Embed Size (px)
Citation preview

BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA, Argentina Torno 59, entregas 1& - 2' Diciembre de 1990
ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE
Emesto Medina *
SYNOPSIS
Based on the anatomy and the prevalence of Bromeliaceae on the humid forest from the neotropical mountains and piedmont regions, different evolutive patems have been recognised.
A bromeliaceous evolutive tree is proposed. The ancestral stock is suposed to be exclusively C3 in their metabolic pathway, belonging from an environment of hihg radiation and humidity. Such environment is, today, restricted to the Guiana and the tropical Andes. From there 3 different lines evolved independently, as it can be infered from adaptation toward arid zones or toward less radiated one.
The most primitive members of each line -identified with subfamilies- bear the common characteristic of being C3 from a more or less humid area with high intensity of radiation. The most evolved types of each line exibit adaptation toward arid zones, developing the CAM pathway, or toward environment with less radiation.
All Tillansioideae are epiphytic, so it is suposed that in their particular ancestral stock the epiphytic habit was allready established. Regarding Pitcairnioideae and Bromelioideae their ancestral stock were from terrestrial habitat. The Bromelioide line has a peculiar derivation from CAM of arid zones toward less arid and most umbrageous areas.
SINOPSIS
Las caracteristicas anatomicas y fisiologicas de las Bromeliaceae actuales, asi como su predominancia en habitats epifiticos de bosques humedos montanos y premontanos del neotropico han servido de base para la fonnulacion de diferentes esquemas evolutivos. En estos ,esquemas ocupan lugar preponderante las caracteristicas asociadas con la resistencia a la desecacion (anatomia foliar) y los mecanismos de incorporacion de agua y nutrientes en ambientes epifiticos (presencia de tricomas peltados altamente especializados). En el traba jo se analizan las prop iedades fisiologicas mas relevantes de las Bromeliaceae relacionadas con su alta demanda de luz, absorcion foliar de agua y nutrientes y la ocurrencia de metabolismo CAM en especies epifitas 0
terrestres de habitats xerofiticos, y en especies restringidas a habitats humedos y frecuentemente sombreados. Se discute el valor de la presencia del metabolismo CAM como un marcador evolutivo de caracter conservador, para evidenciar la diferenciacion de grupos de especies en ambientes de alta exposicion solar y baja disponibilidad de agua. Se propone finalmente un esquema de la evolucion de cada subfamilia. El esquema hace enfasis en la diferenciacion de las subfamilias de ancestros que ocupaban habitats con alta exposicion solar, pero con buena disponibilidad de agua 10 cual probablemente favorecio la seleccion de una anatomia foliar resistente a la desecacion. En las Pitcaimioideae se diferenciaron especies CAM muy xeromorfas solo en areas de aridez marcada en Centro y Suramerica. Las especies mas primitivas, restringidas hoy en dia al escudo de Guayana y los Andes tropicales, son exclusivamente C3. Se supone que en las Tillandsioideae el epifitismo aparece muy tempranamente y las formas de sombra actuales se
* Centro de Ecologia. Instituto Venezolano de Investigaciones Cientificas.

72 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
derivaron de fonnas de sol. sin rnetabolisrno CAM. En las Brornelioideae se postula la existencia de grupos prirnitivos C3 de los cuales se derivarian las especies C3 actuales que se han detectado en el genero Greigia. Las especies epifitas actuales de Brornelioideae se derivaron de especies terrestres con rnetabolisrno CAM. al igual que las especies terrestres de sornbra que se encuentran frecuenternente en nurnerosos generos de esta subfarnilia.
INTRODUCCION
Las Bromeliaceae constituyen un grupo de las monocotiledoneas bien diferenciado de distribucion exclusiva en el neotropico, si se exceptua a una especie de Pitcairnia del Africa tropical. Este grupo ha sido particularmente exitoso en la ocupacion de una extensa area geognifica que abarca desde el sur de los Estados Unidos, alrededor de 352 N, hasta el sur de Chile, alrededor de 4511 S (SMITII Y DOWNS, 1974, 1977, 1979). Dentro de este extenso territorio las bromelhiceas han ocupado practicamente todos los nichos ecologicos disponibles (Fig. 1). Se encuentran especies de Bromeliaceae caracteristicas desde las mesetas de arenisca de la formacion Roraima en el escudo de Guayana entre 1000-3000 m de altura, hasta los bosques pluviales montanos y bajos del tropico y subtropico americano; desde paramos htimedos de los Andes tropicales hasta zonas semi-aridas del norte de Argentina y Chile; en Brasil desde bosques htimedos mesotermicos del sur hasta Caatingas semiaridas tropicales del noreste. Esta diversidad de habitats ha sido ocupada por un conjunto de especies terrestres, saxfcolas y epifitas, con marcadas diferencias en su tolerancia a deficits hfdricos, regimenes de luz y disponibilidad de nutrientes durante el crecimiento.
CARACTERISTICAS ANATOMICAS GENERALES Y DIFERENCIACION TAXONOMICA
Los estudios anatomicos muestran a las Bromeliaceae como una "familia natural y bien diferenciada" caracterizada por la presencia de tricomas foliares peltados de morfologia muy variable los cuales poseen un largo pedicelo uniseriado completamente embutido en la epidermis (TOMLINSON 1969) (Fig. 2). Las celulas epidermicas poseen paredes marcadamente sinuosas, las interiores mas engrosadas que las exteriores, y usualmente incluyen un cuerpo silicoso grande. Las hojas de las Bromeliaceae presentan una hipodermis diferenciable en secciones extema e intema, la ultima desarrolla por 10 general un tejido acuifero profundo en la region adaxial de la hoja (Fig. 3). El tallo incIuye una capa meristematica bien diferenciada asociada con la insercion de trazas radicales entre la corteza y el cilindro central. La corteza incluye numerosas raices intracaulinares.
El desarrollo de tejidos acufferos que determinan la formacion de hojas de anatomia suculenta, y la aparicion de epidermis con superficies impermeabIes (cutinizacion) son caracteres de especial importancia para la evolucion de la familia en habitats secos, cuya significacion no ha sido establecida cuantitativamente (MEDINA .1989).
En las Bromeliaceae se distinguen tres subfamilias bien caracteristicas (SMITII y DOWNS 1974) (Tabla 1). Es notable la tendencia del epifitismo dentro de la familia, habito que llega a ser frecuente en las Bromelioideae, y es

ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 73
muy dominan~ en las Tillandsioideae. Esta tendencia se encuentra asociada con la reduccion de la funcionalidad de las raices como organos de absorcion. Las formas terrestres en Pitcaimioideae y Bromelioideae presentan raices funcionales, mientras que en las formas epifitas las raices funcionan solo como organos de adhesion al substrato. La estructura de las escamas peltadas alcanza su maximo grado de diferenciacion dentro de las Tillandsioideae, donde tambien se ha demostrado una mayor capacidad de absorcion de agua y nutrientes (BENZING 1976). Los tipos de fruto y semilla son tambien caracteristicos de cada subfamilia. En las Bromelioideae los frutos son frecuentemente camosos con semillas desnudas, mientras que en las otras dos subfamilias estos son capsulares y con semillas aladas (Pitcaimioideae) 0 con apendices plumosos bien desarrollados (Tillandsioideae). Los frutos camosos son dispersados predominantemente por animales, mientras que la presencia de apendices seminales favorece la dispersion anemocora.
Las caracteristicas morfologicas y anatomicas antes mencionadas han servido de base para elaborar la mayoria de los esquemas de la diferenciacion evolutiva de la familia Bromeliaceae. La ausencia de un record fosil apropiado, y la dificultad de estudios citogeneticos por 10 diminuto y numeroso de sus cromosomas (MCWILLIAMS, en SMITH y DOWNS 1974, p. 33) ha retardado la aplicacion de estos criterios en la filogenia de la familia. Sin embargo, se ha podido establecer que el numero basico de las espccies existentes es 25 (MARCHANT 1967) Y los valores menores encontrados en la literatura se deben a reduccion por aneuploidia (BROWN y GIU.1ARTIN 1989). Parece claro que las subfamilias Pitcaimioideae y Tillandsioideae son homogeneas y bien diferenciadas entre sf desde el pun to de vista citogenetico (MARCHANT 1967),10 cual se corresponderfa con el contraste entre habitats esencialmente terrestres de las Pitcaimioideae y epifitos de las Tillandsioideae. Las Bromelioideae son citogeneticamente mas heterogeneas y ocupan un lugar intermedio entre las otras dos subfamilias (MARCHANT 1967). Esta ultima sub familia es intermedia desde el punto de vista del grado de desarrollo del epifitismo y la ocupacion de habitats expuestos y secos y/o humedos y sombreados.
Un intento reciente de aplicacion de criterios quimiotaxonomicos basado en la complejidad de los flavonoides ha dado resultados contradictorios. WILLIAMS (1977) establece que la distribuci6n de clases de flavonoides permite diferenciar las 3 subfamilias. Las Bromelioideae se consideran mas primitivas pues solo contienen flavonoles comunes y flavonas, mientras que las Pitcaimioideae poseen adem as flavonas C-glicosidos y G-hidroxiflavonas, y las Tillandsioideae, que aparecen como mas avanzadas, contienen adem as gosipetina, patuletina y derivados metilados de 6-hidroximiricetina. Analisis de otros grupos de pigmentos similares, sin embargo, no permite un orden amiento similar (SAITO y HARBORNE, 1983). Por otra parte, la aplicacion de tecnicas cladisticas para clarificar la filogenia de la familia muestra que las subfamilias Tillandsioideae y Bromelioideae son taxa hermanos mientras que las Pitcaimioideae se encuentran mas cerca del ancestro supuesto de la familia (GILMARTIN y BROWN 1987). Los anal isis conducen tam bien a la proposicion de vias evolutivas que indican el desarrollo de la xeromorfia como caracter derivado en las Tillandsioideae y Pitcaimioideae (GILMARTIN, 1983; GILMARTIN y BROWN, 1986; VARADARAJAN y GILMARTIN 1988). En esta ulti-

74 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
rna subfamilia la separacion de taxa xeromorficos y mesomorficos es bien clara (VARADARAJAN y GILMARTIN 1988) Y se corresponde con la separacion de tipos fisiologicos propuesta por MEDINA (1974).
En este trabajo se utilizan caracteristicas fisiologicas como base para entender la diferenciacion evolutiva de la familia. Para ello analizaremos la informacion disponible sobre distribucion ecologica, caracterfsticas anatomicas, relaciones hidricas y metabolismo fotosintetico. La discusion lOrna en cuenta los argumentos de tipo eco-fisiologico sobre evolucion de la familia Bromeliaceae que han sido desarrollados desde la publicacion del trabajo clasico de SCHIMPER (1888) y que han sido modificados 0 extendidos por -TIETZE (1906), PrTTENDRIGH (1948), BENZING Y RENFROW (1971), MEDINA (1974, 1987), GRIFFITHS y SMITII (1983); MEDINA et al. (1989), BENZING (1989) y SMITH (1989). Tambien se incluyen las recientes contribuciones del amilisis cladistico para el entendimiento de las relaciones de las Bromeliaceae con otras familias de monocotiledoneas y la separacion de las tres subfamilias que la componen (GILMARTIN y BROWN 1987).
ECO-FISIOLOGIA Y DISTRIBUCION
La diferenciacion de las Bromeliaceae parece estar relacionada con el desarrollo de estructuras y mecanismos de conservaci6n de agua y de carbono incorporado y del paso de formas de vida terrestres a epifitas.
Se observa en la familia un alto grado de especializacion anatomica y morfologica, tal como el desarrollo de suculencia foliar (peso fresco/superficie, agua/superficie 0 peso fresco/peso seco), presencia de densa cobertura de escamas peltadas, que permiten incorporar agua y nutrientes eficientemente por 10 men os en las Tillandsioideae, y que a la vez incrementan la reflexion de radiacion solar de las hojas disminuyendo as! el diferencial de temperatura hoja-aire. Ya hemos hecho referencia tambien al desarrollo de semillas con apendices alados 0 plumosos que favorecen la dispersion anemocora de semillas livianas, 0 el desarrollo de frutos baccarios dispersados por animales.
El desarrollo del epifitismo es notable en las Bromelioideae y Tillandsioideae y su evoluci6n parece haber sido independiente en estas subfamilias (BENZING 1989). Este habito aparece asociado con el desarrollo de tricomas peltados, que actuan pasivamente como vruvulas de agua unidireccionales, 0 la formacion de tanques foliares que permiten la acumulacion de agua de lluvia.
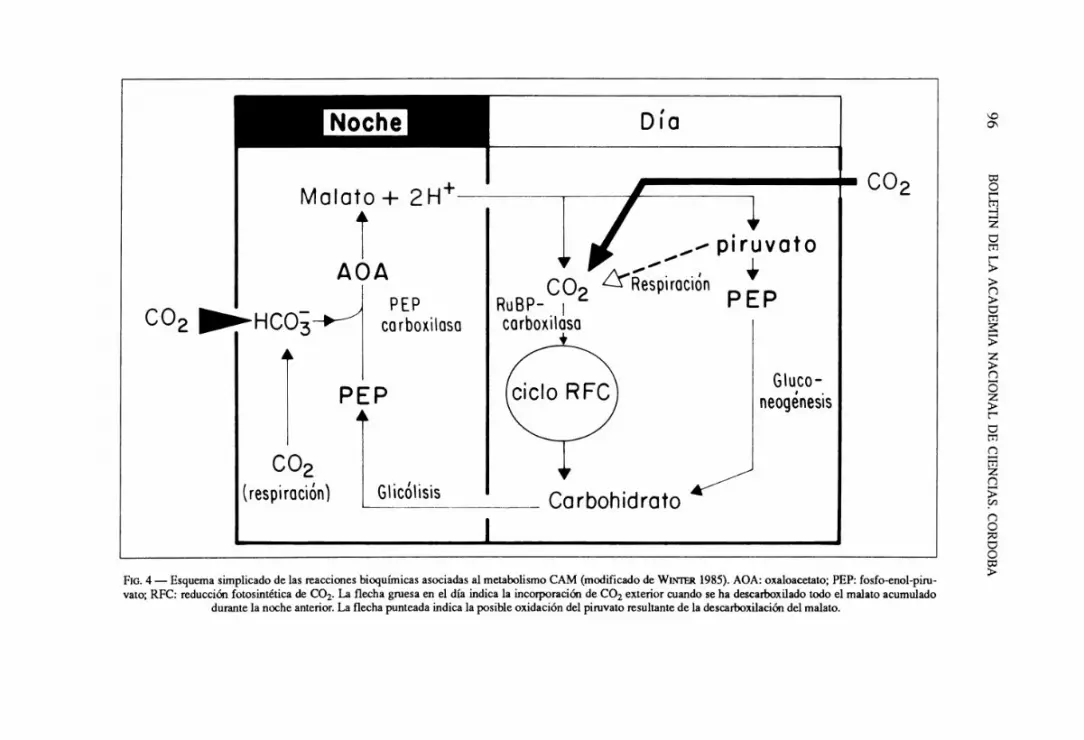
La ocupacion de habitats secos, terrestres 0 epifiticos, ha sido favorecida por el desarrollo de la capacidad de fijacion nocturna de CO2 (Metabolismo Acido de Crassulaceae, CAM). El CAM es un proceso caracterizado por la acumulaci6n de acidos organicos libres, esencialmente acido malico y bajo ciertas condiciones acido citrico, en las vacuolas de tejidos fotosintcticamente activos, como producto de la fijacion noctuma de CO2 mediada por la enzima fosfo-enol-piruvato carboxilasa (OSMOND, 1978; WINTER 1985) (Fig. 4). El acido malico acumulado durante la noche en las vacuolas de celulas fotosinteticas entra al citoplasma al dia siguiente, donde es decarboxilado y el C02 resultante es fijado en los cloroplastos por la enzima ribulosa-difosfato carboxilasa (RUBISCO) e incorporado a carbohidratos en el cido de CALVIN. El resultado es que la apcrtura estomatica y la fijacion de CO2 ocurren durante

ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 75
la noche, cuando la demanda evaporativa de la atmosfera es menor, y la fotosintesis de carbohidratos se produce al dia siguiente. Por ello la eficiencia de uso de agua (cantidad de materia organica sintetizada por unidad de agua transpirada) es mucho mayor en aquellas plantas que poseeen este tipo de metabolismo.
La presencia del metabolismo CAM en una especie dada puede detectarse mediante el fenomeno de fijacion noctuma de CO2 e incremento noctumo de la acidez tisular. Estos procesos esrnn asociados a un ciclo diario invertido de apertura estomatica, es decir, apertura nocturna y cierre al menos durante las primeras horas del dia (WOLF 1960; OSMOND 1978; KLUGE y TING 1978; WINTER 1985; LOTTGE 1988). La fijacion noctuma de CO2 a traves de PEPcarboxilasa, da por resultado que las plantas CAM presenten valores relativamente bajos de discriminacion de carbona 13, esencialmente como resultado del fraccionamiento causado por la difusion del CO2 hasta el interior de la hoja. La discriminacion de carbona 13 en tejidos de plantas se expresa mediante el valor ol3C en por mil (0/00), el cual es negativo, pues corresponde ala comparacion con un patr6n de carbonato f6sil cuyo valor se asume arbitrariamente como cero. Valores de o13C mas negativos representan mayor discriminaci6n respecto a carbona 13 en el proceso de fijaci6n de CO2.
METABOLISMO FOTOSINTETICO
El analisis del metabolismo fotosintetico de Bromeliaceae tiene mucha significaci6n para comprender la evoluci6n de la familia. PITTENDRIGH (1948) infiere de sus estudios sobre la distribuci6n de Bromeliaceae epifitas en Trinidad, que en esta familia predomina una alta demanda de luz, aunque muchas especies son tolerantes a la sombra. La tolerancia a la sombra se deriva de la sensibilidad al deficit hidrico frecuente en habitats epifiticos. Segun PITTENDRIGH esta alta demanda de luz general en la familia seria indicativa de que los ancestros de las bromelias actuales deben haber evolucionado en ambientes de alta exposicion solar. (BENZIl'\G y RENFROW (1971) midieron los requerimientos de luz de varias especies de Tillandsioideae diferenciadas par las caracterfsticas luminicas de sus habitats naturales y por la variaci6n en la densidad de la cubierta de escamas peltadas. Estos autores encuentran que las especies predominantemente de sombra, can hojas delgadas, y con baja densidad de escamas a1canzaban alrededor de 50% de su maxima capacidad fotosintetica a intensidades de luz correspondientes a 1/3 de la maxima luz solar, mientras que las especies mas xeromorfas, con alta densidad de escamas foliares, alcanzaban menos del 30% de su fotosintesis maxima en las mismas condiciones. La explicaci6n mas probable de este resultado es que la cubierta de escamas determina una mayor reflectividad foliar, 10 cual resulta en una menor cantidad de luz absorbida por la hoja (BENZING y RENFROW, 1971).
Parte de las aparentes contradicciones en los resultados de las mediciones de intercambio gaseoso en Bromeliaceae derivan de la comparaci6n de plantas con fijaci6n diuma y fijaci6n nocturna de CO2. COUTIl'\HO (1963, 1969), MCWILLIAMS (1970), MEDINA (1974) MEDIl'\A et al. (1977), y GRIFFITHS Y SMITH (1983) estudiaron la ocurrencia de CAM en numerosas especies de la familia conjuntamente can especies de metabolismo fotosintctico de tipo C3, es decir, solo con fijacion diurna de CO2. Uno de los procesos

76 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
mas resaltantes que ocurre en estas plantas CAM es la acidificacion notable delliquido celular que ocurre durante la noche. El anal isis de la composicion de acidos organicos durante el dfa y la noche muestra que el acido malico es el principal responsable de los cambios de acidez (Fig. 5) llegando a constituir entre 70-100% del cambio de acidez total (Tabla 2). El acido cftrico puede tambien contribuir a los cambios de acidez noctuma, particularmente en Pitcaimioideae y Bromelioideae (Fig. 5) como se observa frecuentemente en especies de Crassulaceae (WOLF, 1960). Recientemente se ha confirmado el papel del acido cftrico en algunas especies de Bromeliaceae, particularmente dentro de la subfamilia Bromelioideae (LEE et al .. 1989). La acumulacion de este ultimo acido representarfa una altemativa energeticamente mas eficiente especialmente en condiciones de sequia (LOTTGE, 1988).
La fijacion noctuma de CO2 tiene consecuencias impartantes para la eficiencia de uso de agua, ya que los estomas se abren cuando la demanda evaparativa de la atmosfera es menor. La capacidad productiva de plantas CAM, sin embargo, es comparativamente menor a las de las plantas C3, ya que la cantidad de CO2 que puede fijarse durante la noche esm limitada por el suministro de carbohidratos que sirven como substrato para la carboxilacion, y la capacidad de la vacuola para acumular acidos organicos. Bajo ciertas condiciones, sin embargo, la capacidad productiva es comparable a la de plantas C3, por ejemplo, en plantas CAM con suministro adecuado de agua y nutrientes, como ocurre en el caso de los cultivos comerciales de Ananas comosus. 0 en condiciones de sombra, donde la fotosfntesis esm limitada por luz (MEDINA et al .. 1989).
CAM COMO MECANISMO DE CONSERVACION DE CO2 BAJO CONDICIONES DE SEQUIA
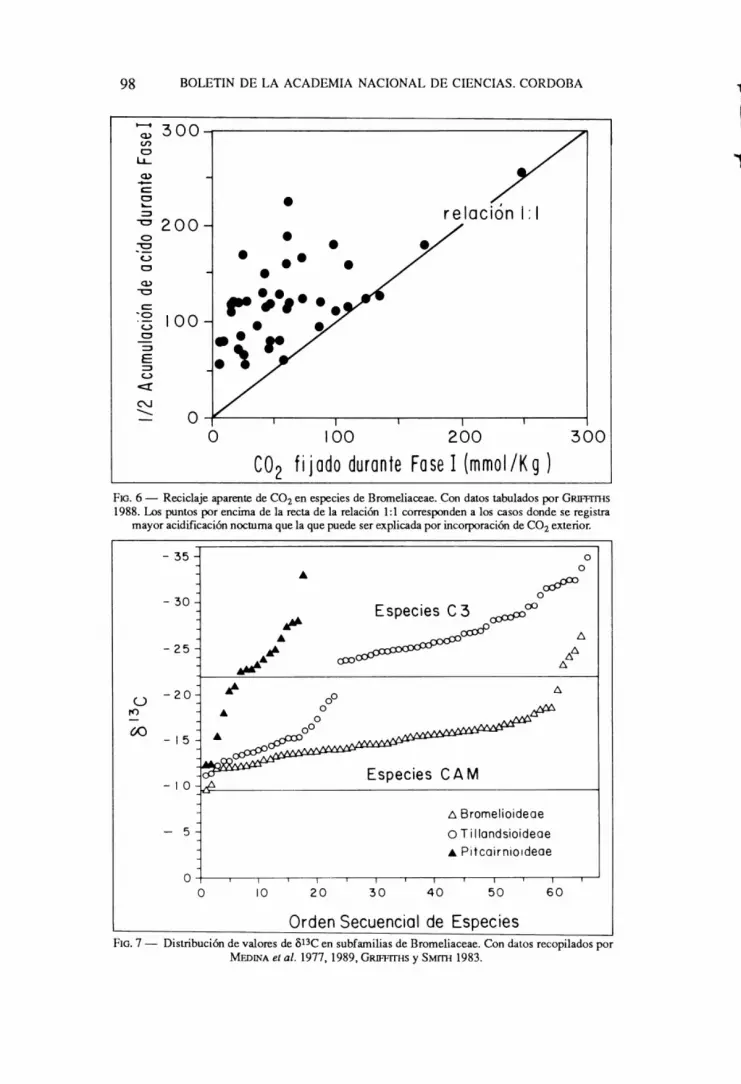
La repeticion de perfodos secos de duracion variable en habitats epiffticos 0 terrestres, particularmente en climas semi-aridos, trae como consecuencia que los estomas permanecen cerrados durante gran parte del dfa. Durante estos perfodos la respiracion continua pues el mantenimiento de la estructura celular rcquicre de un suministro de energia constante. En plantas con fijacion diurna de CO2 esto significa una pcrdida neta de carbono proveniente de las reservas de carbohidratos movilizables. En cacmceas CAM se demostro que ejemplares sometidos a sequias prolongadas se akanzaba un estado donde el intercambio neto de CO2 era nulo, pero se segufan observando cambios diarios en la acidez tisular (SZAREK et al .. 1973). Este mecanismo de reciclaje del CO2 respiratorio tiene especial significacion para la supcrvivencia de plantas en climas secos, pues constituye un efectivo medio de conservacion de reservas de carbohidratos. EI fenomeno de reciclaje de CO2 respiratorio ha sido documentado ampliamente por GRIFFITHS (1988) quien demuestra que el fenomeno se puede detectar en condiciones normales de crecimiento en Bromeliaceae CAM epifitas y terrestres (Fig. 6). Por otra parte, el proceso tiene tam bien significacion para la conservacion de carbona respiratario, pues las plantas CAM tienen una alta demanda de ATP durante la noche para la acumulacion de acido malico en las vacuolas (aproximadamente un mol de ATP par mol de acido malico transportado a la vacuola (LOTTGE y BALL, 1987).

ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 77
La estimacion de la recirculacion de CO2 respiratorio se hace midiendo simultaneamente durante la noche la incorporacion de CO2 y la acumulacion de acidez titulable en tejidos fotosinteticamente activos. EI cociente entre CO2 fijado y 1/2 de la acidez titulable acumulada representa la fraccion del incremento de acidez resultante de la recirculacion de CO2 respiratorio (GRIFFITHS, 1988). Esto es valida cuando cada CO2 fijado genera solo 2 equivalentes de acidez, es decir, cuando el unico acido responsable de la acidificacion es el acido malico (Fig. 4). Si se produce una cierta fraccion de acido citrico, una unidad de CO2 generaria 3 equivalentes de acidez. Por esto es indispensable conocer la proporcion de acidos malico y citrico en la acidificacion noctuma para poder determinar la magnitud de la refijacion de CO2 respiratorio. Una manera de resolver este problema seria la medicion de la fijacion de marcado con 14C y la medida de la actividad especifica del malato y el citrato acumulados durante la noche.
EI analisis bioquimico del fenomeno de reciclaje de CO2 respiratorio es aun incompleto, y todavia no hay una explicacion satisfactoria al hecho de que, con la tecnica descrita anteriormente, en muchos casos el reciclaje de CO2 no puede detectarse (puntos sobre 0 por debajo de la diagonal en la Fig. 6).
RELACIONES HIDRICAS DE BROMELIACEAE EPIFITAS Y TERRESTES
A pesar de las caracteristicas xericas de habitats epiffticos, aun en bosques tropicales Mmedos, las Bromeliaceae epifitas se caracterizan por tensiones xilematicas reducidas cuya magnitud presenta solo pequei'ias oscilaciones diarias (SMITII et aI., 1985, 1986) (Tabla 3). La presion osmotica del juga vacuolar es correspondientemente baja pero cambia significativamente durante el dfa en aquellas especies que presentan acidificacion noctuma de sus tejidos foliares. Las variaciones diumas de la tension del xilema, y la presion osmotica promedio del juga vacuolar es mayor en aquellas especies que crecen bajo exposicion en comparacion con aquellas de habitats parcialmente sombreados por el dosel del bosque. La sequia afecta similarmente los parametros hidricos de especies mesoffticas y xerofiticas. En algunas especies, sin embargo, la sequia induce tambien cambios metabolicos que se reflejan en la acumulacion de acidos organicos en las vacuolas celulares. El estudio de SMITII et al. (1985) confirmo experimentos previos en Guzmania monostachia (MEDINA et al. 1977) que indicaban que bajo condiciones de deficit hidrico esta especie era capaz de desarrollar actividad CAM.
En general, especies CAM epifitas son mas resistentes a la sequfa que las especies C3. Si se grafica la frecuencia de bromelias epifitas C3 y CAM contra la precipitacion promedio a 10 largo de gradientes altitudinales en Trinidad se observa claramente que las bromelias CAM dominan a altitudes menores y habitats expuestos, donde la sequia potencial tiene mayor probabilidad de ocurrir (GRIFFITHS y SMITH, 1983; MEDINA et al. 1987).
En especies de Bromeliaceae terrestres las relaciones hidricas son similares a las de bromelias epifitas, aun en habitats semi-aridos. Las diferencias en presion osmotica del juga celular durante las epocas de lluvia y sequia en poblaciones de Bromelia humilis de habitats expuestos en el norte de Vene-

78 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
zuela se encuentran alrededor de 0,4 MPa para el amanecer y 0,2 MPa para el atardecer (Tabla 4). Por otra parte, las diferencias en tension del xilema al amanecer son muy reducidas.
Es evidente que las Bromeliaceae, tanto C3 como CAM, sc comportan como suculentas, pues su jugo vacuolar tiene concentraciones bajas, 10 que resulta en presiones osmoticas maxim as de alrededor de 1 MPa, mientras que la tension xilematica por 10 general es menor de 1 MPa, aun en ambientes muy secos.
Los estudios de las relaciones hfdricas en Bromeliaceae demuestran que tanto la proteccion contra la desecacion derivada de la anatomia foliar, como el CAM, contribuyen al mantenimiento de un presupuesto hfdrico balanceado. Como se vera mas adelante, la penetracion de habitats sombreados por bromelias CAM resulta en la perdida parcial de las estructuras que protegen ala planta contra perdidas de agua excesivas. Esas especies son mucho mas sensibles al deficit hfdrico a pesar de la actividad CAM.
INTERACCIONES ENTRE DISPONIBILIDAD DE AGUA Y EL AMBIENTE LUMINICO
Las interacciones entre disponibilidad de agua y capacidad productiva de Bromeliaceae CAM ha sido demostrada en especies terrestres que habitan en areas semi-aridas en los tropicos (MEDINA et ai. 1986b), pero las relaciones deben ser similares en plantas epifitas (MEDINA, 1987). Bromelia humilis es una Bromelioideae terrestre que crece en ambientes semi-aridos bajo completa exposici6n solar. Esta especie ha side descripta como una especie formadora de tanque, con rakes que se dcsarrollan y crecen entre las axilas foliares de las rosetas. Estas rakes son capaces de absorber agua y nutrientes del tanque 10 cual hace a la planta virtual mente independiente del suelo (PITTENDRIGH, 1948; MEDINA et al. 1986b; LEE et ai., 1989). Esta capacidad explica su presencia en suelos salinos, los cuales sedan toxicos para la planta si las rakes tuviesen que crecer en ellos. Bajo tales condiciones el agua sc convierte en el principal factor limitante y reduce efectivamente la capacidad productiva y el intercambio gaseoso. Plantas de B. humiiis que crecen bajo sombra parcial de especies lei'iosas en zonas semi-aridas del nor-oeste de Venezuela, presentan mayores contenidos de clorofila y nitrogeno que aquellas de poblaciones que crecen a plena exposici6n (Tabla 5). De igual manera, su capacidad fijacion noctuma de CO2, reprcsentada por la acidificacion nocturna, es mayor en las plantas bajo sombra parcial. El nivel de hexosas disminuye durante la noche, en parte son utilizadas como fuente de piruvato para sfntesis de acidos organicos, y en parte son trasladadas hacia otros organos. La disminucion del contenido de hexosas durante la noche debe ser por 10 menos igual al incremento en los equivalentes de acidez (en rclaci6n 4:1 si es malico el unico acido responsable de la acidificaci6n). Las diferecias entre los dos tipos de planta se reducen durante la epoca de lluvias, cuando el agua no es factor limitante. Durante la epoca de sequia es tambien posible demostrar que la capacidad de acidificacion nocturna se restaura rapidamente despues de irrigar las plantas experimentalcs (MEDINA et ai., 1986b). La conclusion es que en aquellos habitats en los cuales el agua es el factor limilante de la productividad vegetal, plantas CAM crecen mejor bajo condiciones de

ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 79
sombra parcial, en donde la demanda evaporativa de la atmosfera es menor y la disponibilidad de nitrogeno es mayor. Este ejemplo permite visualizar tam bien como podrfa ocurrir la ocupacion de habitats sombreado por plantas CAM esencialmente de sol.
EL METABOLISMO CAM EN LAS BROMELIACEAE COMO MARCADOR DE LA EVOLUCION EN AMBIENTES ARID OS
Es po sible utilizar directamente los valores de 813C para determinar la distribucion de metabolismo CAM en Bromeliaceae, ya que en la familia no se presentan vainas vasculares fotosinteticas como en el caso de las plantas C4 (MEDINA et at., 1977). Esta tecnica es de gran utilidad para analisis de gran numero de especies, particularmente cuando no se pueden hacer mediciones fisiologicas detalladas. En general, valores de 813C comprendidos entre -9 y -22O/w corresponden a plantas CAM en las Bromeliaceae.
La recopilacion de los valores de 813C medidos en especies de Bromeliaceae permite observar un patron que ya habfa sido identificado por MEDINA (1974) (Fig. 7). La mayorfa de las especies de Pitcairnioideae estudiadas poseen metabolismo fotosintetico C3, particularmente las de aquellos generos cuya distribucion esta restring ida al escudo de Guayana (Tabla 6). En las Bromelioideae, por el contrario la gran mayorfa de las especies medidas son CAM, mientras que en las Tillandsioideae, aunque predominan las especies C3, hay un numeroso grupo de especies CAM. En la Fig. 7 hemos colocado el valor de 813C de -22O/w como una delimitacion entre plantas C3 y CAM, pero ella es relativamente arbitraria por dos razones. Primero dentro de cada grupo pueden existir especies en las cuales el metabolismo CAM pueda ser inducido por condiciones de crecimiento determinadas (MEDINA et at., 1977). En segundo lugar el valor especffico medido puede variar de acuerdo a condiciones de crecimiento tales como intensidad de luz, stress de sequfa, y en condiciones de bosques densos la concentraci6n de CO2 y el 813C del CO2 en el aire circundante (MEDINA et at., 1986a).
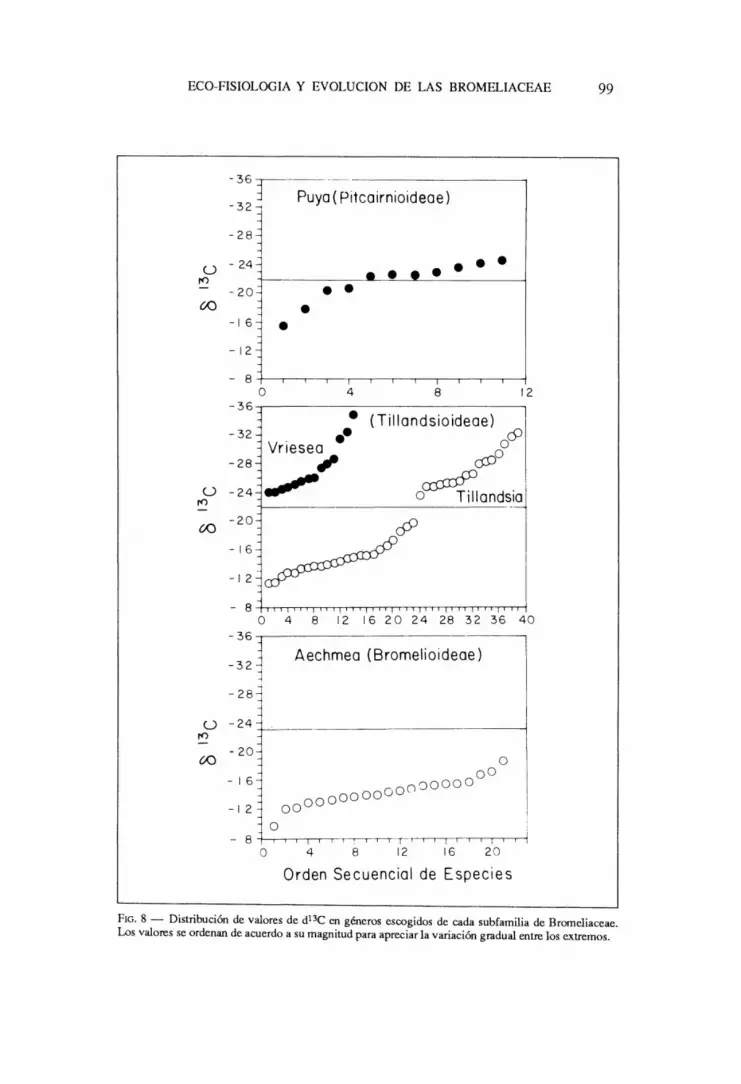
Notese ademas la gran variacion en los valores especfficos dentro de cada grupo que van desde -10 hasta -22 en especies CAM y desde -22 hasta -36 en especies C3. Esta variaci6n es de gran interes ecofisiol6gico pues sugiere diferencias en los patrones de incorporacion de carbona en condiciones naturales. En algunos generos donde se ha medido un numero importante de especies puede tambien observarse esta variacion muy claramente (Fig. 8). En el genero Puya las especies que se encuentran distribuidas en los Andes del norte son claramente C3, mientras que en aquellas de los Andes del sur, en climas mucho mas secos, hay una clara tendencia a valores de 813C indicativos de fijacion nocturna de CO2. En el genero Tillandsia hay una clara diferenciacion entre especies C3 de habitats mas humedos y especies CAM de habitats mas secos. En el genero Vriesea, a pesar de su similitud morfologica con el genero Tillandsia, la mayorfa de las especies estudiadas son {}e tipo C3, aunque los valores disponibles varfan marcadamente desde -24 hasta -36o/w . Recientemente se han encontrado algunas especies dentro del grupo de las llamadas "Vrieseas grises" segun Rauh (1981) presentan fijacion nocturna de CO2 (H. GRIFFITHS, comunicacion personal). Estas especies de Vriesea poseen un habito y una distribuci6n ecologica muy similares a especies del genero Tillandsia

80 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
subgenero Tillandsia. La convergencia morfologica proviene del desarrollo de hojas triangulares densamente cubiertas por tricoma<; peltados. Este grupo 10 constituyen unas 32 especies bien diferenciadas dentro de las secciones X iphion y Vriesea (SMmI Y DOWNS 1977) Y ciertamente merccen un estudio ecofisiologico detallado. El genero Aechmea de las Bromelioidcae presenta solo especies CAM, pero que ocurren en una gran diversidad de habitats, desde semi-aridos hasta bosques humedos, y correspondientemente presentan una fuerte variacion en los valores de 813C.
Las correlaciones entre posicion dentro de la familia, la distribucion ecologica de las especies y las caracterfsticas fisiologicas permite hacer una estimacion del potencial de especies con CAM en cada una de las subfamilias. La distribucion y numero de especies de los generos descriptos por SMITH y DOWN (1974 - 1979) (Tabla 6) y el comportamiento fisiologico de especies caracterfsticas dentro de la mayorfa de los generos permite estimar que en las Pitcairnioideae aproximadamente 173 del total de 731 especies descriptas (24%), y en las Tillandsioideae 202 de las 800 especies conocidas (27%), poseen probablemente CAM. Por el contrario, en las Bromclioideae 465 de las 556 especies descriptas presentan probablemente CAM (84%).
ESPECIALIZACION MORFOLOGICA, EPIFITISMO, XEROFITISMO Y EVOLUCION
La diversidad de formas biologicas, la presencia de numerosas caracterfsticas anatomicas especializadas, y la abundancia de especies en una gran diversidad de habitats, constituyen un caso de especial valor para estudiar las relaciones entre distribucion ecologica y diferenciacion evolutiva dentro de un grupo de plantas taxonomicamente homogeneo. Este ha sido ciertamente el caso en el estudio de las especies epifitas, las cuales constituyen una comunidad muy notoria en los bosques montanos humedos del neotropico. En un estudio clasico SCHIMPER (1888) analiza las restricciones ecologicas que caracterizan el habitat epifftico en bosques tropicales. EI estudio demuestra que dentro de las Tillandsioideae la presencia de tricomas peJtados capaces de absorber agua en forma lfquida y nutrientes disueltos constituye una efectiva sustitucion de la funcion de las rakes en e~os habitats. Las escamas peltadas, principalmente en las Tillansioideae, funcionan como el mecanismo principal para la incorporacion de agua y nutrientes en esos habitats deficientes en nutrientes y con sequfas recurrentes (MHz, 1904; BENZING, 1976).
SCHIMPER (1888) propuso una hipotesis que explica la evolucion del habito epifftico como dependiente primero del modo de dispersion de semilIas, y segundo de la capacidad para sobrevivir con bajos suministros de nutrientes y sequfas eventuales, particularmente durante las primeras etapas del establecimiento de las plantulas. Segun esta hipotesis las epifitas deberfan poseer semillas livianas provistas de apendices que faciliten la dispersion por el viento, y tam bien estructuras que Ie permitiesen a estas semillas adherirse a la superficie de los arboles huesped, 0 las semillas deberfan estar contenidas en frutos camosos, los cuales podrfan ser comidos y dispersados por animales. Las especies epifitas de las subfamilias Tillandsioideac y Bromelioideae poseen exactamente estas caracteristicas. Las Tillandsioideae produ-

ECO-FlSIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 81
cen frutos capsulares que liberan numerosas semillas livianas, con apcndices plumosos bien desarrollados, apropiadas para la dispersion por el viento. Las Bromelioideae poseen por 10 general frutos indehiscentes, baccarios, frecuentemente camosos, apropiados para la dispersion por animales.
SCHIMPER noto tambien que hay una clara diferencia morfologica de las especies epifitas en gradientes verticales en bosques humedos. Aquellas especies del dosel bajo tienen menor densidad de escamas y color predominantemente verde, mientras que aquellas que habitan las ramas superiores, bajo condiciones de alta iluminacion, tendfan a ser mas xeromorfas, con alta densidad de escamas, 10 cual Ie da a la planta un color grisaceo. SCHIMPER concibio al desarrollo del epifitismo como un proceso que solo podria tener lugar bajo condiciones de alta humedad (bajo condiciones sombreadas del bosque) para que pudiese ocurrir un establecimiento exitoso de la plantula en un ambiente en donde el suministro de agua es necesariamente mas limitado. Especies epifitas desarrolladas bajo tales condiciones podrfa subsecuentemente dar origen a las caracterfsticas mas xeromorfas requeridas para la supervivencia en el ambiente mas exigente de las ramas superiores. Las especies epifitas de las ramas expuestas podrfan entonces colonizar bosques mas secos (tales como sabanas en la opinion de SCHIMPER) 0 ambientes terrestres 0 saxfcolas donde la disponibilidad de agua constituye el factor limitante primario.
Basado en la diversificacion de habitats en la flora de bromelias epifitas de Trinidad, PITTENDRIGH (1948) fue el primero en ofrecer una explicacion altemativa del posible desarrollo del epifitismo en especies de Tillandsioideae. Este autor basa sus argumentos en el alto requerimiento de luz de la mayorfoa de las especies de bromelias por el estudiadas. Su presencia en ambientes parcial- 0 totalmente sombreados era la consecuencia de su incapacidad para mantener un presupuesto hfdrico balanceado bajo condiciones de exposici6n completa. Ademas, PITTENDRIGH argiiia que la morfologfa floral de las especies xeroffticas de Tillandsia es mas primitiva que la de las especies tfpicas de los ambientes hUmedos y sombreados. La concepcion de PITTENDRIGH de la evolucion de las Bromeliaceae, particularmente de las Tillandsioideae epifitas, comparte el concepto basico de TIETZE (1906) quien opina que las formas mas primitivas de la familia son "xerofitas terrestres". De este tipo se desarrollo la morfologfa de tanque que permite la retencion de cantidades considerables de agua de precipitacion, por perfodos mas largos, reduciendo con ella la dependencia de la planta de fuentes de agua del suelo. En este proceso se produce el desarrollo de tricoma peltado hasta alcanzar la funcion altamente especializada de organa de absorcion de agua y nutrientes. Tanto la posesion de morfologfa de tan que y el desarrollo de tricomas absorbentes fueron las principales caracterfsticas que permitieron la ocupaci6n exitosa de habitats epiffticos en bosques hUmedos y secos.
MEDINA (1974) llama la atenci6n sobre el desarrollo de una propiedad fisiologica, la capacidad de fijacion noctuma de CO2 (metabolismo CAM), que parece tener una distribucion particular dentro de la familia, relacionada tanto con la filogenia como con la separacion de habitats. Se observa que el CAM no se encuentra en los miembros mas primitivos de la familia, y tampoco ocurre con frecuencia en las especies de sombra tipicas de bosqucs hUmedos. EI CAM aparece como un caracter derivado, el cual pucdc seT utilizado como un marcador evolutivo, conjuntamente con la morfologia floral,

82 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
la estructura foliar, la estructura de las escamas peltadas, y los requerimientos de luz para dilucidar los patrones de evolucion dentro de la familia. El caracter marcador se deriva de la siguiente proposicion: el CAM solo puede haber evolucionado en ambientes secos con alta exposicion solar, y ademas constituye un caractcr conservador, es decir, que se mantiene aun en aquellos casos que los grupos en los que se desarrollo invadan, en el curso de su historia evolutiva, habitats hUmedos y sombreados. Un corolario de esta hipotesis es que la presencia de metabolismo CAM por sf misma no representa una desventaja competitiva respecto a plantas con metabolismo fotosintetico C3 en habitats sombreados (MEDINA et al .. 1989). El potencial de desarrollo de CAM es una caracterfstica de toda la familia, y parece claro que el CAM ha aparecido independientemente dentro de las tres subfamilias.
Para las consideraciones que siguen se acepta la proposicion que la subfamilia Pitcairnioideae es mas primitiva (HARMS 1930, SMITH 1934, VARADAJARAN Y GILMARTIN 1988) aunque probablemente no constituye el grupo ancestral para las otras dos subfamilias (PITTENDRIGH, 1948). Las Tillandsioideae y Bromelioideae son grupos mas evolucionados que aparentemente han seguido lineas de diferenciacion bien distintas.
MODELO HIPOTETICO DE LA EVOLUCION DE LAS BROMELIACEAE
En la evolucion de las Bromeliaceae han interactuado factores biologicos y ambientales que han llevado a la seleccion de las caracterfsticas morfologicas, fisiologicas y a la distribucion del conjunto actual de especies que conocemos. Estos factores son: 1) aparente alta demanda de luz, la cual parece estar asociada con ancestros de la familia distribuidos en habitats expuestos aunque hUmedos y probablemente de altas montai'ias tropicales, 2) fuerte tendencia al desarrollo del epifitismo dentro de la familia, representado particularmente en las dos subfamilias mas diferenciadas morfologicamente, las Tillansioideae y las Bromelioideae y 3) el incremento en aridez por la expansion del area de ocupacion hacia zonas mas secas desde ellugar de origen probablemente mas hUmedo, 10 cuaillevo a la seleccion de caracterfsticas xerom6rficas y al desarrollo del metabolismo CAM en los grupos recientes. Estas tres condiciones que han interactuado en la diferenciacion de la familia se han representado en el esquema de la Fig. 9. Cada sub-familia se presenta separadamente debido a la notable diferenciacion morfologica y de habito (TOMLINSON 1969), asf como por la evidencia de la aparicion independiente del metabolismo CAM en cada una de elIas. Es importante destacar que esta representaci6n deja lugar a mucha especulacion sobre una de las tendencias mas recientes en la diferenciacion de la familia, como es la invasion progresiva de ambientes hUmedos y sombreados de bosques tropicales por grupos con caracterfsticas xeroffticas.
En el modelo de la Fig. 9 se supone que las Bromeliaceae primitivas derivaron de ancestros terrestres que habitaban ambientes de alta exposicion solar, pero con adecuada disponibilidad de agua. El metabolismo fotosintetico de los ancestros y las Bromeliaceae primitivas era C3 sin excepci6n. Las Pitcairnioideae evolucionaron en estos ambientes de alta irradiaci6n donde el stress lumfnico y eventual incidencia de sequia podria haber favorecido la

ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 83
seleccion de una anatomfa foliar escleromorfa 0 suculenta, resistente a la desecacion. La gran mayoria de las Pitcaimioideae actuales son terrestres, pero se enuentran formas genuinamente epifitas en los generos Brocchinia y Pitcairnia. Las Pitcaimioideae actuales, particularmente aquellas de la zona del escudo de Guayana (Tabla 6), poseen todas el tipo fotosintetico C3, mientras que aquellos generos de la subfamilia distribuidos en zonas semiaridas del norte de Argentina, Chile y algunas areas del noroeste de Brasil evolucionaron como xerofitas en toda su extension, incluyendo el desarrollo del CAM (Encholirion, Dyckia. Deuterocohnia). La ocupacion de zonas semi-aridas por Pitcaimioideae tambien ocurrio en la region de Centro America donde se encuentra actualmente el genero HeChtia que posee metabolismo CAM. EI genero Pitcairnia es de gran in teres por su elevado mimero de especies (Tabla 6) las cuales se presume son C3 en su totalidad. Este genero esm bien diferenciado ecologicamente y posee especies de sol, xeromorfas, con metabolismo C3. en microhabitats xericos en areas hUmedas (particularmente dentro del subgenero Pepinia. por ej. P. pruinosa. P. bulbosa), y especies mesomorfas, poco resistentes a la sequfa, tambien con metabolismo fotosintetico C3, que crecen en habitats sombreados de bosques tropic ales (frecuentes dentro del subgenero Pitcairnia. por ej. P. juncoides y P. rubiginosa).
EI anal isis cladfstico de la subfamilia Pitcaimiodeae ha permitido separar 3 grupos de generos basados en caracterfsticas morfologicas (VARADARAJAN y GILMARTIN 1988a): 1) el grupo Brocchinia; 2) cl grupo de Ayensua. Connellia. Cottendorfia. Fosterella. Navia Pepinia (segun VARADARAJAN y GILMARTIN 1988b), Pitcairnia y Steyerbromelia; 3) el grupo de Abromeitiella. Brewcaria, Deuterocohnia. Dyckia. Encholirion, Hechtia y Puya. Los dos primeros grupos estan restringidos en su distribucion a las tierras altas de Guayana (SMITH 1988), mientras que el tercero incluye todos los generos que ocupan habitats xeroffticos en todo cl continente. La excepcion en este ultimo grupo la constituye el genero Brewcaria, de las tierras altas de Guayana pero que morfologicamente esm asociado con las formas mas xeroffticas. La importante de esta diferenciacion cladfstica es que incluye en un grupo a todos los generos xeroffticos, en los cuales se ha comprobado la presencia de metabolismo CAM. Serfa de gran interes eco-fisiologico investigar al genero Brewcaria por su localizacion dentro del grupo xerofftico a pesar de su area de distribucion. La separacion del grupo de Brocchinia tiene tambien especial significacion ecologica y evolutiva por ser este cl unico genero de la subfamilia donde se ha demostrado la presencia de tricorn as con capacidad de absorcion de agua (BENZING et al. 1985).
La probable evolucion de las Tillandsioideae se basa esencialmente en la concepcion original de TIETZE (1906) y PITTENDRIGH (1948), la cual ha sido modificada y ampliada recientemente porestudios ecofisiologicos en Venezuela (MEDINA 1974) Y Trinidad (SMITH y GRIFFITHS, 1983; SMITH et al. 1986a). Las Tillandsioideae, siendo casi exclusivamente una subfamilia de especies epifitas, muestran una clara diferenciacion entre especies CAM y C3, muy bien correlacionada con su distribucion en habitats htimedos y secos. Se presume que el epifitismo surgio muy tempranamente en la diferenciacion de esta subfamilia, y las especies mas primitivas han debido ocurrir como epifitas en areas expuestas del dosel de bosques hUmedos tropicales. En esta etapa las caracterfsticas anatomicas han debido jugar un papel

84 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
cntico en la supervivencia de las especies epifitas, por su papel en la protecci6n contra la desecaci6n y por el desarrollo del tricoma peltado altamente especializado, caracteristico de esta subfamilia. En las Tillandsioideae actuales se encuentran numerosos ejemplos con estas caracteristicas. Un grupo actual esta representado por especies como Tillandsia fendlerU y Vriesea platynema, epifitas de bosques nublados que pueden encontrarse en lugares muy expuestos. No presentan una cubierta densa de tricomas peltados, pero poseen una morfologfa de tanque que les permite acumular agua de lluvia. Un segundo grupo 10 constituyen especies mas xeromorfas, que ocupan habitats similares, como T. longifolia y V. incurva, las cuales no forman tanques de acumulaci6n pero estan cubiertas por densas capas de tricomas peltados. Es concebible el desarrollo de formas de Tillandsioideae mas mesomorfas, mas tolerantes a la sombra, a partir del primer grupo, mientras que formas mas xeromorfas, eventualmente con metabolismo CAM, se desarrollarian a partir del segundo grupo. Es interesante destacar que dentro de las Tillandsioideae se ha reportado una sola especie con metabolismo CAM inducible, Guzmania monostachia (MEDINA et al. 1977).
En las Bromelioideae la mayorfa de las especies son CAM, 10 que sugiere que se han originado de ancestros con alta demanda de luz, y que su diferenciaci6n es mas reciente. Esto ultimo concuerda con la presencia de ovario fnfero en esta subfamilia. Las Bromelioideae primitivas han debido ser terrestres y evolucionado en habitats expuestos penetrando en ambientes de aridez cada vez mayor. Se ha supuesto la existencia de especies C3 primitivas en la subfamilia de las cuales hayan podido derivarse las especies C3 actuales que se encuentran dentro del genero Greigia (MEDINA et al. 1989). Este genero presenta una distribuci6n disyunta en los Andes, con un grupo de especies que se encuentra hacia el norte, desde Ecuador hasta Mexico, y un segundo grupo mas pequei'lo que se encuentra en Chile (SMITH y DoWNS 1979). La presencia de este genero en una subfamilia cuya evoluci6n parece haber ocurrido predominantemente en regiones semi-aridas, en las cuales ocurri6 el desarrollo del metabolismo CAM. es uno de los problemas de mayor interes para clarificar la evoluci6n de la familia. En este sentido sena tambien de gran interes el estudio de los generos Ochagavia y Fascicularia cuya distribuci6n se restringe a zonas extratropicales en el cenuo-sur de Chile.
La evoluci6n de las Bromelioideae CAM terrestres parece haber seguido dos vias: la primera representada por la invasi6n de habitats terrestres en el soto de bosques estacionales 0 hUmedos. Muchas especies de Bromelioideae con metabolismo CAM y marcado grado de suculencia foliar crecen bajo sombra parcial de bosques secos (por ej.: Neoglaziovia. Cryptanthus. Bro -melia) 0 hUmedos (por ej.: Canistrum. Nidularium. Ananas). En este grupo se han encontrado algunas especies en los generos Nidularium y Wittrockia con valores de 513C caracteristicos de plantas C3 (MEDINA et al. 1977). Habria que determinar si estas especies poseen un metabolismo CAM inducible por sequia, 0 si por el contrario, se trata de especies exclusivamente C3.
Una segunda via evolutiva estaria representada por el desarrollo del epifitismo, proceso en el cual primero se ocuparon habitats expuestos en el dosel y progresivamente se invadieron habitats epiffticos mas sombreados. En este proceso de adaptaci6n a la sombra las Bromelioideae retienen su capacidad de fijaci6n noctuma de CO2• pero se hacen menos tolerantes a

ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 85
deficits hidricos. Este proceso puede observarse claramente entre las especies del genero Aechmea, en donde encontramos especies de zonas relativamente secas, como terrestres 0 epifitas (por ej. A. aquilega) y especies que crecen incluso como terrestres en el pi so de bosques nublados de la costa norte de Venezuela (A. fendlerU). Asimismo, A. rubiginosa en el norte de la cuenca del Amazonas, y A. magdalenae de Centro America crecen en el soto de bosques hUmedos y sombreados. Esta ultima especie es capaz de mantener tasas similares de fijaci6n noctuma de CO2 en un rango muy amplio de intensidad de luz, y su actividad de crecimiento parece estar mas limitada por la disponibilidad de agua que por la luz (PFITSCH y SMITII 1988). La presencia de actividad CAM en plantas predominantemente de sombra podrfa interpretarse como un relicto de la evoluci6n del genero en habitats expuestos, probablemente mas xericos.
CONSIDERACIONES FINALES
Las discusiones anteriores han mostrado la combinaci6n de caracteristicas morfol6gicas y fisiol6gicas que explican el exito y la diferenciaci6n evolutiva de 1a familia Bromeliaceae en ambientes neotropicales. La proposici6n que surge como mas resaltante de estos estudios es la posibilidad de utilizar propiedades fisiol6gicas y metab61icas como marcadores evolutivos de la diferenciaci6n de los ancestros de la familia en habitats caracterizados por alta irradiaci6n solar. Debe notarse, sin embargo, que muchas de las general izaciones sefialadas en este trabajo se derivan de estudios precisos en un numero de especies muy reducido, 10 cual puede generar errores en la discusi6n de los patrones evolutivos. Hasta hace poco se pensaba, por ejemplo, que el genero Vriesea estaba constituido exclusivamente por especies C3, pero analisis recientes basados en sugerencias de WERNER RAUII (HEIDELBERG) han mostrado que un grupo de especies xer6ffticas, las llamadas "Vrieseas grises" (RAUII 1981) presentan caracteristicas CAM identic as a las observadas en el genero Tillandsia. Esta observaci6n, sin embargo, no cambia el cuadro general que ya presentamos hace varios afios (MEDINA 1974), Y por el contrario, refuerza el concepto de la potencialidad CAM de la familia y el significado de las condiciones de aridez en el desarrollo evolutivo de esta propiedad.
Muchos nuevos problemas han sido planteados que requieren estudios detallados. Entre los mas importantes deben destacarse:
I) "C6mo se modifican los requerimientos energeticos y la productividad de especies con metabolismo CAM en habitats sombreados de bosques hUmedos?
2) "Cual es la significaci6n de los cambios en la anatomia foliar producidos por la adaptaci6n a la sombra para la resistencia a la desecaci6n de bromelias CAM?
3) "C6mo se diferencian las tasas de crecimiento de bromelias de CAM y C3 que coexisten en condiciones de sombra en bosques humedos tropic ales?
4) "C6mo podria demostrarse la capacidad general de la familia Bromeliaceae para el desarrollo del metabolismo CAM?

86 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
REFERENCIAS BIBLIOGRAFICAS
BENZING, D.H. 1976. Bromeliad trichomes: structure, function, and ecological significance. Selbyana 1: 330-348. 1989. The evolution of epiphytism. En: Liittge, U. (ed.) Vascular plants as epiphytes. Ecological Studies, Vol. 76, pp. 15-41. Springer Verlag. Berlin.
BENZING, D.H. Y RENFROW, A. 1971. The significance of photosynthetic efficiency to habitat preference and phylogeny among tillandsioid bromeliads. Botanical Gazelle 132: 19-30. 1971.
BENZING, D.H. GIVNISH, T.I. y BERMUDES D. 1985. Absorptive trichomes in Brocchinia reducta (Bromeliaceae) and their evolutionary and systematic significance. Syst. Bot. 10: 81-91.
BROWN, G.K. Y GILMAKflN, A.M. 1989. Chromosome numbers in Bromeliaceae. Amer. J. Bot. 76: 657-665.
COlITINHO, LM. 1963. Algumas informa~oes sobre a ocorrencia do "Efeito de DeSaussure" em epifitas e herbaceas terrestres da mata pluvial. Bol. Fac. Filos. Cienc. Letr. V.S.P. Ng 288, Botanica 20: 81-98. 1969. Novas observa~oes sobre a ocorrencia do "efeito de DeSaussure" e suas rela~Oes com a suculencia, a temperatura folhear e os movimentos eSlomaticos. Bol. Fac. Filos. Cienc. Letr. V.S.P. Ng 331, Botanica 79-102.
GILMARTIN, A.I. 1983. Evolution of mesic and xeric habits in Tillandsia and Vriesea (Bromeliaceae). Syst. Bot. 8: 233-242.
GILMAKfIN, A.I. y BROWN, G.K. 1986. Oadistic tests of hypothesis concerning evolution of xerophytes and mesophytes within Tillandsia subg. Phytarrhiza (Bromeliaceae). Am. I. BOT. 73: 387-397. 1987. Bromeliales, related monocots, and resolution of relationships among Bromeliaceae subfamilies. Syst. Bot. 12: 493-500.
GRIFFITHS, H. 1988. Crassulacean acid metabolism: a re-appraisal of physiological plasticity in form and function. Adv. Bot. Res. 15: 43-92. 1989. Carbon dioxide concentrating mechanisms and the evolution of CAM in vascular epiphytes. En: Liittge, U. (ed.) Vascular plants as epiphytes. Ecological Studies, Vol. 76, pp. 42-86. Springer Verlag. Berlin.
GRIFFITHS, H. Y SMITH I.A.C. 1983. Photosynthetic pathways in the Bromeliaceae of Trinidad: relations betweeillife-forms, habitat preference and the occurrence of CAM. Oecologia 60: 176-184.
Harms, H. 1930. Bromeliaceen, En: Engler y Prantl. Die natw-liche PJlanzen/amilien. 2a. ed. 15a, pp. 65-159.
LEE, H.S.1., LOTrGE, U., MEDINA, E., SMITH, I.A.C., CRAM, W.I.C., DIAZ, M., GRIFFITHS, H., POPP, M., SCHAFFER, C., STIMMEL, K.H. y THONKE, B. 1989. Ecophysiology of xerophytic and halophytic vegetation of coastal alluvial plain in northern Venezuela. III. Bromelia humilis, a terrestrial CAM bromeliad. New Phytologist111: 253-271.
LOTrGE, U. 1987. Carbon dioxide and water demand: Crassulacean acid metabolism (CAM), a versatile ecological adaptation exemplifying the need for integration in ecophysiological worle New Phytologist 106: 593-629. 1988. Day-night changes in citric-acid levels in crassulacean acid metabolism: phenomenon and ecophysiological significance. Plant, Cell and Environ. 11: 445-457.
LOTrGE, U. y BALL, E. 1987. Dark respiration of CAM plants. Plant Physiol. Biochem. 25: 3-10. MARCHANT, C.l 1967. Chromosome evolution in the Bromeliaceae. Kew Bulletin 21: 161-169. MCWILLIAMS, E.L. 1970. Comparative rates of dark CO2 uptake and acidification in Bromelia-
ceae, Orchidaceae and Euphorbiaceae. Bot. Gaz. 131: 285-290. MEDINA, E., MONTES, G., CUEVAS, E., ROKZANDIC, Z. 1986a. Profiles of CO2 concentration and
/i!3C values in tropical rain forests of the Upper Rio Negro basin, Venezuela. Journal o/Tropical Ecology 2: 207-217.
MEDINA, E., OLIVARES, E., DIAz, M., VAN DER MERWE, N. 1989. Metabolismo acido de crasulaceas en bosques humedos tropicales. Monographs in Systematic Botany (Missouri Botanical Garden) 27: 56-67.
MEDINA, E., OLIVARES, E., DIAZ, M. 1986b. Water stress and light intensity effecls on growth and nocturnal acid accumulation in a terrestrial CAM bromeliad (Bromelia humilis Iacq.) under natural conditions. Oecologia (Berl) 70: 441-446.

ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 87
MEDINA, E. 1984. On the temperature dependence of dark CO2 fixation in CAM plants: acidity, growth and d13C values. En: Physiological Ecology of CAM Plants (MEDINA E. ed.) pp. 52-72. Ediciones CIET-IVIC. Caracas. 1987. Aspectos ecofisiol6gicos de plantas CAM en los tr6picos. Revista de Biologia Tropical 35 (Supl. 1): 55-70. 1974. Dark CO2 fixation, habitat preference and evolution within the Bromeliaceae. Evolwion 28: 677-686.
MEDINA, E., DELGADO, M., TROUGHTON J.H. AND MEDINA, J.D. 1977. Physiological ecology of CO2 fixation in Bromeliaceae. Flora 166: 137-152.
MEZ, C. 1904. Physiologische Bromeliaceen-Studien. 1. Die Wasserokonomie der extrem atmosphirischen Tillandsien. Jahrbuch Wiss. Botanik 40: 157-229.
OSMOND, C.B. 1978. Crassulacean acid metabolism. A curiosity in context. Ann. Rev. Plant Physiology 29: 379-414.
PFrrsOl, W.A. Y SMrm, A.P. 1988. Growth and photosynthesis of Aechmea magdalenae, a terrestrial CAM plant in a tropical moist forest, Panama. Journal of Tropical Ecology 4: 199-207.
PrrrnNDRIGH C.S. 1948. The bromeliad-Anopheles-malaria complex in Trinidad. 1. The bromeliad flora. Evolwion 2: 58-89.
RAUH, W. 1981. Bromelien. 2a. Edici6n. Eugen Ulmer Verlag. Stuttgart. SAITO, N. y HARBORNE, J.B. 1983. A cyanidin glycoside giving scarlet coloration in plants of the
Bromeliaceae. Phytochemistry 22: 1735-1740. SCHIMPER, A.F.W. 1888. Die epiphytische Vegetation Amerikas. Botanische Mitteilungen aus
den Tropen. ll. Jena, Fischer Verlag. SMIlH, J.A.C. 1989. Epiphytic bromeliads. En: Liittge, U. (ed.) Vascular plants as epiphytes.
&ological Studies, Vol. 76, pp. 109-138. Springer Verlag. Berlin. SMIlH, J.A.C., GRIFFI1HS, H., BASSET, M. Y GRIFFI1HS, N.M. 1985. Day-night changes in the leaf
water relations of epiphytic bromeliads in the rain forests of Trinidad. OECOLOGIA 67: 475-485.
SMITH, J.A.C., GRIFFITHS, H., LOTrGE, U., CROOK, C.E., GRIFFITHS, N.M. Y STIMMEL, K.H. 1986b. Comparative eco-physiology of CAM and C3 bromeliads. IV. Plant water relations. Plant, Cell and Environment 9: 395-410.
SMIlH, J.A.C., GRIFFITHS, H., LOTrGE, U. 1986 a. Comparative eco-physiology of CAM and C3 bromeliads. I. The ecology of Bromeliaceae in Trinidad. Plant, Cell and Environment 9: 359-376.
SMIlH, L.B. 1934. Geographical evidence on the lines of evolution in the Bromeliaceae. Bot. Jahrb. 66: 446-468. 1988. New key to the genera of the Bromeliaceae. Beitr. Bioi. Pflanzen 63: 403-411.
SMIlH L.B. Y DOWNS, R.J. 1974. Flora Neotropica, Monograph No. 14. Part 1. Pitcaimioideae (Bromeliaceae). Hafner Press. New York. 1977. Flora Neotropica, Monograph No. 14. Part 2. Tillandsioideae (Bromeliaceae). Hafner Press. New York. 1979. Flora Neotropica, Monograph No. 14. Part 3. Bromelioideae (Bromeliaceae). Hafner Press. New York. .
SZAREK, S.R., JOHNSON, H.B. Y TING, 1.P. 1973. Drought adaptation in Opuntia basilaris. Significance of recycling carbon through crassulacean acid metabolism. Plant Physicl. 52: 539-541.
TIETZE, M. 1906. Physiologische Bromeliaceen-Studien. II. Die Entwicklung der wasseraufnehmenden Bromeliaceen-Trichome. Z. Naturwiss. 78: 1-49.
TOMLINSON, P.B. 1969. Commelinales-Zingiberales. En: Anatomy of Monocotyledons Vol. III (Metcalf, C.R., ed.). Clarendon Press. Oxford.
VARADAJARAN, G.S. Y GILMAKfIN, A.J. 1988a. Phylogenetic relationships of groups of genera within the subfamily Pitcaimioideae (Bromeliaceae). Syst. Bot. 13: 283-293. 1988b. Taxonomic realignments within the subfamily Pitcairnioideae (Bromeliaceae). Syst. Bot. 13: 294-299.
Wn.LIAMS, C.A. 1978. The systematic implications of the complexity of leaf flavonoids in the Bromeliaceae. Phytochemistry 17: 729-734.
WINTER, K. 1985. Crassulacean Acid Metabolism. In: Photosynthetic Mechanisms and the Environment. Ed. Barber, J. and Baker, N.R. Elsevier Science Publ. B.V. (Biomedical Division). The Hague. pp. 329-387.
WOLF, J. 1960. Der diumale Siurerbythmus. In: Handbuch der Pflanzenphysiologie. Ed. Ruhland, H. Xll/2. Springer Verlag, Berlin. pp. 809-889.

88 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
TABLA 1. - Diferenciaci6n taxon6mica de las Bromeliaceae (segun SMITH y DOWNS, 1974-1979)
Habito
Rakes
Indumento foliar
Ovario
Fruto
Semillas
Diferenciaci6n taxon6mica
Pitcairnioideae
Terrestres 0
saxlcolas
Bien desarrolladas y funcionales
Escamas finamente divididas
Supero (excepto algunas spp. de Pitcairnia y Navia
Capsular
Apendice entero
13 generos 731 especies
Bromelioideae Tillandsioideae
Terrestres 0 Epifitas epifitas
Por 10 general Solo de soporte solo de soporte a veces reducidas
Escamas peltadas Escamas peltadas con celulas dispuestas de simetria radial irregularrnente
Infero Supero (excepto Glomeropitcairnia )
Baccario, 0 seco Capsular
Desnudas Apendices plumosos
27 generos 6 generos 556 especies 800 especies
TABLA 2.- Acumulaci6n de malato y cambios de acidez titulable de extractos acuosos de Bromeliaceae con CAM. Valores en ~mol/g peso
fresco (MEDINA 1974)
Dickya tuberosa Tillandsia utriculala Aechmea bromelifolia am pm am pm am pm
Acidez titulable 218.7 20.8 206.2 25.0 300.0 25.0
t. 198 181 275
Malato 255.8 121.0 194.1 8.8 271.3 24.5
t. 135 185 247
% acidez como malato 68 100 90

ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 89
TABLA 3.- Relaciones hfdricas de bromelias epifitas en Trinidad (SMITH et ai. 1985)
Especies JI(MPa) L1JI Tension Xilema L1 &/+ y localidad Min Max D{a Noche mollm3
Bosque de bajura (6Om) Especies CAM Tilllandsia utricu/ala 0.490 0.860 0.35 0.21 0.36 0.11 207 Aechmea nudicaulis 0.450 0.680 0.23 0.19 0.47 0.28 190
Especies C3 Guzmanin monoslachin 0.505 0.37 0.19 -D.18 n.s. Guzmanin lingulala 0.405 0.43 0.20 -D.23 n.s.
GaTO del bosque (250m) Especies CAM Tillandsin utriculata 0.495 1.160 0.665 0.26 0.48 0.22 365 Aechmea nudicaulis 0.510 1.175 0.665 0.21 0.55 0.34 303
Especies C3 Guzmanin monoslachin 0.460 0.615 0.155 0.51 0.25 -0.26 88
TABLA 4.- Presion osmotica del juga celular y tension del xilema al amanecer y al atardecer para las epocas de lluvia y sequia en poblaciones
de Bromeiia humiiis que crecen a plena exposicion solar (n=3) (modificado de LEE et al. 1989).
presi6n osm6tica (MPa) l!. (manana-tarde)
tensi6n xilematica (MPa) l!. (manana-tarde)
Epoca de lluvias Epoca de sequ{a Manana Tarde Manana Tarde
0.77 (0.09) 0.67 (0.11) 1.17 (0.02) 0.86 (0.06) 0.10 0.31
0.59 (0.07) 0.22 (0.06) 0.59 (0.07) 0.47 (0.12) 0.37 0.12

90 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
TABLA 5.- Variaciones estacionales de la composicion foliar y la actividad CAM en Bromelia humilis (MEDINA et al. 1986).
Eslacion Area Foliar Esp. Agua Clorofila Nilrogeno & Acidez & Hexasa eq. m21kg glm2 mglm2 glm2 meqlm2 mmollm2
Eslacion Seca plantas de sol 4.64 588 59 0.95 4 -25 plantas de sombra 4.87 947 148 2.40 18 -4
Es/acion de Lluvias plantas de sol 6.08 1014 44 0.83 191 -79 plantas de sombra 4.95 1057 227 2.28 219 -99
Final de la Eslacion de Lluvws
plantas de sol 5.33 833 154 1.44 103 -45 plantas de sombra 4.26 1057 599 3.87 196 -64
TABLA 6.- Distribucion por genero del metabolismo CAM dentro de las Bromeliaceae. Entre parentesis el mlmero de especies
descriptas segun SMITII y DOWNS (1974-79). Debajo de generos muy diferenciados se indican los
subgeneros correspondientes.
Pltcalrnloldeae: 173 especies CAM potenciales (24 % del total). No se incluyen los generos Brewcarw, Sleyerbromelw y Lindmannw propuestos por Smith (1988).
Generos presumiblemente solo C3. Plqa
Plqopsis (162) Plqa (6)
Pilcairnw PeJ1lnia (40) Pilcairnw (220)
Connellw (4) COllendoifUl (24) Brocchinia (18) Navia (74)
TIerras altas de Guayana y cordillera Andina Andes del Sur
Principalmente en la alta y baja Guayana Andes tropicales. Guayana, C.A. y sureste de Brasil TIerras altas de Guayana Tierras altas de Guayana TIerras altas de Guayana Tierras altas y bajas de Guayana
Generos con especies CAM comprobadas Deu/erocohnw (7) Noreste de Argentina y Bolivia Dyckw (103) Sureste de Brasil, norte de Argntina Encholirion (12) Noreste de Brasil Hechlw (45) America Central
Generos no estudiados Ayensua (1) Abromeiliella (2) Foslerella (13)
TIerras altas de Guayana (probablemente C3) Norte de Argentina (probablemente CAM) America Central, Andes del Sur

ECO-HSIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 91
TABLA 6 .- (Continuaci6n)
TiIIandsioideae: 184 especies CAM potenciales (23 % del total)
Generos presumiblemente solo C3. Vriesea
X iphion (241) Alcantarea (8)
Guzmania (126) Catopsis (19) Glomeropitcairnia (2)
America Central, norte de S.A., sureste de Brasil sureste de Brasil, Jamaica America Central, norte de S.A. y Andes tropicales America Central, sureste de Brasil, Andes, alta Guayana norte de Suramerica, Antillas
Generos con especies CAM comprobadas Tillandsta ampliamente distribuido
Allardtia (147) predominantemente especies C3 Anoplophy/um (34) predominantemente especies CAM Phy/arrhyza (35) especies mesofiticas C3, xerofiticas CAM Diaphoran/hema (17) presumiblemente todas CAM Tilllandsia (117) presumiblemente todas CAM Pseudoalcanlarea (4) no estudiadas Pseudocalopsis (48) presumiblemente todas C3
Generos no estudiados Mezobromelia (2) Andes de Colombia y Ecuador (probablemente C3)
Bromelioideae: 465 especies CAM potenciales (84 % del total). No inciuye los generos Deinacanlhon y Lymannia propuestos por Smith (1988).
Generos presumiblemente solo C3. Greigia (26) America Central, tropicales y sur de Chile.
Generos con especies CAM comprobadas. Araeococcus (5)
Araeococcus Pseudoaraeococcus
Slreplocalyx (14) Neoregelia (71)
Hylaeaicum Neoregelia
Cryplan/hus (20) Nidularium (23)
Canislropsis Nidularium
Bromelia (47) Acanloslachys (1) Or/ophy/um (17) Canis/rum (7) Will roc kia (7) Hohenbergia (40)
Hohenbergia Willmackiopsis
Aechmea (172) Podaechmea
Guayana, cuenca Amawnas Cuenca Amawnas, Brasil oriental Cuenca Amawnas, Guayana, Brasil oriental
Cuenca Amawnas Sureste de Brasil Sur- y nor-este de Brasil
Sureste de Brasil Sureste de Brasil Centroamerica, norte de S.A., Amawnas, y sur de Brasil Sureste de Brasil Este de Brasil Este y sureste de Brasil Sureste de Bras il
Norte de Suramerica, este de Brasil Antillas mayores
America Central y Peru

92 BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
Lamprococcus Aechmea Ortgiesia Plalyaechm£a Pothuaya Macrochordion Chevaliera
Quesnelia (14) Quesnelia Billbergiopsis
Billbergia (53) Billbergia Helicoidea
Neoglaziovia (2) Portea (7) Ananas (8)
Generos no estudiados Fernseea (1) Ronnbergia (8) Androlepis (1) Ochagavia (3) Andrea (1) Hohenbergiopsis (1) Fascicularia (5) Disteganthus (2) Pseudananas (1)
TABLA 6.- (Continuaci6n)
Sur de Venezuela y Colombia, este de Brasil Centroamerica, Antillas, Suramerica Sureste de Brasil, Paraguay, Argentina Centroamerica, cuenca Amazonas, Bolivia, sureste de Brasil Centroamerica, tropical Andes, sureste de Brasil Centroarnerica, cuenca Amazonas, Central y este de Brasil Centroamerica, cuenca Amazonas, este de Brasil
Sureste de Brasil Sureste de Brasil
Guatemala, este y sureste de Brasil Mexico, Guayana, cuenca Amazonas, Brasil central y oriental Noreste de Brasil Este de Brasil Guayana, cuenca Amazonas, Brasil central y oriental
Sureste de Brasil Centroamerica, occidente de Colombia Centroamerica y Cuba Sur de Chile Sureste de Brasil Guatemala Sur de Chile Guayana Francesa, norte de Brasil Brasil central y oriental, Paraguay

ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 93
FIG. 1 - Habitats de Bromeliaceae en Veneruela. A. Epifitas en bosques pluviales estacionales. Tillandsia elongata, Caucagua. B. Terrestre, bosque nublado Isla de Margarita, 800 m s.n.m. Glomeropiteairnia ereetiflora . C. Terrestre en bosques humedos montanos. Piedra de la Virgen, Gran Sabana. Broeehinia mierantha . D. Saxlcola sobre afloramientos granfticos. Piteairnia pruinosa, P. bulbosa y P. armata en Puerto Ayacucho. E. Terrestre y epifita en zonas semi-aridas del norte de Venezuela. Bromelia humilis (terrestre) y Tillandsia reeurvata (epifita atmosferica). F. Atmosfericas extremas. Tillandsia
flexuosa sobre cables en Hueque, Edo. Falcon.

FIG. 2 _ Fotograffas de superficie foliares con tricorn as peltados tornadas con rnicroscopio electronico de barrido. El aurnento es el rnisrno en los tres casos, de rnanera que pueden cornpararse la distribucion y el tarnano. A. Puya arisleguielae; B. Tillandsia elongala; C. Aechmea aquilega.
CI:l o § z tj tTl r J> J> (") J> tj tTl s: :; Z J> (")
(3 Z J> r tj tTl (")
m z (")
:; (/l
(") o ;;0 tj o w J>
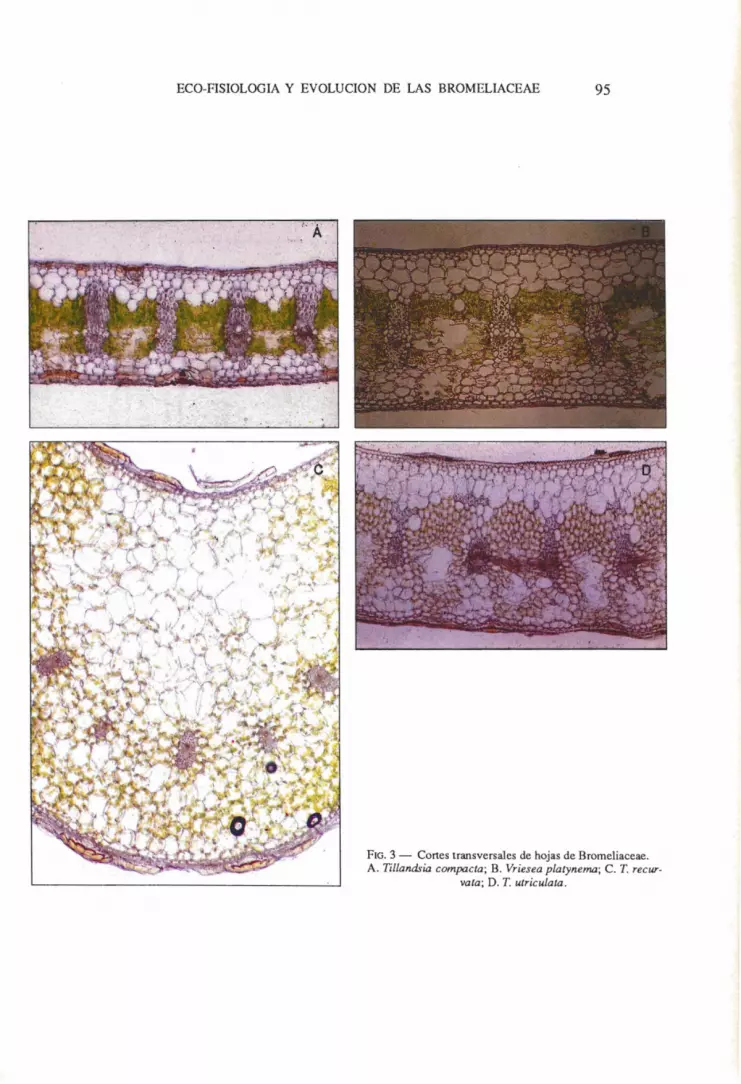
ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 95
FIG. 3 - Cortes transversales de hojas de Bromeliaceae. A. Tillandsia compacta; B. Vriesea platynema; C. T. recur·
vata; D. T. utriculata.
Noche 010
M a I a t 0 + 2 H +-----------...-----1----------...
HC03
CO2 (respirocion)
1 AOA
PEP corboxiloso
PEP
Glicolisis
CO2 RuBP- I corboxiloso
.","" piruvoto .","" I Lr-R ..1 •
esplroclOn PEP
Gluco-1
neogenesIs
Carbohidrato
FIG. 4 - Esquema simplicado de las reacciones bioquimicas asociadas al metabolismo CAM (modificado de WINTER 1985). AOA: oxaloacetato; PEP: fosfo-enol-piruvato; RFC: reducci6n fotosintetica de CO2. La flecha gruesa en el dia indica la incorporaci6n de CO2 exterior cuando se ha descarboxilado todo el malato acumulado
durante la noche anterior. La flecha punteada indica la posible oxidaci6n del piruvato resultante de la descarboxilaci6n del malato.

A echmea bromel! folia
TiJiandsio utnculata am
Dickyo fubero50 am
am
pm pm
pm
180 200 220 200 220 24() 260 280
Temperature DC Temperature °C Temperature °C
FIG. 5 - Composicion de acidos organicos obtenida por cromatografia de gases en extractos de acidos organicos derivados con trimetil-silil (Medina et at. 1977). Los numeros 3 y 5 corresponden a los acidos malico y dtrico respectivamente.
98
......... Q) en 0
lL..
~ c: 0 .... ~ ~
0 ~ u 0
Q)
~
c: '0
u 0
~
E ~ u
c::r N
-----
BOLETIN DE LA ACADEMIA NACIONAL DE CIENCIAS. CORDOBA
300~------------------------------~
200
100
•
• • •• •
• ... ~., ...
• • .. ,. .,
relacion I: I
•
O~----~----~----~------~----~--~
o 100 200 300
CO 2 fijado durante Fase I (mmoI/Kg) FIG. 6 - Reciclaje aparente de CO2 en especies de Bromeliaceae. Con datos tabulados por GRIFAl1IS
1988. Los puntos por encirna de la recta de la relaci6n 1: 1 corresponden a los casos donde se registra mayor acidificaci6n noctuma que la que puede ser explicada por incorporaci6n de CO2 exterior.
U ro -(,()
- 35
- 30
- 25
-20
-15
-10
- 5
o o
o~ Especies C 3 ~($)
(fX)~~
Especies CAM
t::. Bromelioideae
o T i Ilandsioideae .. Pitcairnioldeae
O+--,,--,--.--,---,--.--,---,--.--,,--,--.--,~
o 10 20 30 40 50 60
Orden Secuencial de Especies FIG. 7 - D1StnbuCl6n de valores de a13c en subfamilias de Bromehaceae. Con datos recopllados por
MEDINA et a/. 1977, 1989, GRIFFITHS y SMITH 1983.
ECO-FISIOLOGIA Y EVOLUCION DE LAS BROMELIACEAE 99
u ,.., -c..o
- 36 -.~ --P-U-Y~(Pitcairnioideae) -32
-28
- 24
-20
-16
-12
- 8 0
-36
-32-
-28
• • • •
4
• • • • • • •
8
• (Tiliandsioideae)
12
•• Oc9 Vriesea 0
fII' Qr5
U -24 ,.., ~ o~andsia {--------------------------------
c..o -20 cf9
:::~
U ,.., -c..o
- 8 -J-,...,rrr-rrr-rrrrrrT'T'TTTTTT''T'""TTlrrnrrrrTTrTT''j o 4 8 12 16 20 24 28 32 36 40
::: 1 Aechmea (Bromelioideae)
-28.:1
-24
- 20
-16
-12
- 8 0 4 8 12 16 20
Orden Secuencial de Especies
FIG. 8 - Distribuci6n de valores de d13C en generos escogidos de cada subfamilia de Bromeliaceae. Los valores se ordenan de acuerdo a su magnitud para apreciar la variaci6n gradual entre los extremos.

<l ~
<l CD
N W 0
a::: c:r
<l I--.J <l
GRUPO ANCESTRAL terrestre, alto irradiacion y humedad
I NTENSI DAD DE LU Z _----""'"'=::::::::::::=------__ BAJA ALTA
lr [;3) - ~ .. -~- 11 (C3) =[1... I I
-' \ - 1rU ~ lLl terrestres c::::::::J
epifitas IT [;~~ oo [l
PC3-L
PCAM -L B CAM-L
TILLANDSIOIDEAE PITCA I RN 10 I D EAE BROMELIOIDEAE
FIG. 9 - Esquema hipotetico de la evoluci6n de las Bromeliaceae. Las Ietras p. T y B corresponden a las subfamilias Pitcaimioideae. Tillandsioideae y Bromelioideae. respectivamente. Las designaciones C3 y CAM despues de la primera Ielra indica el tipo fotosinletico. mientras que las letras L y S al final indican habitats expuestos (alta intensidad de luz) y sombra. respectivamente. Las letras I1enas corresponden a forrnas terrestres y las vadas a forrnas epifitas. EI espacio evolutivo de cada subfamilia se divide en 4 cuadrantes. de acuerdo a condiciones de alta y baja inlensidad de luz y alto y bajo indice de aridez. La combinaci6n de baja intensidad de Juz ymar-
cada aridez no existe en condiciones naturales.
..... 8
I:l:l o ~ -l Z o tTl r :> :> n :> o tTl :: :; z :> n o z :> r o tTl n m z n :; C/)
n o ;N o o o:l :>





























