Embed Size (px)
Citation preview

Biochemical Systematics and Ecology, Vol. 19, No. 8, pp. 629-642, 1991. 0305-1978/91 $3.00 + 0.00 Printed in Great Britain. © 1991 Pergamon Press plc.
Effect of some Leaf Essential Oil Phenotypes in Coastal Redwood on the Growth of several Fungi with Endophytic Stages
FRANCISCO J. ESPINOSA-GARCiA* and JEAN H. LANGENHEIMt Department of Biology, University of California Santa Cruz, CA 95064, U.S.A.
Key Word Index--Sequoia sempervirens ; Phomopsis occulta ; Pestalotiopsis funerea ; Cryptosporiopsis abieEna ; Botrytis cinema; Seiridium juniperi; Mefia parkeri; essential oil phenotypes; leaf fungus endophytes; symbiosis.
Abstract--Experiments assessed the effect of four foliar essential oil phenotypes from a coastal redwood (Sequoia sempervirens) population on isolates of six fungus species with leaf endophytic stages. The hypothesis tested was that leaf essential oil phenotypes would have differential effects in vitro on these endophytic species. Another objective was to determine whether differences in response to redwood essential oils existed among pathogens isolated from redwood and a mutualistic endophyte never isolated from redwood. These species were: Botr)ms cinema, an opportunist generalist pathogen able to attack redwood; Pestalotiopsis funerea, Phornopsis occulta and Seiridiurn juniper~ actual or potential conifer pathogens isolated as leaf endophytes in redwood; Cryptosporiopsis abietina, a common conifer endophyte with uncertain ecological status isolated from redwood; and Meria parken; a mutalistic endophyte known only from Douglas fir. The four essential oil phenotypes were uniformly inhibitory for some species and differ- entially so for others. Susceptibility to the four phenotypes varied widely within and among fungus species. The conifer-specific pathogens were the least susceptible and the Douglas fir endophyta the most; the other two displayed intermediate susceptibility. The diversity of responses by these fungus species suggests that these redwood terpenoids may have a differential intra- and interspecific importance in preventing pathogenic activity in the species found within redwood foliage.
Introduction Terpenoids are present in many plants as complex mixtures (i.e. resins, essential oils) that can be accumulated in the plant before or after the attack of herbivores or pathogens (Gibbs, 1968; Raffa and Bern/man, 1982; Gambliel et al., 1985; Miller et aL, 1986). Although the composition of these complex mixtures is under tight genetic control in conifers and other trees (Hanover, 1966; Von Rudloff, 1975; Langenheim, 1984), it may vary ontogenetically within the individual, and within and among populations of the same species (Langenheim and Stubblebine, 1983; Wagner et al., 1989; Hall and Langenheim, 1986a, 1986b, 1987). The variability of composition of resins or essential oils among individuals tends to be discrete in that few compositional types or chemical profiles may be present within and among plant populations (Langenheim and Stubblebine, 1983; Squillance et al., 1985; Leather et al., 1987).
Terpenoids are well known for their activity against fungi (e.g. Hintikka, 1970; De Groot, 1972; V~is~la, 1974; Benjalili et al., 1984; Bridges, 1987; Pauli and Knobloch, 1987; Ennos and Swales, 1988), and have been associated with pathogen resistance in conifers (Rockwood, 1973, 1974; Thibault-Balesdent and Delatour, 1985; Michelozzi eta/., 1990). In spite of the variability of resins and essential oils, and the fact that fungi encounter these mixtures in nature, relatively few studies have investigated the
*Present address: Centro de Ecologia, Universidad Nacional Autbnoma de M~xico, Ciudad Universitaria, 04510, M~xico, D.F. Mexico.
tAuthor to whom correspondence should be addressed.
(Received 15 May 1991)
629

630 F.J. ESPINOSA-GARdA AND J. H. LANGENHEIM
effects of complete resins or essential oils on fungi interacting with the producer plant (Cobb eta/., 1968; Shrimpton and Whitney, 1968; Flodin and Fries, 1978; Franich etaL, 1982; Arrhenius and Langenheim, 1983; Raffa et aL, 1985).
Fungi with endophytic stages producing unapparent infections in aerial tissues are ubiquitous in conifers (Carroll, 1986; Petrini, 1986). They have even been proposed to be "as common among plants as are mycorrhizae" (Carroll, 1988). Although some of these organisms are latent pathogens, many are symbionts that do not cause apparent harm to the plant in their endophytic stages and can be isolated from symptomless tissues. The ecological roles of fungi with endophytic stages can be pathogenic, saprobic or mutualistic (Verhoeff, 1974; Carroll, 1988, 1990; Chapela and Boddy, 1988). In this paper, the term "endophyte" is used to refer to these fungi.
Many leaf endophytes in conifers are thought to remain inactive or apparently inactive after colonization; active fungus growth and reproduction apparently occur when the leaf senesces or is damaged (Carroll, 1986). The mechanisms by which this apparent inactivity is maintained and by which growth is resumed are not known, although mechanisms controlling latent fungus infections, such as secondary chemicals of the host, changes in the nutritional quality of the tissues or a combi- nation ef both factors (Verhoeff, 1974), may be similar for all endophytes.
Both endophytes and essential oils are ubiquitous in leaves of coastal redwood [Sequoia sempervirens (D.Don ex Lamb) Endl.] (Gregonis et aL, 1968; Okamoto et al., 1981; Hall, 1985; Espinosa-Garcia and Langenheim, 1990). Many species of leaf endophytes have been isolated from redwood in its natural range (Rollinger and Langenheim, 1989; Epsinosa-Garcia and Langenheim, 1990) and the variability of the monoterpene fraction of redwood essential oils is well documented (Hall and Langenheim, 1986a, 1986b, 1987). The variability in the monoterpene fraction of leaf essential oils tends to be discrete in redwood, where five compositional patterns were found in a sample of 36 individuals of a population (Espinosa-Garcia, 1991). Because monoterpenes constitute 60-80% of the redwood essential oil (Gregonis eta/., 1968; Hilal et aL, 1977; Okamoto eta/., 1981), these compositional patterns are called "essential oil phenotypes" here. Four of these phenotypes inhibited differently the growth of the suspected redwood mutualist endophyte Pleuroplaconema sp. (Espinosa-Garcia and Langenheim, 1991).
The purpose of this study was to explore some relationships between four essential oil phenotypes and endophytes in redwood leaves. The hypothesis tested was that leaf essential oil phenotypes would have different effects in vitro on six endophytic species. Another objective was to determine whether differences in response to redwood essential oils existed among pathogens isolated from redwood (conifer-specific or generalists), and a mutualistic endophyte never isolated from redwood.
Each of the four leaf essential oil phenotypes in this study (Fig. 1) is a representative of compositional patterns found in a Santa Cruz redwood population. These patterns were characterized by five predominant monoterpenes (sabinene, limonene, ~-phellandrene, y-terpinene and a-pinene), although the sesquiterpene caryophyllene and eight other monoterpenes were included. Sabinene is the pre- dominant compound in three of the groups, followed by similar but characteristic patterns of predominance of either limonene, y-terpinene, ~-phellandrene or ~- pinene (Phenotypes I, II and IV). In the group represented by Phenotype III, ~- phellandrene is the predominant monoterpene, followed by similar concentrations of sabinene, limonene and ~-pinene. There is a relatively high amount of caryophyllene in Phenotype I.
The leaf endophytes tested were: Botrytis cinerea (Fr.) Pers., an opportunist generalist pathogen that can attack redwood and can form latent infections in a number of plants (Verhoeff, 1974); Pestalotiopsis funerea (Desm.) Stey., Phomopsis

ESSENTIAL OILS IN COASTAL REDWOOD 631
occulta Tray. and SeMdium juniperi (All.) Sutton, actual or potential conifer pathogens isolated as leaf endophytes in redwood; Cryptosporiopsis abietina Petrak, a common conifer endophyte with uncertain ecological status, isolated from redwood leaves; and Meria state of Rhabdocline parkeri Sherwood-Pike, Stone and Carroll, a mutualistic endophyte known only from Douglas fir. The frequent species, C. abietina, R funerea and Ph. occulta were represented in the experiment with five isolates each, due to the potential infraspecific variability of fungi in response to terpenoids (De Groot, 1972; Thibault-Balesdent and Delatour, 1985; Ennos and Swales, 1988; Espinosa-Garcia and Langenheim, 1991). Only one isolate per species was available for the other endophytes that are infrequent or non-existent as redwood endophytic symbionts (Espinosa-Garcia and Langenheim, 1990).
Materials and Methods Essential oil extracdon. Four trees with distinctive essential oil phenotypes (Fig, 1 ), were selected in a coastal redwood population of Santa Cruz, California, U.S.A., as source of essential oils. Foliage from each tree was collected in the second week of November, 1989. Leaves one to three years old were detached from twigs and then extracted using two methods: one that does not modify the original terpenoid profile, and another of larger scale that can modify this profile. In the srnall-scale extraction, leaves (2 g per tree) were extracted by grinding in pentane with tetradecane added as internal standard. The extract was dried with sodium sulfate, concentrated to 0.1 ml under a rniid nitrogen stream, and analysed by GC. The terpenoid profiles were compared with those obtained in the larger scale extraction procedure. In the latter, leaves (300-400 g per tree) were put in jars, and soaked in pentane for five days at room temperature in the dark. Each extract from soaked leaves was fractionate-distilled, the residue saved, and the distillate put back in the jars where the soaked leaves remained. This procedure was repeated three times. Each time the residue was stored in amber bottles in the freezer. After the fourth distillation, distillate and liquid from cold traps were combined (ca. 1 I ), dried and concentrated to 50 ml under a mild nitrogen stream. The concentrate was added to the previously collected residues; this, in turn, was dried and distilled to remove pentane. Pentane at concen- trations higher than 0.17 mg pentane ml 1 air decreases the growth of some endophytic fungi (D. Schneck, unpublished results). Complete removal of pentane caused loss of some monoterpenes; therefore, they were replaced with commercially prepared compounds (Aldrich, Co. and ICN Chemicals). The more than 95%
Phenotype I 301
lO
0 A B C D E F G N I J K L M N
Phenotype II
20
10
0 A B C D E F G H I J K L M N
Phenotype III
~" 0 A B C D E F G H i J K L M N
Phenotype IV
20
1
A B C D E F G H I d K L M N
Compound Compound FIG. 1. ESSENTIAL OIL PHENOTYPES USED IN THE EXPERIMENT. Bars represent per cent of individual compounds relative to the total amount of monoterpenes plus caryophyllene. Each terpenoid profile was obtained from an individual representing an essential oil compositional pattern group. Key: A, tricyclene; B, thujene; C, ct-pinene; D, camphene; E, {~-pinene; F, sabinene; G, myrcene; H, a-phellandrene; I, limonene; J, {3-phellandrene; K, y-terpinene; L, p-cymene; M, terpinolene; N, caryophyllene.

632 F. J. ESPINOSA-GARCIA AND J. H. LANGENHEIM
purity of these chemicals was confirmed by GC. Because ~-phellandrene was not available commercially, the remaining amount of this compound in the extracts was used as a base to adjust the relative proportions of the other monoterpenes to simulate the original pattern. These reconstituted concentrates were analysed with GC and their patterns compared with those obtained with the small quantities of leaves. No significant differences were found in the monoterpene fraction, and the pentane concentration was below the inhibitory level. Pentane and volatile terpenoids in the extracts were quantified by adding 5 ~1 of 99% tetradecane to 60 ~1 of extract in a tightly closed mini-vial. After mixing thoroughly, an aliquot was injected and analysed by GC.
Gas chromatographic analysis. A gas chromatograph with glass capillary column 0.9 mm i.d. and 86 m long coated with 12% Carbowax 20 on Chromosorb and flame ionization detector was used. Injections were split 40/1 ratio with He as carrier gas at a 2 ml s ~1 flux, with the following temperature program: 65°C held for 5 min, 6°C min 1 increase up to 180°C and held for 10 rain. Injector temperature 210°C, detector temperature 250°C. Monoterpenes and caryophyllene were identified by retention time and the peak enrichment method.
Fungus isolates. Five isolates of Cryptosporiopsis abietina and Pestalotiopsis funerea and four of Phomopsis occulta were isolated from redwoods growing in the same forest where the tree sources of the essential oils were selected. The isolation procedure used was as follows: leaves one to three years old were collected and placed separately on water-agar slant tubes following surface sterilization with serial immersion in 95% EtOH for 1 min, 65% household bleach for 10 min and 35% EtOH for 30 s ×2. The tubes were incubated at room temperature (20-25°C) for up to three months until fungi sporulated and were identified. When a tube had one of the selected species, spores were transferred to redwood broth agar [macerated redwood foliage extract (equivalent to 50 g foliage I 1), filtered through Whatman 1 paper and autoclaved; filter-sterilized glucose (5 g I=1), ampicillin (100 mg I-1), streptomycin (25 mg 1-1) and autoclaved water agar (15 g I 1) were added]. Tubes were incubated at room temperature. Isolates representing one species were always selected from different trees. An additional Phomopsis occulta isolate was obtained from surface-sterilized redwood seeds collected in Mount Madonna from the Santa Cruz mountains. All representatives of these species were isolated and identified by F.J.E. Seiridiumjuniperiwas isolated as redwood leaf endophyte from Hendy Woods State Park, California by Jeanatte Rollinger; Botrytis cinema was isolated by Dr Arthur McCain growing pathogenically on Pseudotsuga mensiezit~ This isolate is also pathogenic on redwood cuttings and seedlings (Triplett, 1990; Espinosa-Garcia and Rollinger, unpublished results]; and Meria parkeri was isolated as leaf endophyte from Pseudotsuga mensiezii from Santa Cruz by F.J.E. and identified by Dr G. C. Carroll.
All isolates were transferred to redwood broth agar. These cultures were used as inoculum source for colonies which provided plugs from colony edges for tests.
Experimental design and statistical analysis. The experiment had a nested factorial design 5 x 18 (extract type by isolate). All species were exposed to volatile terpenoids from all essential oil phenotypes. In the control, treatment isolates grew with no volatiles. All treatments had three replicates per isolate. Results were analysed using analysis of variance (ANOVA), and means were compared with Duncan's multiple range test. Comparisons among species were performed with ANOVA on the arcsin transformed per cent growth of control response of the isolates to essential oils. Separate ANOVAs were performed for each species with untransformed data.
Statistical analyses were carried out using SAS statistical package (SAS Institute Inc., Cary, NC, U.S.A.). Procedure. Plugs (4 mm diameter) of actively growing colonies were inoculated into 7-ml glass vials with
ca. 1 ml of redwood broth medium that solidified when the vial was horizontal. A filter paper disk (7 mm diameter) was put on the inside of the vial's screw cap, 10-20 I~1 of extract dispensed on the paper and the vial was immediately closed very tightly. When applicable, the vials received ca. 5.0 mg of volatile terpenoids (Table 1), enough to initially saturate the vial's atmosphere. Once the vials were closed, they were grouped according to species and treatment and put in tightly closed 60-ml jars sealed with parafilm.
Cultures were incubated at 25°C in the dark until measurement. Botrytis cinerea was incubated for five days because of its fast growth rate. Meria parkeri was measured after 30 days, because the growth of the control was minimal after 15 days. All the other species were incubated for 12 days. Linear growth was measured to the nearest 0.5 mm under a dissecting microscope.
Results Al l r e d w o o d essent ia l oil p h e n o t y p e s s h o w e d d i f ferent ia l i nh ib i to ry ac t i v i t y on e n d o p h y t i c l inear g r o w t h (Tables 2-4) . Overal l , P h e n o t y p e I w a s the least inh ib i tory , a l t hough its e f fect fo r s o m e e n d o p h y t e spec ies w a s equa l l y as s t rong as those o f o the r p h e n o t y p e s (Table 4, Fig. 2). P h e n o t y p e II w a s the m o s t i nh ib i t o ry fo r all isolates, e x c e p t fo r t hose o f Phomopsis occulta. The o the r t w o p h e n o t y p e s e l ic i ted an inter- m e d i a t e f ungus response w i t h con t ras t ing ef fects on s o m e species. P h e n o t y p e III had m i l d - to -ave rage inh ib i to ry ac t i v i t y aga ins t Pestalotiopsis funerea, and a s t ronger e f fect on Seiridium juniper~ w h e r e a s P h e n o t y p e IV had a reversed ac t i v i t y pa t te rn on these species.
E n d o p h y t e s s h o w e d w i d e intra- and in terspec i f ic va r iab i l i t y in g r o w t h response to

ESSENTIAL OILS IN COASTAL REDWOOD
TABLE 1. CONCENTRATION (rag/VIAL) OF MOST MONOTERPENES, CARYOPHYLLENE AND OTHER VOLATILE COMPOUNDS DELIVERED IN THE EXTRACTS FROM THE REDWOOD FOLIAGE USED IN THE EXPERIMENT
Tree 1 Tree 2 Tree 3 Tree 4
Tricyclene 0.004 0.000 0.003 0.002 Thujene 0.029 0.014 0.026 0.025 ~-Pinene 0.383 0.313 0.439 0.521 Camphene 0.079 0.019 0.016 0.017 ~-Pinene 0.021 0.000 0.038 0.003 Sabinene 0.758 0.827 0.426 0.756 Myrcene 0.350 0.177 0.350 0.217 (~-Phellandrene 0.033 0.021 0.025 0.020 Umonene 0.529 0.492 0.442 0.437 ~-Phellandrene 0.479 0.351 0.686 0.154 ?-Terpinene 0.192 0.386 0.181 0.615 P-Cymene 0.183 0.177 0.056 0.121 Terpinolene 0.096 0.088 0.114 0.125 Caryophyllene 0.462 0.076 0.086 0.079 Monoterpene total 3.137 2.865 2.801 3.013 Other compounds* 1.937 2.138 2.193 1.984
*Total of sesquiterpenes, oxygenated terpenoids and other minor components.
633
TABLE 2. RESULTS OF TWO-WAY ANALYSES OF VARIANCE APPLIED TO ENDOPHY'rlC GROWTH AFTER EXPOSURE TO VOLATILES FROM FOUR REDWOOD ESSENTIAL OIL PHENOTYPES AND A CONTROL WITH NO OIL (TREATMENTS)
Cryptosporiopsis abietina
Source of variation df V% MS F Isolates (I) 4 3.0 6.5 13.4 Treatments (T) 4 84.2 181.8 376.4 I × T 16 10.1 5.4 11.3 Residual 42 0.5
Pestalobbpsis funerea
Source of variation df V% MS F Isolates (I) 4 26.3 133.1 113.1 Treatments (3") 4 63.1 319.5 271.6 I × E 16 7.8 9.8 8.4 Residual 50 1.2
Phomopsis occuffa
Source of variation df V% MS F Isolates (I) 4 20.3 41.1 29.6 Treatments (1") 4 45.9 92.9 67.0 I × E 16 25.2 12.8 9.19 Residual 50 1.4
Five isolates per endophyte species were tested. V%= percentage of variance accounted for by source of variation. P<0.0001.
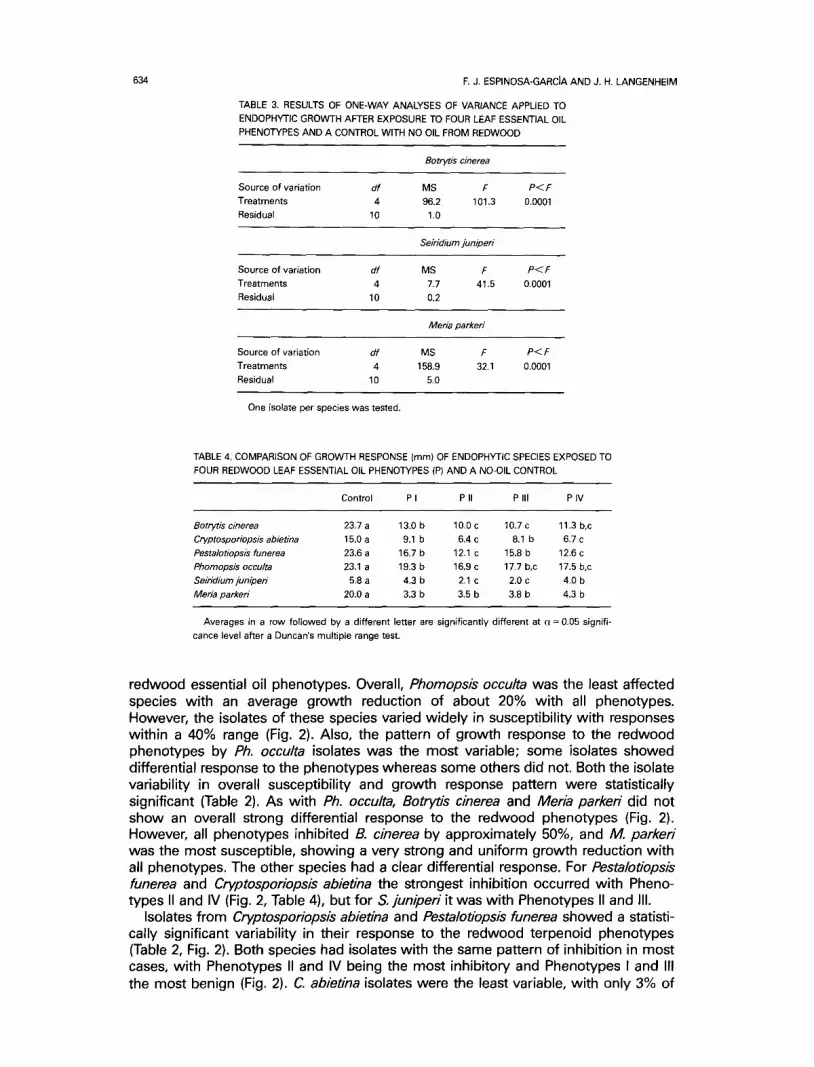
634 F. J. ESPINOSA-GARCJA AND J. H. LANGENHEIM
TABLE 3. RESULTS OF ONE-WAY ANALYSES OF VARIANCE APPLIED TO ENDOPHY-I'IC GROWTH AFTER EXPOSURE TO FOUR LEAF ESSENTIAL OIL
PHENOTYPES AND A CONTROL WITH NO OIL FROM REDWOOD
Botrytis cinerea
Source of variation df MS F P < F Treatments 4 96.2 101.3 0.0001 Residual 10 1.0
Seiridium juniperi
Source of variation df MS F P < F Treatments 4 7.7 41.5 0.0001 Residual 10 0.2
Meria parkeri
Source of variation df MS F P< F
Treatments 4 158.9 32.1 0.0001 Residual 10 5.0
One isolate per species was tested.
TABLE 4. COMPARISON OF GROWTH RESPONSE (mm) OF ENDOPHYTIC SPECIES EXPOSED TO
FOUR REDWOOD LEAF ESSENTIAL OIL PHENOTYPES (P) AND A NO-OIL CONTROL
Control P I P II P III P IV
Botrytis cinerea 23.7 a 13.0 b 10.0 c 10.7 c 11.3 b,c Cryptosporiopsisabietina 15.0a 9.1 b 6.4c 8.1 b 6.7c Pestalotiopsls funerea 23.6 a 16,7 b 12.1 c 15.8 b 12.6 c
Phomopsis occulta 23.1 a 19.3 b 16.9 c 17.7 b,c 17.5 b,c Seiridium juniperi 5.8 a 4.3 b 2.1 c 2.0 c 4.0 b
Meria parkeri 20.0 a 3.3 b 3.5 b 3.8 b 4.3 b
Averages in a row followed by a different letter are significantly different at a = 0.05 signifi- cance level after a Duncan's multiple range test.
redwood essential oil phenotypes. Overall, Phomopsis occulta was the least affected species with an average growth reduction of about 20% with all phenotypes. However, the isolates of these species varied widely in susceptibility with responses within a 40% range (Fig. 2). Also, the pattern of growth response to the redwood phenotypes by Ph. occulta isolates was the most variable; some isolates showed differential response to the phenotypes whereas some others did not. Both the isolate variability in overall susceptibility and growth response pattern were statistically significant (Table 2). As with Ph. occulta, Botrytis cinerea and Meria parkeri did not show an overall strong differential response to the redwood phenotypes (Fig. 2). However, all phenotypes inhibited B. cinerea by approximately 50%, and M. parkeri was the most susceptible, showing a very strong and uniform growth reduction with all phenotypes. The other species had a clear differential response. For Pestalotiopsis funerea and Cryptosporiopsis abietina the strongest inhibition occurred with Pheno- types II and IV (Fig. 2, Table 4), but for S. juniperi it was with Phenotypes II and Ill.
Isolates from Cryptosporiopsis abietina and Pestalotiopsis funerea showed a statisti- cally significant variability in their response to the redwood terpenoid phenotypes (Table 2, Fig. 2). Both species had isolates with the same pattern of inhibition in most cases, with Phenotypes II and IV being the most inhibitory and Phenotypes I and III the most benign (Fig. 2). C. abietina isolates were the least variable, with only 3% of
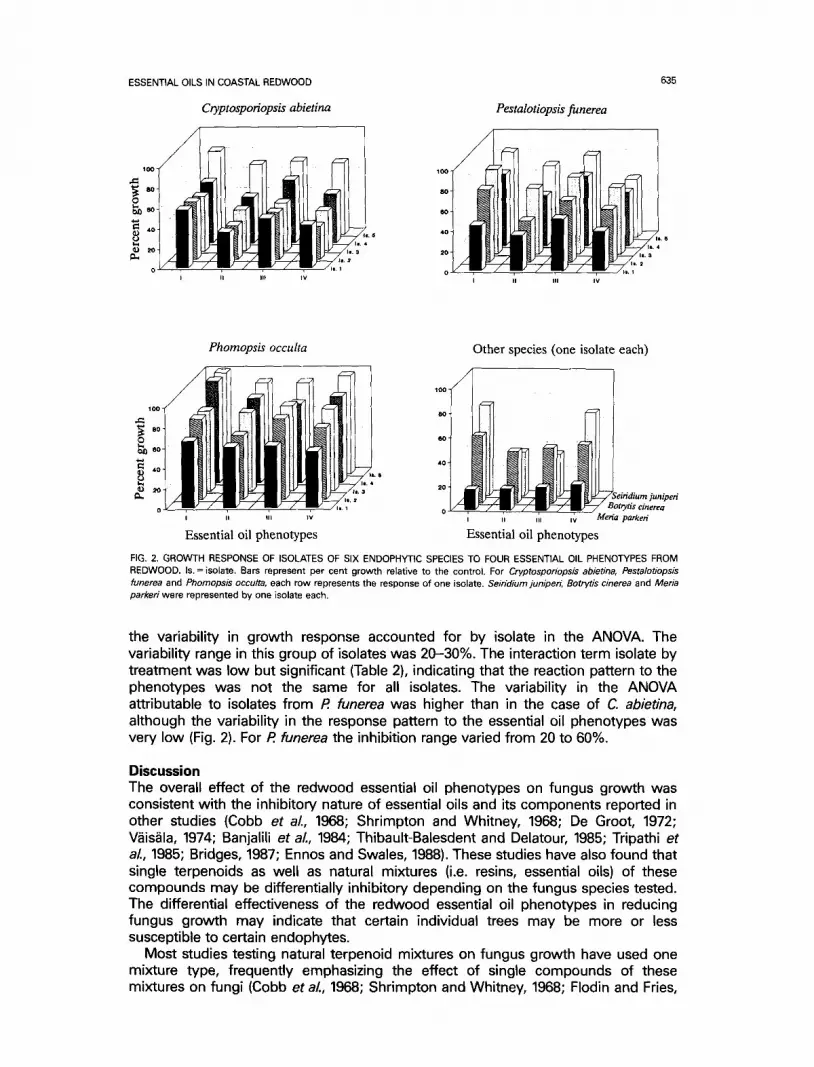
ESSENTIAL OILS IN COASTAL REDWOOD
Cryptosporiopsis abietina
0 V - T - - e - 7 " - e --7-- e " 7 - - ~ t , . l
Pestalotiopsis funerea
4 0
2 0
0 I
l l . a
I I I I I IV
635
Phornopsis occulta
• . ~ I0O~ '°i Z ltll
• °t I 1ttll L 6 I i , 4
tin, 3 Is, It
I I I I V
Essential oil phenotypes
Other species (one isolate each)
4O
2O
eiridium juniperi o , , ~ / Botrye's cinerea
. . i iv Meria parked
Essential oil phenotypes
FIG. 2. GROWTH RESPONSE OF ISOLATES OF SIX ENDOPHYTIC SPECIES TO FOUR ESSENTIAL OIL PHENOTYPES FROM REDWOOD. Is. = isotate. Bars represent per cent growth relative to the control. For Cryptosporiopsis abietina, Pestalotiopsis funerea and Phomopsis occulta, each row represents the response of one isolate. Seiridium juniper~ Botrytis cinerea and Meria parkeri were represented by one isolate each.
the variability in growth response accounted for by isolate in the ANOVA. The variability range in this group of isolates was 20-30%. The interaction term isolate by treatment was low but significant (Table 2), indicating that the reaction pattern to the phenotypes was not the same for all isolates. The variability in the ANOVA attributable to isolates from R funerea was higher than in the case of C. abietina, although the variability in the response pattern to the essential oil phenotypes was very low (Fig. 2). For R funerea the inhibition range varied from 20 to 60%.
Discussion The overall effect of the redwood essential oil phenotypes on fungus growth was consistent with the inhibitory nature of essential oils and its components reported in other studies (Cobb et al., 1968; Shrimpton and Whitney, 1968; De Groot, 1972; V~is~la, 1974; Banjalili et al., 1984; Thibault-Balesdent and Delatour, 1985; Tripathi et a/., 1985; Bridges, 1987; Ennos and Swales, 1988). These studies have also found that single terpenoids as well as natural mixtures (i.e. resins, essential oils) of these compounds may be differentially inhibitory depending on the fungus species tested. The differential effectiveness of the redwood essential oil phenotypes in reducing fungus growth may indicate that certain individual trees may be more or less susceptible to certain endophytes.
Most studies testing natural terpenoid mixtures on fungus growth have used one mixture type, frequently emphasizing the effect of single compounds of these mixtures on fungi (Cobb et al., 1968; Shrimpton and Whitney, 1968; Flodin and Fries,

636 F. J. ESPINOSA-GARCIA AND J. H. LANGENHEIM
1978; Franich et aL, 1982; Tripathi et al., 1985). Although single terpenoids within natural mixtures appear to be solely responsible for growth inhibition in certain fungi (e.g. Arrhenius and Langenheim, 1983), intra- and interpopulation variability in terpenoid profile, naturally encountered by fungi, is a common feature in many terpene producing plants (Rhoades et al., 1976; Langenheim and Stubblebine, 1983; Hall and Langenheim, 1986a, 1986b, 1987; Squillance et al., 1985; Leather et aL, 1987). Furthermore, mixtures frequently contain more than one compound inhibitory to fungi (e.g. Bridges, 1987), and interactions among mixture components may play an important role when fungi or herbivores encounter plants (Chararas et aL, 1982). For example, comparable amounts of various Abies grandis single monoterpenes were inhibitory to the fungus Trichosporium symbioticum, but a comparable amount of total resin was the most inhibitory, indicating synergistic effects or the action of other non- monoterpene compounds (Raffa et al., 1985). The essential oils in redwood had a diverse effect on the endophytes although the distinctive monoterpene profiles were relatively similar. Thus, the differential effects of the phenotypes might be attributed to either a combination of compounds or to the effect of other volatile terpenoids in the mixture that were not identified and quantified singly.
Which compound or combination of compounds in the essential oils was the cause for the differential activity of the phenotypes was not evident. The proportion of sabinene and 3'-terpinene was higher in both Phenotypes IV and II, which produced the highest overall inhibition of the phenotypes. These two monoterpenes were equally inhibitory in single compound trials and acted in an additive fashion on endophytes in a separate experiment (Espinosa-Garcia, 1991). However, when the activities of Phenotypes IV and II are examined in each species, their effects are sometimes comparable with those of other phenotypes. The high proportion of ~-phellandrene in Phenotype III might suggest that this compound is important in reducing growth in Seiridium juniper~ although the combined effect of this mono- terpene and others in Phenotype III is also possible. The extrapolation of results obtained with trials with single compounds of a mixture may not be appropriate to explain growth inhibition with complete mixtures for certain endophytic species. Most phenotypes were mildly inhibitory to Phomopsis occulta, which was the most tolerant endophyte in this experiment. However, this fungus showed intermediate tolerance to sabinene and y-terpinene, alone and in mixtures, in a separate experiment when compared to the endophytes tested here (Espinosa-Garcia, 1991). A relevant finding is that relatively similar but distinctive redwood essential oils occurring naturally had a differential activity on endophytes. This suggests that essential oil variability is important in redwood-endophyte interactions.
Phytopathological studies involving terpenoids and fungi have frequently tested the effect of these compounds on one or few isolates per fungus species (Cobb et al., 1968; Shrimpton and Whitney, 1968; Hintikka, 1970; V~is~la, 1974; Flodin and Fries, 1978; Franich et al., 1982; Tripathi et al., 1985; Bridges, 1987). A disadvantage of this approach is that different strains of the same fungus species from the same or different geographic locations may vary significantly in response to terpenoids (De Groot, 1972; Thibault-Balesdent and Delatour, 1985; Ennos and Swales, 1988; Espinosa-Garcia and Langenheim, 1991). Isolates of Cryptosporiopsis abietina, Pestalotiopsis funerea and Phomopsis occulta in this study also vary significantly. However, fungus pathogens may not be highly variable within a locality, e.g. epidemic infection fronts of Ophiostorna ulm~ the agent of the Dutch Elm Disease, are charac- terized by one or two genotypes of the fungus (Brasier, 1988). Thus, single isolate trials may still provide useful information, and results of this experiment with Botrytis cinema, Meria parkeri and Seiridium juniperi could indicate the response pattern of these species to the redwood phenotypes. However, a larger sample size is needed to determine the variability of response of these species to these essential oils.

ESSENTIAL OILS IN COASTAL REDWOOD 637
The endophytes did not show a uniform response as a group, but had a wide variability in susceptibility to redwood essential oils. Thus, the capability to live endophytically is probably a poor predictor of terpenoid tolerance. Instead, the tightness and type of the fungus-host association may be a better tolerance predictor, as well as a cue in elucidating the roles of terpenoids in these symbioses. A gradient of tolerance was apparent in this study, with the conifer-specific pathogens on or close to one extreme and a non-redwood endophyte on the other. Meria parkeri was the most inhibited by the essential oil phenotypes; it is known only as an endophyte in foliage of Pseudotsuga mensiezii (Sherwood-Pike et al., 1986), and it interacts there with a different terpenoid profile.
Excluding Meria parkeri, all the species were either isolated or reported as endophytes from redwood (Espinosa-Garcia and Langenheim, 1990) and redwood essential oils are not foreign to them. Yet, strong differences in the overall susceptibility to essential oils among redwood endophytes are apparent. Pleuro- placonema sp., the most frequent endophyte and suspected mutualist in redwood (Rollinger and Langenheim, 1989; Espinosa-Garcia and Langenheim, 1990), showed a higher susceptibility to the same redwood phenotypes than the endophytes studied here (Espinosa-Garcia and Langenheim, 1991).
Phomopsis occulta and Pestalotiopsis funerea, species found almost exclusively in conifers (Funk, 1981; Sinclair et aL, 1987), were the least susceptible to redwood essential oils. Both species are known as pathogens and probably can be pathogenic to redwood. Although Ph. occulta has been mentioned as a saprophyte (Farr et al., 1989), its pathogenicity to conifer seedlings as a leaf blight or canker-producing agent is well established (Sanderson and Worf, 1986; Kliejunas, 1988). Ph. occulta can also attack redwood seeds (Espinosa-Garcia, unpublished observation), and a Phomopsis sp. was partially responsible for redwood seedling mortality in natural conditions (Davidson, 1971). Pestalotiopsis funerea is frequently associated with cankers and is able to produce stem and leaf blights in seedlings (Hepting, 1971; Tuset Barrachina, 1972; Mordue, 1976; Affeltranger and Cordell, 1989) and in adult trees when they are not growing under optimal conditions (Luisi and Triggiani, 1977; Funk, 1981). R funerea can also attack redwood seeds (Davidson, 1971). Although the effects of Phornopsis occulta and Pestalotiopsis funerea in a natural redwood population might not be apparent, their potential effects on seedlings and debilitated individuals could be significant.
Seridium cardinale, a close relative of S. juniper~ is a serious leaf blight and canker- producing pathogen in Thuja and in other members of the Cupressaceae. The trees react by exuding resin around the canker wound, although generally this is not sufficient to impede the pathogen (Funk, 1981; Raddi and Paconesi, 1981). Also, a Seiridium (Coryneum) sp. produces twig and branch cankers in redwood, killing seedlings, saplings and young trees (Roy, 1966; Scharpf, 1978).
The relatively high tolerance of Phomopsis occulta, Pestalotiopsis funerea and Seiridiumjuniperi to redwood essential oil phenotypes may reflect their adaptation to the host defensive responses that involve terpenoids (e.g. resin exudation around wounds). However, the differential response of the two last species to the essential oils suggests that redwood individuals with certain phenotypes might be less susceptible to P. funerea and/or S. juniperi than individuals with other phenotypes. The essentially uniform response of Ph. occulta to the four phenotypes may indicate that this fungus may not be affected by the essential oil phenotype variability in redwood.
Botrytis cinerea, a generalist pathogen that can be a serious problem for redwood seedlings and cuttings in nurseries (McCain and Smith, 1978; Mittal et al., 1987), showed an intermediate and almost uniform tolerance to redwood essential oils. A similar inhibition occurred by exposing another isolate of B. cinerea to some redwood

638 F.J. ESPINOSA-GARCiA AND J. H. LANGENHEIM
monoterpenes incorporated into a culture medium (Hall, 1985). Stains of this species that attack woody plants do not show host specificity (Sinclair et al., 1987), and the almost uniform response of this isolate of B. cinerea to redwood essential oils might be representative of other strains. Here again, the B. cinerea-redwood relationship is that of pathogen-host, although probably not as specialized as that of the other endophytes considered here.
Cryptosporiopsis abietina displayed an overall susceptibility to the redwood essential oil phenotypes that was lower than those of all pathogens except B. cinerea. C. abietina has been mentioned as a weak pathogen in Abies (Sutton, 1980), and as secondary pathogen in damaged mycorrhizae of Picea abies (Haug et al., 1988). However, it occurs as an endophyte in various conifers (Carroll eta/., 1977; Sieber, 1989), where disease has not been attributed to it. Instead C. abietina has been reported as antagonist to the butt-rot fungus Heterobasidion annosum (Pratt, 1982) as well as the larch canker pathogen, Tricoscyphella willkommi (Buczaki, 1973). In both cases antagonism was deemed likely in vivo and demonstrated in vitro. Thus, C. abietina may be a fungus leaning towards the mutualistic side of the latent pathogen- mutualistic endophyte continuum proposed by Carroll (1988).
Mutualistic endophytism has been postulated to have evolved from latent pathogens because most proven endophytic mutualists are close relatives of virulent pathogens, and some otherwise harmless endophytes can cause disease when the plant is under stress (Carroll, 1988; Clay, 1988). The possibility of a relationship between increased specialization in endophytic life habit and loss of pathogenicity, with high susceptibility to the host secondary chemicals, has been proposed for the redwood suspected mutualist Pleuroplaconema sp. (Espinosa-Garcia and Langenheim, 1991). Cryptosporiopsis abietina appears to show a similar relationship: because its pathogenicity is weak, it has been found mostly as an endophyte and it showed a higher susceptibility to redwood terpenoids than the conifer-specific pathogens Phomopsis occulta, Pestalotiopsis funerea and Seiridium juniperL Also, it is similar in its overall susceptibility to redwood essential oils to that of Pleuroplaconema sp., which showed an overall inhibition of 65% with essential oil doses similar to those used here (Espinosa-Garcia and Langenheim, 1991).
Redwood essential oils have been proposed as an important factor controlling the apparent inactivity of Pleuroplaconema sp. (Espinosa-Garcia and Langenheim, 1991). However, the diversity of responses by the species studied here suggests that redwood essential oils may have a differential intra- and interspecific influence preventing pathogenic activity within redwood foliage.
Endophytes within a living redwood leaf are probably influenced by factors such as minerals, sugars, other secondary chemicals, etc. that could interact with essential oils. Although there is no evidence for such interactions in living leaves, Kurita and Koike (1982) showed that sodium chloride in the substratum enhanced the inhibitory activity of certain essential oil components against food-decomposing fungi. This suggests that agents stressing fungi under natural conditions may enhance fungus susceptibility to plant secondary chemicals. These potential interactions might enhance the effect of these terpenoids on endophytes in some cases and make an apparently ineffective essential oil effective against some fungi. For Phomopsis occulta and Pestalotiopsis funerea these chemicals may not be very important in preventing fungus growth acting alone at the doses tested; there are probably other factors influencing their apparent dormancy in their endophytic stage.
Plants produce various classes of compounds able to simultaneously affect fungi. Dothiostroma pin~ a foliar pathogen in Pinus radiata, was affected by both foliar volatile terpenoids and benzoic acid from its host (Franich eta/., 1982; Franich et al., 1986). Redwood leaves have several kinds of secondary chemicals, such as diterpene acids, bisflavones and proanthocyanidins (Miura and Kawano, 1968; Ohta and

ESSENTIAL OILS IN COASTAL REDWOOD 639
Nawamiki, 1978; Stafford and Lester, 1986). The redwood endophytes may also be affected by these compounds, or they may be inhibited more effectively by higher concentrations of essential oils.
This experiment was designed to expose endophytes to essential oil doses that fungi would encounter wi th in redwood leaves, either by the disruption of the resin canals or the vapors wi th in the leaf. The essential oil concentration in this experiment was sufficient to initially saturate the atmosphere of the vial where an isolate was exposed to the volatiles. However, leaks, compound degradation and/or absorption in the medium made this saturation ephemeral. Thus, the degree of fungus inhibit ion was probably lower than that which would be obtained by a constantly terpenoid saturated atmosphere (Espinosa-Garcia and Langenheim, 1991).
Total yield of monoterpenes in mature lower canopy leaves varies among redwood trees from 0.1 to 2.2% dry weight (Hall, 1985). Mature leaves from young basal sprouts have two to three times the yield of those of adult trees, and newly developed leaves 50% more compared with the mature leaves (Hall and Langenheim, 1986a). Thus, redwood endophytes are likely to find a wide spectrum of essential oil doses.
Pestalotiopsis funerea showed 80% growth inhibit ion by redwood essential oil doses twice as large as the ones tested here, i.e. wi th a saturated atmosphere held longer (Espinosa-Garcia, 1991). Also, single isolates of Botrytis cinema, Crypto- sporiopsis abietina, R funerea and Phomopsis occulta were totally or strongly inhibited by saturated atmospheres of ~,-terpinene or sabinene, which can be major con- stituents of redwood essential oils (Espinosa-Garcia, 1991). Thus, increased doses of redwood essential oil may augment the influence of these compounds on endo- phytes.
The large variabil ity in susceptibil ity to the essential oil phenotypes shown by the small number of isolates of endophytes such as Ph. occulta, R funerea and C. abietina, as well as the diverse effects of redwood essential oil phenotypes on endophytic species, underlines the importance of experiments that consider intraspecific variabil ity in the fungi as well as the chemistry of the host.
Acknowledgements--We wish to thank Dr G. C. Carroll, Dr F. Cobb, Jr, J. Rollinger and R. Goralka for their comments on the manuscript. We also appreciate E. Garcia-Marquez's help during the set-up of the experi- ment. We are also grateful to G. Hall and S. Wood for the help with statistical analysis and the extraction process of some terpenoids, respectively. We thank Dr G. C. Carroll for the identification of Meria parkeri and to Dr A. C. McCain and J. Rollinger for the isolates of Botrytis cinerea and Seiridium juniper~ respectively. This work was partially funded by a U.C.S.C. Faculty Research grant to J.H.L. and graduate student grants to F.J.E. from the U.C.S.C. Biology Department. F.J.E. wishes to thank the Universidad Nacional Autbnoma de M~xico and Mexico's Consejo Nacional de Ciencia y Tecnologia for supporting him with a fellowship during the period of this study.
References Affeltranger, C. E. and Cordell, C. E. (1989) Eastern white pine foliage blight. In Forest Nursery Pests (Cordell, C.
E., Anderson, R. L., Hoffard, W. H., Landis, T. D., Smith, R. S., Jr, and Toko, H. V., technical coordinators), pp. 38-39. U.S. Department of Agriculture, Forest Service. Agriculture Handbook No. 680.
Arrhenius, S. P. and Langenheim, J. H. (1983) Inhibitory effects of Hyrnenaea and Copaifera leaf resins on the leaf fungus, Pestalotia subcuticularis. Biochem. Syst. Ecol. 11, 361-366.
Benjalili, B., Tantaoui-Elaraki, A., Ayadi, A. and Ihal, M. (1984) Methods to study antimicrobial effects of essential oils: application to the antifungal activity of six Moroccan essences. J. Food Prot. 47, 742-748.
Brasier, C. M. (1998) Rapid changes in genetic structure of epidemic populations of Ophiostoma ulmi. Nature 332, 538-541.
Bridges, J. R. (1987) Effects of terpenoid compounds on growth of symbiotic fungi associated with the southern pine beetle. Phytopathology 77, 83-85.
Buczacki, S. T. (1973) A microecological approach to larch canker biology. Trans. Br. Mycol. Soc. 61, 315-329. Carroll, F. E., Muller, E. and Sutton, B. C. (1977) Preliminary studies on the incidence of needle endophytes in
some European conifers. Sydowia 29, 87-103. Carroll, G. C. (1986) The biology of endophytism in plants with particular reference to woody plants. In

640 F. J. ESPINOSA-GARC[A AND J. H. LANGENHEIM
Microbiology of the Phyllosphere (Fokkema, N. and van den Heuvel, J,, eds), pp. 205-222. Cambridge University Press, London.
Carroll, G. C. (1988) Fungal endophytes in stems and leaves: from latent pathogen to mutualistic symbiont. Ecology 69, 2-9.
Carroll, G. C. (1991) Fungal associates of woody plants as insect-antagonists. In Mediation of Herbivore-Plant Interactions (Barbosa, P., Krischik, Vo A. and Jones, C. G., eds). John Wiley, New York (in press).
Chapela, I. H. and Boddy, L. (1988) The fate of early colonizers in beech branches decomposing on the forest floor. FEMS Microbial Ecology 53, 273-284.
Chararas, C., Revolon, C., Feinberg, M. and Ducauze, C. (1982) Preference of certain scolytidae for different conifers. A statistical approach. J. Chem. Ecol. 8, 1093-1109.
Clay, K. (1988) Clavicipitaceous fungal endophytes of grasses: coevolution and the change from parasitism to mutualism. In Coevolution of Fungi with Plants andAnimals (Pirozinski, K. A. and Hawksworth, D. L., eds), pp. 79-105. Academic Press, London.
Cobb, F. W., Jr, Krstic, M., Zavarin, E. and Barber, H. W. (1968) Inhibitory effects of volatile oleoresin components on Fomes annosus and four Ceratocystis species. Phytopathology 58, 1327-1335.
Davidson, J. G. N. (1971) Pathological Problems in Redwood Regeneration from Seed. Ph.D. Thesis, University of California, Berkeley. 288 pp.
De Groot, R. C. (1972) Growth of wood inhabiting fungi in saturated atmospheres of monoterpenoids. Mycologia 64, 863-870.
Ennos, R. A. and Swales, K. W. (1988) Genetic variation in tolerance to host monoterpenes in a population of the ascomycete canker pathogen Crumenulopsis sororia. Plant Path, 37, 407-416.
Espinosa-Garcia, F. J. (1991) Studies of the Relation of the Fungal Endophytic Community and EssenEal Oils in the Leaves of Coastal Redwood (Sequoia sempervirens). Ph.D. Dissertation, University of California, Santa Cruz, California.
Espinosa-Garcia, F. J. and Langenheim, J. H. (1990) The endophytic fungal community in leaves of a coastal redwood population. Diversity and spatial patterns. New Phytoh 116, 89-97.
Espinosa-Garcia, F. J. and Langenheim, J. H. (1991) Effect of some leaf essential oil phenotypes from coastal redwood on the growth of its predominant endophytic fungus. J. Chem. EcoL 17 (in press).
Farr, D. F., Bills, G. F., Chamuris, G. P. and Rossman, A. Y. (1989) Fungion Plants and Plant Products in the United States. APS Press, St. Paul, Minnesota.
Flodin, K. and Fries, N. (1978) Studies on volatile compounds of Pinus sylvestris and the effect on wood decomposing fungi. II. Effects of some volatile compounds on fungus growth. Eur. J. For Path. 8, 300-310.
Franich, R. A., Gaskin, R. E., Wells, L. G. and Zabkiewicz, J. A. (1982) Effect of Pinus radiata needle mono- terpenes on spore germination and mycelial growth of Dothiostroma piniin vitro in relation to mature tree resistance. PhysioL Plant Path. 21, 55-63.
Franich, R. A., Carson, M. J. and Carson, S. D. (1986) Synthesis and accumulation of benzoic acid in Pinus radiata needles in response to tissue injury by dothiostromin, and correlation with resistance of R radiata families to Dothiostroma pinL Physiol. Plant Path, 28, 267-286.
Funk, A. (1981) Parasitic Microfungiof Western Trees. Canadian Forestry Service, Pacific Forest Research Centre, Victoria, B.C.
Gambliel, H. A., Cates, R. G., Caffey-Moquin, M. K. and Paine, T. D. (1985) Variation in the chemistry of Ioblolly pine in relation to infection by the blue-stain fungus. In Integrated Pest Management Research Symposium: The Proceedings (Branham, S. J. and Thatcher, R. C., eds), pp. 177-185. U.S. Dep. Agric. For. Serv. South. For. Exp. Stn. Gen. Tech. Rep. SO-56.
Gibbs, J. N. (1968) Resin and resistance of conifers to Fomes annosus. Ann. BoL 32, 649-665. Gregonis, D. E., Portwood, R. D., Davidson, W. H., Durfee, D. A. and Levinson, A. S. (1968) Volatile oils from
foliage of coast redwood and big tree. Phytochemistry 7, 975-981. Hall, G. D. (1985) LeafMonoterpenes of Coast Redwood (Sequoia sempervirens). Ph.D. Dissertation. University
of California, Santa Cruz, California. Hall, G. D. and Langenheim, J. H. (1986a) Within-tree spatial variation in the leaf monoterpenes of Sequoia
sempervirens. Biochem. Syst. Ecol. 14, 625-632. Hall, G. D. and Langenheim, J. H. (1986b) Temporal changes in the leaf monoterpenes of Sequoia
sempervirens. Biochem. Syst. Ecol. 14, 61-69. Hall, G. D. and Langenheim, J. H. (1987) Geographic variation in leaf monoterpenes of Sequoia sempervirens.
Biochem. Syst. Ecol. 15, 31-43. Hanover, J. W. (1966) Environmental variation in the monoterpenes of Pinus monticola Dougl. Phytochemistry
5, 713-717. Haug, I., Weber, G. and Oberwinkler, F. (1968) Intracellular infection by fungi in mycorrhizae of damaged
spruce trees. Eur. J. For. Path, 18, 112-120. Hepting, G. H. (1971) Diseases of Forest and Shade Trees of the United States. U.S. Dept. of Agriculture. Forest
Service. Agriculture Handbook No. 386. Hilal, S. H., Zedan, H. H., Haggug, M. Y. and Soliman, F. M. (1977) Volatile oil of Sequoia sempervirans Endl.
Egypt J. Pharmac. ScL 18, 85-95. Hintikka, V. (1970) Selective effects of terpenes on wood decomposing Hymenomycetes. Karstenia 11, 28-32. Kliejunas, J. (1988) Fungicide trials for control of Phomopsis canker of Douglas fir at a Northern California

ESSENTIAL OILS IN COASTAL REDWOOD 641
nursery. Tree Planters'Notes 39, 26--29. Kurita, N. and Koike, S. (1982) Synergistic antimicrobial effect of sodium chloride and essential oil
components. Agric. Biol. Chem. 46, 159-165. Langenheim, J. H. (1984) Role of plant secondary compounds in wet tropical ecosystems. In Physiological
Ecology of Plants in the Wet Tropics (Medina, E., Mooney, H, and Vasquez-Yanes, C., eds), pp. 189-208. W. Junk, The Hague Publ.
Langenheim, J. H. and Stubblebine, W. H. (1983) Variation in leaf resin composition between parent tree and progeny in Hymenaea: implications for herbivory in the humid tropics. Biochem. SysL Ecol. 11, 97-106.
Leather, S. R., Watt, A. D. and Forrest, G. I. (1987) Insect induced chemical changes in young Iodgepole pine (Pinus contorta): the effect of previous defoliation on oviposition, growth and survival of the pine beauty moth, Panolis flammea. Ecol. Entomol. 12, 275-281.
Luisi, N. and Triggiani, O. (1977) Sui recenti casi di sccumi nei cipressi. Informatore fitopatol. 10, 13-16. McCain, A. H. and Smith, P. C. (1978) Evaluation of fungicides for control of Botrytis blight of container-grown
redwood seedlings. Tree Planters'Notes 32, 12-13. Michelozzi, M., Squillance, A. E. and White, T. L. (1990) Monoterpene composition and fusiform rust resistance
in slash pine. For. Sc~ 36, 470-475. Miller, R. H., Berryman, A. A. and Ryan, C. A. (1986) Biotic elicitors of defense reactions in Iodgepole pine.
Phytochemism/ 25, 611-612. Mittal, R. K., Singh, P. and Wang, B. S. P. (1987) Botrytis: a hazard to reforestation. A literature review. Eur. J.
For. Path, 17, 369-384. Miura, H. and Kawano, N. (1968) Sequoiaflavone in the leaves of Sequoia sempervirens and Cunninghamia
lanceolata var. konishii and its formation by partial demethylation. Yakugaku Zasshi 88, 1489-1491. Mordue, J. M. E. (1976) Pestalotiopsis funerea. CMI Descriptions of Pathogenic Fungi and Bacteria. No. 514. Okamoto, R. A., Ellison, B. O. and Kepner, R. E. (1981) Volatile terpenes in Sequoia sempervirens foliage.
Changes in composition during maturation. J. Agric. Food Chem. 29, 324-326. Ohta, K. and Nawamiki, T. (1978) (+)-Polyalthic acid, a repellent against a sea snail Monodonta neritoides. Agric.
Biol, Chem. 42, 1957-1958. Pauli, A. and Knobloch, K. (1987) Inhibitory effects of essential oil components on growth of food con-
taminating fungi. Z. Lebensm Unters Forsch. 185, 10-13. Petrini, O. (1986) Taxonomy of endophytic fungi of aerial plant tissues. In Microbiology of the Phyllosphere
(Fokkema, N. and van den Heuvel, J., eds), pp. 175-187. Cambridge University Press, London. Pratt, J. E. (1982) Fomes annosus butt-rot of Sitka spruce IV. Observations on the distribution of Cryptosporiosis
abietina (Petrak) in the stems of rotted trees. Forestry 55, 183-187. Raffa, K. F. and Berryman, A. A. (1982) Accumulation of monoterpenes and associated volatiles following
inoculation of grand fir with a fungus transmitted by the fir engraver, Scolytus ventralis (Coleoptera: Scolytidae). Can. Ent. 114, 797-810.
Raffa, K. F., Berryman, A. A., Simaski, J., Teal, W. and Wong, B. L. (1985) Effects of grand fir monoterpenes on the fir engraver, Scolytus ventrelis (Coleoptera: Scolytidae), and its symbiotic fungus. Environ. Entomol. 14, 552-556.
Raddi, P, and Paconesi, A. (1981) Cypress canker disease in Italy: biology, control possibilities and genetic improvement for resistance. Eur. J. For. Path. 11, 340-347.
Rhoades, D. G., Lincoln, D. E. and Langenheim, J. H. (1976) Preliminary studies of monoterpenoid variability in Satureja douglasii Biochem. Syst. Ecol. 4, 5-12.
Rockwood, D. L. (1973) Monoterpene-fusiform rust relationships in Ioblolly pine. Phytopathology63, 551-553. Rockwood, D. L. (1974) Cortical monoterpene and fusiform rust resistance relationships in slash pine.
Phytopathology 64, 976-979. Rollinger, J. and Langenheim, J. H. (1989) Geographic variation in fungal endophyte community cmposition in
leaves of Sequoia sempervirens (D. Don ex Lamb) Endl. (Abstract). Proceedings of the Pacific Division, American Association for the Advancement of Science 8, Part 1, 33.
Roy, D. F. (1966) Sylvical Characteristics of Redwood [Sequoia sempervirens (D. Don) End~.]. U.S.D.A. Forest Service. Research Paper PSW-28, 20 pp.
Sanderson, P, G. and Worf, G. L. (1986) Phomopsis shoot blight of colorado blue spruce. J. Environ. Hort. 4, 134-138.
Scharpf, R. (1978) Stem Cankers. In Diseases of Pacific Coast Conifers (Bega, R. V., ed.), pp. 69-90, U.S. Dept of Agriculture, Forest Service, Agricultural Handbook No. 521.
Sherwood-Pike, M., Stone, J. K. and Carroll, G. C. (1986) Rhabdocline parker~ a ubiquitous foliar endophyte of Douglas fir, Can. J. BoL 64, 1849-1855.
Shrimpton, D. M. and Whitney, H. S. (1968) Inhibition of growth of blue stain fungi by wood extractives. Can. J. Bot. 46, 757-761.
Sieber, T. N. (1989) Endophytic fungi in twigs of healthy and diseased Norway spruce and white fir. Mycol. Res. 92, 322-326.
Sinclair, W. A., Lyon, H. H. and Johnson, W. T. (1987) Diseases of Trees and Shrubs, Comstock Publishing, Cornell University Press, Ithaca.
Squillance, A. H., Powers, H. R., Jr and Kossuth, S. V. (1985) Monoterpene phenotypes in Ioblolly pine popu- lations: natural selection trends and implications. Proc. 18th South. For. Tree Improve. Conf 299-308.

642 F. J. ESPINOSA-GARdA AND J. H. LANGENHEIM
Stafford, H. A. and Lester, H. H. (1986) Proanthocyanidins in needles from six genera of the Taxodiaceae. Am. J. Bo~ 73, 1555-1562.
Sutton, B. C. (1980) The Coelomycetes. Commonwealth Mycological Institute, Kew, Surrey, England. 696 pp. Thibault-Balesdent, M. and Delatour, C. (1985) Variabilite du comportament de Heterobasidion annosum (Fr.)
Bref. a trois monoterpenes. Eur. J. For. Path. 15, 301-307. Tripathi, R. D., Banerji, R., Sharma, M. L., Balasubrahmanyam, V. R. and Nigam, S. K. (1985) Toxicity of
essential oil from a new strain of Ocimum gratissimum (Clocimum) against betelvine pathogenic fungi. Agric. Biol. Chem. 49, 2277-2282.
Triplet-t, M. A. (1990) Potential Pathogenic Effects of Some Redwood Endophytes on the Growth and Survival of Redwood Seedlings in vitro. B.S. Thesis, University of California, Santa Cruz, California.
Tuset Barrachina, J. J. (1972) Un marchitamiento y secado de hojas y ramas j6venes de origen micol6gico, en el cipres. An. I.N.I.A. Set. ProL veg. 2, 11-25.
V~is~la, L. (1974) Effects of terpene compounds on the growth of wood-decomposing fungi. Ann. bo~ Fennici 11, 275-278.
Verhoeff, K. (1974) Latent infections by fungi. A. Rev. Phytopath 12, 99-110. Von Rudloff, E. (1975) Volatile leaf oil analysis in chemosystematic studies of North American conifers.
Biochem. Syst. Ecol. 2, 131-167. Wagner, M. R., Clancy, K. M. and Tinus, R. W. (1989) Maturational variation in needle essential oils from
Pseudotsuga menziesii, Abies concolor and Picea engelmanniL Phytochemistry 28, 765-770.





























