Embed Size (px)
Citation preview

The
Pla
nt C
ell
This article is published in
The Plant Cell
Online,
The Plant Cell
Preview Section, which publishes manuscripts accepted for publication afterthey have been edited and the authors have corrected proofs, but before the final, complete issue is published online. Early posting of articles
reduces normal time to publication by several weeks.
The Plant Cell
Preview, www.aspb.org © 2003 American Society of Plant Biologists 1 of 12
Dissection of Arabidopsis Bax Inhibitor-1 Suppressing Bax–, Hydrogen Peroxide–, and Salicylic Acid–Induced Cell Death
Maki Kawai-Yamada, Yuri Ohori, and Hirofumi Uchimiya
1
Institute of Molecular and Cellular Biosciences, University of Tokyo, 1-1-1 Yayoi, Bunkyo-ku, Tokyo 113-0032, Japan
Overexpression of plant Bax Inhibitor-1 (BI-1) was able to suppress Bax-mediated cell death in yeast and Arabidopsis. Here,we demonstrate that reactive oxygen species production induced by the ectopic expression of Bax was insensitive to thecoexpression of AtBI-1. Similarly, H
2
O
2
– or salicylic acid–mediated cell death also was suppressed in tobacco BY-2 cells
overexpressing AtBI-1. To define the functional domain of AtBI-1 as a cell death suppressor, a truncated series of the AtBI-1
protein was analyzed in yeast possessing a galactose-inducible mammalian Bax. The results showed that
�
C-AtBI-1 (with
the C-terminal 14 amino acids deleted) lost the ability to sustain cell growth. Furthermore, a mutant protein in which theC-terminal seven amino acid residues of AtBI-1 were replaced with others lacking a coiled-coil structure failed to inhibit celldeath, suggesting that the C-terminal region is essential for the inhibition of cell death. We also noted that the C-terminalhydrophilic region was interchangeable between animal and plant Bax inhibitors.
INTRODUCTION
Like animal cells, plant cells can respond to various stimuli, in-cluding fungal toxins and biotic and abiotic stresses, by initiat-ing programmed cell death (PCD). Some morphological andbiochemical changes, such as cell shrinkage, chromatin con-densation, and DNA fragmentation, seem to be common fea-tures of cell death because they occur in both plant and animalcells (Lam et al., 2001). In animals, the key regulators of apop-tosis are evolutionarily conserved. For example, the ced-9 pro-tein in
Caenorhabditis elegans
is homologous with the antiap-optotic members of the Bcl-2 family (Hengartner and Horvitz,1994), which is a large family of proteins that act as either acti-vators (e.g., Bax and Bak) or suppressors (e.g., Bcl-2 and Bcl-XL) of PCD (reviewed by Reed, 1994; Kroemer, 1997). Similarly,the main executors of PCD, such as caspases, are conservedin the animal kingdom (Yuan et al., 1993). By contrast, in a Ba-sic Local Alignment Search Tool (BLAST) database search ofthe yeast and plant genomes, no obvious homologs of any cru-cial regulators of metazoan apoptosis (members of the Bax/Bcl-2 family, caspases, Apaf-a/Ced-4, p53) were detected.However, the introduction of mammalian death regulators inyeast and plant cells induced the proper phenotype with regardto the nature of the gene introduced (Lacomme and Cruz,1999; Mitsuhara et al., 1999; Kawai-Yamada et al., 2001).
It has been noted that yeast cells, both
Saccharomyces cere-visiae
and
Schizosaccharomyces pombe
, can undergo celldeath by the expression of the proapoptotic mammalian genesBax and Bak (Bischoff et al., 1992; Sato et al., 1994; Hanada et
al., 1995; Greenhalf et al., 1996). Madeo et al. (2002) demon-strated that Yor197w, a yeast protein with structural homologywith mammalian caspases, has caspase-like processing activ-ity and regulates H
2
O
2
-induced yeast death. Therefore, ele-ments of the PCD pathway conserved in yeast as well as ani-mals should belong to a basic and ancient, evolutionarilyconserved mechanism. In animal cells, the mitochondrion playsa crucial role in PCD. Upon perceiving cell death signals, theproapoptotic protein Bax forms channels in the outer mem-brane of the mitochondrion and triggers the release of cyto-chrome
c
(Green and Reed, 1998). The latter subsequently acti-vates a series of caspases that result in the proteolysis ofproteins essential for the maintenance of cell integrity.
Oxidative stress also was found to be involved in plant PCDprocesses (Lamb and Dixon, 1997; Mittler, 2002). Several in-vestigators have shown that H
2
O
2
induced PCD in suspensioncultures of soybean (Levine et al., 1994), Arabidopsis (Desikanet al., 1998), and tobacco (Houot et al., 2001) cells. In addition,Houot et al. (2001) showed that H
2
O
2
induces PCD through aprocess similar to apoptosis, including cell shrinkage, chroma-tin condensation, and DNA fragmentation. The process in-duced by H
2
O
2
depends on active cellular metabolism and canbe blocked by protease inhibitors (Solomon et al., 1999). Raoand Davis (1999) demonstrated that treatment with salicylicacid (SA) caused plant cell death by enhanced H
2
O
2
produc-tion, lipid peroxidation, and oxidative damage to proteins.
Although no Bax homolog has been identified in plant ge-nomes to date, the overexpression of mammalian Bax in to-bacco (Lacomme and Cruz, 1999) and Arabidopsis (Kawai-Yamada et al., 2001) causes cell death. Thus, when the animalBax gene is expressed in plant cells under a dexamethasone(DEX)-inducible system, such plants exhibit marked cell deathat the whole-plant level, with cell shrinkage, membrane de-struction, and other apoptotic phenotypes (Kawai-Yamada et
1
To whom correspondence should be addressed. E-mail [email protected]; fax 81-3-5841-8466.Article, publication date, and citation information can be found atwww.plantcell.org/cgi/doi/10.1105/tpc.014613.
The
Pla
nt C
ell
2 of 12 The Plant Cell
al., 2001). Recently, Abramovitch and co-workers (2003) dem-onstrated that the
Pseudomonas
type III effecter AvrPtoB sup-presses Bax-induced cell death in yeast. They showed that thisgene induces plant disease susceptibility by inhibiting hostPCD. The morphological and biochemical features resulting fromthe ectopic expression of human death regulators (Lacommeand Cruz, 1999; Mitsuhara et al., 1999; Kawai-Yamada et al.,2001) strongly suggest that death mechanisms in plants areoperational as in animal cells.
Xu and Reed (1998) identified a human cDNA that sup-presses Bax-mediated cell death in yeast, and the correspond-ing protein was named Bax Inhibitor-1 (BI-1). Subsequently, weisolated BI-1 homologs from rice (OsBI-1) and Arabidopsis(AtBI-1) and showed that the overexpression of plant BI-1 alsocould suppress Bax-mediated cell death in yeast (Kawai et al.,1999). The chlorosis caused by Bax expression was retarded intransgenic Arabidopsis expressing both Bax and AtBI-1, sug-gesting that the plant antiapoptotic protein AtBI-1 is biologi-cally active in suppressing mammalian Bax action in planta.The BI-1 protein has seven transmembrane domains and isthought to be localized in the endoplasmic reticulum (ER) mem-brane that includes the nuclear envelope (Xu and Reed, 1998;Kawai-Yamada et al., 2001; Bolduc et al., 2003). Interestingly,the evolutionarily conserved function of both plant and animalBI-1 also was demonstrated in a mammalian cell culture sys-tem (Yu et al., 2002; Bolduc et al., 2003).
Direct evidence for the role of AtBI-1 in PCD was presentedrecently in a study of elicitor-induced hypersensitive response(HR) in rice suspension cells (Matsumura et al., 2003). The elici-tors isolated from rice blast pathogen induced cell deaththrough the stimulation of reactive oxygen species (ROS). Suchcell death was overcome by the overexpression of AtBI-1. Fur-thermore, the role of BI-1 in Mlo-mediated resistance to
Blume-ria graminis
also was demonstrated recently by an overexpres-sion analysis of barley BI-1 (Hückelhoven et al., 2003). Todetermine the mode of action of AtBI-1 in suppressing celldeath, AtBI-1 was overexpressed in tobacco suspension-cul-tured cells. Here, we demonstrate that the C terminus of AtBI-1is essential for the suppression of H
2
O
2
- and SA-induced plantcell death. The precise base substitution analysis of the C-ter-minal region of AtBI-1 clearly demonstrated the critical role ofthe C-terminal amino acid composition of the BI-1 protein asER membrane components, which play a novel role against thedetrimental effects associated with oxidative stresses.
RESULTS
AtBI-1 Does Not Suppress the Bax-Induced
Accumulation of O
2
�
The overexpression of mammalian Bax in Arabidopsis plantstriggers cell death (Kawai-Yamada et al., 2001). To test whetherROS production is involved in this process, transgenic Arabi-dopsis plants were grown in the presence or absence of 5
�
MDEX and leaves were stained with nitroblue tetrazolium (NBT)(Figure 1). Blue formazan precipitates, produced by the reac-tion of NBT with generated O
2
�
, were evident in leaves of Baxtransgenic plants within 7 h after Bax treatment (Figures 1B,
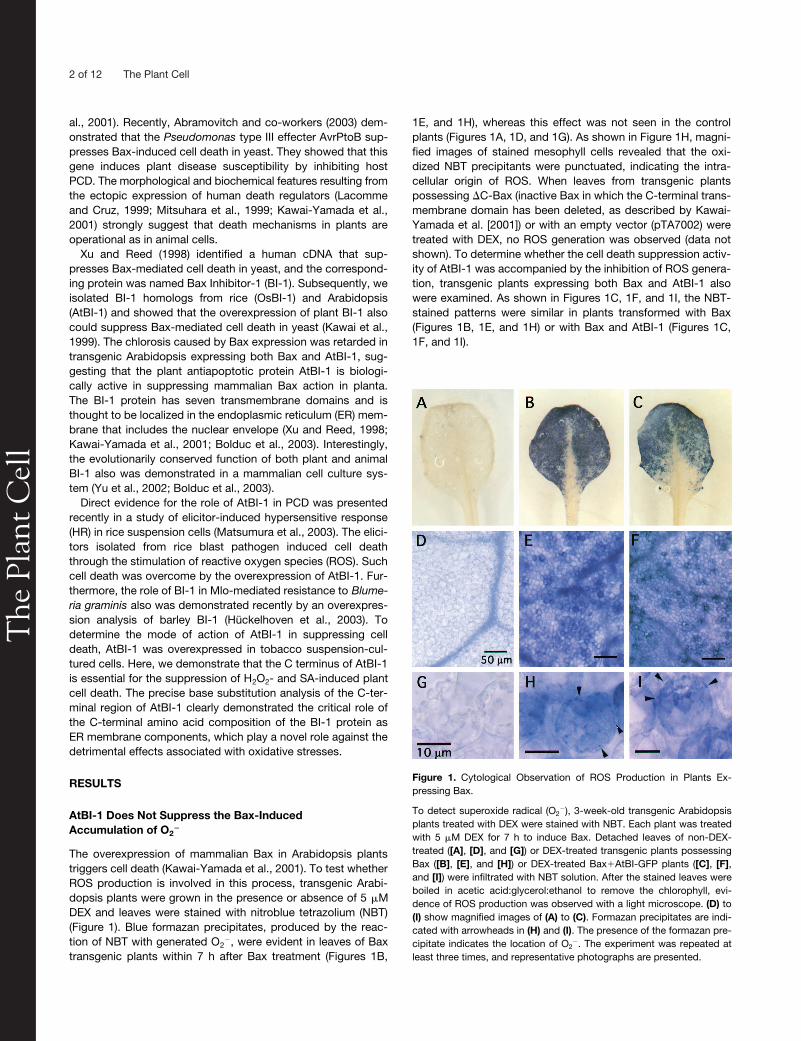
1E, and 1H), whereas this effect was not seen in the controlplants (Figures 1A, 1D, and 1G). As shown in Figure 1H, magni-fied images of stained mesophyll cells revealed that the oxi-dized NBT precipitants were punctuated, indicating the intra-cellular origin of ROS. When leaves from transgenic plantspossessing
�
C-Bax (inactive Bax in which the C-terminal trans-membrane domain has been deleted, as described by Kawai-Yamada et al. [2001]) or with an empty vector (pTA7002) weretreated with DEX, no ROS generation was observed (data notshown). To determine whether the cell death suppression activ-ity of AtBI-1 was accompanied by the inhibition of ROS genera-tion, transgenic plants expressing both Bax and AtBI-1 alsowere examined. As shown in Figures 1C, 1F, and 1I, the NBT-stained patterns were similar in plants transformed with Bax(Figures 1B, 1E, and 1H) or with Bax and AtBI-1 (Figures 1C,1F, and 1I).
Figure 1. Cytological Observation of ROS Production in Plants Ex-pressing Bax.
To detect superoxide radical (O2�), 3-week-old transgenic Arabidopsis
plants treated with DEX were stained with NBT. Each plant was treatedwith 5 �M DEX for 7 h to induce Bax. Detached leaves of non-DEX-treated ([A], [D], and [G]) or DEX-treated transgenic plants possessingBax ([B], [E], and [H]) or DEX-treated Bax�AtBI-GFP plants ([C], [F],and [I]) were infiltrated with NBT solution. After the stained leaves wereboiled in acetic acid:glycerol:ethanol to remove the chlorophyll, evi-dence of ROS production was observed with a light microscope. (D) to(I) show magnified images of (A) to (C). Formazan precipitates are indi-cated with arrowheads in (H) and (I). The presence of the formazan pre-cipitate indicates the location of O2
�. The experiment was repeated atleast three times, and representative photographs are presented.
The
Pla
nt C
ell
AtBI-1 Suppresses Oxidative Cell Death 3 of 12
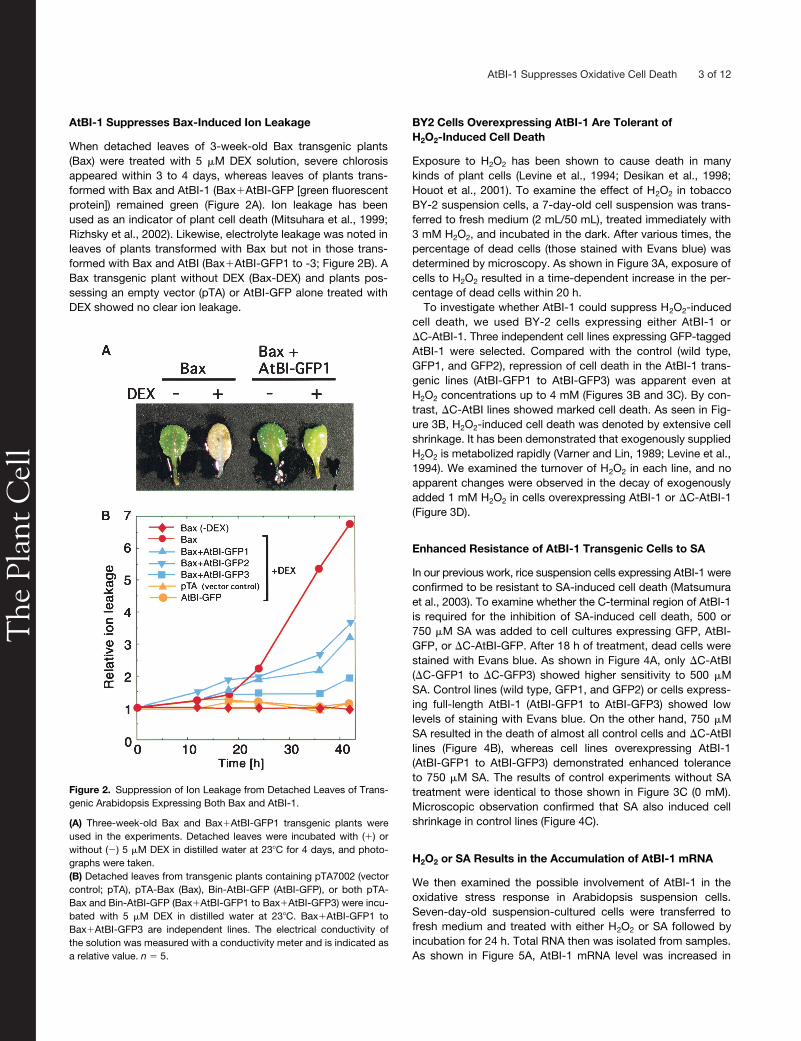
AtBI-1 Suppresses Bax-Induced Ion Leakage
When detached leaves of 3-week-old Bax transgenic plants(Bax) were treated with 5
�
M DEX solution, severe chlorosisappeared within 3 to 4 days, whereas leaves of plants trans-formed with Bax and AtBI-1 (Bax
�
AtBI-GFP [green fluorescentprotein]) remained green (Figure 2A). Ion leakage has beenused as an indicator of plant cell death (Mitsuhara et al., 1999;Rizhsky et al., 2002). Likewise, electrolyte leakage was noted inleaves of plants transformed with Bax but not in those trans-formed with Bax and AtBI (Bax
�
AtBI-GFP1 to -3; Figure 2B). ABax transgenic plant without DEX (Bax-DEX) and plants pos-sessing an empty vector (pTA) or AtBI-GFP alone treated withDEX showed no clear ion leakage.
BY2 Cells Overexpressing AtBI-1 Are Tolerant of H
2
O
2
-Induced Cell Death
Exposure to H
2
O
2
has been shown to cause death in manykinds of plant cells (Levine et al., 1994; Desikan et al., 1998;Houot et al., 2001). To examine the effect of H
2
O
2
in tobaccoBY-2 suspension cells, a 7-day-old cell suspension was trans-ferred to fresh medium (2 mL/50 mL), treated immediately with3 mM H
2
O
2
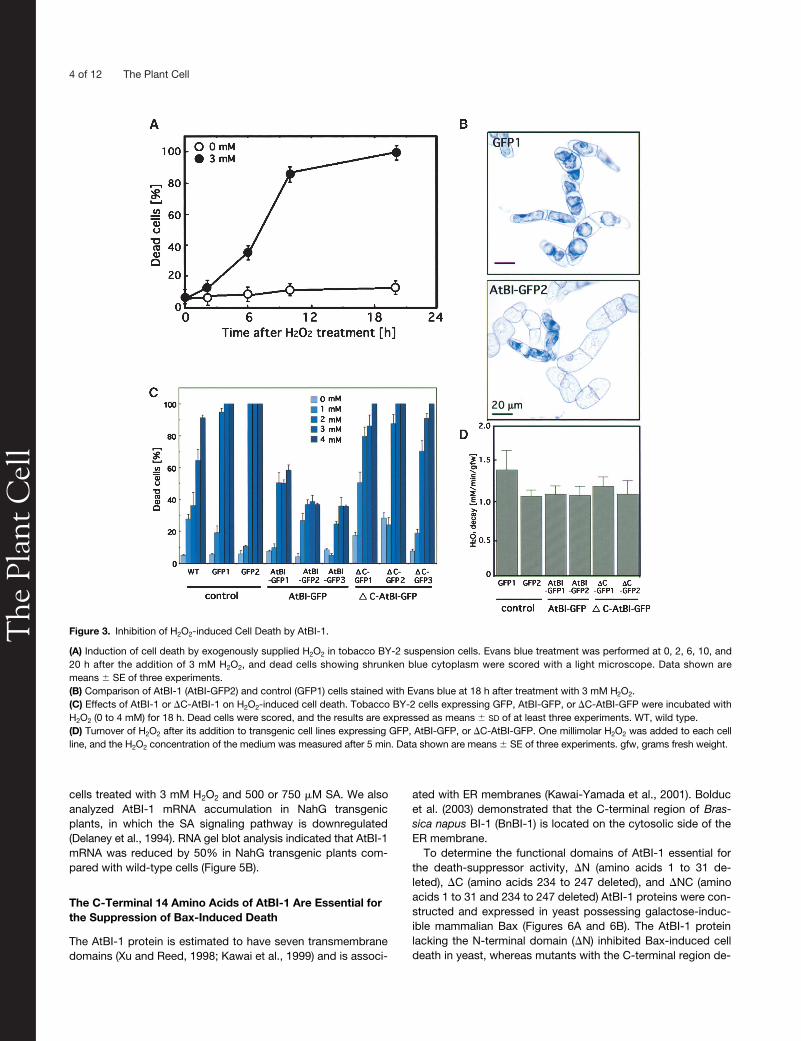
, and incubated in the dark. After various times, thepercentage of dead cells (those stained with Evans blue) wasdetermined by microscopy. As shown in Figure 3A, exposure ofcells to H
2
O
2
resulted in a time-dependent increase in the per-centage of dead cells within 20 h.
To investigate whether AtBI-1 could suppress H
2
O
2
-inducedcell death, we used BY-2 cells expressing either AtBI-1 or
�
C-AtBI-1. Three independent cell lines expressing GFP-taggedAtBI-1 were selected. Compared with the control (wild type,GFP1, and GFP2), repression of cell death in the AtBI-1 trans-genic lines (AtBI-GFP1 to AtBI-GFP3) was apparent even atH
2
O
2
concentrations up to 4 mM (Figures 3B and 3C). By con-trast,
�
C-AtBI lines showed marked cell death. As seen in Fig-ure 3B, H
2
O
2
-induced cell death was denoted by extensive cellshrinkage. It has been demonstrated that exogenously suppliedH
2
O
2
is metabolized rapidly (Varner and Lin, 1989; Levine et al.,1994). We examined the turnover of H
2
O
2
in each line, and noapparent changes were observed in the decay of exogenouslyadded 1 mM H
2
O
2
in cells overexpressing AtBI-1 or
�
C-AtBI-1(Figure 3D).
Enhanced Resistance of AtBI-1 Transgenic Cells to SA
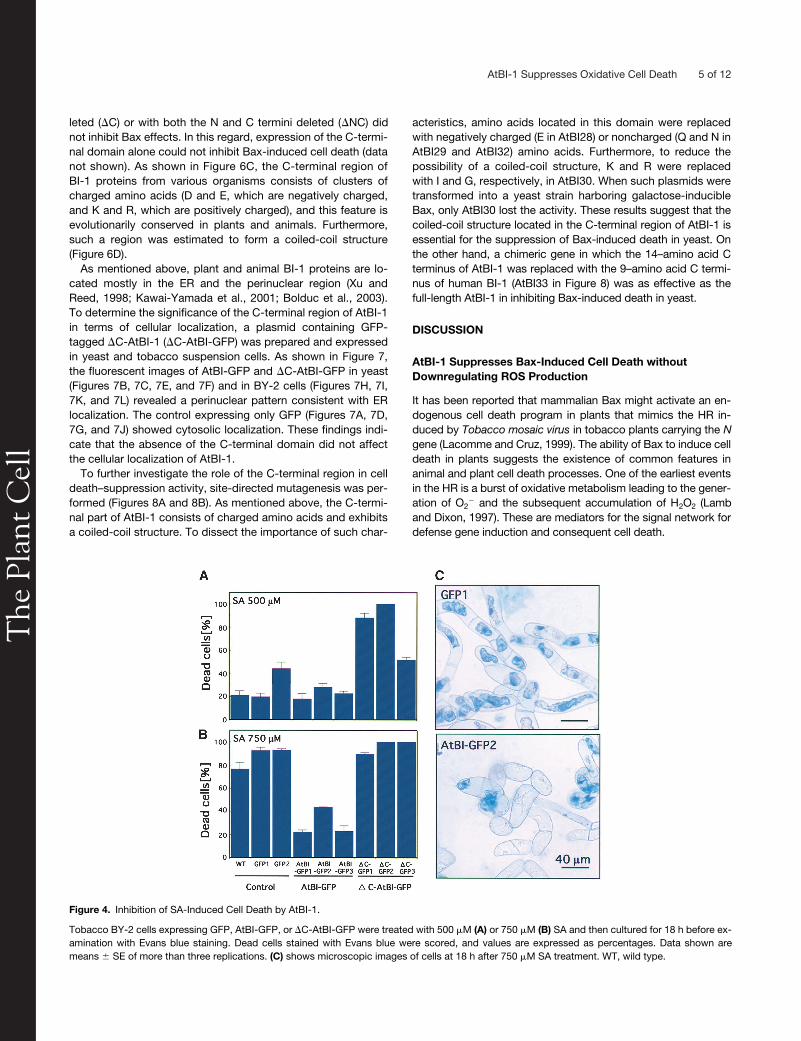
In our previous work, rice suspension cells expressing AtBI-1 wereconfirmed to be resistant to SA-induced cell death (Matsumuraet al., 2003). To examine whether the C-terminal region of AtBI-1is required for the inhibition of SA-induced cell death, 500 or750
�
M SA was added to cell cultures expressing GFP, AtBI-GFP, or
�
C-AtBI-GFP. After 18 h of treatment, dead cells werestained with Evans blue. As shown in Figure 4A, only
�
C-AtBI(
�
C-GFP1 to
�
C-GFP3) showed higher sensitivity to 500
�
MSA. Control lines (wild type, GFP1, and GFP2) or cells express-ing full-length AtBI-1 (AtBI-GFP1 to AtBI-GFP3) showed lowlevels of staining with Evans blue. On the other hand, 750
�
MSA resulted in the death of almost all control cells and
�
C-AtBIlines (Figure 4B), whereas cell lines overexpressing AtBI-1(AtBI-GFP1 to AtBI-GFP3) demonstrated enhanced toleranceto 750
�
M SA. The results of control experiments without SAtreatment were identical to those shown in Figure 3C (0 mM).Microscopic observation confirmed that SA also induced cellshrinkage in control lines (Figure 4C).
H
2
O
2
or SA Results in the Accumulation of AtBI-1 mRNA
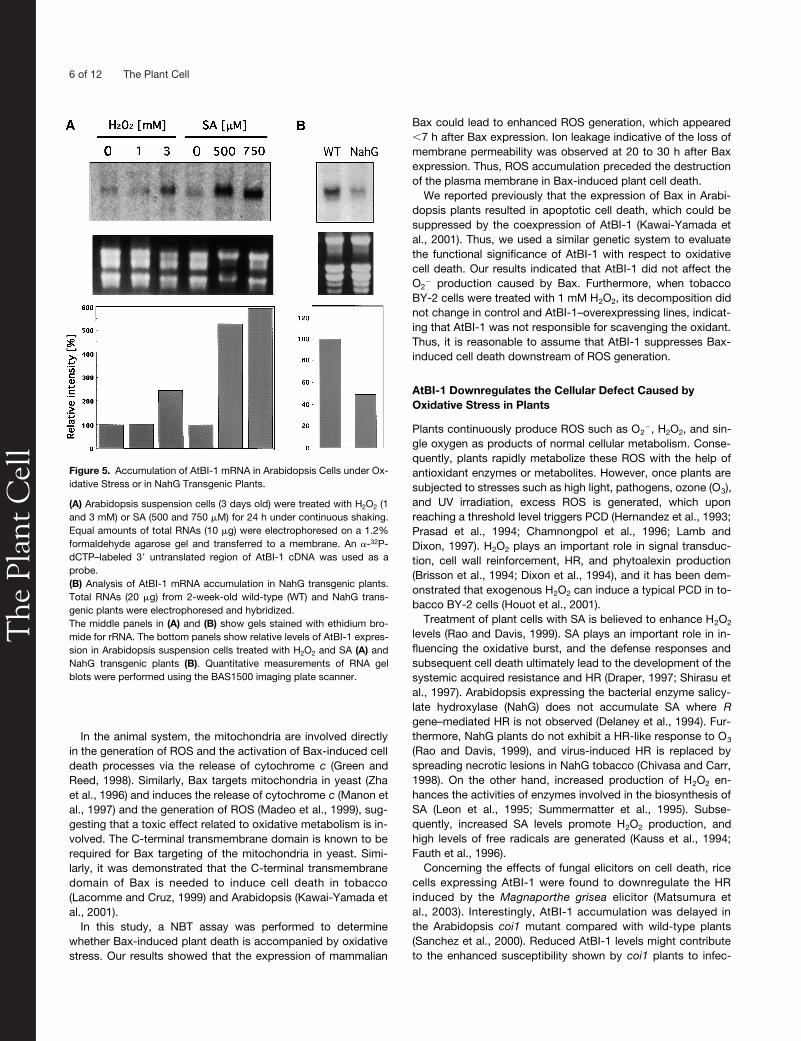
We then examined the possible involvement of AtBI-1 in theoxidative stress response in Arabidopsis suspension cells.Seven-day-old suspension-cultured cells were transferred tofresh medium and treated with either H
2
O
2
or SA followed byincubation for 24 h. Total RNA then was isolated from samples.As shown in Figure 5A, AtBI-1 mRNA level was increased in
Figure 2. Suppression of Ion Leakage from Detached Leaves of Trans-genic Arabidopsis Expressing Both Bax and AtBI-1.
(A) Three-week-old Bax and Bax�AtBI-GFP1 transgenic plants wereused in the experiments. Detached leaves were incubated with (�) orwithout (�) 5 �M DEX in distilled water at 23�C for 4 days, and photo-graphs were taken.(B) Detached leaves from transgenic plants containing pTA7002 (vectorcontrol; pTA), pTA-Bax (Bax), Bin-AtBI-GFP (AtBI-GFP), or both pTA-Bax and Bin-AtBI-GFP (Bax�AtBI-GFP1 to Bax�AtBI-GFP3) were incu-bated with 5 �M DEX in distilled water at 23�C. Bax�AtBI-GFP1 toBax�AtBI-GFP3 are independent lines. The electrical conductivity ofthe solution was measured with a conductivity meter and is indicated asa relative value. n � 5.
The
Pla
nt C
ell
4 of 12 The Plant Cell
cells treated with 3 mM H
2
O
2
and 500 or 750
�
M SA. We alsoanalyzed AtBI-1 mRNA accumulation in NahG transgenicplants, in which the SA signaling pathway is downregulated(Delaney et al., 1994). RNA gel blot analysis indicated that AtBI-1mRNA was reduced by 50% in NahG transgenic plants com-pared with wild-type cells (Figure 5B).
The C-Terminal 14 Amino Acids of AtBI-1 Are Essential for the Suppression of Bax-Induced Death
The AtBI-1 protein is estimated to have seven transmembranedomains (Xu and Reed, 1998; Kawai et al., 1999) and is associ-
ated with ER membranes (Kawai-Yamada et al., 2001). Bolducet al. (2003) demonstrated that the C-terminal region of
Bras-sica napus
BI-1 (BnBI-1) is located on the cytosolic side of theER membrane.
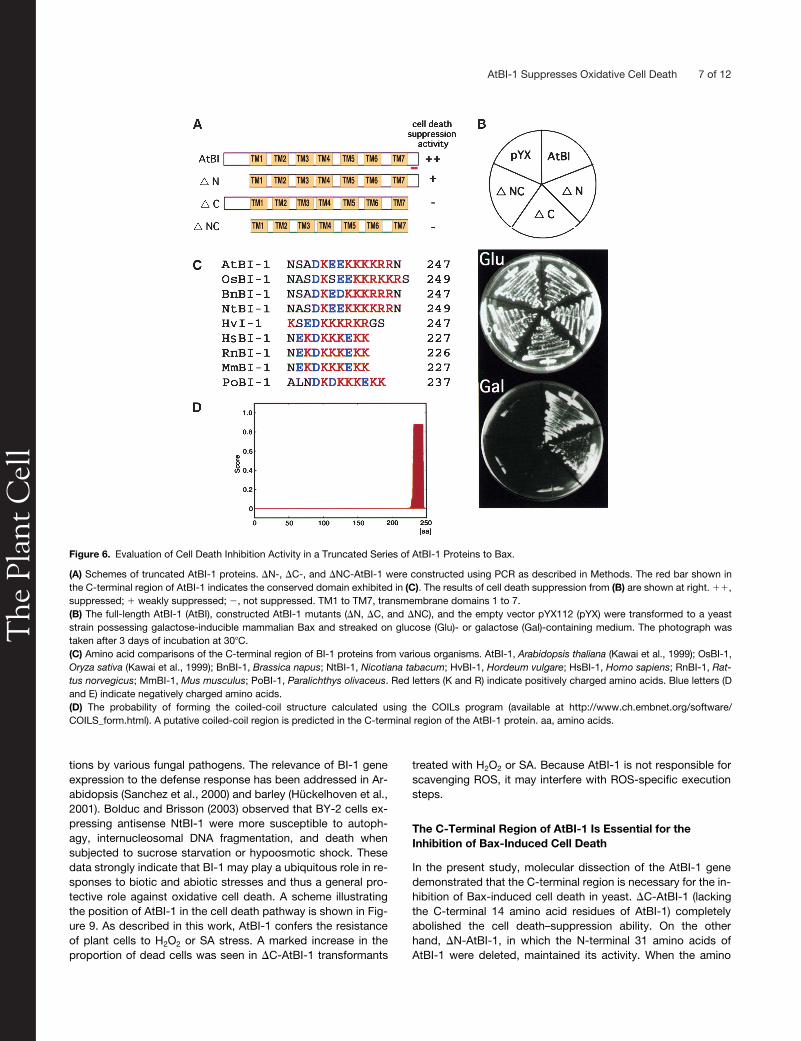
To determine the functional domains of AtBI-1 essential forthe death-suppressor activity,
�
N (amino acids 1 to 31 de-leted),
�
C (amino acids 234 to 247 deleted), and
�
NC (aminoacids 1 to 31 and 234 to 247 deleted) AtBI-1 proteins were con-structed and expressed in yeast possessing galactose-induc-ible mammalian Bax (Figures 6A and 6B). The AtBI-1 proteinlacking the N-terminal domain (
�
N) inhibited Bax-induced celldeath in yeast, whereas mutants with the C-terminal region de-
Figure 3. Inhibition of H2O2-induced Cell Death by AtBI-1.
(A) Induction of cell death by exogenously supplied H2O2 in tobacco BY-2 suspension cells. Evans blue treatment was performed at 0, 2, 6, 10, and20 h after the addition of 3 mM H2O2, and dead cells showing shrunken blue cytoplasm were scored with a light microscope. Data shown aremeans � SE of three experiments.(B) Comparison of AtBI-1 (AtBI-GFP2) and control (GFP1) cells stained with Evans blue at 18 h after treatment with 3 mM H2O2.(C) Effects of AtBI-1 or �C-AtBI-1 on H2O2-induced cell death. Tobacco BY-2 cells expressing GFP, AtBI-GFP, or �C-AtBI-GFP were incubated withH2O2 (0 to 4 mM) for 18 h. Dead cells were scored, and the results are expressed as means � SD of at least three experiments. WT, wild type.(D) Turnover of H2O2 after its addition to transgenic cell lines expressing GFP, AtBI-GFP, or �C-AtBI-GFP. One millimolar H2O2 was added to each cellline, and the H2O2 concentration of the medium was measured after 5 min. Data shown are means � SE of three experiments. gfw, grams fresh weight.
The
Pla
nt C
ell
AtBI-1 Suppresses Oxidative Cell Death 5 of 12
leted (�C) or with both the N and C termini deleted (�NC) didnot inhibit Bax effects. In this regard, expression of the C-termi-nal domain alone could not inhibit Bax-induced cell death (datanot shown). As shown in Figure 6C, the C-terminal region ofBI-1 proteins from various organisms consists of clusters ofcharged amino acids (D and E, which are negatively charged,and K and R, which are positively charged), and this feature isevolutionarily conserved in plants and animals. Furthermore,such a region was estimated to form a coiled-coil structure(Figure 6D).
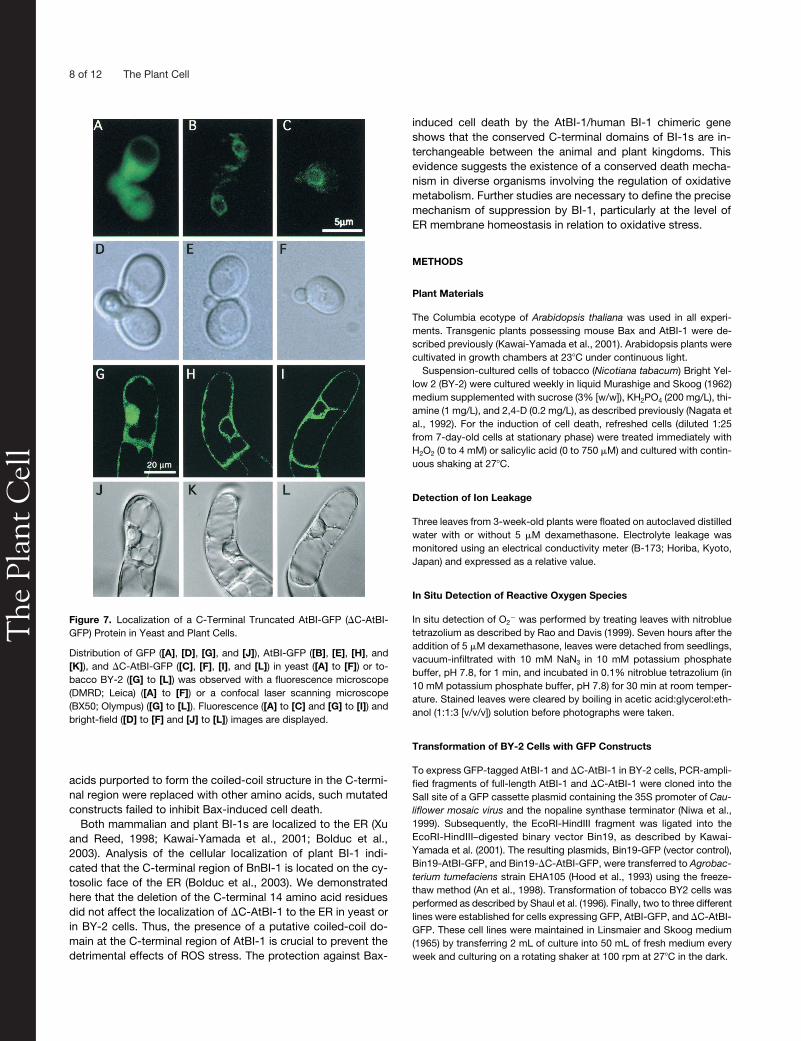
As mentioned above, plant and animal BI-1 proteins are lo-cated mostly in the ER and the perinuclear region (Xu andReed, 1998; Kawai-Yamada et al., 2001; Bolduc et al., 2003).To determine the significance of the C-terminal region of AtBI-1in terms of cellular localization, a plasmid containing GFP-tagged �C-AtBI-1 (�C-AtBI-GFP) was prepared and expressedin yeast and tobacco suspension cells. As shown in Figure 7,the fluorescent images of AtBI-GFP and �C-AtBI-GFP in yeast(Figures 7B, 7C, 7E, and 7F) and in BY-2 cells (Figures 7H, 7I,7K, and 7L) revealed a perinuclear pattern consistent with ERlocalization. The control expressing only GFP (Figures 7A, 7D,7G, and 7J) showed cytosolic localization. These findings indi-cate that the absence of the C-terminal domain did not affectthe cellular localization of AtBI-1.
To further investigate the role of the C-terminal region in celldeath–suppression activity, site-directed mutagenesis was per-formed (Figures 8A and 8B). As mentioned above, the C-termi-nal part of AtBI-1 consists of charged amino acids and exhibitsa coiled-coil structure. To dissect the importance of such char-
acteristics, amino acids located in this domain were replacedwith negatively charged (E in AtBI28) or noncharged (Q and N inAtBI29 and AtBI32) amino acids. Furthermore, to reduce thepossibility of a coiled-coil structure, K and R were replacedwith I and G, respectively, in AtBI30. When such plasmids weretransformed into a yeast strain harboring galactose-inducibleBax, only AtBI30 lost the activity. These results suggest that thecoiled-coil structure located in the C-terminal region of AtBI-1 isessential for the suppression of Bax-induced death in yeast. Onthe other hand, a chimeric gene in which the 14–amino acid Cterminus of AtBI-1 was replaced with the 9–amino acid C termi-nus of human BI-1 (AtBI33 in Figure 8) was as effective as thefull-length AtBI-1 in inhibiting Bax-induced death in yeast.
DISCUSSION
AtBI-1 Suppresses Bax-Induced Cell Death without Downregulating ROS Production
It has been reported that mammalian Bax might activate an en-dogenous cell death program in plants that mimics the HR in-duced by Tobacco mosaic virus in tobacco plants carrying the Ngene (Lacomme and Cruz, 1999). The ability of Bax to induce celldeath in plants suggests the existence of common features inanimal and plant cell death processes. One of the earliest eventsin the HR is a burst of oxidative metabolism leading to the gener-ation of O2
� and the subsequent accumulation of H2O2 (Lamband Dixon, 1997). These are mediators for the signal network fordefense gene induction and consequent cell death.
Figure 4. Inhibition of SA-Induced Cell Death by AtBI-1.
Tobacco BY-2 cells expressing GFP, AtBI-GFP, or �C-AtBI-GFP were treated with 500 �M (A) or 750 �M (B) SA and then cultured for 18 h before ex-amination with Evans blue staining. Dead cells stained with Evans blue were scored, and values are expressed as percentages. Data shown aremeans � SE of more than three replications. (C) shows microscopic images of cells at 18 h after 750 �M SA treatment. WT, wild type.
The
Pla
nt C
ell
6 of 12 The Plant Cell
In the animal system, the mitochondria are involved directlyin the generation of ROS and the activation of Bax-induced celldeath processes via the release of cytochrome c (Green andReed, 1998). Similarly, Bax targets mitochondria in yeast (Zhaet al., 1996) and induces the release of cytochrome c (Manon etal., 1997) and the generation of ROS (Madeo et al., 1999), sug-gesting that a toxic effect related to oxidative metabolism is in-volved. The C-terminal transmembrane domain is known to berequired for Bax targeting of the mitochondria in yeast. Simi-larly, it was demonstrated that the C-terminal transmembranedomain of Bax is needed to induce cell death in tobacco(Lacomme and Cruz, 1999) and Arabidopsis (Kawai-Yamada etal., 2001).
In this study, a NBT assay was performed to determinewhether Bax-induced plant death is accompanied by oxidativestress. Our results showed that the expression of mammalian
Bax could lead to enhanced ROS generation, which appeared7 h after Bax expression. Ion leakage indicative of the loss ofmembrane permeability was observed at 20 to 30 h after Baxexpression. Thus, ROS accumulation preceded the destructionof the plasma membrane in Bax-induced plant cell death.
We reported previously that the expression of Bax in Arabi-dopsis plants resulted in apoptotic cell death, which could besuppressed by the coexpression of AtBI-1 (Kawai-Yamada etal., 2001). Thus, we used a similar genetic system to evaluatethe functional significance of AtBI-1 with respect to oxidativecell death. Our results indicated that AtBI-1 did not affect theO2
� production caused by Bax. Furthermore, when tobaccoBY-2 cells were treated with 1 mM H2O2, its decomposition didnot change in control and AtBI-1–overexpressing lines, indicat-ing that AtBI-1 was not responsible for scavenging the oxidant.Thus, it is reasonable to assume that AtBI-1 suppresses Bax-induced cell death downstream of ROS generation.
AtBI-1 Downregulates the Cellular Defect Caused by Oxidative Stress in Plants
Plants continuously produce ROS such as O2�, H2O2, and sin-
gle oxygen as products of normal cellular metabolism. Conse-quently, plants rapidly metabolize these ROS with the help ofantioxidant enzymes or metabolites. However, once plants aresubjected to stresses such as high light, pathogens, ozone (O3),and UV irradiation, excess ROS is generated, which uponreaching a threshold level triggers PCD (Hernandez et al., 1993;Prasad et al., 1994; Chamnongpol et al., 1996; Lamb andDixon, 1997). H2O2 plays an important role in signal transduc-tion, cell wall reinforcement, HR, and phytoalexin production(Brisson et al., 1994; Dixon et al., 1994), and it has been dem-onstrated that exogenous H2O2 can induce a typical PCD in to-bacco BY-2 cells (Houot et al., 2001).
Treatment of plant cells with SA is believed to enhance H2O2
levels (Rao and Davis, 1999). SA plays an important role in in-fluencing the oxidative burst, and the defense responses andsubsequent cell death ultimately lead to the development of thesystemic acquired resistance and HR (Draper, 1997; Shirasu etal., 1997). Arabidopsis expressing the bacterial enzyme salicy-late hydroxylase (NahG) does not accumulate SA where Rgene–mediated HR is not observed (Delaney et al., 1994). Fur-thermore, NahG plants do not exhibit a HR-like response to O3
(Rao and Davis, 1999), and virus-induced HR is replaced byspreading necrotic lesions in NahG tobacco (Chivasa and Carr,1998). On the other hand, increased production of H2O2 en-hances the activities of enzymes involved in the biosynthesis ofSA (Leon et al., 1995; Summermatter et al., 1995). Subse-quently, increased SA levels promote H2O2 production, andhigh levels of free radicals are generated (Kauss et al., 1994;Fauth et al., 1996).
Concerning the effects of fungal elicitors on cell death, ricecells expressing AtBI-1 were found to downregulate the HRinduced by the Magnaporthe grisea elicitor (Matsumura etal., 2003). Interestingly, AtBI-1 accumulation was delayed inthe Arabidopsis coi1 mutant compared with wild-type plants(Sanchez et al., 2000). Reduced AtBI-1 levels might contributeto the enhanced susceptibility shown by coi1 plants to infec-
Figure 5. Accumulation of AtBI-1 mRNA in Arabidopsis Cells under Ox-idative Stress or in NahG Transgenic Plants.
(A) Arabidopsis suspension cells (3 days old) were treated with H2O2 (1and 3 mM) or SA (500 and 750 �M) for 24 h under continuous shaking.Equal amounts of total RNAs (10 �g) were electrophoresed on a 1.2%formaldehyde agarose gel and transferred to a membrane. An -32P-dCTP–labeled 3� untranslated region of AtBI-1 cDNA was used as aprobe.(B) Analysis of AtBI-1 mRNA accumulation in NahG transgenic plants.Total RNAs (20 �g) from 2-week-old wild-type (WT) and NahG trans-genic plants were electrophoresed and hybridized.The middle panels in (A) and (B) show gels stained with ethidium bro-mide for rRNA. The bottom panels show relative levels of AtBI-1 expres-sion in Arabidopsis suspension cells treated with H2O2 and SA (A) andNahG transgenic plants (B). Quantitative measurements of RNA gelblots were performed using the BAS1500 imaging plate scanner.
The
Pla
nt C
ell
AtBI-1 Suppresses Oxidative Cell Death 7 of 12
tions by various fungal pathogens. The relevance of BI-1 geneexpression to the defense response has been addressed in Ar-abidopsis (Sanchez et al., 2000) and barley (Hückelhoven et al.,2001). Bolduc and Brisson (2003) observed that BY-2 cells ex-pressing antisense NtBI-1 were more susceptible to autoph-agy, internucleosomal DNA fragmentation, and death whensubjected to sucrose starvation or hypoosmotic shock. Thesedata strongly indicate that BI-1 may play a ubiquitous role in re-sponses to biotic and abiotic stresses and thus a general pro-tective role against oxidative cell death. A scheme illustratingthe position of AtBI-1 in the cell death pathway is shown in Fig-ure 9. As described in this work, AtBI-1 confers the resistanceof plant cells to H2O2 or SA stress. A marked increase in theproportion of dead cells was seen in �C-AtBI-1 transformants
treated with H2O2 or SA. Because AtBI-1 is not responsible forscavenging ROS, it may interfere with ROS-specific executionsteps.
The C-Terminal Region of AtBI-1 Is Essential for the Inhibition of Bax-Induced Cell Death
In the present study, molecular dissection of the AtBI-1 genedemonstrated that the C-terminal region is necessary for the in-hibition of Bax-induced cell death in yeast. �C-AtBI-1 (lackingthe C-terminal 14 amino acid residues of AtBI-1) completelyabolished the cell death–suppression ability. On the otherhand, �N-AtBI-1, in which the N-terminal 31 amino acids ofAtBI-1 were deleted, maintained its activity. When the amino
Figure 6. Evaluation of Cell Death Inhibition Activity in a Truncated Series of AtBI-1 Proteins to Bax.
(A) Schemes of truncated AtBI-1 proteins. �N-, �C-, and �NC-AtBI-1 were constructed using PCR as described in Methods. The red bar shown inthe C-terminal region of AtBI-1 indicates the conserved domain exhibited in (C). The results of cell death suppression from (B) are shown at right. ��,suppressed; � weakly suppressed; �, not suppressed. TM1 to TM7, transmembrane domains 1 to 7.(B) The full-length AtBI-1 (AtBI), constructed AtBI-1 mutants (�N, �C, and �NC), and the empty vector pYX112 (pYX) were transformed to a yeaststrain possessing galactose-inducible mammalian Bax and streaked on glucose (Glu)- or galactose (Gal)-containing medium. The photograph wastaken after 3 days of incubation at 30�C.(C) Amino acid comparisons of the C-terminal region of BI-1 proteins from various organisms. AtBI-1, Arabidopsis thaliana (Kawai et al., 1999); OsBI-1,Oryza sativa (Kawai et al., 1999); BnBI-1, Brassica napus; NtBI-1, Nicotiana tabacum; HvBI-1, Hordeum vulgare; HsBI-1, Homo sapiens; RnBI-1, Rat-tus norvegicus; MmBI-1, Mus musculus; PoBI-1, Paralichthys olivaceus. Red letters (K and R) indicate positively charged amino acids. Blue letters (Dand E) indicate negatively charged amino acids.(D) The probability of forming the coiled-coil structure calculated using the COILs program (available at http://www.ch.embnet.org/software/COILS_form.html). A putative coiled-coil region is predicted in the C-terminal region of the AtBI-1 protein. aa, amino acids.
The
Pla
nt C
ell
8 of 12 The Plant Cell
acids purported to form the coiled-coil structure in the C-termi-nal region were replaced with other amino acids, such mutatedconstructs failed to inhibit Bax-induced cell death.
Both mammalian and plant BI-1s are localized to the ER (Xuand Reed, 1998; Kawai-Yamada et al., 2001; Bolduc et al.,2003). Analysis of the cellular localization of plant BI-1 indi-cated that the C-terminal region of BnBI-1 is located on the cy-tosolic face of the ER (Bolduc et al., 2003). We demonstratedhere that the deletion of the C-terminal 14 amino acid residuesdid not affect the localization of �C-AtBI-1 to the ER in yeast orin BY-2 cells. Thus, the presence of a putative coiled-coil do-main at the C-terminal region of AtBI-1 is crucial to prevent thedetrimental effects of ROS stress. The protection against Bax-
induced cell death by the AtBI-1/human BI-1 chimeric geneshows that the conserved C-terminal domains of BI-1s are in-terchangeable between the animal and plant kingdoms. Thisevidence suggests the existence of a conserved death mecha-nism in diverse organisms involving the regulation of oxidativemetabolism. Further studies are necessary to define the precisemechanism of suppression by BI-1, particularly at the level ofER membrane homeostasis in relation to oxidative stress.
METHODS
Plant Materials
The Columbia ecotype of Arabidopsis thaliana was used in all experi-ments. Transgenic plants possessing mouse Bax and AtBI-1 were de-scribed previously (Kawai-Yamada et al., 2001). Arabidopsis plants werecultivated in growth chambers at 23�C under continuous light.
Suspension-cultured cells of tobacco (Nicotiana tabacum) Bright Yel-low 2 (BY-2) were cultured weekly in liquid Murashige and Skoog (1962)medium supplemented with sucrose (3% [w/w]), KH2PO4 (200 mg/L), thi-amine (1 mg/L), and 2,4-D (0.2 mg/L), as described previously (Nagata etal., 1992). For the induction of cell death, refreshed cells (diluted 1:25from 7-day-old cells at stationary phase) were treated immediately withH2O2 (0 to 4 mM) or salicylic acid (0 to 750 �M) and cultured with contin-uous shaking at 27�C.
Detection of Ion Leakage
Three leaves from 3-week-old plants were floated on autoclaved distilledwater with or without 5 �M dexamethasone. Electrolyte leakage wasmonitored using an electrical conductivity meter (B-173; Horiba, Kyoto,Japan) and expressed as a relative value.
In Situ Detection of Reactive Oxygen Species
In situ detection of O2� was performed by treating leaves with nitroblue
tetrazolium as described by Rao and Davis (1999). Seven hours after theaddition of 5 �M dexamethasone, leaves were detached from seedlings,vacuum-infiltrated with 10 mM NaN3 in 10 mM potassium phosphatebuffer, pH 7.8, for 1 min, and incubated in 0.1% nitroblue tetrazolium (in10 mM potassium phosphate buffer, pH 7.8) for 30 min at room temper-ature. Stained leaves were cleared by boiling in acetic acid:glycerol:eth-anol (1:1:3 [v/v/v]) solution before photographs were taken.
Transformation of BY-2 Cells with GFP Constructs
To express GFP-tagged AtBI-1 and �C-AtBI-1 in BY-2 cells, PCR-ampli-fied fragments of full-length AtBI-1 and �C-AtBI-1 were cloned into theSalI site of a GFP cassette plasmid containing the 35S promoter of Cau-liflower mosaic virus and the nopaline synthase terminator (Niwa et al.,1999). Subsequently, the EcoRI-HindIII fragment was ligated into theEcoRI-HindIII–digested binary vector Bin19, as described by Kawai-Yamada et al. (2001). The resulting plasmids, Bin19-GFP (vector control),Bin19-AtBI-GFP, and Bin19-�C-AtBI-GFP, were transferred to Agrobac-terium tumefaciens strain EHA105 (Hood et al., 1993) using the freeze-thaw method (An et al., 1998). Transformation of tobacco BY2 cells wasperformed as described by Shaul et al. (1996). Finally, two to three differentlines were established for cells expressing GFP, AtBI-GFP, and �C-AtBI-GFP. These cell lines were maintained in Linsmaier and Skoog medium(1965) by transferring 2 mL of culture into 50 mL of fresh medium everyweek and culturing on a rotating shaker at 100 rpm at 27�C in the dark.
Figure 7. Localization of a C-Terminal Truncated AtBI-GFP (�C-AtBI-GFP) Protein in Yeast and Plant Cells.
Distribution of GFP ([A], [D], [G], and [J]), AtBI-GFP ([B], [E], [H], and[K]), and �C-AtBI-GFP ([C], [F], [I], and [L]) in yeast ([A] to [F]) or to-bacco BY-2 ([G] to [L]) was observed with a fluorescence microscope(DMRD; Leica) ([A] to [F]) or a confocal laser scanning microscope(BX50; Olympus) ([G] to [L]). Fluorescence ([A] to [C] and [G] to [I]) andbright-field ([D] to [F] and [J] to [L]) images are displayed.

The
Pla
nt C
ell
AtBI-1 Suppresses Oxidative Cell Death 9 of 12
Antioxidant Activity
The antioxidant activity of BY-2 suspension cells was assayed by mea-suring the decay of 1 mM H2O2 with time as described by Tiwari et al.(2002). The media of cells treated for various time intervals (0, 3, 5, and10 min) with 1 mM H2O2 were mixed with 0.1 mL of titanium sulfate andincubated for 15 min at room temperature. The oxidation of titanium sul-fate was recorded by reading OD410. These values were converted tocorresponding concentrations using a standard calibration plot of knownconcentrations of H2O2.
RNA Gel Blot Analysis
Total RNAs were isolated from H2O2– or salicylic acid–treated Arabidop-sis suspension cells using guanidinium thiocyanate as described previ-ously (Kawai et al., 1999). Total RNAs, electrophoresed on a denaturing
1.2% agarose gel, were transferred to a nylon membrane (Biodyne B;Pall, Port Washington, NY), which then was hybridized to the -32P-labeled 3� untranslated region fragment of AtBI-1 (Kawai et al., 1999) in10% dextran sulfate, 1 M NaCl, 1% SDS, and 100 �g/mL heat-dena-tured salmon sperm DNA. Washing was performed at high stringency(0.1� SSC [1� SSC is 0.15 M NaCl and 0.015 M sodium citrate] and0.1% SDS at 65�C) . The membrane was analyzed with a BAS1500 im-aging plate scanner (Fuji Film, Tokyo, Japan).
Yeast Strains and Expression
Yeast strains and plasmids used in this study have been described pre-viously (Kawai et al., 1999; Kawai-Yamada et al., 2001). Truncated AtBI-1genes (�N, amino acids 1 to 31 deleted; �C, amino acids 234 to 247deleted; and �NC, amino acids 1 to 31 and 234 to 247 deleted) taggedwith EcoRI sites by PCR were ligated to the EcoRI site of the expression
Figure 8. Comparison of the Cell Death–Suppression Activity of C-Terminal Mutants of AtBI-1.
(A) Various mutations of the C terminus of AtBI-1 were constructed, and their ability to suppress Bax-induced cell death was analyzed. Mutated seriesof AtBI-1 (AtBI28 to AtBI33) were constructed using PCR as described in Methods. The amino acids replaced are underlined. The estimated charges(�, �, or none) of the replaced regions and calculated scores for the coiled-coil structure also are indicated. In AtBI33, the C-terminal 14 amino acidswere replaced with the 9 amino acids of human BI-1.(B) Spot assay of cell death–suppression activity in yeast. Yeast transformed with plasmids containing mutagenized AtBI-1 genes were cultured in SDmedium containing glucose. After 1 day of shaking, the OD600 of each culture was adjusted to 0.2 and diluted to 0.0002. Each dilution was spotted onSD medium containing glucose (Glu) or galactose (Gal). The results show the growth of each line after 2 days (Glu) and 4 days (Gal) of incubation at30�C. The results of cell death suppression are shown at right: �, suppressed; �, weak suppression or not suppressed.

The
Pla
nt C
ell
10 of 12 The Plant Cell
vector pYX112, possessing a 2-�m replicon. The Saccharomyces cere-visiae strain QX95001, which is BF264-15au (MAT ade1 his2-3, 112trp1-1a ura3) (Xu and Reed, 1998) containing the LEU2-marked mouseBax-encoding plasmid Yep51-Bax, was transformed with the plasmidspYX-AtBI, pYX-�N, pYX-�C, and pYX-�NC using the lithium acetatemethod. Ura�Leu� transformants were streaked on either a syntheticdropout (SD)–glucose (2%) plate or a SD-galactose (5%) plate and incu-bated at 30�C.
For site-directed mutagenesis of the C-terminal region of AtBI-1, thefollowing oligonucleotide primers were used for the amplification of mu-tated fragments by PCR: 27, 5�-AAGCTTGTTTCTCCTTCTCCTCCTCCT-CTCTTC-3�; 28, 5�-AAGCTTGTTTTCCTCTTCCTCCTCCTCCTCTTC-3�;29, 5�-AAGCTTGTTCTTCTTTTGCTGCTGCTGCTCTTC-3�; 30, 5�-AAG-CTTGTTTCCCCCTATAATGATAATCTCTTC-3�; 31, 5�-AAGCTTGTTTCT-CCTTTTCTTCTTCTTCTTCTTTTTCTT-3�; 32, 5�-AAGCTTGTTTCTCCT-TTTCTTCTTCTTCTGTTGGTTATT-3�; and 33, 5�-AAGCTTTTTCTTCTC-TTTCTTCTTATCCTTTTCATTCTTCAACATTATGATGAGAATCCG-3�. Mu-tagenized fragments amplified by PCR were cloned into a pGEM T-Easyvector (Promega, Madison, WI) for sequencing. After digestion withEcoRI and HindIII, each fragment was cloned into the EcoRI-HindIII siteof the pYX112 vector.
In the spot assay, yeast culture was diluted to various concentrations(OD600 � 0.2, 0.02, 0.002, and 0.0002), and an aliquot (5 �L) from eachdilution was spotted onto SD-glucose and SD-galactose medium and in-cubated for 2 days (glucose medium) or 4 days (galactose medium) at30�C.
For the analysis of the cellular localization of �C-AtBI protein in yeast,a chimera gene of AtBI-1 and GFP was constructed in the cassette vec-tor pTS910 (provided by Y. Kikuchi, University of Tokyo, Japan). TheDNA fragment tagged with the EcoRI-HindIII site by PCR was ligated
into the EcoRI-HindIII–digested yeast expression vector pYX112. Theplasmids pYX112-GFP and pYX112-AtBI-GFP were described in theprevious report (Kawai-Yamada et al., 2001).
Cytological Methods
GFP fluorescence was examined at a 488-nm excitation wavelength witha fluorescence microscope (DMRD; Leica, Wetzlar, Germany) or with aconfocal laser scanning microscope (BX50; Olympus, Tokyo, Japan).The viability of cells was measured with the addition of Evans blue(0.05%; Nakalai, Kyoto, Japan), which only penetrates dead cells and re-sults in blue staining of the cellular contents (Kawai et al., 1998; Kawaiand Uchimiya, 2000). The percentage of dead cells in each treatmentwas determined by scoring several hundred cells with a microscope.
Upon request, materials integral to the findings presented in this pub-lication will be made available in a timely manner to all investigators onsimilar terms for noncommercial research purposes. To obtain materials,please contact Hirofumi Uchimiya, [email protected].
Accession Numbers
The accession numbers for the sequences shown in Figure 6 are as fol-lows: AtBI-1, AB025927; OsBI-1, AB025926; BnBI-1, AF390555; NtBI-1,AF390556; HvBI-1, AJ290421; HsBI-1, AF033095; RnBI-1, X75855;MmBI-1, BC005588; and PoBI-1, AF220548.
ACKNOWLEDGMENTS
We thank J. Reed, N.H. Chua, Y. Kikuchi, Y. Niwa, and A. Hasezawa fortheir help and gifts of materials. We thank H. Ogawa-Suzuki for her helpin all experiments. We thank Louise Brisson, Patrick Gallois, and ChrisHawes for editing the manuscript. This research was supported by a Re-search for the Future grant from the Japan Society for the Promotion ofScience.
Received June 12, 2003; accepted October 1, 2003.
REFERENCES
Abramovitch, R.B., Kim, Y.J., Chen, S., Dickman, M.B., and Martin,G.B. (2003). Pseudomonas type III effecter AvrPtoB induces plantdisease susceptibility by inhibition of host programmed cell death.EMBO J. 22, 60–69.
An, G., Ebert, R.R., Mitra, A., and Ha, S.B. (1998). Binary vectors. InPlant Molecular Biology Manual, S.B. Gelvin and R.A. Schilperoort,eds (Dordrecht, The Netherlands: Kluwer Academic Publishers), pp.1–19.
Bischoff, J.R., Casso, D., and Beach, D. (1992). Human p53 inhibitsgrowth in Schizosaccharomyces pombe. Mol. Cell. Biol. 12, 1405–1411.
Brisson, L.F., Tenhanken, R., and Kamb, C. (1994). Function of oxida-tive cross-linking of cell wall structural proteins in plant disease resis-tance. Plant Cell 6, 1703–1712.
Bolduc, N., and Brisson, L.F. (2003). Antisense down regulation ofNtBI-1 in tobacco BY-2 cells induces accelerated cell death uponcarbon starvation. FEBS Lett. 532, 111–114.
Bolduc, N., Ouellet, M., Pitre, F., and Brisson, L.F. (2003). Molecularcharacterization of two plant BI-1 homologues which suppress Bax-induced apoptosis in human 293 cells. Planta 216, 377–386.
Chamnongpol, S., Willekens, H., Langebartels, C., van Montagu, M.,Inze, D., and van Camp, W. (1996). Transgenic tobacco with a re-
Figure 9. Scheme of the Role of the BI-1 Protein in the ROS-InducedCell Death Pathway.
ROS cause mitochondrial permeability transition, ATP depletion, releaseof cytochrome c from the mitochondrial membranes, and lipid peroxida-tion, which cause ion leakage and cell death (Rao and Davis, 1999;Houot et al., 2001; Maxwell et al., 2002; Tiwari et al., 2002). BI-1 pro-teins, as components of the ER membrane, are involved in counteract-ing “oxidative stress.”

The
Pla
nt C
ell
AtBI-1 Suppresses Oxidative Cell Death 11 of 12
duced catalase activity develops necrotic lesions and induces patho-genesis-related expression under high light. Plant J., 10, 491–503.
Chivasa, S., and Carr, J.P. (1998). Cyanide restores N gene–mediatedresistance to tobacco mosaic virus in transgenic tobacco expressingsalicylic hydroxylase. Plant Cell 10, 1489–1498.
Delaney, T.P., Uknes, S., Vernooji, B., Friedrich, L., Weymann, K.,Negrotto, D., Gaffne, T., Gu-Rella, M., Kessmann, H., Ward, E.,and Ryals, J. (1994). A central role of salicylic acid in plant diseaseresistance. Science 266, 1247–1250.
Desikan, R., Reynolds, A., Hancock, J.T., and Neill, S.J. (1998).Harpin and hydrogen peroxide both initiate programmed cell deathbut have differential effects on defense gene expression in Arabidop-sis suspension cultures. Biochem. J. 330, 115–120.
Dixon, R.A., Harrison, M.J., and Lamb, C.J. (1994). Early events in theactivation of plant defense responses. Annu. Rev. Phytopathol. 32,479–501.
Draper, J. (1997). Salicylate, superoxide synthesis and cell suicide inplant defense. Trends Plant Sci. 2, 162–165.
Fauth, M., Merten, A., Hahn, M.G., Jeblick, E., and Kauss, H. (1996).Competence for elicitation of hydrogen peroxide in hypocotyls of cu-cumber is induced by breaching the cuticle and is enhanced by sali-cylic acid. Plant Physiol. 110, 347–354.
Green, D.R., and Reed, J.C. (1998). Mitochondria and apoptosis. Sci-ence 281, 1309–1312.
Greenhalf, W., Steohan, C., and Chaudhuri, B. (1996). Role of mitochon-dria and C-terminal membrane anchor of Bcl-2 in Bax induced growth ar-rest and mortality in Saccharomyces cerevisiae. FEBS Lett. 380, 169–175.
Hanada, M., Aimé-Sempé, C., Sato, T., and Reed, J.C. (1995). Struc-ture-function analysis of Bcl-2 protein: Identification of conserved do-mains important for homodimerization with Bcl-2 and heterodimeriza-tion with Bax. J. Biol. Chem. 270, 11962–11968.
Hengartner, M., and Horvitz, H.R. (1994). C. elegans cell survival geneced-9 encodes a functional homologue of the mammalian proto-oncogene bcl-2. Cell 76, 665–676.
Hernandez, J.A., Corpas, F.J., Gomez, M., Del Rio, L.A., and Sevilla,F. (1993). Salt-induced oxidative stress mediated by activated oxygenspecies in pea leaf mitochondria. Physiol. Plant. 89, 103–110.
Hood, E.E., Gelvin, S.B., Melchers, L.S., and Hoekema, A. (1993).New Agrobacterium vector for plant transformation. Transgenic Res.2, 208–218.
Houot, V., Etienne, P., Petitot, A.S., Barbier, S., Blein, J.P., and Suty,L. (2001). Hydrogen peroxide induces programmed cell death fea-tures in cultured tobacco BY-2 cells, in a dose-dependent manner. J.Exp. Bot. 52, 1721–1730.
Hückelhoven, R., Dechert, C., and Kogel, K.H. (2003). Overexpressionof barley BAX inhibitor 1 induces breakdown of Mlo-mediated pene-tration resistance to Blumeria graminis. Proc. Natl. Acad. Sci. USA100, 5550–5560.
Hückelhoven, R., Dechert, C., Trujillo, M., and Kogel, K.H. (2001).Differential expression of putative cell death regulator genes in near-isogenic, resistant and susceptible barley lines during interaction withthe powdery mildew fungus. Plant Mol. Biol. 47, 739–748.
Kauss, H., Jeblick, W., Zielger, J., and Kraber, W. (1994). Pretreat-ment of parsley (Petroselinum crispum L.) suspension culture withmethyl jasmonate enhances the elicitation of active oxygen species.Plant Physiol. 105, 89–94.
Kawai, M., Pan, L., Reed, J.C., and Uchimiya, H. (1999). Evolutionallyconserved plant homologue of the Bax inhibitor-1 (BI-1) gene capable ofsuppressing Bax-induced cell death in yeast. FEBS Lett. 464, 143–147.
Kawai, M., Samarajeewa, P.K., Barrero, R.A., Nishiguchi, M., andUchimiya, H. (1998). Cellular dissection of the degradation pattern ofcortical cell death during aerenchyma formation of rice roots. Planta204, 277–287.
Kawai, M., and Uchimiya, H. (2000). Coleoptile senescence in rice(Oryza sativa L.). Ann. Bot. 86, 405–414.
Kawai-Yamada, M., Jin, L., Yoshinaga, K., Hirata, A., and Uchimiya,H. (2001). Mammalian Bax-induced plant cell death can be down-regulated by overexpression of Arabidopsis Bax Inhibitor-1 (AtBI-1).Proc. Natl. Acad. Sci. USA 98, 12295–12300.
Kroemer, G. (1997). The proto-oncogene Bcl-2 and its role in regulatingapoptosis. Nat. Med. 3, 614–620.
Lacomme, C., and Cruz, S.S. (1999). Bax-induced cell death in to-bacco is similar to the hypersensitive response. Proc. Natl. Acad. Sci.USA 96, 7956–7961.
Lam, E., Kato, N., and Lawton, M. (2001). Programmed cell death,mitochondria and the plant hypersensitive response. Nature 411,848–853.
Lamb, C., and Dixon, R.A. (1997). The oxidative burst in plant diseaseresistance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 48, 251–275.
Leon, J., Lawton, M.A., and Raskin, I. (1995). H2O2 stimulates salicylicacid biosynthesis in tobacco. Plant Physiol. 108, 1673–1678.
Levine, A., Tenhaken, R., Dixon, R., and Lamb, C. (1994). H2O2 fromthe oxidative burst orchestrates the plant hypersensitive disease re-sistance response. Cell 79, 583–593.
Linsmaier, E.M., and Skoog, F. (1965). Organic growth factor require-ments of tobacco tissue cultures. Phys. Plant. 18, 100–127.
Madeo, F., Fröhlich, E., Ligr, M., Grey, M., Sigrist, S.J., Wolf, D.H.,and Fröhlich, K.U. (1999). Oxygen stress: A regulator of apoptosis inyeast. J. Cell Biol. 145, 757–767.
Madeo, F., Herker, E., Maldener, C., Wissing, S., Lächelt, S., Herlan,M., Fehr, M., Lauber, K., Sigrist, S.J., Wesselborg, S., and Fröhlich,K.U. (2002). A caspase-related protease regulates apoptosis in yeast.Mol. Cell 9, 911–917.
Manon, S., Chaudhuri, B., and Bueerin, M. (1997). Release of cyto-chrome c and decrease of cytochrome c oxidase in Bax expressingyeast cells, and prevention of these effects by coexpression of Bcl-XL. FEBS Lett. 415, 29–32.
Matsumura, H., Nirasawa, S., Kiba, A., Urasaki, N., Saitoh, H., Ito,M., Kawai-Yamada, M., Uchimiya, H., and Terauchi, R. (2003).Overexpression of Bax inhibitor suppresses the fungal elicitor-inducedcell death in rice (Oryza sativa L.) cells. Plant J. 33, 425–434.
Maxwell, D.P., Nickels, R., and McIntosh, L. (2002). Evidence of mito-chondrial involvement in the transduction of signals required for theinduction of genes associated with pathogen attack and senescence.Plant J. 29, 269–279.
Mitsuhara, I., Malik, K.A., Miura, M., and Ohashi, Y. (1999). Animalcell-death suppressors Bcl-x(L) and Ced-9 inhibit cell death in to-bacco plants. Curr. Biol. 9, 775–778.
Mittler, R. (2002). Oxidative stress, antioxidants and stress tolerance.Trends Plant Sci. 7, 405–410.
Murashige, T., and Skoog, F. (1962). A revised medium for rapidgrowth and bioassays with tobacco tissue culture. Physiol. Plant. 15,473–497.
Nagata, T., Nemoto, Y., and Hasezawa, S. (1992). Tobacco BY2 cellline as the “HeLa” cell line in the cell biology of higher plants. Int. Rev.Cytol. 132, 1–30.
Niwa, Y., Hirano, T., Yoshimoto, K., Shimizu, M., and Kobayashi, H.(1999). Non-invasive quantitative detection and applications of non-toxic, S65T-type green fluorescent protein in living plants. Plant J. 18,455–463.
Prasad, T.K., Anderson, M.D., Martin, B.A., and Stewart, C.R. (1994).Evidence for chilling-induced oxidative stress in maize and a regula-tory role for hydrogen peroxide. Plant Cell 6, 65–74.
Rao, M.Y., and Davis, K.R. (1999). Ozone-induced cell death occursvia two distinct mechanisms in Arabidopsis: The role of salicylic acid.Plant J. 17, 603–614.

The
Pla
nt C
ell
12 of 12 The Plant Cell
Reed, J.C. (1994). Bcl2 and the regulation of programmed cell death. J.Cell Biol. 124, 1–6.
Rizhsky, L., Hallak-Herr, E., Van Breusegem, F., Rachmilevitch, S.,Barr, J.E., Rodermel, S., Inze, D., and Mittler, R. (2002). Double an-tisense plants lacking ascorbate peroxidase and catalase are lesssensitive to oxidative stress than single antisense plants lackingascorbate peroxidase or catalase. Plant J. 32, 329–342.
Sanchez, P., de Torres-Zabala, M., and Grant, M. (2000). AtBI-1, aplant homologue of Bax inhibitor-1, suppresses Bax-induced celldeath in yeast and is rapidly up regulated during wounding andpathogen challenge. Plant J. 21, 393–399.
Sato, R., Hanada, M., Bodrug, S., Irie, S., Iwama, N., Boise, L.H.,Thompson, C.B., Golernis, E., Fong, L., Wang, H.G., and Reed,J.C. (1994). Interactions among members of the bcl-2 protein familyanalyzed with a yeast two-hybrid system. Proc. Natl. Acad. Sci. USA91, 9238–9242.
Shaul, O., Nironov, V., Burssens, S., Montague, M.V., and Inze, D.(1996). Two Arabidopsis cyclin promoters mediate distinctive tran-scriptional oscillation in synchronized tobacco BY-2 cells. Proc. Natl.Acad. Sci. USA 93, 4868–4872.
Shirasu, K., Nakajima, H., Rajashekar, K., Dixon, R.A., and Lamb, C.(1997). Salicylic acid potentiates an agonist-dependent gain controlthat amplifies pathogen signals in the activation of defense mecha-nisms. Plant Cell 9, 261–270.
Solomon, M., Belenghi, B., Delledonne, M., and Levine, A. (1999).The involvement of cysteine proteases and protease inhibitor genesin programmed cell death in plants. Plant Cell 11, 431–444.
Summermatter, K., Sticher, L., and Metraux, J.P. (1995). Systemic re-sponse in Arabidopsis thaliana infected and challenged with Pseu-domonas syringae pv. syringae. Plant Physiol. 108, 1379–1385.
Tiwari, B.S., Belenghi, B., and Levine, A. (2002). Oxidative stress in-creased respiration and generation of reactive oxygen species, result-ing in ATP depletion, opening of mitochondrial permeability transition,and programmed cell death. Plant Physiol. 128, 1271–1281.
Varner, J.E., and Lin, L.S. (1989). Plant cell wall architecture. Cell 56,231–239.
Yu, L.H., Kawai-Yamada, M., Naito, M., Watanabe, K., Reed, J.C.,and Uchimiya, H. (2002). Induction of mammalian cell death by aplant Bax inhibitor. FEBS Lett. 512, 308–312.
Yuan, J.Y., Shaham, S., Ledoux, S., Ellis, H.M., and Horvitz, R.(1993). The C. elegans cell death gene ced-3 encodes a protein simi-lar to mammalian interleukin-1 converting enzyme. Cell 75, 641–652.
Xu, Q., and Reed, J.C. (1998). Bax inhibitor-1, a mammalian apoptosissuppressor identified by functional screening in yeast. Mol. Cell 1,337–346.
Zha, H., Fisk, H.A., Yaffe, M.P., Mahajan, N., Herman, B., and Reed,J.C. (1996). Structure-function comparison of the proapoptotic proteinBax in yeast and mammalian cells. Mol. Cell. Biol. 16, 6494–6508.

DOI 10.1105/tpc.014613; originally published online December 11, 2003;Plant Cell
Maki Kawai-Yamada, Yuri Ohori and Hirofumi UchimiyaAcid-Induced Cell Death
Dissection of Arabidopsis Bax Inhibitor-1 Suppressing Bax-, Hydrogen Peroxide-, and Salicylic
This information is current as of January 5, 2015
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists









![Robert Bax & Associates v Cavenham Pty Ltd [2012] QCA 177](https://img.dokumen.tips/doc/110x75/633b615ed575315e540495e5/robert-bax-associates-v-cavenham-pty-ltd-2012-qca-177.jpg)



















