremely unlikely to occur, since most unprotected areas are privately owned. in the western United States, the federal government owns a lot of land, some of which is protected as wilderness areas or national parks, while much of the national forest and national grassland area is leased to private timber
H
xviii+598_Biodiversity_F.indd 301 11/17/11 1:13 PM

first entry302 herbivory 302
and ranching interests. the federal government could consolidate protected areas, removing some of the leased areas from private use. However, the lease-holders have strongly voiced their opinions that the land is effectively theirs, even if they do not own it. in what has been dubbed the “Sagebrush Rebellion,” some Bureau of land Management officials have been intimidated by ranchers.
a possible solution to the problem of migration may be to establish corridors of suitable habitat between the isolated fragments. this would also help to reduce the genetic problems of the isolated populations. this is now an important component of conservation planning. the yellowstone to yukon conservation initiative is intended to connect 2,000 miles (3,200 km) of protected mountain areas. the Mesoamerican Biological corridor is intended to protect 80,000 square miles (208,000 square km), which is 27 percent of the land area in central america. these corridors, however, have an even more pronounced edge effect than the fragments they connect. it remains to be seen whether these corridors can be established and, if so, whether they will have the desired effect of saving biodiversity.
See also communities, ecological; endemic species; federal government role in conser-vation (United States).
Further readingartis, Scott. “combating habitat fragmentation with gar-
den corridors.” available online. URl: http://www.ecosystemgardening.com/gardens-as-wildlife-corri dors.html. accessed august 1, 2010.
cawley, R. McGreggor. Federal Lands, Western Anger: The Sagebrush Rebellion and Federal Politics. lawrence: University Press of Kansas, 1993.
clark, Rulon W., et al. “Roads, interrupted dispersal, and genetic diversity in timber rattlesnakes.” Conservation Biology 24 (2010): 1,059–1,069.
cole, David n. Beyond Naturalness: Rethinking Park and Wilderness Stewardship in an Era of Rapid Change. Washington, D.c.: island Press, 2010.
collinge, Sharon K. Ecology of Fragmented Landscapes. Baltimore, Md.: Johns Hopkins, 2009.
Damschen, ellen i., et al. “corridors increase plant spe-cies richness at large scales.” Science 313 (2006): 1,284–1,286.
Fahrig, lenore. “effects of habitat fragmentation on bio-diversity.” Annual Review of Ecology, Evolution, and Systematics 34 (2003): 587–516.
Ferraz, Gonçalo, et al. “a large-scale deforestation experi-ment: effects of patch area and isolation on amazon birds.” Science 315 (2007): 238–241.
Forman, Richard t. t., and lauren e. alexander. “Roads and their major ecological effects.” Annual Review of Ecology and Systematics 29 (1998): 207–231.
Gu, Weidong, et al. “estimating the consequences of habi-tat fragmentation on extinction risk in dynamic land-scapes.” Landscape Ecology 17 (2002): 699–710.
Hilty, Jodi, et al. Corridor Ecology: The Science and Prac-tice of Linking Landscapes for Biodiversity Conserva-tion. Washington, D.c.: island Press, 2006.
Johnson, Douglas H. “Habitat fragmentation effects on birds in grasslands and wetlands: a critique of our knowledge.” Great Plains Research 11 (2001): 211–231.
letcher, Benjamin H., et al. “Population response to habi-tat fragmentation in a stream-dwelling brook trout population.” Public Library of Science One 2 (2007): e1139.
lindenmayer, David, and Joern Fischer. Habitat Fragmen-tation and Landscape Change: An Ecological and Conservation Synthesis. Washington, D.c.: island Press, 2006.
lópez, alexander, and alicia Jiménez. “environmental con-flict and cooperation: the Mesoamerican Biological corridor as a Mechanism for transborder environmen-tal cooperation.” United nations environmental Pro-gramme. available online. URl: http://www.unep.org/dewa/eccreports/ecPi_latin_america_assessment_Final_Report_Feb21. pdf. accessed august 14, 2010.
Opdam, Paul, and Dirk Wascher. “climate change meets habitat fragmentation: linking landscape and biogeo-graphical scale levels in research and conservation.” Biological Conservation 117 (2004): 285–297.
Peterkin, George. “Reversing the habitat fragmentation of British woodlands.” available online. URl: http://assets.panda.org/downloads/ukforestsfragmentation.pdf. accessed august 1, 2010.
Ribeiro, M. c., et al. “the Brazilian atlantic Forest: How much is left, and how is the remaining forest distrib-uted? implications for conservation.” Biological Con-servation 142 (2009): 1,141–1,153.
tallmon, David a., et al. “Of mice and men and trillium: cascading effects of forest fragmentation.” Ecological Applications 13 (2003): 1,193–1,203.
turner, Monica G. “landscape ecology: What is the state of the science?” Annual Review of Ecology, Evolution, and Systematics 36 (2005): 319–344.
Worboys, Graeme l. Connectivity Conservation: A Man-agement Guide. london: earthscan Publications, 2010.
herbivory the term herbivore usually refers to an animal that eats leaves, stems, roots, or fruits. if an animal consumes and destroys a whole plant (for example, a seed that contains an embryonic plant), it is considered to be a predator.
Herbivorous insects may lay hundreds of eggs and produce several generations of offspring in a single summer, building up huge populations. Popu-
xviii+598_Biodiversity_F.indd 302 11/17/11 1:13 PM

herbivory 303
lations of herbivores could quickly consume every green sprig on the planet. What prevents them from doing this? Why is the world green? the answer is that plants have many defenses that have evolved against herbivores. Plants derive no benefit from having their leaves eaten; natural selection has pro-duced characteristics that protect the leaves from herbivores. Herbivory is necessary to the global eco-system, but plants do everything they can to avoid it.
Plants have numerous characteristics that make them difficult to eat, for example:
● Many plants have physical defenses such as spines and hairs. a cactus in the desert stores water in its stem; its spines protect it from thirsty animals. Hairs can be as effec-tive against tiny insects as spines are against larger animals.
● all plant parts contain large amounts of cellulose, which is the major component of cell walls and which most animals cannot digest. Many stems are woody, and most animals cannot eat them.
● Grass stems and leaves are very rough because they contain silicon compounds.
● Deer will not browse pine needles in winter because the needles are very tough, waxy, and full of sticky resin.
● Plants that store large quantities of starch, such as potatoes, generally do so in places not accessible to most herbivores, such as in underground tubers or the cortex in the middle of the stem.
in addition, most plants contain chemical com-pounds that are harmful to herbivores, especially in their leaves. Some of these chemicals inhibit the efficiency of digestion; others are mildly, or strongly, poisonous. examples include:
● Mildly toxic alkaloids, such as caffeine, nicotine, cocaine, morphine, and theobro-mine (an ingredient in chocolate), are wide-spread in the plant kingdom. Various other chemicals, such as mustard oils and menthol (which gives mint its distinctive smell), also make some leaves unpalatable to herbivores.
● Some plants produce extremely toxic com-pounds. the ricin that is found in the seeds of the castor bean (Ricinus communis) is the third most toxic biological compound known, following the bacterial toxins of botulism and tetanus. Many of these very toxic compounds, however, may not be as toxic to specific herbivores that have evolved the ability to consume the plant.
● Some plants absorb toxic metal ions from the soil and accumulate them in their leaves. locoweeds (genera Astragalus and Oxytro-pis) accumulate selenium.
● Milkweeds contain chemicals that interfere with heart muscle contractions. Some chem-icals, such as those found in members of the nightshade family and the ricin found in castor bean plants, are among the deadliest chemicals known. the pesticide rotenone is extracted from South american and asian leguminous plants.
these chemicals may be toxic not only to herbi-vores but also to bacteria and fungi. therefore many chemicals found in leaves and bark have antibiotic and other pharmaceutical properties.
Plants must use a great deal of energy and material to manufacture defensive structures and chemicals. if a plant makes no such compounds and escapes the herbivores, it can afford to grow larger and produce more seeds. in some cases, plants that grow rapidly and live for only a short period and that live only temporarily in one place do not pro-duce many defensive chemicals. if herbivores find these plants they can eat them easily, but frequently these plants complete their life cycles before the her-bivores find them. in contrast, slow-growing, long-lived plants may benefit from producing defensive compounds. For example, botanist Stanley Rice and his students found that leaves of long-lived bur oak trees (Quercus macrocarpa) were more toxic than those of short-lived chokecherry leaves (Prunus vir-giniana) in the Black Hills.
Plant defense against herbivores can also vary with the seasons. the leaves of trees in climates with cold winters may produce few defensive chemicals in the spring when insects are not yet abundant; during the summer, however, when insect popula-tions increase, the leaves produce defensive chemi-cals. the tree can grow rapidly during the spring because it is not spending its energy on making defensive chemicals. this was the pattern observed by ecologist Paul Feeny in the english oak (Quercus robur). in climates with milder winters, however, herbivorous insects are abundant in early spring, and the trees may produce leaves that are toxic when they are young. Several researchers have found that oak leaves produce more tannins in the spring than in the summer. For example, botanist Stanley Rice and associates have found that the post oak (Quer-cus stellata) in the dry, warm forests of Oklahoma produce more tannins in the spring than in the sum-mer (see figure on page 304). ecologist Juha-Pekka Salminen and associates found the same pattern in Quercus robur (the same species studied by Feeny)
xviii+598_Biodiversity_F.indd 303 11/17/11 1:13 PM
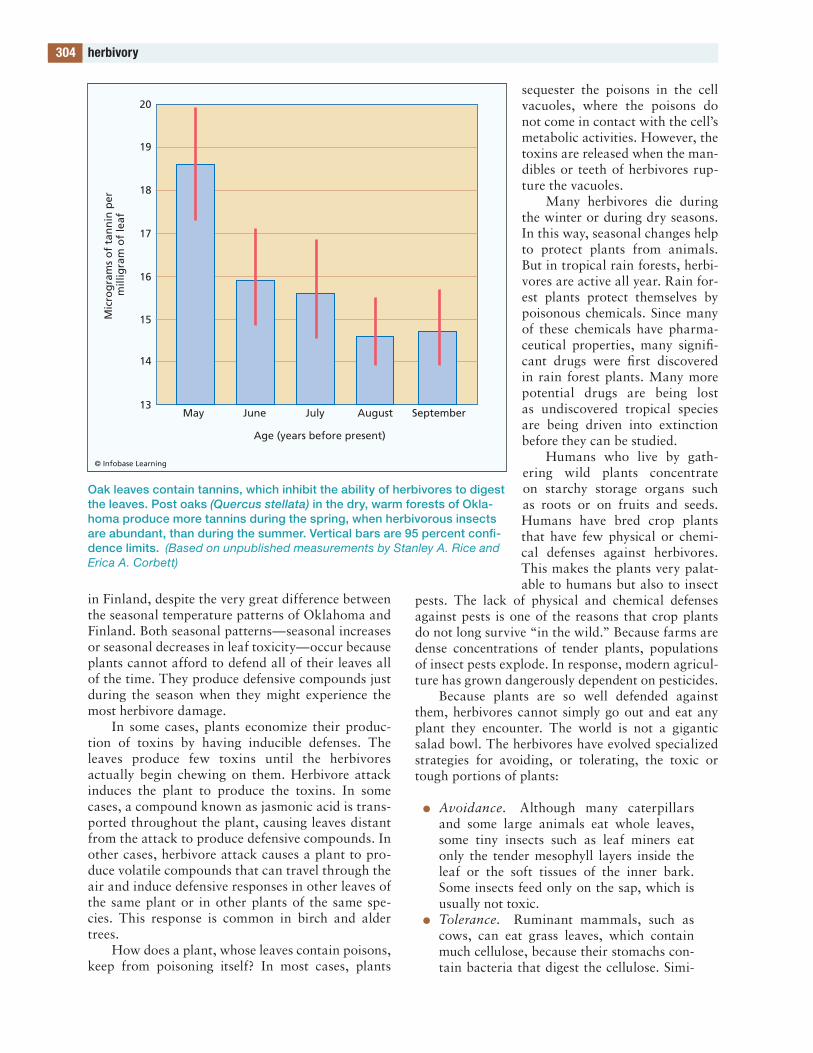
first entry304 herbivory 304
in Finland, despite the very great difference between the seasonal temperature patterns of Oklahoma and Finland. Both seasonal patterns—seasonal increases or seasonal decreases in leaf toxicity—occur because plants cannot afford to defend all of their leaves all of the time. they produce defensive compounds just during the season when they might experience the most herbivore damage.
in some cases, plants economize their produc-tion of toxins by having inducible defenses. the leaves produce few toxins until the herbivores actually begin chewing on them. Herbivore attack induces the plant to produce the toxins. in some cases, a compound known as jasmonic acid is trans-ported throughout the plant, causing leaves distant from the attack to produce defensive compounds. in other cases, herbivore attack causes a plant to pro-duce volatile compounds that can travel through the air and induce defensive responses in other leaves of the same plant or in other plants of the same spe-cies. this response is common in birch and alder trees.
How does a plant, whose leaves contain poisons, keep from poisoning itself? in most cases, plants
sequester the poisons in the cell vacuoles, where the poisons do not come in contact with the cell’s metabolic activities. However, the toxins are released when the man-dibles or teeth of herbivores rup-ture the vacuoles.
Many herbivores die during the winter or during dry seasons. in this way, seasonal changes help to protect plants from animals. But in tropical rain forests, herbi-vores are active all year. Rain for-est plants protect themselves by poisonous chemicals. Since many of these chemicals have pharma-ceutical properties, many signifi-cant drugs were first discovered in rain forest plants. Many more potential drugs are being lost as undiscovered tropical species are being driven into extinction before they can be studied.
Humans who live by gath-ering wild plants concentrate on starchy storage organs such as roots or on fruits and seeds. Humans have bred crop plants that have few physical or chemi-cal defenses against herbivores. this makes the plants very palat-able to humans but also to insect
pests. the lack of physical and chemical defenses against pests is one of the reasons that crop plants do not long survive “in the wild.” Because farms are dense concentrations of tender plants, populations of insect pests explode. in response, modern agricul-ture has grown dangerously dependent on pesticides.
Because plants are so well defended against them, herbivores cannot simply go out and eat any plant they encounter. the world is not a gigantic salad bowl. the herbivores have evolved specialized strategies for avoiding, or tolerating, the toxic or tough portions of plants:
● Avoidance. although many caterpillars and some large animals eat whole leaves, some tiny insects such as leaf miners eat only the tender mesophyll layers inside the leaf or the soft tissues of the inner bark. Some insects feed only on the sap, which is usually not toxic.
● Tolerance. Ruminant mammals, such as cows, can eat grass leaves, which contain much cellulose, because their stomachs con-tain bacteria that digest the cellulose. Simi-
Oak leaves contain tannins, which inhibit the ability of herbivores to digest the leaves. Post oaks (Quercus stellata) in the dry, warm forests of Okla-homa produce more tannins during the spring, when herbivorous insects are abundant, than during the summer. Vertical bars are 95 percent confi-dence limits. (Based on unpublished measurements by Stanley A. Rice and Erica A. Corbett)
xviii+598_Biodiversity_F.indd 304 11/17/11 1:13 PM

herbivory 305
larly, koalas can eat unpalatable eucalyptus leaves because of their complex intestines. this is an example of mutualism. Many cat-erpillars, mammals such as sloths, and birds such as the hoatzin consume massive quanti-ties of leaves and excrete most of what they eat, including the poisons.
in many cases, the herbivores can use the plant defenses for their own benefit. the caterpillars of monarch butterflies are poisonous. inexperienced birds may eat them and then vomit, thereby learning to avoid them. But the caterpillars do not produce the poison themselves. it comes from the milkweeds that they eat. the caterpillars confine the poisons in specialized structures within their bodies, where the poison does not affect them but will harm a predator that, in the process of eating the caterpillar, breaks open the poison sacs. another example involves chim-panzees, which, like humans, can be infected with malaria. the chimps eat the antimalarial leaves of a plant Trichilia rubescens, and they eat clay with the leaves, which enhances antimalarial activity of the leaves. Birds may include sprigs of aromatic plants in their nests which, according to some researchers, reduces the number of parasites on their nestlings.
in many spice plants, chemicals deter herbivores from eating the leaves. the spice compounds in the leaves make them unsuitable as a staple food source for humans, but humans find these chemicals pleas-ant in small quantities. Many human cultures have made use of poisonous plant compounds such as caffeine as stimulants, but only in small quantities.
Some plant species have intricate defenses against herbivores. certain butterflies lay their eggs on leaves of passionflower vines. to prevent over-crowding of their larvae, the butterflies avoid laying eggs on leaves upon which eggs have already been laid. Some species of passionflower vine have evolved yellowish spots or bumps on the leaves, which look (to the eye of the butterfly) like eggs. natural selec-tion proceeded to modify these spots and bumps into structures that convincingly resemble insect eggs, because the butterflies most frequently avoided the plants that had the most egglike spots upon them.
Despite all their defenses, most plants suffer at least some loss to herbivores. However, they have evolved methods of making the best of their losses. consider a plant, for example, that is experienc-ing a mild drought or is growing in poor soil. if a herbivore consumes some of the leaves but does not harm the roots, the remaining leaves now have access to all of the water and minerals that the roots can provide. the additional water and minerals may therefore stimulate photosynthesis in the remaining leaves. this may allow compensatory regrowth, in
which the plant grows back and becomes as large as it would have been if the herbivores had not attacked it.
Plants defend their leaves, stems, and roots from herbivores. Fruits, in contrast, may attract animals rather than driving them away. the function of a fruit is to get the seeds transported to a new loca-tion, away from the parent plant. that is, animals (especially birds and mammals) and plants have coevolved a dispersal relationship that benefits both of them. this is why many fruits attract, rather than repel, animals.
the effects of herbivory are not limited to the plant and the herbivore. they can produce a cascade of effects on ecological communities. For example, when herbivores eat the flowers, they affect the num-ber of flowers available to pollinators. But in addi-tion, the plant releases volatile chemicals, which may themselves affect the behavior of the pollinators. Plant toxins may affect not only the herbivores but the parasites that live upon the herbivores.
When a plant species migrates or is transported by humans to a new location, it often becomes an invasive species. One of the reasons that it may suc-cessfully invade its new habitat is that it has left behind the herbivores that previously kept its popu-lations under control. the plant populations may then evolve a different defensive chemistry suitable to their new biological environment. if the herbivore is subsequently introduced into the invasive plant population, which is sometimes done in an attempt to control the plants, the coevolution of plant and herbivore resumes in an altered form.
Global warming may alter relationships between plants and insect herbivores. Higher temperatures may de-synchronize the germination or leafing of the plants and the hatching of the insects. the car-bon dioxide itself may influence herbivory. carbon dioxide stimulates photosynthesis and the produc-tion of organic compounds but does not enhance the amount of nitrogen fertilizer available to the plants. Some plant defensive compounds contain nitro-gen atoms. therefore, high levels of atmospheric carbon dioxide might cause the plants to produce tissues that are more palatable to herbivores. How-ever, when the herbivores eat more and grow larger, they may attract parasites or predators that actually reduce their populations.
Herbivory is an ecological interaction that can be readily adapted to classroom studies. expertise, chemicals, and equipment are often necessary to directly measure the toxic compounds in plants—or even to know which compounds should be measured. However, there are indirect methods of measuring plant toxicity that students have found to be under-standable and interesting. Brine shrimp (Artemia
xviii+598_Biodiversity_F.indd 305 11/17/11 1:13 PM

first entry306 herbivory 306
salina) are tiny crustaceans that are often used to determine toxicity. thousands of eggs can be cheaply purchased at pet supply stores. the student needs only to add freshly hatched brine shrimp to brack-ish water that contains a plant extract, wait a day, and count how many of the shrimp have survived. From this, the lD50 can be calculated. this is the “lethal dose” concentration that kills 50 percent of the animals and is a standard measurement in toxic-ity studies. a more toxic extract will have a lower lD50. Similarly, the extract can be mixed into a stan-dardized chow which is fed to tobacco hornworm caterpillars (Manduca sexta). Both the chow and the caterpillars are available from biological supply com-panies. each caterpillar can be weighed every few days. a more toxic compound will cause the caterpil-lars to grow more slowly than a less toxic compound.
evolution has produced a prodigious variety of adaptations by which plants, as much as possible, keep animals from eating their leaves, stems, and roots and an equally prodigious variety of adapta-tions by which animals can eat the plants anyway. evolutionary innovation in plant defense stimulates evolutionary innovation in the ability of animals to get around these defenses.
See also biodiversity, resources for medi-cine; coevolution; seed dispersal.
Further readinganderson, D. “Below-ground herbivory in natural com-
munities: a review emphasizing fossorial animals.” Quarterly Review of Biology 62 (1987): 261–286.
Berenbaum, May, arthur Zangerl, and James nitao. “con-straints on chemical coevolution: Wild parsnips Pas-tinaca sativa and the parsnip webworm Depressaria pastinacella.” Evolution 40 (1986): 1,215–1,228.
choat, J. H., and K. D. clements. “Vertebrate herbivores in marine and terrestrial environments: a nutritional ecology perspective.” Annual Review of Ecology and Systematics 29 (1998): 375–403.
creelman, Robert a., and J. e. Mullet. “Jasmonic acid distribution and action in plants: Regulation during development and response to biotic and abiotic stress.” Proceedings of the National Academy of Sciences USA 92 (1995): 4,114–4,119.
Dearing, M. Denise, et al. “the influence of plant secondary metabolites on the nutritional ecology of herbivorous terrestrial vertebrates.” Annual Review of Ecology, Evolution, and Systematics 36 (2005): 169–189.
Donaldson, J. R., and Richard l. lindroth. “Genetics, environment, and their interaction determine efficacy of chemical defense in trembling aspen.” Ecology 88 (2007): 729–739.
Feeny, Paul. “Seasonal changes in oak leaf tannins and nutrients as a cause of spring feeding by winter moth caterpillars.” Ecology 51 (1970): 565–581.
Hanhimäki, Sinikka. “induced resistance in mountain birch: Defense against leaf-chewing insect guild and herbi-vore competition.” Oecologia 81 (1989): 242–248.
Knepp, R. G., et al. “elevated cO2 reduces leaf damage by insect herbivores in a forest community.” New Phy-tologist 167 (2005): 207–218.
lambert, e. c., et al. “tritrophic effects of xanthotoxin on the polyembryonic parasitoid Copidosoma sosares (Hymenoptera: encyrtidae). Journal of Chemical Ecol-ogy 34 (2008): 783–789.
lewinsohn, thomas M., et al. “insects on plants: Diversity of herbivore assemblages revisited.” Annual Review of Ecology, Evolution, and Systematics 36 (2005): 597–620.
núñez-Farfán, Juan, et al. “the evolution of resistance and tolerance in herbivores.” Annual Review of Ecology, Evolution, and Systematics 38 (2007): 541–566.
Ohgushi, takayuki. “indirect interaction webs: Herbivore-induced effects through trait change in plants.” Annual Review of Ecology, Evolution, and Systematics 36 (2005): 81–105.
Peacock, l., and S. Herrick. “Responses of the willow bee-tle Phratora vulgatissima to genetically and spatially diverse Salix spp. plantations.” Journal of Applied Ecology 37 (2000): 821–831.
Rice, Stanley a., and ian B. Maness. “Brine shrimp bioas-says: a useful technique in biological investigations.” American Biology Teacher 66 (2004): 208–215.
———, and Jennifer R. Griffin. “the hornworm assay: Useful in mathematically-based biological investiga-tions.” American Biology Teacher 66 (2004): 487–491.
Salminen, Juha-Pekka, et al. “Seasonal variation in the con-tent of hydrolyzable tannins, flavonoid glycosides, and proanthocyanidins in oak leaves.” Journal of Chemical Ecology 30 (2004): 1,693–1,711.
Schultz, Jack c. “Seasonal and individual variation in leaf quality of two northern hardwoods tree species, sugar maple (Acer saccharum) and yellow birch (Bet-ula allegheniensis). American Journal of Botany 69 (1982): 753–759.
Seldal, tarald, et al. “Wound-induced proteinase inhibitors in gray alder (Alnus incana): a defense mechanism against attacking insects.” Oikos 71 (1994): 239–245.
Stiling, Peter, et al. “elevated cO2 lowers relative and abso-lute herbivore density across all species of a scrub-oak forest.” Oecologia 134 (2003): 82–87.
———, and tatiana cornelissen. “How does elevated carbon dioxide (cO2) affect plant-herbivore interac-tions? a field experiment and meta-analysis of cO2-mediated changes on plant chemistry and herbivore performance.” Global Change Biology 13 (2007): 1,823–1,842.
tscharntke, teja, et al. “Herbivory, induced resistance, and interplant signal transfer in Alnus glutinosa.” Biochemical Systematics and Ecology 29 (2001): 1,025–1,047.
xviii+598_Biodiversity_F.indd 306 11/17/11 1:13 PM

hybridization 307
Zangerl, arthur R., and May R. Berenbaum. “effects of florivory on floral volatile emissions and pollination success in the wild parsnip.” Arthropod-Plant Interac-tions 3 (2009): 181–191.
——— and ———. “Parsnip webworms and host plants at home and abroad: trophic complexity in a geographic mosaic.” Ecology 87 (2006): 3,070–3,081.
———, et al. “Selection for chemical trait remixing in an invasive weed after reassociation with a coevolved specialist.” Proceedings of the National Academy of Sciences USA 105 (2008): 4,547–4,552.
holocene epoch See neogene period, bio-diversity in.
humboldt, alexander von (1769–1859) German Naturalist and explorer Born Septem-ber 14, 1769, in Berlin, Baron Friedrich Wilhelm Heinrich alexander Freierr von Humboldt was one of the most important scientists of the late 18th and early 19th centuries. He began the modern study of biogeography. at an early age, he had a passion for studying plants and animals. at the University of Frankfurt, he studied subjects from commerce to politics to geology. though he rose to a high position as a geologist as a government assessor of mines, he resigned in order to travel and conduct scientific investigation.
in 1799, he began a scientific journey through latin america, including Mexico, the caribbean, and western South america, with French botanist aimé Bonpland. He traveled 1,725 miles (2,776 km) in the amazon Basin, where he was the first european to (painfully) discover the electric eel. He climbed mountains in the andes, nearly reach-ing the top of the 20,565-foot (6,268-m) chimbo-razo volcano in ecuador. He collected plants and, with Bonpland, was the first scientist to realize that temperature patterns (due both to latitude and to elevation) largely determined the distribution of plant species. throughout this expedition he made detailed measurements of temperature and magnetic fields with the finest scientific equipment available at the time, one of the first scientists to insist on such precision. Such measurements allowed him to discover new ocean currents, including one now called the Humboldt current (off the west coast of South america), and that earth’s magnetic field was stronger at the poles and weaker near the equator. after meeting President thomas Jefferson, Hum-boldt returned to Germany in 1804.
Humboldt analyzed his data and published many books about his journey over the next 21 years. as he did so, he proposed that South amer-
ica and africa had once been joined into a single continent, a theory not fully vindicated until con-tinental drift was accepted a century and a half later. then he turned his thought to the unifica-tion of all branches of scientific knowledge in his five-volume Kosmos, which he began to publish in 1845. His thought was strongly influenced by the romantic movement, which idealized nature as being perfect apart from human interference. Because of Humboldt’s enormous influence in science, scientists until recently tended to ignore the important role that native peoples have had in determining natural ecological communities. While writing his books, Humboldt traveled little, until he took a 9,614-mile (15,472-km) journey through Russia in 1829.
Humboldt’s insights and his bold adventures inspired many scientists, most notably charles Dar-win, who saw himself as retracing Humboldt’s jour-ney when he visited South america. Darwin said of Humboldt, “i have always admired him. now i wor-ship him.” Humboldt died in Berlin on May 6, 1859.
See also biogeography; Darwin, charles.
Further readingHumboldt, alexander von. Personal Narrative of a Journey
to the Equinoctial Regions of the New Continent. abridged edition by Jason Wilson. new york: Pen-guin, 1996.
———, and aimé Bonpland. Essay on the Geogra-phy of Plants. Stephen t. Jackson, ed., and Sylvie Romanowski, transl. chicago: University of chicago, 2010.
lack, H. Walter. Alexander von Humboldt: The Botanical Exploration of the Americas. new york: Prestel, 2009.
Rupke, nicolaas a. Alexander von Humboldt: A Metabi-ography. chicago: University of chicago, 2008.
Walls, laura Dassow. The Passage to Cosmos: Alexander von Humboldt and the Shaping of America. chicago: University of chicago, 2009.
hutchinson, g. evelyn See competition, ecological.
hybridization Hybridization is the crossbreed-ing of different species of organisms. the term hybrid has also been used for crosses between vari-eties within a species (as in hybrid corn within the species Zea mays). the offspring with many hetero-zygous traits produced by intraspecific crosses may produce hybrid vigor. However, conservation and evolutionary biologists generally restrict the term hybrid to crosses between species.
according to the biological species concept, distinct species do not normally produce fertile
xviii+598_Biodiversity_F.indd 307 11/17/11 1:13 PM

first entry308 hybridization 308
offspring. this is most obviously the case with crosses between donkeys and horses, resulting in mules, which are very strong but are sterile. crosses between lions and tigers (such as ligers and tigons) have also occurred in zoos (see table). Many other hybrids are possible between feline species. While even fertile hybrids may be pro-duced between different feline species, they are rare in the wild, which suggests that either the spe-cies nearly always avoid one another when mating and/or the hybrid offspring have reduced health or fertility. Hybrids also occur among canine spe-cies. For example, black coat color in forest wolves (Canis lupus) has been traced to hybridization with domesticated dogs (Canis familiaris), a spe-cies that recently evolved from wolves. Hybrids have also been reported between species of fishes. the bluegill (Lepomis macrochirus) can cross with the green sunfish (L. cyanellis). intergeneric hybrids, such as between pumas and leopards and between dromedaries and llamas, have also been reported (see table on page 309). When a scientific name is assigned to a hybrid species, an x often precedes the specific name.
Hybrids between plant species in the same genus occur more often than hybrids between animal spe-cies. Hybridization in plants is not usually success-ful, for at least two reasons:
● the two species may pollinate at differ-ent times of the year. among the oaks of north america, hybridization can occur among species of white oaks and among species of red and black oaks (genus Quer-cus). However, all of the white oaks have acorns that mature during the same year, while most of the red and black oaks have acorns that mature the following year, after pollination. Because of these dif-ferences in reproductive pattern, hybrids between the two groups of oaks cannot occur. Hybridization between species of pine (the bishop pine Pinus muricata and the Monterey pine P. radiata) and between species of alders (the seaside alder Alnus maritima and the hazel alder A. serrulata) cannot occur, as pollination occurs at dif-ferent times of the year.
interspeCifiC Crosses in animals and plants (Within genera)
parent parent hybrid
Jackass (Equus asinus) Mare (E. caballus) Mule
Jenny (Equus asinus) Stallion (E. caballus) Hinny
Male tiger (Panthera tigris) Lioness (P. leo) Tigon
Tigress (Panthera tigris) Male lion (P. leo) Liger
Leopard (Panthera pardus) Lion (P. leo) Leopon
Bluegill (Lepomis macrochirus) Green sunfish (L. cyanellis) Hybrid sunfish
Larkspur (Delphinium hesperium) Larkspur (D. recurvatum) D. gypsophilium
Penstemon (Penstemon centrantifolius) Penstemon (P. grinnellii) P. spectabilis
Japanese larch (Larix kaempferi) European larch (L. decidua) Dunkeld larch
Magnolia (Magnolia denudata) Magnolia (M. liliiflora) M. soulangeana
American sycamore (Platanus occidentalis) European (P. orientalis) London plane
Elm (Ulmus glabra) Elm (U. minor) U. x hollandica
Wingnut (Pterocarya fraxinifolia) Wingnut (P. stenoptera) P. x rehderiana
Linden (Tilia cordata) Linden (T. platyphyllos) T. x europaea
White poplar (Populus alba) Aspen (P. tremula) Gray poplar
Arbutus (Arbutus unedo) Arbutus (A. andrachne) A. x andrachnoides
Horse chestnut (Aesculus hippocastanum) Horse chestnut (A. pavia) A. x carnea
xviii+598_Biodiversity_F.indd 308 11/17/11 1:13 PM
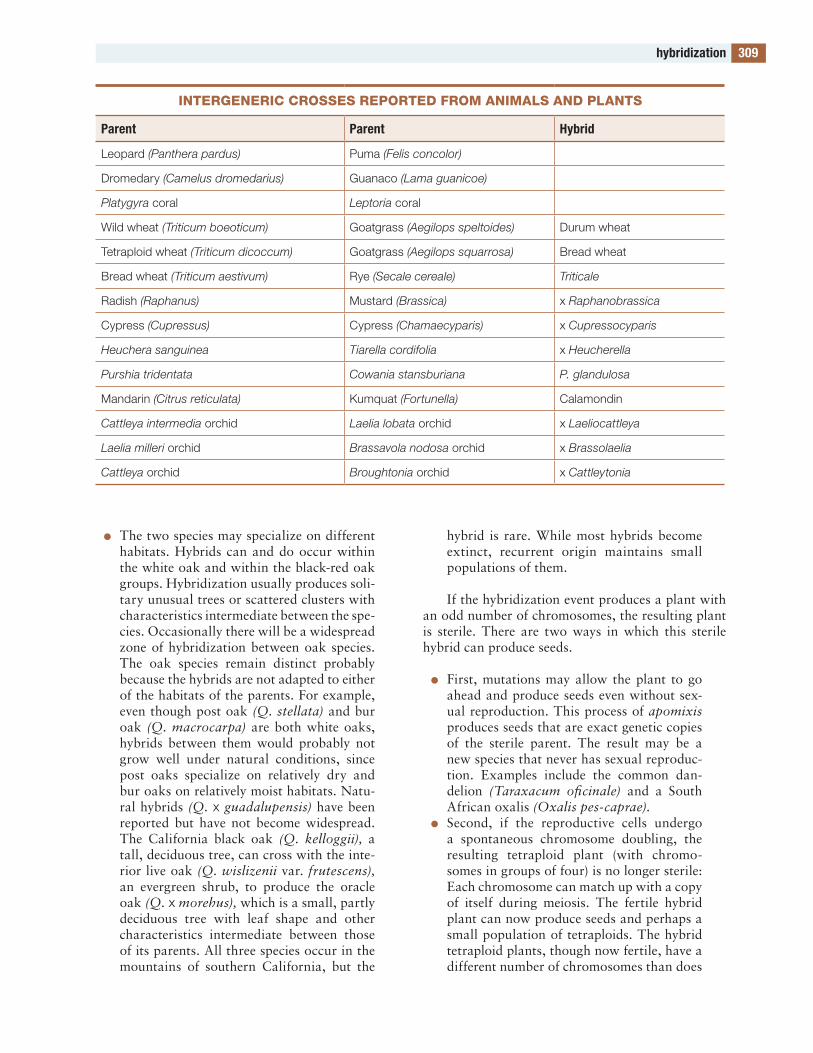
hybridization 309
● the two species may specialize on different habitats. Hybrids can and do occur within the white oak and within the black-red oak groups. Hybridization usually produces soli-tary unusual trees or scattered clusters with characteristics intermediate between the spe-cies. Occasionally there will be a widespread zone of hybridization between oak species. the oak species remain distinct probably because the hybrids are not adapted to either of the habitats of the parents. For example, even though post oak (Q. stellata) and bur oak (Q. macrocarpa) are both white oaks, hybrids between them would probably not grow well under natural conditions, since post oaks specialize on relatively dry and bur oaks on relatively moist habitats. natu-ral hybrids (Q. x guadalupensis) have been reported but have not become widespread. the california black oak (Q. kelloggii), a tall, deciduous tree, can cross with the inte-rior live oak (Q. wislizenii var. frutescens), an evergreen shrub, to produce the oracle oak (Q. x morehus), which is a small, partly deciduous tree with leaf shape and other characteristics intermediate between those of its parents. all three species occur in the mountains of southern california, but the
hybrid is rare. While most hybrids become extinct, recurrent origin maintains small populations of them.
if the hybridization event produces a plant with an odd number of chromosomes, the resulting plant is sterile. there are two ways in which this sterile hybrid can produce seeds.
● First, mutations may allow the plant to go ahead and produce seeds even without sex-ual reproduction. this process of apomixis produces seeds that are exact genetic copies of the sterile parent. the result may be a new species that never has sexual reproduc-tion. examples include the common dan-delion (Taraxacum oficinale) and a South african oxalis (Oxalis pes-caprae).
● Second, if the reproductive cells undergo a spontaneous chromosome doubling, the resulting tetraploid plant (with chromo-somes in groups of four) is no longer sterile: each chromosome can match up with a copy of itself during meiosis. the fertile hybrid plant can now produce seeds and perhaps a small population of tetraploids. the hybrid tetraploid plants, though now fertile, have a different number of chromosomes than does
intergeneriC Crosses reported from animals and plants
parent parent hybrid
Leopard (Panthera pardus) Puma (Felis concolor)
Dromedary (Camelus dromedarius) Guanaco (Lama guanicoe)
Platygyra coral Leptoria coral
Wild wheat (Triticum boeoticum) Goatgrass (Aegilops speltoides) Durum wheat
Tetraploid wheat (Triticum dicoccum) Goatgrass (Aegilops squarrosa) Bread wheat
Bread wheat (Triticum aestivum) Rye (Secale cereale) Triticale
Radish (Raphanus) Mustard (Brassica) x Raphanobrassica
Cypress (Cupressus) Cypress (Chamaecyparis) x Cupressocyparis
Heuchera sanguinea Tiarella cordifolia x Heucherella
Purshia tridentata Cowania stansburiana P. glandulosa
Mandarin (Citrus reticulata) Kumquat (Fortunella) Calamondin
Cattleya intermedia orchid Laelia lobata orchid x Laeliocattleya
Laelia milleri orchid Brassavola nodosa orchid x Brassolaelia
Cattleya orchid Broughtonia orchid x Cattleytonia
xviii+598_Biodiversity_F.indd 309 11/17/11 1:13 PM

first entry310 hybridization 310
either parent species. therefore it cannot crossbreed with either of its parent species; it can only cross-pollinate with other mem-bers of the tetraploid population. the tet-raploid population, reproductively isolated from the parental species, now functions as a new species.
examples of spontaneous chromosome doubling that has restored the fertility of the reproductive cells of the sterile plant are salt grasses, tobaccos, mus-tards, and sunflowers:
● cross-pollination between Spartina mari-tima (native to europe) and S. alterniflora (native to california), two species of salt-marsh grasses, occurred when S. alterniflora was introduced into england. the hybrid, S. x townsendii, was sterile. chromosome doubling in the sterile hybrid resulted in S. anglia, which now grows vigorously in english salt marshes.
● the common tobacco Nicotiana tabacum (with 48 chromosomes) can cross with another tobacco species, N. glutinosa. However, N. glutinosa has only 24 chro-mosomes. a sterile hybrid, with 36 chromo-somes, results. Spontaneous chromosome doubling can occur in this sterile hybrid, resulting in seeds with 72 chromosomes. the result is the hybrid species N. digluta.
● Hybridization among three mustard species in the wild has produced three new species.
● the sunflowers Helianthus deserticola and H. anomalus have been shown to have hybrid origins. Botanist loren Rieseberg has experimentally replicated the hybrid ori-gin of H. anomalus in the greenhouse. By crossing H. annuus with H. petiolaris, Ries-eberg produced three independent hybrid lines. Only four generations were required to select vigorous hybrids. the genetic markers of the greenhouse hybrids closely resemble those in the wild populations of H. anomalus.
in plants, intergeneric hybrids (while still rare) are more common than in animals. Russian botanist Georgi Karpechenko crossed the radish Raphanus sativus with the cabbage Brassica oleracea, produc-ing a Raphanobrassica hybrid in 1927. Unfortu-nately, the hybrid had the leaves of radish and the roots of cabbage. Wild Raphanobrassica hybrids have been reported from california. Triticale, a hybrid of wheat and rye, is reported to have the high yield of wheat and the hardiness of rye but is rarely
grown on a commercial scale. Hybrids between plant genera are known from orchids and cypresses as well (see table on page 309). among orchids, hybrids are possible among three or even more genera, although this almost always occurs under artificial conditions. in natural habitats, pollinator specificity usually pre-vents extensive crossings among orchids. For exam-ple, Brassolaeliocattleya is a cross among the genera Brassavola, Laelia, and Cattleya. When a scientific name is assigned to an intergeneric hybrid, the x may precede the generic name.
natural hybrids tend to be rare, but hybrid plants produced for horticultural purposes can be vigorous. the reduced reproductive capacity of the horticultural hybrid is not a drawback because arti-ficial propagation is available.
Perhaps the plant hybridizations that are most famous in the history of the world are the crosses that produced modern wheat. the wild wheat Trit-icum boeoticum had chromosomes in pairs (dip-loid, 14 chromosomes) and was bred by ancient farmers into the cultivated einkorn wheat Triticum monococcum. However, T. boeoticum accidentally cross-pollinated with the wild goatgrass Aegilops speltoides. the chromosomes from the two parents were incompatible, but chromosome doubling pro-duced a fertile hybrid with chromosomes in groups of four (tetraploid, with 28 chromosomes), the wild T. dicoccoides. this was bred by ancient farmers into the emmer wheat T. dicoccum and into durum (T. durum) and other wheats. T. dicoccum also acci-dentally cross-pollinated with a wild goatgrass, this time A. squarrosa. chromosome doubling turned a sterile hybrid into a species of wheat with chromo-somes in groups of six (hexaploid, 42 chromosomes) that are today’s major wheat species T. spelta (spelt) and T. aestivum (bread wheat) (see table on page 309). One of the distinguishing features of modern bread wheat is the gluten protein, which makes the flour sticky, allowing it to hold in bubbles of carbon dioxide during leavening. the gene for gluten appar-ently came not from the Triticum ancestor but from the Aegilops squarrosa weed with which emmer wheat accidentally hybridized.
Hybridization, aside from producing new plant species, can also facilitate the transfer of genes from one species of plant to another. consider two plant species that hybridize and produce a new hybrid spe-cies. the two original plant species seldom hybridize with one another but may be able to crossbreed more often with the intermediate hybrid species. Genes from the first plant species can enter into the hybrid population by cross-pollination and can then enter into the population of the second plant species, also by cross-pollination. the hybrid species has thus acted as a bridge over which genes have crossed, or
xviii+598_Biodiversity_F.indd 310 11/17/11 1:13 PM

hydrological cycle 311
introgressed, from one species into another. this concept of introgressive hybridization or introgres-sion was first suggested by edgar anderson, an early 20th-century expert on the genetic history of crop species.
the relative rarity of hybridization between spe-cies suggests that a new species, once it evolves, has a coordinated team of genes that would be disrupted by mixing with a different set of genes. this may explain why evolutionary changes may occur so rap-idly when a species first forms, followed by a period of relative stability.
Hybridization raises particularly interesting questions about the conservation status of endan-gered species. are the hybrids, one or both of whose parents may be endangered, also protected? While artificial hybrids are not a concern for conservation biologists, hybrids that occur in nature as a result of human activity, such as global warming, may deserve protection. For example, what about the grolar bear? First reported in northern canada in 2006, this hybrid between a grizzly bear and a polar bear may have come about because global warming has encouraged grizzly bears to migrate northward into polar bear territory. the man who shot the gro-lar bear was worried that he had violated the terms of his hunting license, which permitted him to hunt only one of the parental species.
See also speciation.
Further readinganderson, tovi M., et al. “Molecular and evolutionary
history of melanism in north american gray wolves.” Science 323 (2009): 1,339–1,343.
Gompert, Zachariah, et al. “Homoploid hybrid specia-tion in an extreme habitat.” Science 314 (2006): 1,923–1,925.
Kadereit, J. W., et al. “Reciprocal hybridization at differ-ent times between Senecio flavus and Senecio glaucus gave rise to two polyploid species in north africa and south-west asia.” New Phytologist 169 (2006): 431–441.
Miller, Howard, and Samuel lamb. Oaks of North Amer-ica. Happy camp, calif.: naturegraph Publishers, 1985.
Pfennig, Karin S. “Facultative mate choice drives adaptive hybridization.” Science 318 (2007): 965–967.
Schwarzbach, andrea, lisa a. Donovan, and loren H. Rieseberg. “transgressive character expression in a hybrid sunflower species.” American Journal of Bot-any 88 (2001): 270–277.
hydrological cycle also known as the water cycle, the hydrological cycle is the movement of water molecules through the ecosystems of the earth. the
details of this cycle have an overwhelming impact on which ecological communities live in which loca-tions and on the effects of global warming.
Oceans. about 98 percent of the water on earth is oceanic salt water. another 1.6 percent is ice, with the remaining 0.4 percent freshwater on land surfaces. the volume of water in the oceans does not remain constant. During periods of global warming, ocean levels rise for two reasons. First, the water expands. Second, ice on land surfaces melts. these two processes contribute about equally to sea-level rise during warm periods. During the most recent ice age, ocean levels were more than 300 feet (about 100 m) lower than they are today.
the oceans also have a profound effect on global temperatures. Oceans absorb heat during warm periods and release it during cold periods of history, although this process is too slow to counter-act the current crisis of global warming. land areas near oceans have much less temperature fluctuation during the year than do land areas far from oceans. this is the reason that the winters are warmer and the summers cooler in Seattle than in Minneapolis, despite their similar latitude and amount of heat they receive from the Sun. Ocean currents redistrib-ute heat around the world. One important example is the Gulf Stream, part of a larger “global con-veyor” system of ocean currents. the warm Gulf Stream waters carry heat from the caribbean toward europe. this is the reason that the climate is warmer in england than in labrador, even though both are at similar latitudes.
Evaporation and condensation. evaporation from the ocean surfaces is the major source of water vapor in the atmosphere.
Water vapor and clouds also have a profound effect on global temperatures. Water vapor is a green-house gas. When condensation forms clouds, these clouds have two effects on heat. First, the albedo (reflectivity) of clouds sends much solar energy right back into outer space, preventing it from heating the earth. Second, clouds absorb and emit infrared light, thus preventing the earth’s surface from emit-ting infrared light into outer space. clouds, there-fore, both cool and warm the earth. Different kinds of clouds have different relative amounts of heating and cooling effects on the earth. clouds are the principal method by which water that evaporates from oceans is transported over land surfaces.
Precipitation. at the tops of clouds, water droplets may be supercooled, that is, remain liquid below the freezing point. When a droplet receives a freezing nucleus (for example, a dust or sulfate particle), it freezes. Frozen water can fall in the form of snow, hail, or sleet. Most raindrops are ice that melted on the way down.
xviii+598_Biodiversity_F.indd 311 11/17/11 1:13 PM

fi rst entry312 hydrological cycle 312
the biodiversity and ecological communities of the earth are primarily infl uenced by temperature and precipitation. terrestrial ecological communi-ties with more precipitation have greater productiv-ity and more species. thousands of years ago, the Sahara received more rainfall and was home to many more plant and animal species. the annual timing of precipitation is also important. the highest spe-cies diversity is in the tropical rain forests, where rainfall is uniform. Snow and ice that accumulate in the winter may melt in the spring, supplying water to environments that would otherwise experience a dry summer. Dry summers would probably cause the mountains of western north america to have less forest growth if it were not for the winter snowpack.
Groundwater and runoff. Rain and snow that enter the ground slowly may percolate down into shallow aquifers, which feed creeks and rivers with a relatively uniform fl ow of water. By intercepting raindrops, vegetation cover allows much more water to percolate into the soil than does bare soil. Runoff
water fl oods quickly into creeks and rivers, often carrying much soil with it and occasionally caus-ing mudslides (see fi gure). thicker vegetation cover reduces the amount of runoff (see table on page 313). the fact that deforestation increases runoff has been experimentally demonstrated on whole mountainsides, such as the Hubbard Brook water-shed in new Hampshire where ecologist Gene lik-ens and associates clear-cut some mountain slopes and left adjacent ones intact. Similar results have been found at the coweeta watershed in the south-ern appalachians.
Reservoirs delay the fl ow of runoff into riv-ers and oceans. Reservoirs allow humans to have water for irrigation and hydroelectricity. they do not appear to signifi cantly increase humidity; the mountains near reservoirs in the desert Southwest appear no different from those farther away. the impoundment of runoff in reservoirs has measurably reduced the amount of sea-level rise that would oth-erwise have occurred with global warming.
Deforestation allows rainwater to rush down the mountain slope and cause fl oods, rather than percolating down into the soil. Transpiration from forests also puts a great deal of water vapor into the air.
xviii+598_Biodiversity_F.indd 312 11/17/11 1:13 PM
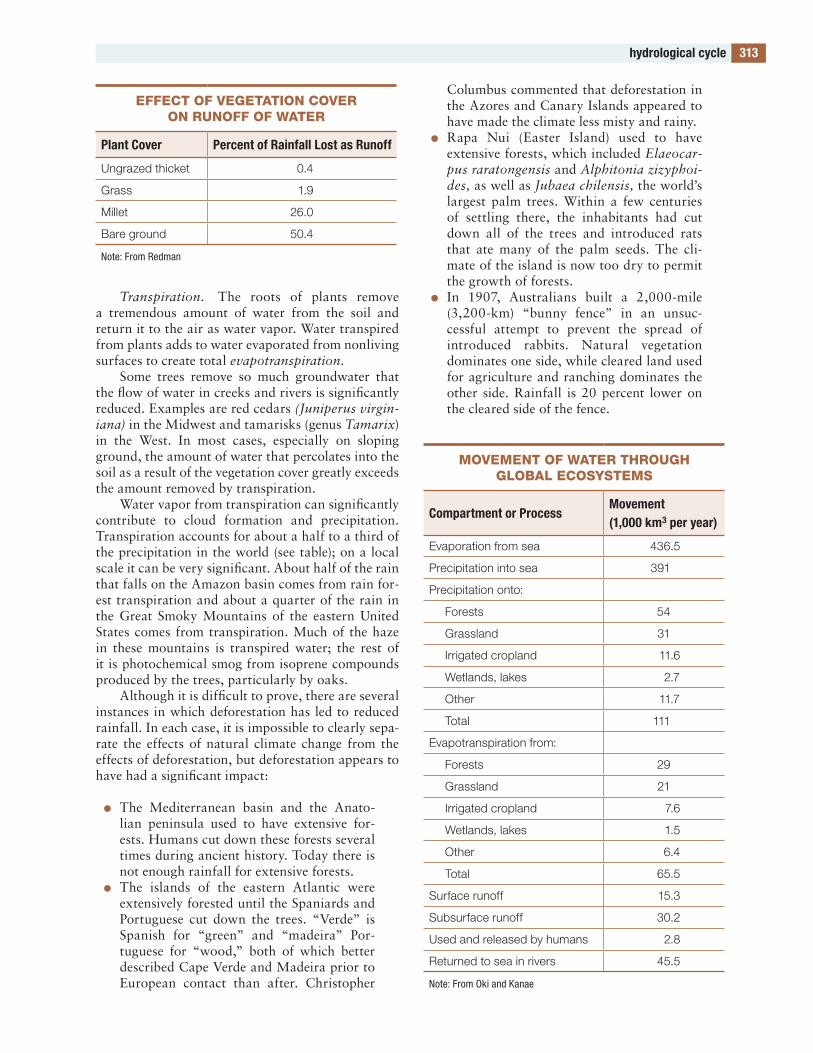
hydrological cycle 313
Transpiration. the roots of plants remove a tremendous amount of water from the soil and return it to the air as water vapor. Water transpired from plants adds to water evaporated from nonliving surfaces to create total evapotranspiration.
Some trees remove so much groundwater that the flow of water in creeks and rivers is significantly reduced. examples are red cedars (Juniperus virgin-iana) in the Midwest and tamarisks (genus Tamarix) in the West. in most cases, especially on sloping ground, the amount of water that percolates into the soil as a result of the vegetation cover greatly exceeds the amount removed by transpiration.
Water vapor from transpiration can significantly contribute to cloud formation and precipitation. transpiration accounts for about a half to a third of the precipitation in the world (see table); on a local scale it can be very significant. about half of the rain that falls on the amazon basin comes from rain for-est transpiration and about a quarter of the rain in the Great Smoky Mountains of the eastern United States comes from transpiration. Much of the haze in these mountains is transpired water; the rest of it is photochemical smog from isoprene compounds produced by the trees, particularly by oaks.
although it is difficult to prove, there are several instances in which deforestation has led to reduced rainfall. in each case, it is impossible to clearly sepa-rate the effects of natural climate change from the effects of deforestation, but deforestation appears to have had a significant impact:
● the Mediterranean basin and the anato-lian peninsula used to have extensive for-ests. Humans cut down these forests several times during ancient history. today there is not enough rainfall for extensive forests.
● the islands of the eastern atlantic were extensively forested until the Spaniards and Portuguese cut down the trees. “Verde” is Spanish for “green” and “madeira” Por-tuguese for “wood,” both of which better described cape Verde and Madeira prior to european contact than after. christopher
columbus commented that deforestation in the azores and canary islands appeared to have made the climate less misty and rainy.
● Rapa nui (easter island) used to have extensive forests, which included Elaeocar-pus raratongensis and Alphitonia zizyphoi-des, as well as Jubaea chilensis, the world’s largest palm trees. Within a few centuries of settling there, the inhabitants had cut down all of the trees and introduced rats that ate many of the palm seeds. the cli-mate of the island is now too dry to permit the growth of forests.
● in 1907, australians built a 2,000-mile (3,200-km) “bunny fence” in an unsuc-cessful attempt to prevent the spread of introduced rabbits. natural vegetation dominates one side, while cleared land used for agriculture and ranching dominates the other side. Rainfall is 20 percent lower on the cleared side of the fence.
effeCt of vegetation Cover on runoff of Water
plant Cover percent of rainfall lost as runoff
Ungrazed thicket 0.4
Grass 1.9
Millet 26.0
Bare ground 50.4
Note: From Redman
movement of Water through global eCosystems
Compartment or processmovement (1,000 km3 per year)
Evaporation from sea 436.5
Precipitation into sea 391
Precipitation onto:
Forests 54
Grassland 31
Irrigated cropland 11.6
Wetlands, lakes 2.7
Other 11.7
Total 111
Evapotranspiration from:
Forests 29
Grassland 21
Irrigated cropland 7.6
Wetlands, lakes 1.5
Other 6.4
Total 65.5
Surface runoff 15.3
Subsurface runoff 30.2
Used and released by humans 2.8
Returned to sea in rivers 45.5
Note: From Oki and Kanae
xviii+598_Biodiversity_F.indd 313 11/17/11 1:13 PM

first entry314 hydrological cycle 314
it is clear that ecological communities not only depend upon precipitation but that they influence it. By destroying vegetation cover, humans may be significantly altering the amount of precipitation in some areas. Global warming will have yet further effects on the hydrological cycle.
See also communities, ecological; ecosystems.
Further readingBarnett, tim P., et al. “Human-induced changes in the
hydrology of the western United States.” Science 319 (2008): 1,080–1,083.
Bosch, J. M., and J. D. Hewlett. “a review of catchment experiments to determine the effect of vegetation changes on water yield and evapotranspiration.” Jour-nal of Hydrology 103 (1982): 232–333.
chao, B. F., et al. “impact of artificial reservoir water impoundment on global sea level.” Science 320 (2008): 212–214.
costa, Marcos Heil, et al. “effects of large-scale changes in land cover on the discharge of the tocantins River, southeastern amazonia.” Journal of Hydrology 283 (2003): 206–217.
coweeta long term ecological Research. “coweeta long term ecological Research.” available online. URl: http://coweeta.ecology.uga.edu. accessed June 25, 2010.
Dransfield, J., et al. “a recently extinct palm from easter island.” Nature 312 (1984): 750–752.
Gordon, line J., et al. “Human modification of global water vapor flows from the land surface.” Proceed-ings of the National Academy of Sciences 102 (2005): 7,612–7,617.
Hubbard Brook Research Foundation. “Hubbard Brook ecosystem study.” available online. URl: http://www.hubbardbrook.org. accessed February 27, 2011.
Kerr, Richard a. “another side to the climate-cloud conun-drum finally revealed.” Science 319 (2008): 889.
Kröpelin, S., et al. “climate-driven ecosystem succession in the Sahara: the past 6000 years.” Science 320 (2008): 765–768.
likens, Gene e., et al. “effects of forest cutting and her-bicide treatment on nutrient budgets in the Hubbard Brook watershed-ecosystem.” Ecological Monographs 40 (1970): 23–47.
noticewala, Sonal. “at australia’s bunny fence, variable cloudiness prompts climate study.” New York Times, august 14, 2007. available online. URl: http://www.nytimes.com/2007/08/14/science/earth/14fenc.html. accessed august 8, 2011.
Oki, taikan, and Shinjiro Kanae. “Global hydrological cycles and world water resources.” Science 313 (2006): 1,068–1,072.
Redman, charles l. Human Impact on Ancient Environ-ments. tucson: University of arizona Press, 2001.
xviii+598_Biodiversity_F.indd 314 11/17/11 1:13 PM

315
ice ages the ice ages are recurring periods of glaciation that began during the Pleistocene epoch of the Quaternary period. During each ice age, glaciers advanced from the north across north america and eurasia. an interglacial period followed each gla-ciation, in which the glaciers melted and retreated. each ice age glaciation plus its subsequent intergla-cial period lasted about 100,000 years. therefore, during the approximately 2 million years of the Pleistocene, about 20 ice ages occurred, with the Holocene epoch consisting of the time since the peak of the most recent glaciation. in many cases, a more recent glaciation has obliterated the geologi-cal evidences of an earlier one. Several of the more extensive glaciations can be directly studied from geological deposits.
the Quaternary period has been one of the cold-est in all of earth’s history, but glaciations also occurred many times in the past. For example, gla-ciations occurred in the southern continent of Gond-wana 355 to 280 million years ago and in antarctica beginning 33 million years ago. Perhaps the most extensive ice ages occurred during the Precambrian time. three times, the earth nearly froze into a per-manent ball of ice popularly called Snowball earth. this entry focuses upon the recurring northern Hemisphere ice ages of the Quaternary period.
By the middle of the 19th century, scientists (led by geologist louis agassiz) had determined that there had been at least one ice age in the northern Hemisphere. But nobody understood what caused them. During the 1860s, scientifi c journals received papers about physics from a certain James croll of anderson’s University in Glasgow. One of these papers suggested that variations in the orbit of the earth caused ice ages to come and go. the calcu-
lations were oversimplifi ed but provided a major insight that turned out to be correct. croll was not a professor, but a janitor who spent a lot of time in the library. the details of how the variations in the earth’s orbit—a combination of changes in the distance from the Sun and the tilt of the earth—caused the ice ages were worked out by the Serbian mathematician Milutin Milanković. the dates that emerged from Milanković’s calculations for when ice ages were most likely to have occurred did not cor-respond to what was then believed to be the times of glaciation. therefore by the time Milanković died he did not have the satisfaction of seeing his theory vin-dicated. now that scientists have radiometric dates of several of the glaciations, they see that Milanković was largely correct.
Variations in earth’s angle and orbit affect the ice ages because they shift the balance of solar energy going into and out of the planet. Short, cool summers resulting from less solar energy—because of a com-bination of distance from the Sun and angle of the sunlight—reduce snowmelt. the increased area cov-ered with snow refl ects more light into outer space, increasing the albedo (refl ected light) of the planet. this further shifts the balance of climate toward an ice age. the accumulation of ice and snow in the northern latitudes eventually pushes the ice south-ward in great rivers of snow (glaciers). if the summers in the latitudes farther south are cool enough that the ice does not melt as much as it advances, then the result is glaciation. the rate of advance was variable but has seldom exceeded three feet (one meter) a year. the return of warmer summers results in glaciers melting more than they advance.
the relatively slight differences in solar radia-tion seem inadequate to explain the drastic changes
I
xviii+598_Biodiversity_F.indd 315 11/17/11 1:13 PM

first entry316 ice ages 316
in temperature that characterize the coming and going of ice ages. But these slight changes in temper-ature set in motion a cascade of effects that amplify glaciation and then amplify the melting of glaciers. as snow begins to accumulate in the north, more sunlight is reflected back into outer space (albedo), which creates even colder conditions, speeding up glaciation. cooler temperatures also reduce the decomposition of dead biomass, reducing the green-house effect. Once the glaciers begin to melt, both albedo and greenhouse effect amplify the onset of the interglacial period.
the most recent maximum glacial extent ended about 14,000 years ago. the arctic Ocean was rela-tively ice-free during the glaciations and was the source of evaporated water for much of the snow that caused them. extensive glaciations did not occur in the Southern Hemisphere, because it contains mostly ocean, which does not change temperature as readily as land. Glaciers covered and still cover antarctica, but there is very little land surface between antarc-tica and the tip of South america. therefore ant-arctica produced, and produces, separate icebergs rather than advancing glaciers.
the Milanković cycle explains why cyclical gla-ciations occur today but does not explain why they did not occur prior to the Pleistocene. the land that is now north and South america had not been connected since the breakup of Pangaea during the Mesozoic era. a gap of ocean between them allowed a vast worldwide circulation of ocean water to flow into the atlantic Ocean, forcing warm water into the arctic Ocean. Water holds more heat than air, and this movement of heat from tropics to the arctic caused temperatures to be sufficiently warm that trees grew right up to the north Pole, even though they were in darkness half of the year. However, the isthmus of Panama gradually arose, first as islands that slowed the flow of water, then as a land bridge that stopped it. today, the Gulf Stream carries warm caribbean water to the north atlantic, causing the climate of europe to be much warmer than the land at the same latitudes in north america (compare labrador and england, for example). But the Gulf Stream is not big enough to warm up the arctic Ocean. the planetary movements that produce the Milanković cycle occurred long before the Pleisto-cene, but until then they had not produced cyclical glaciations.
as the glaciers advanced, they pushed piles of rubble ahead of them. these piles, after the glaciers began to retreat, formed moraines. Some moraines have resulted in major geographical landmarks, such as long island and cape cod. Glaciers tore large chunks of rock from some mountains, while going around others. the isolated mountains that rose
above the sheet of ice were called nunataks. Some-times whole mountain systems, such as the Black Hills of South Dakota, remained above the glaciers. the temperatures were sufficiently cold and dry that very few organisms survived on nunataks.
Glaciations during the Quaternary ice ages influenced the climate of north america very severely. Most of the same vegetation zones that exist today were present during the glaciation, including tropical rain forests, seasonal forests, and savannas. However, because the glaciers came so far southward, the northern vegetation zones were thinner and compressed toward the equator (see figure on page 317). the tropical areas, though slightly cooler, were still warm. However, since so much of the water was removed from meteorologi-cal circulation and stored in glaciers, the global cli-mate was drier. in the dry tropical regions, deserts and grasslands were extensive, while tropical forests consisted of isolated groves. Because so much water was frozen in the glaciers, sea level was almost 300 feet (100 m) lower than it is today. Much of the con-tinental shelf, which today is under shallow water, was dry land. Some “fishing banks” of shallow water in the oceans were exposed islands or penin-sulas during the most recent glaciation. Some plant communities that have no counterpart today, such as spruce savannas, existed during the glaciation.
the most recent glacial retreat began about 12,000 years ago. Retreat was generally faster than advance, sometimes as much as 30 feet (10 m) a year. therefore a person would readily notice the retreat of glaciers during a lifetime, and there were modern humans in both eurasia and in north america to observe this. the return of water to the meteoro-logical cycle resulted in a wetter climate in tropical regions, in some regions even wetter than today. What is now the Sahara Desert, about 9,000 years ago had extensive grasslands and herds of wild ani-mals, which human tribes observed, carving images of them on rocks.
the vast amounts of water from melting gla-ciers had to go somewhere. Many rivers were much larger during the time of glacial melt than they are today. For example, the bluffs along the Missouri River, formed from fine soil blown from land newly exposed by the retreating glaciers, are today more than two miles (over three kilometers) from the current banks of the river, implying that the river was about four miles (seven kilometers) wider as the glaciers were melting upstream. Vast lakes, such as lake agassiz in north america, formed from the meltwater. although the Great lakes are among the largest lakes in the world and contain about a fifth of the planet’s freshwater, they are much smaller than lake agassiz, of which they are remnants. in
xviii+598_Biodiversity_F.indd 316 11/17/11 1:13 PM

ice ages 317
some cases, ice blocked the drainage of the lakes, and when the ice gave way, massive fl oods occurred downstream. this occurred with lake agassiz and produced a fl ood of water down the St. lawrence River. So much freshwater drained during this fl ood that it altered the climate of the entire region for a few hundred years. Since freshwater is lighter than salt water, this cold water remained at the surface of the north atlantic and caused a temporary rever-sal of the warming trend that had been occurring. a similar fl ood, in western north america, pro-duced huge ripples and other deposits known as the Scablands.
Within about 5,000 years (by 7,000 years before the present), the warming trend had reached a max-
imum, known as the hypsithermal. at this time, grasslands covered much of the land that is today forested in north america. this included a new kind of grassland, the tallgrass prairie, consisting of large deep-rooted grasses that grow mostly during warm summers. as conditions began to change once again, leading the northern Hemisphere toward the next ice age, cool moist conditions returned to the north and the forests advanced southward. the tallgrass prairie, however, persisted in regions with a climate capable of supporting trees, because the accumula-tion of dead grass encouraged fi res. the fi res kill tree seedlings but merely create fertilizer for the grasses and wildfl owers that resprout from underground stems and roots.
North American vegetation zones during the most recent ice age
xviii+598_Biodiversity_F.indd 317 11/17/11 1:13 PM

first entry318 insects 318
the ice ages continue. During the last glaciation, 30 percent of north america was covered with ice. But 10 percent of it still is and another 14 percent has permafrost, which is a permanent layer of ice beneath the soil. if past conditions occur again, another ice age is on the way. However, the green-house effect is causing the opposite trend to occur; glaciers and sea ice in north america, as well as antarctica, are becoming smaller and thinner. it would be very nice for human civilization if the greenhouse effect precisely balanced the return of the ice ages; however, global temperature is rising (largely due to human production of carbon dioxide) 10 times faster than it did even during the period of rapid glacial retreat. the greenhouse effect is caus-ing much of the permafrost to melt, resulting in soil subsidence and the buckling of highways.
the ice ages occurred rapidly enough that very little evolution resulted from them. For the most part, species of plants and animals have dispersed to new locations, rather than evolving into new species, as a result of any single ice age. Sometimes, plant species have gotten trapped in limited locations as they have moved northward. the alpine tundra, at the tops of ranges such as the Rocky, cascade, and Olympic Mountains and the Sierra nevada, came into existence when arctic tundra retreated to the tops of mountains rather than northward. Subal-pine forests came into existence when boreal forests retreated up the slopes of mountains. Some individ-ual species have gotten trapped in regions to which they may not be well adapted, resulting in a disjunct distribution pattern. One example is the Monterey cypress (Cupressus macrocarpa) of california. Dur-ing the most recent ice age, it survived in Mexico. as this species dispersed northward during glacial retreat, it migrated westward and ended up con-fined to a small area near the Monterey Peninsula of california. Had it turned eastward rather than west-ward, it might have found a much more extensive region in which to live.
in some cases, plant species dispersed northward after the last ice age more quickly than can be eas-ily explained by the movements of their seeds. in europe, the oak Quercus robur can spread rapidly, as jays carry their seeds, but Q. petraea seeds spread much more slowly. Recent research suggests that Q. petraea spread northward not by its seeds but by its pollen. the two oak species can cross-pollinate. Q. robur spread quickly northward as birds car-ried its seeds, and the wind carried pollen of Q. petraea northward. this pollen fertilized Q. robur individuals once they had matured. the resulting hybrid seeds grew into male-sterile trees, which pro-duced only female flowers. When the hybrid trees matured, they too were pollinated by Q. petraea pol-
len, and some of the offspring were completely fertile Q. petraea trees. in this way, Q. petraea dispersed northward at the rapid pace of their windborne pol-len, not the slow pace of their acorns.
Many extinctions (the Pleistocene extinction) occurred at the end of the last glaciation, far more than at the ends of previous ice ages. in many cases, trees were left without the animals that had previ-ously dispersed their seeds.
in some cases, the overall climatic changes of the Pleistocene may have affected evolution, even if the individual glaciations did not. One example has direct relevance to humans. On at least two occa-sions starting about 5 million years ago, the spread of drier conditions across africa caused forests to retreat and savannas to spread. these cycles of arid-ity may have been important stimuli in the evolution of the human lineage. a drying trend in africa about 5 million years ago may have been an important stimulus in causing australopithecines to evolve the ability to walk on two legs, even though recent evi-dence indicates that human ancestors still lived in forests even after becoming bipedal. another drying trend over a million years ago may have stimulated the earliest members of the genus Homo to make tools, eat more meat, and evolve larger brains. evo-lutionary shifts in other african mammals, such as antelope, occurred at about the same times.
in europe, during the ice age prior to the most recent, neanderthals (Homo neanderthalen-sis) were the only humans. Modern humans (H. sapiens) migrated into europe and displaced the neanderthals during the interglacial period. the neanderthals were extinct by the time the most recent glaciation reached its maximum.
See also biogeography; boreal and sub-alpine forests; disjunct species; greenhouse effect; Pleistocene extinction; tundra.
Further readingcolinvaux, Paul. Amazon Expeditions: My Quest for the
Ice Age Equator. new Haven, conn.: yale University Press, 2008.
Macdougall, Doug. Frozen Earth: The Once and Future Story of Ice Ages. Berkeley: University of california Press, 2006.
Petit, Rémy J., et al. “Hybridization as a mechanism of inva-sion in oaks.” New Phytologist 161 (2003): 151–164.
Pielou, e. c. After the Ice Age: The Return of Life to Glaci-ated North America. chicago: University of chicago Press, 1992.
insects insects are arthropods with six legs and three body parts. like other arthropods, they have an external skeleton (exoskeleton) and jointed legs.
xviii+598_Biodiversity_F.indd 318 11/17/11 1:13 PM
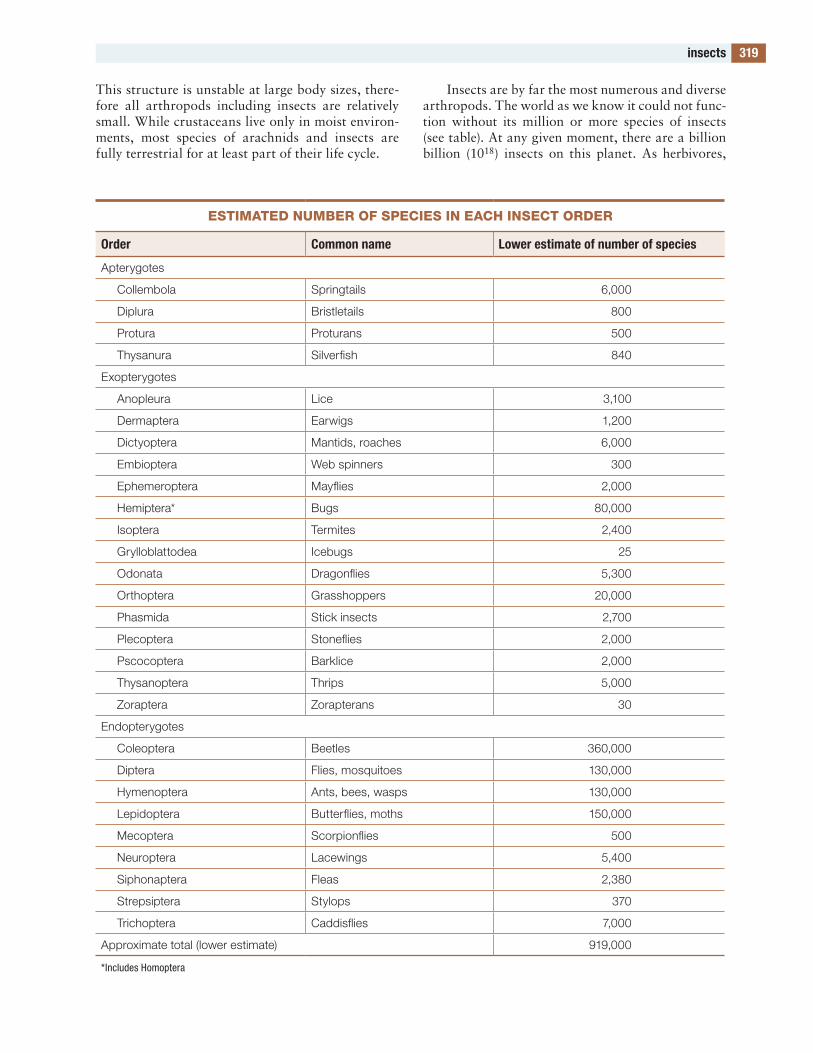
insects 319
this structure is unstable at large body sizes, there-fore all arthropods including insects are relatively small. While crustaceans live only in moist environ-ments, most species of arachnids and insects are fully terrestrial for at least part of their life cycle.
insects are by far the most numerous and diverse arthropods. the world as we know it could not func-tion without its million or more species of insects (see table). at any given moment, there are a billion billion (1018) insects on this planet. as herbivores,
estimated number of speCies in eaCh inseCt order
order Common name lower estimate of number of species
Apterygotes
Collembola Springtails 6,000
Diplura Bristletails 800
Protura Proturans 500
Thysanura Silverfish 840
Exopterygotes
Anopleura Lice 3,100
Dermaptera Earwigs 1,200
Dictyoptera Mantids, roaches 6,000
Embioptera Web spinners 300
Ephemeroptera Mayflies 2,000
Hemiptera* Bugs 80,000
Isoptera Termites 2,400
Grylloblattodea Icebugs 25
Odonata Dragonflies 5,300
Orthoptera Grasshoppers 20,000
Phasmida Stick insects 2,700
Plecoptera Stoneflies 2,000
Pscocoptera Barklice 2,000
Thysanoptera Thrips 5,000
Zoraptera Zorapterans 30
Endopterygotes
Coleoptera Beetles 360,000
Diptera Flies, mosquitoes 130,000
Hymenoptera Ants, bees, wasps 130,000
Lepidoptera Butterflies, moths 150,000
Mecoptera Scorpionflies 500
Neuroptera Lacewings 5,400
Siphonaptera Fleas 2,380
Strepsiptera Stylops 370
Trichoptera Caddisflies 7,000
Approximate total (lower estimate) 919,000
*Includes Homoptera
xviii+598_Biodiversity_F.indd 319 11/17/11 1:13 PM

first entry320 insects 320
carnivores, pollinators, and decomposers, they play a greater role in the flow of energy and matter through the earth’s ecosystem than any other group. about 800 species of insects have become pests of agricultural crops; on the other hand, one-third of all commercial crops depend on bees for pollination. Flight has allowed insects to evolve many different pollination relationships with plants; the evolution of pollination relationships may have been the most important factor contributing to the evolution of insect diversity.
STruCTurE aNd fuNCTIoN of INSECTSinsects have an open circulatory system, in which the blood bathes all of the internal organs inside of the exoskeleton. Most of the blood is watery hemo-lymph, with some blood cells. the blood cells do not have hemoglobin, therefore the circulatory system does not carry oxygen through the body. insects have a single blood vessel, open at both ends. in it, a heart pumps blood along the dorsal (back or upper) side of the body toward the head. Oxygen is delivered to all parts of the insect body not by the blood but by a series of air tubes called tracheae. air enters the tracheae through openings (spiracles) in the thorax and abdomen. Muscular activity during movement helps to squeeze fresh air in and out of the insect’s body. in aquatic stages of the life cycle, outgrowths of the tracheal system form gills that absorb oxygen from surrounding water. insects may also store air bubbles inside or outside of their body. these bubbles serve two functions: first, they allow terrestrial insects to take an air supply with them underwater; secondly, they allow insects that live in dry environments to store an air supply without opening their spiracles, through which they could lose water vapor, very often.
insects have appendages, which are legs or modified legs. evolution modified the functions of these appendages into specific and diverse forms, such as antennae and mandibles. the evidence for this is genetic. certain mutations cause appendages to develop into the wrong kind of appendage; for example, the antennapedia mutation of flies causes legs to develop where antennae are supposed to be.
insects have three body parts, each composed of several segments, as follows:
The head. an insect head has appendages such as antennae and mandibles. the antennae (which may have many segments) have a sensory func-tion, often by reacting to the presence of specific molecules such as sex pheromones. the mandibles manipulate and process food items. therefore insect mandibles (modified from legs) have an entirely dif-ferent developmental origin from vertebrate jaws (modified from gill arches of fishes). Mandibles may
chew or (if modified into a long proboscis) suck nec-tar from a flower or juices from plant and animal tissues. the head also has a pair of compound eyes, which consist of a large number of small lenses or ommatidia (4,000 in the case of the house fly) (see photograph on page 321). each lens registers a pixel of light intensity and color. compound eyes do not produce a sharp image but are good at detecting movement. Between the compound eyes there are usually three ocelli, or single-lensed eyes.
The thorax. the thorax, which is the middle part of the body, contains three pairs of legs. each leg has six segments: the coxa anchors the leg to the thorax; the trochanter is a swivel joint where the leg attaches; the femur, tibia, tarsus, and pretarsus make up the long portion of the leg. Most insects have one or two pairs of wings. in some insect lin-eages, wings did not evolve; other lineages, such as fleas, lost their wings during evolution. the thorax contains a lot of muscles.
The abdomen. the abdomen contains the heart, digestive system, and reproductive system.
the evolution of wings, unique among arthro-pods, is one of the insects’ most important terrestrial adaptations. Wings allow some insects to travel at great speeds. the sphinx moth can fly at 1,000 body lengths per second. a jet flying at three times the speed of sound travels only 100 body lengths per second. in addition to speed, wings allow insects to have tremendous maneuverability.
the earliest insects lived on land during the Devonian period. insects remained rare until the carboniferous period. the earliest insects ate detritus, pollen, or spores; they did not eat leaves until very much later, perhaps as late as the Juras-sic period. this is one reason that plant material from carboniferous swamps built up into large coal deposits: insects did not defoliate them before they died. insect flight evolved during the carbon-iferous period, particularly the large dragonflies. Meganeura monyi had a wingspan of over two feet (75 cm). Such a wingspan seems too large to have allowed flight, and the insect system of respira-tion through spiracles seems incapable of provid-ing enough oxygen to such a large organism. Some scientists speculate that the atmosphere contained more oxygen at that time, which made respiration easier and also made the atmosphere thicker. as a result, the large dragonflies would have moved their wings slowly, mostly gliding through the thick air. By the Devonian period, other modern insect groups such as orthopterans, coleopterans (beetles), and hemipterans (bugs) had evolved. numerous orders of insects died during the Permian extinction, but surviving orders (especially flies and beetles) prolif-erated during the Mesozoic era. By the cretaceous
xviii+598_Biodiversity_F.indd 320 11/17/11 1:13 PM

insects 321
period, insect life consisted mostly of the orders that dominate the world today.
Most modern insects are either amphibious (liv-ing first in the water then on land) or entirely terres-trial. the first insects lived partly or wholly on land; a few modern insects have returned to an entirely aquatic existence.
insects undergo metamorphosis, from juvenile to adult, by passing through a series of molts or instars. Some insects such as grasshoppers have relatively small changes during metamorphosis, in which the juvenile insect (nymph) that hatches from the egg closely resembles the adult (hemimetabolous devel-opment). Wings usually develop only on the final instar. Other insects undergo complete metamor-phosis. they have a larval stage that looks almost completely different from the adult, as “grubs” bear little resemblance to adult beetles or bees, nor cater-pillars to butterflies (holometabolous development). the larvae are almost always adapted as “eating machines,” while the adults (which devote them-selves to reproduction) eat much less, if anything at all. in amphibious insects, such as the dragonfly, the juvenile stage lives underwater, but the adult lives on land and in the air.
ovErvIEw of INSECT dIvErSITythe three major lineages of the insects are as fol-lows: (1) the apterygota are wingless and have not had wings during their evolutionary history. this group includes the springtails (collembolans) and silverfish (thysanurans). they have long antennae that curve backward and long abdomens that end in three bristles. Most of them live in the soil, although some silverfish live in books, which resemble leaf litter. (2) the exopterygota have hemimetabolous development and wings that develop on the outside of the body. (3) the endopterygota have holome-
tabolous development and wings that develop inside of the body. Some scientists classify the apterygota separately from the insects.
the exopterygotes, whose nymphs resemble the adults, include the following:
Order Ephemeroptera. the mayflies are the oldest group of winged insects still in existence. they are common in freshwater worldwide. the lar-vae live underwater, usually eating algae or detritus, for up to two years. they breathe through abdomi-nal gills. after undergoing many instar molts, the mayfly emerges into the atmosphere during its sec-ond to last molt. Mayflies are the only insects to have wings before the final molt. they then land on a surface to undergo their final molt. Males swarm over water and females choose males, mate with them, and lay eggs. they are called ephemeroptera because the adults live very brief (ephemeral) lives.
Order Odonata. the dragonflies and damsel-flies have two pairs of membranous wings. Dragon-flies hold their wings horizontally, and damselflies hold them vertically. Unlike other insects, dragon-flies and damselflies can move the front and back wings independently of one another. this allows them to have tremendous maneuverability. Because the wings are so large, they can beat only one-tenth as fast as those of bees. though they have very small antennae, they have very large eyes, each with as many as 30,000 ommatidia. Odonates were among the first flying insects, as noted previously. Drag-onflies and damselflies are unique among insects in that the male inseminates the female with a sec-ondary sexual apparatus; before mating, he must transfer sperm to this apparatus from his primary sexual apparatus. nymphs are aquatic and adults terrestrial. they are predators, and the nymphs can be cannibalistic.
Order Orthoptera. the grasshoppers and crickets are easily recognized by enlarged back legs, used primarily for jumping. Grasshoppers have short antennae, while crickets have long antennae. they have chewing mouthparts. Grasshoppers are pri-marily herbivores, while crickets are omnivorous; a few orthopterans are predatory. they are renowned for the sounds that they produce. crickets rub their left and right front wings together, while grasshop-pers rub their upper hind legs against their front wings. Some orthopterans produce sounds by rub-bing their mandibles together or by drumming their legs against wood.
Order Phasmida. the stick and leaf insects are either elongated, resembling sticks, or flattened, resembling leaves. they are spectacular examples of mimicry, some species having a remarkable resem-blance to their models and all of them a close enough resemblance to fool most predators. Often, they are
Insects, such as this Condylostylus fly, have compound eyes. (Thomas Shahan)
xviii+598_Biodiversity_F.indd 321 11/17/11 1:13 PM

first entry322 insects 322
also animal mimics. Females of Extatosoma tiara-tum lay eggs in ant nests. the eggs have a protuber-ance that the ants like to eat. the ants carry the eggs inside the nests, eat the protuberance, and throw the eggs into the garbage pile. Being in the ant garbage pile protects the eggs from other predators.
Order Dictyoptera. this order consists of the mantids and cockroaches. the mantids (such as the praying mantis) have enlarged front legs which they keep against their bodies as they sit still for long periods of time (reminding some observers of prayer). When a prey animal comes within reach (usually a smaller arthropod, though sometimes small vertebrates) the mantids quickly extend their legs and catch the prey with the bristles on the legs and voraciously consume it. in some mantid species, the female eats the male during copulation; even with his head removed, the male can copulate. it is not clear whether this is or is not simply a mistake on the part of the female, who may attack almost any small animal within reach. Roaches are flat, with hind wings hidden underneath strong fore-wings, and their shape allows them to seek out and fit into crevices (natural or human). less than 1 per-cent of cockroach species inhabit buildings and are human pests. they are not disease vectors the way fleas and mosquitoes are, although they may carry germs with them from inaccessible and uncleaned areas in a building.
Order Isoptera. termites are renowned for their societies. they live in groups of up to a million individuals, often in large mounds which can be up to 24 feet (8 m) tall, the temperature conditions of which they can control (see photograph). the less specialized termites eat wood, much of which is inside of human buildings, causing a great deal of economic damage. More specialized termites eat other types of food as well, including leaves, seeds, and lichens, much of it (like wood) containing cel-lulose. termites cannot digest wood. they depend upon mutualistic microorganisms living within their digestive systems to perform this task: protists in the less specialized termites and anaerobic bacteria in the more specialized termites. Some of them (like leaf-cutter ants, to which they are not closely related) raise and eat gardens of fungus inside their mounds. all of the reproduction is carried out by a primary female (“queen”) and primary male (“king”), whose hormones suppress reproduction in all the other termites in the mound. the queen becomes an egg-laying machine, with a grossly distended abdomen, laying an egg every three seconds—tens of millions over her lifetime. the queen does not eat wood but is fed by the saliva of the workers. Sterile soldiers defend the nest in several ways, depending on the species. Some soldiers have huge heads and man-
dibles for attacking intruders; other soldiers spit toxic solutions; others have large heads with which they block the entrances to the nest when it is under attack. termites are so abundant that the methane produced by their colonies makes a measurable con-tribution to the global carbon cycle. Most termites are wingless, but winged reproductive individuals can swarm to new locations. Upon arrival, they shed their wings.
Order Anopleura. lice are wingless and hor-izontally flattened parasites of vertebrates, allow-ing them to lie very close to the skin of the host. their flatness makes them difficult to remove by scratching or preening. the lice are divided into two groups. the Mallophaga (biting lice) mostly con-sume skin and feather fragments (and occasionally blood) of birds. the Siphunculata (sucking lice) suck blood from mammals. two louse species live only on humans: the pubic louse and the body louse (with separate subspecies that live on the head or on the trunk of the body).
Order Hemiptera. the true bugs are insects that have pairs of sucking mandibles. Most of them
Termites produce large mounds in tropical areas. This mound in Litchfield National Park, Northern Territory, Australia, is about 16 feet (5 m) tall and may be more than 50 years old. The termites live underground; the aboveground mound is a complex chamber in which fungi partially decompose the grass that the Nasuti-termes triodiae termites eat. (J. Brew)
xviii+598_Biodiversity_F.indd 322 11/17/11 1:13 PM

insects 323
consume plant fluids (thus often causing economic loss), although some are predatory. examples include the shield bugs (or stink bugs) with a large shield over the back of the thorax; the water striders, which can walk upon the surface tension of water; the cica-das; and the aphids.
● Cicada nymphs (within order Hemiptera) feed on plant roots; the adults emerge from the ground and produce sounds from tree branches by buckling special membranes on the foremost abdominal segment. the adults reproduce and die. Periodical cicadas emerge en masse every 13 or 17 years. When they emerge, there are too many of them for predators to deplete their populations, and the predator life cycles cannot easily adjust to those of their cicada prey, which emerge on a prime-number schedule.
● Aphids (within order Hemiptera) are tiny, soft-bodied bugs that insert a proboscis into plant phloem cells, from which they drink sap. Phloem sap is pressurized, therefore aphids do not need to suck or even swallow. the sugary fluid passes through them too fast for efficient digestion; objects under-neath a tree infested with aphids become sticky with slightly-altered plant sap called honeydew. aphids can withdraw so much sap from a plant that it dies. aphids can also transmit plant diseases (often viruses) by the proboscis. less than 10 percent of aphid species are agricultural pests. the sap that aphids consume contains little nitro-gen. aphids obtain the nitrogen that they need (such as amino acids) by processing a large amount of sap and from the mutu-alistic metabolism of bacteria (often genus Buchnera) within their digestive systems. large aphid populations may consist only of females that reproduce asexually by laying unfertilized eggs, but toward the end of the season sexual forms develop and mate. Dur-ing their rapid population growth phase, female aphids have live birth, and the little aphids already have eggs developing into embryos inside of them. Many aphids have a mutualistic association with ants, in which the ants live largely or exclusively from hon-eydew, and the aphids depend upon the ants to transport and protect them. this mutual-ism has evolved independently many times.
in some schemes of classification, bugs such as cicadas and aphids are placed in their own order, the Homoptera.
the endopterygotes, which have a larval stage that pupates, include the following:
Order Siphonaptera. adult fleas suck the blood of birds and mammals. the larvae eat detritus from the bodies or nests of their hosts. among the adaptations that make them successful parasites are the following:
● Fleas are laterally flattened, making them very difficult to remove by scratching or preening.
● Fleas have strong legs that allow them to escape danger and to leap from one host to another, up to 150 times their body length or 80 times their height.
● Fleas have backward-pointing bristles that enhance their ability to escape from other animals.
● Fleas release an anticoagulant when they bite, which not only keeps the blood flowing but also causes the itchy irritation.
● Fleas respond to the hormones in the blood, allowing them to synchronize their repro-duction with that of the host. in this way, a new generation of fleas can infest a new generation of baby mammals or birds.
Fleas are vectors of disease, the most famous of which is the bubonic plague, which has caused numerous plagues including the Black Death in europe in 1345–47. Bubonic plague is spread by rat fleas, Xenopsylla cheopsis, which carry the plague bacteria from rats to humans, and not by the human flea Pulex irritans. the Black Death, as well as other plagues, has changed the course of human history.
Order Diptera Most flying insects have four wings, but flies and mosquitoes have just two—the front pair. “Diptera” is Greek for “two wings.” the hind pair of wings develops into balancing devices, known as halteres, which greatly enhance their maneuverability. the two main groups of dipterans are the nematocera (mosquitoes and relatives) and the Brachycera (flies and relatives).
Mosquito larvae are the aquatic “wrigglers” with complete heads and mouth bristles, but no legs. they eat smaller algae and bacteria, often in the sur-face film of still water. their pupae float, and adult mosquitoes emerge from them into the air. craneflies look like large mosquitoes. their larvae consume decaying vegetation in moist soil. adult mosquitoes and craneflies may not eat or may consume only nectar. Female mosquitoes need a blood meal to provide protein and iron for egg development. the saliva of the female mosquito contains compounds that prevent blood clotting, promote blood flow, and inhibit host immune response. Mosquito bites
xviii+598_Biodiversity_F.indd 323 11/17/11 1:13 PM

first entry324 insects 324
transmit many diseases, such as yellow fever, dengue fever, and viral encephalitis. they do not trans-mit HiV (the virus that causes aiDS). One of the most common and most severe diseases in the world, malaria, spreads through mosquito bites, mostly by the genus Anopheles.
Fly larvae are maggots, with few distinguishable external features, that (depending on the species) live in decaying flesh, fruit, dung, and occasion-ally in living flesh (as with bot flies in cattle). adult flies have very large compound eyes. although there are many thousands of species of flies, one spe-cies (Musca domestica, the house fly) has followed humans throughout the world. adult flies may feed on many different kinds of fermenting liquid food. adult fruit flies and vinegar flies are attracted to ripe or decaying fruit, as well as to wine. Because their mouthparts form a tube, adult flies cannot ingest solid food. Often their saliva helps to soften the food before they ingest it. Because the adults often visit vertebrate feces, house flies can spread many of the same diseases that otherwise spread by sewage contamination of water and food. Flies, however, almost continually scrape contaminants from their body surfaces.
Order Lepidoptera. Butterfly and moth cater-pillars are eating machines, usually consuming large amounts of leaf material, much of it toxic, from which they extract the little bit of nutrition that is available and produce a lot of waste. in most species, with exceptions such as the gypsy moth (Lymant-ria dispar), the caterpillars eat only one species of plant. in some cases, the species may be flexible, but the individual caterpillars are not. Hornworms (the caterpillars of the sphinx moth or hawkmoth, Manduca sexta) can feed on many different spe-cies within the plant family Solanaceae (including tobacco, tomatoes, and jimsonweed) but a caterpil-lar will reject leaves from all solanaceous species except the one which it first ate. Many caterpillars may eat other insects if they are small enough and are encountered by chance. a carnivorous caterpil-lar (genus Eupithecia) has evolved in Hawaii. Some moth caterpillars may eat fungi, decaying material, or even feces. explosions of caterpillar populations may cause severe damage to crops or wild plants; the gypsy moth caterpillar defoliates large acreages of trees in the northeastern United States each summer. insects have six legs, but caterpillars appear to have more. the only true jointed legs are the six toward the front of the caterpillar; the others are prolegs that are not jointed but end in bristly pads that help them clasp to stems and leaves.
Once it has completed its feeding, the caterpillar transforms into a pupa. in moths, the pupa forms a cocoon of silk. in butterflies, the pupa is a chrysa-
lis with exoskeleton plates. the pupating caterpillar of the sphinx moth (hawkmoth) (Manduca sexta) forms a tube inside of which the adult tongue will develop. inside the protected chrysalis or cocoon, the pupa undergoes a striking metamorphosis. caterpil-lars have “buds” of cells that they do not use but that become active during metamorphosis. enzymes digest much of the caterpillar’s body from the inside, and the buds form new structures (such as wings). the meta-morphosis includes the exchange of chewing man-dibles for a long tongue. Pupae are usually immobile, although sphinx moth caterpillars can wriggle if dis-turbed. it is perhaps this stage of the life of a butterfly or moth that has most fascinated humans, to whom it appears to be a death and rebirth.
the adults emerge with four wings covered with scales, some of which refract light into different iridescent colors. Most of them insert their long tongues, like coiled ropes, into long-tubed flowers to drink nectar. Some adult moths do not eat anything. a few will consume animal materials: Lobocraspis griseifusa drinks tears of large mammals, Ophiusa tirhaca pierces the skin of fruits, and Calyptra minu-ticornis actually pierces the skin of animals to drink blood. the tongue may be longer than the body of the butterfly or moth itself. Butterflies often have delicate wings, while moths have more rigid wings. Sphinx moths beat their rigid wings rapidly and can be mistaken for hummingbirds by casual observers. the antennae (feathery in the case of moths) detect pheromones (aerially dispersed sex hormones) which allow males to find females. Butterflies are usually colorful and active during the day, while moths are often dull in color and nocturnal, although there are many exceptions, such as the luna moth (see photo-graph on page 325).
Coleopterans. With more than 360,000 rec-ognized species, the beetles are the largest order of insects and perhaps the most diverse group of animals in the world. about one in four species of animals is a beetle. Someone once asked the famous British biologist J. B. S. Haldane what he learned about God from his studies of nature. He answered that God must be “inordinately fond of beetles.” “coleoptera” means “sheath-winged” because the first pair of wings in the adult form a sheath (the elytra) that covers the flying wings; beetles raise their elytrae when they fly. their species diversity is matched by their diversity of size: Titanus giganteus (South american long-horned beetle) is the largest, at over six inches (16 cm) in length and a fungus beetle Nanosella fungi may be the smallest insect, at one-hundredth of an inch (0.25 mm). the adults have chewing mouthparts.
Some beetles eat plant materials and may be pests of human crops and food. Both the larvae and
xviii+598_Biodiversity_F.indd 324 11/17/11 1:13 PM

insects 325
adults of the colorado potato beetle (Leptinotarsa decemlineata) feed on potatoes and related crops. Weevils (family curculionidae) destroy crops and stored foods. But beetles play essential ecological roles. Many are pollinators of flowers and may have pollinated the earliest flowers. Some beetles make a living from the decomposition of vertebrates or their dung. Dung beetle (scarab beetle) adults may live in or bury dung or roll spheres of dung with their hind legs. they lay eggs in the dung, and the grubs develop there. Burying beetles (genus Nicophorus) find a vertebrate corpse, remove the hair or feathers, cover it with secretions that inhibit decomposition, and dig a crypt underneath it, eventually burying it. the female lays eggs near the carcass, and then they eat the carcass and feed regurgitated food to the grubs that hatch. Both larvae and adult ladybird beetles (family coccinelidae) are predators upon other insects, especially aphids. “lightning bugs” are actually beetles (family lampyridae) that use light, produced by a very efficient enzymatic reaction that generates almost no heat, as mating signals.
Hymenopterans. ants, bees, and wasps have legless larvae that develop in cocoons. the adults have chewing mouthparts, sometimes modified for sucking, two pairs of membranous wings (Hymenop-
tera comes from the Greek for “membrane-winged”), and a very narrow connection of the abdomen to the thorax. the narrow waist allows hymenopterans to bend their abdomens at an acute angle while laying eggs or stinging.
Many species of bees and wasps are solitary, but many of them, as well as nearly all ants, form large societies (in some cases of 300 million individu-als). in these societies, the females are diploid (have pairs of chromosomes) while the males are haploid (have unpaired chromosomes), a system known as haplodiploidy. Queen females lay all of the eggs; in the workers, the ovipositor (egg tube) develops into a stinger. Most hymenopteran females have the ability to sting. the bee’s stinger has barbs. the worker cannot remove its stinger from the flesh of an intruder. the worker bee dies after pulling free from the intruder. each worker ant, having a barb-less stinger, can sting an intruder several times. the haplodiploid system results in a high genetic related-ness among the workers, which in turn has allowed the evolution of altruism to the extent that workers may die in defense of the colony. When fire ants (Solenopsis invicta) attack intruders, many of them crawl onto the intruder and then synchronize their stinging attack.
Many butterflies and moths, such as this luna moth in Arkansas (Actias luna), have colorful wings. (Rutgers University)
xviii+598_Biodiversity_F.indd 325 11/17/11 1:13 PM

first entry326 insects 326
the ecological and economic importance of hymenopterans are almost impossible to overstate. numerous species of wild bees pollinate flowers as they forage for sweet nectar and protein-rich pollen. One species, the european honeybee Apis mellifera, is overwhelmingly important to agriculture; accord-ing to one estimate, every third bite of food comes from plants that require bee pollination or comes from cattle whose alfalfa requires pollination. Para-site and disease outbreaks (and possibly pollutants) threaten honeybee populations, especially since the emergence of “colony collapse disorder” in 2007. Some scientists are seeking to cultivate wild bees, which are often more efficient at pollinating specific kinds of agricultural plants. Some wasps are also pollinators, especially of figs. Many wasps lay eggs in caterpillars, which the wasp grubs eat; wasps can therefore be an important biological control of agri-cultural pests. ant burrows aerate soils.
INSECTS aNd humaNSinsect populations are so massive that, next to bacte-ria and plants, they may be the major players in the ecosystems of the earth. For example, one-third of the animal biomass of the amazon rain forest con-sists of ants and termites.
Many insects become pests when their popula-tions get out of control. Massive use of pesticides has not only failed to control them but has resulted in the evolution of super-pests. Modern agricultural researchers now realize that the best, and only per-manent, control of insect pests is to release diseases, predators, parasites to kill the pests, or release steril-ized adults of the pest species, which mix with the pest population and reduce its reproductive success. in addition, if humans try to spray all of the insect pests, they will also kill the many insects that are not only beneficial but also essential, especially the polli-nators. these are some of the reasons that integrated pest management is beginning to replace the heavy use of pesticides in many regions of the world.
insects may provide great medical benefits. Sometimes medicines are ineffective against infected wounds, but doctors can sometimes use maggots—yes, the larvae of flies—to clean out decaying flesh from a person’s wound, a process known as debride-ment. Furthermore, scientists have wondered why some insects such as ants can live in dense popula-tions without having outbreaks of disease. it appears to be because these insects produce antibiotics that protect them, either directly or by encouraging the growth of beneficial microorganisms. in the contin-ual search for new antibiotics, researchers may have to turn to the insects.
Many people do not like insects, even those they will admit are good for the human economy. they
need someone to popularize them. May Berenbaum, an entomologist at the University of illinois, has done precisely this. She is famous not only for her research in the coevolution of insects and plants and for leading research into the causes of colony collapse disorder (the deaths of honeybee colonies), but also for her popular science books and for start-ing the insect Fear Film Festival at the University of illinois, an annual event for more than a quarter of a century.
See also arthropods; integrated pest management; invasive species; mutualism; pollination.
Further readingaverof, Michalis, and Stephen M. cohen. “evolutionary
origin of insect wings from ancestral gills.” Nature 385 (1997): 627–630.
Bänziger, H. “Skin-piercing blood-sucking moths i: eco-logical and ethological studies on Calpe eustrigata (lepidoptera, noctuidae).” Acta Tropica 32 (1975): 125–144.
Berenbaum, May. Bugs in the System: Insects and Their Impact on Human Affairs. Upper Saddle River, n.J.: addison-Wesley, 1996.
———. Buzzwords: A Scientist Muses on Sex, Bugs, and Rock ‘n’ Roll. Washington, D.c.: national academies Press, 2000.
———. “colony collapse disorder and pollinator decline.” Presented to committee on agriculture, U.S. House of Representatives, March 29, 2007. available online. URl: http://www7.nationalacademies.org/ocga/testimony/ colony_collapse_Disorder_and_Pollinator_Decline.asp. accessed august 10, 2011.
———. The Earwig’s Tail: A Modern Bestiary of Multi-Legged Legends. cambridge, Mass.: Harvard Univer-sity Press, 2009.
———. Ninety-nine Gnats, Nits, and Nibblers. Urbana, ill.: University of illinois Press, 1989.
chernin, e. “Surgical maggots.” Southern Medical Journal 79 (1986): 1,143–1,145.
conner, William e., ed. Tiger Moths and Woolly Bears: Behavior, Ecology, and Evolution of the Arctidae. new york: Oxford University Press, 2008.
eisner, thomas. For Love of Insects. cambridge, Mass: Harvard University Press, 2003.
Franks, n. R. “army ants: a collective intelligence.” Ameri-can Scientist 77 (1989): 139–145.
Glausiusz, Josie. Buzz: The Intimate Bond Between Humans and Insects. San Francisco, calif.: chronicle Books, 2004.
Grimaldi, David, and Michael S. engel. Evolution of the Insects. new york: cambridge University Press, 2005.
Hölldobler, Bert, and edward O. Wilson. The Ants. cam-bridge, Mass: Harvard University Press, 1990.
xviii+598_Biodiversity_F.indd 326 11/17/11 1:13 PM

327integrated pest management
Oldroyd, B. P. “What’s killing american honey bees?” Pub-lic Library of Science Biology 5 (2007): e168. avail-able online. URl: http://www.plosbiology.org/article/info:doi/10.1371/journal.pbio.0050168. accessed May 16, 2011.
Ramel, Gordon. “the insect orders: an introduction to each of the 32 orders of insects.” available online. URl: http://www.earthlife.net/insects/orders.html. accessed May 16, 2011.
Waldbauer, Gilbert. What Good Are Bugs: Insects in the Web of Life. cambridge, Mass: Harvard University Press, 2003.
White, Richard e. Beetles: A Field Guide to the Beetles of North America. a Peterson Field Guide. new york: Houghton Mifflin, 1998.
integrated pest management (IPM) inte-grated pest management (iPM) is a way of control-ling pests, principally insects that damage crops and spread diseases, that recognizes the reality of ecol-ogy and evolution. it is a viewpoint about how to restrict the growth of pest populations rather than a detailed guide to doing so. the “conventional” view of pest control, promoted by large corporations that sell chemicals, is that large amounts of pesticides are the only effective way to control pests. in contrast, iPM (as its name suggests) integrates many different ways of controlling pests, including the use of pesti-cides. Here follow some of the guiding principles of integrated pest management:
Zero pest population levels are unattain-able. no matter how much a farmer may spray on a cotton field to control bollworms or how much a public health official may spray to control mos-quitoes, there is no way to kill all of the pests. as the United States environmental Protection agency says, “Sighting a single pest does not always mean control is needed.” But it is possible to keep pest populations low enough that they do not cause sig-nificant damage to crops or to human health by using a combination of methods.
Overuse of pesticides causes pest resur-gence. the heavy use of pesticides can, in fact, greatly reduce pest populations. However, the pes-ticides also reduce the populations of the natural predators, either directly (as when the pesticides kill spiders or parasitic wasps) or indirectly (as when pes-ticides concentrate in the food chain and kill birds). all of the populations begin to recover after the use of the pesticide. the predator populations grow more slowly than the pest populations. Soon after the pesticide has either degraded or washed away into wetlands and soils, there is a population explo-sion of pests. in order to prevent this, iPM attempts to keep the wild predator populations intact.
Overuse of pesticides in rice fields in indone-sia caused an outbreak of the brown plant hopper Nilaparvata lugens in 1974. the powerful central government decreed restrictions on pesticide use, and advertisements encouraged farmers to not kill spiders. natural predators returned, including the beetles Harmonia octomaculata and Ophionea nigrofasciata and the parasitic insects Tetrastichus schoenobii, Telenomus rowani, and Cotesia angus-tibasis. Pesticides are still necessary and used, but at much lower levels than in 1974.
Overuse of pesticides causes the evolution of resistant pests. natural selection is inevitable in pest populations, just as in every other population. Pests have physiological mechanisms to either avoid or to detoxify pesticides. these mechanisms are the product of millions of years of natural selection that have given the animals the ability to avoid or resist natural toxins, especially those produced by plants, and they work just as well against synthetic toxins. an environment saturated with pesticides creates an ideal stage for the evolution of pest populations that are largely unaffected by the pesticide.
“Natural” pesticides can become ineffective almost as easily as synthetic ones. the problems of pesticide overuse cannot be avoided by simply using natural compounds such as pyrethrin (produced by Chrysanthemum cinerariaefolium flowers, primar-ily grown in Kenya) or Bt (produced by Bacillus thuringiensis bacteria). these compounds are less toxic than most synthetic pesticides, but their over-use can cause resurgence and the evolution of resis-tant insects. Some pest insects have already evolved resistance to Bt, including the Bt produced in the leaves of genetically modified crop plants.
Some harmless chemicals can be used for pest control. One such chemical is soap. When soap is sprayed on aphid infestations, the aphid exoskeletons are disrupted, which kills them. conventional pesti-cides, when mixed with soap, penetrate insects more effectively, which allows a reduced use of pesticides.
Pests can be controlled by reducing host den-sity. in a huge field of corn (a monoculture), every plant is a source of food for corn earworms. in smaller patches of corn, interspersed with patches of other crops such as soybeans, the corn earworm populations cannot grow as rapidly. Pest popula-tions can be controlled in time as well as in space. crop rotation reduces the growth of pest popula-tions. if earworms infest a cornfield, they may hatch the next year and discover that the field is now a soybean field, and many of them will die.
Pests can be controlled by encouraging predator populations. if agricultural fields are surrounded by fencerows of trees or shrubs, predatory insects or birds can live in them and fly out to capture pests
xviii+598_Biodiversity_F.indd 327 11/17/11 1:13 PM

first entry328328 integrated pest management
from the fields before returning to safety. it is impor-tant that the fields be small; predators will not fly long distances to the middle of a large field. the fencerow trees can also help to prevent wind and water erosion, an added benefit beyond pest management. natural predators can be important to an agricultural econ-omy. india banned frog exports in 1987 when the profits of selling them became less than the costs of pesticides to make up for insects the frogs ate.
Pests can be controlled by manipulating their behavior. Pest insects are not very smart and can be easily tricked; they often do whatever their air-borne hormones (pheromones) tell them to do. two important uses of synthetic pheromones in insect control are as follow:
● Mass trapping. Pheromone bait can attract insect pests to box traps. Millions of bark and other beetles have been trapped in this manner in the forests of northwestern north america. Pheromone bait has also been important in controlling codling moths (Cydia pomonella), whose caterpillars are a serious pest of apples and pears. it is esti-mated that pheromone traps killed 4.5 bil-lion spruce bark beetles (Ips typographus) in norway during a major infestation in 1980. Pheromone traps have also been used against the tomato pinworm, Keiferia lycop-ersicella, in Mexico.
● Mating disruption. a dilute suspension of pheromones, when sprayed in an agricul-tural field, can confuse male moths, and they cannot find the females.
Pheromones are effective at extremely low con-centrations, and synthetic pheromones are not pro-hibitively expensive.
The only way for pesticides to remain effective is to use them sparingly. Most environmentalists, following the lead of Rachel carson, did not oppose the frugal use of pesticides. in fact, this is the only way that the pesticides can remain effective: if the pesticides are rarely used, natural selection cannot favor the evolution of pest populations that resist them. no matter how carefully iPM is used, pest outbreaks will happen occasionally; for those times, pesticides are effective because iPM was practiced between pest outbreaks. carson opposed the blan-keting of the entire countryside with mirex, which was a pesticide intended to control invasive fire ants; instead, she called for the use of mirex by workers with backpack sprayers who could deliver the mirex directly to the ant mounds. the mirex concentration on the mound would be high, but the countryside as a whole would have a relatively low concentra-
tion. DDt, a pesticide notorious for accumulating in the food chain, is used safely in africa to control mosquitoes that spread malaria, because it is used just around houses (direct human contact with small amounts of it is not harmful) and because the inhab-itants of the houses also use mosquito nets when they sleep.
iPM is complex. it is not as simple as spraying. it requires the simultaneous use of many different tech-niques and an understanding of biology. it requires that a farmer think of a farm as an ecological com-munity. it requires that the public health official think of nonchemical ways to prevent the growth of mosquito populations. For example, old junk left out in the open can accumulate pools of water that are perfect breeding places for mosquitoes. that is, there is a connection between the cleanup of old junk and the spread of diseases such as the West nile virus. However, the scientific principles are not dif-ficult to understand, and many of them are simply common sense. the principles of iPM are not too difficult to implement in any situation.
Many insect pests have been imported without their natural parasites or predators. it is tempting to think about importing their parasites or predators as biological control agents. this has worked in the past. the cottony cushion scale Icerya purchasi, from aus-tralia, became a citrus pest in california in the early 20th century. it was brought under control by the vidalia beetle Rodolia cardinalis, also imported from australia. there are, however, other not so successful examples of invasive species that were introduced for the purpose of biological control of another invasive species, and this practice is now uncommon.
iPM does not have a perfect record of success. its success, however, is comparable to that of reli-ance on pesticides. agricultural scientist Matthew thomas indicates that many iPM programs have been ineffective because they were not integrated enough: they depended too heavily on just one com-ponent, usually the chemical one. He wrote, “the single solution, ‘magic bullet’ approach has under-mined our basic understanding of how individual components actually work and has oriented iPM toward quick fixes, often underpinned by commer-cial incentives.”
Rare species are particularly vulnerable to the overuse of pesticides. a pesticide application that will decimate the populations of common preda-tory insects and birds only temporarily and of pest insects even more briefly can eradicate a rare species. iPM is therefore not just a good idea for agriculture and for the control of disease but an essential com-ponent of protecting biodiversity.
See also biomagnification; carson, Rachel; invasive species; resistance, evolution of.
xviii+598_Biodiversity_F.indd 328 11/17/11 1:13 PM

invasive species 329
Further readingBianchi, F., et al. “Sustainable pest regulation in agricul-
tural landscapes: a review on landscape composition, biodiversity, and natural pest control.” Proceedings of the Royal Society of London B (Biology) 273 (2006): 1,715–1,727.
Bloomers, l. H. M. “integrated pest management in euro-pean apple orchards.” Annual Review of Entomology 39 (1994): 213–241.
cardé, R. t., and a. K. Minks. “control of moth pests by mating disruption.” Annual Review of Entomology 40 (1995): 559–585.
cuyno, leah c. M., et al. “economic analysis of envi-ronmental benefits of integrated pest management: a Philippine case study.” Agricultural Economics 25 (2005): 227–233.
Donald, P. F., et al. “agricultural intensification and the collapse of europe’s farmland bird populations.” Pro-ceedings of the Royal Society of London B (Biology) 268 (2001): 25–29.
environmental Protection agency. “integrated pest man-agement (iPM) principles.” available online. URl: http://www.epa.gov/pesticides/factsheets/ipm.htm. accessed august 8, 2011.
Flint, Hollis M., and charles c. Doane. “Understanding semiochemicals with emphasis on insect sex pher-omones in integrated pest management programs.” available online. URl: http://ipmworld.umn.edu/chapters/flint.htm. accessed august 2, 2010.
Hendrix, P. F., et al. “Detritus food webs in conventional and no-tillage agroecosystems.” BioScience 36 (1986): 374–380.
Holt, J., et al. “Strategies of insecticide use to avoid resur-gence of Nilaparvata lugens (Homoptera: Delphaci-dae) in tropical rice: a simulation analysis.” Journal of Economic Entomology 85 (1992): 1,979–1,989.
Johnson, David R., et al. “controlling weeds with phytopatho-genic bacteria.” Weed Technology 10 (1996): 621–624.
liu, tong-Xian, and Philip a. Stansly. “insecticidal activ-ity of surfactants and oils against silverleaf whitefly (Bemisia argentifolii) nymphs (Homoptera: aleyrodi-dae) on collards and tomato.” Pest Management Sci-ence 56 (2000): 861–866.
nyffeler, M., and K. D. Sutherland. “composition, abun-dance, and pest control potential of spider communi-ties in agroecosystems: a comparison of european and US studies.” Agriculture Ecosystems and Environment 95 (2003): 579–612.
Ogol, c., et al. “Maize stem borer colonization, establish-ment, and crop damage levels in a maize-leucaena agroforestry system in Kenya.” Agriculture Ecosys-tems and Environment 76 (1999): 1–15.
Paoletti, Maurizio G., and David Pimentel. “environmental risks of pesticides vs. genetic engineering for agricul-tural pest control.” Journal of Agricultural and Envi-ronmental Ethics 12 (2000): 279–303.
Perfecto, ivette, and R. Snelling. “Biodiversity and the transformation of a tropical agroecosystem—ants in coffee plantations.” Ecological Applications 5 (1995): 1,084–1,097.
Pollard, K. a., and J. M. Holland. “arthropods within the woody elements of hedgerows and their distribu-tion pattern.” Agricultural and Forest Entomology 8 (2006): 203–211.
Richards, a. J. “Does low biodiversity resulting from mod-ern agricultural practice affect crop pollination and yield?” Annals of Botany 88 (2001): 165–172.
Rutledge, claire e., et al. “Soybean aphid predators and their use in integrated pest management.” Annals of the Entomological Society of America 97 (2004): 240–248.
Salom, S. M., et al. “effect of an inhibitor-based sup-pression tactic on abundance and distribution of southern pine beetle (coleoptera: scolytidae) and its natural enemies.” Journal of Economic Entomology 88 (1995): 1,703–1,716.
Settle, W. H., et al. “Managing tropical rice pests through conservation of generalist natural enemies and alter-native prey.” Ecology 77 (1996): 1,975–1,988.
Smith, n. J. H., et al. “agroforestry developments and potential in the Brazilian amazon.” Land Degradation and Rehabilitation 6 (1995): 251–263.
thomas, Matthew B. “ecological approaches and the development of ‘truly integrated’ pest management.” Proceedings of the National Academy of Sciences USA 96 (1999): 5,955–5,951.
Vibe-Petersen, H., et al. “effects of predation and dispersal on Mastomys natalensis population dynamics in tan-zanian maize fields.” Journal of Animal Ecology 75 (2006): 213–220.
invasive species invasive species are species that proliferate within an ecological community and disrupt it. Usually invasive species are exotic, which means they are not native to the ecologi-cal community. in some cases, a native species can become invasive if ecological processes that once kept its populations in check are disrupted. impor-tant examples of invasive native species are two spe-cies of cedars in the southern Great Plains of north america.
When a species enters into a community from outside, its most likely fate is extinction. the cli-matic and microclimatic conditions in the new habi-tat will probably be unsuitable for the introduced species. even if the new species survives, its popula-tions may grow slowly, and it may be eliminated by competition with the better adapted native species. in some cases, however, an alien species may find the new habitat to be ideal for wild proliferation. the native range of the introduced species may have
xviii+598_Biodiversity_F.indd 329 11/17/11 1:13 PM

first entry330 invasive species 330
been very small, while its new range can be very extensive. the Monterey pine (Pinus radiata) was native to three small forest patches in california, but upon its introduction to new Zealand it spread over large areas.
the rapid spread of introduced species has often been caused by human activities, as in these examples:
● Most of the major american agricultural and urban weeds, including the ubiquitous dandelion (Taraxacum officinale), came from europe, probably because their seeds mixed with those of crop and garden species.
● Plants and animals may be brought to a new location as livestock, pets, or ornamen-tals. Ring-necked pheasants (Phasianus col-chicus) were brought from china as game birds. Purple loosestrife (Lythrum salicaria) was an ornamental plant, and starlings were pets, brought to america from europe. One of the most persistent of american weeds, the velvetleaf Abutilon theophrasti, was brought to america because its beautiful immature fruits could be used to make dec-orations on slabs of butter, hence its other common names, stampweed and butter-print. in each case, organisms escaped and experienced rapid population growth. Many feral livestock animals now roam free, such as goats and pigs in Hawaii and donkeys in Death Valley (see figure).
● aquatic organisms live in bilgewater in ships that cross the ocean and escape when bilgewater is dumped. eurasian watermil-foil (Myriophyllum spicatum) and the zebra mussel (Dreissena polymorpha), both of which are proliferating in many american waterways, may have been introduced in this way. the asian tiger mosquito, Aedes albopictus, is a vector of viral diseases. it came to north america in pools of water that had accumulated in tires being shipped from asia to the United States.
● new species are sometimes deliberately released because humans want to alter the natural environment. australian melaleuca trees (Melaleuca quinquenervia) were released into the everglades in order to dry them up for development, something they continue to successfully do.
● Deliberate introductions were sometimes intended for recreational purposes. Red foxes (Vulpes vulpes) were introduced into australia so that white immigrants could hunt them, as they had in england; starlings were brought from europe and released in new york city because someone wanted to have all of the birds mentioned in Shake-speare in north america.
● asian carp were raised in enclosed ponds along the Mississippi River. Floods allowed some of them to escape, and they spread northward. electrical barriers failed to pre-
Donkeys run wild in Death Valley, California. Their populations grew from animals that were released by borax miners in the early 20th century. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 330 11/17/11 1:13 PM

invasive species 331
vent their spread, through artificial water-ways, into the Great lakes.
● two species of cedars, the eastern red cedar (Juniperus virginiana) and the ashe juniper (J. ashei) are native to the southern Great Plains but were relatively rare in the grass-lands before fire suppression and deforesta-tion began in the late 19th century. today, eastern red cedar is invading many ranch-lands, and the ashe juniper grows abun-dantly in mountain areas where native oak forests have been cut down. they are con-sidered invasive species even though they are locally native.
in some cases, species were introduced in a sincere but misguided attempt to control other invasive species. cane toads (Bufo marinus) were introduced into australia to control agricultural pests, but they did not do so. coatimundis (Nasua nasua), relatives of the raccoon, were introduced on some Pacific islands, such as the Juan Fernandez islands, to control rats, but the coatis preyed mostly upon ground-nesting native birds. the birds had evolved in the absence of native predators. an afri-can land snail (Achatina fulica) was introduced into the Society islands, near tahiti. then a predatory snail (Euglandina rosea) was introduced to control it. the predatory snail instead mostly ate native Parula snails, causing most of them to become extinct. Mosquito fish (genus Gambusia) have been introduced into many freshwater wetlands so that they would eat mosquito larvae; in some cases, this has been successful, while in others, the fish have themselves caused ecological problems by displac-ing native fishes. the ladybird beetle Harmonia axyridis was introduced into europe to control aphids; it ate aphids but also ate native ladybird beetles.
in some cases, alien species invade new territory by natural means, but those means are enhanced by human activity. Microbes and the spores of fungi that kill agricultural crops can spread with dust in the wind even from one continent to another. Human activity, including global warming, has caused soil erosion and perhaps stronger storms, which has caused more dust, and spores, to travel to new locations.
in nearly all cases, the new species was intro-duced while the predators and parasites that would normally have held its populations in check were not. For example, red imported fire ants (Solenopsis invicta) are native to South america, where native phorid flies are parasitic upon them and limit their population growth. in the southern United States, the fire ants are spreading but the flies have not established permanent populations. Once the exotic (continues)
major invasive speCies in the united states*
Aquatic plants
Alternanthera philoxeroides (alligatorweed)Arundo donax (giant reed)Caulerpa taxifolia (Caulerpa seaweed)Didymosphenia geminata (didymo)Egeria densa (Brazilian waterweed)Eichhornia crassipes (water hyacinth)Hydrilla verticillata (hydrilla)Ipomoea aquatica (water spinach)Lythrum salicaria (purple loosestrife)Melaleuca quinquenervia (melaleuca)Myriophyllum spicatum (watermilfoil)Phragmites communis (common reed)Pistia stratiotes (water lettuce)Salvinia molesta (salvinia)Trapa natans (water chestnut)
Aquatic animals
Alosa pseudoharengus (alewife)Carcinus maenas (European green crab)Channa argus (snakehead fish)Didemnum vexillum (sea squirt)Dreissena bugensis (quagga mussel)Dreissena polymorpha (zebra mussel)Eriocheir sinensis (Chinese mitten crab)Gymnocephalus cernuus (Eurasian ruffe)Monopterus albus (Asian swamp eel)Myocastor coypus (nutria)Neogobius melanostomus (round goby)Oronectes rusticus (rusty crawfish)Petromyzon marinus (sea lamprey)Potamypyrgus antipodarum (New Zealand mud snail)Pterois volitans (lionfish)Pylodictus olivaris (flathead catfish)Rapana venosa (veined rapa whelk)Rana catesbeiana (bullfrog)
Terrestrial plants
Ailanthus altissima (tree of heaven)Alliaria petiolata (garlic mustard)Bromus tectorum (cheatgrass)Carduus nutans (musk thistle)Celastrus orbiculatus (Oriental bittersweet)Centaurea diffusa (diffuse knapweed)Centaurea calcitrapa (purple star thistle)Centaurea solstitialis (yellow star thistle)Centaurea stoebe (spotted knapweed)Cirsium arvense (Canada thistle)Cynoglossum officinale (hounds-tongue)Cystisus scoparius (Scotch broom)Dipsacus fullonum (common teasel)Elaeagnus umbellata (autumn olive)Elaeagnus angustifolia (Russian olive)Elymus repens (quackgrass)
xviii+598_Biodiversity_F.indd 331 11/17/11 1:13 PM

first entry332 invasive species 332
species has arrived, native species may accidentally help the invader to spread; for example, muskrats help spread the underground stems of loosestrife.
the introduced species have many profound influences on the community they invade. they may displace native species or even drive them to extinc-tion, as in these examples:
● Some invasive animals harm native animals. Red imported fire ants (Solenopsis invicta) were introduced into the United States about 1930. they have spread through-out the southern states and have recently entered california as well. they cause pain-ful stings, kill some ground-dwelling wild animals, and damage some equipment and some agricultural crops. Ferocious african honeybees have displaced european honey-bees (both varieties of Apis mellifera) from the tropical zones of the americas, reach-ing as far north as southern texas. african “killer bees” have had a longer history of exploitation by humans and other predators, and their evolutionary response has been a tendency to attack potentially dangerous animals. african bee queens were allowed to escape from a breeding experiment in Brazil in the 1960s and have mixed with european honeybee populations. Unable to tolerate cold winters, the african bees will probably not extend beyond the southern-most tier of states. european honeybees are themselves an invasive species in north and South america, displacing many native bee species. a population explosion of a comb jelly from chesapeake Bay, Mnemiopsis leidyi, has destroyed some fisheries in the caspian Sea. toxic cane toads in austra-lia have nearly driven the native marsupial quoll (Dasyurus hallucatus) to extinction because the marsupial does not recognize the toad as poisonous.
● Some invasive species reproduce faster and outcompete native species. the nile perch (Lates niloticus) was introduced into lake Victoria in africa, where it has driven many of the native species of fish nearly to extinc-tion. introduced birds such as the myna bird have driven the native birds of Hawaii away from the coastlands and many into extinc-tion. the few remaining native Hawaiian birds can only be found in the highlands. european grasses, originally spread in the droppings of horses that escaped from the Spaniards, have displaced the native cali-fornia grasses. Water lettuce and water hya-cinth fill the surface waters and crowd out
major invasive speCies in the united states* (continued)
Euphorbia esula (leafy spurge)Fallopia japonica (Japanese knotweed)Heracleum mantegazzianum (giant hogweed)Hypericum perforatum (Saint-John’s-wort)Imperata cylindrica (cogongrass)Lepidium appelianum (giant whitetop)Lepidium draba (whitetop)Linaria dalmatica (Dalmatian toadflax)Linaria vulgaris (common toadflax)Lonicera japonica (Japanese honeysuckle)Lygodium microphyllum (Old world climbing fern)Onopordum acanthium (Scotch thistle)Paulownia tomentosa (princess tree)Persicaria perfoliata (mile-a-minute weed)Pueraria montana var. lobata (kudzu)Rhaponticum repens (Russian knapweed)Rosa multiflora (multiflora rose)Solanum viarum (tropical soda apple)Sorghum halepense (Johnson grass)Striga asiatica (witchweed)Taeniatherum caput-medusae (medusahead)Tamarix spp. (salt cedar)Triadica sebifera (tallow tree)Vitex rotundifolia (beach vitex)
Terrestrial invertebrates
Achatina fulica (African giant snail)Adelges tsugae (hemlock woolly adelgid)Aedes albopictus (Asian tiger mosquito)Agrilus planipennis (emerald ash borer)Anastrepha ludens (Mexican fruit fly)Apis mellifera scutellata (Africanized honeybee)Anoplophora glabripennis (Asian long-horned beetle)Anoplophora chinensis (Citrus long-horned beetle)Bemisia argentifolii (silverleaf whitefly)Cactoblastis cactorum (cactus moth)Ceratitis capitata (Mediterranean fruit fly)Coptotermes formosanus (Formosan subterranean termite)Diaphorina citri (Asian citrus psyllid)Diuraphis noxia (Russian wheat aphid)Epiphyas postvittana (light brown apple moth)Heterodera glycines (soybean cyst nematode)Homalodisca coagulata (glassy-winged sharpshooter)Ips typographicus (European spruce bark beetle)Lymantria dispar (gypsy moth)Maconellicoccus hirsutus (mealybug)Solenopsis invicta (red imported fire ant)
Terrestrial vertebrates
Boiga irregularis (brown tree snake)Bufo marinus (cane toad)Sturnus vulgaris (starling)Sus scrofa (wild boar)
*From National Agricultural Library. This table does not include agricultural weeds or species whose potential to spread is limited (e.g., burros in Death Valley).
xviii+598_Biodiversity_F.indd 332 11/17/11 1:13 PM

invasive species 333
phytoplankton. Populations of melaleuca and loosestrife outgrow those of native plants because they produce millions of seeds. introduced african grasses inhibit the germination of native Hawaiian tree seed-lings. the american gray squirrel (Sciurus carolinensis) has displaced the native red squirrel (Sciurus vulgaris) in many areas of europe.
● invasive species can alter ecosystems, mak-ing them unsuitable for native species. cedars cast dense shade and compete with other plants for water, which they transpire liberally into the air. native species can-not compete well with them for light and moisture. Fire destroys cedars. But when a fire gets started in the dense cedar brush, it is hot enough to destroy native forest trees. the leaves of the wax myrtle (Myrica cer-ifera), which has invaded Florida wetlands, do not decompose well and have literally clogged up the aquatic food chain. the deep litter of casuarina trees (Casuarina equisetifolia) introduced into Hawaii from Southeast asia inhibits the growth of practi-cally everything under their canopy. Kudzu vine (Puereria montana), introduced from asia, drapes over and kills many trees in the southeastern United States (see photo-graph). Salt cedars (species of Tamarix; not related to junipers) were introduced from the Middle east and have spread along riv-ers throughout the western United States, smothering entire riverbanks and driving out the native plant species.
invading species may create many disturbances. Feral pigs in Hawaii create disturbances in the rain forest, in which the native plant species cannot grow well. However, introduced plants, such as the straw-berry guava, thrive in these disturbances. When grasses are first introduced into a community that previously had none, they can create a fire hazard. Fires deplete the populations of species that are not fire-resistant and further encourage the spread of the grasses, which can grow back after the fires. in places where fires were a common part of the natu-ral ecosystem, fire suppression has encouraged the spread of invasive species. ellwood cooper planted 50,000 Eucalyptus seedlings at his ranch near Santa Barbara, california, in 1876. today eucalyptus groves grow where no trees formerly lived. they draw down the water table and can burn explosively. their fragrant leaf litter accumulates in deep layers because local decomposers cannot consume them. Gum from their flowers clogs the faces and bills of pollinating birds.
invading plant species reduce plant species diver-sity in the community that they invade, and they do so by enhancing the total amount of productiv-ity. invasive plants enhance net primary production (total net photosynthetic weight gain) by 80 percent. this may occur in part because the invading plants stimulate microbes to release nitrogen into a form that can be readily absorbed and used and in part because invading plants have thinner leaves—this allows more photosynthesis to occur with a given amount of leaf tissue.
exotic predators have disrupted whole food chains. For instance, the brown tree snake (Boiga irregularis) has severely depleted native bird popula-tions on Guam.
invading species can have severe economic impacts on agriculture and even on public health, as well as on nature preserves. the eastern red cedar and the salt cedar transpire enough water that underground water supplies are affected. the ashe juniper releases pollen that is particularly allergenic. Zebra mussels can clog water intake systems and interfere with municipal water supplies. cane toads have poisonous skin.
Much money and effort are spent in attempts to control the spread of invasive species. it may be impossible to eradicate any invasive species, but in some cases conservation agencies have limited their spread. Sometimes this requires putting up fences. Fences have limited the spread of feral hogs in Hawaii, allowing native vegetation to grow back on one side of the fence and crowding the hogs onto the other side of the fence where they can be more easily shot from helicopters.
in order to control the spread of invasive spe-cies, it is necessary to monitor them. By the time the invasive organisms are easily visible, it may be too late to do anything about them. the asian carp in
Do not park your truck too long in the American state of Georgia, or kudzu (the invasive Puereria montana, a plant from East Asia) will grow all over it. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 333 11/17/11 1:13 PM

first entry334 invasive species 334
illinois, mentioned previously, were first discovered not by observing carp outside the electric barrier but by detecting carp Dna in the water upstream from the fence.
Governments now inspect incoming airplanes, ships, and cargo as closely as time and funds permit. in the United States, the animal and Plant Health inspection Service (aPHiS) is the agency that carries out these inspections.
Often the means of controlling invasive species, such as the use of pesticides and herbicides, are as dangerous as they are ineffective, such as mass hunt-ing campaigns. australians organize mass citizen hunting campaigns against cane toads, with little visible impact.
Scientists have investigated biological controls as a method of bringing escaped exotic species under control. loosestrife is not a pest in europe, where its populations are controlled by beetles. Scientists are now attempting to introduce those beetles to north america. Biological controls, unlike pesti-cides, can be permanent. as noted previously, bio-logical control agents run the risk of becoming pests themselves.
eventually, other species (either native or them-selves introduced) will evolve ways of eating the invaders, bringing their populations into check. the time required for this process, however, is likely to make it meaningless to human concerns.
See also communities, ecological.
Further readingagricultural Research Service, United States Department
of agriculture. “Biological control sends scurrying scourge scrambling.” available online. URl: http://www.ars.usda.gov/is/aR/archive/may03/biol0503.htm. accessed august 1, 2010.
Burdick, alan. “the truth about invasive species.” Dis-cover, May 2005, 34–41.
coote, t., and e. loeve. “From 61 species to five: endemic tree snails of the Society islands fall prey to an ill-judged biological control programme.” Oryx 37 (2003): 91–96.
cox, William. Alien Species and Evolution: The Evolution-ary Ecology of Exotic Plants, Animals, Microbes, and Interacting Native Species. Washington D.c.: island Press, 2004.
Daehler, curtis c. “Performance comparisons of co-occur-ring native and alien invasive plants: implications for conservation and restoration.” Annual Review of Ecol-ogy, Evolution, and Systematics 34 (2003): 183–212.
Davis, Mark a. Invasion Biology. new york: Oxford Uni-versity Press, 2009.
Duncan, Richard P., et al. “the ecology of bird introduc-tions.” Annual Review of Ecology, Evolution, and Sys-tematics 34 (2003): 71–98.
enserink, Martin. “a mosquito goes global.” Science 320 (2008): 864–867.
Forseth, irwin n., and a. F. innis. “Kudzu (Puereria mon-tana): History, physiology, and ecology combine to make a major ecosystem threat.” Critical Reviews in Plant Sciences 23 (2004): 401–413.
Greenway, Roger. “nOaa solution to lionfish invasion, eat them!” Environmental News Network, august 8, 2010. available online. URl: http://www.enn.com/wildlife/article/41636. accessed august 8, 2011.
Hendrix, Paul F., et al. “Pandora’s box contained bait: the global problem of introduced earthworms.” Annual Reviews of Ecology, Evolution, and Systematics 39 (2008): 593–613.
Hobbs, R. J., and laura F. Huenneke. “Disturbance, diver-sity, and invasion—implications for conservation.” Conservation Biology 6 (1992): 324–337.
isard, S. a., et al. “Principles of the atmospheric pathway for invasive species applied to soybean rust.” BioSci-ence 55 (2005): 851–861.
Keller, Reuben P., et al., eds. Bioeconomics of Invasive Species: Integrating Ecology, Economics, Policy, and Management. new york: Oxford University Press, 2009.
Krajick, Kevin. “Winning the war against island invaders.” Science 310 (2005): 1,410–1,413.
liu, Shu-Sheng, et al. “asymmetric mating interactions drive widespread invasion and displacement in a whitefly.” Science 318 (2007): 1,769–1,772. Summa-rized by Reitz, Stuart R., “invasion of the whiteflies.” Science 318 (2007): 1,733–1,734.
Mooney, Harold a., et al., eds. Invasive Alien Species: A New Synthesis. Washington, D.c.: island Press, 2005.
national agricultural library. “national invasive species information center.” available online. URl: http://www.invasivespeciesinfo.gov. accessed august 8, 2011.
O’Donnell, S., et al. “conditioned taste aversion enhances the survival of an endangered predator imperiled by a toxic invader.” Journal of Applied Ecology 47 (2010): 558–565.
Parker, John D., et al. “Opposing effects of native and exotic herbivores on plant invasions.” Science 311 (2006): 1,459–1,461.
Peterson, a. townsend, et al. “Predicting the potential invasive distributions of four alien plant species in north america.” Weed Science 51 (2003): 863–868.
Pyke, Graham H. “Plague minnow or mosquito fish? a review of the biology and impacts of Gambusia spe-cies.” Annual Reviews of Ecology, Evolution, and Sys-tematics 39 (2008): 171–191.
Romero, Simon. “War in the Pacific: it’s hell, espe-cially if you’re a goat.” New York Times, May 1, 2007. available online. URl: http://www.nytimes.
xviii+598_Biodiversity_F.indd 334 11/17/11 1:13 PM

island biogeography 335
com/2007/05/01/world/americas/01galapagos.html?pagewanted=print. accessed august 8, 2011.
Rout, Marnie e., and Ragan M. callaway. “an invasive plant paradox.” Science 324 (2009): 734–735.
Ruesink, Jennifer l., et al. “introduction of non-native oys-ters: ecosystem effects and restoration implications.” Annual Review of Ecology, Evolution, and Systemat-ics 36 (2005): 643–689.
Sax, Dov F., John J. Stachowicz, and Steven D. Gaines, eds. Species Invasions: Insights into Ecology, Evolution, and Biogeography. Sunderland, Mass.: Sinauer associ-ates, 2005.
Snyder, William e., and edward W. evans. “ecological effects of invasive arthropod generalist predators.” Annual Review of Ecology, Evolution, and Systemat-ics 37 (2006): 95–122.
Suarez, a. V., et al. “effects of fragmentation and invasion on native ant communities in coastal southern califor-nia.” Ecology 79 (1998): 2,041–2,056.
Svoboda, elizabeth. “the unintended consequences of changing nature’s balance.” New York Times, February 17, 2009. available online. URl: http://www.nytimes.com/2009/02/17/science/17isla.html. accessed august 8, 2011.
tennesen, Michael. “When juniper and woody plants invade, water may retreat.” Science 322 (2008): 1,630–1,631.
terrill, Geiridwen. Unnatural Landscapes: Tracking Inva-sive Species. tucson: University of arizona Press, 2007.
todd, Kim. Tinkering With Eden: A Natural History of Exotic Species in America. new york: norton, 2002.
traveset, anna, and D. M. Richardson. “Biological inva-sions as disrupters of plant reproductive mutualisms.” Trends in Ecology and Evolution 21 (2006): 208–216.
United States Department of agriculture. “animal and Plant Health inspection Service: Safeguarding ameri-can agriculture.” available online. URl: http://www.aphis.usda.gov. accessed august 8, 2011.
Vogt, J. t., Stanley a. Rice, and Steven a. armstrong. “Seed preferences of the Red imported Fire ant (Hymenop-tera: Formicidae) in Oklahoma.” Journal of Entomo-logical Science 38 (2003): 696–698.
Vredenburg, V. t. “Reversing introduced species effects: experimental removal of introduced fish leads to rapid recovery of a declining frog.” Proceedings of the National Academy of Sciences USA 101 (2004): 7,646–7,650.
Wiles, G. J., et al. “impacts of the brown tree snake: Patterns of decline and species persistence in Guam’s avifauna.” Conservation Biology 17 (2003): 1,350–1,360.
Williams, Susan l., and Jennifer e. Smith. “a global review of the distribution, taxonomy, and impacts of intro-duced seaweeds.” Annual Review of Ecology, Evolu-tion, and Systematics 38 (2007): 327–359.
island biogeography Biogeography is the study of what kinds of organisms grow in dif-ferent geographical locations. even before charles Darwin and alfred Russel Wallace, islands were of special interest because their isolation has allowed the evolution of many endemic species. it was the uniqueness of island species but also their simi-larity to the species of the nearby mainland that helped charles Darwin formulate the theory of evo-lution. But island biogeography remained largely a collection of interesting evolutionary stories until ecologists Robert H. Macarthur and edward O. Wilson formalized it into a mathematically based theory.
an island may have no species when it first forms. new species disperse to the island from nearby mainland areas and evolve on the island. Meanwhile, species become extinct as well. two factors affect the extent to which these processes occur:
● Distance effect islands that are more dis-tant from the mainland are less likely to receive immigrants.
● Area effect larger islands are able to maintain a greater number of species than smaller islands.
the consequences of these processes are that smaller and/or more distant islands will have fewer species and that an island may reach an equilib-rium number of species. new species will continue to arrive or evolve, and old species will become extinct, leaving the equilibrium number of species unchanged.
the area effect results from the fact that larger islands have a greater number of different kinds of habitats in which different species can live. this results in the species-area relationship. a very small island might force different species to compete, whereas on a larger island the species could special-ize on different ways of obtaining resources. even though the theory of island biogeography mainly concerns the arrival and survival of species that already exist, there is also a longer-term evolution-ary dimension to it. a larger island will allow the evolution of more new species than will a smaller island. the Galápagos islands collectively have enough area that 13 species of finches have been able to evolve different specializations, whereas cocos island does not have enough area for the finches to form separate breeding populations. the cocos island finch remains a single species even while different individuals obtain food in differ-ent ways, some of them similar to the specializa-tions of the different Galápagos finches. the greater
xviii+598_Biodiversity_F.indd 335 11/17/11 1:13 PM

first entry336 island biogeography 336
species capacity of a larger island not only allows more species to evolve but prevents those species from becoming extinct as often. the species-area relationship, however, is not linear. as islands get bigger, the increased area is not associated with an increased number of new habitat types. that is, as islands get larger, the number of species tends to level off. the curve never reaches a maximum num-ber, however, unless the island becomes nearly as large as a continent itself.
it might appear simple enough to test island biogeography theory by comparing species richness on different islands, but modern species richness is produced both by ecological and evolutionary processes. How is it possible to test island bioge-ography on an ecological timescale short enough that new species will not evolve? ecologist Daniel Simberloff, working with edward O. Wilson, tried an experimental approach. they identified differ-ent mangrove islands (islands formed by mangrove tree clumps growing out of shallow water) in the Gulf of Mexico, of many different sizes and dis-tances from the mainland. they then used insecti-cides to kill all of the arthropods on these islands. they waited for arthropods to colonize the islands from the mainland. they then counted the number of arthropod species on each island. they focused on arthropods because vertebrates were unlikely to be permanent residents on such small islands. also, arthropods have short enough generation times that the equilibrium species number could be reached within a manageable length of time. they did find that smaller islands and more dis-tant islands had fewer arthropod species after a few years. Of course, there were other factors that affected the number of species on each island, including the hurricanes that occasionally came through their research area.
the concepts of island biogeography are important for ecological islands as well as for geo-graphical islands. Some habitats, especially those that have many endemic species, are isolated pock-ets surrounded by habitat that is unsuitable for those species. the surrounding area might as well be an ocean, for all the difference it makes to the species in the small habitats. the dry temperate oak forests on mountaintops surrounded by the chihuahuan Desert are called “sky islands.” in a more general sense, nearly every habitat can be thought of as an island surrounded by some other kind of habitats.
the consequences for conservation are impor-tant. Just how big does a protected area need to be? and where should protected areas be located? they need to be big enough to have microhabitat hetero-
geneity that will allow all of the species to survive. if a habitat type is protected by several small areas rather than one large one, these small areas need to be close enough together that a species, should it happen to become extinct in one of them, can be replaced by colonists from another one. One reason that conservation biologists use island biogeogra-phy analysis is that the option of protecting entire habitats usually does not exist. they have to make scientifically based arguments for even the smallest amount of habitat area that they ask the government to protect from development or exploitation. island biogeography theory allows them to make the best decisions for the protection of small areas that may allow as many species as possible to survive. island biogeography theory also helps to explain why habi-tat fragmentation can so severely damage the sur-vival prospects of rare species.
See also biogeography; endemic species; habitat fragmentation.
Further readingDavis, anthony M., and thomas F. Glick. “Urban ecosys-
tems and island biogeography.” Environmental Con-servation 5 (1978): 299–304.
Diamond, Jared M. “the island dilemma: lessons of modern biogeographic studies for the design of nat-ural reserves.” Biological Conservation 7 (1975): 129–146.
Drakare, Stina, et al. “the imprint of the geographical, evolutionary, and ecological context on species-area relationships.” Ecology Letters 9 (2005): 215–227.
Gilbert, F. S. “the equilibrium theory of island biogeog-raphy: Fact or fiction?” Journal of Biogeography 7 (1980): 209–235.
Harris, larry D. The Fragmented Forest: Island Biogeog-raphy Theory and the Preservation of Biotic Diversity. chicago: University of chicago Press, 1984.
Kadmon, Ronen, and H. Ronald Pulliam. “island bioge-ography: effect of geographical isolation on species composition.” Ecology 74 (1993): 977–981.
Macarthur, Robert H., and edward O. Wilson. Theory of Island Biogeography. Princeton, n.J.: Princeton Uni-versity Press, 1967.
Quammen, David. The Song of the Dodo: Island Biogeogra-phy in an Age of Extinctions. new york: Scribner, 1996.
Scheiner, Samuel M. “Six types of species-area curves.” Global Ecology and Biogeography 12 (2003): 441–447.
Simberloff, Daniel S. “equilibrium theory of island bioge-ography and ecology.” Annual Review of Ecology and Systematics 5 (1974): 161–182.
———, and lawrence G. abele. “Refuge design and island biogeographic theory: effects of fragmentation.” American Naturalist 120 (1982): 41–50.
xviii+598_Biodiversity_F.indd 336 11/17/11 1:13 PM

isolating mechanisms 337
Simberloff, Daniel S., and edward O. Wilson. “experi-mental zoogeography of islands: the colonization of empty islands.” Ecology 50 (1969): 278–296.
Whittaker, Robert J., and José María Fernandez-Palacios. Island Biogeography: Ecology, Evolution, and Con-servation. new york: Oxford University Press, 2007.
Zimmerman, B. l., and R. O. Bierregaard. “Relevance of the equilibrium theory of island biogeography and
species-area relations to conservation with a case from amazonia.” Journal of Biogeography 13 (1986): 133–143.
isolating mechanisms See speciation.
xviii+598_Biodiversity_F.indd 337 11/17/11 1:13 PM

338
jackson, Wes (1936– ) American Agricultural ecologist Wes Jackson is the founder and leader of the land institute, a Kansas organization that is the principal proponent of a biodiversity-based agricul-ture (natural systems agriculture).
Born June 15, 1936, on a farm in Kansas, Jack-son received a master’s degree in botany from the University of Kansas and a Ph.D. in genetics from north carolina State University. He established one of the earliest environmental studies programs in the United States while he was at the Sacramento campus of california State University. in 1976, he resigned and returned to Kansas to establish the land institute. the institute began with farmland and intact prairie, and Jackson wanted to use the prairie as a model for how agriculture could be transformed from an industrial mode to a natural and sustainable mode.
the land institute has attracted researchers from around the world who work either at the land institute or at their home institutions. For many years, the land institute sponsored undergraduate interns but now focuses on supporting graduate stu-dent research. Some land institute staff, including Jerry Glover, have received international recognition for their research.
Wes Jackson is widely recognized as one of the major ecological visionaries of modern times. His books tie in agricultural themes with every aspect of modern environmentalism. Life magazine listed Jackson as one of the 100 most important people of the 20th century, and Smithsonian magazine listed him as one of the 35 people who had made the great-est difference in the world in the 20th century.
See also natural systems agriculture.
Further readingJackson, Wes. Altars of Unhewn Stone: Science and the
Earth. Wooster, Ohio: Wooster Books, 2006.———. Becoming Native to This Place. Washington, D.c.:
counterpoint, 1994.———. Consulting the Genius of the Place: An Ecological
Approach to a New Agriculture. Washington, D.c.: counterpoint, 2010.
———. New Roots for Agriculture. lincoln, neb.: Univer-sity of nebraska Press, 1980.
janzen, daniel h. (1939– ) American Ecol-ogist, evolutionary biologist, conservationistDan Janzen is one of the most signifi cant fi gures in modern ecology and conservation. He is famous for his original insights, his creative ways of research-ing and expressing them, and his dynamic lectures about them. among his research contributions have been the following:
● ants (such as Pseudomyrmex ferruginea) protect acacia trees (e.g., Acacia cornigera) from herbivores, one of the prime examples of the evolution of mutualism.
● the seed dispersers of Spondias mombin, a tropical tree, became extinct at the end of the Pleistocene epoch, and today it depends on white-tailed deer to disperse its seeds.
● Herbivory is one of the reasons that there are so many species of trees in tropical forests.
● Because tropical rain forests have uniform temperatures throughout the year, tropi-cal organisms are more likely to experience even the slightly cooler conditions of tropi-
J
xviii+598_Biodiversity_F.indd 338 11/17/11 1:13 PM

Jurassic period, biodiversity in 339
cal mountains as a barrier to dispersal than are temperate organisms. that is, mountain passes “are higher” in the tropics.
● Decomposers produce chemicals that repel herbivores and scavengers that would oth-erwise eat them along with the old or dead plant or animal tissue. this is why “fruits rot, seeds mold, and meat spoils.”
even when later researchers have challenged some of his broad hypotheses, none have denied that they were creative.
Born in 1939 in Milwaukee, Wisconsin, Janzen graduated from the University of Minnesota in 1961 and earned his doctorate from the University of california at Berkeley four years later. Since 1965 he has been one of the leaders of the Organization for tropical Studies, which trains students in tropical biology. He taught at universities in Kansas, chi-cago, Venezuela, and Puerto Rico before joining the faculty at the University of Pennsylvania.
Janzen has been on the faculty of the University of Pennsylvania since 1976, but for many years he has served without remuneration as an adviser to the government of costa Rica. largely through his lead-ership, costa Rica has the best system of ecological reserves, relative to its size, of any country in the world, including a plan that connects the reserves together; as a result, one-quarter of costa Rica is protected or being restored. Janzen has incorporated the need for local inhabitants to use the land sustain-ably, rather than simply keeping people out of the parks. He is also leading an ambitious project to cat-alog the country’s biodiversity, including the use of Dna barcoding to reveal previously unrecognized biodiversity. in 1984 he received the crafoord Prize, given to scientists whose research is not in a category specified for the nobel Prize.
See also essay: Ghosts of Biodiversity Past; bio-diversity, patterns of; Dna studies of biodi-versity; habitat fragmentation; mutualism; Pleistocene extinction; restoration ecology; seed dispersal.
Further readingGhalambour, cameron K., et al. “are mountain passes
higher in the tropics? Janzen’s hypothesis revisited.” Integrative and Comparative Biology 46 (2006): 5–17.
Hebert, P. D. n., et al. “ten species in one: Dna barcoding reveals cryptic species in the neotropical skipper butter-fly Astraptes fulgerator.” Proceedings of the National Academy of Sciences 101 (2004): 14,812–14,817.
Janzen, Daniel H. 1966. “allelopathy by myrmecophytes: the ant Azteca as an allelopathic agent of Cecropia.” Ecology 50 (1969): 147–153.
——— “coevolution of mutualism between ants and acacias in central america.” Evolution 20 (1966): 249–75.
———. “escape of Cassia grandis l. beans from predators in time and space.” Ecology 52 (1971): 964–979.
———. “Gardenification of wildland nature and the human footprint.” Science 279 (1998): 1,312–1,313.
———. “Herbivores and the number of tree species in tropical forests.” American Naturalist 104 (1970): 502–528.
———. “Why fruits rot, seeds mold, and meat spoils.” American Naturalist 111 (1977): 691–713.
———. “Why mountain passes are higher in the tropics.” American Naturalist 101 (1967): 233–249.
——— and Paul S. Martin. “neotropical anachronisms: the fruits the gomphotheres ate.” Science 215 (1982): 19–27.
Sherratt, thomas n., et al. “Why fruits rot, seeds mold, and meat spoils: a reappraisal.” Ecological Modeling 192 (2006): 618–626.
Smith, M. a., et al. “extreme diversity of tropical parasitoid wasps exposed by iterative integration of natural his-tory, Dna barcoding, morphology, and collections.” Proceedings of the National Academy of Sciences USA 105 (2008): 12,359–12,364.
jurassic period, biodiversity in the Juras-sic period (210 to 140 million years ago) was the middle period of the Mesozoic era. it followed the triassic period. the Mesozoic era is also known as the age of Dinosaurs, because dinosaurs were the largest land animals during that time. Dinosaurs first evolved during the triassic period but became the largest, most diverse, and most abundant terres-trial vertebrates during the Jurassic period and into the cretaceous. Dinosaurs became extinct during the cretaceous extinction, which is one of the two most important mass extinctions in the history of the earth.
Climate. in the mild climate of the Jurassic period, swamps and shallow seas covered wide-spread areas. as in the carboniferous, atmospheric levels became very high, even though carbon diox-ide continued to create a greenhouse atmosphere (see table on page 340).
Continents. the continents formed by the breakup of Pangaea continued to separate. the northern continent (laurasia) consisted of what are today the northern continents (north america, europe, asia). the southern continent (Gondwana) consisted of what are today the southern continents (South america, africa, antarctica, australia), with the tethys Sea between them.
xviii+598_Biodiversity_F.indd 339 11/17/11 1:13 PM

first entry340340 Jurassic period, biodiversity in
multiCellular biodiversity in the jurassiC period
ConditionsAtmospheric oxygenAtmospheric carbon dioxideGlobal temperatureSea level
30% above modern level5.5 times modern level5°F (3°C) above modern level300 ft (100 m) higher than today
Approximate number of families*ContinentalMarineTotal
200450650
Examples of marine animalsBelemnobatis morinicusCycloprosopon dobrogeaErmoceras sp.Eryon actiformisGrendelius mordaxHybodus houtiensisLeptolepis talbragarensisMetriorhynchus geoffroyiPlesiosaurus dolichodeirus
ray (shark relative)crabcephalopod mollusklobsterichthyosaur reptilesharkbony fishsea crocodileplesiosaur reptile
Examples of continental animalsAllosaurus fragilisApatosaurus ajaxArchaeolepis maneArchaeopteryx lithographicaBharatagama rebbanensisBrachiosaurus altithoraxCastorocauda lutrasimilisCeratosaurus ingensCompsognathus longipesDiplodocus longusFrutafossor windscheffeliGrimmyia balticaIguanodon bemissartensisMesocordulia borealaMicroraptor zhaoianusMorganucodon watsoniPatarchaea muralisPterodactylus antiquusSiamochelys peninsularisSigmaboilus peregrinusStegosaurus amatusSupersaurus vivianaeTritylodon longaevusVelociraptor mongoliensisXyletoma nigricornis
large herbivorous dinosaurlarge herbivorous dinosaurbutterfly or mothfeathered dinosaurlizardlarge herbivorous dinosaurmammallarge predatory dinosaursmall predatory dinosaurlarge herbivorous dinosaurmammalflymedium herbivorous dinosaurdragonflyfeathered dinosaurmammalspiderpterosaurturtlecricket relativelarge herbivorous dinosaurlarge herbivorous dinosaurmammallike reptilefeathered dinosaurwasp
Examples of land plantsAraucarioxylon arizonicumBucklandia sp.Ginkgo sibiricaKomlopteris nordenskioeldiiSewardiodendron laxumWilliamsoniella coronata
conifercycadginkgoseed fernconiferbennettitalean seed plant
*Based on Benton.
xviii+598_Biodiversity_F.indd 340 11/17/11 1:13 PM

Jurassic period, biodiversity in 341
Marine life. Marine life continued to evolve:
● Invertebrates. the inverte-brates that predominated in the triassic oceans continued to proliferate. among the crustaceans, the fi rst lobsters and crabs evolved. One of the crabs, Cycloprosopon, had no legs.
● Vertebrates. Bony fishes and sharks continued to pro-liferate in the oceans. Rays, relatives of sharks, fi rst evolved during the Jurassic. Marine reptiles that evolved in the triassic continued to fl ourish; in addition, new groups of marine reptiles evolved: plesiosaurs (marine reptiles with very long necks) and sea crocodiles, some of them 10 feet (three m) long.
Continental life. Gymnosperms dominated the terrestrial forests, in which dinosaurs proliferated and in which the fi rst birds evolved.
● Plant life. conifers, such as relatives of the modern monkey puzzle tree, dominated the forests, which also contained many spe-cies of ginkgo and cycads. Araucarioxylon trees made up the forest now preserved at Petrifi ed Forest national Park in arizona. Seed ferns persisted but were becoming less common.
● Invertebrates. terrestrial invertebrates continued to diversify, especially insects such as fl ies, crickets, beetles, wasps, and dragonfl ies. the earliest lepidopterans (but-terfl ies and moths) are known from the Jurassic.
● Vertebrates. Freshwater fi shes, lungfi shes, and amphibians continued to evolve. Some reptiles such as turtles persisted. the fi rst liz-ards appeared during the Jurassic. Mammal-like reptiles were becoming less abundant, as true mammals evolved. although not as diverse as dinosaurs, mammals did come in numerous forms, including some beaverlike aquatic mammals (Castorocauda) and some that ate termites (Frutafossor).
the dominant land vertebrates of the Jurassic were the dinosaurs, which diversifi ed into many forms after the triassic extinction. Some herbivorous dinosaurs grew very large and had long necks that
allowed them to eat foliage from the tops of conifers. examples include Apatosaurus (formerly Bronto-saurus), Diplodocus, and Brachiosaurus; Supersau-rus, almost 100 feet (30 m) long and weighing 100 tons (metric tons), was nearly at the theoretical max-imum size that is possible for a terrestrial animal. Some herbivorous dinosaurs, such as Iguanodon,were smaller and could run on two legs. though there were some large predators such as Ceratosau-rus, many of the predators were small and rapid—and many of them had feathers. Small feathered dinosaurs were, in fact, the fi rst birds, of which 23 Jurassic genera have been found. the most famous is Archaeopteryx, but there were many others as well. it is not known which of them might have been the ancestor of more recent birds. Fossils of Archaeop-teryx were preserved with feather impressions in the surrounding rock. the fl ying pterosaurs (such as Pterodactylus) were a separate lineage of evolution and were not birds.
See also dinosaurs.
Further readingBenton, Michael J. “Dinosaur summer.” in The Book of
Life: An Illustrated History of the Evolution of Life on Earth, edited by Stephen J. Gould. new york: norton, 2001.
———. “Diversifi cation and extinction in the history of life.” Science 268 (1995): 52–58.
carpenter, K., and J. Mcintosh. “Upper Jurassic sauropod babies from the Morrison Formation.” in Dinosaur Eggs and Babies, edited by K. carpenter, K. F. Hirsch, and J. R. Horner. cambridge: cambridge University Press, 2007.
Feldmann, Rodney M., et al. “Ekalakia (Decapoda: Brachyura): the preservation of eyes links cretaceous crabs to Jurassic ancestors.” Journal of Paleontology82 (2008): 1,030–1,034.
Stegosaurus was a large armored dinosaur of the Jurassic period.
xviii+598_Biodiversity_F.indd 341 11/17/11 1:13 PM

first entry342 Jurassic period, biodiversity in 342
Foster, J. Jurassic West: The Dinosaurs of the Morrison For-mation and Their World. Bloomington, ind.: indiana University Press, 2007.
Ji, Qiang, et al. “a swimming mammaliaform from the Middle Jurassic and ecomorphological diver-
sification of early mammals.” Science 311 (2006): 1,123–1,127.
Stevens, Kent a., and J. Michael Parrish. “neck posture and feeding habits of two Jurassic sauropod dino-saurs.” Science 284 (1999): 798–800.
xviii+598_Biodiversity_F.indd 342 11/17/11 1:13 PM

343
keystone species Keystone species play an important role in their ecological communities far out of proportion to their abundance or diversity. the name is borrowed from the keystone of an arch which, while just a single stone, maintains the integ-rity of the arch. ecologist Robert Paine introduced the idea in the 20th century. a keystone species performs an important function that creates or alters conditions upon which many other species depend. therefore, keystone species are often irreplaceable in an ecological community, and their disappearance leads to great disruption.
Many of the large mammals that perished in the Pleistocene extinction were keystone species. at the end of the most recent ice age, about 12,000 years ago, many large mammals became extinct, including grazers and browsers such as mammoths and mastodons. Several major events appeared to have happened simultaneously: the large mammal extinctions; changes in climate, which caused for-ests to spread and grasslands to retreat; an increase in the frequency of fi res; and the arrival of humans with advanced hunting techniques such as the clovis style of arrowheads. Some scientists attrib-uted the large mammal extinctions to vegetation changes caused by climate; others attributed them to overhunting by the clovis people. a more careful study of the sequence of events, as recorded in lake sediments, showed that the large mammal popu-lations began to decline a thousand years before the forests began to spread and the clovis people arrived. Something caused the large herbivores to begin dying. these animals had maintained the grasslands. the decline in keystone herbivores caused the vegetation changes, not the other way
around. in fact, even the increase in fi re frequency was an effect rather than a cause. the spread of for-ests, during the mammoth and mastodon declines, caused a buildup of dead wood that promoted fi res. the authors of the study speculated that the decline in large mammals was due to overhunting by earlier peoples, and that the clovis hunting technology was a response to, rather than a cause of, declining abundance of prey.
consider the following examples of large, abun-dant keystone species:
● large grazing animals eat a lot of grass and other plants, thus playing a major role in the food chain and making a major contri-bution to the cycling of nutrients. in fact, they alter their ecosystems. Grasses grow from underground stems and easily grow back after grazing, while many dicot herbs, shrubs, and tree seedlings do not recover as easily. Heavy grazing pressure can there-fore encourage the spread of grasslands at the expense of forests. elephants can func-tion as keystone species, just as did their ancient mammoth and mastodon relatives. One reason that the african savanna does not become acacia woodland is that ele-phants rip and bulldoze the trees to eat their leaves and seed pods. But african elephants function as keystone species in another way as well. they consume the seeds but do not chew all of them. Most acacia seed pods are infested with insect larvae, which destroy most of the seeds—unless, that is, the seed pods are eaten by an elephant. the
K
xviii+598_Biodiversity_F.indd 343 11/17/11 1:13 PM

first entry344 keystone species 344
elephant’s digestive tract destroys the insect larvae. elephant droppings often contain intact acacia seeds. elephants are one of the major sources of new acacia trees in the african savanna.
● top predators often control the populations of many species of prey. One example is the dusky shark, Carcarhinus obscurus. it preys upon other predatory fish, keeping their populations low. When the shark pop-ulations decline, populations of the middle predators in the food chain increase. they eat a lot of scallops, causing the scallop populations to decline. another example is starfish, which are predators upon mussels. Without starfish predators, mussels would dominate some rocky shorelines. Starfish predation opens up some room on the rocks, where barnacles and seaweeds can grow. the disappearance of top predators may cause trophic cascades, in which bursts of overpopulation and decline pass down each link in the food web, from predators to prey to plants.
● Some keystone species actually create habi-tats in which other species live. Beavers build dams, which can convert an entire terres-trial habitat into a wetland. Some ecologists call beavers “ecosystem engineers.” corals are keystone species in the reef communities that are named after them mainly because their calcium carbonate shells accumulate, creating a new and complex substrate upon which seaweeds can grow and starfish can graze.
● Small herbivores can be keystone species (or sets of species) as well. the applica-tion of pesticides that killed most of the arthropods and mollusks in european meadows caused the plant species diversity to decrease.
it is not necessary that other species entirely depend upon the activities of the keystone species. Prairie dogs (Cynomys spp.) are generally considered keystone species in the shortgrass prairies of north america because of their extensive tunnel systems. they aerate and fertilize the soil and create pockets of disturbed soil in which many plant species grow better than in undisturbed grass sod. it turns out that few if any of the animal and plant species that ben-efit from prairie dog activities actually depend upon them; they are all found in nearby areas, though not as abundantly. Despite this, prairie dogs can still be considered a keystone species because they greatly
enhance the abundance of other species. the same is true of the plateau pika (Ochotona curzoniae) in tibet. in both of these cases, humans extensively poison the keystone species in the mistaken belief that they eat too much of the grass upon which live-stock forage.
Keystone species interactions can get very com-plex. ecologists Gretchen Daily and Paul ehrlich describe a keystone species system in the subalpine forests of colorado, where red-naped sapsuckers are keystone species in two different ways. First, they excavate nest cavities in aspens in which the heartwood has decomposed. two other bird species depend upon these nests, after the sapsuckers have abandoned them. the sapsuckers also drill holes in willow trunks, which collect sap that is eaten by many other animal species. the authors con-clude, “Disappearance of any element of the com-plex could cause an unanticipated unraveling of the community.”
in many cases, a group of species, rather than a single keystone species, may control an eco-logical community. a couple of examples are the following:
● Wolves and deer on the Kaibab plateau. the Kaibab Plateau forms the north Rim of the Grand canyon in arizona. it is a mix-ture of grassland, aspen groves, and tem-perate pine forests. Deer prefer aspen over other kinds of forage, therefore aspen is rare during deer population explosions. One of the pioneers of modern ecological science, aldo leopold, had once participated eagerly in government-sponsored predator eradica-tion campaigns, which aimed to increase the deer population for hunters by removing the predators that competed with the hunters for prey. He changed his mind, however, when he noticed that, in the absence of wolves, hunters did not kill enough deer to keep the populations from exploding. He attributed the deer population explosion in the early 20th century directly to wolf eradication campaigns. as it turns out, very few wolves were killed on the Kaibab Plateau, where mountain lions were the principal predators. it therefore appears that a set of predator species, rather than just one, controls deer populations.
● The American chestnut. chestnuts (Casta-nea dentata) were important forest trees in eastern deciduous forests of north america until a fungus (Cryphonectria parasitica, formerly Endothia parasitica) escaped from
xviii+598_Biodiversity_F.indd 344 11/17/11 1:13 PM

keystone species 345
imported european chestnut trees and killed all of the adult american chestnut trees. the chestnut blight spread from new york down to Georgia, killing almost all large appalachian chestnut trees by the 1950s. the forests were temporarily disrupted, but within decades the spaces formerly filled by chestnuts were taken by other trees, mostly oaks. chestnuts were just one of several spe-cies that can play a keystone role in decidu-ous forests.
Sometimes the keystone effect of a species can change, depending on its behavior. alewives (a spe-cies of fish, Alosa pseudoharengus) eat zooplankton in lakes. Heavy predation by alewives results in zooplankton with smaller bodies. in some lakes, the alewives remain all year, while in others they can migrate to the ocean and back. lakes with landlocked alewives had small-bodied zooplankton year-round, and lakes that had no alewives had large-bodied zooplankton year-round. in contrast, zooplankton communities in lakes with migratory alewives cycled between large-bodied zooplankton in the winter and spring and small-bodied zooplank-ton in the summer.
Because of their importance, keystone species are a special focus of preservation and restoration efforts. Keystone species are essential in a restored habitat, because they create the habitat conditions upon which many other species depend.
Keystone species are merely pursuing their own interests. they do nothing for the benefit of the ecological community that they help to create. they may benefit from the results but they evolved because their way of living, which just happened to have important ecological effects, enhanced their evolutionary fitness.
the most significant keystone species in the world is, of course, humans, who have altered the ecosystems of the earth in numerous ways described throughout this encyclopedia. even some of the mildest human activities can have cascading effects on ecosystems. consider the example of yosemite national Park. Millions of people visit it, and they are forbidden to cause any harm to the natural areas, a rule that is largely respected. However, their pres-ence scares away cougars. Mule deer, in contrast, have learned that people like to watch them. Mule deer populations are therefore quite large, and they graze seedlings of Quercus kelloggii oak seedlings as well as wildflowers such as the evening prim-rose Oenothera hookeri, which were once one of yosemite’s most famous beauties. Oaks and evening primroses are becoming scarce in yosemite Valley,
despite control burns intended to encourage their propagation.
See also ecosystems; leopold, aldo; Pleis-tocene extinction.
Further readingBaum, J. K., et al. “collapse and conservation of shark
populations in the northwest atlantic.” Science 299 (2003): 389–392.
Binkley, Dan, et al. “Was aldo leopold right about the Kaibab deer herd?” Ecosystems 9 (2006): 227–241.
Daily, Gretchen c., Paul R. ehrlich, and n. M. Haddad. “Double keystone bird in a keystone species complex.” Proceedings of the National Academy of Sciences USA 90 (1993): 592–594.
Gill, Jacquelyn l., et al. “Pleistocene megafaunal collapse, novel plant communities, and enhanced fire regimes in north america.” Science 326 (2009): 1,100–1,103. Summarized by Johnson, christopher. “Megafaunal decline and fall.” Science 326 (2009): 1,072–1,073.
Kotliar, natasha B., et al. “a critical review of assumptions about the prairie dog as a keystone species.” Environ-mental Management 24 (1999): 177–192.
Menge, Bruce a., et al. “the keystone species concept: Variation in interaction strength in a rocky inter-tidal habitat.” Ecological Monographs 64 (1994): 249–286.
Mills, l. Scott, et al. “the keystone-species concept in ecology and conservation.” BioScience 43 (1993): 219–224.
Mooney, Kailen a., et al. “evolutionary trade-offs in plants mediate the strength of trophic cascades.” Science 327 (2010): 1,642–1,644.
Morell, Virginia. “yosemite: Protected but not preserved.” Science 320 (2008): 597.
Myers, R. a., and B. Worm. “Rapid worldwide depletion of predatory fish communities.” Nature 423 (2003): 280–283.
———, et al. “cascading effects of the loss of apex preda-tory sharks from a coastal ocean.” Science 315 (2007): 1,846–1,850.
Paine, Robert t. “Food web complexity and species diver-sity.” American Naturalist 100 (1966): 65–75.
Post, David M., et al. “intraspecific variation in a predator affects community structure and cascading trophic interactions.” Ecology 89 (2008): 2,019–2,032.
Ripple, William J., and Robert l. Beschta. “Hardwood tree decline following large carnivore loss on the Great Plains, USa.” Frontiers in Ecology and the Environ-ment 5 (2007): 241–246.
Schmitz, Oswald J. “effects of predatory hunting mode on grassland ecosystem function.” Science 319 (2008): 952–954.
Sharam, Gregory J., et al. “Serengeti birds maintain forests by inhibiting seed predators.” Science 325 (2009): 51.
xviii+598_Biodiversity_F.indd 345 11/17/11 1:13 PM

first entry346 keystone species 346
Smith, andrew t., and J. Marc Foggin. “the plateau pika (Ochotona curzoniae) is a keystone species for biodi-versity on the tibetan Plateau.” Animal Conservation 2 (1999): 235–240.
Stein, claudia, et al. “impact of invertebrate herbivory in grasslands depends on plant species diversity.” Ecol-ogy 91 (2010): 1,639–1,650.
terborgh, John, and James estes, eds. Trophic Cascades: Predators, Prey, and the Changing Dynamics of Nature. Washington, D.c.: island Press, 2010.
Watson, David M. “Mistletoe—a keystone resource in forests and woodlands worldwide.” Annual Review of Ecology and Systematics 32 (2001): 219–249.
Willson, Mary F., and Karl c. Halupka. “anadromous fish as keystone species in vertebrate communities.” Con-servation Biology 9 (2002): 489–497.
Wright, Justin P. “linking populations to landscapes: rich-ness scenarios resulting from changes in the dynam-ics of an ecosystem engineer.” Ecology 90 (2009): 3,418–3,429.
xviii+598_Biodiversity_F.indd 346 11/17/11 1:13 PM

347
lakes and ponds See rivers, streams, ponds, and lakes.
leopold, aldo (1886–1948) American Ecolo-gist, conservationist aldo leopold was one of the founders of modern environmental ethics. Born January 11, 1886, in Burlington, iowa, Rand aldo leopold spent much time exploring the natural world. He not only watched birds but counted them, an attitude that prepared him for a scientifi c view of nature. He was trained in the forestry school that Gifford Pinchot had started at yale University.
Beginning in 1909, leopold worked for the U.S. Forest Service in arizona and new Mexico. Much of his responsibility was to shoot predators such as wolves. One of his transformative moments was when he shot a mother wolf with pups. He later interpreted his experience in this way:
We reached the old wolf in time to watch a fi erce green fi re dying in her eyes. i realized then, and have known ever since, that there was something new to me in those eyes—something known only to her and to the mountain. i was young then, and full of trigger-itch; i thought that because fewer wolves meant more deer, that no wolves would mean hunters’ paradise. But after seeing the green fi re die, i sensed that neither the wolf nor the mountain agreed with such a view.
He then began to develop the modern prac-tices of wildlife management, which recognizes the important role of predators in ecological communi-ties. He developed these ideas when he joined the faculty of the University of Wisconsin in 1933.
He also began to develop what he called “the land ethic,” in which he called for the preservation of entire ecological communities as an ethical imper-ative. His ideas were developed in essays, published posthumously as A Sand County Almanac, one of the most famous environmental books. in one essay he wrote, “a thing is right when it tends to preserve the integrity, stability, and beauty of the biotic com-munity. it is wrong when it tends otherwise.” in this way, he diverged from Pinchot’s view of nature as resources to be managed for human use. He also referred to the ecological view of nature as “thinking like a mountain.” leopold died of a heart attack on april 21, 1948, while doing a control burn to man-age his rural property.
See also conservation biology; keystone species.
Further readingaldo leopold Foundation. “the aldo leopold Founda-
tion.” available online. URl: http://www.aldoleopold.org. accessed august 15, 2010.
Knight, Richard l., and Suzanne Riedel, eds. Aldo Leopold and the Ecological Conscience. new york: Oxford University Press, 2002.
leopold, aldo. A Sand County Almanac. new york: Oxford University Press, 2001.
lorbiecki, Marybeth. Aldo Leopold: A Fierce Green Fire.Helena, Mont.: Falcon Press, 1996.
newton, Julianne lutz. Aldo Leopold’s Odyssey. Washing-ton, D.c.: island Press, 2006.
lichens lichens are a mixture of two very dif-ferent kinds of organisms: fungi and single-celled photosynthetic organisms.
L
xviii+598_Biodiversity_F.indd 347 11/17/11 1:13 PM

first entry348 lichens 348
● the photosynthetic cells (phycobionts) are usually green algae of the kingdom Protista; the genus Trebouxia is the phycobiont of about 40 percent of lichen species. cyano-bacteria, usually of the genus Nostoc, may also be associated with fungi in lichens. Only about 8 percent of lichens have cyano-bacterial phycobionts. it is the phycobionts that give lichens their diverse and often bril-liant colors.
● Many different species of fungi (myco-bionts) participate in the lichen symbiosis. about 20 percent of all fungal species (40 percent of the sac fungi) are lichen myco-bionts. almost 98 percent of lichens have sac fungus mycobionts.
each lichen is a mutualistic association, and since they grow and reproduce as single individuals, they can be considered examples of symbiogenesis. Both partners benefit from the association:
● lichens can withstand severe conditions of drought, cold, and heat, because the fun-gus shields the algae from sunlight when conditions are too dry, cold, or hot to allow photosynthesis. For example, some lichens live on rock surfaces and in the nearly bar-ren lands of the far north; reindeer lichens (Cladina spp.) are in fact abundant enough that reindeer graze upon them. the fungus also absorbs and retains water that may be briefly available from rain or fog. Because of the protection provided by the fungus, the algae can live in places where no other algae can survive.
● Fungi require a source of organic molecules as food. the algae provide food to the fungi. therefore these fungi can live in places where no other fungi can survive.
although the fungus strands penetrate and occa-sionally destroy the photosynthetic cells, the photo-synthetic cells reproduce rapidly enough to maintain the symbiosis. the ability to survive on bare rock surfaces is the reason that lichens were among the earliest organisms to live on dry land.
Because the fungi determine the structures of lichens, lichens are classified as fungi. the phy-cobionts, however, are classified independently. Both phycobionts and mycobionts can be cultured independently, but they do not grow independently in nature. each phycobiont species can be associ-ated with different fungus species, and each myco-biont species can be associated with different algal species.
the fungus strands form the structure of the lichen body; the algae are sandwiched between and intermingled with the strands. lichens are most commonly crustose (forming a crust like paint), foliose (leaflike), or fruticose (appearing to have branches) (see figure). like corals that have photo-synthetic cells in their tissues, lichens spread their phycobionts out into a layer that can absorb the maximum amount of sunlight.
lichens usually disperse to new locations either by fragmenting into smaller pieces when they are dry or by forming and releasing small structures in which phycobiont cells are wrapped in mycobiont strands. in some cases, the mycobionts produce spores as if they were free-living fungi. the spores germinate into a new fungus in a new location, where they must find a suitable phycobiont if they are to survive.
lichens are dependent upon the absorption of nutrients from dust and rainwater. as a result they absorb a great deal of air pollution from both sources. they also have no deciduous parts and cannot shed tissue that may be damaged by pollution. lichens are
Lichens, such as these from the Black Hills of South Dakota, have single-celled “algae” inside a fungus structure. The resulting structure is a mutualistic association between two species. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 348 11/17/11 1:13 PM

life history, diversity of 349
therefore sensitive to air pollution, some more than others. lichens are frequently studied as biological monitors of air pollution in natural habitats.
See also bacteria; fungi; mutualism; protists.
Further readingesseen, Per-anders, and Karl-erik Renhorn. “edge effects
on an epiphytic lichen in a fragmented forest.” Conser-vation Biology 12 (2008): 1,307–1,317.
Gargas, a., et al. “Multiple origins of lichen symbioses in fungi suggested by SSU rDna phylogeny.” Science 268 (1995): 1,492–1,495.
lesica, Peter, et al. “Differences in lichen and bryophyte communities between old-growth and managed sec-ond-growth forests in the Swan Valley, Montana.” Canadian Journal of Botany 69 (1991): 1,745–1,755.
lutzoni, François, et al. “Major fungal lineages are derived from lichen symbiotic ancestors.” Nature 411 (2001): 937–940.
nash, t. H., ed. Lichen Biology. new york: cambridge University Press, 1995.
Pettersson, Roger B., et al. “invertebrate communities in boreal forest canopies as influenced by forestry and lichens with implications for passerine birds.” Biologi-cal Conservation 74 (1995): 57–63.
Rose, c. i., and D. l. Hawksworth. “lichen recolonization in london’s cleaner air.” Nature 289 (1981): 289–292.
Sancho, l. G., et al. “lichens survive in space: Results from the 2005 licHenS experiment.” Astrobiology 7 (2007): 443–454.
Sillett, Stephen c., et al. “Dispersal limitations of epiphytic lichens result in species dependent on old-growth for-ests.” Ecological Applications 10 (2000): 789–799.
yuan, X., et al. “lichen-like symbiosis 600 million years ago.” Science 308 (2005): 1,017–1,020.
life history, diversity of life history refers to the patterns of growth, reproduction, and death in organisms. Differences in life history are among the most important differences among species. certain sets of life history characteristics, called strategies, function successfully together; however, seldom does an environment or habitat have a single best life history strategy. Below are examples, rather than a complete overview.
phySIoloGy aNd GrowThOrganisms have limits. in particular, they have lim-ited resources and must allocate those resources among functions. Frequently, to do more of one thing means to do less of another. Organisms accordingly invest resources into those functions that will allow them to obtain more resources and to reproduce more successfully.
consider this simplified example of plant growth. the plant uses its food resources to produce leaves; this is an investment, because the leaves then allow the plant to make more food. However, it must also produce stems, to supply the leaves with water and to hold the leaves up higher than the leaves of other plants, and roots, which obtain water and minerals for the leaves. if the plant allocates most of the food that the leaves make into the production of new leaves, neglecting its stems (for example, by making them flimsy) and producing few new roots, it may be able to grow very fast but will probably be unable to survive the winter. this is part of the life history strategy of a weed; it grows fast because it invests all its resources in growth rather than main-tenance. if the plant allocates more of the food made by the leaves into the production of wood and peren-nial roots, it will grow more slowly but will be able to maintain a system of trunk, stems, and roots for many years. this is part of the life history strategy of a tree. tree species differ from one another in their life history patterns. trees that grow quickly after an ecological disturbance (early successional trees) produce cheap wood and do not live very long; trees that grow slowly and dominate old forests (late suc-cessional trees) produce expensive wood and live a long time. the wood of early successional trees con-tains more of the large vessels (pores) that transport water and allows rapid growth than does the wood of later successional trees (see figure on page 350). an animal example of different life histories would be to compare a butterfly and a turtle. the butterfly has wings that cannot repair themselves, but this does not matter, as the butterfly dies very quickly. the turtle, however, has a body built to last.
consider the interactions of organisms with other species. if the plant produces lots of defensive chemicals in its leaves, it cannot grow as fast, since it has expended many of its resources on defense rather than growth. Often, the plants that live longer produce leaves that are more toxic, while short-lived plants have less toxic leaves. if insects attack, the plant with the defenses will not suffer as great a loss of leaf material; however, the faster-growing plant may rapidly grow back after the insects attack it. the plants and animals have coevolved in response to one another.
rEproduCTIoNinvestment and allocation patterns can allow organ-isms to grow large and strong, but if they do not reproduce, natural selection eliminates their adap-tations. the timing, amount, and pattern of repro-duction are important parts of life history; in fact, they make the growth and physiology worthwhile from an evolutionary viewpoint. Reproductive
xviii+598_Biodiversity_F.indd 349 11/17/11 1:13 PM
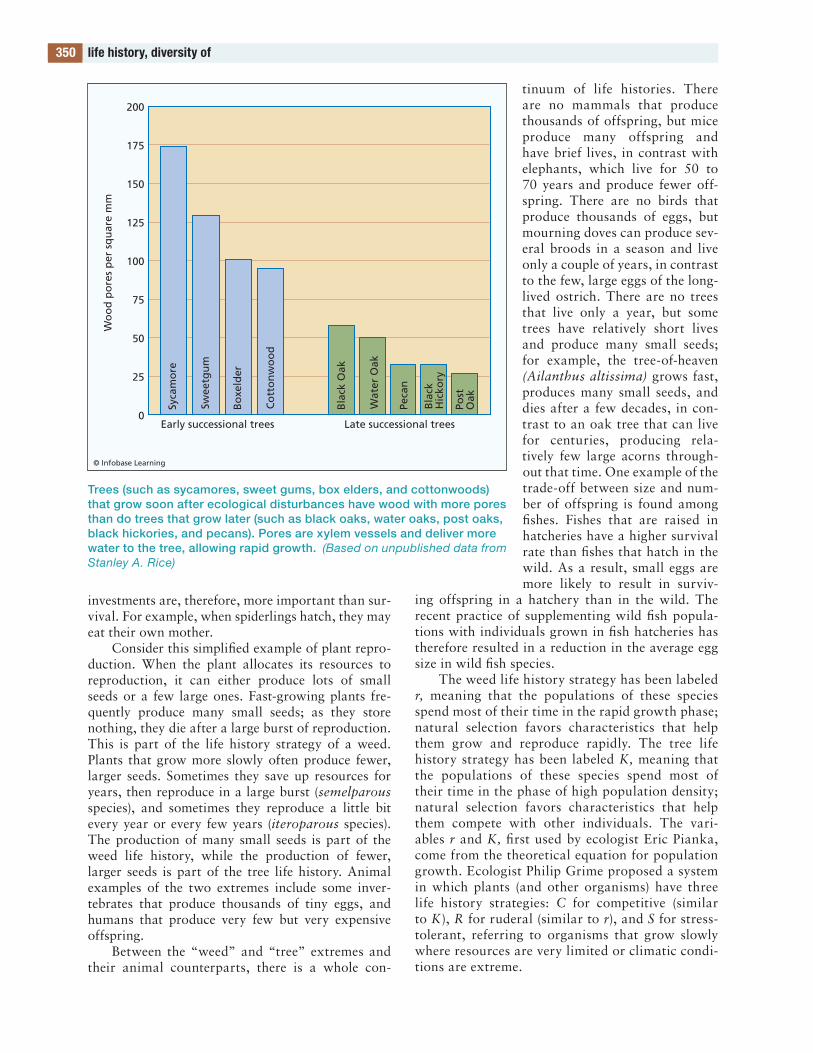
first entry350 life history, diversity of 350
investments are, therefore, more important than sur-vival. For example, when spiderlings hatch, they may eat their own mother.
consider this simplified example of plant repro-duction. When the plant allocates its resources to reproduction, it can either produce lots of small seeds or a few large ones. Fast-growing plants fre-quently produce many small seeds; as they store nothing, they die after a large burst of reproduction. this is part of the life history strategy of a weed. Plants that grow more slowly often produce fewer, larger seeds. Sometimes they save up resources for years, then reproduce in a large burst (semelparous species), and sometimes they reproduce a little bit every year or every few years (iteroparous species). the production of many small seeds is part of the weed life history, while the production of fewer, larger seeds is part of the tree life history. animal examples of the two extremes include some inver-tebrates that produce thousands of tiny eggs, and humans that produce very few but very expensive offspring.
Between the “weed” and “tree” extremes and their animal counterparts, there is a whole con-
tinuum of life histories. there are no mammals that produce thousands of offspring, but mice produce many offspring and have brief lives, in contrast with elephants, which live for 50 to 70 years and produce fewer off-spring. there are no birds that produce thousands of eggs, but mourning doves can produce sev-eral broods in a season and live only a couple of years, in contrast to the few, large eggs of the long-lived ostrich. there are no trees that live only a year, but some trees have relatively short lives and produce many small seeds; for example, the tree-of-heaven (Ailanthus altissima) grows fast, produces many small seeds, and dies after a few decades, in con-trast to an oak tree that can live for centuries, producing rela-tively few large acorns through-out that time. One example of the trade-off between size and num-ber of offspring is found among fishes. Fishes that are raised in hatcheries have a higher survival rate than fishes that hatch in the wild. as a result, small eggs are more likely to result in surviv-
ing offspring in a hatchery than in the wild. the recent practice of supplementing wild fish popula-tions with individuals grown in fish hatcheries has therefore resulted in a reduction in the average egg size in wild fish species.
the weed life history strategy has been labeled r, meaning that the populations of these species spend most of their time in the rapid growth phase; natural selection favors characteristics that help them grow and reproduce rapidly. the tree life history strategy has been labeled K, meaning that the populations of these species spend most of their time in the phase of high population density; natural selection favors characteristics that help them compete with other individuals. the vari-ables r and K, first used by ecologist eric Pianka, come from the theoretical equation for population growth. ecologist Philip Grime proposed a system in which plants (and other organisms) have three life history strategies: C for competitive (similar to K), R for ruderal (similar to r), and S for stress- tolerant, referring to organisms that grow slowly where resources are very limited or climatic condi-tions are extreme.
Trees (such as sycamores, sweet gums, box elders, and cottonwoods) that grow soon after ecological disturbances have wood with more pores than do trees that grow later (such as black oaks, water oaks, post oaks, black hickories, and pecans). Pores are xylem vessels and deliver more water to the tree, allowing rapid growth. (Based on unpublished data from Stanley A. Rice)
xviii+598_Biodiversity_F.indd 350 11/17/11 1:13 PM

life history, diversity of 351
no single life history strategy is always better than another. the weed strategy and the tree strat-egy both work but in different ways. Both minnows and sturgeons and both mice and elephants success-fully make their living in the world. although no one life history is superior over others, certain environ-ments may favor one strategy over another.
● the tundra has few weeds. the growing season is so brief and chilly that a plant could not grow rapidly enough to reproduce successfully after only one year. if the plant must survive the winter, in order to have two or more growing seasons, it must have adaptations such as strong stems or under-ground storage organs, which force it to have slower growth.
● Deserts may have many plant species with a weedy life history. the brief wet spring-time of a desert is warm enough that many plants can, in fact, complete their growth and reproduction within a few weeks. these plants may be extremely small. this is not the only successful strategy for a des-ert plant. cactuses store water, and desert bushes such as mesquite and creosote grow very deep roots that allow them to survive the dry season so that they can grow for many years.
● in areas such as a forest disturbed by fires or storms, many weeds grow. their small seeds germinate into small seedlings, but there is little competition from other plants (most of which were killed by the fire or storm). the seedlings grow rapidly and reproduce soon; they complete their lives before the forest grows back. Once the forest has grown back, however, weedy species are at a dis-advantage, since their tiny seeds could not grow well in the shade of the trees; now the K or C plants have the advantage.
Just as the human economy has evolved a great diversity of different ways of making a living, so the “economy of nature” has produced communities of plants and animals that make their living by means of many different life history strategies.
charles Darwin, following the lead of economist thomas Malthus, noted that all organisms have the capacity to reproduce far in excess of the space and resources available. Darwin calculated that even a pair of elephants, if they bred as much as possible and all the offspring survived, could produce 19 million individuals in 750 years. the elephant repre-sents the K end of the life history spectrum. Species at the r end of the spectrum have almost ridicu-
lously large potential reproductive outputs, because the vast majority of their offspring die. if resources were unlimited and all offspring survived, a pair of houseflies could produce a population of 200 quin-tillion in five months and a pair of starfish could produce a population in 16 years as great as the number of electrons in the visible universe. a single bacterium, dividing every few minutes, could cover the earth two meters deep in bacterial slime in less than two days. these organisms use their prodigious reproductive capacity only during brief times when resources are abundant.
Organisms may increase or decrease their repro-ductive investments, depending on the immediate circumstances. During stressful years with inade-quate resources, plants may produce fewer seeds, and animals smaller litters than during years of abundance. adjustments can be made even after the next generation has been initiated. Under conditions of inadequate resources or pollination, plants may abort seeds or fruits. Spontaneous abortions are common in animals. in golden hamsters, a mother may even eat some of her own offspring if resources become inadequate.
the life history of Homo sapiens and its imme-diate evolutionary ancestors is a particularly striking example of the K strategy. One of the major features of the evolution of the hominin lineage has been the prodigious increase in brain size; both relatively and absolutely, the modern human brain is outstand-ing in the animal kingdom. the first hominins to experience the evolutionary explosion in brain size were the early members of the genus Homo. Once large brains and their associated intelligence had evolved, they provided an advantage that justified their tremendous cost. Much of this brain growth occurs during fetal development and continues well past childhood.
a human fetus, because of its large head, is born only with great difficulty. the passage of the head through the birth canal is the most prolonged, painful, and dangerous part of the birth process in humans. the birth canal cannot evolve to be much larger than it is, for otherwise a woman would not be able to walk. therefore natural selection favored an increase in brain size but also put constraints upon it. the evolutionary solution to this dilemma was for the fetus to be born early, while the head was still small. in chimpanzees, the ratio of the gesta-tion period (in weeks) to the total lifespan (in years) is about 0.8. if the human gestation period were the same length of time, relative to the life span, as it is in chimp primates, the human gestation period would be about 15 months and birth would be impossible (see table on page 352). By being born after nine months’ gestation, the baby’s head is small enough to
xviii+598_Biodiversity_F.indd 351 11/17/11 1:13 PM
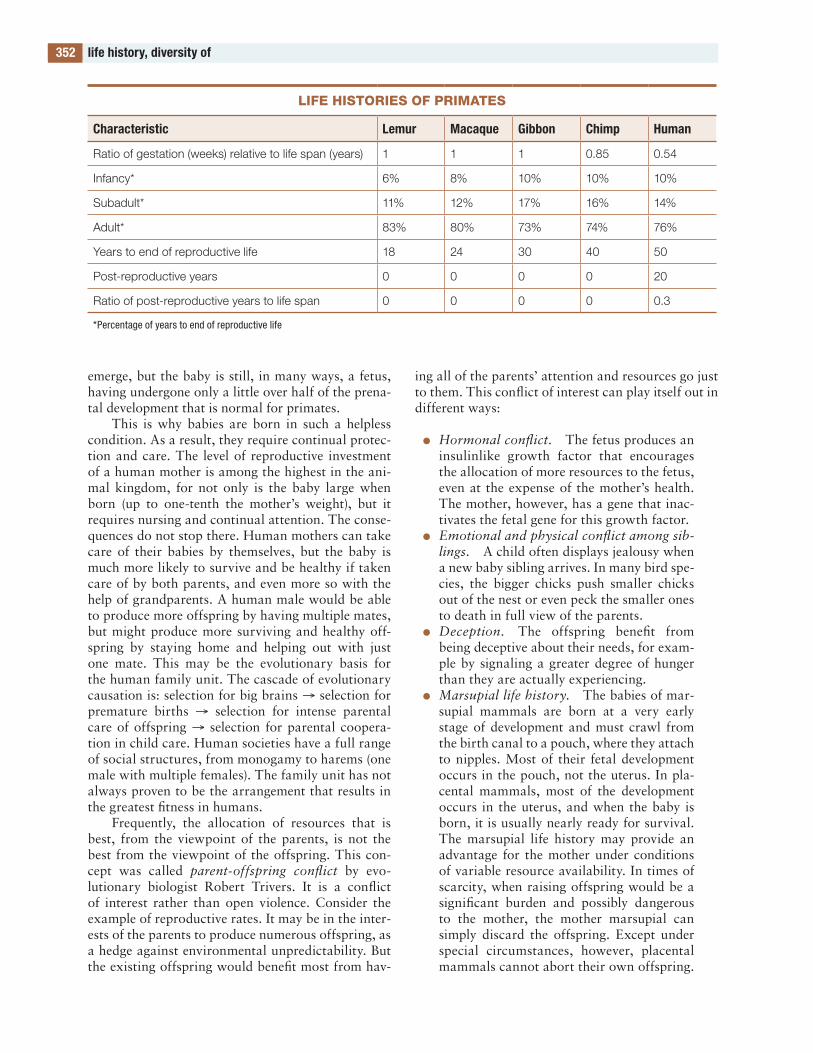
first entry352 life history, diversity of 352
emerge, but the baby is still, in many ways, a fetus, having undergone only a little over half of the prena-tal development that is normal for primates.
this is why babies are born in such a helpless condition. as a result, they require continual protec-tion and care. the level of reproductive investment of a human mother is among the highest in the ani-mal kingdom, for not only is the baby large when born (up to one-tenth the mother’s weight), but it requires nursing and continual attention. the conse-quences do not stop there. Human mothers can take care of their babies by themselves, but the baby is much more likely to survive and be healthy if taken care of by both parents, and even more so with the help of grandparents. a human male would be able to produce more offspring by having multiple mates, but might produce more surviving and healthy off-spring by staying home and helping out with just one mate. this may be the evolutionary basis for the human family unit. the cascade of evolutionary causation is: selection for big brains → selection for premature births → selection for intense parental care of offspring → selection for parental coopera-tion in child care. Human societies have a full range of social structures, from monogamy to harems (one male with multiple females). the family unit has not always proven to be the arrangement that results in the greatest fitness in humans.
Frequently, the allocation of resources that is best, from the viewpoint of the parents, is not the best from the viewpoint of the offspring. this con-cept was called parent-offspring conflict by evo-lutionary biologist Robert trivers. it is a conflict of interest rather than open violence. consider the example of reproductive rates. it may be in the inter-ests of the parents to produce numerous offspring, as a hedge against environmental unpredictability. But the existing offspring would benefit most from hav-
ing all of the parents’ attention and resources go just to them. this conflict of interest can play itself out in different ways:
● Hormonal conflict. the fetus produces an insulinlike growth factor that encourages the allocation of more resources to the fetus, even at the expense of the mother’s health. the mother, however, has a gene that inac-tivates the fetal gene for this growth factor.
● Emotional and physical conflict among sib-lings. a child often displays jealousy when a new baby sibling arrives. in many bird spe-cies, the bigger chicks push smaller chicks out of the nest or even peck the smaller ones to death in full view of the parents.
● Deception. the offspring benefit from being deceptive about their needs, for exam-ple by signaling a greater degree of hunger than they are actually experiencing.
● Marsupial life history. the babies of mar-supial mammals are born at a very early stage of development and must crawl from the birth canal to a pouch, where they attach to nipples. Most of their fetal development occurs in the pouch, not the uterus. in pla-cental mammals, most of the development occurs in the uterus, and when the baby is born, it is usually nearly ready for survival. the marsupial life history may provide an advantage for the mother under conditions of variable resource availability. in times of scarcity, when raising offspring would be a significant burden and possibly dangerous to the mother, the mother marsupial can simply discard the offspring. except under special circumstances, however, placental mammals cannot abort their own offspring.
life histories of primates
Characteristic lemur macaque Gibbon Chimp human
Ratio of gestation (weeks) relative to life span (years) 1 1 1 0.85 0.54
Infancy* 6% 8% 10% 10% 10%
Subadult* 11% 12% 17% 16% 14%
Adult* 83% 80% 73% 74% 76%
Years to end of reproductive life 18 24 30 40 50
Post-reproductive years 0 0 0 0 20
Ratio of post-reproductive years to life span 0 0 0 0 0.3
*Percentage of years to end of reproductive life
xviii+598_Biodiversity_F.indd 352 11/17/11 1:13 PM

life history, diversity of 353
From the viewpoint of the offspring, the marsupial life history is never an advantage; a womb is always safer than a pouch.
Most animals are either males or females. in many fishes, however, an individual animal can change from one gender to another. this makes the life histories of these fishes very unusual. Sex-ual selection favors large males; therefore a small, young male would be at a disadvantage in com-petition with the few, large, dominant males. in these fishes, there are many small and medium-sized females. Some females, when they become large, change into males. these individuals are maximiz-ing their reproductive output by being female when they are young and small and male when they are older and larger.
dEaThevolution cannot produce immortal organisms. every organism runs the risk of accidents leading to injury or death. Furthermore, mutations accumulate in the chromosomes of organisms. Radiation in the environment and even the very oxygen in the air induce mutations. the longer an organism lives, the greater risk it runs of death.
it is quite clear, however, that cells and organ-isms ought to be able to live much longer than they actually do:
● animal populations contain genetic vari-ability that would allow an increased life span. Researchers have found the indy (“i’m not dead yet”) gene in the fruit fly Drosoph-ila melanogaster and a similar gene in the roundworm Caenorhabditis elegans. this gene extends life span by reducing metabolic rate. Fruit flies have been experimentally bred that have life spans twice as long as those of wild fruit flies.
● Germ (germline) cells in animals and most plant cells are potentially immortal. early in embryonic development, animal cells dif-ferentiate into somatic cells, which become nonreproductive tissues and organs and which eventually die, and germ cells, which are passed on to the next generation in the form of eggs, sperm, or spores. (these are not to be confused with pathogens such as bacteria, which are popularly called germs.) among the somatic cells, stem cells may also be potentially immortal. White blood cells, the basis of the immune system, have a much greater capacity to reproduce than most other cells of vertebrates. cancerous white blood cells (as in leukemia) may have
no physiological constraints on life span. a leukemia patient, Henrietta lacks, died in 1951 but some of her “Hela” white blood cells are still alive in medical laboratories all over the world.
the inescapable conclusion is that natural selec-tion has actually favored organisms that die. the reason seems to be that reproduction itself poses a survival cost on the parent. in order to reproduce more, organisms must take metabolic and behav-ioral risks.
● Metabolic risks. a higher metabolic rate may allow greater reproductive output. animal species with faster metabolism do not live as long as species with slower metabolism. experimental breeding of fruit flies has shown that the longer-lived strains have lower metabolism and have lower reproductive output. Food restric-tion can cause an individual, whether a fly or a human, to live longer. an exception is that most bats have rapid metabolism and long life spans.
● Behavioral risks. an animal that takes risks may be injured but may reproduce more than a cautious animal.
the expense of producing a long-lived body may so greatly impair reproduction that natural selection will favor shorter-lived organisms, if this allows them a greater total reproductive output. For animals, the continual renewal of components that are damaged, such as the beak of a woodpecker, would be enormously expensive—and that expense would come at the cost of reproductive fitness. Since very old individuals would be rare even in a popula-tion of biologically immortal animals (most would already have died from accidents), natural selection has not favored the evolution of adaptations that prolong the lives of very old individuals. the rep-tiles with some of the longest life spans are turtles, and the mammals with some of the longest life spans (aside from humans) are porcupines, both of which are relatively sluggish and well protected from injury. therefore natural selection will not eliminate mutations that bring harm only to older individuals. One reason is that, in most popula-tions, few individuals live to be old. another reason is that even those individuals that do live to be old will have very little reproductive life left ahead of them. Scientists such as biologist Peter Medawar have pointed out that pleiotropic genes, which have more than one effect, may be responsible for aging (senescence). Such genes may enhance fitness in
xviii+598_Biodiversity_F.indd 353 11/17/11 1:13 PM

first entry354 life history, diversity of 354
younger individuals even if they also reduce the sur-vival of older individuals.
those few organisms that do live for a very long time (all of them trees) either possess very expensive adaptations or else grow slowly. an example of the former is the giant sequoia (Sequoiadendron gigan-teum), many of which are more than 2,000 years old and which produce very thick fire-retardant bark. an example of the latter is the bristlecone pine (Pinus longeava) in california. Many of these pines are more than 4,500 years old but have grown less than 30 feet (10 m) tall during that time, because of the harshly cold and dry environment in which they live. Many of the world’s oldest and largest trees, such as the ancient lime (linden) trees of england and Germany, are not actually the original trees but are the outgrowth of branches or roots that the orig-inal tree produced, from which the original trunks have been lost.
Virginia opossums (Didelphus virginiana) on the mainland of the southeastern United States have evolved in the presence of predators, but on Sapelo island there are no predators. the island opossums have evolved delayed old-age senescence (as mea-sured by the breaking time of their collagen fibers). they also reproduce relatively more in their second year of life and less in their first year, compared to mainland opossums. this pattern suggests that under more dangerous conditions, in this case when predators are present, natural selection favors earlier reproduction and favors (or at least does not disfa-vor) more rapid aging.
Humans are unusual not only in the enormous amount of care they give to offspring but also in the enormously prolonged post-reproductive life span. in most animals, once reproduction is finished, the individuals die. a comparison among different pri-mates shows that chimps and humans have slightly longer infancy and juvenile periods, relative to the life span up to the end of reproduction, than do other primates. But the most striking difference is the human post-reproductive period, which adds another one-third to the life span (see table on page 352). in humans, the post-reproductive life span can be almost as long as the juvenile and reproductive life spans combined. the human with the longest life span verified by a birth certificate was Jeanne calment, who died in arles, France, at the age of 122 in 1997.
the explanation usually given for this is that the older individuals possess a wealth of knowledge that can prove immensely valuable to individual families and to the tribe as a whole. this body of knowledge can be so large that it takes a lifetime to teach. a bird can learn its repertory of songs and how to forage in a single season, but human
knowledge fills a lifetime longer than the reproduc-tive life span. Since the invention of writing, this knowledge can be passed from one generation to another impersonally, but through most of human evolutionary history old people, not books or data-bases, were the repository of tribal knowledge, especially about the uses of the many wild species. Science writer natalie angier calls grandmothers the “alexandrian libraries for preliterate tribes.” this cultural knowledge would directly benefit the wise old person’s descendants.
evolutionary scientists have had a particularly difficult time explaining the origin of menopause. Many evolutionary scientists consider menopause to be a nonadaptive side effect of the evolution of human life history. Menopause evolved acciden-tally, and since very few old women survived in prehistoric societies, there was no selection against menopause. in contrast, some evolutionary biolo-gists, such as Kristin Hawkes, say that menopause is adaptive. Dubbed the “grandmother hypothesis,” this theory says that a woman may have greater inclusive fitness by devoting herself to the nurture of her grandchildren than by continuing to have her own children, once she is old. this may explain why older women seldom reproduce, but how could natural selection have favored the sudden enforced cessation of reproduction? the reason may be that continued reproduction, even continued reproduc-tive ability, in older women may present physiologi-cal risks:
● Breast cancer. there is a correlation between the number of menstrual cycles and the risk of breast cancer, since the hormones that the woman’s body produces after ovu-lation (estrogen and progesterone) stimulate breast cell division. continuation of men-strual cycles into old age could therefore represent a risk. Some evolutionary scien-tists conclude from this that menopause reduced breast cancer incidence in older women. Other evolutionary scientists point out that women have far fewer menstrual cycles in primitive societies than in civilized societies, because they were usually preg-nant or lactating. Breast cancer incidence is much lower in societies with high reproduc-tive rates. in early human societies, even old women would not have had enough men-strual cycles to create a risk of breast cancer.
● Difficulty during childbirth. an older woman would be at greater risk of difficulty during childbirth and would be less likely to live long enough to nurture her own child to maturity.
xviii+598_Biodiversity_F.indd 354 11/17/11 1:13 PM

linnaeus, Carolus 355
Older men experience reduced sperm production but no discrete point in their lives at which sexual reproduction ceases. this may reflect the fact that sperm production is less risky than childbirth.
natural selection has favored, under some con-ditions, characteristics (including a brief life span) that go with the r life history strategy. But natural selection has also favored limited life spans for spe-cies with the K life history strategy. the short lives of weeds and mayflies, therefore, just like the long but finite life of a human, are not evolutionary short-comings but are products of natural selection.
ecological communities nearly always consist of species with very different life histories, each one of them specializing in a different way of making a living: living rapidly in briefly available resources or surviving for a long time through times when resources are scarce.
See also coevolution; natural selection; sexual selection; succession, ecological.
Further readingcharlesworth, Brian. “Fisher, Medawar, Hamilton and the
evolution of aging.” Genetics 156 (2000): 927–931.clutton-Brock, t. H. The Evolution of Parental Care.
Princeton, n.J.: Princeton University Press, 1991.De Magalhães, João Pedro. “Why are we allowed to age?”
available online. URl: http://senescence.info/evolu-tion.html. accessed april 20, 2011.
elgar, Mark a. “evolutionary compromise between a few large and many small eggs: comparative evidence in teleost fish.” Oikos 59 (1990): 283–287.
ellison, P. t. On Fertile Ground: A Natural History of Human Reproduction. cambridge, Mass.: Harvard University Press, 2003.
Hawkes, Kristin. “Grandmothering, menopause, and the evolution of human life histories.” Proceedings of the National Academy of Sciences USA 95 (1998): 1–4.
Hrdy, Sarah Blaffer. Mother Nature: Maternal Instincts and How They Shape the Human Species. new york: Bal-lantine, 2000.
Kirkwood, thomas B. l., and S. n. austad. “Why do we age?” Nature 408 (2000): 233–238.
Medawar, Peter. An Unsolved Problem in Biology. london: H. K. lewis, 1952.
trivers, Robert l. “Parent-offspring conflict.” American Zoologist 14 (1974): 249–264.
Westendorp, R., and thomas B. l. Kirkwood. “Human longevity at the cost of reproductive success.” Nature 396 (1998): 743–746.
Williams, George c. “Pleiotropy, natural selection and the evolution of senescence.” Evolution 11 (1957): 398–411.
———. Sex and Evolution. Princeton, n.J.: Princeton Uni-versity Press, 1975.
linnaeus, Carolus (1707–1778) Swedish Botanist carolus linnaeus was the Swedish bota-nist who invented modern taxonomic classification of biodiversity using latin names. He was born on May 23, 1707, as carl linnaeus, and his family name was originally linné; both of his names are lati-nized versions. the linnaean system brought orga-nization into the exuberant world of biodiversity.
like many other important intellectuals in world history, linnaeus showed little promise as a stu-dent. His father apprenticed him to a cobbler, which inspired carl to go back to school and improve his grades. He studied medicine, but his passion, start-ing in his twenties, was to catalogue all the known species of organisms.
linnaeus was passionate about two other things as well. First, he was passionate about his own greatness. He wanted Princeps Botanicorum (prince of botanists) on his tombstone and consid-ered his method of classification to be “the greatest achievement in the realm of science.” Second, he was passionate about sex. He used a sexual system, based upon anthers and pistils, to classify plants. But he went further. His terms for the anatomy of mollusks included vulva, labia, pubes, anus, hymen, even though the structures he described had no rela-tion to the corresponding parts of human anatomy. among the names he gave plants were Clitoria, For-nicata, and Vulva. His descriptions of plant repro-duction were very florid. He described the parts of the flower, correctly, as sexual organs, but went fur-ther: “the flowers’ leaves serve as a bridal bed . . . the bridegroom with his bride might there celebrate their nuptials . . . then it is time for the bridegroom to embrace his beloved bride and surrender himself to her.” Predictably, some scholars disliked lin-naeus’s sexual classification system. a professor in St. Petersburg, Johann Siegesbeck, wrote that God would not have created the natural world based on this “shameful whoredom.” linnaeus’s response was to name a small, ugly weed Siegesbeckia (now Sigesbeckia) in his honor. in his personal life, lin-naeus was very conservative, despite his florid writings.
the order that linnaeus brought into biology by this system, however, inspired nearly all scientists. in particular, a number of young men were his disciples and explored all over the world at great personal risk to find new species of plants to classify with the linnaean system. Peter Kalm explored canada dur-ing the height of the tensions between France and england; Frederik Hasselquist explored egypt; Peter Osbeck went to china; Pehr löfling went to Spain; and lars Montin went to lapland, all spending their own fortunes to collect plants and send them back to linnaeus. though not a disciple of linnaeus,
xviii+598_Biodiversity_F.indd 355 11/17/11 1:13 PM

first entry356 living fossils 356
botanist John Bartram went into the wilderness of the american southeast looking for plants and was one of the first white visitors to the cherokee tribe.
another reason that linnaeus’s system caused a flurry of botanical exploration is that his sys-tem allowed naturalists to name plants after them-selves or other naturalists whom they admired. the genus name of a small herbaceous plant of the north woods, Linnaea borealis, honors linnaeus, and the species name of Abutilon theophrasti, the velvetleaf weed, honors the Greek philosopher theophrastus. Both amateur naturalists and professional scientists yearned for the immortality that would result.
even though he did not even think about evolu-tion, linnaeus included in his writings some inter-mediate forms based on hearsay. He was willing to believe them because he accepted the concept of the Great chain of Being (scala naturae), in which God had not only created every type of creature but all of the intermediates between them. For example, he wrote about strange species of humans such as Homo ferus, a feral human on all fours and unable to talk, and Homo caudatus, man with a tail.
linnaeus has gained scientific immortality, even though his classification system has become unwieldy and may possibly be superseded by an evo-lutionary (phylogenetic) system. linnaeus died on January 10, 1778.
See also biological classification; cladistics.
Further readingconniff, Richard. “Happy birthday, linnaeus.” Natural
History, December 2006, 42–47.Gould, Stephen Jay. “Ordering nature by budding and
full-breasted sexuality.” chap. 33 of Dinosaur in a Haystack: Reflections in Natural History. new york: Harmony, 1995.
Schiebinger, londa. “the loves of the plants.” Scientific American, February 1996, 110–115.
———. “the Private lives of Plants: Sexual Politics in carl linnaeus and erasmus Darwin.” in Science and Sensibility: Gender and Scientific Enquiry 1780–1945, edited by M. Benjamin, 121–143. new york: Oxford University Press, 1991.
living fossils Living fossils is the popular term given to modern multicellular organisms that closely resemble forms that have been known from the fos-sil record for at least several million years (see table on page 357). Most of them, such as the ginkgoes and dawn redwoods in china, persist in habitats that have not changed very much for millions of years. Most are members of evolutionary lineages that have not rapidly diversified; for example, there
are few living fossils among the flowering plants, tele-ost bony fishes, or tetrapod vertebrates. Horseshoe crabs have changed little for millions of years (see fig-ure), and there are few species of them today. Within those groups, the living fossils resemble some of the most ancient species. Amborella trichopoda, which is often considered a living fossil flowering plant, is also the modern species that most closely resembles the ancestor of all flowering plants. the lake sturgeon, often considered a living fossil, is one of the most primitive of the ray-finned fishes, and tuataras repre-sent one of the most ancient reptile lineages.
there is no precise definition of a living fossil. cockroaches are often called living fossils because representatives of the order Dictyoptera can be found in deposits as old as the carboniferous period. However, it appears that, although the cockroach genus Cryptocercus appears to be very ancient, most modern cockroach species are not living fossils. it is often impossible to determine whether a species rep-resented in the fossil record is the same as a modern species, no matter how closely it resembles it.
an online search for living fossils will connect to dozens of creationist Web sites. they claim, based
Modern horseshoe crabs (bottom) closely resemble the fossils of their ancient ancestors (top). (David Goehring [top] and U.S. Geological Survey [bottom])
xviii+598_Biodiversity_F.indd 356 11/17/11 1:13 PM
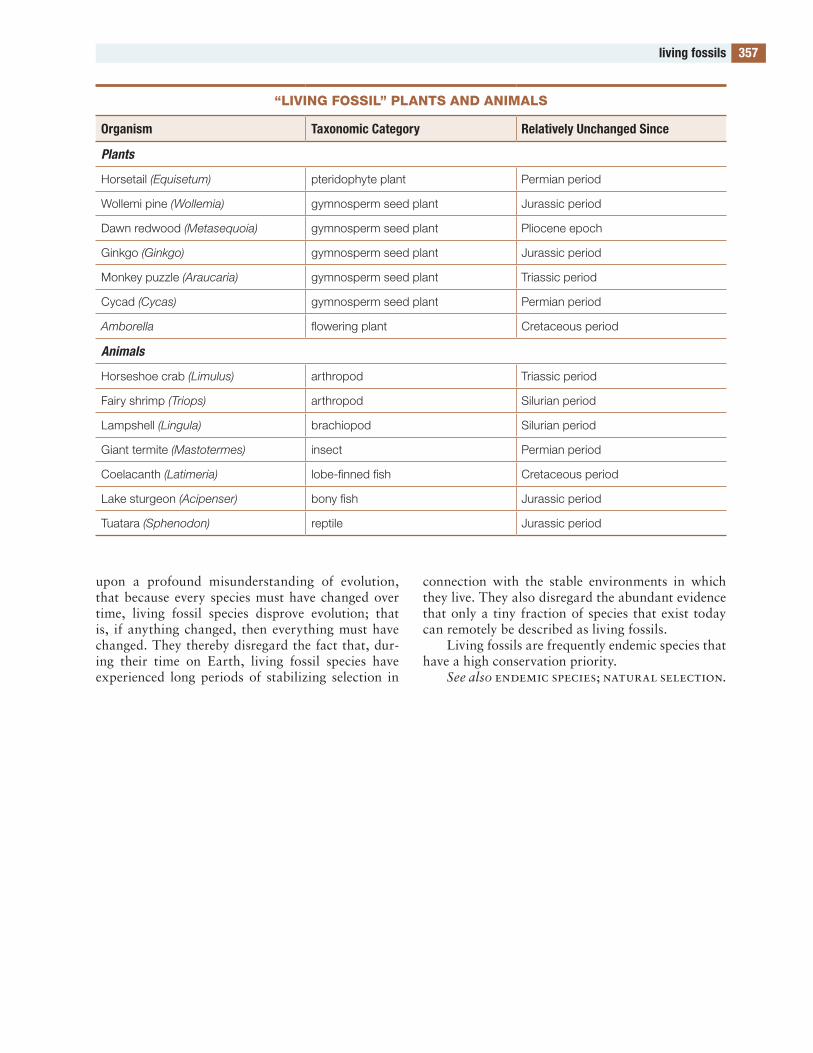
living fossils 357
upon a profound misunderstanding of evolution, that because every species must have changed over time, living fossil species disprove evolution; that is, if anything changed, then everything must have changed. they thereby disregard the fact that, dur-ing their time on earth, living fossil species have experienced long periods of stabilizing selection in
connection with the stable environments in which they live. they also disregard the abundant evidence that only a tiny fraction of species that exist today can remotely be described as living fossils.
living fossils are frequently endemic species that have a high conservation priority.
See also endemic species; natural selection.
“living fossil” plants and animals
organism Taxonomic Category relatively unchanged Since
Plants
Horsetail (Equisetum) pteridophyte plant Permian period
Wollemi pine (Wollemia) gymnosperm seed plant Jurassic period
Dawn redwood (Metasequoia) gymnosperm seed plant Pliocene epoch
Ginkgo (Ginkgo) gymnosperm seed plant Jurassic period
Monkey puzzle (Araucaria) gymnosperm seed plant Triassic period
Cycad (Cycas) gymnosperm seed plant Permian period
Amborella flowering plant Cretaceous period
Animals
Horseshoe crab (Limulus) arthropod Triassic period
Fairy shrimp (Triops) arthropod Silurian period
Lampshell (Lingula) brachiopod Silurian period
Giant termite (Mastotermes) insect Permian period
Coelacanth (Latimeria) lobe-finned fish Cretaceous period
Lake sturgeon (Acipenser) bony fish Jurassic period
Tuatara (Sphenodon) reptile Jurassic period
xviii+598_Biodiversity_F.indd 357 11/17/11 1:13 PM

358
maathai, Wangari (1940–2011) Kenyan Con-servationist Wangari Muta Maathai (see photo-graph on page 359) founded the Green Belt Movement in Kenya. this movement enlisted thousands of Kenyan women in an effort to plant trees and to campaign for the rights of poor women. the quality of life in rural Kenyan villages depends strongly upon the availability of forests, particularly for fi rewood. Maathai’s work, for which she received the nobel Peace Prize in 2004, showed that improving the lives of the poor is inseparable from conservation and rec-lamation. She was the fi rst african woman, and the fi rst environmentalist, to win the nobel Peace Prize.
Wangari Muta was born april 1, 1940, in the rural village of ihithe. at a time when it was uncom-mon for girls to go to school, Muta attended a cath-olic missionary primary school and high school. Her superlative grades qualifi ed her as one of about 300 Kenyans to attend college in the United States under a program sponsored by Senator John F. Kennedy, in 1960. She earned a bachelor’s degree in biology at Mt. St. Scholastica college (now Benedictine col-lege) in Kansas and then a master’s degree in biol-ogy at the University of Pittsburgh. She returned to Kenya as a research assistant in veterinary science. She earned a Ph.D. in anatomy at University col-lege in nairobi in 1971, where she became a senior lecturer, department chair, and an associate profes-sor, the fi rst woman at nairobi to attain any of these positions.
Wangari Muta’s political activism began with seeking equal rights for women faculty, but her lead-ership in local organizations associated with the United nations environment Programme (UneP) opened her eyes to realize that most of Kenya’s social problems were associated with environmental deg-
radation. She organized local women volunteers to plant trees in nairobi, grown from seeds of local forest species rather than the imported species that were commonly used for tree plantations. She mar-ried Mwangi Mathai, who won a seat in the Kenyan parliament. When he sued for divorce because of her outspoken devotion to political issues, he insisted she drop his name, but instead she changed it to Maathai. She wanted to run for parliament herself, and she resigned from the university to do so but was opposed by the autocratic leader Daniel arap Moi. She was left without home or employment.
about this same time, Maathai’s tree-planting activity caught the attention of the norwegian For-estry Service and then the United nations, which provided support for her volunteers to continue planting trees, which led to the formal beginning of the Green Belt Movement. But the Green Belt Movement did more than plant trees. When she discovered that Moi’s government planned to use nairobi’s large public park to build a private busi-ness complex, Maathai led protests. Moi denounced her and her associates as crazy and even accused her of masterminding an aphid infestation. When she discovered that she was on an assassination list of government enemies, she organized media pro-tests and barricaded herself in her house until gov-ernment forces broke in and arrested her, charging her with treason. By now, Maathai’s struggle had gained much international attention. in 1992, eight american senators, including al Gore and edward Kennedy, warned Moi that relations with the United States would be severely harmed if he pursued his unsubstantiated accusations. Maathai continued leading protests, during which she was beaten by police, until all political prisoners were released.
M
xviii+598_Biodiversity_F.indd 358 11/17/11 1:13 PM

mammals 359
When, in 1998, Moi’s government again attempted to expropriate public forest land (this time for a golf course), the Green Belt Movement, along with parliament members and international observers, protested and were attacked—but the visual images of the attack spread around the world. then it hap-pened again in 2001. this time, however, Maathai was released without charge.
Maathai and her political supporters did well in the 2002 elections, when Moi lost power. She was elected to parliament and then appointed as assistant minister of the environment and natural resources. She was awarded the nobel Peace Prize in 2004. She continued to lead environmental restoration and edu-cation programs, such as the United nations Billion tree campaign that began in 2006. the Green Belt Movement has planted more than 30 million trees in Kenya. She gained world admiration for promot-ing conservation and tree planting as an essential component of democracy in nations where poor peo-ple suffer the most from environmental destruction. Maathai began the Green Belt Movement, not simply because she loved trees, but because she saw that trees were essential to the livelihoods of the poor: trees prevented floods, soil erosion, and droughts and provided essential materials such as firewood. Maathai was not so much planting trees as empower-ing the poor people of Kenya. She had seen forested hillsides in the village where she grew up and fresh-water in streams, and she saw the forests cut down and the streams dry up. Her passion may have been
best expressed in a statement she made to national Public Radio on September 27, 2007, where she indi-cated that watching a poor person cut down the last tree for firewood to cook her last meal is a situation we need to avoid by planting trees now. Maathai died in Kenya on September 25, 2011.
See also ecosystem services; restoration ecology.
Further readingGreen Belt Movement. “Wangari Maathai.” avail-
able online. URl: http://greenbeltmovement.org/w.php?id=59. accessed May 16, 2011.
Maathai, Wangari. Unbowed: A Memoir. new york: Ran-dom House, 2006.
mammals the class Mammalia consists of vertebrates that have live birth, hair, are warm-blooded, and produce milk. carolus linnaeus clas-sified mammals into one group and even invented the name, based on the latin mammae, referring to breasts. Mammals also have four-chambered hearts and teeth clearly differentiated into incisors, canines, premolars, and molars. Mammals constitute a van-ishingly small percentage of animal species—about 1 to 2 percent as many species as beetles—but are of intense interest to human observers.
CharaCTErISTICS of mammalSSome of the distinctive characteristics of mammals are explained below:
● Live birth. Monotremes such as the duck-billed platypus lay eggs. the eggs have rub-bery shells and remain glued to the mother’s underside for incubation while she is in her burrow. in all other mammals, the embryo begins to develop into a fetus inside the uterus of the mother. in marsupial mam-mals such as opossums and kangaroos, the very young fetus is born and crawls out of the birth canal into the pouch, inside of which they locate and nurse from nipples. in placental mammals, a placenta develops in which nutrients and oxygen from the mother’s blood diffuses into the developing blood system of the fetus. therefore pla-cental fetuses remain inside the uterus far longer than marsupial fetuses.
● Hair. Hairs are cylindrical structures made primarily of keratin, the same protein that makes up vertebrate skin, reptile scales, and bird feathers. Humans have about as many hair follicles per unit area of skin (about 1,500 per square inch or 250 per cm2) as
Wangari Maathai led thousands of Kenyan women to plant native trees in places where forests had been destroyed. (United Nations Environment Programme)
xviii+598_Biodiversity_F.indd 359 11/17/11 1:13 PM

first entry360 mammals 360
other mammals, but on most of the skin sur-face the hairs do not develop to full size. this produces the appearance of hairless skin in humans. Keratin is strong and lightweight. a single hair can hold almost a quarter of a pound (100 grams) of weight and stretch to twice its length before it severs. cells in the hair follicle adhere to the growing hair shaft and die; the hair shaft grows from the base and consists of dead cells. Hair is an excel-lent insulator, holding pockets of air against the skin. tiny muscles (arerectores pilorum; singular arrector pili) attach to each follicle and control the angle of each hair. When a mammal is cold, the muscles make the hairs stand straight up, holding warm air next to the skin. When a mammal is in danger, it may contract these muscles, allowing the vertical hairs to make it appear larger. On human skin surface that appears hairless, these muscles produce goose bumps. Goose bumps are therefore one of the vestigial characteristics of evolution.
● Warm blood. Mammals and birds are warm-blooded, because they have the high-est metabolic rates of all vertebrates. Mam-mals are endothermic (Greek for “heat from within”), which means they produce enough body heat to create a temperature that is usually warmer than the environment, and homeothermic (Greek for “same heat”), which means they regulate their body tem-perature at a relatively constant level. this allows mammals to have high metabolic rates even in cold environments or during winter.
● Milk. Mammae, possessed by both males and females, are modified sweat glands that produce milk, a liquid with a high content of sugars and fats and used to feed new-borns. Mammae do not develop in males except under unusual hormonal conditions, for example in starving men who suddenly have all the food that they can eat. in mono-tremes, the most primitive group of mam-mals, the mammae are scarcely more than milk glands on the abdomen. in marsupials and placentals, the mammae have nipples that deliver milk to newborns when sucked. Mammae swell during lactation. Uniquely among mammals, mammae of women store fats on a continual basis and function as indicators of fitness in sexual selection. the number of mammae on a female mammal is roughly proportional to the number of offspring she produces at one time. Some australian rats have at least 22 nipples and
may give birth to 17 pups at a time; in con-trast, humans have two nipples and usually give birth to one child at a time.
● Hearts. Mammal hearts have four cham-bers, which allow them to have two separate circuits of blood flow. the first circuit is the pulmonary circulation: Blood returns to the heart from throughout the body in veins, which deliver deoxygenated (dark) blood to the right atrium of the heart. the right ven-tricle pushes this blood to the lungs, where the red blood cells absorb oxygen and the blood becomes bright red. the red blood from the lungs enters the left atrium of the heart. the left ventricle pushes the red blood through the aorta and out into the rest of the body, which forms the second circuit, the systemic circulation. Mammal blood there-fore flows in a figure 8: a pulmonary loop alternating with a systemic loop. this sys-tem allows mammalian blood to be well and quickly oxygenated, which allows mammals to have high metabolic rates. Birds have even higher metabolic rates than mammals and also have four-chambered hearts. Most reptiles, except for crocodiles and their rela-tives, have three-chambered hearts and a slower metabolism.
● Teeth. Mammals have differentiated teeth. incisors allow mammals to bite their food, canines allow them to tear it, and premolars and molars allow them to grind it. Rep-tiles, in contrast, have teeth that are all more or less alike. Modern birds have no teeth, although their ancestors had reptil-ian teeth. not only does the differentiation of teeth allow mammals to have close con-trol over the processing of food but allows the developmental basis for specialization to different diets. Grazing animals have large molars, allowing them to grind coarse plant materials such as grass; they may possess no canine teeth at all. carnivores have large canine teeth that they use to capture and consume their prey (and to fight with one another). Rodents have incisors that grow continually, as they are worn down by pierc-ing the tough outer coverings of fruits and seeds. Humans have teeth less differenti-ated than those of most mammals, reflecting their omnivorous diet.
orIGIN of mammalSMammals evolved from synapsids, one of the lin-eages of reptiles. Many vertebrates have openings in the right and left temporal bones of the skull;
xviii+598_Biodiversity_F.indd 360 11/17/11 1:13 PM

mammals 361
synapsids have a single opening, unlike many other reptiles. One synapsid reptile of the late carbonifer-ous period was Dimetrodon, which had a huge sail of bony extensions along its back. although Dime-trodon itself is unlikely to have been the ancestor of mammals, it may have shared a characteristic that may have been widespread among synapsids: warm blood. Dimetrodon may have used its sail to warm its blood in the morning (by absorbing sunshine) and cool it off in midday (by radiating heat through the skin surface). this may have helped Dimetro-don to be, while not homeothermic, at least partly endothermic.
By the Permian period, the dominant lineage of synapsids was the therapsids (from the Greek for “nurse”). While earlier synapsids had a sprawling gait similar to that of modern reptiles, the therap-sids had legs underneath the body, allowing them to run more rapidly. Scientists infer that a higher meta-bolic rate (and more efficient body heat production) was necessary for this way of life. compared to ear-lier reptiles, therapsids had fewer skull bones, teeth that were more differentiated, and multiple sets of replacement teeth rather than just juvenile “milk teeth” replaced by adult teeth. One group of therap-sids that survived the Permian extinction was the cynodonts (from the Greek for “dog teeth”), with teeth even more differentiated (doglike) than those of earlier therapsids. Small pits in the facial bones of cynodonts may have been openings for extensive blood vessels, and this has led some scientists to conclude that cynodonts were fully warm-blooded. cynodonts such as Morganucodon and Sinocodon, which lived during the Jurassic period, represent nearly perfect “missing links” (actually not miss-ing) between earlier synapsid reptiles and modern mammals. Morganucodon is considered one of the earliest mammals, even though it is intermediate between synapsid reptiles and modern mammals. the very fact that so many “missing links” have been discovered means that the line between reptiles and mammals is somewhat arbitrary. like mod-ern mammals, Morganucodon had juvenile milk teeth replaced by adult teeth; Sinocodon retained the ancestral reptilian pattern of multiple replace-ment sets of teeth.
the intermediate status of the cynodonts is illus-trated in three ways:
● Lower jaw. in early synapsid reptiles, as in modern reptiles, the lower jaw consists of several bones, one of which is the den-tary bone; but in mammals the lower jaw consists entirely of the dentary bone. in cyn-odonts, the dentary bone was intermediate in size.
● Ear bones. in early synapsid reptiles, as in modern reptiles, the inner ear has a single bone, the stapes, which conducts vibrations to the cochlea. the mammalian inner ear consists of three bones: the incus, which corresponds to the reptilian quadrate bone; the malleus, which corresponds to the rep-tilian articular bone; and the stapes. in cynodonts, these bones are intermediate between the upper jawbones of reptiles and the inner ear bones of mammals. in fact, juvenile opossums (genus Didelphis) have inner ear bones that closely resemble those of Morganucodon.
● Jaw joint. in early synapsid reptiles, as in modern reptiles, the quadrate bone articu-lates with the articular bone of the skull, but in mammals it is the dentary bone that articulates with the skull. this means that during the evolution of mammals, the loca-tion of the jaw joint had to change from quadrate-articular to dentary-skull. in cyn-odonts, both jaw joints were functional.
From the carboniferous to the triassic periods, synapsids were the dominant reptiles on the earth, in terms of size and species diversity. By the end of the triassic period, synapsids had become less abundant and were represented mainly by small individuals. the thecodont lineage of reptiles, which evolved into crocodiles, dinosaurs, and birds, became dominant. Most of the evolutionary history of mammals occurred during the Mesozoic era, alongside the dinosaurs. though none was large or abundant, there were numerous species of mammals at that time. the dinosaurs were, on the average, larger and ruled the day, while most mammals were smaller and were probably most active at night. While carnivorous dinosaurs must have often eaten mammals, occasionally a large mammal would eat a small dinosaur. Scientists have found the fossil of a three-foot-long (1-m-long) Kepenomamus robus-tus mammal that had eaten a baby dinosaur 130 million years ago.
By the Jurassic period, mammals had backbones and shoulders that allowed them to have a more flexible, mammalian way of running. three major lineages of mammals survived by the cretaceous period: the monotremes, the multituberculates, and the therians. the modern descendants of the Juras-sic monotremes are the duck-billed platypus and the spiny anteater (echidna) of australia and nearby islands (see photograph on page 362). Monotremes retain some primitive features shared by most (but not all) reptiles. For example, they lay eggs. they also possess, like reptiles, a common opening for
xviii+598_Biodiversity_F.indd 361 11/17/11 1:13 PM
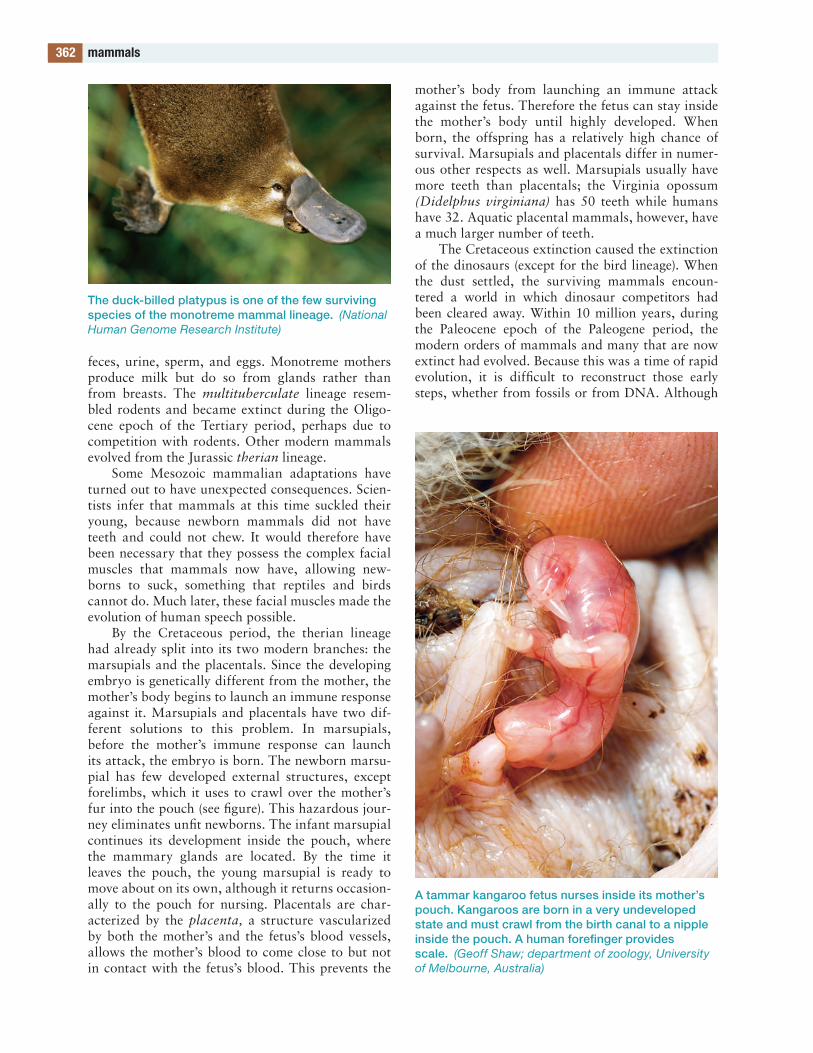
first entry362 mammals 362
feces, urine, sperm, and eggs. Monotreme mothers produce milk but do so from glands rather than from breasts. the multituberculate lineage resem-bled rodents and became extinct during the Oligo-cene epoch of the tertiary period, perhaps due to competition with rodents. Other modern mammals evolved from the Jurassic therian lineage.
Some Mesozoic mammalian adaptations have turned out to have unexpected consequences. Scien-tists infer that mammals at this time suckled their young, because newborn mammals did not have teeth and could not chew. it would therefore have been necessary that they possess the complex facial muscles that mammals now have, allowing new-borns to suck, something that reptiles and birds cannot do. Much later, these facial muscles made the evolution of human speech possible.
By the cretaceous period, the therian lineage had already split into its two modern branches: the marsupials and the placentals. Since the developing embryo is genetically different from the mother, the mother’s body begins to launch an immune response against it. Marsupials and placentals have two dif-ferent solutions to this problem. in marsupials, before the mother’s immune response can launch its attack, the embryo is born. the newborn marsu-pial has few developed external structures, except forelimbs, which it uses to crawl over the mother’s fur into the pouch (see figure). this hazardous jour-ney eliminates unfit newborns. the infant marsupial continues its development inside the pouch, where the mammary glands are located. By the time it leaves the pouch, the young marsupial is ready to move about on its own, although it returns occasion-ally to the pouch for nursing. Placentals are char-acterized by the placenta, a structure vascularized by both the mother’s and the fetus’s blood vessels, allows the mother’s blood to come close to but not in contact with the fetus’s blood. this prevents the
mother’s body from launching an immune attack against the fetus. therefore the fetus can stay inside the mother’s body until highly developed. When born, the offspring has a relatively high chance of survival. Marsupials and placentals differ in numer-ous other respects as well. Marsupials usually have more teeth than placentals; the Virginia opossum (Didelphus virginiana) has 50 teeth while humans have 32. aquatic placental mammals, however, have a much larger number of teeth.
the cretaceous extinction caused the extinction of the dinosaurs (except for the bird lineage). When the dust settled, the surviving mammals encoun-tered a world in which dinosaur competitors had been cleared away. Within 10 million years, during the Paleocene epoch of the Paleogene period, the modern orders of mammals and many that are now extinct had evolved. Because this was a time of rapid evolution, it is difficult to reconstruct those early steps, whether from fossils or from Dna. although
The duck-billed platypus is one of the few surviving species of the monotreme mammal lineage. (National Human Genome Research Institute)
A tammar kangaroo fetus nurses inside its mother’s pouch. Kangaroos are born in a very undeveloped state and must crawl from the birth canal to a nipple inside the pouch. A human forefinger provides scale. (Geoff Shaw; department of zoology, University of Melbourne, Australia)
xviii+598_Biodiversity_F.indd 362 11/17/11 1:13 PM

mammals 363
all modern orders of mammals existed in the early Paleogene, the dominant animals were in families that no longer exist, and the representatives of mod-ern orders did not look like their modern counter-parts. that is, modern orders existed, but modern families and genera did not. in contrast, most of the genera of early Paleogene trees would have been rec-ognizable to an experienced naturalist. in contrast to most mammals, most birds evolved into light-weight, airborne forms. there was, therefore, little direct competition between birds and mammals in the post-dinosaur world. One exception was in South america, where until only about 2 million years ago the largest vertebrate was a predatory phorusrhacid bird that stood almost 10 feet (three meters) tall.
mammal dIvErSITythere are or have been in the recent past about 5,000 genera of mammals. the surviving 1,135 gen-era contain about 4,700 species. this is far less than the almost one-and-a-quarter million species of insects, three-quarter million of which are beetles. though mammals have less species diversity than insects or even nematodes, they are overwhelm-ingly important in natural ecosystems and in the human perception of nature. Many are large grazers or predators, and some species have large popula-tions. their activities can transform entire ecologi-cal communities. almost 160 years ago, american naturalist Henry David thoreau made numerous careful observations that allowed him to conclude that the main reason that oak trees grew up under-neath pines was the activity of squirrels that buried the acorns. Grazing animals (harboring cellulytic bacteria in their digestive systems) transform a huge amount of plant material into wastes on which many insects feed and which cycle more rapidly through grassland ecosystems than would be the case with simple decomposition. Humans, being mammals themselves, are more fascinated by wild mammals than by any other group of organisms. this is especially true of the big ones, referred to as charismatic megafauna, a fact that conservation organizations know very well: they raise money to save pandas, not roundworms.
the five species of monotreme mammals (see above) are the last survivors of the most ancient mammal lineage still in existence. the duck-billed platypus and four species of echidnas lay eggs and have a single cloacal opening. the adults are tooth-less. Male monotremes have spurs on their hind legs that can inject poison into other animals.
there are about 315 species of marsupial mam-mals (see table on page 364). these include the 95 species of opossums, all of them in South america except for the Virginia opossum. these, together
with the six species of shrew opossums and Dromi-ciops gliroides (which more closely resembles aus-tralian marsupials), are the new World marsupials.
there are many species of australian marsupi-als. the red kangaroo (Macropus rufus) lives in arid regions of australia and can be six to seven feet (two meters) tall and weigh 200 pounds (90 kg). three other species of kangaroos are smaller and live in other parts of australia. Kangaroos have adapted well to human activity, especially ranching. they are the ecological equivalent of placental mammals such as deer and cattle. Unlike cattle, kangaroos do not release methane during the digestion of coarse vegetation in their diet; instead they produce citric acid, which provides further energy to the animal rather than being a waste product. thirty species of wallabies are small relatives of kangaroos. there are about 20 species of bandicoots, smaller than wallabies, which often live in forests. Because of their size, and because of their varied diet, they are the ecological equivalent of placental rodents. the tasmanian devil Sarcophilus harrisii lives today only on tasmania and resembles a stocky and ferocious placental dog. Wombats are medium-sized burrow-ing marsupials that eat underground vegetative mat-ter. there are two species of marsupial moles that, like placental moles, are blind burrowers. the koala is a slow-moving marsupial that consumes almost nothing but eucalyptus leaves; its slow metabolism allows it to subsist on these leaves, which are toxic and offer little nutrition. thylacines were called marsupial wolves because of their external resem-blance to dogs; the last thylacine became extinct in captivity in the early 20th century. these examples clearly demonstrate that most surviving or recently extinct marsupials closely resemble placental mam-mals, even though they evolved as entirely separate evolutionary lineages.
Modern placental mammals include the follow-ing groups (as well as others in the table on page 364).
Carnivores (order carnivora) (see photograph on page 365) have sharp canine teeth, sharp claws, and binocular vision, and most of them are preda-tors. they also have carnassial teeth (the last pre-molars and the first molars) that are sharp and come together in a scissorlike fashion that tears meat. cats, pinnipeds, and weasels are exclusively carnivorous, while raccoons and bears are omnivorous. even the giant panda bear (Ailuropoda melanoleuca), which primarily eats bamboo, will eat eggs or insects if they encounter them. they range in size from the least weasel (less than an ounce, or 25 g) to the elephant seal (11,000 pounds, or 5,000 kg). Most carnivores are terrestrial, although otters spend a lot of time in the water. Pinnipeds, however, are poor walkers and live mostly in water. their name (which means
xviii+598_Biodiversity_F.indd 363 11/17/11 1:13 PM
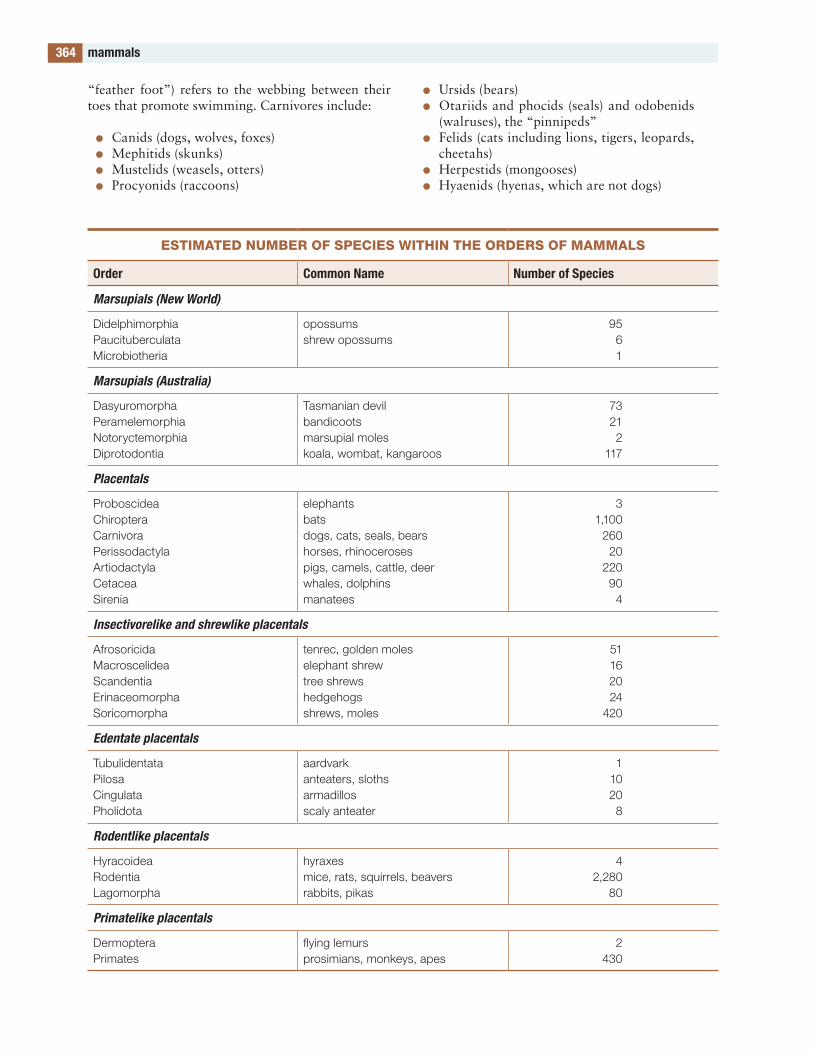
first entry364 mammals 364
“feather foot”) refers to the webbing between their toes that promote swimming. carnivores include:
● canids (dogs, wolves, foxes) ● Mephitids (skunks) ● Mustelids (weasels, otters) ● Procyonids (raccoons)
● Ursids (bears) ● Otariids and phocids (seals) and odobenids
(walruses), the “pinnipeds” ● Felids (cats including lions, tigers, leopards,
cheetahs) ● Herpestids (mongooses) ● Hyaenids (hyenas, which are not dogs)
estimated number of speCies Within the orders of mammals
order Common Name Number of Species
Marsupials (New World)
DidelphimorphiaPaucituberculataMicrobiotheria
opossumsshrew opossums
9561
Marsupials (Australia)
DasyuromorphaPeramelemorphiaNotoryctemorphiaDiprotodontia
Tasmanian devilbandicootsmarsupial moleskoala, wombat, kangaroos
73212
117
Placentals
ProboscideaChiropteraCarnivoraPerissodactylaArtiodactylaCetaceaSirenia
elephantsbatsdogs, cats, seals, bearshorses, rhinocerosespigs, camels, cattle, deerwhales, dolphinsmanatees
31,100
26020
22090
4
Insectivorelike and shrewlike placentals
AfrosoricidaMacroscelideaScandentiaErinaceomorphaSoricomorpha
tenrec, golden moleselephant shrewtree shrewshedgehogsshrews, moles
51162024
420
Edentate placentals
TubulidentataPilosaCingulataPholidota
aardvarkanteaters, slothsarmadillosscaly anteater
110208
Rodentlike placentals
HyracoideaRodentiaLagomorpha
hyraxesmice, rats, squirrels, beaversrabbits, pikas
42,280
80
Primatelike placentals
DermopteraPrimates
flying lemursprosimians, monkeys, apes
2430
xviii+598_Biodiversity_F.indd 364 11/17/11 1:13 PM

mammals 365
● Viverrids (civet cats, which are not cats) ● euplerids (Madagascar fossas)
Cetartiodactyls are the cetaceans (order ceta-cea) and even-toed ungulates (order artiodactyla).
● cetaceans (see photograph) have numerous adaptations that permit them to live entirely in water (all of them marine, except for four species of freshwater dolphins). the nostril (blowhole) is on top of the head, and the muscles contain a much greater amount of myoglobin, which holds oxygen, than those of other mammals. (Rabbit muscle may be 0.06 percent myoglobin by fresh weight, while some whale muscles are 6 percent myo-globin.) these adaptations allow cetaceans to dive deeply and for a long time. their ears are adapted for hearing underwater, where sounds can travel for long distances. the blue whale (Balaenoptera musculus) is the largest animal that has ever lived: it can be over 100 feet (more than 30 m) in length and weigh 190 tons (more than 170 metric tons). cetaceans feed either on plankton by baleen filters, as do the largest whales, or by hunting animals with their numerous teeth, as do the dolphins and smaller whales. the front legs of cetaceans are flippers, and they have no hind legs except for occasional evo-lutionary vestiges. cetaceans live in com-plex societies and migrate thousands of miles. they communicate by songs that are highly structured and can be heard almost a thousand miles (over a thousand kilometers) away. Dolphins have larger brains than most other mammals.
● the artiodactyls are herbivores that browse on herbs and shrubs or graze on grasses. When they walk, the weight is primarily on the third and fourth toes (even-toed). they include pigs; camels; cattle, sheep, and goats; deer and antelope; hippopota-muses; giraffes; and pronghorns. Pigs are omnivorous, and their stomachs have only two chambers; fermentation of coarse plant material occurs in the caecum, a pouch in the intestine. in contrast, ruminants (deer, giraffes, pronghorns, cattle, sheep, goats, antelope, and camels) have several stomachs in the first of which bacteria break down cellulose, a process that is aided by regurgi-tation of the undigested roughage (the cud) for further chewing.
Few scientists seriously considered that ceta-ceans and artiodactyls were closely related until fos-sil transitional forms from the early cenozoic era showed the relationship, which was confirmed by Dna comparisons. Manatees (order Sirenia) are, like the cetaceans to which they are not closely related, entirely aquatic, living in shallow waters rather than the open sea. they have front flippers and only remnants of hind legs.
Odd-toed ungulates (order Perissodactyla) includes horses, tapirs, and rhinoceroses. like artio-dactyls, they browse or graze. Unlike artiodactyls, their legs put most of the weight on the third (mid-dle) toe. in modern horses, the hoof is a single toe. they have simple stomachs; as in pigs, fermentation of coarse plant material in perissodactyls occurs in the intestine.
Bats (order chiroptera) are the only flying mam-mals, although several groups of mammals can glide. Bat wings, unlike bird wings, consist of a membrane
Carnivores, such as this Bengal tiger, have large sharp teeth that allow them to capture and tear the flesh of their prey. (iStockPhoto.com)
Cetaceans, such as this breaching whale, are fully adapted to marine existence. (SeaTurtle.org)
xviii+598_Biodiversity_F.indd 365 11/17/11 1:13 PM

first entry366 mammals 366
that connects elongated fingers; bat and bird flight represent an evolutionary convergence from separate nonflying ancestors. Most of the larger bats (also called flying foxes) and many smaller ones have large eyes and feed on fruit and pollen. the giant golden-crowned flying fox (Acerodon jubatus) has a wing-span of five feet (a meter and a half). Most of the smaller bats (about 70 percent of species) eat insects, which they locate using very high-pitched sounds. their brains construct something like a visual map using sound, very similar to human sonar imaging equipment. insectivorous bats, while relying primar-ily on sound for navigation, have rudimentary eyes and are not blind. Bats can form enormous colonies of a million or more individuals, which roost and nest in caves and emerge en masse at nightfall. a single cave can contain a million nesting bats, yet a mother can find her own offspring by distinguishing its vocalizations from those of others nearby.
Rodents (order Rodentia) constitute 40 percent of mammal species (see photograph). they eat pri-marily seeds (including those stored by humans) and
other plant material. their two long incisor teeth in each jaw are very sharp and allow them to penetrate some of the hardest seeds. these teeth continually grow, and rodents continually wear them down. they range in size from the african pygmy mouse (Mus minutoides) at a quarter of an ounce (seven grams) to the South american capybara (Hydro-choerus hydrochaerus) at 180 pounds (80 kg). Rodents include mice, rats, squirrels, chipmunks, gophers, porcupines, beavers, and guinea pigs. in contrast to the rodents, the rabbits, hares, and pikas (order lagomorpha) have four continually growing incisors in their upper jaws. Hyraxes (order Hyra-coidea) look like small rodents but are more closely related to elephants.
Primates (order Primates) usually have binocular (stereoscopic) vision and opposable thumbs. they have arboreal adaptations, for example the ability to jump or swing among branches, even if they do not live in trees. Primates have the largest brains rela-tive to body size of any order of mammals. Many have three-color vision, which is useful in identi-fying fruits amidst leaves; primates rely on vision much more than on other senses for locating food. they range in size from the pygmy mouse lemur (Microcebus myoxinus) at one ounce (30 g) to the mountain gorilla (Gorilla beringei) at 440 pounds (200 kg). the Strepserrhini (“curly-nosed”) primates include the lemurs, aye-ayes, and lorises. the Hap-lorrhini (“dry-nosed”) primates include the tarsiers, the platyrrhines (“flat-nosed”) new World monkeys, and the catarrhines (“downward-nosed”) Old World monkeys and apes. new World monkeys include capuchin, howler, and squirrel monkeys. Old World monkeys include macaques and baboons. apes include gibbons, orangutans, gorillas, chimpanzees, and humans. Humans are most closely related to the chimpanzee-bonobo lineage, from which they separated just more than 5 million years ago. the colugos (order Dermoptera) used to be considered primates. they have large surfaces of skin between all of their limbs and are the best gliding mammals.
Elephants (order Proboscidea) consist of only three species (two in africa, one in asia) but extinct mammoths, mastodons, and gomphotheres were close relatives of elephants. they are remarkable for their long noses, with which they can manipulate food and drink water, and for their long canine teeth, curled into tusks. they are the largest terres-trial mammals. they consume a great deal of leaves, stems, and fruits, sometimes bulldozing small trees to do so. they contribute importantly to the repro-duction of those very trees. elephants have large brains and are nearly unique among nonprimate mammals in being able to recognize their own reflec-tions. elephants can remember the voices of other
Some rodents, such as this black-tailed prairie dog (Cynomys ludovicianus) from the badlands of South Dakota, form complex social networks. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 366 11/17/11 1:13 PM

mammals 367
elephants from whom they have been separated a long time.
Edentates live primarily on small invertebrates. the ones that live on ants and termites (anteaters and aardvarks) have long snouts and long tongues that allow them to invade nests and few or no teeth. armadillos have epidermal scales that form a leath-ery armor. the giant armadillo (Priodontes maxi-mus) can be five feet (one and a half meters) long and weigh 130 pounds (60 kg). the only armadillo in north america is the nine-banded armadillo (Dasy-pus novemcintus). Sloths (see photograph) consume leaves that contain a lot of toxins, from which they obtain little nourishment. their very slow movement is an adaptation to this diet. the edentate manner of living has evolved separately several times; therefore, the old order edentata has been split into Pilosa (anteaters and sloths), tubulidentata (the aardvark Orycteropus afer), Pholidota (scaly anteaters), and cingulata (armadillos).
Insectivores are a diverse group of small mam-mals that live in forests and eat small invertebrates or are omnivorous. Moles dig burrows with their
large front legs and consume small invertebrates that fall into the burrows. they can dig toward animals whose vibrations they can feel with their tubular snouts. they have small eyes but are not blind. the insectivore manner of living has evolved separately several times; therefore, the old order insectivora has been split into Soricomorpha (shrews and moles), Scandentia (tree shrews), afrosoricida (tenrecs and golden moles), Macroscelidea (elephant shrews), and erinaceomorpha (hedgehogs).
Some placental mammals retain presumed ancestral characteristics. the hyraxes, for example, are only primitively homeothermic; they must occa-sionally bask in the Sun in order to maintain a constant body temperature. tree shrews have skel-etal features and feeding habits that resemble those of the earliest mammals. in contrast, several other orders of placental mammals have evolved amazing specializations to their environments and modes of living. consider the two extremes of size and habitat represented by the bats and the cetaceans.
Competition between marsupials and placentals. Wherever marsupials and placentals have been in contact, the placentals usually prevail:
● in the Mesozoic era, marsupials were wide-spread in the world. after the Oligocene epoch, however, most marsupials were found in australia and South america. Pla-cental mammals may have outcompeted them in other places.
● about 2.5 million years ago, the Panama land bridge formed, connecting north and South america for the first time since the breakup of Pangaea during the Mesozoic era. South america had many marsupial mammals, as well as some placentals such as sloths, armadillos, anteaters, and some rodents. When the land bridge formed, many placental mammals migrated south from north america and outcompeted marsupi-als in South america. Few South american marsupials successfully moved northward, the opossum being the lone north american marsupial success story.
● When modern humans arrived in australia thousands of years ago, they brought dogs, which became the wild dingos; europeans brought many placental mammals such as dogs, cats, and rodents a couple of hundred years ago. competition with placentals has driven most australian marsupials (except kangaroos) near or into extinction.
this should not, however, be taken as evidence of inherent inferiority of the marsupial reproductive
Sloths eat very low-quality food (toxic plant leaves) in tropical rain forests. One of their adaptations to a low-energy diet is to move very slowly and hang from branches by their claws. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 367 11/17/11 1:13 PM

first entry368 margulis, lynn368
system. Some observers think that marsupials lost in competition because they had not been, in southern continents, subjected during their evolution to the climatic extremes that had occurred in the northern continents. the fact that the marsupial reproductive system seems well suited to variable and occasion-ally harsh conditions seems to contradict this idea, however. in placentals, short of spontaneous abor-tion, the mother has to carry the fetuses to full term, even under stressful conditions such as famine. the infants may die soon after birth, but not until after the placental mother has made a tremendous prena-tal investment in them. in marsupials, in contrast, the fetus is born at a very early stage, and when food is scarce this is the stage at which they can be aban-doned. Marsupials, in addition, have a lower meta-bolic rate during rest, which may help them conserve scarce energy. Because marsupials appear superior under the stressful conditions that all lineages have experienced at some time or other, the reasons why the placentals have largely prevailed over the marsu-pials are not clear.
See also convergence; primates.
Further readingagustí, J., and M. antón. Mammals, Sabertooths, and Hom-
inids: 65 Million Years of Mammalian Evolution in Europe. new york: columbia University Press, 2002.
angielczyk, Kenneth D. “Dimetrodon is not a dinosaur: Using tree thinking to understand the ancient relatives of mammals and their evolution.” Evolution Educa-tion and Outreach 2 (2009): 257–271.
Benton, Michael. “Four Feet on the Ground.” chap. 4 of The Book of Life: An Illustrated History of the Evolu-tion of Life on Earth, edited by Stephen Jay Gould. new york: norton, 1993.
Frick, Winifred F., et al. “an emerging disease causes regional population collapse of a common north american bat species.” Science 329 (2010): 679–683.
Gibson, Robert a., et al. “Fatty acids of the milk and food of the platypus (Ornithorhynchus anatinus). Lipids 23 (1988): 377–379.
Gould, Stephen Jay. “Sticking up for marsupials.” chap. 28 in The Panda’s Thumb. new york: norton, 1980.
Hopson, J. a. “the mammal-like reptiles: a study of tran-sitional fossils.” American Biology Teacher 49 (1987): 16–26.
Jehle, Martin. “Marsupials: a southern success story.” in Paleocene Mammals of the World. available online. URl: http://www.paleocene-mammals.de/marsupials.htm. accessed august 3, 2010.
Kemp, t. S. The Origin and Evolution of Mammals. Oxford, U.K.: Oxford University Press, 2005.
Kielan-Jaworowska, Zofia, et al. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. new york: cambridge University Press, 2004.
Prothero, Donald R. “evolutionary transitions in the fossil record of the terrestrial hoofed mammals.” Evolution Education and Outreach 2 (2009): 289–302.
Rich, thomas H., et al. “independent origins of middle ear bones in monotremes and therians.” Science 307 (2005): 910–914.
Rubidge, Bruce S., and christian a. Sidor. “evolution-ary patterns among Permo-triassic therapsids.” Annual Review of Ecology and Systematics 32 (2001): 449–480.
teague, Matthew. “camel beauty queens.” National Geo-graphic, august 2009, 114–125.
turner, a., and M. antón. Evolving Eden: An Illustrated Guide to the Evolution of the African Large-Mammal Fauna. new york: columbia University Press, 2004.
margulis, lynn (1938– ) American Evolution-ary biologist lynn Margulis is one of the few scientists alive today who has changed some of the basic assumptions of a whole field of study. She persistently defended the endosymbiotic theory of the origin of mitochondria and chloroplasts that had been proposed decades earlier by Russian bota-nists and by american biologist ivan Wallin. She gathered evidence from many studies that finally established this theory. Mitochondria and chloro-plasts are the evolutionary descendants of bacteria. She is currently testing the theory that eukaryotic cilia, flagella, and microtubules are also the evolu-tionary descendants of bacteria. She has presented evidence that symbiogenesis (the symbiotic origin of cellular structures) is an important source of evolutionary novelty and diversification. Symbiosis (for example, with microorganisms living inside the roots of plants) made plant life on land possible, and plants then created the habitats in which animals lived. Margulis says, “Symbiogenesis was the moon that pulled the tide of life from its oceanic depths to dry land and up into the air.” She also explains that, because all life-forms interact so closely, they produce a global living system that regulates some physical conditions (such as temperature and atmo-spheric gas composition) of the entire planet, an idea called the Gaia hypothesis. to Margulis, sym-biosis is pervasive throughout the living world, from the smallest cellular components to the global eco-system. She says, “Symbiosis is not a marginal or rare phenomenon. it is natural and common. We abide in a symbiotic world.”
Margulis has always been a creative thinker and willing to take bold steps to pursue new theories. Born on March 5, 1938, lynn alexander was an early entrant, at age 14, into the University of chi-cago. She pursued a general liberal arts degree that emphasized independent study and the reading of
xviii+598_Biodiversity_F.indd 368 11/17/11 1:13 PM

marshes and swamps 369
original publications rather than textbooks. While at chicago she met another precocious and cre-ative student of science. astronomer carl Sagan was already a graduate student at age 19. Both she and Sagan, who later married, were to become two of the most creative thinkers and prolific writers in modern science, and both were to contribute in different ways to an understanding of planetary atmospheres as well as of evolution.
lynn and carl Sagan moved to Wisconsin. She studied genetics at the University of Wisconsin, Madison, then continued her studies at the Univer-sity of california at Berkeley, where she completed her doctorate in 1965. She became especially inter-ested in cytoplasmic inheritance, in which genetic traits were passed from one generation to another in the cytoplasm rather than by the nucleus. She found out that early 20th-century geneticists Hugo DeVr-ies and carl correns knew that not all of a plant cell’s genes were in the nucleus and had discovered that chloroplasts also had genes. But they and most geneticists since them had divided genetic inheri-tance into “nuclear v. unclear,” dismissing cytoplas-mic inheritance as unimportant. after 15 rejections and losses, lynn Sagan’s paper was published in 1967. Her persistence and her refusal to accept a simplistic doctrine eventually led to a revolution in biology.
lynn Margulis joined the biology faculty of Bos-ton University in 1966. the departmental chair, biologist George Fulton, described Margulis as “the only instructor who was paid half time and worked time and a half.” Margulis left Boston University, and in 1988 joined the faculty, first in biology then in geosciences, at the University of Massachusetts at amherst, where she is currently a distinguished professor. She believes strongly that students in her courses should go outdoors and directly observe organisms in their contexts of symbiotic interaction. Margulis is a member of the national academy of Sciences and has written numerous books and articles with her son, the writer Dorion Sagan.
See also Gaia; symbiogenesis.
Further readingMargulis, lynn. “Mixing it up: How i became a scientist.”
Natural History, September 2004, 80.——— Symbiosis in Cell Evolution: Microbial Communi-
ties in the Archaean and Proterozoic Eons, 2nd ed. new york: W. H. Freeman, 1993.
———. Symbiotic Planet: A New Look At Evolution. new york: Basic Books, 1998.
Margulis lynn, and Dorion Sagan. “the beast with five genomes.” Natural History, June 2001, 38–41.
———. Mystery Dance: On the Evolution of Sexuality. new york: touchstone, 1992.
———. Slanted Truths: Essays on Gaia, Symbiosis, and Evolution. new york: copernicus, 1997.
Margulis, lynn, and Karlene V. Schwartz. Five Kingdoms: An Illustrated Guide to the Phyla of Life on Earth, 3rd ed. new york: W. H. Freeman, 1998.
Sagan, lynn. “On the origin of mitosing cells.” Journal of Theoretical Biology, 14 (1967): 225–274.
Sciencewriters. “Sciencewriters: about the authors.” avail-able online. URl: http://www.xsnrg.com/sciencewriters/ authors.htm. accessed august 15, 2010.
marshes and swamps Marshes and swamps are terrestrial and coastal wetlands. they are flooded by shallow water most of the time; the dominant plants are rooted in the soil and emerge above the water surface. Grasslike plants dominate marshes, while shrubs or trees dominate swamps (see pho-tograph on page 370). in the absence of ecological disturbance, ponds become swamps or marshes. if a river changes course and takes a shorter route, the old riverbed becomes an oxbow swamp or marsh.
SomE TypES of wETlaNdSFreshwater marshes. the most abundant marsh plants are the grasslike sedges, rushes, cattails (or tules), and bur reeds in the shallow water and water lilies in the deeper water (see table on page 371). Many of these plants form thick, clonally spread-ing mats that cannot easily be penetrated by other plants. Surprisingly enough, it is often fire that keeps trees from growing in marshes. Fires burn plant tis-sues above water level or during the dry season; this kills trees and shrubs but allows the grasslike plants to resprout. One of the largest freshwater marshes in north america is the everglades of Florida. During the wet season, water flows from lake Okeechobee across a limestone shelf (which prevents the water from percolating into the ground) in a slow river about 60 miles (almost 100 km) wide into Florida Bay. it is dominated by sawgrass, a sedge. the cen-tral Valley of california (the Sacramento and San Joaquin Valleys) was until the 20th century the larg-est marsh in the United States. Snowmelt from the Sierra nevada fed the marshes, and the water was blocked by the coast Range. all the major rivers have been dammed and the entire marsh converted to irrigated agriculture.
trees may grow on hummocks of raised soil in marshes. live oak (Quercus virginiana) and gumbo limbo (Bursera simaruba) trees and Roystonea palms dominate the hummocks of the everglades.
Freshwater swamps. the most famous swamp tree is the bald cypress, a tall tree that dominates swampland throughout most of the eastern United States. cypress swamps are relatively undisturbed.
xviii+598_Biodiversity_F.indd 369 11/17/11 1:13 PM

first entry370 marshes and swamps 370
in many cases, new plants have no place to germi-nate except the stumps of old cypress trees. Swamps with more disturbance may be dominated by shrubs such as the hazel alder, which has numerous trunks continually sprouting from the rootstock as distur-bances destroy the older trunks.
Many marsh and swamp plants, such as saw-grass, grow best when oxygen can reach the roots. However, thick layers of anaerobic organic ooze accumulate at the bottom of many marshes and swamps. Bacteria in these deposits may produce methane, which can ignite and form a ghostly blu-ish glow known as will-o’-the-wisp or foxfire. the roots of swamp trees and shrubs also build up mounds of soil.
Estuaries. Brackish marshes or salt marshes have ocean water diluted by freshwater to varying degrees. the most abundant estuary plant is cord-grass. Salinity is one of the most important factors that produce zonation within estuaries. in San Fran-cisco Bay, cordgrass (Spartina foliosa) dominates the
zone between mean sea level and the average high tide level, while pickleweed, a small succulent (Sali-cornia virginica), dominates the zone above average high tide, where the soil contains more salt due to evaporation.
Mangrove swamps. around the world, man-groves form thick woodlands along beaches.
Wetland animals. Many species of animals live primarily in wetlands. Herbivores include muskrats (Ondatra zibethicus), northern river otters (Lontra canadensis), and beavers (Castor canadensis). Pred-ators include alligators (Alligator mississippiensis) and snapping turtles (Macrochelys temminckii). Bea-vers are a keystone species, as they construct dams that can transform a creek into a marsh or swamp. among the birds that are common in marshes are red-winged blackbirds (Agelaius phoeniceus), which live in the aerial vegetation, and wading birds such as the great blue heron (Ardea herodius), which hunt fishes. the larvae of many insect species live in wetlands.
Trees dominate swamps, such as this bald cypress swamp in the National Audubon Society’s Corkscrew Swamp Sanctuary in Florida. Emergent herbaceous plants are rooted to the soil under the water. (Marc Ryckaert)
xviii+598_Biodiversity_F.indd 370 11/17/11 1:13 PM
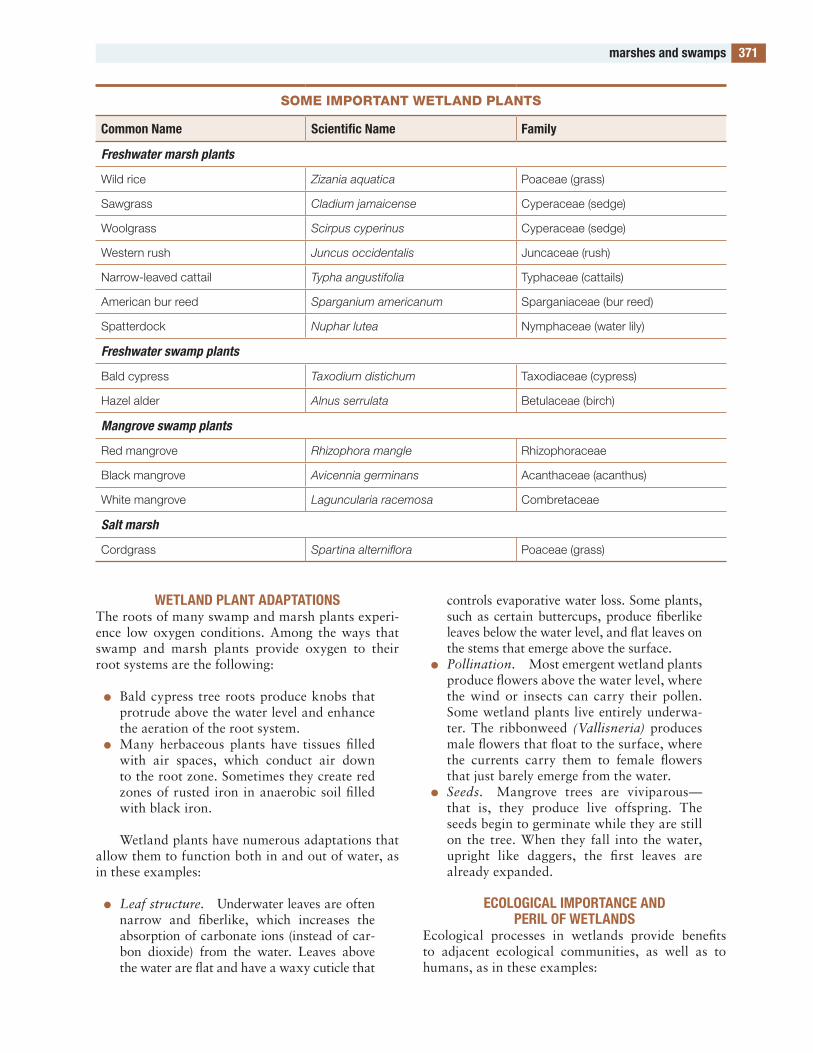
marshes and swamps 371
wETlaNd plaNT adapTaTIoNSthe roots of many swamp and marsh plants experi-ence low oxygen conditions. among the ways that swamp and marsh plants provide oxygen to their root systems are the following:
● Bald cypress tree roots produce knobs that protrude above the water level and enhance the aeration of the root system.
● Many herbaceous plants have tissues filled with air spaces, which conduct air down to the root zone. Sometimes they create red zones of rusted iron in anaerobic soil filled with black iron.
Wetland plants have numerous adaptations that allow them to function both in and out of water, as in these examples:
● Leaf structure. Underwater leaves are often narrow and fiberlike, which increases the absorption of carbonate ions (instead of car-bon dioxide) from the water. leaves above the water are flat and have a waxy cuticle that
controls evaporative water loss. Some plants, such as certain buttercups, produce fiberlike leaves below the water level, and flat leaves on the stems that emerge above the surface.
● Pollination. Most emergent wetland plants produce flowers above the water level, where the wind or insects can carry their pollen. Some wetland plants live entirely underwa-ter. the ribbonweed (Vallisneria) produces male flowers that float to the surface, where the currents carry them to female flowers that just barely emerge from the water.
● Seeds. Mangrove trees are viviparous—that is, they produce live offspring. the seeds begin to germinate while they are still on the tree. When they fall into the water, upright like daggers, the first leaves are already expanded.
EColoGICal ImporTaNCE aNd pErIl of wETlaNdS
ecological processes in wetlands provide benefits to adjacent ecological communities, as well as to humans, as in these examples:
some important Wetland plants
Common Name Scientific Name family
Freshwater marsh plants
Wild rice Zizania aquatica Poaceae (grass)
Sawgrass Cladium jamaicense Cyperaceae (sedge)
Woolgrass Scirpus cyperinus Cyperaceae (sedge)
Western rush Juncus occidentalis Juncaceae (rush)
Narrow-leaved cattail Typha angustifolia Typhaceae (cattails)
American bur reed Sparganium americanum Sparganiaceae (bur reed)
Spatterdock Nuphar lutea Nymphaceae (water lily)
Freshwater swamp plants
Bald cypress Taxodium distichum Taxodiaceae (cypress)
Hazel alder Alnus serrulata Betulaceae (birch)
Mangrove swamp plants
Red mangrove Rhizophora mangle Rhizophoraceae
Black mangrove Avicennia germinans Acanthaceae (acanthus)
White mangrove Laguncularia racemosa Combretaceae
Salt marsh
Cordgrass Spartina alterniflora Poaceae (grass)
xviii+598_Biodiversity_F.indd 371 11/17/11 1:13 PM

first entry372 marsupial mammals 372
● Breeding grounds for ocean fishes. Many marine fishes begin their lives in estuaries, where plankton food is abundant and the young fishes can hide from larger fishes.
● Cleaning the runoff water. Wetlands serve as a filter in which organic compounds (including some of human origin) decom-pose in the soil.
● Storm surge protection. Wetland plants, particularly mangroves, hold down the soil and protect inland areas from storm surges. Scientists frequently attribute the high lev-els of damage that follow tsunamis and hurricanes to the destruction of mangrove swamps, often to use the land for shrimp farms.
not only are wetlands threatened by habitat destruction, but they are frequently the recipients of pollution, which flows downstream and concentrates in them. Many of the examples of biomagnification occur in wetlands. Several invasive plant species are taking over large areas of wetlands as well. Many areas of open water in the southern United States have been covered with water hyacinth (Eichhornia crassipes) or water lettuce (Salvinia molesta). large areas of the everglades have been invaded by shrubs such as the pepper tree (Schinus terebinthinifolius) or large trees such as Melaleuca quinquenervia. Mela-leuca trees, which transpire huge amounts of water vapor into the air, were imported from australia spe-cifically to help drain the everglades, a job that they are now doing all too well. Purple loosestrife is an escaped ornamental wetland plant that is spreading throughout the marshes of the United States. it dis-places the wetland plants that serve as food for native animals. Because of their importance to other habi-tats, wetlands are a frequent focus of habitat mitiga-tion, in which an agency or business must establish a new wetland if they destroy or disturb an old one.
See also biomagnification; disturbances, ecological; keystone species.
Further readingBarnett, cynthia. Mirage: Florida and the Vanishing Water
of the Eastern U.S. ann arbor, Mich.: University of Michigan Press, 2007.
carter, W. Hodding. Stolen Water: Saving the Everglades from its Friends, Foes, and Florida. new york: Simon & Schuster, 2004.
Grunwald, Michael. The Swamp: The Everglades, Florida, and the Politics of Paradise. new york: Simon & Schuster, 2006.
Kathiresan, R. M. “allelopathic potential of native plants against water hyacinth.” Crop Protection 19 (2000): 705–708.
Mahall, Bruce e., and Roderic B. Park. “the ecotone between Spartina foliosa trin. and Salicornia virgi-nica l. in salt marshes of northern San Francisco Bay: i. Biomass and production.” Journal of Ecology 64 (1976): 421–433.
Myers, Randell S., et al. “Baldcypress (Taxodium distichum (l.) Rich.) restoration in southeast louisiana: the relative effects of herbivory, flooding, competition, and macronutrients.” Wetlands 15 (1995): 141–148.
naiman, Robert J., et al. “ecosystem alteration of boreal forest streams by beaver (Castor canadensis).” Ecol-ogy 67 (1986): 1,254–1,269.
Saiki, Michael K., and t. Peter lowe. “Selenium in aquatic organisms from subsurface agricultural drainage water, San Joaquin Valley, california.” Archives of Environmental Contamination and Toxicology 16 (1987): 657–670.
Valiela, ivan, et al. “Macroalgal blooms in shallow estuar-ies: controls and ecophysiological and ecosystem con-sequences.” Limnology and Oceanography 42 (1997): 1,105–1,118.
marsupial mammals See mammals.
mass extinctions Mass extinction occurs when many species become extinct at roughly the same time. extinction in general may result if popu-lations do not have the genes that allow them to adapt to environmental changes, that is, from “bad genes”; but extinction may also occur suddenly as a result of a worldwide disaster, for which no popula-tions possess genetic resistance, that is, from “bad luck,” to use paleontologist David Raup’s terminol-ogy. Global mass extinctions are usually considered to have resulted from “bad luck.” therefore true mass extinction events may result from rare cata-strophic events rather than an acceleration of normal extinction patterns.
Most of the geological periods ended with global changes in climate and the origin of recog-nizably different sets of species. that is, there were global extinctions at the end of every geological period. Five of these events were mass extinctions (see table on page 373). two of the mass extinction events brought the first two eras of the Phanero-zoic eon: the Permian extinction ended the Paleo-zoic era, and the cretaceous extinction ended the Mesozoic era. a mass extinction, especially of the ediacaran organisms, may have occurred at the end of the Precambrian time. the Ordovician, Permian, and cretaceous mass extinctions occurred against a background of rapid evolution and therefore were caused by global catastrophes, whereas the Devo-nian and triassic mass extinctions occurred against
xviii+598_Biodiversity_F.indd 372 11/17/11 1:13 PM
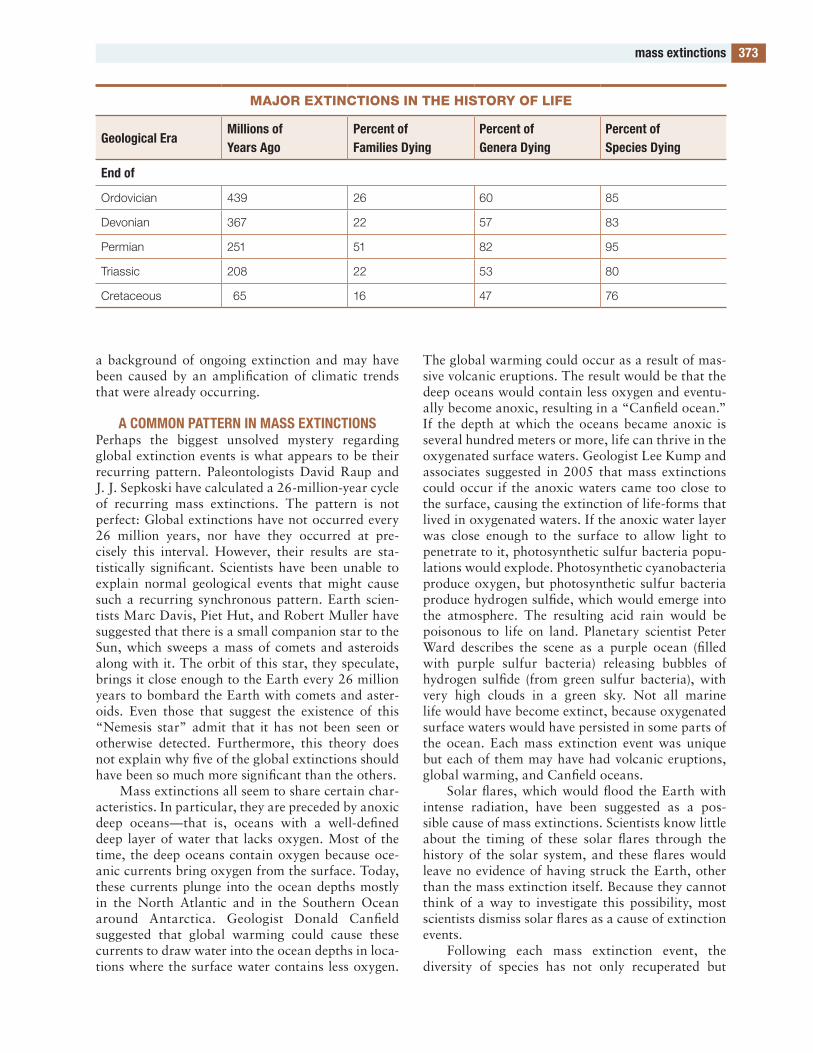
mass extinctions 373
a background of ongoing extinction and may have been caused by an amplification of climatic trends that were already occurring.
a CommoN paTTErN IN maSS ExTINCTIoNSPerhaps the biggest unsolved mystery regarding global extinction events is what appears to be their recurring pattern. Paleontologists David Raup and J. J. Sepkoski have calculated a 26-million-year cycle of recurring mass extinctions. the pattern is not perfect: Global extinctions have not occurred every 26 million years, nor have they occurred at pre-cisely this interval. However, their results are sta-tistically significant. Scientists have been unable to explain normal geological events that might cause such a recurring synchronous pattern. earth scien-tists Marc Davis, Piet Hut, and Robert Muller have suggested that there is a small companion star to the Sun, which sweeps a mass of comets and asteroids along with it. the orbit of this star, they speculate, brings it close enough to the earth every 26 million years to bombard the earth with comets and aster-oids. even those that suggest the existence of this “nemesis star” admit that it has not been seen or otherwise detected. Furthermore, this theory does not explain why five of the global extinctions should have been so much more significant than the others.
Mass extinctions all seem to share certain char-acteristics. in particular, they are preceded by anoxic deep oceans—that is, oceans with a well-defined deep layer of water that lacks oxygen. Most of the time, the deep oceans contain oxygen because oce-anic currents bring oxygen from the surface. today, these currents plunge into the ocean depths mostly in the north atlantic and in the Southern Ocean around antarctica. Geologist Donald canfield suggested that global warming could cause these currents to draw water into the ocean depths in loca-tions where the surface water contains less oxygen.
the global warming could occur as a result of mas-sive volcanic eruptions. the result would be that the deep oceans would contain less oxygen and eventu-ally become anoxic, resulting in a “canfield ocean.” if the depth at which the oceans became anoxic is several hundred meters or more, life can thrive in the oxygenated surface waters. Geologist lee Kump and associates suggested in 2005 that mass extinctions could occur if the anoxic waters came too close to the surface, causing the extinction of life-forms that lived in oxygenated waters. if the anoxic water layer was close enough to the surface to allow light to penetrate to it, photosynthetic sulfur bacteria popu-lations would explode. Photosynthetic cyanobacteria produce oxygen, but photosynthetic sulfur bacteria produce hydrogen sulfide, which would emerge into the atmosphere. the resulting acid rain would be poisonous to life on land. Planetary scientist Peter Ward describes the scene as a purple ocean (filled with purple sulfur bacteria) releasing bubbles of hydrogen sulfide (from green sulfur bacteria), with very high clouds in a green sky. not all marine life would have become extinct, because oxygenated surface waters would have persisted in some parts of the ocean. each mass extinction event was unique but each of them may have had volcanic eruptions, global warming, and canfield oceans.
Solar flares, which would flood the earth with intense radiation, have been suggested as a pos-sible cause of mass extinctions. Scientists know little about the timing of these solar flares through the history of the solar system, and these flares would leave no evidence of having struck the earth, other than the mass extinction itself. Because they cannot think of a way to investigate this possibility, most scientists dismiss solar flares as a cause of extinction events.
Following each mass extinction event, the diversity of species has not only recuperated but
major extinCtions in the history of life
Geological Eramillions of years ago
percent of families dying
percent of Genera dying
percent of Species dying
End of
Ordovician 439 26 60 85
Devonian 367 22 57 83
Permian 251 51 82 95
Triassic 208 22 53 80
Cretaceous 65 16 47 76
xviii+598_Biodiversity_F.indd 373 11/17/11 1:13 PM

first entry374 mass extinctions 374
increased. the best fossil record that is available is that of marine invertebrates. this record indicates a steady increase in worldwide species diversity, espe-cially after mass extinction events. One example of evolution following a mass extinction is the adaptive radiation of mammal species after the extinction of the dinosaurs.
Some extinction events, although not considered mass extinctions, have still had an important effect on the evolutionary history of life. the Hemphillian extinction event about 5 million years ago drove relatively few species to extinction, but among them were many grazing animals. a diversity of horse spe-cies was reduced to just one.
terrestrial plants may not respond to mass extinction events as rapidly or as markedly as marine and terrestrial animals. Most plant extinctions have been caused by long-term climatic changes. While many plants became extinct during the five mass extinction events, their response was delayed until several million years after each of the extinction events.
the Permian mass extinction that brought the Permian period and the whole Paleozoic era to an end 250 million years ago was, in the words of paleontologist Douglas erwin, the “mother of all extinctions.” at this time, at least half of all families of organisms became extinct. Because some families contain many species, it has been estimated that up to 95 percent of all species became extinct at that time! Biodiversity recovered after the Permian extinction, but it took 100 million years to regain its former level. and the Permian extinction had long-term consequences. During the Permian period, the oceans contained many filter feeders that remained attached to rock surfaces. after the Permian extinc-tion, many motile animals evolved in the oceans. therefore the post-Permian increase in diversity was not merely the evolution of new species but new ways of obtaining resources. Furthermore, during the Permian period, mollusks (Phylum Mollusca) and lampshells (Phylum Brachiopoda) were of roughly equal abundance, but (perhaps due to chance) the lampshells suffered greater extinction than the mol-lusks. today, mollusks are abundant and lampshells remain rare.
ThE CrETaCEouS ExTINCTIoN waS uNIquEthe most famous example of a mass extinction event is the cretaceous extinction event 65 million years ago. at that time, many species of organisms, most famously the dinosaurs, became extinct. exception-ally high levels of the element iridium in the geo-logical deposits 65 million years ago suggest that the earth was hit by an asteroid; asteroids usually con-tain higher levels of this element than do the rocks
of the earth’s crust. the greatly eroded remnants of this impact can still be seen in the chicxulub crater of yucatán. even though some evidence for an aster-oid collision at the end of the Permian period has been presented, it has failed to convince most scien-tists. therefore the cretaceous extinction may have been the only one caused by an asteroid.
even the cretaceous extinction event, however, had more than one cause. By the time the asteroid hit the earth, populations of many dinosaur and other species had already been in decline for millions of years. (However, many other species flourished up until the moment of the extinction event.) Moreover, a massive volcanic eruption occurred about the same time in what is now india, producing a 200,000 square mile (over 500,000 km2) lava flow called the Deccan traps. Several interacting causes, the rela-tive importances of which are still debated, brought about the cretaceous extinction.
asteroid collisions and other planetary disasters have been relatively rare for the past 1 or 2 bil-lion years. according to earth scientists Peter Ward and David Brownlee, humans should, so to speak, thank their lucky stars for this: the Sun is a rela-tively stable star, compared to most in the universe, and the planet Jupiter (which can be considered a star that never ignited) has swept part of the solar system free of most asteroids except those in the asteroid belt. craters on the Moon appear to be mostly 3 to 4 billion years old, indicating that dur-ing this time period comets and asteroids were very common in the solar system. even though bacterial life appeared on earth soon after earth’s formation, frequent bombardment of the earth during that period may have delayed the appearance of complex cells until about a billion and a half years ago. the evolution of complex life-forms would have been impossible on a planet subjected to frequent mass extinctions.
ThE SIxTh maSS ExTINCTIoNMany scientists describe the extinctions that are now occurring throughout the world, as a result of human activities such as the destruction of natural habitats, as the “sixth mass extinction.” extinctions are in fact now occurring at a rate that would justify this description. Before the appearance of humans, the global extinction rate was about one per million species per year; now the global extinction rate is one per thousand per year.
it is possible that global warming, caused by human carbon emissions, may eventually make its own contribution to the sixth mass extinction. it is possible that the most recent canfield ocean event was the Paleocene-eocene thermal Maxi-mum, a brief period of global warming following
xviii+598_Biodiversity_F.indd 374 11/17/11 1:13 PM

monocots 375
a sharp rise in atmospheric carbon dioxide that occurred 55 million years ago. although it was not a mass extinction, this event was associated with a tremendous turnover of animal species, par-ticularly the extinction of early cenozoic mammals and the origin of modern mammalian groups. the atmospheric carbon dioxide concentration was only about 800 parts per million, a level that the atmo-sphere may reach again by the year 2100 if current rates continue.
Writer Wendell Berry comments, regarding the countless extinctions that are now occurring, “We can less afford [these] losses than nature can.” it is humanity that is perhaps harmed most by the mass extinction that it is causing. nor can we simply blame big business or government for destroying nature. all participants in the world economy are partly responsible. Berry noted that humans are all complicit in the murder of creation.
See also acid rain and fog; bacteria; cre-taceous extinction; extinction; Permian extinction.
Further readingcourtillot, Vincent. Evolutionary Catastrophes: The Sci-
ence of Mass Extinction. transl. Joe Mcclinton. new york: cambridge University Press, 2002.
Davis, M., P. Hut, and R. Muller. “extinction of species by periodic comet showers.” Nature 308 (1984): 715–717.
Hallam, anthony. Catastrophes and Lesser Calamities: The Causes of Mass Extinctions. new york: Oxford Uni-versity Press, 2004.
——— and P. B. Wignall. Mass Extinctions and Their Aftermath. new york: Oxford University Press, 1997.
Horner, Jack, and James Horman. How to Build a Dino-saur: Extinction Doesn’t Have to Be Forever. new york: Dutton, 2009.
Krug, andrew Z, et al. “Signature of the end-cretaceous mass extinction in the modern biota.” Science 323: (2009): 767–771. Summarized by crame, J. alistair. “time’s stamp on modern biogeography.” Science 323 (2009): 720–721.
Kump, lee, et al. “Massive release of hydrogen sulfide to the surface oceans and atmosphere during intervals of oceanic anoxia.” Geology 33 (2005): 397–400.
leakey, Richard, and Roger lewin. The Sixth Extinction: Patterns of Life and the Future of Humankind. new york: anchor Books, 1996.
lidgard, Scott, et al. “the search for evidence of mass extinction.” Natural History, September 2009, 26–32.
Raup, David M. Extinction: Bad Genes or Bad Luck? new york: norton, 1991.
Villier, loïc, and Dieter Korn. “Morphological disparity of ammonoids and the mark of Permian mass extinc-tions.” Science 306 (2004): 264–266.
Ward, Peter D. Under a Green Sky: Global Warming, the Mass Extinctions of the Past, and What They Can Tell Us About Our Future. new york: Harpercollins, 2008.
——— and David Brownlee. Rare Earth. new york: copernicus Books, 2000.
——— and alexis Rockman. Future Evolution. new york: Freeman, 2001.
mimicry See camouflage and mimicry.
mineral cycles See biogeochemical cycles.
miocene epoch See neogene period, biodi-versity in.
monocots Monocots are an evolutionary lineage of flowering plants. there are at least 90,000 species of monocots, ranging from tiny herbaceous plants to trees. along with the conifers, the flowering plants have transformed the landscape of the earth into a living surface of green. in addition, flowering plants have undergone coevolution with animals that pol-linate their flowers and disperse their seeds. as a result, the flowering plants have filled the world with an astounding diversity of form, color, and scent.
Flowering plants have flowers and fruits. Flow-ers may contain:
● sepals: leaflike protective structures; ● petals: often colorful structures that attract
pollinators; ● stamens: male structures that produce pol-
len; and ● pistils: female structures that produce seeds.
the term monocot refers to the embryo of the plant: when it emerges from the seed, the embryo has just one cotyledon, or embryonic leaf. Most mono-cots also have veins that are mixed with starch stor-age cells in the middle of the roots and in the young stems and are parallel to one another in the leaves, in contrast to the arrangement of veins in the dicots, the embryos of which have two cotyledons.
moNoCoTS wITh Showy flowErSSome families of monocots have brightly colored flowers that attract pollinators (see photographs on page 376). Below are some important examples.
the lilies (liliaceae) are a large family of monocots, which includes lilies, daffodils, onions, and agaves. Some taxonomists divide this family
xviii+598_Biodiversity_F.indd 375 11/17/11 1:13 PM

first entry376 monocots 376
into several smaller ones. they are all herbaceous, except the succulent agaves. the flowers have three sepals and three petals. Sometimes the sepals resem-ble the petals, giving the appearance of six petals. Six stamens surround a pistil of three united seed chambers.
the irises (iridaceae) have flowers similar to those of lilies, except that the three stamens sur-round a pistil whose seed chambers develop below the flower.
the orchids (Orchidaceae) are very diverse, with at least 20,000 species, many of them pollinated by only one kind of insect. Orchid flowers have three petals, one of which (the labellum) is larger and more ornate than the others (see figure). in some orchids, the labellum forms an intricate mechanism that deposits packets of pollen on the body of the pollinator, perhaps temporarily entrapping it in the process. Many orchids are epiphytes, living in the branches of tropical trees and obtaining water from rain. Because epiphytic orchids cannot obtain water from the soil, they often have succulent leaves that store water. they produce a large number of very
tiny seeds, some of which consist of only a few cells and cannot germinate unless they form a mutualistic partnership with a fungus.
Palm trees (arecaceae) are the largest monocots. their trunks would not be able to support large branches. the large leaves emerge from a single crown at the top of the tree.
the arums (araceae) such as jack-in-the-pulpit produce many tiny staminate and pistillate flowers on a spike, which is surrounded by a large, usually colorful leaf. it is this leaf, rather than the flowers themselves, that attracts pollinators.
moNoCoTS wIThouT Showy flowErSSeveral families of monocots have flowers the sepals and petals of which are greatly reduced. these monocots rely on wind pollination.
the grass family (Poaceae) is one of the most important families of flowering plants on earth, one of two families of seed plants (along with the pines) that can be seen from outer space. their roots hold the soil, and their leaves and stems produce
The flowers of monocots, such as this Tradescantia ohiensis, often have three petals and either three or (in this case) six stamens. The leaves often have par-allel veins. (Stanley A. Rice)
Orchids are recognizable because one of their three petals is fancy and colorful relative to the others. (Weddingaces.com)
xviii+598_Biodiversity_F.indd 376 11/17/11 1:13 PM

muir, John 377
most of the food, in grassland ecosystems that cover almost one-quarter of the earth’s dry land surface. they have coevolved with grazing mammals, largely because their leaves have cells that allow them to grow from the base of the blade; the leaves can there-fore keep growing after the tip has been eaten. each tiny flower is enclosed within two specialized leaves (bracts), and the flowers are compressed into a spike-let. Spikelets, in turn, can be arranged on reproduc-tive stems in many ways. the flowers are visible only when the stamens and stigmas emerge from between the bracts during pollination. in addition to their ecological importance, the economic importance of grasses can hardly be overstated: wheat, maize, rice, sugarcane, oats, rye, and barley are all grasses.
the sedge family (cyperaceae) contains many species that live in and maintain the ecological integrity of wetlands. they closely resemble grasses, except that the tiny flowers of their spikelets are pressed against the reproductive stem by a single bract, rather than between two. although the only economically important sedge today is the water chestnut, this family played an important role in human history. leaves of the papyrus sedge were used by the ancient egyptians to make the first paper.
the rush family (Juncaceae) is also aquatic and resembles grasses and sedges. its tiny flowers, how-ever, have sepals and petals.
the cattail family (typhaceae) has long suc-culent leaves that emerge from an underwater stem. the reproductive stalk produces thousands of tiny staminate and pistillate flowers. When the pistillate flowers form seeds, they remain on the stem, pro-ducing the characteristic appearance of a hot dog on a stick.
From simple ancestors that lived near the beginning of the cretaceous period, the flowering plants have diversified into hundreds of thousands of species, produced many specialized reproductive structures, and adapted to every habitat on earth, including coastal antarctica. Of the two flower-ing plant species that live in antarctica, one is a monocot and one is a dicot. Most of these flowering plants are dicots. no other group of multicellular organisms, except the insects, have evolved such tre-mendous diversity. explorers continue to discover new species of monocots, usually but not always in tropical forests. Many species are driven to extinc-tion, often before they have been studied by scien-tists, when humans destroy their habitats. this is a tremendous loss to the ecological communities of which they are the basis, to scientific research, and to the human economy.
See also dicots; flowering plants (angio-sperms); pollination.
Further readingBrown, lauren. Grasses: An Identification Guide. Boston:
Houghton Mifflin, 1992.Dafni, a., and y. ivri. “Floral mimicry between Orchis isra-
elitica Baumann and Dafni (Orchidaceae) and Belleva-lia flexuosa Boiss. (liliaceae).” Oecologia 49 (1981): 229–232.
Darwin, charles R. On the Various Contrivances by which British and Foreign Orchids are Fertilised by Insects. london: John Murray, 1862.
Johnson, S. D., and K. e. Steiner. “long-tongued fly pol-lination and evolution of floral spur length in the Disa draconis complex (Orchidaceae).” Evolution 51 (1997): 45–53.
neiland, Mary Ruth M., and christopher c. Wilcock. “Fruit set, nectar reward, and rarity in the Orchi-daceae.” American Journal of Botany 85 (1998): 1,657–1,671.
Riffle, Robert lee. The Timber Press Pocket Guide to Palms. Portland, Ore.: timber Press, 2008.
monocultures and polycultures (agri-culture) See biodiversity, resources for agriculture.
monotremes See mammals.
montane forests See temperate coniferous forests.
mosses See seedless plants.
muir, john (1838–1914) American Conser-vationist Born april 21, 1838, in Scotland, John Muir grew up as part of a large family of farmers in Wisconsin. While growing up, Muir acquired a deep love of the land and was disturbed to see the widespread destruction of the eastern forests. He studied botany at the University of Wisconsin, but did not finish a degree. He went to canada during the american civil War, apparently to avoid the draft, and worked in a sawmill. While he worked in a factory in indiana, an accident left Muir tempo-rarily blinded in one eye, and when he regained his sight, he decided to walk everywhere he could to see and appreciate the natural world. He walked from indiana to Florida. He went to california to recover from malaria he contracted in Florida. in 1868, he first saw yosemite Valley, in the Sierra nevada, and he stayed there for six years, working mostly as a shepherd.
xviii+598_Biodiversity_F.indd 377 11/17/11 1:13 PM

first entry378 mutualism 378
While wandering in the Sierras, Muir wrote articles about nature and what he had learned on his many hikes. He had a deeply spiritual reverence for all of the species in the forests and meadows (he often talked with wildflowers) and understood their interdependence. He valued the preservation not just of natural resources but of wilderness and of wilder-ness not just for recreation but for its spiritual value. Muir became a celebrity in yosemite Valley for his knowledge of biodiversity and as a guide. He loved the writings of Ralph Waldo emerson and Henry David thoreau, whose writings promoted the kind of life that Muir was living. emerson went to yosem-ite and met Muir in 1871 and offered him a profes-sorship at Harvard—which Muir declined.
Muir dedicated himself to wilderness preserva-tion. in 1892 he cofounded the Sierra club, which remains one of the principal voices for conservation of wilderness. Muir was also the principal voice behind the formation of yosemite national Park in 1899.
Muir met Gifford Pinchot, who was to become the most important force for government conserva-tion in the early 20th century, while hiking in the adirondacks, and they became friends and trav-eled extensively through western forests. although they shared a deep love of nature, their views on conservation differed markedly. Pinchot believed that the purpose of conservation was the efficient management of natural resources for the long term, while Muir defended wilderness preservation. in 1907, Pinchot (by then head of the United States Forest Service under President theodore Roosevelt) defended the creation of Hetch Hetchy reservoir in the Sierras by building a dam and flooding a valley near yosemite, while Muir opposed it vigorously. Pinchot and Muir spent their later lives debating their conflicting views of what conservation was supposed to accomplish.
Muir married into a wealthy family in 1878 and became the manager of an orchard and a ranch. as Muir’s health began to fail, his wife encouraged him to go back to hiking in the mountains in the Pacific northwest. On one of these hikes, in 1914, Muir got pneumonia. He died on December 24, 1914.
See also environmental ethics; federal government role in conservation (United States); Pinchot, Gifford; thoreau, Henry David.
Further readingegan, timothy. The Big Burn: Teddy Roosevelt and the
Fire that Saved America. Boston: Houghton Mifflin, 2008.
Worster, Donald. A Passion for Nature: The Life of John Muir. new york: Oxford University Press, 2008.
mutualism Mutualism is a symbiotic relation-ship between two species that benefits both of them. each provides some service, often a nutrient, to the other. Many thousands of examples of mutualism are known, of which a few are here described.
dIGESTIvE aNd oThEr muTualISmS ThaT maINTaIN vErTEBraTE hEalTh
cows cannot digest grass, which is odd because they walk around and moo and eat grass all day. it is not the cow itself but the bacteria that live in its rumen (the first stomach) that digest cellulose, one of the major components of grass. the cow chews the grass and delivers it to the rumen. the bacteria live inside the rumen, where they are protected in warm, moist, anaerobic conditions, and digest food that has been broken into small pieces by the cow’s teeth. after a while, the grass returns to the cow’s mouth, where it “chews the cud” to further break down the grass, then swallows it again. the bacteria digest cellu-lose into sugar for their own benefit, but the cow’s stomach absorbs much of the sugar that the bacteria release. Many other ruminant animals have a simi-lar relationship with digestive bacteria. eucalyptus leaves are not very nutritious, but koalas eat large amounts of them and allow bacteria to release nutri-ents from them. leaves of rain forest plants contain many toxins and are not very nutritious, but animals such as sloths and colobus monkeys eat them, relying on digestive bacteria. Marine iguanas of the Galápa-gos islands are the only reptiles to eat seaweeds, and they rely on intestinal bacteria to help them digest their food.
Most human intestinal bacteria are commensals, but some of them produce waste products that, to humans, are vitamins. the main human source of vitamin K and an important source of B vitamins are intestinal bacteria. One-quarter of the bacteria in the human intestine are Bacteroides thetaiotaomi-cron, which competes with food-borne pathogenic bacteria and produce protective compounds such as lactate, peroxide, and bacteriocins. they even stimu-late the production of blood vessels in the lining of the intestine.
in order to demonstrate the importance of intes-tinal bacteria in humans, it would be necessary to raise a human in completely sterilized conditions, which would be unethical even if it were possible. Such experiments can be done upon rats, which are physiologically similar to humans. Since a newborn mammal is exposed to bacteria during the birth process, scientists deliver baby rats by cesarean sec-tion and raise them in sterilized cages on steril-ized rat chow. these gnotobiotic rats, never having been exposed to bacteria, are unhealthy. they show symptoms of vitamin deficiency and eat 30 percent
xviii+598_Biodiversity_F.indd 378 11/17/11 1:13 PM

mutualism 379
more calories, perhaps in an attempt to compensate for the lack of vitamins. they also have impaired immune function.
Proper intestinal function in many animals requires a balanced community of bacterial species, a balance that can be disturbed by the use of broad-spectrum oral antibiotics. Pathogenic bacteria may thrive in the depleted community following the use of these antibiotics. typically, however, some of the resident bacteria have remained alive in the lining of the intestine and recolonize it. Disruption of diges-tive bacteria is, however, one of the reasons that the use of antibiotics should be reserved for emergency situations.
there appears to be mutualism among the intestinal bacteria as well. Most of the intestinal bacteria are anaerobic—they die upon exposure to oxygen. But Escherichia coli (the famous “E. coli”) are microaerophiles, which means that their growth is enhanced by low levels of oxygen. E. coli lives mostly near the lining of the intestine, where oxygen diffuses out of the tiny blood vessels, and in so doing they use up the oxygen before it reaches the middle of the intestine, where the anaerobes live.
there are many other examples of protective bacteria in the human body. One of them is the Lac-tobacillus bacteria that live in the vaginas of women, where they convert sugar to lactic acid. the acid inhibits the growth of Candida albicans fungi that
might otherwise cause infections. Some skin bacteria produce antimicrobial compounds. Some microbes in the human mouth produce antimicrobial com-pounds that help prevent gum disease. they do so as a weapon against other microbes with which they compete for food, but the indirect result is the main-tenance of human health.
another mutualistic association that contributes to vertebrate health occurs when animals remove external parasites from the skins of other animals. Several species of birds remove arthropod parasites from mammals and even from reptiles (see figure). the birds get to eat the parasites, and the larger animals are relieved from those parasites. cleaning mutualisms also occur in the ocean.
TErmITE aNd aNT GardENStermites cannot digest wood. this is odd because termites spend all of their time eating wood. instead, they depend on microbes and fungi to do it for them. Some termites have microbes in their intes-tines. Many termites, however, build collections of gigantic mounds that rise like Stonehenges from the african savanna. the termites do not live and breed in the mounds; they live underground. instead the mounds are their flow-through composters. Worker termites bring a chewed wood slurry to the top of the mound, where the fungi begin to digest it. the termites ingest the product at the bottom, and then
A bird picks a parasite off a giant Galápagos tortoise. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 379 11/17/11 1:13 PM

first entry380 mutualism 380
their gut microbes proceed to digest it further. the wind ventilates the mounds, promoting the metabo-lism of the fungi. the termites even maintain a physiological homeostasis within their fungus gar-dens. the more metabolically active mounds require more oxygen, and they get it, because they are taller and protrude farther from the boundary layer of the ground. if the worker termites sense that carbon dioxide levels are too high within the mound, they will chew open more ventilation holes.
leaf-cutter ants (genus Atta) cannot digest leaves. this is odd because they spend most of their time gnawing leaves off of tropical trees and carry-ing them down into their nests. Some of the ant colo-nies are gigantic and can strip an entire tree of its leaves in a few hours. they chew up the toxic leaves, making a compost that feeds a fungus. the fungus consumes the compost but is not itself toxic. the ants eat structures produced by the fungus. When a queen flies off to form a new colony, she takes some of the fungus with her. Some of the workers maintain colonies of actinobacteria on their bodies. these bacteria produce antibiotics that suppress the growth of pathogenic fungi in the fungus gardens. a species of yeast also eats the bacteria. the result, as demonstrated by evolutionary biologist cameron currie, is a five-way symbiosis: ant, food fungus, parasitic fungus, actinobacteria, and yeast.
Wood and leaves may be a good source of car-bohydrates, thanks to fungi, but they are not good sources of nitrogen, which termites and ants, like all animals, need for the production of proteins. Many termites and leaf-cutter ants have nitrogen-fixing bacteria in their digestive systems, which convert atmospheric nitrogen gas into forms that the insects can use.
in the tropical seasonal forests, many acacia trees (genus Acacia) are infested with Pseudomyr-mex ants. the acacia tree has structures that feed
the ants: nectaries that produce nectar and clusters of protein-rich food at the tips of immature leaflets. the ants chew holes in immature thorns and hol-low them out (see photograph). the ants, including the queen, live inside of the thorns. in turn, the ants defend the acacia trees from herbivores and competi-tors. if a herbivorous insect or mammal begins to eat the foliage, the ants swarm and attack. if a vine begins to grow on the acacias, the ants chew it off. By eating the insects that land on the leaves, the ants are not merely defending the tree but are obtaining an extra source of food. the acacia trees remain green well into the dry season, when most other trees have lost their leaves. this makes the acacia trees a very visible target to herbivores, from which the ants protect them. a similar association is found in Old World tropics between ants of the genus Cre-matogaster and trees of the genus Macaranga.
agriculture is not merely a mutualism between a food species and an animal species that promotes its growth. it is an obligate mutualism. the termite and leaf-cutter ant gardens and the acacia trees may therefore qualify as agriculture. the termites, leaf-cutter ants, and Pseudomyrmex ants have no other food, and the fungi in their gardens live nowhere else. Without their ants, the acacia trees are quickly covered with vines.
Some ants appear to be ranchers rather than farmers. Phloem sap in plants is sweet and is not toxic. aphids have small mouthparts that they can insert into a single phloem cell. aphids can therefore tap into pressurized sap. the pressure pushes more sap through the aphids than they can consume. ants cannot tap into the phloem, but they can eat the leftover sap from the aphids. they do more than this, however. they protect the aphids, as a rancher would protect milk cows, from predators.
oCEaN muTualISmSMutualisms are as ubiquitous in the oceans as on land. Here are some examples:
● coral reef fish line up at cleaning stations, where small fish crawl inside their mouths and remove parasites. cleaner fishes of the species Labroides dimidiatus can clean 2,300 individuals of 132 fish species each day.
● Healthy embryos of the shrimp Palaemon macrocadtylus are covered with Alteromo-nas bacteria. the bacteria produce com-pounds that defend them against the fungus Lagenidium callinectes. if the bacteria are experimentally removed, the fungus kills the shrimp embryos. the antibacterial com-pound also works by itself in experimental treatments.
Pseudomyrmex spinicola ants live in the thorns of acacia trees in tropical seasonal forests. (Alex Wild)
xviii+598_Biodiversity_F.indd 380 11/17/11 1:13 PM

mutualism 381
● Up to 40 percent of a sponge’s weight is symbiotic bacteria.
● Fish guts, like those of marine iguanas, con-tain microbes that help them to digest algae.
● the crab Trapezia ferruginea and the shrimp Alpheus lottini protect reef corals from predation by the starfish Acanthaster planci. in return, the crab and the shrimp not only live safely in the crevices of the reef but they feed on mucus produced by the corals.
EColoGICal rulES of muTualISmMutualism presents many interesting stories, but these stories add up to a tremendous impact upon the ecosystems of the world. One of the most com-mon examples of mutualism is the lichen. a lichen is a sandwich of single-celled algae inside of layers of fungus. the algae produce food for themselves and the fungus through photosynthesis, while the fungus appears to protect the algae from light damage when they are dormant. is a lichen one organism or two? Biologists have decided not to decide. they have assigned species names to the lichens, as well as to the fungal and algal symbionts.
lichens are able to grow in places where few other organisms can survive. For example, they grow on tree trunks in deciduous and coniferous forests and on rocks in grasslands. they are also the major ground cover in some environments such as the dry, northern edges of the tundra. in such places they are the major biological components of food production and the cycling of nutrients. “Reindeer moss” is actually a lichen (Cladonia rangiferina) and an important food source for reindeer. lichens, which are very sensitive to toxic compounds, are also important indicators of air pollution. lichens are therefore much more than an interesting story of symbiosis, but play a major role in some ecosystems.
Mycorrhizae are fungi, of many different spe-cies, that live in plant roots. the plant feeds them with sugars from the root storage tissues. in return, the fungi extend their strands out into the soil, func-tioning as an extra set of tiny roots. they are espe-cially effective at absorbing phosphorus from the soil, which then diffuses into the plant root. the mutualists are not deliberately helping one another out. the fungus obtains the phosphorus for its own use and cannot prevent the phosphorus from dif-fusing into the root. the ecological importance of mycorrhizae is difficult to overestimate. Many plant species rely on them to enhance their nutrient uptake and in some cases could not survive in poor soil without them. Many of the nutrients that pass through the food web begin as soil ions absorbed by mycorrhizae.
Mutualisms are also important in the food web of the oceans. cells of the diatom Hemiaulus haukii contain a photosynthetic bacterium, Richelia. the diatom carries out its own photosynthesis and does not need Richelia for this reason. However, like other cyanobacteria, Richelia can fix atmospheric nitrogen gas into a form that the diatom can use as fertilizer. in the portions of the ocean where this diatom is dominant, the mutualism stimulates growth by a factor of three and contributes 25 percent of the nitrogen in the food web. “to visual-ize the magnitude of this nitrogen addition,” says ecologist Mark Hay, “think of it as the mass of 1.5 million Harley-Davidson road king motorcycles, 7.3 million undergraduate students, or 157 million lap-top computers.”
it is possible to have mutualisms within mutu-alisms. Myxotricha paradoxa is a protist that lives in termite guts and allows termites to digest wood. Myxotricha appears to have cilia that allow it to swim. What appear to be cilia, however, are mutual-istic bacteria embedded in the protist’s surface.
CoNSErvaTIoN of muTualISmin order to protect the biodiversity of mutualists, it is necessary to protect the other species that interact with them. Here is one example. When there are no giraffe or elephant grazers, the acacia trees stop producing nectar, protein, and large thorns. Unless there are grazers to threaten the acacias, the mutual-ism begins to break down.
it is also necessary to protect the habitats in which the mutualists live. if the habitat is suitable for one of the mutualists but not the other, both are doomed. this may be the case with cacti that sur-vive but cannot reproduce without their mutualistic pollinators.
muTualISm aNd humaNSaside from the previous examples in which mutual-ism preserves human health, there are at least two major types of mutualism that humans have created. the first is agriculture. Humans have domesticated crop plants and livestock animals. Humans use them as food, but the crops and livestock also benefit, because humans plant, feed, and protect them. as science writer Michael Pollan points out, maize is one of the most successful plants in the world, and its success is due to the fact that humans have destroyed large areas of forest and prairie land and planted a gigantic lawn of corn. Moreover, as in the other examples of agriculture noted previously, humans and agricultural species have an obligatory mutual dependence. agriculture is one of the main ways in which humans have transformed almost a third of the surface of the earth into managed habitats.
xviii+598_Biodiversity_F.indd 381 11/17/11 1:13 PM

first entry382 mutualism 382
the second way is by having pets. Humans keep billions of pet animals. the food that pets eat and the wastes that they produce have a major impact on the ecosystems of the earth. Pets can be considered mutualists, because their companionship (and, in some cases, the protection that they offer) is a major psychological resource for humans.
See also agriculture; biogeochemical cycles; coevolution; herbivory; lichens; pollination.
Further readingarnold, a., et al. “Fungal endophytes limit pathogen dam-
age in a tropical tree.” Proceedings of the National Academy of Sciences USA 100 (2003): 15,649–15,654.
Banning, Jenifer l., et al. “antifungal skin bacteria, embry-onic survival, and communal nesting in four-toed sala-manders, Hemidactylium scutatum.” Oecologia 156 (2008): 423–439.
Bascompte, Jordi, and Pedro Jordano. “Plant-animal mutu-alistic networks: the architecture of biodiversity.” Annual Review of Ecology, Evolution, and Systemat-ics 38 (2007): 567–593.
Batra, l. R., ed. Insect-Fungus Symbiosis: Mutualism and Commensalism. new york: John Wiley, 1979.
Becker, Matthew H., et al. “the bacterially produced metabolite violacein is associated with survival of amphibians infected with a lethal fungus.” Applied and Environmental Microbiology 75 (2009): 6,635–6,638.
Bshary, R., and D. Schäffer. “choosy reef fish select cleaner fish that provide high-quality service.” Animal Behav-iour 63 (2002): 557–564.
clement, lars W., et al. “Strategies of a parasite of the ant-acacia mutualism.” Behavioral Ecology and Sociobiol-ogy 62 (2008): 953–962.
comstock, l., and M. coyne. “Bacteriodes thetaiotaomi-cron: a dynamic, niche-adapted human symbiont.” Bioessays 25 (2003): 926–929.
Dangerfield, J. M., et al. “the mound-building termite Macrotermes michaelseni as an ecosystem engineer.” Journal of Tropical Ecology 14 (1998): 507–520.
eckburg, Paul B., et al. “Diversity of the human intestinal microbial flora.” Science 308 (2005): 1,635–1,638.
Fiala, B., et al. “Studies of a South east asian ant–plant association: protection of Macaranga trees by Crema-togaster borneensis.” Oecologia 79 (1989): 463–470.
Fischer, R. c., et al. “Plants feed ants: food bodies of myr-mecophytic and their significance for the interaction with Pheidole bicornis ants.” Oecologia 133 (2002): 186–192.
Gaume, l., et al. “ant-plant conflicts and a novel case of castration parasitism in a myrmecophyte.” Evolution-ary and Ecological Research 7 (2005): 435–452.
Grice, elizabeth a., et al. “topographical and temporal diversity of the human skin microbiome.” Science 324
(2009): 1,190–1,192. Summarized by Marx, christo-pher J. “Getting in touch with your friends.” Science 324 (2009): 1,150–1,151.
Hay, Mark e., et al. “Mutualisms and aquatic community structure: the enemy of my enemy is my friend.” Annual Review of Ecology, Evolution, and Systemat-ics 35 (2004): 175–198.
Heil, Martin, and Doyle McKey. “Protective ant-plant interactions as model systems in ecological and evolu-tionary research.” Annual Review of Ecology, Evolu-tion, and Systematics 34 (2003): 425–454.
Hill, M. “intestinal flora and endogenous vitamin synthe-sis.” European Journal of Cancer Prevention 6 (1997): S43–S45.
Hongō, yuichi, et al. “Genome of an endosymbiont cou-pling n2 fixation to cellulolysis within protist cells in termite gut.” Science 322 (2008): 1,108–1,109.
izzo, t. J., and H. l. Vasconcelos. “cheating the cheater: domatia loss minimizes the effects of ant castration in an amazonian ant-plant.” Oecologia 133 (2002): 200–205.
Jaenike, John, et al. “adaptation via symbiosis: Recent spread of a Drosophila defensive symbiont.” Science 329 (2010): 212–215.
Janzen, Daniel H. “coevolution of mutualism between ants and acacias in central america.” Evolution 20 (1966): 249–275.
Oliver, Kerry M., et al. “Bacteriophages encode factors required for protection in a symbiotic mutualism.” Sci-ence 325 (2009): 992–994.
Palmer, todd M., et al. “Breakdown of an ant-plant mutu-alism follows the loss of large herbivores from an african savanna.” Science 319 (2008): 192–195. Sum-marized: leslie, Mitch. “the importance of being eaten.” Science 319 (2008): 146–147.
Pinto-tomás, adrián a., et al. “Symbiotic nitrogen fixation in the fungus gardens of leaf-cutter ants.” Science 326 (2009): 1,120–1,123.
Raine, n. e., et al. “Guards and thieves: antagonistic inter-actions between two ant species coexisting on the same ant-plant.” Ecological Entomology 29 (2004): 345–352.
Roth, R. R., and W. D. James. “Microbial ecology of the skin.” Annual Reviews of Microbiology 42 (1988): 441–464.
Scott, Jarrod J., et al. “Bacterial protection of beetle-fun-gus mutualism.” Science 322 (2008): 63. Reviewed in: Berenbaum, May R., and thomas eisner. “Bugs’ bugs.” Science 322 (2008): 52–53.
Stadler, Bernhard, and anthony F. G. Dixon. “ecology and evolution of aphid-ant interactions.” Annual Review of Ecology, Evolution, and Systematics 36 (2005): 345–372.
turner, J. Scott. “On the mound of Macrotermes michaelseni as an organ of respiratory gas exchange.”
xviii+598_Biodiversity_F.indd 382 11/17/11 1:13 PM

mycorrhizae 383
Physiological and Biochemical Zoology 74 (2001): 798–822.
Usher, Kayley M., et al. “exploring cyanobacterial mutu-alisms.” Annual Review of Ecology, Evolution, and Systematics 38 (2007): 255–273.
Van der Heijden, M. G. a., et al. “Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability, and productivity.” Nature 396 (1998): 69–72.
Woodhams, Douglas c., et al. “Symbiotic bacteria con-tribute to innate immune defenses of the threatened mountain yellow-legged frog, Rana muscosa.” Bio-logical Conservation 138 (2007): 390–398.
Xu, J., and J. i. Gordon. “Honor thy symbionts.” Proceed-ings of the National Academy of Sciences USA 100 (2003): 10,452–10,459.
youngsteadt, elsa. “all that makes fungus gardens grow.” Science 320 (2008): 1,006–1,007.
yu, D. W. “Parasites of mutualisms.” Biological Journal of the Linnean Society 72 (2001): 529–546.
———, and n. e. Pierce. “a castration parasite of an ant-plant mutualism.” Proceedings of the Royal Society of London B (Biology) 265 (1998): 375–382.
mycorrhizae See mutualism.
xviii+598_Biodiversity_F.indd 383 11/17/11 1:13 PM

384
natural selection natural selection is the process by which evolution produces adaptation. Other processes, such as genetic drift, can cause evolution to occur, but in a random direction. Only natural selection guides the evolutionary process in a direction that produces adaptation to environmental and social conditions. evolution is the source of all biodiversity.
natural selection is the process that was fi rst clearly explained by charles Darwin, although some earlier writers such as W. c. Wells, edward Blyth, and Patrick Mathew had presented fragments of the idea. natural selection was independently discov-ered by alfred Russel Wallace.
Darwin not only demonstrated that evolution had occurred but also explained how it occurred. as a result, evolution became believable for the fi rst time. Darwin’s contemporaries largely accepted his demonstration that evolution had occurred, but the scientifi c community did not embrace natural selec-tion for another 80 years.
Sexual selection, which was also fi rst elucidated by charles Darwin, operates in a manner similar to natural selection. However, sexual selection usually produces arbitrary and sometimes outlandish char-acteristics that are not usually considered adaptation.
Both Darwin and Wallace alluded to artifi cial selection, the process by which humans breed crops and livestock in desired directions, in crafting the phrase natural selection. in both cases, some indi-viduals are selected to propagate and others are not; in natural selection, it is nature rather than humans that does the selecting. Both artifi cial and natu-ral selection can produce genetic changes within populations and can cause divergence of popula-tions into different varieties. either could eventu-
ally cause varieties to be different enough that they would constitute different species. artifi cial selec-tion, however, has not gone on long enough for this to happen.
natural selection occurs within populations. individual organisms cannot evolve because they cannot undergo whole-body genetic changes. if mutations occur in somatic cells, which make up most of the body of an animal, the cells that contain the mutations will die when the organism dies. Only those mutations that occur in germline cells, either eggs or sperm or the cells that produce them, can be passed on to the next generation. therefore the genetic effects of evolution can only be observed in the next generation of the population.
natural selection occurs because individuals within populations differ from one another in how successfully they reproduce. evolutionary theories before Darwin, such as lamarckism, were incor-rectly based upon the transformation of an entire population or species, and/or as a result of changes within the individuals themselves. But Darwin’s insight was that, because some individuals in the population reproduce more successfully than oth-ers, over time the population begins to resemble the successful individuals. Biologist ernst Mayr said that by formulating the theory of natural selection, Darwin introduced population thinking into evolu-tion, to replace typological thinking—that is, evo-lution occurs in populations of diverse individuals, rather than being the transformation of one type into another.
proCESS of NaTural SElECTIoNernst Mayr summarized the process of natural selec-tion as fi ve facts and three inferences that can be
N
xviii+598_Biodiversity_F.indd 384 11/17/11 1:13 PM

natural selection 385
derived from those facts. all of these facts are part of the science of ecology, the study of the relationship between organisms and their living and nonliving environments. therefore by elucidating the process of natural selection, Darwin was not only mak-ing evolution believable for the first time, but also inventing the science of ecology while he was at it.
Fact 1. Populations can grow exponentially. Because reproductive groups of individuals can mul-tiply, populations grow in curves, not in lines. in a population of bacteria, one bacterium can produce two, but 1 million bacteria can produce 2 million bacteria during cell division. this allows popula-tions to grow explosively. this will occur, of course, only in an unlimited environment. as Darwin wrote in The Origin of Species:
there is no exception to the rule that every organic being naturally increases at so high a rate, that, if not destroyed, the earth would be covered by the progeny of a single pair. even slow-breeding man has doubled in twenty-five years, and at this rate, in less than a thousand years, there would literally not be standing-room for his progeny. linnaeus has calculated that if an annual plant produced only two seeds—and there is no plant so unpro-ductive as this—and their seedlings next year pro-duced two, and so on, then in twenty years there should be a million plants.
Darwin did some calculations for the Origin of Species (incorrectly, as it turned out) on the repro-ductive rate and population growth of elephants in an unlimited environment:
the elephant is reckoned the slowest breeder of all known animals, and i have taken some pains to estimate its probable minimum rate of natu-ral increase; it will be safest to assume that it begins breeding when thirty years old, and goes on breeding till ninety years old, bringing forth six young in the interval, and surviving till one hundred years old; if this be so, after a period of from 740 to 750 years there would be nearly nineteen million elephants alive, descended from the first pair.
Fact 2. Populations sometimes grow, and sometimes become smaller, but tend to remain sta-ble. therefore something must be preventing popula-tion growth, either by repressing reproduction or by causing death. Darwin wrote:
lighten any check, mitigate the destruction ever so little, and the number of the species will almost instantaneously increase to any amount.
Darwin insisted on evidence and on experimen-tation where possible. He dug and cleared a small plot of ground at his estate and laboriously marked each seedling that germinated. the total was 357. Of these, 295 seedlings died, mainly because slugs and insects ate them. He also counted the number of bird nests on his property in the spring of 1854 and again the spring of 1855 and estimated that during the intervening winter nearly four-fifths of the birds had died.
Fact 3. Resources are limited. Ultimately, if reproduction continued unabated, the world could not contain the offspring. However, resources are limited by much more than simply room in the world. Organisms need resources and ecosystem services that provide food, water, shelter, oxygen, and process their wastes. each kind of organism is adapted to live in only a limited range of conditions; therefore, for a polar bear, it does not much matter how big the world is, but only how big the arctic is, and for species that are limited to a single island or mountain or swamp, the world is pretty small. the disparity between limited resources and exponential popula-tion growth in human societies had been famously pointed out by economist thomas Robert Malthus, and Darwin simply applied it to nature: “it is the doctrine of Malthus applied with manifold force to the whole animal and vegetable kingdoms . . .”
these three facts lead to inference 1, which is the struggle for existence. in nearly all natural populations, there are not enough resources for all of the individuals to have all of the resources they need. today scientists say that competition occurs in nearly all natural populations nearly all of the time. Sometimes competition is strong enough that scien-tists can actually observe the struggle. in a jungle, vines grow over trees, and trees crowd together, as if they are fighting for access to the light. in a desert, the bushes are evenly spaced across the landscape, because there is not enough water for any bush to grow close to another bush. Predators fight over a kill, and birds chase one another out of the choice territories. But even when it cannot be seen, competi-tion can be inferred. competition is the unavoidable result of exponential population growth and limited resources. as Darwin wrote:
nothing is easier than to admit in words the truth of the universal struggle for life, or more diffi-cult—at least i have found it so—than constantly to bear this conclusion in mind . . . We behold the face of nature bright with gladness, we often see superabundance of food; we do not see or we forget, that the birds that are idly singing round us live on insects or seeds, and are thus con-stantly destroying life; or we forget how largely
xviii+598_Biodiversity_F.indd 385 11/17/11 1:13 PM

first entry386 natural selection 386
these songsters, or their eggs, or their nestlings, are destroyed by birds or beasts of prey; we do not always bear in mind, that, though food may now be superabundant, it is not so at all seasons of each recurring year.
Darwin’s theory of evolution by means of nat-ural selection severely shook the complacency of european science, which had comfortably assumed the divine creation and long-term stability of spe-cies. His invention of ecology had at least as great an effect on scientific and popular thought. Many people thought of the natural world as God’s vast and orderly garden, full of life. Darwin showed the natural world to be violent and full of death. to his readers, Darwin had shaken their view of the world—and they had not even gotten past chapter 3 (the Struggle for existence) of his book, On the Origin of Species! the poet alfred, lord tennyson, reflected the sadness not only of the death of indi-viduals but the extinction of species in his poem In Memoriam, which is the origin of the famous phrase “red in tooth and claw.” (tennyson published the poem before Darwin wrote his book.) the fact of extinction may have forced scholars to think about the fact that disease and death—often the result of the struggle for existence—were much more painful than they needed to be in a perfectly ordered world. Darwin experienced this firsthand through the death of his mother when he was young. even without evolution, religious scholars had, and still have, to struggle with the “problem of natural evil.”
Fact 4. each individual in a population is unique. each population contains a tremendous amount of diversity. Humans can easily observe this diversity in their own populations, but it is true of the populations of all other species as well, even if humans cannot recognize it. Darwin learned this not from other scientists, but from animal breed-ers, particularly pigeon fanciers. all the different breeds of pigeon had been selected from ancestral stocks of rock pigeons (Columba livia). these breeds included some with outlandish feathers, anatomy (for instance, the ability to inflate their crops mas-sively), or behavior (such as the ability to tumble during flight). Since all of these breeds had come from rock doves, then all of their characteristics had to be present, but hidden, within the rock dove pop-ulations. Breeding brought out these traits, distilling them as one might distill brandy from wine. the first two chapters of the Origin described how artificial selection and the observation of natural populations showed the tremendous amount of diversity that they contained.
Fact 5. Much of the variability in populations is heritable. Darwin knew nothing about genetics.
He was apparently unaware of the writings of Gregor Mendel, which explained how traits could be passed from one generation to another. But every plant and animal breeder knew that this was the case. there was confusion about the mechanism. Many scientists held theories about varieties of lamarckism, or the inheritance of acquired characteristics. Many also believed in “blending inheritance” in which parental traits blended permanently in their offspring, like blending different colors of paint. Mendel, in con-trast, was one of the researchers who demonstrated that traits were inherited in a particulate rather than a blending fashion; that is, a trait could disappear in one generation and reappear in another, like mix-ing different colors of marbles rather than of paint. Darwin himself ended up accepting a variation of lamarckism, which he called pangenesis, a theory which most evolutionary scientists today consider to be his only major blunder.
the first inference, plus facts 4 and 5, give rise to inference 2, which is natural selection itself. in order to reproduce, an individual needs to obtain resources beyond just what is needed for survival. Because of the struggle for existence (inference 1), not all indi-viduals in the population can reproduce or even sur-vive. if all individuals in the population were alike, then the ones that prevailed would be due either to chance or to historical contingency, which might just be another form of chance. But since individuals are unique, then some of the individuals have what it takes to survive and reproduce more than others. For example, in a moist environment, some of the plants may have larger leaves and shallower roots than oth-ers. the plants with larger leaves could grow faster because they would have greater photosynthesis, and they would not waste their resources producing deep roots which are not needed in a moist environment. But if the trait cannot be passed on to the next gen-eration, the superior survival and reproduction of the plants with larger leaves would not matter, since their offspring would not have leaves any larger than those of other plants.
natural selection is the differential reproduction of organisms in a population because of their genes. that is all that it is. if some genetically distinct individuals consistently reproduce more than oth-ers, natural selection occurs. Scientists may be able to determine what caused the superiority of some individuals over others, as in the case of Darwin’s finches, or they may not, as in the famous British study of the peppered moths. either way, natural selection has occurred. as Darwin described it,
Owing to [the] struggle [for existence], varia-tions, however slight and from whatever cause proceeding, if they be in any degree profitable
xviii+598_Biodiversity_F.indd 386 11/17/11 1:13 PM

natural selection 387
to the individuals of a species, in their infinitely complex relations to other organic beings and to their physical conditions of life, will tend to the preservation of such individuals, and will generally be inherited by the offspring . . . i have called this principle, by which each slight varia-tion, if useful, is preserved, by the term natural Selection . . .
inference 2 leads directly to inference 3, which is that if natural selection continues long enough, evolutionary change results in the population.
the traits that confer superiority in natural selection, therefore in evolution, are not the same everywhere. there are countless pathways to suc-cess, in the natural world as in the human economy. the above example described successful plants in moist environments as having large leaves and shal-low roots. in an arid environment successful plants would have small leaves (which waste less water than large leaves) and deep roots (that may be able to tap into new sources of water). in most cases, evolution can go in more than one direction, depending on the circumstances. to use one example close to home, the major pattern of evolution in the human lineage has been the increase in brain size, but in at least one case (the Flores island people, Homo floresiensis), human brain size decreased during evolution.
TypES of NaTural SElECTIoNevolutionary scientists generally classify natural selection into three types (see figure on page 388):
1. in stabilizing selection, the average indi-viduals reproduce better than the extreme individu-als. For example, in an environment with medium humidity, very large-leaved and very small-leaved individuals in a plant population would grow more slowly and reproduce less. Stabilizing selection reinforces the mean or average trait value by elimi-nating outlying variation. Stabilizing selection may be common in populations once a particularly suc-cessful combination of genes has been assembled. according to some evolutionary scientists, stabi-lizing selection has been the experience of nearly every species during most of its existence, result-ing in long periods of evolutionary stasis between speciation events, a pattern known as punctuated equilibria.
2. in directional selection, individuals at one of the extremes reproduce better than either the average individuals or the individuals at the other extreme. if the environment of the population of plants described above became wetter, the larger-leaved individuals would grow and reproduce better than the individuals with either medium or small leaves. a drier environment would favor the indi-
viduals with small leaves over all the others. either of these would be directional selection.
3. in diversifying selection, the average individu-als are at a disadvantage and the extreme individu-als reproduce the most. During intense competition, the average individuals may compete most strongly. individuals that are very small or very large might escape direct competition. as a result, the popula-tion may begin evolving in two directions at once. this process cannot continue as long as the individu-als can all interbreed. if a reproductive barrier arises between the small and large individuals, they may begin to diversify into two species. Darwin neatly explained how a population could evolve, perhaps eventually becoming a new species. natural selec-tion does not itself explain how one species can evolve into two or more species. Reproductive isola-tion must also occur.
Frequency-dependent selection is a special instance in which an individual has greater repro-ductive success not because of better adaptation to environmental conditions but simply from being rare. the most commonly cited example of fre-quency-dependent selection is that of sex ratios. if males are rare in a population, the average male will leave more offspring than the average female. con-versely, rare females will leave more offspring, on the average, than common males. Whichever sex is rare enjoys greater reproductive success. Frequency-dependent selection is considered to be the main process by which most animal populations maintain an equal mix of males and females.
lImITaTIoNS of NaTural SElECTIoNnatural selection is not all-powerful. Below are examples of limitations upon the power of natural selection:
● Physical constraints. natural selection cannot cause the evolution of characteristics that violate the laws of physics and chemis-try. For example, natural selection cannot produce a large animal with skinny legs. also, the tallest trees in the world are about 350 feet (about 70 m) tall. trees much taller than this are not possible, because there is a maximum height to which wood can con-duct water, and the tallest redwoods may be very close to this maximum height. large animals with wheels cannot evolve, for the wheel could not receive nutrition from the rest of the body through the axle.
● Phyletic constraints. Once an evolution-ary lineage has adopted a certain adapta-tion, this adaptation itself puts constraints on evolutionary change. it is not possible
xviii+598_Biodiversity_F.indd 387 11/17/11 1:13 PM
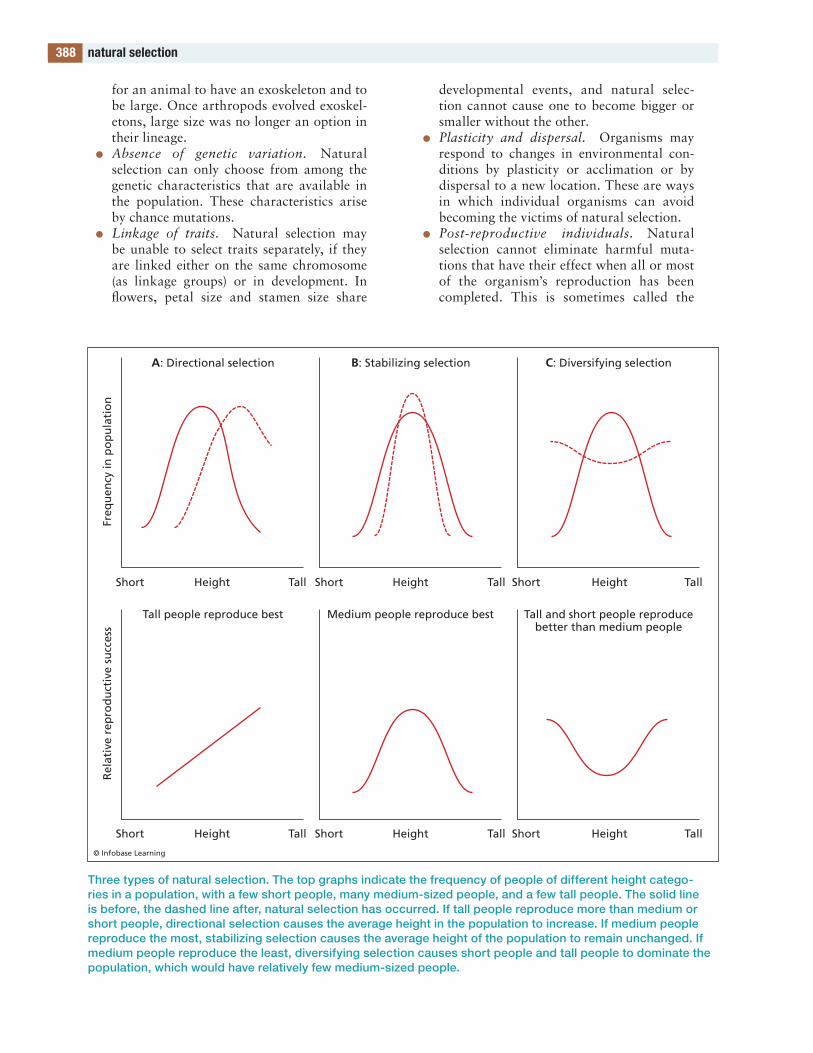
first entry388 natural selection 388
for an animal to have an exoskeleton and to be large. Once arthropods evolved exoskel-etons, large size was no longer an option in their lineage.
● Absence of genetic variation. natural selection can only choose from among the genetic characteristics that are available in the population. these characteristics arise by chance mutations.
● Linkage of traits. natural selection may be unable to select traits separately, if they are linked either on the same chromosome (as linkage groups) or in development. in flowers, petal size and stamen size share
developmental events, and natural selec-tion cannot cause one to become bigger or smaller without the other.
● Plasticity and dispersal. Organisms may respond to changes in environmental con-ditions by plasticity or acclimation or by dispersal to a new location. these are ways in which individual organisms can avoid becoming the victims of natural selection.
● Post-reproductive individuals. natural selection cannot eliminate harmful muta-tions that have their effect when all or most of the organism’s reproduction has been completed. this is sometimes called the
Three types of natural selection. The top graphs indicate the frequency of people of different height catego-ries in a population, with a few short people, many medium-sized people, and a few tall people. The solid line is before, the dashed line after, natural selection has occurred. If tall people reproduce more than medium or short people, directional selection causes the average height in the population to increase. If medium people reproduce the most, stabilizing selection causes the average height of the population to remain unchanged. If medium people reproduce the least, diversifying selection causes short people and tall people to dominate the population, which would have relatively few medium-sized people.
xviii+598_Biodiversity_F.indd 388 11/17/11 1:13 PM

natural selection 389
“kingpin principle,” based on a story about automobile manufacturer Henry Ford. He had his engineers study old Ford cars and find which part of the old cars was in the best condition. they reported that the king-pins were just like new. Ford told his factory managers to stop wasting their time making such good kingpins. there was no need for kingpins to last forever in a car that did not. Similarly, in organisms, there is no advan-tage for genes that promote the health and vigor of post-reproductive individuals.
● The future. natural selection cannot anticipate the future and prepare for it.
● Group selection. natural selection cannot favor characteristics that benefit the group but reduce the fitness of the individual that possesses them. Some adaptations result in harm for the individual (for example, worker bees that die after stinging a vic-tim, and mother spiders that die when her spiderlings eat her) but these are compen-sated by an increase in inclusive fitness, an example of altruism. When pathogens infect a host, natural selection may favor those selfish lineages of pathogens that reproduce most rapidly, even if the host dies as a result. natural selection may work against such selfish pathogens if the death of the host results in the death of the entire pathogen population or if the illness of the host causes other hosts to stay away from the sick individual, thus preventing the dis-persal of any pathogens of that population to new hosts. But natural selection cannot result in individuals sacrificing their own fitness within a population.
NaTural SElECTIoN oBSErvEd IN NaTurEBelow are just a few examples of natural selection that have been observed in nature.
Stabilizing selection. evolutionary biolo-gists arthur Weis and Warren abrahamson studied insects that lay their eggs inside of plant stems. the grub releases a compound that causes the stem to produce a round swelling called a gall. the grub can develop, well fed, inside this gall. there are dangers from predators, however. Gall wasps can recognize galls and pierce their ovipositors into the gall, laying their eggs on the grub, and the wasp larvae eat the grub. Birds can also recognize galls and tear them open, to eat the protein-rich grub. Gall wasps prefer smaller galls; wasp parasitism therefore selects for larger galls. Birds prefer larger galls; bird predation therefore selects for smaller galls. the result is what looks like stabilizing selection for medium-sized
galls but is actually a compromise of two processes of directional selection.
Directional selection. Soapberry bugs (Jadera haematoloma) pierce fruits of certain plants with their long beaks. they are native to the keys of Florida, where they obtained their food from native balloon vines. after 1925, landscapers introduced the golden rain tree, an asian relative of the balloon vine but with smaller fruits, into central Florida. Soapberry bugs began to use the new trees as a food source. in central Florida, natural selection favored bugs with shorter beaks. Over the next few decades, the central Florida bug populations evolved shorter beaks, while the bugs in the keys retained their long beaks. evolutionary scientists Scott carroll and christin Boyd documented this change by examin-ing bug collections that had been made before and after the introduction of golden rain trees to central Florida.
Diversifying selection. in populations of afri-can black-bellied seedcrackers (a type of bird), indi-viduals have either large or small beaks. these two beak sizes correspond to two different kinds of seeds (large v. small) that the birds eat. Juvenile birds have greater variation in beak size than adults. evolutionary biologist thomas Bates Smith found that the juveniles that survived were those that specialized most narrowly upon the available seed sizes. among the large-beaked individuals, those with beaks too large or too small died, and among the small-beaked individuals, those with beaks too large or too small also died. Diversifying selection thus reinforced the large and the small beak sizes in this bird population.
NaTural SElECTIoN ExpErImENTSOccasionally, natural selection can be verified in natural populations by manipulative experiments, as in these examples:
Bacteria. artificial selection of bacteria has produced strains of bacteria able to tolerate condi-tions very different from those to which the original bacterium was adapted.
Anole lizards. anoles are lizards that live in the caribbean. Some very small caribbean islands have no natural populations of anoles. evolutionary biologist Jonathan losos introduced anole lizards onto some of these islands. the islands differed in the relative amount of plant cover. Within 10 to 14 years, recognizably different forms of anole had apparently evolved on different islands.
Guppies. Guppies live in freshwater pools on the island of trinidad. in some pools, there are rela-tively large predators that eat guppies of all sizes; in other pools, there are only small predators, which eat only small guppies. the guppies that live with
xviii+598_Biodiversity_F.indd 389 11/17/11 1:13 PM

first entry390 natural systems agriculture 390
large predators have a more rapid life cycle, growing faster and reaching reproductive maturity sooner, than those that live with small predators. evolution-ary biologist David Reznick found some pools that had small predators, no large predators, and also no guppies. this was the perfect opportunity for an experiment. He predicted that if fast-growing gup-pies that had previously experienced large predators were exposed to this new environment, they would evolve into slower-growing forms. He performed the experiment and obtained the expected results.
ImporTaNCE of NaTural SElECTIoN To all of SoCIETy
evolution in general, and natural selection in par-ticular, has been a subject of little interest to many students (and professors) in the health sciences and agriculture. Recent developments, however, have shown that it was disastrous for medical and agri-cultural professionals to ignore the process of natu-ral selection. in particular, natural selection has led directly to the evolution of antibiotic-resistant bacte-ria. natural selection in pathogenic bacteria is fast-paced, outrunning the ability of the medical industry to produce new antibiotics; it costs a lot of money and it kills thousands of people. the evolution of pesticide resistance in mosquitoes, flies, ticks, lice, and other disease vectors presents yet another public health menace. Quite literally, what you do not know about evolution can kill you. natural selection also occurs everywhere and all the time in agricultural fields, as insects evolve resistance to pesticides and weeds evolve resistance to herbicides. the response of many farmers is to spray more, with less success, which results in both the poisoning of the environ-ment and the loss of agricultural production. Medical and agricultural professionals, however, have found some degree of success by using the process of natu-ral selection in their favor rather than letting it work against them. By using antibiotics and pesticides spar-ingly rather than abundantly, and by using mixtures of several types rather than a single type, they have been able to slow down the evolution of resistance. as ecologist Rachel carson indicated, Darwin could not have imagined such a vindication of his theory.
evolutionary biologist Richard Dawkins referred to the “devastating elegance” by which natural selec-tion explains complex and seemingly perfect adapta-tions. the evolution of such adaptations may seem highly improbable, but natural selection breaks the problem of improbability into small pieces. instead of multiplying the improbabilities of evolution, it allows the accumulation of probabilities. Because of natural selection, evolution makes sense.
See also adaptation; Darwin, charles; Dar-win’s finches; ecology; founder effect and
genetic drift; life history, diversity of; sex-ual selection; speciation; Wallace, alfred Russel.
Further readingBalanyá, Joan, et al. “Global genetic change tracks global
climate warming in Drosophila pseudoobscura.” Sci-ence 313 (2006): 1,773–1,775.
Bates Smith, thomas. “Disruptive selection and the genetic basis of bill size polymorphism in the african finch Pyrenestes.” Nature 363 (1993): 618–620.
carroll, Sean P. and christin Boyd. “Host race radiation in the soapberry bug: natural history with the history.” Evolution 46 (1992): 1,052–1,069.
conrod, William, and erica Bree Rosenblum. “a desert Galapagos.” Natural History, May 2008, 15–18.
Jones, Kristina n., and Jennifer S. Reithel. “Pollinator-medi-ated selection on a flower color polymorphism in experi-mental populations of Antirrhinum (Scrophulariaceae).” American Journal of Botany 88 (2001): 447–454.
losos, Jonathan B., K. i. Warheit, thomas W. Schoener. “adaptive differentiation following experimental island colonization in Anolis.” Nature 387 (1997): 70–73.
Mayr, ernst. What Evolution Is. new york: Basic Books, 2001.
Reznick, David n., et al. 1997. “evaluation of the rate of evolution of natural populations of guppies (Poecilia reticulata).” Science 275 (1997): 1,934–1,937.
Sultan, Sonia. “What has survived of Darwin’s theory? Phenotypic plasticity and the neo-Darwinian legacy.” Evolutionary Trends in Plants 6 (1992): 61–71.
turner, Paul e. “cheating viruses and game theory.” Ameri-can Scientist 93 (2005): 428–435.
Weiner, Jonathan. “evolution in action.” Natural History, november 2005, 47–51.
Weis, arthur e. and Warren G. abrahamson. “evolution of host plant manipulation by gall makers: ecological and genetic factors in the Solidago-Eurosta system.” American Naturalist 127 (1986): 681–695.
Zimmer, carl. “Fast-reproducing microbes provide a win-dow on natural selection.” New York Times, June 26, 2007. available online. URl: http://www.nytimes.com/2007/06/26/science/26lab.html. accessed august 8, 2011.
natural systems agriculture natural sys-tems agriculture is an approach to food production that builds upon the principles of biodiversity and ecology, rather than ignoring them. it is the opposite of industrial agriculture.
industrial agriculture, which has predominated food production in industrial countries for more than a century, seeks to eliminate biodiversity and ecology as much as possible from food production. an industrial farm consists, as far as possible, of one
xviii+598_Biodiversity_F.indd 390 11/17/11 1:13 PM

natural systems agriculture 391
species—a single variety of a single crop. all other species are eliminated, as far as possible, by chemi-cal poisons. the soil exists only to temporarily hold onto chemical fertilizers. Soil loss through wind and water erosion is considered an acceptable expendi-ture; the use of more fertilizer can compensate for it. Some kinds of organic agriculture are very similar to industrial agriculture except that toxic chemicals are not used.
it has been increasingly obvious even to indus-trial farmers that industrial agriculture cannot con-tinue very far into the future. it is totally dependent upon fossil fuels, not only to make and apply the chemicals but to process and transport food from huge centralized farms. if soil loss continues, the only alternative will be hydroponic food production, in which food is grown, very expensively, in water, usually in greenhouses. it has also become obvious that the laws of ecology and evolution will not go away just by being ignored. industrial agriculture itself creates perfect conditions for the continued eruption of pest populations and of resistance of those pests to the chemicals intended to control them.
natural systems agriculture was the insight of Wes Jackson, a geneticist who started the land institute in Salina, Kansas, in the 1970s. the insti-tute acquired some farmland and some adjacent intact tallgrass prairie. Jackson and later other sci-entists undertook the task of learning about food production by studying the nearby prairie. the fol-lowing principles emerged, among others:
● the prairie built up soil instead of losing it, for two reasons. First, the deep roots of the grasses and forbs prevented the min-eral components from eroding and con-tinually added humus. Second, there was never any season of the year in which the prairie soil was exposed to erosion. even immediately after a fire, the prairie sod was thick with roots. industrial agriculture, in contrast, was based on plants with shallow roots which lived only for part of a year, leaving bare soil exposed during late fall, winter, and early spring. any agriculture that intends to continue indefinitely into the future must use perennial crops to prevent soil erosion.
● the prairie did not have pest outbreaks in the way that industrial farms did. Prai-ries have few weeds, since the sod is thick. Further, the mixture of species in the prai-rie (a polyculture) prevented any special-ist herbivore populations from building up to excessive levels. industrial agricultural
monocultures encourage pest population explosions. While no farm may withstand a horde of generalist herbivores such as grass-hoppers that can eat anything, a polyculture farm resists specialist herbivores much more readily than a monoculture farm. and while these pests can evolve resistance to chemical pesticides, they cannot evolve resistance to a low density of their food base.
● the prairie does not need inputs of chemical fertilizer. the prairie not only holds onto its soil nutrients better than an industrial farm, but the mixture of species includes legu-minous plants, which have nitrogen-fixing bacteria in their roots.
Research on natural systems agriculture has repeatedly shown that perennial polyculture farms are superior to industrial farms in all of the ways stated above. the big challenge for natural systems agriculture is to develop perennial crops and poly-culture cropping systems that have a consumer mar-ket. there are virtually no wild prairie species that very many people want to eat. the land institute and affiliated researchers at a few universities in var-ious countries are working on hybridization between major crops and their wild perennial relatives (for example, between annual wheat Triticum aestivum and wild wheatgrass Thinopyrum intermedium). the researchers select the hybrids that produce the most marketable seeds and that have perennial traits. Because industrial agriculture receives so much more funding (at public expense) and has so many more researchers compared to natural systems agriculture, a marketable alternative to industrial agriculture cannot be expected to emerge very soon.
Many farmers use one or more parts of natural systems agriculture. Very few farmers (as opposed to orchardists) grow perennial crops, but many of them use conservation tillage, which allows stubble to remain on the surface of the soil after harvest, which reduces soil erosion and encourages the buildup of humus. Many farmers use contour plowing, in which the plowed ridges slow down the movement of water and thus reduces water erosion of soil. Many farmers now plant more than one kind of crop in adjacent strips, a process called strip-cropping, that somewhat mimics natural polycultures. these practices have been partly successful, but still retain enough industrial elements that they cannot make agriculture sustainable in the long run.
See also biodiversity, resources for agri-culture; communities, ecological; grass-lands and savannas; Jackson, Wes; nitrogen cycle; population biology; resistance, evolu-tion of.
xviii+598_Biodiversity_F.indd 391 11/17/11 1:13 PM

first entry392 Neogene period, biodiversity in 392
Further readingcox, t. S., et al. “Prospects for developing perennial
grains.” BioScience 56 (2006): 649–659.Glover, Jerry D., et al. “Future farming: a return to roots?”
Scientific American, august 2007, 82–89.———, et al. “increased food and ecosystem security via
perennial grains.” Science 328 (2010): 1,638–1,639.Scherr, Sara J., and Jeffrey a. Mcneely, eds. The Science
and Practice of Ecoagriculture. Washington, D.c.: island Press, 2007.
neogene period, biodiversity in the neo-gene period (23 to 2.5 million years ago) was the second period of the cenozoic era. it followed the Paleogene period; the Quaternary period followed it. the neogene period is the second part of what has traditionally been called the tertiary period, consisting of the Miocene and Pliocene epochs. Scientists have divided the tertiary into Paleogene and neogene periods largely because the climates of the two periods were different on a worldwide scale (see table).
Climate. though climatic conditions were warmer and wetter than those of today and for-ests remained widespread, the neogene period was cooler and drier than the Paleogene period. ice sheets covered much of antarctica, which was for-ested in the Paleogene. Oxygen and carbon dioxide in the atmosphere were nearly at their modern levels; carbon dioxide was at preindustrial levels, which were lower than today. as more and more water was frozen in glaciers, ocean levels fell. about 5 mil-lion years ago, the Mediterranean Sea dried up and became a salt flat and the climate of much of africa became arid.
Continents. the continents assumed their modern arrangement, with slight exceptions. For example, north and South america were not yet connected by the isthmus of Panama. this slight difference was important, however. Ocean currents flowed between north and South america, allowing warm tropical water to reach the arctic Ocean and prevent extensive ice formation.
Marine life. life in the neogene oceans closely resembled that of modern times (see figure):
● Invertebrates. Ocean invertebrates were very similar to those of today.
● Vertebrates. Bony fishes and sharks con-tinued to diversify in the oceans, as did the toothed whales, baleen whales, porpoises, and dolphins. the largest known shark, Car-charodon megalodon, swam in the neogene oceans. it grew up to 49 feet (15 m) in length.
multiCellular biodiversity in the neogene period
Conditions
Atmospheric oxygen 8% above modern level
Atmospheric carbon dioxide At preindustrial levels (20% below modern level)
Global temperature same as today
Sea level similar to today
Approximate number of families*
Continental 1,300
Marine 1,100
Total 2,400
Examples of marine animals
Carcharodon megalodon shark
Eurhinodelphis longirostris dolphin
Gadella sp. bony fish
Hoploparia gazdzicki lobster
Lepas kuwayamai barnacle
Zygophyseter varolai toothed whale
Examples of continental animals
Australopithecus afarensis bipedal ape
Bison paleosinensis bison
Callicovatellus petrodytes beetle
Capricamelus gettyi camel
Coluber dolnicensis racer snake
Equus simplicidens horse
Gigantocamelus spatulus giraffe
Hipparion forcei horse
Lyssomanes sp. spider
Mertensiella mera salamander
Orrorin tugenensis bipedal ape
Paranthropus robustus bipedal ape
Pliohippus sp. horse
Voltinia dramba butterfly
xviii+598_Biodiversity_F.indd 392 11/17/11 1:13 PM

Neogene period, biodiversity in 393
Continental life. Plants and animals continued to thrive and diversify not only in the extensive forests but also in the world’s fi rst grasslands and deserts:
● Plant life. Many modern genera of trees were in existence in the extensive forests. Species of cactus and grass, as well as many herbaceous wildfl owers, spread in the grasslands and deserts. not only did grasses as a whole expand their range, but in particular the c4 grasses, which have a type of photosynthesis that is particu-larly effective in hot, dry climates. these grasses may also have spread because c4 photosynthesis is effective at low concentrations of carbon dioxide. atmospheric carbon levels were lower during the neogene than they had ever been in the history of the earth.
● Invertebrates. all major groups of terrestrial inverte-brates had evolved before the neogene period.
● Ve r t e b r a t e s . M o d e r n groups of freshwater fi shes, amphibians, and reptiles continued to diversify. Mam-mals and birds were in the evolutionary lineages that are most common today. Horses adapted to the spreading grasslands. Hippa-
rion migrated from north america, where all previous horses had lived, into eurasia, becoming the ancestor of all Old World horse species. like earlier horses, Hip-parion had three toes, but the two outer ones were vestigial and did not touch the ground. Modern humans evolved during this time, fi rst in the dry forests, then in the savannas, of africa.
Further readingarnold, caroline. Giant Shark: Megalodon, Prehistoric
Super Predator. new york: Houghton Miffl in, 2000.Benton, Michael J. “Diversifi cation and extinction in the
history of life.” Science 268 (1995): 52–58.Bianucci, Giovanni, and Walter landini. “Killer sperm
whale: a new basal physeteroid (Mammalia, cetacea) from the late Miocene of italy.” Zoological Journal of the Linnaean Society 148 (2006): 103–131.
Fortelius, Mikael, et al. “late Miocene and Pliocene large land mammals and climatic changes in eurasia.” Pal-aeogeography, Palaeoclimatology, and Palaeoecology238 (2006): 219–227.
Govers, Rob. “choking the Mediterranean to dehydration: the Messinian salinity crisis.” Geology 37 (2009): 167–170.
lindsay, everett H., et al. “Pliocene dispersal of the horse Equus and late cenozoic mammalian dispersal events.” Nature 287 (1980): 135–138.
Osborne, colin P. “atmosphere, ecology, and evolution: What drove the Miocene expansion of c4 grasslands?” Journal of Ecology 96 (2008): 35–45.
Examples of land plants
Acer pohangense maple
Alnus kefersteinii alder
Artemisia sp. sagebrush
Carya protojaponica hickory
Celtis rusingensis hackberry
Distichlis sp. grass
Ginkgo biloba ginkgo
Lophocereus schottii cactus
Pinus griffi thii pine
Quercus lonchitis oak
*Based on Benton
Diprotodon, a mammal of the Neogene period, was the largest marsupial that ever lived.
xviii+598_Biodiversity_F.indd 393 11/17/11 1:13 PM

first entry394 niche 394
Retallack, Gregory J. “Middle Miocene fossil plants from Fort ternan (Kenya) and evolution of african grass-lands.” Paleobiology 18 (1992): 383–400.
———. “neogene expansion of the north american Prai-rie.” Palaios 12 (1997): 380–390.
Stanley, Steven M. Children of the Ice Age: How a Global Catastrophe Allowed Humans to Evolve. new york: W. H. Freeman, 1998.
niche See competition, ecological.
nitrogen cycle the nitrogen cycle is the move-ment of nitrogen atoms through the ecosystems of the earth: through the atmosphere, food webs, and soils. it is a cycle because it begins and ends with nitrogen gas (n2) in the atmosphere. Organisms con-trol the cycling of nitrogen atoms.
the atmosphere consists mostly of nitrogen gas (n2). Because of this, there would seem to be no problem with the availability of nitrogen atoms to any organism on earth. n2, however, is virtually useless to most organisms. nearly the only way that nitrogen atoms become available to organisms is through nitrogen fixation, a process carried out only by a few kinds of bacteria. in this process, nitro-gen gas is made into ammonia (nH3). Once the ammonia is formed, microorganisms and plants can change it into compounds such as nitrates (nO3
-). Plants absorb and use ammonia and nitrates from the soil or from the nitrogen-fixing bacteria in the nodules of the roots of some plants.
Biological nitrogen fixation is carried out by the nitrogenase enzyme. nitrogenase operates most efficiently in the presence of relatively low concen-trations of O2. among the microorganisms that fix nitrogen are the cyanobacteria and the nitrogen-fixing bacteria:
● cyanobacteria are photosynthetic. Most of them live in rivers, lakes, and oceans. nitrogenase is often found in specialized cells called heterocysts, which shield the nitrogenase from high concentrations of O2 found in atmosphere and water. Because the growth of cyanobacteria is not limited by the availability of nitrates or ammonia in the water, their populations may grow explosively (a process called eutrophica-tion) if phosphates become abundant, as when water is polluted by even nontoxic concentrations of phosphate-containing detergents.
● Most nitrogen-fixing bacteria (in the genera Nitrobacter and Nitrosomonas) live inside
of the roots of certain plants, although many live independently in the soil. Many of these plants (beans, clover, alfalfa, and locust and acacia trees) are in the legume family Fabaceae. the roots of these plants have lumps called nodules which contain many nitrogen-fixing bacteria. leguminous plants can obtain ammonia directly from these nodules. Other plants, such as alders, contain a different group of nitrogen-fixing bacteria, in the genus Frankia.
When the leguminous plants die, decompos-ers consume them, breaking down their molecules. Major waste products of this decomposition are nH4
+ and nO3-, which accumulate in the soil. Plants
that do not have nitrogen-fixing bacteria in their roots obtain nitrogen atoms from these soil reserves of nH4
+ and nO3-. animals obtain the nitrogen
atoms that they need by eating leguminous or other plants or by eating other animals. When these plants or animals die, decomposers release nH4
+ and nO3-
from them as well. the nH4+ and nO3
- released by decomposers enter the soil slowly, whereas the nH4
+ and nO3
- fertilizers used by farmers are applied in large quantities. Much of the nO3
- that is applied to farmlands washes through the soil and into rivers or down into the underground water supplies from which people drink.
if nitrogenous ions accumulated indefinitely in the soil, they would reach toxic levels. this does not occur, for two major reasons. First, water washes nitrates out of the soil, and they travel through streams and rivers to the ocean. there they accu-mulate, eventually ending up in sediments which are made into rocks. these rocks, millions of years later, can contribute nitrates to the soil that is formed from them. ammonium clings more tightly to clay particles in the soil, but also eventually leaches from soil. Second, denitrifying bacteria can change soil nitrogen compounds back into nitrogen gas, which eventually returns to the atmosphere, completing the cycle.
Farmers sometimes grow leguminous plants such as clover during the season when their principal crop is not under cultivation. Because they have nitrogen-fixing bacteria in their roots, the clover is rich in nitro-gen compounds. the farmers then plow the clover into the soil during spring planting. two advantages result from this practice. First, the leguminous plants protect the soil from erosion during the late fall, winter, and early spring. Second, when the plants decompose in the soil, abundant nitrates are released. More often, farmers will practice crop rotation in which they plant a leguminous crop one year and another crop the next. they harvest the leguminous crop, thus remov-
xviii+598_Biodiversity_F.indd 394 11/17/11 1:13 PM

nitrogen cycle 395
ing some of the nitrate that would otherwise have returned to the soil. However, the leaf litter and the dead roots from the leguminous crop contribute some nitrate to the soil. this practice reduces the amount of fertilizer the farmers need to apply.
Humans have had a tremendous impact on the global nitrogen cycle, primarily by the application of artificial fertilizers. Some bacteria produce nitrous oxide (n2O), which is a pollutant and greenhouse gas; the overuse of fertilizers has caused atmospheric levels of nitrous oxide to increase.
See also biogeochemical cycles; ecosys-tems; rivers, streams, ponds, and lakes.
Further readingFalkowski, Paul G. “evolution of the nitrogen cycle and its
influence on the biological sequestration of cO2 in the ocean.” Nature 387 (1997): 272–275.
Galloway, James n. “the global nitrogen cycle: changes and consequences.” Environmental Pollution 102 (1998) supplement: 15–24.
leigh, G. J. The World’s Greatest Fix: A History of Nitro-gen and Agriculture. new york: Oxford University Press, 2004.
Vitousek, Peter M., et al. “Human alteration of the global nitrogen cycle: Sources and consequences.” Ecological Applications 7 (2007): 737–750.
xviii+598_Biodiversity_F.indd 395 11/17/11 1:13 PM

396
oligocene epoch See Paleogene period, biodiversity in.
ordovician period, biodiversity in the Ordovician period (510 to 440 million years ago) was the second period of the Paleozoic era. it fol-lowed the cambrian period and preceded the Silu-rian period. an average of about 400 families of marine organisms existed during the Ordovician, compared to 1,000 marine families (and 2,200 total families) today.
Climate. Oceans were warm around equato-rial continents, but cool around the mass of south-ern continents. there was little multicellular life on the continents to be affected by terrestrial cli-mate. atmospheric oxygen levels were much lower, and carbon dioxide levels much higher, than today. Global tem-peratures were also higher than today (see table on page 397).
Continents. Oceans sepa-rated the continents of lauren-tia, Baltica, Siberia (these three continents today dominating the northern Hemisphere) and Gond-wana (today, the southern conti-nents). Sea level was much higher than today—almost 700 feet (over 200 meters) higher than modern oceans.
Marine life. almost all organisms lived in the oceans and freshwaters at this time (see fi g-ure). By the end of the Ordovician period, the diversity of marine
animals more than tripled over that of the cambrian period. this was partly due to an increasing special-ization in the ways that the animals obtained food. it was also partly due to the formation of the fi rst reefs, though the corals that formed them were of classes that are now extinct, and many reefs were formed by sponges. When the reefs formed, they created many new habitats for other swimming and crawling and sessile animals.
● Invertebrates. the most abundant group of arthropods was the trilobites; modern forms of arthropods such as crabs were rare. the early Ordovician period, with about 60 trilobite families, was the peak of trilobite diversity. the fi rst bivalve and cephalopod mollusks evolved. large cephalopod mol-
O
Eurypterids (also called sea scorpions) were some of the largest predators in the Ordovician oceans. They were larger than, and may have preyed upon, some early fi shes.
xviii+598_Biodiversity_F.indd 396 11/17/11 1:13 PM

ordovician period, biodiversity in 397
lusks such as Endoceras had cone-shaped shells. among the echinoderms were the stalked crinoids such as Caleidocrinus.
● Vertebrates. the only vertebrates were fishes, jawless descendants of the early fishes of the cambrian period. Astrapsis had no jaws or fins; only its tail protruded outside a
bony armor that covered its body. this four-inch (10-centimeter) fish presumably used its simple tubelike mouth to filter food from the ocean floor. larger jawless armored fishes included Pycnapsis.
Continental life. terrestrial life-forms have not been clearly confirmed from the Ordovician period. Spores characteristic of primitive land plants and trace fossils of animal activity have been found from the later part of this period.
Extinctions. One of the great mass extinctions occurred at the end of the Ordovician period. it is estimated that 26 percent of families, representing 83 percent of species, became extinct by the end of the Ordovician. While the continent that later became north america and europe was equatorial and surrounded by tropical reefs, the large Gondwa-nan continent (which today exists as South amer-ica, africa, australia, antarctica, and fragments of other land masses) was at the South Pole. not only did Gondwanan weather become colder, but ocean levels dropped as glaciers formed around the pole, and this devastated tropical reefs. the return of warm weather and the melting of glaciers toward the end of the Ordovician may also have caused stagna-tion of deep-sea waters.
Further readingadachi, natsuko, et al. “early Ordovician reef construc-
tion in anhui Province, South china: a geobiologi-cal transition from microbial- to metazoan-dominant reefs.” Sedimentary Geology 220 (2009): 1–11.
Benton, Michael. “Diversification and extinction in the his-tory of life.” Science 268 (1995): 52–58.
Buatois, luis a., et al. “the changing face of the deep: colonization of the early Ordovician deep-sea floor, Puna, northwest argentina.” Palaeogeography, Pal-aeoclimatology, and Palaeoecology 280 (2009): 291–299.
cope, John c. W. “Diversification and biogeography of bivalves during the Ordovician period.” Geological Society, London, Special Publications 194 (2002): 35–52.
Webby, Barry D., et al., eds. The Great Ordovician Biodi-versification Event. new york: columbia University Press, 2004.
multiCellular biodiversity in the ordoviCian period
Conditions
Atmospheric oxygen 68% of modern level
Atmospheric carbon dioxide
12 times modern level
Global temperature 3°F (2°C) higher than today
Sea level 650 feet (200 m) higher than today
Approximate number of families*
Continental 0
Marine 400
Total 400
Examples of marine animals
Astrapsis splendens jawless fish
Amplexograptus sp. hemichordate
Asaphus kotlukovi trilobite arthropod
Caleidocrinus multiramus crinoid echinoderm
Cysaster stellatus echinoderm
Endoceras sp. cephalopod mollusk
Glyptarca sinensis bivalve mollusk
Panderodus gracilis conodont fish
Platystrophia ponderosa brachiopod
Protaraea richmondiensis coral
Pycnapsis splendens ostracoderm fish
*Based on Benton
xviii+598_Biodiversity_F.indd 397 11/17/11 1:13 PM

398
paleocene epoch See Paleogene period, biodiversity in.
paleogene period, biodiversity in the Paleogene period (65 to 23 million years ago) was the fi rst period of the cenozoic era. it followed one of the mass extinctions which occurred at the end of the cretaceous period; the neogene period fol-lowed it. the cenozoic era is also known as the age of Mammals, because after the extinction of the dinosaurs the mammals had opportunity to evolve into tremendous diversity, including many large forms, including the fi rst fully aquatic mammals. the Paleogene period is the fi rst part of what has traditionally been called the tertiary period, consist-ing of the Paleocene, eocene, and Oligocene epochs. Scientists have divided the tertiary into Paleogene and neogene periods largely because the climates of the two periods were different on a worldwide scale (see table on page 399).
Climate. Warm moist conditions that allowed extensive forests were widespread in the Paleogene period. the temperature difference between the equa-tor and the poles was only half as great as it is today. Forests grew near the north and South Poles. Scien-tists estimate that conditions were warm enough that the trees in the far north did not need to be deciduous to avoid snowfall in winter. they were deciduous, scientists have concluded, because of the half year of darkness rather than because of cold temperatures. the land surface of the earth was virtually covered with forests at this time. Before the end of the Paleo-gene, ice sheets had begun to form in antarctica.
about 55 million years ago, at the end of the Paleocene epoch, the earth experienced a brief and
large increase in temperature (the Paleocene-eocene thermal Maximum). Due to its brevity and the lack of evidence of other causes, this event may have resulted from the sudden emergence of meth-ane from continental shelves, similar to one of the factors that contributed to the Permian extinction. Fossils from that time indicate a sudden increase in the carbon isotope that is common in organisms. it took only 20,000 years for the global average tem-perature to increase almost 11°F (6°c). this is about the same rate that global temperatures are increasing today, which is one reason that the greenhouse effect is considered the most important ecological crisis of modern times. the rapid increase in global atmo-spheric and oceanic carbon also made the oceans acidic. this may help to explain why there was so much extinction of shelled marine organisms at that time, although other groups of organisms were not greatly affected. the regrowth of forests all over the world brought the high levels of atmospheric carbon back down.
Continents. the northern and southern con-tinents were largely separate from one another dur-ing the Paleogene. as the continents continued to move, connections began to form between the north-ern and southern continents. the subcontinent of india collided with the eurasian continent before the end of the Paleocene, about 34 million years ago, and began to form the Himalayas. europe and north america were just beginning to separate at the beginning of the Paleogene.
Marine life. life in the Paleogene oceans closely resembled that of modern times:
● Invertebrates. all modern groups of inver-tebrates were not only in existence but were
P
xviii+598_Biodiversity_F.indd 398 11/17/11 1:13 PM

paleogene period, biodiversity in 399
dominant. ammonites (large cephalopod mollusks with coiled shells) had become extinct during the cretaceous extinction.
● Vertebrates. all of the completely marine reptiles had become extinct during the cre-taceous extinction. at the beginning of the Paleogene, the dominant marine ver-tebrates were bony fishes and sharks. Very soon after the cretaceous extinction, the first whales began to evolve. Pakicetus ina-chus was a terrestrial mammal that spent a lot of time in the water beginning about 56 million years ago. Ambulocetus natans lived in shallow waters about 50 million years ago and had hind legs more adapted to swimming than to walking. like mod-ern whales, Ambulocetus had no external ears—an aquatic adaptation. Rodhocetus, which lived about 47 million years ago, was almost exactly intermediate between Pakicetus and modern whales; it still had front and hind legs. Basilosaurus cetoides was a whale that lived about 40 million years ago. its hind legs were becoming ves-tigial: they were only two feet (0.6 m) long compared to the animal’s body length of 60 feet (18 m). Modern whales do not have hind legs at all, although they have vestigial hip bones. these animals form a perfect series of intermediates in whale evolution. By the end of the Paleogene, both lineages of modern whales (baleen and toothed) had differentiated. the toothed whale Squalo-don used echolocation, like modern whales and dolphins. Aetiocetus cotylaveus was a baleen whale, but its blowhole was only halfway up its face rather than on the top of the head as in modern whales.
Continental life. Plants and animals thrived and diversified in the worldwide forests of the continents:
● Plant life. Many modern genera of trees were in existence in the worldwide forests. all modern groups of conifers, including pines, existed by this time. Bananas grew in what is now Oregon. there were not any exten-sive grasslands or deserts until the neogene period, but species of cactus and grass existed during the Paleogene: the grasses probably lived along disturbed riversides, and cactuses may have been epiphytes, as many are today.
● Invertebrates. all major groups of terres-trial invertebrates, such as ants and butter-flies, had evolved by the Paleogene period.
multiCellular biodiversity in the paleogene period
Conditions
Atmospheric oxygen 30% above modern level
Atmospheric carbon dioxide
40% above modern level
Global temperature 7°F (4°C) above modern level
Sea level 180 feet (110 m) higher than today
Approximate number of families*
Continental 1,200
Marine 1,000
Total 2,200
Examples of marine animals
Acanthonemus sp. bony fish
Aetiocetus cotylaveus baleen whale
Aturia sp. cephalopod mollusk
Basilosaurus cetoides whale ancestor
Enoploclytia gardnerae crustacean
Heliobatis radians ray
Notorynchus serratissimus shark
Rodhocetus balochistanensis
whale with legs
Squalodon sp. toothed whale
Examples of continental animals
Ambulocetus natans whale ancestor
Aquilavis hypogaeus hawk
Arfia opisthotoma small carnivorous mammal
Borealosuchus formadibilis crocodile
Brontotherium gigas large grazing mammal
Champsosaurus gigas freshwater reptile
Folisaurus boutersemensis lizard
Gastornis giganteus large predatory ground bird
Hyracotherium leporinum horse ancestor
Leptoganga sepultus dove
Mesohippus latidens primitive horse
Miacis rosei carnivore mammal
Monotrematum sudamericanum
monotreme mammal
(continues)
xviii+598_Biodiversity_F.indd 399 11/17/11 1:13 PM

first entry400 paleogene period, biodiversity in 400
● Vertebrates. Modern groups of freshwater fishes, amphibians, and reptiles continued to diversify (see figure). the snake Titanoboa was as long as a bus and weighed as much as a car. at the beginning of the Paleogene, most of the mammals were in groups that are now extinct, such as the multitubercu-lates. the ferretlike Tritemnodon, the dog-like Arfia, the catlike Patriofelis, and the hyenalike Oxyaena were not related to mod-ern ferrets, dogs, cats, or hyenas, but were all creodonts—an evolutionary lineage that is now extinct. the largest land mammals to ever exist, such as the rhinoceros-like Bron-totherium, evolved into the role vacated by the extinction of the dinosaurs. the largest was Paraceratherium, which was 16 feet (five meters) tall at the shoulder; its head was 25 feet (7.5 meters) above the ground, and its skull was almost four and a half feet long (1.3 meters). the earliest horses evolved—Hyracotherium in the early Paleogene and Mesohippus in the late Paleogene. By the end of the Paleogene, modern mammal groups had evolved, including bats, rodents, and doglike carnivores. in addition to some mod-ern types of birds such as pigeons, hawks, and owls, there were large predatory birds that ran on the ground. there were swifts, but small hummingbirds (their close rela-tives) had not yet evolved.
See also cretaceous extinction; green-house effect.
Further readingBains, S., et al. “termination of global warmth at the Pal-
aeocene/eocene boundary through productivity feed-back.” Nature 407 (2000): 171–174.
Benton, Michael J. “Diversification and extinction in the history of life.” Science 268 (1995): 52–58.
Dickens, G. R., et al. “a blast of gas in the latest Paleo-cene: Simulating first-order effects of massive dissocia-tion of oceanic methane hydrate.” Geology 25 (1997): 259–262.
Gingerich, Philip D., et al. “Hind limbs of eocene Basi-losaurus: evidence of feet in whales.” Science 249 (1990): 154–157.
———, et al. “new whale from the eocene of Pakistan and the origin of cetacean swimming.” Nature 368 (1994): 844–847.
Head, J. J., et al. “Giant boid snake from the Palaeocene neotropics reveals hotter past equatorial tempera-tures.” Nature 457 (2009): 715–717.
Jahren, a. Hope. “the arctic forest of the Middle eocene.” Annual Review of Earth and Planetary Sciences 35 (2007): 509–540.
multiCellular biodiversity in the paleogene period (continued)
Necrobyas harpax owl
Oxyaena lupina small carnivorous mammal
Pakicetus inachus whale ancestor
Palaeochiropteryx tupaiodon
bat
Paraceratherium transouralicum
large grazing mammal
Patriofelis ferox small carnivorous mammal
Priscacara serrata freshwater bony fish
Protochelydra zangerli turtle
Pseudoloris godinoti primate
Ptilodus sp. multituberculate mammal
Simimys simplex rodent
Titanoboa cerrejonensis snake
Tritemnodon agilis small carnivorous mammal
Vanessa amerindica butterfly
Xenopus romeri frog
Examples of land plants
Acacia mahengense acacia
Alnus parvifolia alder
Betula leopoldae birch
Ensete oregonense banana
Eopuntia douglassii cactus
Fagus pacifica beech
Ginkgo dissecta ginkgo
Juglans clarnensis walnut
Larix altoborealis larch
Macginitiea gracilis sycamore
Metasequoia occidentalis redwood
Pharus sp. grass
Pinus andersoni pine
Populus cinnamomoides poplar
Quercus nevadensis oak
Ulmus woodii elm
*Based on Benton
xviii+598_Biodiversity_F.indd 400 11/17/11 1:13 PM

parasitism 401
Janis, christine. “Victors by default: the mammalian succession.” in The Book of Life: An Illustrated History of the Evolution of Life on Earth, edited by Stephen J. Gould. new york: norton, 2001.
Katz, M. e., et al. “Uncorking the bottle: What triggered the Paleo-cene/eocene thermal maximum methane release.” Paleoceanogra-phy 16 (2001): 667.
Köhler, M., and S. Moyà-Solà. “a fi nding of oligocene primates on the european continent.” Pro-ceedings of the National Acad-emy of Sciences USA 96 (1999): 14,664–14,667.
labandeira, conrad c., et al. “a den-droctonous bark engraving (cole-optera: Scolytidae) from a Middle eocene Larix (coniferales: Pina-ceae): early or delayed coloni-zation?” American Journal of Botany 88 (2001): 2,026–2,039.
Shellito, cindy J., and lisa c. Sloan. “Reconstructing a lost eocene paradise: Simulating the change in global fl oral distribution at the initial eocene thermal maxi-mum.” Global and Planetary Change 50 (2006): 1–17.
Simmons, nancy B., et al. “Primitive early eocene bat from Wyo-ming and the evolution of fl ight and echolocation.” Nature 451 (2008): 818–821.
Sluijs, a., et al. “Subtropical arctic Ocean temperatures during the Palaeocene/eocene thermal max-imum.” Nature 441 (2006): 610–613.
thewissen, J. G. M., et al. “Skeletons of terrestrial ceta-ceans and the relationship of whales to artiodactyls.” Nature 413 (2001): 277–281.
thomas, D. J., et al. “Warming the fuel for the fi re: evi-dence for the thermal dissociation of methane hydrate during the Paleocene-eocene thermal maximum.” Geology 30 (2002): 1,067–1,070.
Zachos, J. c., and l. R. Kump. “carbon cycle feedbacks and the initiation of antarctic glaciation in the earliest Oligocene.” Global and Planetary Change 47 (2005): 51–66.
———, et al. “Rapid acidifi cation of the ocean during the Paleocene-eocene thermal Maximum.” Science 308 (2005): 1,611–1,615.
———, et al. “an early cenozoic perspective on green-house warming and carbon-cycle dynamics.” Nature451 (2008): 279–283.
parasitism Parasitism occurs when individuals of one species (the parasite) obtain their living from individuals of another species (the host). Parasites have a negative impact on the host, either by con-suming directly from it, by consuming some of its food (as when intestinal worms harm the host’s nutrition), by producing toxic waste products (as do many bacterial and viral parasites such as chol-era and bubonic plague), or by causing the host’s own immune system to have harmful effects on the host. it is sometimes diffi cult to distinguish para-sitism from other kinds of ecological interactions. For example, aphids insert their mouthparts into the pressurized sugar sap of plant phloem; are they parasites or herbivores? an organism that entirely consumes another is a predator. an organism that makes its living off a host without harming the host is a commensal. the distinctions among parasite,
Two mammals of the Paleogene period. (A) Uintatherium was a large browsing mammal, one of the largest mammals that ever lived. (B) Eurotamandua had adaptations for eating ants, even though it was prob-ably not an ancestor of the modern anteater.
xviii+598_Biodiversity_F.indd 401 11/17/11 1:13 PM

first entry402 parasitism 402
predator, and commensal are sometimes unclear. a predator may not consume the entire body of its prey, and a parasite can kill its host. a commensal that takes just a little too much from its host may shift into becoming a parasite.
the evolution of parasites has been strongly influenced by the hosts. For example, the life cycles of fleas that live on rabbits are coordinated with hormone levels in the rabbits’ blood, which ensures that the young fleas hatch at the same time as the baby rabbits are born. the evolution of the hosts has been influenced by parasites; the very existence of immune systems may be considered proof of this.
Because parasite and host species influence one another’s evolution, parasitism is an example of coevolution. it is also, along with commensalism and mutualism, an example of symbiosis in which two species do not merely influence one another’s evolution but are intimately connected with one another. in parasitism, the parasite harms the host; in commensalism, the commensal has no effect on the host; in mutualism, the two mutualists benefit one another.
Parasitism has contributed to biodiversity in at least three ways. First, many thousands of species are parasites. Some entire animal phyla (Myxozoa, Orthonectida, nematomorpha, and acanthoceph-ala) consist of parasites. Second, parasitism—either parasites finding new hosts, or hosts finding new ways to avoid parasites—has often been the stimu-lus for speciation. adaptations of parasites and hosts are among the most complex in the natural world. third, parasitism may be the principal force behind some kinds of sexual selection. according to many evolutionary scientists, colorful feathers on male birds may be fitness indicators that dem-onstrate to the female that they are free of para-sites. Sexual selection, in turn, leads to even more speciation.
Parasites make their living off hosts in numer-ous ways. Many parasites live inside of their hosts. nearly all infectious diseases of plants, animals, and humans are examples of this, as in the following examples:
● Most bacteria are harmless to multicellu-lar organisms, although the list of bacterial diseases in animals is long and depressing, such as bubonic plague and cholera. Per-haps because plant tissues are mildly acidic, something that most bacteria cannot tol-erate, bacteria cause relatively few plant diseases.
● Most fungi are harmless to multicellular organisms as well, although many of them
cause plant diseases, such as smuts and rusts. Relatively few fungi cause animal dis-eases, and these usually occur only in weak animals.
● Many protists also cause diseases, such as malaria in animals and late blight in potatoes.
● Viruses do not have their own molecular machinery for replication, and must infect many kinds of multicellular organisms. animal examples include influenza and aiDS. Viruses called bacteriophages even infect bacteria, replicating inside of them and eventually causing them to burst open, releasing hundreds of new viruses.
● Parasitic worms such as tapeworms live in various parts of animal bodies. Many live in the digestive system, where they have undergone evolutionary simplification. they attach to the host gut lining and absorb the food that the host digests. Others get into organs such as eyes, causing blind-ness. Worms that live in the lymph system may block the passages and cause swelling. an example is elephantiasis, which causes extensive swelling of the legs and scrotum (see figure). Some parasitic worms even live in brain tissue.
● a few kinds of flowering plants live entirely inside the tissues of other flowering plants. Plants in the genus Rafflesia have no leaves
Elephantiasis is caused by a parasitic worm that stim-ulates the human immune system to produce massive swelling. (Centers for Disease Control and Prevention; Public Health Image Library)
xviii+598_Biodiversity_F.indd 402 11/17/11 1:13 PM

parasitism 403
or roots and live entirely within grape vines of the genus Tetrastigma. their presence can be seen only when the large flowers emerge from the host vine.
● Some insects lay their eggs in plant tissue. When the larval insect hatches, it releases a chemical that induces the plant to produce a gall that both feeds and protects the larval insect (see photograph above).
Some parasites live on the outside of their hosts. examples include the following:
● ticks, fleas, and lice are arthropods that drink blood from the skin of vertebrates. they have structures that allow them to attach very firmly to the host, and their bod-ies are flattened, which makes it difficult for the host to scratch them away.
● Some plants are partly or entirely para-sitic upon other plants. examples include the dodder (genus Cuscuta), indian pipe (Monotropa), and some orchids (Corallo-rhiza). they have no chlorophyll and obtain water, minerals, and sugar from the xylem and phloem of their host plants.
● Some plants are partly parasitic upon other plants. examples include mistletoe species, which have chlorophyll and make their
own sugar, but obtain water and minerals from the xylem of their host trees. Many herbaceous plants attach to the roots of other plants, often grasses, and obtain water and minerals from them. examples include asian sandalwood tree (genus San-talum) and its north american herbaceous relative, the bastard toadflax (Coman-dra). Many members of the toadflax fam-ily, such as indian paintbrushes (Castilleja) and witchweed (Striga), are root parasites. Witchweed parasitizes large grasses, includ-ing maize, and can completely kill corn-fields in africa.
Some parasites only occasionally visit their hosts, usually to obtain blood from mammals. examples include mosquitoes and vampire bats. Surprisingly, blood (which transports but does not store nutrients) is not a very nutritious food source. Female mosquitoes use vertebrate blood to promote egg development. Vampire bats must drink blood daily; successful bats share blood with unsuccessful ones. there are even some finches on the Galapagos islands that peck the feet of other birds to obtain blood.
Hosts have numerous ways of responding to parasites. Some hosts tolerate them by continuing to function despite a parasite load. Others resist the parasites by means such as an immune system, in which white blood cells attack microbes. in many cases, hosts use a combination of tolerance and resistance. Genomic studies have shown that mouse adaptations conferring tolerance are different from those conferring resistance; mice that are more toler-ant are less resistant, and vice versa.
Parasites usually have very effective meth-ods of dispersing to new hosts. this includes extremely high reproductive rates. intestinal para-site bacteria or eggs can fill polluted waters. a single witchweed plant can produce 50,000 seeds. But parasites often have other ways of enhancing their dispersal. Many contagious diseases spread because the parasites cause the host to cough, thus projecting droplets of saliva that contain mil-lions of bacteria or viruses. Often the disease is most contagious prior to the stage when symp-toms become obvious, and the host continues to mingle with other members of its species. Parasites can even change the bodies and behavior of their hosts. Some parasitic worms cause mollusks, nor-mally reclusive, to climb out on the tips of leaves where birds will see and eat them, while others cause ant abdomens to swell and become red, resembling berries, which birds eat (see photo-graph on page 404). the virus that causes rabies
This spherical gall on a post oak leaf was produced by an insect larva. The larva produced a waste product that functioned as a plant growth hormone; the oak leaf produced the spherical structure, which protected and fed the grub. The main defense that oak trees have against gall-producing insects is to shed the infected leaves. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 403 11/17/11 1:13 PM

first entry404 parasitism 404
causes the vertebrate host to become very aggres-sive, spreading the virus by its bites.
in many cases, animal vectors spread the para-site. Usually the vectors are themselves parasites. examples include mosquitoes. Viral and protist par-asites live in mosquito salivary glands and spread to new hosts when the mosquitoes bite them. Fleas and ticks are also disease vectors. Sometimes more than one vector species is involved in spreading a disease. For example, the mosquito Culex tarsalis spreads West nile viruses in bird populations, while C. pipi-ens spreads the virus to humans. in california, the deer tick Ixodes spinipalpis spreads the lyme dis-ease bacterium Borrelia burgdorferi among rodents, but I. pacificus spreads the bacteria to humans. epi-demiologists are focusing much of their attention upon stopping the spread of diseases by interfering with the activity of the vector. the bacterium Wol-bachia kills Aedes aegypti mosquitoes, which spread malaria, but does not do so right away. But since it is usually the older mosquitoes that spread most of the malaria, infecting mosquito populations with Wolbachia may reduce the amount of malaria that they spread.
Many multicellular parasites have complex life cycles, some examples of which are provided in the entry about animals. they change form as they travel from one host to another and some-times a third. Parasites may also have adaptations that allow them to escape from the host species’
behavioral protection. Some arthropods live in ant nests. Some, such as the caterpillars of Maculinea butterflies, have skin surface chemicals that entice the ants to take care of them. Some spiders, which have eight legs and no antennae, imitate the behav-ior of ants, which have six legs and antennae, by walking along with their front legs held up like antennae.
Parasites can adapt to new host species or populations, often in conjunction with a genetic mutation that alters their host preference or effects. Several human diseases (known as zoonotic dis-eases) have come from livestock. One of the worst human diseases in history, smallpox (which has been eradicated from human populations by vac-cination), apparently evolved from cowpox. a few diseases have transferred the other way; apparently cattle caught tuberculosis from humans. the bac-terium Treponema pallidum causes both syphilis and yaws. Researchers found a variant form of yaws (T. p. pertenue) in a remote South american tribe that had characteristics similar to syphilis (T. p. pallidum). apparently, europeans carried the vari-ant form of yaws to europe, where it evolved into syphilis.
Some birds, most notably cowbirds and cuckoos, are nest parasites: they lay their eggs in the nests of other bird species (see photograph on page 405). the parent birds of the host species feed the parasitic nestlings as if they were their own. the host birds do not recognize that the parasitic nestling is not their own, even when it is much larger and a differ-ent color than their own offspring and the parasite pushes the host offspring out of the nest.
paraSITISm aNd CommENSalISmcommensal populations living on hosts often help to protect the host from parasites. Most of the trillions of bacteria on or in the host body are commensals, and their populations often crowd out the populations of parasites. commensalis-tic bacteria in chicken intestines usually crowd out deadly Salmonella bacteria, and chicks obtain these commensalistic bacteria from their moth-ers. chickens raised in vast industrial populations, however, usually do not have an opportunity to inherit commensalistic bacteria from the mothers who laid the eggs. Parasites also spread more read-ily through dense populations of hosts. Huge “fac-tory” populations of chickens therefore suffer from Salmonella infections more than do “free-range” chickens that grow up in smaller populations with their mothers.
the distinction between parasitism and com-mensalism is a shifting border. normally commen-salistic microbes can become parasitic under certain
A parasite has caused this Cephalotes aratus ant to have a bright red abdomen. Birds, mistaking the ant for a berry, will consume the ants and ingest the para-site. The parasite develops into a new form and lives inside the bird. Parasite eggs in bird droppings are ingested by foraging ants. (Steve Yanoviak; University of Arkansas)
xviii+598_Biodiversity_F.indd 404 11/17/11 1:13 PM

parasitism 405
conditions. Probably all commensalistic microbes are prevented from becoming parasites by the host immune system, as in these examples:
● the human mouth, throat, and bronchial passages are filled with millions of bacte-ria and also some protists. the only harm they normally cause is to produce acid under anaerobic conditions, leading to tooth decay, which can be prevented by proper hygiene. these bacteria do not normally infect the lungs because the host immune system prevents them from doing so. When the host immune system is weakened after a long bout of influenza or during infection with HiV, they can become parasitic and cause potentially deadly pneumonia.
● the human skin is covered with millions of bacteria, including staph. these bacteria are harmless if they remain on the outside of the skin, where they consume dead cells and oils. their worst effect is to produce body odor, once again controllable by hygiene. if they get inside the skin, when the skin is cut, they can cause serious and potentially deadly skin infections.
● the human intestine is filled with trillions of harmless (or in some cases beneficial) bacteria. if these bacteria get into the lining of the intestine, they can cause an infection. infection of the appendix causes appendici-tis, which can cause the appendix to burst and spread deadly bacteria throughout the body cavity. Gunshot and stab wounds can release bacteria from the intestines into the blood, causing blood poisoning, or into tis-sues, causing gas gangrene.
the border between parasitism and commen-salism is also an evolutionary one. natural selection often favors the parasites that have the least effect upon the host as well as the hosts that are most resistant to the parasites. that is, natural selection often leads to milder bacteria and stronger hosts, a process known as balanced pathogenicity. this process occurs mostly with parasites that spread by direct contact from one host to another, because the parasites that spread most effectively are those that allow their hosts to mingle with the rest of the population. People tend to stay away from others who display severe disease symptoms. Microbes that spread by insect vectors or sewage, however, do not evolve to become milder. this is because rel-atively deadly and relatively mild microbes spread equally well in these cases. Humans cannot stay away from water or mosquitoes that carry virulent
microbes any better than they can from those that carry mild microbes.
paraSITISm aNd ENvIroNmENTSeveral human practices increase the parasite load of natural populations. Parasitic diseases often spread through fish raised on fish farms, and when these fish escape into the wild they carry the parasites with them. it is also very likely that the green-house effect will cause parasitic diseases to spread, a fact that has caught the attention of many public health experts. Some diseases such as dengue fever are tropical mainly because the mosquitoes that serve as their vectors live only in tropical climatic zones. as temperatures increase, so will the tropical zones, allowing the mosquitoes (and the parasites) to spread to locations previously free of them. chol-era bacteria persist in resting stages when they are not actively moving through human populations but can become active again under warmer conditions. Global warming might, in this way, encourage the spread of cholera.
newly introduced parasites are less likely to spread through intact ecosystems. apparently bio-diversity itself is an important component of ecosys-tems that helps to prevent the spread of diseases.
See also animals; coevolution; commensal-ism; greenhouse effect; mutualism; sexual selection.
A common cuckoo laid an egg in the nest of a great reed warbler, Acrocephalus arundinaceus. The cuckoo egg hatched before those of the warbler, and the cuckoo nestling pushed warbler eggs out of the nest. (Image from Anderson et al., PLoS ONE)
xviii+598_Biodiversity_F.indd 405 11/17/11 1:13 PM

first entry406 parasitism 406
Further readinganatolin, Michael F. “Unpacking β: Within-host dynamics
and the evolutionary ecology of pathogen transmis-sion.” Annual Reviews of Ecology, Evolution, and Systematics 39 (2008): 415–437.
anderson, Michael G., et al. “egg eviction imposes a recov-erable cost of virulence in chicks of a brood parasite.” PLoS ONE 4 (2009): e7725.
andersson, anders F., and Jillian F. Banfield. “Virus pop-ulation dynamics and acquired virus resistance in natural microbial communities.” Science 320 (2008): 1,047–1,050.
Blanford, Simon, et al. “Fungal pathogen reduces poten-tial for malaria transmission.” Science 308 (2005): 1,638–1,641.
Burdon, Jeremy J., and Peter H. thrall. “coevolution of plants and their pathogens in natural habitats.” Sci-ence 324 (2009): 755–756.
cantor, norman F. In the Wake of the Plague: The Black Death and the World it Made. new york: Free Press, 2001.
carrell, Jennifer lee. The Speckled Monster: A Historical Tale of Battling Smallpox. new york: Penguin, 2003.
chambouvet, aurelie, et al. “control of toxic marine dino-flagellate blooms by serial parasitic killers.” Science 322 (2008): 1,254–1,257.
crawford, Dorothy H. Deadly Companions: How Microbes Shaped Our History. Oxford, U.K.: Oxford University Press, 2007.
Davies, nicholas B., and Justin a. Welbergen. “Social trans-mission of a host defense against cuckoo parasitism.” Science 324 (2009): 1,318–1,320.
Desowitz, Robert S. The Malaria Capers: More Tales of Parasites and People, Research and Reality. new york: norton, 1991.
Duplaix, nicole. “Fleas: lethal leapers.” National Geo-graphic, May 1988, 67–94.
Fenner, Frank, et al. Smallpox and Its Eradication. Geneva, Switzerland: World Health Organization, 1988.
Finlay, B. Brett. “the art of bacterial warfare.” Scientific American, February 2010, 56–63.
Fisher, Jeffrey a. The Plague Makers: How We Are Cre-ating Catastrophic New Epidemics—and What We Must Do to Avert Them. new york: Simon & Schus-ter, 1994.
Fox, Maggie. “Parasite makes ants into ‘berries’ to entice birds.” Reuters, January 16, 2008. available online. URl: http://uk.reuters.com/article/2008/01/16/uk-ant-berry-idUKn1662901820080116. accessed august 8, 2011.
Garrett, laurie. The Coming Plague: Newly Emerging Dis-eases in a World Out of Balance. new york: Farrar, Straus and Giroux, 1994.
Hayes, K. S., et al. “exploitation of the intestinal micro-flora by the parasitic nematode Trichuris muris.” Sci-ence 328 (2010): 1,391–1,394.
Henig, Robin Marantz. A Dancing Matrix: Voyages along the Viral Frontier. new york: Knopf, 1993.
Kambris, Zakaria, et al. “immune activation by life-short-ening Wolbachia and reduced filarial competence in mosquitoes.” Science 326 (2009): 134–136.
Krkošek, Martin, et al. “epizootics of wild fish induced by farm fish.” Proceedings of the National Academy of Sciences USA 103 (2006): 15,506–15,510.
leroy, e. M., et al. “Fruit bats as reservoirs of ebola virus.” Nature 438 (2005): 575–576.
li, Jianxiong, and Michael P. timko. “Gene-for-gene resis-tance in Striga-cowpea associations.” Science 325 (2009): 1,094.
li, W. D., et al. “Bats are natural reservoirs of SaRS-like coronaviruses.” Science 310 (2005): 676–679.
lyon, Bruce e., and John Mca. eadie. “conspecific brood parasitism in birds: a life-history perspective.” Annual Reviews of Ecology, Evolution, and Systematics 39 (2008): 343–363.
Marriott, edward. Plague: A Story of Science, Rivalry, and the Scourge that Won’t Go Away. new york: Henry Holt, 2002.
Matossian, Mary Kilbourne. Poisons of the Past: Molds, Epidemics, and History. new Haven, conn.: yale Uni-versity Press, 1989.
May, Robert M. “Population biology of infectious dis-eases.” Nature 280 (1979): 361–367.
Mccormick, Joseph B., and Susan Fisher-Hoch. Level 4: Virus Hunters of the CDC. atlanta: turner, 1996.
McMeniman, conor J., et al. “Stable introduction of a life-shortening Wolbachia infection into the mosquito Aedes aegypti.” Science 323 (2009): 141–144. Sum-marized by Read, andrew F., and Matthew R. thomas. “Mosquitoes cut short.” Science 323 (2009): 5–52.
nash, David R., et al. “a mosaic of chemical coevolution in a large blue butterfly.” Science 319 (2008): 88–90.
Parker, ingrid M., and Gregory S. Gilbert. “the evolu-tionary ecology of novel plant-pathogen interactions.” Annual Review of Ecology, Evolution, and Systemat-ics 35 (2004): 675–700.
Preston, Richard. The Demon in the Freezer. new york: Random House, 2002.
Råberg, lars, Derek Sim, and andrew F. Read. “Disentan-gling genetic variation for resistance and tolerance to infectious diseases in animals.” Science 318 (2007): 812–814.
Rabsch, W., et al. “competitive exclusion of Salmonella enteritidis by Salmonella gallinarum in poultry.” Emerging Infectious Diseases 6 (2000): 443–448.
Scholte, ernst-Jan, et al. “an entomopathogenic fungus for control of adult african malarial mosquitoes.” Science 308 (2005): 1,641–1,642.
Schutt, Bill. Dark Banquet: Blood and the Curious Lives of Blood-Feeding Creatures. new york: Harmony, 2009.
tucker, Jonathan B. Scourge: The Once and Future Threat of Smallpox. new york: atlantic Monthly Press, 2001.
xviii+598_Biodiversity_F.indd 406 11/17/11 1:13 PM

permian extinction 407
Worobey, Michael, et al. “Point, counterpoint: the evolu-tion of pathogenic viruses and their human hosts.” Annual Review of Ecology, Evolution, and Systemat-ics 38 (2007): 515–540.
Zimmer, carl. “isolated tribe gives clues to the origin of syphilis.” Science 319 (2008): 272.
permian extinction the Permian extinction was the biggest mass extinction event in the his-tory of life and as such the greatest challenge to earth’s biodiversity. it has been known since the work of geologist John Phillips in the 1840s that the end of the Permian period was marked by a major turnover in marine animal species; only recently, however, have the magnitude and the causes of this turnover become apparent. the Permian extinction occurred 252 million years ago and marked the end of the Permian period and the Paleozoic era. Paleon-tologist Douglas erwin called it the “mother of all extinctions.” approximately 90 percent of species died during and immediately after this event in the early triassic period. in contrast, the cretaceous extinction, which exterminated the dinosaurs and many other species, killed only about 50 percent of the species on earth. life on earth recovered from the Permian extinction, but it took between 50 and 100 million years to regain its former diver-sity. though the major groups of organisms that had dominated the earth during the Permian (such as seed plants, mollusks, fishes, amphibians, and reptiles) survived, most of the species did not. the triassic period began with a vastly impoverished set of species, and the later triassic had an almost entirely new set of species within these groups. in fact, in some of the earliest triassic deposits, a single species constitutes 90 percent of the vertebrates: a pig-sized, mammal-like reptile, Lystrosaurus. in the earliest triassic marine deposits, only a brachio-pod called Lingula remained abundant. the Perm-ian extinction was the only mass extinction to have significantly affected insects. about half of insect families became extinct. the Paleozoic assemblage of insects went into decline, and modern groups of insects began to predominate in the Mesozoic era. the Permian extinction has left a permanent mark on biodiversity today.
Geological deposits that span the boundary between the Permian and triassic periods are found in china, the Karoo Basin of South africa, and in Russia. the interpretation of the information from these deposits needed to await the refinement of radiometric dating methods. there was worldwide evidence of a massive turnover in species makeup, but only by precise radiometric dating could scien-tists be sure that this turnover had happened at the
same time all over the world at or immediately after the Permo-triassic boundary. it also awaited geo-political developments that allowed scientific teams from Russia and america, and china and america, to work together on detailed studies.
consider the geological evidence of what hap-pened at the end of the Permian, as indicated in the following paragraphs:
Anoxic deposits. One geological indicator is the dark-colored deposits at the Permo-triassic boundary. the dark color is an indicator of the lack of oxygen gas (O2) at the time and place the sediments were deposited: the iron is dark green instead of bright red, and undecomposed organic matter darkens it further to black. Such depos-its occur today at the bottoms of many ponds and swamps. Some of the deposits at the Permo-triassic boundary contain pyrites, an iron-sulfur mineral that forms only in the absence of oxygen. the worldwide black slime layer suggests world-wide anoxia, a radical event in earth’s history. today, oxygen levels in the atmosphere remain very steady at about 21 percent. Some estimates put the atmospheric oxygen content as high as 35 percent during the carboniferous. toward the end of the Permian period, however, the oxygen content may have dropped as low as 15 percent. according to geologists Raymond Huey and Peter Ward, this would have restricted vertebrates to very limited habitats at low elevation, where air pressure was highest. almost half of the land area of the earth would have been, they claim, unin-habitable by large animals. the black “death bed” layer at the end of the Permian represents between 10,000 and 60,000 years of time. Moreover, the Permo-triassic boundary is also characterized by a “chert gap.” chert is a rock formed from the ooze of billions of microscopic shelled animals accu-mulating on the ocean floor. the ooze from which chert forms is found somewhere in the deposits of almost every geological age—except at the Permo-triassic boundary.
Coarse sediments. another geological indica-tor is the deposit, during the very early triassic, of coarse sediments that were washed down violent rivers; this sometimes includes very large boulders that were moved several hundred miles or kilome-ters from their point of origin. the structure of river channels also suggests that rapid erosion was taking place, more rapid than practically anyplace on the earth now, and it was occurring everywhere. this would indicate a worldwide reduction of forest cover.
Evidence from isotopes. another geological indicator is the worldwide shift in oxygen and car-bon isotope ratios. the ratio of oxygen isotopes acts
xviii+598_Biodiversity_F.indd 407 11/17/11 1:13 PM

first entry408 permian extinction 408
as a permanent record of temperature. the seem-ingly worldwide decrease in oxygen isotope ratio suggests a massive global warming, averaging 9°F (16°c). there was also a global shift in carbon iso-tope ratios. Plant photosynthesis prefers one carbon isotope over another as it removes carbon dioxide from the air to make sugar; thus organic matter has a different carbon isotope ratio from inorganic car-bon-containing molecules. the worldwide decrease in carbon isotope ratios at the Permian boundary suggests a worldwide decrease in plant growth.
Death of terrestrial forests. During the Perm-ian, there had been fossils of plant parts from the ferns and conifers that were the dominant plants at that time. Fossil pollen was most abundant, since pollen is produced in large quantities and spreads far from the source plant and has a coat that resists decay. But at the Permo-triassic boundary, fungus spores are very abundant. this may mean that the world was rotting. Something had killed most of the plants, and mold was growing all over them. During the early triassic recovery period, plant pollen reap-peared, but it was mostly pollen from small plants such as club mosses that could grow in recently rav-aged landscapes. Usually, in every geological period (until recently), coal was forming from vast swamps of plants somewhere in the world. the sediments of the first 20 million years of the triassic, however, stood out for the absence of coal deposits. the “spike” of fungus spores and the “coal gap” are biotic indicators of a severe devastation of forest growth.
Reappearance of stromatolites. Stromatolites reappeared in the fossil record immediately after the Permo-triassic boundary. Stromatolites are lay-ers of cyanobacteria and were common in Precam-brian time but became scarce after the evolution of aquatic grazing animals. today stromatolites are found only in shallow seas where the water is too salty for most of the animals that would otherwise eat them. the reappearance of stromatolites at the end of the Permian suggests that many aquatic graz-ing animals had died.
the Permo-triassic deposits in china were formed under shallow marine (continental shelf) conditions. they contain a record of what happened to marine organisms. Most species of marine organ-isms that are preserved as fossils are microscopic plankton with calcium or other hard shells. their shells preserve well, and the organisms themselves were already living in or near the sediments. When scientists reconstructed the durations of more than 300 marine animal species from the chinese rocks, they found that 94 percent of them vanished from the record soon before, at, or soon after the boundary. Furthermore, the deposits right before the bound-
ary show evidence of extensive burrowing by a large number of marine worms; no such evidence occurs at or immediately after the boundary. therefore, toward the end of the Permian and at the beginning of the triassic, there was a collapse in marine life.
Scientists who studied the chinese deposits have pointed out that many species may appear to have vanished before the end of the Permian, simply because, by chance, their fossils were not preserved during the last few thousand years of their existence; and that many species may appear to have vanished after the end of the Permian, simply because, by chance, their fossils were washed upward from the shallow ocean floor and got mixed in with later tri-assic deposits. these arguments would suggest that the Permian extinction was a relatively sudden event, rather than spread out over a long period.
Regardless of how sudden the event was, it had a drastic effect on marine animal life. a Permian seafloor community full of coral reefs, plankton, and many invertebrate animals, including the last of the fading dynasty of trilobites, was replaced by an early triassic seafloor community consisting of very few species. For example, whole early triassic rocks are formed from small shells of just a few species of mollusks that lived right after the disaster. One of the types of animals that survived was the Lin-gula brachiopod shell mentioned earlier. its survival tells a story. it has survived 500 million years and survives today because it burrows deep in the mud, and can survive long periods with severely restricted food and oxygen. among the swimming animals, many species of bony fishes and sharks also died, to be replaced by other, smaller fish species. Both among the mollusks and the fishes, the survivors were smaller than those that had dominated the Permian seas. in short, according to paleontolo-gist Michael Benton, the survivors appeared to be those that, partly because they were small, may have needed less oxygen. Paleontologist Douglas erwin disputes the correlation between ability to tolerate anoxia and success at surviving the Permian extinction.
Meanwhile, what was happening on land? the Karoo Basin deposits in South africa formed from shallow lake sediments that washed northward from a mountain range, now worn away, in what is now antarctica. the vertebrate fossils in the late-Permian Karoo deposits include two amphibian and 72 reptile species. these diverse reptiles included some that ate plants, some that ate insects, some that ate one another; some large, some small; all part of a complex community of interacting reptile species. However, the early triassic deposits have only 28 terrestrial vertebrate species. Since most of them are not the same species that existed in the late
xviii+598_Biodiversity_F.indd 408 11/17/11 1:13 PM

permian extinction 409
Permian, this represents a drastic turnover of spe-cies composition. and rather than a diverse commu-nity of interacting species, many of the vertebrate skeletons from the early triassic are of one kind of reptile mentioned earlier: Lystrosaurus, whose generalist eat-anything niche has led some scien-tists to call it the pig of the ancient reptile world. the Karoo animal world soon after the Permian extinction was crawling with Lystrosaurus and had a few other rare reptiles. this pattern is seen in deposits not only from South africa, but also from South america, antarctica, india, china, and Rus-sia. a complex Permian community of interacting species that specialized on their means of living was replaced by a worldwide triassic landscape dominated by one kind of animal that did a little bit of everything. Overall, 27 of 29 reptile families and six of nine amphibian families became extinct. there was a similar turnover in plant species: 140 plant species, in 11 families, from the Permian were replaced by fewer than 50 species, in only three families, during the triassic.
therefore, at the end of the Permian, something killed most of the plants; partly as a result, the atmo-sphere nearly ran out of oxygen, and the landscape eroded severely. What could have caused such a severe event? Several causes have been suggested:
Loss of continental shelf area. One early sug-gestion, from the 1950s, was that smaller continents collided together to form the world supercontinent of Pangaea. Since many species of organisms known from the fossil record lived in shallow water, the collision of several continents into one would have drastically reduced the amount of coastline envi-ronment in which they lived. a massive continent, furthermore, would have had more severe terrestrial climates. Oceans ameliorate temperature extremes, which is why the summers are hotter and winters are colder in Minneapolis than in Seattle. the inte-rior of Pangaea, then, would have had extreme cli-matic conditions that would have caused at least some extinctions. Finally, species that had evolved separately might have been thrown together into a cosmopolitan mix, in which a few “winners” would outcompete “losers” that had formerly been domi-nant in their own separate lands. While it is beyond dispute that Pangaea formed during Permian times, many scientists have concluded that the resulting changes in terrestrial climate and marine habitat would not have been great enough to cause the Permian extinction. it certainly would not account for the worldwide loss of oxygen and massive death of vegetation.
Asteroid impact. could the Permian extinction have been caused by an asteroid, as was the case with the cretaceous extinction? Scientists are much
less likely to discover evidence of such an impact in deposits 250 million years old than in deposits 65 million years old, which is the time at which an asteroid almost certainly hit the earth, causing the cretaceous extinction. However unlikely it might seem, evidence of what may have been a large aster-oid impact was recently found: the Bedout crater off the coast of australia. the date of this crater corre-sponds closely to the time of the Permian extinction. Most scientists are not, however, convinced that Bedout is an impact crater.
Volcanic eruptions. a massive set of volcanic eruptions known as the Siberian traps occurred at the Permo-triassic boundary. (Traps comes from the Swedish for staircase or steps, referring to the successive layers of lava flow.) these eruptions con-tinued for a few million years. they cover an area of Siberia larger than the entire european commu-nity: 77,200 square miles (20,000 square km). the gases ejected from these eruptions could have caused worldwide devastation. Sulfur dioxide (SO2) reacts with water to produce sulfuric acid, a component today of acid rain. Severe acid rain may have killed much of the vegetation on land and photosynthetic organisms in the oceans, causing the collapse of food chains in both. Other large volcanic eruptions dur-ing earth’s history were not associated with mass extinctions, but the Siberian traps eruptions may have had a worldwide impact because they ejected more sulfur than other volcanic eruptions. the lava from these eruptions is rich in sulfur minerals, sug-gesting that these eruptions would have ejected even more sulfur dioxide than most volcanic eruptions observed today. Volcanoes also eject large amounts of carbon dioxide (cO2), a greenhouse gas that leads to global warming. the resulting death of plants could have contributed to the worldwide plunge in oxygen levels and increase in cO2. Some scientists even suggest that there was enough cO2 in the water to poison aquatic animals directly.
Release of methane. Paleontologist Michael Ben-ton suggests, though without direct evidence, that the moderate global warming caused by the volca-noes caused yet another set of catastrophes to occur: gigantic global burps. Deep underneath continental shelf sediments, especially in polar regions, there are today (and may have been during the Permian) very large deposits of methane hydrate, which is an icelike combination of water and natural gas. Pressure and cold temperature keep the methane in a solid state, but if the ocean waters became warm, the methane might have evaporated explosively, bubbling quickly through the ocean and into the atmosphere. there is evidence that such methane releases have occurred in the geologically recent past: a Paleocene erup-tion 55 million years ago apparently caused a global
xviii+598_Biodiversity_F.indd 409 11/17/11 1:13 PM

first entry410 permian extinction 410
warming of about 3–4°F (5–7°c) over a 10,000-year period. Methane, though short-lived in the atmo-sphere, is a strong greenhouse gas. By enhancing the global warming that was already going on, these methane eruptions could have started a positive-feed-back loop in which more global warming caused the release of even more methane. Methane reacts with oxygen, and this would have worsened the already dire problem of anoxia. When methane reacts with oxygen, it produces carbon dioxide, which is also a greenhouse gas.
the Permian extinction had some permanent effects on the history of life. During the Permian, brachiopod shells were relatively common and mol-lusk shells were relatively rare. However, a few million years after the extinction event, bivalve mol-lusk shells evolved into an explosive radiation of new species, while the brachiopods remained, and remain, rare. this occurred not because, under nor-mal seafloor conditions, mollusks were superior to brachiopods, but because mollusks either got lucky or because most of them were better adapted to the postdisaster world of the earliest triassic.
the implications of the Permian extinction for the future of life on earth are both good and bad. Many biologists believe that human activity is causing a sixth mass extinction. Humans are caus-ing species to become extinct at a rate as great as what occurred in previous mass extinctions. the bad news is that (1) continued destruction of veg-etation can, in fact, lead to massive soil erosion and, if this destruction continues yet further, can even contribute to a depletion of oxygen from the atmosphere. at the end of the Permian, fossil evi-dence of plants (e.g., pollen) nearly vanished and so did oxygen. there is no need to speculate that life on earth is strongly dependent on the work of plants: the “experiment” has in fact been done. the bad news is also that (2) events in the his-tory of the earth—as, in this case, normal volcanic activities—can have a severe impact on life. it does not take an asteroid to kill almost everything off. the good news is that the earth can recover from an almost total extinction event. no matter what humans do to the earth—and probably no matter what happens to it from other causes except for the final explosion of the Sun several billion years from now—the earth will recover. But this good news is also bad. it would take, from the viewpoint of human history, forever for the earth to recover. the human economy is dependent upon the continued stability and smooth operation of natural systems. For humans, even a slight change in global tempera-ture would spell agricultural and economic disaster. Because civilization depends upon the exploitation of natural resources, even the slightest interruption of these resources would send humankind, very
few of whom know how to survive in the wild, into a desperate tailspin. Humans would probably not have survived the Permian extinction.
See also greenhouse effect; mass extinc-tions; Paleogene period, biodiversity in; Permian period, biodiversity in.
Further readingBecker, luann, et al. “Bedout: a possible end-Permian
impact crater offshore of northwestern australia.” Sci-ence 304 (2004): 1,469–1,476.
Benton, Michael J. When Life Nearly Died: The Greatest Mass Extinction of All Time. london: thames and Hudson, 2003.
———, and R. J. twitchett. “How to kill (almost) all life: the end-Permian extinction event.” Trends in Ecology and Evolution 18 (2003): 358–365.
Brayard, arnaud, et al. “Good genes and good luck: ammo-noid diversity and the end-Permian mass extinction.” Science 325 (2009): 1,118–1,121.
erwin, Douglas H. Extinction: How Life on Earth Nearly Ended 250 Million Years Ago. Princeton, n.J.: Princ-eton University Press, 2006.
Huey, Raymond B., and Peter D. Ward. “Hypoxia, global warming, and terrestrial late Permian extinctions.” Sci-ence 308 (2005): 398–401.
Knoll, andrew H., et al. “comparative earth history and late Permian mass extinction.” Science 273 (1996): 452–456.
looy, c. V., et al. “life in the end-Permian dead zone.” Proceedings of the National Academy of Sciences USA 162 (2005): 653–659.
Payne, J. l., et al. “large perturbations of the carbon cycle during recovery from the end-Permian extinction.” Science 305 (2004): 506.
Retallack, Gregory J., et al. “Vertebrate extinction across Permian-triassic boundary in Karoo Basin, South africa.” Bulletin of the Geological Society of America 115 (2003): 1,133–1,152.
———, et al. “Search for evidence of impact at the Perm-ian-triassic boundary in antarctica and australia.” Geology 26 (1998): 979–982.
Visscher, H., et al. “the terminal Paleozoic fungal event: evidence of terrestrial ecosystem destabilization and collapse.” Proceedings of the National Academy of Sciences 93 (1996): 2,155–2,158.
Ward, Peter D. Under a Green Sky: Global Warming, the Mass Extinctions of the Past, and What They Can Tell Us about the Future. new york: Harpercollins, 2008.
———. “impact from the deep.” Scientific American, Octo-ber 2006, 64–71.
———, et al. “altered river morphology in South africa related to the Permian-triassic extinction.” Science 289 (2000): 1,740–1,743.
———, et al. “abrupt and gradual extinction among late Permian land vertebrates in the Karoo Basin, South africa.” Science 307 (2005): 709–714.
xviii+598_Biodiversity_F.indd 410 11/17/11 1:13 PM

permian period, biodiversity in 411
White, R. V. “earth’s biggest ‘whodunnit’: Unravelling the clues in the case of the end-Permian mass extinction.” Philosophical Transactions of the Royal Society of London 360 (2002): 2,963–2,985.
permian period, biodiversity in the Perm-ian period (290 to 250 million years ago) was the sixth and final period of the Paleozoic era. it fol-lowed the carboniferous period. at the beginning of the Paleozoic era, almost all life was aquatic; by the end of the Paleozoic era, forests filled with vertebrates and insects covered large areas of dry land. it ended with the Permian extinction, the most important of the mass extinctions in the history of the earth.
Climate. the climate of the Permian was cooler and drier than that of earlier periods of the Paleo-zoic, especially in the center of the massive continent of Pangaea. today, deserts with extreme tempera-tures are found in the middle of the eurasian con-tinent and were probably found in the middle of Pangaea as well. Glaciation occurred in the south-ern portions of Pangaea. then global warming, caused initially by volcanic eruptions, occurred near the end of the Permian Period, which contributed to massive extinctions (see table). in connection with the Permian extinction, atmospheric oxygen levels became low enough that, according to some investigators, large terrestrial animals had difficulty breathing.
Continents. all of the continents collided to form one massive continent called Pangaea.
Marine life. Diversity of marine animal species remained as high as it had been during the carbon-iferous period, but declined rapidly with the onset of the Permian extinction. the groups of organisms that recuperated from the extinction more rapidly became the dominant groups up through the pres-ent time. the Permian extinction therefore largely determined the pattern of biodiversity for the entire subsequent history of the earth.
● Invertebrates. Some invertebrates that are now rare, such as stalked echinoderms, shelled cephalopods, and brachiopods, were common in the Permian period, but their diversity was strongly reduced by the extinc-tion event. Bivalves, cephalopods without shells, and echinoderms without stalks became common after the Permian extinc-tion, while brachiopods, shelled cephalo-pods, and stalked echinoderms did not. the trilobites had gradually declined from being the dominant marine invertebrates from the cambrian period onward and did not sur-vive the extinction event.
multiCellular biodiversity in the permian period
Conditions
Atmospheric oxygen 15% above modern level before extinction event
Atmospheric carbon dioxide 2.5 times modern level
Global temperature 3°F (2°C) above modern level
Sea level 170 feet (60 m) higher than today before extinction event
Approximate number of families*
Continental 200
Marine 450
Total 650
Examples of marine animals
Acropyge sp. trilobite
Caricula salebrosa brachiopod
Cheirolepis sp. bony fish
Cyclolobus walkeri ammonite mollusk
Helicoprion bessonovi shark
Mamayocaris sp. crustacean
Myalina lamellosa bivalve mollusk
Panduralimulus babcocki horseshoe crab
Platycrinites omanensis crinoid echinoderm
Vermiforinchus sp. annelid
Examples of continental animals
Apthoroblattina sulcata cockroach
Archosaurus rossicus archosaur-like reptile
Dimetrodon grandis mammal-like reptile
Ectosteorhachis nitidus lobe-fin fish
Eryops megacephalus amphibian
Gnathorhiza sp. lungfish
Permocoleus wellingtonensis beetle
Permotipula patricia fly
Zygosaurus lucius amphibian
Examples of land plants
Cassinisia orobica conifer
Crossozamia sp. cycad
Emplecopteris triangularis seed fern
Ginkgo baiera ginkgo
Glossopteris sp. seed fern
Majonica alpina conifer
Titea singularis tree fern
*Based on Benton.
xviii+598_Biodiversity_F.indd 411 11/17/11 1:13 PM

fi rst entry412 permian period, biodiversity in 412
● Vertebrates. Diversity continued to be high among the fi shes. Sharks were abundant and diverse, including Helicoprion, which had a coil of teeth in its lower jaw. Bony fi shes continued to diversify.
Continental life. Forests and wetlands covered much of the land surface at the beginning of the Permian period.
● Plants. the tree-sized horsetails and club mosses that had characterized the “coal swamps” of the carboniferous were close to extinction by the middle of the Permian, replaced by seed ferns (a type of plant now extinct) and by conifers, which fi rst became abundant during the Permian. this proba-bly occurred because the conifers were bet-ter adapted to the cooler, drier conditions of the Permian. the seed fern Glossopterisdominated entire forests. Some seed ferns were woody vines. the fi rst cycads evolved during the Permian, as did the ginkgoes, although both groups became much more abundant and diverse during the Mesozoic era. there were no fl owering plants.
● Invertebrates. among the most abundant terrestrial invertebrates were cockroaches. large dragonfl ies remained common, as in the preceding carboniferous period. though they were rare, the fi rst beetles and fl ies evolved during the Permian.
● Vertebrates. Freshwater bony fi shes, lung-fi shes, and amphibians continued to evolve. Eryops was a large Permian amphibian. the drier conditions encouraged the diversifi ca-tion and spread of reptiles, which had fi rst appeared in the carboniferous (see fi gure). there were two major groups of Permian reptiles: the synapsids and the diapsids. there were no dinosaurs, but some of the reptiles of the Permian were rather large. none of these reptiles were of recognizably modern groups; however, the diapsids were the ancestors of most of the modern ani-mals commonly called reptiles, as well as extinct reptiles such as archosaurs and dino-saurs. the diapsids were also the ancestors of birds. the synapsids are today extinct except for one line of descendants: mam-mals. Some Permian synapsids grew as large as hippos. the most famous Permian synap-sid was Dimetrodon, whose ribs formed a huge “sail” along its back. Some synapsids had many mammal-like characteristics; for example, unlike diapsids and modern rep-tiles, they had several kinds of specialized teeth. they may also have had whiskers: their skulls have small nerve openings where these whiskers would have been. if they had whiskers, they had hair, and therefore were probably warm-blooded. the dissipation of body heat may have been the main function of the sail along the back of Dimetrodon.
in other words, although these synapsids were not mammals, they closely resembled them. a nearly complete set of synap-sid transitional forms has been found between reptiles and mammals.
Extinctions. the Permian extinction brought many evo-lutionary lineages to an end and severely reduced the abundance of many others. Scientists will never know what might have happened if evolution could have continued uninterrupted. Would the synap-sids have developed into the domi-nant vertebrates, ushering in the age of mammals 250 rather than 65 million years ago? Would this have prevented the evolution of the dinosaurs? these questions will never be answered.
See also Permian extinction.
Dimetrodon was a large mammal-like reptile that lived during the Perm-ian period. The large sail of skin tissue may have allowed Dimetrodon to absorb heat from sunlight during cold weather and radiate body heat dur-ing warm weather.
xviii+598_Biodiversity_F.indd 412 11/17/11 1:13 PM

pinchot, Gifford 413
Further readingBenton, Michael J. “Diversification and extinction in the
history of life.” Science 268 (1995): 52–58.———. “Four feet on the ground.” in The Book of Life: An
Illustrated History of the Evolution of Life on Earth, edited by Stephen J. Gould. new york: norton, 2001.
Brayard, arnaud, et al. “Good genes and good luck: ammo-noid diversity and the end-Permian mass extinction.” Science 325 (2009): 1,118–1,121.
Krings, Michael, et al. “How Paleozoic vines and lianas got off the ground: On scrambling and climbing car-boniferous–early Permian pteridosperms.” Botanical Review 69 (2003): 204–224.
pinchot, gifford (1865–1946) American Con-servationist Gifford Pinchot was born august 11, 1865, into a wealthy Pennsylvania family that had made its fortune from clear-cutting the eastern deciduous forest. it is ironic that he became one of the major figures in forest and biodiversity conser-vation. it was at the suggestion of his father, who regretted the damage his family had done to the land, that Gifford Pinchot attended yale University and studied forestry. Gifford Pinchot became chief of the Division of Forestry in the Department of the interior in 1898. the Pinchot family endowed the yale School of Forestry in 1900 to train professional forest managers.
at that time, the U.S. federal government owned thousands of square miles of western forests that had recently been taken from native american ownership and was engaged primarily in giving for-est lands to private railroad, lumber, and mining corporations at minimal charge. When progressive Republican theodore Roosevelt became president in 1901, after the assassination of William McKin-ley, he decided to end the practice of wholesale pub-lic support of private corporations. Both Roosevelt and Pinchot extensively toured western forests and were acquaintances of conservationist John Muir. Based on field information, Pinchot and Roosevelt selected thousands of square miles of forestland to set aside in permanent federal ownership. in 1905, Roosevelt established the U.S. Forest Service of the Department of agriculture to manage these for-ests by leasing them to private corporations, who could make a profit from the renewable resources in return for paying a fee to the government. Pinchot was its head.
although Pinchot is considered one of the found-ing figures in american conservation, his approach was quite different from that of preservationists such as John Muir. Unlike Muir, Pinchot was not inter-ested so much in the preservation of natural habi-tats and wildlands as in the management of federal
lands for sustainable production of timber and other industrial resources. Muir’s view was that conserva-tion was the right thing to do; Pinchot’s view was that it could be profitable in the long term. this was perhaps best illustrated by Pinchot’s support for the damming of Hetch Hetchy Valley near yosemite in the Sierra nevada of california, creating a reservoir for San Francisco, which Muir vigorously opposed. nevertheless, Pinchot had a passion, not merely a business appreciation, for conservation: He deplored the destruction of forests as what he called a gigan-tic massacre, in which waste was a virtue. largely because of Pinchot’s early leadership, the Forest Ser-vice has proven to be an invaluable force in support of conservation and biodiversity, not just of sustain-able timber production.
although Pinchot and Roosevelt set aside much land for the express purpose of leasing it to private timber and mining corporations, these corporations and the politicians whom they funded wanted to cut down all the forests and sell the products as quickly as possible. Roosevelt and Pinchot, progres-sive Republicans, received much political opposi-tion from the reactionary branch of the Republican Party, who led vigorous efforts to restrict funding to the Forest Service. in 1907, a congressional bill forbade the further creation of government forest preserves. in the last days before the bill took effect, Roosevelt and Pinchot set aside 16 million acres (65,000 square km) of forestland.
Under Pinchot’s leadership, yale forestry gradu-ates received not only scientific but also practical and physical training. nevertheless, these forestry graduates from the eastern states were not entirely prepared for the bars-and-brothels culture of the frontier towns of idaho and Montana. the miners and lumberjacks did not take the foresters or the professors who trained them seriously. a hunter shot and killed a government forester in Montana in 1907. One congressman called the Forest Service “google-eyed, bandy-legged dudes from the east and sad-eyed, absent-minded professors and bugologists.” Furthermore, private lumber and mining corpora-tions continued to take control of government for-estlands by two means: First, they simply trespassed, knowing that an underfunded Forest Service would not be able to document the illegal exploitation of government land; second, they paid homesteaders to take possession of tracts of land (which remained legal on government property) and immediately sell the land to the corporations. the foresters, who were nearly powerless to do their jobs, as well as the administrators of the Forest Service sometimes had to depend on personal contributions from Pinchot’s family wealth in order to get paid. Pinchot himself worked without a salary.
xviii+598_Biodiversity_F.indd 413 11/17/11 1:13 PM

first entry414 placental mammals 414
theodore Roosevelt did not run for reelection in 1908. His handpicked successor, William Howard taft, who had promised to continue the work of con-servation, won the election. taft, however, did not carry through on his promise, but instead supported the politicians who wanted the Forest Service to be made as powerless as possible. congress even passed a law in 1910 forbidding the Forest Service from publicizing itself. Pinchot published articles critical of taft, who fired him in 1910. the U.S. Forest Ser-vice seemed destined for obscurity.
then, in summer 1910, a huge forest fire broke out in the mountains of Montana and idaho, rapidly burning hundreds of square miles and threatening all of the major cities of the region. it was up to the Forest Service, with some help from a contingent of black soldiers, to bring the fire under control and protect the human inhabitants. Both the foresters and the soldiers undertook heroic efforts to prevent the spread of the fire and to evacuate the inhabitants. about 100 foresters lost their lives in the fire. Once the fire was contained, publicity of the foresters’ heroism raised the esteem of the Forest Service and may have guaranteed the survival of the agency. the heroism of the black soldiers is also credited with advancing racial equality in the United States. a few critics of the Forest Service, and of conservation in general, remained. Senator Weldon Heyburn, who had long been the Forest Service’s most bitter enemy, said that there would have been no fire at all if the lumber corporations had been allowed to cut down all of the trees.
Gifford Pinchot may have been the first promi-nent person to articulate a philosophy of conser-vation from a government perspective. the whole point of conservation, to Pinchot, was to protect natural resources from immediate plunder by a few, so that they would be available for succeeding gen-erations. the earth, said Pinchot, “belongs of right to all its people, and not to a minority, insignificant in numbers but tremendous in wealth and power.” later, theodore Roosevelt expanded Pinchot’s view, referring to “the central task of leaving this land even a better land for our descendants than it is for us.” conservation, said Roosevelt, “is a great moral issue!” their political adversary, Senator William a. clark, openly proclaimed the opposite view: “those who succeed us can well take care of themselves.” President taft said that Pinchot was “championing the cause of the oppressed and the downtrodden, and harassing the wealthy and the greedy and the dishonest.” taft apparently intended this as a criti-cism, but Pinchot’s view prevailed.
Pinchot remained active throughout his life as a speaker and writer in support of conservation. Pinchot, largely from his experience with the Big
Burn of 1910, believed that all forest fires were bad and had to be stopped, an approach that ecolo-gists now know was mistaken. nevertheless, his influence remained powerfully important, and he advised President Franklin Delano Roosevelt about conservation. He proposed, and Franklin Roosevelt approved, plans for an international conservation conference, to take place during the reconstruction after the Second World War. Both Roosevelt and Pinchot died before these plans could be put into action. Pinchot twice became governor of Pennsylva-nia. He died October 4, 1946.
See also disturbances, ecological; federal government role in conservation (United States); Muir, John; succession, ecological.
Further readingBalogh, Brian. “Scientific forestry and the roots of the
modern american state: Gifford Pinchot’s path to Pro-gressive reform.” Environmental History 7 (2002): 198–225.
egan, timothy. The Big Burn: Teddy Roosevelt and the Fire That Saved America. Boston: Houghton Mifflin, 2009.
placental mammals See mammals.
pleistocene epoch See Quaternary period, biodiversity in.
pleistocene extinction During the latter part, or immediately after, the Pleistocene epoch of the neogene period, many species of large mam-mals became extinct at various locations around the world. not without controversy, this extinction event has been attributed at least in part to hunting by humans.
at the end of the most recent ice age, north america was the home of a much greater range of large mammals than is known from historical times (see table on page 415). For example:
● in addition to grizzly, black, and brown bears, there was also a short-faced bear.
● in addition to cougars, there were lions and cheetahs resembling african species and a saber-toothed cat.
● in addition to the extant species of beavers, bison, and wolves, there was a larger species of each.
● there were large mammal species otherwise known only from the Old World, such as camels and horses.
xviii+598_Biodiversity_F.indd 414 11/17/11 1:13 PM
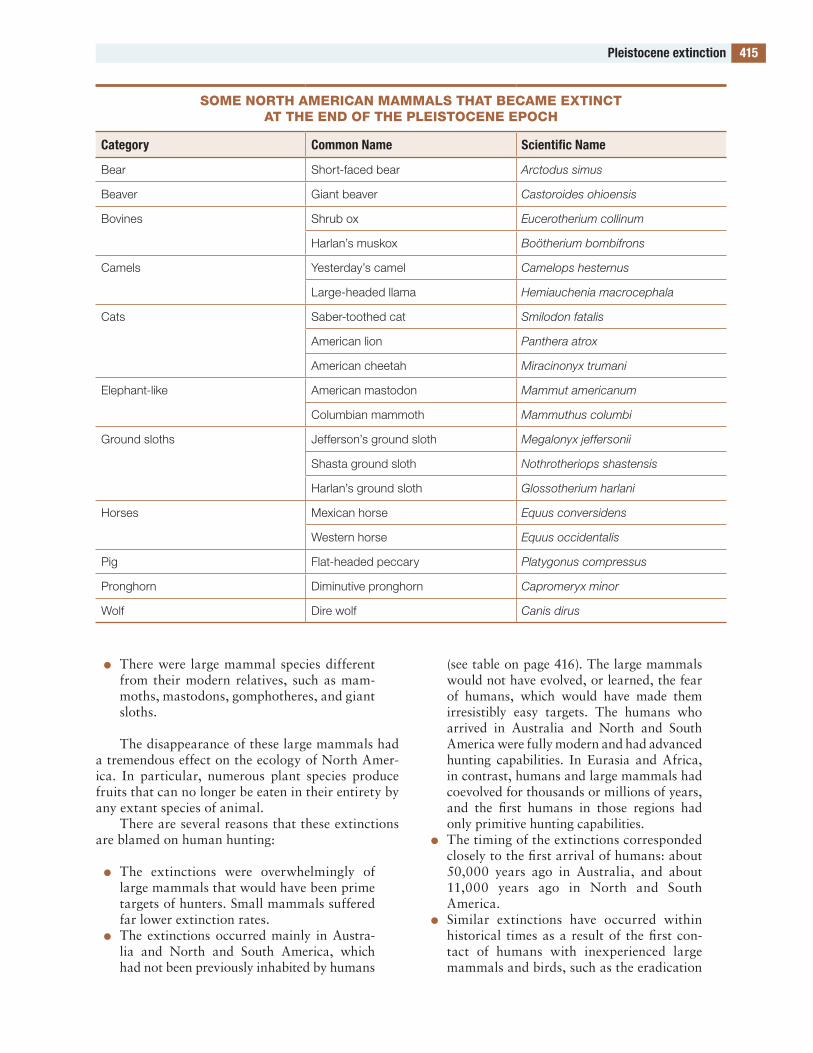
pleistocene extinction 415
● there were large mammal species different from their modern relatives, such as mam-moths, mastodons, gomphotheres, and giant sloths.
the disappearance of these large mammals had a tremendous effect on the ecology of north amer-ica. in particular, numerous plant species produce fruits that can no longer be eaten in their entirety by any extant species of animal.
there are several reasons that these extinctions are blamed on human hunting:
● the extinctions were overwhelmingly of large mammals that would have been prime targets of hunters. Small mammals suffered far lower extinction rates.
● the extinctions occurred mainly in austra-lia and north and South america, which had not been previously inhabited by humans
(see table on page 416). the large mammals would not have evolved, or learned, the fear of humans, which would have made them irresistibly easy targets. the humans who arrived in australia and north and South america were fully modern and had advanced hunting capabilities. in eurasia and africa, in contrast, humans and large mammals had coevolved for thousands or millions of years, and the first humans in those regions had only primitive hunting capabilities.
● the timing of the extinctions corresponded closely to the first arrival of humans: about 50,000 years ago in australia, and about 11,000 years ago in north and South america.
● Similar extinctions have occurred within historical times as a result of the first con-tact of humans with inexperienced large mammals and birds, such as the eradication
some north ameriCan mammals that beCame extinCt at the end of the pleistoCene epoCh
Category Common Name Scientific Name
Bear Short-faced bear Arctodus simus
Beaver Giant beaver Castoroides ohioensis
Bovines Shrub ox Eucerotherium collinum
Harlan’s muskox Boötherium bombifrons
Camels Yesterday’s camel Camelops hesternus
Large-headed llama Hemiauchenia macrocephala
Cats Saber-toothed cat Smilodon fatalis
American lion Panthera atrox
American cheetah Miracinonyx trumani
Elephant-like American mastodon Mammut americanum
Columbian mammoth Mammuthus columbi
Ground sloths Jefferson’s ground sloth Megalonyx jeffersonii
Shasta ground sloth Nothrotheriops shastensis
Harlan’s ground sloth Glossotherium harlani
Horses Mexican horse Equus conversidens
Western horse Equus occidentalis
Pig Flat-headed peccary Platygonus compressus
Pronghorn Diminutive pronghorn Capromeryx minor
Wolf Dire wolf Canis dirus
xviii+598_Biodiversity_F.indd 415 11/17/11 1:13 PM

first entry416 pleistocene extinction 416
of the moa birds by the first Polynesians in new Zealand, the eradication of elephant birds and giant lemurs by the first immi-grants to Madagascar, and the eradication of passenger pigeons and near-eradication of bison by white americans.
Difficulties have been noted, and some responses formulated, regarding this Pleistocene overkill hypothesis:
● in north america, the arrival of humans was not the only thing that was happen-ing 11,000 years ago. the most recent ice age was also ending, accompanied by rapid climatic changes. tundra and grass-lands retreated, and spruce forests spread. Fires also became more frequent. Defend-ers of the overkill hypothesis point out that the climate was becoming warmer, which should have encouraged the survival, not the extinction, of animals. the abundance and diversity of animals during the ice ages, however, suggest that a warmer climate was not necessarily an improvement for these animals. any climatic change that would have altered the food base for these animals may have undermined their populations.
● evidence is lacking for massive kills of large mammals. arrowheads have been found in mammoth bones, but only occasionally, and mass grave sites of large animals are very uncommon.
● native americans, on whom the Pleistocene overkill in the new World is blamed, do not have a mass-slaughter approach to hunt-ing. in fact, some native american tradi-tions are famous for spiritual preparation and gratitude before killing an animal and careful use of the animal carcass. However, the native americans that the europeans encountered beginning just before the 16th century were perhaps very different from those that arrived in america at the end of the last ice age. in particular, native amer-icans may have developed a conservation ethic in response to the collapse of the hunt-ing stock caused by their careless ancestors.
Paleontologist anthony Barnosky performed a thorough analysis of the Pleistocene extinctions around the world. He confirmed that large mam-mals died in disproportionately large numbers and at times that corresponded (within a thousand years) to human arrival. He concluded that neither cli-mate change nor overhunting could by themselves
have caused the extinctions. together, however, they could have produced the Pleistocene extinctions. it would not have been necessary for either climate or hunters to have killed every last animal. Once a population becomes sufficiently small, it becomes vulnerable to extinction. Overhunted animal popu-lations were vulnerable to extinction from climate changes, and climate changes may have reduced ani-mal populations to the point that hunting could send them spiraling to oblivion.
in 2008, some scientists announced the discov-ery of diamond fragments in Pleistocene sediments. they claimed that only an impact such as that of an asteroid could have produced such diamonds. this evidence raised the possibility that an aster-oid impact may have contributed to the Pleistocene extinction. although this event, if it occurred, may have had considerable local impact and may have been responsible for a sudden reversal of warming at the end of the last ice age, it is unlikely to explain the worldwide pattern described by Barnosky. after the initial publication, further evidence of a Pleistocene asteroid was not forthcoming, and most scientists have lost interest in it.
all of the above processes depend upon the assumption that the precipitous decline in large mammal populations began after the climate and vegetation began to change and after the arrival of modern humans that had advanced stone tools for hunting, in particular the clovis arrowhead points (named after a site in new Mexico). Research pub-lished by paleoecologist Jacquelyn Gill and associ-ates in 2009, however, took a careful look at the sediment deposits in lakes in the eastern United States. they quantified three components of the sed-iments: pollen, an indicator of vegetation type; char-coal, an indicator of fire; and spores of the fungus Sporormiella, an indicator of the abundance of graz-ing mammals. this fungus is known to grow only in the dung of herbivorous mammals. the research-
perCent of mammal genera beComing extinCt upon arrival
of first humans*
Continent Extinction
Africa 18%
Australia 88%
Eurasia 36%
North America 72%
South America 83%
*From Barnosky
xviii+598_Biodiversity_F.indd 416 11/17/11 1:13 PM

pollination 417
ers found that the decline in mammal abundance began before the changes in vegetation, not after, and it began before the arrival of the clovis people. they concluded that it was the decline of the mam-moths and mastodons that caused the vegetation change, not the other way around. the mammoths and mastodons were keystone species that had kept the trees browsed down, while grasses could readily sprout back after grazing. they also speculated that the decline in large mammals was caused by hunt-ing, but not by the clovis people. their predecessors had killed many large mammals. the clovis people developed advanced arrowheads and more efficient hunting because the large mammals were becoming rare, not the other way around.
as the earth enters into what many scientists consider the sixth mass extinction as a result of human activity, the lessons of the Pleistocene extinc-tions may be essential. now, as then, it is not neces-sary to kill many animals to drive them to extinction. at the end of the Pleistocene, climate change reduced their habitats and made them vulnerable to hunt-ing. today, habitat destruction for agriculture and human habitation is the principal threat to the sur-vival of most endangered species.
See also mass extinctions; neogene period, biodiversity in.
Further readingBarnosky, anthony D. “assessing the causes of late Pleis-
tocene extinctions on the continents.” Science 306 (2004): 70–75.
Donlan, c. J., et al. “Pleistocene rewilding: an optimistic agenda for twenty-first century conservation.” Ameri-can Naturalist 168 (2006): 660–681.
Gill, Jacquelyn l., et al. “Pleistocene megafaunal collapse, novel plant communities, and enhanced fire regimes in north america.” Science 326 (2009): 1,100–1,103. Summarized by Johnson, christopher. “Megafaunal decline and fall.” Science 326 (2009): 1,072–1,073.
Guthrie, R. Dale. “Rapid body size decline in alaskan Pleis-tocene horses before extinction.” Nature 426 (2003): 169–171.
Harris, John M. “Bones from the tar pits.” Natural History, June 2007, 18–22.
Kennett, Douglas J., et al. “Shock synthesized hexago-nal diamonds in younger Dryas boundary sediments.” Proceedings of the National Academy of Sciences USA 106 (2009): 12,623–12,628.
Martin, Paul S. Twilight of the Mammoths: Extinctions and the Rewilding of America. Berkeley: University of california Press, 2005.
———, and christine R. Szuter. “Revising the ‘wild’ west: Big game meets the ultimate keystone species.” chap-ter 3 of The Archaeology of Global Change: The Impact of Humans on Their Environment, edited by
charles l. Redman et al. Washington, D.c.: Smithson-ian institution Press, 2004.
Miller, Gifford H., et al. “ecosystem collapse in Pleistocene australia and a human role in megafaunal extinction.” Science 309 (2005): 287–290.
Mosimann, J. e., and Paul S. Martin. “Simulating over-kill by Paleoindians.” American Scientist 63 (1975): 304–313.
Pielou, e. c. After the Ice Age: The Return of Life to Glaci-ated North America. chicago: University of chicago Press, 1992.
Steadman, David W., et al. “asyncronous extinction of late Quaternary sloths on continents and islands.” Proceedings of the National Academy of Sciences USA 102 (2005): 11,763–11,768.
pliocene epoch See neogene period, biodi-versity in.
pollination Pollination occurs when pollen is car-ried from the male cones or stamens of one seed plant to the female cones or stigmas of another plant. Seed plants benefit from cross-pollination for the same rea-sons that all species benefit from cross-breeding within genetically diverse populations. even though the pol-len may land on the female reproductive structures of the same plant, pollen will often not germinate and grow unless it lands on the female reproductive struc-tures of a genetically different individual.
cones of most conifers, and flowers that are pollinated by the wind, often release their pollen in the early springtime before the leaves of most plant species have expanded. For example, in trees such as cottonwood and oak, flowers emerge from buds first, and leaves emerge only after pollination is completed. in this way the leaves do not block the movement of pollen to the stigmas. Male cones and wind-pollinated flowers must produce massive amounts of pollen, as the wind does not carry the pollen in any particular direction.
animal-pollinated flowers, in contrast, have a more reliable vector for pollination, as animals can consciously direct their movement from one flower to another of the same species. Because they do not waste as much pollen, these flowers do not produce as much pollen. animal-pollinated flowers have pet-als, often brilliantly colored, and attract animals with nectar and pollen for food. the flowers may have conspicuous lines (sometimes visible only in the ultraviolet) that guide the pollinating insect to the middle of the flower where the nectar is located (see photograph on page 418). Wind-pollinated flow-ers do not generally produce nectar or have petals, which would only block the movement of pollen.
xviii+598_Biodiversity_F.indd 417 11/17/11 1:13 PM

first entry418 pollination 418
it is important that pollen be carried from one flower to another of the same species. if flowers of many species were indiscriminately pollinated by animals, much of a flower’s pollen would be wasted on the stigmas of other species. Some species of flow-ers have characteristics that make them available to a wide range of pollinators. these characteristics include a round shape (radial symmetry), reproduc-tive organs that are easily accessible, and a hori-zontal orientation. But many other species of plants have evolved flowers that mainly attract specialized pollinators. For example:
● Some flowers consist of a long tube too nar-row for bees to climb into; the nectar can be reached only by animals with long tongues, such as butterflies or hummingbirds.
● Many flowers that hummingbirds special-ize on are bright red, a color that attracts hummingbirds.
● the tubular petals of some flowers consist of an upper and lower lip that remain tightly closed unless a bee lands on the lower lip, pushing the tube open with its weight. a small fly would not weigh enough to push such a tube open.
● Flowers pollinated by moths or bats open at night and are light colored to reflect light (see figure).
these specializations are not perfect, however. the red, tubular flowers of Silene virginica look perfectly like hummingbird flowers, but it turns out that bumblebees are their most common pollinators.
animals, by specializing on certain flowers, are able to exploit a reliable source of nectar and pollen. therefore pollinator specialization benefits both the plant and the animal. natural selection has favored plants whose flowers successfully attract pollina-tors as well as pollinators that successfully obtain food from flowers, resulting in coevolution. Many flowers can be pollinated by more than one kind of animal. Bees have been observed to remove pol-len from protruding stamens at the same time that moths obtained nectar from deep inside the flowers of the evening primrose Oenothera rhombipetala in Oklahoma.
Flowers often have intricate adaptations to assure that pollen gets properly carried by animals. For example:
● in some flowers, the weight of the pollina-tor causes the stamens to bend over and rub pollen onto the insect’s back.
● in some species of plants, the flowers come in two forms. in one form, the stamens are
Many flowers have nectar guides, which are colored lines directing the attention of pollinators to the center of the flower, where pollen and nectar are found. The beetle visiting this Geranium viscosissiumum in the Black Hills of South Dakota may be a pollinator or may just be drinking nectar without carrying pollen to another flower. (Stanley A. Rice)
Cactus flowers are often pollinated by bats. (Merlin D. Tuttle, Bat Conservation Society)
xviii+598_Biodiversity_F.indd 418 11/17/11 1:13 PM

pollination 419
short and the styles are long; in the other form, the stamens are long and the styles are short (see fi gure). a fl ower with long stamens places pollen on the rear of the insect, while a fl ower with short stamens places pollen on the front of the insect. When the insect visits fi rst a fl ower with short stamens and then a fl ower with short styles, the front of the insect will rub pollen onto the second fl ower’s short style. if the insect visits fi rst a fl ower with long stamens and then a fl ower with long styles, the rear of the insect will rub pollen onto the second fl ower’s long style.
● Some fl owers have pollen in packets, which they attach to the pollinator insect.
animal pollinators would benefi t from fl owers that provide copious amounts of nectar. However, fl owers do not typically do this. the reproductive interests of the fl owers are best served by keep-ing the pollinators on the move. not fi nding quite enough nectar in one fl ower, the pollinator moves on to the next, perhaps on a different plant. the nectar of some fl owers contains chemicals that irri-tate the pollinator. For example, the wild tobacco Nicotiana attenuata produces nectar with benzyl-acetone, which attracts pollinators, and nicotine,
which repels them. the nicotine keeps the pollina-tors from staying too long at one fl ower.
nectar is not the only fl oral product that pol-linators use. Bees scrape much of the pollen off their bodies and use it to make bee bread. Bees that polli-nate a central american bucket orchid, Coryanthes panamensis, gather wax that is a component of their own sex attractants.
Flowers and pollinators do not always benefi t one another. Some animals are nectar thieves, and some plants have evolved fl oral mimicry.
Nectar thieves. animals visit fl owers to obtain food; they pollinate the fl owers more or less by accident. in some cases, insects will bore holes into the backs of fl owers to obtain nectar that would be unavailable to them had they tried to crawl in from the front. Such “nectar thieves” consume a large share of the nectar in many species of fl owers. natural selection favors pollinators that successfully obtain food from fl owers, whether they pollinate them or not. in some cases, an insect may be both a pollinator and a thief. nectar-thief bumblebees have been observed to pierce the bases of Oenothera rhombipetala fl owers in Oklahoma and drink the nectar. But because they approached the fl owers from the front, their abdomens came in contact with the stamens and stigma anyway.
Some species of fl owers have two or more forms. In this example, short stamens deposit pollen on the back of the bee, which carries it to a fl ower that has short pistils. The short-pistil fl owers have long stamens, which deposit pollen on the bee’s abdomen. When the bee visits a short-stamen fl ower, its abdomen deposits pollen on the long pistils.
xviii+598_Biodiversity_F.indd 419 11/17/11 1:13 PM

first entry420 pollination 420
Floral mimicry. Sometimes flowers deceive ani-mals into transporting their pollen without reward-ing them. For example:
● Flies are attracted by the stench of rotting meat, and fly-pollinated flowers produce nectar with just such an aroma. Many fly-pollinated flowers open near the ground and have markings that resemble those of a car-cass. the flies visit and pollinate these flow-ers, but receive no food. numerous flower species, representing several unrelated plant families, have evolved this mode of pollination.
● Several species of orchids are pollinated by male wasps. the flowers of most of these orchids visually resemble female wasps and even produce nectar that resembles the female wasp’s sex attractant. a young male wasp copulates with the orchid, believing it to be a female wasp, and gets dusted with pollen. the wasp then makes the same mis-take again, copulating with another orchid, delivering pollen from the first orchid to the stigma of the second. in some orchids, the flowers resemble hovering male wasps, and these are pollinated by the real male wasps that attack them.
● Some species of orchids, which themselves do not produce nectar, resemble lilies that do produce nectar. young bees, which have not yet learned the details that distinguish the flowers that have nectar from those that do not, will often visit these orchids.
● the small, drab flowers of the lily, Scoliopus bigelovii, grow in dark, moist forests. they smell like mushrooms and are pollinated by fungus gnats.
these are called floral mimicry because the flowers imitate the appearance and even chemistry of corpses, female insects, or other flowers. this pro-cess is reminiscent of the evolutionary mimicry by which one animal species mimics another. natural selection favors flowers that successfully attract pol-linators, whether they reward them or not.
in some cases, the bad luck of the deluded pol-linator may contribute yet further to the benefit of the flowers. Male Lyssopimpla excelsa wasps eject sperm into Cryptostylis tongue orchids. When these males subsequently mate with female wasps, little or no sperm remain. in the reproductive system of many ants, bees, and wasps, unfertilized eggs develop into males. the female wasps therefore lay eggs that develop into males, which hatch and seek out more tongue orchids.
ThE STudy of pollINaTIoNPollination is one of the most intensively studied nat-ural phenomena. the traditional method of study-ing pollination is for a human observer to watch a flower and record data about how many pollinators, of which species, visit a flower during a period of time. a pollinator visit does not mean successful pol-lination, however. in many pollination studies, the investigator captures the pollinator as it leaves and microscopically examines it to estimate how much pollen, and of which plant species, it carries. in some cases, as in the study by Karen Bowers, the inves-tigator uses fluorescent powder in place of pollen, thereby tracing the exact route of the pollinator. the effectiveness of pollination can be experimentally manipulated by surrounding flowers with a mesh bag that keeps the pollinators away.
More recently, researchers have begun to use genetically modified plants to study pollination. ecologist Danny Kessler and associates used geneti-cally modified wild tobacco plants (Nicotiana attenuata) that either produced or did not produce benzylacetone (an attractant) or nicotine (a repel-lant) (see above). as a control, they used genetically modified plants that had normal production of these two chemicals. in this way, they experimentally demonstrated the effectiveness of the two nectar components.
pollINaTIoN aNd EvoluTIoNPollination has had a tremendous impact upon the evolution of biodiversity. the flowering plants evolved near the beginning of the cretaceous period. this event opened up new possibilities for coevolution. Flowering plants and pollinating insects (especially the hymenopterans and lepidop-terans) underwent explosive speciation. the coni-fers, which dominated many landscapes prior to the origin of flowering plants, are usually pollinated by the wind, but scorpion flies pollinated some early conifers as long as 320 million years ago, 200 million years before the origin of flowering plants. the origin of flowering plants was not the origin of animal pollination, but of countless new pollina-tion symbioses.
Pollination may also contribute to speciation. a slight change in floral nectar chemistry might cause a small population of flowers to be attractive to a different pollinator, thus isolating it from other members of its species. Fig populations that are pol-linated by different species of wasps are reproduc-tively isolated from one another.
pollINaTorS IN aGrICulTurE aNd CoNSErvaTIoNPollination is not just something that happens out in the wild. Many important agricultural crops (such
xviii+598_Biodiversity_F.indd 420 11/17/11 1:13 PM

pollination 421
as fruits and nuts) depend on pollinators. about every third bite of food comes from a domesticated plant that depends on animal pollination. the hon-eybee Apis mellifera, a species introduced from the Old World, pollinates most american crops. in 2007, many honeybee hives suffered colony collapse disorder (ccD), in which the workers left and did not return, leaving the queen and grubs to starve. a significant association was found between ccD and a type of virus from eurasia, but it was not clear whether the virus causes ccD or whether ccD was caused by parasitized honeybees that were already suffering from some other stress. Because honeybee hives are trucked all over the United States (about half of them are taken to california during the almond pollination season), diseases can spread rap-idly among their populations.
an alternative to reliance on eurasian honey-bees is to encourage wild pollinator species to take the place of honeybees. Wild pollinators can be more effective than honeybees because they are often spe-cialists, while honeybees are generalists. For exam-ple, about 750 blue orchard bees (Osmia lignaria) can pollinate the same number of apple or almond trees as roughly 100,000 honeybees. the leaf-cut-ting bee (Megachile rotundata) is a very effective pollinator of alfalfa, and the bee Osmia ribifloris is especially effective in pollinating blueberries. Fur-thermore, wild pollinators work in a greater range of weather conditions than do honeybees. nearly every animal-dependent crop species, even those not native to north america, is visited by a native (wild) pollinator.
Honeybees will remain the major agricultural pollinator. they have strength in numbers, as there can be 100,000 bees in one hive, compared to wild pollinators, which may be solitary breeders. On the other hand, diseases are less likely to spread among wild pollinators, and there is less chance that any one disease can disrupt crop production if many wild pollinator species are available. However, to increase reliance upon wild pollinators, it will be necessary to reduce the use of pesticides. Pesticides do not harm honeybees as much as wild pollinators, since honey-bee hives can be closed up and moved out of the way when pesticides are applied. Other relatively simple measures can be taken to promote the populations of wild bees. the bee Centris tarsata pollinates wild cashews but not cashews in commercial orchards. this bee needs floral oils that cashew trees cannot provide. But if caribbean acerola cherries (which produce a lot of vitamin c and are themselves a commercial crop) are interspersed with the cashews, the wild bees survive very well. Wild pollinators can also be encouraged by having smaller, more diverse farms. Wild species perform up to 62 percent of the
pollination on the small farms of the Delaware River Valley in the northeastern United States, but are very rare in the central Valley of california, where fields are large and drenched with pesticides. Populations of wild pollinators can be supported by the pres-ence of hedgerows and wildflower belts around farm fields. Smaller and more diverse farms offer many economic and ecological benefits, such as reduced dependence on pesticides, in addition to the encour-agement of wild pollinators.
Wild pollinators are also essential for the sur-vival of many wild plant species. conservationists have preserved giant cactuses in national parks in the southwest, but their pollinators have become rare. the survival of these cactuses into their next generation, therefore, is endangered.
See also coevolution; flowering plants (angiosperms); mutualism; speciation.
Further readingBeismeijer, J. c., et al. “Parallel declines in pollinators and
insect-pollinated plants in Britain and the nether-lands.” Science 313 (2006): 351–354.
Bowers, Karen a. W. “the pollination ecology of Solanum rostratum.” American Journal of Botany 62 (1975): 633–638.
Buchmann, Stephen l., and Gary Paul nabhan. The For-gotten Pollinators. covelo, calif.: island Press, 1997.
DeWeerdt, Sarah. “Pollinator panic.” Worldwatch, novem-ber/December 2008, 24–29.
Dunn, Robb. “love is in the air.” National Geographic, December 2009, 120–133.
Fenster, charles B., et al. “Pollination syndromes and floral specialization.” Annual Review of Ecology, Evolution, and Systematics 35 (2004): 375–404.
Galen, candace. “Why do flowers vary? the functional ecology of variation in flower size and form within natural plant populations.” BioScience 49 (1999): 631–640.
———, and J. cuba. “Down the tube: Pollinators, preda-tors, and the evolution of flower shape in the alpine sky pilot Polemonium viscosum.” Evolution 55 (2001): 1,963–1,971.
Gaskett, anne c., et al. “Orchid sexual deceit provokes ejaculation.” American Naturalist 171 (2008): e206–e212.
Goulson, Dave. “effects of introduced bees on native eco-systems.” Annual Review of Ecology, Evolution, and Systematics 34 (2003): 1–26.
James, Rosalind, and theresa l. Pitts-Singer, eds. Bee Polli-nation in Agricultural Ecosystems. new york: Oxford University Press, 2008.
Kearns, c. a., et al. “endangered mutualisms: the con-servation of plant-pollinator interactions.” Annual Review of Ecology, Evolution, and Systematics 29 (1998): 83–112.
xviii+598_Biodiversity_F.indd 421 11/17/11 1:13 PM

first entry422 pollution 422
Kessler, Danny, et al. “Field experiments with transformed plants reveal the sense of floral scents.” Science 321 (2008): 1,200–1,202. Reviewed by Raguso, Robert a. “the ‘invisible hand’ of floral chemistry.” Science 321 (2008): 1,163–1,164.
Knight, tiffany M., et al. “Pollen limitation of plant repro-duction: Pattern and process.” Annual Review of Ecol-ogy, Evolution, and Systematics 36 (2005): 467–497.
Pollan, Michael. “love and lies: Orchids.” National Geo-graphic, September 2009, 100–121.
Proctor, Michael, Peter yeo, and andrew lack. The Nat-ural History of Pollination. Portland, Ore.: timber Press, 1996.
Raguso, Robert a. “Wake up and smell the roses: the ecol-ogy and evolution of floral scent.” Annual Reviews of Ecology, Evolution, and Systematics 39 (2008): 549–569.
Ratnieks, Francis l. W., and norman l. carreck. “clarity on honey bee collapse?” Science 327 (2010): 152–153.
Ren, Dong, et al. “a probable pollination mode before angiosperms: eurasian, long-proboscid scorpionflies.” Science 326 (2009): 840–847. Summarized by Oller-ton, Jeff, and emma coulthard. “evolution of animal pollination.” Science 326 (2009): 808–809.
Richards, a. J. “Does low biodiversity resulting from mod-ern agricultural practice affect crop pollination and yield?” Annals of Botany 88 (2001): 165–172.
Ricketts, t. H. “tropical forest fragments enhance pollina-tor activity in nearby coffee crops.” Conservation Biol-ogy 18 (2004): 1,262–1,271.
Sessions, laura a., and Steven D. Johnson. “the flower and the fly.” Natural History, March 2005, 58–63.
Steffan-Dewenter, i., and t. tscharntke. “effects of habitat isolation on pollinator communities and seed set.” Oecologia 121 (1999): 432–440.
Whitney, Heather M., et al. “Floral iridescence, produced by diffractive optics, acts as a cue for animal pollina-tors.” Science 323 (2009): 130–133.
Wilcock, c., and R. neiland. “Pollination failure in plants: Why it happens and when it matters.” Trends in Plant Science 7 (2002): 270–277.
pollution Despite decades of national legislation, pollution by toxic chemicals remains an important problem, not just in areas with dense human popula-tions, but also in protected areas and for endangered species. Below are some examples of serious pollu-tion problems.
Pesticide residues. about 5.7 billion pounds (2.6 billion kg) of pesticides are released into the envi-ronment every year worldwide. the crop on which the most pesticide is used is cotton, in which (since it is not eaten by people) pesticide residues are not considered an important problem. the United States imports many vegetables and fruits from Mexico.
the warmer climate in Mexico allows vegetables and fruits to be produced during the winter season. Mexico also has less stringent policies regarding the use of, and exposure of farm workers to, pesticides. Sociologist angus Wright explains how the ameri-can demand for winter vegetables and fruits results in the exposure of Mexican children and adults to levels of pesticide that can cause nerve damage and death. the evolution of insect resistance to pesti-cides has prompted the owners of large Mexican farms to increase their application of pesticides. this has made some Mexican produce, including from the highly productive culiacán region, unaccept-able to the american market. Many consumers now demand organic produce, which is grown without the use of pesticides or inorganic fertilizers.
Feedlot runoff. Many livestock feedlots produce as much sewage as a large city of humans. Feed-lot sewage treatment is often less careful than the treatment of human sewage: Bacterial decomposi-tion occurs in open pools from which floods can wash the sludge into streams and rivers. in addition to these specific point sources of pollution, pollu-tion from generalized nonpoint sources may occur in runoff from livestock ranches. even when properly treated, feedlot runoff is high in phosphates, which promotes eutrophication in freshwaters. Oklahoma sued arkansas in 2005, claiming that poultry feedlot runoff was damaging Oklahoma rivers. in 2009, a federal judge recognized that the cherokee nation, into whose traditional territory much of the pollution drained, also had a legal interest in this ongoing suit. litigation continues at press time.
Mining runoff. Mining companies have tradi-tionally left mine tailings piled up without contain-ment, allowing acid and metal ions (such as cadmium and zinc) to leach into nearby creeks and ground-water. the contaminated water is often in primarily native american communities, such as the notorious case of Picher, Oklahoma, most of whose residents are Miami and cherokee, and where tar creek flows orange with acidic mine runoff. in the case of gold mines, it is not the minerals present in the ore so much as the cyanide and mercury used to extract the metal that is a cause of serious contamination.
Persistent organic pollutants (POPs). Many organic contaminants decompose quickly, but some of them resist oxidation and decomposition long enough to contaminate whole food chains as a result of biomagnification. Many of these chemicals are fat-soluble and accumulate in animal bodies. Some of them mimic hormones, suppress immune func-tion, and harm the nervous system. in the Gulf of St. lawrence, the poisoned corpses of beluga whales were themselves considered to be toxic waste because of organic contamination.
xviii+598_Biodiversity_F.indd 422 11/17/11 1:13 PM

population biology 423
Pharmaceutical wastes. normally, antibiotic resistance evolves in populations of bacteria that live in the bodies of people who overuse antibiot-ics. However, six times as much antibiotic is used for livestock as for people. Populations of bacteria can evolve resistance in the livestock, and feedlot runoff can contaminate freshwaters with these resis-tant bacteria—which can, in turn, transfer resis-tance genes to bacteria that infect humans. Bacteria resistant to antibiotics used by humans are routinely found in streams and rivers. in addition, pharma-ceutical hormones (such as estrogen used for birth control) can persist in and contaminate freshwaters, even after sewage treatment. estrogen can steril-ize male trout at concentrations as low as 10 parts per trillion. Feminized fish are frequently found downstream from wastewater treatment facilities. the herbicide atrazine can feminize male frogs at a concentration thousands of times lower than occurs in agricultural runoff and even at a concentration 400 times lower than is found in rainwater in some agricultural areas. Because atrazine, at concentra-tions that might possibly be found in water supplies, can cause human fetuses to grow more slowly and to deliver early, this herbicide is banned in France, Germany, italy, Sweden, and norway.
Pollutants permeate regions in which humans live and the natural areas surrounding them. eco-logical processes such as biomagnification can make the problems even worse for humans.
See also biomagnification; resistance, evo-lution of; rivers, streams, ponds, and lakes.
Further readingBadgley, c., and ivette Perfecto. “can organic agriculture
feed the world?” Renewable Agriculture and Food Systems 22 (2007): 80–85.
Bank, M. S., et al. “Mercury bioaccumulation in northern two-lined salamanders from streams in the north-eastern United States.” Ecotoxicology 14 (2005): 181–191.
Batty, lesley c., and Kevin B. Hallberg. Ecology of Indus-trial Pollution. new york: Oxford University Press, 2010.
Blauenstein, a. R., et al. “UV repair and resistance to solar UV-B in amphibian eggs: a link to population declines.” Proceedings of the National Academy of Sci-ences USA 91 (1994): 1,791–1,795.
———, et al. “Ultraviolet radiation, toxic chemicals, and amphibian population declines.” Diversity and Distri-butions 9 (2003): 123–140.
chavez, Will. “cherokee nation appeals to join lawsuit against poultry industry.” Cherokee Phoenix, Decem-ber 30, 2009. available online. URl: http://www.cher okeephoenix.org/20991/article.aspx. accessed august 14, 2010.
Damsgaard, i., et al. “Persistent pesticides in human breast milk and cryptorchidism.” Environmental Health Per-spectives 114 (2006): 1,113–1,118.
Gosling, P., et al. “arbuscular mycorrhizal fungi and organic farming.” Agriculture Ecosystems and Envi-ronment 113 (2006): 17–35.
Green, R. e., et al. “Farming and the fate of wild nature.” Science 307 (2005): 550–555.
Hayes, t. B., et al. “Herbicides: Feminization of male frogs in the wild.” Nature 419 (2002): 895.
———, et al. “Hermaphroditic, demasculinized frogs after exposure to the herbicide atrazine at low ecologically relevant doses.” Proceedings of the National Academy of Sciences USA 99 (2002): 5,476–5,480.
Hole, D., et al. “Does organic farming benefit biodiver-sity?” Biological Conservation 122 (2005): 113–130.
lotter, D., et al. “the performance of organic and con-ventional cropping systems in an extreme climate year.” American Journal of Alternative Agriculture 18 (2003): 146–154.
Mäder, P., et al. “Soil fertility and biodiversity in organic farming.” Science 296 (2002): 1,694–1,697.
Oklahoma Department of environmental Quality. “tar creek native american issues final report.” avail-able online. URl: http://www.environment.ok.gov/documents/tarck/naissuesFinalReport.pdf. accessed august 14, 2010.
Pierce, B. “the effects of acid precipitation on amphib-ians.” Ecotoxicology 2 (1993): 65–77.
U. S. Water news Online. “Oklahoma readies for another court fight, blaming river’s decline on poultry.” available online. URl: http://www.uswaternews.com/archives/arcrights/5oklaread7.html. accessed august 14, 2010.
Wright, angus. The Death of Ramón González: The Mod-ern Agricultural Dilemma. Rev. ed. austin: University of texas Press, 2005.
population biology a population is all of the individuals of a species within a defined area, within which the individuals could potentially interbreed. a population consists of demes. Within each deme, interbreeding is not only possible but occurs with some regularity. the delimitation of a population is somewhat arbitrary in practice. One can speak of the human population of the earth, or of california, or of San Diego. One could study the population of dandelions in one yard or in all the yards in the country. to preserve biodiversity, it is necessary to rescue the small populations of rare species, making population biology an intensely important subject for conservationists.
all individuals in a population are members of the same species, but that is where the uniformity ends. there are several sources of diversity within populations:
xviii+598_Biodiversity_F.indd 423 11/17/11 1:13 PM

first entry424 population biology 424
● Populations usually contain individuals of many different ages and sizes. adult indi-viduals reproduce, while juveniles do not; and adults may produce and/or consume more resources than do juvenile individuals. thus two populations of one species may differ from one another even if the popula-tion size is the same: One of the populations may have many small, young individuals, while the other has many large, older ones. a population that consists of 1,000 buffalo of which 300 are calves may consume less grass than a population of 1,000 buffalo of which only 50 are calves. in plant popula-tions, individuals can differ greatly in size even though they have the same age. this occurs in part because plants are capable of surviving on very limited resources for months or years; therefore some old plants are frequently smaller than young plants in the same population.
● among most animals, populations consist of a mixture of males and females, usually but not always in roughly equal proportions. in plants, the sexual characteristics of a popu-lation can be much more complex. in some species of plants the populations consist of individual plants that have bisexual flow-ers. in other species, some plants have flow-ers that have only male parts, while others have flowers with female parts. Many other sexual combinations exist in plant species.
● Populations almost always consist of indi-viduals of many different genotypes, result-ing in a diversity of phenotypes. Often a human observer cannot see the differences among individuals in a population. Scien-tists can detect genetic differences among individuals in plant and animal populations only by comparing the enzymes or Dna of the individuals.
the fact that crop and domesticated animal pop-ulations contain heritable diversity has been known since ancient times. ancient herdsmen assumed live-stock offspring would have characteristics like those of the parents. charles Darwin recognized that if populations of domesticated species have heritable diversity, then populations of wild species probably do also, for all domesticated species were once wild. as a result, he was able to explain that the process of evolution was caused largely by means of natural selection, just as the evolution of domesticated spe-cies was caused by artificial selection, or breeding, imposed by humans. Darwin studied animal breed-ing, particularly pigeon breeding. the various breeds
of pigeon that Darwin studied differed strikingly from one another in appearance. Some had hoods of feathers, others had fanlike tails; some did aerial somersaults, some made unusual sounds. One had an “enormously developed crop, which it glories in inflating, [and which] may well excite astonishment and even laughter,” as he wrote in the first chapter of The Origin of Species. Darwin then wrote,
altogether a score of pigeons might be chosen, which, if they were shown to an ornithologist, and he were told that they were wild birds, would certainly be ranked by him as well-defined species.
Despite these enormous differences between breeds of pigeon, Darwin was convinced (as were most of his fellow scientists, even before he proposed the general theory of evolution) that they had all been derived from a single ancestral species of wild pigeon. Wild pigeons may all look alike to humans. certainly wild pigeons do not have fanlike tails or inflatable throats. yet the genetic basis for big tails and inflatable throats existed and probably still exists within the wild pigeon populations. the wild pigeon populations, like nearly all populations, con-tain a great deal of hidden genetic variation.
in some species, what appear to be individual members of the population are asexually propagated copies of one individual—that is, they are a clone. Some such clones can be extremely large. the larg-est organisms in the world may be clones of aspen trees, in which many trunks are connected by under-ground stems that can form a network over hun-dreds of acres.
how populaTIoNS Grownew individuals are added to a population by birth and by the immigration of individuals from other populations. individuals are removed from the pop-ulation by death and when individuals leave the population (emigration). Populations grow if more individuals are added than removed; populations decline if fewer are added than removed.
imagine a pond in which frogs live. this imagi-nary frog population doubles in size every day and fills the pond after 30 days of growth. On what day is the pond half-filled with frogs? the pond is half full of frogs on the 29th day. On the 29th day, a frog could look around the pond and see that there is still a lot of free, open water not yet filled. an optimistic frog would say that the pond is still half empty. yet the crisis of the pond being completely filled is only one day away from happening.
this story depicts quite accurately the way pop-ulations can grow. in the simplest case, one bacterial
xviii+598_Biodiversity_F.indd 424 11/17/11 1:13 PM

population biology 425
cell (in generation 0) divides to become two cells (in generation 1), and these two cells divide to become four (in generation 2), and so forth. in this example, the bacterial population grows from a single cell to over a billion in just 30 generations. in some bacte-rial species, under ideal conditions, such popula-tion growth can occur overnight in a laboratory. in another 30 generations, this number could rise to more than a billion billion (1018) cells. this kind of growth is called exponential, because the super-script, called an exponent, indicates how many times the base number (in this case 2) is multiplied by itself. On a graph of population size as a function of time, exponential growth produces a curve in which not only the size of the population but also the rate of increase increases. in contrast, linear growth would occur if the population increased at a constant rate, resulting in a straight line on a population growth graph. all populations are capable of exponential growth. the exponential growth of populations can take humans by surprise because the human mind generally thinks in linear terms.
Most populations are more complex than those of bacteria. For instance, most animals reproduce in pairs, not as individuals. Furthermore, in bac-teria the parent “dies” because it splits in half and literally becomes the offspring, whereas in animal and plant populations the parents persist and per-haps continue to reproduce even after their off-spring reach adulthood. these circumstances make the mathematical calculations of population growth for animals and plants more complex than for bac-teria. However, these complexities do not alter the exponential shape of the growth curve. even a very slow reproductive rate can result in exponential population growth.
One of the first people to write about the fact that human populations cannot grow indefinitely was economist thomas Malthus in 1798. after read-ing his writings, Darwin applied the same principle to plant and animal populations:
it is the doctrine of Malthus applied with mani-fold force to the whole animal and vegetable kingdoms. . . . although some species may be now increasing . . . in numbers, all cannot do so, for the world would not hold them.
a mathematical equation that describes popula-tion growth is:
N2 = N1ert
● N1 is the population size at time 1. ● N2 is the population size at time 2. ● e is the natural base of logarithms.
● r is the rate of increase as a decimal frac-tion, per year. this variable is also called the Malthusian parameter after thomas Malthus.
● t is the amount of elapsed time.
exponential population growth can occur tem-porarily when a few plants or animals enter a new habitat in which there is a superabundance of food and space for them. this has often happened when humans have carried plants or animals away from their native habitat and introduced them into new regions. Darwin noticed this also:
Still more striking is the evidence from our domestic animals of many kinds which have run wild in several parts of the world; if statements of the rate of increase of slow-breeding cattle and horses in South america, and latterly in austra-lia, had not been well authenticated, they would have been incredible.
two male and six female pheasants were intro-duced to an island off the coast of Washington. Within five years, there were 1,325 pheasants—an increase of 180 percent per year. Sheep populations also increased rapidly when they were introduced to tasmania.
the human population has experienced an exponential increase in recent centuries. Within the last few years, the growth rate of the human popula-tion is no longer increasing as fast as it once was (see below), even though it is still increasing. as a result, the exponential phase of human population growth may be coming to an end—but not any time soon.
a population with a higher fertility rate can grow faster than a population with a lower fertil-ity rate. the fertility rate is the average number of offspring per reproductive individual or, in many animals, per female. a Princess anne tree (Paulow-nia tomentosa) can easily produce 50,000 small wind-blown seeds in a single year. a population of such trees has a very high average fertility rate, and its population would grow very fast if all of its seeds fell in an area with unlimited space, light, water, and minerals. contrast this with a black walnut tree (Juglans nigra), which can produce only a few hundred seeds at a time. this species has a much lower average fertility rate, and its population would grow much more slowly in a region with unlimited resources.
not only do different species have different aver-age fertility rates, but different populations within a species may have different fertility rates. consider human populations as an example. as of 2011, the world average fertility rate is 2.5. that is, the average
xviii+598_Biodiversity_F.indd 425 11/17/11 1:13 PM

first entry426 population biology 426
woman in the world has 2.5 children during her life-time. the fertility rate in niger is 7.0 children per female, almost three times the world average, while in Belgium the fertility rate is 1.4 children per female. Partly as a result, niger’s population has been grow-ing 18 times faster than Belgium’s: niger’s population is 3.6 percent bigger each year, while in Belgium it is only 0.2 percent bigger each year.
all around the world, human fertility rates are declining slightly. For example, the fertility rate in Bangladesh was 6.7 in 1953, while it was 2.4 in 2010. as a result, the world population growth rate is declining as well. among the reasons for this appear to be:
● Greater availability and reliability of food and medicine. this has resulted in a world-wide reduction in infant mortality. When parents have a greater assurance that their offspring will survive, they often choose to have fewer offspring.
● Greater availability of birth control options. this allows parents (especially mothers) to translate their desire to have fewer offspring into practice.
● Greater availability of education and oppor-tunities for women. in most countries, women now have more choices as alterna-tives to childbearing.
Despite this decline in birth rate, world popu-lation is still growing rapidly. the current rate of world population increase, 1.2 percent, in a world population of 7 billion people means that each year the world contains about 84 million more people than the year before!
a population with a lower age at first reproduc-tion can grow more rapidly than a population that has a higher age at first reproduction. Most farm weeds, such as velvetleaf, ragweed, and foxtail, are annuals, which reproduce during their first and only year of life. Perennial plants live for two or more years. Princess anne trees have a much later age of first reproduction than weeds. not surprisingly, annual weed populations can grow more rapidly than tree populations, even though a Princess anne tree can produce, in a given year, more seeds than most annual weeds. Moreover, many trees do not reproduce heavily every year. they may produce few or no seeds for several years, saving up for a mast year during which they dump seeds all over the ground.
the human age at which reproduction first becomes possible has decreased in some countries. in europe, the average age of first menstruation was 17 in the mid-1800s, while today it is 13. this has
probably occurred because children receive better nutrition today than in the past.
Fertility rate and age at first reproduction both affect population growth. a female cockroach can lay a hundred eggs in a single egg case and therefore has a higher fertility rate than a bacterium in which the “parent” can produce only two “offspring.” However, the bacterial population can grow faster because the bacteria can divide every half hour, while at least two weeks is required for a female cockroach to produce an egg case.
Some species are adapted to live and grow in places in which some disturbance such as fire or human activity has removed the plant cover. Weeds that live in these disturbed areas grow quickly, pro-duce many offspring, and have short life spans. this allows them to grow abundantly in habitats that persist for only a short time. Weeds cannot compete against the bushes and trees that later grow in these disturbed areas. natural selection has favored rapid growth and early reproduction in weeds because their populations are usually well below the number that the resources can support. Other species, such as walnut trees, are adapted to live in stable con-ditions. evolution has favored slower growth and later reproduction in these plants, allowing them to compete well against other species, because their populations are usually at or near the number that the resources can support.
Small, juvenile individuals generally do not reproduce, nor do very old individuals. therefore, to understand a population’s growth rate, one must know the size and age differences among individu-als within the population. consider an example from human populations. niger has about 16.1 million people and Belgium has about 11 million people. in Belgium, 17 percent of the population is under the age of 15, and 17 percent of the popula-tion is over the age of 65. this represents a stable population. in niger, however, 49 percent of the population is under 15 and only 2 percent of the population is over 65 years of age. thus, when we compare the age structure of the two populations, we see that a large number of youngsters in niger will soon be entering their reproductive years, whereas in Belgium this is not the case. even if both countries had low fertility rates, niger would soon experience an explosive population growth and Belgium would not.
Because of the very large number of children in the world (27 percent of the world population is under age 15), world population will continue to grow rapidly for many years, despite the decline in the world average fertility rate.
Some organisms produce a large number of small offspring, most of which die. Small offspring—for
xviii+598_Biodiversity_F.indd 426 11/17/11 1:13 PM

population biology 427
instance, the small seeds from a Princess anne tree, which may weigh 7-millionths of an ounce (200 μg)—have very little chance of survival. there is very little food stored in the seed to nourish the seedling as it germinates. if this seed lands in a thick sward of grass or underneath bushes or trees, it has very little chance of surviving even if it germinates. the Princess anne tree, however, produces a large number of these seeds. Perhaps one of these seeds will land in a suitably open, sunny, fertile spot. the parent tree can easily afford to produce many seeds, since each seed requires few resources and is “cheap” to produce.
Other organisms produce a small number of large, expensive offspring, more of which survive. Walnut trees produce large fruits, each one with a large embryo, a dense hard shell, and a thick husk. each one can weigh more than three ounces (100 g), 500,000 times as much as a Princess anne tree seed. Such a large seed has a greater food reserve that allows it to survive as it germinates amidst a dense growth of other plants. the seedling itself is large enough to outgrow many of its neighboring seedlings. the walnut tree, however, cannot afford to produce very many such “expensive” seeds.
Both trees make investments that may be equally successful. the Princess anne tree is gambling: each seed is a lottery ticket that is almost guaranteed to lose, but the plant invests in a very large number of tickets. the walnut is making fewer, safer invest-ments. animal examples also abound. Some insects, for example, lay a large number of eggs, most of which die. these animals produce a large number of cheap offspring. cattle, in contrast, produce very few offspring, but take very good care of them. Most of their offspring survive. Humans are among the most extreme examples of animals that put much investment into each individual offspring. each off-spring is carried in the mother’s womb for nine months, is quite large when born, is fed and pro-tected for years of helpless childhood, and educated for many more years.
Darwin also recognized that a species with a high fertility rate could end up having a population growth rate no greater than a species with a low fertility rate:
the condor lays a couple of eggs and the ostrich a score, and yet in the same country the condor may be the more numerous of the two; the Ful-mar petrel lays but one egg, yet it is believed to be the most numerous bird in the world. One fly deposits hundreds of eggs, and another, like the hippobosca, a single one; but this difference does not determine how many individuals of the two species can be supported in a district. it would
suffice to keep up the full number of a tree, which lived on an average for a thousand years, if a sin-gle seed were produced once in a thousand years, supposing that this seed were never destroyed, and could be ensured to germinate in a fitting place. So that, in all cases, the average number of any animal or plant depends only indirectly on the number of its eggs or seeds.
the number of surviving offspring that are pro-duced by an organism is a measure of its fitness. this term fitness as used by population and evolutionary ecologists should not be confused with the term physical fitness as used by physiologists and exercise scientists.
lImITS To populaTIoN GrowThexponential population growth cannot continue in a world with limited food, space, and other resources. economists sometimes assume that population growth and resource use can continue essentially forever. as economist Kenneth Boulding wrote, “anyone who believes exponential growth can go on forever in a finite world is either a madman or an economist.” Recently, ecological economics, which recognizes the limits to growth, has become more widely accepted.
Survivorship is how many offspring survive from the total number born and how this number of survi-vors changes over time. Populations differ from one another in their patterns of survivorship. in some populations, most individuals survive until old age; in others, a relatively constant proportion of individuals die at all ages; in others, most deaths occur among the youngest individuals. Humans are an example of the first pattern, in which most deaths are of old individuals. around the world, only 4.6 percent of human infants die within their first year. in some countries without adequate nutrition, sanitation, and medical care, up to 15 percent of the infants die dur-ing their first year. in most industrialized countries, such as the United States and Japan, infant mortality rate is less than 1 percent. the third pattern seems to be the most common in nonhuman species, because the youngest and smallest individuals are the ones least capable of responding to the dangers of the envi-ronment, such as drought, disease, or crowding. in plants, seedlings are also close to the ground, which can be very hot and dry, yet at the same time they have very small roots. it is the young plants also that are most tender and vulnerable to fungal infection. Darwin not only noticed this but actually counted the numbers of seedlings that died:
Seedlings . . . are destroyed in vast numbers by various enemies; for instance on a piece of
xviii+598_Biodiversity_F.indd 427 11/17/11 1:13 PM

first entry428 population biology 428
ground three feet long and two wide, dug and cleared, and where there could be no choking from other plants, i marked all the seedlings of our native weeds as they came up, and out of 357 no less than 295 were destroyed, chiefly by slugs and insects.
Many plant populations contain a large number of seeds that lie dormant in the soil for months, years, or centuries. they form a seed bank from which a few seeds may germinate every few years or which may remain dormant until some event, such as a forest fire, removes the vegetation that has been shading them. a plant population can therefore begin growing again even if all above-ground evidence of its existence has disappeared. in other plant species, such as in some tropical trees, many seedlings will germinate in the shade, grow a few centimeters, then wait for years without growing very much, until the branches above them are cleared away by a disturbance. When bright light becomes available, these small seedlings will quickly grow.
Some of the factors that limit population growth are unaffected by the size or density of the popula-tion—for instance, a flood or a freeze. Such a factor may destroy more organisms in a large population than in a small population, for the simple reason that a larger population has more individuals to get in the way. However, the factor itself is density-independent, for it occurs regardless of the presence of populations.
Many other factors, however, reduce the growth of populations with high density much more than populations with low density. if a factor has a greater effect at higher population densities, it is density-dependent. examples include:
● Intraspecific competition. individuals that are far apart have no effect on one another, and each individual has access to an unlim-ited amount of resources. Competition results when crowded individuals do not have access to unlimited resources (see fig-ure). in this case, the competition is intraspe-cific (within a species). competition can have many effects on a population, one of which is a reduction in the number of offspring that the average individual produces.
● Attack by animals. Sphinx moths look for tomato plants, on which they lay eggs that hatch into voracious hornworm caterpil-lars. a dense grouping of tomato plants may be easier for a sphinx moth to find than a single, isolated tomato plant. a big herd of animals may be more easily visible
to a predator than is an isolated individual; a herd cannot as easily hide. crowded pop-ulations may therefore suffer more from attacks by animals that eat them. insect pest populations can grow more rapidly in large, dense crop populations than in small, sparse populations. Modern agricul-ture requires large, dense populations of crops, with the inevitable result that pest populations explode.
● Infectious disease. Diseases will spread much more rapidly through a dense human population, as individuals come in contact with one another directly or with each other’s breath or wastes. Some human diseases require a certain minimum population size to persist. Smallpox was eradicated not because literally everybody in the world was vaccinated, but because there were so few unvaccinated people in any given locality that the population of viruses could no longer spread. the chest-nut blight, a fungal disease that destroyed all of the adult chestnut trees in the for-ests of eastern north america, did not spread to isolated chestnut trees that had been planted in other parts of the country before the outbreak of the blight. individu-als in dense populations may experience stress and therefore be more susceptible to disease. Diseases spread more rapidly in large, dense populations of crop plants than in small, sparse populations, just as is the case with insect pests.
Cones of bishop pines (Pinus muricata) open only after a fire and produce a thick growth of seedlings the next year. In this photograph, taken seven years after a fire in the Purísima Hills of coastal California, the seedlings have grown into a dense population of trees, each much smaller than trees that grow at lower density. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 428 11/17/11 1:13 PM

population biology 429
although most density-dependent factors are biological, some physical factors can also be influ-enced by density. a fire, for instance, is more likely to start and spread in a dense than in a sparse plant population, particularly if the dense plant popula-tion has much litter or many dead or dying stems.
Darwin realized that such density-dependent factors customarily prevented populations from hav-ing exponential growth:
a plant which annually produces a thousand seeds, of which only one of an average comes to maturity, may be . . . said to struggle with plants of the same and other kinds which already clothe the ground. . . . lighten any check, mitigate the destruction ever so little, and the number of the species will almost instantaneously increase to any amount.
While very high densities can have negative effects on individuals within populations, there can also be beneficial effects of moderate crowding. While a large population of prey might be more easily spotted by a predator, it might also be able to defend itself more effectively than an isolated individual. For example, a herd of animals can carry out defensive maneuvers to ward off predators and protect the young. among plants, an isolated individual in a bare field might suffer the consequences of soil erosion and hot dry winds, from which a moderate density of other plants might be able to shield it. the benefits of moderate population density, relative to low population density, were first pointed out by ecologist Warder c. allee and are called the allee effect.
the maximum number of individuals that can survive, given the available resources at a particu-lar time and place, is called the carrying capacity. Density-dependent factors may cause populations to grow more slowly as they approach the carry-ing capacity. if the population exceeds its carrying capacity, density-dependent factors may cause the population to decrease back down to the carrying capacity. in agricultural situations, the ideal den-sity is that which allows each individual to repro-duce at its maximum level, resulting in optimum yield.
it is not always possible to calculate an exact carrying capacity, because the carrying capacity will be higher if the average individual in the population uses fewer resources. this is especially true of the carrying capacity of humans. americans use about one-third of the energy and material resources of the world. if everybody in the world used as many resources as americans, the carrying capacity of the human species would be less than 900 million people. clearly, the world cannot support a human
population of several billion people living at ameri-can standards.
Occasionally, populations increase until they reach the carrying capacity, at which density the population levels off. Sheep were first introduced to tasmania about 1820. their population density was low, and there were no native mammals that were able to compete with them for food. the sheep pop-ulation underwent exponential growth, then leveled off at about 2 million, where it has remained ever since. it appears that 2 million is their approximate carrying capacity in tasmania.
examples of such smooth population growth patterns are, however, very rare. it is more frequent to find drastic fluctuations from year to year. Popu-lations that grow far beyond their carrying capacity may suffer a severe crash. in some cases, populations rise and fall in a cyclical pattern, for reasons that are not entirely clear. Darwin noticed that populations underwent dramatic fluctuations:
We behold the face of nature bright with glad-ness, we often see superabundance of food; . . . we do not always bear in mind, that, though food may be now superabundant, it is not so at all sea-sons of each recurring year.
Population growth can level off at the carrying capacity in several ways. Density-dependent factors may cause the death of individuals. Density-depen-dent factors may cause fertility rates to decline, with-out necessarily causing death. if birth rates decline, while organisms continue to die at the same rate, the population growth will decline as surely as if the organisms suffered increased death. Finally, indi-viduals may emigrate from dense populations.
as noted above, the human population growth rate is declining slowly because there are more opportunities for reliable food, medicine, education, and opportunity. in this way, humans are differ-ent from most animal species, whose population growth rates decline mainly due to death and emi-gration. Some observers, such as biologist Garrett Hardin and economist Kenneth Boulding, expected the human population to respond to resources the same way as populations of other animal species. in 1964, Boulding published his “utterly dismal theo-rem”: “if the only thing which can check the growth of population is starvation and misery, then the ultimate result of any technological improvement is to enable a larger number of people to live in misery than before and hence to increase the total sum of human misery.” Fortunately, this has proven to not be the case.
Because modern agriculture promotes dense populations of single crop species, the eruption of
xviii+598_Biodiversity_F.indd 429 11/17/11 1:13 PM

first entry430 population genetics 430
pest population outbreaks is nearly inevitable. By reducing the density of any one crop species, for example by planting alternating strips of each crop species rather than entire fields of each crop, pest out-breaks can be reduced in frequency and severity. this reduces the need for the heavy use of pesticides. in this way, human activities such as agriculture would follow the same laws of nature (in this case, of popu-lation biology) as do the activities of all other species.
See also Darwin, charles; natural selec-tion; population genetics.
Further readingBrown, lester R., Gary Gardner, and Brian Halwell.
Beyond Malthus: Sixteen Dimensions of the Popula-tion Problem. Worldwatch Paper 143. Washington, D.c.: Worldwatch institute, 1998.
cohen, Joel e. “ten myths of population.” Discover, april 1996, 42–47.
Darwin, charles. “the struggle for existence.” chap. 3 in The Origin of Species by Means of Natural Selection or the Preservation of Favored Races in the Struggle for Life. new york: new american library, 1958.
Hardin, Garrett. Living within Limits: Ecology, Econom-ics, and Population Taboos. new york: Oxford Uni-versity Press, 1993.
livi-Bacci, Massimo. a Concise History of World Popula-tion. 3rd ed. london: Blackwell, 2001.
Population Reference Bureau. “2011 World Population Data Sheet.” available online. URl: http://prb.org/pdf11/2011population-data-sheet_eng.pdf. accessed august 8, 2011.
United States census Bureau. “international Data Base.” available online. URl: http://www.census.gov/popu lation/international/data/idb/informationGateway.php accessed august 8, 2011.
population genetics a population is defined as a set of organisms, all of the same species, that can interbreed with one another because there are no reproductive barriers and because they all live in the same place. a local population in which interbreed-ing is not only possible but frequent is considered a deme. the genes of the organisms in a deme, or in a population, are therefore considered to be a single gene pool.
Population genetics is the study of genetic varia-tion in populations. Genes, encoded in Dna, are the only information that passes from one generation to the next in most species, although many animal species, especially humans, transmit cultural infor-mation from one generation to the next. in cells of eukaryotes, most of the genes are in the nucleus, although there are also many cytoplasmic factors (genes in chloroplasts and mitochondria). Genes (the
genotype) affect the characteristics of organisms (the phenotype); however, phenotype changes that occur during the life of an organism cannot be transferred back to the genotype. that is, acquired characteris-tics cannot be inherited. evolution occurs in popula-tions. therefore, the study of genetic variation in a population is the study of the raw material available for evolution in that population. the only evolution that can occur is what the genetic variation permits.
the study of population genetics is essential to the preservation of biodiversity, because a species can only survive if enough of its populations survive, and a population can survive only if there is enough genetic variation.
populaTIoNS havE GENETIC varIaBIlITyin most species, an individual organism can have at most two alleles for a given locus (plural loci) or location on a chromosome: it has two copies of one allele if it is homozygous, and one copy of each of two alleles if it is heterozygous for that locus. a population can have many alleles for each locus. in many laboratory populations, one allele predomi-nates (the wild type) over a few rare mutant alleles, which produce defects in the phenotype. in many wild populations, for many loci, there is more than one common allele (a polymorphism). in contrast, populations of agricultural crops and livestock have very little genetic variability (see figures on page 431).
Populations can differ from one another in the types of alleles that they have for a locus. even if they have the same alleles, they can differ in the relative frequencies of these alleles. For exam-ple, in the Mn blood protein group in humans, the n allele strongly predominates in australian aboriginal populations, and the M allele in navaho populations, while the two alleles occur at approxi-mately equal frequency among caucasians in the United States. the three populations differ geneti-cally, even though all of them have the same two alleles for the locus.
Because evolution depends upon the variation in the gene pool, one would expect that evolution would proceed more rapidly in a population that has greater genetic variability. this is the idea behind the fundamental theorem of natural selection proposed by evolutionary biologist R. a. Fisher. Fruit flies (genus Drosophila) have a short generation time and can be raised easily in the laboratory. Geneticists such as thomas Hunt Morgan and theodosius Dob-zhansky pioneered the use of fruit flies for genetic and evolutionary studies. experiments with Dro-sophila indicate that populations that have greater genetic variability grow faster than populations with less genetic variability when that population is exposed to new environmental conditions (for
xviii+598_Biodiversity_F.indd 430 11/17/11 1:13 PM

population genetics 431
example, a change in temperature), which is consis-tent with Fisher’s theorem.
One difficulty that geneticists often encounter when studying populations is how to define an indi-vidual. the etymology of the word itself suggests that an individual is an entity that cannot be divided without killing it. With vertebrates, it is easy enough to recognize an individual. With plants and some invertebrates, however, it is not as easy, as in these examples:
● Plants and starfish can be cut up into pieces, and a new organism can grow from each piece. are plants and starfish individuals?
● Many plants and invertebrates produce seeds and eggs asexually, resulting in numer-ous organisms that are genetically identical. Should these clonal organisms be considered individuals?
● Some plants spread by asexual means, for example by underground branches. in this way, an entire field of goldenrod plants (ramets) might be a single genetic individual (genet). the connections among the ramets might be maintained or might not, depend-ing upon the species and circumstances. Should ramets be considered individuals?
● it may make more sense for a tree to be con-sidered a population of branches rather than an individual. Somatic mutations can occur in buds. a branch that grows from a bud that has a somatic mutation is genetically different from the other branches of the tree. each branch of a tree may differ in its abil-ity to resist herbivores. insect herbivores, which have several generations in a single year, can evolve much faster than trees. it has been suggested that the generation of somatic mutations allows trees to keep up with the evolutionary pace of the insects that eat their leaves. if this branch produces seeds, the genetic differences can be passed on to the next generation. Does a tree func-tion as a genetic and even as an evolutionary population?
mEaSurING GENETIC varIaBIlITy IN populaTIoNS
it has often proven very difficult to measure vari-ability for any given gene or overall genetic vari-ability in populations. Several approaches have been used:
Phenotypic traits. One approach is to estimate variability of a measurable phenotypic trait (e.g., area of a leaf or length of a bird’s beak). this vari-ability can be expressed as a variance, as a standard
deviation, as a standard error, or as a coefficient of variation. this variability among individuals, how-ever, is caused partly by genetic differences among the individuals (the heritability) and partly by dif-ferences among the individuals in the environmen-tal conditions they have experienced. Variance, as originally defined by R. a. Fisher, is the average squared deviation from the average value; because of the square component, variance is symbolized as s2. the heritability component of the variance is symbolized as h2. Genetic and environmental
Every wild population contains genetic variability. This variability is usually invisible. Genetic variation for the production of floral pigment, however, is visible in this population of Sabatia campestris in Oklahoma (top). Agricultural populations, in contrast, contain very little genetic variability. This entire field of onions in California may be genetically identical (bottom). (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 431 11/17/11 1:13 PM

first entry432 population genetics 432
variability is also symbolized by the letter V. thus, total phenotypic variability (VP) consists of genetic variability (VG) and environmental variability (Ve). Genetic variability can itself be divided into two components: the variability that results from diver-sity of alleles (additive genetic variability, Va) and the variability that results from one allele being dominant over another (dominance variability, VD). therefore VG = Va + VD. Heritability can be expressed as either broad-sense or narrow-sense heritability:
● Broad-sense heritability is VG/VP = VG/(VG+Ve), the proportion of phenotypic vari-ation that is genetic.
● Narrow-sense heritability quantifies only the additive genetic variation and is there-fore Va/VP = Va/(Va+VD+Ve). narrow-sense heritability can also be quantified as the proportion of total variability in offspring that can be explained by the variability in the parents in a statistical analysis.
Heritability calculations assume that parental and offspring environments are uncorrelated; if they are correlated, environmental effect can be mistaken for genetic inheritance. Researchers can sometimes avoid this problem experimentally. they can, for example, switch eggs among bird nests, with the result that the offspring of each parent experiences each of the nest environments. this is what Peter and Rosemary Grant did to establish heritability for beak traits in populations of Darwin’s finches on the Galápagos islands. this cannot, however, be done in all experimental systems.
Heritability is a measure of genetic variation among members of a population; it is not the same as inheritance. if there is only one allele for a locus in the entire population, and that allele has a strong effect on a trait, then the trait is very strongly inherited but has a heritability of zero. Heritabil-ity is not a fixed number for a population, since it depends upon the amount of plasticity. consider a genetic disease that is caused by a single allele and that is invariably fatal; the heritability would be 100 percent. But as soon as a medicine is discovered that cures the disease, the heritability drops to 0 percent.
Proteins. another approach is to measure the variability of proteins. Differences among proteins can be very important, as enzyme proteins control all the chemical reactions in an organism. Different forms of the same enzyme, determined by different alleles at the same locus, are called allozymes. an individual that is homozygous for one of the alleles of the gene that encodes this protein will produce
just one allozyme; an individual heterozygous for this gene can produce two allozymes; the individu-als in the population can collectively produce more than two allozymes. However, two allozymes, while they differ in structure, may not differ in function. that is, they may have exactly the same effect on the phenotype of the organism and on its ability to survive and reproduce. therefore, the study of allo-zymes may reveal greater variability than the study of phenotypic traits.
Proteins can be separated from one another by electrophoresis. a mixture of proteins is placed at one end of a gelatinlike sheet (usually composed of the complex sugar agarose). an electric current cre-ates positive and negative poles on this sheet. Pro-teins, which have a slight negative charge, diffuse through the molecules of the gel toward the positive pole. Because they must diffuse through an obstacle course of agarose molecules, the smaller proteins move faster than the larger proteins. therefore, electrophoresis separates proteins on the basis of size as well as of charge. if a type of protein from an individual is placed on a gel, the protein will show up as a band, after the appropriate staining is used to make it visible. if the individual is homo-zygous for that allozyme, one band shows up; an individual heterozygous (producing two allozymes) will have two bands. electrophoresis was first used to study the protein variability of populations by geneticists Richard lewontin and John Hubby in the 1960s.
DNA. another approach is to measure the vari-ability of the Dna itself. Measuring the overall vari-ability of the Dna itself would seem to be an ideal quantification of genetic variability. However, since most of the Dna in a eukaryotic cell is noncoding Dna, a measure of overall Dna variability may be meaningless as an indicator of how much evolution may be possible in that population. even measuring the variability in the Dna of a specified gene may not be very meaningful, as much of this variabil-ity may not produce differences in proteins. How-ever, differences between populations, or differences between species, in their Dna (whether it codes for proteins or not) is routinely used as a measure of how much evolutionary divergence has occurred between them.
Measurements of genetic variability can be used to reconstruct population and species history. For example, biologists took blood samples from 20 birds representing six of the 13 species of Darwin’s finches on the Galápagos islands. they measured the number of alleles in these 20 birds for a single locus, for the MHc ii B class 1 gene. in 20 birds, there can be at most 40 alleles. these birds had 21 alleles. Because the rate at which this gene mutates
xviii+598_Biodiversity_F.indd 432 11/17/11 1:13 PM

population genetics 433
has been estimated, the researchers concluded that the common ancestor of all these birds lived at least 10 million years ago. But the islands are not that old. the original immigrant fi nches may have lived on one of the islands that has eroded away and migrated to their current locations. Furthermore, for the fi nches on the Galápagos islands to have this much genetic variability, the original population would have had to contain at least 40 genetically distinct individual birds.
GENETIC ChaNGES IN populaTIoNSto determine whether or not natural selection is occurring in a population, biologists need to deter-mine what the frequency of each allele (for a given locus) would be in the absence of natural selection, that is, if the population was at equilibrium rather than being infl uenced by natural selection.
consider a simple example in a species of ani-mal, such as a kind of fi sh, that has external fer-tilization and in which a certain locus has just two alleles, A and a. Of the sperm and eggs released into the water by the fi sh in this population, 40 percent carry allele A, while 60 percent of the sperm and eggs carry allele a. the frequency of allele A is p = 0.4, and the frequency of allele a is q = 0.6. Because there are only two alleles, p + q = 1. if the eggs and the sperm come together randomly, you would expect the following results (see fi gure):
1. Because 40 percent of the eggs and 40 percent of the sperm carry allele A, there is a 40% × 40% = p2 = 16% chance that the zygote will have the homozygous AA genotype.
2. Because 60 percent of the eggs and 60 percent of the sperm carry allele a, there is a 60% × 60% = q2 = 36% chance that the zygote will have the homozygous aa genotype.
3. there is a 40% × 60% = pq = 24% chance that a sperm carrying allele a will meet an egg carrying allele A; there is also a 24 percent chance that the opposite will happen: a sperm carrying allele A will meet an egg carrying allele a. there is therefore a 24% + 24% = 2pq = 48% chance that the zygote will have the homozygous Aa genotype.
4. all of the zygotes will have one of the three genotypes
AA, Aa, or aa. therefore p2 + 2pq + q2 = 16% + 48% + 36% = 100%.
in a population that is at equilibrium, one would expect these results simply due to genetic recombina-tion: 16% AA, 48% Aa, and 36% aa. this principle was fi rst explained by G. H. Hardy, a mathematician, and W. Weinberg, a biologist, and is therefore called the Hardy-Weinberg equilibrium. if the population differs from this pattern, the departure from equilib-rium could be caused by a number of possibilities:
1. the population may not be very large. Ran-dom departures from equilibrium can occur in small populations. this is called genetic drift.
2. the mixture of gametes may not be occur-ring randomly among individuals in the population. this could be caused by non-random mating patterns, in which individu-als within the population prefer to mate with one genotype more than with another.
3. Mutations have occurred.4. Migration (of one genotype more than
another) has occurred into or out of the population.
5. natural selection has occurred.
in this last possibility, one of the alleles may be less frequent than its equilibrium value because
If allele A is present in 40 percent of the gametes (eggs and sperm) of a population and allele a is in 60 percent of the gametes, the resulting fre-quencies of the organisms under conditions of Hardy-Weinberg equilib-rium will be 16 percent AA, 48 percent Aa, and 36 percent aa.
xviii+598_Biodiversity_F.indd 433 11/17/11 1:13 PM

first entry434 population genetics 434
natural selection has “selected against” individuals that carry this allele. the Hardy-Weinberg equilib-rium is the background against which natural selec-tion, defined as changes in allele frequencies in a population, might be detected.
if natural selection operates against an allele, the allele will become less common in the popula-tion. Strong selection against a dominant allele (an allele whose presence always shows up in the pheno-type) may quickly eliminate that allele. (this rarely occurs, because an allele that produces an inferior phenotypic trait would probably never have become dominant in the first place.) Strong selection against a recessive allele (such as a lethal mutation), how-ever, may never eliminate the allele from the popula-tion. this is because the recessive allele can hide in the heterozygotes; the heterozygotes are therefore carriers of this allele. a lethal recessive will decline in frequency according to this formula:
q0qn = — 1 + nq0
in which q is the frequency of the lethal recessive allele, q0 is the initial frequency of the allele, and qn is the frequency after n generations. Starting with a frequency of 25 percent, the allele will decline to a frequency of 8 percent after eight generations. even after a hundred generations, the frequency of this recessive allele will still be just under 1 percent. When the allele is at very low frequency, it may be lost from the population by random events such as genetic drift.
Sometimes, a recessive allele might be lethal in the homozygous form, but might actually confer a survival or reproductive advantage in the het-erozygous form. this can lead to a balanced poly-morphism that maintains both alleles because the balance between advantage and disadvantage can cause more than one form of the allele (a poly-morphism) to be maintained in the population. the most famous example is sickle-cell anemia in humans, which is caused by a recessive mutation in one of the hemoglobin genes. the mutation causes anemia, because the blood cells that have this hemoglobin are sickle-shaped and are destroyed. However, the malaria parasite cannot reproduce in these blood cells. therefore, in regions (usually cen-tral africa) where malaria is common, people who are homozygous for the mutant die from anemia, but people who are homozygous for normal hemo-globin often die of malaria. People who are hetero-zygous for the mutation strike a balance between anemia and malaria, avoiding malaria and having relatively mild anemia. natural selection therefore favors this mutation, because there are many more
heterozygous people than homozygous recessive people.
another possible example of a balanced poly-morphism involves cystic fibrosis. individuals homo-zygous for this mutation usually die at a young age; the carriers of the mutation, however, are largely unaffected. Population geneticists have been puzzled as to why this mutation is so common in certain (often northern european) populations. they con-cluded that there must be a heterozygote advantage, and it probably is connected with resistance to cer-tain diseases.
why do populaTIoNS havE So muCh varIaBIlITy?
When natural selection began to gain popularity as the mechanism of evolution, geneticists were attracted to the idea that natural selection main-tained this genetic variability. Diversifying selec-tion in a heterogeneous environment would be able to do so. they considered that there had to be a good evolutionary reason why there was so much genetic variation. the reason cannot be that populations need it so that they can evolve. natu-ral selection can produce only those characteristics that provide a benefit to individuals, not to whole populations. the primacy of natural selection in producing and maintaining genetic variability was central to the contributions of mathematicians R. a. Fisher and J. B. S. Haldane to the mod-ern synthesis. in contrast, geneticist Sewall Wright emphasized random events such as genetic drift in the course of evolution.
With the advent of electrophoresis, it became apparent that there was a lot more genetic variabil-ity in populations than could be seen in the pheno-type. Populations appeared to accumulate neutral variation—genetic variation that was neither ben-eficial nor harmful. Beginning in the late 1960s, the neutral theory of evolution, proposed by geneti-cist Motō Kimura, emphasized the primacy of neu-tral genetic variation in populations. this offered a challenge to the selectionist arguments of Fisher and Haldane and was more consistent with the genetic drift of Wright.
INBrEEdINGinbreeding occurs when close genetic relatives inter-breed. Most individual organisms have a few lethal recessive mutations that originated in relatively recent ancestors. in the heterozygous state, these mutations have no effect on the phenotype. So long as the individual does not mate with another individual that carries the same mutation (that is, a close rela-tive descended from the recent ancestor that had the mutation), the mutation will not appear in the homo-
xviii+598_Biodiversity_F.indd 434 11/17/11 1:13 PM

population genetics 435
zygous form. When two carriers interbreed, there is a one-in-four chance that the offspring will receive a double dose of the mutation and die as a result.
Small populations often have reduced genetic variability. there is a greater risk that individuals in these small populations will crossbreed with another individual that carries the same lethal mutation. this is one of the principal problems with species that are close to extinction. even if they are rescued and given a safe habitat in which to live, these spe-cies (such as the Hawaiian goose) will have poor reproductive success. the genetic variability that they lost is impossible to replace in the short run and perhaps forever. this is one of the major problems in rescuing endangered species.
inbreeding may occur in humans for cultural reasons. in isolated villages, populations are often founded by a small number of people with limited genetic variability. Most people mate with other people in their village, with the result that they often share lethal recessive mutations with their mates. One example comes from the Domini-can Republic village of Salinas (population about 4,000), where about one out of every 90 girls turns into a boy when reaching puberty. this happens so frequently that the villagers have a name for it: guevedoche, or “penis at twelve.” a similar phe-nomenon occurs among the Sambia tribe of Papua new Guinea, where they call it kwolu-aatmwol in their native tongue or “turnim-men” in pidgin. Women have two X chromosomes, while men are Xy. Gender, however, turns out to be due to more than just sex chromosomes. the guevedoche and kwolu-aatmwol boys had y chromosomes and pro-duced testosterone like regular boys; however, they lacked an enzyme called 5-α-reductase. the gene encoding this enzyme was defective. this enzyme changes testosterone into a related molecule (DHt) that stimulates genital development in the fetus. therefore, during gestation and childhood, these boys had genitals that appeared female. However, at puberty, testosterone itself stimulates and maintains masculinity; therefore, at puberty, the boys devel-oped male genitalia. all of the guevedoche boys in the Dominican Republic can trace their ancestry to one woman, altagracia carrasco; she was probably the person who carried the mutation. inbreeding can also occur in religious communities that remain apart from the rest of society. Some religious sects such as the amish have elevated levels of occasion-ally lethal genetic diseases.
Summarynatural populations have genetic variability. even those species that produce offspring without sexual recombination have at least some genetic variability
due to mutation. Genetic variability in populations can be measured on phenotypic traits or on pro-teins. the Hardy-Weinberg equilibrium expresses the mixture of genes that will result in the absence of natural selection. Recessive lethal mutations can be eliminated only slowly from populations; lethal mutations may be selected if they provide a benefit to heterozygotes; and lethal mutations can show up more frequently in small populations, often of endangered species, than in large populations. inbreeding in small populations, which have less genetic variability, is one of the major problems in the survival of endangered species.
See also Darwin’s finches; founder effect and genetic drift; natural selection.
Further readingavise, John c. Population Genetics in the Wild. Washing-
ton D.c.: Smithsonian, 2002.ayala, Francisco J. Population and Evolutionary Genetics:
A Primer. Menlo Park, calif.: Benjamin-cummings, 1982.
Bradshaw, corey J. a., et al. “low genetic diversity in the bottlenecked population of endangered non-native banteng in northern australia.” Molecular Ecology 16 (2007): 2,298–3,008.
Fallon, Sylvia M. “Genetic data and the listing of species under the U.S. endangered Species act.” Conservation Biology 21 (2007): 1,186–1,195.
Freeman, Scott, and Jon c. Herron. “Mechanisms of evolu-tionary change.” Part 2 of Evolutionary Analysis. 4th ed. Upper Saddle River, n.J.: Pearson Prentice Hall, 2007.
Ge, Song, et al. “comparisons of genetic diversity in the endangered Adenophora lobophylla and its wide-spread congener, A. potaninii.” Conservation Biology 13 (1999): 509–513.
Gibson, J. Phil, Stanley a. Rice, and clare M. Stucke. “comparison of population genetic diversity between a rare, narrowly distributed species and a common, widespread species of Alnus (Betulaceae). American Journal of Botany 95 (2008): 588–596.
Godoy, José a., et al. “Phylogeography, genetic struc-ture, and diversity in the endangered bearded vulture (Gypaetus barbatus l.) as revealed by mitochondrial Dna.” Molecular Ecology 13 (2004): 371–390.
Hughes, Jennifer B., Gretchen c. Daily, and Paul R. ehrlich. “Population diversity: its extent and extinction.” Sci-ence 278 (1997): 689–692.
Kaneko, Shingo, et al. “Genetic differentiation among pop-ulations of an oceanic island: the case of Metrosideros boninensis, an endangered endemic tree species in the Bonin islands.” Plant Species Biology, 23 (2008): 119–128.
lawrence, Hayley a., et al. “new genetic approach to detecting individuals of rare and endangered species.” Conservation Biology 22 (2008): 1,267–1,276.
xviii+598_Biodiversity_F.indd 435 11/17/11 1:13 PM

first entry436 predation 436
leRoi, armand. Mutants: On Genetic Variety and the Human Body. new york: Viking, 2003.
lewontin, Richard. The Genetic Basis of Evolutionary Change. new york: columbia University Press, 1974.
liu, P., et al. “ecological risk assessment using RaPD and distribution pattern of a rare and endangered species.” Chemosphere 68 (2007): 1,497–1,505.
Ruder, Kate. “Genomics in amish country.” Genome news network. available online. URl: http://www.genomenewsnetwork.org/articles/2004/07/23/sids.php. accessed august 8, 2011.
Williams, Barry l. “conservation genetics, extinction, and taxonomic status: a case history of the regal fritillary.” Conservation Biology 16 (2002): 148–157.
predation a predator is an organism that kills another organism, its prey, in order to consume part or all of it. Most familiar examples of preda-tion are of predatory animals eating other animals. However, an animal can be a predator even when it eats plants. animals that eat and digest seeds are often considered to be “seed predators,” since the seed contains an entire embryonic plant. Plants such as the Venus flytrap can be predators upon animals.
Predatory animals have evolved numerous adaptations that allow them to pursue their prey (see photograph on page 437). in general, preda-tors have the swiftest movement, sharpest eyesight, and most intelligence in the animal kingdom. not only do predators have keen eyesight, but their brains allow them to instantly recognize the move-ment of prey distinct from the movement of, for example, leaves in the wind. nocturnal predators have particularly keen senses. Some populations of predators, such as wolves, work together in packs to locate and capture prey.
CoEvoluTIoN of prEdaTorS aNd prEyPrey species have numerous defenses against preda-tors. examples include the following:
● Some prey species have camouflage. they may have coloring that resembles the leaves, stems, litter, or rocks of their environment. For example, a certain species of insect in Madagascar is bright red and forms clus-ters on branches, thereby resembling flowers and escaping the notice of predators. Prey may hide themselves not only visually but by olfactory means as well. Some califor-nia ground squirrels have been observed to chew on rattlesnake skins and smear the scent on their fur, which masks their own scent from predators.
● in contrast, some prey animals have col-ors that make them very conspicuous to predators. Rather than making them likely targets for predation, such coloration pro-tects them, because this warning coloration announces the fact that the prey is poison-ous. Poisonous frogs, unlike other frogs, are brightly colored and are active during the day rather than the night. Warning col-oration usually evolves in poisonous prey, because the poison confers no benefit on the prey if the ignorant predator eats it and discovers too late that it is poisonous. the adult monarch butterfly retains the poisons that it consumed from milkweeds when it was a caterpillar. it has orange and black warning coloration.
● the prey may use a startle display to tem-porarily confuse the predator while they escape. Some insects have large eyespots on their bodies that can briefly make them appear to be a large animal.
● Some prey produce chemicals that irritate or even endanger predators. Skunks are notori-ous for this defense. Many amphibians are famous for having poisons in their skins. the only poisonous bird (Pitohni dichrous) has the same kind of toxin in its feathers as do certain poisonous frogs. Whip scorpions produce a spray that contains oily caprylic acid that penetrates the skin or exoskeleton of a predator and watery acetic acid that burns it. an insect known as the devil’s rider walking stick sprays streams of terpenes at its attackers. Some millipedes produce pro-gesterones, which reduce the fertility of ver-tebrates that eat them.
● Some prey animals have structural defenses such as the quills of the porcupine or the poison glands of the zebra fish. certain liz-ards have tails that detach when a predator grabs them by the tail.
● Some animals such as the hippopotamus and the elephant are safe from predation because of their large size. large size may have evolved in these species because the larger individuals in the populations were more resistant to attack by predators.
● Prey animals may even individually attack predators, if they can outmaneuver them. it is a common sight to see small birds attack-ing crows and hawks by diving on them from above.
● Some prey animals are protected by large numbers. Prey animals are often found in large single-species groups such as schools
xviii+598_Biodiversity_F.indd 436 11/17/11 1:13 PM

predation 437
of fish or herds of grazing animals. a lion is less likely to attack a whole group of wil-debeest than an isolated individual. Some herds have sentries that watch for preda-tors and sound an alarm if one is sighted. Flocks of birds make rapid coordinated movements that may startle the predator. When a large number of prey animals is born in one year, the predators cannot pos-sibly eat all of them; this predator satia-tion allows many of the prey to survive. the 17-year cicada hatches in massive numbers every 17 years; because 17 is a prime number, predator population cycles have not adjusted to the time of emergence of this prey.
● though, in general, prey animals have less intelligence than predators, they still have adaptations that allow them to detect the presence of predators and react quickly. they, too, have eyes and brains that allow them to recognize the movement of a predator against a background of leaves in the wind.
Some researchers have even suggested that migration is a prey adaptation to escape from preda-tors. they suggest that some birds fly all the way up into the arctic during the summer because there are fewer predators to disturb their nests.
Perhaps the most important adaptation that prey species have is reproduction. Prey animals are notorious for producing large numbers of small offspring, most of which die by predation and by many other means. Prey animals also reproduce at a young age, often in their first year of life. in the
presence of predators, natural selection favors a life history of early and abundant reproduction. in the absence of predators, these same animals may have later reproduction and produce fewer, larger offspring. a population of opossums (Didelphus virginiana) became isolated on an island off the coast of eastern north america, without preda-tors. these opossums lived longer and reproduced when older, compared to mainland populations of opossums susceptible to predator attack. On the island of trinidad, most freshwater pools con-tain guppies (Poecilia reticulata). in some of these pools, there are also predatory fishes; in other pools, the predators are absent. the guppies that live in pools without predators live longer and reproduce when they are older.
the old and young prey animals, along with any that are inferior in their defenses, are most often caught by predators. in this way, predators unintentionally help to maintain the health of the prey population. this results in natural selection for prey that defend themselves well against preda-tors as well as for predators that successfully catch prey.
in some cases, both predators and prey have evolved tricks that maximize their success. these adaptations are called mimicry because one spe-cies has characteristics or behavior patterns that resemble (mimic) those of another species. examples include the following:
● the angler fish attracts smaller fish with an extension from its head that resembles a worm.
● One species of spider eats female moths and then uses their scent to attract male moths.
● certain spiders can enter ants’ nests with impunity because they can imitate the touch responses and the chemical signals that the ants use to recognize the other members of their own colony.
● Sometimes the prey species has warning col-oration, even though it is not poisonous. the viceroy butterfly, for instance, looks very much like the poisonous monarch but-terfly; even though it is not poisonous, most predatory birds avoid eating it.
adaptations such as camouflage and mimicry do not need to be perfect in order to be favored by natu-ral selection. natural selection has produced some adaptations that appear almost perfect to human observers—for example, a stick-insect that really looks like a stick. However, even an adaptation that only partially camouflages an animal, or gives it only a partial mimicry of another species, can be
Ospreys (Pandion haliaetus) prey upon fish in North America. (Dunedin Osprey Cam)
xviii+598_Biodiversity_F.indd 437 11/17/11 1:13 PM

first entry438 predation 438
favored by natural selection. camouflage and mim-icry that appear unconvincing to a human observer may be perfectly convincing to a predatory insect with a small brain or to a predator with imper-fect eyesight or under conditions of limited visibility (such as dusk).
Plants and even fungi can be predators:
● the leaf of the Venus flytrap closes around an insect when its movement triggers tiny hairs on the leaf; once the insect is inside, the leaf secretes digestive enzymes.
● in the pitcher plant, insects fall into a pool of water and digestive enzymes and cannot escape, partly because hairs inside of the pitcher point downward.
● in the sundew plant, insects adhere to the sticky hairs on the leaves, where enzymes digest them.
● there is even a fungus (Arthrobotrys oligos-pora) that captures and consumes nematodes.
carnivorous plants live in bogs, in which the plants have access to very few mineral nutrients. the plants make their own energy compounds by photo-synthesis, and they use the insects solely as a source of fertilizer, especially nitrogen compounds.
Predation is a good source of high quality food. in contrast to fruits and leaves and even seeds, ani-mals are rich in protein and fat. Most birds will feed insects to their nestlings, even if they primarily eat seeds. even herbivores will occasionally eat meat if the opportunity presents itself; they are simply not well adapted to catch prey. Many people have seen squirrels eating roadkill, even of other squirrels. People who raise chickens know that they have to protect them from deer and even cows will eat birds that are trapped in ornithologists’ mist nets.
the coevolution of predators and prey can lead to an “evolutionary arms race.” For example, some populations of newts (a type of amphibian) contain enough toxin in their skins to kill dozens of preda-tors. this would seem to be an excessive expendi-ture. But they are not defending themselves against predators in general but against garter snakes. Some garter snakes produce a large amount of enzyme that detoxifies the newt’s defensive chemical. the more poisonous the newts become, the better the snakes become at detoxifying them, in a spiral of coevolu-tion. there is a price to pay. the garter snakes that can detoxify the newts are more sluggish than other garter snakes.
in some cases, a predator can use the prey’s defensive chemicals to defend itself. the snake Rhabdophis tigrinus consumes venomous toads but uses the toad’s toxin to make its own skin poisonous.
prEdaTIoN aNd populaTIoNSPredation plays an immensely important role in con-trolling prey populations. the number of insects eaten by birds and bats is virtually incalculable. the extinction of a top predator (which is an example of a keystone species) can result in a prey popula-tion explosion. Some conservationists claim that the hunting of wolves and mountain lions has resulted in explosions of deer populations in the mountains of western north america, although this interpretation remains controversial.
Perhaps the most famous example of predators affecting populations of prey is the story of the canada lynx (Lynx canadensis) and the snowshoe hare (Lepus americanus). approximately once a decade, the hare populations explode. a few years later, the lynx populations also increase, in response to the increased availability of their food. the hare population then declines. the lynx populations also decline, in response to a lack of food. these cycles of explosion and decline have been going on for many decades, according to data from companies that buy furs of both species from trappers. What is unclear is whether the predators are actually in control of the prey population cycles. Does the lynx popula-tion bring the hare population back down, or do the hares experience a population crash due to disease or because they run out of food? it is possible that the lynx are merely responding to, rather than con-trolling, the hare populations.
humaNS aS prEdaTorSthe earliest members of the human lineage were gatherers with brains no larger than those of the chimpanzee relatives from which they had recently diverged. a rapid evolutionary increase in human brain size occurred at the same time as the develop-ment of stone tools and the consumption of other animals. the stone tools of early members of the human genus (such as Homo habilis and H. ergaster) were probably not used for hunting. archaeological evidence indicates that these early members of the genus Homo were scavengers. Only later, with the development of thrusting spears by neanderthals and of bows and arrows by early modern humans, did hunting became a major source of food. even then, meat constituted a smaller portion of the diet than did seeds, fruits, and roots, and small animals made up most of that portion. Many evolution-ary scientists believe that hunting evolved because it rewarded successful hunters with social status and therefore more mating opportunities.
in regions where humans and their prey animals evolved together for thousands or millions of years, humans did not threaten most prey populations with extinction. However, in places where fully
xviii+598_Biodiversity_F.indd 438 11/17/11 1:13 PM

predation 439
modern humans arrived with advanced hunting technologies such as bows and arrows, prey exter-mination sometimes resulted. the major example is the Pleistocene extinction in north america at the end of the most recent ice age, probably caused by overhunting of prey that had not evolved the instinct to run from humans. (Recent evidence has called this theory into question.) the evidence that overhunting caused extinctions in australia and Madagascar is better.
after a few thousand years, humans in these regions developed social traditions that allowed a rough equilibrium of prey populations to develop. even if their Pleistocene ancestors had caused mass extinctions, native americans were excellent man-agers of prey populations at the time of european contact. Before european contact, native americans hunted many animals (such as bison and passenger pigeons) and caught a lot of fish. european diseases decimated native populations, often even before they came in contact with the europeans. this may have been the reason that, during the 18th century, some of these prey animals experienced popula-tion explosions. the huge populations of bison and passenger pigeons of the 18th and 19th centuries are legendary. they were probably also unnatural. When botanist William Bartram traveled in what is now the southeastern United States in the late 18th century, he saw huge populations of fishes, and large numbers of alligators eating them. it did not occur to him, nor to virtually anyone else until recently, that this may have resulted from the plagues that depopu-lated native villages.
White american hunters proceeded to celebrate the enormous populations of prey animals by shoot-ing as many of them as possible. Billions of passen-ger pigeons had darkened the skies of the eastern United States in the 19th century, but by the early 20th century, the last passenger pigeon had died in a zoo. Many white american hunters of the 19th century aspired to “shoot the last buffalo” (see pho-tograph). the extermination of the bison was part of a deliberate, government-backed campaign to undermine the livelihood of the last native ameri-can tribes and bring them into subjugation. this goal was achieved before the bison became extinct, and today bison populations are growing in wild-life preserves (such as Badlands national Park in South Dakota and the nature conservancy’s tall-grass nature Preserve in Oklahoma) and on private ranches.
prEdaTorS aNd CoNSErvaTIoNWildlife management developed in the 20th century as a way of making hunting sustainable, allowing it to continue without depleting prey populations. in this way, humans once again play an important role in stabilizing prey populations, because hunters are allowed only a limited number of prey, a limit that is adjusted each year. the role of hunters is now essential, since hunters in the late 19th and early 20th centuries nearly exterminated other large pred-ators such as wolves and bears that had previously kept prey populations in check. Populations of large predators are now recovering, in some cases after the deliberate reintroduction of animals from zoos (in the case of european bears) or from other areas in which the animals were still numerous (as in the case of wolves in the Rocky Mountains). these large predators normally stay away from livestock, due to a fear of humans, but not always, and ranchers often oppose the return of wild predators.
Because of the “ten percent law” of the transfer of energy and matter through the food chain, predator populations tend to be smaller than prey populations. Herbivorous prey animals consume food that does not move, but predators often need to search a large area for their prey. For both of these reasons, it is often the critical area for a viable predator population that determines the area of a nature preserve. Because their populations grow relatively slowly, predator spe-cies are often the most vulnerable to environmental disruptions. Because predators are at the top of the food chain, they are the most frequent victims of the biomagnification of toxic materials. together with the primordial human reverence for predators, these rea-sons make predators the “poster children” of many conservation campaigns. it is not without reason that conservationists believe that the health of a predator
In the 1870s, white Americans shot huge numbers of bison, partly in a deliberate attempt to destroy the food source of unconquered Native American tribes. In this image, a pile of bison skulls is waiting to be ground for fertilizer. (Burton Historical Collection; Detroit Public Library)
xviii+598_Biodiversity_F.indd 439 11/17/11 1:13 PM

first entry440 predation 440
population is a good indicator of the health of the entire ecosystem of which it is a part.
See also biomagnification; camouflage and mimicry; carnivorous plants; coevolution; conservation biology; ecosystems; keystone species; life history, diversity of; Pleistocene extinction.
Further readingaustad, Steven n. “Retarded senescence in an insular pop-
ulation of Virginia opossums (Didelphus virginiana).” Journal of Zoology 229 (1993): 697–708.
Barbosa, Pedro, and ignacio castellanos, eds. Ecology of Predator-Prey Interactions. new york: Oxford Uni-versity Press, 2005.
Brodie, edmund D., Jr., et al. “the evolutionary response of predators to dangerous prey: Hotspots and cold-spots in the geographic mosaic of coevolution between garter snakes and newts.’ Evolution 56 (2002): 2,067–2,082.
Brodie, edmund D., iii, et al. “Parallel arms races between garter snakes and newts involving tetrodotoxin resis-tance as the phenotypic interface of evolution.” Jour-nal of Chemical Ecology 31 (2005): 343–356.
Brodie, edmund D., iii, and edmund D. Brodie, Jr. “evo-lutionary response of predators to dangerous prey: reduction of toxicity of newts and resistance of garter snakes in island populations.” Evolution 45 (1991): 221–224.
———. “Predator-prey arms races.” Bioscience 49 (1999): 557–568.
carlson, S. M., et al. “Does variation in selection imposed by bears drive divergence among populations in the size and shape of sockeye salmon?” Evolution 63 (2009): 1,244–1,261.
———, et al. “Predation by bears drives senescence in natural populations of salmon. Public Library of Sci-ence ONE 2 (2007): e1,286.
corcoran, aaron J., et al. “tiger moth jams bat sonar.” Sci-ence 325 (2009): 325–327.
Dicke, Marcel, and Paul Grostal. “chemical detection of natural enemies by arthropods: an ecological perspec-tive.” Annual Review of Ecology and Systematics 32 (2001): 1–23.
eisner, thomas e. “Beetle’s spray discourages predators.” Natural History 75 (1966): 42–47.
Feldman, c. R., et al. “the evolutionary origins of beneficial alleles during the repeated adaptation of garter snakes to deadly prey.” Proceedings of the National Academy of Sciences USA 106 (2009): 13,415–13,420.
Genovart, M., et al. “the young, the weak, and the sick: evidence of natural selection by predation.” Public Library of Science ONE 5 (2010): e9,774.
Godeau, J.-F., et al. “Reaction of ants to, and feeding biology of, a congeneric myrmecophilous and non-
myrmecophilous ladybird.” Journal of Insect Behavior 22 (2009): 173–185.
Gosler, a. G., et al. “Predation risk and the cost of being fat.” Nature 377 (1995): 621–623.
Kenward, R. e. “Hawks and doves: Factors affecting success and selection in goshawk attacks on woodpigeons.” Journal of Animal Ecology 47 (1978): 449–460.
McKinnon, l., et al. “lower predation risk for migratory birds at high latitudes.” Science 327 (2010): 326–327. Summarized by Gilg, Olivier, and nigel G. yoccoz. “explaining bird migration.” Science 327 (2010): 276–277.
Møller, anders Pape, and J. erritzøe. “Predation against birds with low immunocompetence.” Oecologia 122 (2000): 500–504.
neves, R., et al. “Does a shell matter for defence? chemical deterrence in two cephalaspidean gastropods with cal-cified shells.” Journal of Molluscan Studies 75 (2009): 127.
nilsen, erlend B. “a slow life in hell or a fast life in heaven: Demographic analyses of contrasting roe deer popula-tions.” Journal of Animal Ecology 78 (2009): 585–594.
nordbring-Hertz, Birgit, and Bo Mattiasson. “action of a nematode-trapping fungus shows lectin-mediated host-microorganism interaction.” Nature 281 (1979): 477–479.
Osborn, Karen J., et al. “Deep-sea, swimming worms with luminescent ‘bombs.’ ” Science 325 (2009): 964.
Pafilis, Panayiotis, et al. “tail shedding in island lizards (lacertidae, Reptilia): Decline of antipredator defenses in relaxed predation environments.” Evolution 63 (2009): 1,262–1,278.
Penteriani, V., et al. “Owls and rabbits: Predation against substandard individuals of an easy prey.” Journal of Avian Biology 39 (2008): 215–221.
Reznick, David, and John a. endler. “the impact of preda-tion on life history evolution in trinidadian guppies (Poecilia reticulata).” Evolution 36 (1982): 160–177.
Servedio, M. R. “the effects of predator learning, forget-ting, and recognition errors on the evolution of warn-ing coloration.” Evolution 54 (2000): 751–763.
Sih, andrew. “evolution, predator avoidance, and unsuc-cessful predation.” American Naturalist 125 (1985): 153–157.
Stanford, craig B. Chimpanzee and Red Colobus: The Ecology of Predator and Prey. cambridge, Mass.: Harvard University Press, 1998.
Stenseth, nils c., et al. “Population regulation in snow-shoe hare and canadian lynx: asymmetric food web configurations between hare and lynx.” Proceedings of the National Academy of Sciences USA 94 (1997): 5,147–5,152.
Stolzenburg, William. Where the Wild Things Were: Life, Death, and Ecological Wreckage in a Land of Vanish-ing Predators. new york: Bloomsbury, 2008.
xviii+598_Biodiversity_F.indd 440 11/17/11 1:13 PM

primates 441
Svensson, e. i., and M. Friburg. “Selective predation on wing morphology in sympatric damselflies.” American Naturalist 170 (2007): 101–112.
Williams, Becky l., et al. “a resistant predator and its toxic prey: Persistence of newt toxin leads to poisonous (not venomous) snakes.” Journal of Chemical Ecology 30 (2004): 1,901–1,919.
young, Kevin V., et al. “How the horned lizard got its horns.” Science 304 (2004): 65.
primates Primates constitute the class of mam-mals that includes lemurs, monkeys, and apes. Humans are apes, therefore primates. the term pri-mate, used by scientists since the time of carolus linnaeus, suggests that this group, which includes humans, is the most important among mammalian classes. While scientists no longer rank classes by their importance relative to humanity, the term pri-mate continues in use.
Biologists classify primates into approximately 230 living species of prosimians, tarsiers, and anthropoids (see figure on page 442). Primates, in general, share these characteristics:
● they have the largest brains among mam-mals, after adjustment for body size.
● they often live in large, complex social groups. large brains aid survival but are considered to be most important for social interactions.
● they take a longer time to reach maturity than most other mammals, a characteristic related to the evolution of intelligence.
the first primates lived in trees, and most still do, except for a few groups such as baboons and humans that have (independently from one another) evolved the ability to live on the ground. this adaptation is complete enough that in humans, uniquely among primates, the lower limbs (legs) are relatively long. Some, like galagos, have remarkable leaping abilities: though less than a foot and a half (50 cm) in length, galagos can leap more than six feet (2 meters). leap-ing from branch to branch in trees has encouraged the evolution of important characteristics that pri-mates possess more than most other mammals:
● The ability to grasp. this results primarily from an opposable thumb, which is rare in other mammals. Primates not only have the ability to grasp branches in trees, but the juvenile primates can efficiently cling to the fur of their mothers. Most primates also have opposable big toes, although primates
that have evolved a ground-based existence have lost this characteristic. nonhuman apes have opposable big toes; humans do not; the australopithecines (human ances-tors) had big toes that were intermediate in their adaptation for grasping branches.
● Excellent vision. nocturnal primates have big eyes. For example, each of the tarsier’s eyes is larger than its brain. Diurnal pri-mates have inherited big eyes from noctur-nal ancestors. Visual acuity is important not only for night vision, but for jumping around in trees. For prosimians, smell remains an important sense, while most anthropoids rely primarily on vision.
● Depth perception. Primates have both eyes on the front of the face, a characteristic most people notice immediately in all pri-mates. this allows binocular vision and depth perception, which allows primates to grasp branches while leaping.
● Color vision. While not the only mam-mals with color vision, primates (primar-ily anthropoids) benefit from color vision because it allows some primates to recog-nize ripe fruits in colors such as red against a background of green leaves, in the absence of a well-developed sense of smell.
Prosimians are the more primitive group of pri-mates and are generally smaller than anthropoids. Most prosimians are lemurs that live in Madagas-car, but others live on the african continent and in southeast asia. Lemurs range in size from the pygmy mouse lemur (weighing about an ounce, or 30 g) to the indri (weighing about 20 pounds, or 9 kg). Some lemurs are diurnal, others nocturnal. Galagos and lorises live on the african continent; lorises also live in india and Southeast asia. Many zoologists clas-sify prosimians into the subclass Strepsorhini.
Tarsiers (see photograph on page 443) are small, nocturnal primates that live in Southeast asia. though they look to many people very similar to the prosimians, they share some features with the anthropoids, such as skull characteristics, and big hands and feet. Many zoologists classify tar-siers together with anthropoids into the subclass Haplorhini.
Anthropoids (monkeys and apes) have the larg-est brains among primates and include two fami-lies of new World monkeys, found in the tropical rain forests of central and South america, as well the Old World monkeys, the lesser apes, and the great apes, all of which originated in africa or asia. Most monkeys and apes have longer arms than do
xviii+598_Biodiversity_F.indd 441 11/17/11 1:13 PM

fi rst entry442 primates 442
prosimians, which allow them to not merely jump around in trees but to swing from branches, a unique kind of locomotion called brachiation. Most anthro-poids are diurnal. Most have highly developed social patterns, which may be the principal reason for their braininess. For example, when howler monkeys of the amazon recognize a threat (for instance, in the form of a human intruder), they may form into a circle in the branches over the intruder’s head, and it is time for the human to leave.
new World monkeys have evolved separately from other primates for at least 20 million years, largely as a result of the separation of the african and South american continents. all live in trees. their nostrils are fl at and face sideways, which is why they are called platyrrhines (fl at-nosed). the most noticeable distinction is that they have long prehensile tails that have muscles and even fi n-gerprint skin that allows these monkeys to grasp branches with their tails. new World monkeys have
smaller thumbs (spider monkeys have none at all) than Old World monkeys and apes. the marmosets and tamarins of South america differ from other new World monkeys in having only two, rather than three, molars and are classifi ed separately.
the Old World monkeys and the apes are cat-arrhines, so named because their nostrils are nar-row and face downward. Old world monkeys live primarily in trees, except baboons, but spend part of their time on the ground. they are much less likely to have long tails, and these tails are not prehensile. their rump skin pads help them sit on the ground but are also useful in sexual signal-ing: colors indicate species and estrus, which is the female readiness for mating. the colobine subfam-ily of Old World monkeys includes the langurs of asia and the colobus monkeys of africa. they have relatively longer legs, tails, and thumbs than other Old World monkeys. although they eat a variety of foods, their large multichambered stomachs allow
the digestion of leaf material. a diet of leaves, most of which are toxic, requires the assistance of microbes in mammals such as the colobus monkey, the cow, and the marsupial koala; even the marine iguanas of the Galápagos islands, which live on seaweed, require microbial symbionts. the cerco-pithecine subfamily of Old World monkeys includes the macaques of africa and asia (some extend their range a little bit into europe) and the baboons and mangabeys of africa. cercopithecines have less complex stomachs and eat less leaf material. the Japanese macaque monkeys live farther north than any other nonhuman primate. their intelligence and cultural propagation of knowl-edge has been studied as a model for the evolution of intelligence.
Apes have no tails and have larger brains than other anthro-poids. apes have at least a limited ability to walk on two legs (biped-alism). Lesser apes include the gibbons and siamangs of South-east asia, which have extremely long arms with which they brachi-ate. “lesser” refers to the smaller size of these apes than the great apes. lesser apes, unusual among animals, have a lifelong monog-amous mating system. Great
A commonly accepted classifi cation system (which does not include all the families)
xviii+598_Biodiversity_F.indd 442 11/17/11 1:13 PM

primates 443
apes (family Hominidae; the hominids) include the orangutans of Southeast asia as well as the gorilla, chimpanzee, bonobo, and humans that originated in africa. Because great apes do not have rump pads, they generally build nests in trees or on the ground for sleeping.
With males weighing as much as 160 pounds (about 80 kg), the orangutans of Borneo and Suma-tra are the largest arboreal primates. Orangutans are the least social of apes. they forage separately for fruit. Males and females interact only to mate, and the only social bond among orangutans is between mother and infants. With males weighing as much as 350 pounds (about 180 kg), the gorillas are the largest primates (see photograph on page 444). the three subspecies of gorillas (the mountain gorilla and the eastern and western lowland gorillas) live in the rain forests of central africa in small groups that eat plant materials. in both orangutans and gorillas, males weigh about twice as much as females, which is characteristic of the harem. On at least two occa-sions, gorillas have been observed to use wooden tools.
the apes commonly called chimpanzees consist of two species: the true chimpanzees (Pan troglo-dytes) and the pygmy chimpanzee or bonobo (Pan paniscus). true chimps range from dry woodlands to rain forests throughout central africa; pygmy chimps live in a small area south of the congo River. they eat a wide variety of foods, includ-ing leaves, fruits, seeds, insects, and occasionally red meat. they spend part of their time in trees and part on the ground (bonobos less than true chimps). chimps exhibit behaviors that scientists once thought were found only in humans, such as the use of tools (although they do not fash-ion stones into tools) and warfare. Most of these
behaviors were revealed only when researchers such as Jane Goodall spent long periods of time with chimp societies. true chimps are very aggressive, in contrast to bonobos, who live in small bands and engage in frequent, humanlike copulation. as primatologist Frans DeWaal says, “the chimpan-zee resolves sexual issues with power; the bonobo resolves power issues with sex.” Humans share a common ancestor with chimps and bonobos that lived only about 5 million years ago.
Primates similar to today’s prosimians evolved shortly after the cretaceous extinction that wiped out the dinosaurs and other large reptiles. Some other highly specialized orders of mammals, such as bats and whales, also evolved in the Paleocene epoch of the cenozoic period. When dinosaurs were no longer the dominant land animals, mammalian evolution exploded in diversity. One of the earliest primates, Purgatorius, may have evolved before the end of the cretaceous. Purgatorius, and the plesi-adapiform primates that evolved during the Paleo-cene, lived in europe and north america (which were still connected at that time). they resembled shrews. During the ensuing eocene epoch, adapid and onomyid primates evolved. it is unclear which of these groups may have been the ancestors of monkeys and apes. Both groups of eocene primates had larger brains, shorter faces, and a better ability to grasp than Paleocene primates and relied on sight more than smell. the first true monkeys evolved dur-ing the ensuing Oligocene epoch in the Old World. Oligocene primates from what is now the Fayyum deposit in egypt (such as Proconsul) may represent the first true apes.
Sometime after 7 million years ago, perhaps associated with the spread of drier conditions across africa, the human evolutionary lineage (now usually called the hominins) separated from the chimpanzee lineages. the earliest distinctive char-acteristic of the hominins was upright posture and walking on two legs, although numerous distin-guishing features (such as very large brains, fash-ioning stones into tools, use of fire, use of language) arose later in the hominin line. the earliest known hominins were Orrorin and Sahelanthropus from west africa and Australopithecus primarily from east africa. Many australopithecines (and related hominins such as Kenyanthropus) are known from the period between 5 and 2 million years ago in east africa. the plenitude of fossil information reveals a complex bush of hominin lineages, rather than a clear line of human ancestry. about the only thing regarding human ancestry of which anthro-pologists are certain is that the robust australopith-ecines (genus Paranthropus) did not lead to modern humans. From one of the gracile australopithecines,
The Philippine tarsier (Tarsius syrichta), about five inches (13 cm) long, is one of the smallest primates. (Roland Birkle)
xviii+598_Biodiversity_F.indd 443 11/17/11 1:13 PM

first entry444 primates 444
either known or yet to be discovered, the genus Homo evolved about 2 million years ago in east africa. today, Homo sapiens is the only surviving species of hominin.
the fact that humans are primates means that the physiology of the other primates most nearly resembles that of humans. it is for this reason that primates, especially monkeys and (even better) chimps, are favored for medical experimentation. For example, primates are the best subjects to use to study the transmission of viral diseases such as ebola, which apparently emerged from primate pop-ulations, and to try out malaria vaccines. Some pri-mates have simian immune deficiency viruses (SiV), similar to human immune deficiency viruses (HiV), and experimentation on monkeys and apes may allow researchers to find out how other primates prevent spread of the virus in their bodies. Primates can have or be induced to have human neurodegen-erative diseases such as Parkinson’s and alzheimer’s diseases. Because primate brains are the most simi-lar to humans, the study of their brain functions may help medical researchers understand appetite, aggression, depression, and anxiety.
Because monkeys and nonhuman apes have large brains and complex behavior, they may also experience such experimentation as cruelty, more so than other laboratory animals such as mice, rats, and dogs. For both reasons, medical experi-mentation upon primates has become particularly controversial.
Most nonhuman primates survive in remote areas away from human contact, and a large pro-portion of them are endangered. Most primates are endangered from habitat destruction, but they are
also endangered by predation by humans as “bush-meat” and for traditional medicine. Only a few hundred mountain gorillas survive, in the Virunga Mountains of east africa. Because of their restricted habitats, they are vulnerable to natural disasters from which they could easily recover if they still had their original ranges. Drought and forest fires in 1997–98 resulted in the deaths of many orangutans (Pongo pygmaeus) in Borneo, either directly, or by reducing their habitat, or because villagers shot them as they fled from the fires. their populations decreased from 23,000 before the fires to 15,400 after. there is good reason why saving primates is a high conservation priority.
See also mammals.
Further readingBeck, B., et al., eds. Great Apes and Humans: The Ethics of
Coexistence. Washington, D.c.: Smithsonian institute Press, 2001.
Brashares, J. S., et al. “Bushmeat hunting, wildlife declines, and fish supply in West africa.” Science 306 (2004): 1,180–1,183.
calvin, William H. A Brain for All Seasons: Human Evolu-tion and Abrupt Climate Change. chicago: University of chicago Press, 2002.
De Waal, Frans B. M. My Family Album: Thirty Years of Primate Photography. Berkeley: University of califor-nia, 2003.
Hart, Donna l., and Robert W. Sussman. Man the Hunted: Primates, Predators, and Human Evolution. new york: Westview, 2005.
Johanson, Donald, lenora Johanson, and Blake edgar. Ancestors: In Search of Human Origins. new york: Villard, 1994.
Kalpers, J., et al. “Gorillas in the crossfire: Population dynamics of the Virunga mountain gorillas over the past three decades.” Oryx 37 (2003): 326–337.
Keele, B. F., et al. “chimpanzee reservoirs of pandemic and nonpandemic HiV-1. Science 313 (2006): 523–526.
leakey, Richard, and Roger lewin. Origins Reconsidered: In Search of What Makes Us Human. new york: Doubleday, 1992.
Pennisi, elizabeth. “Genomicists tackle the primate tree.” Science 316 (2007): 218–221.
Primate Photo Gallery. “Primate Photo Gallery: chimpan-zees, gorillas, orang-utans, monkeys, and apes.” avail-able online. URl: http://www.primates.com. accessed august 3, 2010.
Rowe, noel. The Pictorial Guide to the Living Primates. east Hampton, n.y.: Pogonias Press, 1996.
———. “Primate conservation, inc.” available online. URl: http://www.primate.org. accessed august 8, 2011.
Rumbaugh, Duane M., and David a. Washburn. Intelligent Apes and Other Rational Beings. new Haven, conn.: yale University Press, 2003.
Like humans, gorillas are apes and provide exten-sive care to their offspring. (NOAA/Department of Commerce)
xviii+598_Biodiversity_F.indd 444 11/17/11 1:13 PM

protists 445
Sapolsky, Robert M. A Primate’s Memoir: A Neuroscien-tist’s Unconventional Life among the Baboons. new york: Simon and Schuster, 2001.
Seiffert, erik R. “Basal anthropoids from egypt and the antiquity of africa’s higher primate radiation.” Science 310 (2005): 300–304.
Stewart, caro-Beth, and todd R. Disotell. “Primate evolu-tion—in and out of africa.” Current Biology 8 (1998): R582–R588.
Stringer, chris, and Peter andrews. The Complete World of Human Evolution. london: thames and Hudson, 2005.
tattersall, ian, and Jeffrey Schwartz. Extinct Humans. new york: Westview, 2000.
templeton, alan R. “Out of africa again and again.” Nature 416 (2002): 45–51.
van Schaik, c. P., et al. “Orangutan cultures and the evolu-tion of material culture.” Science 299 (2003): 102–105.
Walker, alan, and Pat Shipman. The Ape in the Tree: An Intellectual and Natural History of Proconsul. cam-bridge, Mass.: Harvard University Press, 2005.
———. The Wisdom of the Bones: In Search of Human Origins. new york: Knopf, 1996.
Walsh, P. D., et al. “catastrophic ape decline in western equatorial africa.” Nature 422 (2003): 611–614.
Winckler, Wendy, et al. “comparison of fine-scale recombi-nation rates in humans and chimpanzees.” Science 308 (2005): 107–111.
Wood, Bernard, and M. collard. “the human genus.” Sci-ence 284 (1999): 65–71.
protists Protists are eukaryotic organisms that do not produce complex tissues. Protists are gener-ally considered to be simpler in structure than the “crown eukaryotes” (animals, plants, and fungi). Many protists are single-celled, though these single cells (such as Tetrahymena) can have very complex structures. if multicellular, they do not have highly organized tissues, although large seaweeds come pretty close to the complexity of plants, animals, and fungi.
all eukaryotes appear to have evolved from a common ancestor that had a nucleus and mito-chondria. according to the reconstruction used by evolutionary scientist Sandie Baldauf, eight major lineages of eukaryotes evolved. in one of the lin-eages, plants evolved; in another lineage, animals and fungi evolved. “Protists,” therefore, refers to all eukaryotes other than plants, animals, and fungi, making it a paraphyletic cladistic group. Kingdom Protista is therefore a conceptual category, not a distinct evolutionary group. Because they are not animals, and because seven of the eight lineages do not include animals, protists are no longer called protozoans (Greek for “original animals”).
Protists do not have adaptations for living on land, although many can survive desiccation. Pro-tists are found in every moist region of the planet and fulfill nearly every ecological role. Some are decomposers; some are predators; some cause dis-eases; and some are photosynthetic. in all of these ways, they are important components of natural ecosystems. Protists are most reliably classified on the basis of Dna similarities. Such a classification results in evolutionary lineages that are difficult to define in terms of anatomy and life cycle. Perhaps the best way to understand the enormous diversity of protists is to survey the lineages (see table).
ExCavaTE aNd dISCICrISTaTE proTISTSExcavates are single-celled protists with flagella and an “excavated” feeding groove on one surface. Some excavates live in the intestines of insects. a few excavates are intestinal pathogens of animals, including humans, such as Giardia lamblia, which causes giardiasis. Giardia is one of the diplomo-nads, which appear to be two cells fused together: Giardia has two nuclei, each associated with four flagella, and duplicated organelles. excavates lack mitochondria, but some of them have mitosomes and hydrogenosomes, which are vestigial remnants of mitochondria.
Discicristates are also single-celled protists with flagella. their name indicates that they have disc-shaped cristae (membranes) within their mito-chondria. they often, like excavates, have a feeding groove. examples include Euglena, which swims in water, guided by an eyespot that detects the direc-tion of light. Euglena makes its food by photosyn-thesis with its large chloroplasts. the chloroplasts
major lineages of protists
lineage Examples
Excavates and discicristates
Giardia, Euglena, acrasid slime molds
Heterokonts Diatoms, brown algae, water molds
Alveolates Ciliates, dinoflagellates, Plasmodium
Cercozoans Foraminifers
Amoebozoans Plasmodial slime molds
Green protists Red algae, green algae (and plants)
Opisthokonts Choanoflagellates (and fungi and animals)
xviii+598_Biodiversity_F.indd 445 11/17/11 1:13 PM

first entry446 protists 446
of plants and their ancestors (such as green algae) evolved from endosymbiotic cyanobacteria—that is, plant and green algae cells are “cells within cells.” in contrast, the chloroplasts of Euglena appear to have evolved from entire green algae cells, making the chloroplasts the evolutionary descendants of “cells within cells within cells,” an example of symbiogen-esis. Euglena can also ingest food particles in the same manner as animal cells. Some discicristates are intestinal pathogens of animals, such as the Try-panosoma that cause sleeping sickness. the acrasid slime molds are in this lineage also. acrasid cells are amoeba-like, which means that they flow across moist surfaces into extensions called pseudopods. they spend most of their time independently forag-ing for food, but under certain conditions some of them will release a hormone that causes the cells to gather together into a single amoeba-like organism that undergoes sexual reproduction and produces spores. the slime-mold life cycle has independently evolved in the amoebozoan lineage (see below). Some evolutionary scientists consider discicristates to be a lineage within the excavates.
hETErokoNT proTISTSHeterokonts (also called stramenopiles) are another major lineage of eukaryotes. their name is Greek for “different flagella,” because their flagella have hollow, three-pointed hairs unlike the flagella of other eukaryotes. Most of them are photosynthetic “algae,” but not all algae are heterokonts—just the diatoms, golden algae, and brown algae. their chlo-roplasts appear to be the evolutionary vestiges of sin-gle red algae cells (see below) that were incorporated by symbiogenesis (they are cells within cells within cells). Heterokont chloroplasts have chlorophyll a, just as do cyanobacteria and all other chloroplasts, and chlorophyll c, unique to this group. also unique is the fucoxanthin that gives these algae their golden or brown color. Brown algae store food in the forms of laminarin, a carbohydrate, and mannitol, a kind of alcohol, rather than as starch, as do green plants. Diatoms store oil, which is lighter than water and keeps them afloat.
Diatoms and golden algae are single-celled. Diatoms have shells made of silicate, the same min-eral found in glass; some of these shells can be intri-cately sculpted. Besides being major components of the phytoplankton on which aquatic food webs depend, diatoms can accumulate at the bottoms of shallow seas. Millions of years of such accumu-lation, followed by compression, results in thick layers of diatomaceous earth, an extremely light rock that can be mined as a filtration material. One such deposit is in the Purísima Hills near lompoc, california.
Brown algae (phaeophytes) are multicellular and mostly live in the oceans. they are the most famous big seaweeds. they can have complex structures that include:
● holdfasts, which look like roots but do not absorb and transport water; like the rhi-zoids of mosses, they anchor the organism to rocks
● stipes, which look like stems but do not transport water as do the true stems of plants; the tissues inside the stipes do, how-ever, have a limited ability to transport food molecules from one part of the organism to another
● blades, which look like leaves ● pneumatocysts, which are balloonlike struc-
tures that provide buoyancy to the organism ● reproductive structures (see below)
each part of the seaweed (except the holdfasts) is photosynthetic and largely on its own for nutrition.
the heterokont lineage also includes the oomy-cetes, or “water molds.” as their name suggests, they superficially resemble fungi: they form a myce-lium of strands and digest their way through organic food. they used to be classified as fungi, despite the major differences on the cellular level: Water molds have diploid nuclei, while fungi have haploid nuclei, in their mycelia; and water molds have cell walls made of cellulose, unlike the chitin cell walls of fungi. Water molds also produce flagellated spores, like other heterokonts. it eventually became clear that the oomycetes were heterokonts that had lost their chloroplasts and evolved into parasites. Water molds are one of the best examples of how appear-ances, without help from phylogenetic analysis, can be deceiving: they look and act like fungi but are more closely related to seaweeds. Some water molds have great economic and ecological importance, such as the following:
● Phytophthora infestans is the “mold” that often kills whole fields of potatoes. it spread through european and american potato fields in the 19th century and in 1845 erupted in ireland and destroyed nearly the entire crop. the irish lived almost exclusively on potatoes, and the collapse of the potato crop resulted in the Potato Famine, in which a million irish starved to death (despite the availability of food for those who could buy it) and another million emigrated.
● Phytophthora ramorum causes sudden oak death, which is killing thousands of oak trees in the coast Ranges of california.
xviii+598_Biodiversity_F.indd 446 11/17/11 1:13 PM

protists 447
● Pythium species kill seedlings of many kinds of plants, especially in greenhouses.
Some oomycetes (such as the two Phytophthora species above) are specialist parasites, while oth-ers such as Phytophthora megakarya and Pythium infect a range of hosts. P. megakarya is a parasite both of cocoa trees and of the trees in whose shade the cocoa bushes grow.
Heterokonts have motile, flagellated cells at some point in their life cycle. in some species, these cells are sperm, which unite with eggs to produce zygotes, while in others they are zoospores, which grow into haploid structures. Single-celled hetero-konts reproduce asexually most of the time, but occasionally they produce eggs or sperm, which unite to form a genetically different cell. Multi-cellular heterokonts (brown algae and oomycetes) produce antheridia (which produce sperm) and archegonia (which produce eggs). Most brown algae have a multicellular diploid phase (the sporo-phyte) that produces spores by meiosis. each spore is capable of growing into a multicellular haploid phase (the gametophyte) that may be indistinguish-able from the diploid phase. Male gametophytes release sperm, and female gametophytes produce eggs. Sporophytes of the genus Fucus directly pro-duce eggs and sperm, dispensing with the gameto-phyte phase altogether.
“Kelp forests,” such as the dense growth of Macrocystis off the coast of california, look like forests because the bulbs hold the stipes upright. the stipes can be more than 200 feet (up to 70 meters) in length. like a forest, the kelp create a complex habitat in which many other species live. like estu-aries and seagrass beds, kelp forests are places where young fishes can grow before heading out into the ocean. Kelp forests are threatened by population explosions of sea urchins, which graze upon young kelp. Sea urchin populations are often large because human activities have reduced the numbers of preda-tors (large fishes and sea otters) that normally con-sume them or their larvae. Kelp from these forests is commercially harvested by trawlers that cut off the tops of the organisms, which grow back. alginates from kelp are used in products such as soap and ice cream. the Sargasso Sea, below the Gulf Stream in the atlantic Ocean, is filled with brown algae of the genus Sargassum.
Haptophytes are single-celled algae related to the heterokonts and produce calcium carbonate coc-coliths. the white cliffs of Dover in england formed from the massive accumulation of coccoliths. Many evolutionary scientists classify the cryptomonads, which resemble single-celled brown algae, and the haptophytes into a larger group, the chromists.
alvEolaTE proTISTSAlveolates have cavities (similar to the alveoli of lungs) underneath the membrane that can form a pellicle or form armored plates. the three major lin-eages of alveolates are the following:
the ciliates have large numbers of cilia, which are like flagella, but shorter. they use cilia for move-ment, feeding, attachment, and sensation. they have two kinds of nuclei: micronuclei, which can replicate by means of accurate mitosis, and large macronuclei, composed of lots of copies of some of the genes. Macronuclei replicate less accurately and must be regenerated occasionally by micronuclei. Some ciliates, like Paramecium and Tetrahymena, are single complex cells that swim with their cilia. Others, like Stentor, attach themselves to a surface and grab food with their cilia. large ciliates such as Didinium may ingest smaller ciliates such as Paramecium.
the apicomplexans are protists that often have an apical complex, which is a degenerate complex chloroplast that no longer carries out photosynthe-sis. the apical complex produces enzymes that help parasitic apicomplexans to enter host cells. the most famous apicomplexans are the species of Plasmo-dium that cause malaria, one of the most widespread and devastating diseases in the world, by living inside of red blood cells.
the dinoflagellates are covered with hard cel-lulose plates and have two flagella. they often have complex chloroplasts (cells within cells within cells), just as do heterokonts and the other alve-olates. they swim in water and make food by photosynthesis. Some dinoflagellates do not have chloroplasts and live by consuming small food items. Dinoflagellate means “terrible flagellate,” in reference to the fact that they can be poisonous to vertebrates. Under certain conditions, such as warm weather and abundant nutrients, dinofla-gellates have population explosions, forming “red tides.” Mollusks can consume large numbers of dinoflagellates without difficulty, but if vertebrates eat these mollusks, the toxins may cause nerve damage. this is why fishermen who gather shellfish traditionally avoid summer months (the months without the letter R in their name). Some blooms of neurotoxic algae are caused by diatoms, not dinoflagellates. Some dinoflagellates are the zoo-xanthellae that live as mutualists inside the skin of aquatic animals. these animals, which include cor-als, get most of their food from the zooxanthellae that live in them.
CErCozoaN proTISTScercozoans are amoebae that produce long, fil-amentous pseudopods, with which they capture
xviii+598_Biodiversity_F.indd 447 11/17/11 1:13 PM

first entry448 protists 448
food items, and often have hard outer shells. they include the foraminiferans and the radiolarians. “Forams” have calcium carbonate (limestone) shells that can be very complex and occasionally large—some have been reported to be almost 7 inches (19 cm) in diameter. Because of their abun-dance, they are major contributors to the buildup of lime sediments at the bottom of the ocean, and their shells have been abundant fossils for millions of years. the carbon isotope ratios of the limestone in fossil foram shells are one of the major sources of information about climate changes in the history of the earth. although they are not photosynthetic, they form mutualistic associations with photosyn-thetic protists such as green algae, red algae, golden algae, diatoms, and dinoflagellates. Some of them ingest algae but keep the chloroplasts alive in their tissues. Radiolarians are similar to foraminiferans, but their shells are usually composed of silicate rather than lime.
GrEEN proTISTSthe major lineages of green protists are the red algae and the green algae. they are the only photo-synthetic eukaryotes to have simple chloroplasts—evolutionary descendants of cyanobacteria (cells within cells).
the red algae (Rhodophyta) are marine sea-weeds. Some of them have calcium carbonate reinforcements to their cell walls and contribute to the buildup of reefs. they have chlorophyll a and a unique chlorophyll d. their red color comes from phycobilins. they live at greater depths in the oceans than other algae; their red color allows them to absorb blue light, which is the color that penetrates most deeply into the water. Most edible seaweeds (such as Japanese nori) are red algae. Despite their superficial resemblance to brown seaweeds, the red algae evolved separately from primordial eukaryotic ancestors and are closely related to green algae.
the green algae (chlorophyta) are considered to be the ancestors of all land plants, even though they are not the most complex algae. Many are unicel-lular; some of them form sheets; and others form more complex structures; but none of them rival the gigantic kelp in size or complexity. However, they have important similarities to land plants on the cel-lular level:
● Green algae include many freshwater spe-cies. almost all brown and red algae, for example, are marine, as are many of the green algae. land plants evolved from fresh-water, not marine, ancestors.
● the cells of green algae are chemically and structurally similar to those of land plants. they have the same kinds of chlorophyll, store the same kinds of carbohydrates, and have the same kinds of cell walls.
the green algal origin of land plants, long known from the fossil record, has been confirmed by Dna studies in which green algae of the family charophyceae have been found to be more similar to land plants than are any other algae. the charo-phytes have some reproductive structures that are similar to the embryonic enclosures found in land plants. land plants, therefore, are a lineage within the green algae.
Freshwater algae often live in shallow water, and shallow water frequently dries up. algae that had adaptations that allowed them to continue to func-tion when water became mud would be favored by evolution. the first land plants, as seen in the fossil record, were small plants in wetlands, consistent with this explanation.
amoEBozoaN proTISTSMost of the amoebalike protists are in the amebo-zoan lineage. amoebalike cells move along surfaces when their cytoplasm flows into pseudopods. the pseudopods may also surround and engulf food items. amoebalike cells have evolved independently in the discicristate and cercozoan lineages, although the amoebozoans have lobe-shaped pseudopods, as opposed to the filamentous ones in the cercozoans. Plasmodial slime molds are amoebozoans. cells of Dictyostelium discoideum crawl in the soil and eat bacteria. When their food supply is scarce, they release cyclic aMP, which acts as a hormone that causes them to integrate into a multicellular sluglike creature that crawls along the surface of the ground. the head end of the slug attaches to the ground, and the rear end of the slug heaves up into the air (slowly) and lengthens into a stalk with a capsule on the end. the capsule produces sexual spores, which blow away to new locations. although plasmodial slime molds superficially resemble cellular (discicristate) slime molds, they evolved this mode of living independently.
opISThokoNT proTISTSOpisthokonts have flagellated cells with a single, basal flagellum. One of the opisthokont lineages is the fungi. another lineage has produced flagellated protists called choanoflagellates, and the animals.
the name choanoflagellate refers to the flagellum that emerges from a collarlike structure. the flagel-lum draws water into the collar, where small cellular protrusions capture bacteria and detritus. choano-
xviii+598_Biodiversity_F.indd 448 11/17/11 1:13 PM

protostomes 449
flagellates can swim, but can also adhere to surfaces, sometimes forming small clusters or colonies.
the structure of choanoflagellates closely resem-bles that of choanocytes, the cells that line the pas-sageways of sponges. choanocytes of sponges create water currents and capture food items. the close similarity of choanoflagellates and sponges was noticed, and choanoflagellates have been considered the closest single-celled relatives of animals since the 19th century. this relationship has been reinforced by Dna comparisons.
independently in most of these lineages, multi-cellularity has at least begun to evolve: Slime molds form temporary multicellular reproductive units in two of the lineages; heterokonts include the brown algae; the plant lineage includes red algae as well as green algae and land plants; and animals and fungi are opisthokonts that independently evolved multi-cellularity. there is no overarching law of progress toward complexity in evolution, but multicellular-ity allows adaptations that single cells cannot have and has been favored by natural selection over and over again. even single-celled organisms (including bacteria) benefit from forming colonies with some division of labor among their components.
See also cladistics; eukaryotes; symbiogenesis.
Further readingByrnes, J., et al. “Predator diversity strengthens trophic
cascades in kelp forests by modifying herbivore behav-ior.” Ecology Letters 9 (2006): 61–71.
Dayton, P. K. “ecology of kelp communities.” Annual Review of Ecology and Systematics 16 (1985): 215–245.
estes, James a., and D. O. Duggins. “Sea otters and kelp forests in alaska: Generality and variation in a com-
munity ecological paradigm.” Ecological Monographs 65 (1995): 75–100.
Foster, M. S., and D. R. Schiel. “the ecology of giant kelp forests in california: a community profile.” U.S. Fish and Wildlife Service Report 85 (1985): 1–152.
Graham, M. H. “effects of local deforestation on the diversity and structure of Southern california giant kelp forest food webs.” Ecosystems 7 (2004): 341–357.
———, et al. “Deep-water kelp refugia as potential hot-spots of tropical marine diversity and productivity.” Proceedings of the National Academy of Sciences USA 104 (2007): 16,576–16,580.
Halpern, B. S., et al. “Strong top-down control in South-ern california kelp forest ecosystems.” Science 312 (2006): 1,230–1,232.
King, n., et al. “the genome of the choanoflagellate Mono-siga brevicollis and the origin of metazoans.” Nature 451 (2008): 783–788.
Opoku, i. y., et al. “Shade trees are alternative hosts of the cocoa pathogen Phytophthora megakarya.” Crop Pro-tection 21 (2002): 629–634.
Patterson, D. J. “the diversity of eukaryotes.” American Naturalist 154 (1999, supplement): S96–S124.
Pearse, J. S., and a. H. Hines. “expansion of a central california kelp forest following the mass mortality of sea urchins.” Marine Biology 51 (1987): 83–91.
Sala, e., et al. “Fishing, trophic cascades, and the struc-ture of algal assemblages: evaluation of an old but untested paradigm.” Oikos 82 (1998): 425–439.
Steneck, Robert S., et al. “Kelp forest ecosystems: Biodiver-sity, stability, resilience, and future.” Environmental Conservation 29 (2002): 436–459.
protostomes See animals.
xviii+598_Biodiversity_F.indd 449 11/17/11 1:13 PM

450
quaternary period, biodiversity in the Quaternary period (starting 2.5 million years ago) is the third period of the cenozoic era. it follow the neogene period. the Quaternary period consists of the Pleistocene epoch, which ended about 12,000 years ago, and the current Holocene epoch.
Climate. the ice ages began in the Quaternary period. the motion of the earth relative to the Sun causes slight changes in sunlight intensity, which are magnifi ed by the earth’s climatic systems. lower sun-light intensity encourages the expansion of glaciers, while higher sunlight intensity encourages the glaciers to melt. these Milankovic cycles have been going on for much of earth’s history, but until the Quaternary period they did not cause ice ages. During times of gla-cial expansion, atmospheric car-bon dioxide was relatively scarce and sea levels were low; during times of glacial melt, atmospheric carbon levels increased and oceans rose (see table on pages 451 and 452). never before in earth’s his-tory had there been such rapid shifts in global climate. However, the greenhouse effect is currently causing more rapid warming of the earth than occurred even at the end of the most recent glaciation.
Continents. the conti-nents assumed their modern arrangement in the Quaternary. especially important was the for-mation of the isthmus of Panama. Ocean currents had previously fl owed between north and South america, allowing warm tropi-
cal water to reach the arctic Ocean and prevent extensive ice formation. When the isthmus weak-ened ocean currents, huge ice sheets could build up around the very cold arctic Ocean. Since the beginning of the Quaternary, the arctic Ocean has been mostly covered with an ice cap. During periods of glaciation, conditions in the tropical areas were drier, largely because so much water was frozen in the ice caps.
Marine life. life in the Quaternary oceans closely resembled that of modern times:
● Invertebrates. Ocean invertebrates were very similar to those of today.
Q
Glyptodonts were large armadillos, now extinct, of the early Quaternary period.
xviii+598_Biodiversity_F.indd 450 11/17/11 1:13 PM
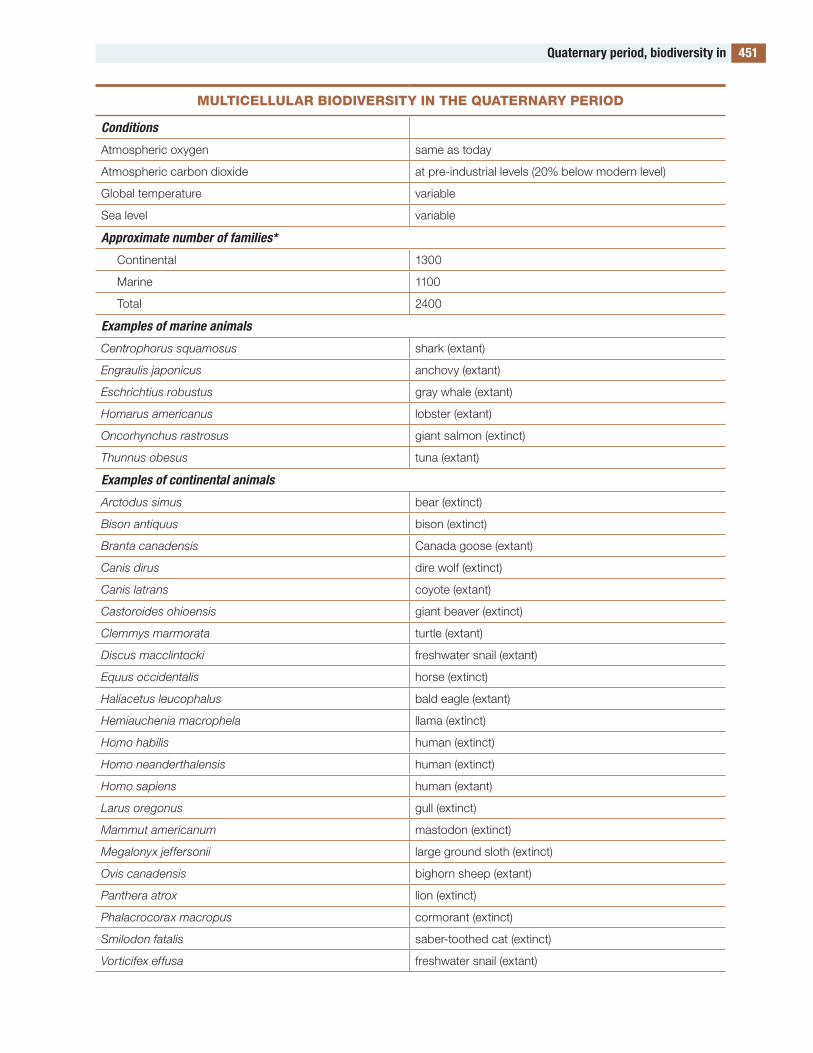
quaternary period, biodiversity in 451
multiCellular biodiversity in the quaternary period
Conditions
Atmospheric oxygen same as today
Atmospheric carbon dioxide at pre-industrial levels (20% below modern level)
Global temperature variable
Sea level variable
Approximate number of families*
Continental 1300
Marine 1100
Total 2400
Examples of marine animals
Centrophorus squamosus shark (extant)
Engraulis japonicus anchovy (extant)
Eschrichtius robustus gray whale (extant)
Homarus americanus lobster (extant)
Oncorhynchus rastrosus giant salmon (extinct)
Thunnus obesus tuna (extant)
Examples of continental animals
Arctodus simus bear (extinct)
Bison antiquus bison (extinct)
Branta canadensis Canada goose (extant)
Canis dirus dire wolf (extinct)
Canis latrans coyote (extant)
Castoroides ohioensis giant beaver (extinct)
Clemmys marmorata turtle (extant)
Discus macclintocki freshwater snail (extant)
Equus occidentalis horse (extinct)
Haliacetus leucophalus bald eagle (extant)
Hemiauchenia macrophela llama (extinct)
Homo habilis human (extinct)
Homo neanderthalensis human (extinct)
Homo sapiens human (extant)
Larus oregonus gull (extinct)
Mammut americanum mastodon (extinct)
Megalonyx jeffersonii large ground sloth (extinct)
Ovis canadensis bighorn sheep (extant)
Panthera atrox lion (extinct)
Phalacrocorax macropus cormorant (extinct)
Smilodon fatalis saber-toothed cat (extinct)
Vorticifex effusa freshwater snail (extant)
xviii+598_Biodiversity_F.indd 451 11/17/11 1:13 PM

first entry452 quaternary period, biodiversity in 452
● Vertebrates. Bony fishes and sharks con-tinued to diversify in the oceans, as did the toothed whales, baleen whales, porpoises, and dolphins.
Continental life. With each ice age cycle, plants and animals in the northern Hemisphere migrated south when glaciers advanced and north when gla-ciers retreated. the ice ages resulted in some extinc-tions, but speciation also occurred as populations were alternately separated and rejoined.
● Plant life. nearly all modern genera of plants were in existence, and probably most species. Grassland, boreal, and desert plants spread during glacial advances, while tropi-cal and deciduous forests spread during interglacial periods.
● Invertebrates. all major groups of terres-trial invertebrates had evolved before the Quaternary period.
● Vertebrates. Most modern genera of fresh-water fishes, amphibians, and reptiles were in existence. Many large mammals did not survive the Pleistocene extinction (see fig-ure on page 450). Modern horses of the genus Equus (which includes donkeys and zebras) became common during the Qua-ternary. Human ancestors evolved, first in dry forests then in the savannas of africa toward the end of the neogene period, and modern humans evolved during the Quaternary.
Extinction. the Pleistocene epoch ended with the Pleistocene extinction. it was not one of the mass extinctions in the history of life, but it is famous because many species of large terrestrial vertebrates became extinct at that time. the fact that this extinction event had an impact mostly on large land animals is the reason that it was not a mass extinc-tion: large land animals have always represented a minority of species on earth. the Pleistocene extinc-tion is the only extinction that selectively struck a certain group of organisms, which is one of several reasons that many scientists conclude that it was caused by human overhunting.
See also greenhouse effect; ice ages; Pleis-tocene extinction.
Further readingBarnosky, anthony D. “assessing the causes of late Pleis-
tocene extinctions on the continents.” Science 306 (2004): 70–75.
Benton, Michael J. “Diversification and extinction in the history of life.” Science 268 (1995): 52–58.
elias, Scott a. “Fossil insect evidence for late Pleistocene paleoenvironments of the lamb Spring site, colo-rado.” Geoarcheology 1 (2007): 381–387.
Harris, John M. “Bones from the tar pits.” Natural History, June 2007, 18–22.
Kenchington, ellen l., et al. “Pleistocene glaciation events shape genetic structure across the range of the ameri-can lobster, Homarus americanus.” Molecular ecol-ogy 18 (2009): 1,654–1,667.
Kürschner, Wolfram M., et al. “Oak leaves as biosensors of late neogene and early Pleistocene paleoatmospheric
Examples of land plants
Acer pleistocenicum maple (extinct)
Agropyron cristatum grass (extant)
Alnus glutinosa alder (extant)
Asimina triloba pawpaw (extant)
Dacrycarpus carpenterii podocarp conifer (extinct)
Deschampsia cespitosa grass (extant)
Fraxinus quadrangulata ash (extant)
Ginkgo biloba ginkgo (extant)
Picea abies spruce (extant)
Picea critchfieldi spruce (extinct)
Pinus monophylla piñon pine (extant)
Quercus palmeri oak (extant)
Yucca brevifolia Joshua tree (extant)
*Based on Benton.
xviii+598_Biodiversity_F.indd 452 11/17/11 1:13 PM

quaternary period, biodiversity in 453
cO2 concentrations. Marine Micropaleontology 27 (1996): 299–312.
liu, Jin-Xian, et al. “late Pleistocene divergence and subse-quent population expansion of two closely related fish species, Japanese anchovy (Engraulis japonicus) and
australian anchovy (Engraulis australis).” Molecular Phylogenetics and Evolution 40 (2006): 712–723.
Miller, Gifford H., et al. “ecosystem collapse in Pleistocene australia and a human role in megafaunal extinction.” Science 309 (2005): 287–290.
xviii+598_Biodiversity_F.indd 453 11/17/11 1:13 PM

454
reptiles Reptiles are primarily terrestrial ver-tebrates, with amniotic eggs but without feathers or hair. an amniotic egg has a shell and a series of fl uid-fi lled membranes that feed and protect the developing embryo. largely because of the amni-otic egg, most reptiles are very well adapted to life on dry land. there are about 8,000 species of rep-tiles (see table).
the category reptiles includes some but not all of the lineages that diverged from a common ancestor. as bird and mammal lineages evolved from reptile lineages, they would also be considered members of the reptile group. Many evolutionary biologists now distinguish between the synapsids and the reptiles. the synapsid group is today repre-sented only by mammals, while the reptile group is represented by birds and those animals commonly called reptiles. the reptiles, mammals, and birds (amniotes) represent a lineage within the tetrapodsor four-legged vertebrates. all other tetrapods, are referred to as amphibians. Of the major lineages of reptiles:
● the anapsids are turtles, fi rst known from the Jurassic period. “anapsid” refers to the lack of any openings in the dermal bone on the sides of the skull; the skulls have openings only for eyes and nostrils. turtles are either the sole survivors of the primitive anapsid form, or else they evolved from diap-sids when the skull openings fused shut.
● the synapsids included pelycosaurs, such as Dimetrodon of the Permian period and the therapsids of the triassic and Jurassic peri-ods. the mammals, fi rst known from the Jurassic period, evolved within this lineage. the name refers to the single opening of the dermal bones of their skulls. Mammals are the only surviving synapsids.
● the diapsids include all other reptiles. the birds evolved from within this lineage. the name refers to the two openings of the der-mal bones of their skulls.
By the Mesozoic era, the diapsids had diverged into the archosaurs and the lepidosaurs.
● the archosaurs diverged into the crocodiles and their relatives, fi rst known from the Jurassic period; the pterosaurs, fl ying reptiles of the Jurassic period and cretaceous period; and the dinosaurs. the only surviving archo-saurs are the crocodile lineage and birds, which are the surviving lineage of dinosaurs.
● the lepidosaurs diverged into the tuataras,of which only one lineage survives on islands off of new Zealand, and the squamates(lizards and snakes), fi rst known from the Jurassic period.
R
estimated number of reptile speCies
Group Number of Species
Turtles 325
Crocodilians 22
Squamates
Lizards 4,000
Snakes 3,000
xviii+598_Biodiversity_F.indd 454 11/17/11 1:13 PM

reptiles 455
phySIoloGICal adapTaTIoNS of rEpTIlESReptiles neither generate very much body heat nor regulate their body temperature. therefore they are both ectothermic and heterothermic. Many researchers believe dinosaurs were endothermic.
Most reptiles have three-chambered hearts. the two atria of the reptile heart allow a partial sep-aration of blood that circulates to the lungs and back (pulmonary circulation) from the blood that circulates through the rest of the body (systemic circulation). as a result, oxygenation of the blood is less efficient in most reptiles than it is in birds and mammals, which have four-chambered hearts. crocodilians have four-chambered hearts, but the structure of the heart still allows blood to cross between systemic and pulmonary circulation; their hearts evolved four chambers independently of those in birds and in mammals. Some lizards and snakes have heart muscles that can contract in such a way as to create the same blood circulation pattern that is found in mammals and birds, even though their hearts have only three chambers.
Reptiles have numerous features that allow com-plete adaptation to terrestrial life. their scaly skins resist water loss. Both juvenile and adult reptiles breathe by means of lungs. Most reptiles excrete most of their nitrogenous wastes in the form of uric acid, which is less toxic than ammonia (which amphibians excrete) and requires little water to elim-inate. turtles, which live in water, excrete urea, just as mammals do. these wastes, together with intes-tinal wastes and gametes, exit the body through a single opening, the cloaca.
Reptiles have teeth that are all more or less alike. Many reptiles replace their teeth throughout their lives. crocodilian teeth are hollow, with new teeth developing inside of old ones; as a result, when they lose a tooth (they can lose 3,000 in a lifetime), a new one is already in place. Only in the synap-sid lineage did teeth differentiate into the mamma-lian pattern of incisors, canines, and grinding teeth. therefore reptiles do not chew their food very well before swallowing it. crocodilians swallow stones (stomach stones or gastroliths), which help to grind the food in their muscular gizzard (the first chamber of the stomach).
the reproductive life cycle of reptiles also allows complete adaptation to terrestrial life. Most reptiles lay eggs on land. even marine turtles return to land to lay their eggs. in some snakes, the eggs hatch inside the mother’s body, allowing the live birth of young. Reptiles have internal fertilization. Males and females press their cloacae together, allowing the male’s sperm to travel to the female’s reproduc-tive tract without being exposed to dry air. in many reptiles such as turtles and crocodilians, the gender
of the offspring is determined by the temperature at which the eggs are incubated.
TurTlES aNd TorToISESthe turtles and tortoises (see figure) are classified as chelonians or testudines. the common names have no clear definition: in many cases, these animals are called turtles if they live in water and tortoises if they live on dry land. “terrapin” often refers to Malaclemys terrapin, which lives in brackish water; the name comes from a native american word.
the largest turtle is the leatherback sea turtle (Dermochelys coriacea), which can reach six and a half feet (2 m) in length and weigh a ton (900 kg). Sea turtles, particularly this species, have been the focus of worldwide conservation efforts not only because of their ecological importance in marine food webs, but also because of their dramatic egg-laying and hatching events on numerous beaches around the world. Giant tortoises were widespread on land until the arrival of modern humans, who presumably hunted them to extinction. this was nearly the fate of the giant tortoises of the Seychelles and Galápagos islands. turtles have shells (upper carapace and lower plastron) in which ribs are fused with dermal tissue. Most turtles have hard, thick shells, but members of the family trionychidae are the “soft-shell turtles” that defend themselves mostly by camouflage. turtle jaws, without teeth, have beaks that (in species such as the snapping turtle) can serve as fierce weapons. Many turtles and tor-toises are predators, although Galápagos tortoises eat vegetation (see photograph on page 456).
ancestral turtles could not withdraw their heads inside of their shells. Modern turtles can withdraw their heads in two different ways: the cryptodires (most turtles) pull their heads straight in, underneath
Painted turtles (Chrysemys picta) are cold blooded but warm up by lying in the sun. (Mary Moore Ferracci)
xviii+598_Biodiversity_F.indd 455 11/17/11 1:13 PM

first entry456 reptiles 456
the spine, while the pleurodires (the side-neck tur-tles) bend their necks sideways.
CroCodIlES aNd ThEIr rElaTIvES“crocodilians” include alligators, crocodiles, cai-mans, and gharials. the largest crocodilian (and the largest reptile) is the saltwater crocodile (Crocodylus porosus), which can reach 3,300 pounds (1,500 kg) in weight and 17 feet (5 m) in length. Prehistoric crocodiles were often much larger. though their sprawling gait suggests that of primitive amphibians, crocodiles can walk with their legs erect underneath their bodies, as some prehistoric crocodiles did. crocodiles and their relatives have webbed feet and a vertically flattened tail that aids them in swimming through their aquatic habitats. the secondary palate that isolates the breathing passages from the mouth allows them to immerse in water except for eyes and nostrils as they wait for their prey.
alligators and caimans have wider and shorter heads, with a snout that is more U-shaped than V-shaped, as in crocodiles. crocodiles are most easily recognizable because the fourth tooth of the lower jaw is visible outside of the mouth when the jaw is closed. crocodiles are usually lighter in color and more aggressive than alligators. there are only two species of alligators: one in the southeastern United States (Alligator mississippiensis) and one in the yangtze River of china. there are only two species of gharials, with elongated snouts, native to india and Malaysia.
Because of their role as top predators in many freshwater habitats, protection of crocodilians has been a focus of many conservationists. Researchers were surprised to discover that crocodilian blood has strong antiviral properties. this is just one more example of the unexpected human benefits that may result from the preservation of biodiversity.
lIzardS aNd SNakES (SquamaTES)Snakes are legless reptiles that lack eyelids and exter-nal ears, and have broad plates on the ventral (lower) surface of the body, the movement of which propels them forward. Snakes have no necks, very long bod-ies with 200 to 400 vertebrae, and very short tails. Snakes have adaptations that allow their bodies to be as narrow as possible while still having a lot of muscle. For example, many snakes have only a right lung. Some snakes have primitive properties: Pythons and boas have rudimentary hip bones, indicating that their ancestors had legs, and their left lungs are almost as large as their right lungs. Many snakes are venom-ous. Venom is modified saliva, delivered by fangs that can be grooved or hollow. More than 700 species of snakes are venomous, 250 of which can kill a human with a single bite. Some snakes squeeze and suffocate (but do not crush) their victims. Snakes smell with their tongues, which, being forked, can distinguish the direction from which a particular scent comes. Because they lack external ears, snakes cannot hear very well, but they are very sensitive to vibrations.
Because they cannot manipulate large food items, snakes must swallow their prey whole. their very flexible jaws allow them to eat animals larger than themselves (see figure). the lack of legs allows snakes to move through narrow crevices that few other animals could penetrate.
examples of snakes include:
● Boas (family Boidae) and pythons (family Pythonidae) asphyxiate their victims.
● elapids (family elapidae) include many of the most venomous snakes. Naja naja, the
Giant tortoises live on the Galápagos Islands off the coast of Ecuador. Most of them now live in protected areas. (Stanley A. Rice)
Snakes can eat very large prey: This Afghan snake is eating a lizard that is almost as large as itself. (Wikimedia)
xviii+598_Biodiversity_F.indd 456 11/17/11 1:13 PM

reptiles 457
indian cobra, raises itself from the ground and spreads out its upper body in a threat display. Dendroaspis polylepis, the black mamba of africa, has venom that shuts down both nerve and heart activity in its victims. the spitting cobra (Hemachatus haemachatus) can spit venom over 6 feet (2 m) into the eyes of prey or attackers. north american coral snakes are poisonous, though not aggressive. nonpoisonous king snakes have red, black, and yellow bands, apparently in mimicry of coral snakes. they can be distinguished by the red and yellow bands adjoining one another in coral snakes (“Red and yellow, kill a fellow”).
● Vipers (family Viperidae), with their dis-tinctive triangular heads, can inject venom through hollow, retractable fangs. they can open their mouths nearly 180° when bit-ing. One group of vipers, the pit vipers, has infrared-sensitive pits behind their eyes that allow them to sense the presence of a warm-blooded victim in the dark. Vipers in the United States include the copperhead Agkis-trodon contortrix, the cottonmouth Agkis-trodon piscivorus, and rattlesnakes such as the sidewinder Crotalus cerastes. Shed scales remain attached to the tail, which rattle-snakes vibrate to produce a warning rattle.
● Scolecophid snakes are small snakes that burrow in the ground. One of them, Lep-totyphlops carlae from Barbados, is only 4 inches (10 cm) long.
Other squamates include the following:
● Chameleons (family chameleonidae) have several unique features. their eyes can move independently of one another, allowing a range of vision unmatched in most ani-mals. in Madagascar, where chameleons are diverse and numerous, the native Malagasy people say that chameleons keep one eye on the past and one on the future. they also have very long tongues, which may be as long as the body. the tongue has a bone that moves the tongue quickly and a suction cup that grabs the prey (in 30-thousandths of a second). chameleons are perhaps most famous for being able to change the color of their skin to camouflage them against their backgrounds and, perhaps more impor-tantly, as a social signal. One layer of skin contains cells with yellow or red pigment granules; if the granules are concentrated in the center of a cell, it is nearly transparent,
but if the granules are spread throughout the cell, it has intense color. another layer of skin reflects blue light. Green skin results from blue light reflected back through cells with dispersed yellow pigment granules.
● Komodo dragons (Varanus komodoensis), up to 10 feet (3 m) long and weighing 150 pounds (70 kg), are the largest lizards in the world. they live on a few indonesian islands. there are no large native mammals on the islands they inhabit, and as a result they have evolved to a large size. they are poisonous but are too slow to frequently hunt; they mostly eat carrion.
● Gila monsters (Heloderma suspectum) are the only venomous lizards in the United States. they are very sluggish and produce little venom. they use the venom mostly for defense and must chew in order to get the venom into the victim’s tissues.
● Geckos are numerous species of small, often brightly colored lizards that are unique in being able to make sounds and in their abil-ity to walk on vertical surfaces. their foot pads have numerous tiny hairlike structures that adhere to surfaces not with chemicals but by displacing electrons on their surfaces, creating an electrical attraction. technolo-gists are studying gecko feet to produce non-chemical adhesive structures, making geckos one of the best examples of biomimicry.
● Skinks are small lizards with long tails, short necks, and short legs.
Snakes are not as distinct from lizards as might first appear. Some lizards lack movable eyelids. Per-haps most importantly, many separate lineages of lizards are fully or partly legless (a major example of evolutionary convergence), as in the following examples:
● Glass snakes (genus Ophisaurus, Greek for “snake-lizard”) are legless lizards that, unlike snakes, have movable eyelids, exter-nal ear openings, and the ability to drop parts of their tails (like broken glass) to distract predators. the dropped portion of the tail continues to move as a decoy to the predator, while the glass snakes make their bodies immobile.
● Grass lizards (genus Chamaesaura) have rudimentary hind limbs and no forelimbs. they “swim” through the grass with snake-like movements.
● Fogs (family Pygopodidae) have rudimen-tary hind limbs and no forelimbs, and are
xviii+598_Biodiversity_F.indd 457 11/17/11 1:13 PM

first entry458 reptiles 458
very elongated in shape, like snakes. they are close relatives of geckos.
● Dibamid lizards are not only legless but, liv-ing in burrows, are nearly blind.
● Genus Anniella contains two species of american legless lizards.
● Several species of spectacled lizards (family Gymnophthalmidae) have reduced or absent hind limbs; they pull themselves forward with their forelimbs.
● Genus Typhlosaurus is a skink without legs; other genera of skinks have reduced legs.
● Some species of plated lizards (family Ger-rhosauridae) have only rudimentary hind limbs.
● Amphisbaeneans are another lineage of leg-less lizards. they are sometimes considered to be a lineage distinct from the lizards and snakes.
TuaTaraSthe two species of tuataras (genus Sphenodon) look like lizards, but they are the only surviving mem-bers of a separate lineage that may closely resemble primitive diapsids. tuataras were more common and diverse than lizards 200 million years ago. today they live only in new Zealand. For example, the vertebrae are hourglass-shaped, as in fishes and a few amphibians, and tuataras are the only amniotes to have this type of vertebrae. their abdominal ribs also have a primitive structure that is unique among amniotes. tuataras also have a third eye on the top of the head: the parietal eye has its own lens, cornea, and retina. its optic nerve connections have degener-ated, and it is more closely associated with the pineal gland, which regulates the biological clocks of verte-brates. the third eye is visible only in newly hatched tuataras, and is soon covered by skin and scales.
See also biodiversity, resources for medi-cine; biomimicry; convergence; dinosaurs; Galápagos islands.
Further readingadams, clark e., and John K. thomas. Texas Rattlesnake
Roundups. college Station, tex.: texas a and M Press, 2008.
apesteguía, Sebastián, and Hussam Zaher. “a cretaceous terrestrial snake with robust hindlimbs and a sacrum.” Nature 440 (2006): 1,037–1,040.
Barrow, Mark. “Dragons in distress: naturalists as bioac-tivists in the campaign to save the american alligator.” Journal of the History of Biology 42 (2009): 267–288.
Browne, constance l., and Stephen J. Hecnar. “Species loss and shifting population structure of freshwater turtles despite habitat protection.” Biological Conservation 138 (2007): 421–429.
cree, a., et al. “Reproduction of a rare new Zealand rep-tile, the tuatara Sphenodon punctatus, on rat-free and rat-inhabited islands.” Conservation Biology 9 (1995): 373–383.
Dubois, y., et al. “thermoregulation and habitat selec-tion in wood turtles Glyptemys insculpta: chasing the sun slowly.” Journal of Animal Ecology 78 (2009): 1,023–1,032.
ernst, c. H., and R. W. Barbour. Turtles of the World. Washington, D.c.: Smithsonian institution Press, 1989.
Fry, Brian, et al. “early evolution of the venom system in lizards and snakes.” Nature 439 (2006): 584–588.
Girard, charlotte, et al. “Post-nesting migrations of log-gerhead sea turtles in the Gulf of Mexico: Dispersal in highly dynamic conditions.” Marine Biology 156 (2009): 1,827–1,839.
Glaucas, X., et al. “timing and reproduction of a cold desert viperid snake from north america, the Great Basin rattlesnake (Crotalus lutosus).” Journal of Arid Environments 73 (2009): 719–725.
Greene, Harry W. Snakes: The Evolution of Mystery in Nature. Berkeley: University of california Press, 1997.
Henderson, Donald M. “effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: a computational analysis.” Canadian Journal of Zool-ogy 81 (2003): 1,346–1,357.
Kelly, Shannon M., et al. “accumulation and maternal transfer of polychlorinated biphenyls in snapping tur-tles of the upper Hudson River, new york, U.S.a.” Environmental Toxicology and Chemistry 27 (2008): 2,565–2,574.
Kenow, Kevin P., et al. “Predation of radio-marked mallard (Anas platyrhynchos) ducklings by eastern snapping turtles (Chelydra serpentina serpentina) and western fox snakes (Pantherophis vulpinus) on the upper Mis-sissippi River.” Journal of Herpetology 43 (2009): 154–158.
Maladen, Ryan D., et al. “Undulatory swimming in sand: Subsurface locomotion of the sandfish lizard.” Science 325 (2009): 314–318.
Mazzotti, Frank J., et al. “alligators and crocodiles as indi-cators for restoration of everglades ecosystems.” Eco-logical Indicators 9 (2009, supplement): S137–S149.
Merchant, Mark e., et al. “antiviral activity of serum from the american alligator (Alligator mississippiensis).” Antiviral Research 66 (2005): 35–38.
Motani, Ryosuke. “the evolution of marine reptiles.” Evo-lution Education and Outreach 2 (2009): 224–235.
Mullin, Stephen J., and Richard a. Seigel. Snakes: Ecology and Conservation. ithaca, n.y.: cornell University Press, 2009.
navas, carlos a., et al. “Morphological and physiologi-cal specialization for digging in amphisbaenians, an ancient lineage of fossorial vertebrates.” Journal of Experimental Biology 207 (2004): 2,433–2,441.
xviii+598_Biodiversity_F.indd 458 11/17/11 1:13 PM

resistance, evolution of 459
Pianka, eric R., and l. J. Vitt. Lizards: Windows to the Evolution of Diversity. Berkeley: University of califor-nia Press, 2003.
Pyron, R. alexander, and Frank t. Burbrink. “lineage diversification in a widespread species: Roles for niche divergence and conservatism in the common king-snake, Lampropeltis getula.” Molecular Ecology 18 (2009): 3,443–3,457.
Ramstad, Kristina M., et al. “Species and cultural conser-vation in new Zealand: Maori traditional ecologi-cal knowledge of tuatara.” Conservation Biology 21 (2007): 455–464.
Santidrián tomillo, Pilar, et al. “influence of emergence suc-cess on the annual reproductive output of leatherback turtles.” Marine Biology 156 (2009): 2,021–2,031.
Shine, Richard. “life history evolution in reptiles.” Annual Review of Ecology, Evolution, and Systematics 36 (2005): 23–46.
———, et al. “Windows of embryonic sexual lability in two lizard species with environmental sex determina-tion.” Ecology 88 (2007): 1,781–1,788.
Stuart-Fox, D., and a. Moussalli. “Selection for social signaling drives the evolution of chameleon color change.” Public Library of Science Biology 6 (2008): e25.
Vincent, S. e., et al. “convergence in trophic morphology and feeding performance among piscivorous natricine snakes.” Journal of Evolutionary Biology 22 (2009): 1,203–1,211.
Walker, Deette, and John c. avise. “Principles of phy-logeography as illustrated by freshwater and terres-trial turtles in the southeastern United States.” Annual Review of Ecology and Systematics 29 (1998): 23–58.
Wang, Xianyan, et al. “Response specificity to advertise-ment vocalization in the chinese alligator (Alligator sinensis). Ethology 115 (2009): 832–839.
Warner, Daniel a., and Richard Shine. “the adaptive sig-nificance of temperature-dependent sex determination in a reptile.” Nature 451 (2008): 566–568.
White, Mel. “When crocs ruled.” National Geographic, november 2009, 128–139.
resistance, evolution of Microbes, insects, and weeds have evolved resistance to the chemical methods used to control them. the most impor-tant modern example of natural selection—both in terms of the clear and easily studied process, and of importance to human health and the economy—is the evolution of antimicrobial, pesticide, and herbi-cide resistance. as biologist Rachel carson wrote in 1962, “if Darwin were alive today the insect world would delight and astound him with its impressive verification of his theories of the survival of the fit-test. Under the stress of intensive chemical spraying the weaker members of the insect populations are
being weeded out. now, in many areas and among many species only the strong and fit remain to defy our efforts to control them.” this entry will focus on the evolution of resistance to herbicides in popu-lations of weeds and of resistance to pesticides in populations of animal pests.
Within populations of pest animals and weeds, some individual organisms may be resistant to the chemicals that kill most of the other organisms. examples include the following:
● Within a population of insects, certain indi-viduals may have a genetically based abil-ity to resist pesticides that will kill most members of their species. this resistance may result from an increased production of defensive enzymes, the production of an improved defensive enzyme, or a behavioral tendency to detect and avoid the pesticide. For example, some cockroach populations have evolved an aversion to sugar—not a good thing for them, under normal condi-tions, but it does cause them to avoid sugar-baited pesticide traps.
● Within a population of weeds, certain indi-viduals may have a genetically based abil-ity to resist herbicides that would kill most members of their species. this resistance may result from a mutation that alters the membranes of the chloroplast, the structure within plant cells that carries out photosyn-thetic food production.
Under normal conditions (in the absence of pes-ticides and herbicides), the mutant (resistant) indi-viduals have inferior reproduction:
● insects that expend much of their metabolic reserves producing resistance enzymes have less energy left over for reproduction than normal insects.
● Rats that are resistant to the rat poison war-farin suffer a high incidence of bleeding and require higher levels of vitamin K in their diets; therefore, under normal conditions, they bleed to death and die of vitamin defi-ciency more frequently than normal rats.
● Weeds with mutated chloroplasts have infe-rior photosynthetic food production com-pared to normal weeds.
Under these conditions, natural selection oper-ates against the resistant individuals. But when pes-ticides or herbicides are common and persistent in the environment of the populations, natural selec-tion operates in favor of these very same resistant
xviii+598_Biodiversity_F.indd 459 11/17/11 1:13 PM

first entry460 restoration ecology 460
individuals. the heavy and persistent use of chemi-cals, therefore, creates the perfect environment for the evolution of animals and weeds that resist the very chemicals that humans use to control their populations.
individual animals and weeds generally cannot change their resistance. they either are resistant or are not. the individuals survive and reproduce or they do not survive; therefore only the populations and species can evolve resistance.
the extensive and heavy use of pesticides has selected pesticide-resistant insects and rats; the extensive and heavy use of herbicides has selected herbicide-resistant weeds. the first DDt-resistant mosquitoes in Pakistan were detected in 1965, just five years after DDt use began in the region. the swift evolution of resistance occurs with each new pesticide. in the last decade, more than 520 spe-cies of insects and mites, 273 weed species, 150 plant diseases, and 10 rodent species have developed genetic resistance to at least one pesticide or herbi-cide. Many species are now becoming resistant to all major classes of insecticides. an insect that is resistant to one pesticide is often resistant to other related, chemically similar pesticides. in addition to promoting the evolution of resistant strains of ani-mals and weeds, the overuse of chemicals in insect control and agriculture has resulted in the buildup of toxicity in the environment and in the food chain.
the evolution of pesticide resistance in insects and rat populations has implications both for public health and for agriculture. Pesticide sprays are heav-ily used to control many species of insects, such as the mosquitoes that spread diseases like malaria and yellow fever. insect pests (such as weevils and cater-pillars) cause millions of dollars of agricultural yield loss. When the sprays become less effective (because many, though not all, of the insect populations evolve resistance), public health officials may respond by increasing the application of sprays, which has two effects: greater environmental contamination and an even greater evolution of resistance.
the evolution of herbicide resistance in weeds also portends greater agricultural yield loss (due to competition between weeds and crops) and greater environmental contamination from stepped-up her-bicide applications. in recent years, crops (such as maize, soybeans, and potatoes) that have been genet-ically engineered to resist herbicides have come on the market, and large acreages are now devoted to them. Farmers can now spray massive amounts of herbicides on their fields, to kill the weeds, without fear of killing the crops. this encourages the heavy use of herbicides—which is precisely what the com-panies that sell the seeds of these crops want, since the herbicides are manufactured by the same compa-
nies that sell the seeds. Unfortunately, this will result in even greater environmental contamination.
there is some concern that the genetic basis of herbicide resistance, which has been put into crops by genetic engineering, could spread to wild weed species by crossbreeding. While this has not hap-pened extensively, it has occurred: Resistance to the herbicide glyphosate has been transferred from crops to weeds within the crucifer (mustard) family by cross-pollination. Once the resistance genes are in the wild weed populations, natural selection will favor their spread whenever herbicides are used. to minimize this process, researchers have also bred crops that resist the herbicide Dicamba. Because the mechanism of resistance occurs in the chloroplasts, and pollen contains no chloroplasts, it is unlikely that this form of herbicide resistance will spread through cross-pollination.
an evolutionary understanding of public health and agriculture demands that pesticides and herbi-cides be used sparingly to minimize environmental contamination and to prevent the evolution of resis-tance in the very species of organisms that humans are trying to control. if the use of chemical control agents is restrained, they will remain effective when they are really needed in an emergency. the overuse of chemical control agents is a direct result of over-looking the laws of ecology and evolution.
See also natural selection.
Further readingBehrens, Mark R., et al. “Dicamba resistance: enlarging
and preserving biotechnology-based weed manage-ment strategies.” Science 316 (2007): 1,185–1,188. Summarized by Service, Robert F. “a growing threat down on the farm.” Science 316 (2007): 1,114–1,117.
carson, Rachel. Silent Spring. new york: Houghton Mif-flin, 1962.
Smith, P., et al. “a new aspect of warfarin resistance in wild rats: Benefits in the absence of poison.” Functional Ecology 7 (1993): 190–194.
tardif, F. J., et al. “a mutation in the herbicide target site acetohydroxyacid synthase produces morphologi-cal and structural alterations and reduces fitness in Amaranthus powellii.” New Phytologist 169 (2006): 251–264.
WeedScience.org. “international Survey of Herbicide Resis-tant Weeds.” available online. URl: http://www.weed science.org/in.asp. accessed august 8, 2011.
restoration ecology Restoration ecology is the application of ecological principles to restore a damaged ecosystem to something close to its original integrity. Scientific principles of ecology allow current problems to be identified and allow
xviii+598_Biodiversity_F.indd 460 11/17/11 1:13 PM

restoration ecology 461
researchers to determine what the goal should be—the characteristics that the restored ecosystem should have.
One major problem in restoration ecology is that it is often impossible to determine what the characteristics of the “original” ecosystem were. throughout most of the world, prehistoric peoples have in fact altered natural ecosystems on large and small scales. the most noticeable large-scale alteration has been the maintenance of grasslands. native americans started fires that burned large areas of central north america, encouraging the spread of grasslands (and the game animals that lived in them) at the expense of forests. although the tallgrass prairie was not an artificial ecosystem, its geographic extent was artificially enhanced. On a smaller scale, native americans built up artificial embankments of rich soil on which they raised crops and lived. the soils of the amazon Basin are (except along rivers and near the volcanic andes) notori-ously poor in nutrients and will not hold nutrients for very long after fertilizer (such as the ashes of a burned forest) is applied. amazonian tribes built compost piles that included pottery shards, which persist today as pockets of rich soil known as terra preta de Indio. the Mississippian culture covered much of eastern north america about 700 years ago, and the tribes (united by extensive trade net-works) built artificial mounds not only for their buildings (thus earning the name Mound Builders), but also for agriculture. When botanist William Bartram traveled through Florida and what is now Mississippi and alabama in the late 18th century, he saw many of these mounds, regarding which the natives that he met knew nothing. there is evidence that australian aborigines used fire to transform the arid regions of that continent into grassland. Few if any restoration ecologists would claim that ecosystems should be restored to the conditions that existed prior to the fire regimes and other transfor-mations of the original inhabitants. the standard map of potential natural vegetation in the cotermi-nous United States, drawn up by a. W. Küchler in 1964, depicts plant communities prior to european, not native, habitation.
in some cases, aboriginal inhabitants pro-moted extensive ecological degradation. Outstand-ing examples are Pleistocene and recent extinctions and land degradation. there is evidence that the Hohokam culture of the american southwest, the Mississippian culture of eastern north america, and the Mayan culture of central america caused deforestation and soil erosion extensive enough to contribute to their own collapse, although the immediate precipitating events were droughts sev-eral centuries ago. in semiarid regions, such as the
american Southwest, ecological degradation caused by aboriginal inhabitants may have contributed to a nearly permanent change in vegetation. the land around chaco canyon, new Mexico, is now desert, but when it was inhabited by people who lived in large apartment complexes (see photograph), piñon pine forest was extensive, and there was enough water to raise corn for hundreds of people. it is pos-sible that soil degradation, caused by those inhabit-ants, contributed to the failure of the pine forest to regenerate. Restoration ecologists, therefore, would aim to restore ecosystems to the condition in which native peoples originally lived before they caused ecological degradation.
Some situations that observers of nature have assumed to be “natural” may have been the product of indirect human alteration of habitats. Perhaps the best example of this is the huge populations of bison (Bison bison) and passenger pigeons (Ectopistes migratorius) just prior to european and american conquest of north america. every child is taught that prairies covered with millions of bison and the air blackened with millions of passenger pigeons that would break limbs from trees in which they roosted, was “natural” and brought to an end by deliber-ate overhunting. the second half of this statement is undeniably true. But how natural were the huge populations? Scholars have suggested that large pop-ulations of native americans kept both bison and pigeon populations under control by hunting. then european diseases such as smallpox, introduced (in most cases) accidentally, reduced native american populations by up to 90 percent. in some cases, entire agriculture-based civilizations (such as those of southeastern north america) were destroyed and replaced by scattered villages. With only 10
There was a large city in Chaco Canyon of New Mex-ico about 1000 c.e. This civilization collapsed during a prolonged drought that was made worse by defores-tation. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 461 11/17/11 1:13 PM

first entry462 restoration ecology 462
percent as many hunters as before, game populations exploded. this may also be the reason that William Bartram saw hundreds of alligators in nearly every pool and stretch of river that he explored in the late 18th century. Most of the native fishermen had died; fish populations exploded and so did the popula-tions of alligators that ate them. it would therefore be unrealistic for a restoration ecologist to attempt to bring back huge populations of bison, pigeons, or alligators.
an even more important problem is that it may be impossible to restore the “original” ecosystem characteristics even if they were known. Reasons include the following:
● Some of the original species have become extinct. this would apply not only to all of the species lost in the Pleistocene extinction, but those driven to extinction by hunting much more recently, such as the elephant birds (family aepyornithidae) extirpated by the original inhabitants of Madagascar, the moas (bird family Dinornithidae) by the Maori of new Zealand, and the dodos (Raphus cucullatus) killed by europeans on the island of Mauritius. extinctions of some species can cause a cascade of other extinctions, although this has not been doc-umented to occur. it appears that dodos were important dispersers of the seeds of the tambalacoque tree (Sideroxylon grandiflo-rum), and that the extinction of the dodos has reduced the reproduction of this tree. in most cases, however, other species move into the role vacated by the extinct species and at least partly compensate for the disap-pearance. Dodos were not the only dispers-ers of tambalacoque seeds (although most of the other animals that ate the seeds are also extinct). When a fungal disease killed almost all adult american chestnut trees in the eastern deciduous forest of north amer-ica, various species of oaks primarily took their place.
● Invasive species have been introduced. it is nearly impossible to eradicate invasive species. Restoration ecologists have consid-ered themselves successful when they have reduced invasive species populations to the extent that they do not significantly depress the populations of the native species.
● Climatic conditions have changed and will continue to change. ecological degradation is not the only reason that piñon forests will not grow in chaco canyon today; a drier climate also contributes. as the greenhouse
effect intensifies, it may be impossible to restore vegetation cover that depends on the climatic conditions that prevailed even a few decades ago. if global warming con-tinues at its current rate, sugar maples will grow only in canada. a certain amount of global warming is inevitable. therefore, some of the southern part of the sugar maple’s range will inevitably be lost. tun-dra conditions may not exist, and tundra restoration may be impossible, in the next century. Sometimes these factors interact with one another. introduced tree species such as eucalyptus often use a lot of water from underground sources and cause the water table to drop, thus creating an effec-tively drier climate.
● Agriculture covers a large area of the Earth. almost a third of the earth’s dry land sur-face has been transformed into farmland or managed pasture. intensification of agri-cultural practices has barely kept pace with increases in human population. it is nearly inconceivable that the area of land now used for agriculture will ever be reduced or that those areas will be restored to the condition they were in prior to modern agriculture.
● Human habitation has created a demand for fire suppression. Fires are an essential component of nearly every natural ecosys-tem, even though they may play a minor role in those with high rainfall. Fire suppres-sion has allowed ponderosa pine to spread into land formerly occupied by shortgrass prairie in the Black Hills of South Dakota. trees that live in wetter conditions, such as shumard oaks (Quercus shumardi), grow underneath and replace post oaks (Quer-cus stellata) in the cross timbers forest that have not been burned, and these trees have actually created wetter conditions, a process known as mesophication. large-scale fires for habitat restoration would be vigorously opposed by local residents in all of these places.
● The scale of restoration is simply too large. the aquatic habitats of chesapeake Bay have been severely damaged by human activities, including the nutrients and sediments in the watershed runoff. the previously abundant oysters find it difficult to regenerate as sedi-ment builds up. Fertilizer in the runoff from cities and farms has caused eutrophication. the result has been large “dead zones” of anoxic water at the bottom of the bay. there have been some recent successes at ecologi-
xviii+598_Biodiversity_F.indd 462 11/17/11 1:13 PM

restoration ecology 463
cal restoration in chesapeake Bay. ecologist David Schulte led a project to create artifi-cial reefs on one of the tributaries to the bay. Within five years, this project had allowed the regeneration of 185 million native oys-ters (Crassostrea virginica). But saving the entire bay may prove impossible. although chesapeake Bay is very large, it is dwarfed by the size of the heavily populated water-shed around it.
● The causes of environmental degrada-tion are ongoing. Perhaps the best example of this problem is coral bleaching. Some bleached coral reefs have been restored by creating new surfaces upon which larval corals can settle or by simply having local divers break off pieces of healthy coral in one location and wedge them into bleached coral reefs, where they act as sources of regeneration. the sometimes spectacular (though spatially limited) regrowth of coral on reefs will be nullified by the same condi-tions that produced bleaching in the first place, which have been intensified by global warming.
One example of a natural habitat that has been changed beyond the possibility of restoration (on any timescale meaningful to modern civilization) is the San Joaquin Valley of california. Much of this valley used to be covered by a large shallow lake, tulare lake, because snowmelt from the Sierra nevada came down the Kern, tule, and Kaweah Rivers and was blocked by the coast Ranges. Dur-ing the 1930s, a large system of irrigation canals was built by the federal Bureau of Reclamation to use the snowmelt for irrigation. the San Joaquin Valley is now almost entirely agricultural. the cli-mate is much less humid, and the valley is saturated with pesticides. nearly every plant species in the valley is introduced, either by agriculture (e.g., cit-rus and olive orchards) or as weeds, except some of the spring wildflowers. the dominant grasses are annuals introduced from europe and asia (such as Avena and Bromus); the native perennial grasses are rare. eucalyptus trees, introduced from aus-tralia, can tolerate the dry conditions, and when they establish a forest, their toxic leaves inhibit the growth of other plants. there has been remarkably little extinction, though many animal species persist only in nearby foothills.
Restoration ecologists generally focus on restoring ecosystem function rather than return-ing ecosystems to a pristine, precivilized condition. indicators of damaged ecosystems are conditions such as excessive runoff of water, soil, and nutri-
ents or the proliferation of pests and introduced species. a “managed ecosystem,” which functions as agricultural land or as a source of wood, will have more, but perhaps not much more, runoff than a natural ecosystem. in many places, such as china, intact forests are cut down and replaced by tree plantations of single species, such as euca-lyptus (the low-quality wood of which is used for paper production), rubber, or oil palm. the profit, much of which goes to poor local people, is nearly irresistible. But research has shown that a mixture of native tree species can yield almost as much profit and have much less soil erosion and runoff. in a mixture of tree species, the faster-growing trees can be selectively harvested, leaving behind the slower-growing trees for later harvest; the trees left behind hold down the soil. a mixed-tree plan-tation produces better humus and water retention than a single-species plantation. a rubber planta-tion in Southeast asia has three times as much run-off and 45 times as much soil erosion as an intact forest. a managed forest can easily have less runoff and erosion than a rubber plantation. a replanted forest of large native trees, with a shade crop of tea underneath them, is not a natural ecosystem but functions like one and can be profitable to local people.
in a few cases, habitat restoration is as easy as discontinuing the disturbance that damaged it in the first place. Many species return on their own without needing to be seeded or stocked. Some-times they can do so with a little technological help. coral reef regeneration is enhanced by expos-ing the damaged reef to a chemical that induces coral larvae to settle on the exposed surfaces. Usu-ally, however, substantial work is needed to create conditions for the success of the returning species. Deforested land is often too hot and dry for native trees to grow back. Shrubs may need to be planted as “nurse plants” under whose shade the native trees can establish themselves. Sometimes these nurse plants are the invasive species themselves, which do not need to be planted, and many of which will die in the shade of the replanted trees once they have grown. Frequently, invasive plants must be laboriously and repeatedly removed, at least until the restored species become established. in addition, it is necessary to ensure that the popu-lations involved in habitat recovery have sufficient genetic variation. care must be taken to ensure that pollinators and other mutualists return to the restored ecosystem.
One of the most ambitious restoration projects is taking place in the aral Sea in the former Soviet Union. the aral Sea was once the fourth largest inland body of water. Under Soviet control, the water
xviii+598_Biodiversity_F.indd 463 11/17/11 1:13 PM

first entry464 restoration ecology 464
from rivers that kept the aral Sea full was diverted to irrigate cotton. One result was that large areas of the aral Sea dried up, destroying the fishing industry that it had once supported. the sight of large fishing vessels sitting stranded on vast dry plains became an international icon of environmental degradation (see figure). another effect was from the irrigation itself. Salt buildup (salinization) degraded the fertility of the agricultural soil, and salt in the dust contrib-uted to respiratory problems of the local residents. the northern section of the aral Sea had shrunk by 75 percent by the time a joint project of the World Bank and the government of Kazakhstan built the Kok-aral Dam between the northern and southern sections of the aral Sea. Between 2003 and 2008, the northern part of the sea grew 30 percent larger and 40 percent deeper. the next phase of reclamation will remediate salty soils into sustainable pasturelands.
Perhaps the biggest threat to the success of res-toration ecology is that the causes of the original problem are ongoing. Global warming will continue to promote coral bleaching and the decline of for-ests. Poor people will continue to devastate natural habitats in their pursuit of mere survival. they need an alternative to practices that are destructive to natural ecosystems, such as deforestation for farming and the blast fishing that has destroyed many coral reefs. Meanwhile, the new techniques that are being developed by restoration ecologists will allow habitat managers to quickly take advantage of any improve-ments in the current situation that may come along.
there remains a lot of room for improvement. a recent study that combined the results of 89 res-toration assessments showed that the restored eco-systems had only 44 percent as much biodiversity, and provided only 25 percent as many ecosystem services, as intact ecosystems.
See also extinction; grasslands and savan-nas; invasive species; marshes and swamps; Pleistocene extinction; temperate deciduous forests.
Further readingBartram, William. Travels through North and South
Carolina, Georgia, East and West Florida, the Cher-okee Country, the Extensive Territories of the Mus-cogulges or Creek Confederacy, and the Country of the Chactaws. Philadelphia: James and Johnson, 1791; reprinted Savannah, Ga.: Beehive Press, 1973.
Behre, Karl-ernst. “the role of man in european vegeta-tion history.” in Vegetation History, edited by B. Hunt-ley and t. Webb iii, 633–672. Dordecht, netherlands: Kluwer Scientific Publications, 1988.
Benayas, José M. Rey, et al. “enhancement of biodiversity and ecosystem services by ecological restoration: a meta-analysis.” Science 325 (2009): 1,121–1,124.
chazdon, Robin l. “Beyond deforestation: Restoring for-ests and ecosystem services on degraded lands.” Sci-ence 320 (2008): 1,458–1,460.
comin, Francisco a., ed. Ecological Restoration: A Global Challenge. new york: Oxford University Press, 2010.
Dean, Jeffry S. “anthropogenic environmental change in the Southwest as viewed from the colorado Plateau.” chap. 10 in The Archaeology of Global Change: The Impact of Humans on Their Environment, edited by charles l. Redman et al. Washington, D.c.: Smithson-ian institution Press, 2004.
Dixon, Kingsley W. “Pollination and restoration.” Science 325 (2009): 571–573.
environmental news Service. “the Kazakh Miracle: Recov-ery of the north aral Sea.” august 1, 2008. avail-able online. URl: http://www.ens-newswire.com/ens/aug2008/2008-08-01-01.asp. accessed august 4, 2010.
Harris, Jim. “Soil microbial communities and restora-tion ecology: Facilitators or followers?” Science 325 (2009): 573–574.
Henson, Kate S. e., et al. “the restoration of parasites, parasitoids, and pathogens to heathland communi-ties.” Ecology 90 (2009): 1,840–1,851.
Jackson, Stephen t., and Richard J. Hobbs. “ecological restoration in the light of ecological history.” Science 325 (2009): 567–569.
Koenig, Robert. “Unleashing an army to repair alien-rav-aged ecosystems.” Science 325 (2009): 562–563.
Mann, charles c. 1491: New Revelations of the Americas before Columbus. new york: Knopf, 2005.
———. “addicted to rubber.” Science 325 (2009): 564–566.Manning, adrian D., and David lindenmayer. “Paddock
trees, parrots, and agricultural production: an urgent need for large-scale, long-term restoration in south-eastern australia.” Ecological Management and Resto-ration 10 (2009): 126–135.
This fishing vessel is stranded on dry land that used to be under the surface of the Aral Sea in western Asia. (Lukasz Kruk)
xviii+598_Biodiversity_F.indd 464 11/17/11 1:13 PM

rivers, streams, ponds, and lakes 465
Maran, tiit, et al. “the survival of captive-born animals in restoration programmes: case study of the endan-gered european mink, Mustela lutreola.” Biological Conservation 142 (2009): 1,685–1,692.
Martin, Katherine l., and Katherine l. Kirkman. “Man-agement of ecological thresholds to re-establish disturbance-maintained herbaceous wetlands of the south-eastern USa.” Journal of Applied Ecology 46 (2009): 906–914.
nishihiro, Jun, et al. “toward the restoration of a sus-tainable population of a threatened aquatic plant, Nymphoides peltata: integrated genetic/demographic studies and practices.” Biological Conservation 142 (2009): 1,906–1,912.
normile, Dennis. “Bringing coral reefs back from the living dead.” Science 325 (2009): 559–561.
norton, David a. “Species invasions and the limits to res-toration: learning from the new Zealand experience.” Science 325 (2009): 569–571.
nowacki, Gregory J., and Marc D. abrams. “the demise of fire and the ‘mesophication’ of forests in the eastern United States.” BioScience 58 (2008): 123–138.
Palmer, Margaret a., and Solange Filoso. “Restoration of ecosystem services for environmental markets.” Sci-ence 325 (2009): 575–576.
Redman, charles l. “environmental degradation and early Mesopotamian civilization.” chap. 8 in The Archae-ology of Global Change: The Impact of Humans on Their Environment, edited by charles l. Redman et al. Washington, D.c.: Smithsonian institution Press, 2004.
Rey, Pedro J., et al. “community-level restoration profiles in Mediterranean vegetation: nurse-based vs. tradi-tional reforestation.” Journal of Applied Ecology 46 (2009): 937–945.
Schou, Majbritt Overgård, et al. “Restoring lakes by using artificial plant beds: Habitat selection of zooplankton in a clear and a turbid shallow lake.” Freshwater Biol-ogy 54 (2009): 1,520–1,531.
Schulte, David M., et al. “Unprecedented restoration of a native oyster metapopulation.” Science 325 (2009): 1,124–1,128.
Stokstad, erik. “Oysters booming on new reefs, but can they survive disease?” Science 325 (2009): 525.
Stone, Richard. “nursing china’s ailing forests back to health.” Science 325 (2009): 556–558.
tennesen, Michael. “Black gold of the amazon.” Discover, april 2007, 46–52.
Vieira, Daniel M., et al. “agro-successional restoration as a strategy to facilitate tropical forest recovery.” Restora-tion Ecology 17 (2009): 451–459.
Willis, K. J., et al. “How ‘virgin’ is virgin rain forest?” Sci-ence 304 (2004): 402–403.
Witmer, M. c., and a. S. cheke. “the dodo and the tam-balacoque tree: an obligate mutualism reconsidered.” Oikos 61 (1991): 133–137.
rivers, streams, ponds, and lakes Most of the water in the smallest creeks comes from underground aquifers, often from springs. Rainfall contributes to creeks only briefly after heavy rain events. When two of the smallest (first-order) creeks converge, they form a second-order creek. Of the 3.2 million miles (5.2 million km) of streams and rivers in the United States, half consist of first-order streams and 85 percent are first- to third-order. the columbia River is a ninth-order river, and the Mis-sissippi is a tenth-order river. the major drainage basins in the United States are the following:
● atlantic and Gulf coastal system, in which the major rivers drain into the atlantic Ocean and the Gulf of Mexico
● Great lakes system, which drains into the atlantic Ocean through the St. lawrence River
● Rio Grande system, in which major rivers (the longest of which is the Rio Grande) drain into the Gulf of Mexico
● Mississippi River basin, in which the rivers of most of the central United States join to form the Mississippi River, which drains into the Gulf of Mexico
● Red River basin, in which water from Min-nesota and the Dakotas enters a river that ultimately drains into Hudson Bay
● colorado River basin, in which several riv-ers of the american West join to form the colorado River, which drains into the Gulf of california
● Great Basin system, most of whose rivers do not reach the ocean
● columbia River basin, in which several riv-ers on the landward side of the cascades join to form the columbia River, which drains into the Pacific Ocean
● Pacific coast system, in which the rivers drain into the Pacific Ocean, many of them through San Francisco Bay
With each increase in order, the creek or river undergoes changes such as the following:
● the average velocity of the water increases. the most rapid water is in the middle of the creek or river, because the drag imposed by rocks, soil, and vegetation slows down the water near the banks.
● the water may become cooler, because the average depth of the water increases.
● the oxygen content of the water may increase, if bubbles of air enter the water through the action of waterfalls or rapids.
xviii+598_Biodiversity_F.indd 465 11/17/11 1:13 PM

first entry466 rivers, streams, ponds, and lakes 466
in a large river, especially if it is broad and shal-low, the temperature may have risen and oxygen con-tent decreased, due to the accumulation of eroded soil and the microbial life that comes along with it. creeks may form ponds, and rivers may form lakes, on their way to (usually) the ocean. therefore, rivers, streams, ponds, and lakes offer a great diversity of microhabitats in which different aquatic species of plants and animals live. creeks experience frequent floods, which temporarily disturb the ecological communities. in contrast, lakes may experience very stable conditions for long periods.
One of the major differences between different freshwater habitats is the nutrient content of the water. Oligotrophic water has clear water because it contains very few dissolved nutrients and therefore has few suspended single-celled organisms. Most of the nutrients are within the cells of organisms. the low level of nutrients is associated with the lack of suspended silt and clay. Eutrophic water, in con-trast, has turbid water because abundant nutrients promote the growth of single-celled organisms, and the water may also contain silt and clay. Despite the abundance of nutrients, eutrophic habitats may have lower biodiversity because a few species of algae and other protists may dominate the food web. the abundance of single-celled organisms uses up much of the dissolved oxygen. eutrophic habitats often have slower and warmer water, which holds less oxygen than cool water.
in many cases, freshwater habitats may be eutro-phic because of human influences. Wastewater, even when treated, may contain abundant nutrients. Sources of these nutrients include:
● Nonpoint sources. Soil and organic mate-rial from all along the riverbanks, which often have farms and ranches, may pro-mote eutrophic conditions. Farms fre-quently have fertilizer runoff, especially when chemical fertilizers are heavily used. Ranches and concentrated animal feed-ing operations (caFOs) may produce an immense amount of animal waste, which is rich in nutrients.
● Point sources. Municipalities send storm water and sewage to defined locations on riverbanks. in nearly every american municipality, the sewage is treated; the solid material (sludge) ferments before being dis-posed of, and bacteria break down organic molecules in the dissolved material (efflu-ent), which is also exposed to oxygen by various mechanical means. the purpose of sewage treatment is to eliminate poten-tially dangerous bacteria. Older municipali-
ties have combined water sources in which the storm water and treated effluent come together. newer municipalities may have the storm water, which does not need treatment, separate from the effluent. the advantage of the newer system is that floodwater gener-ally cannot overwhelm the sewage treatment system.
nutrients from human activity often cause eutrophication, which is the artificial production of eutrophic conditions. eutrophication causes freshwater habitats to be very green, the surface layers filled primarily with cyanobacteria. these photosynthetic bacteria can fix their own nitrogen compounds from atmospheric nitrogen gas, and the nitrogen content of the water is not a limiting resource to their growth. it is primarily phosphorus (from soaps and detergents, as well as from human wastes) that causes eutrophication. eutrophication results in freshwater habitats that are typically far more eutrophic than any conditions found in the wild.
While cyanobacteria predominate in conditions of eutrophication, most unaltered freshwaters have food webs based on eukaryotic photosynthetic pro-tists such as desmids and diatoms. Single-celled animal-like protists and small floating animals such as rotifers eat the algae. Small invertebrates, such as copepod crustaceans, eat the smallest animals. larger invertebrates and small fishes eat them. the top predator may be large fishes, such as sturgeons or paddlefishes, or a fish-eating mammal such as the otter.
there are many examples of insects that have aquatic larvae and terrestrial or aerial adults. these include stoneflies; caddisflies, whose larvae con-struct protective cases from sand and organic litter; dobsonflies, whose larvae are called hellgrammites; mosquitoes; dragonflies and damselflies; and some beetles, whose larvae are called water pennies. Drag-onfly nymphs are large enough that they eat small fishes. the insects may spend several years in a lar-val stage and emerge in adult form for a relatively brief mating period, which in the case of mayflies is only a single day. While the larvae are important prey (and are often predators themselves) in the water, the adults are important prey items for terres-trial animals such as birds. in this way, the streams and rivers help to support the food web of the sur-rounding land.
Mammals such as otters, and birds such as kingfishers, bald eagles (Haliaeetus leucocepha-lus), and ospreys (Pandion haliaetus) hunt in the water by diving. the dipper or water ouzel (Cinclus mexicanus) may walk right into the water. Some
xviii+598_Biodiversity_F.indd 466 11/17/11 1:13 PM

rivers, streams, ponds, and lakes 467
animals, such as crayfish, live both underwater and in tunnels in wet soil on the margins of creeks and ponds.
Beavers are important keystone species because they often build dams that transform creeks into ponds. they use their front teeth as chisels to chop down trees, often ones like cottonwood or alder that have soft wood, and they construct dams by inter-weaving the trunks or branches. they live in beaver lodges that are above the water level but have an underwater entrance.
Plants and animals that live underwater have adaptations allowing them to obtain carbon dioxide and oxygen, respectively.
● Underwater plants often have very finely dissected leaves, such as those of the par-rot feather (Miriophyllum aquaticum), or very thin leaves, such as those of the Elo-dea canadensis. emergent plants, including some buttercups, may produce thin or dis-sected leaves underwater but larger, thicker leaves on their stems above water. the very high ratio of surface area to leaf volume allows rapid absorption of bicarbonate ions and dissolved carbon dioxide, without any risk of excessive water loss as would occur with such leaves in the air. Other emergent plants, such as Nelumbo lutea water lilies, Typha angustifolia cattails, and Pontede-ria cordata pickerelweeds, have underwater stems that spread along the bottom of the pond and from which leaves and flowering stems emerge. these plants may have tissues that conduct air down into the submerged stems and roots.
● animals have adaptations that allow their blood to absorb oxygen from the water. Gills of underwater animals such as fishes bring their blood in close contact with the oxygen dissolved in the water. Segmented Tubifex tubifex worms embed themselves in sediments and wave their tails in the water. the abundant hemoglobin in the tails, and the close contact with water that results from their movement, helps to absorb oxy-gen from water that may contain little of it. turbellarian flatworms, which eat detritus and smaller animals and protists on the bot-tom of creeks and ponds, are small and flat, allowing the whole body to absorb scarce oxygen from the water.
Some plants and aquatic insects have adap-tations that are unique to the air-water interface. Duckweeds (family lemnaceae) are monocot flower-
ing plants, each of which consists almost entirely of a tiny floating green plate. Wolffia duckweeds are the smallest seed plants. Some larger aquatic plants, such as water hyacinth (Eichhornia crassipes) and water lettuce (Salvinia molesta), float on the water. Water striders (bugs of the family Gerridae) are light enough that they can actually walk on the surface of the water without breaking the surface tension. Others, such as whirligig beetles (family Gyrinidae), are buoyant and float as they swim half in and half out of the water. the top half of the body is water-repellent, and the top half of each eye is suited for aerial vision, while the bottom half is suited for underwater vision. Backswimmers (bugs of the fam-ily notonectidae) swim upside down in the surface layer of water. Some air-breathing adult insects take air with them as they dive: Water boatmen (bugs of the family corixidae) take a bubble of air with them on the underside of the body, while diving beetles (family Dytiscidae) hold air underneath their hard wing covers. Water scorpions (which are actually bugs, Nepa cinerea) and rat-tailed maggots (the lar-vae of the hoverfly, Eristalis tenax) have a hollow tube protruding from their abdomens that siphons air from above the surface while they feed under-water. Fishing spiders (genus Dolomedes) usually walk on the water’s surface in search of prey (which includes small fishes), but when they dive their bod-ies are surrounded by bubbles of air that allow them to breathe.
animals of streams and rivers have adaptations that allow them to move with or against the cur-rent. Many fishes are strong swimmers. Sculpins (fishes of several different families) brace themselves against the current by embedding their strong fins in the bottom of the creek. Freshwater mussels have larvae that are not strong enough to swim upstream. What is it that keeps the last freshwater mussels from washing out to sea? Freshwater female bivalves of the species Lampsilis perovalis produce floating clumps of larvae that look like small fishes. the clump remains attached to the mother by a thin line, causing the clump to move like a fish in the currents. When a predatory fish tries to eat the larvae, they hitch a ride on the fish and may thereby disperse upstream. Several other kinds of mollusks have a similar strategy.
Several species of fishes are anadromous, that is, they hatch in streams, travel downriver to live in the oceans, then swim back upstream (often to the same stream in which they hatched) to fertilize or lay eggs. Most famous are the chinook, coho, chum, and pink salmon of the Pacific northwest (Oncorhynchus tshawytscha, O. kisutsh, O. keta, and O. gorbuscha). chinook and sockeye salmon return to central idaho, over 900 miles (1,400 km)
xviii+598_Biodiversity_F.indd 467 11/17/11 1:13 PM
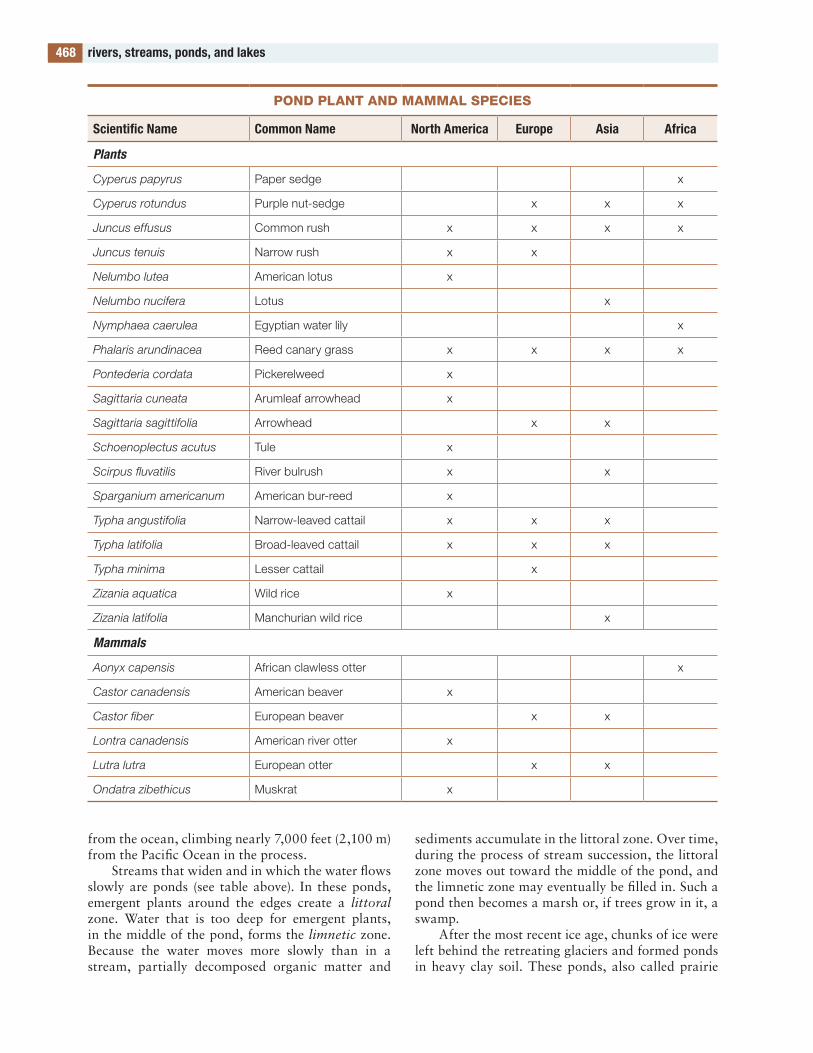
first entry468 rivers, streams, ponds, and lakes 468
from the ocean, climbing nearly 7,000 feet (2,100 m) from the Pacific Ocean in the process.
Streams that widen and in which the water flows slowly are ponds (see table above). in these ponds, emergent plants around the edges create a littoral zone. Water that is too deep for emergent plants, in the middle of the pond, forms the limnetic zone. Because the water moves more slowly than in a stream, partially decomposed organic matter and
sediments accumulate in the littoral zone. Over time, during the process of stream succession, the littoral zone moves out toward the middle of the pond, and the limnetic zone may eventually be filled in. Such a pond then becomes a marsh or, if trees grow in it, a swamp.
after the most recent ice age, chunks of ice were left behind the retreating glaciers and formed ponds in heavy clay soil. these ponds, also called prairie
pond plant and mammal speCies
Scientific Name Common Name North america Europe asia africa
Plants
Cyperus papyrus Paper sedge x
Cyperus rotundus Purple nut-sedge x x x
Juncus effusus Common rush x x x x
Juncus tenuis Narrow rush x x
Nelumbo lutea American lotus x
Nelumbo nucifera Lotus x
Nymphaea caerulea Egyptian water lily x
Phalaris arundinacea Reed canary grass x x x x
Pontederia cordata Pickerelweed x
Sagittaria cuneata Arumleaf arrowhead x
Sagittaria sagittifolia Arrowhead x x
Schoenoplectus acutus Tule x
Scirpus fluvatilis River bulrush x x
Sparganium americanum American bur-reed x
Typha angustifolia Narrow-leaved cattail x x x
Typha latifolia Broad-leaved cattail x x x
Typha minima Lesser cattail x
Zizania aquatica Wild rice x
Zizania latifolia Manchurian wild rice x
Mammals
Aonyx capensis African clawless otter x
Castor canadensis American beaver x
Castor fiber European beaver x x
Lontra canadensis American river otter x
Lutra lutra European otter x x
Ondatra zibethicus Muskrat x
xviii+598_Biodiversity_F.indd 468 11/17/11 1:13 PM

rivers, streams, ponds, and lakes 469
potholes, frequently had no creeks flowing into or out of them and received limited water from under-ground and from rain. Because the water contained few nutrients and because tundra species (includ-ing sphagnum moss) colonized them, most of these ponds developed into bogs, which may be very dif-ferent from regular ponds.
if a lake is large enough, the water may become stratified, with warm oxygenated water at the top and cool, less-oxygenated water at the bottom. nutrients tend to accumulate in the bottom layer as well. the layers are frequently mixed, whether by wind or by the flow of river water into and out of the lake. in addition, in lakes in the temperate climatic zone, there may be an annual overturn of water. this occurs because water is most dense at about 40°F (4°c). as ice melts in the spring, the water becomes denser, and when it exceeds 40°F it sinks toward the bottom of the lake, forcing the bottom water up.
the world’s oldest and deepest lake is lake Bai-kal in Siberia, with an average depth of 2,440 feet (744 m); the greatest depth is 5,380 feet (1,642 m). More than 80 percent of the 1,550 species and vari-eties of animals in the lake live nowhere else. the dominant species of zooplankton is a species found nowhere else. lake Baikal is the home of the Baikal seal or nerpa (Phoca sibirica), one of only three freshwater seal species.
Most large lakes persist for a long time, because sediments do not build up very much within them. the inflow and outflow of river water keeps them cleaned out. large reservoirs, created by large dams, accumulate sediment, since the outflow of water is either through tunnels with turbines or over the spill-way. there are more than 36,000 dams in the world
that are greater than about 50 feet (15 m) in height, and they collectively impound 2,050 cubic miles (8,400 cubic km) of water. this is seven times the volume of all the rivers in the world. artificial reser-voirs are therefore collectively important enough to affect the hydrological cycle of the entire planet.
See also bogs, life in; hydrological cycle; insects; keystone species.
Further readingallan, J. David. “landscapes and riverscapes: the influence
of land use on stream ecosystems.” Annual Review of Ecology, Evolution, and Systematics 35 (2004): 257–284.
Boulton, andrew J., et al. “the functional significance of the hyporheic zone in streams and rivers.” Annual Review of Ecology and Systematics 29 (1998): 59–81.
carpenter, Stephen R., et al. “cascading trophic interac-tions and lake productivity.” BioScience 35 (1985): 634–639.
Dudgeon, D., et al. “Freshwater biodiversity: importance, threats, status, and conservation challenges.” Biologi-cal Reviews 81 (2006): 163–182.
Haag, Wendell R., et al. “an extraordinary reproductive strategy in freshwater bivalves: Prey mimicry to facili-tate larval dispersal.” Freshwater Biology 34 (2006): 471–476.
Paul, Michael J., and Judy l. Meyer. “Streams in the urban landscape.” Annual Review of Ecology and Systemat-ics 32 (2001): 333–365.
Persson, lennart, et al. “Density dependent interactions in lake ecosystems: Whole lake perturbation experi-ments.” Oikos 66 (1993): 193–208.
Keddy, Paul. Wetland Ecology, 2nd ed. new york: Oxford University Press, 2010.
xviii+598_Biodiversity_F.indd 469 11/17/11 1:13 PM

470
seed dispersal Dispersal usually refers to the transport of seeds from the parent plant to a new location. it can also refer to the transport of spores. transport of pollen from one plant to another is called pollination. it is important for seeds to dis-perse to new locations for at least two reasons:
● if seeds did not disperse, they would grow into dense populations near the parent plant. Plants in these dense clumps would com-pete with one another for resources, such as light, water, and nutrients. Pests and diseases would also spread rapidly through such dense plant populations, particularly in the trop-ics, where pest and disease populations are not reduced each year by winter conditions.
● if plants remained in localized clumps, the entire clump might be destroyed by an eco-logical disturbance, such as a fi re or a fl ood. natural selection favors plants that disperse their seeds, not only because the genetic lineage will have a greater chance of sur-vival by traveling to new locations, but such plants will also be able to spread quickly into newly disturbed areas.
Many plants rely on the wind to transport their seeds. the seeds themselves may have fl at surfaces, such as the seeds of catalpa trees, or the seeds may be surrounded by a fruit that provides a thin, fl at surface, as in the seeds of maple trees, whose sur-faces function as glider wings. Dandelion seeds have a thin fruit covering that forms a delicate parachute. these adaptations would not be considered exam-ples of coevolution, since the wind, not other organ-isms, have selected them.
a few species of plants have ballistic seed dis-persal, that is, the fruits explode when touched by an animal or jostled by the wind. Jewel weeds (genus Impatiens) has spring-loaded tissue inside of its fruits, while squirting cucumbers (Ecballium elat-erium) have fruits that hold in water pressure.
Some seeds and fruits fl oat in the water. Because water always fl ows downhill, any upland plant spe-cies that relied solely upon water dispersal would eventually fi nd all of its seeds out in the ocean. But many of these species rely on wind as well as water. Other water-dispersed seeds are transported back uphill during occasional storms. coconuts already live by the seashore. their seeds can fl oat thousands of miles on ocean currents. these seeds often land on sandy shores that have poor soil. coconut seeds have a large and rich supply of food available to the tiny embryonic seedling inside.
Some plants produce dry fruits that have prickles or barbs, which attach to mammal fur. cockleburs are a well-known example. in some cases, as with the puncture-vine, the fruit has sharp spikes that imbed in the soft feet of mammals or fl ightless birds. the unicorn plant, despite its name, has dry fruits with two long horns, which clasp onto the legs of animals or fl ightless birds. the seeds eventually fall out of the dry fruits in a new location. in all of these cases, the animal either attempts to scratch away the fruits or the fruits fall off of the animal. Puncture-vine seeds can even be transported by car tires. these adapta-tions are not often considered examples of coevolu-tion, since few if any animals have evolved special adaptations to either facilitate the dispersal of these fruits or prevent them from attaching.
Many fruits are highly attractive to animals because of their bright color, sugar content, and
S
xviii+598_Biodiversity_F.indd 470 11/17/11 1:13 PM

seed dispersal 471
flavors. the animals (particularly birds and mam-mals) eat the fruits, but the seeds, which have hard seed coats, survive the passage through the intes-tines. the seeds emerge from the animal, having been transported to a new location and receiving a patch of fertilizer in which to begin growing. in many of these species, such as the wild persim-mon, passage of the seed through an animal’s diges-tive tract may be necessary to soften the seed coat enough to allow germination to occur.
in many plant species, it is the seed rather than the fruit that is soft and nourishing to animals. Many seeds have eliasomes, which are fat-producing structures on the seed coat. ants carry the seeds from the ground, where they might otherwise be eaten by mammals or birds, and bury them in the nest, where they can eat the eliasome at their leisure. Once the ants have eaten the eliasome, the seed ger-minates in good soil, which has been aerated and fertilized by ants.
these plants, in effect, pay the animals for the work of dispersing the seeds by providing them with food. However, they expend only as much as is nec-essary. Most wild fruits have much less pulp, relative to the seeds, than do domesticated fruits, which humans have bred to produce excessive amounts of pulp. Furthermore, the pulp of wild fruits attaches tightly to the seed. therefore, it is much easier for the animal to just swallow the seeds than to spit them out. Domesticated fruits, in contrast, have seeds that are easily avoided by humans.
Plants that produce soft fruits discourage ani-mals from consuming fruits in which the seeds are not ready to germinate. this is the reason that unripe fruits are green, hard, starchy, and distasteful. Only after the seeds are mature will the fruit also mature, becoming colorful, soft, sugary, and fragrant. there is, however, competition among animals to eat the fruits as early as possible, that is, earlier than other animals. animals may, therefore, eat fruits that are not quite ripe. the fruits, at this stage, contain astrin-gent digestion inhibitors, which the animals counter-act by ingesting clay. Some parrots consume unripe fruits and then congregate at exposed banks of earth to consume the avian version of kaopectate. (Kaopec-tate is the brand name of an antidiarrheal medication that was originally made from kaolin clay.)
if the animal chews up the seeds along with the fruit pulp, the seeds are killed and dispersal fails. this is why animals that pick seeds out of a fruit and eat them—such as when squirrels eat bois d’arc seeds—are considered seed predators, not dispers-ers. in many cases, the seeds inside of soft fruits are poisonous. Seeds of several fruits in the rose family, for example, contain cyanide. this adaptation tips the balance of natural selection away from animals
that chew up the seeds and toward the animals that simply swallow them.
in some plant species, the seed predator also functions in seed dispersal. Squirrels hoard caches of acorns for the winter, placing the hoards in dif-ferent locations so that they do not lose an entire supply of food. this activity is therefore known as scatterhoarding. the squirrels eat many of the nuts but fail to locate all of them. these acorns germi-nate, having been dispersed to a new location by the squirrel. Some birds disperse the seeds of pines in this fashion. cones of subalpine pines remain tightly closed and are sticky with resin; plundering squirrels cannot open them to get the seeds. Jays, however, can pry open the cones and take the seeds. the jays hide the seeds in rock crevices, often in locations at some distance from the parent tree. Some of these seeds germinate into new pine trees.
Most of the plant species that live on oceanic islands are the descendants of plants whose seeds can disperse long distances. Once the plants are on the island, however, natural selection may favor the loss of dispersal ability, because the plants reproduce more successfully by remaining on the island than by launching out once again over the seas.
the dispersal of soft fruits and scatterhoarding are examples of coevolution, in which each partner provides resources to the other partner, even while investing primarily in its own reproductive success.
See also coevolution; pollination.
Further readingandresen, e., and D. J. levey. “effects of dung and seed
size on secondary dispersal, seed predation, and seed-ling establishment of rain forest trees. Oecologia 139 (2004): 45–54.
cody, Martin l., and J. M. Overton. “Short-term evolution of reduced dispersal in island plant populations.” Jour-nal of Ecology 84 (1996): 53–61.
corlett, R. t. “Frugivory and seed dispersal by vertebrates in the Oriental (indomalayan) Region.” Biological Reviews 73 (1998): 413–448.
Garrison, W. J., et al. “Ballistic seed projection in two herbaceous species.” American Journal of Botany 87 (2000): 1,257–1,264.
lengyel, S., et al. “ants sow the seeds of global diversifica-tion in flowering plants.” Public Library of Science ONE 4 (2009): e5480.
levin, Simon a., et al. “the ecology and evolution of seed dispersal: a theoretical perspective.” Annual Review of Ecology, Evolution, and Systematics 34 (2003): 575–604.
levine, Jonathan M., and David J. Murrell. “the commu-nity-level consequences of seed dispersal patterns.” Annual Review of Ecology, Evolution, and Systemat-ics 34 (2003): 549–574.
xviii+598_Biodiversity_F.indd 471 11/17/11 1:13 PM

first entry472 seedless plants 472
Manzano, Pablo, and Juan e. Malo. “extreme long-dis-tance seed dispersal via sheep.” Frontiers in Ecology and the Environment 4 (2006): 244–248.
narbona, e., et al. “explosive seed dispersal in two peren-nial Mediterranean Euphorbia species (euphor-biaceae).” American Journal of Botany 92 (2005): 510–516.
Sorenson, a. e. “Seed dispersal by adhesion.” Annual Review of Ecology and Systematics 17 (1986): 443–463.
tiffney, Bruce H. “Vertebrate dispersal of seed plants through time.” Annual Review of Ecology, Evolution, and Systematics 35 (2004): 1–30.
Wenny, D. G., and D. J. levey. “Directed seed disper-sal by bellbirds in a tropical forest.” Proceedings of the National Academy of Sciences USA 95 (1997): 6,204–6,207.
seedless plants Seedless plants are bryo-phytes (mosses and their relatives) and pterido-phytes (ferns, horsetails, and club mosses, and their relatives). (algae are sometimes considered to be seedless plants, but they are actually photosynthetic protists.) Seedless plants must live in moist environ-ments on land, for one or both of these two reasons:
● Vascular tissue is the plumbing system of higher plants. Vascular tissue consists of xylem, which brings water up from the soil to the leaves, and phloem, which carries nutrients (dissolved in water) throughout the plant. Bryophytes do not have vascular tis-sue; because they must soak up water rather than transporting it in vascular tissue, they must live in moist environments. Pterido-phytes have vascular tissue.
● Both bryophytes and pteridophytes repro-duce sexually by releasing sperm or similar cells that swim or are carried to eggs. this form of sexual reproduction can only occur in a wet environment, and this is why pte-ridophytes also must live in moist environ-ments. When seed plants evolved, sexual reproduction could be carried out in the absence of a wet environment.
Because life began in water, the earliest plants were aquatic. the first plants that lived on land were seedless plants similar to those that are in moist ter-restrial environments today. an analogous situation occurred with animals, in which the earliest animals on the earth were aquatic, and the first animals that lived on land were restricted to moist environments. as with plants, this was due both to a reduced abil-ity to survive dry conditions and to the inability to sexually reproduce in dry conditions. in both plants
and animals, evolution has produced progressively more species that live in dry environments.
Freshwater green algae, the ancestors of land plants, often live in shallow water, and shallow water frequently dries up. algae that had adaptations that allowed them to continue to function when water became mud would be favored by evolution. the first land plants, as seen in the fossil record, were small plants in wetlands, consistent with this explanation.
nearly all organisms have a sexual life cycle in which a haploid phase alternates with a diploid phase. cells with unpaired chromosomes are hap-loid; cells with paired chromosomes are diploid. a type of cell division known as meiosis separates chromosome pairs and produces haploid cells; in turn, some haploid cells come back together, in a process called fertilization, to restore the original diploid number. in animals, the haploid phase con-sists only of eggs and sperm. But in plants, the hap-loid (gametophyte) phase is multicellular. as green algae evolved into land plants, they diverged:
● Some of them evolved into bryophytes, in which the gametophyte phase is relatively large and independent, while the diploid mul-ticellular phase (the sporophyte) is relatively small and depends on the haploid cells to supply its needs. they reproduce when sperm swim through a film of water to eggs. none of these plants grows taller than a few centi-meters. these plants were already established on the earth by at least 350 million years ago.
● Some of them evolved into plants in which the sporophyte phase was relatively large and independent, while the gametophyte phase was relatively small and sometimes dependent on the diploid cells to supply its needs. these plants were the earliest known land plants. they lived in marshlike habitats during the Silurian period 435 million years ago, although a few fossil spores have been found from the previous Ordovician period. Some of their fossils have been so well pre-served that the vascular pipes can actually be seen in rock cross sections. they were the ancestors of pteridophytes and seed plants.
the earliest land plants, in the swamps of the Silurian period, were only about a meter tall or less and were hardly more than branching stems with reproductive structures at the tips. they had no roots or leaves. they included:
● rhyniophytes, such as Cooksonia. they consisted mostly of branching stems that produced sporangia. Vascular tissue can be
xviii+598_Biodiversity_F.indd 472 11/17/11 1:13 PM

seedless plants 473
recognized in the fossil remains. the rhynio-phytes apparently became extinct without descendants.
● zosterophylls. Some zosterophylls evolved into the ancestors of the club mosses.
● trimerophytes. Some trimerophytes evolved into the ancestors of horsetails, whisk ferns, and true ferns. these plants were common during the Silurian and Devonian periods. their evolutionary descendants included large forest trees of the carboniferous period (see below) as well as smaller plants, which are today’s ferns and fernlike plants.
By the carboniferous period, many of these plants had grown very tall. Some of them, such as Lepido-dendron, were relatives of modern club mosses, while others such as Calamitales were relatives of modern horsetails. then as now, some ferns reached the size of small trees. their leaves were long, narrow, and very simple. their water pipes allowed them to grow tall, but their upward growth was limited by their inability to continue producing new wood. they dominated the carboniferous swamps. they could only live in moist environments, such as swamps, because their sexual reproduction required sperm to swim through water, just as seedless plants do today. therefore, the forests of the carboniferous period were found only in swamps; the hillsides and dry plains were probably barren. Fortunately for these plants, swamp conditions were extremely widespread on earth during the carboniferous period. When the plants died, they did not completely decompose. the piles of organic matter that they produced constitute today’s coal deposits. although coal itself contains no fossils, mineral inclusions (coal balls) often do. these coal balls provide most of the information that scientists have about these plants.
Beginning in the carboniferous period, seed plants started their evolutionary diversification and eventually dominated most of the earth. in very
few places on land do the seedless plants dominate (though the ocean is still the realm of seaweeds, including forests of giant kelp). Seedless plants, how-ever, still have a fair measure of species diversity (see table). though they primarily hide in the shade of the seed plants, they are still important to the biodi-versity of the living world.
dIvErSITy of BryophyTESLiverworts (division Marchantiophyta) are one of the bryophyte lineages. they are a separate lineage from all other plants. their name refers to the flat, roundish, liver-shaped lobes of the gametophytes of some common species that grow on the surfaces of moist soil. Most liverwort species, however, have gametophytes with leaflike and stemlike structures. Because liverworts have no vascular tissue, their flat photosynthetic structures are not really leaves, and the stalks that support these structures are not really stems, although they may look like them. liverworts can be distinguished from mosses (see below) because their leaflike structures are often lobed and have hairlike projections. they have rhi-zoids that look like tiny roots, but their function is to hold the plant in place rather than to absorb water, since they, also, have no xylem. liverworts often live in wet areas and on rock surfaces, to which the rhizoids anchor them. it is the game-tophyte phase that has the leaflike, stemlike, and rhizoid structures.
Male gametophytes of liverworts produce sacs called antheridia underneath the caps of umbrella-like stalks. each antheridium produces many sperm cells. Female gametophytes produce sacs called archegonia under the caps of their stalks. each arche-gonium typically produces one large egg cell. Sperm (which have two flagella) emerge from the anther-idia, swim down the male stalk, over the ground, and up the female stalk to an archegonium, through a layer of water from spring rains or snowmelt. the sperm cells enter the archegonium through an opening, and one of them fertilizes the egg cell. (in some liverworts, antheridia and archegonia are on the same gametophyte.) the egg cell, now a zygote (fertilized egg cell), develops into a sporophyte that contains a capsule (often football-shaped), which produces haploid spores and releases them into the wind. the spores, if they land on a suitable moist surface, grow into new gametophytes.
Mosses (division Bryophyta) are another lineage of bryophytes. like many liverworts, mosses can generally be recognized by the presence of leaflike and stemlike structures and rhizoids. Moss rhizoids, unlike those of liverworts, are multicellular. the rhizoids look like tiny roots, but their function is to hold the moss plant in place rather than to absorb
approximate number of speCies in seleCted groups
of seedless plants
Group Number of Species
Mosses 12,000
Liverworts 8,000
Club mosses 1,200
Horsetails 30
Ferns 20,000
xviii+598_Biodiversity_F.indd 473 11/17/11 1:13 PM

first entry474 seedless plants 474
water, since they have no xylem. Mosses often live in wet areas and on rock surfaces, to which the rhi-zoids anchor them. it is the gametophyte phase that has the leaflike, stemlike, and rhizoid structures. Many mosses have hydroid cells that are similar to xylem and can conduct water from the base to the tip of a gametophyte, though not as well as the xylem cells of vascular plants. although they can metabolize and grow only under wet conditions, most mosses have the ability to tolerate complete and prolonged desiccation. the gametophytes usu-ally grow together in clumps.
Most mosses grow in shaded, moist conditions in forests and along waterways. Peatmoss (genus Sphagnum) forms thick mats on tops of bogs. Bog water is acidic and poor in nutrients. the mats keep the bog water cool, largely isolate it from oxygen, and contribute to its acidity. the few vascular plants that can tolerate these conditions grow with their roots in the mat of moss. Sphagnum bogs are wide-spread in the boreal forests.
Male moss gametophytes produce antheridia at their tips. each antheridium produces many sperm cells. Female gametophytes produce archegonia at their tips. each archegonium typically produces one large egg cell. Sperm emerge from the antheridia, swim down the male stalk, over the ground, and up the female stalk to an archegonium, through a layer of water from spring rains or snowmelt. the sperm cells enter the archegonium through an opening, and one of them fertilizes the egg cell. (in some mosses, antheridia and archegonia are on the same game-tophyte.) the egg cell, now a zygote (fertilized egg cell) develops into a long stalk, the sporophyte (see photograph). (the liverwort sporophyte, in contrast,
does not do this.) the sporophyte stalk grows from, and is fed by, the female gametophyte. the stalk may extend several inches into the air. a capsule at the end of the stalk produces spores and releases them into the wind. the spores, if they land on a suitable moist surface, grow into new gametophytes.
Hornworts are similar to mosses and liverworts, being distinguished by such features as the structure of their spores. they are the evolutionary lineage from which the vascular plants evolved.
dIvErSITy of pTErIdophyTESPteridophytes often superficially resemble bryo-phytes (as club mosses resemble mosses) but have important differences, such as the following:
● the most visible phase of the life cycle is the sporophyte phase. the gametophytes are very small, living on or in the soil.
● the sporophytes have vascular tissue, which allows them to grow taller than any bryophytes.
Club mosses and quillworts (division lycopodi-ophyta) are small plants that grow mainly in shaded, moist conditions (club mosses) or at the edges of lakes and ponds (quillworts). club moss sporophytes are often branched and covered with small leaves (see photograph on page 475). Some Selaginella club mosses have an iridescent layer of cells that make their leaves appear deep blue, an adaptation that apparently helps them to absorb red light. Sporangia (often superficially resembling cones) grow at the tips of many of the branches and release numerous tiny spores into the air. if the spores land on a moist surface, they may germinate and grow into very small (but not microscopic) gametophytes. Game-tophytes of some lycopods are photosynthetic and grow on the surface of the soil, while others grow underground and depend upon a symbiotic associa-tion with fungi to provide food. the male gameto-phytes release sperm, each with two flagella, which swim to a female gametophyte, where they enter the archegonia and fertilize the eggs. the resulting sporophyte grows from and quickly obliterates the female gametophyte. Quillwort sporophytes have long, narrow leaves that grow from an underground corm and among which the sporangia grow. though modern club mosses are small plants, some of them were trees up to 100 feet (30 m) in height, such as the Lepidodendron of the Paleozoic era—the largest plants of the early Paleozoic forests.
Horsetail (division equisetophyta) sporophytes spread by underground rhizomes in wet areas. Single stems grow up out of the rhizomes, which branch out and absorb water and nutrients; they do
Mosses, which do not have roots or water-conducting tubes, grow low to the ground, where they absorb water. They release their reproductive spores into the wind from capsules at the end of stalks. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 474 11/17/11 1:13 PM

seedless plants 475
not produce an extensive root system. the hollow stems have segments that fit inside one another, not unlike drinking straws. Unlike seed plants, in which stems grow at the tips, horsetail stems can grow as each segment becomes longer. Whorls of needlelike leaves emerge from each segment, giving the whole stem the appearance of a bottle brush or the tail of a horse. Sporangia (often superficially resembling cones) grow at the tips of the branches and release numerous tiny spores into the air. if the spores land on a moist surface, they may germinate and grow into very small (but not microscopic) underground gametophytes. the male gametophytes release sperm, which swim to a female gametophyte, where they enter the archegonia and fertilize the eggs. the resulting sporophyte grows from and quickly oblit-erates the female gametophyte. Modern horsetails are relatively small (though some can reach almost 10 feet [3 m] in height), but during the Paleozoic era some horsetails such as Calamites grew much taller.
True fern (division Pteridophyta) sporophytes produce leaves (fronds) that are often large and divided into many smaller surfaces connected by stalks that contain the vascular tissues. in Adian-tum maidenhair ferns, the stalks are dark and nar-
row, suggesting the appearance of human hair. the fronds emerge from underground rhizomes and unfurl, resulting in the appearance of a “fiddlehead.” Staghorn ferns produce two kinds of fronds: flat basal fronds and large fronds that remind people of antlers. the rhizomes produce fibrous underground roots. tree ferns produce trunks, but the trunks do not grow by producing new wood, as do the trees of seed plants. instead, the trunks are reinforced by the growth of aboveground mats of roots. Because of this weak structure, most tree ferns do not grow more than about 20 feet (6 m) tall, though some grow up to 65 feet (20 m) in height.
in some fern species, sporangia develop on the undersides of the fronds in regular patterns along the veins or the edges of the blades. in other fern species, sterile fronds are green and photosynthetic, while fertile fronds are covered with sporangia and do not produce blades (see figure). if a spore lands on a suitable surface, it germinates into a game-tophyte, which is aboveground and green, though very inconspicuous. the male gametophytes release sperm, each with two flagella, which swim to a
Club mosses, such as this Selaginella plana, resemble mosses, but they have roots and water-conducting tubes. (Forest and Kim Starr)
Many ferns produce spores on the undersides of their photosynthetic leaves. This fern, however, produces spores in reddish structures on fronds that are sepa-rate from the photosynthetic leaves. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 475 11/17/11 1:13 PM

first entry476 seedless plants 476
female gametophyte, where they enter the arche-gonia and fertilize the eggs. the resulting sporo-phyte grows from and quickly obliterates the female gametophyte.
Ferns are usually found in moist, shaded areas, although they can also grow in the cracks of rocks (see figure). Filmy ferns (family Hymenophyllaceae) have very thin leaves and grow where they are con-tinually moistened by mist from waterfalls. But, as with other pteridophytes, many ferns are restricted to moist areas mainly by the gametophyte stage. the sporophytes can be quite resistant to dry con-ditions. Bracken fern (Pteridium aquilinum) has spread extensively in acidic soils around the world and fills whole fields (where gametophytes could not survive) just by the growth of its rhizomes. Fronds of Pleopeltis polypodioides, the resurrection fern that grows on tree branches in the southeastern United States, can expand and become green upon exposure to water after shriveling in dry conditions. the stag-horn fern Platycerium veitchii, which also lives on
tree branches, even has the same kind of photosyn-thesis that is found in succulents.
Some ferns, such as the climbing fern Lygodium, spread when the tip of a frond touches the ground and grows into a new plant. Ferns of the genus Sal-vinia and Azolla float on the surface of water; some of them have become invasive species, covering the surfaces of rivers. Azolla ferns have symbiotic cyano-bacteria that fix nitrogen into fertilizer; Azolla ferns growing in plots of aquatic crops such as taro act as a natural fertilizer.
Whiskferns (division Psilophyta) and the adder’s-tongue ferns (Ophioglossophyta) are lin-eages of pteridophytes that were once classified with the true ferns. like horsetails, they are rela-tively small plants that grow from underground rhizomes; in whiskferns, these rhizomes serve in place of roots. the aboveground stems branch and produce sporangia, which release numerous tiny spores into the air. if the spores land on a moist sur-face, they may germinate and grow into very small (but not microscopic) gametophytes. Unlike fern gametophytes, they grow underground and depend upon a symbiotic association with fungi to provide food. the male gametophytes release sperm, each with several flagella, which swim to a female game-tophyte, where they enter the archegonia and fertil-ize the eggs. the resulting sporophyte grows from and quickly obliterates the female gametophyte. Genus Psilotum does not produce leaf blades, and the others produce just one leaf at a time. Despite its superficial resemblance to some of the earliest land plants (such as Rhynia), Psilotum evolved from the fern lineage.
See also seed plants.
Further readingconard, Henry S., and Paul l. Redfearn, Jr. How to Know
the Mosses and Liverworts. Dubuque, iowa: William c. Brown, 1979.
Hebant, c., and David W. lee. “Ultrastructural basis and developmental control of blue iridescence in Selagi-nella leaves.” American Journal of Botany 71 (1984): 216–219.
Holtum, J. a. M., and K. Winter. “Degrees of crassulacean acid metabolism in tropical epiphytic and lithophytic ferns.” Australian Journal of Plant Physiology 26 (1999): 749–757.
Gensel, Patricia G. “the earliest land plants.” Annual Reviews of Ecology, Evolution, and Systematics 39 (2008): 459–477.
Graham, linda e., Martha e. cook, and J. S. Busse. “the origin of plants: Body plan changes contributing to a major evolutionary radiation.” Proceedings of the National Academy of Sciences USA 97 (2000): 4,535–4,540.
The fern Asplenium septentrionale grows over a wide area of North America and Europe, living in crevices on boulders, but is very rare in each location. In the Black Hills of South Dakota, there are only a few individuals, all of them on granite boulders near the base of Mount Rushmore. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 476 11/17/11 1:13 PM

seed plants 477
Kenrick, Paul, and Peter crane. “the origin and early evo-lution of plants on land.” Nature 389 (1997): 33–39.
Moran, Robbin c. A Natural History of Ferns. Portland, Ore.: timber Press, 2004.
niklas, Karl J. The Evolutionary Biology of Plants. chi-cago: University of chicago Press, 1997.
Qiu, y. l., et al. “the deepest divergences in land plants inferred from phylogenomic evidence.” Proceedings of the National Academy of Sciences USA 103 (2006): 15,511–15,516.
Shear, William a. “the early development of terrestrial ecosystems.” Nature 351 (1993): 283–289.
seed plants Seed plants are plants that have vascular tissue and produce pollen and seeds, as explained below:
● Vascular tissue. Vascular tissue consists of xylem and phloem. Xylem tissue conducts water up from roots into stems and leaves. Phloem tissue conducts water with sugar and other organic molecules, usually down from the leaves where the sugar is made to the roots. Some such as ferns also have vascular tissue. Roots, stems, and leaves are defined partly by the presence of vascular tissue. Mosses and their relatives do not have vascular tissues and do not have true roots, stems, or leaves. Because of vascular tissue, seed plants are well adapted to life on dry land.
● Pollen. Pollen grains contain one or more sperm nuclei inside a hard protein coat that protects the sperm and other male cells while they are transported through the air by the wind or by pollinators. in seedless plants, sperm must swim through layers of water from one plant to another. Because pollen carries sperm through the air, seed plants are well adapted to life on dry land.
● Seeds. Seeds contain an embryonic plant and a food supply, surrounded by a pro-tective coat. Because the embryo is already partly grown and has a food supply, a plant can germinate from a seed and grow quickly, whereas seedless plants must begin their growth from single spores.
Most species of eukaryotes have life cycles in which meiosis alternates with fertilization. Meiosis eventually produces haploid eggs and sperm that fuse together during fertilization. Plant life cycles differ from those of protists, animals, and fungi by having a multicellular haploid phase called the gametophyte stage. the gametophytes produce eggs
and/or sperm. the haploid structures of the simplest land plants, which evolved earliest, live in water or moist soil. in seed plants, however, the male game-tophytes are pollen grains, which contain sperm and travel through the air from one plant to another; the female gametophytes remain within the immature seed. Pollen grains land on female tissue and ger-minate, producing pollen tubes that grow toward the female gametophytes. Sperm nuclei, instead of swimming through a layer of water in moist soil, travel through pollen tubes. the fertilization of egg and sperm nuclei produce a zygote, which is the beginning of the diploid, or sporophyte, phase of the plant life cycle. Seeds contain, feed, and protect embryonic sporophytes.
the two major groups of seed plants are the gymnosperms and the angiosperms.
GymNoSpErmSGymnosperms constitute a broad category that includes all extant seed plants that are not flower-ing plants, as well as many extinct forms. it is, therefore, not a monophyletic category. in gymno-sperms, pollen grains usually develop inside of small male cones, and seeds often develop inside of larger female cones. Seeds of gymnosperms (Greek for “naked seed”) develop without being surrounded by parental tissue.
the first gymnosperms evolved during the Paleozoic era. the earliest gymnosperms were Elkinsia and Archaeosperma, which appeared late in the Devonian period. Gymnosperms remained a relatively minor component of the forest vegeta-tion during the carboniferous period, when seedless plants grew to a very large size, such as tree-sized club mosses. During the Permian period, as cooler and drier conditions spread, the large seedless trees declined in abundance, and gymnosperms domi-nated the forests. among these gymnosperms were the trees known as Cordaites. two modern groups of gymnosperms, the conifers and the cycads, evolved during the Permian period. also common in the late Mesozoic era were seed ferns (pteridosperms), which were not ferns and were probably not related to modern gymnosperms.
in the cooler, drier conditions of the Mesozoic era, gymnosperms (especially conifers and cycads) dominated the forests of the triassic and Jurassic periods. Many of the conifers resembled the mod-ern norfolk island pine, which is not really a pine. the Wollemi pine, which is also not really a pine, was thought to be extinct until it was discovered in australia in 1994. true pines and their relatives also evolved during the Mesozoic.
another group of Mesozoic gymnosperms was the bennettitalean plants. the details of their leaf
xviii+598_Biodiversity_F.indd 477 11/17/11 1:13 PM

first entry478 sexual selection 478
and wood anatomy, as well as of their reproduc-tive structures, suggest that they may have been the ancestors of the flowering plants. Some bennet-titalean plants had female conelike structures sur-rounded by male reproductive structures inside of bracts, which is very similar to the structure of a flower. When the flowering plants evolved at or before the beginning of the cretaceous period, they displaced the gymnosperms from the tropical areas.
Modern gymnosperms spread during the ceno-zoic era to their present habitats. Modern gymno-sperms include these groups:
● Conifers. Most conifers have stiff needlelike or scalelike evergreen leaves. Some primitive conifers, such as the podocarps of new Zea-land and South america, and the auracarians of chile, are found in the Southern Hemi-sphere. Most modern conifers are junipers, cypresses, pines, spruces, firs, hemlocks, and larches. Pines evolved during the Mesozoic era but have spread extensively since, espe-cially in cool regions with poor soils.
● Cycads. cycads look like small palm trees. they produce seeds on specialized leaves rather than in cones.
● Ginkgoes. there is only one modern spe-cies of ginkgo (the maidenhair tree, Ginkgo biloba) that closely resembles ginkgoes that lived millions of years ago. this species may have been saved from extinction by chinese monks who grew them in monasteries. Gink-goes have leaves with veins that diverge rather than branching out. they produce seeds sin-gly on stalks rather than inside of cones.
● Gnetales. this is a loosely defined group of plants with gymnosperm reproductive struc-tures, some of which have thin, flat leaves that resemble those of flowering plants. One member of this group is the desert ephedra bush.
aNGIoSpErmSangiosperms (the flowering plants) are one of the largest groups of plants, with at least 260,000 living species in 453 families. angiosperms live in nearly every habitat except the deep oceans. they range in size from large trees to tiny floating duckweeds. Despite their tremendous diversity, angiosperms are a monophyletic group, which means that they all evolved from a common ancestral species.
the major lineages of the angiosperms are as follows:
● basal angiosperms, which include plants such as magnolias, water lilies, and butter-
cups, whose flowers often have many sepa-rate parts
● monocots, such as grasses and lilies ● eudicots, which include most of the other
angiosperms, such as roses, sunflowers, and oak trees
angiosperms have flowers, which facilitate the transport of pollen to the immature seeds and fruits, which facilitate the dispersal of seeds to new locations. extensive coevolution between flowering plants and animals (such as pollinating insects and mammals that disperse fruits) has contributed to the explosive speciation of flowering plants and animals that has occurred since the beginning of the creta-ceous period.
See also cladistics; coevolution; conifers; flowering plants (angiosperms); seedless plants.
Further readingBaskin, carol c., and Jerry M. Baskin. Seeds: Ecology, Bio-
geography, and Evolution of Dormancy and Germina-tion. San Diego: academic Press, 2001.
Bewley, J. D., M. Black, and P. Halmer, eds. The Encyclope-dia of Seeds: Science, Technology, and Uses. Oxford, U.K.: Oxford University Press, 2006.
Henry, Robert J. Plant Diversity and Evolution: Genotypic and Phenotypic Variation in Higher Plants. Oxford, U.K.: Oxford University Press, 2005.
Kesseler, Rob, and Wolfgang Stuppy. Seeds: Time Capsules of Life. Richmand Hill, Ont.: Firefly Books, 2006.
leubner, Gerhard. “the seed biology place.” available online. URl: http://www.seedbiology.de. accessed May 16, 2011.
Museum of Paleontology, University of california, Berke-ley. “introduction to the Bennettitales.” available online. URl: http://www.ucmp.berkeley.edu/seed plants/bennettitales.html. accessed May 16, 2011.
Richardson, Sarah. “Seeds: How flowering plants beat out the competition on ancient earth.” Discover, October 2006, 36–41.
Wollemi Pine international. “the Wollemi Pine.” available online. URl: http://www.wollemipine.com. accessed May 16, 2011.
sexual selection Sexual selection is the evolu-tionary selection of characters that enhance repro-ductive success rather than (even at the expense of) survival. the individuals in which the characteristic is well developed will reproduce more often and leave more offspring than individuals in which the characteristic is absent or less well developed.
Sexual selection is similar to natural selection, in that a characteristic evolves because it causes the
xviii+598_Biodiversity_F.indd 478 11/17/11 1:13 PM

sexual selection 479
individuals that possess the trait to leave more off-spring than individuals that do not possess it. the difference is that sexual selection favors characteris-tics that directly enhance reproductive success rather than characteristics that enhance it indirectly through survival or obtaining resources. charles Darwin pro-posed natural selection as the major mechanism of evolution in his world-changing 1859 book On the Origin of Species by Means of Natural Selection. He also proposed sexual selection in his 1871 book The Descent of Man and Selection in Relation to Sex. Sexual selection may be responsible for even more of the biodiversity in the world than is natural selection. this is because in natural selection, certain adapta-tions work and others do not, whereas in sexual selec-tion, almost anything can happen.
SExual SElECTIoN IN aNImalSin most animal species, males can produce more off-spring than can females. this is partly because sperm cells are small and cheap, while egg cells are large and expensive. in animals with external fertilization, such as most fishes, this may be almost the entire difference in reproductive cost between males and females. in species with internal fertilization (such as most terres-trial vertebrates), at least part of the embryonic devel-opment occurs within the body of the female. Bird and reptile eggs are relatively large by the time the female lays them. in species with live birth (vivipary), it is the female in which embryonic development occurs. in most animal species in which newborns receive paren-tal care, it is the female that provides a disproportion-ately large amount of that care.
Sexual selection can operate at the level of the reproductive cell or of the reproductive individual, as in these examples:
Sexual selection upon sperm. in many animal species, males produce millions of sperm, while females produce far fewer eggs. this can lead to an intense selection process occurring upon the sperm themselves. in humans, very few of the many mil-lions of sperm reach the egg. this is why a sperm count of 50 million, although that sounds like a large number, may indicate infertility.
● the acidic environment of the vagina kills many sperm.
● the sperm must also be strong swimmers to climb up through the cervix into the uterus.
● Because the sperm are genetically different from the cells of the woman’s body, the woman’s immune system begins to launch an attack on the sperm.
Selection of strong sperm may also result in the selection of strong offspring, because many of the
metabolic genes that promote sperm success also promote the vigor of the offspring. Sexual selec-tion has not only favored the production of strong sperm but also the production of chemicals in the semen that partially suppress the woman’s immune response. Selection for successful sperm is a very important part of reproductive success. each sperm is the product of a long series of cell divisions, each one an opportunity for mutations. there are there-fore many mutant sperm that need to be “weeded out.” this explanation seems inadequate to explain all of the characteristics of sperm cells in all animal species. no convincing explanation has yet been offered for why the sperm of Drosophila bifurca, a species of fruit fly that is about one-20th of an inch (1 mm) long, can have tails almost two inches (6 cm) long, as described by evolutionary biologists adam Bjork and Scott Pitnick.
Sexual selection at the individual level. Because one male can produce so many sperm, and since in many animal species the act of mating is the male’s only contribution to the success of the offspring, a female animal cannot typically increase her repro-ductive success by mating more often. therefore, it is usually males that compete with other males for access to females; it is females that choose which males they allow to fertilize their eggs.
Females may choose among males on the basis of genetically based characteristics that have no con-nection with the quality of the male. according to the “sexy sons” hypothesis, females in a species may choose a male with any characteristic, perhaps an outlandish color pattern. the sons of these females will also have this characteristic, which will enhance their chances of being chosen by females in the next generation. evolutionary theorist R. a. Fisher first pointed out that this could result in a positive feed-back loop of runaway sexual selection.
Females may also choose among males on the basis of genetically based characteristics that indi-cate the quality of the male:
● the best genes may be those that allow males to obtain resources most effectively.
● the best genes may be those that allow resistance to parasites and diseases. Because parasites can evolve so much more rapidly than their hosts, most animal species are in a continual race to keep ahead of their parasites.
there are many fitness indicators that females can use to choose the best males. a fitness indica-tor is any trait that reliably displays the strength of the state of health of the male. a healthy male is likely to be superior in resource acquisition and
xviii+598_Biodiversity_F.indd 479 11/17/11 1:13 PM

first entry480 sexual selection 480
in resistance to parasites, therefore likely to have higher potential fitness. a fitness indicator must, therefore, be expensive—and the more expensive it is, the better a fitness indicator it is. this is the “handicap principle” proposed by evolutionary biologist amotz Zahavi. evolutionary biologists William Hamilton and Marlene Zuk proposed that the most valuable fitness indicators are those that show the male to be free from parasites. Fitness indicators include:
● a male that defends a large territory in prime quality habitat not only has more resources to offer to the female but is prob-ably healthier and therefore able to drive away other males. it is not only expensive for the male to obtain and defend the terri-tory, but to advertise it to females. a male mockingbird may sing all day for several weeks, which uses a lot of energy and makes him noticeable to predators.
● Bodily ornamentation is another fitness indi-cator. consider a bird species that uses col-ored feathers as fitness indicators. any male, even an inferior one, can (potentially) pro-duce a few small colored feathers, but only a healthy one, free of parasites, is likely to produce many long colorful feathers. High testosterone levels, which enhance aggres-sion in birds and mammals, also decrease the function of the immune system. the males that manage to have both the longest, most colorful feathers as well as the most aggressive territorial defense may be able to leave the most offspring. Feathers will con-tinue to evolve to be longer and more color-ful until the feathers become too expensive or burdensome. in many bird species, male ornamentation consists of only a few feath-ers or a crest, but in some, such as peacocks and birds of paradise, the ornamentation is so excessive that it constitutes a signifi-cant drain of resources and puts the male in greatly enhanced danger of attack by preda-tors. Researchers such as evolutionary biolo-gist Malte andersson have experimentally augmented the feathers of some males and found that the males with artificially long feathers have more offspring.
● in many bird species, males congregate in leks to display their plumage and their mat-ing dances, and females stroll around and choose from among them.
● Symmetry is a fitness indicator in many species, including humans, who show a preference for faces and bodies that are sym-
metrical. Poor health often results in asym-metry between the right and left sides of a vertebrate.
expensive fitness indicators among mammals include antlers (which in the now-extinct irish elk could reach more than six feet [2 m] in expanse). natural selection can put a stop to sexual selection, but not before excesses of ornamentation, some of them bacchanalian, are produced. the more expen-sive the better, up to a point, because if the orna-mentation were cheap, less healthy males could cheat by producing an ornament that merely made them look healthy. aside from being expensive, and being truthful representations of health, fitness indicators may be arbitrary, as reflected in the tremendous diversity of feather ornamentation and courtship dances among the more than 10,000 bird species.
Once fitness indicators come into existence, as a result of sexual selection, they can serve as reproduc-tive isolating mechanisms that help to separate one population into two. this can lead to speciation, when males with novel traits are chosen by females with a preference for those same novel traits.
in many animal species, sexual selection has produced particularly marked characteristics that facilitate competition between males:
● Size dimorphism. in many animal species, the male is larger than the female even though it is the females, not the males, that have most of the costs of reproduction. large size gives males a competitive advan-tage in fighting with one another.
● Weapons. the same antlers that serve as fit-ness indicators for female choice also serve as weapons for fighting other males.
● Impulsiveness. in most species, it is the males that are more impulsive, and some-times more violent. this is why, as Dar-win noted in 1871, it is usually males (even among the invertebrate animals) that are, as he said in his inimitable Victorian prose, more “pugnacious” than females. in their hurry to mate, males can sometimes be absurdly undiscriminating: Male turkeys, for example, will attempt to mate with a replica of a female turkey head.
● Interference with the reproduction of other males. there are numerous instances in which males interfere with the opportu-nity for other males to inseminate females. in some stick insects such as Necroscia sparaxes, the male will copulate with the female for weeks at a time, thus keeping rival males away. Whether in flies or in
xviii+598_Biodiversity_F.indd 480 11/17/11 1:13 PM

sexual selection 481
primates, males may have very large repro-ductive organs that deliver a large amount of sperm. this helps to outcompete the sperm that a previous male may have left. Male voles produce more sperm if there are other males in the vicinity than they do if they are alone. the males of some species have penises with bristles, which are believed to scrub away sperm left by other males. in other cases, males produce secretions that plug up the female’s reproductive tract after he has mated with her, thus preventing the sperm of other males from entering. How-ever, the males have also evolved the ability to remove the plugs inserted by other males. evolutionary biologist Rebecca Dean and colleagues explain that when young or old males copulate, they may not be successful at producing offspring but may succeed in keeping other males from copulating.
● Stealth. in many cases, males have evolved to be stealthy in their approach to females. in some salamanders, such as Plethodon jordani, the female follows the male until the male drops a spermatophore, a struc-ture that contains sperm; the female then sits on the spermatophore and receives the sperm. Sometimes a third male will intrude between the couple, waddle along as if he is a female, and then when the first male drops his spermatophore, the second runs off with the female. in some fishes, some males imi-tate females, infiltrate themselves into the harems controlled by dominant males, and mate with some of the females. Perhaps the ultimate example is when certain male bats (Myotis lucifugus) creep around amidst hibernating females and inseminate them.
● Chemical components of semen. Some male houseflies even produce semen that contains proteins that reduce the female’s interest in copulation—after he has finished with her, of course. this reduces the chance that she will accept the advances of another male. in some cases, the compounds in the semen are even slightly poisonous. in one experiment, in which male flies were allowed to evolve in competition but female flies were not, the semen from the male flies actually killed the females.
Male-male competition may in some cases have promoted speciation. in dragonflies and damselflies, males clasp females behind their heads to mate with them, thus keeping other males away. their penises scrub out the sperm of previous males. this pro-
cess can be so violent that the females get severely injured. But the post-reproductive survival of the female does not matter to the male. according to evolutionary biologist Ola Fincke, the female evolu-tionary response to this conflict of interest may have been new shapes and colors, which make it more dif-ficult for the males to recognize the females.
For animals with either monogamous or harem mating systems, in which the male can be reasonably confident of his exclusive access to the female, males have small reproductive organs. Gorilla males, for example, have penises even smaller than those of some ducks. in these instances, the male reproduc-tive investment may be completed upon successful copulation, and thus the male may be expendable. in the cases of many spiders and praying mantises (Mantis religiosa), females often eat the males—in the case of the mantis, even while copulation is occurring.
there are cases in which the males have the larger reproductive investment, and females choose among them. Females may provide eggs to males, who take care of them; often, males receive eggs from more than one female:
● in the rhea (Rhea Americana), a South american bird similar to the ostrich, the male incubates the eggs and takes care of the hatchlings.
● in some wading birds, such as the red-necked phalarope (Phalaropus lobatus), it is the males that raise the young. the red-necked phalarope is also one of the few bird species in which the females are more brightly colored than the males.
● in some stickleback fishes such as Gasteros-teus aculeatus, females prefer to provide eggs to males who already have eggs in the nests they have built. this has led to the evolution of stealing behavior, in which males will steal eggs from the nests built by other males.
● Males of the fish species Cyrtocara eucinos-tomus, though only 0.4 inches (10 cm) in length, build enormous sand castles a meter wide at the base, one mouthful of sand at a time. the female chooses the male that con-structs the best castle.
● in some frogs and fishes (such as the sea-horse), the female lays the eggs, and then the male scoops them up into his mouth to protect them while they hatch, a process called mouth-brooding. this activity ben-efits the male in that he can be certain the offspring are his. However, during this pro-longed period of time, the male cannot fer-tilize any other eggs. this is a considerable
xviii+598_Biodiversity_F.indd 481 11/17/11 1:13 PM

first entry482 sexual selection 482
expense for the male. Because it typically takes more than one male to mouth-brood the eggs produced by one female, it is the females that compete with one another for access to mouth-brooding males.
therefore, while sperm are individually cheap, male reproductive expenditure can be expensive.
in some instances, the females may be so widely dispersed that a male is lucky to ever find one, and a female is unlikely to be found by more than one male. in these cases, there is no advantage for a male to be large. in some species of invertebrates (such as some barnacles) as well as some fishes (such as deep sea angler fishes), the males are small, and even merge into the flesh of the females so that they resemble an organ or a lump on the female.
Sexual selection has operated strongly during human evolution. Men have evolved numerous fit-ness indicators to influence female choice. Some of these fitness indicators are visible characteristics such as muscularity; as a result, sexual selection may have caused men to become more muscular than they need to be. Most male fitness indicators have less to do with appearance than with function. the human penis is larger than that of some other primates. there may be two reasons for this. One reason is, as noted above, a large penis is associated with promiscuous mating systems, as in chimpanzees, while a small penis is associated with harem mating systems, as in gorillas; humans are intermediate. another reason is that large penis size may have been selected because of the tactile pleasure that it provides to the females, who would then choose to keep company with the man who could provide this pleasure. this explanation would make no sense if pregnancy often occurred after a single copulation (as in cats), because by the time the female deter-mined that a certain male gave less pleasure than others it would be too late to reject him. However, because numerous copulations are typically neces-sary for pregnancy to occur in humans, pleasure is a useful piece of information on which a woman could partly base her decision. therefore female choice that maintains a relationship, as well as the choice that initiates it, has been important in human evolution.
as noted above, males sometimes select among competing females. this is especially the case in animal species that have expensive offspring, such as humans. Human reproduction is so expensive that there is a definite survival benefit to the offspring if both parents help to raise them. the genetic fit-ness of the men, as well as the women, would be enhanced if the man stays with the woman and helps with the provision and protection of the offspring.
if the woman has fitness indicators that will attract the man and entice him to stay, her reproductive suc-cess may be enhanced, as well as his. the man, in turn, may want his genes to be carried by a woman who is most likely to raise the children to adult-hood. Women have, therefore, also evolved fitness indicators that show that they are healthy and well nourished. a primary indicator of health is the face. a face that is symmetrical and unblemished is an indicator of past and current health. another indica-tor is fat accumulation. Fat accumulation serves not only as a fitness indicator, but as a direct contribu-tor to the ability to raise children—through nurs-ing and later the ability to survive food shortages during which feeding the child or children receives top priority. Fat accumulation throughout the body would be less obvious than fat accumulation in vis-ible places such as breasts and buttocks. to be effec-tive, a fitness indicator must be expensive and show variation among individuals. Faces, breasts, and but-tocks not only indicate health and nutrition but also age. if skin did not wrinkle and if breasts did not sag with advancing age, they would not have served as reliable indicators of female fitness.
evolutionary scientists have been slow to catch on to Darwin’s insight that sexual selection has been paramount in human evolution. they have instead focused on the more practical survival aspects of life as paramount in human evolution, for example: lan-guage as a means of communicating the locations of resources or sorting out social hierarchies; tools as means of obtaining resources or as weapons against competing tribes; and the relative hairlessness of the human body as an adaptation that allows warm-blooded mammals to cool off in the hot african savannas. in general, all of the many adaptations that are collectively called intelligence have been attributed to natural selection for the serious busi-nesses of resource acquisition and war. in many cases, these explanations were and are believable. tools certainly have enhanced the ability of humans to exploit natural resources, and complex social interactions would be impossible without language (even though chimpanzee societies are quite complex despite the lack of true language).
But even the believable explanations seem insuf-ficient. consider, for example, language, which some scientists consider the major adaptation that distin-guishes human from animal intelligence. theories proposed early in the 20th century attempted to explain the origin of language in terms of practical natural selection. What should have been obvious even at that time is that languages are far more com-plex than they need to be for such simple practical requirements. Just how complex does language have to be in order to say “lion!” or “time for lunch!”?
xviii+598_Biodiversity_F.indd 482 11/17/11 1:13 PM

sexual selection 483
Perhaps the best explanation for the origin of language is that it was an indicator of the quality of intelligence and of the brain. People who could not just talk but speak with complex grammar and poetical allusions obviously had good brains. Many other characteristics of human intelligence, such as the production of art and music, serve as clear indi-cators of the quality of the brain and could therefore function as sexual fitness indicators. evolutionary biologist Geoffrey Miller explains that the human brain itself is a fitness indicator, since it is such an expensive organ (consuming tremendous amounts of energy and oxygen) and at least half of the genes are involved in brain development. therefore, prac-tically anything that the brain does can be a fitness indicator. as such, it can be the subject of sexual selection. By choosing intelligent mates, both men and women could be reasonably certain that their mate had good genes.
another example of a characteristic often con-sidered to have evolved for its practical benefits is hunting. the practical explanation of hunting is that men bring home the meat while women gather nuts, berries, and grains. While plant materials may pro-vide most of the calories, the meat brought home by the men provides essential nutrients that would be missing from a vegetarian diet. even if it is true that a purely vegetarian diet would be inadequate, this does not explain why men hunt big game in nearly every tribe that has been studied and continue to do so today in civilized nations. Men could provide adequate meat by hunting small game. in fact, they could provide more meat in this manner and with considerably less danger to themselves. Simply put, hunting big game is not a practical way for a man to provide for his family. it appears that hunting, in all societies, has been and is a sport. its purpose is to be a fitness indicator: Only very strong males with good brains can show off their qualities by undergo-ing the physical exertion and having the intelligence to track down, or trick, dangerous game animals and then kill them. By choosing good hunters, the women could be reasonably sure that their mates were strong and intelligent.
Many human characteristics, not the least of which is the large human brain, are part of a pattern called neoteny, in which juvenile characteristics are retained into adulthood. this explains many unique human features as part of a suite of neotenous traits but does not explain why neoteny occurred in the first place. What, if anything, caused neoteny to operate during human evolution? it is possible that the retention of juvenile characteristics was selected because they made older people look younger, thus deceptively enhancing fitness indicators of youth in both sexes.
Some scientists question the pervasiveness of sexual selection in the animal kingdom. evolution-ary biologist Joan Roughgarden points out that if sexual selection were the most important explana-tion of sexual reproductive behavior among animals, then such behavior would be mostly heterosexual. Other forms of sexual behavior are too common to be dismissed as mere exceptions to the rule. Sexual behavior plays other roles, such as dominance or the enhancement of cooperation. Other scientists point out that, while this is true, it was sexual selection that produced the reproductive systems in the first place, and they were then available for other func-tions, such as dominance and cooperation.
SExual SElECTIoN IN plaNTSMate choice, resulting in sexual selection, can occur in organisms other than animals. evolutionary biol-ogists Mary Willson and nancy Burley pointed out not only that male plants, or the male function of bisexual plants, compete with one another in terms of sperm or pollen production (as explained previ-ously) but also that female plants, or the female function of bisexual plants, may be able to discrimi-nate among sperm or pollen. Male competition in plants occurs for the same reason that it does in ani-mals: a plant can produce more sperm, or more pol-len, than a female plant can produce eggs, or ovules, and the female plant, or female function within the plant, not only produces eggs or ovules, but also nourishes the resulting embryo, often within a seed, and provides for its dispersal, often within a fruit. instances of sexual selection in plants may include the following:
● Male competition can cause plants to undergo male reproduction earlier than female reproduction, since the first plant to produce sperm or pollen can fertilize numer-ous female plants. Botanist Stanley Rice and his students have shown that in cot-tonwoods (Populus deltoides), which have separate male and female trees, the male trees open their flower buds earlier, on the average, than female trees—although, of course, their flowering times must overlap or else pollination would fail. Male trees that open earlier may pollinate many female flowers before other male trees get a chance to do so. this could be interpreted as sexual selection among male trees.
● Plants do not copulate. Pollen must be car-ried from the male parts of one plant to the female parts of another. Plants rely on the wind, or on many species of pollinators, to accomplish this. the plants that more
xviii+598_Biodiversity_F.indd 483 11/17/11 1:13 PM

first entry484 sexual selection 484
effectively attract pollinators are more effec-tive at sexual reproduction. all of the char-acteristics of plants related to pollination can therefore be considered the products of sexual selection. in animal-pollinated dioe-cious plant species, in which some plants are male and others are female, male flowers are usually larger and more colorful than female flowers. this can be interpreted as male-male competition for sperm delivery, and therefore sexual selection.
● Once a flower has been pollinated, the pol-len must grow a tube down into the flower, making contact with an ovule, which may be a great distance away. in maize, for example, the tube from a pollen grain must grow up to eight inches (20 cm), a thousand times the length of the pollen grain, to reach the ovule. the sperm nucleus then descends through this tube to complete the act of fer-tilization. the structure through which the pollen tube must grow, the style, can be said to exist for the sole purpose of “weeding out” inferior pollen grains. Botanist allison Snow has found that in the bush Hibiscus moscheutos, pollen grains from some plants grow tubes faster than pollen grains from other plants. Because flowers often receive more pollen than they need, there is often a race between the different pollen tubes to be the first ones to arrive at the immature seeds—which may constitute sexual selec-tion among the pollen tubes.
● in terms of differences among pollen grains in the rate of growth, the style functions just as the food source through which the tube grows. But the pistil is more than just a food source for the pollen tube. the pistil itself may chemically encourage the growth of some pollen tubes and discourage others. this is difficult to demonstrate. it has been shown that there is greater variation in pol-len tube growth rate in the pistils of healthy plants than of plants that are experiencing stresses such as drought. this implies but does not prove that a healthy plant chemi-cally distinguishes among pollen tubes. the haploid cells surrounding the egg cell, in the immature seed, have genes that control the release of sperm from the pollen tubes. this implies the possibility of sexual selec-tion in which the female cells are in control of which sperm are allowed to fertilize the immature seeds.
● Botanist Diane Marshall found that rad-ish plants may provide more food to those
developing fruits that contain seeds that are more genetically variable (the result of a greater diversity of pollen donors) than to fruits that contain genetically more uniform seeds. Botanists K. Searcy and M. Mcnair found that monkeyflower plants (Mimulus guttatus) that grow in soil that contains mildly toxic levels of copper selectively abort seeds that had been fertilized by pollen from donors that were not resistant to copper. these may be examples of sexual selection in which the maternal plant selects among pollen sources.
● the endosperm is the food supply for the embryo inside the seed. it is triploid, hav-ing formed from two sets of chromosomes from the plant that produces it but only one set from the pollen. Several researchers, such as botanist Mark Westoby, have inter-preted this as sexual selection in which the maternal plant gains more control than the paternal plant in controlling the growth of the embryo.
therefore, while natural selection explains the diversity of adaptations that allow organisms to sur-vive and reproduce in different environments, and in response to other species, it is sexual selection that primarily explains much of the seemingly arbitrary characteristics of animals and perhaps many charac-teristics of plants as well. natural selection has made the world of organisms efficient; sexual selection has made it wildly beautiful and extremely diverse.
See also coevolution; life history, diver-sity of; natural selection.
Further readingalbert, arianne y. K., and Sarah P. Otto. “Sexual selection
can resolve sex-linked sexual antagonism.” Science 310 (2005): 119–121.
alcock, John. An Enthusiasm for Orchids: Sex and Decep-tion in Plant Evolution. new york: Oxford University Press, 2005.
andersson, Malte. “evolution of classical polyandry: three steps to female emancipation.” Ethology 111 (2005): 1–23.
———, and leigh W. Simmons. “Sexual selection and mate choice.” Trends in Ecology and Evolution 21 (2006): 296–302.
Bernasconi, G., et al. “evolutionary ecology of the prezy-gotic stage.” Science 303 (2004): 971–974.
Bjork, adam, and Scott Pitnick. “intensity of sexual selec-tion along the anisogamy-isogamy continuum.” Nature 441 (2006): 742–745.
chaine, alexis S., and Bruce e. lyon. “adaptive plastic-ity in female mate choice dampens sexual selection
xviii+598_Biodiversity_F.indd 484 11/17/11 1:13 PM

sexual selection 485
on male ornaments in the lark bunting.” Science 318 (2008): 459–462.
crump, Marty. Headless Males Make Great Lovers: And Other Unusual Natural Histories. chicago: University of chicago Press, 2007.
cuervo, J. J., and R. M. De ayala. “experimental tail shortening in barn swallows (Hirundo rustica) affects haematocrit.” Functional Ecology 19 (2005): 828–835.
Darwin, charles. The Descent of Man, and Selection in Relation to Sex. london: John Murray, 1871. Reprint, with introduction by James Moore and adrian Des-mond, new york: Penguin classics, 2004.
Dean, Rebecca, Michael B. Bonsall, and tommaso Piz-zari. “aging and sexual conflict.” Science 316 (2007): 383–384.
DelBarco-trillo, Javier, and Michael H. Ferkin. “Male mammals respond to a risk of sperm competition conveyed by odours of conspecific males.” Nature 431 (2004): 446–449.
Diamond, Jared. Why Is Sex Fun? new york: Basic Books, 1997.
Gomendio, Montserrat, et al. “Male fertility and sex ratio of birth in red deer.” Science 314 (2006): 1,445–1,447.
Haig, David, and Mark Westoby. “Parent-specific gene expression and the triploid endosperm.” American Naturalist 134 (1989): 147–155.
———. “Seed size, pollination costs and angiosperm suc-cess.” Evolutionary Ecology 5 (1991): 231–247.
Hamilton, William D., and Marlene Zuk. “Heritable true fitness and bright birds: a role for parasites?” Science 218 (1982): 384–387.
Heinsohn, Robert, Sarah legge, and John a. endler. “extreme reversed sexual dichromatism in a bird without sex role reversal.” Science 309 (2005): 617–619.
Holland, Jennifer S. “Feathers of seduction.” National Geographic, July 2007, 82–101.
Judson, Olivia. Dr. Tatiana’s Sex Advice to All Creation. new york: Henry Holt, 2002.
Kappeler, Peter M., and carel P. van Schaik. Sexual Selec-tion in Primates: New and Comparative Perspec-tives. cambridge, U.K.: cambridge University Press, 2004.
lehrer, Jonah. “the gay animal kingdom: the effeminate sheep and other problems with Darwinian sexual selection.” available online. URl: http://seedmagazine.com/news/2006/06/the_gay_animal_kingdom.php. accessed august 3, 2010.
Marshall, Diane l., and M. W. Folsom. “Mate choice in plants: an anatomical to population perspective.” Annual Review of Ecology and Systematics 22 (1991): 37–63.
———, and Pamela K. Diggle. “Mechanisms of differ-ential pollen donor performance in the wild radish,
Raphanus sativus (Brassicaceae).” American Journal of Botany 88 (2001): 242–257.
Miller, Geoffrey. The Mating Mind: How Sexual Choice Shaped the Evolution of Human Nature. new york: Doubleday, 2000.
Miller, Paige M., et al. “Sexual conflict via maternal-effect genes in ZW species.” Science 312 (2006): 73.
nielsen, M. G., and W. B. Watt. “interference competi-tion and sexual selection promote polymorphism in Colias (lepidoptera, Pieridae).” Functional Ecology 14 (2000): 718–730.
Pomiankowski, a., and y. iwasa. “Runaway ornament diversity caused by Fisherian sexual selection.” Pro-ceedings of the National Academy of Sciences USA 95 (1998): 5,106–5,111.
Reed, Wendy. “Physiological effects on demography: a long-term experimental study of testosterone’s effects on fitness.” American Naturalist 167 (2006): 667–683.
Roughgarden, Joan. Evolution’s Rainbow: Diversity, Gen-der, and Sexuality in Nature and People. Berkeley: University of california Press, 2004.
Searcy, K. B., and M. R. Mcnair. “Developmental selec-tion in response to environmental conditions of the maternal parent in Mimulus guttatus.” Evolution 47 (1993): 13–24.
Snow, allison a. “Postpollination selection and male fit-ness in plants.” American Naturalist 144 (1994): 569–583.
———, and timothy Spira. “Pollen-tube competition and male fitness in Hibiscus moscheutos.” Evolution 60 (1996): 1,866–1,870.
trivers, Robert. “Parental investment and sexual selec-tion.” in Sexual Selection and the Descent of Man, edited by B. campbell, 136–179. chicago: aldine, 1972.
Van Oort, H., and R. D. Dawson. “carotenoid ornamenta-tion of adult male common redpolls predicts probabil-ity of dying in a salmonellosis outbreak.” Functional Ecology 19 (2005): 822–827.
West, Peyton M. “the lion’s mane.” American Scientist 93 (2005): 226–235.
Williamson, K. a., et al. “yolk antioxidants and mate attractiveness in the zebra finch.” Functional Ecology 20 (2006): 354–359.
Willson, Mary F., and nancy Burley. Mate Choice in Plants: Tactics, Mechanisms, and Consequences. Princeton, n.J.: Princeton University Press, 1983.
Zahavi, amotz. The Handicap Principle: A Missing Piece of Darwin’s Puzzle. new york: Oxford University Press, 1997.
Zimmer, carl. “in ducks, war of the sexes plays out in evolution of genitalia.” New York Times (May 1, 2007). available online. URl: http://www.nytimes.com/2007/04/30/science/01duck.html?pagewanted=all. accessed May 16, 2011.
xviii+598_Biodiversity_F.indd 485 11/17/11 1:13 PM

first entry486 shrublands 486
shrublands the shrubland ecological commu-nity consists of dense shrubs from about a foot to about nine feet (about one-half meter to 3 m) in height and the other species living in association with them. Shrubs, rather than trees or grasses, dominate these communities, usually for the follow-ing reasons:
● Low rainfall. Shrublands typically receive little rainfall (and only very rarely snow), and most of this occurs in the winter. Domi-nant shrubs have adaptations such as small, thick leaves that allow them to survive the long, dry summers. One of the reasons that most of the shrubs are evergreen is that most of the rain falls during mild winters.
● Poor soil. Shrubland soils are typically very poor and often very rocky. in some shrub-lands, the soil is acidic. another reason that most of the shrubs are evergreen is that the soil is poor, and the shrubs retain their leaves because of the nutrients in them.
● Fire. Fires occur in shrublands every few decades because the shrubs grow densely and because of summer drought. the accu-mulation of dry dead leaves promotes the chances of fire, especially during windy con-ditions. Many shrub and wildflower species cannot persist in the shrublands unless a fire has burned away the shrub cover, at which time many seeds germinate and some of the shrubs resprout. Fires also convert plant material into ash, which fertilizes the poor soil.
Some shrublands have ecological conditions different from these. For example, the moorlands of england are wet and may even have layers of peat moss. artificial shrublands are found in many areas scattered around the world. Humans cleared forests thousands of years ago and have kept the forest trees out by fire and by continued clearing in these areas.
Chaparral and related shrublands. the chap-arral community dominates some areas with a Mediterranean climate of mild, wet winters and hot, dry summers. this kind of climate occurs in california, where most of the chaparral is located; in the Mediterranean Basin, where the shrublands are called maquia or garrigue; in central chile and central Mexico, where they are called matorral; in southwestern australia, where they are called mallee; and in the cape region of South africa, where they are called fynbos. the shrubs, often about six feet (2 m) tall, are very dense and all but
some important dominant speCies in shrublands
Chaparral (California)
Bush monkeyflower Mimulus aurantiacus
Chamise Adenostoma fasciculatum
Holly-leaf cherry Prunus ilicifolia
Large-fruited ceanothus Ceanothus megacarpus
Laurel sumac Malosma (Rhus) laurina
Pointleaf manzanita Arctostaphylos pungens
Scrub oak Quercus dumosa
Toyon Heteromeles arbutifolia
Maquia and Garrigue (Mediterranean)
Holm oak Quercus ilex
Kermes oak Quercus coccifera
Myrtle Myrtus communis
Strawberry tree Arbutus unedo
Tree heath Erica arborea
Matorral (Mexico)
Desert ceanothus Ceanothus greggii
Pointleaf manzanita Arctostaphylos pungens
Scrub oak Quercus sebifera
Texas madrone Arbutus xalapensis
Matorral (Chile)
Coastal daisy Bahia ambrosioides
Colliguay Colliguaya odorifera
Soapbark tree Quillaja saponaria
Litre Lithraea caustica
Fynbos (South Africa)
Cape Flats erica Erica verticillata
Broad-leaved sugarbush Protea eximia
Laurel protea Protea laurifolia
Renosterbos Elytropappus rhinocerotis
Sunshine conebush Leucadendron saligna
Mallee (Southwestern Australia)
Firewood Banksia Banksia menziesii
Golden Dryandra Banksia nobilis
xviii+598_Biodiversity_F.indd 486 11/17/11 1:13 PM

shrublands 487
impenetrable to humans and large animals. chap-arral and similar ecological communities usually have many dominant shrub species, which contrib-ute to the hot spot diversity in their geographical areas.
Due to massive buildup of combustible leaf litter and dead branches, fires become nearly inevi-table in the chaparral after a few decades. Without fire, the chaparral would probably become temper-ate evergreen forest, for example, of evergreen oaks in california or the Mediterranean or of eucalypts in australia. (Oaks and eucalypts also resprout or germinate after fires.) With excessive fire, the chap-arral would probably become grassland. Because the thick branches create a very hot fire, because the fires spread rapidly, and because they are inevi-table, people who build homes in chaparral regions need to be aware of the risk they are taking—more of a fire risk than in perhaps any other natural habi-tat on earth.
Some wildflower species are rare except imme-diately after a chaparral fire. examples from cali-fornia include the giant-flowered phacelia (Phacelia grandiflora) and the fire poppy (Papaver californi-cum). Other wildflowers, such as the ground pink (Linanthus dianthiflorus) and the california poppy (Eschscholzia californica), grow in disturbed areas and, for this reason, are abundant after chaparral burns.
the northern Hemisphere shrub communi-ties share some genera of dominant plants, most notably scrub oaks (Quercus). Both california and Mexico share the manzanitas (Arctostaphylos) and Ceanothus bushes. a completely different set of plants dominates the Southern Hemisphere shrub-lands (see table).
Mediterranean sage scrub. Short bushes dom-inate sage scrublands. in california and in the Mediterranean Basin, many of these shrubs are in the mint family (lamiaceae) and have strongly scented leaves. One role that the volatile chemicals from these leaves may play is allelopathy, that is, the chemicals inhibit the growth of other species of plants.
Heather. the heather of england and Scotland are dominated by a relatively small number of shrub species in the genus Erica and Calluna. thin acidic soil covers chalk (basic) soil in these areas, where shallow-rooted species tolerate acid, deeper rooted ones do not, and some species tolerate both acidic and basic conditions. it is likely that the heather is an artificial shrubland that resulted from the prehistoric clearing of forests.
See also competition, ecological; distur-bances, ecological.
Further readingarmstrong, Wayne P. “ashes to Wildflowers: a Promise of
Renewal Springs from Destruction.” available online. URl: http://waynesword.palomar.edu/ww0604.htm. accessed July 8, 2010.
arroyo, J., and t. Marañón. “community ecology and dis-tributional spectra of Mediterranean shrublands and heathlands in southern Spain.” Journal of Biogeogra-phy 17 (1990): 163–176.
california chaparral institute. “california chaparral insti-tute.” available online. URl: http://www.california chaparral.org. accessed July 8, 2010.
christensen, norman l., and cornelius H. Muller. “effects of fire on factors controlling plant growth in Adenos-toma chaparral.” Ecological Monographs 45 (1975): 29–55.
Holmgren, Milena, et al. “Why have european herbs so successfully invaded the chilean matorral? effects of herbivory, soil nutrients, and fire.” Journal of Arid Environments 44 (2000): 197–211.
Jaksić, F. M., and e. R. Fuentes. “Why are native herbs in the chilean matorral more abundant beneath bushes: Microclimate or grazing?” Journal of Ecology 68 (1980): 665–669.
Jiménez, Héctor e., and Juan J. armesto. “importance of the soil seed bank of disturbed sites in chilean mator-ral in early secondary succession.” Journal of Vegeta-tion Science 3 (1992): 579–586.
Prickly toothbrush Grevillea armigera
Quandong Santalum acuminatum
Sage Scrub (California)
Black sage Salvia mellifera
California buckwheat Eriogonum fasciculatum
California sagebrush Artemisia californica
White sage Salvia leucophylla
Sage Scrub (Mediterranean)
Common sage Salvia officinalis
Lavender Lavandula spica
Rosemary Rosmarinus officinalis
Thyme Thymus vulgaris
Heather (Britain)
Bell heather Erica cinerea
Ling heather Calluna vulgaris
xviii+598_Biodiversity_F.indd 487 11/17/11 1:13 PM

first entry488 Silurian period, biodiversity in 488
Keeley, Jon e., et al. “Role of allelopathy, heat, and charred wood in the germination of chaparral herbs and suf-frutescents.” Journal of Ecology 73 (1985): 445–458.
———, et al. “Reexamining fire suppression impacts on brushland fire regimes.” Science 284 (1999): 1,829–1,832.
———, et al. “Fire suppression impacts on postfire recov-ery of Sierra nevada chaparral shrublands.” Interna-tional Journal of Wildland Fire 14 (2005): 255–265.
lavorel, Sandra. “ecological diversity and resilience of Mediterranean vegetation to disturbance.” Diversity and Distributions 5 (1999): 3–13.
Minnich, R. a. “Fire mosaics in southern california and northern Baja california.” Science 219 (1983): 1,287–1,294.
Moritz, M. a., et al. “testing a basic assumption of shrubland fire management: How important is fuel age?” Frontiers in Ecology and the Environment 2 (2004): 67–72.
Muller, cornelius H., Walter Muller, and Bruce Haines. “Volatile growth inhibitors produced by aromatic shrubs.” Science 143 (1966): 471–473.
Valiente-Banuet, alfonso, et al. “the chaparral vegeta-tion in Mexico under non-Mediterranean climate: the convergence and Madrean-tethyan hypotheses reconsidered.” American Journal of Botany 85 (1998): 1,398–1,408.
silurian period, biodiversity in the Silu-rian period (440 to 410 million years ago) was the third period of the Paleozoic era. it followed the Ordovician period and preceded the Devonian period. an average of about 450 families of marine organisms and 50 families of continental organisms existed during the Ordovician, compared to 2,200 total families today.
Climate. the climate of the continents was generally warm and moist, except the landmasses directly over the South Pole, during the Silurian period. Some southern glaciers from the Ordovician period melted and raised the sea level. atmospheric oxygen levels were much lower, and carbon dioxide levels much higher, than today. Global temperatures were also higher than today (see table on page 489).
Continents. as in the preceding and following periods, most of the landmass was near the South Pole (Gondwana). Other landmasses that today constitute eurasia, Siberia, and china were near the equator. Sea level was much higher than today—just under 600 feet (about 180 m) higher than modern oceans.
Marine life. During the Silurian period, aquatic animals continued expanding from shallow marine waters into deeper waters and into freshwater.
● Invertebrates. the trilobites continued to evolve new forms as they declined in diver-
sity. among the earliest crustaceans from this period was Ceratiocaris. Mollusks continued to be abundant and diverse in shallow seas, including bivalves. However, brachiopods (which superficially resemble bivalves) were much more common than bivalves. among the echinoderms were the stalked crinoids such as Ampheristocystus.
● Vertebrates. the only vertebrates were fishes, jawless descendants of the early fishes of the Ordovician period. armored jawless fishes were common. the first jawed fishes evolved at the end of the period but were rare.
Life on land. the earliest complete specimens of continental organisms date from the Silurian period.
● Plants. the first complete specimens of land plants were small vascular plants, such as Cooksonia, that resembled modern club mosses. they grew only in freshwater wet-lands, extending a few inches into the air rather than being completely underwater. But they had the plumbing cells (xylem) that would later allow plants to live on dry land. the most enigmatic fossil of the Silurian period is Prototaxites. it was originally con-sidered to be a vascular plant that towered over 25 feet (8 m), high above the other plants. then it was suggested, based on a chemical analysis of the fossils, that Proto-taxites was actually a giant fungus. Most recently, botanist linda Graham and asso-ciates have questioned whether it was an organism that stood upright at all. they claim that these fossils may be rolled-up mats of liverworts!
● Invertebrates. the first terrestrial animals were relatives of modern millipedes, scor-pions, and insectlike collembolans. these arthropods fed mostly on one another or on detritus. the plants contained lignins and toxins; it is likely that the animals could not digest the plant material until it had been softened and detoxified by fungi. While today the food chain is based largely upon the consumption of fresh green plant tis-sue, the Silurian continental food chain may have depended only indirectly on plants.
● Vertebrates. the first bony fishes evolved during the Silurian period. little is known about Silurian bony fishes. the earliest bony fishes may have evolved in freshwater. Bones may have served primarily to store calcium, of which freshwater can be deficient.
xviii+598_Biodiversity_F.indd 488 11/17/11 1:13 PM

speciation 489
Extinctions. the Silurian period began after the Ordovician extinction, which was one of the five mass extinctions in the history of the earth.
See also Devonian period, biodiversity in; Ordovician period, biodiversity in.
Further readingBenton, Michael. “Diversification and extinction in the his-
tory of life.” Science 268 (1995): 52–58.Botella, Hector, et al. “Jaws and teeth of the earliest bony
fishes.” Nature 448 (2007): 583–586.ericksson, Mats e., et al. “Vertebrate extinctions and reor-
ganizations during the late Silurian lau event.” Geol-ogy 37 (2009): 739–742.
Graham, linda e., et al. “Structural, physiological, and stable carbon isotopic evidence that the enigmatic Paleozoic fossil Prototaxites formed from rolled liverwort mats.” American Journal of Botany 97 (2010): 268–275.
Jeram, andrew J., et al. “land animals in the Silurian: arachnids and myriapods from Shropshire, england.” Science 250 (1990): 658–661.
speciation Speciation occurs when two or more species diverge from a single ancestral population. this process has produced all of the species-level biodiversity in the world. natural selection and sexual selection can produce evolutionary change within lineages of organisms over time. However, as many writers have pointed out, charles Darwin’s book Origin of Species did not actually explain how two or more species could evolve from one species but only how one species could change into another. Something more is needed, besides natural and sexual selection, to explain speciation. a certain amount of reproductive isolation is necessary. if one population of organisms is separated into two populations, the two groups cannot interbreed. this could result from geographical isolation, in which the two populations are isolated from one another by impassible barriers such as deserts or oceans. Or it could result from characteristics of the organisms themselves: Isolating mechanisms are negative pro-cesses that prevent interbreeding between groups, and specific mate recognition systems are positive processes that encourage breeding within groups.
Once even partial reproductive isolation occurs, the genes of the two populations no longer mix completely together. the two populations may even-tually become two species because new genetic com-binations, and new mutations, that occur in one population will be different from those in the other, and the two populations could then diverge along different evolutionary lines. Divergence may then occur:
● natural selection may favor different char-acteristics in each group strongly enough to compensate for interchange of genes (gene flow) between the populations. after the populations have begun to diverge, natural selection may favor isolating mechanisms
multiCellular biodiversity in the silurian period
Conditions
Atmospheric oxygen 70% of modern level
Atmospheric carbon dioxide
12.9 times modern level
Global temperature 3°F (2°C) higher than today
Sea level 580 feet (180 m) higher than today
Approximate number of families*
Continental 50
Marine 450
Total 500
Examples of marine animals
Amperistocystus sp. crinoid echinoderm
Ceratiocaris sp. crustacean
Coelocaulus sp. gastropod mollusk
Dalmanites limuliroides trilobite arthropod
Doryapsis nathorsti armored jawless fish
Eurypteris sp. sea scorpion
Gavosites gothlandicus coral
Pentameroides subrectus brachiopod
Phragmoceras exag-geratus
cephalopod mollusk
Examples of continental animals
Andreolepis hedei bony fish
Pneumodesmus newmani millipede
Proscorpius sp. scorpion
Rhyniognatha hirsti insect relative
Example of land plant and fungus
Cooksonia pertoni club moss relative
Prototaxites loganii fungus**
*Based on Benton.**Identity as fungus now disputed.
xviii+598_Biodiversity_F.indd 489 11/17/11 1:13 PM

first entry490 speciation 490
and specific mate recognition systems that complete the process of speciation.
● Different and random genetic changes may occur in each group. even if natural selec-tion favors the same characteristics in both populations, they will probably diverge because they will not have the same genes upon which natural selection can act.
evolutionary biologists distinguish species on the basis of either a biological or a phylogenetic species concept. the biological species concept recognizes the potential to interbreed under natu-ral conditions as the definition of species member-ship. even though this concept has some difficulties (hybridization between recognized species occurs frequently, and investigators cannot know the inter-breeding potential of geographically isolated popu-lations or fossil species), it is the concept that most closely reflects the evolutionary process itself. the phylogenetic species concept recognizes that species are different if they have evolved along distinct evo-lutionary lines.
natural or sexual selection may occur rapidly after reproductive isolation, as the incipient species adjusts by directional selection to the new environ-mental conditions or the new set of species with which it is in contact. this may be followed by a long period of stabilizing selection. this has occurred fre-quently enough, according to many paleontologists, to produce a pattern of punctuated equilibria in the fossil record, in which long periods of equilibrium are punctuated by brief spurts of speciation.
paTTErNS of SpECIaTIoNSpeciation can occur in different patterns, as described in the following sections.
Allopatric speciation. allopatric speciation occurs when populations are geographically or eco-logically isolated. this has occurred frequently on islands, where dispersal of plants and animals from the mainland is rarely successful, and once it occurs, the island population is unlikely to disperse back to the mainland. this is the process that has pro-duced the vast number of unique endemic species found only on certain islands. the presence of many endemic species on islands—most notably the Galá-pagos islands—was one of Darwin’s chief evidences that evolution has, in fact, occurred. Most species of nettles are small, stinging herbs, but in Hawaii, a stingless nettle bush has evolved, in isolation from mainland nettles. Hawaii has or had many species of birds, snails, insects, plants, and many other spe-cies found nowhere else in the world. not surpris-ingly, Hawaii has the longest list of threatened and endangered species of any state in the United States.
Given long enough time, the geographic separation of populations almost inevitably produces new spe-cies through allopatric speciation.
Peripatric speciation. Peripatric speciation is similar to allopatric speciation but involves the geo-graphical isolation of small populations from the main population. Small populations can be impor-tant in speciation, since the genetic makeup of small populations can change more rapidly than that of large populations. a classic experimental demon-stration of rapid evolution in small populations is the study by theodosius Dobzhansky of evolution-ary change in fruit flies over the course of a year and a half. Dobzhansky established 20 large and 20 small populations of fruit flies, each with a cer-tain chromosome at 50 percent frequency. in most populations, this chromosome became less frequent. in all of the large populations, the chromosome became less frequent to about the same extent; they all evolved similarly. in the small populations, some hardly evolved at all, and some evolved a great deal; the small populations had a much greater range of evolutionary outcomes than the large populations.
Parapatric speciation. a large population may evolve into two or more species by specialization on different habitats within the geographical range of the ancestral species.
Sympatric speciation. Reproductive isolation can also occur between two populations that live in the same location. examples include:
● if two species of plants open their flowers in different seasons (for example, the autumn-flowering v. the spring-flowering species of alder trees of the genus Alnus), they can-not interbreed, even if they live in the same habitat.
● if two species of plants depend upon dif-ferent pollinators (such as the humming-bird-pollinated v. the bumblebee-pollinated monkeyflowers of the genus Mimulus), they will not interbreed, even if they are right next to one another (see below). Genera of plants with bilaterally symmetrical flowers tend to have more species than genera that are otherwise similar but have radially sym-metrical flowers, perhaps because bilaterally symmetrical flowers often have more spe-cialized relationships with pollinators.
● if two species of animals specialize on dif-ferent food plants (especially if they live on and mate on these food plants as well), they will not interbreed very often, even if their respective food plants are right next to one another. it has been reported that some european corn borer larvae now consume
xviii+598_Biodiversity_F.indd 490 11/17/11 1:13 PM

speciation 491
hops. this has resulted in adult insects that emit different mating pheromones. adult borers whose larvae ate hops preferentially mate with other such adults, rather than adults whose larvae ate corn.
● if two species of animals have different mat-ing calls or rituals, they will not interbreed very often, even if they are in contact. this has been observed with Darwin’s finches on the Galápagos islands (see below). indi-gobirds are nest parasites, which means that they lay their eggs in the nests of other bird species. cladistic analysis indicates that indigobirds evolved into several species more recently than did the host species. this apparently happened because the young indigobirds learned songs and habits from their hosts, and when they grew up, they tended to mate with other indigobirds that had similar songs and habits and to parasit-ize the nests of the same species of birds as their parents.
● if two species of plants can cross-pollinate, but the pollen grains do not successfully fertilize the egg nuclei—or if two animal species can mate with one another but the sperm from one population has inferior per-formance in the female reproductive tracts of the other—then the proximity of the two populations does not matter.
● in some cases, symbiogenesis may cause reproductive isolation if one population that consists of individuals in which a mutualis-tic microbe is living cannot crossbreed with a population whose individuals lack the microbe.
isolating mechanisms (see below) can sepa-rate two populations and allow them to evolve into two species. the two populations might as well be separated by an ocean. Given long enough time, the reproductive separation of populations almost inevitably produces new species through sympatric speciation.
Speciation by hybridization. Hybrids between species may result in offspring with incompatible chromosomes. if the chromosomes within the cells of the offspring are unable to match up with one another during meiosis, the offspring are sterile. among animals, hybrids are generally the end of the evolutionary line. a strong mule, for example, cannot produce offspring. Many species of animals hybridize, but the hybrid offspring are often inferior in survival and reproduction. nevertheless, an occa-sional hybridization can be an important source of new genes for a species. the gene for organophos-
phate insecticide resistance originated in the mos-quito species Culex pipiens, but it spread worldwide, even across mosquito species boundaries, probably because of rare but important hybridizations.
However, among plants, sterile individuals pro-duced by hybridization may have options that are not available to animals. Many plants reproduce without sex, a process called apomixis. the embryo produced inside the seed is an exact genetic copy of the parent, and the plant that grows from it is a clone of the parent. an example is the wildflower Oxalis pes-caprae in the shamrock family. Many individuals in this species have chromosomes in groups of five rather than in groups of two; they pro-duce beautiful flowers, but because meiosis cannot handle unpaired chromosomes, the flowers cannot produce either pollen or egg nuclei. they reproduce only by making underground bulbs, by which they survive the dry season. each lineage of these plants is reproductively isolated from all the rest. Muta-tions may occur, and if the mutations are passed on through the bulbs, each lineage can evolve into what looks like a different species. in their native South africa, there are a few Oxalis pes-caprae with evenly paired chromosomes that can reproduce sexually, but they are reproductively isolated from the sterile Oxalis individuals. to call each sterile lineage of Oxalis a separate species would be mad-deningly complex, and yet each sterile lineage does function as if it were a separate species. Dandeli-ons (Taraxacum officinale) have chromosomes in groups of three and are also unable to reproduce sexually. they produce seeds, but the seeds contain copies of the parental genes. Pollination is necessary to stimulate seed production but does not mix the genes of pollen and egg nuclei. each clump of dan-delion, then, functions as if it were a separate spe-cies. a single population of dandelions can contain dozens of distinct genetic lineages, which may have arisen by the multiple evolution of apomixis, or by somatic mutation, or both.
apomixis is not the only way that evolution can allow sterile plants to reproduce. When the repro-ductive cells of animals experience spontaneous chromosome doubling, the cell usually dies. Most animals tolerate only the most minor changes in the number of chromosomes; even then, as in Down syndrome (trisomy of chromosome 21) in humans, a single extra chromosome results in sterility and retardation. Plants, in contrast, frequently undergo spontaneous doubling in the number of chromo-somes. a plant egg cell with doubled chromosomes may survive. a sexually sterile plant, if it undergoes chromosome doubling, may produce offspring that are not sterile, because the doubled chromosomes now form pairs, which allows meiosis to occur. each
xviii+598_Biodiversity_F.indd 491 11/17/11 1:13 PM

first entry492 speciation 492
chromosome can now match up with a copy of itself. these plants can now reproduce sexually. the popu-lation of plants with doubled chromosomes, how-ever, may not be able to interbreed with either of the ancestral populations. the hybrid is then instantly isolated from both parents and has characteristics different from both; it becomes an “instant species.” new species have arisen by hybridization in the goatsbeard Tragopogon and in the sunflower Heli-anthus. this form of speciation is common in the plant kingdom. the study of chromosomes suggests that almost half of all plant species, and 95 percent of fern species, originated by chromosome doubling following hybridization.
ISolaTING mEChaNISmSisolating mechanisms are biological processes, as opposed to geographical features, that keep popula-tions apart and allow them to evolve into different species. they are particularly important in sym-patric speciation (see above). Prezygotic isolating mechanisms prevent interbreeding from occurring in the first place, that is, the zygote (fertilized egg) does not form. Postzygotic isolating mechanisms operate after the formation of the zygote.
Prezygotic isolating mechanisms include the fol-lowing examples:
Differences in pollination mechanisms. if a mutation occurs in a population of flowering plants that changes the characteristics of the flowers, the new kind of flower may not be recognized by the same pollinator that visits the old kind of flower.
this is apparently what happened in califor-nia populations of monkeyflowers (genus Mimulus). Most species of Mimulus, including M. lewisii, have yellow flowers, which attract bees. Mimulus cardina-lis, however, has red flowers, which attract humming-birds. all of the monkeyflowers have the ability to produce red carotenoids, but in most of them a gene prevents carotenoid production in the flowers (except to form isolated spots, not enough to attract hum-mingbirds). a simple mutation occurred, causing a sudden surge in red carotenoid production. the flow-ers that had this mutation, though side by side with the yellow flowers, were reproductively isolated from them, because bees (which cannot see red) ignored them, and hummingbirds visited them instead.
Subsequent to the reproductive isolation, fur-ther evolutionary changes occurred in the red- and yellow-flowered species. For example, the popula-tion of red flowers evolved a long, narrow tubular shape that was appropriate for hummingbirds, while the yellow flowers retained a short, wide tube that was appropriate for bees. the changes in flower shape reinforced the initial reproductive isolation that had occurred because of a change in flower
color. While the change in flower color was sufficient to separate one species into two, natural selection subsequently favored changes in flower shape that enhanced the success of pollination in each species separately. What was once one species of monkey-flower evolved into two, side by side.
although nobody was there to observe the origi-nal formation of the red-flowered species, evolution-ary biologists Douglas Schemske and toby Bradshaw have experimentally re-created the sequence of events. they crossed the two monkeyflower spe-cies and produced a whole range of hybrids with intermediate shapes and colors. they found that the Dna sequences that enhanced carotenoid produc-tion, not the genes that controlled the shapes of the flowers, had an immediate and strong effect on pol-linator preference.
another example of a change in pollination mechanism involves Schiedea salicaria, a small shrub that is native to Hawaii, investigated by evolutionary biologists ann Sakai and Steve Weller. a few relatively minor genetic mutations appear to have caused some individuals in the population to rely more on pollination by wind and less on pol-lination by insects. these two groups can cross-pollinate, but because they rely largely on different methods of pollination, they do not frequently do so. the individuals that have greater wind-pollina-tion characteristics tend to live in drier locations, where insect pollinators are less common. though not completely isolated from one another, these two groups of plants have apparently started on the road to separation, perhaps eventually into two different species.
natural selection often reinforces the depen-dence on different methods of pollination. For exam-ple, flowers that have characteristics intermediate in their appeal to hummingbirds and to bumblebees will probably not be pollinated very well by either hummingbirds or bumblebees.
Differences in timing of reproduction. each species has its own reproductive season. Mutations that cause some individuals in a population to have a different reproductive season will prevent those individuals from interbreeding with others. One example is the alder trees (genus Alnus) mentioned above. early in their evolution, alder populations diverged into some that pollinated in the autumn (subgenus Clethropsis) and those that pollinated in the spring (subgenus Alnus). these subgenera can no longer interbreed. another example is the flies that live on apple and hawthorn trees. Rhagole-tis pomonella flies, native to north america, lay their eggs on the fruits of hawthorns, a wild bush that produces fruits that resemble small crabapples. When europeans and white americans began plant-
xviii+598_Biodiversity_F.indd 492 11/17/11 1:13 PM

speciation 493
ing apples in eastern north america, some popula-tions of this fly species began to lay their eggs in apples. the populations of flies that lived in apple trees could potentially interbreed with the popula-tions that lived on hawthorns but seldom did so, even if wild hawthorns and cultivated apples were side by side. an immediate reproductive isolation occurred because apple fruits mature three to four weeks earlier than those of hawthorns, with the result that the flies that hatch from apples are ready for reproduction weeks earlier than those that hatch from hawthorns.
Differences in animal mating behavior. animal populations may be reproductively isolated because of differences in mating rituals. evolutionary biolo-gists Peter and Rosemary Grant have extensively studied the beak sizes and mating preferences of several species of Darwin’s finches on some uninhab-ited islands in the Galápagos archipelago. they have found that hybrids between finch species can occur, but occur rarely. young male birds learn their mat-ing songs from their fathers, and females use male mating songs to guide their choice of mates. Occa-sionally, a young male bird overhears and learns the song of a different species of finch nearby. When the young male bird grows up and sings, he attracts females of the wrong species.
Hawaii has hundreds of species of fruit flies (genus Drosophila), all of them unique to the Hawai-ian islands, and all of them having recently evolved there (the islands are from 5 to one-half million years old). there may be scores of species living on a single island within the archipelago and close enough to one another to readily interbreed. they do not interbreed, however, because they have evolved different mating dances. the females of one spe-cies will not recognize the males of another species as being potential mates. laboratory investigations of fruit flies (some species of which are the most intensively studied animals in the history of biol-ogy) indicate that small genetic mutations can cause major changes in mating behavior. therefore, the reproductive isolation between two groups of flies may have occurred quickly after a relatively minor genetic change.
Subsequent to the reproductive isolation caused by differences in mating dances in the Hawaiian flies, the populations evolved yet other differences, making them into recognizably different species. the species now differ in the anatomy of mating and reproductive anatomy, and other characteristics that make them look different to one another (for example, one species has its eyes on long stalks). these anatomical differences between species may or may not have anything to do with adaptation to their environments.
Specialization on different food resources. Spe-cialization of animals on different food resources will result in reproductive isolation only if individ-uals cross-breed primarily with other individuals that have the same specialization. the most famous examples of this are of insect populations within the same species that specialize on new food plants. they not only eat the plants but live and mate upon them, isolated from the insects on a different kind of food plant. a major example of this is the Rhagoletis flies that lay eggs on apples and hawthorns, as described previously. after a couple of centuries of isolation, the apple flies and the hawthorn flies will no longer interbreed, even when given the opportunity to do so in the laboratory. this process was observed by natu-ralist Benjamin Walsh, an american acquaintance of charles Darwin, in the 19th century, and was more fully investigated in the 20th century.
another example of reproductive isolation occurring on newly introduced plant species involves the soapberry bug. Soapberry bugs have snouts about 0.36 inch (9 mm) in length that penetrate the fruit and consume contents from the seeds of soapberry trees, native to the southern United States. However, in southern Florida, golden rain trees have been imported from asia and used as ornamentals, in the vicinity of soapberry trees. Golden rain tree fruits are smaller, thus a smaller snout is needed to penetrate it and get to the seed. Some populations of soapberry bugs have specialized on golden rain tree and seldom interbreed with the populations that still live on soapberry trees. the populations of bugs living on the golden rain trees have begun to evolve shorter beaks, about 0.28 inch (7 mm) in length. this process occurred recently, as golden rain trees were not extensively planted in Florida until the 1950s. in another example, fruit flies that resulted from the hybridization of a species of fly that lives and reproduces on blueberries and a species that lives and reproduces on snowberries have become reproductively isolated from both parental species by living on Japanese honeysuckle, which has been in north america for less than three centuries.
Sometimes the isolation of populations on dif-ferent food resources can result from just a small number of genetic changes. two species of flies in the Seychelle archipelago of the indian Ocean are Dro-sophila simulans and Drosophila sechellia. the first species avoids the poisonous morinda fruit, while the second species is attracted to it, for laying their eggs. Researchers found that the species differed by only a small number of genes—just enough to pro-duce tolerance of and attraction to morinda fruits in D. sechellia.
examples are not, however, limited to insects that live on and eat plants. Marine stickleback fishes
xviii+598_Biodiversity_F.indd 493 11/17/11 1:13 PM

first entry494 speciation 494
invaded freshwater lakes of British columbia after the most recent of the ice ages. Some of them had a tendency, apparently genetically based, to eat small plankton near the surface of the lakes; oth-ers preferred eating material that had settled to the bottoms of the lakes. Males and females mated preferentially in the same locations where they ate, which set them on the road to becoming two sepa-rate populations. now, the surface- and the bottom-dwelling populations have begun to evolve unique adaptations to their new modes of life: the mouths of the bottom dwellers are larger and turned down-ward, those of the surface dwellers smaller and not downturned.
in each of these cases, natural selection has prob-ably favored the isolating mechanism. interbreeding between populations or species that specialize on different food resources would probably result in offspring that are inefficient at exploiting either of the parental food resources.
Specialization on different symbionts. Symbio-genesis is the origin of new species as a result of the symbiotic merger of two or more species. For example, a population of bacteria may take up resi-dence in a new kind of host cell, such as an amoeba, and reproduce inside that host cell. Whenever the amoeba reproduces, the bacteria are passed on to the new amoebae. the amoeba represents a new environment for this population of bacteria. this population is isolated from the bacteria from which it originally came and adapts to a new set of condi-tions. the amoebae, which have evolved a tolerance for the bacteria, may also be isolated from other amoebae if the other amoebae cannot tolerate the bacteria. By depending on each other, the amoebae and the bacteria are isolated from other amoebae and bacteria. the result can be a new species of amoeba and a new species of bacteria. in time, the bacteria may degenerate to such an extent that they are no longer recognizable as a distinct species. the first stages of this process were actually observed to occur in a laboratory setting.
Differential success at fertilization. even if individuals from two populations succeed in mat-ing, their reproduction may still not be successful if the male reproductive cells (sperm of animals, or pollen nuclei of plants) are inferior at fertilizing the female reproductive cells (eggs of animals, or ovules of plants). the sperm cells from one popula-tion may swim faster and suffer fewer deaths on their way to the egg than the sperm cells from another population; this will tend to isolate the two populations. in flowering plants, pollen grains land on the stigma of the flower, germinate, and grow a pollen tube down into the ovary of the flower. Some pollen grains are better at doing this than others.
Frequently, the stigma of a flower will not even permit the wrong kind of pollen—either pollen that is genetically identical to it, or pollen that comes from the wrong species—to germinate in the first place. the female reproductive tract of many ver-tebrates represents a hostile environment through which only the hardiest sperm can travel—that is, the sperm that are best adapted to those particular conditions.
as noted previously, the most famous of the postzygotic isolating mechanisms is the production of sterile hybrid offspring, as when horses and don-keys hybridize and produce sterile mules.
INvESTIGaTIoNS of SpECIaTIoNSpeciation has been investigated as it occurs, as described in the following sections.
Experimental speciation. Speciation is difficult to observe in the wild or to demonstrate experi-mentally. Perhaps the best example is from fruit flies of the genus Drosophila. Researchers can raise many generations of fruit flies under laboratory conditions. Beginning with a single population of flies, they separated them into two populations: one kept in warm conditions, the other kept in cool conditions. after many generations, the two popu-lations of flies had diverged enough that they did not interbreed when they were placed back together. this may have occurred partly from the gradual accumulation of genetic differences, but may also have involved symbiogenesis, since one of the popu-lations lost microbes that had lived in the digestive system of its ancestors. in another fruit fly experi-ment, researchers kept some flies in the dark and others in the light. not surprisingly, the flies in the dark lost the genetic ability to use visual cues for mating; they evolved mating cues based upon touch and vibration. the visual flies could not interbreed with the tactile flies.
Reconstructing speciation in the wild. in natu-ral populations where speciation may be impossible to observe in action, researchers have frequently found convincing evidence regarding how the spe-ciation occurred. evolutionary biologists Douglas Schemske and toby Bradshaw (see above) have found the trait that was almost certainly respon-sible for the evolution of two species of Mimulus from a single ancestor, and botanist loren Ries-eberg has identified the two parental species of sun-flowers whose chromosomes have come together to form the hybrid species Helianthus paradoxa. evolutionary biologists Peter and Rosemary Grant have carefully measured divergence in beak size in two groups of Darwin’s finches, which, if it contin-ues for a long time, may result in the formation of new species.
xviii+598_Biodiversity_F.indd 494 11/17/11 1:13 PM

speciation 495
Speciation occurring in the wild. in some cases, two incipient species (not quite yet differentiated enough to be considered separate species) can exist in adjacent habitats. if there is a series of such spe-cies, each one grading into the next, it is possible that the species at the two ends of the series cannot inter-breed. this would constitute a ring species. examples include gulls of the genus Larus across the northern continents, salamanders of the genus Ensatina in the mountains of california, and the greenish warbler Phylloscopus trochiloides around the Himalayas.
evolutionary scientists have found numerous examples of speciation that is occurring in wild populations (see table). evolution has produced far more species than any human observer would con-sider necessary. For example, there are well over 350,000 species of beetles. if evolution produced species only in response to environmental condi-tions, there would be far fewer species of organisms in the world: a few species of animals and plants best suited to moist, cool conditions, a few other species best suited to hot, dry conditions, etc. But speciation usually results when species evolve in response to one another after geographic and reproductive isola-tion has occurred in each.
See also adaptation; coevolution; Darwin’s finches; Galápagos islands; hybridization; island biogeography; natural selection; sex-ual selection; symbiogenesis.
Further readingabrahamson, Warren G., et al. “Gall flies, inquilines, and
goldenrods: a model for host-race formation and sympatric speciation.” American Zoologist 41 (2001): 928–938.
arnqvist, G., et al. “Sexual conflict promotes speciation in insects.” Proceedings of the National Academy of Sci-ences USA 97 (2000): 10,460–10,464.
Blondel, Jacques, et al. “Selection-based biodiversity at a small spatial scale in a low-dispersing insular bird.” Science 285 (1999): 1,399–1,402.
carroll, Scott P., et al. “Genetic differentiation of fitness-associated traits among rapidly evolving popula-tions of the soapberry bug.” Evolution 51 (1997): 1,182–1,188.
condon, Marty a., et al. “Hidden neotropical diversity: Greater than the sum of its parts.” Science 320 (2008): 928–931.
Dreckmann, U., and M. Doebeli. “On the origin of spe-cies by sympatric speciation.” Nature 400 (1999): 354–357.
Freeman, Scott, and Jon c. Herron. “Mechanisms of speciation.” chap. 16 of Evolutionary Analysis. 4th ed. Upper Saddle River n.J.: Pearson Prentice Hall, 2007.
Futuyma, Douglas. Evolution. Sunderland, Mass.: Sinauer associates, 2005.
Grant, Peter, and Rosemary Grant. How and Why Species Multiply: The Radiation of Darwin’s Finches. Princ-eton, n.J.: Princeton University Press, 2008.
Grant, Verne. Plant Speciation, 2nd ed. new york: colum-bia University Press, 1981.
Gray, D. a., and W. H. cade. “Sexual selection and spe-ciation in field crickets.” Proceedings of the National Academy of Sciences USA 97 (2000): 14,449–14,454.
Howard, D. J., and Stuart H. Berlocher, eds. Endless Forms: Species and Speciation. new york: Oxford University Press, 1998.
Hunt, Dana e., et al. “Resource partitioning and sympatric differentiation among closely related bacterioplank-ton.” Science 320 (2008): 1,081–1,085.
irwin, Darren e., et al. “Speciation by distance in a ring species.” Science 307 (2005): 414–416.
Jiggins, chris D., et al. “Reproductive isolation caused by colour pattern mimicry.” Nature 411 (2001): 302–305.
Martin, Robert a. Missing Links: Evolutionary Concepts and Transitions through Time. Sudbury, Mass.: Jones and Bartlett, 2004.
Mayr, ernst. What Evolution Is. new york: Basic Books, 2001.
a feW examples of speCiation
organism divergence That Is occurring
Flowering plants Coevolution with pollinators
Hawaiian plants Coevolution with birds for seed dispersal
Fireweeds Polyploidy
Soapberry bugs Different host plants
Goldenrod gall insects Different host plants
Heliconius butterflies Different warning colorations
Field crickets Male courtship song
Stickleback fish Bottom-dwelling vs. open-water-dwelling popula-tions
Sockeye salmon Populations that return from the sea vs. remained in lakes
Blue tits Populations that breed in evergreen vs. deciduous oaks
Bowerbirds Male courtship patterns
xviii+598_Biodiversity_F.indd 495 11/17/11 1:13 PM

first entry496 spiders 496
Ortíz-Barrientos, Daniel, and Mohamed a. F. noor, “evi-dence for a one-allele assortative mating locus.” Sci-ence 310 (2005): 1,467.
Palumbi, Stephen R. The Evolution Explosion: How Humans Cause Rapid Evolutionary Change. new york: norton, 2001.
Price, Jonathan P., and Warren l. Wagner. “Speciation in Hawaiian angiosperm lineages: cause, consequence, and mode.” Evolution 58 (2004): 2,185–2,200.
Retchless, adam c., and Jeffrey G. lawrence. “temporal fragmentation of speciation in bacteria.” Science 317 (2007): 1,093–1,096.
Rieseberg, loren, and John H. Willis. “Plant speciation.” Science 317 (2007): 910–914.
Rundle, H. D., et al. “natural selection and parallel specia-tion in sympatric sticklebacks.” Science 287 (2000): 306–308.
Ryan, Peter G., et al. “ecological speciation in South atlan-tic island finches.” Science 315 (2007): 1,420–1,423.
Sargent, Risa D. “Floral symmetry affects speciation rates in angiosperms.” Proceedings of the Royal Society of London B (Biology) 271 (2004): 603–608.
Schilthuizen, Menno. Frogs, Flies, and Dandelions: The Making of Species. new york: Oxford University Press, 2001.
Uy, J. a. c., and G. Borgia. “Sexual selection drives rapid divergence in bowerbird display traits.” Evolution 54 (2000): 273–278.
Via, Sara. “Sympatric speciation in animals: the ugly duck-ling grows up.” Trends in Ecology and Evolution 16 (2001): 381–390.
Wake, D. B. “incipient species formation in salamanders of the Ensatina complex.” Proceedings of the National Academy of Sciences USA 94 (1997): 7,761–7,767.
Weir, Jason t., and Dolph Schluter. “the latitudinal gradi-ent in recent speciation and extinction rates of birds and mammals.” Science 315 (2007): 1,574–1,576.
Whibley, annabel c. “evolutionary paths underlying flower color variation in Antirrhinum.” Science 313 (2006): 963–966.
Zimmer, carl. “What is a species?” Scientific American, June 2008, 72–79.
spiders Spiders are the largest group of arach-nids, which is the largest group of chelicerates, in the arthropod phylum. arthropods have an external skeleton made primarily of chitin. this exoskeleton includes jointed appendages, which includes anten-nae, mouthparts, wings, and legs. Spiders have eight legs and no wings. Many arthropods have head, thorax, and abdomen body regions; in spiders, the head and thorax are fused into a cephalothorax. therefore, a spider can be easily distinguished from an insect, because a spider has two body parts and
eight legs, while an insect has three body parts and six legs. Spiders live everywhere except antarctica and the oceans (the small, delicate sea-spiders are arachnids but not spiders).
Spiders have mouthpart appendages called che-licerae that have venom glands. the chelicerae of spiders have poison fangs, which retract inside a groove like the blade of a pocketknife. almost all spiders are predators that paralyze their victims with venom and then inject enzymes that digest the victim; they suck the digested products. Spiders can-not chew. Because of this method of eating, spiders usually consume other arthropods, although spiders eat vertebrates (see photograph). if a small bird gets caught in a spider web, a large spider will not neces-sarily reject it. Most spider attacks on vertebrates, however, are defensive. One species of jumping spider, Bagheera kiplingi, is primarily vegetarian. Behind the chelicerae, ahead of the walking legs, are pedipalps that manipulate food.
Spiders, like other arachnids, have an open circulatory system in which the blood bathes all of
Spiders usually eat insects, but this large tropical spi-der has captured a gecko lizard. (Aidan Jones)
xviii+598_Biodiversity_F.indd 496 11/17/11 1:13 PM

spiders 497
the internal organs. the blood contains blue, cop-per-based hemocyanin that carries oxygen. Some spiders, like most terrestrial arthropods, have tra-cheae, which are breathing passages that open to the outside. Some spiders, like most aquatic arthro-pods, have gills, but these gills are enclosed inside a body chamber; these chambers are called book lungs.
like most arthropods, spiders have a pair of nerve cords running along the lower (ventral) surface of the body. in most spiders, the nerves form a large ganglion in the cephalothorax—the spider version of a brain. Spiders have eight eyes. the smaller ones are capable only of detecting movement, but the large ones can, unlike the eyes of most arthropods, form images. Some jumping spiders have eyes with with four visual pigments, and may therefore discriminate colors more effectively than mammals, which have only three visual pigments.
Spiders produce silk from an abdominal gland. Spider silk protein is one of the strongest materials known. Some spiders produce complex orb-shaped webs with sticky silk, in which small animals become entangled. the spider then paralyzes the animal and wraps it in silk. the spider knows which strands of the web are sticky and which are not, walking only on the latter. Other spiders produce cobwebs. Some spider webs are funnel-shaped, encouraging the prey to fall into the chamber of the waiting spider. the trapdoor spiders (family ctenizidae) live in burrows that are covered with a hinged door made of soil, vegetation, and silk. When a prey animal comes by, the spider quickly opens the door and grabs the vic-tim. Bolas spiders swing a line of silk, with adhesive compounds on the end, in the air to capture insects. they lure male moths within range by producing the female moth pheromone. Spiders often consume and reuse their own silk.
all spiders produce silk, but many spiders do not use silk to obtain prey. Many, such as tarantulas, wander in search of prey. Jumping spiders (family Salticidae), the largest family of spiders, jump up to 80 times the distance of their own body lengths, first securing themselves with a strand of silk. to do this, they use not only strong muscles, but also pressurized body fluid. crab spiders wait in vegeta-tion, often near flowers the coloration of which they closely mimic, and grab prey with strong front legs (see photograph).
Fishing spiders (see above) walk on the surface tension of water, and when they detect movements of prey underwater, they grab the prey and eat it. Spiders of the genus Argyroneta build an underwa-ter structure of silk, which they fill with air that they carry from the surface.
Some spiders live in societies of up to 50,000 individuals that cooperatively forage for prey and live together in common webs. they do not have social organization like that of the hymenopteran social insects. However, many spiders take advantage of the social organization of ants. Some spiders live near, or even inside of, ant colonies. they thereby gain protec-tion from predators. Sometimes they feed on the ants or on the aphids that the ants tend as sources of hon-eydew. these spiders are ant mimics, and mimic ants in numerous ways, such as the following:
● Spiders have two body parts, while ants have three. ant-mimic spiders often have constrictions in the middle of the cephalo-thorax, giving the appearance of three body parts.
● Spiders do not have antennae, but ants do. ant-mimic spiders often wave their front legs in apparent imitation of antennae.
● Spiders do not have large compound eyes, but ants do. ant-mimic spiders often have large color patches around the largest two of their eight eyes, making them look larger.
● ant-mimic spiders often imitate the behav-ior of ants. For example, jumping spiders that imitate ants do not jump, and ant-mimic spiders walk in a zig-zag, ant-like fashion. Synemosyna spiders imitate Pseu-domyrmex ants by walking on the outside edges of leaves. While Amyciaea spiders do not closely resemble Oecophylla ants, they imitate the behavior of dying ants, thus attracting workers. Some ant-mimic spiders hold an ant that they have just killed in their jaws, concealing their identity behind that of the dead ant.
This crab spider waits for prey on a Mexican poppy (Eschscholzia mexicana) in the saguaro desert of Arizona. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 497 11/17/11 1:13 PM

first entry498 spiders 498
● the spider Habronestes bradleyi releases 6-methyl-5-hepten-2-one, the same chemical that the ant Iridomyrmex purpureus uses as an alarm signal. this chemical attracts worker ants to the spider, which then eats them.
ant-mimic spiders also produce a pheromone that makes the ants react to them as if they were ants that belong in the home colony.
although spiders are excellent predators, they are often prey. in particular, “tarantula hawk” wasps (family Pompilidae) sting tarantulas but do not kill them, drag them into burrows, and lay eggs in them.
Sexual reproduction in spiders is a touchy and dangerous operation. after the final molt, the adult male stops eating. the male spider, often much smaller than the female, puts sperm on one of his front appendages and goes searching for a female. He uses special approach signals so that he is not mistaken for prey. the male inserts the append-age, with sperm, into a pore in the female’s abdo-men. Mating in scorpions is even more risky than in spiders, involving complex dances and the transfer of sperm into the female’s abdomen by one of the male’s legs. in some species, the female spider eats the male after mating.
Female spiders frequently take very good care of their offspring. they often protect the egg case. Female wolf spiders (family lycosidae) carry their egg cases, and when the spiderlings hatch, the mother may carry them on her back until they are large enough to live by themselves (see photograph). Fre-quently, a female spider will paralyze a small animal, not killing it, and lay her eggs in it; her offspring eat the still-living victim. in some species, the mother spider allows her hatchlings to eat her body.
the most primitive spiders (that is, those that most closely resemble the common chelicerate ances-tor) are the tarantulas, which are large wandering predators. new World tarantulas have irritating, defensive hairs that can be poisonous to small ani-mals. their chelicerae point downwards. Most spiders, in contrast, have chelicerae that point diago-nally forward. tarantulas produce silk but use it for building nests or lining burrows. Most range in size from 1 to 4 inches (2.5 to 10 cm), although the Goli-ath birdeater (Theraphosa blondi) can be 10 inches (28 cm) in length (including the legs). tarantulas prey mainly on other arthropods, but some can kill mice or birds.
nine of the most venomous spiders in the world live in australia. two of them are large funnel-web spiders. the Sydney funnel-web (Atrax robustus), found only within about 60 miles (100 km) of Syd-ney, is responsible for most deadly spider bites in
australia, especially the males that wander into sub-urban areas. the northern tree funnel-web spider (Hadronyche formidabilis) is also dangerous. the australian redback spider (Lactrodectus hasseltii) is a relative of the american black widow spider.
in the United States, the two native species of poisonous spiders are the black widow (Lactrodectus mactans) and the brown recluse (Loxosceles reclusa). Venom from female black widow spiders causes a buildup of the neurotransmitter acetylcholine. this neurotransmitter normally stimulates muscle con-tractions; when it builds up, excessive muscle con-tractions and high blood pressure result. the toxin of the brown recluse destroys red blood cells and induces the white blood cells to attack surrounding tissues. Fortunately, these bites are seldom fatal. Most spiders bite in defense, although the Brazil-ian wandering spider Phoneutria nigriventer readily attacks larger animals, including humans. Despite the fear that humans have of spiders (exacerbated by the recognized psychological term arachnophobia), there were only about 100 confirmed cases of human deaths by spider bite in the 20th century.
See also arthropods.
Some spiders take care of their offspring. This mother wolf spider carries baby spiders on her abdomen in the Badlands of South Dakota. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 498 11/17/11 1:13 PM

succession, ecological 499
Further readingallen, Rachel a., et al. “exploitation of an ant chemical
alarm signal by the zodariid spider Habronestes brad-leyi Walckenauer.” Proceedings of the Royal Society of London B (Biology) 263 (1996): 69–73.
andrade, Maydianne c. B. “Sexual selection for male sac-rifice in the australian redback spider.” Science 271 (1996): 70–72.
Dalton, Stephen. Spiders: The Ultimate Predators. Rich-mond Hill, Ont.: Firefly Books, 2008.
elgar, Mark a., and Rachel a. allan. “chemical mimicry of the ant Oecophylla smaragdina by the myrme-cophilous spider Cosmophasis bitaeniata: is it colony-specific?” Journal of Ethology 24 (2006): 239–246.
Gastreich, K. R. “trait-mediated indirect effects of a theri-diid spider on an ant-plant mutualism.” Ecology 80 (1999): 1,066–1,070.
Hillyard, Paul. Book of the Spider. new york: Harper Perennial, 1998.
———. The Private Life of Spiders. Princeton, n.J.: Princ-eton University Press, 2008.
Johnson, J. chadwick, and andrew Sih. “Fear, food, sex, and parental care: a syndrome of boldness in the fish-ing spider, Dolomedes triton.” Animal Behaviour 74 (2007): 1,131–1,138.
Maddison, Wayne, and Jerzy Proszynski. “Jumping spiders of the world: a confederation of projects on salticid systematics and diversity.” available online. URl: http://salticidae.org/jsotw.html. accessed May 16, 2011.
nelson, Ximena J., and Robert R. Jackson. “collective Batesian mimicry of ant groups by aggregating spi-ders.” Animal Behaviour 78 (2009): 123–129.
———. “anti-predator crèches and aggregations of ant-mimicking jumping spiders (araneae: Salticidae). Biological Journal of the Linnean Society 94 (2008): 475–481.
———, et al. “living with the enemy: Jumping spiders that mimic weaver ants.” Journal of Arachnology 33 (2005): 813–819.
Pechmann, Matthias, and nikola-Michael Prpic. “append-age patterning in the South american bird spider Acanthoscurria geniculata (araneae: Mygalomor-phae). Development Genes and Evolution 219 (2009): 189–198.
Vollrath, Fritz, and Paul Selden. “the role of behavior in the evolution of spiders, silks, and webs.” Annual Review of Ecology, Evolution, and Systematics 38 (2007): 819–846.
Wise, David H. Spiders in Ecological Webs. cambridge, U.K.: cambridge University Press, 1995.
succession, ecological areas disturbed by fire, storms, or landslides (ecological disturbances) are temporary microhabitats. these areas quickly fill with plant growth and are no longer open to
new plant colonists that require open spaces to ger-minate. in early-successional habitats, soon after the disturbance, plants and animals may thrive in conditions of reduced competition, but as the areas fill with more species and become late-successional habitats, competition becomes more intense.
consider the case in which a plot of land from the deciduous forest of eastern north america has been cleared for farming (an example of an ecological disturbance), then abandoned (see the bottom photo-graph on page 504). Most of the first plants to live in the disturbed area are annuals, which are plants that live only one year (see table on page 503). they grow quickly in the abundant nutrients and light. these col-onizing plants are often the familiar weeds of roadside and farm. they reproduce during their first year, then they die. if the seeds are small, they may disperse over a wide range, and a few of them may land in a newly disturbed area. Weed seeds may remain dormant but alive in the soil in that place for decades or even centu-ries, until the next disturbance occurs there.
During the first season, the disturbed area is thick with annuals. in their shade, seedlings of herbaceous perennials (plants that live for several to many years, for example the goldenrod) grow slowly. although they grow slowly during their first year, they store nutrients in their stems or roots. at the beginning of the second season, many seeds of the annuals may germinate, but the perennials grow faster, due to the food reserves they accumulated the previous year.
During the second year, the annuals grow slowly in the shade and usually die without reproducing. Starting the second year or soon thereafter, the peren-nials dominate the field. these herbaceous perenni-als sprout from underground buds each year. Bushes and trees grow up through their shade, becoming more common until they in turn dominate.
the first trees to dominate the field are the shade-intolerant trees, for example ashes, cherries, and locusts. they are called shade-intolerant because their seedlings must germinate in the bright light of an open field. Some shade-intolerant shrubs and trees spread not so much by seeds as by underground stems. they form a large clump of trunks (see figure on page 504). the seedlings cannot grow in the shade of the parents; therefore, shade-intolerant trees dominate the habitat only temporarily. the seedlings of shade-tolerant trees, such as maples, grow up in the shade of the shade-intolerant trees. Forest floor herbs that cannot tolerate open fields now grow in the shade. the plants in each stage shade out the plants of the preceding stage.
Finally, the old forest clearing may be filled with the same kind of forest that was present prior to disturbance. a maple forest, for example, can perpetuate itself, since maple seedlings can grow up in the shade of adult maples. no further changes in
xviii+598_Biodiversity_F.indd 499 11/17/11 1:13 PM

fi rst entry500 succession, ecological 500
species composition usually occur, in the absence of further disturbance, in such a community. ecolo-gists often use metaphors to describe the self-per-petuating forest: it is called climax forest as if it is the culmination of a story, or mature forest, as if succession were the growth stages of an organism.
in the eastern forests of the United States, between 100 and 200 years must pass before a climax species composition is reached. as the climax forest gets older, its dominant trees get larger. in the eastern deciduous forest zone of the United States, few people will ever see a true “virgin forest.” Practically all of the decidu-ous forests have been cut down within the past two centuries. although many of these forests have grown back and once again contain their original species (see the bottom photograph on page 504), they no longer contain the huge old trees that the native americans knew and that were described by the fi rst european explorers.
although some early-successional species die out during the process of succession, others persist near the edge of the developing forest. Many shade-toler-ant forest-fl oor plants can survive only in the shade of the climax trees. net species diversity, therefore, increases during succession in these cases.
along with plant succession, there is also a suc-cession of animals. Being mobile, animals are not
as confi ned to a particular successional stage as are most plants. Many mammals and birds may fi nd abundant food in early-successional zones but can also fl y or run into the deep woods. Many insects specialize on certain food plants and are therefore restricted in the successional habitat in which they live. caterpillars of the monarch butterfl y, for exam-ple, live exclusively upon milkweeds that grow in recently disturbed areas. Many species of small ani-mals, and fungi, may be able to survive only in the cool, moist soils of the forest fl oor.
During succession, the early colonists change the soil, light, temperature, and humidity conditions, which may facilitate the survival of species that arrive later. they might also inhibit the survival of species that arrive later. in other words, the early-succes-sional species set the stage for many of the later-successional species. this is not true, however, of all of the later-successional species. Shade-tolerant tree species may begin to grow the very fi rst season after disturbance, surviving in the shade of the weeds. they may dominate the habitat later not because they arrived later but because they grew more slowly.
Other kinds of disturbances result in different patterns of succession. if the forest burned instead of being converted into a farm, succession would fol-low a different pattern. Some trees might escape the
During secondary succession, plants grow in disturbed areas with intact soil. Annual weeds grow most rapidly and are replaced in turn by perennial weeds, shrubs, and then trees. The trees that grow latest in the succes-sional series are the species whose seedlings can grow in the shade of the adults.
xviii+598_Biodiversity_F.indd 500 11/17/11 1:13 PM

succession, ecological 501
what would happen to the world if humans vanished from it?
Such a thing is very unlikely to happen. Humans are so numerous and so adapt-able that there will probably be at least a few people on the Earth until the Sun enters its final throes of death. What is more likely to happen is that human civili-zation could collapse. The modern human economy and system of food production depend entirely upon fossil fuels and advanced technology. While there would still be sunshine and photosynthesis, there would not be fuel and machinery for plant-ing, growing, and harvesting crops, for processing food, or for transporting food to where people can obtain it. Even the avail-ability of food does not guarantee survival, as was evidence during the Irish Potato Famine, when people starved because they could not buy the plentiful food in the marketplace. The human population of nearly 7 billion cannot conceivably be supported by hunting and gathering, espe-cially by a mob of hungry people pour-ing out of cities and towns. Even rural people survive on groceries. The few that are lucky enough to find wild hickory nuts (or to even know what they are) will dis-cover that the shells are too hard to break except with a very large hammer. To sur-vive by cracking hickory nuts uses more calories than the nuts would provide. The few naturalists who know that greenbriar shoots are edible would find them only in the spring. As soon as the ammunition ran out, few hunters would know what to do, aside from wrestling a few turtles. Even though the extinction of humans is extremely unlikely, such a disappearance is a useful scenario for understanding human impact upon Earth.
Much of what humans have done to the Earth would be quickly erased. Mod-ern buildings, less sturdy than ancient ones, would soon collapse. Mayan build-ings have been almost buried by soil and vegetation during the past millennium. Ancient buildings in dry environments have survived a bit longer.
Nearly all of the plants and animals that humans have bred as crops and livestock would quickly become extinct. They largely depend upon humans for their survival. Tomatoes, for example, cannot produce seeds unless humans keep their fruits from touching the ground and rotting. Ears of corn would sprout into a dense mass of seedlings, all of which would probably die. Poodles would die instantly. Humans have bred crops and livestock for purposes other than survival in the wild. Some live-stock, pets, and plants would become feral, as has happened with horses, swine, cats, and kudzu. Pest animals and weeds have adapted to the condi-tions that humans have created. Some of them, such as rats, would continue to thrive. But as humans are the major source of ecological disturbances, the weeds and pests that live in such condi-tions would eventually become rare. The weeds that thrive in agricultural fields are generally not the same ones that colonize after disturbances, such as floods and fires, in the natural world. For example, the velvetleaf weed (Abutilon theophrasti), which today thrives in soy-bean fields and in recently abandoned farmland, would grow only occasionally on the banks of streams and lakes. The human contribution to the diversity of species and, especially, varieties within species would vanish quickly. The new rulers of the Earth would be species that existed before humankind.
Some disturbances would become more common after the disappearance of most humans than they now are. Fire suppression would cease, and fires might once again clear vast areas in which grassland and chaparral plant species, and the animals that live with them, could grow.
Human population collapse has occurred before. When Europeans intro-duced diseases such as smallpox to North American native populations, the germs spread faster than the Europeans,
and annihilated up to 90 percent of the natives. At that time, population explo-sions of wild animal species occurred. Natives in the middle of North Amer-ica had hunted bison and passenger pigeons; with the collapse of hunting, populations of pigeons and bison vastly increased. Natives in the southeastern part of North America had fished exten-sively; with their disappearance, popu-lations of fish—and of alligators that ate them—exploded. This explains the observations of Europeans and Ameri-cans, such as botanist William Bartram, who marveled at the hundreds of alliga-tors in every lake and river in Florida; and they assumed North America had always been this way. If humans vanished today, a similar phenomenon would probably not occur, since most sustenance comes from helpless livestock rather than wild game.
One human artifact would persist for millennia: the high levels of carbon diox-ide in the atmosphere, contributing to a greenhouse effect. The Earth would be warmer for thousands of years because humans had been on it. However, this greenhouse effect would not be very devastating, except for tundra species whose habitat would vanish forever. The main reason that the greenhouse effect threatens biodiversity is because humans have blocked the corridors of movement by which, in previous millen-nia, plants and animals have adjusted to global climate changes by migrat-ing. With humans out of the way, global warming would probably cause even fewer extinctions than the end of the most recent ice age. Indeed, the leading explanation for the Pleistocene extinc-tion is the overkill hypothesis, in which humans killed enough of the large ani-mals in North America that their popula-tions could not persist. Even coral reefs, already dying because carbon diox-ide is acidifying ocean waters, would
“if huMAnS vAniShed . . .”
(continues)
xviii+598_Biodiversity_F.indd 501 11/17/11 1:13 PM

first entry502 succession, ecological 502
fire and serve as a source of seeds that would hasten the regrowth of that species. Many perennial plants in the forest may sprout from surviving underground parts. in many cases, the events that follow a fire are not succession at all. if a grassland burns, most of the plants that grow after the fire are simply sprouts from the plants that were there before. if a savanna burns, the grasses may simply spread underneath the charred tree trunks.
each region of the world has its own set of annu-als, herbaceous perennials, shade-tolerant trees, and shade-intolerant trees and has its own unique trajec-tory of succession. in the Rocky Mountains, aspen (Populus tremuloides) is a common early-succes-sional tree, while spruce (Picea glauca) is a common late-successional tree. in the Sierra nevada, giant sequoia (Sequoiadendron giganteum) is an early-successional tree, and white fir (Abies concolor) is a late-successional tree. Giant sequoias grow to prodi-gious sizes and live for millennia but do not repro-duce well except in sunny openings. Fire suppression has resulted in reduced regeneration of sequoias and the spread of white firs. in tropical rain forests, large natural disturbances are rare. When a canopy tree falls, tree seedlings grow rapidly and fill the gap in a decade or less. in some cases, there are many seed-lings of canopy trees growing slowly in the under-
story. if a disturbance does not happen soon, these seedlings will die. But if a disturbance does occur, these seedlings begin to grow rapidly. they win the competitive race by waiting in the shade as seedlings rather than seeds.
Furthermore, within any given area, the plant species that dominate each successional stage will depend on the species of seeds that are in the soil or brought in by wind or animals. if there are no nearby maple forests, it may take a plot of land a very long time to grow back into a maple forest, for the seeds must travel from a great distance, and it may be two or three generations before maples form a continu-ous cover. Moreover, soil plays an important role in the makeup of the climax community. in Michigan forests on good soil, the climax species composition includes maples, beeches, and hemlocks; however, on glacial sand and gravel, the mature forest is domi-nated by jack pines.
Secondary succession occurs when the veg-etation is destroyed but the soil, even if disturbed, remains in place. Because soil is present, colonizing plants can grow immediately. the soil contains seeds that begin to grow immediately. Disturbances such as fires and floods do not usually remove the soil.
Some kinds of disturbance, however, create new land surface, on which soil must be built before
recuperate in a few hundred years after human disappearance.
Perhaps the human artifact that would persist the longest, for several million years, would be the toxins that industry manufactures. Heavy metals and persistent organic molecules already recycle through the food web over and over. And then there is plastic. Plastic is forever. No known natural process breaks it down. Save for a little bit of plastic destroyed during incineration, every bit of plastic ever manufactured still exists. Ultraviolet radiation and oxy-gen in the air can make it brittle, but it simply fractures into smaller and smaller pieces. This makes plastic trash even more dangerous. When the fragments are small, many animals consume it with the plankton that they normally eat. Nat-ural selection has not produced seabirds that can distinguish between a fish and a
floating piece of plastic. In some places, each handful of sand is 20 percent plas-tic, and plastic fragments are up to six times as abundant as plankton. There are seven major gyres of ocean current that trap even large pieces of plastic into large, and almost permanent, garbage dumps. Long after the lights of the cities cease to be visible on the dark side of the Earth, long after the last reservoirs fill with silt and their rivers rush over the dams like natural waterfalls, and entropy has scrambled the last of the digital data, flakes of plastic will still be swirling through the ecosystems of the Earth.
See also disturBances, ecological; greenHouse effect; greenHouse effect and Biodiversity; invasive species; pleisto-cene extinction; succession, ecological.
Further readingCronon, William, ed. Uncommon Ground:
Rethinking the Human Place in Nature. New York: Norton, 1995.
Donlan, Josh, et al. “Re-wilding North Amer-ica.” Nature 436 (2005): 913–914.
Quammen, David. “The weeds shall inherit the Earth.” The Independent, November 22, 1998, 30–39.
Rathje, William, and Cullen Murphy. Rubbish! The Archaeology of Garbage. Tucson: Uni-versity of Arizona, 2001.
Sanderson, E. W., et al. “The human footprint and the last of the wild.” BioScience 52 (2002): 891–904.
Tegmark, Max, and Nick Bostrum. “How unlikely is a doomsday catastrophe?” Nature 438 (2005): 754.
Ward, Peter, and Donald Brownlee. The Life and Death of Planet Earth. New York: Henry Holt, 2002.
Weisman, Alan. The World without Us. New York: Picador, 2008.
Zalasiewicz, Jan. The Earth after Us. Oxford, U.K.: Oxford University Press, 2009.
(continues)
xviii+598_Biodiversity_F.indd 502 11/17/11 1:13 PM
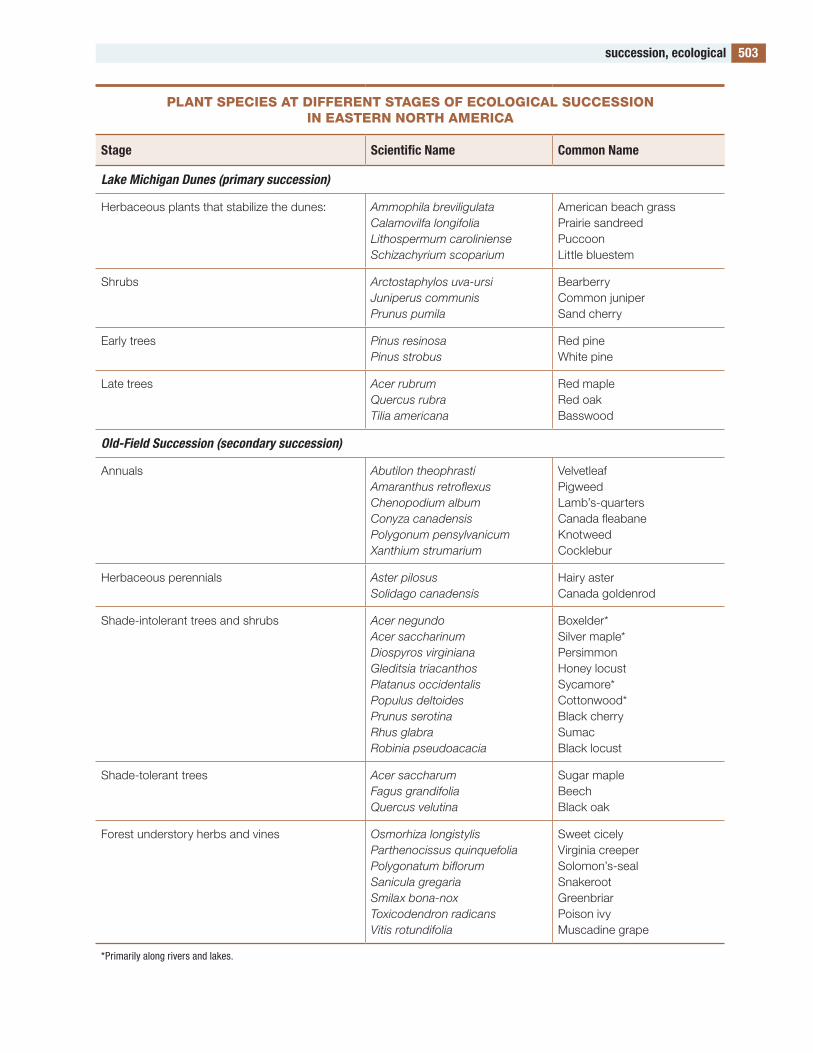
succession, ecological 503
plant speCies at different stages of eCologiCal suCCession in eastern north ameriCa
Stage Scientific Name Common Name
Lake Michigan Dunes (primary succession)
Herbaceous plants that stabilize the dunes: Ammophila breviligulataCalamovilfa longifoliaLithospermum carolinienseSchizachyrium scoparium
American beach grassPrairie sandreedPuccoonLittle bluestem
Shrubs Arctostaphylos uva-ursiJuniperus communisPrunus pumila
BearberryCommon juniperSand cherry
Early trees Pinus resinosaPinus strobus
Red pineWhite pine
Late trees Acer rubrumQuercus rubraTilia americana
Red mapleRed oakBasswood
Old-Field Succession (secondary succession)
Annuals Abutilon theophrastiAmaranthus retroflexusChenopodium albumConyza canadensisPolygonum pensylvanicumXanthium strumarium
VelvetleafPigweedLamb’s-quartersCanada fleabaneKnotweedCocklebur
Herbaceous perennials Aster pilosusSolidago canadensis
Hairy asterCanada goldenrod
Shade-intolerant trees and shrubs Acer negundoAcer saccharinumDiospyros virginianaGleditsia triacanthosPlatanus occidentalisPopulus deltoidesPrunus serotinaRhus glabraRobinia pseudoacacia
Boxelder*Silver maple*PersimmonHoney locustSycamore*Cottonwood*Black cherrySumacBlack locust
Shade-tolerant trees Acer saccharumFagus grandifoliaQuercus velutina
Sugar mapleBeechBlack oak
Forest understory herbs and vines Osmorhiza longistylisParthenocissus quinquefoliaPolygonatum biflorumSanicula gregariaSmilax bona-noxToxicodendron radicansVitis rotundifolia
Sweet cicelyVirginia creeperSolomon’s-sealSnakerootGreenbriarPoison ivyMuscadine grape
*Primarily along rivers and lakes.
xviii+598_Biodiversity_F.indd 503 11/17/11 1:13 PM

first entry504 succession, ecological 504
plants can grow on it. During primary succession, plants contribute to the buildup of the soil.
One of the earliest examples of primary succes-sion to be studied, and still the most famous, is the development of a deciduous forest on top of sand dunes along lake Michigan. Botanist Henry chan-dler cowles, like many other people, had visited the southern shores of lake Michigan, near chicago, an area now preserved in the indiana Dunes national lakeshore. However, cowles saw more than just the beautiful sand and the oak forest. a graduate stu-dent and later a professor of botany at the University of chicago, he realized that these dunes and forests were constantly changing. the changes were too slow to actually observe. Because cowles used the present as the key to the past, he was able to conduct the first scientific study of ecological succession, which was published about 1900.
Over thousands of years, sand dunes have accu-mulated along the southern shore of lake Michi-gan. even though the weather in this region is moist, the sand dunes are a dry microenvironment, because the sand cannot hold very much water. Plants that live in the open dunes near the lake must tolerate a wide range of challenges: the sand is dry, becomes very hot in the sun, and shifts in the wind, sometimes burying the leaves and sometimes exposing the roots of the plants. Only a couple of species of grass are able to colonize these sand dunes.
the grasses not only survive in the dunes but also begin to change the dunes. their stems and roots hold down the sand, while their dead leaves, stems, and roots contribute organic matter and humus to it, enriching it with nutrients (see table and figure). the humus begins to hold water from the
abundant rains. lack of rain prevents this process from occurring very much in desert sand dunes.
Other plants, such as bushes, grow in the sand once the grasses have stabilized and enriched it. the dune is no longer merely sand, although it is still mostly sand. Pine trees begin to grow amidst the bushes. in their shade, the dunes are cooler and do not dry out as much. Pine needles further enrich the soil. Other bushes and trees begin to grow that could not survive in the bright sun and dry sandy soil. Mean-while, the original grasses have been shaded out.
Forest trees such as oak and basswood grow up under and shade out the pines. after about a century, an oak forest is growing on what was once a sand dune (see photograph on page 506). Sugar maple (Acer saccharum) and beech (Fagus grandi-folia) dominate the general region, but oak forests dominate the old dunes because the soil consists only of sand and leaf litter and holds less water than the loam soils in which maples grow.
Because dunes accumulate along the shore, the dunes become progressively older inland. a visitor can walk away from the shore and, in effect, be walking along a time axis of succession. in just a
A persimmon tree (Diospyros virginiana) grows in a grassland, beginning the process of transforming the grassland into forest. The “tree” looks like numerous small trees, but they are all connected underground and are a singe individual. (Stanley A. Rice)
A forest of maples and hemlocks grows in what was a farm during Revolutionary War times at Teatown Lake, north of New York City. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 504 11/17/11 1:13 PM

succession, ecological 505
hundred meters, a visitor or a scientifi c investigator can walk through centuries of succession.
Primary succession also occurs on lava fl ows in Hawaii. When the lava cools into rock, it is a barren, sterile surface. Gradually, dust (with mineral nutrients) accumulates. Seeds and spores blow in, accumulat-ing in the deeper fi ssures, where the most water is found. Ferns and ohia trees (Metrosideros polymor-pha) begin to grow in these fi ssures (see photograph on page 507). their roots help crack the lava into smaller pieces, and their dead leaves help to build up a soil. Ferns and ohia trees will eventually dominate the mature forest. Primary succession depends upon the amount of rainfall; rain forest cannot grow on old lava fl ows in those areas of Hawaii that receive little rain.
in some examples of succession, new land sur-face is formed, but this surface may consist of rich soil brought in from elsewhere. Ponds, for instance, gradually fi ll in with eroded soil and with dead plant material. the new soil thus created is rich and supports the growth of emergent plants such as cat-tails and rushes that grow in water but are rooted in soil. these plants also help build up the soil by contributing their dead leaves and by trapping silt. Once enough soil has accumulated that the standing
water has been replaced by mud, seedlings of trees such as willow and boxelder (Acer negundo) can begin growing. these kinds of trees are typically found only along streams, rivers, and pond margins (see table on page 503). they are shade-intolerant, however, and may be replaced by maples that grow in their shade. a pond is a temporary habitat, and may eventually be replaced by a maple forest, unless further disturbances (for example, fl ooding) occur. Succession can occur as a bog fi lls in, just as in a pond. However, when a bog becomes a forest, the soil remains acidic because the soil consists mostly of dead peatmoss. the acidity of the soil restricts the types of plants that can grow in the climax for-est that grows where a bog once existed.
Disturbance and succession continually occur along rivers, as soil erodes from or is deposited along the bank. in the new soil, seedlings of boxelder and other streamside trees germinate and grow, begin-ning the process of succession all over.
ExCESSIvE dISrupTIoNS of EColoGICal CommuNITIES
a million years ago, when there were only a million people in the world, it did not really matter what
During primary succession, soil must build up before most plants can grow. The plants help to build up the soil. In the dunes south of Lake Michigan, grass rhizomes hold down the sand and contribute organic material to it, allowing shrubs and then trees to grow in turn. The plants themselves change the environmental conditions of moisture, nutrients, light, organic matter, and wind.
xviii+598_Biodiversity_F.indd 505 11/17/11 1:13 PM

first entry506 succession, ecological 506
humans did to the earth. When a caveman threw out a stone tool, it was just a rock. But today, there are 7 billion people, and when they throw out dead batteries, it is toxic waste. Both because of prodi-gious population and toxic technology, every little thing humans do now seems to be harmful to the earth.
Some disturbances caused by humans are more disruptive than others. Some human activities—particularly carrying species to new locations, causing extinctions, and creating new kinds of pollution—have disrupted ecological communities in ways that may require millions of years for recovery. Severe disturbances have occurred during the earth’s history, for instance the giant asteroid believed to have struck the earth 65 million years ago, causing the cretaceous extinction. Severe dis-turbances continue to occur, for instance volcanic eruptions. the 1883 eruption of Krakatoa in South-east asia produced a noise that could be heard—and a tidal wave that reached—thousands of miles. ecological communities have recovered from each of these disturbances, however. communities do not so easily recover from repeated disturbances, such as mowing, soil compaction, and other distur-bances caused by humans.
Human technology creates many disturbances that are not only repeated but also quite different from natural ones. During the brief period of earth history in which human technology has existed, spe-cies have not adapted to these new kinds of dis-turbances, particularly the discharge of poisonous pollutants. around the world, extensive disruption of soil is occurring due to deforestation and overgrazing. the result is often the spread of deserts. extensive soil erosion resulting from deforestation has contributed to the downfall of civilizations all over the world, for instance those of the Greeks and Romans, of the ana-sazi of western north america, and of the Polyne-sians on easter island in the Pacific Ocean. no species has disturbed as many square miles of land surface or polluted as much ocean as have humans.
See also essay: “if Humans Vanished . . .”; com-petition, ecological; disturbances, ecological.
Further readingBazzaz, Fakhri a. “Succession on abandoned fields in the
Shawnee Hills, southern illinois.” Ecology 49 (1968): 924–936.
———. “the physiological ecology of plant succession.” Annual Review of Ecology and Systematics 10 (1979): 351–371.
On the sand dunes south of Lake Michigan, only a few seedlings can sprout, and most of these die (left). Once the seedlings and grass rhizomes have stabilized the soil, trees can grow. A forest of linden (Tilia americana) and red oak (Quercus rubra) grows on what was once a sand dune (right). (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 506 11/17/11 1:13 PM

symbiogenesis 507
———. Plants in Changing Environments: Linking Physi-ological, Population, and Community Ecology. new york: cambridge University Press, 1996.
christensen, norman l., and Robert K. Peet. “conver-gence during secondary forest succession.” Journal of Ecology 72 (1984): 25–36.
cowles, Henry chandler. “the ecological relations of the vegetation on the sand dunes of lake Michigan.” Botanical Gazette 27 (1899): 95–117, 167–202, 281–308, 361–391.
Dale, Virginia H., Frederick J. Swanson, and charles M. crisafulli, eds. Ecological Responses to the 1980 Erup-tion of Mount St. Helens. new york: Springer, 2005.
Foster, David R., and John D. aber, eds. Forests in Time: The Environmental Consequences of 1,000 Years of Change in New England. new Haven, conn.: yale University Press, 2004.
Gill, David S., and Peter l. Marks. “tree and shrub seed-ling colonization of old fields in central new york.” Ecological Monographs 61 (1991): 183–205.
Hopfensperger, Kristine n. “a review of similarity between seed bank and standing vegetation across ecosystems.” Oikos 116 (2007): 1,438–1,448.
Körner, christian, et al. “Small differences in arrival time influence composition and productivity of plant com-munities.” New Phytologist 177 (2008): 698–705.
lichter, John. “colonization constraints during primary succession on coastal lake Michigan sand dunes.” Journal of Ecology 88 (2000): 825–839.
Macleish, William H. The Day before America: Changing the Nature of a Continent. new york: Houghton Mif-flin, 1994.
Pickett, Steward t. a. “Succession: an evolutionary interpretation.” American Naturalist 110 (1976): 107–119.
———. “Population patterns through twenty years of old-field succession.” Plant Ecology 49 (1982): 45–59.
Pielou, e. c. After the Ice Age: The Return of Life to Glaci-ated North America. chicago: University of chicago Press, 1992.
Rice, Stanley a. “Plants heal the landscape.” chap. 9 in Green Planet: How Plants Keep the Earth Alive. Pisca-taway, n.J.: Rutgers University Press, 2009.
Rosenzweig, Michael. Win-Win Ecology: How the Earth’s Species Can Survive in the Midst of Human Enter-prise. new york: Oxford University Press, 2003.
Wardle, David a., et al. “the response of plant diversity to ecosystem retrogression: evidence from contrast-ing long-term chronosequences.” Oikos 117 (2008): 93–103.
yoon, carol Kaesuk. “as Mt. St. Helens recovers, old wisdom crumbles.” New York Times, (May 16, 2000). available online. URl: http://www.nytimes.com/2000/05/16/science/as-mt-st-helens-recovers-old-wisdom-crumbles.html?pagewanted=all. accessed May 16, 2011.
symbiogenesis Symbiogenesis is the generation of biodiversity and of evolutionary novelty through symbiosis. Symbiosis is the physiological associa-tion of members of two or more distinct kinds of organism, and it results from coevolution. When populations of two species form a close symbiotic partnership, natural selection may favor genetic changes that enhance the partnership of the two species. if these genetic changes are associated with reproductive isolation, then the partners may be considered new species distinct from their immedi-ate ancestors. Further, the partners may merge their activities so closely, as when photosynthetic bacte-ria bocame chloroplasts or when algae merge with fungi to become lichens, that they are often named as a single new species. as a result, symbiogenesis can result in the origin of new species by merger of some populations of previously independent species. the principal modern scientist who has promoted awareness of symbiogenesis is lynn Margulis of the University of Massachusetts at amherst.
the principal image that has dominated Darwin-ian evolutionary theory is the “tree of life,” in which ancestral species branch out to form new species. One
Ohia trees (Metrosideros polymorpha) grow from the cracks of pahoehoe lava on the Kona coast of the Big Island of Hawaii. In this unusual pattern of succession, the first plants to grow on the lava are also the plants that dominate the forests that will later cover the lava. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 507 11/17/11 1:13 PM

first entry508 symbiogenesis 508
of the earliest images of the tree of life is the single illustration that is found in charles Darwin’s land-mark 1859 book Origin of Species. in this image, the branches grow farther and farther apart but never merge. When symbiogenesis occurs, however, the tree of life experiences grafts, sometimes between branches that are very far apart from one another on the tree.
the earliest statements of the idea of symbiogen-esis were made by two Russian botanists, Konstantin S. Mereschkovsky and andrey S. Famitsyn, prior to the Russian Revolution. after Mereschkovsky died in 1918 and Famitsyn in 1921, another Russian bota-nist, Boris M. Kozo-Polyansky, kept the idea alive. it was never, however, a major theory in the Soviet Union, especially in the Stalinist era, when the theo-ries of lysenkoism dominated genetics and evolution, and remained practically unknown in the United States until recently, when their ideas were promoted by Margulis and science writer Dorion Sagan. inde-pendently of the Russian scientists, an american anatomist, ivan e. Wallin (at the University of colo-rado Medical center in Denver), formulated an idea of symbiogenesis, which he called symbionticism in a 1927 book. His ideas were ignored, and he finished his career without pursuing symbiosis any further.
lynn Margulis investigated the inheritance pat-terns of some genetic traits that appeared to be passed on not through the nucleus but through the cytoplasm of the cell. in sexual reproduction, most of the cytoplasm of the fertilized egg cell comes from the unfertilized egg, rather than from the sperm; the cytoplasmic factors showed a pattern of inheritance that was passed on solely through the maternal line. this indicated that there was Dna in the cytoplasm, not just in the nucleus. and she had an idea where that Dna was located.
all independently living cells (and most viruses as well) have genetic information encoded in Dna. in contrast, organelles (specialized struc-tures within the cell) generally do not have their own Dna; their production and operation are dic-tated by the Dna in the cell’s nucleus. Margulis wrote a paper in 1967 in which she proposed that chloroplasts and mitochondria had used their own Dna. the Dna was there because these organ-elles are the evolutionary descendants of bacteria that had merged with ancestral eukaryotic cells. the bacteria remained, without killing their host cells, and the host cell lineages evolved processes that accommodated and made use of the bacte-ria. the bacteria became symbionts. if chloroplasts and mitochondria were merely cell structures, one would not expect them to have their own Dna. the chloroplasts had evolved from photosynthetic bacteria (often called blue-green algae), while the mitochondria had evolved from aerobic bacteria that consumed organic materials.
these symbiotic events happened more than a billion years ago. Frequently in cells, segments of Dna can move from one location to another. Many of the genes of the mitochondria and chloroplasts were transposed from the symbiotic bacteria to the nuclei of the host cells. the simplified symbionts were now incapable of surviving on their own, and they became the chloroplasts and mitochondria that are today found only in eukaryotic cells.
though not accepted by many scientists at first, Margulis’s proposal of symbiogenesis quickly advanced in the scientific community. as better chemical and microscopic techniques became avail-able, convincing evidence of the bacterial origin of chloroplasts and mitochondria emerged. chloro-plasts and mitochondria:
● have their own Dna, which resembles bac-terial Dna more than it resembles the Dna of eukaryotic chromosomes.
● have ribosomes, which are structures that use genetic information in Dna to produce pro-teins. as a result, mitochondria and chloro-plasts produce some of their own proteins. in the case of an important photosynthetic enzyme called rubisco, part of the enzyme (the large subunit, rbcl) is produced by the chloroplasts, while the small subunit is pro-duced by the ribosomes out in the chloroplast, using information not found in the nucleus.
● reproduce themselves, rather than being constructed by the cell. Small, immature mitochondria and chloroplasts may abound in eukaryotic cells; some of the immature structures mature into functional mito-chondria and chloroplasts, which cannot reproduce, while other immature structures reproduce themselves.
in addition, some marine invertebrates consume algae but keep the algal chloroplasts alive in some of their cells. this strongly suggests that chloroplasts and mitochondria may have arisen in a similar way, as symbiotic bacteria that lost most of their genetic independence—but not quite all of it, thus betraying their symbiogenetic origins.
as techniques became available to actually determine the sequence of nucleotides within Dna and other nucleic acids, it became possible to make comparisons between the Dna of different species to determine their evolutionary relationships. Fol-lowing the initial lead of carl R. Woese of the Uni-versity of illinois, biologists constructed a tree of life showing the branching patterns of evolution. the tree was based solely upon living species from which Dna and Rna could be obtained. Mitochondrial genes turned out to be very close to aerobic bacteria
xviii+598_Biodiversity_F.indd 508 11/17/11 1:13 PM

symbiogenesis 509
on the tree of life, and not at all similar to the genes in the nuclei of the cells in which mitochondria live. chloroplast genes turned out to more closely resemble cyanobacteria on the tree of life than the genes in the nuclei of the plant cells in which they now live. this is precisely what would be expected as confirmation of Margulis’s symbiogenetic theory of their origins.
While nearly all biologists accept Margulis’s symbiogenetic explanation of the origin of chloro-plasts and mitochondria, some of her subsequent proposals have not met with widespread acceptance. Perhaps the most famous example is her proposal that the motile structures of eukaryotic cells were originally symbiotic bacteria as well.
Margulis uses the term undulipodia (“undulat-ing feet” in Greek) to denote structures in eukary-otic cells that are usually called cilia and eukaryotic flagella. cilia are hairlike structures on the outside of many eukaryotic cells, for example free-living pro-tists and the cells that line animal respiratory pas-sages, and that have the power of movement. cilia assist in the movement of protists such as Parame-cium and some small aquatic animals. Flagella are whiplike structures with which many eukaryotic cells can swim, for example protists like Euglena and sperm cells. Many bacteria also have flagella with which they propel themselves, but the smaller rotary structure of bacterial flagella is totally unlike the more complex structure of eukaryotic flagella, which is why Margulis does not use the same term for both of them.
Margulis claims that the protein strands within undulipodia are the remnants of spirochete bacteria. How might this have occurred? today, there are examples of spirochete bacteria that burrow part-way into protist cells. the protruding parts of the bacteria flap in a coordinated rhythm, propelling the protist cell. these protists, when viewed under the microscope, look just like ciliated or flagellated cells; one must look closely and watch the process of their formation to realize that they are not cilia at all but bacteria! in other words, spirochetes are, today, doing the very thing that Margulis claimed they did back when undulipodia came into existence.
another structure that may have had a symbio-genetic origin is the hydrogenosome, a structure that metabolizes hydrogen and is found in some protist cells. in some cases, the genes that control hydro-genosome activity are all together in the host cell nucleus and resemble bacterial rather than eukary-otic genes, as if they were transposed to the nucleus from the ancestral hydrogenosome, which may have been a mitochondrion or a bacterium.
Margulis reconstructs the history of eukaryotic cells in four stages, which she calls Serial endosym-biotic theory:
1. the nucleus had a bacterial origin. the most likely ancestor for the nucleus is an archae-bacterium similar to modern Thermoplasma. Some archaebacteria have proteins that resemble the histones associated with the Dna in eukaryotic chromosomes. the ances-tral cell was now a eukaryote, with a nucleus.
2. Spirochetes associated with this cell and eventually became undulipodia.
3. aerobic bacteria moved into some of these cells and eventually became mitochondria. Some of the protists that had mitochondria became the ancestors of animals, fungi, and plants.
4. Photosynthetic cyanobacteria were con-sumed, but not digested, by cells that already had mitochondria, and eventually became chloroplasts. Some of the protists that had chloroplasts became the ancestors of plants.
One of the strongest proofs of symbiogenesis is the fact that the structures now called chloroplasts have originated more than once. Most chloroplasts, like those of all terrestrial plants, resemble bac-teria and are surrounded by a simple membrane. But the chloroplasts of some protists, such as dino-flagellates, have multiple membranes. this is also true of the apicoplast of the Plasmodium falciparum protist that causes malaria, which is a degenerate, multiple-membraned chloroplast. the Plasmodium does not use its apicoplasts for photosynthesis. the multiple membranes of these chloroplasts and apico-plasts suggest that they evolved when photosynthetic eukaryotic cells invaded other eukaryotic cells, stayed there, and simplified. in other words, regular chloroplasts are degenerate cells inside a cell; dino-flagellate chloroplasts and plasmodial apicoplasts represent degenerate cells inside of degenerate cells inside of a cell! there is even microscopic evidence that these chloroplasts have nucleomorphs, which appear to be small, vestigial nuclei.
a dramatic experimental demonstration of sym-biogenesis (albeit unintended) occurred when biologist Kwang Jeon of the University of tennessee at Knox-ville noticed that his cultures of the protist Amoeba proteus looked sick. Under the microscope, he could see that each amoeba was infected with thousands of rod-shaped bacteria. the amoebae usually digested bacteria, but these bacteria could resist the digestive enzymes of the amoebae. Most of the amoebae died from the bacterial infection. However, a few amoebae could resist the bacteria and survive. Jeon cultured the survivors over and over. He was artificially selecting for the amoebae that could best resist the bacterial infection, as well as for the bacteria that had the mildest effects on the amoebae. after about a year and a half, the amoebae and bacteria had developed
xviii+598_Biodiversity_F.indd 509 11/17/11 1:13 PM

first entry510 symbiosis 510
a mutually beneficial relationship. after several years, the coevolution of amoebae and bacteria had pro-gressed so far that neither of the species could survive without molecules supplied by the other. they had become an essential part of each other’s environment. Might the amoebae now be considered a new species of protist? Might the bacteria now be considered a new species as well? Perhaps they could even be con-sidered one new species of organism?
there are many examples of symbiogenetic pro-cesses that involve bacteria and insects. the bacte-rium Buchnera aphidicola infects aphids. like an organelle, the Buchnera bacterium can be transmit-ted from one generation to the next through eggs. Because Buchnera manufactures certain amino acids that aphids cannot get from the sap that they eat, it can be considered mutualistic. the bacterium Cinara cedri does a similar thing. the endosymbi-otic bacterium Carsonella ruddii has a set of Dna only about a quarter as large as that found in plant mitochondria. about 10 percent of insect species have endosymbiotic bacteria with reduced genomes that produce nutrients upon which they depend. Did mitochondria begin their evolution in a similar way?
evolutionary biologists noriko Okamoto and isao inouye have found evidence of a secondary symbiosis that appears to be in progress. there is a green-algal symbiont that lives in certain protist host cells. the symbiont still has a nucleus, mitochondria, and a large green plastid, but the flagella, cytoskel-eton, and internal membranes are gone. When the symbiont divides, it produces two cells: one with the plastid and one without. the cell without the green plastid then develops a feeding apparatus and engulfs the cell with the green plastid. this may rep-resent a direct observation of a “missing link” in the evolution of chloroplasts by symbiosis.
the symbiogenetic event that produced chloro-plasts and other similar structures was not a single, freak event; symbiogenesis cannot, therefore, be safely ignored in evolutionary theory as a weird aberration. Symbiogenesis has occurred, and continues to occur, a lot. in fact, according to Margulis, it occurs so often that earth should be considered a “symbiotic planet.” the worldwide tendency of organisms to associate with one another may, according to Margulis, help to explain why earth operates as a single system.
Symbiogenesis can occur more than once even in a single cell. One of the best examples is a protist that lives in the intestines of termites in australia. this protist appears to have several different types of undulipodia. actually, they are four different kinds of spirochetes that form a symbiotic association with the protist. and the symbiosis does not end there. the protist itself is a symbiont inside of termite intestines, and it helps the termites to digest wood. termites
could not survive if they did not have protists living in their guts that aid with the digestion of wood.
Whenever coevolution produces symbiosis, new opportunities for evolution result. Small mutualists living inside of larger cells may lose some of their genes to the host cell. When this occurs, symbiogen-esis may very quickly produce new biodiversity.
See also coevolution; eukaryotes; Margu-lis, lynn; natural selection; protists.
Further readingakhmanova, a., et al., “a hydrogenosome with a genome.”
Nature 396 (1998): 527–528.Boxma, B., et al. “an anaerobic mitochondrion that pro-
duces hydrogen.” Nature 434 (2005): 74–79.chapman, Michael J., Michael F. Dolan, and lynn Margu-
lis. “centrioles and kinetosomes: Form, function, and evolution.” Quarterly Review of Biology 75 (2000): 409–429.
Douglas, Susan, et al. “the highly reduced genome of an enslaved algal nucleus.” Nature 410 (2001): 1,091–1,096.
Dyall, Sabrina, Mark t. Brown, and Patricia J. Johnson. “ancient invasions: From endosymbionts to organ-elles.” Science 304 (2004): 253–257.
Falkowski, Paul G., et al. “the evolution of modern eukary-otic phytoplankton.” Science 305 (2004): 354–727.
Jeon, K. W. “integration of bacterial endosymbionts in amoebae.” International Review of Cytology, Suppl. 14 (1983): 29–47.
Margulis, lynn. Symbiotic Planet: A New Look at Evolu-tion. new york: Basic Books, 1998.
———. “Serial endosymbiotic theory (Set) and com-posite individuality: transition from bacterial to eukaryotic genomes.” Microbiology Today 31 (2004): 172–174.
Margulis, lynn, and R. Fester, eds. Symbiosis as a Source of Evolutionary Innovation. cambridge, Mass.: Mit Press, 1991.
Margulis, lynn, and Dorion Sagan. “the beast with five genomes.” Natural History, June 2001, 38–41.
———. Acquiring Genomes: A Theory of the Origins of Species. new york: Basic Books, 2002.
nakabachi, natsushi, et al. “the 100-kilobase genome of the bacterial endosymbiont Carsonella.” Science 314 (2006): 267.
Okamoto, noriko, and isao inouye. “a secondary symbio-sis in progress?” Science 310 (2005): 287.
Pérez-Brocal, Vicente, et al. “a small microbial genome: the end of a long symbiotic relationship?” Science 314 (2006): 312–313. Summarized by andersson, Siv G. e. “the bacterial world gets smaller.” Science 314 (2006): 259–260.
symbiosis See coevolution; commensalism; mutualism; parasitism.
xviii+598_Biodiversity_F.indd 510 11/17/11 1:13 PM

511
temperate coniferous forests the temper-ate coniferous forests are found primarily in mod-erately dry mountainous regions. they have cold, snowy winters and warm summers. they receive less summer rain than the temperate deciduous forests, and this is one important reason that the temper-ate coniferous forests are found in drier regions of western north america while deciduous forests are found in the wetter regions of the east. they receive more rain than the temperate dry evergreen oak forests.
in western north america and Mexico, three zones within the temperate coniferous forests may be distinguished (see the following table):
● the driest coniferous forests are the piñon-juniper woodlands, which are found on the ranges in the Great Basin and on the slopes of the Rockies and the Sierra Madre.
● coniferous forests that receive a little more precipitation are mostly dominated by pon-derosa pines in the Great Basin and the Rockies, arizona pines in the Sierra Madre, and Jeffrey pines in california.
● the temperate coniferous forests with the greatest amount of precipitation are domi-nated by trees such as Douglas fi rs, white fi rs, lodgepole pines, and sugar pines.
temperate coniferous forests are also found throughout europe and asia. Small patches of hardwood trees may occur in temperate coniferous forests. in the Black Hills of South Dakota and Wyo-ming, bur oaks (Quercus macrocarpa) may grow in moist microhabitats. Understory hardwood trees include chokecherry (Prunus virginiana).
temperate coniferous forests intergrade into boreal and subalpine forests. Some trees, such as the Scots pine (Pinus sylvestris) and the norway spruce (Picea abies), grow in boreal forests as well as temperate ones. temperate coniferous forests also intergrade into temperate deciduous forests. Many species of pines are abundant in moist deciduous forests. examples include white pines (P. strobus)and hemlocks (Tsuga canadensis) in eastern north america and Japanese black pines (P. thunbergii) in asia. in dry regions, such as western north amer-ica, they can intergrade into cool desert sagebrush.
in some moist coves of the Pacifi c slope of the Sierra nevada, giant sequoia trees (Sequoiadendron giganteum) dominate the temperate coniferous for-ests. On the average, giant sequoias are the larg-est trees in the world. Many of them live more than 2,000 years. Because of their thick bark, giant sequoias survive fi res that kill many other trees. these same fi res create the sunny conditions in which sequoia seedlings grow.
See also conifers; temperate deciduous forests; temperate dry forests.
Further readingcoultrap, D. e., et al. “Relationships between western
juniper (Juniperus occidentalis) and understory veg-etation.” Invasive Plant Science and Management 1 (2008): 3–11.
Delucia, evan H., and William H. Schlesinger. “Resource-use effi ciency and drought tolerance in adjacent Great Basin and Sierran plants.” Ecology 72 (1991): 51–58.
Rehfeldt, Gerald e., et al. “Physiologic plasticity, evolution, and impacts of a changing climate on Pinus contorta.” Climatic Change 50 (2001): 355–376.
T
xviii+598_Biodiversity_F.indd 511 11/17/11 1:13 PM
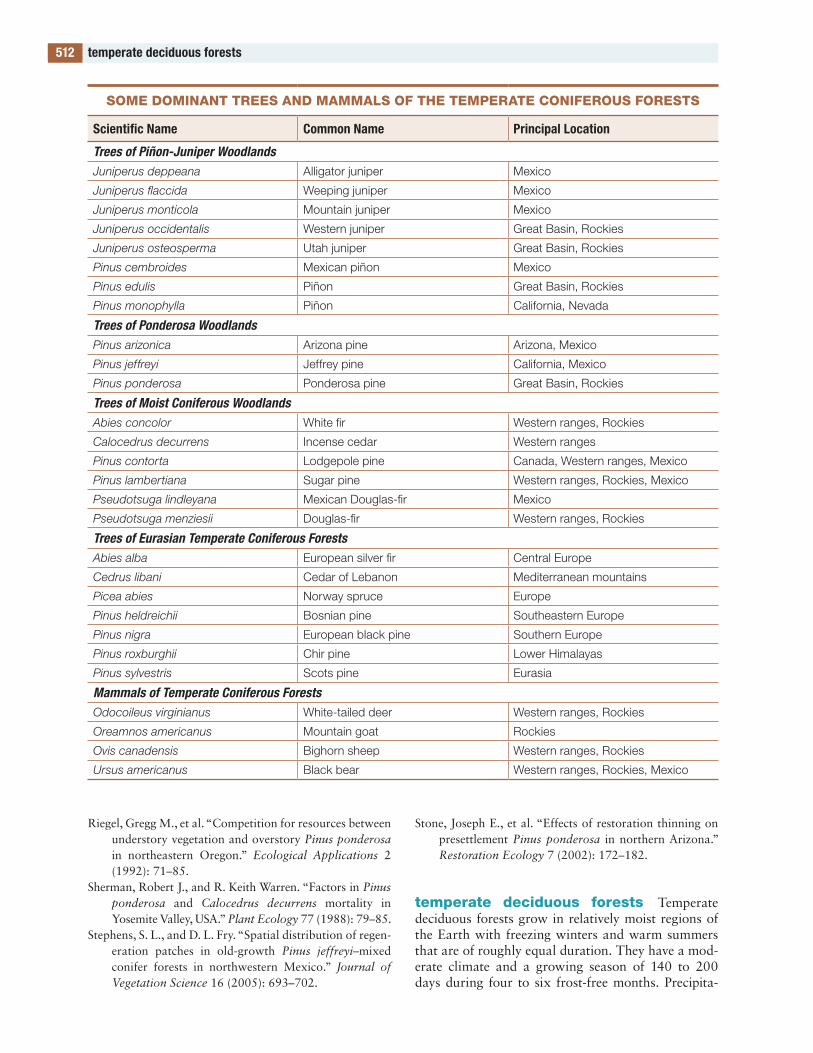
first entry512 temperate deciduous forests 512
Riegel, Gregg M., et al. “competition for resources between understory vegetation and overstory Pinus ponderosa in northeastern Oregon.” Ecological Applications 2 (1992): 71–85.
Sherman, Robert J., and R. Keith Warren. “Factors in Pinus ponderosa and Calocedrus decurrens mortality in yosemite Valley, USa.” Plant Ecology 77 (1988): 79–85.
Stephens, S. l., and D. l. Fry. “Spatial distribution of regen-eration patches in old-growth Pinus jeffreyi–mixed conifer forests in northwestern Mexico.” Journal of Vegetation Science 16 (2005): 693–702.
Stone, Joseph e., et al. “effects of restoration thinning on presettlement Pinus ponderosa in northern arizona.” Restoration Ecology 7 (2002): 172–182.
temperate deciduous forests temperate deciduous forests grow in relatively moist regions of the earth with freezing winters and warm summers that are of roughly equal duration. they have a mod-erate climate and a growing season of 140 to 200 days during four to six frost-free months. Precipita-
some dominant trees and mammals of the temperate Coniferous forests
Scientific Name Common Name principal location
Trees of Piñon-Juniper WoodlandsJuniperus deppeana Alligator juniper Mexico
Juniperus flaccida Weeping juniper Mexico
Juniperus monticola Mountain juniper Mexico
Juniperus occidentalis Western juniper Great Basin, Rockies
Juniperus osteosperma Utah juniper Great Basin, Rockies
Pinus cembroides Mexican piñon Mexico
Pinus edulis Piñon Great Basin, Rockies
Pinus monophylla Piñon California, Nevada
Trees of Ponderosa WoodlandsPinus arizonica Arizona pine Arizona, Mexico
Pinus jeffreyi Jeffrey pine California, Mexico
Pinus ponderosa Ponderosa pine Great Basin, Rockies
Trees of Moist Coniferous WoodlandsAbies concolor White fir Western ranges, Rockies
Calocedrus decurrens Incense cedar Western ranges
Pinus contorta Lodgepole pine Canada, Western ranges, Mexico
Pinus lambertiana Sugar pine Western ranges, Rockies, Mexico
Pseudotsuga lindleyana Mexican Douglas-fir Mexico
Pseudotsuga menziesii Douglas-fir Western ranges, Rockies
Trees of Eurasian Temperate Coniferous ForestsAbies alba European silver fir Central Europe
Cedrus libani Cedar of Lebanon Mediterranean mountains
Picea abies Norway spruce Europe
Pinus heldreichii Bosnian pine Southeastern Europe
Pinus nigra European black pine Southern Europe
Pinus roxburghii Chir pine Lower Himalayas
Pinus sylvestris Scots pine Eurasia
Mammals of Temperate Coniferous ForestsOdocoileus virginianus White-tailed deer Western ranges, Rockies
Oreamnos americanus Mountain goat Rockies
Ovis canadensis Bighorn sheep Western ranges, Rockies
Ursus americanus Black bear Western ranges, Rockies, Mexico
xviii+598_Biodiversity_F.indd 512 11/17/11 1:13 PM

temperate deciduous forests 513
tion is usually distributed evenly throughout the year, ranging from 39 to 59 inches (75 to 150 cm), falling as snow in winter and rain in summer. Soils are typi-cally fertile and enriched with decaying litter (humus). temperate forests occur in eastern north america, northeastern asia, and western and central europe.
the dominant tree species shed their leaves every autumn (see figure). if they did not do so, a heavy weight of snow would accumulate on the broad leaves and damage the branches. they produce new leaves every spring. trees of boreal forests do not shed their leaves each autumn, primarily because the boreal summers are so short that there is insuf-ficient time for a new crop of leaves to grow every year. trees of tropical forests do not seasonally shed their leaves, primarily because tropical regions have year-round summer. Deciduous forests, therefore, live in climatic zones with winters that are interme-diate in length between those of boreal and tropical zones. the broad leaves of the deciduous trees have higher rates of photosynthesis than do needle-shaped leaves. trees of boreal forests have needle-shaped leaves, despite their lower photosynthesis, because needle-shaped leaves are better than broad leaves at surviving the winter. needle leaves result in less accumulation of snow than broad leaves. the leaves of deciduous trees are adapted to rapid photosynthe-sis in the summer, and the leaves of boreal trees are adapted to surviving the winter.
BIodIvErSITy IN dECIduouS forESTSDeciduous forests do not have as much biodiversity as the tropical rain forests, for reasons such as the following:
Winter and wind. as in the rain forests, a great deal of coevolution occurs between plants and their
herbivores and their pollinators. Selection for chemi-cal defenses against herbivores, which can lead to speciation, is less intense in the deciduous forest because many insect populations are reduced by win-ter death. Selection for pollinator specialization is not as strong because many temperate plants, especially dominant forest trees, depend on wind pollination.
Broad ranges of tolerance. Because each spe-cies has to tolerate wide fluctuations of temperature each year, and often moisture as well, each species is able to grow over a more extensive geographical range than is often the case with rain forest species.
Specialization on microhabitats. Deciduous for-est species are able to specialize within their habitat. For example, when the snow melts in the spring, sunlight penetrates the bare branches and warms the ground, allowing shrubs to burst their buds and spring ephemeral wildflowers to grow. these wildflowers, such as the spring beauty (Claytonia virginica), are perennials that resprout each year from underground food storage organs. Meanwhile, the wind remains chilly and the buds of the trees remain dormant. For about a month, the spring ephemerals receive abundant sunlight and relative warmth. Once the tree buds begin to open, casting shade, the ephemerals begin to senesce, until the next year. then the shade-adapted summer wild-flowers grow and dominate the forest floor. there are also lianas, such as grapes, in deciduous for-ests, as in rain forests. Drier conditions, however, make epiphytes (plants whose roots obtain moisture and minerals from accumulated leaf litter up in the branches of trees) very rare, in contrast to their tropical exuberance.
Deciduous forests contain a few evergreen spe-cies. Some, such as hemlocks (Tsuga canadensis), are trees. Many, like the hollies (Ilex opaca and I. americana), are shrubs. they may be protected from winter conditions by the larger deciduous trees around them, and they make use of the sunlight of the early spring before the deciduous trees have opened their leaves. the deciduous forests of asia, having been less disrupted by the ice ages than the forests of north america and europe, have many coniferous tree species, including the dawn redwood (Metasequoia glyptostroboides) and the Japanese cypress (Cryptomeria japonica).
Within the deciduous forest zone, there are dif-ferences in climatic conditions, and, therefore, species composition; the deciduous forest species have evolved separately in north america, europe, and asia for millions of years, as indicated in the table on pages 514–515. the major north american deciduous for-ests are the following:
● the beech-maple forests of north america are dominated by american beeches (Fagus
Deciduous forests are renowned for their autumn colors. Not all the colorful deciduous trees are broad-leafed trees: The bright red trees growing in the water at Beaver’s Bend State Park in Oklahoma are bald cypresses (Taxodium distichum). (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 513 11/17/11 1:13 PM
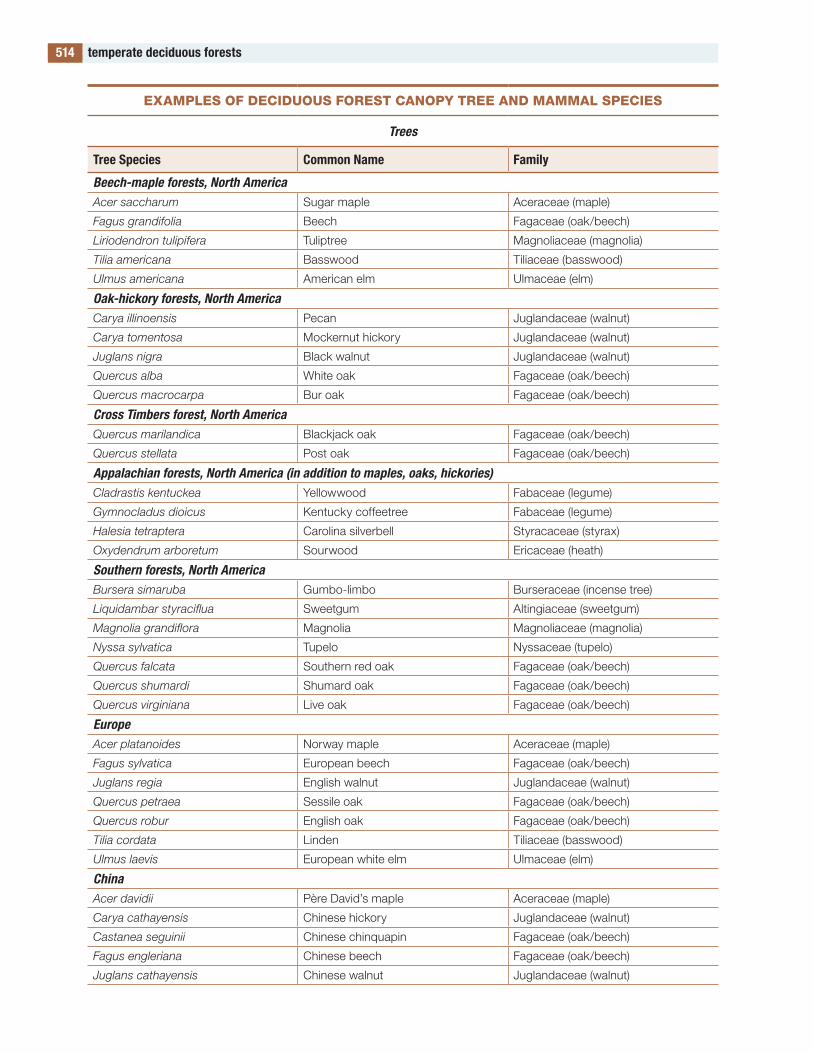
first entry514 temperate deciduous forests 514
examples of deCiduous forest Canopy tree and mammal speCies
Trees
Tree Species Common Name family
Beech-maple forests, North AmericaAcer saccharum Sugar maple Aceraceae (maple)
Fagus grandifolia Beech Fagaceae (oak/beech)
Liriodendron tulipifera Tuliptree Magnoliaceae (magnolia)
Tilia americana Basswood Tiliaceae (basswood)
Ulmus americana American elm Ulmaceae (elm)
Oak-hickory forests, North AmericaCarya illinoensis Pecan Juglandaceae (walnut)
Carya tomentosa Mockernut hickory Juglandaceae (walnut)
Juglans nigra Black walnut Juglandaceae (walnut)
Quercus alba White oak Fagaceae (oak/beech)
Quercus macrocarpa Bur oak Fagaceae (oak/beech)
Cross Timbers forest, North AmericaQuercus marilandica Blackjack oak Fagaceae (oak/beech)
Quercus stellata Post oak Fagaceae (oak/beech)
Appalachian forests, North America (in addition to maples, oaks, hickories)Cladrastis kentuckea Yellowwood Fabaceae (legume)
Gymnocladus dioicus Kentucky coffeetree Fabaceae (legume)
Halesia tetraptera Carolina silverbell Styracaceae (styrax)
Oxydendrum arboretum Sourwood Ericaceae (heath)
Southern forests, North AmericaBursera simaruba Gumbo-limbo Burseraceae (incense tree)
Liquidambar styraciflua Sweetgum Altingiaceae (sweetgum)
Magnolia grandiflora Magnolia Magnoliaceae (magnolia)
Nyssa sylvatica Tupelo Nyssaceae (tupelo)
Quercus falcata Southern red oak Fagaceae (oak/beech)
Quercus shumardi Shumard oak Fagaceae (oak/beech)
Quercus virginiana Live oak Fagaceae (oak/beech)
EuropeAcer platanoides Norway maple Aceraceae (maple)
Fagus sylvatica European beech Fagaceae (oak/beech)
Juglans regia English walnut Juglandaceae (walnut)
Quercus petraea Sessile oak Fagaceae (oak/beech)
Quercus robur English oak Fagaceae (oak/beech)
Tilia cordata Linden Tiliaceae (basswood)
Ulmus laevis European white elm Ulmaceae (elm)
ChinaAcer davidii Père David’s maple Aceraceae (maple)
Carya cathayensis Chinese hickory Juglandaceae (walnut)
Castanea seguinii Chinese chinquapin Fagaceae (oak/beech)
Fagus engleriana Chinese beech Fagaceae (oak/beech)
Juglans cathayensis Chinese walnut Juglandaceae (walnut)
xviii+598_Biodiversity_F.indd 514 11/17/11 1:13 PM
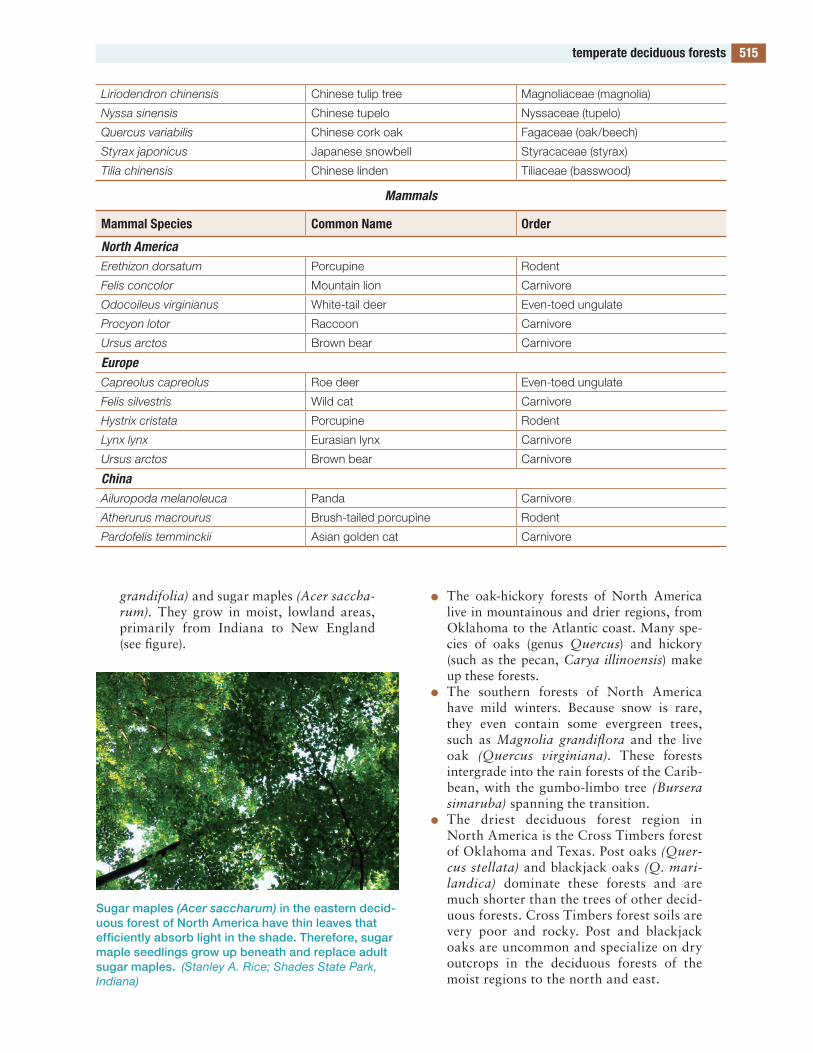
temperate deciduous forests 515
grandifolia) and sugar maples (Acer saccha-rum). they grow in moist, lowland areas, primarily from indiana to new england (see figure).
● the oak-hickory forests of north america live in mountainous and drier regions, from Oklahoma to the atlantic coast. Many spe-cies of oaks (genus Quercus) and hickory (such as the pecan, Carya illinoensis) make up these forests.
● the southern forests of north america have mild winters. Because snow is rare, they even contain some evergreen trees, such as Magnolia grandiflora and the live oak (Quercus virginiana). these forests intergrade into the rain forests of the carib-bean, with the gumbo-limbo tree (Bursera simaruba) spanning the transition.
● the driest deciduous forest region in north america is the cross timbers forest of Oklahoma and texas. Post oaks (Quer-cus stellata) and blackjack oaks (Q. mari-landica) dominate these forests and are much shorter than the trees of other decid-uous forests. cross timbers forest soils are very poor and rocky. Post and blackjack oaks are uncommon and specialize on dry outcrops in the deciduous forests of the moist regions to the north and east.
Liriodendron chinensis Chinese tulip tree Magnoliaceae (magnolia)
Nyssa sinensis Chinese tupelo Nyssaceae (tupelo)
Quercus variabilis Chinese cork oak Fagaceae (oak/beech)
Styrax japonicus Japanese snowbell Styracaceae (styrax)
Tilia chinensis Chinese linden Tiliaceae (basswood)
Mammals
mammal Species Common Name order
North AmericaErethizon dorsatum Porcupine Rodent
Felis concolor Mountain lion Carnivore
Odocoileus virginianus White-tail deer Even-toed ungulate
Procyon lotor Raccoon Carnivore
Ursus arctos Brown bear Carnivore
EuropeCapreolus capreolus Roe deer Even-toed ungulate
Felis silvestris Wild cat Carnivore
Hystrix cristata Porcupine Rodent
Lynx lynx Eurasian lynx Carnivore
Ursus arctos Brown bear Carnivore
ChinaAiluropoda melanoleuca Panda Carnivore
Atherurus macrourus Brush-tailed porcupine Rodent
Pardofelis temminckii Asian golden cat Carnivore
Sugar maples (Acer saccharum) in the eastern decid-uous forest of North America have thin leaves that efficiently absorb light in the shade. Therefore, sugar maple seedlings grow up beneath and replace adult sugar maples. (Stanley A. Rice; Shades State Park, Indiana)
xviii+598_Biodiversity_F.indd 515 11/17/11 1:13 PM

first entry516 temperate deciduous forests 516
european forests have evolved separately from north american forests for at least 50 million years. european forests have maples, beeches, and oaks, just as do american deciduous forests, but not the same species. the same is true of the deciduous forests of china and Japan, whose species also dif-fer from one another. Fagus crenata, the Japanese beech, is not found in china.
the deciduous forests of north america, europe, and asia also contain similar types of mam-mals, although usually different species of each (see table on pages 514–515). the brown bear Ursus arc-tos lives in both europe and north america.
Before north america separated from europe, and before the ice ages began, the deciduous for-ests formed a more or less continuous band across the northern continents. ancestral populations of forest trees and other plants evolved separately in these places, but the large animals continued to migrate across the Bering land bridge until the ice ages began, and in some cases until the end of the most recent ice age.
the north american deciduous forests with the greatest diversity of species are found in the south-ern appalachians, where abundant rainfall and mild winters allow the coexistence of hundreds of tree species from both the beech-maple and oak-hickory forests, and many species, such as Kentucky coffee-tree (Gymnocladus dioicus), which are not abun-dant anywhere else.
Many species of deciduous trees grow rapidly and specialize on growing in disturbed areas before they are eventually replaced by the dominant tree species. examples include black cherry (Prunus sero-tina), cottonwood (Populus deltoides), and syca-more (Platanus occidentalis).
chestnut trees (Castanea americana) once dom-inated the oak-hickory forests, until a disease, the chestnut blight, was introduced into new york about 1900. this fungal disease killed all adult chestnut trees, except for very rare isolated stands, reaching Georgia by the 1950s. today, chestnuts can be found only as resprouts from surviving rootstocks.
today, only scattered remnants of original old-growth temperate forests remain. Because of their proximity to major centers of civilization, most of these forests were cut early in civilization: the Medi-terranean forests before 2000 b.c.e., the central european forests by about 1000 c.e., and the north american deciduous forests by about 1900. Many of these forests have grown back, though with smaller trees and lower diversity of tree species. Hemlocks and maples have grown back where there were once farms in new england, but most modern people will never see examples of the huge trees that the first european settlers encountered on the north
american continent. Since 1970, however, decidu-ous forest area in the United States has declined by about 4 percent, mainly due to the expansion of suburbs and to mountaintop-removal coal mining.
the animal that had perhaps the most signifi-cant impact on these forests immediately before white american settlement was the passenger pigeon. When white americans were traveling and homesteading across the eastern forest, they saw flocks of billions of passenger pigeons. they assumed that this is the way it had always been. Recent scholarship has suggested, however, that hunting by native americans had kept the pigeon populations under control for thousands of years, and the pigeon population exploded only after dis-eases such as smallpox reduced native populations by as much as 90 percent. a similar explanation has been offered for why there were so many mil-lions of bison in the prairies just prior to white american expansion. according to the earliest explorers who visited native tribes prior to their decimation by smallpox, native americans exten-sively transformed the landscape by agriculture and fire. though they farmed and burned much of what the poet Henry Wadsworth longfellow called the “primeval forest,” they had only stone tools and did not cut down the largest trees.
Perhaps the most extensive deciduous forest in the world that has largely escaped recent destruc-tion lies on the border of Belarus (where it is called Belovezhskaya Pushcha) and Poland (where it is called Białowieża Puszcza). as in appalachian and asian forests, there are some abundant conifers, such as norway spruce and Scotch pine. the decidu-ous tree species include alders, oaks, birches, ashes, basswoods, maples, aspen, elms, and hornbeams. Many of the trees are quite large. this forest is the home of some of the last wisent (Bison bonasus) herds, which are separated by the fence that divides Belarus and Poland. Despite the centuries of protec-tion that this forest has received, it remains exten-sively disturbed by humans; the average age of a forest stand in the protected zone is only 130 years.
See also temperate coniferous forests; temperate dry forests.
Further readingabrams, Marc D. “the red maple paradox.” BioScience 48
(1998): 355–364.association for ecotourism Development (Poland). “in the
heart of Bialowieza national Park.” available online. URl: http://www.puszczabialowieska.org/eng/flora.htm. accessed July 25, 2010.
Bonnicksen, thomas M. America’s Ancient Forests: From the Ice Age to the Age of Discovery. new york: John Wiley, 2000.
xviii+598_Biodiversity_F.indd 516 11/17/11 1:13 PM

temperate dry forests 517
clebsch, edward e. c., and Richard t. Busing. “Secondary succession, gap dynamics, and community structure in a southern appalachian cove forest.” Ecology 70 (1989): 728–735.
Delcourt, Hazel R. Forests in Peril: Tracking Deciduous Trees from Ice-Age Refuges into the Greenhouse World. Granville, Ohio: McDonald and Woodward, 2002.
Duffy, David cameron, and albert J. Meier. “Do appalachian herbaceous understories ever recover from clearcut-ting?” Conservation Biology 6 (2003): 196–201.
Francaviglia, Richard V. The Cast Iron Forest: A Natural and Cultural History of the North American Cross Timbers. austin: University of texas Press, 2000.
Macleish, William H. The Day before America: Changing the Nature of a Continent. new york: Houghton Mif-flin, 1994.
Mann, charles c. 1491: New Revelations of the Americas before Columbus. new york: Knopf, 2005.
Mccormick, J. Frank, and Robert B. Platt. “Recovery of an appalachian forest following the chestnut blight or catherine Keever—you were right!” American Mid-land Naturalist 104 (1980): 264–273.
Parker, G. R., et al. “tree dynamics in an old-growth, deciduous forest.” Forest Ecology and Management 11 (1985): 31–57.
Rice, Stanley a. “Plants create habitats.” chap. 8 in Green Planet: How Plants Keep the Earth Alive. Piscataway, n.J.: Rutgers University Press, 2009.
Whigham, Dennis F. “ecology of woodland herbs in tem-perate deciduous forests.” Annual Review of Ecology, Evolution, and Systematics 35 (2004): 583–622.
yahner, Richard H. Eastern Deciduous Forests: Ecology and Wildlife Conservation, 2nd ed. Minneapolis: Uni-versity of Minnesota Press, 2000.
temperate dry forests temperate dry for-ests consist of evergreen broadleaf trees. the sum-mers are hot and dry, and the winters are cool and wet. these conditions are usually called “Mediter-ranean” because they are widespread in southern europe, the Middle east, and north africa. these conditions are also found in california, chile, South africa, and australia. the trees are short and seldom form a continuous canopy. temperate dry forests are also found in the dry mountains of northern Mexico, where, because of summer rain-fall, the climate is not Mediterranean. african dry forests are subtropical and are considered tropical seasonal forests.
the dry evergreen forests of the Mediterra-nean Basin, california, and Mexico are dominated by oaks (see table). the dry evergreen forests of australia are dominated by eucalypts. Unlike the other dry forests here mentioned, eucalypt forests regenerate abundantly after fires. Moist forests
in the Mediterranean Basin and california con-tain oak species as well, some of them deciduous. Moist forests in australia are also dominated by evergreen eucalypts, but they grow taller. Pines may grow among or rise above the small evergreen oaks, such as Pinus halepensis and P. pinea in the Mediterranean, and P. coulteri and P. sabiniana in california.
temperate dry forests intergrade with other eco-logical communities, such as the following:
● Dry forests have a grass layer and may inter-grade into grasslands.
● chaparral, matorral, and other fire-cycle shrublands are often adjacent to dry forests. in such cases, fires maintain the shrublands at the expense of the forests. Some oak trees
some temperate dry forest trees
Scientific Name Common Name
Mediterranean
Quercus calliprinos Palestine oak
Quercus coccifera Kermes oak
Quercus ilex Holm oak
Quercus suber Cork oak
California
Quercus agrifolia Coast live oak
Quercus chrysolepis Canyon live oak
Quercus dumosa Scrub oak
Quercus englemannii Englemann oak
Quercus wislizeni Interior live oak
Mexico
Quercus arizonica Arizona white oak
Quercus chihuahuaensis Chihuahua oak
Quercus emoryi Emory oak
Quercus rugosa Netleaf oak
Australia
Corymbia (Eucalyptus) calophylla
Marri
Eucalyptus globulus Tasmanian blue gum
Eucalyptus marginata Jarrah
Eucalyptus radiata Narrow-leaved peppermint
xviii+598_Biodiversity_F.indd 517 11/17/11 1:13 PM

first entry518 temperate rain forests 518
are found both in dry forests and shrub-lands. examples include Quercus wislizeni and Q. dumosa in california and Q. coc-cifera in the Middle east.
● in Mexico, temperate dry forests may inter-grade with temperate coniferous forests. Mexican piñon (Pinus cembroides) domi-nate the drier coniferous forests, while the ocote blanco (P. leiophylla) and Mexican subspecies of sugar pine (P. lambertiana) and ponderosa pine (P. ponderosa) domi-nate the coniferous forests that receive more rainfall and snow.
● in california, temperate dry forests inter-grade with a mixture of deciduous broad-leaf trees (such as the california black oak, Quercus kelloggii; the blue oak, Q. doug-lasii; the valley oak, Q. lobata; and the tanbark oak, Lithocarpus densiflorus) and conifers (such as ponderosa pine, Pinus pon-derosa; Douglas fir, Pseudotsuga menziesii; Santa lucia fir, Abies bracteata; and incense cedar, Calocedrus decurrens).
● in the Mediterranean, temperate dry forests may intergrade with temperate deciduous forests, often dominated by deciduous oaks, such as Quercus robur.
● Dry eucalypt forests in australia intergrade into temperate rain forests dominated by other eucalypts and by kauri trees (Agathis australis).
temperate dry forests often grow on dry, nutri-ent-poor soil. the dry conditions and the thick ever-green leaves discourage rapid decomposition and humus formation.
the similarity in climatic conditions between australia, california, and new Zealand has allowed an exchange of invasive species. Railroad workers planted Eucalyptus globulus from australia in cali-fornia in the 19th century, after which it took over large areas that were formerly shrubland and grass-land. the native range of the Monterey pine (Pinus radiata) is just three small groves in california, but after being planted in new Zealand, it displaced much of the native forest.
Unlike tropical seasonal forests and savannas in africa, the dry evergreen forests are not well known for wildlife. White-tailed deer (Odocoileus virginianus) and black bears (Ursus americanus) live in the dry forests in california and Mexico. Many marsupials, most famously the koalas (Phasolarc-tos cinereus), live in dry eucalyptus forests of aus-tralia. thousands of years of human contact have eliminated many game animals from Mediterranean forests. However, cork oak forests in north africa
are the home of the endangered Barbary macaque (Macaca sylvanus).
See also invasive species; shrublands; tem-perate coniferous forests; temperate decidu-ous forests.
temperate rain forests temperate rain for-ests are cool coastal forests that receive moisture, much of it in the form of fog, all year. Because the winters are mild, the dominant trees are evergreen conifers or broadleaf trees. Because the air is usually humid, water can rise to the tops of very tall trees. the three tallest tree species in the world—the coast redwood, the Douglas fir, and the Sitka spruce—all grow in the temperate rain forest of the Pacific northwest of north america. the tallest tree in the world is a coast redwood (Sequoia sempervirens) 378 feet (116 m) tall. the range of these redwoods extends inland only as far as the coastal fog. the almost continuous moisture also allows many epi-phytes, such as ferns and mosses, to grow in the branches of the large conifers and broadleaf trees. Some temperate rain forests in australia and taiwan are in coastal mountains.
Because of thick undergrowth and wet soil, seed-lings often germinate more readily on fallen logs (“nurse logs”) than on the ground. long after the log has rotted away, the roots of large trees still form a tunnel formerly occupied by the log (see photograph).
the relatively mild, wet conditions of temperate rain forests resemble climate conditions that were widespread earlier in the cenozoic era. they have therefore served as refugia for tree species, especially conifers other than pines, which used to be much more widespread in temperate regions. the domi-nant broadleaf trees of temperate rain forests tend to be relatives of oaks (tanbark oaks, southern beeches)
These trees in the Olympic rain forest in Washington began their growth on a log that decomposed long ago. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 518 11/17/11 1:13 PM
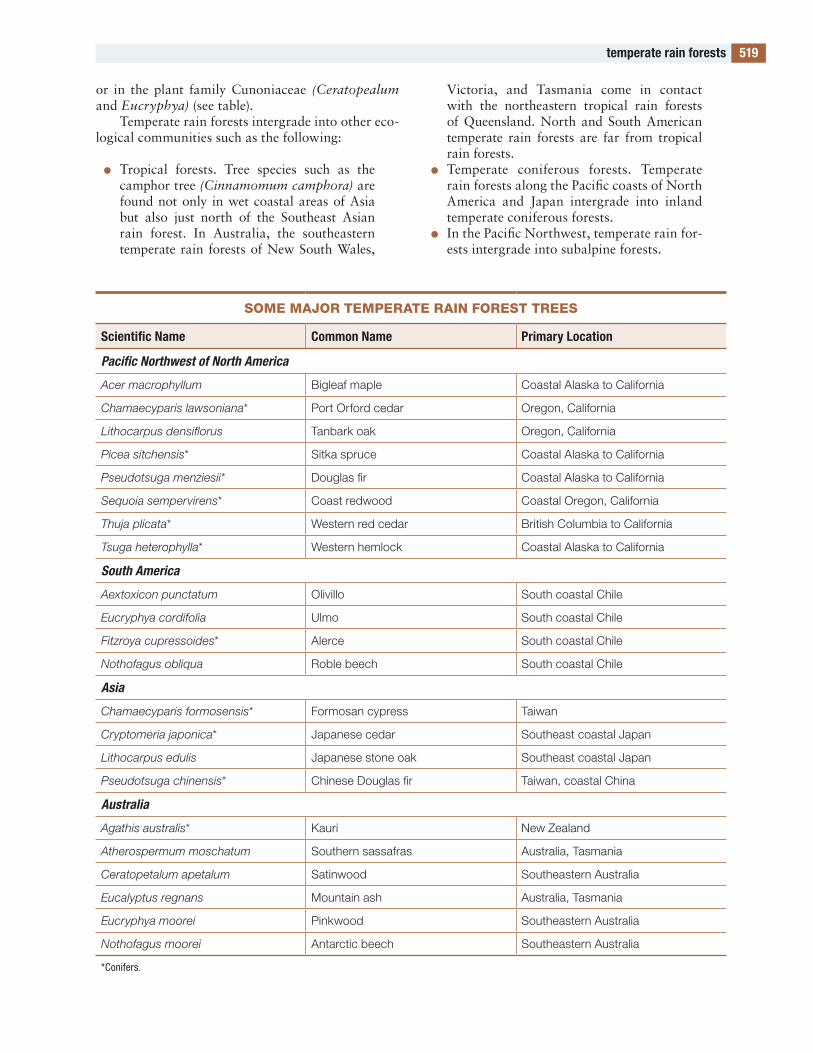
temperate rain forests 519
or in the plant family cunoniaceae (Ceratopealum and Eucryphya) (see table).
temperate rain forests intergrade into other eco-logical communities such as the following:
● tropical forests. tree species such as the camphor tree (Cinnamomum camphora) are found not only in wet coastal areas of asia but also just north of the Southeast asian rain forest. in australia, the southeastern temperate rain forests of new South Wales,
Victoria, and tasmania come in contact with the northeastern tropical rain forests of Queensland. north and South american temperate rain forests are far from tropical rain forests.
● temperate coniferous forests. temperate rain forests along the Pacific coasts of north america and Japan intergrade into inland temperate coniferous forests.
● in the Pacific northwest, temperate rain for-ests intergrade into subalpine forests.
some major temperate rain forest trees
Scientific Name Common Name primary location
Pacific Northwest of North America
Acer macrophyllum Bigleaf maple Coastal Alaska to California
Chamaecyparis lawsoniana* Port Orford cedar Oregon, California
Lithocarpus densiflorus Tanbark oak Oregon, California
Picea sitchensis* Sitka spruce Coastal Alaska to California
Pseudotsuga menziesii* Douglas fir Coastal Alaska to California
Sequoia sempervirens* Coast redwood Coastal Oregon, California
Thuja plicata* Western red cedar British Columbia to California
Tsuga heterophylla* Western hemlock Coastal Alaska to California
South America
Aextoxicon punctatum Olivillo South coastal Chile
Eucryphya cordifolia Ulmo South coastal Chile
Fitzroya cupressoides* Alerce South coastal Chile
Nothofagus obliqua Roble beech South coastal Chile
Asia
Chamaecyparis formosensis* Formosan cypress Taiwan
Cryptomeria japonica* Japanese cedar Southeast coastal Japan
Lithocarpus edulis Japanese stone oak Southeast coastal Japan
Pseudotsuga chinensis* Chinese Douglas fir Taiwan, coastal China
Australia
Agathis australis* Kauri New Zealand
Atherospermum moschatum Southern sassafras Australia, Tasmania
Ceratopetalum apetalum Satinwood Southeastern Australia
Eucalyptus regnans Mountain ash Australia, Tasmania
Eucryphya moorei Pinkwood Southeastern Australia
Nothofagus moorei Antarctic beech Southeastern Australia
*Conifers.
xviii+598_Biodiversity_F.indd 519 11/17/11 1:13 PM

first entry520 Thoreau, henry david520
there are cloud forests in some mountainous areas in the tropical latitudes. they are treated in a separate entry in this encyclopedia.
temperate rain forests are not noted for large animals. Besides bears and foxes, the main keystone predator in the Pacific northwest rain forest is the spotted owl (Strix occidentalis). temperate rain for-ests, like tropical rain forests, have many animals that require continuous moisture, such as amphib-ians and mollusks. the banana slug (genus Arioli-max) is a famous inhabitant of the Pacific northwest rain forest.
the temperate rain forest of the Pacific north-west has been heavily logged, since the trees are so large and each log is worth a lot of money. Only about one-tenth of the old-growth northwest for-est remains. timber corporations have frequently insisted on logging the old-growth forests, even though they have for decades been raising (and boasting about) sustainable tree plantations. Protest-ers and loggers have sometimes had violent confron-tations. environmentalist Julia Butterfly Hill spent 738 days in one of the largest trees to prevent log-gers from cutting it down. the loggers spared the tree but a vandal attacked it later. the spotted owl, instead of the entire ecological community, became the focus of much of the controversy.
See also boreal and subalpine forests; cloud forests; temperate coniferous forests; temperate deciduous forests.
Further readingantos, Joseph a., and Donald B. Zobel. “Habitat rela-
tionships of Chamaecyparis nootkatensis in southern Washington, Oregon, and california.” Canadian Jour-nal of Botany 64 (1986): 1,898–1,909.
Bourne, Joel. “the super trees.” National Geographic, august 2009.
Burgess, S. S. O., and t. e. Dawson. “the contribution of fog to the water relations of Sequoia sempervi-rens (D. Don): Foliar uptake and prevention of dehy-dration.” Plant, Cell, and Environment 27 (2004): 1,023–1,034.
chen, Jiquan, et al. “Vegetation responses to edge environ-ments in old-growth Douglas-fir forests.” Ecological Applications 2 (1992): 387–396.
christy, e. Jennifer. “effect of root competition and shad-ing on growth of suppressed western hemlock (Tsuga heterophylla).” Plant Ecology 65 (1986): 21–28.
collins, Brian D., and David R. Montgomery. “Forest development, wood jams, and restoration of flood-plain rivers in the Puget lowland, Washington.” Res-toration Ecology 10 (2002): 237–247.
Dietrich, William. The Final Forest: The Battle for the Last Great Trees of the Pacific Northwest. new york: Pen-guin, 1993.
Fonda, R. W. “Forest succession in relation to river terrace development in Olympic national Park, Washington.” Ecology 55 (1974): 927–942.
Halpern, charles B., and thomas a. Spies. “Plant spe-cies diversity in natural and managed forests of the Pacific northwest.” Ecological Applications 5 (1995): 913–934.
Hill, Julia Butterfly. The Legacy of Luna: The Story of a Tree, a Woman, and the Struggle to Save the Red-woods. new york: Harper, 2001.
lyons, Betsy, nalini M. nadkarni, and Malcolm P. north. “Spatial distribution and succession of epiphytes on Tsuga heterophylla (western hemlock) in an old-growth Douglas-fir forest.” Canadian Journal of Bot-any 78 (2000): 957–968.
Preston, Richard. The Wild Trees: A Story of Passion and Daring. new york: Random House, 2008.
Rodríguez-cabal, Mariano a., et al. “Habitat fragmen-tation disrupts a plant-disperser mutualism in the temperate forest of South america.” Biological Con-servation 139 (2007): 195–202.
traveset, anna, and Mary F. Willson. “effect of birds and bears on seed germination of fleshy-fruited plants in temperate rain forests of southeast alaska.” Oikos 80 (1997): 89–95.
Willson, Mary F., et al. “avian communities of fragmented south-temperate rain forests in chile.” Conservation Biology 8 (2002): 508–520.
thoreau, henry david (1817–1862) American Naturalist, writer Henry David thoreau is widely revered as a “prophet of environmentalism” because the ideas he wrote and put into action appear to have been at least a century ahead of their time. He is less often mentioned as one of the leading scientists of early 19th-century america. He was an amateur scientist, but he laid the groundwork for the ecologi-cal study of seed dispersal and ecological succession. He collected data that constitute one of the earliest studies of the adjustment of organisms to chang-ing seasons. His data sets have been used by mod-ern scientists to document climate change that has occurred since his time.
David Henry thoreau (who never changed his legal name) was born on July 12, 1817, in concord, Massachusetts, where he spent his entire life. He attended Harvard University from 1833 to 1837. He studied classics, philosophy, and science, earn-ing but not bothering to accept a degree that Har-vard awarded to students for persistence rather than merit. He became a schoolteacher at the concord academy, run by his brother John, but was dis-missed from the faculty for refusing to administer corporal punishment. Several Massachusetts writers were transcendentalists, who believed that the world
xviii+598_Biodiversity_F.indd 520 11/17/11 1:13 PM

Thoreau, henry david 521
of nature reflected an inner spiritual reality that also manifests itself in the human mind. One of them, Ralph Waldo emerson, became his lifelong friend. For most of the period from 1841 to 1844, thoreau lived with emerson’s family and was the tutor of emerson’s children. thoreau’s father owned a pen-cil-making factory, where the son worked on and off for the rest of his life, in addition to odd jobs as sur-veyor and carpenter. thoreau invented an improved method of preparing graphite for pencils and might have been headed for wealth in the pencil business. He rejected this path when, in 1845, he decided to live in the woods.
thoreau went out into a woodlot owned by emerson (one of the few forest tracts near concord that had not been cut down) and built a cabin near Walden Pond. the woodlot was not wilderness but was a second-growth forest and was less than two miles (a little more than 2 km) from concord. He started his work on July 4 of that year, intending it as a statement of independence from the pursuit of wealth (which, he said, led most people into “lives of quiet desperation”), just as July 4 marked american independence and just as concord was the site of the first battle of the Revolutionary War. But he also intended his cabin as an experiment. He kept a jour-nal the rest of his life, some of which he published in the book Walden; or, Life in the Woods, which has become the environmental classic of the 19th century. in the journal, and book, thoreau wrote detailed records of how much it cost him to build the cabin, and how he raised or caught much of his own food, going to town only for a few staple food items. He even built the cabin from wood and nails left over from a laborer family’s shanty. He spent only a couple of years in the cabin. He did not intend it as a permanent change in lifestyle but to demonstrate that it could be done.
thoreau was also famous for his essay “Resis-tance to civil Government,” later known as “On civil Disobedience,” which was based on an experi-ence he had in 1846. For six years, he had refused to pay poll taxes because of his opposition to slavery and to the Mexican-american War. Massachusetts had no slaves but profited from goods produced by slaves, and the federal Fugitive Slave act required all citizens to aid in the capture of slaves who had escaped from the South. He also considered the war to be nothing more than aggression. the con-cord tax collector liked thoreau but was obligated to arrest him. thoreau was ready to spend a long period of time in jail as a protest, but his aunt paid his taxes, and he was released after one night.
thoreau’s civil disobedience inspired generations of social reformers, such as Russian writer leo tol-stoy, indian leader Mohandas Gandhi, and american
reformer Martin luther King, Jr. But his insistence that frugal living (what today would be called “hav-ing a small ecological footprint”) is not only possible but enjoyable contributed to his modern reputation as the prophet of environmentalism. Since the most ancient times, prophets have predicted disastrous outcomes to the way most people in their society lived, called for repentance from that way, and them-selves lived in a way that was a constant reminder of the way of repentance. this is exactly what Henry David thoreau did. like all prophets, thoreau was an inconvenient man. though by no means a hermit (he went to town every couple of days even during his cabin phase), he was always separated from the normal crowd: When in town, he observed people as might an anthropologist from another planet. His presence was a prophetic denouncement of materi-alistic society. thoreau was particularly vivid (and virtually alone) in his criticism of people who did not take care of their land. thoreau wrote that Flint’s Pond (also near concord) was named after an “unclean and stupid” farmer who laid its shores bare, loved a shiny dollar more than a shiny pond, reflecting “his own brazen face; who regarded even the wild ducks . . . as trespassers. . . . Who never saw it, who never bathed in it, who never loved it, who never protected it, who never spoke a good word for it, nor thanked God that He had made it. . . . Who exhausted the land around it. . . . He would carry his God to market; . . . whose fields bear no crops, whose meadows no flowers, whose trees no fruits, but dollars.” thoreau defended biodiversity against an agricultural mindset that, then as now, reduced a farm or a woodlot to its economic value and evalu-ated every plot of land on earth, however different each was from the other, according to just one scale of value.
thoreau was also a scientist, though without formal training. His observations of nature sug-gested hypotheses to him, which he (however imper-fectly) investigated. He was passionate about making observations (for example, the colors of Walden Pond ice and the stages by which it thawed) and measurement (the depths of Walden Pond). Scholars puzzle that his last writings were all mere observa-tions of seed dispersal and spring budburst dates of plants. But his observations were the basis upon which important aspects of ecological science were later based, such as the following:
● Seed dispersal. in his long essays “the Dis-persion of Seeds” and “the Succession of Forest trees,” thoreau presented many detailed observations to prove that trees grew only where their seeds had been planted by wind, water, or animals. as unbelievable
xviii+598_Biodiversity_F.indd 521 11/17/11 1:13 PM

first entry522 Thoreau, henry david522
as it may sound today, some intellectuals (none of them scientists) believed that trees just sprang up from the ground. thoreau demonstrated that wild cherry trees grew where birds had carried their seeds, and that cherry seeds would not grow unless they had passed through a bird’s digestive system. He tested a hypothesis that mice dispersed hazelnut seeds by microscopically comparing tooth marks on a hazel nut with the teeth of a mouse skull.
● Forest succession. He also systematically surveyed tree stumps, counting the rings to reconstruct forest history by determin-ing when each tree had begun to grow. He applied his observations to an understand-ing of the successional processes by which forests changed over time: Oaks grew up underneath pines, not underneath oaks, not only because oak forests cast too much shade upon the seedlings and oak leaf litter contained toxins, but also because animals such as squirrels and jays carried acorns preferentially into pine forests. thoreau also knew about natural disturbances, as well as human disturbances (he noted that native americans deliberately set fire to forests).
● Stochastic processes. thoreau recognized what scientists today call stochastic processes. ecological succession is not a deterministic process, always occurring in the same way, but by stochastic chance, depending upon which species of plants as a seed source might happen to be near the disturbance.
● Spatial patterns of plants. thoreau also studied spatial patterns in plants, noting that species whose seeds were dispersed by wind tended to grow in clumps, while animal-dispersed plants were more evenly spread on the landscape.
● Seasonal patterns. thoreau also kept copi-ous notes about the dates on which trees burst their buds in the spring, on which birds migrated, on which the ice thawed, and on which wild fruits matured. Modern scien-tists, such as botanist a. J. Miller-Rushing, have used thoreau’s data to document that springtime comes earlier, and winter comes later, in new england today than in tho-reau’s day. thus thoreau started the science of phenology, which is the study of seasonal adaptations of the life cycles of plants and animals.
the most famous thoreau quote, which reflects his important insights into the environment and sci-
ence, was “in wildness is the preservation of the world,” often misquoted as “wilderness.” to tho-reau, wildness could be found in a well-managed woodlot as easily as in the Maine woods that he visited as a young man.
thoreau was inspired by the writings of scientists who closely observed the world during their travels, particularly american botanist William Bartram, who traveled among the cherokee of the appala-chians in the late 18th century, and charles Darwin’s Voyage of the Beagle. thoreau strongly believed in the complete connectedness of humans and nature (“am i not partly leaves and vegetable mold myself?”) but did not fully understand it until he read Darwin’s Origin of Species shortly before his death. Previous to this, he had believed, as a transcendentalist, that all of nature reflected spiritual patterns. He was very serious when he observed leaf-like patterns in thaw-ing clay and said that it was the same natural law that produced the shapes of real leaves and also of human internal organs. the earth had a body, but its organs were on the outside: “What is man but a mass of thawing clay?” When he saw a mouse girdling a pine tree (chewing a ring of bark), he thought it was the earth’s way of thinning out the pines. Darwin’s Origin of Species undermined thoreau’s faith, and thoreau spent his last years making scientific obser-vations with few transcendentalist insights.
Without Ralph Waldo emerson, there would have been no remembrance of thoreau. it was emer-son’s woodlot in which thoreau briefly lived. emer-son popularized thoreau after the latter’s death. But they were very different. emerson would write long flowery-tongued passages about things, whether about the world of nature or the breathlessness of love, which he had not closely observed. to thoreau, nature was a living world from which to learn; to emerson, it was a canvas upon which to paint his grand ideas. For example, emerson said that “sav-age” languages were simple and consisted mostly of nouns. Had he even bothered to ask anyone who had learned native american languages—and there were plenty in his scholarly circle—he would have known this was wrong. But thoreau was fascinated by what he could learn from native americans. (His last words were “moose” and “indian.”)
thoreau caught pneumonia when he was in the woods counting tree rings. He was only 44 years old when he died on May 6, 1862.
See also disturbances, ecological; seed dis-persal; succession, ecological.
Further readingcain, William, ed. A Historical Guide to Henry David
Thoreau. cambridge, U.K.: Oxford University Press, 2006.
xviii+598_Biodiversity_F.indd 522 11/17/11 1:13 PM

Triassic period, biodiversity in 523
cramer, Jeffrey S., ed. Walden: A Fully Annotated Edition. new Haven, conn.: yale University Press, 2004.
Dassow, laura. Seeing New Worlds: Henry David Thoreau and 19th Century Science. Madison: University of Wisconsin, 1995.
Dean, Bradley P., ed. Faith in a Seed: The First Publication of Thoreau’s Last Manuscript. covelo, calif.: island Press, 1993.
Henry David thoreau Online. “the Succession of For-est trees.” available online. URl: http://www.tho reau-online.org/the-succession-of-forest-trees.html. accessed May 16, 2011.
Miller-Rushing, abraham J., et al. “Photographs and her-barium specimens as tools to document phenological changes in response to global warming.” American Journal of Botany 93 (2006): 1,667–1,674.
Richardson, Robert D. Henry Thoreau: A Life of the Mind. Berkeley: University of california Press, 1988.
Robinson, David M. Natural Life: Thoreau’s Worldly Tran-scendentalism. ithaca, n.y.: cornell University Press, 2004.
Sullivan, Robert. The Thoreau You Don’t Know: What the Prophet of Environmentalism Really Meant. new york: Harper, 2009.
triassic period, biodiversity in the trias-sic period (250 to 210 million years ago) was the first period of the Mesozoic era. it followed the Permian period, which ended with the Permian extinction.
Climate. Because the continent of Pangaea was so large, and centered on the equator, the middle of the continent had hot, arid conditions. the edges of the continent were warm and humid. low oxygen levels and high levels of carbon dioxide created a greenhouse world (see table).
Continents. a single worldwide continent, Pan-gaea, had formed during the Permian period. the Panthalassian (“world ocean”) Sea surrounded it, and the tethys Sea penetrated into it. the widening of the tethys Sea separated Pangaea into the north-ern laurasia and the southern Gondwana by the end of the triassic period.
Marine life. Many lineages of marine animals had become extinct at the end of the Permian period, but the surviving lines began to evolve into many new forms.
● Invertebrates. the trilobites became extinct at the end of the Permian period. Brachio-pods survived the extinction, but since the triassic period they have been much less common than the superficially similar bivalve mollusks.
● Vertebrates. Both sharks and bony fishes continued to be abundant in the oceans. a
multiCellular biodiversity in the triassiC period
Conditions
Atmospheric oxygen 80% of modern level
Atmospheric carbon dioxide 5 times modern level
Global temperature 5°F (3°C) higher than today
Sea level same as today
Approximate number of families*
Continental 150
Marine 300
Total 450
Examples of marine animals
Acrodus cuneocostatus shark
Anasibirites sp. cephalopod mollusk
Mysidiella sp. bivalve mollusk
Shonisaurus popularis ichthyosaur reptile
Traumatocrinus caudex crinoid echinoderm
Examples of continental animals
Cleithrolepis granulatus bony fish
Coelophysis bauri dinosaur
Dinodontosaurus oliveirai mammal-like reptile
Eudimorphodon ranzii pterosaur reptile
Ornithosuchus longidens crocodile-like reptile
Mollenia rieki hymenopteran insect
Parasuchus sp. crocodile
Plateosaurus longiceps dinosaur
Proganochelys sp. turtle
Tanystropheus conspicuus archosaur reptile
Tetracoleus minimus beetle
Thrinaxodon brasiliensis reptilelike mammal
Triadobatrachus massinoti frog
Triassaraneus andersonorum spider
Triassopsychoda olseni fly
Examples of land plants
Antarcticycas schopfi cycad
Caytonia sp. seed fern
Telemachus elongatus conifer
*Based on Benton.
xviii+598_Biodiversity_F.indd 523 11/17/11 1:13 PM

fi rst entry524 Triassic period, biodiversity in 524
new group of bony fi shes, the teleosts, fi rst appeared in the triassic period. they were lighter and faster than earlier bony fi shes. On the continents, reptiles proliferated into many forms, and some of these rep-tiles evolved into marine forms. Tanystro-pheus lived on the seashore, and half of its 22-foot (7 m) length was neck, with which it captured fi sh. the fi rst ichthyosaurs (such as Shonisaurus) evolved during the trias-sic. these reptiles were so fully adapted to marine life that they looked very much like fi sh and could not come onto dry land.
Continental life. Scientists speculate that the dry conditions in the middle of Pangaea provided an advantage to seed plants over plants that reproduced by spores and to reptiles over amphibians.
● Plant life. the seed fern Glossopteris domi-nated vast forest areas at the beginning of the triassic. Seed plants later dominated over seedless plants, forming the fi rst exten-sive forests of conifers, which had fi rst evolved in the late Paleozoic era.
● Invertebrates. insect lineages that evolved in the Permian, such as fl ies and beetles, continued to diversify during the triassic. the fi rst hymenopterans (today represented by ants, bees, and wasps) evolved during the triassic but represented only a single family.
● Vertebrates. Some lineages of amphib-ians survived the Permian extinction and evolved into new forms. Frogs evolved dur-ing the triassic. at the beginning of the
triassic period, the mammal-like reptile Lystrosaurus was overwhelmingly abundant (see fi gure). this small gen-eralist reptile could tolerate the low oxygen of, and fi nd food in, the post-extinction world. after about 10 million years, reptiles began to evolve into numerous forms. the fi rst turtles are known from this time. the mammal-like reptiles continued to evolve, and the fi rst reptilelike mam-mals (such as Thrinaxodon) evolved during the triassic. Other mammal-like reptiles evolved along a separate path, becoming very large ani-mals, such as Dinodontosau-rus, but they became extinct
at the end of the triassic. archosaur rep-tiles evolved legs that were underneath the body, which allowed them to run faster; this included the early crocodile Parasuchus and the dinosaur-like Ornithosuchus, which could run on two legs. the fi rst pterosaurs (fl ying reptiles) evolved at this time also. the fi rst dinosaurs evolved during the triassic but were a minor component of biodiver-sity. Some, like Coelophysis, were small, rapid carnivores that ran on two legs, while some herbivorous dinosaurs (such as Plateo-saurus) were large (up to 26 feet, or 8 m), though not as large as the ones that would evolve in the Jurassic period.
Extinction. another of the mass extinctions occurred at the end of the triassic period, which may have allowed the dinosaurs to become the over-whelmingly most abundant terrestrial vertebrates.
See also mass extinctions; Permian extinction.
Further readingBenton, Michael J. “Diversifi cation and extinction in the
history of life.” Science 268 (1995): 52–58.———. “Four feet on the ground.” in The Book of Life: An
Illustrated History of the Evolution of Life on Earth,edited by Stephen J. Gould. new york: norton, 2001.
Kubo, tai, and Michael J. Benton. “tetrapod postural shift estimated from Permian and triassic trackways.” Pal-aeontology 52 (2009): 1,029–1,037.
Marshall, charles R., and David K. Jacobs. “Flourishing after the end-Permian mass extinction.” Science 325 (2009): 1,078–1,080.
During the early Triassic period, right after the Permian extinction, there were few animal species. Lystrosaurus, a mammal-like reptile, was abun-dant in many parts of the world.
xviii+598_Biodiversity_F.indd 524 11/17/11 1:13 PM

tropical rain forests and deforestation 525
Mcelwain, Jennifer c., et al. “Fossil plant relative abun-dances indicate sudden loss of late triassic biodi-versity in east Greenland.” Science 324 (2009): 1,554–1,556.
tropical rain forests and deforestation tropical forests have no winter and receive abundant rainfall during at least part of the year. Tropical sea-sonal forests have a rainy season and a dry season, while tropical rain forests receive rain all year (see figure). in some rain forests, the rain comes pre-dictably every afternoon. tropical forests exist just north and south of the equator and are called tropi-cal because they are found between the tropic of cancer (23.5° n) and the tropic of capricorn (23.5° S). temperatures remain relatively constant, within
the range of 68–77°F (20–25°c). Precipitation (all of it rain) exceeds 78 inches (200 cm) per year. along with the lack of temperature variation, the tropi-cal forests have relatively little wind. tropical rain forests cover less than 5 percent of the land surface of the earth but contain nearly half of its species and can therefore be considered the most important repositories of biodiversity in the world. However, only a few of the small biodiversity hotspots are tropical.
BIodIvErSITy IN TropICal forESTStropical rain forest trees are evergreen. Up to half of the rain that falls comes from the transpiration of water vapor from the leaves of the trees themselves. Deforestation interrupts transpiration and allows water that would otherwise have transpired back
A rainbow appears as rain retreats from a forest in the Ecuadorian Amazon. In many rain forests, rain is a daily occurrence. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 525 11/17/11 1:13 PM

first entry526 tropical rain forests and deforestation 526
up into the sky to flow out into the ocean instead. extensive deforestation of rain forests may therefore produce a drier climate that will not support the regrowth of those forests, resulting in a destruction of the rain forest that is permanent over a time span meaningful to the human economy.
tropical rain forests have the greatest produc-tivity of plant growth (in weight of plant material produced per area per year) as well as the highest biodiversity of species of any region of the earth. a few acres or hectares of rain forest near Manaus, Peru, may contain as many tree and bird species as all of europe. each tree species may be separated from the nearest other member of its species by up to a mile (1.4 km). Borneo has 2,500 species of orchids, and it has at least 60 different tree species in an acre of forest. among the reasons that rain forests have such prodigious species diversity are the following:
● No winter. the lack of winter allows eco-logical interactions and thus coevolution to occur all year long. coevolution not only produces evolutionary change but can also result in speciation. Plants, for example, cannot rely upon winter to diminish the populations of herbivorous insects. each species of plant evolves its own set of chem-ical and other defenses, and each species of herbivorous animal evolves its own way of avoiding or tolerating those defenses. Many of these toxins, when used in small quantities, function as pharmaceutical com-pounds. Because of their high biodiversity and because so many species (especially of plants) produce toxins, tropical forests have been a gold mine in the discovery of phar-maceutical compounds.
● Lack of wind pollination. the lack of wind, and the persistent moisture, makes wind pollination relatively ineffective. each flow-ering plant specializes upon an animal that can carry its pollen reliably over the con-siderable distance that separates it from the next plant of the same species.
● Specialization on microhabitats. the incredible abundance of moisture, warmth, and sunlight allows plants to specialize on different vertical microhabitat layers within the rain forest. the forest canopy contains many tree species, while some tree species (such as Cecropia sp.) emerge above the can-opy. the dark understory contains relatively few plants, but there are many woody vines (lianas). they are rooted in the soil, but their
branches coil amidst the canopy branches of the trees. Many epiphytes, especially bro-meliads and orchids, are rooted in litter and tree bark up in the tree branches and obtain all their water from rain and nutrients from dead leaves shed by the trees around them (see figure). lianas are less common, and epiphytes rare, in temperate forests, where there is not enough rain to keep epiphytes alive. Because the plants divide up the layers of rain forest so finely among themselves, they also create a lot of different habitats for animals, most of which—with the excep-tion of a few such as tapirs—spend most of their lives up in the tree branches. However, there is little evidence for horizontal special-ization in rain forests; many tropical trees grow, widely spaced, over a large geographi-cal area.
not only do tropical rain forests have incred-ible diversity within them, but tropical forests differ markedly in species composition from place to place. evolution has produced patterns of biogeography among different tropical forests, just as it has with other ecological communities. tropical rain forests are famous for their endemic species. For example, Borneo has the world’s largest orchid, the largest moth, and the largest carnivorous plant; it also has squirrels, lizards, colugos, frogs, and even snakes that glide through the air. Forty-four of its 222 mammal species, 74 of its 420 bird species, 91 of its 254 reptile species, 114 of its 149 amphibian species, 160 of its 430 fish species, and 155 of its 265 spe-cies of dipterocarp trees are endemic to the island.
Epiphytes on a tree near Santa Elena, Costa Rica. They are common in tropical rain forest trees. (Dirk van der Made)
xviii+598_Biodiversity_F.indd 526 11/17/11 1:13 PM

tropical rain forests and deforestation 527
the vine that produces the world’s largest flower, Rafflesia arnoldii, is found only on the slopes of Mt. Kinabalu on Borneo. the rain forests of Madagascar consist mostly of species found nowhere else, includ-ing a little species of periwinkle (Catharanthus roseus) that was the original source of the important antileukemia drugs vincristine and vinblastine. con-sider these patterns of canopy trees and mammals (see table on pages 528–530):
● the rain forest tree species of the new World (the neotropics) are completely dif-ferent from those of the Old World (the paleotropics), with few exceptions. (West africa, like the amazon, contains the kapok tree Ceiba pentandra, although its subspe-cies are unique to the neotropics.)
● Within the neotropics and the paleotropics, the rain forests are also largely unique. the atlantic rain forest of South america, a nar-row belt of forest in the vicinity of Rio de Janeiro and São Paulo, is distinct from the amazon rain forest, although they share some tree species such as Slonea guianen-sis. tree species in the Pacific rain forest of colombia and coastal ecuador differ from those of the amazon, which is just on the other side of the andes. the yuca-tán and central american rain forests are largely distinct from the amazon rain for-est, though they also share some species. the indonesian and Malaysian rain forests are overwhelmingly dominated by tree spe-cies in the plant family Dipterocarpaceae, a family that is rare elsewhere in the Old World. the forests of indonesia have a dif-ferent set of species from those of india and Sri lanka and those of northern australia. the rain forest of eastern Madagascar is likewise unique.
● the plant families that are important in the tropics are those that evolved there and tend to be less common in temperate zones. the ebony family (ebenaceae), common in the tropics, is represented in the United States by the persimmon tree (Diospyros virginiana). the mulberry family, which has thousands of fig and other species in the rain forests, is represented in the United States by mulberries (such as Morus rubra) and by the bois d’arc (Maclura pomifera), none of which are dominant forest tree species. Kapok trees (genus Ceiba) are in the mallow family, represented mainly by shrubs and wildflowers outside the tropics.
animal species differ markedly from one tropi-cal forest to another, as in these examples:
● Primates. the new World monkeys evolved separately from the Old World monkeys. each neotropical rain forest has different monkeys. the golden lion tamarin is unique to the atlantic rain forest, while capuchin monkeys range more widely. in the paleo-tropics, proboscis monkeys live in Sumatra, rhesus monkeys in and around india, and colobus monkeys in africa. the neotropical rain forests have no apes. among the apes, gibbons and orangutans live only in asian rain forests, and chimpanzees and gorillas only in african rain forests. Madagascar is famous for having no monkeys or apes but only lemurs.
● Other placental mammals. neotropi-cal mammals are completely distinct from paleotropical mammals. Some neotropi-cal mammals, such as sloths, jaguars, and tapirs, live in all neotropical rain forests. Some mammals are unique to certain rain forests, such as the okapi in africa. Once again, Madagascar is unique. its top preda-tor is the fossa, which is a type of mongoose.
● Marsupials and monotremes. Perhaps the most distinctive feature of tropical mam-mal distribution is the prevalence of mar-supials, and the presence of monotremes, in australia and nearby regions such as new Guinea. the 19th-century naturalist alfred Russel Wallace first noticed that placental mammals dominated indonesia and that marsupials dominated the islands near aus-tralia and proposed the now-famous Wal-lace’s line. Scientists now understand that this line corresponds closely to the junc-tion of the eurasian and australian crustal plates. Mammals evolved separately in asia and australia, which came close together only recently in geological history. the dip-terocarps that dominate indonesian forests are also rare in the australian zone. Both trees and mammals make australia and new Guinea a world unto itself in terms of biodiversity.
Similar patterns of tropical biodiversity are found in all kinds of fungi, plants, and animals—and no doubt protists and bacteria also, although these have not yet been studied as much as animals and plants.
(continues on page 530)
xviii+598_Biodiversity_F.indd 527 11/17/11 1:13 PM
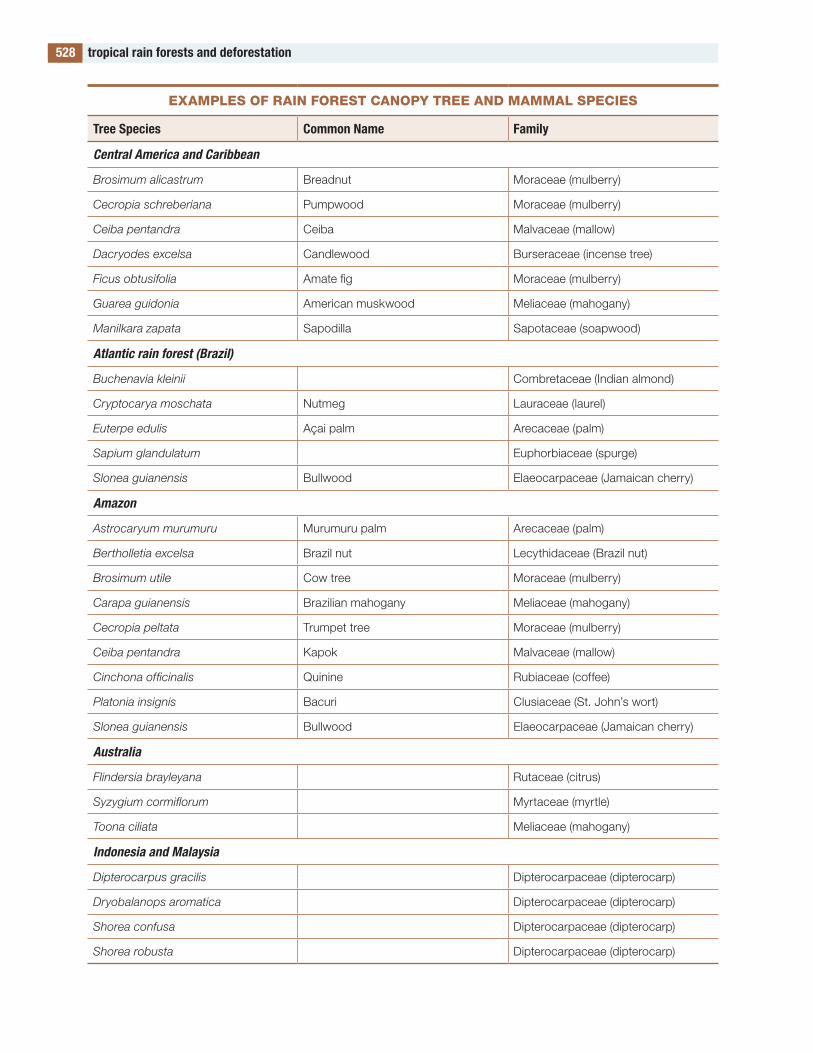
first entry528 tropical rain forests and deforestation 528
examples of rain forest Canopy tree and mammal speCies
Tree Species Common Name family
Central America and Caribbean
Brosimum alicastrum Breadnut Moraceae (mulberry)
Cecropia schreberiana Pumpwood Moraceae (mulberry)
Ceiba pentandra Ceiba Malvaceae (mallow)
Dacryodes excelsa Candlewood Burseraceae (incense tree)
Ficus obtusifolia Amate fig Moraceae (mulberry)
Guarea guidonia American muskwood Meliaceae (mahogany)
Manilkara zapata Sapodilla Sapotaceae (soapwood)
Atlantic rain forest (Brazil)
Buchenavia kleinii Combretaceae (Indian almond)
Cryptocarya moschata Nutmeg Lauraceae (laurel)
Euterpe edulis Açai palm Arecaceae (palm)
Sapium glandulatum Euphorbiaceae (spurge)
Slonea guianensis Bullwood Elaeocarpaceae (Jamaican cherry)
Amazon
Astrocaryum murumuru Murumuru palm Arecaceae (palm)
Bertholletia excelsa Brazil nut Lecythidaceae (Brazil nut)
Brosimum utile Cow tree Moraceae (mulberry)
Carapa guianensis Brazilian mahogany Meliaceae (mahogany)
Cecropia peltata Trumpet tree Moraceae (mulberry)
Ceiba pentandra Kapok Malvaceae (mallow)
Cinchona officinalis Quinine Rubiaceae (coffee)
Platonia insignis Bacuri Clusiaceae (St. John’s wort)
Slonea guianensis Bullwood Elaeocarpaceae (Jamaican cherry)
Australia
Flindersia brayleyana Rutaceae (citrus)
Syzygium cormiflorum Myrtaceae (myrtle)
Toona ciliata Meliaceae (mahogany)
Indonesia and Malaysia
Dipterocarpus gracilis Dipterocarpaceae (dipterocarp)
Dryobalanops aromatica Dipterocarpaceae (dipterocarp)
Shorea confusa Dipterocarpaceae (dipterocarp)
Shorea robusta Dipterocarpaceae (dipterocarp)
xviii+598_Biodiversity_F.indd 528 11/17/11 1:13 PM
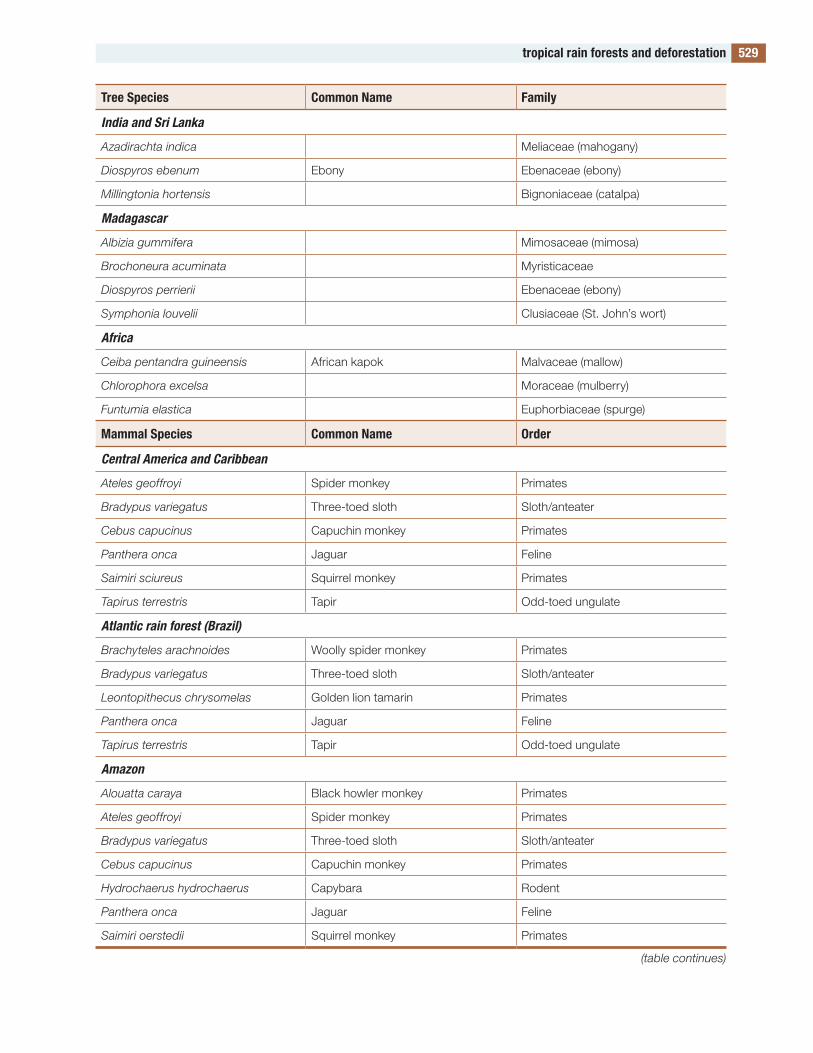
tropical rain forests and deforestation 529
Tree Species Common Name family
India and Sri Lanka
Azadirachta indica Meliaceae (mahogany)
Diospyros ebenum Ebony Ebenaceae (ebony)
Millingtonia hortensis Bignoniaceae (catalpa)
Madagascar
Albizia gummifera Mimosaceae (mimosa)
Brochoneura acuminata Myristicaceae
Diospyros perrierii Ebenaceae (ebony)
Symphonia louvelii Clusiaceae (St. John’s wort)
Africa
Ceiba pentandra guineensis African kapok Malvaceae (mallow)
Chlorophora excelsa Moraceae (mulberry)
Funtumia elastica Euphorbiaceae (spurge)
mammal Species Common Name order
Central America and Caribbean
Ateles geoffroyi Spider monkey Primates
Bradypus variegatus Three-toed sloth Sloth/anteater
Cebus capucinus Capuchin monkey Primates
Panthera onca Jaguar Feline
Saimiri sciureus Squirrel monkey Primates
Tapirus terrestris Tapir Odd-toed ungulate
Atlantic rain forest (Brazil)
Brachyteles arachnoides Woolly spider monkey Primates
Bradypus variegatus Three-toed sloth Sloth/anteater
Leontopithecus chrysomelas Golden lion tamarin Primates
Panthera onca Jaguar Feline
Tapirus terrestris Tapir Odd-toed ungulate
Amazon
Alouatta caraya Black howler monkey Primates
Ateles geoffroyi Spider monkey Primates
Bradypus variegatus Three-toed sloth Sloth/anteater
Cebus capucinus Capuchin monkey Primates
Hydrochaerus hydrochaerus Capybara Rodent
Panthera onca Jaguar Feline
Saimiri oerstedii Squirrel monkey Primates
(table continues)
xviii+598_Biodiversity_F.indd 529 11/17/11 1:13 PM
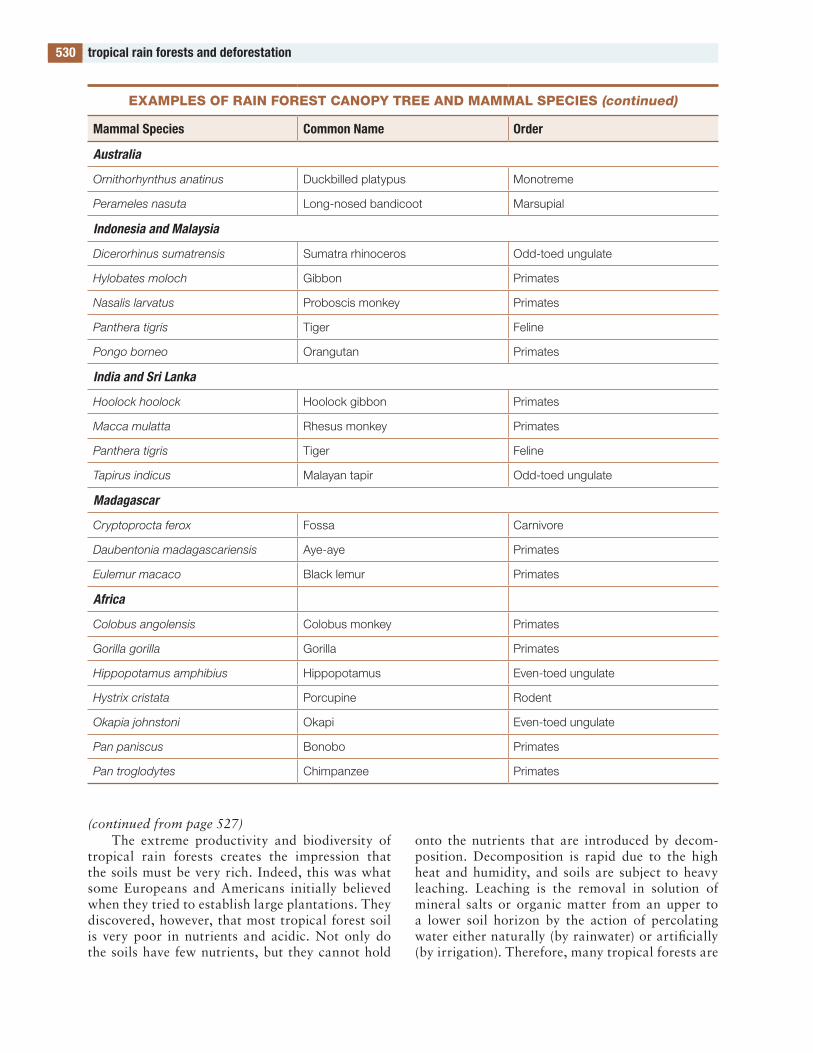
first entry530 tropical rain forests and deforestation 530
the extreme productivity and biodiversity of tropical rain forests creates the impression that the soils must be very rich. indeed, this was what some europeans and americans initially believed when they tried to establish large plantations. they discovered, however, that most tropical forest soil is very poor in nutrients and acidic. not only do the soils have few nutrients, but they cannot hold
onto the nutrients that are introduced by decom-position. Decomposition is rapid due to the high heat and humidity, and soils are subject to heavy leaching. leaching is the removal in solution of mineral salts or organic matter from an upper to a lower soil horizon by the action of percolating water either naturally (by rainwater) or artificially (by irrigation). therefore, many tropical forests are
examples of rain forest Canopy tree and mammal speCies (continued)
mammal Species Common Name order
Australia
Ornithorhynthus anatinus Duckbilled platypus Monotreme
Perameles nasuta Long-nosed bandicoot Marsupial
Indonesia and Malaysia
Dicerorhinus sumatrensis Sumatra rhinoceros Odd-toed ungulate
Hylobates moloch Gibbon Primates
Nasalis larvatus Proboscis monkey Primates
Panthera tigris Tiger Feline
Pongo borneo Orangutan Primates
India and Sri Lanka
Hoolock hoolock Hoolock gibbon Primates
Macca mulatta Rhesus monkey Primates
Panthera tigris Tiger Feline
Tapirus indicus Malayan tapir Odd-toed ungulate
Madagascar
Cryptoprocta ferox Fossa Carnivore
Daubentonia madagascariensis Aye-aye Primates
Eulemur macaco Black lemur Primates
Africa
Colobus angolensis Colobus monkey Primates
Gorilla gorilla Gorilla Primates
Hippopotamus amphibius Hippopotamus Even-toed ungulate
Hystrix cristata Porcupine Rodent
Okapia johnstoni Okapi Even-toed ungulate
Pan paniscus Bonobo Primates
Pan troglodytes Chimpanzee Primates
(continued from page 527)
xviii+598_Biodiversity_F.indd 530 11/17/11 1:13 PM

tropical rain forests and deforestation 531
sitting on top of, rather than growing in, the soil. as soon as leaf litter decomposes, the roots of the trees absorb the nutrients. When the forests are cut down, the nutrients are lost, leaving a ravaged landscape in which relatively few plants, including few agricultural plants, can grow. Recent evidence suggests that some small regions of “black soil” (terra preta de Indio) in South american rain for-ests were artificially produced by composting by native tribes in Precolumbian times to produce soil suitable for agriculture. Some tropical for-ests near volcanoes, such as those of the western amazon Basin and the Philippines, have rich soils continually replenished by nutrients from volcanic ash. Rain forest soils near major rivers may receive nutrients from river silt. However, in most tropi-cal forests, it takes a very long time for forests to grow back, so long that tropical forest destruction may be permanent on time scales that matter to the human economy.
in many tropical rain forests, there is sufficient rainfall that many plant species can grow without being rooted in the soil. these epiphytes include vines, bromeliads, and orchids. the competition for light is so intense that most of the plant species and most of the leaf area of the trees are found in the canopy. the understory of a tropical rain forest may have extensive areas without very much plant growth, except for tree trunks. the popular image of the “jungle” being nearly impenetrable without machetes is true of the edges of these forests and of areas where the original forest has been disturbed. Most animal species also live in the canopy. this includes many of the vectors of disease, such as mosquitoes, which grow in small pools of water in
tree trunks and bromeliad leaf bases, and parasit-ize treetop animals living in the canopy. Destruc-tion of these forests often has the unintended side effect of bringing humans and mosquitoes into more intense contact, thereby enhancing the spread of diseases such as malaria and yellow fever. Scientists who study rain forests now often work in treetop perches, traveling by rope bridges or cables from one tree to another, and often coming and going by means of helicopters, largely ignoring the shade underneath them.
dESTruCTIoN of TropICal forESTStropical rain forests are being lost at an alarm-ing rate: 46,300 square miles (120,000 km2) per year. Half of the original 5.4 to 7 million square miles (14–18 million km2) of rain forest has been destroyed (see photograph).
tropical rain forests are just as capable of responding to natural disturbances as are any other ecological communities. in temperate forests, many seeds remain dormant in the soil until a distur-bance occurs, at which time they germinate. in tropical forests, seeds have little dormancy and will often germinate in the shade and grow slowly. if a disturbance clears away some of the canopy over-head, they begin growing quickly. in the amazon rain forest, some native american tribes and peas-ants of european descent make their living partly by raising crops in small clearings. tribal peoples in rain forests all over the world have done the same. in this slash and burn or shifting agriculture, a small plot of rain forest trees is cut, allowed to dry, and burned. the ashes contribute nutrients to the poor soil, allowing a rich growth of crops. the poor soil, however, does not hold the nutri-ents, which soon wash away. the farmers then abandon the field and allow secondary succession to occur. the rain forest recovers from these small disturbances, as leaf litter and tree seeds come in from the nearby forest. Within decades, the rain forest reclaims the opening (see figure on page 532). this kind of shifting agriculture is not permanently dangerous to tropical forests unless it occurs at a high frequency due to the activity of dense human populations.
However, when a very large rain forest clearing is created and then abandoned, most of the clearing is far away from the forest edge from which seeds and leaf litter would come to begin the successional process. Seeds of rain forest species, unlike the colo-nizing species of temperate zones, do not remain dormant in the soil, but die within months of being shed. Moreover, while a small clearing is partially shaded by the nearby forest, the middle of a large
Tropical deforestation in Brazil. The red soil, where trees have been cut down, is poor in nutrients and will raise meager crops. Biodiversity has been degraded in the remaining forest fragment. (Alberto Cesar; Greenpeace; courtesy Rechargenews.com)
xviii+598_Biodiversity_F.indd 531 11/17/11 1:13 PM

fi rst entry532 tropical rain forests and deforestation 532
clearing is baked by the tropical sun, which kills seedlings and dries the thin soil despite frequent rains (see fi gure). the few plants that live in the dry open area are more prone to fi re, which further sets back the successional process. Soil erosion occurs severely in large clearings, making the chances of recovery even slimmer. Gradually, succession occurs along the edges of the clearing. if the extensive destruction of forests causes a local reduction in rainfall, the large clearing may become a permanent arid zone, and the reduced rainfall may harm even the rain forest that remains uncut. the good news is that when forest succession does occur, most of the plant species (more than 90 percent, according to plant ecologist Robin chazdon) return.
Just a few decades ago, the rain forests of the world seemed not only eternal but impenetrable. Many people believed that “the jungle” was merely a hindrance to the development of useful land. calling it a “green hell,” the Brazilian government launched a campaign to conquer the amazon rain forest, even moving their capital from the coast to the jungle outpost of Brasília. During the 1970s and even more during the 1980s, rain forests all over the world were demolished.
Governments of tropical countries could not believe that the forests would ever run out. However, forests even larger than the amazon rain forest (such as the eastern deciduous forest of the United States) have been cut down. therefore, the destruction of the amazon rain forest is not impossible. certainly, smaller tropical forests, such as the atlan-tic forest of Brazil, are vulnerable. conservationists proclaimed that rain forest destruction would lead to disaster, not only for tropi-cal countries but for the whole world. about 1990, governments all over the world began to real-ize that the conservationists had been right: Rain forests were threatened with annihilation, a prospect with consequences for the whole earth. By then, the forces of destruction had become so great that they can hardly be slowed, much less stopped. the Brazilian government, which had once waged war on its green hell, now used satellite imagery to help government offi cials track down and stop people from cutting the
forests. But even this has not rescued the rain forests. Here are some of the seemingly unstoppable forces of destruction:
● Pastures. Many central and South ameri-can rain forests have been cut down to pro-duce pastureland for cattle. Some american and multinational fast-food corporations have imported beef from pastures that had once been central american rain forest. Because the soil is bad, lands that had once been the world’s richest forests now became the world’s poorest pastures.
● Agriculture. the large coastal cities of Bra-zil teem with poor people. the government believed that the solution to their poverty was to bus them out into the amazon rain forest and give them free land. this solu-tion has not worked, for the poor soil of the rain forest makes terrible farmland, leav-ing the peasants as poor in the rain forest as they had been back in the city. Further, the population of poor in the cities has not diminished noticeably. Peasants destroy only about one-quarter as much forest as
Seedling regrowth and seed dispersal occur rapidly in a small gap distur-bance in a tropical forest but much more slowly in a large disturbed area.
xviii+598_Biodiversity_F.indd 532 11/17/11 1:13 PM

tropical rain forests and deforestation 533
pastures in Brazil. But peasant agriculture is a small force compared to large-scale com-mercial agriculture. in Borneo, one-third of the rain forest that existed in 1985 had been destroyed by 2005, a great deal of it to cre-ate plantations of oil palm. in Brazil, soy-bean plantations are having a similar effect on the amazon rain forest.
● Logging. Some tropical trees provide the world’s best wood for carpentry and musi-cal instruments. logging of these trees, however, accounts for very little deforesta-tion. Most trees that are cut by tropical loggers are simply burned; those that are used mostly end up as wood chips for paper, a tragic waste of some of the world’s best wood. Many Southeast asian rain forests have been destroyed largely to produce card-board. Selective logging removes trees with high economic value and leaves the others. although selective logging does not destroy a rain forest, it may alter it considerably by reducing the canopy cover and allowing drier conditions.
● Extraction of underground resources. the rain forests of ecuador are rich in petro-leum. Much rain forest has been destroyed during the exploration for and extraction of oil, as well as by the spilling of toxic materials. Rich gold deposits in Brazilian rain forests have tempted thousands of min-ers to strip away whole mountainsides of soil. Gold extraction uses mercury, which severely pollutes rivers downstream far from the actual site of destruction. in africa, rain forests have been destroyed for diamond mining. On Borneo, large tracts of rain for-est are destroyed for coal mining.
● War. a great deal of rain forest destruction occurred during the Vietnam War, much of it by the aerial spraying of defoliant chemi-cals. Some destruction of the amazon rain forest was apparently intended to bring independent native tribes under the control of the central government.
● Dams. Some tropical countries have built dams for hydroelectric power. in low-lying tropical areas, these reservoirs are often shal-low and very extensive, destroying much rain forest. Many farmers cut down forests on slopes surrounding the reservoir, leading to soil erosion, which clogs the reservoirs and makes them useless soon after they are built.
can anything be done to stop the direct destruc-tion of the rain forests? a few promising trends are
occurring, although they may be too little too late. they include the following:
Policy decisions. Governments of tropical coun-tries, such as Brazil, are beginning to prohibit defor-estation. costa Rica has set aside almost one-quarter of its area as land managed for conservation. the World Bank has begun requiring tropical countries to protect their rain forests before it agrees to lend money to them. in several cases, some debts that tropical countries owe to the major industrialized countries have been waived in return for rain forest preservation projects. Political leaders around the world are beginning to discuss the possibility that industrial countries, which benefit because tropical rain forests remove carbon dioxide from the atmo-sphere, should pay the developing tropical countries for these important environmental services. Perhaps the major example is the United nations ReDD Pro-gram (Reducing emissions from Deforestation and Forest Degradation). While government and bank policy decisions are not enough by themselves, there would be no hope of rain forest conservation without them. Meanwhile, many countries with a lot of rain forest land continue to have corrupt governments. indonesia has emerged from the Suharto regime, in which concessions to “develop” rain forest land were given almost for free to the dictator’s associates. at local levels, corruption continues almost unabated. agreements to save rain forests mean absolutely nothing in countries with corrupt governments.
Sustainable use of rain forest lands and securing land rights. Many countries have realized that the intact rain forest is more profitable than one that has been cut down. Brazil has established several extrac-tive preserves in which people live in the rain forest, have small garden plots, and hunt a limited amount of game. they collect sap from native rubber trees and make it into rubber, a process that does not kill the trees. they also gather and sell many native forest nuts (including Brazil nuts), fruits, and cocoa pods. these preserves have been successful; however, the people who live in them are poor, and not many peo-ple have chosen to move into the rain forest preserves to pursue this career. extractive preserves cannot by themselves save the rain forests. as noted above, many pharmaceutical compounds were first discov-ered in the leaves of rain forest plants. Pharmaceuti-cal companies formerly obtained plant specimens for free but now must pay for the privilege of pharma-ceutical prospecting. their money helps make the preservation of intact rain forests profitable.
Reclaiming degraded tropical lands. Former rain forest lands that have been severely degraded seem almost beyond hope for reclamation. However, in some cases, trees have been replanted. Some rain forest seedlings can survive bright sunlight and can
xviii+598_Biodiversity_F.indd 533 11/17/11 1:13 PM

first entry534 tropical rain forests and deforestation 534
be planted in the middle of the denuded wasteland. these seedlings require much care from humans to survive. However, once these trees become large enough to survive on their own, birds and bats will roost in them and drop seeds under them. the seeds grow into yet more tree seedlings. the large trees cast shade in which seedlings of shade-tolerant tree spe-cies can survive. these replanted trees, though few in number, may serve as nuclei for natural successional processes.
the legal basis of the extractive preserves and pharmaceutical prospecting is perhaps more impor-tant than their profitability. the rights of indigenous tribes and of rain forest peasants to their lands must be recognized. in the past, governments acted as if the rain forests were free, empty land, whereas in reality they were filled with tribal peoples. this is the same policy that the United States took toward the native american lands in the 19th century. Only now in Brazil and Southeast asia are tribal peoples beginning to receive the right to pursue their tradi-tional styles of life, on their own lands, safe from agriculture, pastures, mining, and lumbering by out-siders. it is widely understood that there is no way to save the rain forests unless the poor people of tropi-cal countries have a way of making a living.
in a few places, rain forest destruction has been slowed, as tropical governments and citizens all over the world have begun to recognize the importance of rain forests. But deforestation rates seemed to be more closely connected to the economy than to conservation sentiments. in Brazil, for example, deforestation decreased during times of recession and increased during times of recovery. Because a great deal of rain forest destruction in Brazil is due to creating soybean fields, deforestation rates rise and fall with soybean prices. Brazilian deforesta-tion in 2005–2009 was only 36 percent as great as its 1996–2005 average, largely because the interna-tional timber market now demands that rain forest timber come from certified plantations rather than wild rain forests. the Brazilian government has the power to make banks cancel credit extended to large landowners who are logging rain forest land ille-gally. although 51 percent of the remaining rain forest in Brazil is protected, enforcement is difficult and imperfect. Much destruction will continue even after everyone agrees that the rain forests should be saved, for the forces of destruction may have taken on a momentum of their own.
TropICal dEforESTaTIoN aNd GloBal warmING
Recent estimates have indicated that the rates of tropical deforestation are less than had previously been estimated and that fewer species are threat-
ened with extinction than had been thought. at the same time that this good news emerged, how-ever, scientists began making predictions about the effects of global warming on ecosystems around the world. like every other ecosystem, tropical forests will experience climatic shifts. the consequences are almost certain to be more dramatic than any that could have resulted from deforestation. Some tropical forests, such as those of the southern ama-zon Basin, will experience warmer, drier conditions, which will not only cause the death of many trees, smaller plants, and animals that require a moist habitat but will encourage the spread of fires, which will encourage the spread of fire-adapted vegetation. Fires have already become more common in rain forest lands. tropical trees, with thin bark, are very easily killed by fires. Some tropical forests, however, such as those of the northern amazon Basin, will experience warmer, wetter conditions, which is a set of conditions found no place in the world today. as a result, the consequences for these forests cannot be predicted. as with nearly every other environmental challenge described in this book, the protection of the tropical rain forests will primarily depend upon the prevention of an excessive greenhouse effect.
tropical deforestation is one of the major con-tributors to the greenhouse effect. On Borneo, 11 percent of the land is peat swamp, which has organic matter up to 60 feet (almost 20 m) deep. When an acre of regular rain forest land is cleared and burned, about 300 tons of carbon dioxide (702 metric tons per hectare) are released into the air; clearing of peat swamp puts about 725 tons per acre (1,650 metric tons per hectare) into the air. every year thereafter, an acre of peat soil puts almost 25 tons of carbon dioxide (55 metric tons per hectare) into the air just because the peat decomposes in the sunny dry con-ditions. this contrasts with the one ton (2.5 metric tonnes) per year that a typical american household releases. these are the principal reasons that indone-sia is the number-three emitter of carbon, after the United States and china.
the tropical rain forests collectively have the greatest biodiversity of any ecological community on the planet and are also being destroyed more rapidly than any other. their destruction not only reduces global biodiversity but contributes to the greenhouse effect, both by releasing carbon and by reducing photosynthesis. it is in the political and economic interest, not just the scientific interest, of the devel-oped world to reward developing nations for pro-tecting their rain forests, in a manner pioneered by the United nations ReDD program.
See also biodiversity, resources for medicine; biogeography; coevolution; dis-turbances, ecological; endemic species;
xviii+598_Biodiversity_F.indd 534 11/17/11 1:13 PM

tropical seasonal forests 535
greenhouse effect and biodiversity; specia-tion; succession, ecological; tropical sea-sonal forests.
Further readinganderson, anthony B. Alternatives to Deforestation: Steps
toward Sustainable Use of the Amazon Rain Forest. new york: columbia University Press, 1990.
cole, R. J., K. D. Holl, and R. a. Zahawi. “Seed rain under tree islands planted to restore degraded lands in a tropical agricultural landscape.” Ecological Applica-tion, 20 (2010): 1,255–1,269.
cowell, adrian. The Decade of Destruction: The Crusade to Save the Amazon Rain Forest. new york: Henry Holt, 1990.
Dauvergne, Peter. Shadows in the Forest: Japan and the Politics of Timber in Southeast Asia. cambridge, Mass.: Mit Press, 1997.
De Onis, Juan. The Green Cathedral: Sustainable Devel-opment of Amazonia. new york: Oxford University Press, 1992.
Klanderud, Kari, et al. “Recovery of plant species richness and composition after slash-and-burn agriculture in a tropical rain forest in Madagascar.” Biodiversity and Conservation 19 (2010): 187–204.
laborde, Javier, et al. “tree and shrub seed dispersal in pastures: the importance of rain forest trees outside of forest fragments.” Ecoscience 15 (2008): 6–16.
laurance, William F., and carlos a. Peres, eds. Emerging Threats to Tropical Forests. chicago: University of chicago Press, 2006.
leigh, egbert Giles. Tropical Forest Ecology: A View from Barro Colorado Island. Oxford, U.K.: Oxford Univer-sity Press, 1999.
Guevara, Sergio, et al. “the role of remnant forest trees in tropical secondary succession.” Plant Ecology 66 (1986): 77–84.
Hecht, Susanna, and alexander cockburn. The Fate of the Forest: Developers, Destroyers, and Defenders of the Amazon. Updated ed. chicago: University of chicago Press, 2010.
Hooper, elaine R., et al. “Factors affecting composition of forest regeneration of deforested, abandoned land in Panama.” Ecology 85 (2004): 3,313–3,326.
Marcus, Bernard. Tropical Forests. Sudbury, Mass.: Jones and Bartlett, 2008.
Martin, Patrick H., Ruth e. Sherman, and timothy J. Fahey. “Forty years of tropical forest recovery from agriculture: Structure and floristics of secondary and old-growth riparian forests in the Dominican Repub-lic.” Biotropica 36 (2004): 297–317.
Montagnini, Florencia, and carl F. Jordan. Tropical Forest Ecology: The Basis for Conservation and Manage-ment. new york: Springer, 2005.
nepstad, Daniel, et al. “the end of deforestation in the Brazilian amazon.” Science 326 (2009): 1,350–1,351.
Phillips, Oliver l., et al. “Drought sensitivity of the ama-zon rain forest.” Science 323 (2009): 1,344–1,347.
Plotkin, Mark J. Tales of a Shaman’s Apprentice: An Eth-nobotanist Searches for New Medicines in the Ama-zon Rain Forest. new york: Viking, 1993.
Rice, Stanley a. “an injured paradise.” chap. 1 in Green Planet: How Plants Keep the Earth Alive. new Bruns-wick, n.J.: Rutgers University Press, 2009.
Royte, elizabeth. The Tapir’s Morning Bath: Mysteries of the Tropical Rain Forest and the Scientists Who Are Trying to Solve Them. Boston: Houghton Mifflin, 2001.
Schlawin, Justin R., and R. a. Zahawi. “ ‘nucleating’ suc-cession in recovering neotropical wet forests: the leg-acy of remnant trees.” Journal of Vegetation Science 19 (2008): 485–492.
Spray, Sharon l., and Matthew D. Moran. Tropical Defor-estation. lanham, Md.: Rowman and littlefield, 2006.
Stokstad, erik. “Debate continues over rain forest fate—with a climate twist.” Science 323 (2009): 448.
terborgh, John. Requiem for Nature. covelo, calif.: island Press, 1999.
United nations. “Un-ReDD Programme: the United nations collaborative Programme on Reducing emis-sions from Deforestation and Forest Degradation in Developing countries.” available online. URl: http://www.un-redd.org. accessed august 8, 2011.
Vazquez-yanes, c., and a. Orozco-Segovia. “Patterns of seed longevity and germination in the tropical rain forest.” Annual Review of Ecology and Systematics 24 (1993): 69–87.
Vieira, i. c. G., christopher Uhl, and Daniel nepstad. “the role of the shrub Cordia multispicata cham. as a succession ‘facilitator’ in an abandoned pas-ture, Paragominas, amazonia.” Vegetatio 115 (1994): 91–99.
White, Mel. “Borneo’s moment of truth.” National Geo-graphic, november 2008, 36–63.
Willson, Mary F., and F. H. J. crome. “Patterns of seed rain at the edge of a tropical Queensland rain forest.” Jour-nal of Tropical Ecology 5 (1989): 301–308.
Zahawi, R. a. “instant trees: Using giant vegetative stakes in tropical forest restoration.” Forest Ecology and Management 255 (2008): 3,013–3,016.
tropical seasonal forests tropical forests, found between the tropic of cancer and the tropic of capricorn, do not have winters. Most tropical forests are rain forests, in which there is no dry sea-son. the year-round rainfall occurs because water vapor condenses and rains down upon the tropics. near the northern and southern edges of the tropi-cal zone, however, air from which the moisture has rained out descends toward the surface of the earth, causing drier conditions. at about 30° north
xviii+598_Biodiversity_F.indd 535 11/17/11 1:13 PM
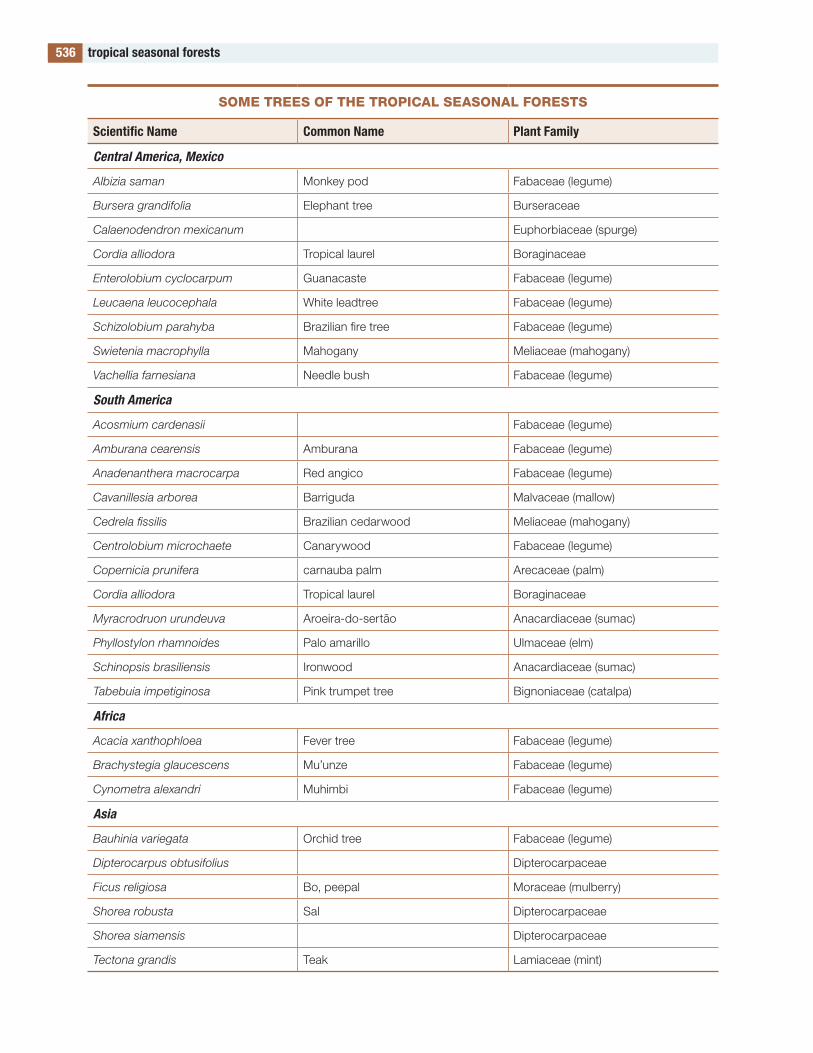
first entry536 tropical seasonal forests 536
some trees of the tropiCal seasonal forests
Scientific Name Common Name plant family
Central America, Mexico
Albizia saman Monkey pod Fabaceae (legume)
Bursera grandifolia Elephant tree Burseraceae
Calaenodendron mexicanum Euphorbiaceae (spurge)
Cordia alliodora Tropical laurel Boraginaceae
Enterolobium cyclocarpum Guanacaste Fabaceae (legume)
Leucaena leucocephala White leadtree Fabaceae (legume)
Schizolobium parahyba Brazilian fire tree Fabaceae (legume)
Swietenia macrophylla Mahogany Meliaceae (mahogany)
Vachellia farnesiana Needle bush Fabaceae (legume)
South America
Acosmium cardenasii Fabaceae (legume)
Amburana cearensis Amburana Fabaceae (legume)
Anadenanthera macrocarpa Red angico Fabaceae (legume)
Cavanillesia arborea Barriguda Malvaceae (mallow)
Cedrela fissilis Brazilian cedarwood Meliaceae (mahogany)
Centrolobium microchaete Canarywood Fabaceae (legume)
Copernicia prunifera carnauba palm Arecaceae (palm)
Cordia alliodora Tropical laurel Boraginaceae
Myracrodruon urundeuva Aroeira-do-sertão Anacardiaceae (sumac)
Phyllostylon rhamnoides Palo amarillo Ulmaceae (elm)
Schinopsis brasiliensis Ironwood Anacardiaceae (sumac)
Tabebuia impetiginosa Pink trumpet tree Bignoniaceae (catalpa)
Africa
Acacia xanthophloea Fever tree Fabaceae (legume)
Brachystegia glaucescens Mu’unze Fabaceae (legume)
Cynometra alexandri Muhimbi Fabaceae (legume)
Asia
Bauhinia variegata Orchid tree Fabaceae (legume)
Dipterocarpus obtusifolius Dipterocarpaceae
Ficus religiosa Bo, peepal Moraceae (mulberry)
Shorea robusta Sal Dipterocarpaceae
Shorea siamensis Dipterocarpaceae
Tectona grandis Teak Lamiaceae (mint)
xviii+598_Biodiversity_F.indd 536 11/17/11 1:13 PM

tundra 537
and south latitude, the dry air results in a band of deserts. in the transitional zone between tropics and deserts, between about 10° and 20° north and south latitude, there are tropical forests that have dry seasons: the tropical seasonal forests. the dom-inant trees of these forests are usually deciduous, losing their leaves during the dry season. evergreen broadleaf trees may dominate tropical seasonal for-ests with access to groundwater supplies, or that have infertile soil. in asia, where the wet season comes from monsoon rains, the forests are called monsoon forests.
tropical seasonal forests are common in Mexico and central america, where leguminous trees pre-dominate (see table on page 536). When the larger trees shed their leaves at the beginning of the dry season, many smaller trees of the genus Acacia retain their leaves. Retaining leaves during the dry season may make a tree the highly visible target of herbi-vores, but these acacia trees are protected by ferocious ants with which they have a mutualistic relationship.
tropical seasonal forests also dominate areas around the amazon rain forest. the caatinga of northeastern Brazil has short deciduous trees mixed with evergreen palm trees. cacti may grow in the openings between these trees. Some areas of Bolivia also have tropical dry forests.
acacias and related leguminous trees also domi-nate the tropical dry forests of africa, which are between the rain forests and the savannas. acacias are also the most important trees in the savannas.
the dry forests of india and indochina contain trees that are relatives of South asian rain forest species, for example the dipterocarps. teak, a tree whose timber was very important in shipbuilding in earlier centuries, is one of the few tree-sized species in the mint family. Many mammal and bird species live in these forests, including gibbons (Hylobates concolor), monkeys such as the langur Pygathrix nemaeus, asian elephants (Elephas maximus), and tigers (Panthera tigris), which also live in rain forests.
See also tropical rain forests and deforestation.
Further readingBecerra, Judith X. “timing the origin and expansion of
the Mexican tropical dry forest.” Proceedings of the National Academy of Sciences USA 102 (2005): 10,919–10,923.
Bowden, charles, and Jack W. Dykinga. The Secret Forest. albuquerque: University of new Mexico Press, 1993.
Bullock, Stephen H., and J. arturo Solis-Magallanes. “Phe-nology of canopy trees of a tropical deciduous forest in Mexico.” Biotropica 22 (1990): 22–35.
espiríto-Santo, Mário M., et al. “Sustainability of tropi-cal dry forests: two case studies in southeastern and
central Brazil.” Forest Ecology and Management 258 (2009): 922–930.
Högberg, Peter. “Root symbioses of trees in african dry tropical forests.” Journal of Vegetation Science 25 (2009): 393–400.
Justiniano, Marcos Joaquin, and todd S. Fredericksen. “Phenology of tree species in Bolivian dry forests.” Biotropica 32 (2000): 276–281.
Kennard, Deborah K. “Secondary forest succession in a tropical dry forest: Patterns of development across a 50-year chronosequence in lowland Bolivia.” Journal of Tropical Ecology (2002) 18: 53–66.
Marod, Dokrak. “effects of drought and fire on seedling survival and growth under contrasting light conditions in a seasonal tropical forest.” Journal of Vegetation Science 15 (2004): 691–700.
Swaine, M. D. “characteristics of dry forest in West africa and the influence of fire.” Journal of Vegetation Science 8 (1992): 365–374.
Vieira, Daniel l. M., et al. “effects of habitat, cattle grazing and selective logging on seedling survival and growth in dry forests of central Brazil.” Biotropica 39 (2006): 269–274.
———, and aldicir Scariot. “Principles of natural regenera-tion of tropical dry forests for restoration.” Restora-tion Ecology 14 (2006): 11–20.
tundra tundra is the plant community that lives in the coldest parts of the earth that are capable of supporting a plant cover. it consists of short perennial plants, both herbaceous and woody. Only the polar deserts, in which very few plants grow, are colder.
arctic tundra is found in a ring around the arctic Ocean: northern alaska and canada, extreme northern europe, and northern Siberia. alpine tundra is found on the tops of the tallest mountains in north america, europe, and asia. During the most recent ice age, tundra was found in a latitudinal line across the middle of what is now the United States. as the glaciers retreated, many tundra plants moved northward by seed dis-persal to their present arctic locations. Some of the plants, however, migrated up to the tops of mountains, where they are found today in the high-est elevations of the Rocky Mountain system (see photograph on page 538). the alpine tundra of the Sierra nevada of california also has some of the same species of plants as the arctic tundra. Because the alpine tundra of the Sierra nevada is drier than the tundra of the cascades and the arctic, it has a much less extensive plant cover and includes some plant species whose evolutionary ancestors came from the desert rather than from the arctic tundra. extensive alpine tundra can be found in Sequoia, yellowstone, and Rocky Mountain national Parks.
xviii+598_Biodiversity_F.indd 537 11/17/11 1:13 PM

first entry538 tundra 538
it is the overwhelmingly cold conditions that define nearly every characteristic of the tundra. the winters are long and extremely cold. the only plants that can survive are those that are short enough to be protected by the snow in the winter. the effect is also a cause: there are no tall trees to slow down the wind. in the brief summer, the snow melts, as does the surface layer of soil. However, below this depth, the soil remains permanently fro-zen (permafrost). the shallow soil is another reason that tall trees do not live in the tundra. the tundra plants include sedges, wildflowers, and short wil-lows (see table on page 539). nearly all are peren-nials, for the growing season is too short to allow an annual plant to complete its life cycle in a single season. the permafrost also inhibits the ability of melted water to drain away. the tundra is very wet, even though in some cases the amount of precipi-tation is low enough that the habitat would, were it not for the permafrost, qualify as a desert. the cold, wet conditions mean that what little soil is available to the plants is poor and acidic. the pools
of water also hatch massive hordes of mosquitoes. the alpine tundra does, however, provide food for large herds of grazing mammals, such as caribou and musk oxen. the mammals of the alpine tundra are small, such as the pika (see table on page 539).
Some tundra plant species, such as the moun-tain avens (Dryas octopetala) and the dwarf azaelea (Loiseleuria procumbens), grow in arctic and alpine tundra worldwide. Other genera of plants, such as the Ranunculus buttercups and Polemonium sky pilots, have different species in each tundra region. this has occurred because speciation has proceeded to different extents in different plant genera. a com-pletely different set of plant species has evolved in the páramo tundra of South america, because this region was never connected with the alpine tundra during the ice ages.
Despite its low productivity, the tundra supports many animals, such as reindeer and musk oxen. the large size of these mammals (and also of the now-extinct mammoths and mastodons) is beneficial for retaining body heat during cold winters. Smaller tundra mammals, such as pikas, hibernate. Some tundra mammals, such as the reindeer (called cari-bou in north america), range across the entire arctic tundra. Musk oxen are native only to north ameri-can arctic tundra. Pikas, which are small relatives of the rabbit, live in alpine tundra worldwide, but each region has different species.
in some drier spots in the tundra, wildflowers occur in the form of cushion plants, such as the dwarf azalea. composed of several different species, cush-ion plants really look like little cushions or pillows on the ground. this condensed growth form protects the stems from the effects of the wind and allows dead leaves to build up a rich soil directly underneath the plants. they therefore create tiny zones of good growing conditions immediately around them.
Pollination is a risky business in the tundra, as the weather conditions do not reliably allow insect pollinators to be present. Some plant species, such as the bistort Polygonum viviparum and the grass Poa alpina var. vivipara, have evolved the ability to pro-duce bulblets where they would otherwise produce flowers. Because the bulblets fall to the ground after they have already begun to grow, these plants are said to be viviparous (having “live birth”).
Despite their similarities in temperature con-ditions, arctic and alpine tundra experience very different sunlight conditions. in the arctic tundra, the winter has long and often continuous nights, whereas the summer has long days when the sun may not set for several weeks. in the alpine tundra, days and nights are of more equal length: the ecua-dorean páramo region and the alpine grasslands of Papua new Guinea experience approximately
Flowers bloom abundantly in the alpine tundra during the brief summer in the Rocky Mountains of Colorado. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 538 11/17/11 1:13 PM
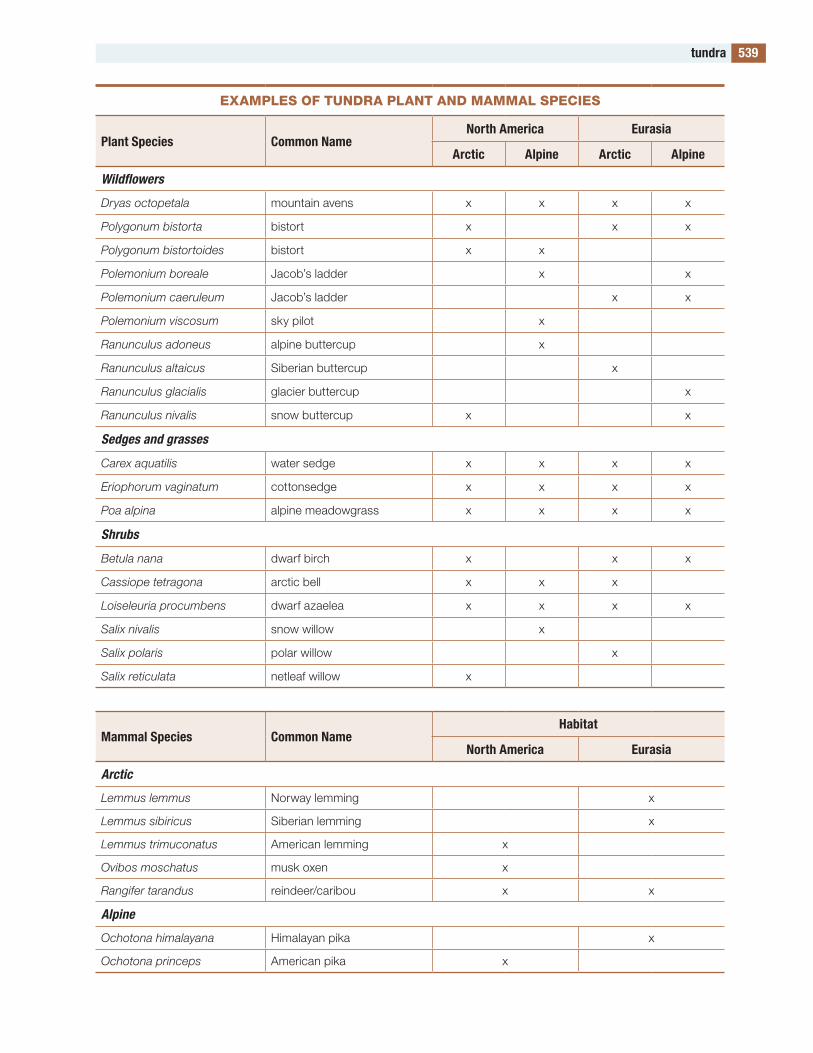
tundra 539
examples of tundra plant and mammal speCies
plant Species Common NameNorth america Eurasia
arctic alpine arctic alpine
Wildflowers
Dryas octopetala mountain avens x x x x
Polygonum bistorta bistort x x x
Polygonum bistortoides bistort x x
Polemonium boreale Jacob’s ladder x x
Polemonium caeruleum Jacob’s ladder x x
Polemonium viscosum sky pilot x
Ranunculus adoneus alpine buttercup x
Ranunculus altaicus Siberian buttercup x
Ranunculus glacialis glacier buttercup x
Ranunculus nivalis snow buttercup x x
Sedges and grasses
Carex aquatilis water sedge x x x x
Eriophorum vaginatum cottonsedge x x x x
Poa alpina alpine meadowgrass x x x x
Shrubs
Betula nana dwarf birch x x x
Cassiope tetragona arctic bell x x x
Loiseleuria procumbens dwarf azaelea x x x x
Salix nivalis snow willow x
Salix polaris polar willow x
Salix reticulata netleaf willow x
mammal Species Common Namehabitat
North america Eurasia
Arctic
Lemmus lemmus Norway lemming x
Lemmus sibiricus Siberian lemming x
Lemmus trimuconatus American lemming x
Ovibos moschatus musk oxen x
Rangifer tarandus reindeer/caribou x x
Alpine
Ochotona himalayana Himalayan pika x
Ochotona princeps American pika x
xviii+598_Biodiversity_F.indd 539 11/17/11 1:13 PM

first entry540 tundra 540
12-hour days all year. tropical alpine tundra, unlike the arctic tundra, is bombarded with intense ultra-violet radiation.
tundra intergrades with boreal forest. a tran-sitional zone that can be 60 miles (100 km) in width connects the arctic tundra and the boreal forest in canada. the boreal forest dominates the warm south-facing well-drained slopes in this zone. the transitional zone between alpine tundra and subalpine forest is characterized by stunted subalpine trees (krummholz) that are often short enough to be protected by snow in the winter and which may have branches only on the leeward side where winter winds have not killed them (see fig-ure). the krummholz trees form a vaguely defined timberline.
Because alpine tundra is wet, acidic, and poor in nutrients, many arctic tundra plant species found refuge in bogs as the glaciers retreated northward. examples include sphagnum moss and the sedges Eriophorum vaginatum and Carex aquatilis.
Global warming threatens the tundra in two ways. alpine tundra can only survive on cold moun-taintops; if mountaintops become even a little bit
warmer, as a consequence of global warming, forests may grow all the way to the top, resulting in the elimination of the alpine tundra. in addition, arctic tundra now experiences warmer and drier condi-tions. in 2007, a fire spread through the tundra of alaska’s north Slope, burning more area than all north Slope fires back to 1950 combined. Fires are sufficiently rare in the tundra that recovery from them is very slow. Because global warming is occur-ring more rapidly in the arctic than anywhere else in the world, the arctic tundra is being closely studied. the deep organic matter in the soils, accumulated and frozen over millennia, will almost certainly be a major source of carbon dioxide and methane in the atmosphere.
the arctic tundra on the north slope of alaska is also threatened by oil extraction. While contro-versy surrounded the opening of the arctic national Wildlife Refuge to oil extraction, the federal govern-ment went ahead and leased petroleum rights on the adjacent land and offshore areas. the reason that oil extraction may prove disastrous is that the slow growth of tundra plants will not allow significant recovery from any disturbances from construction. Ruts caused by vehicles driving through tundra mud often look as fresh as new even decades after they are formed.
See also boreal and subalpine forests; deserts, polar; disturbances, ecological; ice ages.
Further readingaerts, Rien. “nitrogen-dependent recovery of subarctic
tundra vegetation after simulation of extreme winter warming damage to Empetrum hermaphroditicum.” Global Change Biology 16 (2010): 1,071–1,081.
auger, Sarah, and Serge Payetfe. “Four millennia of wood-land structure and dynamics at the arctic treeline of eastern canada.” Ecology 91 (2010): 1,367–1,379.
Blok, D., et al. “Shrub expansion may reduce summer permafrost thaw in Siberian tundra.” Global Change Biology 16 (2010): 1,296–1,305.
campbell, Barbara J., et al. “the effect of nutrient deposition on bacterial communities in arctic tun-dra soil.” Environmental Microbiology 12 (2010): 1,842–1,854.
Forbes, Bruce c., et al. “Russian arctic warming and ‘green-ing’ are closely tracked by tundra shrub willows.” Global Change Biology 16 (2010): 1,542–1,554.
Hallinger, Martin, et al. “establishing a missing link: Warm summers and winter snow cover promote shrub expansion into alpine tundra in Scandinavia.” New Phytologist 186 (2010): 890–899.
Jones, Michael t., and Scott D. Smyers. “Occurrence of pond-breeding amphibians at alpine ponds in the
A whitebark pine (Pinus albicaulis) grows in a krumm-holz form near Lassen Peak in California. By growing near the ground, the tree is protected by the snow-pack from winter winds. (Stanley A. Rice)
xviii+598_Biodiversity_F.indd 540 11/17/11 1:13 PM

tundra 541
White Mountains, new Hampshire.” Northeastern Naturalist 17 (2010): 161–166.
nagy, laszlo, and Georg Grabherr. The Biology of Alpine Habitats. Oxford, U.K.: Oxford University Press, 2009.
Rice, Stanley a. “Plants create habitats.” chap. 8 in Green Planet: How Plants Keep the Earth Alive. Piscataway, n.J.: Rutgers University Press, 2009.
Sturm, Matthew. “arctic plants feel the heat.” Scientific American, May 2010, 66–73.
Sullivan, Patrick F. “Snow distribution, soil temperature, and late winter cO2 efflux from soils near the arc-tic treeline in northwest alaska.” Biogeochemistry 99 (2010): 65–77.
thomas, D. n., et al. The Biology of Polar Regions, 2nd ed. Oxford, U.K.: Oxford University Press, 2008.
xviii+598_Biodiversity_F.indd 541 11/17/11 1:13 PM

542
Wallace, alfred russel (1823–1913) British Evolutionary biologist alfred Russel Wallace (see photograph on page 543) is famous principally as the codiscoverer, along with charles Darwin, of natural selection as the principal mechanism of evolution. However, he was famous also for his travel writings, his contributions to the study of what is now called biodiversity, and his social activism. He was perhaps the most famous British naturalist of the late 19th century.
Born January 8, 1823, Wallace came from an intellectually active and sometimes fi nancially chal-lenged family. He worked with his brother as a surveyor, and then taught surveying skills at a small British college. He found increasingly more time for the study of local natural history (insects, plants, geology), especially after he began a friendship with another young naturalist, Henry Walter Bates.
Wallace and Bates decided to travel to South america to collect plant and animal specimens. Since neither was rich, they planned to sell the specimens to pay for the expedition. they went separate ways in South america. Both amassed large collections. Bates remained many years, studying the butterfl ies that were the fi rst and still most famous example of mimicry, a type still called Batesian mimicry. Wal-lace spent four years in South america and traveled farther in some of the river systems of South amer-ica than any european had ever gone. His studies in the amazon basin formed the basis of his developing thoughts about biogeography and about the process of evolution. He lived among, and closely studied, the ways and languages of the native peoples. in so doing, he developed an appreciation that tribal peoples have an intellectual capacity equal to that of civilized peoples, a belief that few european intel-
lectuals held at that time. as he returned to england with his collections, the ship caught fi re, and he lost everything except some of his drawings and notes. Upon returning home, he published and spoke about some of his observations to scientifi c societies and the general public.
Wallace’s accomplishments as a naturalist were not yet famous but were suffi cient for him to obtain support from the Royal Geographic Society for a journey to indonesia and new Guinea (at that time called the Malay archipelago). He spent nearly eight years there, from 1854 to 1862. He traveled 14,000 miles (22,530.8 km) and collected 125,660 biologi-cal specimens (including more than 1,000 species new to science). the book that he wrote upon his return, The Malay Archipelago, ranks as one of the most popular pieces of travel writing from the 19th century, partly because he published some of the earliest information about birds of paradise, orang-utans, and the native peoples of new Guinea. the book is still in print.
Wallace’s work in indonesia would have been suffi cient to guarantee him lifelong fame as a natu-ralist; he certainly had a lifetime of work awaiting him upon his return just to analyze and write about his specimens and travels. But it was an event that occurred at the midpoint of his journey, in 1858, that changed the course of scientifi c history. He had contracted malaria and was lying ill with it when his fevered mind came up with an explanation for how the process of evolution could work, a process that we today call natural selection. When he was well enough, he wrote his ideas in an article entitled “On the tendency of Varieties to Depart indefi -nitely from the Original type.” it was the word indefi nitely (meaning “without limit”) that made
W
xviii+598_Biodiversity_F.indd 542 11/17/11 1:13 PM

wallace, alfred russel 543
Wallace’s proposal distinct from orthodox scientific views. Wallace was ready to send the article back to england for publication. He knew, but did not much care, that his views might be controversial. How-ever, he decided to let another, more experienced, scientist review his article before publication. He sent his article to charles Darwin.
Wallace could not have known what an effect his article would have. like Wallace, Darwin had traveled extensively in tropical regions of the world and had spent much time (while ill) thinking about evolution. like Wallace, Darwin had come up with natural selection as a way of explaining how evolu-tion could occur. However, Darwin had done all of this almost exactly 20 years earlier. Darwin had not published any of his hypotheses on evolution, however, because he knew they would be controver-sial, and he wanted to amass all available evidence into a huge book that would answer every possible objection and settle the question. therefore, Darwin delayed 20 years, while Wallace was ready to publish right away.
Darwin was astounded that, after this long delay, another scientist had come up with the same ideas, even using the same term to describe the process. Darwin said, “i never saw a more striking coinci-dence. if Wallace had my manuscript sketch written out in 1842, he could not have made a better short abstract.” in consultation with the famous geolo-gist Sir charles lyell, Darwin decided that his 1842 summary of natural selection should be presented and published jointly with Wallace’s article. Darwin was ill, so lyell presented the joint Darwin-Wallace paper to the linnaean Society in 1858. Strangely, the scientists in attendance took little notice of the paper, nor did the president of the linnaean Society, who summarized the events of 1858 as unremark-able. Wallace, still in indonesia, knew nothing of this.
While in indonesia, Wallace noticed that the animals on the island of lombok differed strik-ingly from the animals of Bali, and those of Bor-neo differed from those of Sulawesi. the mammals of lombok and Borneo included many marsupials and few large predators, while the animals of Bali and Sulawesi were mostly placental mammals and included large predators, such as tigers. He noticed a difference also in the birds. in some cases, the islands with different animals were almost within sight of one another. the line he drew on the map to separate the two faunas is now known as Wal-lace’s line, and that part of indonesia is frequently called Wallacea. Wallace interpreted this, correctly, as resulting from the separate evolution of the two groups of animals. He could not have known that a deep ocean trench separated the two groups, which
were on different continental plates and therefore had little contact throughout geological history. Wallace’s line is considered one of the earliest major contributions to the science of biogeography.
after returning to england in 1862, Wallace spent three years organizing his specimens, present-ing papers to scientific meetings, and writing. He earned money from his famously interesting public presentations; one speaking tour took him as far as california. His striking appearance (he was tall, had a long beard, and snow-white hair for the last 40 years of his life) contributed to his public success. He married a woman almost 20 years younger than he but who shared his passion for gardening and natural history. later, they journeyed overseas on botanizing expeditions. although his collections and writings proved profitable, his investments did not. He had to take on small editing and speaking assign-ments for pay and move his family progressively far-ther from london to find cheaper accommodations. eventually, Darwin intervened to convince the Brit-ish government to give Wallace an annual stipend in recognition of his services to science.
Wallace continued his studies of evolution and advanced into other areas of science. His paper,
Alfred Russel Wallace in old age (The Linnean Society)
xviii+598_Biodiversity_F.indd 543 11/17/11 1:13 PM

first entry544 wallace, alfred russel544
“the Origin of Human Races and the antiquity of Man Deduced from the theory of natural Selec-tion,” presented in 1864, preceded Darwin’s book The Descent of Man by seven years. Wallace pub-lished a paper in 1863 that gave an evolutionary explanation for the hexagonal construction of the cells in beehives, a topic Darwin incorporated into later editions of the Origin of Species. Wallace also published a theory of glaciation and evolutionary explanations of mimicry, protective coloration and color vision in animals, bird migration, and the necessity of aging. He was one of the first to present evidence that the Pleistocene extinction may have been caused by overhunting by humans. His writings were among the earliest that contrasted different kinds of life history in animals. His ideas about the evolution of reproductive isolation have even been called the Wallace effect. He wrote in 1907 that the ice caps of Mars were carbon dioxide (dry ice), not water.
Wallace also expanded his studies into many different areas throughout the remainder of his life. He began to give serious time and attention to social issues, in which he defended the causes of the work-ing man and the oppressed. His views may be mainly attributed to his experiences with amazonian, indo-nesian, and Papuan peoples and to his exposure to socialism when he worked as a surveyor. among the social issues he championed are:
● Land nationalization. Wallace called for the protection of rural lands and histori-cal monuments, as well as the formation of greenbelts and parks, for the preservation of nature but also for the recreation of the lower and middle classes.
● Labor reform. Rather than calling for strikes, Wallace in 1899 called for working men to lay aside a portion of their salaries into a fund that would allow them to buy stock in the company and thereby partly or wholly control the company. He also called for double-overtime pay rates. later in life he fully converted to socialism.
● Opposition to vaccination. this issue puz-zles many modern scientists: Why would one of the most prominent scientists in europe oppose vaccination? Wallace had both scientific and social reasons for it. Scientifically, he could demonstrate with epidemiological data that the incidence of contagious diseases began to decline before the introduction of vaccination. today, sci-entists know that he was correct: advances in public hygiene have proven at least as important as vaccination in improving pub-
lic health. Socially, Wallace saw vaccina-tion as the government’s way of trying to improve the lot of the poor without having to spend the money necessary for public sanitation. it was much cheaper and easier to jab poor people in the arm than to build sewers and provide safe drinking water.
● Women’s suffrage. He was one of the earli-est proponents of voting rights for women in male intellectual society.
● Consumer safety. Wallace proposed in 1885 that all manufactured items have labels that identify their component materials and that the government regulate standards for these materials—practices that are standard in all industrialized countries today.
● Eugenics. Wallace criticized the attempt, then popular among scientists, including Darwin’s nephew Francis Galton, to attri-bute social problems to the supposed genetic inferiority of the lower classes and nonwhite races. Wallace saw this as an abuse of sci-ence and an attempt to justify oppression.
in the late 1860s, Wallace became an adherent of spiritualism, which he remained for the rest of his life. as a scientist, he sought experimental confirma-tion, but he was satisfied with demonstrations that more skeptical scientists would not accept. He wrote more than 100 publications on spiritualism. His lecture on life after death, delivered mainly during his california speaking tour, was one of his most popular. His outspoken association with spiritual-ism, however, did not prevent him from receiving a continuous stream of scientific awards and honors.
Related to his belief in spiritualism was his accep-tance of what is now called the anthropic principle, in which he believed that the universe was designed for man—a perfect universe in which humans were intended to evolve. the rebuttal written by Samuel clemens (Mark twain), “Was the World Made for Man?” has become much more famous than Wal-lace’s original writings on the subject.
the publication of Wallace’s first article about natural selection jointly with charles Darwin in 1858 created the impression that Darwin and Wal-lace believed the same things about evolution. Wal-lace’s publications about evolution were to prove that this was not the case, at least with human evolution. Wallace distinguished between the material process of natural selection that produced the human body and a spiritual one that produced the human mind, a distinction Darwin never made. today, nearly all evolutionary scientists explain the origin of human behavior patterns in terms of evolutionary processes acting upon the brain. Many scholars assume that
xviii+598_Biodiversity_F.indd 544 11/17/11 1:13 PM

wilson, Edward o. 545
Wallace’s rejection of natural selection as an expla-nation for the origin of the human mind resulted from his irrational attraction to spiritualism. How-ever, evolutionary biologist Stephen Jay Gould and others have pointed out a different possibility. Both Darwin and Wallace were aware that tribal peoples had intelligence equal to that of civilized peoples: Darwin knew this from his experience with the Fue-gians, and Wallace from the amazonians and indo-nesians. But Wallace pointed out, further, that the tribal peoples did not need or use the high levels of intelligence of which they proved themselves capable when given european education. Wallace reasoned that if natural selection were the sole explanation of the origin of the human mind, then scientists would see tribal peoples with low intelligence. Why did these people have so much more intelligence than they used? (this argument is now rejected by anthropologists, who recognize that tribal peoples have very complex languages, customs, and oral tra-ditions.) Wallace, in other words, was taking natural selection more seriously, not less seriously, than Dar-win. Of course, Wallace’s acceptance of spiritualism made his rejection of the evolution of mind much easier.
Wallace remained intellectually active well into his old age. Between 1898 and 1910, mostly in his ninth decade, he published more than 4,000 pages of material. Wallace died november 7, 1913.
See also Bates, Henry Walter; biodiversity, concept of; biogeography; camouflage and mimicry; Darwin, charles; natural selection; Pleistocene extinction.
Further readingBrackman, arnold c. A Delicate Arrangement: The Strange
Case of Charles Darwin and Alfred Russel Wallace. new york: times Books, 1980.
camerini, Jane, ed. The Alfred Russel Wallace Reader: A Selection of Writings from the Field. Baltimore, Md.: Johns Hopkins University Press, 2001.
Fichman, Martin. An Elusive Victorian: The Evolution of Alfred Russel Wallace. chicago: University of chicago Press, 2004.
Raby, Peter. Alfred Russel Wallace: A Life. Princeton, n.J.: Princeton University Press, 2002.
Smith, charles H. “the alfred Russel Wallace Page.” avail-able online. URl: http://www.wku.edu/~smithch/index1.htm. accessed august 4, 2010.
Wallace, alfred Russel. The Malay Archipelago. Oxford, U.K.: John Beaufoy Publishing, 2009.
Wilson, edward o. (1929– ) American Evo-lutionary biologist edward Osborne Wilson has been at the leading edge of breakthroughs in evo-
lutionary science, entomology, biogeography, and the study of the diversity of species. He even helped to invent some of the terms that are now central to these subjects, such as biodiversity, biophilia, island biogeography, and sociobiology. He has also been an immensely popular writer on these subjects: On Human Nature and The Ants (with entomolo-gist Bert Hölldobler) have won Pulitzer Prizes, and The Diversity of Life and The Future of Life have been immensely popular books about biodiversity. in 2010 he published a novel, Anthill, that explores similarities between human and ant societies in an environmental context.
Born June 10, 1929, in Birmingham, alabama, Wilson grew up primarily in alabama and Flor-ida. at a young age he became fascinated with the natural world, and as a Boy Scout he spent a lot of time exploring the marshes and swamps, collecting insects. He became an amateur expert on things that most people did not see even if they looked straight at them. it was Wilson who, at about 12 years of age, was the first to report the presence of the red imported fire ant, which is now a major problem, in the United States. His youthful passion for under-standing the diversity of the natural world was the force that impelled his entire subsequent career.
He earned bachelor’s and master’s degrees at the University of alabama and worked as an ento-mologist for the alabama Department of conserva-tion, primarily studying the fire ants whose invasion he had discovered. it was at the university that he learned about evolutionary science, and from that point onward he questioned the religious founda-tions he had embraced as a youth. He completed his Ph.D. at Harvard University in 1955. He was invited to join the Harvard faculty in 1956, and he is still there, as Pellegrino University professor emeritus. He works as honorary curator in entomology at the Museum of comparative Zoology.
Receipt of a Harvard fellowship allowed him to travel soon after he began his faculty appointment in 1956. He studied ants in new Guinea and other Pacific islands. He was the first outsider to climb Mt. Boana, the crest of the central Sarawaget Mountains of new Guinea. He also traveled in the american tropics. He collected lots of ants, always with the goal of understanding the evolutionary history of the entire ant family Formicidae.
Wilson’s observations also made him think about how species diversity developed on islands. curious as always, when he returned to Harvard, he sat in on mathematics classes, recognizing that he needed to know more math if he was to develop general theories of population biology. He shared his understanding with the world in A Primer of Popu-lation Biology (with geneticist William H. Bossert),
xviii+598_Biodiversity_F.indd 545 11/17/11 1:13 PM

first entry546 wilson, Edward o.546
which is still one of the fundamental texts on the subject. With Robert H. Macarthur, an ecologist at Princeton University, Wilson developed the theory of island biogeography, which tied species immigra-tion and extinction to the area of islands and their distance from the mainland.
But is it possible to study island biogeography experimentally? in the late 1960s Wilson, with ecologist Daniel Simberloff, removed all of the ani-mals from mangrove islands of various sizes and distances from the mainland in the Florida Keys—a difficult task, especially when a hurricane is com-ing—and then documented the colonization of these islands by animals. His work transformed the study of island species from stories to a science and made island biogeography theory useful to conser-vation efforts.
Wilson’s studies of ants created breakthroughs of understanding in how insects communicate. He and collaborators worked out the principles of how insects communicate by chemicals. Subsequently, many researchers have found examples of chemical communication among vertebrates and even among plants.
Wherever he traveled, at home or abroad, he could not help but notice that human activity was destroying natural habitats and, in the process, replacing high-diversity ecological communities with low-diversity artificial habitats. He could see, before most scientists had thought about it, that species were being driven into extinction faster than anyone could even recognize them let alone study them. He knew that there was yet a lot to be learned from wild species of ants. For example, ants have high population densities yet they hardly ever expe-rience epidemic diseases. this is because their meta-pleural glands and some symbiotic bacteria produce chemicals that kill bacteria. Humanity needs new antibiotics, yet humans are driving many ant spe-cies into extinction. Wilson’s 1984 Biophilia argued that humans have a psychological need for contact with the natural world. in The Diversity of Life and The Future of Life, Wilson explained the causes and consequences of the sixth mass extinction, which humans are bringing upon the world, and what humans might be able to do to slow or stop it. not content with writing, Wilson has been active with the american Museum of natural History, conser-vation international, the nature conservancy, and the World Wildlife Fund. the destruction of biodi-versity, he writes, “is the folly our descendants are least likely to forgive us.”
Wilson was largely responsible for synthesiz-ing the modern understanding of insect behavior in his 1971 book, The Insect Societies. He applied these concepts across the entire animal kingdom
in his 1975 book, Sociobiology: The New Syn-thesis. there was only a little bit, the last chapter, about humans, but it sparked a vigorous contro-versy about the role of evolutionary biology in human behavior and created a field of study now sometimes called evolutionary psychology. Wil-son did not hesitate to make his message clear to the general public in his 1979 book, On Human Nature. Strong disagreement came from other sci-entists, including Harvard colleagues Stephen Jay Gould and Richard lewontin. Outside the aca-demic world, many activists believed that socio-biology provided an intellectual justification for oppression. at a 1978 meeting of the american association for the advancement of Science in Washington, D.c., Wilson was one of the platform scientists at a discussion of human evolution. Pro-testers chanted, “Racist Wilson you can’t hide, we charge you with genocide!” a protester jumped up on stage with a pitcher of water and poured it all over Wilson, saying, “Wilson, you are all wet!” evolutionary anthropologist napoleon chagnon helped push the protester off the stage, while Ste-phen Jay Gould took the microphone to denounce the protesters’ tactics. Meanwhile, Wilson just wiped off the water and continued with the meet-ing. no one who has met Wilson can believe that he in any way condones the misuse of sociobiology as a rationalization for political oppression.
if anyone could handle the challenge of try-ing to bring together all fields of knowledge into a single unified structure, it would be edward Wilson. His 1998 Consilience called for a return to origi-nal enlightenment ideals for bridging the sciences and the humanities. college education is a smor-gasbord of largely disconnected subjects that leaves undergraduates more annoyed than enlightened; the typical divisions of a newspaper (news, business, sports, leisure) are even more disconnected. Rather than to have science as just one area of study—equal perhaps to music, or as an occasional page in a newspaper—Wilson wants science, particularly evolutionary science, to be understood as the foun-dation of all human history and activity. naturally this approach has drawn attacks from scholars in the humanities who do not want their field to be seen as a subset of science. Wilson writes, “My truths, three in number, are the following: first, humanity is ulti-mately the product of biological evolution; second, the diversity of life is the cradle and greatest natural heritage of the human species; and third, philoso-phy and religion make little sense without taking into account these first two conceptions.” although Wilson admits his approach to consilience may be wrong, it is widely recognized as one of the most concise and organized attempts ever made.
xviii+598_Biodiversity_F.indd 546 11/17/11 1:13 PM

wilson, Edward o. 547
Wilson has received many of the highest awards that are available. there is no nobel Prize for evo-lutionary biology, but Wilson has received the cra-foord Prize, the award given by the Royal Swedish academy of Sciences for areas not covered by the nobel Prize, as well as the U.S. national Medal of Science. in addition to his two Pulitzer Prizes, Wilson has received prestigious prizes in Japan, France, italy, and Saudi arabia and from world conservation organizations. as much as he cherishes these prizes, he is still very grateful for the teaching awards he has received from the students of Har-vard University.
even though most people his age have decided to take it easy, Wilson still has major projects. His primary goal is to work on the “encyclopedia of life,” a project he helped launch, which will use modern internet technology to speed up the process of documenting the earth’s rapidly disappearing bio-diversity. His motto might be reflected in this quote: “love the organisms for themselves first, then strain for general explanations, and, with good fortune, discoveries will follow. if they don’t, the love and the pleasure will have been enough.” even though he is one of the most respected scientists of modern times, he will not hesitate to crouch down on the ground to look at an ant. He is still, at heart, a Boy Scout working on an insect merit badge.
See also biodiversity, concept of; invasive species; island biogeography.
Further readingcowley, Geoffrey. “Wilson’s world.” Newsweek, June 22,
1998, 58–62.Macarthur, Robert H., and edward O. Wilson. Theory of
Island Biogeography. Princeton, n.J.: Princeton Uni-versity Press, 1967.
Wilson, edward O. Anthill: A Novel. new york: norton, 2010.
———. Biophilia. cambridge, Mass.: Harvard University Press, 1984.
———. Consilience: The Unity of Knowledge. new york: Vintage Press, 1998.
———. The Creation: An Appeal to Save Life on Earth. new york: norton, 2006.
———. The Diversity of Life. cambridge, Mass.: Harvard University Press, 1992.
———. “the encyclopedia of life.” Trends in Ecology and Evolution 18 (2003): 77–80.
———. The Future of Life. new york: Vintage Press, 2002.———. Naturalist. Washington, D.c.: island Press, 1994.———. Nature Revealed: Selected Writings, 1949–2006.
Baltimore, Md.: Johns Hopkins University Press, 2006.———. On Human Nature. cambridge, Mass.: Harvard
University Press, 1978.———. Sociobiology: The New Synthesis. cambridge,
Mass.: Harvard University Press, 1975.———, and William H. Bossert. A Primer of Population
Biology. Sunderland, Mass.: Sinauer associates, 1971.———, and Bert Hölldobler. The Ants. cambridge, Mass.:
Harvard University Press, 1990.
xviii+598_Biodiversity_F.indd 547 11/17/11 1:13 PM

548
Appendix IChronology
ca. 350 B.C.E. aristotle attempts to classify all animals
ca. 250 B.C.E. theophrastus describes about 500 species of plants in his 10-volume Historia Plantarum
ca. 50 Pliny the elder publishes 37-volume Historia Naturalis
1682 John Ray defi nes the concept of a species and describes monocots and dicots in Methodus Plantarum Novae
1736 carolus linnaeus publishes the fi rst of 12 editions of Systema Naturae, a book that outlined a system for classifi cation of plants, animals, and minerals
1749 Georges-louis leclerc, comte de Buffon, publishes fi rst volume of His-toire Naturelle, the series in which he states that species are mutable
1753 carl linnaeus publishes Species Plantarum, introducing bionomial nomenclature
1798 thomas Malthus publishes Essay on the Principle of Population, which later stimulates the theory of natu-ral selection by charles Darwin and alfred Russel Wallace
1799 alexander von Humboldt begins sci-entifi c exploration of latin america
1802 Jean-Baptiste lamarck presents a theory of evolution based on the inheritance of modifi ed traits
1831 charles Darwin begins a scientifi c journey around the world on HMS Beagle
1848 Henry Walter Bates and alfred Rus-sel Wallace begin an insect-collecting expedition through amazonia, discovering astonishing levels of biodiversity
1859 charles Darwin publishes On the Origin of Species
1864 ernst Haeckel publishes two vol-umes of Generelle Morphologie der Organismen, in which he proposes a new kingdom, Protista; George Perkins Marsh publishes Man and Nature, one of the earliest books of environmentalism
1865 Gregor Mendel investigates inheri-tance patterns in peas
1866 Haeckel introduces the term ecology
1872 Sir c. Wyville thomson begins the Challenger expedition, during which thousands of new marine organisms are discovered; yellowstone becomes the fi rst United States national Park; fi rst tree-Planting Day (now arbor Day)
1875 congress prohibits unauthorized cut-ting of trees on federal government property
1878 John Wesley Powell publishes Report on the Lands of the Arid Regions of the United States, which calls for environmentally responsible develop-ment of western lands
1891 establishment of fi rst Forest Reserve (later called national Forests)
1892 John Muir founds the Sierra club, devoted to wilderness preservation
1898 Gifford Pinchot becomes chief of the Division of Forestry of the U.S. Department of agriculture and begins modern conservation-based forest management
1903 President theodore Roosevelt estab-lishes the fi rst federal wildlife refuge at Pelican island, Florida
1914 Death of the last passenger pigeon
1916 national Park Service created
xviii+598_Biodiversity_F.indd 548 11/17/11 1:13 PM

Chronology 549
1935 arthur tansley introduces the ecosys-tem concept; due to efforts of Hugh Hammond Bennett, the U.S. Soil conservation Service is established
1936 Death in australian zoo of the last thylacine (tasmanian tiger)
1949 aldo leopold’s Sand County Alma-nac proposes the land ethic
1951 the nature conservancy is estab-lished to promote ecological preserva-tion of private lands
1959 G. evelyn Hutchinson publishes “Santa Rosalia” article that focuses attention on processes that determine biodiversity
1962 Rachel carson publishes Silent Spring
1964 Wilderness act designates millions of acres of wilderness area in United States
1970 First earth Day; national environ-mental Policy act (nePa) estab-lishes the environmental Protection agency; clean air act
1972 clean Water act; pesticide DDt banned
1973 endangered Species act
1977 Robert Ballard and his team discover ecological communities based on
bacterial chemosynthesis surrounding hydrothermal vents; carl Woese pro-poses a third domain of life, archaea
1988 James Hansen brings evidence of the greenhouse effect to the attention of U.S. congress
1992 Un convention on Biological Diver-sity and Un Framework convention on climate change are adopted in Rio de Janeiro, Brazil, at the Un earth Summit meeting
1994 Discovery of newest animal phylum, cycliophora
1997 Kyoto Protocol adopted as first step in implementation of the Un Frame-work convention on climate change
2009 United nations climate change con-ference in copenhagen negotiated targets for carbon emission reduc-tions to succeed the Kyoto Protocol
2010 nagoya Protocol adopted, updating the Un convention on Biological Diversity
2010 Deepwater Horizon oil spill (BP) in Gulf of Mexico, largest oil spill to date
2011 Major earthquake in Japan causes meltdown of nuclear plants and release of radiation
xviii+598_Biodiversity_F.indd 549 11/17/11 1:13 PM

550
Appendix IIGlossary
acclimatization physiological adjustments by an individual organism that enhance its survival and reproduction
adaptation a characteristic of organisms in a pop-ulation that allows the organisms to have greater survival and reproduction, produced by natural selection acting upon that characteristic
adaptive radiation evolution of diverse species from a common ancestor
agriculture deliberate cultivation of fungi or plants by animals
allelopathy production by plants of toxins that inhibit the growth of other plants
allometry differences between organisms due to their different sizes, which are necessary to allow structural support or the exchange of matter and energy with the physical environment
amniotes vertebrates that produce eggs with shells that potentially resist desiccation. Reptiles, birds, and mammals are amniotes.
amphibian a vertebrate that usually has an aquatic juvenile phase and a terrestrial adult phase
angiosperm a seed plant that produces fl owers and fruits
animal a multicellular eukaryote that ingests chunks of food
archaea prokaryotes with unique cellular charac-teristics and that are usually extremophiles
arthropod an animal with an exoskeleton and jointed legs
autecology ecology of organisms responding to their physical environments
avoidance a method of acclimatization, plasticity, or adaptation that allows organisms to maintain their physiological processes by producing inter-nal conditions that are more favorable to their function than are the external conditions
bacteria prokaryotes with unique cellular charac-teristics and that are usually not extremophiles
benthic dwelling at the bottom of a body of waterbiodiversity the total genetic and species diversity
of organismsbiodiversity hotspot a small area with an unusu-
ally high number of species, many of them endemic
biogeochemical cycles movement of atoms or molecules (e.g., water, carbon, nitrogen, minerals) from the physical environment, through a food web, and back into the physical environment
biogeography the patterns of which kinds of organisms live in which locations
biomagnifi cation the concentration of toxic min-erals persistent organic pollutants (POPs) at higher trophic levels
biomimicry the conscious imitation of biological adaptations by human technology
biophilia human innate psychological love of nature and biodiversity
bioremediation use of organisms (primarily plants) to remove toxic compounds from natural environments, usually soil
birds terrestrial two-legged vertebrates with feathers
bog a wetland with acidic water that is low in nutrients
boreal forest a coniferous forest that grows in high latitude regions, in which summers are too short to allow the production of a new set of leaves every year
camoufl age adaptations that make animals dif-fi cult for their predators or their prey to see
carnivore organism (usually an animal) that obtains food by eating animals
chaparral dry shrubland maintained by a fi re cyclecharacter displacement process by which compe-
tition between two species causes lower fi tness of individuals of at least one of the species that are most similar to the individuals of the other
chordate animal that has a notochord along its back, associated with the main nerve bundle, at least during its embryonic development
cladistics classifi cation of species of organisms based on the pattern of evolutionary branches that produced them
class (taxonomic) in traditional classifi cation, the level into which phyla or divisions may be divided. each class consists of one or more orders.
cloud forests forests of short trees with abundant epiphytes that live in misty mountains
xviii+598_Biodiversity_F.indd 550 11/17/11 1:13 PM

Glossary 551
coevolution evolution of two or more species in response to one another
commensalism a symbiotic relationship between two species in which the commensal species ben-efits and the host species is unaffected
community, ecological all of the species in a given location and all of their interactions
competition, ecological interactions among two or more species that depend on a single resource, which is not abundant enough for all of them
conifer a seed plant that produces pollen in male cones and seeds in female cones
coniferous forest a forest dominated by conifer-ous trees, such as boreal and subalpine forests, or the forests of the relatively dry mountain regions of western north america
conservation human activities intended to protect wild species and their habitats, while allowing human access and limited use
consumers organisms that obtain their food from other organisms, living or dead, in contrast with producers, which use photosynthesis to produce their food
continental shelf edge of a continent that is below sea level
convergence evolution of similar adaptations by different lineages of organisms
coral bleaching loss of symbiotic photosynthetic organisms from corals, as a result of adverse envi-ronmental conditions, which causes the corals to starve
crustacean an arthropod that usually has numer-ous legs and numerous visible body segments
deciduous forest a forest dominated by trees that lose their leaves every autumn and grow a new set of leaves every spring
decomposition consumption of dead organisms, or the organic molecules released by other decom-posers, usually by microorganisms
deserts ecological communities with low rainfall and long rainless periods
deuterostomes animals with radial cleavage dur-ing embryonic development; includes chordates
dicot an angiosperm whose seeds have embryos with two leaves
dinosaurs reptiles that were the dominant verte-brates during much of the Mesozoic period and whose modern descendants are birds
disjunct a species geographically distant from closely related species, or a population sepa-rated from other populations of the same spe-cies by a distance that is sufficient to prevent interbreeding
dispersal (1) movement of offspring away from the parent; (2) movement of organisms that results in a new population in a distant location
disturbance, ecological damage to or destruction of the dominant plants in an ecological commu-nity, which may allow resources to become avail-able to surviving plants, or which may be severe enough to destroy an ecological community
DNA the molecule that stores genetic information in the cells of an organism. Dna can also be ana-lyzed, allowing the identification of the species, and can mutate, producing genetic diversity upon which evolution can act.
domain (taxonomic) in traditional classification, the highest category, which contains one or more kingdoms. the eukaryotic domain contains pro-tist, fungus, plant, and animal kingdoms.
ecological footprint the area of productive land that would be necessary to produce the resources that a person, or a nation, consumes
ecology interactions of organisms with their phys-ical or biological environments
ecotone the zone in which one ecological commu-nity intergrades into another
ecosystem an ecological community, together with the matter and energy with which it interacts
ecosystem services ecological processes that happen to benefit the survival and prosperity of human societies
ectotherm an animal that absorbs most of its body heat from its environment
edaphic factors physical and chemical factors related to the surface (e.g., soil or rocks) upon which plants grow and animals live
endemic species a species that lives only in one limited area
endotherm an animal that generates most of its body heat by internal physiological processes
energy pyramid loss of stored energy as the energy is transferred from one trophic level to the next
ephemeral organisms organisms that are dor-mant most of the time and flourish during briefly favorable periods
epigenetics patterns of inheritance of character-istics from one generation to another that result from some genes being inactivated
era a duration of geological time that ends with a major extinction event. each geological era con-sists of several geological periods.
estuary a marine brackish or salt marsh, often at the mouth of a river, in which many marine ani-mal species breed
eukaryote an organism whose cells contain nuclei, usually contain mitochondria, and may contain chloroplasts
eutrophication an explosive growth of photosyn-thetic bacteria as a result of phosphate pollution
evolution genetic changes over time in populations
xviii+598_Biodiversity_F.indd 551 11/17/11 1:13 PM

first entry552 Glossary552
exaptation a characteristic of organisms in a pop-ulation that allows the organisms to have greater reproduction, produced by natural selection that acts upon a different characteristic
exoskeleton a hard outer covering of an animal, usually an arthropod, that provides the principal support for its body weight
extinction death of the last member of a speciesextremophile an organism that lives in very hot,
very salty, or very acidic conditions, or under conditions of high pressure or radiation. Most extremophiles are archaea.
family (taxonomic) in traditional classification, the level into which orders may be divided. each family consists of one or more genera.
feedback the effect of one process upon another. Positive feedback occurs when one process makes another process occur more rapidly; negative feedback, when it makes it occur more slowly.
fishes aquatic vertebrates without legsfitness, evolutionary the successful production of
offspringfood chain a series of organisms, usually in which
herbivores eat plants and carnivores eat herbi-vores, and all of the organisms, upon death, are metabolized by decomposers
food web a complex set of food chains, in which each organism eats and/or is eaten by organisms of more than one species
fossilization partial or complete transformation of an organism, or something produced by that organism (e.g., a footprint), into mineral form
founder effect a genetic pattern in a population that results from the genetic characteristics of the individuals that started the population
fungi multicellular eukaryotes that digest cells in their environment and absorb the digested molecules
Gaia an image for understanding the entire earth as a living ecosystem whose physical characteris-tics are modified by its organisms
genetically modified organisms (GMO) organ-isms, primarily bacteria, crop plants, and live-stock animals, whose genes have been modified by laboratory techniques
genetic bottleneck loss of genetic diversity in a population as a result of a reduction in popula-tion size
genetic drift genetic changes that occur by chance, rather than by selection, in small populations
genetics patterns of inheritance of characteristics from one generation to another that result from different alleles (versions) of genes
genus (pl. genera) in traditional classification, the level into which families may be divided. each genus consists of one or more species.
grassland an ecological community in which grasses make up most of the biomass, even if other kinds of plants constitute most of the species
greenhouse effect global warming that results from the accumulation of greenhouse gases (e.g., carbon dioxide) in the atmosphere
gymnosperm a seed plant that produces seeds that are not enclosed in a fruit
habitat a local ecological community in which an organism lives
habitat fragmentation division of a habitat into small portions by barriers (e.g., roads) that many organisms cannot readily cross
Hardy-Weinberg equilibrium the pattern of alleles (versions of genes) that occurs in populations when selection, genetic drift, nonrandom mating, mutation, immigration, and emigration are not occurring
herbivore an animal that subsists primarily by eat-ing plant material
homeotherm an organism that maintains a rela-tively constant body temperature
humus organic matter in the soil that results pri-marily from the decomposition of dead plant parts
hybridization the process by which individuals of (usually) two different species produce offspring by sexual reproduction
hydrostatic skeleton structural support for an animal that results from sacs of fluid in which muscles can produce pressure
hydrothermal vents deep-ocean volcanic erup-tions that release molecules such as hydrogen sul-fide, which bacteria can metabolize and upon which a food web can be based
ice age a period of glaciation, in which the accu-mulation of snow produces glaciers that cover large areas. Glaciation alternates with glacial melting.
inbreeding production of offspring by sexual reproduction occurring between close genetic rel-atives. the offspring often reveal otherwise hid-den genetic mutations.
insect an arthropod with six legs and three body parts; insects are the most diverse group of ani-mals on earth
integrated pest management (IPM) control of agricultural pests primarily by natural means, with a limited amount of pesticide
invasive species species that enter a new habitat and undergo explosive population growth
invertebrate an animal without an internal skel-eton or backbone
island biogeography the pattern by which new species arrive at or evolve on a geographical or ecological island, and by which they become extinct, as a result of the size of the island and
xviii+598_Biodiversity_F.indd 552 11/17/11 1:13 PM

Glossary 553
its distance from the mainland or from larger populations
isolating mechanism a biological process that prevents two populations or two species from interbreeding, even if they are in close proximity
keystone species a species that significantly affects the survival of many other species in an ecological community and whose extinction would greatly alter the community
lichen an organism that is a symbiotic combination of single photosynthetic cells and strands of fungus
life history the pattern of growth, reproduction, and longevity of an organism
mammal a vertebrate with hair in which the females can produce milk
marsh a wetland dominated by herbaceous plants, primarily grasses and sedges
marsupial mammal a mammal in which very young embryos are born and which must crawl into a pouch to find a nipple
meiosis sexual cell division that separates pairs of chromosomes from one another and reduces the number of chromosomes by half
mesophication the process by which secondary succession, in the absence of fire, produces a for-est that is dominated by species of trees that require wetter (more mesic) conditions
metamorphosis the development of an animal larva into a very different adult form, usually dur-ing a period of pupation
microhabitat a small area within a larger habitat, within which individuals of a species spend most of their time
mimicry characteristics of an organism that imi-tate some aspect of its environment (e.g., a dif-ferent species of organism) in such a way that it is protected from danger or otherwise gains a reproductive advantage
monocot an angiosperm whose seeds have embryos with one leaf
monoculture an ecological community (usually an agricultural one) that consists almost entirely of a single dominant species or even a single dominant breed
monophyletic a cladistic category of organisms, all of which trace their ancestry to a single ances-tral population
monotreme mammal a mammal whose females lay eggs but feed their young with milk
mutualism a symbiotic relationship between two species in which both species benefit
mycorrhizae fungi that live within plant roots and absorb minerals from the soil, thus supplementing the mineral supply available to the plant
natural selection the process by which some indi-viduals in a population reproduce more than oth-
ers as a result of their genetic characteristics, in response to the physical environment or to other species
natural systems agriculture a system of agricul-ture that imitates natural ecosystems, particularly with respect to soil conservation, soil minerals, and pest control
niche the set of conditions in which an individual, population, or species survives and reproduces best
nitrogen fixation bacterial transformation of gas-eous nitrogen into ammonia
omnivore an animal that eats food from both plant and animal sources
order (taxonomic) in traditional classification, the level into which classes may be divided. each order consists of one or more families
organic agriculture agriculture that avoids the use of industrially produced chemicals
paraphyletic a cladistic category of organisms, some but not all of which trace their ancestry to a single ancestral population
parasitism a symbiotic relationship between two species in which the parasite species benefits but the host species is harmed
pelagic dwelling in deep ocean watersperiod a duration of geological time that ends with
a minor extinction event. each geological era con-sists of several geological periods.
permafrost permanently frozen soil in the tundrapersistent organic pollutants (POPs) organic
molecules, usually of industrial origin, that do not readily decompose but remain in the environment for long periods
photosynthesis the process by which some bacte-ria, or the chloroplasts of some eukaryotes, trans-fer sunlight energy into the production of sugar molecules; the basis of most food webs
phylum (taxonomic) in traditional classification, the level into which domains may be divided. each phylum consists of one or more classes. this level is called “division” in plants, fungi, protists, and prokaryotes.
placental mammal a mammal in which the fetus remains inside the body of the mother for a pro-longed period and is fed by the mother through a placenta
plankton floating (usually aquatic) organismsplasticity, phenotypic development of different
characteristics in response to different environ-mental conditions by organisms with the same types of genes
poikilotherm an animal whose body temperature fluctuates with environmental temperature
pollination transfer of pollen from the flower or cone of one seed plant to the flower or cone of another seed plant, usually of the same species
xviii+598_Biodiversity_F.indd 553 11/17/11 1:13 PM

first entry554 Glossary554
polyculture an ecological community (usually an agricultural one) that consists of more than one dominant species
polymorphism differences within a population of individual characteristics, resulting from differ-ences in alleles (versions of genes)
polyphyletic a cladistic category of organisms that trace their ancestry to more than one ancestral population
population all the organisms of a species that are in close enough proximity to one another to cus-tomarily interbreed
predator an organism (usually an animal) that eats other organisms (such as seeds or prey)
primates an order of placental mammals with acute vision and opposable thumbs, which includes monkeys and apes (such as humans)
producers organisms that produce their food by photosynthesis
productivity the total amount of photosynthetic production of organic molecules in an ecologi-cal community (gross primary productivity), or the amount that remains after cellular respiration (net primary productivity), or the production of organic molecules at any trophic level
prokaryotes cells (usually very small) whose Dna is not contained within a nucleus
protists organisms that consist of a single eukary-otic cell or of eukaryotic cells that are not orga-nized into tissues
protostomes animals with spiral cleavage during embryonic development
rain forest a forest that receives a great deal of rain, usually uniformly throughout the year
rain shadow interception of atmospheric moisture on the windward side of a mountain range, result-ing in reduced rainfall on the leeward side
reef a large mineral deposit that results from secre-tion of minerals (mostly calcium carbonate) by aquatic organisms
relictual population a small isolated popula-tion that was once part of a larger, widespread population
reptile an amniote other than a bird or mammalrespiration, cellular the process by which cells
(or mitochondria within cells) release metabolic energy from sugar
resurgence rapid growth of a pest population after the pesticide that was used to control it is no longer in active circulation within the ecosystem
root nodules root structures, primarily in legu-minous plants and alders, within which bacteria carry out nitrogen fixation
salinization buildup of salt in soil, usually agricul-tural soil, as a result of evaporation of water (usu-ally irrigation water) that contains salt
savanna a grassland with scattered treesseed a reproductive structure in which a plant
embryo, with a stored food supply, is surrounded by a protective coat
sexual selection the process by which some indi-viduals in a population reproduce more than oth-ers as a result of their genetic characteristics and of success in gaining access to members of the opposite gender
speciation evolution of two or more species from a single ancestral population
species a set of naturally interbreeding populationsspider an arthropod with eight legs and two body
parts, which usually obtains its food by injecting digestive enzymes into a prey animal and con-suming the resulting molecules
spores single reproductive cells produced by meio-sis in protists, fungi, and plants, or the resting phase of a bacterium that can resist adverse envi-ronmental conditions
subalpine forest a coniferous forest that grows at high elevations, in which summers are too short to allow the production of a new set of leaves every year
succession, ecological changes in ecological communities that result after disturbance, with some species replacing others as the dominant species
succulents plants that store water in their tissuesswamp a wetland dominated by woody species
(e.g., mangroves)symbiogenesis the origin of a new evolutionary
lineage as the result of the symbiotic merger of different species, usually on the cellular level
symbiosis the interaction of two species in which the interaction is of overwhelming importance to the fitness of at least one of the species
symmetry the balance of an organism’s structure around one or more axes
synecology ecology of organisms responding to other organisms
taxonomy the classification of organisms on the basis of their adaptations and/or evolutionary history
tetrapods vertebrates with four limbs (legs and arms)
timberline the ecotone between tundra and foresttolerance a method of acclimatization, plasticity,
or adaptation that allows organisms to maintain their physiological processes despite adverse inter-nal conditions
transpiration evaporation of water from leavestrophic cascade effects of populations of a key-
stone species on other trophic levels in the eco-logical community
trophic levels levels of a food chain, such as pro-ducers, herbivores, and predators
xviii+598_Biodiversity_F.indd 554 11/17/11 1:13 PM

Glossary 555
tundra an ecological community with long winters and permafrost and which consists of small plants
vascular plant a plant that can transport water through pipelike cells such as xylem
vertebrate an animal with an internal skeleton, including a backbone
vicariance divergence of the species in an ecologi-cal community as a result of adaptive radiation
xviii+598_Biodiversity_F.indd 555 11/17/11 1:13 PM

556
Appendix IIIFurther Resources
BooKsBarrow, Mark V., Jr. Nature’s Ghosts: Confronting
Extinction from the Age of Jefferson to the Age of Ecology. University of chicago Press, 2009. this book reviews the changing concept of extinction, from the enlightenment era, when many scholars believed extinction was impossible, to a modern understanding of the role of extinction in evolu-tion and conservation.
Bazzaz, Fakhri a. Plants in Changing Environments: Linking Physiological, Population, and Com-munity Ecology. new york: cambridge Univer-sity Press, 1996. this book explains how plants respond to one another within populations and within ecological communities, in conditions that change over ecological time and as a result of human activity.
Begon, Michael, colin R. townsend, and John l. Harper. Ecology: From Individuals to Ecosystems,4th ed. Oxford, U.K.: Blackwell, 2006. this text-book is a thorough overview of all aspects of ecology.
carson, Rachel. Silent Spring. new york: Houghton Miffl in, 1962/2002. this book, more than any other, started the modern ecological movement. it thoroughly documented the effects of pesticides on entire ecosystems, including what we today call biomagnifi cation, and the evolution of pesti-cide resistance, whereby humans were polluting their environments in order to ineffectively control pests. yet it is so delightfully written that millions of nonscientists read it.
chivian, eric, and aaron Bernstein, eds. Sustaining Life: How Human Health Depends on Biodiver-sity. new york: Oxford University Press, 2008. this book, richly illustrated, shows nearly every possible way in which the protection of biodiver-sity is essential to human health and nutrition. the authors are at the Harvard Medical School.
cole, David n. Beyond Naturalness: Rethinking Park and Wilderness Stewardship in an Era of Rapid Change. Washington, D.c.: island Press, 2010. this book explains that national parks and wil-
derness areas cannot be kept intact by merely keeping people out of them.
collinge, Sharon K. Ecology of Fragmented Land-scapes. Baltimore, Md.: Johns Hopkins, 2009. this book explains how habitat fragmentation disturbs the ecosystems and biodiversity of pro-tected areas.
comin, Francisco a., ed. Ecological Restoration: A Global Challenge. new york: Oxford Univer-sity Press, 2010. this book explains how natural areas need to not only be preserved but restored throughout the world and the challenges associ-ated with doing this.
cox, c. Barry, and Peter D. Moore. Biogeogra-phy: An Ecological and Evolutionary Approach.Hoboken, n.J.: John Wiley, 2010. this textbook presents an up-to-date overview of the science of biogeography.
Darwin, charles. On the Origin of Species by Means of Natural Selection, 1st ed. london: John Mur-ray, 1859. available online. URl: http://darwin-online.org.uk/content/frameset?itemiD=F373&viewtype=side&pageseq=1. accessed august 2, 2010. this is perhaps the most important scientifi c book ever written. it lays the foundation for not just evolution but for ecology as well. Six editions were published during Darwin’s lifetime, each one addressing criticisms of previous editions, criti-cisms that are now considered to be inaccurate, and Darwin’s responses to them bogged down the initial insight of his fi rst edition.
Golley, Frank Benjamin. A History of the Ecosystem Concept in Ecology: More than the Sum of the Parts. new Haven, conn.: yale University Press, 1993. this book reviews the development of the ecosystem concept from before its formal proposal by tansley in 1935 to modern times when the emergent properties of ecosystems are recognized.
Hagen, Joel B. An Entangled Bank: The Origins of Ecosystem Ecology. new Brunswick, n.J.: Rut-gers University Press, 1992. this book is a history of important ecological concepts and the lives of the scientists who developed them. the name
xviii+598_Biodiversity_F.indd 556 11/17/11 1:13 PM

further resources 557
refers to the metaphor used in Darwin’s Origin of Species.
Hubbell, Stephen P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton, n.J.: Princeton University Press, 2001. this book explains what ecological communities would look like if the processes usually used to explain them were not operating.
Jackson, Wes. New Roots for Agriculture. lincoln, neb.: University of nebraska Press, 1980. Wes Jackson, considered one of the most important thinkers of modern times, proposes a whole new agriculture based on biodiversity and perennial crops. a great deal of contemporary research is emerging from his initial insights.
leopold, aldo. A Sand County Almanac: With Essays on Conservation from Round River. new york: Sierra club/Ballantine Books, 1974. Originally published in 1949, after leopold’s death, this set of essays presented some of the foundational con-cepts of modern environmental ethics.
louv, Richard. Last Child in the Woods: Saving Our Children from Nature Deficit Disorder. chapel Hill, n.c.: algonquin Books, 2008. in this book, louv explains the numerous personal and social problems that result from kids not having the chance to play outside in natural environments and the challenges that communities face in trying to solve this problem.
lovelock, James. The Vanishing Face of Gaia: A Final Warning. new york: Basic Books, 2009. lovelock is the original proponent of the Gaia hypothesis, and in this book, written late in his life, he explains that human activities may have pushed earth beyond the ability of its internal regulatory systems to maintain its stability. it is not an upbeat message of hope.
Mann, charles c. 1491: New Revelations of the Americas before Columbus. new york: Vintage, 2006. Mann explains how native americans had transformed the landscape before european con-tact and how much of what modern americans consider to be “natural” resulted from the decline of native american populations.
———. 1493: Uncovering the New World Columbus Created. new york: Knopf, 2011. Mann explains how the exchange of organisms, including disease parasites, between america and the rest of the world altered the entire earth ecosystem.
Marcus, Bernard. Tropical Forests. Sudbury, Mass.: Jones and Bartlett, 2008. this book is an overview of tropical forests, which contain more biodiver-sity than any other kind of ecological community, and each of which differs from the others.
Mccarthy, Dennis. Here Be Dragons: How the Study of Animal and Plant Distributions Revolution-
ized Our Views of Life and Earth. new york: Oxford University Press, 2009. this book traces the historical development of human realization that every part of the earth had different plants and animals.
McGavin, George c. Endangered: Wildlife on the Brink of Extinction. Richmond Hill, Ont.: Fire-fly Books, 2006. this book introduces readers to some important endangered species.
McKibben, Bill. The End of Nature. new york: Ran-dom House, 1990/2006. this book was the first popular science book about global warming. the author explains how, by enhancing the natural greenhouse effect, humans have changed the entire natural world, so that it is an extension of human activity rather than being “nature.”
Menino, Holly. Darwin’s Fox and My Coyote: Why Science Alone Can’t Win the Race to Save Wild Animals. charlottesville, Va.: University of Vir-ginia Press, 2008. this book is an appeal to envi-ronmental and conservation ethics, without which scientific efforts cannot succeed.
Merchant, carolyn. American Environmental His-tory: An Introduction. new york: columbia Uni-versity Press, 2007. this textbook is an overview of the development of conservation and environ-mentalism in the United States.
Plotkin, Mark J. Tales of a Shaman’s Apprentice: An Ethnobotanist Searches for New Medicines in the Amazon Rain Forest. new york: Viking, 1993. the author relates his stories of finding medicinal plants in the rain forest by listening to the people who live there.
Primack, Richard B. Essentials of Conservation Biol-ogy. Sunderland, Mass: Sinauer associates, 2006. this textbook relates the basic scientific concepts of conservation.
Quammen, David. Song of the Dodo: Island Bio-geography in an Age of Extinctions. new york: Scribner, 1996. the author explains island bioge-ography and introduces the scientists who have studied it.
Rice, Stanley a. Green Planet: How Plants Keep the Earth Alive. new Brunswick, n.J.: Rutgers Uni-versity Press, 2009. this book presents the argu-ment that plants keep the earth alive by removing carbon dioxide from the air, regulating tempera-ture and the water cycle, making all the food, cre-ating soil, and determining and restoring habitats and why plant diversity is essential.
———. Life of Earth: Portrait of a Beautiful, Mid-dle-Aged, Stressed-Out World. amherst, n.y.: Prometheus Books, 2010. this book presents the evolutionary saga from the viewpoint of humans, who are the most extreme example of many evo-lutionary processes from sex to altruism and who
xviii+598_Biodiversity_F.indd 557 11/17/11 1:13 PM

first entry558 further resources558
are also the biggest threat to the Gaia system of planetary interaction.
Rosenzweig, Michael. Win-Win Ecology: How the Earth’s Species Can Survive in the Midst of Human Enterprise. new york: Oxford University Press, 2003. the author presents the argument that endangered species can be preserved along with human prosperity.
terborgh, John. Requiem for Nature. covelo, calif.: island Press, 1999. this book explains that most of our efforts to protect the natural world will prove futile.
thomas, Peter, and Rob Mcalpine. Fire in the For-est. new york: Oxford University Press, 2010. this book explains the many roles of fire in ecology.
todd, Kim. Tinkering with Eden: A Natural History of Exotic Species in America. new york: norton, 2002. this book presents many examples of inva-sive species in north america and the ecological damage they have inflicted.
Volk, tyler. Gaia’s Body: Toward a Physiology of Earth. new york: copernicus, 1998. the author presents the Gaia hypothesis as an imperfect system of negative feedbacks rather than as a perfectly integrated organism, but a system that nevertheless has a physiology.
Whittaker, Robert J. Island Biogeography: Ecology, Evolution, and Conservation. new york: Oxford University Press, 1998. an overview of the science of island biogeography and the conservation chal-lenges associated with endemic species on ecologi-cal and geographical islands.
Wilson, edward O. The Diversity of Life. cam-bridge, Mass.: Harvard University Press, 1992. the founder of the modern concept of biodiversity presents an important overview of the exuberant diversity of life on earth—and the continuing sur-prises that come from the discovery of yet more diversity—and how the human future is threat-ened by the extinction of wild species.
———. The Creation: An Appeal to Save Life on Earth. new york: norton, 2006. the founder of the modern concept of biodiversity sends an open letter to an unspecified conservative christian preacher about why christians should care about the protection of endangered species and the eco-systems of earth.
Internet resourcesagricultural Research Service, United States Depart-
ment of agriculture. “Phytoremediation: Using Plants to clean Up Soils.” available online. URl: http://www.ars.usda.gov/is/ar/archive/jun00/soil0600.htm. accessed august 8, 2011. this gov-
ernment Web site gives examples of how plants can clean up toxic wastes from soils.
american chestnut Foundation. “american chestnut Foundation.” available online. URl: http://www.acf.org. accessed august 8, 2011. this foundation is attempting to restore the american chestnut to its former prominence in appalachian forests.
amphibiaweb. “amphibiaweb.” available online. URl: http://www.amphibiaweb.org. accessed august 8, 2011. this Web site is a resource for information about amphibians.
animal Welfare institute. “endangered Species Handbook.” available online. URl: http://www.endangeredspecieshandbook.org. accessed august 8, 2011. this Web site presents stories about endangered species and habitats that can be used by teachers.
Blue Ocean institute. “Guide to Ocean Friendly Seafood.” available online. URl: http://www.blueocean.org/seafood/seafood-guide. accessed august 8, 2011. this is an online guide to which seafood has been harvested in a manner that will not cause the collapse of ocean populations.
Botanical Society of america. “carnivorous Plants/insectivorous Plants.” available online. URl: http://www.botany.org/carnivorous_Plants. accessed august 8, 2011. this Web site introduces viewers to every species of carnivorous plant.
Bushmeat crisis task Force. “Bushmeat crisis task Force.” available online. URl: http://www.bush-meat.org. accessed august 8, 2011. this Web site provides information about and coordinates actions to reduce the destruction of african pri-mates as a food source.
california Oak Mortality task Force. “california Oak Mortality task Force.” available online. URl: http://www.suddenoakdeath.org. accessed august 8, 2011. this Web site monitors the spread of the Phytophthora infestations that are killing california oak forests.
charles Darwin Foundation. “charles Darwin Foundation.” available online. URl: http://www.darwinfoundation.org/english/pages/index.php. accessed august 8, 2011. this is the Web site for scientific research and conservation on the Galá-pagos islands.
chicago Wildnerness. “leave no child inside.” available online. URl: http://www.kidsoutside.info. accessed august 8, 2011. this Web site coor-dinates activities to get kids to experience natural habitats.
children and nature network. “children and nature network: Building a Movement to Reconnect chil-dren and nature.” available online. URl: http://www.childrenandnature.org. accessed august 8, 2011. this Web site coordinates activities around
xviii+598_Biodiversity_F.indd 558 11/17/11 1:13 PM

further resources 559
the United States to get kids to experience natural habitats.
convention on international trade in endangered Species (citeS). “convention on international trade in endangered Species of Wild Fauna and Flora.” available online. URl: http://www.cites.org. accessed august 8, 2011. this is the Web site for the international agreement that protects endangered species by prohibiting the transport and sale of endangered organisms, living or killed.
coral Reef alliance. “coral Reef alliance.” available online. URl: http://www.coral.org. accessed august 8, 2011. this Web site documents the destruction of coral reefs and what can be done to stop it.
cornell lab of Ornithology. “all about Birds.” avail-able online. URl: http://www.allaboutbirds.org. accessed august 8, 2011. this Web site is the place to go to find out about all kinds of birds and their classification.
earth’s endangered creatures. “World endan-gered Species list.” available online. URl: http://www.earthsendangered.com/full_list_html.asp. accessed august 8, 2011. all in one place, here is a list (without links) to all endangered species in the world, by latin and common names.
encyclopedia of life. “encyclopedia of life.” available online. URl: http://www.eol.org. accessed august 8, 2011. the encyclopedia of life is an attempt to establish a Web page for every species on earth.
environmental Protection agency. “integrated Pest Management (iPM) Principles.” available online. URl: http://www.epa.gov/pesticides/factsheets/ipm.htm. accessed august 8, 2011. the federal government presents guidelines for how iPM can be effectively practiced.
———. “Persistent Organic Pollutants: a Global issue, a Global Response.” available online. URl: http://www.epa.gov/oia/toxics/pop.html. accessed august 8, 2011. this is the definitive list of long-lasting pollutants in the environment and what is being done to control them.
Froese, R., and D. Pauly, eds. “FishBase.” available online. URl: http://www.fishbase.org. accessed august 8, 2011. this is the place to go to find out about all kinds of fishes.
Galápagos conservancy. “Galápagos conservancy.” available online. URl: http://www.galapagos.org/2008/index.php?id=68. accessed august 8, 2011. this is the Web site for an organization that is working to restore the damaged habitats of the Galápagos islands.
Global Footprint network. “national footprints.” available online. URl: http://www.footprintnet work.org/gfn_sub.php?content=national_foot prints. accessed august 1, 2010. this Web site compares the ecological footprints of all nations.
Hooked on nature. “tools, Resources, and Workshops to Help People Deepen their Relationship with the natural World.” available online. URl: http://www.hookedonnature.org. accessed august 8, 2011. ideas for education in schools and for the general public, to promote awareness of biodiversity.
integrated taxonomic information System. “inte-grated taxonomic information System.” available online. URl: http://www.itis.gov. accessed august 8, 2011. the authoritative Web site for the clas-sification of all american species.
international Society for Phylogenetic nomenclature. “international Society for Phylogenetic nomencla-ture.” available online. URl: http://phylonames.org. accessed august 8, 2011. this Web site explains and defends the cladistic system of classification.
international Society of Organic agriculture Research. “international Society of Organic agri-culture Research.” available online. URl: http://www.isofar.org. accessed august 8, 2011. this organization promotes improvements in organic agriculture.
international Union for the conservation of nature. “iUcn.” available online. URl: http://www.iucn.org. accessed august 8, 2011. this Web site pro-vides access to the “iUcn Red list,” which is infor-mation about endangered species all over the world, but also provides links to information about climate change and how to develop a “green” economy.
land institute. “the land institute.” available online. URl: http://www.landinstitute.org. accessed august 8, 2011. the land institute is the world center for developing natural Systems agriculture.
leopold center for Sustainable agriculture, iowa State University. “Welcome to the leopold cen-ter.” available online. URl: http://www.leopold.iastate.edu. accessed august 8, 2011. the leop-old center promotes the benefits of biodiversity within agriculture.
local Harvest. “local Harvest.” available online. URl: http://www.localharvest.org. accessed august 8, 2011. this Web site provides informa-tion about where consumers can buy food directly from local farmers.
Martin, R. aidan. “ReefQuest center for Shark Research.” available online. URl: http://www.elasmo-research.org. accessed august 8, 2011. this is the place to go to find out about sharks.
Millennium ecosystem assessment. “Millennium ecosystem assessment.” available online. URl: http://www.maweb.org/en/index.aspx. accessed august 8, 2011. this Web site presents reports of ongoing studies of how ecosystem changes are affecting human well-being.
Missouri Botanical Garden. “MBGnet.” available online. URl: http://www.mbgnet.net. accessed
xviii+598_Biodiversity_F.indd 559 11/17/11 1:13 PM

first entry560 further resources560
august 8, 2011. this Web site introduces the diversity of plant species for children.
Monterey Bay aquarium. “Seafood Watch.” avail-able online. URl: http://www.montereybayaquar ium.org/cr/seafoodwatch.aspx. accessed august 8, 2011. this is an online guide to which seafood has been harvested in a manner that will not cause the collapse of ocean populations.
national agricultural library. “national inva-sive Species information center.” available online. URl: http://www.invasivespeciesinfo.gov. accessed august 8, 2011. this government Web site provides comprehensive invasive species infor-mation for the United States.
national Oceanic and atmospheric administra-tion. “nOaa coral Reef conservation Program.” available online. URl: http://coralreef.noaa.gov. accessed august 8, 2011. this Web site describes the U.S. government’s plans for saving and restor-ing coral reefs.
natural Resource conservation Service, United States Department of agriculture. “Plants Data-base.” available online. URl: http://www.plants.usda.gov. accessed august 8, 2011. this Web site provides information about and photographs of nearly all american plant species.
Ornithological council, national Museum of nat-ural History, Smithsonian institution. “Birdnet: the Ornithological information Source.” avail-able online. URl: http://www.nmnh.si.edu/BiRDnet. accessed august 8, 2011. this Web site is the place to go for information about bird species.
Pelagic Shark Research Foundation. “Pelagic Shark Research Foundation.” available online. URl: http://www.pelagic.org. accessed July 9, 2010. this Web site is the place to go for information about sharks.
Population Reference Bureau. “2011 World Popula-tion Data Sheet.” available online. URl: http://prb.org/pdf11/2011population-data-sheet_eng.pdf. accessed august 8, 2011. this Web site pro-vides population and economic information for every country in the world.
Ratnasingham, Sujeevan, and Paul D. n. Hebert. “Barcoding of life Data System.” available online. URl: http://www.barcodinglife.org. accessed august 8, 2011. this Web site explains the proto-cols for using Dna to identify every species.
Realclimate. “Realclimate: climate Science from climate Scientists.” available online. URl: http://www.realclimate.org. accessed august 8, 2011. this is a scientific source of information about global warming.
Rowe, noel. “Primate conservation, inc.” available online. URl: http://www.primate.org. accessed
august 8, 2011. this organization promotes the protection of endangered primate species.
Solomon, Susan, and the intergovernmental Panel on climate change. “climate change 2007: the Physical Science Basis.” available online. URl: http://www.ipcc.ch/publications_and_data/ar4/wg1/en/contents.html. accessed august 8, 2011. this organization is the recognized world author-ity on global warming.
tree of life Web Project. “explore the tree of life.” available online. URl: http://www.tolweb.org. accessed august 8, 2011. this Web site provides a constantly expanding cladistic classification of all species.
United nations. “convention on Biological Diversity.” available online. URl: http://www.cbd.int. accessed July 11, 2010. this Web site explains the role of the United nations in protecting biodiversity around the world.
———. “Un-ReDD Programme: the United nations collaborative Programme on Reducing emissions from Deforestation and Forest Degra-dation in Developing countries.” available online. URl: http://www.un-redd.org. accessed august 8, 2011. this Web site explains the United nations effort to reduce deforestation by paying countries for the ecosystem services provided by leaving their forests intact.
United nations environment Programme. “Over-fishing: a threat to marine biodiversity.” avail-able online. URl: http://www.grid.unep.ch/product/publication/download/ew_overfishing.en.pdf. accessed august 8, 2011. UneP explains the crisis of overfishing in the oceans.
United nations Food and agriculture Organization. “Biodiversity for a World Without Hunger.” avail-able online. URl: http://www.fao.org/biodiver sity/en/. accessed august 8, 2011. this Web site explains the efforts of the Food and agriculture Organization to protect the genetic diversity upon which agriculture depends.
United States census Bureau. “international Data Base.” available online. URl: http://www.census.gov/population/international/data/idb/informa-tionGateway.php accessed august 8, 2011. this Web site provides access to information about the population characteristics of every country in the world, from 1950 onward, including projections into the future.
United States Department of agriculture. “animal and Plant Health inspection Service: Safeguard-ing american agriculture.” available online. URl: http://www.aphis.usda.gov. accessed august 8, 2011. this Web site explains the work of aPHiS, which protects the United States from the impor-tation of invasive species.
xviii+598_Biodiversity_F.indd 560 11/17/11 1:13 PM

further resources 561
University of illinois at Urbana-champaign. “land-scapes and Human Health laboratory.” avail-able online. URl: http://lhhl.illinois.edu. accessed august 8, 2011. this Web site presents informa-tion about the importance of green landscapes in human mental health.
University of Michigan at Dearborn. “native ameri-can ethnobotany.” available online. URl: http://herb.umd.umich.edu. accessed august 8, 2011. this Web site provides access to a database about the native american uses of medicinal plants.
Weed Science Society of america. “international Sur-vey of Herbicide Resistant Weeds.” available online. URl: http://www.weedscience.org/in.asp. accessed august 8, 2011. this Web site is the definitive list of weeds that are resistant to herbicides.
PerIodIcalsAmerican ForestsPublished by american ForestsBox 2000Washington, Dc 20013telephone (202) 737-1944www.americanforests.org
American Journal of BotanyPublished by the Botanical Society of americaP.O. Box 299St. louis, MO 63166-0299telephone (314) 577-9566www.botany.org
Annual Review of Ecology, Evolution, and Systematics
Published by annual Reviews, inc.P.O. Box 10139Palo alto, ca 94303-0139www.annualreviews.org
AudubonPublished by the national audubon Society225 Varick Street, 7th floornew york, ny 10014telephone (212) 979-3172www.audubon.org
The AukPublished by american Ornithological Union5400 Bosque Boulevard, Suite 680Waco, tX 76710telephone (254) 399-9636www.aou.org
Biodiversity and ConservationPublished by Springer
223 Spring Streetnew york, ny 10013www.springer.com
BioSciencePublished by american institute of Biological
Sciences1900 campus commons Drive, Suite 200Reston, Va 20191telephone (703) 674-2500www.aibs.org
BiotropicaPublished for association for tropical Biology and
conservation by John Wiley111 River StreetHoboken, nJ 07030-5774telephone (201) 748-6000www.wiley.com
Conservation BiologyPublished by Society for conservation Biology1017 O Street nWWashington, Dc 20001-4229telephone (202) 234-4133www.conbio.org
Ecological ApplicationsPublished by the ecological Society of america1990 M Street nW, Suite 700Washington, Dc 20036telephone (202) 833-8773www.esa.org
EcologyPublished by the ecological Society of america1990 M Street nW, Suite 700Washington, Dc 20036telephone (202) 833-8773www.esa.org
EcospherePublished by the ecological Society of america1990 M Street nW, Suite 700Washington, Dc 20036telephone (202) 833-8773www.esa.org
Endangered Species ResearchPublished by inter-Research Science centernordbünte 23 (+ 3, 5, 28, 30)21385 Oldendorf/luhe, Germanywww.int-res.com/journals/esr/esr-home
Environmental EntomologyPublished by the entomological Society of america
xviii+598_Biodiversity_F.indd 561 11/17/11 1:13 PM

first entry562 further resources562
10001 Derekwood lane, Suite 100lanham, MD 20706-4876telephone (301) 731-4535www.entsoc.org
Frontiers in Ecology and the EnvironmentPublished by the ecological Society of america1990 M Street nW, Suite 700Washington, Dc 20036telephone (202) 833-8773www.esa.org
Functional EcologyPublished by the British ecological Societycharles Darwin House12 Roger Streetlondon, Wc1n 2JUtelephone 0207 685 2500www.britishecologicalsociety.org
Journal of Applied EcologyPublished by the British ecological Societycharles Darwin House12 Roger Streetlondon, Wc1n 2JUtelephone 0207 685 2500www.britishecologicalsociety.org
Journal of BiogeographyPublished by the international Biogeography Society133 Basswood courtWestlake Village, ca 91361-2003www.biogeography.org
Journal of EcologyPublished by the British ecological Societycharles Darwin House12 Roger Streetlondon, Wc1n 2JUtelephone 0207 685 2500www.britishecologicalsociety.org
Journal of Integrated Pest ManagementPublished by the entomological Society of america10001 Derekwood lane, Suite 100lanham, MD 20706-4876telephone (301) 731-4535www.entsoc.org
Journal of MammalogyPublished by the american Society of MammalogistsP.O. Box 7060lawrence, KS 66044telephone (800) 627-0326www.mammalsociety.org
Landscape EcologyPublished by Springer223 Spring Streetnew york, ny 10013www.springer.com
OecologiaPublished by Springer223 Spring Streetnew york, ny 10013www.springer.com
Proceedings of the National Academy of Sciences USA
Published by the national academy of Sciences500 Fifth Street nWWashington, Dc 20001telephone (202) 334-2000www.nas.edu
Restoration EcologyPublished for Society for ecological Restoration
international by John Wiley111 River StreetHoboken, nJ 07030-5774telephone (201) 748-6000www.wiley.com
SciencePublished by the american association for the
advancement of Science1200 new york avenue nWWashington, Dc 20005telephone (202) 326-6417www.sciencemag.org
Scientific American415 Madison avenuenew york, ny 10017telephone (212) 754-0550www.sciam.com
Trends in Ecology and EvolutionPublished by cell Press600 technology Squarecambridge, Ma 02139telephone (617) 661-7057
socIetIes and organIzatIonsamerican association for the advancement of Sci-
ence (www.aaas.org), 1200 new york avenue nW, Washington, Dc 20005. telephone: (202) 326-6400
xviii+598_Biodiversity_F.indd 562 11/17/11 1:13 PM

further resources 563
american Forests (www.americanforests.org), Box 2000, Washington Dc 20013. telephone: (202) 737-1944
american institute of Biological Sciences (www.aibs.org), 1900 campus commons Drive, Suite 200, Reston, Va 20191. telephone: (703) 674-2500
american Ornithological Union (www.aou.org), 5400 Bosque Boulevard, Suite 680, Waco, tX 76710. telephone: (254) 399-9636
american Society of Mammalogists (www.mammal society.org), P.O. Box 7060, lawrence, KS 66044. telephone: (800) 627-0326
association for tropical Biology and conservation (www.tropicalbio.org)
Botanical Society of america (www.botany.org), Box 299, St. louis, MO 63166-0299. telephone: (314) 577-9566
British ecological Society (www.britishecologicalso ciety.org), charles Darwin House, 12 Roger Street, london, Wc1n 2JU United Kingdom. telephone: 0207 685 2500
ecological Society of america (www.esa.org), 1707 H Street nW, Suite 400, Washington, Dc 20006-3915. telephone: (202) 833-8775
entomological Society of america (www.entsoc.org), 10001 Derekwood lane, Suite 100, lanham, MD 20706-4876. telephone: (301) 731-4535
international Biogeography Society (www.biogeogra phy.org), 133 Basswood court, Westlake Village, ca 91361-2003
international Society for ecological economics (www.ecoeco.org)
linnean Society of london (www.linnean.org), Burl-ington House, Picadilly, london, W1J 0BF United Kingdom. telephone: +44 020 7434 4479
national academy of Sciences (www.nas.edu), 500 Fifth Street nW, Washington, Dc 20001. tele-phone: (202) 334-2000
national audubon Society (www.audubon.org), 225 Varick Street, 7th floor, new york, ny 10014. telephone: (212) 979-3172
national Science Foundation (www.nsf.gov), 4201 Wilson Boulevard, arlington, Va 22230. tele-phone: (703) 292-5111
nature conservancy (www.nature.org), 4245 north Fairfax Drive, Suite 100, arlington, Va 22203. telephone: (703) 841-5300
Royal entomological Society (www.royensoc.co.uk), the Mansion House, chiswell Green lane, St. albans, al2 3nS United Kingdom. telephone: +44 (0) 1727 899387
Society for conservation Biology (www.conbio.org), 1017 O Street nW, Washington, Dc 20001-4229. telephone: (202) 234-4133
Society for ecological Restoration international (www.ser.org), 1017 O Street nW, Washington Dc 20001. telephone: (202) 299-9518
Society for the Study of amphibians and Reptiles (www.ssarherps.org)
World Wildlife Fund (www.worldwildlife.org), 1250 twenty-Fourth Street nW, P.O. Box 97180, Washington, Dc 20090-7180. telephone: (202) 983-4800
xviii+598_Biodiversity_F.indd 563 11/17/11 1:13 PM

564
note: this system does not include all evolutionary lineages.
Domain archaea
Domain eubacteria thermophilic bacteria chemolithoautotrophic bacteria Proteobacteria (purple bacteria) Green sulfur bacteria cyanobacteria Obligate anaerobic bacteria aerobic bacteria Pathogenic bacteria
Domain eucarya (eukaryotes)Kingdom Protista = Protoctista includes all eukaryotes.
chromista = stramenopiles Oomycota (water molds) Bacillariophyta (diatoms) Xanthophytes and phaeophytes (golden and brown algae) chrysophytaalveolates Dinofl agellata ciliopoda (ciliates) apicomplexachlorophyta includes all green plants (see below)Rhodophyta (red algae)Opisthokonts Fungi (see below) choanofl agellates and animals (see below)amoebasForaminifersRadiolariansDiplomonada (archezoans)euglenasMyxomycota (plasmodial slime molds)acrasiomycota (cellular slime molds)
Kingdom Fungi (see above)
Basidiomycota (mushrooms)ascomycota (sac fungi)
Appendix IVClassification System Used in Encyclopedia of Biodiversity*
xviii+598_Biodiversity_F.indd 564 11/17/11 1:13 PM

565Classification System used in Encyclopedia of Biodiversity
Glomeromycota (vesicular arbuscular mycorrhizae)Zygomycota (bread molds)chytrids
Kingdom Plantae (see above)
Zygnematales (conjugating green algae)charalesembryophytes Marchantiomorpha (liverworts) anthocerotophyta (hornworts) Bryophyta (mosses and liverworts) Rhyniophytes (extinct) lycophytes (extinct) Filicopsida (ferns) Spermatopsida (seed plants) cycads Ginkgos Gnetales conifers Family Pinaceae (pines, spruces) Family taxaceae (yews) Family taxodiaceae (sequoias, junipers) angiosperms Primitive dicots Family nymphaceae (water lilies) Family Magnoliaceae (magnolias) Family Ranunculaceae (buttercups) Monocots liliales Family liliaceae (lilies) commelinanae Family Bromeliaceae (bromeliads) Family cyperaceae (sedges) Family typhaceae (cattails) Family Poaceae (grasses) eudicots Rosids: Fagales Family Fagaceae (oaks and beeches) Family Betulaceae (alders and birches) Family Juglandaceae (hickories and
walnuts) Rosids: Rosales Family Rosaceae (roses) Family Ulmaceae (elms) Moraceae group (mulberries, nettles,
hackberries, and cannabis) Rosids: Fabales Family Fabaceae (leguminous plants) Malphigiales Family Salicaceae (willows) Family euphorbiaceae (spurges) Myrtales Family Myrtaceae (myrtles, eucalyptus) Brassicales Family Brassicaceae (mustards)
xviii+598_Biodiversity_F.indd 565 11/17/11 1:13 PM

first entry566566 Classification System used in Encyclopedia of Biodiversity
Malvales Family Malvaceae (mallows, hibiscus) Sapindales Family Sapindaceae (maples, soapberries) Rutaceae (citrus) Family Vitaceae (grapes) caryophyllales Polygonales Family Polygonaceae (buckwheat) Family cactaceae (cactus) Family caryophyllaceae (carnations) cornales Family cornaceae (dogwoods) ericales Family ericaceae (heaths) Solanales Family Solanaceae (tomatoes, etc.) lamiales Family lamiaceae (mints) Gentianales Family apocynaceae (milkweeds) asterales Family asteraceae (composites) apiales Family apiaceae (umbellifers)
Kingdom animalia (see above)
ctenophora (comb jellies)Porifera (sponges)PlacozoanscnidariaBilateria Protostomes Rotifera (rotifers) Platyhelminthes (flatworms, tapeworms, flukes) lophotrochozoans annelida (segmented worms) Mollusca (mollusks) ecdysozoans nematoda (roundworms) arthropoda (arthropods) chelicerates (e.g., arachnids) Spiders ticks Scorpions Myriapods (e.g., centipedes and millipedes) crustaceans (crabs, lobsters, shrimp,
barnacles) Hexapods (e.g., insects) Odonata (dragonflies) Hemipteroids (bugs and lice) Orthopterans (grasshoppers, roaches, crickets) coleoptera (beetles) Diptera (flies) Siphonaptera (fleas)
xviii+598_Biodiversity_F.indd 566 11/17/11 1:13 PM

567Classification System used in Encyclopedia of Biodiversity
lepidoptera (butterflies and moths) Hymenoptera (ants, bees, wasps) Deuterostomes echinodermata (starfish) chordates Hagfishes Vertebrates lampreys Gnathostomata (jawed vertebrates) chondrichthyes (sharks, rays) Ray-finned fishes (e.g., teleosts) Sturgeons Gars eels carps and catfishes Salmons and trouts Pikes cods Seahorses Sticklebacks Flying fishes Puffers Flatfishes anglerfishes Sarcopterygians lungfishes amphibians Frogs Salamanders caecilians amniotes turtles Diapsids lepidosaurs: lizards, snakes archosaurs crocodiles Dinosaurs long-necked theropods Birds:
Birds:Paleognaths (ostriches)neognaths Galliformes (fowl) anseriformes (ducks) Passeriformes (perching birds) Piciformes (woodpeckers) columbiformes (doves) apodiformes (hummingbirds) caprimulgiformes (nighthawks, swifts) Strigiformes (owls) ciconiiformes (storks, herons, pelicans) Falconiformes (birds of prey) charadriiformes (shorebirds) Sphenisciformes (penguins)
xviii+598_Biodiversity_F.indd 567 11/17/11 1:13 PM

first entry568568 Classification System used in Encyclopedia of Biodiversity
Synapsids Mammals Monotremes Marsupials eutherians (placentals):
Placental mammals:edentates (anteaters, sloths, armadillos)lagomorphs (rabbits)Rodents Muroids (mice, rats, etc.) Sciurids (squirrels)insectivorescarnivores Feliformia (cats) caniformia canidae (dogs) Ursidae (bears) Procyonidae (raccoons) Mustelidae (weasels, skunks) Pinnepedia (seals)cetartiodactyls cetaceans (whales, dolphins) artiodactyls Deer Hippos camels Ruminants Bovids cervids GiraffesPerissodactyls (horses, rhinos)Sirenians (manatees)Proboscideans (elephants)Primates and relatives lemurs new World monkeys (platyrrhines) Old World monkeys and apes (catarrhines) cercopithecids (Old World monkeys) Hylobatids (gibbons) Hominids Orangutans Gorillas chimps Hominins
*Based on: tree of life Web Project. “explore the tree of life.” available online. URl: http://www.tolweb.org/tree. accessed February 1, 2011.
xviii+598_Biodiversity_F.indd 568 11/17/11 1:13 PM

569
note: Boldface page numbers denote main articles on a topic. Italic page numbers indicate illustrations. Page numbers followed by c denote chronology entries; page numbers followed by t denote tables, charts, or graphs; page numbers followed by m denote maps.
aaardvarks 367Abies 92, 137Abies bracteata 222Abies concolor 502Abies fraseri 94Abies magnifi ca 92abrahamson, Warren 389Abutilon theophrasti 3, 4, 330, 356, 501acacia
coevolution 122deserts, warm 175, 177dicots 186Janzen, Daniel H. 338keystone species 343–344mutualism 380
Acacia 9, 380, 537Acacia fumosa 56Acacia tortilis 278Acanthaster planci 149, 381acanthocephala 28, 31Acanthodis curvidens 98acanthopterygii 248Acanthostega 17Acanthostega gunnari 16accipitriformes 89acclimatization 3, 388Acer 73Acer negundo 505Acerodon jubatus 366Acer saccharinum 187Acer saccharum 187, 504, 513, 513Achatina fulica 331achelates 157acid Deposition act 2acidifi cation 150, 297Acidithiobacillus ferooxidans 43acid rain and fog 1–2, 18, 94, 152, 373,
409acid Rain Peer Review Panel 2acoelomate 20acorns 185acorn woodpeckers 90acorn worms 29Acrocephalus arundinaceus 405Acromyrmex 8Actias luna 325actinobacteria 41, 42, 380adapid primates 443adaptation 2–6, 5
amphibians 17animals 20–21biodiversity, patterns of 47
biodiversity, resources for agriculture 58
biogeography 76biomimicry 82–83competition, ecological 132convergence 144, 146–148cultural diversity 158–160deserts, warm 176–182ecology 206fi shes 249–250herbivory 306insects 323life history, diversity of 353–354mammals 362, 365, 366natural selection 388–389parasitism 404pollination 418–419predation 437reptiles 455, 456rivers, streams, ponds, and lakes 467seed dispersal 470–471sexual selection 482
adaptive radiation 6–8, 73, 169, 374adder’s-tongue ferns 476Adelges tsugae 2adelgid bugs 2adhesion 83Adiantum 475adipose fi ns 248adnate fusion 254advanced cell technologies 220Aedes aegypti 404Aedes albopictus 330Aegilops speltoides 310Aegilops squarrosa 310Aequorea victoria 69aerobic bacteria 32, 509aerobic respiration 42Aetiocetus cotylaveus 399afghan snakes 456afl atoxins 262africa
adaptive radiation 6agriculture 8–9biodiversity, resources for agriculture
62biogeography 73, 74biomagnifi cation 81birds 87camoufl age and mimicry 98carson, Rachel louise 109coevolution 122cretaceous period, biodiversity in 154deep oceans, biodiversity in 172deserts, warm 175, 177, 180, 181dicots 187environmental ethics 223extinction 235grasslands and savannas 277t, 278–279greenhouse effect 286Humboldt, alexander von 307
ice ages 318Maathai, Wangari 358–359Pleistocene extinction 415primates 443Quaternary period, biodiversity in 452reptiles 457tropical rain forests and deforestation
533tropical seasonal forests 537
african cherry 66african honeybees 332african land snail 331african meerkats 159agassiz, lake 316, 317agassiz, louis 195, 315Agathis australis 136agaves 375, 376Agelaius phoeniceus 370age of Dinosaurs 153–156, 339–341Agkistrodon contortrix 457Agkistrodon piscivorus 457Aglaophyton major 263agnathan fi shes 246agriculture x, 8–15, 10t, 11, 14
biodiversity, resources for agriculture 62–63
biomagnifi cation 81biomimicry 83coevolution 122convergence 148greenhouse effect 290herbivory 304insects 326invasive species 333mutualism 380, 381natural systems agriculture 390–392pollination 420–421population biology 429–430resistance, evolution of 460restoration ecology 462, 463temperate deciduous forests 516tropical rain forests and deforestation
531–533agriculture, U.S. Department of (USDa)
59, 238, 239, 242, 548cAilanthus altissima 350Ailuropoda melanoleuca 363air pollution 1–2, 289alaska 94, 537alaska’s north Slope 540albedo 205, 211, 286, 311, 315, 316Albumares 217Alcanivorax borkumensis 84alders
boreal and subalpine forests 92conservation biology 139convergence 145dicots 186herbivory 304hybridization 308speciation 490, 492
INDEX
xviii+598_Biodiversity_F.indd 569 11/17/11 1:13 PM

first entry570 tundra 570 first entry570 Index570
alewives 345algae
animals 23competition, ecological 134coral reefs 149eukaryotes 232fungi 264mutualism 381protists 447, 448seedless plants 472
alkaloids 67, 303allee, Warder c. 132, 429allee effect 132, 429alleles 258, 430, 432–434allelochemicals 133allelopathy 132, 267–268, 487alligator(s) 370, 456, 462Alligator mississippiensis 370, 456alligator snapping turtle 99Allium cepa 144allometry 5allopatric speciation 490allozymes 432Alnus 92, 186, 490, 492Alnus maritima 186, 194–195, 308Alnus maritima ssp. georgiensis 221Alnus maritima ssp. maritima 221Alnus maritima ssp. oklahomensis 139, 221Alnus serrulata 194, 308Alosa pseudoharengus 345alpha diversity 125–126Alphadon 156Alpheus lottini 381alpine tundra 76, 194, 296, 537, 538, 538alps 76, 154Alteromonas 380altruism 389alvarez, luis 152alvarez, Walter 152alveolates 447Amanita muscaria 262amaranth 62amazon 548c
amphibians 17Bates, Henry Walter 45biodiversity, patterns of 49fishes 248Humboldt, alexander von 307hydrological cycle 313restoration ecology 461tropical rain forests and deforestation
527, 531, 532, 534tropical seasonal forests 537Wallace, alfred Russel 542
amazonian tribes 461amber 256Ambolucetus natans 399Amborella 254Amborella trichopoda 75, 80, 221, 254, 356Ambystoma mexicanum 18amebozoan protists 448american beeches 513, 515american chestnuts 186, 234, 344–345american gray squirrels 333ammonia 18, 42, 394ammonites 182, 399ammonoids 151amniotic egg 454amoebae 447–448, 509–510Amoeba proteus 509Ampheristocystus 488amphibians 15–19, 17t
biodiversity, resources for medicine 66, 67
carboniferous period, biodiversity in 105–106
Jurassic period, biodiversity in 341neogene period, biodiversity in 393Permian period, biodiversity in 412predation 436Quaternary period, biodiversity in 452
amphipods 158amphisbaeneans 458amsterdam Declaration (2001) 266Amyciaea 497Amytornis goyderi 181anacardiaceae 187anadromous fishes 248, 251, 467anaerobic archaea 32anaerobic bacteria 41, 379anaerobic fungi 261anal fin 246anapsids 454anatolian peninsula 313ancestral traits 110anchovies 248andersson, Malte 480Andrias 17Andropogon 278angier, natalie 354angiosperms. See also flowering plantsangler fishes 99, 171, 248, 437animal adaptations 180–182animal and Plant Health inspection Service
(aPHiS) 334animals 19–31, 20, 21t, 22–24, 26, 30. See
also specific animalsAnisocelis flavolineata 98Ankylosaurus 155annelida 26–27, 172Anniella 458annuals 499anole lizards 389Anomalocaris 96, 97, 257–258Anopheles 324anoplura 38, 322anoxic deep oceans 373anoxic deposits 407anseriformes 89ant(s) 380, 404
agriculture 8–9arthropods 38coevolution 122convergence 145, 148cretaceous period, biodiversity in 155fungi 264insects 323, 325–326Janzen, Daniel H. 338mutualism 380seed dispersal 471spiders 497–498Wilson, edward O. 545, 546
antarcticaarthropods 36–37birds 89deep oceans, biodiversity in 172deserts, polar 174greenhouse effect and biodiversity 296ice ages 315, 316monocots 377
antarctic ice 282, 283, 285antarctic Ocean 171anteaters 361, 367antennae 34, 37, 320antheridia 447, 473, 474Anthill (Wilson) 545anthrax 114anthropic principle 544anthropoids 441–442antibiotic resistance 43, 66–68, 390, 423antibiotics 64, 66–68, 173anticoagulants 66anticonvulsants 66antienvironmentalists 270antifreeze proteins 147antifungals 59anti-inflammatory compound 67Antilocapra americana 173antimicrobials 67, 459–460antlers 480ant-mimic spiders 497–498Ants, The (Wilson and Hölldobler) 545
anxiety 230anza-Borrego Desert 179apalachicola River area 222Apatosaurus 341apes 366, 442, 444apex chert 33, 42, 256aphids 35, 38, 52, 323, 380aPHiS. See animal and Plant Health
inspection Serviceapiaceae 188apicomplexans 447apicoplasts 509Apis mellifera 215, 326, 421apocalyptic christianity 225apocynaceae 188apodiformes 89apomixis 309, 491Apotosaurus 191appalachian Mountains 2, 72, 234, 312,
516appendages 35, 37, 320appetite suppressant 66apples 117–118, 186apterygota 321aquaculture 172–173aquatic arthropods 156aquatic environment 17, 18aquatic habitats 49, 90–91aquatic invertebrates 257aquatic plants 472Aquifex 32arabian Peninsula 175Arabidopsis thaliana 186araceae 376arachnids 35, 496–499arachnophobia 498aral Sea 463–464, 464Araucaria araucana 136araucariaceae 136, 221Araucaria heterophylla 136Araucarioxylon trees 341arbor Day 548carboreal adaptations 366arbuscular mycorrhizae 263arcata, california 61archaea 32–34, 549c
bacteria 41, 42, 44biodiversity, concept of 47biological classification 79decomposers 170deep oceans, biodiversity in 171eukaryotes 230–232
archaean era 33, 42archaebacteria 79, 509Archaeopteris 183Archaeopteryx 84, 85, 193, 193, 257, 341Archaeosperma 477Archaeothyris 106archegonia 447, 473–476Architeuthis dux 171archosaurs 87, 454, 524arctic ice cap 284, 286arctic national Wildlife Refuge 540arctic Ocean 171, 174, 316, 537arctic tern 89arctic tundra 76, 194, 195, 537Arctostaphylos bush 487Ardea herodius 370Ardipithecus ramidus 258arecaceae 376arerectores pilorum muscles 360Arfia 400argentina 13, 53, 173Argyroneta 497aristotle 548carizona 76, 173, 175, 341, 344arizona bark scorpion 36arizona pines 511Arkarua 217Armadillidium 158armadillos 297, 367, 450
xviii+598_Biodiversity_F.indd 570 11/17/11 1:13 PM

tundra 571Index 571
arm bones 15, 17armies 10, 11Armillaria 262Armillaria ostoyae 262–263armored jawless fishes 183army ants 145army corps of engineers 242Army Survival Manual, U.S. 60arrow worms 28arroyos 177, 179arsenic 84Artemia 158Artemia salina 305–306Artemisia annua 67artemisinin 67Arthrobotrys oligospora 438Arthropleura 105arthropods 34–39, 34t, 36, 79
animals 27biodiversity, concept of 46cambrian period, biodiversity in 96carboniferous period, biodiversity
in 105crustaceans 156decomposers 170deep oceans, biodiversity in 172insects 318–327island biogeography 336Ordovician period 396Silurian period, biodiversity in 488spiders 496–499
articular bone 361artificial evolution 58artificial selection 11–12, 165, 384artiodactyla 365arums 376Asclepias 117Asclepias viridis 133–134ascomycota 261–262ascus (asci) 261, 262asexual reproduction 35, 261–263asexual sporangia 260ashe juniper 200, 331ashfall Beds State Historic Park 258asia
biogeography 73dicots 186grasslands and savannas 276–278temperate coniferous forests 511temperate deciduous forests 513, 516temperate rain forests 519tundra 537
asian carp 330–331, 333–334asian elephants 537asian tiger mosquitoes 330asiatic salamanders 17Asimina triloba 119Asipenser baerri baicalensis 222aspens 73, 92, 186, 502Aspergillus 264Aspergillus flavus 262Aspergillus oryzae 262aspirin 66, 67Asplenium septentrionale 476asteraceae 74, 75, 180, 188, 188, 272asterids 188asteroid collisions 373, 374, 409, 416Astragalus 84, 186, 303Astrapsis 397aswan Dam 30asymmetry 22–23atacama Desert 175, 176athlete’s foot 262atlantic cod 172atlantic horseshoe crabs 37, 46, 67atlantic Ocean 141, 154, 172atlantic rain forest 527atmosphere 103, 269–270, 393, 394, 407atmospheric carbon dioxide. See carbon
dioxide, atmosphericAtrax robustus 498
atrazine 18, 423Atriplex hymenelytra 177Atta 8, 380auracarians 136australia
biogeography 75birds 87boreal and subalpine forests 92continental shelves, biodiversity on 141deserts, warm 175, 177, 180, 181disjunct species 196ediacaran organisms 217grasslands and savannas 276, 278greenhouse effect 286hydrological cycle 313invasive species 330, 331mammals 361, 363, 367Pleistocene extinction 415shrublands 486, 487spiders 498symbiogenesis 510temperate dry forests 517, 518temperate rain forests 519tropical rain forests and deforestation
527australian aborigines 461australian rats 360Australopithecus 443–444Australopithecus afarensis 258autapomorphy 110autecology 205–206Avena barbata 12Avena fatua 12Avena sativa 12Aventa fatua 278avermectins 59avocados 118avoidance 177, 180, 304, 305axolotl salamander 18axores 313ayrean grasswren 181Azadirachta indica 59azaelea 174Azolla 476azores 196
Bbacilli 40Bacillus 43Bacillus thuringiensis 59backswimmers 467bacteria 40–45, 41t, 44t
adaptation 5archaea 32–33biodiversity, concept of 47biodiversity, resources for agriculture
59biodiversity, resources for medicine
64–68biological classification 78, 79bioremediation 84coevolution 121–122commensalism 123, 124competition, ecological 133, 134convergence 143, 145, 148decomposers 170deep oceans, biodiversity in 171ecosystems 212eukaryotes 230–232insects 323Margulis, lynn 368mutualism 378–381natural selection 385, 389, 390nitrogen cycle 394parasitism 402, 404, 405pollution 423population biology 424–426symbiogenesis 508–510
bacterial chemosynthesis 549cbacteria-like organisms 32–33
Bacteriodes thetaiotaomicron 378Bactrian camel 173Bagherra kiplingi 496Baikal, lake 222, 469Baikal seal 469Baja, california 52–53baker’s yeast 262Balaenoptera musculus 365balanced pathogenicity 121, 405balanced polymorphism 434balance of nature 130Balanus 133Bald and Golden eagle Protection act 243Baldauf, Sandie 445bald cypresses 137, 369, 371, 512, 513bald eagle 108, 109, 219, 243Ballard, Robert 549cBalsamorhiza sagittata 188Baltica 396Bambi syndrome 198banana 62bandicoots 363Banks, Joseph 54, 56Barbary macaque 518barbs 83, 85, 86, 470barcode of life 203bark 254bark scorpion 36barley 10barnacles 35, 124, 133, 147, 158, 165Barnosky, anthony 416barrier reefs 149Barro colorado island 52, 56, 129Bartman, William 501Barton, andrew 52Barton, Joe 281Bartow county, Georgia 194, 221Bartram, John 233, 356Bartram, William 233, 439, 461, 462, 522basal angiosperms 255, 478Basidiomycota 261, 262basil 59Basilosaurus cetoides 399bastard toadflax 403Bates, Henry Walter 45–46, 50, 99, 542, 548cBatesian mimicry 45, 99Batrachochytrium 19Batrachochytrium dendrobatidis 18batrachotoxin 17bats 147, 418
biodiversity, patterns of 48convergence 146mammals 365–366parasitism 403pollination 418sexual selection 481
Bayer 68Bazzaz, Fakhri 298bdelloid rotifers 28Beagle, H.M.S. 162–163, 167, 271–273,
548cbeaks 86, 168, 169, 455beans 74bears 66, 67, 249, 311, 516, 518beavers 344, 370, 467bee(s)
arthropods 38camouflage and mimicry 98–99coevolution 120, 121convergence 145insects 325–326pollination 418, 419, 421
beeches 73, 78, 185, 504, 513, 515beef tapeworm 30–31Beelzebufo 155beer 262beetles 418
arthropods 37biodiversity, patterns of 51biomimicry 83camouflage and mimicry 99
xviii+598_Biodiversity_F.indd 571 11/17/11 1:13 PM

first entry572 tundra 572 first entry572 Index572
convergence 145decomposers 170deserts, warm 181greenhouse effect and biodiversity 297insects 324–325invasive species 331iPM 328Permian period, biodiversity in 412rivers, streams, ponds, and lakes 467
Belarus 516Benepe, adrian 217Bengal tiger 365Bennett, Hugh Hammond 242, 549cBennettitales 253benthic waters 171–172Benton, Michael 52, 408, 409Beringia 74Bering land bridge 74Berry, Wendell 13, 14, 226, 375beta diversity 126–128Betula 73, 92Betulaceae 186Betula glandulosa 91Betula occidentalis 91Betula pendula 264between-habitat diversity 48bewnnettitalean plants 477–478Bible 224–226bichirs 247bilateral symmetry 20, 20, 24–29bile 66binary code 83binary fission 41binocular vision 363, 366binomial nomenclature 77bioaccumulation 80–82biodiversity, concept of 46–47, 47tbiodiversity, patterns of 47–54, 47t, 129biodiversity, quantification of 54–57, 56tbiodiversity, resources for agriculture
57–64, 61, 63, 83, 215biodiversity, resources for medicine 64–69,
65t, 83, 215biodiversity hot spots 7, 52–53, 56, 75,
221, 234biofilms 41biogeochemical cycles 69–72, 71, 170, 212biogeography 72–77, 74t
deserts, warm 179–180Humboldt, alexander von 307Wallace, alfred Russel 543Wilson, edward O. 546
bioluminescence 41, 148, 171, 248biomagnification 80–82, 81t, 82t, 212, 372biomass 81, 209biomimicry 82–83bionomial nomenclature 548cbiophilia 228Biophilia (Wilson) 546bioprospecting 42bioremediation 42, 83–84biotechnology 58–59, 69bioterrorism 114bipedalism 442birches 73, 92, 186, 264, 304bird eggs 21bird-hipped dinosaurs 191birds 84–90, 85, 86, 88t, 147, 379
biomagnification 80–81camouflage and mimicry 99–100carson, Rachel louise 108, 109cladistics 112coevolution 118–119, 122commensalism 124communities, ecological 126, 128competition, ecological 134convergence 146, 147cretaceous extinction 151cretaceous period, biodiversity in 155cultural diversity 159Darwin’s finches 167–169
deserts, cool 173deserts, warm 181dinosaurs 193disjunct species 196extinction 235fossils and fossilization 257greenhouse effect and biodiversity 297,
298herbivory 305Jurassic period, biodiversity in 341keystone species 344life history, diversity of 350mammals 360, 363marshes and swamps 370mutualism 379natural selection 386, 389neogene period, biodiversity in 393parasitism 404predation 436–438reptiles 454seed dispersal 470–471sexual selection 480, 481tropical seasonal forests 537
bishop pines 128, 136, 196, 199–200, 200, 308, 428
bison x, 198, 439disturbances, ecological 198extinction 234federal government role in conservation
(U.S.) 239grasslands and savannas 279predation 439restoration ecology 461temperate deciduous forests 516
Bison bison 461bivalves 25, 26, 396, 467, 523Bjork, adam 479black bears 518black-bellied seedcrackers 389blackberries 186blackbirds 370black cherry trees 186, 516Black Death 323Black Forest 2Black Hills, South Dakota 91, 127, 196,
303, 511black honey-eater 134black ironwood 196blackjack oaks 515black locust 186black mamba 457black market 235black oaks 185, 308.309black pines 511black poplar 186black soil 531black sunbird 134black-tailed prairie dogs 366black vulture 124black walnut tree 113, 425black widows 498bladderworts 106–107blas spiders 497Blastocladiomycota 261blending inheritance 386blindness 31blister beetles 98–99blood 21, 250blood cells 320, 353, 360blood fluke 30blood vessels 257blueberries 91blue crabs 297bluefin tuna 172, 250bluegills 308blue heron 370blue jays 471blue oaks 6, 128, 278blue orchard bees 421blue poison dart frogs 18blue whale 365
Blyth, edward 384boas 456body shape 23bog birch 91bog people 91bogs, life in 90–91
boreal and subalpine forests 92ecosystems 208, 209, 212rivers, streams, ponds, and lakes 469seedless plants 474succession, ecological 505tundra 540
Boidae 456Boiga irregularis 333bois d’arc 118, 119, 186, 527bolas spider 99Bolivia 537Bomakellia 217bones 15, 16, 86, 193, 257, 258, 360–361,
454, 456bonobo 443Bonpland, aimé 307bony armor 246bony fishes
carboniferous period, biodiversity in 105
cretaceous period, biodiversity in 154fishes 247–250Jurassic period, biodiversity in 341neogene period, biodiversity in 392Paleogene period, biodiversity in 399Permian period, biodiversity in 412Quaternary period, biodiversity in 452Silurian period, biodiversity in 488triassic period, biodiversity in 523–
524book lungs 497boreal and subalpine forests 91–94, 92,
93tbiogeography 76communities, ecological 125disjunct species 196greenhouse effect 292greenhouse effect and biodiversity 297temperate coniferous forests 511tundra 540
Borneo 116, 526–527, 533, 534Borrelia burgdorferi 40, 404Bossert, William H. 545Bos taurus 12Botswana 175botulism 43Boulding, Kenneth 226–227, 427, 429Bouteloua 278Bouteloua curtipendula 198Bouteloua gracilis 198Bowers, Karen 420bowfins 247, 250bow-legged bug 98Bowler, Peter 165boxelder 505Boyd, christin 389BP (British Petroleum) Deepwater Horizon
oil spill (Gulf of Mexico, 2010) 549ccontinental shelves, biodiversity on
142–143ecology 206ecosystem services 214, 215, 217federal government role in conservation
(U.S.) 243, 244brachiation 442brachiopods
animals 27carboniferous period, biodiversity
in 105Devonian period, biodiversity in 182mass extinctions 374Permian extinction 407, 408, 410Permian period, biodiversity in 411Silurian period, biodiversity in 488triassic period, biodiversity in 523
xviii+598_Biodiversity_F.indd 572 11/17/11 1:13 PM

tundra 573Index 573
Brachiosaurus 341Brachycera 323bracken ferns 476bracts 135, 377Bradshaw, toby 492, 494brain 351–352, 365, 366brakefern 84branchiopods 158Branta sandiviensis 258Brassica campestris 12Brassicaceae 74, 186Brassica napus 12Brassica oleracea 186, 310Brazil 498, 531, 532–534, 537Brazilian rubber tree 188breaching whales 365bread 262bread mold 261breast cancer 354brewer’s yeast 262brine shrimp 305–306bristlecone pines 92, 137, 138, 281, 281, 354British columbia 494British Petroleum. See BP Deepwater
Horizon oil spillbroadleaf trees 513, 518broad-sense heritability 432broad-spectrum antibiotics 68brome 278bromeliad plants 52, 526Bromus japonicus 278Bromus tectorum 278Brontosaurus 191, 341brooding frogs 68–69brown algae 232, 446, 447brown bears 516Brownlee, David 374brown plant hopper 327brown recluse spiders 498brown tree snake 333bryophytes 472–474Bryozoa 27Bt toxin 59bubonic plague 38, 323Bubulcus ibis 124Buchloë dactyloides 274, 276Buchnera 323Buchnera aphidicola 510bucket orchid 419budding 262buffalo. See bisonbuffalo gourds 61buffalo grass 274, 276Buffon, Georges-louis leclerc, comte de
77, 548cBufo alvarius 17Bufo marinus 331Bufo periglenes 18, 116, 235bugs 467bulblets 538bull nettle 133–134bumblebees 418, 419bundleflower 61buoyancy 250Buphagus africanus 122Bureau of Reclamation 242Burgess Shale 257Buri, Peter 258Burley, nancy 483bur oaks 6, 303, 309, 511bursage bushes 175Bursera simaruba 369, 515burying beetles 170, 325bush(es) 66, 91, 168, 175, 206, 233Bush, George H. W. 2, 219–220Bush, George W. 220, 225bushmeat 235buttercups 185butterflies
arthropods 37–38Bates, Henry Walter 45
coevolution 120, 121Dna studies of biodiversity 202–203herbivory 305insects 324life history, diversity of 349predation 437
butterfly effect 270butternuts 113butterworts 106
Cc4 grasses 393c4 photosynthesis 393caatinga 537cabbage 310cactaceae 179–180, 188–189cacti 180, 189, 418
biodiversity, resources for agriculture 61
coevolution 119communities, ecological 127deserts, warm 175, 177, 179–180dicots 188–189Galápagos islands 272herbivory 303neogene period, biodiversity in 393tropical seasonal forests 537
cactus finch 169caddisflies 466caecilians 17caecum 365Caenorhabditis elegans 27, 68, 353calamitales 473Calamites 105, 475calanolide a 68calcium 70, 72, 212calcium carbonate 101–102calcium carbonate shells 256calcium silicate 102Caleidocrinus 397california x
adaptive radiation 6agriculture 12biodiversity, patterns of 52–53biodiversity, resources for agriculture
61, 62biogeochemical cycles 72biogeography 73, 75, 76birds 90boreal and subalpine forests 92communities, ecological 127–129conifers 136, 137deserts, cool 173deserts, warm 175, 176, 179, 181disjunct species 196disturbances, ecological 199, 200endemic species 221, 222fossils and fossilization 258greenhouse effect and biodiversity 296marshes and swamps 369restoration ecology 463shrublands 486–487temperate dry forests 517, 518tundra 537
california black oak 309california poppy 487Callinectes sapidus 297Calluna 487calment, Jeanne 354Calocedrus decurrens 137Calochortus 144Calophyllum lanigerum 68caltrop 180. See also puncture vineCalyptra minuticornis 324Camarhynchus pallidus 169cambrian explosion 95–96cambrian period, biodiversity in 96–97, 97,
97t, 245, 257camels 112, 173, 181Camelus bactrianus 173
Camelus dromedarius 181camouflage and mimicry 17, 98–101, 99,
100t, 248–249, 436–438, 455, 457Campsis radicans 120canada 92, 196, 257, 537canada lynx 438canary islands 313cancer 66, 353, 354Candida 262Candida albicans 262, 379cane toads 331, 332canfield, Donald 373canfield ocean 373canines 308Canis familiaris 12, 78, 308Canis latrans 78Canis lupus 46, 78, 308canola 12canopy 526, 531canopy trees 528t–529tcap and trade 2cape Verde 313Capra aegagrus 12capsule 40carapace 37, 157, 455Carassius auratus 248carbon 70, 212carbon cycle 71, 101–103, 102tcarbon dioxide
amphibians 17animals 23communities, ecological 126coral reefs 149decomposers 170fungi 262hybridization 310rivers, streams, ponds, and lakes 467
carbon dioxide, atmospheric 102t, 284t, 288t
agriculture 14biogeochemical cycles 70, 71cambrian period, biodiversity in 96,
97tcarbon cycle 101–103carboniferous period, biodiversity
in 104coevolution 117coral reefs 150cretaceous period, biodiversity in 153decomposers 170desert(s), warm 177Devonian period, biodiversity in 182,
183ecology 205, 206ecosystems 211, 212ecosystem services 214–216federal government role in conservation
(U.S.) 243, 244fossils and fossilization 258Gaia 267–270greenhouse effect 279, 284t, 287–293herbivory 305Jurassic period, biodiversity in 339,
340tmass extinctions 375neogene period, biodiversity in 392,
393Ordovician period 396, 397Paleogene period, biodiversity in 399Permian extinction 408–411Quaternary period, biodiversity in
450, 451Silurian period, biodiversity in 488,
489tsuccession, ecological 501triassic period, biodiversity in 523tropical rain forests and deforestation
533, 534tundra 540
carbon emissions 293t, 549c. See also car-bon dioxide, atmospheric
xviii+598_Biodiversity_F.indd 573 11/17/11 1:13 PM

first entry574 tundra 574 first entry574 Index574
carbon footprint 293carboniferous period, biodiversity in 103–
106, 104t, 105amphibians 16insects 320mammals 361seedless plants 473seed plants 477
carbon isotope ratios 258, 408carbon sequestration 290carbon sinks 102, 102tcarbon sources 102Carcarhinus obscurus 344Carcharodon megalodon 247, 392Carcinoscorpus rotundicaudatus 46Cardiocaryon 113Carex aquatilis 540caribou 76, 538carnassial teeth 363Carnegeia gigantea 180carnivora 78, 363–365carnivores 365
adaptation 5biological classification 78competition, ecological 132dinosaurs 191ecology 206ecosystems 208, 209mammals 360, 361, 363–365triassic period, biodiversity in 524
carnivorous diets 210carnivorous plants 91, 106, 106–107, 107t,
438carp 248, 330–331, 333–334carpels 252, 252, 254Carpinus putoensis 235carrion feeders 151carroll, Scott 389carrots 188carrying capacity 429carson, Rachel louise 107–109, 108, 549c
biomagnification 82federal government role in conservation
(U.S.) 242iPM 328natural selection 390resistance, evolution of 459
Carsonella ruddii 510cartilaginous fishes 246–247, 250Carya 186Carya illinoensis 515cascade Mountains 76cassowaries 87Castanea 73Castanea americana 186, 516Castanea dentata 234, 344–345castor bean 187–188, 303Castor canadensis 370Castorocauda 341Casuarina equisetifolia 333casuarina trees 333cat(s) 12, 112catalogue of life (internet database) 57catarrhines 442caterpillars 37, 98, 117, 305, 324, 500catfish 248Catharanthus roseus 66, 67, 527Cathartes aura 124catkins 185, 186cattails 61, 78, 377cattle 13, 210cattle egrets 124, 427caudal fin 246cavendish banana 62cBD. See convention on Biological
DiversitycBOl. See consortium for the Barcode of
lifeccD. See colony collapse disorderCeanothus bush 487cedar lake Bog, Minnesota 208, 208t, 209
cedars 137, 200, 313, 331, 333Ceiba 527Ceiba pentandra 527cell fossils 42cell membranes 32, 40, 230cellulose 303cell vacuoles 304cell walls 32, 40, 230, 260Cenchrus 144Cenchrus spinifex 145cenozoic era 6, 8, 365, 392–394, 398–401,
450–452, 478centipedes 27, 35central america 9, 189, 296, 527, 532, 537central asian deserts 173central Mexico 486central Valley, california 369Centris tarsata 421Centruroides exilicauda 36cephalocarida 158cephalopods
animals 25, 26carboniferous period, biodiversity
in 105cretaceous period, biodiversity in 154deep oceans, biodiversity in 171Devonian period, biodiversity in 182Ordovician period 396–397Permian period, biodiversity in 411
Cephalotes aratus 404cephalothorax 35, 37, 157, 496, 497Ceratiocaris 488Ceratopealum 519ceratopsians 192Ceratosaurus 341cercopithecine 442cercozoans 447–448cereals 263Cerris 73Cervus canadensis 221cetaceans 365, 365cetartiodactyls 365cetorhinidae 171chaco canyon, new Mexico 461, 461chaetognatha 28chagnon, napoleon 546challenger Deep 171Challenger expedition 548cChamaesaura 457Chamaesyce 168chambers, Robert 165chameleonidae 457channel islands 12, 75chaparral 129, 199, 486–487, 517character displacement 169charadriiformes 89charcoal 416charismatic megafauna 363Charniodiscus 217, 218charophyceae 448cheatgrass 278cheese 262cheetahs 119chelicerae 34–35, 496–499chelonians 455–456chemical defenses 117, 144, 147, 436chemosynthetic bacteria 41chernobyl 82cherokee nation 196, 238cherry trees 66, 186chert gap 407chesapeake Bay 462–463chestnut(s) 73, 234, 344–345, 516chestnut blight 262, 428, 516chicago 229chickadees 69chickens 89, 210, 404chicxulub crater 374chiggers 36chihuahuan Desert 175, 180childbirth 354–355
child care 352chile 486, 517Chiloglottis trapeziformis 99chilopoda 35chimaeras (ghost sharks) 247chimeras (cells) 232chimpanzees 235, 305, 351, 354, 443china
agriculture 9cloud forests 115conifers 137continental shelves, biodiversity on 141endemic species 222extinction 233, 235fishes 245living fossils 356Permian extinction 408reptiles 456temperate deciduous forests 516
chine 49chinook 467–468chiroptera 365–366chitin 260chlorophyll 22, 448chlorophyta 448chloroplasts
bacteria 42biomimicry 83coevolution 122eukaryotes 230–232Margulis, lynn 368, 369protists 445–446, 448symbiogenesis 508–510
chloroquine 68choanocytes 22, 449choanoflagellates 20, 448–449chocolate 60chocolate mint 59chokecherry 303, 511cholla 180chondrichthyes 246chordata 29, 78, 79chordates 29, 244–251christian environmentalism 225christianity 163, 164, 224, 225christmas island 156chromists 447chromosome(s) 90, 230, 232chromosome doubling 309–310, 491–492chrysalis 37, 324Chrysemys picta 455chrysler corporation 84Chthamalus 133chum 467chytridiomycota 261chytrid protists 18cicada 323cichlid fish 6–7ciconiiformes 89cilia 26, 29, 509ciliates 447Cinara cedri 510Cinclus mexicanus 466cingulata 367circulation 360, 455circulatory system 20–21, 26, 34, 147, 320,
496–497citeS. See convention on the international
trade in endangered Speciesclades 80, 112–113Cladina 348cladistics 109–114, 111t, 113t, 114tcladograms 109–110, 111t, 112, 114Cladonia rangiferina 381clams 25, 171, 172clark, William a. 414clarkia, idaho 256clark’s nutcrackers 137class (taxonomic) 78Claviceps 262Claviceps purpurea 264
xviii+598_Biodiversity_F.indd 574 11/17/11 1:13 PM

tundra 575Index 575
claws 363clean air act (1970) 243, 549cclean air act amendments (1990) 2clean Water act 243, 549cclements, Frederic 79, 124Clethropsis 492cleveland, Grover 239click beetles 99climate
biodiversity, patterns of 48–49boreal and subalpine forests 92cambrian period, biodiversity in 96carboniferous period, biodiversity
in 103cretaceous period, biodiversity in 154Devonian period, biodiversity in 182fossils and fossilization 258ice ages 316–317Jurassic period, biodiversity in 339neogene period, biodiversity in 392Ordovician period 396Paleogene period, biodiversity in 398Permian period, biodiversity in 411Pleistocene extinction 416Quaternary period, biodiversity in 450Silurian period, biodiversity in 488triassic period, biodiversity in 523
climate change 416. See also global warm-ing
climatic shifts 295, 534climax forest 500climbing fern 476clinton, Bill 219–220clitellum 27cloaca 18, 86, 87, 250, 361–363, 455clonal organisms 431clones 262cloning 159, 220closed circulatory systems 21closed-cone pine forests 199–200Clostridium 43Clostridium botulinum 43Clostridium dificile 68cloud forests 115–116, 115t, 222, 520cloud formation 313clouds 211, 311clovis people 343, 416, 417club mosses 105, 472–474, 475cnidaria 23, 24, 149, 162, 218Cnidoscolus texanus 133–134coal
acid rain and fog 1biogeochemical cycles 72biomagnification 82carbon cycle 102carboniferous period, biodiversity
in 103environmental ethics 227fossils and fossilization 256greenhouse effect 289greenhouse effect and biodiversity 295insects 320Permian extinction 408seedless plants 473
coal balls 256, 473coal gap 408coast Guard, U.S. 142coatimundis 331cobra 457cobra lilies 106coburn, tom 109cocci 40, 41cockleburs 83, 470cockroaches 105, 322, 356, 412, 426, 459cocoa 60coconuts 470cocoon 37, 324cocos island 335cocos island finch 169codling moths 328coelacanths 249
coelenterates 23, 24coelom 20Coelophysis 524coelourosaurians 192Coenobita clypeatus 156coevolution 116–123, 116tcoffee trees 60coho fish 467cold seeps 171–172cold War 263coleoptera 37, 324–325collembolans 321Colobanthus quitensis 174colobines 442colombia 527colonies 38colony collapse disorder (ccD) 421colorado 258, 344colorado Desert 126, 175, 179colorado potato beetle 325colorado River basin 465colorado River toad 17coloration 17, 98, 436, 437color blindness 258color vision 441colugos 366Columba livia 89, 386columbia River basin 465columbiformes 89columbus, christopher 313Comandra 403comb jelly 23–24, 332Comephorus baikalensis 222Comephorus dybowskii 222comets 373comita, liza 52commensalism 121, 123–124, 207, 401,
402, 404–405commensals 378communication 365communities, ecological x, 124–130, 126m,
127m, 129, 130, 549cbiodiversity, patterns of 51biodiversity, quantification of 54biological classification 79competition, ecological 134–135deserts, cool 173disturbances, ecological 198–200ecology 207ecosystems 207herbivory 305leopold, aldo 347life history, diversity of 355
comoro islands 249compass plant 276compensatory regrowth 305competition 5, 481–483competition, ecological 49, 129, 131–135,
531competition, species 75, 169, 367–368,
384–390competitive enterprise institute 291composite family 74, 188composting 461, 531compound eyes 320computers 83Conartium ribicola 297condensation 177, 311Condylostylus 321cones 135, 417cone snails 26, 66Confusciusornis 156congress, U.S. 548c, 549c
adaptation 2carson, Rachel 109environmental psychology 229federal government role in conservation
(U.S.) 238, 239, 242, 243greenhouse effect 281, 290, 292Pinchot, Gifford 413, 414
coniferous forests 92, 296
conifers 92, 135–138, 136–138acid rain and fog 1biogeography 75carboniferous period, biodiversity
in 105endemic species 222Jurassic period, biodiversity in 341pollination 417, 420seed plants 477, 478temperate coniferous forests 511temperate deciduous forests 513temperate rain forests 518
connate fusion 254connell, Joseph 133conodonts 97, 245–246, 258Conraua goliath 17conservation
deep oceans, biodiversity in 172–173deserts, warm 181disturbances, ecological 200–201federal government role in conservation
(U.S.) 238–244fishes 250–251fungi 264Gaia 269habitat fragmentation 302island biogeography 336Janzen, Daniel H. 338–339Maathai, Wangari 358–359Muir, John 378mutualism 381Pinchot, Gifford 413–414pollination 421reptiles 456tropical rain forests and deforestation
532conservation biology 138–141, 439–440conservation tillage 391Consilience (Wilson) 546consortium for the Barcode of life (cBOl)
203consumers 208continental drift 72continental shelves, biodiversity on 49,
141–143, 142, 316, 409contour feathers 85–86contour plowing 391Conus 66convention on Biological Diversity (cBD)
140, 235convention on the international trade in
endangered Species (citeS) 218, 235, 244
convergence 73, 88, 110, 143–149, 145, 146t, 147, 366
conway, erik 280conway Morris, Simon 148–149cook, James 54cookie-cutter shark 247Cooksonia 472, 488cooper, ellwood 333cope, edward Drinker 190copenhagen conference 549ccopepods 158copper 71copperhead snake 457coprolites 257Copsychus sechellarum 234Coragyps atratus 124coral bleaching 23, 149, 150, 463Corallorhiza 144, 170, 403coral reef(s) 149–151, 150
animals 29biodiversity, patterns of 49, 52continental shelves, biodiversity on
141–142greenhouse effect and biodiversity 297mutualism 381succession, ecological 501–502
coral reef fish 380coral reef shrimp 145
xviii+598_Biodiversity_F.indd 575 11/17/11 1:13 PM

first entry576 tundra 576 first entry576 Index576
coral-root orchid 170corals 67, 344coral snakes 457Cordaites 477cordgrass 297, 370Cordia alliadora 144core eudicots 185Corema album 196Corema conradii 196corixidae 467cork oak 6, 73, 518corkscrew Swamp Sanctuary, Florida 370cormorant 76corn. See maizecorn borer larvae 490–491correns, carl 369Coryanthes panamensis 419costanza, Robert 217costa Rica 339, 526cottonmouth snake 457cotton sedge 91cottonwoods 73, 186, 187, 483, 516cottony cushion scale 328cotyledons 184, 185, 375courtenay-latimer, Marjorie 249cowbirds 99–100coweeta watershed 312cowles, Henry chandler 504cows 11–12, 30–31, 111–112, 122, 304–
305, 378coxa 320coyote 78crabs
arthropods 37biodiversity, concept of 46biodiversity, resources for medicine 67crustaceans 156deep oceans, biodiversity in 172Jurassic period, biodiversity in 341living fossils 356mutualism 381
crab spiders 497, 497crafoord Prize 339cranberries 91craneflies 323Crassostrea virginica 463Crassula erecta 128Crataegus 119crayfish 157creationism 166, 225–226, 356creeks 465, 466Crematogaster 380Crenarcheota 33creosote bushes 175cretaceous extinction 151–153
carbon cycle 102, 103Gaia 268–269mammals 362mass extinctions 372, 374succession, ecological 506
cretaceous period, biodiversity in 153–156, 153t–154t, 155
birds 85, 87deep oceans, biodiversity in 171dinosaurs 193flowering plants 252–253insects 320–321mammals 361, 362
crickets 321crime 229crinoids 397crocodiles 221, 341, 454–456, 524Crocodylus mindorensis 221Crocodylus porosus 456croll, James 315crop plants 10t, 12, 381, 501crop production 13, 304crop rotation 13, 63, 327, 394crossopterygians 247, 249cross-pollination 310, 460cross timbers forest 515
crotalinae 181Crotalus cerastes 457crowberries 75, 196crown-of-thorns starfish 149crows 118crucifers 186crustaceans 31, 35, 154, 156–158, 157, 341crustose lichens 348Cryphonectria parasitica 234, 262, 344–345Cryptocerus 356Cryptococcus 262cryptodires 455–456cryptograms 77Cryptomeria japonica 137cryptomonads 447Cryptostylis 420Cryptotethya crypta 67ctenizidae 497ctenophores 23–24cuckoos 99–100, 405cucumbers 470cud 365Culex pipiens 404, 491Culex tarsalis 404cultivated mustards 12cultivated oats 12cultural diversity 158–160, 159cunoniaceae 519cupressaceae 137Cupressus macrocarpa 318Curatella americana 278curculionidae 325Curcurbita foetidissima 61curly-nosed primates 366currie, cameron 380curtis, John t. 54, 55Cuscuta 110, 403cushion plants 538cuyahoga River, Ohio, fire (1969) 242cyanobacteria
agriculture 13animals 22–23bacteria 41–44deep oceans, biodiversity in 170eukaryotes 232fossils and fossilization 256fungi 264Gaia 267lichens 348nitrogen cycle 394rivers, streams, ponds, and lakes 466symbiogenesis 509
cycads 341, 477, 478cyclic aMP 448cycliophora 21, 28, 549ccyclones 286Cycloprosopon 341Cyclorana platycephalus 180cyclosporine 66, 262Cydia pomonella 328cynodonts 361Cynomys 344Cynomys ludovicianus 366cyperaceae 377Cyperus alternifolius 222cypresses 137, 369–371Cyprinodon diabolis 301Cyprinodon nevadensis nevadensis 301Cyprinodon salinus 180Cyprinus carpio 248Cyrtocara eucinostomus 481cystic fibrosis 434cysts 29–30cytoplasm 40, 230, 231cytoplasmic inheritance 369
ddaddy longlegs 35daffodils 375Daily, Gretchen 344
dairy products 42daisies 177daisy trees 272Daktulosphaira vitifoliae 60dams 251, 469, 533damselflies 321, 466, 481Danaus plexippus 117dandelions 330, 491Daphne Major 273Daphnia 158Darlingtonia californica 106Darwin, charles 161–167, 163, 548c
Bates, Henry Walter 45biodiversity, patterns of 50biodiversity, quantification of 54biogeography 72biological classification 78carnivorous plants 107cladistics 113–114communities, ecological 130competition, ecological 131Darwin’s finches 167–169disjunct species 195flowering plants 253Galápagos islands 271–273Humboldt, alexander von 307island biogeography 335life history, diversity of 351natural selection 384–390population biology 424, 427, 429sexual selection 479, 480speciation 489, 490symbiogenesis 507–508thoreau, Henry David 522Wallace, alfred Russel 542–545
Darwin, Robert 161, 162Darwin, Susannah Wedgewood 161Darwin’s finches 8, 134, 167–170, 168t,
432–433, 491, 493, 494Dasypus novemcintus 367date palms 62, 63, 179Davis, Marc 373Dawkins, Richard 390dawn redwoods 51, 136–137, 222, 356DDt 81t, 549c
biomagnification 80–82carson, Rachel louise 108, 109federal government role in conservation
(U.S.) 242, 243iPM 328resistance, evolution of 460
Dean, Rebecca 481death 353–355Death Valley, california 176, 176, 180,
301, 330debridement 38decapods 35, 157Deccan traps 374deception 352deciduous forests 513, 515
adaptive radiation 6biodiversity, patterns of 49, 52communities, ecological 126greenhouse effect and biodiversity 296keystone species 344–345Paleogene period, biodiversity in 398temperate coniferous forests 511temperate deciduous forests 512–516tropical seasonal forests 537
deciduous oaks 185deciduous white oaks 6, 7decomposers 71, 170
animals 27bacteria 42carbon cycle 101commensalism 124ecology 205, 206ecosystems 208fungi 261–263Gaia 267, 268Janzen, Daniel H. 339
xviii+598_Biodiversity_F.indd 576 11/17/11 1:13 PM

tundra 577Index 577
decompositionagriculture 14bogs, life in 91fossils and fossilization 256, 257Gaia 269greenhouse effect 286–287, 290nitrogen cycle 394tropical rain forests and deforestation
530Deep ecology 226deep oceans, biodiversity in 170–173deep ocean vents 32, 41, 43Deepwater Horizon oil spill (2010). See BP
Deepwater Horizon oil spilldeer 12, 56, 111, 338, 344, 345, 518deer ticks 36, 404Defense, Department of 239, 241–242defense mechanisms 76, 146defoliant chemicals 533deforestation 312, 312, 313, 461, 525–526,
531, 533, 534Deinococcus radiodurans 43Delaware 194, 195Delmarva Peninsula 194, 196, 221demes 423Demodex folliculorum 36Dendroaspis polylepis 457Dendrobates azureus 18dendrobatid frogs 67Dendroctonus ponderosae 297Dendroctonus rufipennis 297dengue fever 324denitrification 42, 394dentary bone 361depth perception 441derived traits 110–112Dermochelys coriacea 455Dermoptera 366Descent of Man, The (Darwin) 166, 479,
544Deschampsia antarctica 174, 296desert(s) 130
adaptation 6arthropods 36biodiversity, resources for agriculture
61biogeography 73convergence 144ecology 206life history, diversity of 351tropical seasonal forests 537
desert(s), cool 174–175, 174tdesert(s), polar 175desert(s), warm 175–182, 176, 177,
178t–179t, 180desert frogs 6, 180desert rhubarb 179desert shrubs 129Desmanthus illinoensis 61detergents 466detritivores 126Deuteromycota 261deuterostomes 28–29Devil’s Hole 301devil’s rider walking stick 436devil’s thorn 180Devonian mass extinction 372–373Devonian period, biodiversity in 15, 182–
184, 183t, 184, 320, 473, 477DeVries, Hugo 369DeWaal, Frans 443Diamond, Jared 11, 134diapsids 87, 412, 454diatomaceous earth 128, 258diatoms 381, 446dibamid lizards 458Dicamba 460dicots 113t, 114, 114t, 144, 184–190, 185,
187–189, 254, 255Dictyoptera 322, 356Dictyostelium discoideum 448
Didelphus virginiana 354, 362, 437digestion 86, 260, 378–379digestive bacteria 43digestive systems 5, 23, 24, 27, 29digital code 83dikaryotic cells 260Dimetrodon 361, 412, 412dingos 367dinoflagellates 46, 232, 447Dinosaur national Monument 258dinosaurs 190–194, 191–193
adaptive radiation 8birds 84–87cladistics 112–113cretaceous extinction 151cretaceous period, biodiversity in
153–156fossils and fossilization 257, 258Jurassic period, biodiversity in 339–
341mammals 361, 362mass extinctions 374reptiles 454triassic period, biodiversity in 524
Dionaea muscipula 106Dioscaryon 113Diospyros kaki 60Diospyros virginiana 60, 504, 527diplococci 41Diplodocus 341diploid cells 260diploid phase 472, 477diplomacy 140Diplopoda 35Dipodomys 181dipper ouzel 466Diprotodon 393Diptera 323dipterans 38Dipterocarpaceae 527, 537Dipylidium caninum 31directional selection 387, 388t, 389discicristates 445–446disease x
agriculture 11animals 29–31arthropods 38bacteria 42biodiversity, resources for agriculture
58, 62coevolution 121ecosystems 210fungi 263greenhouse effect and biodiversity 298insects 323, 324parasitism 404pollination 421population biology 428restoration ecology 461succession, ecological 501temperate deciduous forests 516
disjunct species 194–197, 195disk florets 188dispersal 73–75, 194, 196, 348, 388, 403“Dispersion of Seeds, the” (thoreau)
228–229, 521Distichlis palmeri 60distraction mimicry 98distribution 47–48, 55disturbances, ecological 197–202, 198–
200, 201tbiodiversity, patterns of 51biodiversity, quantification of 56biogeography 77ecology 207extinction 233marshes and swamps 370succession, ecological 499–502,
505–506Diversity of Life, The (Wilson) 545, 546diversity selection 387, 388t, 389
diving beetles 467divisions 78Dna
animals 27archaea 32–33bacteria 40, 43, 44biomimicry 83eukaryotes 230–232fossils and fossilization 256fungi 262mammals 365
Dna sequencing 110–114Dna studies of biodiversity 202–204
biodiversity, resources for agriculture 58–59
biodiversity, resources for medicine 69deep oceans, biodiversity in 171population genetics 432–433speciation 492symbiogenesis 508
dobsonflies 466Dobzhansky, theodosius 119, 430, 490dodder 403dodos 234, 462dogs 12, 75, 78, 112, 308dog tapeworm 31Dolomedes 467dolphins 365domains (taxonomic) 79domesticated grapes 187dominance-diversity curves 55dominant alleles 434donkeys 12, 173, 308, 330dorsal fins 248dorsal nerve cord 29dorsal surface 20double fertilization 252Douglas firs 137, 511, 518dove(s) 89, 350Dover, england 258down feathers 86dragonfishes 248dragonflies 105, 105, 320, 321, 412, 466,
481drainage basins 465Dreissena polymorpha 26, 330Drina River Valley 222dromedary camels 181Dromiciops gliroides 363Drosera capensis 106Drosera rotundifolia 91, 106Drosophila 430, 493, 494Drosophila bifurca 479Drosophila melanogaster 68, 353Drosophila sechellia 493Drosophila simulans 493drought 168, 169, 286, 297drought adaptations 176–180drought resistance 234dry-nosed primates 366dry soils 61dry summers 312duck-billed platypus 362, 363ducks 89duckweeds 467dung beetle 325Dunkleosteus 183DuPont 68Dürer, albrecht xidusky shark 344dust bowl 223, 223, 239, 241, 242Dutch elm disease 262dwarf azaelea 174dwarf lantern shark 246Dytiscidae 467
Eear bones 16, 361early birds 87early-successional habitats 499, 500
xviii+598_Biodiversity_F.indd 577 11/17/11 1:13 PM

first entry578 tundra 578 first entry578 Index578
earth Day 549cearthquake (Japan, 2011) 549cearth’s endangered creatures 219earth Summit 549cearthworms 26–27, 166, 170easter island 66, 313eastern red cedar 200, 331ebenaceae 527ebola virus 121ebony 527Ecballium elaterium 470ecdysis 27–28echidnas 363echinoderms 28–29, 149, 154, 397, 411, 488echiura 28echolocation 146Ecklonia 141ecological degradation 461ecological footprint 292–293, 292t, 521ecological islands 221ecological rules 381ecological succession. See succession, eco-
logicalecology xi, 205–207, 548c. See also restora-
tion ecologyeconomy of nature 351ecosystems x, 207–213, 208t, 209t, 210m,
211tagriculture 12bacteria 42, 43biogeochemical cycles 69–72biomimicry 83fungi 262–264hydrological cycle 311–314invasive species 333keystone species 343–345mammals 363natural selection 385restoration ecology 460–464
ecosystem services 213–217, 216tecotones 125ectomycorrhizae 263Ectopistes migratorius 234ectoprocta 27ectotherms 17, 249, 455ecuador 12, 52, 68, 167–169, 527, 533, 538ecuadorean amazon 525edaphic factors 128edentates 367edge effect 139, 301ediacara Hills 217ediacaran organisms 95–97, 217–218, 372edmondstone, John 162eels 172, 247–238effluent 466egg(s)
amphibians 18animals 20, 21, 25, 29–31arthropods 38birds 87dinosaurs 190reptiles 454, 455
egg capsule 246egg cells 473, 474, 479egypt 30ehrlich, Paul 344Eichhornia crassipes 84, 372, 467eiseley, loren 165elapidae 456–457elapids 456–457elateridae 99electric eels 248, 307electrolocation 146electrophoresis 432Electrophorus electricus 248electroreception 246elephant(s) 276
coevolution 119keystone species 343–344life history, diversity of 350, 351mammals 366–367
natural selection 385tropical seasonal forests 537
elephant birds 87elephantiasis 31, 402Eleutherodactylus coqui 18elevation 176eliasomes 471elk 221Elkinsia 477ellesmere island 16, 174ellstrand, norm 12elms 73, 262el niño 168, 271el niño Southern Oscillation (enSO) 171,
284Elodea canadensis 467embryo(s) 28, 362, 477embryonic development 479emerson, Ralph Waldo 378, 521, 522emperor scorpion 36empetraceae 75, 196Empetrum 196Empusa muscae 261emus 87encephalitis 324encyclopedia of life (internet database)
47, 57endangered species 218–221, 219t, 220t,
235, 242, 244endangered Species act (eSa) 219, 220,
243, 549cendemic species 221–222, 526Endoceras 397endomycorrhizae 263endopterygotes 323endosperm 484endospores 43endosymbiotic bacteria 510endotherms 86, 249, 361energy balance 205–206, 211energy efficiency 210, 292energy flow 48, 207–211energy pyramid 209energy storage 209england. See Great Britainenglish oak 6, 303Ensatina 494enSO. See el niño Southern Oscillationentoprocta 27environmental degradation 463, 464environmental ethics 215, 223, 223–228,
347environmental impact 220environmentalism 520–522environmental Protection agency (ePa)
243, 327, 549cenvironmental psychology 228–230environmental terrorism 226enzymes 32, 231Eomaia 156Eoraptor 191ePa. See environmental Protection agencyephemeral plants 177, 513ephemeroptera 321epibatidine 17, 67, 68epidermal scales 367epigenetics 3Epipedobates tricolor 17, 67epiphytes 52, 376, 513, 518, 526, 526, 531Epischura baikalensis 222equador 116equilibrium 433–434equisetophyta 474–475equitability 56Equus 452Equus asinus 12Equus caballus 12Equus hemionus 173ergot 262, 264Erica 196, 487ericaceae 91
Eriophorum vaginatum 91, 195, 540Eristalis tenax 467erosion 13eruasian walnuts 113erwin, Douglas 148, 374, 407, 408Eryops 412eSa. See endangered Species actEscherichia coli 43, 143, 379Eschscholzia californica 487Eschscholzia mexicana 497española 169Essay on the Principle of Population
(Malthus) 164, 548cEstopistes migratorius 461estuaries 370ethical systems 223ethiopia 56, 60ethnobotanists 66Etmopterus perryi 246etterson, Julie 3eubacteria 79eucalyptus 92, 333, 463, 487, 517, 518Eucalyptus globulus 518Eucalyptus pauciflora 92Eucryphya 519eudicots 185–189, 255, 478eugenics 544Euglandina rosea 331Euglena 445–446eukaryotes 47t, 230–233, 232t
animals 19–31bacteria 42, 44coevolution 122population genetics 430protists 445–449species diversity 47tsymbiogenesis 508–509
eukaryotic flagella 232Euphorbia 187euphorbiaceae 187Euphorbia marginata 133–134Eupithecia 324eurasia 11–12, 73–76, 174, 415eurasian watermilfoil 330europe
acid rain and fog 2adaptive radiation 6agriculture 9biodiversity, patterns of 49biogeography 73, 76boreal and subalpine forests 92Devonian period, biodiversity in 182dicots 186greenhouse effect 285ice ages 318invasive species 331, 333Paleogene period, biodiversity in 398temperate coniferous forests 511temperate deciduous forests 513, 515,
516temperate dry forests 517tundra 537
european blackcap 297, 298european grapevines 60european honeybees 326, 332european rock dove 89Eurotamandua 401Euryarcheota 33eurypterids 396Eusthenopteron 249Eusthenopteron foordi 15eutrophication 13, 150, 394, 466eutrophic water 466Evangeline (longfellow) 201evaporation 206, 311evapotranspiration 313evening primrose 345, 418everglades 369, 372evergreen(s) 92, 513, 515, 525evergreen broadleaf trees 537evergreen oaks 185
xviii+598_Biodiversity_F.indd 578 11/17/11 1:13 PM

tundra 579Index 579
evergreen palm trees 537evergreen white oak 6evolution 548c
adaptation 2–6adaptive radiation 6–14agriculture 8–12amphibians 15–17bacteria 43–44biodiversity, concept of 46biodiversity, patterns of 49biodiversity, resources for agriculture 58biogeography 72, 75biological classification 78birds 85cladistics 113–114convergence 148crustaceans 157Darwin, charles 161–166Darwin’s finches 167–169disjunct species 195ecology 206environmental ethics 226eukaryotes 231fishes 246, 249flowering plants 254fossils and fossilization 258Galápagos islands 271–273Grant, Peter and Rosemary 273–274herbivory 306ice ages 318life history, diversity of 351–355mammals 362–363Margulis, lynn 368–369mass extinctions 374natural selection 384–390parasitism 402pollination 420population biology 426population genetics 430, 434protists 445resistance, evolution of 459–460sexual selection 478–484speciation 494symbiogenesis 507–508tropical rain forests and deforestation
526Wallace, alfred Russel 542–545
evolutionary diversity 261–262evolutionary medicine 58exaptation 4–5excavates (protists) 445exopterygota 321exoskeleton 34, 96, 156, 318, 496exponential population growth 425, 427,
429Expression of Emotions in Men and
Animals, The (Darwin) 166Extatosoma tiaratum 322external digestion 260external fertilization 18, 21, 26, 249external parasites 379external skeletons 28extinction x, 233–237, 236t
agriculture 13amphibians 18–19biodiversity, resources for agriculture
59–62biogeography 75birds 87cambrian period, biodiversity in 97coevolution 118–119conservation biology 139cretaceous extinction 151–152cretaceous period, biodiversity in 156Devonian period, biodiversity in 184dinosaurs 193disjunct species 196ecology 207grasslands and savannas 278–279greenhouse effect and biodiversity
296, 297
ice ages 318mass extinctions 372–375Ordovician period 397Permian extinction 407–410Permian period, biodiversity in 412Pleistocene extinction 414–417Quaternary period, biodiversity in 452restoration ecology 462Silurian period, biodiversity in 489succession, ecological 501, 506triassic period, biodiversity in 524
extractive preserves 533, 534extraterrestrial impact 152extremophiles 32, 33, 43, 134eyes
animals 26arthropods 35biomimicry 83convergence 145, 148fishes 246, 249insects 320, 324mammals 366reptiles 457, 458spiders 497
eyespots 24
fFabaceae 145, 186, 394Fagaceae 73, 74, 185Fagus 73Fagus crenata 516Fagus grandifolia 185–186, 504, 513Falconiformes 89falcons 89Falkowski, Paul 42family (taxonomic) 78Famitsyn, andrey S. 508fan palms 179, 195FaO. See United nations Food and
agriculture Organizationfarming 10–11fat accumulation 482feathers 5, 84–86, 85, 86, 193, 480federal government role in conservation
(United States) 238–244, 240t–241tfederal wildlife refuges 548cfeedlot runoff 422, 423Feeny, Paul 303feet 16Felis catus 12fen 91feral animals 12, 501feral livestock 330feral pigs 12, 333Fergusobia curriei 27fermentation 262ferns 475, 476
carboniferous period, biodiversity in 105
deserts, warm 177Permian period, biodiversity in 412seedless plants 472, 473, 475–476succession, ecological 505triassic period, biodiversity in 524
fertile hybrids 310fertility rates 425–427fertilization
agriculture 13amphibians 18animals 21, 26, 27arthropods 35birds 87fishes 246, 249flowering plants 252, 253reptiles 455seedless plants 472–476sexual selection 479, 484speciation 491, 494
fertilizer runoff 466Ficus 186
fiddleheads 475Field, christopher 291figs 52, 186filaments 41filarial worms 31filmy ferns 476filter feeders 22, 28, 171finches 8, 167–169, 271–274, 335Fincke, Ola 481Finland 304fins 15, 16, 246–249fir(s) 2, 92, 94, 137, 511, 518fire ants 325, 328, 331, 332, 545fire cycle 128, 201tfirefly beetle 99fire poppy 487fires
grasslands and savannas 274, 276greenhouse effect 286, 287restoration ecology 461shrublands 486, 487succession, ecological 500–502temperate deciduous forests 516
fire suppression 200, 201, 462firewood 289Fischbach, Michael 67, 68fish(es) 244–251, 245t, 246, 248–250
adaptive radiation 6–7amphibians 15, 16, 18animals 30biomagnification 81cambrian period, biodiversity in
96–97camouflage and mimicry 99carboniferous period, biodiversity
in 105cladistics 113coevolution 122convergence 146, 147cretaceous period, biodiversity in 154deep oceans, biodiversity in 170, 171Devonian period, biodiversity in
182–184disjunct species 196endemic species 222greenhouse effect and biodiversity 297hybridization 308invasive species 331Jurassic period, biodiversity in 341keystone species 345life history, diversity of 350, 353living fossils 356mutualism 380, 381neogene period, biodiversity in 393Ordovician period 397Paleogene period, biodiversity in 399Permian extinction 408Permian period, biodiversity in 412restoration ecology 462rivers, streams, ponds, and lakes 467sexual selection 481Silurian period, biodiversity in 488succession, ecological 501
Fish and Wildlife Service, U.S. (FWS) 219, 220, 241
fish breeding grounds 372Fisher, R. a. 430, 431, 434, 479fishing 150fishing spiders 467, 497fishopod 16, 16fitness indicators 479–480, 482–483FitzRoy, Robert 54, 162, 163flagella 22, 32, 40, 261, 448, 509flatfishes 249flatworms 24, 29, 467fleas 38, 323, 403, 404fleshy-finned fishes 15, 247, 249flies 321
arthropods 38cretaceous period, biodiversity in 155insects 323, 324
xviii+598_Biodiversity_F.indd 579 11/17/11 1:13 PM

first entry580 tundra 580 first entry580 Index580
Permian period, biodiversity in 412pollination 420sexual selection 481speciation 492
flight 85, 146–147, 193, 320flight feathers 86, 86tflightless birds 87, 173flight muscles 86Flint’s Pond 521flippers 365flood control 242floodplain habitats 200flood prevention 215, 216floods 465, 466flora 57Flores island people 75, 387florets 188Florida 75, 137, 222, 369, 389, 493Florida Keys 546flounder 249flower(s) 188, 252, 252, 418, 419, 538flowering plants (angiosperms) 252, 252–
256, 253, 255adaptation 6biodiversity, patterns of 51, 52biodiversity, quantification of 56biogeography 74t, 75biological classification 80camouflage and mimicry 99carnivorous plants 106–107cladistics 109, 110, 113t, 114coevolution 119–121convergence 144cretaceous period, biodiversity in 154deserts, polar 174deserts, warm 177dicots 184–189dinosaurs 193endemic species 221greenhouse effect and biodiversity 298living fossils 356marshes and swamps 371monocots 375–377parasitism 402–403pollination 417–420seed plants 478speciation 494
flukes 24–25, 29–30Flustra 162flying fishes 248flying foxes 366flying mammals 365–366fog 177, 518fogslimbs 457–458foliose lichens 348follicle mite 36food chain 71, 208t
bacteria 42biodiversity, patterns of 48, 50biogeochemical cycles 72biomagnification 80–82cretaceous extinction 151, 152deep oceans, biodiversity in 171–172ecology 205, 206ecosystems 212invasive species 333keystone species 343predation 439
food chain disruption 298food energy 210food processing 42food production x, 10, 14, 381, 390–392food resource specialization 493–494food web 101, 170, 205, 209, 466foraging 51foraminiferans 448Forbes, andrew 49, 50forbs 127–128, 199, 274, 275t, 276Ford, Henry 389forest(s) 504. See also specific forests, e.g.:
temperate coniferous forests
acid rain and fog 1–2biogeography 73, 76boreal and subalpine forests 91–94carbon cycle 103cretaceous period, biodiversity in
154–155ecosystem services 213greenhouse effect 292Permian extinction 408succession, ecological 500
forest destruction 289–290Forest Reserve 548cForestry, Division of 548cForest Service, U.S. 239, 347, 413, 414forest succession 522fossa 527fossil fuels 14, 103, 210, 225, 227, 280,
289, 291, 293t, 501. See also coal; petro-leum
fossil record 148, 408fossils and fossilization 42, 163, 190, 191,
217–218, 256–259, 257founder effect 4, 75founder effect and genetic drift 259–260foxes 330foxfire 370foxtail pines 137Fraenkel, Gottfried 116Fragaria 186France 60Frankia 394Frankia bacteria 42Franklinia alatamaha 233Franz Josef land 174Fraser firs 94frequency-dependent selection 387freshwater fish 196, 341, 452freshwater marshes 369freshwater mussels 467freshwater pools 437freshwater swamps 369–370fringing reefs 149Frizte, Jessica 229frogs x, 18
adaptation 6amphibians 15, 17, 18biodiversity, resources for medicine
66–69cretaceous period, biodiversity in 155deserts, warm 180predation 436sexual selection 481triassic period, biodiversity in 524
fronds 475, 475, 476frugivorous animals 133fruit
biodiversity, resources for agriculture 61
coevolution 118–119competition, ecological 133dicots 186, 187flowering plants 252herbivory 305seed dispersal 470–471
fruit fliesbiodiversity, resources for medicine
68founder effect and genetic drift 258insects 324life history, diversity of 353population genetics 430sexual selection 479speciation 490, 493, 494
fruit toxins 117–118Frutafossor 341fruticose lichens 348Fucus 447Fuegian people 163Fugitive Slave act 521Fukashi, tadami 134Fulton, George 369
fungi 259t, 260–265, 260t, 261, 262, 262t, 263t
agriculture 8–9biodiversity, resources for agriculture 62biodiversity, resources for medicine
64–67biological classification 78, 79camouflage and mimicry 98coevolution 122competition, ecological 133decomposers 170Dna studies of biodiversity 202eukaryotes 232extinction 234greenhouse effect and biodiversity 297keystone species 344–345lichens 347–349mutualism 379–381parasitism 402predation 438temperate deciduous forests 516
fungus beetle 324funnel-web spiders 498Fusarium fungus 297Fusarium oxysporum 62Future of Life, The (Wilson) 545, 546FWS. See Fish and Wildlife Service, U.S.fynbos 486
GGadsden Purchase 238Gadus morhua 172Gaia 223, 266–271galagos 441Galápagos islands 271–273, 272t
adaptive radiation 8agriculture 12biogeography 76competition, ecological 134Darwin, charles 163Darwin’s finches 167–169endemic species 221Grant, Peter and Rosemary 273–274island biogeography 335speciation 490, 491, 493
Galápagos tortoise 379, 455, 456gall bladder 66Galliformes 89Gallornis 156Galloway, laura 3galls 403Gallus gallus 89gall wasps 389Galton, Francis 544Gambusia 331gametes 20gametophytes 447, 472–477gamma diversity 126, 128–129Gandhi, Mohandas 521Garden of eden 224garrigue 486gars 247, 250garter snakes 438Gasterosteus aculeatus 481gastric brooding frogs 68–69gastroliths 86, 190, 257, 455gastropods 25, 26Gastrotricha 28Gaviiformes 89gazelles 173Gecarcoidea natalis 156, 157geckos 83, 457geese 89gene(s) 17, 69, 231, 353–354gene flow 489–490generalists 208General land Office 238, 239General Sherman tree 136Generelle Morphologie der Organismen
(Haeckel) 548c
xviii+598_Biodiversity_F.indd 580 11/17/11 1:13 PM

tundra 581Index 581
genetically modified organisms (GMO) 12–14
genetic bottleneck 4, 234, 258–259genetic diversity 233, 258genetic drift 4, 75. See also founder effect
and genetic driftgenetic engineering 59genetic extinction 59–60genetic pollution 12genetics 430–435genetic variability 3–5, 301, 353, 388,
430–435gene transfer 310–311gene variation 203genotypes 430Genovesa 169, 273genus (pl. genera) 78geographical isolation 301, 489, 490geological deposits 407geometric series 55Georgia 137, 194, 195, 221, 222, 233Geospiza conirostris 169Geospiza fortis 168, 169Geospiza magnirostris 169Geranium viscosissiumum 418Germany 2, 257germ cells 353germline cells 384Gerridae 467gestation period 351–352GFP (green fluorescent protein) gene 69gharials 456GHGs. See greenhouse gasesghost shark 247giant armadillo 367giant-flowered phacelia 487giant golden-crowned flying fox 366giant moray eel 248giant redwood 222giant sequoia 136, 354, 502, 511giant tortoises 221, 455, 456Giardia lamblia 445giardiasis 445Gibberella zeae 263gibbons 537Gibson, J. Phil 194gila monsters 457Gill, Jacquelyn 416gills 16, 17, 34–35, 467gill slits 29, 245, 247, 250Ginkgo biloba 222, 233, 478ginkgoes 222, 233, 341, 356, 478giraffes 5, 5gizzards 86glacial retreat 316glaciations 49, 95–96, 411glaciers 76, 201, 282–286, 289, 315–316,
450, 452glands 5glass snakes 457glaucoma 67Gleason, Henry 125Gleditsia aquatica 119Gleditsia triacanthos 118, 119, 186global climate change. See global warmingglobal conveyor current 284global temperature 279–284, 280t, 282t,
311, 398global warming x
amphibians 18, 19boreal and subalpine forests 94coral reefs 150deserts, polar 174ecosystems 211ecosystem services 216environmental ethics 224extinction 234, 235Gaia 269, 270greenhouse effect 279–280, 282,
285–287, 290
greenhouse effect and biodiversity 296–298
herbivory 305mass extinctions 373–375Permian extinction 408–410Permian period, biodiversity in 411restoration ecology 462, 464tropical rain forests and deforestation
534tundra 540
Glomeromycota 263Glossopteris 412, 524glue 66glutin 310glyphosate 460glyptodonts 450GMO. See genetically modified organismsGnathostomulida 28Gnetales 253, 478goats 12, 273goatsbeard 492Gobi Desert 173gobies 249goblet worms 27God 224, 225golden eagle 219golden rain tree 389goldenrod 431golden toad 18, 116, 235, 297gold extraction 533goldfish 248Goliath birdeater 498goliath frog 17Gondwana/Gondwanaland
carboniferous period, biodiversity in 105
cretaceous period, biodiversity in 154Devonian period, biodiversity in 182ice ages 315Jurassic period, biodiversity in 339Ordovician period 397triassic period, biodiversity in 523
Goodall, Jane 443Goosen, Hendrick 249Gore, al 358gorillas 223, 235, 443, 444, 481Gould, John 167Gould, Stephen Jay 33, 42, 148, 546Graham, linda 488grains 10, 60–62, 209grama grasses 198gram-negative bacteria 40gram-positive bacteria 40Gram’s stain 40Grand canyon 344grandmother hypothesis 354Grant, Peter and Rosemary 168–169, 271,
273–274, 432, 493, 494Grant, Robert 161grapes 60, 187grasses 129
agriculture 12biodiversity, resources for agriculture 61biogeography 74coevolution 117, 122communities, ecological 127–128convergence 144deserts, cool 173deserts, warm 180disturbances, ecological 198grasslands and savannas 277tgreenhouse effect and biodiversity 296hybridization 310invasive species 332–333monocots 376–377neogene period, biodiversity in 393succession, ecological 504
grasshoppers 13, 38, 181, 321grasslands and savannas 199, 274, 274–
279, 275t, 276, 277t–278t, 504biodiversity, patterns of 49
biogeography 76carbon cycle 103deserts, cool 173dicots 186disturbances, ecological 198–199greenhouse effect 292keystone species 343monocots 377restoration ecology 461temperate dry forests 517
grass lizards 457grass stems 303Gray, asa 166grazing animals 276, 278–279, 343, 360,
363, 377, 538great ages 442–443Great Basin Desert 173, 511Great Basin system 465Great Britain 76, 171, 217, 258, 296, 298,
486, 487Great chain of Being 78, 356Great lakes 316, 331Great lakes system 465Great Outback 175Great Oxidation event 95Great Pacific Garbage Patch. See Pacific
trash VortexGreat Plains 76, 241, 279Great Smoky Mountains 55, 56t, 313great white sharks 247, 249green algae 109, 110, 348, 448, 472Green Belt Movement 358–359greenhouse effect 279–295, 280t, 281,
282t, 283, 284t, 288t, 292t, 293t, 549cgreenhouse effect and biodiversity 295–
300biogeography 76carbon cycle 102, 103cloud forests 116ecology 205environmental psychology 229Gaia 268–270ice ages 318restoration ecology 462succession, ecological 501tropical rain forests and deforestation
534greenhouse gases (GHGs) 14, 205, 286–
290, 311, 395, 409–410. See also carbon dioxide, atmospheric; methane
Greenland 174Greenland ice sheet 283–285green landscaping 228, 229green protists 448green spider orchid 99green sulfur bacteria 43, 44Grime, Philip 350grizzly bear 311grolar bear 311Grosberg, Rick 52Gros Michel banana 62gross primary production 210mground finches 168, 169ground pink (wildflower) 487groundwater 312–313groundwater recharge 215growth patterns 354grubs 389guanacos 173guars 220guevedoche 435Gulf coastal system 214, 465Gulf of Mexico 142–143, 172Gulf of Mexico oil spill (2010). See BP
Deepwater Horizon oil spillGulf Stream 171, 284, 285, 311, 316gulls 89gumbo limbo tree 369, 515guppies 389–390, 437Gymnocladus dioicus 516Gymnophthalmidae 458
xviii+598_Biodiversity_F.indd 581 11/17/11 1:13 PM

first entry582 tundra 582 first entry582 Index582
gymnosperms 135, 154–155, 180, 253, 341, 477–478
Gymnothorax javanicus 248gypsy moth 324Gyrinidae 467
hhabitat
animals 20biodiversity, patterns of 52bogs, life in 90–91disturbances, ecological 200, 201endemic species 222population biology 426rivers, streams, ponds, and lakes 466succession, ecological 499
habitat conservation 139, 251, 372habitat creation 344habitat destruction x, 12–13, 60, 103, 234,
374, 463habitat disruption 139habitat expansion 297habitat fragmentation 12–13, 301–302Habronestes bradleyi 498Habropoda pallida 98–99Hackett, Shannon J. 87Hadley cells 176Hadronyche formidabilis 498hadrosaurs 155Haeckel, ernst 548chagfish 69, 246Haikouichthys 97, 245hair(s) 303, 359–360hairgrass 174Haldane, J. B. S. 324, 434halibut 249Hallucigenia 97hallucinogens 264halophiles 33halteres 147, 323Hamilton, William 480hammerhead sharks 246handicap principle 480hands 16Hansen, James V. 220, 285, 549chaplodiploidy 325haploid cells 252, 260, 325, 472, 477Haplorrhini primates 366haptophytes 447Hardin, Garrett 429hard skeletons 95, 96Hardy, G. H. 433Hardy-Weinberg equilibrium 433–435, 433tHarmonia axyridis 331Hartnett, David 264harvestmen 35Hasselquist, Frederik 355hatchetfishes 248Hautier, yann 49Hawaiian goose 234, 258Hawaiian islands
adaptive radiation 7–8agriculture 12biogeography 76continental shelves, biodiversity on 142endemic species 221invasive species 332–333speciation 490, 492, 493succession, ecological 505
Hawkes, Kristin 354Hawkins, Bradford 48hawkmoth 324hawthorns 119Hay, Mark 381Hayden, Ferdinand V. 239hazel alder 194, 308hazelnuts 78heading 11–12heads 24, 37, 456, 457health 11, 58, 229, 378–379, 460, 482
heartbeat 86hearts 360, 455heath bushes 91heather 487heather shrub 196heath plants 110Heavenly city 225heavy metals 72, 84, 224, 502hedgerows 13heirloom varieties 59Helianthus 492Helianthus annuus 310Helianthus anomalus 310Helianthus deserticola 310Helianthus maximiliani 61Helianthus paradoxa 494Helianthus petiolaris 310heliconid butterflies 45Heliconius 117Helicoprion 412Heloderma suspectum 457Hemachatus haemachatus 457Hemiaulus haukii 381hemichordates 29hemimetabolous development 321hemiparasitic figs 52Hemiptera 322–323hemlocks 128, 137, 504, 511, 516Hemochordata 29hemocyanin 26hemoglobin 15hemolymph 320Hemphillian extinction 374Hennig, Willi 109Henslow, John Stevens 162herbaceous plants 254–255, 296, 375–376,
403herbicides 13–14, 18, 423, 459–460herbivory 302–307, 304t
adaptation 5agriculture 12coevolution 117, 122communities, ecological 126, 130competition, ecological 132dinosaurs 191ecology 205ecosystems 208–209Janzen, Daniel H. 338Jurassic period, biodiversity in 341keystone species 343, 344mammals 365marshes and swamps 370triassic period, biodiversity in 524
herbs 187herds 436–437heritability 431heritable diversity 424hermaphroditism 18, 252hermit crabs 156Herrerasaurus 191herrings 248Hesperornis 156Hetch Hetchy reservoir 378heterocysts 394heterokonts 446–447heterotherms 17, 250, 455heterozygous organisms 258, 430, 432, 434Hevea brasiliensis 188Heyburn, Weldon 414hibernation 206Hibiscus liliflorus 235Hibiscus moscheutos 484hickories 186, 515high-elevation tropical forests 115–116high-input system 13Hill, Julia Butterfly 520Hillis, Daniel 114Himalayas 89, 398Hines, Bob 108Hipparion 393Hippocampus 249
Hippocampus bargibanti 250hirudin 66Histoire Naturelle (leclerc) 548chistones 32, 230, 231Historia Naturalis (Pliny the elder) 548cHistoria Plantarum (theophrastus) 548chistrionicotoxin 17HiV/aiDS 67, 68, 114, 262Hogberg, Peter 264Hohokam culture 461Hölldobler, Bert 545holometabolous development 321homeotherms 86, 147, 250, 360, 367Homestead act 238Hominidae 443hominins 443–444Homo 318, 444Homo caudatus 356Homo erectus 75Homo ergaster 438Homo ferus 356Homo floresiensis 75, 387Homo habilis 114, 438Homo neanderthalensis 318homoplasy 110, 111Homoptera 38, 323Homo sapiens 58, 227, 318, 351–352, 444homozygous organisms 430, 432, 434–435honeybees 38, 215, 326, 332, 421honeydew 323honey locusts 118, 119, 186hooded tickspiders 36Hoodia gordonii 182Hooker, Sir Joseph 165Hoplostethus atlanticus 172hopping-mouse 181horizontal gene transfer 40, 231hormonal conflict 352hormones 231horns 470hornworms 324hornworts 474horse(s) 12, 56, 308, 393, 400, 452horse apples 119horsehair worms 28horseshoe crabs 37, 46, 67, 356, 356horseshoe worms 27horsetails 105, 472–475horticultural therapy 228host density 327hosts 403hot spots. See biodiversity hot spotsHottonia inflata 125house fly 324hox genes 17Hubbard Brook, new Hampshire 197, 312Hubbell, Steven 52, 129human activities x
biogeochemical cycles 72biogeography 76–77boreal and subalpine forests 94carbon cycle 101, 103ecosystems 211, 212environmental psychology 228extinction 235Gaia 269greenhouse effect 289–290habitat fragmentation 301mass extinctions 374–375Permian extinction 410, 417succession, ecological 501–502, 506
human body 42–43human brain 110, 483, 545human civilization 298human evolution 166, 482–483human flea 38human population 425–426Humboldt, alexander von 47, 50, 307, 548cHumboldt current 307humidity 280hummingbirds 86, 89, 120, 418
xviii+598_Biodiversity_F.indd 582 11/17/11 1:13 PM

tundra 583Index 583
hunter-gatherers 9, 10, 57–58hunting 220, 234, 414, 415, 417, 438–439,
483hurricanes 286Hut, Piet 373Hutchinson, G. evelyn 50–51, 132, 549cHuxley, Julian 18Huxley, thomas Henry 165, 166Hyalymenus tarsatus 98hybridization 12, 61–62, 169, 307–311,
308t–309t, 391, 491hybrid vigor 307hydrogenosome 509hydrogen sulfide 42, 269hydroid cells 474hydrological cycle 212, 311–314, 312, 313thydrostatic skeleton 147hydrothermal vents 172, 549chyenas 132–133Hylonomus 106Hymenophyllaceae 476Hymenoptera 38, 325–326, 524hyomandibular bone 16hypertonic 250hyphae 260–264hypotonic 250hypsithermal 317hyraxes 367Hyrocotherium 400
Iice 96, 175, 218, 282, 285, 312ice ages 315–318, 317m
biodiversity, patterns of 49biogeography 76–77boreal and subalpine forests 92disjunct species 195, 196disturbances, ecological 197, 201ediacaran organisms 218fossils and fossilization 258Gaia 270grasslands and savannas 274, 278greenhouse effect 282, 289Quaternary period, biodiversity in
450, 452tundra 537
ice cores 152, 282–284, 283, 284tIcerya purchasi 328ice sheets 268ice shelf 283Ichthyornis 156ichthyosaurs 154, 524Ichthyostega 17Ichthyostega stensioei 16idaho 256iguanas 272Iguanodon 341illinois 128illness 164iltis, Hugh 234immune system 69, 362, 405Impatiens 470inbreeding 164, 434–435incense cedar 137incipient speciation 169incus 361india
biogeography 73cretaceous period, biodiversity in 154deserts, warm 175iPM 328Paleogene period, biodiversity in 398reptiles 456tropical rain forests and deforestation
527tropical seasonal forests 537
indiana Dunes national lakeshore 504indian cobra 457indian Ocean 249indian pipe 403
indigobirds 491indochina 537indonesia
biodiversity, patterns of 52biogeography 75fishes 249iPM 327reptiles 457tropical rain forests and deforestation
527, 534Wallace, alfred Russel 542, 543
industrial Revolution 280indy gene 353infant mortality 426, 427infections 68, 262infectious disease 428infrared photons 205, 206infrared radiation 211, 287inheritance 369, 386, 548cinhofe, James 226In Memoriam (tennyson) 386inouye, isao 510insect eggs 21insecticides 59insectivores 366, 367insects 318–327, 319t, 321, 322, 325
adaptation 6agriculture 13animals 27arthropods 35, 37–38Bates, Henry Walter 45–46biodiversity, patterns of 52biodiversity, quantification of 54camouflage and mimicry 98, 100carboniferous period, biodiversity
in 105coevolution 116, 117, 120–121convergence 145–147cretaceous period, biodiversity in 155Dna studies of biodiversity 203flowering plants 253fossils and fossilization 256herbivory 302–303Jurassic period, biodiversity in 341marshes and swamps 370natural selection 389parasitism 403Permian extinction 407pollination 419population biology 427predation 436resistance, evolution of 459, 460rivers, streams, ponds, and lakes 466,
467sexual selection 480succession, ecological 500triassic period, biodiversity in 524Wilson, edward O. 545, 546
Insect Societies, The (Wilson) 546insect vectors 405instars 37, 321insurance companies 214integrated pest management (iPM) 327–
329integrated taxonomic information System
57interference competition 132, 133intergovernmental Panel on climate change
(iPcc) 281, 282, 287, 290interior, Department of the 238, 239, 241,
242, 244internal fertilization
amphibians 18animals 21, 26, 27arthropods 35birds 87fishes 246reptiles 455
internal membranes 230international Society for Phylogenetic
nomenclature 80
international Union for the conservation of nature (iUcn) 218, 235
interspecific competition 132intertidal zones 133, 142intestinal bacteria 378–379intestines 5, 32, 43intraspecific competition 132, 428introgression 311invasive species 329–335, 330, 331t–332t,
333agriculture 12amphibians 18herbivory 305iPM 328marshes and swamps 372restoration ecology 462, 463temperate dry forests 518
invertebratesanimals 20, 29–31carboniferous period, biodiversity
in 105continental shelves, biodiversity on
142convergence 147cretaceous period, biodiversity in 154,
155deep oceans, biodiversity in 170Devonian period, biodiversity in 182ediacaran organisms 217fossils and fossilization 256, 257Jurassic period, biodiversity in 341neogene period, biodiversity in 392,
393Ordovician period 396–397Paleogene period, biodiversity in
398–399Permian period, biodiversity in 411,
412population genetics 431Quaternary period, biodiversity in
450, 452Silurian period, biodiversity in 488triassic period, biodiversity in 523,
524iodine 71iPcc. See intergovernmental Panel on
climate changeiPM. See integrated pest managementireland 446iridaceae 376iridium anomaly 152, 374Iridomyrmex purpureus 498irises 376irish Potato Famine 501iron 71, 212irrigation 463, 464irwin, terry 54islam 224, 225island biogeography 75–76, 221, 335–337,
471, 490, 546isolating mechanisms 489–490, 492–494isopods 158isoprene compounds 101isoptera 38, 322isotonic balance 250isotope ratios 258, 407–408iteroparous species 350iUcn. See international Union for the
conservation of natureIxodes damminnii 36Ixodes pacificus 404Ixodes spinipalpis 404
JJablonski, David 148jack-in-the-pulpit 376jack pine 92jackrabbits 181Jackson, andrew 238Jackson, Wes 62, 83, 226, 338, 391
xviii+598_Biodiversity_F.indd 583 11/17/11 1:13 PM

first entry584 tundra 584 first entry584 Index584
Jackson, William Henry 239Jadera haematoloma 389Janzen, Daniel H. 52, 338–339Japan 52, 81–82, 137, 141, 263, 516Japanese beech 516Japanese black pines 511Japanese brome 278Japanese persimmons 60Java Man 75jawless fishes 96–97, 154, 183, 245–246,
397jaw worms 28jays 118–119Jefferson, thomas 307Jeffrey pines 511jellyfish 23, 23, 24Jenkin, Fleeming 165Jeon, Kwang 509Jesus christ 225jet propulsion 26jewel weeds 470jock itch 262Johnson grass 12, 61jojoba 61Jomon Sugi tree 137Joshua tree 175Judaism 224, 225Judson, Olivia 166Juglandaceae 186Juglans 113, 186Juglans nigra 425jumping spider 496, 497Juncaceae 377juniper shrubs 137Juniperus 137Juniperus ashei 200, 331Juniperus virginiana 200, 313, 331Jurassic period, biodiversity in 84, 85, 87,
193, 339–342, 340t, 341, 361–362, 454, 477
Justice Department, U.S. 142
kKaibab Plateau 344Kalahari Desert 175Kalm, Peter 355kangaroo 362, 363kangaroo mice 6kangaroo rats 181Kansas 61kapok tree 527Karoo Basin deposits 408–409Karpechenko, Georgi 310katydids 98, 99Kauai 7Keeling, charles 288Keiferia lycopersicella 328kelp forests 141, 142, 447Kennedy, edward 358Kentucky coffee trees 516Kenya 358–359Kepenomamus robustus 361Kessler, Danny 420Kettlewell, H. B. D. 168keystone species 343–346
biodiversity, concept of 46continental shelves, biodiversity on 141marshes and swamps 370Pleistocene extinction 417rivers, streams, ponds, and lakes 467temperate rain forests 520
Kimura, Mot[o] 434kinetic energy 206King, Martin luther, Jr. 521kingdom (taxonomic) 78–79kingpin principle 389king snakes 457King tut’s curse 264Kinorhynchia 28Kirschenmann, Fred 13
kiwis 87–89knockout mice 68Knoll, andrew 95knotweed 133knowledge 354Knowlton, nancy 150koalas 304–305, 363, 518Kok-aral Dam 464Komodo dragons 457Korarcheota 33Kosmos (Humboldt) 307Kozo-Polyansky, Boris M. 508krill 157–158, 170krummholz trees 92, 540Küchler, a. W. 461kudzu 333, 333Kump, lee 373Kuo, Frances e. 229kwolu-aatmwol 435Kyoto Protocol 549c
llabellum 376laboratory animals 27labrador 171Labroides dimidiatus 380lack, David 167, 271lacks, Henrietta 353lactation 360Lactobacillus 43, 379lactose digestion 148lactrodectus hasseltii 498Lactrodectus mactans 498ladybird beetle 331ladybug beetles 325Lagenidium callinectes 380lagerstätten 257lakes. See rivers, streams, ponds, and lakeslake sturgeon 356Lama guanicoe 173lamarck, Jean-Baptiste de 161, 548cLamarckia aurea 128lamarckian evolution 161, 162, 386Lamellibrachia luymesi 172lamiaceae 188, 487Laminaria 141lampreys 69, 246, 246, 250Lamprocyphus augustus 83lampshells 27, 374Lampsilis perovalis 467lampyridae 325lanai 7lancelets 29, 245land crab 156land degradation 461land ethic 226, 347land iguanas 272land institute, the 61, 62, 226, 338land Management, Bureau of 241land nationalization 544land rights 533, 534landscaping 228la niña 171, 280, 284larches 91, 137, 195Larix 137Larix decidua 91larsen B ice shelf 283Larus 494lassen Peak, california 540late Precambrian 95–96Lates niloticus 332late-successional habitats 499, 500latex 144Laticauda colubrine 98Latimeria chalumnae 249Latimeria menadoensis 249latin america 62, 548claurasia 103, 105, 339, 523laurentia 396lava 505
lava lizards 272lD50 (lethal dose, 50%) 306leaching 530lead 84leaf area 177leaf-cutter ants 8–9, 122, 264, 380leaf-cutter bee 421leaf insects 321–322leaf litter frog 98leaf structure 371leakey, Mary 258learning 158–159least tern 196leatherback sea turtle 455leaves
adaptation 3biodiversity, resources for medicine 66boreal and subalpine forests 92conifers 135–136convergence 144, 145deserts, warm 177dicots 185–187ecology 206flowering plants 254fossils and fossilization 256, 258herbivory 303, 304marshes and swamps 371monocots 376–377rivers, streams, ponds, and lakes 467temperate deciduous forests 513tropical seasonal forests 537
leclerc, Georges-louis. See Buffon, Georges-louis, comte de
leeches 66leg bones 15, 17legumes 13, 42, 61–63, 145, 186, 394leks 480lemnaceae 467lemurs 441leonardo da Vinci 256leopold, aldo 224, 226, 344, 347, 549cLepidodendron 105, 473, 474lepidoptera 37–38, 324, 341. See also but-
terflies; mothslepidosaurs 87, 454lepirudin 66Lepomis cyanellis 308Lepomis macrochirus 308Leptinotarsa decemlineata 325Leptotyphlops 457Lepus americanus 438Lepus californicus 181lesser apes 442lethal recessive allele 434, 435Leucaena leucocephala 63leucaena tree 63Leucochrysum fitzgibbonii 177leukemia 66, 353lewis, Martin 227lewontin, Richard 546lianas 513, 526lice 38, 322, 403lichens 84, 262, 264, 347–349, 348, 381life history, diversity of 349–355, 350t, 352tLife magazine 338likens, Gene 2, 312lilacs 296liliaceae 375–376lilies 375, 420limber pine 92, 196lime sediments 448limestone 1, 102, 257limnetic zone 468limonene 144, 148Limulus polyphemus 37, 46, 67Linanthus dianthiflorus 487Linanthus montanus 253lindeman, Ray 208, 212linden 506Lingula 27, 407, 408Linnaea borealis 356
xviii+598_Biodiversity_F.indd 584 11/17/11 1:13 PM

tundra 585Index 585
linnaean system of classification 77–80, 113–114, 232
linnaeus, carolus 77, 355–356, 548clions 112, 132, 133, 308Lithops 98little ice age 281littoral zone 468live birth 147, 246, 359, 455live oak 6, 185, 309, 369, 513liver 80, 81, 250liver flukes 29–30liverworts 473Lives of a Cell (thomas) 266livestock 12, 13, 63, 210, 381, 501livestock feed 13, 209, 210living fossils 356, 356–357, 357tlizard-hipped dinosaurs 191lizards 83, 106, 272, 341, 389, 454–458Lobatae 73lobe-finned fishes 105, 183–184Lobocraspis griseifusa 324lobsters 157, 341locoweeds 84, 186, 303locus 430locusts (insect) 13, 38locust trees 118, 119, 186lodgepole pines 200, 511löfling, Pehr 355logging 94, 239, 520, 533log-normal distribution 55Loiseleuria procumbens 174lompoc, california 258longfellow, Henry Wadsworth 201long-horned beetle 324long-term carbon cycle 101–103long-term mineral cycle 70, 71, 212Lontra canadensis 370loosestrife 333Lophelia pertusa 149lophophores 27loricifera 21, 28lorises 441losos, Jonathan 389louisiana Purchase 238lovelock, James 223, 266lower jaw 361Loxosceles reclusa 498lSD (lysergic acid diethylamide) 264lubchenco, Jane 72luna moths 325lungfishes 105, 183–184, 247, 249, 341, 412lungs 16, 17, 86, 250, 455, 456Lycopersicon esculentum 188lycopodiophyta 474lycosidae 498lyell, Sir charles 163, 165, 542–543Lygodium 476Lymantria dispar 324lyme disease 40, 404lynx 438Lynx canadensis 438lysates 67Lyssopimpla excelsa 420Lystrosaurus 409, 524, 524Lythrum salicaria 330
mMaathai, Wangari 358–359, 359Macaca sylvanus 518macaque 518Macaranga 380Macarthur, Robert H. 51, 335, 546Macarthur distribution 55MacDonald, Julie 220Maclura pomifera 118, 186, 527Macrochelys temminckii 370Macrocystis 141, 447Macropus rufus 363Madagascar 66, 87, 221, 222, 457, 527mad cow disease 210
Madeira 313maggots 38, 170Magnaporthe grisea 263magnesium 70, 72Magnolia grandiflora 515magpie robin 234maidenhair ferns 475maidenhair tree 478maize 9, 11, 12, 58, 60, 234, 381, 484Malaclemys terrapon 455Malacostraca 157–158Malagasy people 457malaria 67, 305, 324Malawi, lake 6Malay Archipelago, The (Wallace) 542Malaysia 456, 527mallee 486malleus 361Mallophaga 322Malthus, thomas 164, 165, 351, 425, 548cMalus 186mammals 79, 359–368, 362, 364t, 365–
367, 393, 401. See also specific mammals, e.g.: bison
adaptation 3adaptive radiation 8biodiversity, patterns of 48, 49biogeography 74–76biological classification 78boreal and subalpine forests 93tcladistics 110convergence 145, 146t, 147cretaceous extinction 151cretaceous period, biodiversity in 156deserts, warm 181flowering plants 253grasslands and savannas 276, 278–279greenhouse effect and biodiversity 297Jurassic period, biodiversity in 341keystone species 343life history, diversity of 350, 353mass extinctions 374neogene period, biodiversity in 393Paleogene period, biodiversity in 400Pleistocene extinction 414–417, 415t,
416tpond species 468tQuaternary period, biodiversity in 452reptiles 454rivers, streams, ponds, and lakes 466seed dispersal 470–471sexual selection 480temperate deciduous forests 516triassic period, biodiversity in 524tropical rain forests and deforestation
529t–530ttropical seasonal forests 537
mammary glands 5mammoths 75, 76, 343, 417managed ecosystems 463managed forest 463Man and Nature (Marsh) 548cmanatees 365mandibles 34, 320, 322–323Manduca sexta 306, 324mangrove islands 336mangrove swamps 370mangrove trees 371manifest destiny 238Manihot esculentum 188manioc 188maniraptorians 193Mann, Michael 281, 282Mantgem, Phillip van 296mantids 98, 322Mantis religiosa 481mantis shrimp 158manzanitas bush 487maples 73, 187, 462, 499–500, 504, 513,
515, 516maquia 486
Marchantiophyta 473Margulis, lynn 123, 231, 232, 266, 270,
368–369, 507–510Marianas trench 171marine ecosystems 209marine habitats 49marine life
animals 29cambrian period, biodiversity in
96–97carboniferous period, biodiversity
in 105cretaceous period, biodiversity in 154Devonian period, biodiversity in 182ediacaran organisms 217–218Jurassic period, biodiversity in 341neogene period, biodiversity in 392Ordovician period 396Paleogene period, biodiversity in 398Permian period, biodiversity in 411Quaternary period, biodiversity in
450, 452Silurian period, biodiversity in 488triassic period, biodiversity in 523–
524marine preserves 142marine reptiles 151Marsh, George Perkins 548cMarsh, Othniel charles 190Marshall, Diane 484Marshall, John 238marshes and swamps 274, 369–372, 370,
371tMarsilea drummondii 177marsupials 393
biogeography 74–75convergence 146, 146t, 147life history, diversity of 352–353mammals 359, 360, 362, 363, 367–368temperate dry forests 518tropical rain forests and deforestation
527Martin, Paul 119Martin, Robert a. 16Mason, John 217Massachusetts 520–522mass extinctions 372–375, 373t
cretaceous extinction 151–152Devonian period, biodiversity in 184extinction 233, 235Ordovician period 397Permian extinction 407–410Pleistocene extinction 414–417Wilson, edward O. 546
Mastigias papua 23mastodons 76, 343, 417mast seeding 118Mathew, Patrick 384mating dances 493mating disruption 328matorral shrublands 486, 517matrix 256matter, cycling of 207, 211–213Maui 7Mauritius 234Mawsonites 217maxillopods 158Maximilian sunflower 61May, Robert M. 52Mayan culture 461, 501mayflies 321Mayr, ernst 384–385McKenney, Daniel W. 296McKinley, William 239McMenamin, Mark 217–218McMurdo dry valleys 174Mcnair, M. 484meadows 130meat 210Medawar, Peter 353Medea Hypothesis 223, 268
xviii+598_Biodiversity_F.indd 585 11/17/11 1:13 PM

first entry586 tundra 586 first entry586 Index586
medical research 68, 444medicine x, 38, 58, 114, 215, 326. See also
biodiversity, resources for medicine; phar-maceuticals
Medieval Warm Period 281Mediterranean Basin 313, 486–487, 517, 518Mediterranean Sea 76, 196medusa stage 22meerkats 159, 159Megachile rotundata 421Meganeura 105, 105Meganeura monyi 320meiosis 20, 260, 261, 472, 477Melaleuca quinquenervia 330, 372melaleuca trees 330, 333, 372Melanerpes formicivorus 90Meloe fransiscanus 98–99Mendel, Gregor 386, 548cmenopause 354mental health 230mercury 72, 80–82, 206, 423, 533Mereschkovsky, Konstantin S. 508Mesoamerican Biological corridor 302Mesohippus 400mesophication 201Mesopotamia 9, 11Mesozoic era
adaptive radiation 6, 8cretaceous period, biodiversity in
153–156dinosaurs 190–193ice ages 316Jurassic period, biodiversity in 339–341mammals 361, 362, 367mass extinctions 372reptiles 454seed plants 477–478
mesquite 175Mestral, George de 83metagenomics 203metamorphic rocks 256metamorphosis 15, 37, 321, 324Metasequoia glyptostroboides 51, 136–137,
222methane 14, 101–103, 205, 287, 370,
409–410Methanobrevibacter smithii 32Methanococcus janaschii 33methanogens 33Methodus Plantarum Novae (Ray) 548cmethyl mercury 72Metrosideros polymorpha 505, 507Mexican piñon 518Mexican poppies 177, 497Mexico
agriculture 12biodiversity, resources for agriculture
60, 62cloud forests 115deserts, cool 173deserts, warm 175endemic species 222pollution 422shrublands 487temperate coniferous forests 511temperate dry forests 517, 518tropical seasonal forests 537
mice 6, 68, 350Michigan 262Michigan, lake 128, 504microaerophiles 379microclimates 52, 90–91Micrognathozoa 21, 28microhabitats
biodiversity, patterns of 51communities, ecological 127, 128rivers, streams, ponds, and lakes 466temperate coniferous forests 511temperate deciduous forests 513tropical rain forests and deforestation
526
micronutrients 71–72microorganisms 47, 64–66, 171Microsporidia 261microwhip scorpions 36Middle east 9–10, 12, 36, 60, 175, 179,
180, 517mid-domain effect 48migration
birds 89boreal and subalpine forests 92fishes 248greenhouse effect and biodiversity
295–297habitat fragmentation 301–302herbivory 305predation 437
Migratory Bird treaty 243Milanković, Milutin 315Milanković cycles 270, 316, 450mildew 262military bases 241–242milk 360, 362milkweeds 117, 133–134, 180, 188, 303Miller, Geoffrey 483Miller-Rushing, a. J. 296, 522millipedes 27, 35, 105, 436Mimetica viridifolia 98mimicry 45, 98, 321–322, 420, 437–438Mimulus 490, 492, 494Mimulus bigelovii 177, 177Mimulus cardinalis 492Mimulus dudleyi 128Mimulus guttatus 484Mimulus lewisii 492Mimus longicaudatus 272mineral cycle 71mineral deposits 41–42, 256mineralization 256minerals 70–72, 206, 212Minerals Management Service (Department
of the interior) 225, 244mining 422Minnesota 208, 209minnows 248mint 59, 188, 487mirex 328Miriophyllum aquaticum 467missing-link fish-amphibians 15missing links 361Mississippian culture 461Mississippi River basin 465Missouri River 316mistletoe 403Mitchell, Mount 2mites 35, 36, 123–124, 126–127, 184,
460mitigation banking 139mitochondria 42, 122, 230, 231, 368,
508–510mitosis 261Mivart, St. George Jackson 165mixed-grass prairies 276Mnemiopsis leidyi 332moas 87, 119Mobulidae 171mockingbirds 272Moi, Daniel arap 358, 359Mojave Desert 175, 181, 301Mola mola 247molds 446, 448, 449mole(s) 363, 367molecules 69–72mole rats 145–146mollusks 25
animals 25–26cretaceous period, biodiversity in 154Devonian period, biodiversity in 182linnaeus, carolus 355mass extinctions 374Ordovician period 396–397Paleogene period, biodiversity in 399
Permian extinction 410Silurian period, biodiversity in 488triassic period, biodiversity in 523
Molokai 7molts 37, 321monarch butterfly 117, 305, 500moncots 255Mongolian steppe 173mongoose 527monkeyflowers 177, 484, 490, 492monkey puzzle tree 136monkeys 112, 441–442, 537monocots 113t, 114, 254, 375–377, 376,
467, 478monoculture 13, 62, 391monotreme mammal 110, 361–363, 527Monotropa 144, 403Monterey canyon 172Monterey cypress 318Monterey pine 138, 308, 330, 518Montin, lars 355moorlands 486Moraceae 186moraines 316Moran, thomas 239moray eels 149, 248Morgan, thomas Hunt 430Morganucodon 361morning glories 110morphology 40mortality 426Morus 186Morus alba 60mosquitoes
arthropods 38biomagnification 81greenhouse effect and biodiversity 298insects 323–324invasive species 330, 331iPM 328parasitism 403, 404resistance, evolution of 460rivers, streams, ponds, and lakes 466speciation 491tundra 538
mosquito fish 331moss animals 27mosses 91, 381, 473–474, 474, 540moth caterpillars 324moths 325
arthropods 37–38biodiversity, resources for agriculture
62biomimicry 83coevolution 120, 121Darwin’s finches 168Dna studies of biodiversity 203pollination 418
Mound Builders 461mountain birch 91mountains 128–129, 176Mount Rainier national Park 239mourning doves 350mouth-brooding 481mudpuppies 18Muir, John 239, 377–378, 413, 548cmulberries 60, 118, 186, 527mule deer 345Müller, Fritz 99Muller, Robert 373Mullerian mimicry 45, 99Mundy, n. i. 148Munson, t. V. 60Muraenidae 149Muricella 250Musca domestica 324muscles 86, 362mushrooms 261, 262muskeg 92musk oxen 538muskrats 370
xviii+598_Biodiversity_F.indd 586 11/17/11 1:13 PM

tundra 587Index 587
mussels 26, 99, 467mustard plants 12, 74, 84, 186, 310, 460mutations
biodiversity, patterns of 49biodiversity, resources for medicine 68disjunct species 194Dna studies of biodiversity 202extinction 233–234founder effect and genetic drift 258hybridization 309insects 320life history, diversity of 353natural selection 384, 388–389population genetics 431, 434–435
mutualism 378–383, 379, 380agriculture 8bacteria 41, 43coevolution 121–122communities, ecological 126convergence 144–145ecology 207fungi 263–264Gaia 267insects 323Janzen, Daniel H. 338lichens 347–349
mycelium 260mycobionts 264, 347–349Mycoplasma 32mycoplasmas 67mycorrhizae 122, 263, 264, 381Myllokunmingia 97, 245myna bird 332Myotis lucifugus 481myriapods 35Myrica cerifera 333Myriophyllum spicatum 330Myxococcus xanthus 41Myxotricha paradoxa 381Myzomela pammelaena 134
Nnabhan, Gary 58nagoya Protocol 549cNaja naja 456–457naked mole rat 145–146nalidixic acid 68namibia 175, 217Nanoarcheota 33Nanosella fungi 324nardoo fern 177narrow-sense heritability 432Nasua nasua 331Nasutitermes triodiae 322national academy of Sciences xi, 2national environmental Policy act (nePa)
243, 549cnational forests 238, 239, 241, 548cnational Marine Fisheries Service Office of
Protected Resources 219national Oceanic and atmospheric
administration (nOaa) 280national parks 239, 240t–241t, 548cnational Park Service (nPS) 2, 220, 239,
548cnational wildlife refuges 241native american lands 238native americans
agriculture 11biodiversity, resources for agriculture
62disjunct species 196disturbances, ecological 201grasslands and savannas 279Pleistocene extinction 416predation 439restoration ecology 461succession, ecological 501temperate deciduous forests 516
natural gas 102
Natural History of Selborne (White) 162natural hybrids 309, 310Naturalist on the River Amazons, The
(Bates) 46naturalists 542–545natural pesticides 327natural resource destruction 239natural Resources conservation Service
242natural selection 384–390, 388t, 548c
adaptation 3, 5agriculture 11–12Bates, Henry Walter 45biodiversity, concept of 46biodiversity, patterns of 50biogeography 72biomimicry 82–83cambrian explosion 96competition, ecological 131cultural diversity 158Darwin, charles 164, 165Darwin’s finches 167–169environmental ethics 226environmental psychology 228Gaia 267, 268herbivory 305life history, diversity of 351–355parasitism 405pollination 418population biology 426population genetics 433–434predation 437–438resistance, evolution of 459–460sexual selection 478–480, 482speciation 489–490, 492, 494Wallace, alfred Russel 542–545
natural systems agriculture (nSa) 61, 62, 83, 338, 390–392
Natural Theology (Paley) 162the nature conservancy 549cnature preserves 298nautilus 25n-dimensional hyperspace 132neanderthals 318nebraska 258neck 16, 191Necroscia sparaxes 480nectar 169, 419, 420nectar guides 418Nectarinia sericea 134nectar thieves 120, 419Necturus maculosus 18needle-leaved evergreen trees 92neem tree 59negative feedback 267nematocera 323nematocysts 23nematodes 27, 68, 124nematomorpha 28nemertea 28nemesis star 373nene 234, 258neocallimastigomycota 261neogene period, biodiversity in 392–394,
392t–393t, 393neognaths 89neopterygii 247neoteny 18, 483neotropics 527Neozeleboria cyroptoides 99nePa. See national environmental Policy
actNepa cinerea 467Nepenthes lowii 107Nerine (fishing trawler) 249nerpa seal 469nervous system 24Nesochen sandiviensis 234net production 55nettles 75, 76, 490neurogenesis 69
neutral theory of evolution 434nevada 173new caledonia 75, 221, 254newfoundland 53, 217new Guinea 9, 527, 542new Hampshire 197, 312new Mexico 175, 461, 461New Roots for Agriculture (Jackson) 226newton, Sir isaac 166newts 438new World monkeys 366, 442, 527new World tropics 49new World vultures 89new Zealand 87, 89, 119, 136, 138, 141,
458, 518nGOs. See nongovernmental organizationniche 132Nicophorus 325Nicotiana attenuata 419, 420Nicotiana digluta 310Nicotiana glutinosa 310Nicotiana tabacum 310Nicrophorus americanus 170nightshade family 188niihau 7Nilaparvata lugens 327nile perch 332nine-banded armadillo 367nipples 360nitrates 42, 394–395nitric acid 1nitrites 42Nitrobacter 394nitrogen 42, 70, 206, 212nitrogenase 394nitrogen cycle 71, 267, 394–395nitrogen fixation 42, 394nitrogen-fixing bacteria 13, 63, 122, 186,
212, 380nitrogen gas 394Nitrosomonas 394Nitrospira moscoviensis 43nitrous oxide 287, 395nixon, Richard 243–244nOaa. See national Oceanic and
atmospheric administrationnoah 224nobel Peace Prize 358, 359no child left inside 229nodules 394nonavian dinosaurs 113, 190noncoding Dna 111nongovernmental organization (nGOs) 59nonrenewable resources 212norfolk island pine 136, 477north africa 36, 175, 517, 518north america
adaptive radiation 6biodiversity, patterns of 49, 52biodiversity, resources for agriculture
61biogeography 73–76boreal and subalpine forests 91, 92communities, ecological 128conifers 137deserts, cool 173deserts, polar 174deserts, warm 175–177, 179–180Devonian period, biodiversity in 182dicots 185–187, 189disjunct species 194–196grasslands and savannas 274–279habitat fragmentation 301ice ages 316–318invasive species 330keystone species 344–345mammals 367Paleogene period, biodiversity in 398Pleistocene extinction 414–416reptiles 457temperate coniferous forests 511
xviii+598_Biodiversity_F.indd 587 11/17/11 1:13 PM

first entry588 tundra 588 first entry588 Index588
temperate deciduous forests 513, 515, 516
temperate rain forests 519tundra 537
north atlantic Ocean 285northern Hemisphere
biodiversity, patterns of 48biogeography 73–74carboniferous period, biodiversity
in 103communities, ecological 128–129competition, ecological 133greenhouse effect 282ice ages 315Quaternary period, biodiversity in 452shrublands 487
northern river otters 370norway 59norway spruce 511Nostoc 348Notaden bennetti 66nothofagaceae 73Nothofagus 73notochord 29, 245Notomys alexis 181notonectidae 467novotny, Vojtek 50nPS. See national Park ServicenSa. See natural systems agricultureNucifraga columbiana 137nuclear membrane 231nuclear plants 549cnuclei 447nucleic acid sequences 32–33nucleus 230–232null models 48nunataks 316nunavat people 16nurse logs 518nurse plants 463nutrient cycling 381nutrients 170, 171, 263–264, 348, 466,
469, 530–531nutrition 11, 212nuts 185, 186nymphs 321
oOahu 7oaks x, 7, 129, 403, 506
adaptive radiation 6biogeography 73biological classification 77–79communities, ecological 128, 129dicots 185disturbances, ecological 200endemic species 222grasslands and savannas 278greenhouse effect and biodiversity 296herbivory 303hybridization 308.309, 309ice ages 318keystone species 345mammals 363protists 446shrublands 487succession, ecological 504temperate deciduous forests 515temperate dry forests 517–518temperate rain forests 518
oases 179oats 12Obama, Barack 220, 292obligate mutualism 380ocean acidification 297ocean currents
deep oceans, biodiversity in 171deserts, warm 176greenhouse effect 284–285Humboldt, alexander von 307
hydrological cycle 311neogene period, biodiversity in 392Quaternary period, biodiversity in 450
ocean floor 172ocean mutualisms 380–381oceans 101–102, 170–173, 292, 311, 381,
396ocean sunfish 247ocean temperature 286ocean vents 32, 41, 43ocelli 35Ochotona curzoniae 344ocote blanco 518ocotillo bush 177octopus 25, 26, 154, 171Oculina varicosa 149odd-toed ungulates 365Odocoileus virginianus 518Odonata 321Odysseus 223Oecophylla ants 497Oenothera caespitosa 128, 199Oenothera hookeri 345Oenothera rhombipetala 418, 419Office of Science and technology Policy 2offshore oil drilling 215. See also BP
Deepwater Horizon oil spillohia trees 505, 507oil. See petroleumoil fish 222oil palm plantations 13oilseeds 61, 62oil spills 42, 242. See also BP Deepwater
Horizon oil spillOkamoto, noriko 510okapis 56Oklahoma 191, 194, 196, 221, 242, 303,
304, 515Old testament 224Old World monkeys 366, 442, 527Old World vultures 89Olea europaea 196Olea laurifolia 196Oligocene epoch 367oligotrophic water 466olive trees 196Olympic rain forest (Washington) 518ombrotrophic bogs 91ommatidia 320omnivores 151, 208, 365onagers 173“On civil Disobedience” (thoreau) 521Oncorhynchus gorbuscha 467Oncorhynchus keta 467Oncorhynchus kisutsh 467Oncorhynchus tshawytscha 467Ondatra zibethicus 370On Human Nature (Wilson) 545, 546onions 144, 375, 431onomyid primates 443On the Origin of Species (Darwin) 479,
489, 508, 544, 548cBates, Henry Walter 45biogeography 72cladistics 113–114communities, ecological 130Darwin, charles 165natural selection 385, 386population biology 424thoreau, Henry David 522
Onychophora 28oomycetes 446–447Opabinia 97open circulatory systems 21Ophioglossophyta 476Ophiostoma 262Ophisaurus 457Ophiusa tirhaca 324opisthokont protists 448–449Opisthorchis sinensis 29–30opossums 354, 362, 437
opportunistic infections 68opposable thumbs 366, 441Opuntia basilaris 189Opuntia ficus-indica 61Opuntia megasperma 273oracle oaks 309orange roughy 172oranges 62, 63, 262orangutans 443orbit of earth 315Orchidaceae 376orchids 376
biodiversity, patterns of 52coevolution 120decomposers 170hybridization 310monocots 376parasitism 403pollination 419, 420tropical rain forests and deforestation
526order (taxonomic) 78Ordovician extinction 372Ordovician period, biodiversity in 246,
396, 396–397, 397t, 472, 488Oregon 136, 263Oreskes, naomi 280organelles 230, 231organic agriculture 13Organization for tropical Studies 339organ pipe (cactus) 180organ transplants 66, 262“Origin of Human Races and the antiquity
of Man, the” (Wallace) 544Origin of Species. See On the Origin of
Speciesornamentation 480ornithischians 191, 192Ornithopods 192Ornithosuchus 524Orrorin 443Orthoptera 38, 321Orycteropus afer 367Osage oranges 118
adaptation 6biodiversity, patterns of 50–52camouflage and mimicry 98conifers 136convergence 148cretaceous period, biodiversity in 155Darwin, charles 166ecology 207flowering plants (angiosperms) 253flowering plants and pollinators
119–121herbivory 305monocots 375, 377mutualism 121–123parasitism 402parasitism to commensalism 121plants and herbivores 116–117predation 436–438predators and prey 117–119seed dispersal 471seed plants 478symbiogenesis 510temperate deciduous forests 513tropical rain forests and deforestation
526Osbeck, Peter 355Osmia lignaria 421Osmia ribifloris 421ospreys 80–81, 437, 466ostariophysians 248Osteichthyes 246ostracods 158ostriches 87, 88otters 370ovaries 87overfishing 141, 172, 250overhunting 234, 343, 416, 417, 439, 461
xviii+598_Biodiversity_F.indd 588 11/17/11 1:13 PM

tundra 589Index 589
overkill hypothesis 416, 501overpopulation 164, 165Oviraptor 155, 193Ovis aries 12Owen, Sir Richard 166, 190Owens Valley Desert 173owls 89, 520Oxalis pes-caprae 491oxazolidinones 68Oxyaena 400oxygen
animals 26biogeochemical cycles 69–71cambrian explosion 95carboniferous period, biodiversity
in 103deep oceans, biodiversity in 171ecology 205ecosystems 212ecosystem services 214Gaia 267–269mass extinctions 373Ordovician period 396Permian extinction 407–408rivers, streams, ponds, and lakes 465,
466oxygen depletion 13oxygen isotope ratios 407–408Oxytropis 84, 303oysters 25, 463
pPachycephalosaurs 192Pacific northwest 296, 519, 520Pacific Ocean 141, 172Pacific Rim 141Pacific trash Vortex 172, 250–251Pacific yew 66Paedocypris progenetica 247Paine, Robert 343painkillers 66, 67painted turtles 455Pakicetus inachus 399Pakistan 175, 460Palaemon macrocadtylus 380palate 456Palau 23Paleocene-eocene thermal Maximum
102–103, 374–375, 398Paleogene period, biodiversity in 362–363,
398–401, 399t–400t, 401paleognaths 87–89paleotropics 527Paleozoic era
amphibians 15birds 87cambrian period, biodiversity in 96–97carboniferous period, biodiversity in
103–106Devonian period, biodiversity in
182–184fishes 245–246, 249fossils and fossilization 258Gaia 269–270mass extinctions 372, 374seedless plants 474, 475seed plants 477Silurian period, biodiversity in 488–489
Paley, William 162Palin, Sarah 68palms 255
biodiversity, patterns of 52biodiversity, resources for agriculture
62, 63deserts, warm 179endemic species 222flowering plants 254–255marshes and swamps 369monocots 376
palolo worms 26
pampas 276–278Panama 52, 284Panama, isthmus of 73, 74, 171, 284, 316,
450Panama canal region 56, 129Panama disease 62Panama land bridge 367pandas 5, 363Panderichthys rhombolepis 15Pandion haliaetus 437, 466Pangaea 73, 154, 409, 411, 523pangenesis 386Pan paniscus 443Panthalassic Ocean 105Pan troglodytes 443Panulirus argus 157Papaveraceae 185Papaver californicum 487papaya 62–63paper daisy 177paper parks 244Papua new Guinea 87, 538, 540papyrus sedge 377Paracanthopterygii 248Paraceratherium 400Parameciuim 447páramo region, ecuador 538, 540parapatric speciation 490paraphyletic cladistic group 112–114, 185,
245parasite resistance 480parasitism 401–407, 402–405
amphibians 18, 19animals 24–25, 27–31arthropods 38camouflage and mimicry 99–100coevolution 121, 122commensalism 124communities, ecological 126convergence 144, 148ecology 207fungi 261herbivory 305insects 322mutualism 379protists 447
Parasuchus 524Parmesan, camille 296, 297parrot feathers 467parrots 471Parthenociccus quinquefolia 187Parus atricapillus 69passenger pigeons 234, 235, 461, 516, 548cPasseriformes 90passerines 90Passiflora 117passionflower 117, 305pastures 532Patagonian Desert 173pathogenic bacteria 43, 390pathogenic fungi 263, 263t, 380pathogens 263, 446–447Patriofelis 400Paulownia tomentosa 425–427Pauropoda 35Pavo crisatus 85pawpaws 119pax gene 148peaches 186peacock 85peanut worms 28pearlwort 174pears 186peas 548cpeatmoss 474pecan trees 515pectoral fins 246, 247, 249pedipalps 35, 36, 496Pelagia panopyra 23pelagic waters 170–171Pelecaniformes 89
Pelican island, Florida 241, 548cpelvic fins 246, 249pelvis 192pelycosaurs 454penguins 89penicillin 67–68, 262Penicillium 262Penicillium chrysogenum 262Penicillium italicum 262penis 35, 87, 481, 482penis worms 28Pentastomes 157peppered moths 168pepper tree 372perches 249perciform fishes 249peregrine falcons 89perennials 61–62, 426, 499, 502, 513, 538peripatric speciation 490perismmons 504Perissodactyla 365periwinkle (flower). See rosy periwinkleperiwinkle snails 297permafrost 538Permian extinction 407–411
carbon cycle 102, 103environmental ethics 223–224Gaia 268–269insects 320mass extinctions 372, 374
Permian period, biodiversity in 361, 411–413, 411t, 412, 454, 477
permineralization 256Permo-triassic boundary 407–409Persea americana 118persimmon 60, 471, 504, 527persistent organic pollutants (POPs) 80,
81t, 82t, 212, 422, 502Peru 175pesticides
agriculture 13arthropods 38biomagnification 80–82carson, Rachel louise 107–109ecology 207federal government role in conservation
(U.S.) 242iPM 327, 328pollination 421pollution 422resistance, evolution of 459–460
pest management 62, 215, 326–328pests 36, 38petals 184, 186, 188, 252, 252, 254, 375,
376, 376, 418Petraitis, Peter 135Petrified Forest nation Park 341petrified wood 257petroleum. See also BP Deepwater Horizon
oil spillagriculture 14bacteria 42bioremediation 84carbon cycle 102carboniferous period, biodiversity
in 103continental shelves, biodiversity on
142–143ecology 206ecosystems 209, 212ecosystems services 217federal government role in conservation
(U.S.) 242–243greenhouse effect 289tropical rain forests and deforestation
533tundra 540
pets 382phacelia 487Phacelia campanularia 177Phacelia cicutaria 128
xviii+598_Biodiversity_F.indd 589 11/17/11 1:13 PM

first entry590 tundra 590 first entry590 Index590
Phacelia grandiflora 487phaeophytes 446Phalaropus lobatus 481Phanerozoic eon 96–97, 372pharmaceuticals
bacteria 42biodiversity, resources for medicine 66deserts, warm 181–182ecosystem services 215herbivory 304pollution 423tropical rain forests and deforestation
526Phasianus colchicus 330Phasmida 321–322Phasolarctos cinereus 518pheasants 330phenology 522phenotypes 424, 430, 434phenotypic plasticity. See plasticity,
phenotypicphenotypic traits 431–432pheromones 328, 498Philippine islands 62, 221Philippine tarsier 443Phillips, John 407phloem 254, 255, 472, 477Phoca sibirica 222, 469Phoenix dactylifera 179Pholidota 367Pholistoma auritum 133Pholistoma membranaceum 128Phoneutria nigriventer 498Phoronida 27phosphate 263phospholipids 32phosphorus 70, 72, 264, 381, 466photonic crystals 83photons 205, 206photosynthesis
bacteria 42biogeochemical cycles 70carbon cycle 101convergence 144deep oceans, biodiversity in 170deserts, warm 180ecology 205ecosystem services 214eukaryotes 231fossils and fossilization 258Gaia 267–270greenhouse effect 291neogene period, biodiversity in 393
photosynthetic bacteria 41, 381photosynthetic symbionts 23, 347–349phycobiont 264, 347–349phyletic constraints 387–388Phyllium giganteum 98Phylloscopus trochiloides 494phylloxera bug 60Phylocode 80, 113phylogenetic analysis. See cladisticsphylogenetic species concept 490phylum (taxonomic) 78Phymatotrichum omnivorum 263Physalia physalis 23physiological adaptation 147–148, 455Phytophthora infestans 446Phytophthora megakorya 447Phytophthora ramorum 446–447phytoplankton 52–53, 209Pianka, eric 350Picea 92, 137Picea glauca 76, 196, 502Picea omorika 222Piciformes 89–90pickleweed 60, 176Pielou, e. c. 72–73pigeons
birds 89Darwin, charles 165
extinction 234, 235natural selection 386population biology 424predation 439restoration ecology 461temperate deciduous forests 516
pigments 22–23pigs 12, 333, 365pikas 344, 538pillbugs 158Pilobolus 261pilocarpine 67Pilocarpus jaborandi 67Pilosa 367Pimentel, David 209Pinaceae 137Pinaroloxias inornata 169Pinatubo, Mount 285Pinchot, Gifford 239, 241, 347, 378, 413–
414, 548cpine(s) 200, 281
biogeography 76biological classification 79boreal and subalpine forests 92coevolution 118–119communities, ecological 127, 128conifers 136, 137disjunct species 196disturbances, ecological 199–200,
201tfungi 264greenhouse effect 281hybridization 308life history, diversity of 354seed dispersal 471seed plants 477, 478succession, ecological 504temperate coniferous forests 511temperate dry forests 517, 518
pineal eye 246pineapples 62, 63pine bark beetles 297pine needles 303Pingelap atoll 258Pinguicula vulgaris 106pink pygmy sea horse 250pink salmon 467pinnipeds 363–364piñon-juniper woodlands 511Pinus 79, 137Pinus albicaulis 136, 137, 297, 540Pinus aristida 92Pinus banksiana 92Pinus cembroides 518Pinus contorta 200Pinus coulteri 517Pinus flexilis 137, 196Pinus garryana 92Pinus halepensis 517Pinus lambertiana 518Pinus longeava 92, 137, 138, 281, 354Pinus muricata 128, 136, 196, 199, 200,
308, 428Pinus pinea 517Pinus ponderosa 136, 200, 518Pinus radiata 138, 308, 330, 518Pinus rigida 200Pinus sabiniana 517Pinus strobus 511Pinus sylvestris 264, 511Pinus thunbergii 511Pinus torreyi 137pipefishes 248–249piranhas 248pistillate cones 135–137pistillate flower 252pistils 419
biological classification 77dicots 184, 188flowering plants 252, 254monocots 375, 377
pitcher plants 91, 106, 107, 438pitch pines 200Pitnick, Scott 479Pitohni dichrous 436pit vipers 181, 457placenta 362placental mammals
biogeography 74–75convergence 146, 146tmammals 359, 360, 362, 367–368tropical rain forests and deforestation
527placoderms 183, 246placozoans 22planarians 24planctomycetes 231, 232plankton 151, 170, 171plant biogeography 179–180plant breeders 58–59, 61plant defenses 144–145, 303, 349plant diseases 460plant growth 296, 349plant migration 296plant range shift 296plant reproduction 350plants
acid rain and fog 1adaptation 3, 5–6biodiversity, patterns of 49, 52biodiversity, quantification of 54–57biodiversity, resources for medicine 66biogeochemical cycles 70, 72biogeography 73–76biological classification 77–79bioremediation 84bogs, life in 91camouflage and mimicry 98–99carboniferous period, biodiversity
in 105carnivorous plants 106–107coevolution 116–117commensalism 124communities, ecological 130competition, ecological 132–134convergence 144–145cretaceous extinction 151cretaceous period, biodiversity in
154–155Darwin, charles 166decomposers 170deserts, polar 174deserts, warm 175, 178t–179tDevonian period, biodiversity in 183disjunct species 194–195disturbances, ecological 197–198Dna studies of biodiversity 202ecology 205, 206ecosystems 208, 212ecosystem services 213–217endemic species 222eukaryotes 232extinction 234, 235fossils and fossilization 257, 258fungi 263–264Galápagos islands 272tgreenhouse effect 289–293herbivory 303–306hybridization 308–311Jurassic period, biodiversity in 341life history, diversity of 351, 353linnaeus, carolus 355marshes and swamps 369mass extinctions 374neogene period, biodiversity in 393Paleogene period, biodiversity in 399parasitism 403Permian period, biodiversity in 412pond species 468tpopulation biology 424, 426–428population genetics 431predation 438
xviii+598_Biodiversity_F.indd 590 11/17/11 1:13 PM

tundra 591Index 591
Quaternary period, biodiversity in 452restoration ecology 463rivers, streams, ponds, and lakes 467seed dispersal 470–471sexual selection 483–484Silurian period, biodiversity in 488speciation 491–492temperate deciduous forests 512–516thoreau, Henry David 522triassic period, biodiversity in 524tropical rain forests and deforestation
526, 527, 531tundra 537–540tundra species 539t
plasmids 40plasmodial slime molds 448Plasmodium falciparum 509plastic 172, 227, 251, 502plasticity, phenotypic 3, 4, 132, 388, 432plastron 455Platanaceae 185Platanus occidentalis 516plateau pika 344plated lizard 458Plateosaurus 524Platycerium veitchii 476Platyhelminthes 24platypus 361, 363platyrrhines 442pleiotropic genes 353–354Pleistocene epoch 315, 450–452Pleistocene extinction 414–417, 415t, 416t
biodiversity, concept of 46coevolution 118–119extinction 234keystone species 343Quaternary period, biodiversity in 452Wallace, alfred Russel 544
Pleistocene overkill hypothesis 416, 501Pleopeltis polypodioides 476plesiadapiform 443plesiosaurs 341Plethodon jordani 481plethodont salamanders 18pleurodires 456Pliny the elder 548cplovers 89plowing 391plums 186Pneumocystis 262Poaceae 74, 376–377Podocarpaceae 136Poecilia reticulata 437poison dart frogs 17, 18, 67poison ivy 187, 298poison oak 187poisonous frogs 68, 436poison sumac 187Poland 516polar bears x, 311Pollan, Michael 13, 381pollen 258, 416, 477, 484pollen tubes 484pollination 417–422, 418, 419
adaptation 6coevolution 119–121conifers 135convergence 144Darwin, charles 166ecology 206–207ecosystem services 215greenhouse effect and biodiversity 298hybridization 308insects 326marshes and swamps 371monocots 376, 377sexual selection 483–484speciation 492temperate deciduous forests 513tundra 538
pollinators x, 119, 120–121, 132, 253, 421
pollution 422–423acid rain and fog 1–2agriculture 12, 13amphibians 18–19biomagnification 80–82bioremediation 83–84deep oceans, biodiversity in 172–173ecosystem services 214environmental ethics 224, 227federal government role in conservation
(U.S.) 242, 243fishes 250–251greenhouse effect 289lichens 348–349marshes and swamps 372
pollution credits 2polychelids 157polyculture 62–63, 63, 391Polygonum pensylvanicum 133polymerase 69polymorphism 430polyps 23Pombo, Richard 220Pompilidae 498ponderosa pines 127, 136, 200, 511ponds. See rivers, streams, ponds, and lakesPongo pygmaeus 444poplar 186poppies 177, 185, 497POPs. See persistent organic pollutantspopulation biology 423–430, 428
disjunct species 194–197ecology 206founder effect and genetic drift
258–259natural selection 384
population control 438population density 132population genetics 139, 430–436, 431, 433tpopulation growth 10, 48–49, 164, 385,
424–427population variability 165Populus deltoides 73, 187, 276, 483, 516Populus tremuloides 92, 502Populus trichocarpa 186porcupines 353pores 230–231, 350tPorifera 22, 22pork tapeworms 31Portugal 196, 298Portuguese man-of-war 23positive feedback 267, 268, 270post oaks 200, 278, 296, 303, 304t, 309,
515potassium 70, 212potato beetle 325potatoes 188Potato Famine 446Powell, John Wesley 548cpower plant scrubbers 1prairie dogs 198, 198, 344, 366prairie potholes 468–469prairies 274
biodiversity, resources for agriculture 62
biogeography 76biomimicry 83communities, ecological 128deserts, cool 173disturbances, ecological 198–200grasslands and savannas 274–276ice ages 317natural systems agriculture 391restoration ecology 461
prawns 157praying mantis 322, 481Precambrian 268–270, 315, 372precipitation
deserts, warm 175ecology 206greenhouse effect 285–286
greenhouse effect and biodiversity 298hydrological cycle 311–314shrublands 486temperate deciduous forests 513tropical rain forests and deforestation
525, 531tropical seasonal forests 535tundra 538
predation 436–441, 437, 439biodiversity, patterns of 51cambrian explosion 96camouflage and mimicry 99coevolution 116–119fishes 246–248
predatorsagriculture 13animals 27, 28arthropods 36birds 89communities, ecological 126, 130competition, ecological 132coral reefs 149deep oceans, biodiversity in 170, 171ecology 206Jurassic period, biodiversity in 341keystone species 344leopold, aldo 347mammals 363marshes and swamps 370parasitism 401, 402population biology 428reptiles 456
predator satiation 437predator switching 51predatory insects 327–328premature birth 351–352preservation 345, 378pressure 32, 171prey 117, 206, 436–438. See also predationprey mimicry 98–99Priapulida 28prickly pear cactus 61, 127primary productivity 209primary succession 504–505, 505primates 352t, 366, 441–445, 442t, 444, 527Primer of Population Biology, A (Wilson
and Bossert) 545–546Princess anne tree 425–427Principles of Geology (lyell) 163Priodontes maximus 367prions 210Pristionchus pacificus 124Proboscidea 366–367Procapra gutturosa 173Procellariiformes 89Prochlorococcus 42progymnosperms 183prokaryotes 32–33, 40, 230, 231prolegs 37promiscuous mating systems 482pronghorns 119, 173Prontosil 68prosimians 441prostate cancer 66Protacanthopterygii 248protected areas 139, 301–302proteins 147, 231, 432–433Proterozoic 33, 42Protista 79, 348, 548cprotists 445–449, 445t
amphibians 18animals 20biological classification 79decomposers 170deep oceans, biodiversity in 170eukaryotes 232lichens 348mutualism 381parasitism 402, 404symbiogenesis 509, 510
protostomes 24
xviii+598_Biodiversity_F.indd 591 11/17/11 1:13 PM

first entry592 tundra 592 first entry592 Index592
Prototaxites 488Prunus 186Prunus africana 66Prunus serotina 186, 516Prunus virginiana 303, 511Psammolepis 183Psaronius 105Pseudomonas 124Pseudomonas aeruginosa 41Pseudomyrmex ants 380, 497Pseudomyrmex spinicola 380pseudopods 446Pseudopterogorgia elisabethae 67pseudoscorpions 36Pseudotsuga menziesii 137Psilophyta 476Psilotum 476Psudomyrmex 380Pteridinium 217Pteridium aquilinum 476Pteridophyta 472, 474–476pteridosperms 477Pteris ensiformis 84pterosaurs 155–156, 454, 524Pterospora 110, 144public health 333, 390, 460public lands 239–242, 244Puccinia 263Puereria montana 333, 333Puerto Rican tree frogs 18puffballs 260Pulex irritans 38, 323pulmonary circulation 360, 455pumiliotoxin 17punctuated equilibria 387, 490puncture vine 145, 168, 180, 470pupae 323, 324pupfishes 180, 301Purgatorius 156, 443Purísima Hills, california 128, 428purple loosestrife 330, 372purple-striped jellyfish 23Putuo island 235Pycnogonida 36–37pygmy forests 128pygmy sea horse 250pygmy shrew 196Pygopodidae 457–458pyrethrin insecticides 59pyrite 71Pyrolobus fumarii 32Pyrus 186Pythium 447Pythonidae 456
qquadrate bone 361quartz 71, 102Quaternary period, biodiversity in 315,
450, 450–453, 451t–452tQueensland, australia 196Quercus
adaptive radiation 6biogeography 73biological classification 79dicots 185hybridization 308shrublands 487temperate deciduous forests 515
Quercus agrifolia 129Quercus alba 185Quercus carmenensis 222Quercus deliquescens 222Quercus douglasii 6, 7, 128, 129, 278Quercus dumosa 6, 7Quercus kelloggii 309, 345Quercus macrocarpa 6, 7, 303, 309, 511Quercus marilandica 515Quercus nigra 185, 185Quercus petraea 318
Quercus robur 6, 7, 303, 318Quercus rubra 506Quercus stellata 304t
dicots 185disturbances, ecological 200grasslands and savannas 278greenhouse effect and biodiversity 296herbivory 303hybridization 309temperate deciduous forests 515
Quercus suber 6, 7Quercus virginiana 6, 7, 185, 369, 515Quercus wislizenii var. frutescens 309Quercus x guadalupensis 309Quercus x morehus 309quillworts 474quinoa 62quinolone 68quorum sensing 41
rradial cell division 28radial symmetry 20, 20, 23–24radiation 549cradioactive waste 82radiolarians 448radish 310radula 26Raffesia 402–403Rafflesia arnoldii 527ragweed 298railroads 238rainfall. See precipitationrain forest(s) 17
agriculture 8–9biodiversity, patterns of 48, 52competition, ecological 133Darwin, charles 163dicots 186herbivory 304hydrological cycle 313Janzen, Daniel H. 338–339succession, ecological 502tropical rain forests and deforestation
525rain shadows 130, 176, 176rainy season 168, 169ramets 431range shift of plants 296rank-free classification 80Ranunculaceae 185rapamycin 66Rapa nui 66Raphanobrassica 310Raphanus sativus 310Raphus cucullatus 234rat(s) 68, 145–146, 273, 360, 459, 460rat fleas 38, 323rat-tailed maggots 467rattlesnakes 457Rattus rattus 273Raup, David 372, 373Ravenala madagascariensis 222raw fish 30Ray, John 548cray-finned fishes 247–249, 356rays 171, 246–247, 341razorback pigs 12rbc gene 231Reagan, Ronald, and administration 2recessive allele 434recreation 229recurring patterns 373red algae 232, 448redback spider 498red blood cells 360red cedars 200, 313red crab 156, 157ReDD Program 533red fir 92
red foxes 330red jungle fowl 89red kangaroo 363red-naped sapsuckers 344red-necked phalarope 481red oaks 73, 185, 308, 309, 506Red River basin 465red squirrel 333red-winged blackbirds 370redwoods 136, 222, 356, 518regeneration 27reindeer 82, 538reindeer lichens 348reindeer moss 381reinsurance industry 214relative density 55relative dominance 55relative frequency 54–55relictual population 194, 196religion 10, 163, 164, 224–226, 228Remipedia 158renewable production 217Repenomamus robustus 156Report on the Lands of the Arid Regions of
the United States (Powell) 548creproduction
animals 24–26, 28, 31arthropods 35bacteria 41birds 87conifers 136conservation biology 139fishes 249fungi 260, 262insects 322life history, diversity of 349–354linnaeus, carolus 355population biology 426predation 437protists 447reptiles 455speciation 492–493spiders 498
reproductive expenditure 481–482reproductive interference 480reproductive isolation 78, 489–493, 544reproductive organs 26, 87, 481, 482reproductive patterns 308reproductive rates 352reproductive success 387reproductive systems 145–146reptiles 454–459, 454t, 455, 456
amphibians 16carboniferous period, biodiversity
in 106cladistics 110, 112, 113cretaceous extinction 151cretaceous period, biodiversity in 154dinosaurs 190–193Jurassic period, biodiversity in 341life history, diversity of 353living fossils 356mammals 360, 361neogene period, biodiversity in 393Permian period, biodiversity in 412Quaternary period, biodiversity in 452triassic period, biodiversity in 524
Republican Party 244, 270, 292reservoirs 312resistance, evolution of 327, 390, 459–460resource allocation patterns 349–352resource competition 47–48, 132, 134resource extraction 533resource investment patterns 349–352resource limitations 385resources 212respiration 20–21, 42, 86, 101, 267, 268restoration ecology 345, 460–465, 461, 464resurgence 207resurrection fern 476retroviruses 67, 68
xviii+598_Biodiversity_F.indd 592 11/17/11 1:13 PM

tundra 593Index 593
Reznick, David 390Rhabdophis tigrinus 438Rhagoletis pomonella 492, 493Rhea americana 173, 481rheas 87, 173, 481Rheobatrachus silus 68–69Rheobatrachus vitellinus 68–69Rheum palaestinum 179Rhincodon typus 246Rhinella margaritifera 98rhinoceros 235Rhinocodontidae 171Rhizobium 42Rhizoctonia 263Rhizodus hibberti 105rhizoids 473–474rhizomes 474–476rhizomorphs 262Rhizopus nigricans 261Rhodophyta 448rhubarb 179Rhynia 183, 476rhyniophytes 472–473Rhysocaryon 113ribbonweed 371ribbon worms 28ribosomes 40, 230, 231, 508ribs 15, 16, 192, 458rice 62, 263Rice, Stanley 194, 303, 483Richelia 381ricin 187, 303Ricinus communis 187–188, 303Rieseberg, loren 310Riftia pachyptila 122, 172ring-necked pheasants 330ring species 494ringworm 262Rio de Janeiro, Brazil 549cRio Grande system 465Rio Summit 549criparian forests 200river blindness 31rivers, streams, ponds, and lakes 1, 316–
317, 465–469, 468triwoche horses 56Rna 231Robinia pseudoacacia 186robins 234rock pigeons 386Rocky Mountain national Park 537Rocky Mountains 538
biogeography 73, 76boreal and subalpine forests 92conifers 137disjunct species 196disturbances, ecological 200succession, ecological 502temperate coniferous forests 511tundra 537
Rodentia 366rodents 74, 360, 366, 459, 460Rodhocetus 399Rodrigues island 235Rohrabacher, Dana 103, 290Roosevelt, theodore 239, 241, 378, 413,
414, 548croots 177, 376, 381Rosaceae 186rose family 186rosids 185–188rosinweed 61rosy periwinkle 66, 67, 67, 527rotenone 303Rotifera 28rotifers 21, 28Roughgarden, Joan 483roundworms 27, 29, 31, 353Roystonea 369rubber tree 188rubisco 111, 231, 508
Rubus 186ruminants 365, 378runoff
amphibians 18biogeochemical cycles 70disturbances, ecological 197ecosystem services 215federal government role in conservation
(U.S.) 243hydrological cycle 312–313marshes and swamps 372pollution 422, 423restoration ecology 462, 463rivers, streams, ponds, and lakes 466
rushes 377rye 310
SSabatia campestris 431Sabbath of the Fields 224Saccharomyces cerevesiae 262Sacculina 158Sagan, carl 369Sagan, Dorion 508sagebrush desert 173sage scrub 487saguaro 180Sahara Desert 175, 176, 312, 316Sahelanthropus 443St. Francis of assisi 224, 225St. Helena island 75Sakai, ann 492salamanders 16–18, 481Salicaceae 186Salicornia 176Salicornia bigelovii 60salicylic acid 67saline pools 180saline soils 60salinity 23, 32, 370salinization 464Salix 74Salminen, Juha-Pekka 303salmon 248, 249, 251, 467Salmonella 404salt cedars 333Salticidae 497salt marsh blue crabs 297salt marshes 370salt marsh grasses 310saltwater crocodile 456Salvinia 476Salvinia molesta 372, 467Samoa 26sandalwood tree 403sandburs 145Sand County Almanac, A (leopold) 226,
347, 549csand dollars 28sand dunes 504–505, 506sand grasshoppers 181sandhill cane 180sandpaper trees 278sandpipers 89San Francisco Bay 370San Francisco Peaks 76San Joaquin Valley, california 463Santa ana winds 200Santa Barbara oil spill (1969) 242Santa lucia fir 222Santalum 403Santa Rosa island 75“Santa Rosalia” (Hutchinson) 549cSapelo island 354Sapindaceae 187Saprolegnia ferax 18Saratoga Springs 301Sarcophilus harrisii 363sarcopterygian fishes 15, 249Sargasso Sea 171
Sargassum seaweeds 171Sarracenia purpurea 91, 106sashimi 30saurischians 191–192sauropods 191Sauroposeidon proteles 191savannas 318. See also grasslands and
savannasSavolainen, Vincent 203Saxifraga oppositifolia 174Scablands 317scala naturae 50, 78, 356scales 16, 455, 457Scalesia trees 272scallops 25, 26Scandinavia 91Scarabaeidae 170scarab beetle 170, 325scatterboarding 471scavengers 208Schemske, Douglas 492, 494Schiedea salicaria 492Schierenbeck, christina 12Schinus terebinthinifolius 372Schistosoma 30schistosomes 30schistosomiasis 30Schluter, Dolph 169Schulman, edward 281Schulman log 281Schulte, David 463Schweitzer, Mary Higby 190Sciurus carolinensis 333Sciurus vulgaris 333Scleroctinia 263scolecophid snakes 457Scoliopus bigelovii 420scorpion flies 420scorpions 35, 36, 36, 184, 436Scotland 487Scots pines 264, 511scramble competition 131scrubbers (power plant smoke) 1scrub oaks 6, 487sculpins 467scurvy 11, 58sea crocodiles 341sea fan coral 250seafood 82seagulls 89seahorses 248–249, 250sea ice 285sea krait 98sea levels 285, 396seals 222, 469Sea of Japan 172sea pens 218Searcy, K. 484sea scorpions 396seaside alder 139, 194–195, 221, 222, 308seasonal forests 525seasonal patterns 296, 304, 522sea spiders 36–37sea turtle 455sea urchins 28, 29seaweed 141–142, 171, 446secondary palate 456secondary succession 500, 502, 504sedges 91, 195, 377, 538, 540sedimentary layers 152sedimentary rocks 256sediments 70, 91, 407, 408, 416seed bank 428seed dispersal 92, 118, 119, 470–472,
520–522, 532seed ferns 412, 477, 524seedless plants 203, 472–477, 473t,
474–476seedlings 532seed plants 105, 135–138, 252, 258,
477–478
xviii+598_Biodiversity_F.indd 593 11/17/11 1:13 PM

first entry594 tundra 594 first entry594 Index594
seed predators 117–118, 471seeds 371, 477Seed Savers exchange 59seed vaults 59segmented bodies 34Seilacher, Dolf 217–218Şekercio, Çaǧan 297Selaginella 474Selaginella plana 475selective logging 533selenium 84Selenocosmia stirlingi 181semelparous species 350semen 481Senate, U.S. 109senescence 353, 354sepals 184, 252, 252, 254, 375, 376Sepkoski, J. J. 373sequoia 130, 136Sequoiadendron giganteum 136, 222, 354,
502, 511Sequoia national Park 239, 537Sequoia sempervirens 136, 518Serbian spruce 222Serial endosymbiotic theory 509serotinous pine forests 199–200serpentine soils 72sewage 13, 30, 61, 172, 466sex hormones 99sex ratios 387, 424sexual adornment 5sexual classification system 355sexual mimicry 98–99sexual reproduction
amphibians 16animals 21, 29arthropods 35birds 87fungi 260, 261, 263seedless plants 472spiders 498symbiogenesis 508
sexual selection 478–485adaptation 3, 5biodiversity, patterns of 50competition, ecological 134Darwin, charles 166natural selection 384parasitism 402
sexy sons hypothesis 479Seychelle islands 234, 493shade 214, 216shade-intolerant trees 499shade-tolerant trees 499, 500shafts (feathers) 85shallow seas 339Shannon, claude 55Shannon-Wiener diversity index 55sharks
biodiversity, resources for medicine 66carboniferous period, biodiversity
in 105cretaceous period, biodiversity in 154deep oceans, biodiversity in 171fishes 246–247, 250Jurassic period, biodiversity in 341keystone species 344neogene period, biodiversity in 392Paleogene period, biodiversity in 399Permian period, biodiversity in 412Quaternary period, biodiversity in 452triassic period, biodiversity in 523
sheath-winged beetles 37sheep 12, 429shells 25, 455Shenandoah national Park 2shield bugs 323shield shrimp 180shifting agriculture 531–532shortgrass prairies 173, 198–200, 274–276short-term carbon cycle 101
short-term mineral cycle 70, 71shrew opossums 363shrews 195–196, 367shrimp 156–158, 172, 180, 305–306, 380,
381shrublands 173, 486–488, 486t–487tshrubs 129, 174, 174t, 176, 177, 180, 187,
196Shubin, neil 15–16Shuvuuia 156Siberia
biogeography 75boreal and subalpine forests 91, 92carboniferous period, biodiversity
in 105Devonian period, biodiversity in 182endemic species 222Ordovician period 396rivers, streams, ponds, and lakes 469tundra 537
Siberian traps 409sibling conflict 352sickle-cell anemia 434Sideroxylon grandiflorum 462sidewinder 457Siegesbeck, Johann 355Sierra club 378, 548cSierra Madre 511Sierra nevada 129, 130
biogeography 76boreal and subalpine forests 92communities, ecological 127conifers 136, 137disjunct species 196endemic species 222Muir, John 377–378succession, ecological 502temperate coniferous forests 511tundra 537
Silene virginica 418Silent Spring (carson) 82, 108, 109, 242,
549csilicon 71silk 83, 497, 498Silphium integrifolium 61Silphium laciniatum 276Silphium terebintinaceum 276Silurian period, biodiversity in 472–473,
488–489, 489tsilverfish 321silver maples 187Simberloff, Daniel 134, 336, 546Simmondsia chinensis 61Simpson’s diversity index 55Singer, S. Fred 2single-species groups 436–437Sinocodon 361Siphonaptera 38, 323Siphunculata 322Sipuncula 28Sirenia 365Sitka spruce 518sixth mass extinction 228, 233, 235, 269,
374–375, 410, 417size 366size dimorphism 480skeletons 20, 22, 95, 96, 246, 247, 249, 258skin 17, 18, 455, 457skin bacteria 379skinks 457, 458skull bones 360–361, 454sky islands 336slash and burn agriculture 531–532SliMe (subsurface lithoautotrophic micro-
bial ecosystems) 43slime molds 446, 448, 449sloths 367, 367slugs 25smallpox 404, 428, 501Smith, thomas Bates 389Smithsonian magazine 338
smokestacks 1snails 25, 25, 26, 66, 172, 331snakes 155, 181, 333, 400, 438, 454–458,
456snapping shrimp 156–157snapping turtles 370snow 312Snow, allison 484Snowball earth 218, 269–270, 315snow gum 92snowshoe hare 438soap 327, 466soapberry bug 389, 493soapberry trees 187social mimicry 99–100social parasitism. See social mimicrysocial structures 352social systems 38sociobiology 546Sociobiology: The New Synthesis (Wilson)
546sockeye salmon 467–468sodium 212soft-shell turtles 455soft tissue 190soil
acid rain and fog 1biodiversity, resources for agriculture
60–61biogeochemical cycles 72bioremediation 84boreal and subalpine forests 92communities, ecological 128decomposers 170deserts, warm 179disturbances, ecological 198fungi 263restoration ecology 461shrublands 486succession, ecological 502temperate deciduous forests 515tropical rain forests and deforestation
530–531tundra 538
soil bacteria 41, 66Soil conservation Service, U.S. 242, 549csoil erosion 13, 128, 242, 391, 461, 463soil nutrients 42Solanaceae 188Solanum tuberosum 188solar flares 373solar power 83, 181solar radiation 270, 315sole 249Solenopsis invicta 325, 331, 332solifugids 35–36Solnhofen limestone 257Solomon, Susan 280somatic cells 353, 384somatic mutations 384, 431Sonoran Desert 175, 181, 301soot 1Sorex minutus 196sorghum 12Sorghum bicolor 12, 61Sorghum halepense 12, 61South africa
continental shelves, biodiversity on 141deserts, warm 180greenhouse effect and biodiversity 296Permian extinction 408–409shrublands 486speciation 491temperate dry forests 517
South americaagriculture 9Bates, Henry Walter 45biodiversity, resources for agriculture
62biogeography 74, 75birds 87
xviii+598_Biodiversity_F.indd 594 11/17/11 1:13 PM

tundra 595Index 595
cretaceous period, biodiversity in 154Darwin, charles 163deep oceans, biodiversity in 171deserts, cool 173deserts, warm 175, 177, 179–180dicots 189grasslands and savannas 276–278Humboldt, alexander von 307mammals 363, 367Pleistocene extinction 415temperate rain forests 519tropical rain forests and deforestation
527, 531, 532Wallace, alfred Russel 542
South Dakota 91, 127, 128, 196, 198, 199, 303, 511
Southeast asia 12, 62, 519, 533, 534Southern Hemisphere 73–74, 87, 92, 136,
316Soviet Union 263, 463–464soybeans 534Spain 196, 296, 298Spartina alterniflora 297, 310Spartina foliosa 370Spartina maritima 310Spartina x townsendii 310spatial patterns 522spawning grounds 251specialists (consumers) 208specialization 126, 418, 513, 526speciation 489–496, 495t
adaptive radiation 8biogeography 73Darwin’s finches 169ecology 207parasitism 402pollination 420Quaternary period, biodiversity in 452tropical rain forests and deforestation
526species xispecies-area relationship 48, 335species diversity 373–374species interactions 206–207, 269Species Plantarum (linnaeus) 548cspecies richness 54, 56specific mate recognition system 489–490spectacled lizards 458Spencer, Herbert 165sperm 20, 23, 29spermatophore 481sperm cells 473–476, 479, 481sperm nuclei 477Sphaeralcea coccinea 128Sphagnum 474sphagnum moss 91, 540Spheniciformes 89Sphenodon 458sphinx moth 37, 62, 324spice plants 305spices 116, 117, 188spicules 22spiders 496–498, 496–499
animals 27arthropods 35deserts, warm 181Devonian period, biodiversity in 184predation 437rivers, streams, ponds, and lakes 467sexual selection 481
spider silk 83spikes 376, 377, 470spines 144, 303spinifex hopping-mouse 181spiny anteater 361spiny-headed worms 28spiny lobster 157spiracles 320spiraea bush 66Spiraea ulmaria 66spirilla 40
spiritualism 544–545spirochetes 40, 231, 232, 509spitting cobra 457split-middle whip scorpions 36Spondias mombin 338sponges 22, 22, 66, 67, 69, 231, 381, 449spongouridine 67spoon worms 28sporangia 260, 261, 474, 475spores 260, 261, 348, 397, 473–475, 475sporophytes 472–477Sporormiella 416spotted owl 520Sprigg, Reginald 217Spriggina 217, 218spring budburst 296springtails 263, 321spruce bark beetles 297, 328spruce trees
biogeography 76boreal and subalpine forests 92conifers 137endemic species 222fossils and fossilization 258succession, ecological 502temperate coniferous forests 511temperate rain forests 518
spurges 133–134, 177, 180, 187squalamine 66Squalodon 399squamates 454, 456–458squids 25–26, 171squirrels 117, 333, 471squirting cucumbers 470Sri lanka 527stabilizing selection 387, 388t, 389staghorn ferns 475, 476stalk 261, 377stalked crinoids 397stamens 252, 376, 419
biological classification 77dicots 184, 186, 188flowering plants 252, 254monocots 375–377
Stamets, Paul 264staminate cones 135, 136staminate flower 252Stanford, alice 113stapes 16, 361staphlinid beetles 145staphylococci 41starfish 28, 29, 149, 154, 431starlings 330stealth 481Stegosaurus 341stem cells 353stems 376–377, 467stem veins 254Stenocereus thurberi 180stenopods 157Stenopterygii 248Stensen, niels 256Stentor 447steppes 199, 276–278Sterna paradisaea 89Sternula antillarum 196sternum 86Stethacanthus mirabilis 105steward 225Stewart, caro-Beth 110stick and leaf insects 321–322stickleback fish 481, 493–494stigmas 377stinging tentacles 23stink bugs 323stinkhorns 260Stipa 278stochastic processes 522Stockholm convention 82stomach 365stoneflies 466
strata 154, 256stratification 469strawberries 186streams. See rivers, streams, ponds, and
lakesstreamside habitats 200Strepserrhini 366streptococci 41Streptococcus mitis 43Streptomyces avermitilis 59stress 229Strigiformes 89strip-cropping agriculture 391Strix occidentalis 520strobilurins 59Strobilurus tenacellus 59stromatolites 408strontium-90 72structural defense 436–437sturgeons 222, 247, 356subalpine forests. See also boreal and subal-
pine forestsacid rain and fog 2biodiversity, quantification of 55, 56tboreal and subalpine forests 92–94communities, ecological 125disjunct species 196keystone species 344temperate rain forests 519tundra 540
succession, ecological 499–507, 500, 503t, 504–507
bogs, life in 91competition, ecological 134–135ecology 207thoreau, Henry David 520, 522
“Succession of Forest trees, the” (thoreau) 521
succulents 98, 128, 144, 177, 179–180, 376sugar maples 187, 462, 504, 515, 515sugar molecules 208sugar pines 511, 518sulfa drugs 68sulfate ions 71sulfur 70, 71sulfur dioxide 1, 2, 409sulfuric acid 1, 43, 71sumac family 186Sumatra 116sumpweeds 9sundews 91, 106, 438sunfish 308sunflowers 9, 60–62, 61, 84, 188, 310, 492,
494sunlight
bacteria 41biodiversity, patterns of 48deep oceans, biodiversity in 171ecology 205ecosystems 208, 210, 211greenhouse effect 286tundra 538, 540
sunscorpions 35–36sunspots 285superior traits 387Supersaurus 191, 191, 192, 341Supreme court, U.S. 238surgery 229survival of the fittest 165survivorship 427–428Sus scrofa 12Svalbard islands 174Svalbard seed vault 59swamp locusts 119swamps 274, 339, 473. See also marshes
and swampsswans 89sweat glands 5swifts 89Switzerland 2sycamores 185, 516
xviii+598_Biodiversity_F.indd 595 11/17/11 1:13 PM

first entry596 tundra 596 first entry596 Index596
symbiogenesis 507–510bacteria 42ecology 207eukaryotes 231, 232lichens 347–349Margulis, lynn 368–369speciation 491, 494
Symbion pandora 28symbiosis 116t
coevolution 116commensalism 123–124ecology 207ediacaran organisms 218lichens 347–349Margulis, lynn 368mutualism 380parasitism 402symbiogenesis 507
symbiotic cyanobacteria 22–23symmetry 20, 20, 480sympatric speciation 490–491Symphyla 35symplesiomorphy 110synapomorphy 110–112synapsids 360–361, 412, 454, 455synecology 206–207Synemosyna 497syphilis 404Systema Naturae (linnaeus) 548csystemic circulation 360, 455
TTachypleus gigas 46Taenia rhynchus saginatus 30–31Taenia solium 31taft, William Howard 414taiga 91, 92tail feathers 86tailless whip scorpions 36taiwan 221taklamakan Desert 173tallgrass prairies 274
biodiversity, resources for agriculture 62
biogeography 76biomimicry 83deserts, cool 173disturbances, ecological 198–200grasslands and savannas 274–276ice ages 317restoration ecology 461
tamarisks 179, 313Tamarix 179, 313, 333tammar kangaroo 362tanganyika, lake 6tannins 303, 304ttansley, arthur 549cTanystropheus 524taoism 226tapeworms 24–25, 30, 30–31taq polymerase 69tarantula hawk wasps 498tarantulas 497, 498Taraxacum officinale 330, 491tarballs 142tardigrades 27tarrant Regional Water District 242tarsiers 441Tarsius syrichta 443tasmania 12tasmanian devil 363tasmanian tiger 549ctaxaceae 136taxidermy 162taxodiaceae 136Taxodium distichum 137, 512, 513taxol 66, 67taxonomy 77, 355Taxus brevifolia 66teak 188, 537
Tectona grandis 188teeth
dinosaurs 191, 192fishes 246, 248fossils and fossilization 256–257mammals 360–363, 366reptiles 455
teleosts 247, 248, 524telomeres 231temperate coniferous forests 511–512, 519temperate deciduous forests 185–186,
512–517, 513, 514t–515t, 515temperate dry forests 517–518, 517t, 518temperate rain forests 518, 518–520, 519ttemperate zones 13, 49, 115–116temperature 49, 176, 206, 465, 466temperature extremes 32temperature patterns 115, 116, 279–284,
307, 316temperature regulation 86–87tennessee 55tennyson, alfred, lord 386ten percent law 50, 209, 439tentacles 23teosinte grass 11, 60termites 38, 322, 322, 379–381, 510terns 89, 196terrapin 455terra preta de Indio 461, 531terrestrial ecosystems 209terrestrial plants 374terrestrial vertebrates 454k–458territory 480terrorism 226testudines 455–456tethys Sea 523Tetrahymena 447tetraploids 309tetrapods 16, 17, 249, 454Tetrastigma 403tetrodotoxin 17Tevnia jerichonana 172texas 76, 175, 515thar Desert 175Thaumaptilon 218Thaumarcheota 33thecodont 361Theobroma cacao 60theophrastus 356, 548cTheraphosa blondi 498therapsids 361, 454therian 362thermodynamics, second law of 270Thermoplasma 32, 509Thermotoga 32Thermus aquaticus 43Thermus aquatilis 69theropods 87, 192Thinopyrum intermedium 61Thiobacillus concretivorans 43Thiomargarita namibiensis 40thomas, lewis 266thomas, Matthew 328thomson, Sir c. Wyville 548cthorax 37, 320thoreau, Henry David 228–229, 363,
520–523Thorius 17thorns 144three-color vision 366“three sisters” crops 62thrush 262thumb 366, 441Thunnus thynnus 172, 250thylacines 363, 549cthyreophores 192thysanurans 321tibet 56, 344ticks 35, 36, 403, 404tickspiders 36tidepools 142
Tidestromia oblongifolia 134, 176, 176, 177
tigers 308, 365, 537tiger sharks 247Tiktaalik roseae 15–16, 16, 184Tilia americana 506tillage 391timberline 540tinamous 87Tinea 262tipping points 286tissue 190Titanoboa 400Titanus giganteus 324toadfishes 248toadflax 403toads 18, 235, 331, 332tobacco 310tobacco hornworm caterpillars 306tolerance 49, 177, 180–181, 304–305, 513tolstoy, leo 521Tolypocladium inflatum 66Tolypocladium niveum 262tomatoes 188, 501tomato pinworm 328tongue 17, 456, 457tools 482tooth enamel 258Torreya 222Torreya taxifolia 137, 222torrey pine 137tortoises 272, 273, 455–456, 456toxic chemicals/compounds 42, 80–82,
133, 303, 304toxic ions 84, 303Toxicodendron 187toxic peptides 66toxic plant chemicals 187–188toxins
amphibians 18animals 23biodiversity, resources for medicine
66, 67ecology 207herbivory 304succession, ecological 502tropical rain forests and deforestation
526trace fossils 258, 397tracheae 21, 35, 36, 320, 497Trachycaryon 113Tradescantia ohiensis 376Tragopogon 492trait linkage 388traits 110–112, 166, 369transcendentalism 520–522transitional zones 537, 540transmutation 165, 166transpiration 313, 525trapdoor spider 497Trapelia 84Trapezia ferruginea 381trapping 328traveler’s palm 222Trebouxia 348tree(s)
biodiversity, quantification of 56commensalism 124Devonian period, biodiversity in 183dicots 185–188environmental psychology 228, 229fungi 263greenhouse effect 291–293hybridization 309life history, diversity of 349–351marshes and swamps 369mutualism 380succession, ecological 499, 500, 502wood pores 350t
tree cactuses 272tree ferns 475
xviii+598_Biodiversity_F.indd 596 11/17/11 1:13 PM

tundra 597Index 597
tree finches 167tree frogs 17, 18tree hibiscus 235tree-of-heaven 350tree of life 507–509tree of life project 79–80tree planting 358–359tree rings 137, 257, 281tree shrews 367trematodes 24Treponema pallidum 404triassic mass extinction 372–373triassic period, biodiversity in 454, 477,
523–525, 523t, 524tribal cultures 66, 159–160, 534Tribrachidium 217Tribulus cistoides 180Tribulus terrestris 144, 145, 180Triceratops 155Trichilia rubescens 305Trichinella spiralis 31trilobites 96, 105, 182, 396, 488trimerophytes 473trionychidae 455Triops australiensis 180Tripsacum 234Tritemnodon 400Triticale 310Triticum aestivum 12, 310Triticum boeoticum 310Triticum dicoccoides 310Triticum dicoccum 310Triticum monococcum 310Triticum spelta 310trivers, Robert 352trochanter 320Trombicula 36trophic cascade 130, 344trophic levels 208, 209tropical alpine tundra 540tropical biodiversity 47–49, 52tropical countries 29–31tropical fish 150tropical rain forests and deforestation 525,
525–535, 526, 528t–530t, 531, 532agriculture 8–9biodiversity, patterns of 47, 49, 52biodiversity, resources for medicine 66cloud forests 115–116competition, ecological 133Darwin, charles 163dicots 186, 188herbivory 304Janzen, Daniel H. 338–339succession, ecological 502temperate rain forests 519
tropical seasonal forests 380, 525, 535–537, 536t
tropic of cancer 535tropic of capricorn 535trout 248true bugs 322–323true ferns 475–476true shrimp 157truffles 260–261trumpet creepers 120Tsuga 137Tsuga canadensis 128, 511tuataras 356, 454tube feet 29tuberculosis 404tubeworms 171, 172Tubifex tubifex 467tubulidentata 367tulare lake 463tule elk 221tunas 249tundra 537–541, 538, 539t, 540
biogeography 74, 76boreal and subalpine forests 92communities, ecological 125
life history, diversity of 351restoration ecology 462succession, ecological 501
tunicates 29turbellaria 467turbot 249turkestan Desert 173turkey vultures 124turtles 349, 353, 370, 454–456, 455, 524twain, Mark 544typhaceae 377Typhlosaurus 458typhoons 286Tyrannosaurus rex 191, 192, 192, 257
uUintatherium 401Ukraine 82Ulmus 73ultraviolet radiation 18, 502, 540Umbellularia californica 129umbrella palm 222understory 526, 531understory birches 264underwater plants 467undulipodia 509UneP. See United nations environment
ProgrammeUneScO (United nations educational,
Scientific and cultural Organization) 273ungulates 365unicorn plant 470unidirectional lungs 86unique traits 110United nations 140, 533United nations Billion tree campaign 359United nations climate change conference
549cUnited nations convention on Biological
Diversity 549cUnited nations environment Programme
(UneP) 235, 358–359United nations Food and agriculture
Organization (FaO) 58United nations Framework convention on
climate change 549cuniverse, age of 268Ural Mountains 105uranium 84urban planners 228urea 18, 455uric acid 455Urnisa guttulosa 181ursodeoxycholic acid 66Ursus americanus 518Ursus arctos 516Urticaceae 75USDa. See agriculture, U.S. Department ofUtah 173Utricularia vulgaris 106–107
vvaccination 544vaginal yeast infections 262Vallisneria 371vampire bats 403Vampyroteuthis infernalis 171vanes (feathers) 85Varanus komodoensis 457variability 431–435vascular system 29vascular tissue 472–473, 477vectors 404, 405, 417vegetarian diets 209–210vegetation zones 317mveins 254, 375Velcro 83Velociraptor 156velocity 465
velvetleaf 330, 356, 501velvet worms 28venom 17, 456–57venomous spiders 496, 498Venter, J. craig 171ventral surface 20, 34Venus flytrap 106, 438Verbenaceae 145Verkoyansk, Siberia 92Vermeij, Geerat 52vertebrae 458vertebrate amphibians 147vertebrate health 378–379vertebrates
animals 20, 29birds 84–90carboniferous period, biodiversity in
105–106cladistics 110cretaceous period, biodiversity in 154,
155Devonian period, biodiversity in
182–184fishes 244–251Jurassic period, biodiversity in 341neogene period, biodiversity in 392,
393Ordovician period 397Paleogene period, biodiversity in 399,
400Permian period, biodiversity in 412Quaternary period, biodiversity in
452Silurian period, biodiversity in 488triassic period, biodiversity in 523–
524vestimentiferan worm 122Vibrio fisheri 41vicariance 73–75, 194, 195viceroy butterfly 437Victoria, lake 6–7, 332Vietnam 56, 533vinblastine 66, 67, 527vincristine 66, 67, 527vinegar flies 324vinegaroons 36vines 124Viperidae 457vipers 457viral encephalitis 324Virgil 13, 63virgin forest 500Virginia creeper 187Virginia opossums 354, 362viruses 114, 231, 402–404vision 363, 366, 441Vitaceae 187vitamin c deficiency 58vitamins 378–379Vitis 187Vitis rotundifolia 60, 187Vitis vinifera 60Vitousek, Peter 72vivipary 371, 479, 538volcanic ash 531volcanoes
adaptive radiation 7cretaceous extinction 151–152deep oceans, biodiversity in 172greenhouse effect 285mass extinctions 374Permian extinction 409succession, ecological 506
voles 481Volk, tyler 267Vonnegut, Kurt 165Vostok ice core 282, 283Voyage of HMS Beagle, The (Darwin) 163,
522Vulpes vulpes 330vultures 89, 132
xviii+598_Biodiversity_F.indd 597 11/17/11 1:13 PM

first entry598 tundra 598 first entry598 Index598
wWagner, Warren 109Walden, or Life in the Woods (thoreau) 521wallabies 363Wallace, alfred Russel 542–545, 543, 548c
Bates, Henry Walter 45biodiversity, patterns of 50biogeography 72Darwin, charles 165natural selection 384tropical rain forests and deforestation
527Wallace’s line 527, 543Wallin, ivan e. 368, 508walnuts 113, 186, 427Walsh, Benjamin 493Walsh, christopher 67, 68Waltheria ovata 169wandering spider 498warblers 405, 494, 533Ward, Peter 223, 268, 373, 374warm-blooded animals. See homeothermsWashingtonia filifera 179, 195wasps
arthropods 38camouflage and mimicry 99coevolution 120, 121convergence 145cretaceous period, biodiversity in 155insects 325–326natural selection 389pollination 420spiders 498
wastewater 466“Was the World Made for Man?” (twain)
544water balance 86water bears 27water birds 89water boatmen 467water chestnut 377water cycle. See hydrological cyclewater-holding frog 180water hyacinth 84, 332–333, 372, 467water lettuce 332–333, 372, 467water molds 446water oaks 185, 185water ouzel 466water pressure 171water resources 242water scorpions 467watersheds 129water storage 177water striders 467water temperature 465water vapor 287, 311, 313water vascular system 29water velocity 465wax myrtle 333weapons 480web(s) 497webbed feet 456Wedgwood, emma 163, 164Wedgwood, Josiah 161, 162weeds xi
agriculture 12–14deserts, warm 180life history, diversity of 349–351population biology 426resistance, evolution of 459–460succession, ecological 501
weevils 325Weinberg, W. 433Weis, arthur 389Weller, Steve 492Wells, W. c. 384Welwitschia mirabilis 180werewolf 264West africa 527West nile virus 404Westoby, Mark 484
wetland animals 370wetland oxygen depletion 13wetland plants 371, 371tWeyerhauser corporation 239whales 111, 124, 171, 365, 365, 399whale shark 246wheat 10, 12, 58, 60, 263, 310wheatgrass 61whip scorpions 36, 436whirligig beetles 467whisk ferns 476whistling spider 181White, Gilbert 162White, lynn, Jr. 224, 225whitebark pines 136, 137, 297, 540white blood cells 353white firs 502, 511white oaks 6, 7, 73, 185, 308, 309white pines 511white spruce 76, 196white-tailed deer 338, 518Whittaker, Robert 55, 125Wiener, norbert 55wild barley 10wild cattails 61wild coffee trees 60wild donkeys 173Wilderness act 549cWilderness areas 242wilderness preservation 378wildfires
boreal and subalpine forests 92carboniferous period, biodiversity
in 103deserts, cool 173disturbances, ecological 198–201greenhouse effect 286, 287keystone species 343temperate dry forests 517tropical rain forests and deforestation
534tundra 540
wildflowers 177, 199adaptation 5–6communities, ecological 128deserts, warm 177dicots 186disjunct species 194–195disturbances, ecological 198neogene period, biodiversity in 393shrublands 487speciation 491tundra 538
wild grains 10wild grapes 60, 187wildlife management 347wild mustard 12wild oats 12, 278wild plant species 57–60wild pollinators x, 421wild tobacco 419, 420wild type 430wild wheat 10, 60wild wheatgrass 61will-o’-the-wisp 370willow bark 66willows 74, 186, 505, 538Willson, Mary 483Wilson, edward O. x, 57, 225–228, 235,
247, 335, 336, 545–547wind dispersal 253, 470wind pollination
coevolution 119–120conifers 135convergence 144flowering plants 253monocots 376pollination 417temperate deciduous forests 513tropical rain forests and deforestation
526
wine industry 60wings 37, 147, 320, 321, 365–366witchcraft 264witchweed 403within-habitat diversity 48Woese, carl R. 32–33, 508, 549cWolbachia 203, 404Wolffia 467wolf spiders 498, 498Wollemia nobilis 51, 136Wollemi pine 51, 136, 477wolves 46, 78, 220, 308, 344, 363wombats 363wood 253woodpecker finch 134, 169woodpeckers 89–90, 134wood pores 350twoolly adelgid bugs 2World War ii 149, 263worms 24, 26–29, 31, 122, 172, 402, 403,
467Wrangel island 75Wright, angus 109, 422Wright, Sewall 434Wright, Shane 49wrist 16Wyoming 220, 511
xXenopsylla cheopsis 38, 323Xenopus laevis 18Xenoturbellida 28xiphosuran arthropods 46Xylaria 264xylem 183, 254, 255, 472, 477Xyphosura 37
yyakushima island 137yaws 404yeast 262, 380yeast infections 262yellow-billed oxpeckers 122yellow fever 324yellowstone national Park 220, 239, 537,
548cyellowstone to yukon conservation
initiative 302yews 66, 67, 136yosemite national Park 239, 345, 378yosemite Valley 377–378younger Dryas 285yucatán rain forest 527yucca 175
zZahavi, amotz 480Zaire 56Zea diploperennis 234Zea mays 12, 234zebra mussels 26, 330zebras 98Zhang, Xiaoyang 296zoo animals 159, 234zoonotic diseases 404zooplankton 345, 469zoospores 447zooxanthellae 122, 149, 447zosterophylls 473Zuk, Marlene 480Zygochloa paradoxa 180Zygomycota 261Zygophyllaceae 180zygosporangium 261zygote 473, 474, 477
xviii+598_Biodiversity_F.indd 598 11/17/11 1:13 PM





























