Embed Size (px)
Citation preview

POPULATION ECOLOGY
Distribution Patterns of Imported Fire Ants(Hymenoptera: Formicidae) on a Sheep and Goat Farm in Oklahoma
JAMES T. VOGT,1 MARGARET L. ALLEN,1 BRADLEY WALLET,2 DEBBIE BOYKIN,3 AND
WAYNE A. SMITH4
Environ. Entomol. 38(3): 551Ð560 (2009)
ABSTRACT Imported Þre ant colonies were quantiÞed in 1,000-m2 circular subplots spaced �125m apart on a sheep and goat farm in Oklahoma. Social form (percent polygyny), mound density,cumulative above-ground mound volume, and average mound volume were subjected to multipleregression analyses to examine trends related to landscape metrics and habitat characteristics. Mo-nogyne populations were spatially autocorrelated, and polygyne mounds tended to be smaller andmore numerous. A model incorporating the effects of percent polygyny, canopy cover, and 1-dcumulative incident solar radiation explained 34% of the variation in mound density. Percent polygynywas not signiÞcantly related to cumulative mound volume, which provides a better estimate of overallant biomass. A model incorporating the effects of 1-d cumulative incident solar radiation on thesummer solstice, elevation, canopy cover, distance from cisterns, distance from water, and distancefrom trees explained 42% of the variation in cumulative mound volume. Monogyne mounds in areasthat were ßat and close to water in low-lying areas were largest. Results indicate that remotely senseddata in combination with publicly available U.S. Geological Survey data may be useful in predictingareas of high and low Þre ant abundance at a Þeld scale.
KEY WORDS Solenopsis invicta, remote sensing, landscape ecology
The large, earthen mounds of the red imported Þre ant(Solenopsis invicta Buren) are a common sight in pas-tures throughout the southeastern United States. Al-though Þre ants can attain impressive population den-sities under the right circumstances (upward of 600 ormore mounds/ha; Vogt et al. 2003), mound density canvary greatly between and within pastures and otherlandscapes. Predictive models of the invasive potentialof S. invicta on a continental or global scale have beenthe subjectof several studies (Stokeret al. 1994,Killianand Grant 1995, Korzukhin et al. 2001, Morrison et al.2004, Sutherst and Maywald 2005, Fitzpatrick et al.2007, Peterson and Nakazawa 2008). However, fewauthors have attempted to describe Þre ant popula-tions at a Þeld scale as a function of landscape metricsor habitat characteristics.
Allen et al. (2006) constructed two predictive mod-els of Þre ant distribution for the state of South Caro-lina based on habitat characteristics. Landscape met-
rics were not important predictors of Þre antabundance, but several habitat characteristics exerteda signiÞcant effect on Þre ant population densities inlogistic models. Fire ants were most likely to be en-countered in freshwater marsh, longleaf pine, andcleared land habitats (probability � 1.00, 0.82, and0.60, respectively) and least likely to be encounteredin mesic mixed, maritime, and saltwater marsh habitats(probability � 0.05. 0, and 0, respectively). A state-wide effort in Louisiana to determine factors inßu-encing population density and social form of Þre antsshowed differences in mound size caused by habitattype as well as soil texture and chemistry (Milks et al.2007). Multivariate models only explained �25% ofthe variation in ant and nest density; however, thisstudy was the Þrst to examine potential effects of soilchemistry on Þre ant abundance and social form. Fireant density and mound size were positively correlatedwith soil characteristics that favor plant growth (phos-phorus and organic matter). As the authors men-tioned, these associations may be caused by positiveeffects on prey abundance and resulting increases inÞre ant abundance or may actually be cumulativephysico-chemical effects of Þre ant mound buildingactivity within areas over time. Porter et al. (1991)found polygyne colonies at 54% of sites surveyed inTexas but found no signiÞcant relationships betweenpolygyny and habitat or environmental conditions. Ata Þner scale, Stiles and Jones (1998) examined Þre antmound density in linear habitats (roads and powerline
Mention of trade names in this publication is for information onlyand does not constitute endorsement by the U.S. Department ofAgriculture or Automated Decisions.
1 Corresponding author: USDAÐARS Biological Control of PestsResearch Unit, PO Box 67, Stoneville, MS 38776 (e-mail:[email protected]).
2 Automated Decisions, 801 Woodcroft Pkwy. 1823, Durham, NC27713.
3 USDAÐARS Mid South Area, 141 Experiment Station Rd., JWD-SRC, Stoneville, MS 38776.
4 Oklahoma Cooperative Extension Service, 1901 S. 9th St., Durant,OK 74702.

rights-of-way) in South Carolina. They found signif-icant edge effects, with colonies generally being moreabundant along road edges or forest edges. Mounds onnorthern edges of linear habitats tended to be larger.They hypothesized that the differences observedmight be caused by physical soil characteristics, dif-ferences in frequency of disturbance (less mowingalong edges), and differential shading. Menzel andNebeker (2008) examined hybrid imported Þre antdistribution in Mississippi forest habitats. Locationswhere hybrid imported Þre ants existed generally hadless canopy cover, more woody and herbaceous plantground cover, and greater proportion of mature pine(measured as percent of diameter at breast height[dbh] of all trees �5 cm dbh in an area) than locationswhere they were absent. These differences werefound to be caused by local conditions rather thancompetitive interactions with other ant species.Tschinkel (1988) conducted a detailed study of thepresence and abundance of S. invicta and a native Þreant, S. geminata, across habitat types with varyinglevels of disturbance in Florida. This study presenteda compelling case for soil disturbance as an importantfactor in establishment and dominance of the invasiveS. invicta. In another study, a relatively undisturbedBlack Belt Prairie remnant in Oktibbeha County, MS,was nearly devoid of Þre ants despite containing muchapparently suitable, open habitat, whereas nearby pas-ture areas contained hundreds of Þre ant colonies perhectare (J.T.V., unpublished data).
Other authors have examined spatial distributionpatterns of individual colonies on a small scale. Forexample, Adams and Tschinkel (1995) examined com-petitive interactions of young Þre ant colonies bymanipulating and measuring their spatial distributionand measured dispersion of colonies in a mature pop-ulation. Naturally occurring incipient colonies weresigniÞcantly clumped, whereas mature populationswere distributed regularly. Brood raiding in youngcolonies resulted in spatially random mortality ratherthan expected distance-dependent mortality; severalpossible explanations were given for this result.Baroni-Urbani and Kannowski (1974) examined spa-tial distribution of colonies in a 20,800-m2 pasture inLouisiana and determined that the population wasaggregated. Tschinkel and Howard (1983) observedthat new nests of founding S. invicta queens werestrongly associated with slightly (a few inches) higherterrain in a study site, where rain runoff would notinundate the new nests; they were unable, however, todetermine the relative importance of topography andsoil characteristics in determining nest site prefer-ence.
Additional studies have focused on one or a fewhabitat variables and their impact on Þre ant colonies.Structures may play a role in location preference orsurvivability of Þre ant colonies. Thorvilson et al.(1992) noted increased overwintering survival of col-onies located next to a curb and a wall with southernexposure compared with colonies located in an openarea near Lubbock, TX. They also detected signiÞcantdifferences in nest temperature for different nest lo-
cations, indicating that building mounds next to struc-tures confers some thermal advantage to the colony.Canopy and resultant degree of shading can inßuenceÞre ant population densities, presumably because ofdecreased solar insolation. Fleetwood et al. (1984)documented increased mortality of lone star tick (Am-blyomma americanum) caused by Þre ant predation inopen areas compared with thicket (canopied) areas inTexas. In southeastern Louisiana longleaf pine savan-nas, Þre ants can dominate open, grassy areas (Colbyand Prowell 2006).
Soil moisture may play a critical role in the abun-dance and/or size of Þre ant colonies. In an early studyin Brazil, Allen et al. (1974) found large colonies of S.invicta to be restricted to moist or protected areas;however, Williams and Whitcomb (as cited in Allen etal. 1974) noted mounds in open areas in Brazil inFebruary, during the rainy season. Fire ant moundsexhibit seasonal change, becoming lower and less con-spicuous during dry periods (Tschinkel 1993), so aperceived lack of colonies in dry areas may be caused,at least in part, by reduced visibility and detectability.
In pastures, discrete extrinsic and intrinsic featuressuch as roads, buildings, and ponds may inßuence Þreant abundance. Variation in habitat elements such asvegetation density, slope and aspect (and resultantinsolation), elevation, soil moisture, and soil distur-bance may also inßuence Þre ant populations on a Þeldscale. In an attempt to gain a better understanding ofÞre ant abundance and distribution in animal produc-tion agroecosystems, we measured Þre ant density andmound size on a sheep and goat farm in Oklahomaalong with landscape metrics and habitat character-istics. SpeciÞcally, we wanted to determine whetherreadily available and/or easily extracted data acquiredthrough multispectral remote sensing and U.S. Geo-logical Survey sources might be useful for modelingÞre ant abundance at a relatively coarse spatial scale(120-m2 sampling grid) within pasture. We were es-pecially interested in examining effects of slope, as-pect, and resultant insolation, because these have re-ceived little attention in previous studies, with theexception of Porter et al. (1991), and the data arereadily available. A better understanding of variationin Þre ant abundance at this scale may yield informa-tion useful for imputing the presence and abundanceof Þre ant mounds based on a subset of detectablemounds in remotely sensed data and ultimately forusing precision application of chemical or biologicalcontrol measures. Other uses may include low-costidentiÞcation of high Þre ant density areas for researchor biological control efforts before Þeld data collec-tion or identiÞcation of suitable sentinel sites for reg-ulatory surveys.
Materials and Methods
Sampling Site. The study site was a sheep and goatfarm located in Johnston County, OK (�96�34�20� W,34�11�38� N; Fig. 1). The area sampled was �1.67 km2
and consisted primarily of gently rolling grazed pas-ture, ranging from heavily managed paddocks (pri-
552 ENVIRONMENTAL ENTOMOLOGY Vol. 38, no. 3

marily Bermuda grass, Cynodon sp., with some fescue,Festuca sp.) to unmanaged areas dominated at the timeof sampling by ragweed (Ambrosia sp.). Large areas ofhardwood forest were present in the northern half ofthe site, and many of the pasture areas also containedscattered hardwoods and cedars. Several buildings,cisterns, and ponds were also present.Data Acquisition. Fire ant mounds were quantiÞed
and measured within 1,000-m2 circular plots (n� 108)centered on grid points spaced �120 m apart. Each
mound was measured (long axis, short axis, height)and assessed for activity by probing with a small stickand observing workers rushing out to defend the col-ony. Above-ground mound volume was estimated us-ing a formula for the volume of an oblique cone, witha correction factor (Vogt 2007). Social status of col-onies across the Þeld was determined by collecting asample of 20Ð30 worker ants from each of three ran-domly selected mounds in each plot where moundswere present. Samples were placed in 70% ethanol and
Fig. 1. Aerial digital true color composite of study site in southern Oklahoma. White plots contained no imported Þreant mounds, green plots contained polygyne colonies, orange plots contained both monogyne and polygyne colonies, andred plots contained monogyne colonies.
June 2009 VOGT ET AL.: HABITAT AND FIRE ANT DENSITY 553

transported to Stoneville, MS, to determine social sta-tus using the method described by Chen and Allen(2006), yielding an estimate of percent polygyne andpercent monogyne mounds.
Aerialmultispectral imagery(red, green,blue,near-infrared) of the site was acquired in May 2006 throughGeoData Airborne Mapping and Measurement, Weir,MS. Visual resolution of the imagery was just under0.16 m. The camera system is described in detail inVogt (2004). Because ground truth data were notobtained at the time of the ßight, data were calibratedto yield “reßectance-like” data using the log residualstool in ENVI (ITT Industries, Boulder, CO) (Greenand Craig 1985). The resulting data were used tocalculate the ratio vegetation index (RVI; near-infra-red/red) (Jordan 1969). Landscape metrics derivedfrom the imagery for each plot included Euclideandistance from cisterns, ponds, trees, and buildings.Trees were classiÞed using Feature Analyst for ArcGIS(Visual Learning Systems, Missoula, MT), and percentcanopy cover was calculated for each plot.
Digital elevation models and additional orthoimag-ery of surrounding areas were obtained from the U.S.Geological Survey, EROS Data Center, Sioux Falls, SD(http://seamless.usgs.gov). Slope, aspect, and 1-d cu-mulative incident solar radiation (w � h/m2) for eachsampling plot were extracted from digital elevationmodels using 3D Analyst and Spatial Analyst tools inArcMap (ESRI, Redlands, CA). Cumulative insolationwas calculated for the Þrst day of sampling (samplingtook 3 consecutive d to complete). To test for poten-tial effects of thermal history on Þre ant distribution,additional calculations were performed to determinecumulative insolation on the winter solstice (21 De-cember) when sun angle is lowest, and on the summersolstice (21 June), when sun angle is highest.
Data Analysis. An r2 selection method was appliedto the data to determine the best potential predictivevariables for polygyny (expressed as percent polygynemounds detected), log10-transformed Þre ant abun-dance data (mound density and cumulative moundvolume), and log10-transformed average mound vol-ume. Stepwise regression was used to eliminate non-signiÞcant variables. Analysis of commonality (Emigh1984) was used to determine the relative and com-binedcontributionsof signiÞcantvariables in themod-els. Regression and commonality analyses were per-formed in SAS (Ver. 8.02; SAS Institute, Cary, NC).Additional visualization techniques (semivariograms,scatter plots, contour plots) were used to examinetrends in the data. Graphics were produced usingSigmaPlot 8.0 (Systat Software, San Jose, CA).
Results
Social Form. The majority of plots across the studysite were dominated by polygyne mounds; some plotshad mixed populations, with only a few dominated bymonogyne mounds. Plots with monogyne moundswere strongly aggregated (Fig. 1). In a model assum-ing spatial independence, polygyny was signiÞcantlyrelated to elevation and RVI (F� 11.65, df � 2,92; P�0.0001, r2 � 0.2021). Residuals for the model werespatially autocorrelated, however, and examination of2 residual log likelihood indicated that a model in-corporating an exponential spatial error term wasmore appropriate; in this model, elevation and RVIwere not signiÞcant, but polygyny was related tomound density and cumulative mound volume (F �5.49; df � 2,92; P � 0.0056; Table 1). Plots that weredominated by polygyne colonies contained large num-bersofmounds,withcorrespondinglyhighcumulative
Table 1. Regression coefficients for landscape metrics and habitat variables influencing polygyne red imported fire ant populationdensity in Oklahoma pasture
Dependent variable Explanatory variable Estimate SE P � Fa
Percent polygyny (spatial independence model) Intercept 72.838 81.921 0.3763Elevation (m) 0.818 0.352 0.0224RVIb 0.263 0.064 �0.0001
Percent polygyny (spatial dependence model) Intercept 79.227 7.907 ÑMounds/plot 0.869 0.262 0.0013Cumulative mound vol 0.165 0.074 0.0275
Mound density Intercept 20.362 8.172 0.0145Insolation at samplingc 0.004 0.001 0.0115Canopy cover 0.004 0.002 0.0262Percent polygyny 0.006 0.001 0.0001
Cumulative mound vol Intercept 94.191 24.712 0.0002Summer insolation 0.017 0.004 0.0001Elevation 0.019 0.008 0.0216Canopy cover 0.013 0.003 �0.0001Distance to a cistern 0.0004 0.0002 0.0291Distance to water 0.002 0.001 0.0198Distance to trees 0.007 0.003 0.0296
Average mound vol Intercept 2.953 0.677 �0.0001Slope (degrees) 0.041 0.017 0.0188Elevation 0.008 0.003 0.0083Distance to water 0.0007 0.0002 0.0013Percent polygyny 0.003 0.001 �0.0001
a P value based on partial sums of squares.b Ratio vegetation index: �NIR/�Red.c Insolation values expressed as w � h/m2.
554 ENVIRONMENTAL ENTOMOLOGY Vol. 38, no. 3

mound volume, whereas plots dominated by mono-gyne colonies had low numbers of mounds with widevariation in cumulative mound volume. Some mono-gyne areas with relatively low numbers of mounds hadcumulative mound volume comparable to polygyneareas with two to three times as many mounds (Fig. 2).Overall, social formwasmoreclosely related tomounddensity than to cumulative mound volume.Mound Density. Mound density (log10-trans-
formed) was best predicted using a model includinginsolation at time of sampling, canopy cover, and per-cent polygyny (F � 15.7; df � 3,91; P � 0.0001; r2 �0.3411; Table 1). Polygyny was the most importantpredictor of mound density, accounting for �42% ofthe variation explained by the model, followed byinsolation at sampling (14%) and canopy cover (11%)(Fig. 3). These percentages are measured by analysisof commonality of the regression equation. The per-centages indicate unique contribution (partial sums ofsquares) of the independent variable, alone or in com-bination, as well as information explained in common(collinear) with other independent variables. Mounddensities ranged from �50 (20 mounds/acre) to 220mounds/ha (88 mounds/acre) for monogyne and po-lygyne areas, respectively. The effect of both insola-tion at sampling and canopy cover in combinationwith polygyny were important in the model, explain-ing �18% of the variation. Plots with monogyne ormixed populations did not approach the high mounddensities of the polygyne plots, even under conditionsof high insolation and no canopy cover. There were noindications of potential problems with collinearity,and residuals for the model were not spatially auto-correlated.
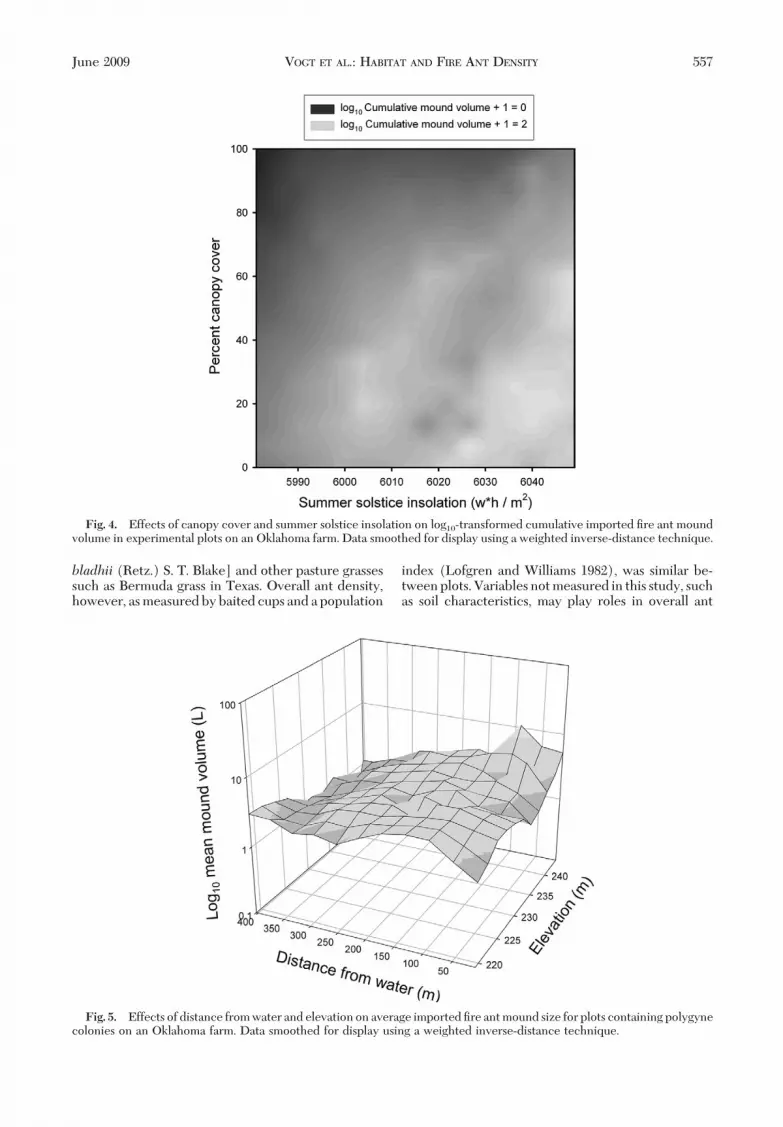
Mound Volume. A model incorporating the effectsof summer solstice insolation, elevation, canopy cover,distance to a cistern, distance to water, and distance totrees explained �42% of the variation in log10-trans-formed cumulative mound volume (F � 12.17; df �6,101; P� 0.0001; r2 � 0.4196). The models above forsocial form and mound density had fewer degrees offreedom because of exclusion of plots with zeromounds; in the case of cumulative mound volume,social form was not signiÞcant, and all plots wereincluded in the model. Canopy cover and summersolstice insolation explained the largest proportions ofthe variation (37 and 22%, respectively; Fig. 4), witheach remaining explanatory variable accounting for�8%. The combined effect of canopy cover and sum-mer solstice insolation accounted for an additional16%. Other combinations of variables made relativelyminor contributions to the model. Not surprisingly,collinearity was noted among some paired habitat andlandscape variables including insolation and elevation,canopy and tree distance, and insolation and distancefrom water, but the variation explained in commonwas generally �6%.
We also examined log10-transformed averagemound volume (calculated for each plot) to detectpossible trends caused by local conditions or socialform. Slope, elevation, distance from water, and per-cent polygyny all had signiÞcant negative effects onaverage mound volume (F � 10.14; df � 4,86; P �0.0001; r2 � 0.3204; Table 1). Polygyny accounted for�59% of the variation explained by the model, andelevation and distance from water each accounted for�19%. The combined effect of elevation and polygynyaccounted for another 17%. Areas with higher slope
Fig. 2. Relationship between mound density, cumulative mound volume, and percent polygyny in experimental plots onan Oklahoma farm.
June 2009 VOGT ET AL.: HABITAT AND FIRE ANT DENSITY 555

tended to have slightly smaller mounds. Polygynemounds within 100Ð150 m of water (a cistern or pond)tended to be about twice as large as mounds 300Ð400m from water. For plots close to water, the effect ofelevation was negligible, but average mound size de-creased with increasing elevation at greater distancesfrom water (Fig. 5). Additionally, mounds very closeto water tended to be smaller than mounds at a mod-erate distance from water. Trends were similar formounds in monogyne and mixed plots, but moundswere larger overall.
Our analyses make the assumption that Þre antdensity (mounds or cumulative mound volume) perplot can be approximated using linear models. Al-though count data frequently follow a Poisson distri-bution when the average count is close to zero, ourdata do not have an abundance of zeros, the log10-transformed values are used, and inference is aboutthe mean (predicted) values. We conclude that theassumption of normality is valid but caution that ap-plying our models to data that have an abundance ofzeros may be problematic because of prediction out-side of the range of data and difÞculties caused by useof the wrong distribution.
Discussion
Social Form.Monogyne colonies in our study werestrongly aggregated in areas of low elevation and high
vegetation density, but these trends were not signif-icant in a model taking spatial autocorrelation intoaccount. Clearly, additional data are needed to seewhether elevation, vegetation density, or other habitatcharacteristics inßuence social form of imported Þreant colonies at a Þeld scale. Porter et al. (1991) foundno differences in social form caused by habitat char-acteristics (proximity to trees, vegetation type, soiltype, climatic conditions, topography, ground cover,and solar orientation) during an extensive survey forpolygyne populations in Texas. Resampling severalmonths later showed that a small percentage of sites(�18%) switched from monogyny to polygyny, or viceversa; however, error associated with determination ofsocial form and exact site location made interpretationof results difÞcult. Their study was not designed todetect differences in social form or abundance on aÞne scale within Þelds; rather, it provided regionaldata on social form and abundance across severalTexascounties.Porter(1993)observed littlechange insocial form at Þeld sites over a 1- to 3-yr period,whereas Greenberg et al. (1992) documented an in-crease in polygyny in a Texas pasture over a period of9 yr, although some monogyne colonies persisted inthe same area. Neither study included detailed char-acterization of habitat variables or landscape metrics.Sternberg et al. (2006) documented possible differ-ences in social form and mound density in adjoiningplots of a variety of Old World bluestem [Bothriochloa
Fig. 3. Relationship between percent canopy cover, insolation, and mound density for experimental plots containingdifferent social forms of imported Þre ants on an Oklahoma farm.
556 ENVIRONMENTAL ENTOMOLOGY Vol. 38, no. 3
bladhii (Retz.) S. T. Blake] and other pasture grassessuch as Bermuda grass in Texas. Overall ant density,however, as measured by baited cups and a population
index (Lofgren and Williams 1982), was similar be-tween plots. Variables not measured in this study, suchas soil characteristics, may play roles in overall ant
Fig. 4. Effects of canopy cover and summer solstice insolation on log10-transformed cumulative imported Þre ant moundvolume in experimental plots on an Oklahoma farm. Data smoothed for display using a weighted inverse-distance technique.
Fig. 5. Effects of distance from water and elevation on average imported Þre ant mound size for plots containing polygynecolonies on an Oklahoma farm. Data smoothed for display using a weighted inverse-distance technique.
June 2009 VOGT ET AL.: HABITAT AND FIRE ANT DENSITY 557

density or social form (Milks et al. 2007). Remotelysensed data such as vegetation indices may show pat-terns that are not immediately obvious to researcherson the ground. Although it is generally acknowledgedthat polygyne populations are expanding in the UnitedStates, if monogyne colonies persist in discrete areas,it is possible that certain environmental conditionsfavor one social form over another. We intend torevisit this study site to examine stability of monogyneand polygyne populations in the future.Mound Density. It is evident from Fig. 3 that im-
ported Þre ant colonies can exist in partially canopiedareas, provided insolation is sufÞcient. This is in agree-ment with recent work by Menzel and Nebeker(2008), who described the effects of local environ-mental conditions on hybrid imported Þre ant densityand interspeciÞc competition in Mississippi forests.Indicators of forest disturbance (open canopy, highvegetative ground cover, and high percentage ofpines) were positively associated with abundance ofimported Þre ants, but they were collected under arange of canopy cover conditions. Our results indi-cated that the ability for imported Þre ants to invadepartially canopied areas on a local scale is inßuencedby overlying patterns of insolation. Insolation at thetime of sampling was a better predictor of mounddensity than insolation during the winter or summersolstices, when considered with canopy cover andpolygyny. Differences between the models weresmall, and all models were signiÞcant. Imported Þreant nests are not permanent structures, and it is likelythat they move in response to local habitat variations.Some ant species are known to relocate their nest sitesseasonally; for example, the invasive Argentine ant[Linepithema humile (Mayr)] is seasonally polydo-mous, moving to and from nest sites in response tomicroclimate changes (Heller and Gordon 2006). Sev-eral other ant species are known to move with varyingfrequency (Smallwood 1982 and references therein).S. richteri abandon their mounds and construct newones every 3.1 mo on average in Argentina, wheremovement is stimulated by rainfall (Briano et al.1995). It would be useful to collect imported Þre antdistribution data and calculate corresponding insola-tion data at different times of year to determinewhether colonies move in a predictable fashion inresponse to local changes in insolation.MoundVolume.Mound volume is closely related to
colony biomass (Tschinkel 1993, Macom and Porter1996) and provides a good estimate of overall im-ported Þre ant density in areas. Polygyny was notsigniÞcant in our predictive model, suggesting thatenvironmental or habitat factors inßuence total antbiomass in similar ways, regardless of social form. Sum-mer solstice insolation was positively correlated withcumulative volume, whereas increased elevation andincreasing distance from trees and water sources werenegatively correlated. In the study of Tschinkel (1988)of monogyne S. invicta, imported Þre ants seemed tobe limited in distribution to pond edges in some hab-itats, but it was unclear whether this was caused by ahigh water table and less xeric conditions, frequency
of ßooding (to which S. invicta is adapted), or negativeimpacts on competing ant species. Early observationsby Lyle and Fortune (1948) indicated that importedÞre ant mounds were located primarily in low areassuch as ditches or near bodies of water in southernAlabama and Mississippi. There has been speculationamong entomologists that imported Þre ant alates maybe attracted to bodies of water, but to our knowledge,this has not been borne out with formal observations.In our study, close proximity to artiÞcial water sources(cisterns) and natural water sources (ponds) bothfavored high ant density. The negative effect of in-creased distance from trees is more difÞcult to explain.Imported Þre ants readily forage in tree canopies, andtrees may represent an important resource in terms ofinsect prey and honeydew, enabling nearby areas tosupport higher ant populations.
Average mound volume was also negatively corre-lated with elevation and distance from water; thus, wecannot say whether the effects of elevation and dis-tance from water sources in the model for cumulativemound volume above were a direct result of lowerpopulation density or a result of local conditions in-ßuencing the nature and/or size of mounds. In a studyby Macom and Porter (1996), 77Ð85% of variation inimported Þre ant mound volume was explained byworker number, total number of individuals, workerbiomass, and colony biomass. No differences in theserelationships were detected between monogyne andpolygyne colonies. Given the close relationship be-tween mound volume and colony size, the secondexplanation above is unlikely, but should not be dis-missed without further study.
This study yielded information on habitat charac-teristics that tend to favor (increased insolation, prox-imity to water) or suppress (canopy cover, elevation)imported Þre ant populations. Of the habitat variablesexamined, canopy cover and insolation were the mostimportant predictors of Þre ant population density.Combined effects of these two variables suggest thatÞre ants can inhabit areas with some canopy cover,provided that sufÞcient sun exposure occurs in thoseareas. A better understanding of habitat characteris-tics and landscape metrics affecting Þre ant popula-tions is a Þrst step in predicting population densitybased on these variables. Our results may be useful inplanning Þeld trials or other research projects thatrequire high Þre ant abundance by allowing research-ers to concentrate on those areas within a larger areathat are likely to have numerous ant colonies based notonly on canopy cover but insolation and other vari-ables. For regional detection efforts using remotesensing (Vogt 2004), these data would be useful forßight planning to concentrate on areas with poten-tially high Þre ant abundance, thus reducing costs, orareas with potentially larger mounds, increasing de-tection rates. Additional habitat characteristics thatmight inßuence Þre ant abundance include abun-dance of other arthropods and plant species compo-sition, neither of which is readily acquired using re-motely sensed data or internet sources. As mentionedabove, additional soil characteristics (soil moisture
558 ENVIRONMENTAL ENTOMOLOGY Vol. 38, no. 3

and chemical and physical properties) may be impor-tant predictors of Þre ant abundance. Finally, land usepatterns such as grazing intensity and managementpractices are likely to affect Þre ant abundance. Workis underway to apply these models to unique sites indifferent geographic regions to test predictive valueand to determine the effects of potential predictivevariables at increased spatial resolution inpastures andother areas.
Acknowledgments
We thank S. Coy and A. Martin (USDAÐARS), K. Varner(Oklahoma Cooperative Extension Service), and C. Mackey(Wes Watkins Agricultural Research and Extension Center)for assistance in the Þeld; C. Smith (social form determina-tions) for expert laboratory assistance; M. Aubuchon and J. L.Thomson for helpfully reviewing the manuscript; and C.Mitchell for access to his farm.
References Cited
Adams, E. S., andW.R.Tschinkel. 1995. Spatial dynamics ofcolony interactions in young populations of the Þre antSolenopsis invicta. Oecologia (Berl.) 102: 156Ð163.
Allen,C.R.,A.R. Johnson, andL.Parris. 2006. Aframeworkfor spatial risk assessments: potential impacts of non-indigenous invasive species on native species. Ecol.Soc. 11: 39.
Allen,G.E.,W.F.Buren,R.N.Williams,M. deMenezes, andW. H. Whitcomb. 1974. The red imported Þre ant, So-lenopsis invicta; distribution and habitat in Mato Grosso,Brazil. Ann. Entomol. Soc. Am. 67: 43Ð46.
Baroni-Urbani, C., and P. B. Kannowski. 1974. Patterns inthe red imported Þre ant settlement of a Louisiana pas-ture: some demographic parameters, interspeciÞc com-petition and food sharing. Environ. Entomol. 3: 755Ð760.
Briano, J. A., R. S. Patterson, andH. A. Cordo. 1995. Colonymovement of the black imported Þre ant (Hymenoptera:Formicidae) in Argentina. Environ. Entomol. 24: 1131Ð1134.
Chen, J., and M. L. Allen. 2006. SigniÞcance of digging be-havior to mortality of red imported Þre ant workers,Solenopsis invicta, in Fipronil-treated sand. J. Econ. En-tomol. 99: 476Ð482.
Colby, D., and D. Prowell. 2006. Ants (Hymenoptera: For-micidae) in wet longleaf pine savannas in Louisiana. Fla.Entomol. 89: 266Ð269.
Emigh, T. H. 1984. Path analysis, correlation, and the anal-ysis of variance, pp. 213Ð230. In K. Hinkelmann (ed.),Experimental design, statistical models, and genetic sta-tistics: essays in honor of Oscar Kempthorne. MarcelDekker, New York.
Fitzpatrick, M. C., J. F. Weltzin, N. J. Sanders, and R. R.Dunn. 2007. The biogeography of prediction error: whydoes the introduced range of the Þre ant over-predict itsnative range? Global Ecol. Biogeogr. 16: 24Ð33.
Fleetwood,S.C.,P.D.Teel, andG.Thompson. 1984. Impactof imported Þre ant on lone star tick mortality in open andcanopied pasture habitats of east central Texas. South-west. Entomol. 9: 158Ð163.
Green, A. A., and M. D. Craig. 1985. Analysis of aircraftspectrometer data with logarithmic residuals. Jet Propul-sion Laboratory Publication 85Ð41, pp. 111Ð119.
Greenberg, L., S. B. Vinson, and S. Ellison. 1992. Nine-yearstudy of a Þeld containing both monogyne and polygyne
red imported Þre ants (Hymenoptera: Formicidae). Ann.Entomol. Soc. Am. 85: 686Ð695.
Heller, N. E., and D. M. Gordon. 2006. Seasonal spatial dy-namics and causes of nest movement in colonies of theinvasive Argentine ant (Linepithema humile). Ecol. En-tomol. 31: 499Ð510.
Jordan,C.F. 1969. Derivation of leaf area index from qualityof light on the forest ßoor. Ecology 50: 663Ð666.
Killian,M. J., andW.E.Grant. 1995. A colony growth modelfor the imported Þre ant: potential geographic range of aninvading species. Ecol. Model. 77: 73Ð84.
Korzukhin, M. D., S. D. Porter, L. C. Thompson, and S.Wiley. 2001. Modeling temperature-dependent rangelimits for the Þre ant Solenopsis invicta (Hymenoptera:Formicidae) in the United States. Environ. Entomol. 30:645Ð655.
Lofgren, C., and D. F. Williams. 1982. Avermectin B1: ahighly potent inhibitor of reproduction by queens of thered imported Þre ant. J. Econ. Entomol. 75: 798Ð803.
Lyle,C., and I.Fortune. 1948. Notes on an imported Þre ant.J. Econ. Entomol. 41: 833Ð834.
Macom, T. E., and S. D. Porter. 1996. Comparison of poly-gyne and monogyne red imported Þre ant (Hymenop-tera: Formicidae) population densities. Ann. Entomol.Soc. Am. 89: 535Ð543.
Menzel, T. O., and T. E. Nebeker. 2008. Distribution ofhybrid imported Þre ants (Hymenoptera: Formicidae)and some native ant species in relation to local envi-ronmental conditions and interspeciÞc competition inMississippi forests. Ann. Entomol. Soc. Am. 101: 119Ð127.
Milks, M. L., J. R. Fuxa, A. R. Richter, and E. B. Moser.2007. Multivariate analyses of the factors affecting thedistribution, abundance and social form of LouisianaÞre ants, Solenopsis invicta. Insect. Soc. 54: 283Ð292.
Morrison, L. W., S. D. Porter, E. Daniels, and M. D. Kor-zukhin. 2004. Potential global range expansion of theinvasive Þre ant, Solenopsis invicta.Biol. Invas. 6: 183Ð191.
Peterson, A. T., and Y. Nakazawa. 2008. Environmental datasets matter in ecological niche modeling: an example withSolenopsis invicta and Solenopsis richteri. Global Ecol.Biogeogr. 17: 135Ð144.
Porter, S. D. 1993. Stability of polygyne and monogyne Þreant populations (Hymenoptera: Formicidae) in theUnited States. J. Econ. Entomol. 86: 1344Ð1347.
Porter, S. D., A. Bhatkar, R. Mulder, S. B. Vinson, and D. J.Clair. 1991. Distribution and density of polygyne Þreants (Hymenoptera: Formicidae) in Texas. J. Econ. En-tomol. 84: 866Ð874.
Smallwood, J. 1982. Nest relocations in ants. Insectes Soc.29: 138Ð147.
Sternberg, T., G. Perry, and C. Britton. 2006. Grass repel-lency to the red imported Þre ant. Rangeland Ecol.Manag. 59: 330Ð333.
Stoker, R. L., D. K. Ferris,W. E. Grant, and L. J. Folse. 1994.Simulating colonization by exotic species: a model of thered imported Þre ant (Solenopsis invicta) in North Amer-ica. Ecol. Model. 73: 281Ð292.
Stiles, J. H., and R. H. Jones. 1998. Distribution of the redimported Þre ant, Solenopsis invicta, in road and power-line habitats. Landscape Ecol. 335: 335Ð346.
Sutherst, R. W., and G. Maywald. 2005. A climate model ofthe red imported Þre ant, Solenopsis invicta Buren (Hy-menoptera: Formicidae): implications of new regions,particularly Oceania. Environ. Entomol. 34: 317Ð355.
Thorvilson, H. G., J. C. Cokendolpher, and S. A. Phillips, Jr.1992. Survival of the red imported Þre ant (Hymenop-
June 2009 VOGT ET AL.: HABITAT AND FIRE ANT DENSITY 559

tera: Formicidae) on the Texas high plains. Environ. En-tomol. 21: 964Ð968.
Tschinkel, W. R. 1988. Distribution of the Þre ants Solenop-sis invicta and S. geminata (Hymenoptera: Formicidae) innorthern Florida in relation to habitat and disturbance.Ann. Entomol. Soc. Am. 81: 76Ð81.
Tschinkel, W. R. 1993. Sociometry and sociogenesis of col-onies of the Þre ant Solenopsis invicta during one annualcycle. Ecol. Monogr. 64: 425Ð457.
Tschinkel,W.R., andD. F.Howard. 1983. Colony foundingby pleometrosis in the Þre ant, Solenopsis invicta. Behav.Ecol. Sociobiol. 12: 103Ð113.
Vogt, J. T. 2004. Quantifying imported Þre ant (Hymenop-tera: Formicidae) mounds with airborne digital imagery.Environ. Entomol. 33: 1045Ð1051.
Vogt, J.T. 2007. Three-dimensional sampling method for char-acterizing ant mounds. Fla. Entomol. 90: 553Ð558.
Vogt, J. T., D. A. Streett, R.M. Pereira, andA.M.A. Callcott.2003. Mississippi areawide Þre ant suppression pro-gram: unique aspects of working with black and hybridimported Þre ants. J. Agric. Urban Entomol. 20: 105Ð111.
Received 3 December 2007; accepted 13 February 2009.
560 ENVIRONMENTAL ENTOMOLOGY Vol. 38, no. 3





























