Embed Size (px)
Citation preview

Accepted Manuscript
Effect of Oxygen on Multidrug Resistance in Term Human Placenta
M. Javam, M.C. Audette, M. Iqbal, E. Bloise, W. Gibb, S.G. Matthews
PII: S0143-4004(14)00084-8
DOI: 10.1016/j.placenta.2014.02.010
Reference: YPLAC 2949
To appear in: Placenta
Received Date: 3 December 2013
Revised Date: 10 February 2014
Accepted Date: 23 February 2014
Please cite this article as: Javam M, Audette MC, Iqbal M, Bloise E, Gibb W, Matthews SG, Effectof Oxygen on Multidrug Resistance in Term Human Placenta, Placenta (2014), doi: 10.1016/j.placenta.2014.02.010.
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service toour customers we are providing this early version of the manuscript. The manuscript will undergocopyediting, typesetting, and review of the resulting proof before it is published in its final form. Pleasenote that during the production process errors may be discovered which could affect the content, and alllegal disclaimers that apply to the journal pertain.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPTEffect of Oxygen on Multidrug Resistance in Term Human Placenta M. Javama, M.C. Audettea,d, M. Iqbala, E. Bloisea, W. Gibbb, S.G. Matthewsa,c-e a Department of Physiology, University of Toronto, Toronto, Canada b Depts Ob-Gyn and Cellular & Molecular Medicine, University of Ottawa, Ottawa, Canada c Department of Ob-Gyn, University of Toronto, Toronto, Canada d Department of Medicine, University of Toronto, Toronto, Canada e Fraser Mustard Institute for Human Development, University of Toronto, Toronto, Canada
Short Tile: Oxygen tension and multidrug resistance in the term placenta
Keywords: P-glycoprotein, BCRP, third-trimester placenta, hypoxia, multidrug resistance
Grant Support: This study was funded by the Canadian Institutes for Health Research (grant: FRN-
57746; to S.G.M. and W.G.).
Disclosure Summary: Authors have nothing to declare
Correspondence and reprint requests:
Dr. Stephen G. Matthews, Department of Physiology, Faculty of Medicine, Medical Sciences
Building, Rm 3302, University of Toronto, 1 King's College Circle, Toronto, ON Canada, M5S 1A8.
Tel: (416) 978-2025 Fax:(416) 978-4940 e-mail: [email protected]

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPTAbstract 1
2
Introduction: The placenta contains efflux transporters, including P-glycoprotein (P-gp) and breast 3
cancer resistance protein (BCRP), that limit the passage of xenobiotics, certain hormones and 4
nutrients from the maternal to the fetal circulation. The expression of these transporters changes with 5
gestational age, yet the mechanisms involved remain unknown. However, the changes in P-gp and 6
BCRP transporter expression coincide with those of oxygen tension in the placenta, and oxygen 7
tension has been shown to modulate P-gp and BCRP expression in other tissues. The objective of this 8
study was to investigate the effects of oxygen tension on P-gp and BCRP expression in the term 9
human placenta. 10
Methods: Following equilibration in culture (96h), term placental explants (n=7) were cultured in 3% 11
or 20% oxygen for 24 and 48 h. Culture medium was collected every 24 h to measure lactate 12
dehydrogenase (LDH; explant viability) and human chorionic gonadotropin (hCG; 13
syncytiotrophoblast function). P-gp (encoded by ABCB1) and BCRP (encoded by ABCG2) protein and 14
mRNA, as well as VEGFA mRNA were measured using western blot and qRT-PCR. P-gp localization 15
was determined using immunofluorescence. 16
Results: Oxygen tension had a significant effect on P-gp expression, with ABCB1/P-gp mRNA and 17
protein levels increased in the hypoxic condition (3% O2) after 48 h (p<0.05). VEGFA mRNA was 18
elevated by hypoxia at both 24 and 48h (p<0.05). In contrast, placental ABCG2/BCRP mRNA and 19
protein expression were stable with changes in oxygen tension. We identified profound differences in 20
the glycosylation of P-gp between cultured and non-cultured placental tissue, with cultured explants 21
expressing deglycosylated P-gp. 22
Conclusions: These findings demonstrate that, at term, the expression of placental P-gp, is regulated 23
by oxygen tension. This suggests that changes in oxygenation of the placenta in the third trimester 24
may alter levels of placental P-gp, and in doing so alter fetal exposure to P-gp substrates, including 25
xenobiotics and certain hormones. 26

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT1. Introduction 1
One of the many roles of the placenta throughout gestation is to act as a protective barrier. In 2
addition to acting as a barrier that separates maternal blood from fetal blood, the placenta also 3
contains a variety of efflux transporters that are localized to the syncytiotrophoblast and fetal capillary 4
endothelium, which limit the passage of molecules from the maternal to the fetal circulation [1]. Many 5
of these efflux transporters belong to the ATP-binding cassette (ABC) family of transporters, which 6
decrease intracellular substrate concentrations through an active, energy-dependent mechanism [2]. 7
Two of these important ABC efflux transporters are P-glycoprotein (P-gp encoded by ABCB1) and 8
breast cancer resistance protein (BCRP encoded by ABCG2) [1]. Prior studies have shown these 9
transporters play a critical role in fetal protection throughout pregnancy [3-6]. For example, all the 10
offspring of mice deficient in placental P-gp showed the cleft palate phenotype when exposed to 11
Avermectin, a known substrate of P-gp, whereas the offspring from wild type animals were normal 12
[3]. In BCRP-deficient pregnant mice there was a five-fold increase in fetal concentrations of the 13
BCRP substrate, nitrofurantoin, compared with wild type animals [5]. 14
We have previously shown that the expression of these transporters in the human placenta 15
changes throughout pregnancy [7, 8]. ABCB1 mRNA and P-gp protein expression decrease with 16
advancing gestation [7, 8]. This decrease in P-gp levels has a potential to leave the fetus susceptible to 17
teratogens that may be present in the maternal circulation. The mechanism behind this decrease in P-18
gp expression remains unknown. However, the decrease in P-gp expression coincides with changes in 19
oxygen tension in the placenta. In the first trimester, the fetus is highly sensitive to molecular oxygen 20
and a low oxygen environment helps protect against oxidative stress [9]. At this early stage of 21
pregnancy, the partial pressure of oxygen is about 20 mmHg (~3% O2) due to the occlusion of the 22
terminal portion of maternal spiral arteries by the invading extravillous trophoblast (EVT) [9]. In the 23
early second trimester, the de-plugging of the spiral arteries allows maternal blood to enter the 24
intervillous space, causing an increase in oxygen tension to about 60 mmHg (~8% O2) [9]. However, 25
improper or inadequate conversion of spiral arteries in the first trimester can lead to chronic placental 26
ischemia and hypoxia later in gestation, which can cause deleterious outcomes such as intrauterine 27

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
2
growth restriction (IUGR), preeclampsia, unexplained miscarriage, and pre-term labor [10-13]. 28
Furthermore, these hypoxic placentas may have altered transporter expression patterns compared to 29
placentas from normal pregnancies [14]. 30
Studies in other cell types, especially cancer cells, have provided valuable insight into the 31
relationship between oxygen tension and transporter expression. Both P-gp and BCRP are highly 32
expressed in a number of cancer cells and expression has been shown to increase under conditions of 33
low oxygen tension [15-18]. This increase in expression is mediated through the binding of hypoxia 34
inducible factor (HIF-1α) to the HIF responsive element (HRE) in the ABCB1 and ABCG2 promoters 35
[15, 18]. We have recently shown that oxygen tension affects P-gp and BCRP transporter expression 36
in first trimester human placental explants [19]. An increase in oxygen tension resulted in an increase 37
in ABCG2 expression with no change in ABCB1 mRNA expression. At the protein level, hypoxia 38
induced P-gp and BCRP expression in proliferating cytotrophoblasts [19]. 39
Term placenta is subjected to higher oxygen tension than first trimester placenta and it is not 40
known whether oxygen tension regulates P-gp and BCRP expression at term. Moreover, 41
uteroplacental hypoxia, as a result of occlusion or inadequate trophoblast invasion of maternal spiral 42
arteries, can lead to chronic placental ischemia and hypoxia later in gestation [20]. Altered P-gp and 43
BCRP expression in response to hypoxia would impact fetal exposure to xenobiotics and hormones 44
present in the maternal circulation. The purpose of the present study was to examine the effect of 45
oxygen on the expression of these transporters in term placenta using an in vitro model of intact 46
placental explants. We hypothesized that a decrease in oxygen tension would result in an increase in 47
P-gp and BCRP levels. 48
49
2. Methods 50
2.1 Placental Collection and Tissue Culture 51
Term C-section placentas (n=7) were collected within 30 minutes (min) of delivery from the Research 52
Centre for Women’s and Infants’ Health (RCWIH) BioBank program at Mount Sinai Hospital. 53
Written informed consent was obtained from all subjects and ethical approval was obtained from the 54

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
3
Mount Sinai Hospital and the University of Toronto Research Ethics Board. Placental tissue biopsies 55
(1 cm3) were further dissected in the laboratory into approximately 8 mm3 fragments. Villous explants 56
were then cultured in individual Costar Netwells (Corning, New York) in CMRL-1066 media 57
(Invitrogen, Carlsbad, California, USA), supplemented with NaHCO3 (2.2 mg/ml; Sigma-Aldrich, St. 58
Louis, MO, USA), antibiotics-antimycotics (Invitrogen), insulin (1 mg/ml; Sigma), retinol acetate (1 59
mg/ml; Sigma), L-glutamine (100 mg/ml; Sigma) and fetal bovine serum (5%; Wisent Inc., St. Bruno, 60
Quebec, Canada) as previously described [21]. We and others [21, 22] have shown that in placental 61
explant cultures, the syncytiotrophoblast sheds within the first 2 days of culture and regenerates by 62
days 3-4, using both measures of syncytiotrophoblast function and histological analysis of 63
synctiotrophoblast integrity. Therefore, placental explants were cultured for 4 days (20% O2, 5% CO2, 64
37 °C) as previously described [21, 22], which allowed syncytiotrophoblast shedding and regeneration 65
to take place. On day 4, oxygen tension was changed to 3% (5% CO2, 37 °C) or maintained at 20% 66
for a further 24 and 48h. Culture media was changed every 24h during the culture period and care was 67
taken throughout the culture period for explants to remain out of the incubators for the same length of 68
time. Lactate dehydrogenase (LDH) and human chorionic gonadotropin (hCG) was measured in the 69
media to assess explant viability and syncytiotrophoblast function, respectively [21, 22]. 70
71
2.2 Lactate Dehydrogenase (LDH) and Human Chorionic Gonadotropin (hCG) 72
LDH and hCG were measured as described previously [21]. Briefly, LDH was quantified using the 73
Cytotoxicity Detection kit (Roche Applied Science, Indianapolis, USA) according to the 74
manufacturer’s instructions [19, 21]. A standard curve for the LDH assay was generated using L-lactic 75
dehydrogenase from rabbit muscle (Sigma) [19, 21]. hCG was measured by ELISA (DRG 76
Diagnostics, Germany) according to the manufactures’ instructions [21]. 77
78
2.3 Quantitative real time PCR 79
Total RNA was extracted from the placental explants using TRIzol reagent (Invitrogen) as described 80
in the manufacturer’s protocol. Contaminating genomic DNA was removed from RNA samples using 81

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
4
a DNA-free kit (Ambion, Austin, Texas, USA). RNA purity (A260/A280 ratio of 1.8-2.0) and 82
concentration was assessed using spectrophotometric analysis, and RNA integrity was verified by 83
agarose gel electrophoresis. RNA was reverse-transcribed to cDNA using High Capacity cDNA 84
Reverse Transcription kit (Applied Biosystems, Carlsbad, California, USA) according to the 85
manufacturer’s instructions and stored (-80°C). 86
Levels of ABCB1, ABCG2, and VEGFA mRNA were measured by real time RT-PCR using 87
SsoFast EvaGreen Supermix (Bio-Rad Laboratories, Hercules, CA, USA) and the CFX 96 Real-Time 88
system C1000 Thermal Cycler (Bio-Rad). Relative gene expression was measured using [∆∆c(t)] 89
method and normalized to the geometric mean of three housekeeping genes: glyceraldehyde 3-90
phosphate dehydrogenase (GAPDH), TATA-binding protein (TBP), and zeta polypeptide (YWHAZ) 91
[19]. Expression of these genes was stable during the experiments (data not shown). Data was 92
analyzed using CFX Manager Software (Bio-Rad). Primer sequences for the genes analyzed are listed 93
in Table 1. 94
95
2.4 Western Blotting 96
Western blotting was performed as described previously [23]. Briefly, placental explants were 97
homogenized in RIPA lysis buffer (Cell Signaling Technology, Inc., Danvers, MA), centrifuged 98
(10000g, 10min, 4°C), and the supernatant collected. Protein (40µg) separation using SDS-PAGE was 99
performed using polyacrylamide gels (7%) and transferred to nitrocellulose membranes using iBlot 100
transfer apparatus (Invitrogen). Membranes were blocked in BSA (5% w/v of tris buffered saline 101
[TBS] with Tween 20: TBS-T, 1h, 25°C). Membranes were then incubated with primary antibody 102
overnight (4°C). The primary antibodies used were: mouse anti-P-gp (1:5000; clone 5A12.2; EMD 103
Millipore Corporation, Billerica, MA, USA), mouse anti-BCRP (1:500; clone BXP-21; EMD 104
Millipore Corporation), and rabbit anti-actin (1:10000; Sigma). Membranes were washed and 105
incubated with horseradish peroxidase conjugated secondary antibody (1:15000; PerkinElmer, 106
Waltham, MA) against the corresponding primary antibody, followed by incubation (1min) in 107
enhanced chemiluminescence substrate (Invitrogen) and exposed to chemiluminescence film (General 108

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
5
Electric Healthcare, Baie d’Urfe, Quebec, Canada). The relative optical density (ROD) of the bands 109
was measured using computerized image analysis and was standardized against the β-actin signal 110
(MCID Core 7.0, Imaging Research Inc, Interfocus Imaging Ltd, Cambridge, England). 111
112
2.5 Immunofluorescence 113
Placental explants were fixed in neutral buffered formalin (10%; Sigma), washed with PBS and 114
cryosectioned (10µm). Sections were mounted onto microscope slides and stored (-80°C). Prior to 115
immunofluorescence, slides were brought to room temperature and washed with PBS. Antigen 116
retrieval was performed using boiling sodium citrate (10mM, 10min). Sections were cooled on ice 117
(10min) and washed with PBS before blocking in normal goat serum (5%, 1h; Invitrogen). Slides 118
were incubated overnight with mouse anti-P-gp primary antibody (1:50, D-11, Santa Cruz 119
Biotechnology, Santa Cruz, CA, USA) [19], then washed and incubated (1h, 25°C) with Alexa Fluor 120
488-conjugated goat secondary antibody (1:500; Invitrogen). Negative controls were stained by 121
substitution of primary antibody with non-immune mouse IgG (Dako, Burlington, ON, Canada). After 122
final washes, slides were mounted and cover-slipped using mounting media with 4′, 6-diamidino-2-123
phenylindole (Vector Laboratories, Burlingame, CA). Imaging was performed using Zeiss spinning 124
disk confocal microscope (Zeiss Observer.Z1). 125
126
2.6 Deglycosylation 127
P-gp deglycosylation was carried out using Peptide-N-Glycosidase F (PNGase F; Sigma) according to 128
the manufacturer’s instructions. Briefly, denatured and non-denatured protein (30ug) from fresh 129
placental tissue was incubated with PNGase F (4h, 37°C). Both denaturing and non-denaturing 130
conditions were used because some proteins require denaturation prior to PNGase F digestion. 131
Laemmli sample buffer (Bio-Rad) was added, and samples boiled (5min) to inactivate the PNGase F. 132
Western-blot analysis was performed as described above. 133
134
2.7 Statistical Analysis 135

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
6
Data are expressed as mean ± SEM and all statistical analyses were performed using Prism (GraphPad 136
Software, Inc., San Diego, CA). hCG and LDH concentrations were log transformed and analyzed 137
using one-way repeated measures ANOVA with Newman-Keuls post-hoc test. Differences in mRNA 138
and protein expression over time of culture and oxygen tension were assessed by two-way ANOVA 139
with Bonferonni post-hoc test. Protein and mRNA expression were also analyzed using one-way 140
ANOVA, followed by Dunnett post-test for comparisons against the control group. Statistical 141
significance was set at p<0.05. 142
143
3. Results 144
3.1 Tissue Viability and Syncytiotrophoblast Function 145
LDH release from placental explants across the 6-day culture period was measured to assess tissue 146
viability [21, 22]. LDH levels in the media decreased gradually, reaching a stable phase on day 4 (Fig. 147
1A). In order to monitor syncytial shedding and regeneration, hCG secretion was measured, a marker 148
of syncytiotrophoblast endocrine function [21, 22]. Between day 1 and day 2 there was a steep decline 149
in hCG release into the media (p<0.05) (Fig. 1B). From day 2 to day 3 there was a rapid increase in 150
hCG levels (p<0.05). The elevated hCG levels were sustained throughout the rest of the culture 151
period. This hCG pattern was indicative of syncytiotrophoblast shedding and regeneration [21, 22]. 152
Therefore, placental explants were cultured for 4 days in 20% oxygen, which allowed 153
syncytiotrophoblast shedding and regeneration to take place. The oxygen tension was then reduced to 154
3% or maintained at 20% for 24 and 48 h. Day 4 was considered the control group. 155
156
3.2 Effects of Oxygen Tension on ABCB1, ABCG2, and VEGFA mRNA Expression 157
ABCB1 and ABCG2 mRNA expression was measured in the control (day 4 of culture), 24h (day 5), 158
and 48h (day 6) samples from the same placentae using qRT-PCR. There was a significant effect of 159
oxygen tension on ABCB1 expression, assessed by two-way ANOVA (p<0.05) (Fig. 2A), with no 160
effects of time in culture or interaction of the two variables. There was a significant increase in 161
ABCB1 expression after 48 hours in 3% oxygen compared to the control group (p<0.05) (Fig. 2A). 162

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
7
Oxygen tension had no effect on ABCG2 expression, assessed by two-way ANOVA (Fig. 2B), with 163
no effects of time in culture or interaction of the two variables. VEGFA mRNA expression was 164
measured to assess activation of hypoxic pathways [19]. There was a significant increase in VEGFA 165
expression in the hypoxic condition (3% O2) (p<0.05) (Fig. 2C), with no effects of time in culture or 166
interaction of the two variables. VEGFA expression was also significantly increased at both 24h and 167
48h hypoxic conditions compared to the control group. 168
169
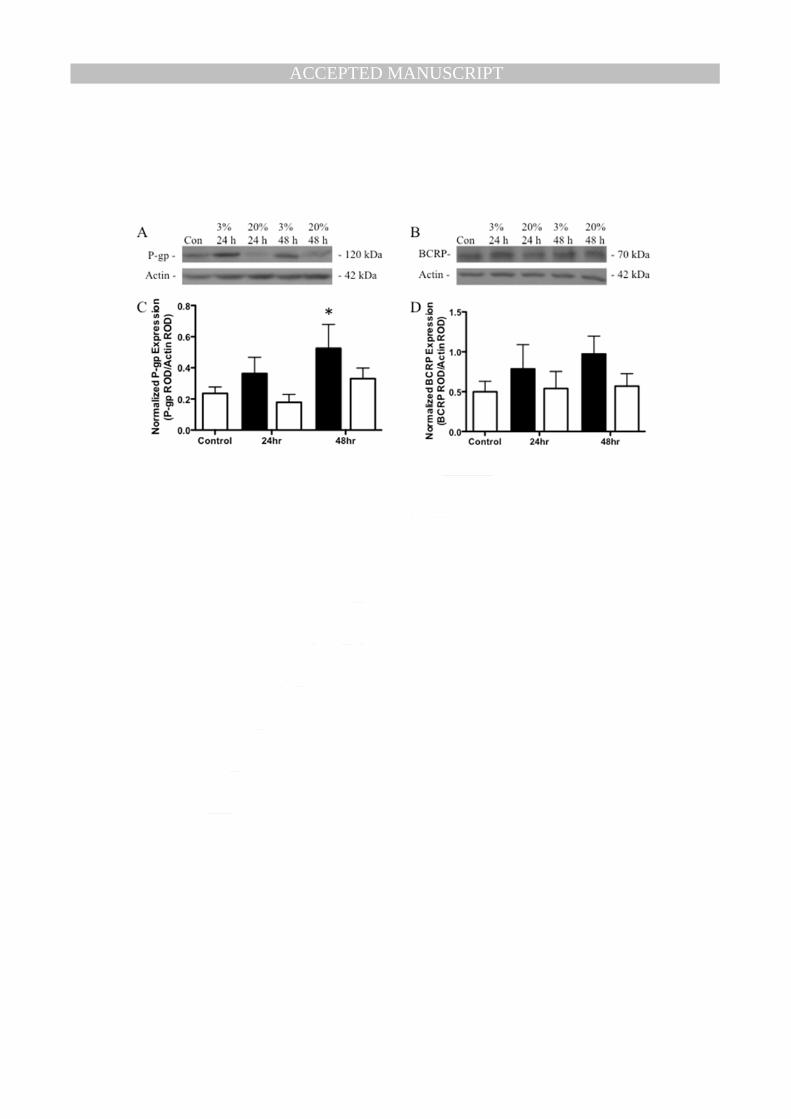
3.3 Effects of Oxygen on P-gp and BCRP Expression 170
There was no significant overall effect of oxygen tension on P-gp protein levels, assessed by two-way 171
ANOVA (Fig. 3C), with no interaction between the two variables. However, there was a significant 172
effect of time in culture on P-gp levels (p<0.05), such that there was an increase in P-gp levels with 173
increased time in culture. Further analysis using repeated measures one-way ANOVA revealed a 174
significant increase in P-gp levels after 48h of culture in 3% oxygen compared to control (p<0.05) 175
(Fig. 3C). Oxygen tension had no effect on BCRP protein levels, assessed by two-way ANOVA (Fig. 176
3D), with no effects of time in culture or interaction of the two variables. 177
178
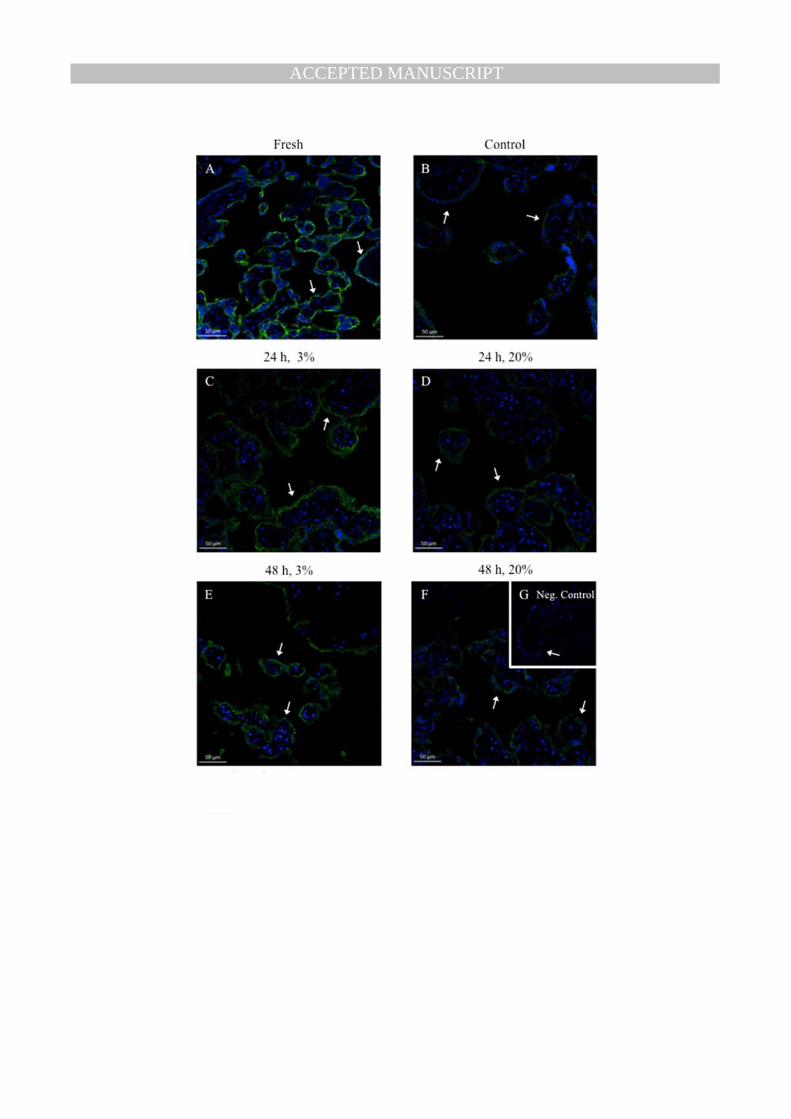
3.4 Effect of Tissue Culture on P-gp Glycosylation and Localization 179
Western blot analysis performed using fresh placental tissue, showed a predominant 150 kDa band for 180
P-gp (Fig. 4A). In contrast, P-gp in cultured placental tissue was approximately 120 kDa (Fig. 4A). To 181
assess whether the difference in the molecular weight of P-gp was due to deglycosylation, protein 182
from fresh placental tissue was treated with PNGase F (this cleaves the link between asparagine and 183
N-acetylglucosamines). PNGase F treatment resulted in a shift in the molecular weight of P-gp from 184
150kDa to 120kDa (Fig. 4B). Changes in oxygen tension had no effect on the glycosylation state of P-185
gp (data not shown). 186
To assess whether deglycosylation altered P-gp localization in the tissue, fresh and cultured explants 187
were analyzed using immunofluorescent staining. In both instances P-gp was localized to the 188

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
8
syncytiotrophoblast (Fig. 5) (n=4). Furthermore, decreasing oxygen tension did not alter P-gp 189
localization to the syncytiotrophoblast (Fig. 5). 190
191
4. Discussion 192
We have shown, for the first time, that changes in oxygen tension in the term human placenta 193
have significant but different effects on P-gp and BCRP expression. Decreased oxygen tension 194
resulted in an increase in ABCB1/P-gp mRNA and protein expression in term placental explants but 195
had no effect on ABCG2/BCRP mRNA or protein expression. We also report the novel finding that 196
while P-gp is glycosylated in fresh placental tissue, it becomes fully deglycosylated in cultured 197
placental explants. This de-glycosylation had no effect on the localization of P-gp to the 198
syncytiotrophoblast. This novel observation has important implications for our understanding of the 199
post-translational regulation of P-gp in the placenta and warrants further detailed investigation. 200
In the human placenta, ABCB1/ P-gp mRNA and protein expression decrease with advancing 201
gestation [7], which correlates closely with changes in oxygen tension throughout pregnancy. In the 202
present study, we found that 48hr exposure to low oxygen tension (3%) resulted in a significant 203
increase in ABCB1/ P-gp mRNA and protein levels, compared to the control group (20% O2). This 204
increase in ABCB1 and P-gp expression in response to hypoxia is consistent with studies performed 205
using other cell types [17, 18]. Furthermore, the responsiveness of P-gp expression to a decrease in 206
oxygen tension provides a possible explanation for the high P-gp levels in the low oxygen tension 207
environment of the first trimester placenta compared to term placenta. 208
Studies investigating the expression pattern of BCRP in the human placenta throughout 209
gestation are inconsistent. We have previously shown that ABCG2 mRNA expression does not change 210
throughout pregnancy, while BCRP protein levels increase towards term [8]. On the other hand, 211
Meyer zu Shwabedisson et al. demonstrated that both ABCG2 mRNA and BCRP protein expression 212
increase with advancing gestation [24] but Mathias et al. found that ABCG2 mRNA and BCRP 213
protein expression did not change with gestational age [25]. The reason for the discrepancy in BCRP 214
expression between studies may result from variability in sampling, as BCRP is present multiple 215

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
9
placental cell-types. In the present study oxygen tension had no effect on ABCG2 mRNA or BCRP 216
protein expression. This would be consistent with the stable ABCG2/BCRP expression pattern during 217
pregnancy, despite the increase in oxygen tension that occurs with advancing gestation. However, a 218
steady level of BCRP expression does not necessarily indicate no change in BCRP function, as no 219
study has measured BCRP activity throughout gestation. In fact, there are numerous studies 220
illustrating posttranslational modifications, such as glycosylation and phosphorylation, changing 221
BCRP function [26-28]. The same is true for P-gp [29-31], as Thews et al have shown a more than 222
doubling of P-gp activity in prostate carcinoma cells in acidic environments with no change in P-gp 223
expression [31]. Future studies are required to measure BCRP and P-gp activity in the placenta with 224
advancing gestation. 225
Compared to the third trimester, the placenta in the first trimester is exposed to lower oxygen 226
levels [12]. Early in the first trimester the extra villous trophoblast (EVT) invade and occlude the 227
terminal portion of spiral arteries, inhibiting maternal blood from entering the intervillous space and 228
resulting in a low oxygen environment (~3% O2) [10]. In late first trimester, the endovascular EVT 229
causes physiological changes in placental spiral arteries, forming funnel-shaped flaccid arterioles with 230
increased vascular compliance and circumference [10]. This leads to the flow of maternal blood into 231
the intervillous space and a dramatic increase in oxygen levels for the remainder of pregnancy (~8% 232
O2) [10]. However, improper or inadequate conversion of spiral arteries in the first trimester can lead 233
to chronic placental ischemia and hypoxia later in gestation [10-13]. Moreover, embryogenesis in the 234
first trimester already takes place under anaerobic conditions, and in the second and third trimester 235
oxygen becomes more important for normal fetal organogenesis and growth [32]. Protection from 236
hypoxia at this stage of development is established via up-regulation of various genes, including 237
superoxide dismutase, insulin-like growth factor (IGF)-2, and heat shock protein 70 [33-35]. Our data 238
indicate that ABCB1 may be one of the protective genes activated in response to hypoxia in the third 239
trimester placenta. In fact, P-gp has been suggested to play a critical role in cell protection by 240
extruding toxins and harmful products of oxidative stress [36]. Therefore, P-gp may also play a role in 241
protecting the fetus from toxic products of oxidative stress in third trimester hypoxic placentas. 242

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
10
We have recently shown that in the first trimester placenta, ABCG2 mRNA expression 243
increased significantly in hyperoxic (20%) conditions after 48hrs in culture with no change in ABCB1 244
mRNA expression [19]. At the protein level, hypoxia resulted in increased immunoreactive BCRP in 245
cytotrophoblasts and in the microvillous membrane of the syncytium, and increased P-gp 246
immunoreactivity in proliferating cytotrophoblasts. This contrasts with the results of the current study 247
and suggests that the placenta in the different stages of pregnancy responds quite differently to 248
changes in oxygen tension. This is understandable, as oxygen tension that is physiological in the first 249
trimester would be considered hypoxic at term. 250
Interestingly, we found that cultured placental tissue contained P-gp that was less glycosylated 251
compared to fresh placental tissue. Post-translational modifications, such as glycosylation, have been 252
shown to alter P-gp activity in various ways, depending on the cell line examined [29]. In general, 253
glycosylation can affect a transporter’s activity, folding, stability, subcellular trafficking, localization, 254
and turnover [37, 38]. P-gp is glycosylated on three recognition sites (Asn 91, 94, and 99) in the first 255
extracellular loop [37]. Deglycosylation of P-gp in fresh placenta decreased the molecular weight 256
from approximately 150 kDa to 120 kDa. This 30 kDa decrease is in agreement with previous studies 257
[39, 40]. Since the effect of deglycosylation on P-gp localization is cell type specific [30, 37, 40, 41], 258
we examined whether deglycosylation affects P-gp localization in term placental tissue. Using 259
immunofluorescent staining, the glycosylation state of P-gp was not found to affect P-gp localization 260
in the explants. Furthermore, oxygen tension had no effect on P-gp glycosylation or its localization to 261
the syncytiotrophoblast. Since the impact of glycosylation on P-gp activity is cell-specific [29, 30, 262
41], future studies are required to determine the effects of deglycosylation on P-gp activity in the 263
placenta. It is also important to highlight the potential limitation of the present study. For example, in 264
the immunofluorescent staining we were unable to definitively distinguish between the 265
syncytiotrophoblast and unfused cytotrophoblasts in the outer trophoblast layer. Also, it is possible 266
that molecular changes in syncytiotrophoblast function occur in pathways that indirectly modulate 267
multidrug resistance, following long term explant cultures. Future studies are required to determine 268
the precise molecular pathways involved in P-gp and BCRP regulation. 269

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
11
In conclusion, oxygen tension affects multidrug resistance in term human placenta. These 270
changes could alter fetal exposure to drugs, xenobiotic and endogenous substrates. Notably, improper 271
or inadequate conversion of spiral arteries in the first trimester can lead to chronic placental ischemia 272
and hypoxia later in gestation, which can cause deleterious outcomes such as intrauterine growth 273
restriction (IUGR) and preeclampsia. Glycosylation is an important regulatory mechanism of P-gp 274
function and expression, and should be considered so that we can better understand the various roles 275
of P-gp both in normal pregnancy progression and in disease states. Moreover, future studies should 276
focus on the expression profile and function of drug transporters in oxygen-related placental 277
pathologies since changes in the activity of these transporters could have a profound impact on fetal 278
drug exposure. 279
280
References 281
282
[1] Iqbal M, Audette MC, Petropoulos S, Gibb W and Matthews SG. Placental drug transporters 283
and their role in fetal protection. Placenta. 2012;33(3):137-42. 284
[2] Vahakangas K and Myllynen P. Drug transporters in the human blood-placental barrier. Br J 285
Pharmacol. 2009;158(3):665-78. 286
[3] Lankas GR, Wise LD, Cartwright ME, Pippert T and Umbenhauer DR. Placental P-glycoprotein 287
deficiency enhances susceptibility to chemically induced birth defects in mice. Reprod Toxicol. 288
1998;12(4):457-63. 289
[4] Smit JW, Huisman MT, van Tellingen O, Wiltshire HR and Schinkel AH. Absence or 290
pharmacological blocking of placental P-glycoprotein profoundly increases fetal drug exposure. 291
J Clin Invest. 1999;104(10):1441-7. 292
[5] Zhang Y, Wang H, Unadkat JD and Mao Q. Breast cancer resistance protein 1 limits fetal 293
distribution of nitrofurantoin in the pregnant mouse. Drug Metab Dispos. 2007;35(12):2154-8. 294
[6] Prouillac C and Lecoeur S. The role of the placenta in fetal exposure to xenobiotics: 295
importance of membrane transporters and human models for transfer studies. Drug Metab 296
Dispos. 2010;38(10):1623-35. 297
[7] Sun M, Kingdom J, Baczyk D, Lye SJ, Matthews SG and Gibb W. Expression of the multidrug 298
resistance P-glycoprotein, (ABCB1 glycoprotein) in the human placenta decreases with 299
advancing gestation. Placenta. 2006;27(6-7):602-9. 300
[8] Yeboah D, Sun M, Kingdom J, Baczyk D, Lye SJ, Matthews SG and Gibb W. Expression of breast 301
cancer resistance protein (BCRP/ABCG2) in human placenta throughout gestation and at term 302
before and after labor. Can J Physiol Pharmacol. 2006;84(12):1251-8. 303
[9] Schneider H. Oxygenation of the placental-fetal unit in humans. Respir Physiol Neurobiol. 304
2011;178(1):51-8. 305
[10] Patel J, Landers K, Mortimer RH and Richard K. Regulation of hypoxia inducible factors 306
(HIF) in hypoxia and normoxia during placental development. Placenta. 2010;31(11):951-7. 307
[11] Tuuli MG, Longtine MS and Nelson DM. Review: Oxygen and trophoblast biology--a source 308
of controversy. Placenta. 2011;32 Suppl 2:S109-18. 309

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
12
[12] Murray AJ. Oxygen delivery and fetal-placental growth: beyond a question of supply and 310
demand? Placenta. 2012;33 Suppl 2:e16-22. 311
[13] Burton GJ and Jauniaux E. Placental oxidative stress: from miscarriage to preeclampsia. J 312
Soc Gynecol Investig. 2004;11(6):342-52. 313
[14] Mason CW, Buhimschi IA, Buhimschi CS, Dong Y, Weiner CP and Swaan PW. ATP-binding 314
cassette transporter expression in human placenta as a function of pregnancy condition. Drug 315
Metab Dispos. 2011;39(6):1000-7. 316
[15] Krishnamurthy P, Ross DD, Nakanishi T, Bailey-Dell K, Zhou S, Mercer KE, Sarkadi B, 317
Sorrentino BP and Schuetz JD. The stem cell marker Bcrp/ABCG2 enhances hypoxic cell survival 318
through interactions with heme. J Biol Chem. 2004;279(23):24218-25. 319
[16] Vaupel P and Mayer A. Hypoxia in cancer: significance and impact on clinical outcome. 320
Cancer Metastasis Rev. 2007;26(2):225-39. 321
[17] Callaghan R, Crowley E, Potter S and Kerr ID. P-glycoprotein: so many ways to turn it on. J 322
Clin Pharmacol. 2008;48(3):365-78. 323
[18] Comerford KM, Wallace TJ, Karhausen J, Louis NA, Montalto MC and Colgan SP. Hypoxia-324
inducible factor-1-dependent regulation of the multidrug resistance (MDR1) gene. Cancer Res. 325
2002;62(12):3387-94. 326
[19] Lye P, Bloise E, Dunk C, Javam M, Gibb W, Lye SJ and Matthews SG. Effect of oxygen on 327
multidrug resistance in the first trimester human placenta. Placenta. 2013;34(9):817-23. 328
[20] Pringle KG, Kind KL, Sferruzzi-Perri AN, Thompson JG and Roberts CT. Beyond oxygen: 329
complex regulation and activity of hypoxia inducible factors in pregnancy. Hum Reprod Update. 330
2010;16(4):415-31. 331
[21] Audette MC, Greenwood SL, Sibley CP, Jones CJ, Challis JR, Matthews SG and Jones RL. 332
Dexamethasone stimulates placental system A transport and trophoblast differentiation in term 333
villous explants. Placenta. 2010;31(2):97-105. 334
[22] Siman CM, Sibley CP, Jones CJ, Turner MA and Greenwood SL. The functional regeneration 335
of syncytiotrophoblast in cultured explants of term placenta. Am J Physiol Regul Integr Comp 336
Physiol. 2001;280(4):R1116-22. 337
[23] Iqbal M, Gibb W and Matthews SG. Corticosteroid regulation of P-glycoprotein in the 338
developing blood-brain barrier. Endocrinology. 2011;152(3):1067-79. 339
[24] Meyer zu Schwabedissen HE, Grube M, Dreisbach A, Jedlitschky G, Meissner K, Linnemann 340
K, Fusch C, Ritter CA, Volker U and Kroemer HK. Epidermal growth factor-mediated activation of 341
the map kinase cascade results in altered expression and function of ABCG2 (BCRP). Drug Metab 342
Dispos. 2006;34(4):524-33. 343
[25] Mathias AA, Hitti J and Unadkat JD. P-glycoprotein and breast cancer resistance protein 344
expression in human placentae of various gestational ages. Am J Physiol Regul Integr Comp 345
Physiol. 2005;289(4):R963-9. 346
[26] Xie Y, Xu K, Linn DE, Yang X, Guo Z, Shimelis H, Nakanishi T, Ross DD, Chen H, Fazli L, Gleave 347
ME and Qiu Y. The 44-kDa Pim-1 kinase phosphorylates BCRP/ABCG2 and thereby promotes its 348
multimerization and drug-resistant activity in human prostate cancer cells. J Biol Chem. 349
2008;283(6):3349-56. 350
[27] Sugiyama T, Shuto T, Suzuki S, Sato T, Koga T, Suico MA, Kusuhara H, Sugiyama Y, Cyr DM 351
and Kai H. Posttranslational negative regulation of glycosylated and non-glycosylated BCRP 352
expression by Derlin-1. Biochem Biophys Res Commun. 2011;404(3):853-8. 353
[28] Draheim V, Reichel A, Weitschies W and Moenning U. N-glycosylation of ABC transporters is 354
associated with functional activity in sandwich-cultured rat hepatocytes. Eur J Pharm Sci. 355
2010;41(2):201-9. 356
[29] Wojtowicz K, Szaflarski W, Januchowski R, Zawierucha P, Nowicki M and Zabel M. 357
Inhibitors of N-glycosylation as a potential tool for analysis of the mechanism of action and 358
cellular localisation of glycoprotein P. Acta Biochim Pol. 2012;59(4):445-50. 359

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
13
[30] Gribar JJ, Ramachandra M, Hrycyna CA, Dey S and Ambudkar SV. Functional 360
characterization of glycosylation-deficient human P-glycoprotein using a vaccinia virus 361
expression system. J Membr Biol. 2000;173(3):203-14. 362
[31] Thews O, Gassner B, Kelleher DK, Schwerdt G and Gekle M. Impact of extracellular acidity 363
on the activity of P-glycoprotein and the cytotoxicity of chemotherapeutic drugs. Neoplasia. 364
2006;8(2):143-52. 365
[32] Hutter D, Kingdom J and Jaeggi E. Causes and mechanisms of intrauterine hypoxia and its 366
impact on the fetal cardiovascular system: a review. Int J Pediatr. 2010;2010:401323. 367
[33] Clifton VL, Vanderlelie J and Perkins AV. Increased anti-oxidant enzyme activity and 368
biological oxidation in placentae of pregnancies complicated by maternal asthma. Placenta. 369
2005;26(10):773-9. 370
[34] Trollmann R, Klingmuller K, Schild RL, Rascher W and Dotsch J. Differential gene expression 371
of somatotrophic and growth factors in response to in vivo hypoxia in human placenta. Am J 372
Obstet Gynecol. 2007;197(6):601 e1-6. 373
[35] Richter HG, Camm EJ, Modi BN, Naeem F, Cross CM, Cindrova-Davies T, Spasic-Boskovic O, 374
Dunster C, Mudway IS, Kelly FJ, Burton GJ, Poston L and Giussani DA. Ascorbate prevents 375
placental oxidative stress and enhances birth weight in hypoxic pregnancy in rats. J Physiol. 376
2012;590(Pt 6):1377-87. 377
[36] Huls M, Russel FG and Masereeuw R. The role of ATP binding cassette transporters in tissue 378
defense and organ regeneration. J Pharmacol Exp Ther. 2009;328(1):3-9. 379
[37] Greer DA and Ivey S. Distinct N-glycan glycosylation of P-glycoprotein isolated from the 380
human uterine sarcoma cell line MES-SA/Dx5. Biochim Biophys Acta. 2007;1770(9):1275-82. 381
[38] Vagin O, Kraut JA and Sachs G. Role of N-glycosylation in trafficking of apical membrane 382
proteins in epithelia. Am J Physiol Renal Physiol. 2009;296(3):F459-69. 383
[39] Beaulieu E, Demeule M, Ghitescu L and Beliveau R. P-glycoprotein is strongly expressed in 384
the luminal membranes of the endothelium of blood vessels in the brain. Biochem J. 1997;326 ( 385
Pt 2):539-44. 386
[40] Schinkel AH, Kemp S, Dolle M, Rudenko G and Wagenaar E. N-glycosylation and deletion 387
mutants of the human MDR1 P-glycoprotein. J Biol Chem. 1993;268(10):7474-81. 388
[41] Seres M, Cholujova D, Bubencikova T, Breier A and Sulova Z. Tunicamycin depresses p-389
glycoprotein glycosylation without an effect on its membrane localization and drug efflux 390
activity in l1210 cells. Int J Mol Sci. 2011;12(11):7772-84. 391
392 393
394

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Table 1. Primers used for real-time PCR. Fig. 1. Placental explant viability and syncytiotrophoblast function. (A) Lactate dehydrogenase (LDH) and (B) Human chorionic gonadotropin (hCG) released from explants into the culture media over the 6-day culture period. Results shown as mean ± S.E.M. Three separate tissues assayed in triplicate. hCG and LDH concentrations were log transformed and analyzed using one-way repeated measures ANOVA with Newman-Keuls post-hoc test. # represents significant difference from day 1 (p<0.05). * represents significant difference from all other days (p<0.05). Fig. 2. Effects of oxygen tension on ABCB1, ABCG2, and VEGF expression. (A) ABCB1 (B) ABCG2 (C) VEGFA mRNA expression in term placental explants normalized to the geometric mean of three housekeeping genes. Placental explants were cultured for 4 days in 20% oxygen, which allowed syncytiotrophoblast shedding and regeneration to take place. The oxygen tension was then reduced to 3% or continued at 20% for 24 and 48 h. Day 4 was considered the control group. There was a significant effect of oxygen tension on ABCB1 and VEGFA expression. Solid bars represent 3% oxygen tension, while open bars represent 20% oxygen tension. * Represents significant difference from control (p<0.05). Results shown as mean ± S.E.M. (n=7) Fig. 3. Effects of oxygen tension on P-gp and BCRP protein expression. Representative western blot of (A) P-gp and (B) BCRP protein from a single placenta. Protein expression of (C) P-gp and (D) BCRP in term placental explants normalized to β-actin. Placental explants were cultured for 4 days in 20% oxygen, which allowed syncytiotrophoblast shedding and regeneration to take place. The oxygen tension was then reduced to 3% or continued at 20% for 24 and 48 h. Day 4 was considered the control group. There was a significant effect of oxygen tension on P-gp expression. Solid bars represent 3% oxygen tension, while open bars represent 20% oxygen tension. * Represents significant difference from control (p<0.05). Results shown as mean ± S.E.M. of relative optical density (ROD). (n=7; *p < 0.05) Fig. 4. Deglycosylation of placental P-gp. (A) Representative western blot comparing P-gp expression in fresh term placental tissue and cultured placental explants. (B) Deglycosylation of fresh placental tissue. (i) Fresh term placental tissue (ii) Denatured protein from fresh placental tissue treated with PNGase F (iii) Non-denatured protein from fresh placental tissue treated with PNGase F (iv) cultured placental explant on Day 6. Fig. 5. Representative immunofluorescent images showing P-gp (green) localization to the syncytiotrophoblast in placental explants exposed to different oxygen tensions. Nuclei were stained by 4’,6-diamidino-2-phenylindole (blue) (n=4). Negative controls were stained by substitution of primary antibody with non-immune mouse IgG. Arrows point to syncytiotrophoblast.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Table 1. Primers used for real-time PCR. Gene Symbol Sequence References ABCB1 Forward: AGC AGA GGC CGC TGT TCG TT 19 Reverse: CCA TTC CGA CCT CGC GCT CC ABCG2 Forward: TGG AAT CCA GAA CAG AGC TGG GGT 19 Reverse: AGA GTT CCA CGG CTG AAA CAC TGC VEGFA Forward: CGG GCC TCC GAAA CCA TGA ACT T 19 Reverse: CCC TCC TCC TTC TGC CAT GGG T TBP Forward: CAC GAA CCA CGG CAC TGA TT 19 Reverse: TTT TCT TGC TGC CAG TCT GGA C GAPDH Forward: AGA TCA TCA GCA ATG CCT CC 19 Reverse: CAT GAG TCC TCC CAC GAT AC YWHAZ Forward: ACT TTT GGT ACA TTG TGG CTT CAA 19 Reverse: CCG CCA GGA CAA ACC AGT AT

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT





























