Embed Size (px)
Citation preview

Clin. exp. Immunol. (1990) 81, 200-206
Effect of phorbol myristate acetate on T cell colony formation,interleukin-2 (IL-2) receptor expression and IL-2 production by
cells from patients at all stages of HIV infection
M. ALLOUCHE*t, Y. LUNARDI-ISKANDAR*, C. VARELA-MILLOT*, M. ITZHAKIJ, P. CORNUET*,P. MEYERT, C. JASMIN* & V. GEORGOULIAS*§ *Unite' d'Oncoge'ne'se Appliquek, H6pital Paul Brousse, Villejuif,
tCentre de Recherche de Biochimie et de Gknktique Cellulaire du CNRS, Toulouse, l~nstitut de Canct~rologie etd'Immunoge'netique (ICIG), H6pital Paul Brousse, Villejuif, France, and §School of Medicine,
Division of Haematology-Oncology, University of Crete, Iraklion, Greece
(Acceptedfor publication 13 February 1990)
SUMMARY
We and others have shown that several T cell responses induced by the mitogen phytohaemagglutinin(PHA), including T cell colony formation, IL-2 receptor (IL-2R) expression, and IL-2 production areimpaired in patients with AIDS and lymphadenopathy syndrome (LAS). We investigated whetherphorbol myristate acetate (PMA) could act in synergy with PHA (as it does in healthy subjects) toenhance in vitro T cell responses of patients at all stages of infect-ion by HIV. In AIDS patients withopportunistic infections (AIDS/QI), PHA + IL-2 +PMA led to a total disappearance of T cellcolonies in 10/11 patients, among whom six already displayed very low numbers of colonies inducedby PHA + IL-2 (less than 50 colonies/5 x 104 cells). In contrast, T cell colony formation induced byPHA + IL-2 +PMA was maintained or increased, compared with that induced by PHA + IL-2, in fiveout of six AIDS patients with Kaposi's sarcoma (AIDS/KS), 10/14 LAS and six out of seven HIV-seropositive asymptomatic (HIVe/AS) homosexuals. In these three groups of patients, a lowpercentage of colony cells induced by PHA + IL-2 +PMA expressed CD3 and CD4 molecules, but50-89% of cells were IL-2R (Tac) positive, as in healthy controls. Studies on T cell activation andIL-2 production were performed on a selected group of 12 HIV-infected patients for whom sufficientnumbers of lymphocytes could be obtained. PMA induced CD4 down-modulation in controls and inHIV-infected patients. However, CD3 down-modulation and induction of the Tac chain of IL-2R byPMA were significantly impaired in patients, compared with controls, and these two parameters werecorrelated. Although PHA alone induced virtually normal levels ofTac antigen on patients' cells, Tacinduction by PHA +PMA was significantly decreased in patients versus controls. Cells from five outof 10 patients tested failed to produce detectable amounts of IL-2 after PHA stimulation, whereasIL-2 production increased significantly in all patients tested (n = 9) after PHA + PMA, with a level ofIL-2 activity significantly higher than in controls. No correlation was found in this group of patientsbetween the effects ofPMA +PHA on T cell colony formation, Tac expression, or IL-2 production,as compared with PHA alone. Taken together, our results indicate that in vitro T cell functionalstudies with PMA may be useful to evaluate better the defects of T cell activation in HIV-infectedpatients.
Keywords HIV phorbol myristate acetate interleukin-2 T cell activation T cell colonies
INTRODUCTION abnormalities of T lymphocytes, including a decrease of abso-lute numbers of CD4+ cells (Fauci et al., 1984), of T cell
Patients with AIDS, persistent lymphadenopathy syndrome proliferative responses to alloantigens and mitogens (Talal &(LAS), as well as HIV-positive, asymptomatic (HIVe/AS) Shearer, 1983), of IL-2 production (Murray et al., 1985) and IL-homosexual men have significant quantitative and qualitative 2 receptor (IL-2R) expression by mitogen-stimulated T lympho-
cytes (Gupta, 1986; Creemers, O'Shaughnessy & Boyko, 1987;Correspondence: Dr Mich~1e Allouche, Centre de Recherche de Prince & Czaplicki, 1988).
Biochimie et de G~n~tique Cellulaires du CNRS, 118 route de Assays for T cell colony growth in semi-solid mediaNarbonne, 31062 Toulouse Cedex, France. (Rozenszajn, Shohaw & Kalecheman, 1975; Georgoulias et al.,
200

PMA-induced T cell responses in HIV-infected patients
1985) are useful to study the proliferation and differentiation ofperipheral blood T cell colony-forming cells (T-CFC). NormalT cell colonies, generated in the presence of phytohaemaggluti-nin (PHA) and IL-2 derive from both mature (CD2+) andimmature (CD2-) T-CFC, and are composed of well-differen-tiated (CD3+CD4+ and CD3+CD8+) T cells (Georgoulias etal., 1985; Triebel et al., 1981). Recently, we and others havereported that T cell colony formation induced by PHA+ IL-2was significantly decreased in AIDS, LAS and, to a lesser extent,HIV+/AS homosexual men (Lunardi-Iskandar et al., 1985,1986; Winkelstein et al., 1985, 1988). However, it is yet unclearwhether this anomaly is due to a decrease of the pool of T cellprogenitors or to a deficient capacity of activation of Tlymphocytes.
Previous studies have shown that the tumour promoterphorbol myristate acetate (PMA) is a potent enhancing factor(together with PHA+ IL-2) for normal T cell colony growth(Klein et al., 1983; Kitamura et al., 1983; Foa et al., 1984;Winkelstein et al., 1986). PMA apparently acts in synergy withPHA, enhancing the expression of IL-2R on normal T lympho-cytes and the production of IL-2 by these cells (Depper et al.,1984; Sando et al., 1981). PMA seems to bypass the first steps ofT cell activation by mimicking a second messenger, diacylgly-cerol, and directly activating protein kinase C (PKC) (Nishi-zuka, 1984). In addition, PMA can down-modulate expressionof T cell surface antigens, such as CD3 (Cantrell, Davies &Crumpton, 1985; Matsuoka et al., 1986) and CD4 (Solbach,1982; Richie et al., 1988), which can play a role in cell activationand/or proliferative signal transduction. Therefore, the presentstudies were conducted to determine whether PMA could act insynergy with PHA to restore and/or enhance, as in normalsubjects, T cell colony formation, IL-2 production and IL-2Rexpression in patients at all stages of HIV infection.
PATIENTS AND METHODS
PatientsHeparinized peripheral blood was obtained from 17 AIDSpatients, presenting either opportunistic infections (01) (11patients), or Kaposi's sarcoma (KS) (six patients, none ofwhompresented 01 during the course of our study); 14 patients withLAS; eight HIV+/AS homosexual men; and 11 healthy hetero-sexual blood donors. All AIDS and LAS patients were adulthomosexual men who were studied at presentation, before anytreatment was applied. All patients, but not the healthy donors,had serum antibodies against HIV-1 proteins as detected byWestern blot. Informed consent for these studies was obtainedfrom all participants.
Cell separationPeripheral blood mononuclear cells (PBMC) were obtained byFicoll/Hypaque (Pharmacia Fine Chemicals, Uppsala, Sweden)density gradient centrifugation (p = 1-077 g/cm3). Interface cellswere washed twice with HBSS (Institut Pasteur Production,Marnes la Coquette, France) and resuspended in growthmedium (alpha-modified Eagle's medium (oe-MEM), fromGIBcO, Grand Island, NY).
T cell colony assayA standard method, described previously for optimal growth ofnormal T cell colonies (Georgoulias et al., 1985; Lunardi-Iskandar et al., 1985, 1986) was used. Briefly, 5 x 105 PBMC/mlwere seeded in methylcellulose (0-8% v/v) in a-MEM supple-mented with 15% (v/v) heat-inactivated fetal calf serum (FCS)(GIBCO), 2 mM L-glutamine, antibiotics, with PHA-P (0 1% v/v,Difco, Detroit, IL) and 0-5 U/ml recombinant IL-2 (rIL-2, a giftfrom Biogen, Switzerland; 1 U IL-2= 40 U BRMP, as defined inthe Biological Responses Modifiers Program of the NationalCancer Institute, Bethesda, MD). PMA (Sigma, St Louis, MO)was added (or not) at a concentration of 10 ng/ml. Cultures wereincubated for 5 to 7 days at 37°C in 5% CO2 in air, andaggregates containing more than 50 cells were counted ascolonies under an inverted microscope. T cell colony growthinferior or equal to 10 colonies/5 x 104 cells is considered as nonsignificant.
Liquid culturesLiquid cultures were only performed for patients whose periph-eral blood samples yielded enough viable mononucleated cells,thus limiting the study to four AIDS/KS, four LAS and fourHIV+/AS patients. PBMC (106 cells/ml) were cultured for 48 hin RPMI 1640 (GIBCO), 10% FCS, with PMA (0-100 ng/ml)and/or PHA (0- 1% v/v) at 37°C in 5% CO2. After extensivewashes, viable cells were counted and used for phenotyping.Cell-free supernatants were collected and kept at - 80°C.
Dosage ofIL-2 activity and of viral reverse transcriptase activitySupernatants from PHA- and/or PMA-stimulated cells weretested for the presence of IL-2 activity using the CTLL2 murineIL-2-dependent cell line proliferation assay (Gillis et al., 1978).The Mg++-dependent viral reverse transcriptase activity ofthese supernatants was also assayed, according to Barr&Sinoussi et al. (1983).
Phenotypic studiesThese were performed on fresh PBMC (day 0), and on cellsincubated with PHA and/or PMA in liquid or semi-solidcultures (on day 2 or 5, respectively). The following murinemonoclonal antibodies (MoAbs) were used: OKT3 (CD3,Ortho, Raritan, NJ); IOT4 (CD4), and IOT8 (CD8) fromImmunotech (Luminy, France); Ti 1 (CD2, Coultronics); andtwo MoAbs recognizing the p55 chain of the IL-2R: anti-Tac (agift from Dr T. Waldmann, NIH, Bethesda, MD) and IOT14(Immunotech). Cells were stained by indirect immunofluores-cence, using FITC-coupled goat anti-mouse immunoglobulin(Nordic, Tilburg, the Netherlands) as second reagent. Cellpreparations were examined using an epifluorescence micro-scope (Leitz).
Statistical analysisStudent's t-test was used to compare the mean of variousparameters between groups of patients; the correlation coeffi-cient and the Wilcoxon test to study the relationship betweentwo variables concerning the same patient; and the Mann-Whitney U-test was used to compare the values of oneparameter between patients (all HIV-infected patients availablewere pooled, regardless of the initial diagnosis) and controls.
201
M. Allouche et al.
RESULTS
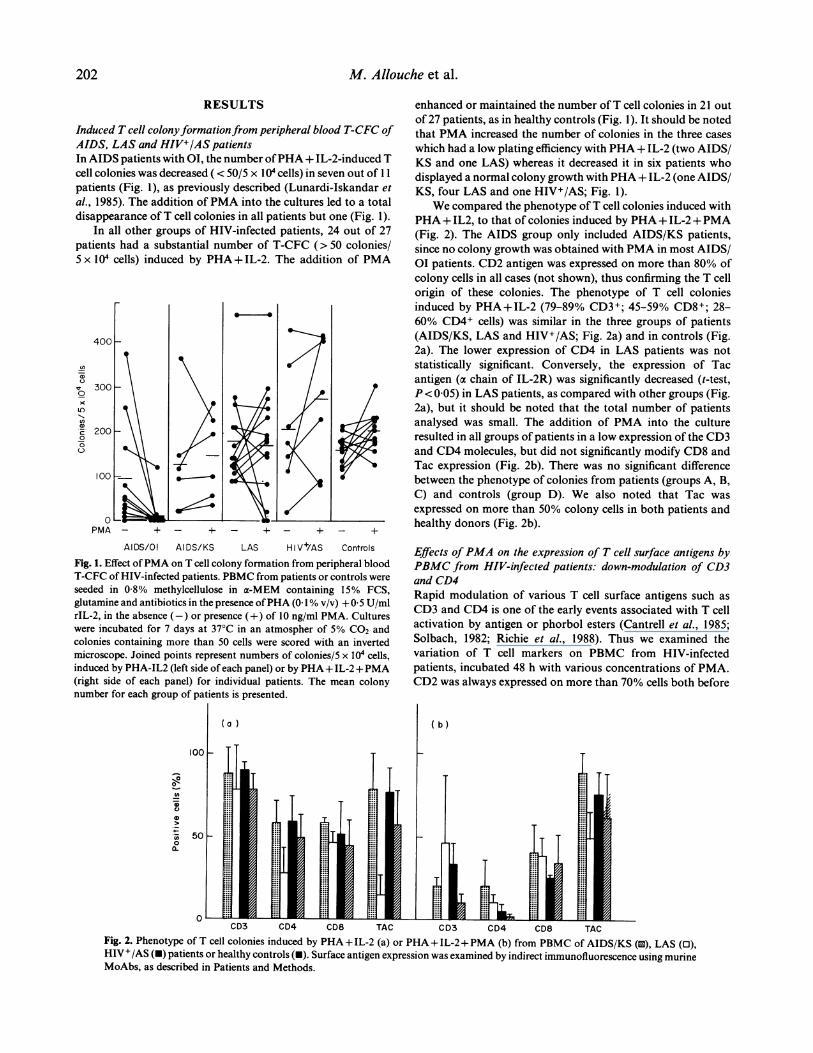
Induced T cell colonyformationfrom peripheral blood T-CFC ofAIDS, LAS and HIV+/AS patientsIn AIDS patients with OI, the number ofPHA+ IL-2-induced Tcell colonies was decreased ( < 50/5 x 104 cells) in seven out of 11patients (Fig. 1), as previously described (Lunardi-Iskandar etal., 1985). The addition ofPMA into the cultures led to a totaldisappearance ofT cell colonies in all patients but one (Fig. 1).
In all other groups of HIV-infected patients, 24 out of 27patients had a substantial number of T-CFC (> 50 colonies/5 x I04 cells) induced by PHA +IL-2. The addition of PMA
400 - + - + - + - + - +
-T ~ ~0-
0
20 200-0
00
100~ + - -
0-~~~~~~~
AIDS/O AIDS/KS LAS HIV+/AS Controls
Fig. 1. Effect ofPMA on T cell colony formation from peripheral bloodT-CFC of HIV-infected patients. PBMC from patients or controls wereseeded in 0 8% methylcellulose in a-MEM containing 15% FCS,glutamine and antibiotics in the presence ofPHA (0 1% v/v) +0 5 U/mlrIL-2, in the absence (-) or presence (+) of 10 ng/ml PMA. Cultureswere incubated for 7 days at 37 C in an atmospher of 5% CO2 andcolonies containing more than 50 cells were scored with an invertedmicroscope. Joined points represent numbers of colonies/5 x 104 cells,induced by PHA-IL2 (left side of each panel) or by PHA+ IL-2 +PMA(right side of each panel) for individual patients. The mean colonynumber for each group of patients is presented.
(a)
00 ibhM
enhanced or maintained the number ofT cell colonies in 21 outof 27 patients, as in healthy controls (Fig. 1). It should be notedthat PMA increased the number of colonies in the three caseswhich had a low plating efficiency with PHA+ IL-2 (two AIDS/KS and one LAS) whereas it decreased it in six patients whodisplayed a normal colony growth with PHA+ IL-2 (one AIDS/KS, four LAS and one HIV+/AS; Fig. 1).
We compared the phenotype ofT cell colonies induced withPHA+ IL2, to that of colonies induced by PHA+ IL-2 +PMA(Fig. 2). The AIDS group only included AIDS/KS patients,since no colony growth was obtained with PMA in most AIDS/01 patients. CD2 antigen was expressed on more than 80% ofcolony cells in all cases (not shown), thus confirming the T cellorigin of these colonies. The phenotype of T cell coloniesinduced by PHA+IL-2 (79-89% CD3+; 45-59% CD8+; 28-60% CD4+ cells) was similar in the three groups of patients(AIDS/KS, LAS and HIV+/AS; Fig. 2a) and in controls (Fig.2a). The lower expression of CD4 in LAS patients was notstatistically significant. Conversely, the expression of Tacantigen (a chain of IL-2R) was significantly decreased (t-test,P < 0 05) in LAS patients, as compared with other groups (Fig.2a), but it should be noted that the total number of patientsanalysed was small. The addition of PMA into the cultureresulted in all groups of patients in a low expression of the CD3and CD4 molecules, but did not significantly modify CD8 andTac expression (Fig. 2b). There was no significant differencebetween the phenotype of colonies from patients (groups A, B,C) and controls (group D). We also noted that Tac wasexpressed on more than 50% colony cells in both patients andhealthy donors (Fig. 2b).
Effects ofPMA on the expression of T cell surface antigens byPBMC from HIV-infected patients: down-modulation of CD3and CD4Rapid modulation of various T cell surface antigens such asCD3 and CD4 is one of the early events associated with T cellactivation by antigen or phorbol esters (Cantrell et al., 1985;Solbach, 1982; Richie et al., 1988). Thus we examined thevariation of T cell markers on PBMC from HIV-infectedpatients, incubated 48 h with various concentrations of PMA.CD2 was always expressed on more than 70% cells both before
° 1-:1. E-.VI 2 E--V/1-:.:IPA I. --I WA FIEU- I3AI F-24ETU2 oim VAdICD3 CD4 CD8 TAC CD3 CD4 CD8 TAC
Fig. 2. Phenotype of T cell colonies induced by PHA +IL-2 (a) or PHA + IL-2+PMA (b) from PBMC of AIDS/KS (u), LAS (0),HIV+/AS (U) patients or healthy controls (-). Surface antigen expression was examined by indirect immunofluorescence using murineMoAbs, as described in Patients and Methods.
202
PMA-induced T cell responses in HIV-infected patients
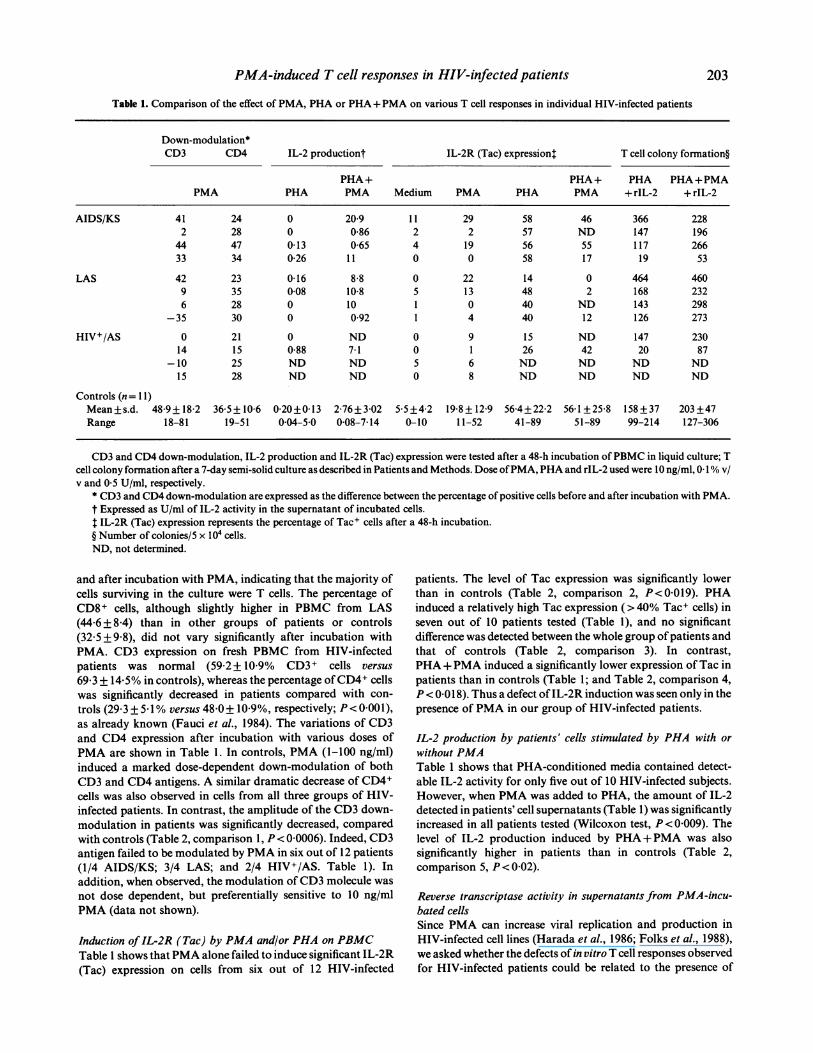
Table 1. Comparison of the effect of PMA, PHA or PHA+PMA on various T cell responses in individual HIV-infected patients
Down-modulation*CD3 CD4 IL-2 productiont IL-2R (Tac) expression: T cell colony formation§
PHA+ PHA+ PHA PHA+PMAPMA PHA PMA Medium PMA PHA PMA + rIL-2 + rIL-2
AIDS/KS 41 24 0 20-9 11 29 58 46 366 2282 28 0 0-86 2 2 57 ND 147 196
44 47 0-13 0 65 4 19 56 55 117 26633 34 0-26 11 0 0 58 17 19 53
LAS 42 23 0-16 8-8 0 22 14 0 464 4609 35 0-08 10-8 5 13 48 2 168 2326 28 0 10 1 0 40 ND 143 298
-35 30 0 0 92 1 4 40 12 126 273
HIV+/AS 0 21 0 ND 0 9 15 ND 147 23014 15 0-88 7-1 0 1 26 42 20 87
-10 25 ND ND 5 6 ND ND ND ND15 28 ND ND 0 8 ND ND ND ND
Controls (n = 11)Mean+s.d. 48-9+18-2 36 5+10-6 0-20+0-13 2-76+3-02 5 5+4 2 19-8+12-9 56 4+22 2 56-1+25 8 158+37 203+47Range 18-81 19-51 0-04-5 0 0-08-7-14 0-10 11-52 41-89 51-89 99-214 127-306
CD3 and CD4 down-modulation, IL-2 production and IL-2R (Tac) expression were tested after a 48-h incubation ofPBMC in liquid culture; Tcell colony formation after a 7-day semi-solid culture as described in Patients and Methods. Dose ofPMA, PHA and rIL-2 used were 10 ng/ml, 01% v/v and 0.5 U/ml, respectively.
* CD3 and CD4 down-modulation are expressed as the difference between the percentage of positive cells before and after incubation with PMA.t Expressed as U/ml of IL-2 activity in the supernatant of incubated cells.$ IL-2R (Tac) expression represents the percentage of Tac+ cells after a 48-h incubation.§ Number of colonies/5 x I10 cells.ND, not determined.
and after incubation with PMA, indicating that the majority ofcells surviving in the culture were T cells. The percentage ofCD8+ cells, although slightly higher in PBMC from LAS(44-6+ 8 4) than in other groups of patients or controls(32-5+9 8), did not vary significantly after incubation withPMA. CD3 expression on fresh PBMC from HIV-infectedpatients was normal (59-2 +10-9% CD3+ cells versus69 3 + 14-5% in controls), whereas the percentage ofCD4+ cellswas significantly decreased in patients compared with con-trols (293 + 51% versus 48-0 + 10-9%, respectively; P < 0001),as already known (Fauci et al., 1984). The variations of CD3and CD4 expression after incubation with various doses ofPMA are shown in Table 1. In controls, PMA (1-100 ng/ml)induced a marked dose-dependent down-modulation of bothCD3 and CD4 antigens. A similar dramatic decrease of CD4+cells was also observed in cells from all three groups of HIV-infected patients. In contrast, the amplitude of the CD3 down-modulation in patients was significantly decreased, comparedwith controls (Table 2, comparison 1, P < 0-0006). Indeed, CD3antigen failed to be modulated by PMA in six out of 12 patients(1/4 AIDS/KS; 3/4 LAS; and 2/4 HIV+/AS. Table 1). Inaddition, when observed, the modulation ofCD3 molecule wasnot dose dependent, but preferentially sensitive to 10 ng/mlPMA (data not shown).
Induction ofIL-2R (Tac) by PMA and/or PHA on PBMCTable I shows thatPMA alone failed to induce significant IL-2R(Tac) expression on cells from six out of 12 HIV-infected
patients. The level of Tac expression was significantly lowerthan in controls (Table 2, comparison 2, P<00019). PHAinduced a relatively high Tac expression (> 40% Tac+ cells) inseven out of 10 patients tested (Table 1), and no significantdifference was detected between the whole group of patients andthat of controls (Table 2, comparison 3). In contrast,PHA+PMA induced a significantly lower expression of Tac inpatients than in controls (Table 1; and Table 2, comparison 4,P< 0-018). Thus a defect ofIL-2R induction was seen only in thepresence ofPMA in our group of HIV-infected patients.
IL-2 production by patients' cells stimulated by PHA with orwithout PMATable I shows that PHA-conditioned media contained detect-able IL-2 activity for only five out of 10 HIV-infected subjects.However, when PMA was added to PHA, the amount of IL-2detected in patients' cell supernatants (Table 1) was significantlyincreased in all patients tested (Wilcoxon test, P < 0-009). Thelevel of IL-2 production induced by PHA+PMA was alsosignificantly higher in patients than in controls (Table 2,comparison 5, P < 0 02).
Reverse transcriptase activity in supernatants from PMA-incu-bated cellsSince PMA can increase viral replication and production inHIV-infected cell lines (Harada et al., 1986; Folks et al., 1988),we asked whether the defects of in vitro T cell responses observedfor HIV-infected patients could be related to the presence of
203

M. Allouche et al.
Table 2. Statistical comparisons between various parameters of T cellresponse ofPBMC from HIV-infected patients versus PBMC of healthy
heterosexual controls
Comparisonno. Variable pa Interpretation
I CD3 down-modulation < 0 0006 Patients < Controlswith PMA
2 Tac induction <0 019 Patients <Controlswith PMA
3 Tac induction NS Patients = Controlswith PHA
4 Tac induction <0 018 Patients < Controlswith PHA+PMA
5 IL-2 production < 0-02 Patients > Controlswith PHA+PMA
6 T cell colony formation NS Patients = Controlswith PHA + IL-2
7 T cell colony formation NS Patients = Controlswith PHA+PMA+ IL-2
* Mann-Whitney U-test.NS, Not significant.
virus in the culture. In eight AIDS (six KS and two 01), five LASand five HIV+/AS patients, the 48-h culture supernatants ofPMA- or PHA-stimulated PBMC did not display any reversetranscriptase activity: 3H-thymidine incorporation rangedbetween 150 and 560 ct/min with a negative control of 546 ct/min and a positive control of 26314 ct/min, respectively.
DISCUSSION
We have studied the effect of PMA and/or PHA on variousparameters of T cell activation, i.e. CD3 and CD4 down-modulation, IL-2R (Tac) induction, and IL-2 production bycells from 12 HIV-infected patients (four AIDS/KS, four LASand four HIV+/AS). We could not study T cell activation inAIDS/0I patients, because they all presented with a profoundlymphopenia. In addition, we observed a decreased cell viabilityof HIV-infected patients' PBMC from all diagnosis groups aftera 48-h culture with PMA and/or PHA as compared withcontrols' cells (P< 0-001, data not shown). However, this rapidcell death could not be attributed to an acute infection by HIVduring the culture, leading to virus release and cytotoxic for Tcells, since no reverse transcriptase activity was detected insupernatants from PHA- or PMA-incubated patients' cells.
We found defects, both ofCD3 down-modulation and of IL-2R (Tac) induction by PMA in patients' cells. Moreover, therewas a positive correlation between these two parameters of Tcell activation (r=0-63, P<0-028). Recently, defects of CD3down-modulation during T cell stimulation through the CD3/Tcell receptor complex, have been demonstrated in T cellsinfected by HIV both in vitro (Linette et al., 1988) and in vivo, i.e.in AIDS-related complex (ARC) patients' lymphocytes (Gurleyet al., 1989). Our results thus show that PMA stimulation mayreveal an early defect of signal transduction during HIVinfection.
In contrast, the down-modulation ofCD4 induced by PMAin HIV-infected patients was the same as in healthy subjects,
suggesting that CD3 and CD4 down-modulations are regulatedby different mechanisms. Hoxie et al. (1988) showed thatanother phorbol ester, phorbol dibutyrate, could still down-modulate CD4 in HIV-infected lymphoid cells. Since CD4 is areceptor for HIV (Klatzmann et al., 1984; Dalgleish et al., 1984),we could hypothesize that the modulation of CD4 may protectuninfected cells from viral infection. Indeed, Firestein et al.(1988) have shown that pre-treatment with PMA induced both adown-modulation of CD4 and a decrease of subsequent in vitroinfection by HIV in monocytoid cell lines.
We found that PHA induced IL-2R (Tac) at normal levels inmost patients, as also reported by others (Prince & Czaplicki,1988; Hofmann et al., 1989). However, we observed a selectivedefect of Tac induction by PMA (in the presence or absence ofPHA). This could be due at least partially to the secretion ofinhibitory factors by the patients' PBMC. Indeed, our group hasshown that peripheral blood adherent cells from HIV-infectedpatients spontaneously release a factor inhibiting Tac ex-pression on normal stimulated T cells, and that PMA enhancesthe production of this activity (Lunardi-Iskandar et al., 1987;A. Ammar et al., submitted for publication).
We observed a defect ofPHA-induced IL-2 production but asignificant increase of IL-2 activity after PHA +PMA stimula-tion of patients' PBMC, as also reported for PMA+ calciumionophore (Hofmann et al., 1989). This may be due to anenhanced IL-2 production, and/or to a lack ofabsorption of IL-2 by patients' T cells, related to the decreased Tac expression bythese cells. These results suggest that the stimulation of IL-2production, and the modulation of surface molecules such asCD3 or Tac by PMA, may be mediated through differentpathways of activation which may depend or not of PKCtranslocation (Kelleher, Pandol & Kagnoff, 1988).
We used the T cell colony assay as a sensitive measure ofimmunological function, allowing to study the proliferation anddifferentiation of T cell progenitors from peripheral blood.Because this assay requires relatively few cells, we includedAIDS/GI patients. The finding that PMA failed to enhance andeven suppressed T cell colony formation in these patientssuggests that they already have a decreased pool ofT-CFC, andthat no further T-CFC can be drawn to proliferate by PMA.Moreover, recent findings from our group suggest that T-CFCcould be destroyed by a mechanism of lytic infection by HIV- 1;indeed, in vitro infection of immature T cell precursors fromnormal human bone marrow leads to a decrease ofT cell colonyformation (Lunardi-Iskandar et al., 1989). Conversely, in theother groups of patients, T-CFC would be present in theperipheral blood, which could be recruited into cell cycle byPMA.
Analysing the data from Table 1, we could not correlate thevariation ofT cell colony formation to that of IL-2 production,or of IL-2R expression, respectively, in the presence ofPHA +PMA as compared with PHA alone. Thus, PMAtogether with PHA can only improve some T cell responses fromHIV-infected patients (such as IL-2 production and, sometimes,T cell colony formation). These results also suggest thatalthough IL-2R (Tac) induction is impaired, T-CFC, whichrepresent only a small subset ofT cells (about I in 500 cells) maybe normally activated by PMA.
In vitro T cell functional studies with PMA may be useful todelineate better the defects ofT cell activation involving the IL-2/IL-2R proliferation pathway and thus evaluate the immuno-
204

PMA-induced T cell responses in HIV-infected patients 205
logical status of HIV-infected patients. We already know that alow number of PHA-induced T cell colonies (<50/5 x 104PBMC) in LAS patients is ofpoor prognosis (Lunardi-Iskandaret al., 1988). Therefore these studies could potentially haveprognostic and therapeutic implications.
ACKNOWLEDGMENTS
This work was supported by grants from the Association de laRecherche contre le Cancer (ARC). We wish to thank Drs J. J. Fourni6and J. Breard for helpful suggestions, and Mrs N. Balliet and Mrs J.Maurel for expert secretarial assistance.
REFERENCES
BARRt-SINOUSSI, F., CHERMANN, J.C., REY, F., NUGEYRE, M.T., CHA-MARET, S., GRUEST, J., DAUGUET, C., AXLER-BLIN, C., VEZINET-BRUN,F., Rouzioux, C., ROZENBAUM, W. & MONTAGNIER, L. (1983)Isolation of a T lymphotropic retrovirus from a patient at risk foracquired immunodeficiency syndrome. Science, 220, 868.
CANTRELL, D.A., DAVIES, A.A. & CRUMPTON, M.J. (1985) Activators ofprotein kinase C down-regulate and phosphorylate the T3/T cellantigen receptor complex of human T lymphocytes. Proc. nail Acad.Sci. USA, 82, 8158.
CREEMERS, P.C., O'SHAUGHNESSY, M. & BOYKO, W.J. (1987) Increasedinterleukin-2 receptor expression after mitogen stimulation on CD4and CD8 positive lymphocytes and decreased interleukin-2 produc-tion in HTLV-III antibody-positive individuals. Immunology, 59,627.
DALGLEISH, A.G., BEVERLEY, P.C., CLAPHAM, P.R., CRAWFORD, D.H.,GREAVES, M.F. & WEISS, R.A. (1984) The CD4 (T4) antigen is an
essential component of the receptor for the AIDS retrovirus. Nature,312, 763.
DEPPER, J.M., LEONARD, W.J., KR6NKE, M., NOGUCHI, P.D., CUNN-INGHAM, R.E., WALDMANN, T.A. & GREENE, W.C. (1984) Regulationof interleukin 2 receptor expression: effects of phorbol diester,phospholipase C and reexposure to lectin or antigen. J. Immunol. 133,3054.
FAUCI, A.S., MACHER, A.M., LONGO, D.L., LANE, H.C., ROOK, A.H.,MASUR, H. & GELMANN, E.P. (1984) NIH Conference: Acquiredimmunodeficiency syndrome: epidemiologic, clinical, immunologicand therapeutic considerations. Ann. intern. Med. 100, 92.
FIRESTEIN, G.S., REIFLER, D., RICHMAN, D. & GRUBER, H.E. (1988)Rapid and reversible modulation ofT4 (CD4) on monocytoid cells byphorbol myristate acetate: effect on HIV susceptibility. Cell Immunol.113, 63.
FOA, R., Lusso, P., TERESA FIERRO, M., CRISTINA GIUBELLINO, M.,LUISA FERNANDO, M. & PEGORARO, L. (1984) Effects of 12-0tetradecanoyl-phorbol-13-acetate (TPA) on the colony growth ofhuman T lymphocytes. Clin. exp. Immunol. 56, 377.
FOLKS, M., JUSTEMENT, J., KINTER, A., SCHNITTMAN, S., ORENSTEIN, J.,POLI, G. & FAUCI, S. (1988) Characterization of a promonocyte clonechronically infected with HIV and inducible by 1 3-phorbol-12-myristate Acetate. J. Immunol. 140, 1117.
GEORGOULIAS, V., MARION, S., CONSOLINI, R. & JASMIN, C. (1985)Characterization of normal peripheral blood T- and B-cell colonycells: growth factor(s) and accessory cell requirements for their invitro proliferation. Cell Immunol. 90, 1.
GILLIS, S., FERM, M.M., Ou, W. & SMITH, K.A. (1978) T-cell growthfactor: parameters of production and a quantitative microassay foractivity. J. Immunol. 120, 2027.
GUPTA, S. (1986) Study of activated T cells in man. II. Interleukin-2receptor and transferrin receptor expression on T cells and produc-tion of interleukin-2 in patients with acquired immune deficiency[AIDS] and AIDS-related complex. Clin. Immunol. Immunopathol.38,93.
GURLEY, R.J., IKEUCHI, K., BYRN, R.A., ANDERSON, K. & GROOPMAN,
J.E. (1989) CD4+ lymphocyte function with early human immunode-ficiency virus infection. Proc. natl Acad. Sci. USA, 86, 1993.
HARADA, S., KOYANAGI, Y., NAKASHIMA, H., KOBAYASHI, N. &YAMAMOTO, N. (1986) Tumor promoter, TPA, enhances replicationof HTLV III/LAV. Virology, 154, 249.
HOFMANN, B., MOLLER, J., LANGHOFF, E., JAKOBSEN, K.D., ODUM, N.,DICKMEISS, E., RYDER, P., THASTRUP, O., SCHARFF, O., FODER, B.,PLATZ, P., PETERSEN, C., MATHIESEN, L., HARTVIG-JENSEN, T.,SKINHOJ, P. & SVEIJGAARD, A. (1989) Stimulation of AIDS lympho-cytes with calcium ionophore (A23187) and phorbol ester (PMA):studies of cytoplasmic free Ca, IL-2 receptor expression, IL-2production and proliferation. Cell Immunol. 119, 14.
HOXIE, A., RACKOWSKI, J.L., HAGGARTY, B.S. & GAULTON, G.N. (1988)T4 endocytosis and phosphorylation induced by phorbol esters butnot by mitogen or HIV infection. J. Immunol. 140, 786.
KELLEHER, D., PANDOL, S.J. & KAGNOFF, M.F. (1988) Phorbol myristateacetate induces IL-2 secretion by HUT 78 cells by a mechanismindependent of protein kinase C translocation. Immunology, 65, 351.
KITAMURA, K., URABE, A., OZAWA, K., KIMURA, Y. & TAKAKU, F.(1983) Enhancement of human T lymphocyte colony formation by12-0-tetradecanoylphorbol-13-acetate (TPA). Exp. Hematol. 11,1014.
KLATZMANN, D., CHAMPAGNE, E., CHAMARET, S., GRUEST, J., GUETARD,D., HERCEND, T., GLUCKMAN, J.C. & MONTAGNIER, L. (1984) Tlymphocyte T4 molecule behaves as the receptor for human retro-virus. Nature, 312, 767.
KLEIN, B., REY, A., JOURDAN, M., DONNADIEU, M.H. & SERROU, B.(1983) Induction ofhuman T colony formation by phorbol myristateacetate. Scand. J. Immunol. 17, 329.
LINETTE, P., HARTZMAN, J., LEDBETTER, A. & JUNE, H. (1988) HIV-1-infected T cells show a selective signaling defect after perturbation ofCD3/antigen receptor. Science, 241, 573.
LUNARDI-ISKANDAR, Y., GEORGOULIAS, V., ALLOUCHE, M., ROZEN-BAUM, W., KLATZMANN, D., CAVAILLE-COLL, M., MEYER, P., GLUCK-MAN, J.C., GENTILINI, M. & JASMIN, C. (1985) ABNORMAL in vitroproliferation and differentiation of T colony forming cells in AIDSpatients and clinically normal male homosexuals. Clin. exp. Immun.60, 285.
LUNARDI-ISKANDAR, Y., GEORGOULIAS, V., ROZENBAUM, W., KLATZ-MANN, D., CAVAILLE-COLL, M., MEYER, P., GENTILINI, M., GLUCK-MAN, J.C. & JASMIN, C. (1986) Abnormal in vitro proliferation anddifferentiation ofT colony forming cells in patients with lymphadeno-pathy syndrome. Blood, 67, 1063.
LUNARDI-ISKANDAR, Y., GEORGOULIAS, V., VITTECOQ, D., NUGEYRE,M.T., AMMAR, A., CLtMENCEAU, C., BARRt-SINOUSSI, F., CHERMANN,J.C., SCHWARTZENBERG, L. & JASMIN, C. (1987) Peripheral bloodadherent cells from AIDS patients inhibit normal T-colony growththrough decreased expression of interleukin 2-receptors and produc-tion of interleukin 2. Leukemia Res. 11, 753.
LUNARDI-ISKANDAR, Y., GEORGOULIAS, V., ViTTECOQ, D., AMMAR, A.,ROZENBAUM, W., MEYER, P. & JASMIN, C. (1988) Prognostic value ofblood and bone marrow T-colony forming cells (T-CFC) in patientswith lymphadenopathy syndrome (LAS). Biomed. Pharmacother. 42,21.
LUNARDI-ISKANDAR, Y., NUGEYRE, M.T., GEORGOULIAS, V., BARRE-SINOUSSI, F., JASMIN, C. & CHERMANN, J.C. (1989) Replication of thehuman immunodeficiency virus I and impaired differentiation of Tcells after in vitro infection of bone marrow immature T cells. J. clin.Invest. 83, 610.
MATSUOKA, M., HATTORI, T., CHOSA, T., TSUDA, H., KUWATA, S.,YOSHIDA, M., UCHIYAMA, T. & TAKATSUKI, K. (1986) T3 surfacemolecules on adult T cell leukemia cells are modulated in vivo. Blood,67, 1070.
MURRAY, H.W., WELTE, K., JACOBS, J.L., RUBIN, B.Y., MERTELSMANN,R. & ROBERTS, R.B. (1985) Production of and in vitro response tointerleukin-2 in the acquired immunodeficiency syndrome. J. clin.Invest. 76, 1959.

206 M. Allouche et al.
NIsHIZUKA, Y. (1984) The role of protein kinase C in cell surface signaltransduction and tumour promotion. Nature, 308, 693.
PRINCE, H.E. & CZAPLICKI, C.D. (1988) In vitro activation of Tlymphocytes from HIV-seropositive blood donors. II. Decreasedmitogen-induced expression of interleukin 2 receptor by both CD4and CD8 cell subsets. Clin. Immunol. Immunopathol. 48, 132.
RICHIE, E.R., McENTIRE, B., PHILLIPS, J. & ALLISON, J.P. (1988) Alteredexpression of lymphocyte differentiation antigens on phorbol ester-activated CD4+ 8 + T cells. J. Immunol. 140, 4115.
ROZENSZAJN, L.A., SHOHAW, D. & KALECHEMAN, I. (1975) Clonalproliferation of PHA-stimulated human lymphocytes in soft agarculture. Immunology, 29, 1041.
SANDO, J.J., HILFIKER, M.L., SALOMON, D.S. & FARRAR, J.J. (1981)Specific receptor for phorbol esters in lymphoid cell populations. Rolein enhanced production ofT cell growth factor. Proc. natl Acad. Sci.USA, 78, 1189.
SOLBACH, W. (1982) Tumor-promoting phorbol esters selectively abro-gate the expression of the T4 differentiation antigen expressed onnormal and malignant (Sezary) T helper lymphocytes. J. exp. Med.156, 1250.
TALAL, N. & SHEARER, G. (1983) A clinical and scientist look at acquiredimmunodeficiency syndrome (AIDS). Immunol. Today, 4, 180.
TRIEBEL, F., ROBINSON, W.A., HAYWARD, A.R. & GOUBE DE LAFOREST,P. (1981) Existence of a pool of lymphocyte colony forming cells (T-CFC) in human bone marrow and their place in the differentiation ofthe T lymphocyte lineage. Blood, 58, 91 1.
WINKELSTEIN, A., KLEIN, R.S., EVANS, T.L., DIXON, B.W., HOLDEN,W.L. & WEAVER, L. (1985) Defective in vitro T cell colony formationin the acquired immunodeficiency syndrome. J. Immunol. 134, 151.
WINKELSTEIN, A., SIMON, P.L., MYERS, P.A. & WEAVER, L.D. (1986)Comparisons of 12-tetradecanoylphorbol-13-acetate (TPA) andPHA as mitogens in the T-lymphocyte colony assay. Exp. Hematol.14, 1023.
WINKELSTEIN, A., KINGSLEY, L.A., KLEIN, R.S., LYTER, D.W., EVANS,T.L., RINALDO, C.R. JR, WEAVER, L.D., MACHEN, L.L. & SCHADLE,R.C. (1988) Defective T-cell colony formation and IL-2 receptorexpression at all stages ofHIV infection. Clin. exp. Immunol. 71, 417.





























