Embed Size (px)
Citation preview

Changes in specific lipids regulate BAX-inducedmitochondrial permeability transitionE. Martınez-Abundis1, N. Garcıa1, F. Correa1, M. Franco2 and C. Zazueta1
1 Departamento de Bioquımica, Instituto Nacional de Cardiologıa Ignacio Chavez, Mexico
2 Departamento de Nefrologıa, Instituto Nacional de Cardiologıa Ignacio Chavez, Mexico
BAX is a pro-apoptotic member of the Bcl-2 protein
family that resides in an inactive state in the cytoplasm
of normal cells. Following an apoptotic stimulus, BAX
undergoes conformational changes [1–4], becoming a
mediator of the intrinsic phase of apoptosis by its
insertion into mitochondrial membranes, a process that
culminates in the release of cytochrome c and activa-
tion of effector caspases [5]. Evidence from the
literature indicates that BAX is activated by other
BH3-only proteins, particularly Bid protein and Bim
peptide [6,7]. In this respect, it has been shown that
Bid activation depends on its proteolytic processing
into tBid and the translocation of tBid to mitochon-
dria [8]. In the mitochondria, tBid may form oligomers
by itself and induce oligomerization of BAX and Bak
[9]. Although it remains unclear how tBid triggers such
oligomerization, there is strong evidence to suggest
that Bid-induced apoptosis depends on the presence of
BAX and Bak [10]. However, it has also been demon-
strated that tBid accumulation in mitochondria is an
essential, but not sufficient, event leading to mito-
chondrial disruption for cytochrome c leakage [6]. The
Keywords
BAX; cholesterol; gangliosides; lipid
microdomains; mitochondrial permeability
transition pore
Correspondence
C. Zazueta, Instituto Nacional de Cardiologıa,
I. Ch., Departamento de Bioquımica, Juan
Badiano No. 1, Colonia Seccion XVI,
Mexico 14080, D.F
Fax: +52 55 5573 0926
Tel: +52 55 5573 2911 (1465)
E-mail: [email protected]
(Received 13 August 2007, revised 17
October 2007, accepted 25 October 2007)
doi:10.1111/j.1742-4658.2007.06166.x
Recent evidence suggests the existence of lipid microdomains in mitochon-
dria, apparently coexisting as structural elements with some of the mito-
chondrial permeability transition pore-forming proteins and members of
the Bcl-2 family. The aim of this study was to investigate the relevance of
the main components of membrane microdomains (e.g. cholesterol and
sphingolipids) in activation of the mitochondrial permeability transition
pore (mPTP) by recombinant BAX (rBAX). For this purpose, we used
chemically modified renal cortex mitochondria and renal cortex mitochon-
dria from hypothyroid rats that show a modified mitochondrial lipid com-
position in vivo. Oligomeric rBAX induced an enhanced permeability
conformation in the mPTP of control mitochondria. rBAX failed to induce
mPTP opening when the cholesterol and ganglioside content of mitochon-
dria were modified with the chelator methyl-beta-cyclodextrin. Accordingly,
hypothyroid mitochondria, with endogenously lower cholesterol and gan-
glioside content, showed resistance to mPTP opening induced by rBAX.
These observations suggest that enriched cholesterol and ganglioside
domains in the mitochondrial membranes may determine BAX interaction
with the mPTP. An intriguing observation was that chemical extraction of
cholesterol and ganglioside in control mitochondria did not have an effect
on rBAX insertion. Conversely, in hypothyroid mitochondria, rBAX inser-
tion was diminished dramatically compared with control mitochondria.
The membrane and protein changes associated with thyroid status and
their possible role in rBAX docking into the membranes are discussed.
Abbreviations
ANT, adenine nucleotide translocator; CSA, cyclosporine A; GST, glutathione S-transferase; MbCD, methyl-beta-cyclodextrin; mPT,
mitochondrial permeability transition; mPTP, mitochondrial permeability transition pore; rBAX, recombinant BAX; VDAC, voltage-dependent
anion channel; Dw, transmembrane potential.
6500 FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS

molecular mechanism underlying such leakage is still a
matter of debate. One proposal is that oligomeric
BAX forms pores in the outer mitochondrial mem-
brane, providing a mechanism for cytochrome c release
[11]; a second proposal is that BAX induces opening
of the permeability transition pore (mPTP) [12]. The
opening of this mega-channel would lead to mitochon-
drial swelling and rupture of the outer membrane, thus
explaining the release of cytochrome c from mitochon-
dria. Although several reports have suggested an inter-
action between BAX and the components of the
mPTP, e.g. the adenine nucleotide translocator (ANT)
[13] and ⁄or the voltage-dependent anion channel
(VDAC) [14], the factors involved in BAX induction
of mitochondrial permeability transition (mPT) are
unknown.
By contrast, lipid microdomains or ‘lipid rafts’ were
first described in the plasma membrane. It is known
that they are composed mainly of sphingolipids and
cholesterol, and have been considered to be architec-
tural domains, where specific proteins could interact to
exert specific regulatory mechanisms [15].
Relevant to this issue, there is recent evidence sug-
gesting the existence of lipid microdomains in mito-
chondria which coexist as structural elements with
some of the mPTP-forming proteins [16]. Recent
reports indicate that the disialoganglioside (GD3) con-
tributes directly to the opening of the permeability
transition pore complex in isolated mitochondria
[17,18] and furthermore, that GD3-induced damage in
mitochondrial membranes is confined to sites that can
be restrained by Bcl-2 [19]. Also, several lines of evi-
dence converge on the assumption that the mPTP prop-
agates the downstream stage of apoptosis, mediated by
the lipid-dependent pathway, which includes ceramide
and GD3 [20]. In addition, cholesterol is known to
induce changes in the phospholipid packing of the lipid
bilayer and has been suggested as critical for micro-
domain formation [21]. Finally, Keller et al. [22]
reported that changing the nature of the sterol in cellu-
lar rafts leads to changes in the protein composition of
those domains. Our results suggest that cholesterol and
GD3 are relevant for BAX interaction with the mPTP.
These data provide evidence that specific lipids play a
key role in cross-talk between rBAX and the mPTP.
Results
rBAX-induced mitochondrial calcium release and
modified cytochrome content
The addition of oligomeric rBAX to isolated mito-
chondria induced the release of accumulated calcium
in a dose-dependent manner (Fig. 1A). Figure 1 also
shows that 1 lm carboxyatractyloside, a well-known
mPTP inducer, promoted immediate depletion of the
intramitochondrial calcium content. The recombinant
protein associated with mitochondrial membranes was
resistant to alkaline extraction, indicating that it was
deeply embedded in the bilayer (Fig. 1B). A clear cor-
respondence between cytochrome c release and calcium
extrusion from the mitochondria was observed by
inducing opening of the mPTP with either carboxya-
tractyloside or rBAX (Fig. 1C).
Cyclosporin A prevented cytochrome c release
and mPTP opening induced by rBAX
The immunosuppressant cyclosporin A (CSA) effec-
tively inhibits mPTP opening under almost all condi-
tions, and is therefore considered a marker for mPT.
To determine whether the altered mitochondrial per-
meability observed in the presence of rBAX in rat
kidney mitochondria was related to the opening of
this mega-channel, we measured the effect of CSA
on calcium release induced by the addition of recom-
binant protein (Fig. 2A). CSA inhibited calcium
release, although rBAX remained attached to the
membranes (Fig. 2B). Interestingly, mitochondria
incubated with rBAX only released cytochrome c
when the permeability transition pore was opened,
and this was abolished in a medium supplemented
with CSA (Fig. 2C). These results suggest active par-
ticipation of the mPTP on cytochrome c release
induced by rBAX.
The effect of rBAX on the mPT was entirely
dependent on calcium. Typical traces of mitochon-
drial swelling with and without calcium are shown in
Fig. 3A. An important decrease in light scattering
was detected when the medium was supplemented
with a final calcium concentration of 50 lm, and was
inhibited completely by 1 lm CSA. Also, pore open-
ing can be estimated by following the discharge
kinetics of the transmembrane potential (Dw). Fig-
ure 3B shows that the membrane potential developed
by mitochondria added to the medium without any
previous treatment (trace a) was similar to that devel-
oped in mitochondria incubated with rBAX in the
absence of calcium (trace b). Conversely, addition of
50 lm CaCl2 to the medium led to a decrease in the
Dw of mitochondria incubated with rBAX, which
eventually resulted in its total collapse (trace c). We
also determined mitochondrial NAD+ content, which
has been related to mPTP gating in situ. The NAD+
content of mitochondria incubated for 15 min with
rBAX, but not exposed to the calcium-containing
E. Martınez-Abundis et al. Microdomain components – effect on BAX-induced mPT
FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS 6501

medium, was similar to control mitochondria that
had undergone incubation and further exposure to a
medium supplemented with calcium, but not incu-
bated with rBAX (9.66 versus 9.38 nmol NAD+Æmg
protein)1, n ¼ 2). Conversely, NAD+ content
decreased in rBAX-treated mitochondria when mPTP
was induced in a medium supplemented with calcium
(2.08 nmol NAD+Æmg protein)1, n ¼ 2).
Lipid modification in rat kidney mitochondria
produced resistance to rBAX-induced
permeability transition
Methyl-beta-cyclodextrin (MbCD) is known to induce
cholesterol efflux from membranes and, consequently,
to promote microdomain disruption. We evaluated the
effect of this chelator on the ability of rBAX to induce
M
Time Course Graph
750
Control,
Cal
cium
rel
ease
rBAX + CSA
0 1500Time (seconds)
0.35Mitos ± rBAX
A 0.18
0.02
rBAX
BAX
Cytc
A B
Con
trol
rBA
X
rBA
X +
CS
A
Std
SS
M
Fig. 2. CSA prevents cytochrome c release
and mPTP opening induced by rBAX. (A)
After treatment of mitochondria with
150 nM rBAX, mPTP opening was evaluated
as calcium release under the conditions
described in Fig. 1. Where indicated, the
assay medium was supplemented with
1 lM CSA. (B) rBAX and cytochrome c con-
tent was detected in mitochondria, as
described in Fig. 1. Traces and blots are rep-
resentative of at least four different experi-
ments. M, mitochondria; SS, supernatants;
Std, purified rBAX and commercial cyto-
chrome c, respectively.
M
SS
CAT 1 μM
(nM)
200
100
50
M± rBAX
Time course graph
0 1090 2200 Time (seconds)
0.30
0.16
0.02
Cal
cium
rele
ase
rBAX (nM)
200 100 - 1
CAT (μM)
ANT
200
rBAX (nM)
BAX
A B
Cytc M
A
100 50 -
Fig. 1. Recombinant BAX induces mitochondrial calcium extrusion and cytochrome c release from isolated mitochondria. (A) Calcium uptake
in isolated mitochondria in the presence of different rBAX concentrations. Mitochondrial protein (1.3 mg) was incubated with oligomerized
rBAX, as described in Experimental procedures, for 15 min and then added to 2 mL of a medium containing 125 mM KCl, 10 mM Tris,
10 mM succinate, 3 mM Pi, 200 lM ADP, 50 lM CaCl2, 3 lgÆmL)1 rotenone and 50 lM Arsenazo III, pH 7.4. (B) BAX detection in mitochon-
dria. After maximal calcium release, 1.8 mL of the suspension was withdrawn and centrifuged at 18 000 g for 10 min. Mitochondrial sam-
ples were treated with Na2CO3 as described in Experimental procedures. Mitochondria were dissolved in 50 lL NaCl ⁄ Pi, pH 7.0, and mixed
with 25 lL of a 3· Laemmli’s loading buffer, boiled for 15 min, subjected to SDS ⁄ PAGE and evaluated by western blot. Protein loading was
determined by using anti-ANT polyclonal IgG. (C) Cytochrome c content in mitochondria (M) and in supernatants (SS) recovered after rBAX
incubation. Proteins in the supernatants were precipitated with trichloroacetic acid and evaluated for cytochrome c content along with mito-
chondrial samples. Results are representative of three independent experiments.
Microdomain components – effect on BAX-induced mPT E. Martınez-Abundis et al.
6502 FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS
opening of the permeability transition pore in rat
kidney mitochondria. MbCD at doses of 0.5–1.5 mm
prevented opening of the mPTP induced by 150 nm
oligomeric rBAX (Fig. 4A). Interestingly, rBAX
remained inserted into MbCD-treated mitochondria
(Fig. 4B). These results indicate that altered cholesterol
levels in mitochondrial membranes are important for
rBAX cross-talk with components of the mPTP, and
do not influence rBAX insertion.
Kidney mitochondria from hypothyroid rats were
refractive to mPTP-opening induced by rBAX
To gain insight into the existence of putative mito-
chondrial lipid microdomains in vivo, we evaluated the
inducing effect of rBAX on opening of the mPTP
in kidney mitochondria from hypothyroid rats. We
hypothesized that if cholesterol and gangliosides are
required for BAX-induced mPTP opening, and if such
0 500 1000Time (seconds)
Mem
bran
e po
tent
ial
(–)
(+)
Time course graph
0.30
0.25
0.20
0.15
0.10
A
d
a
b
c
A B
0 500 1000Time (seconds)
0.80
0.70
0.60
a, bcd
e
M±rBAX
Time course graph
Mito
chon
dria
l sw
ellin
g
A
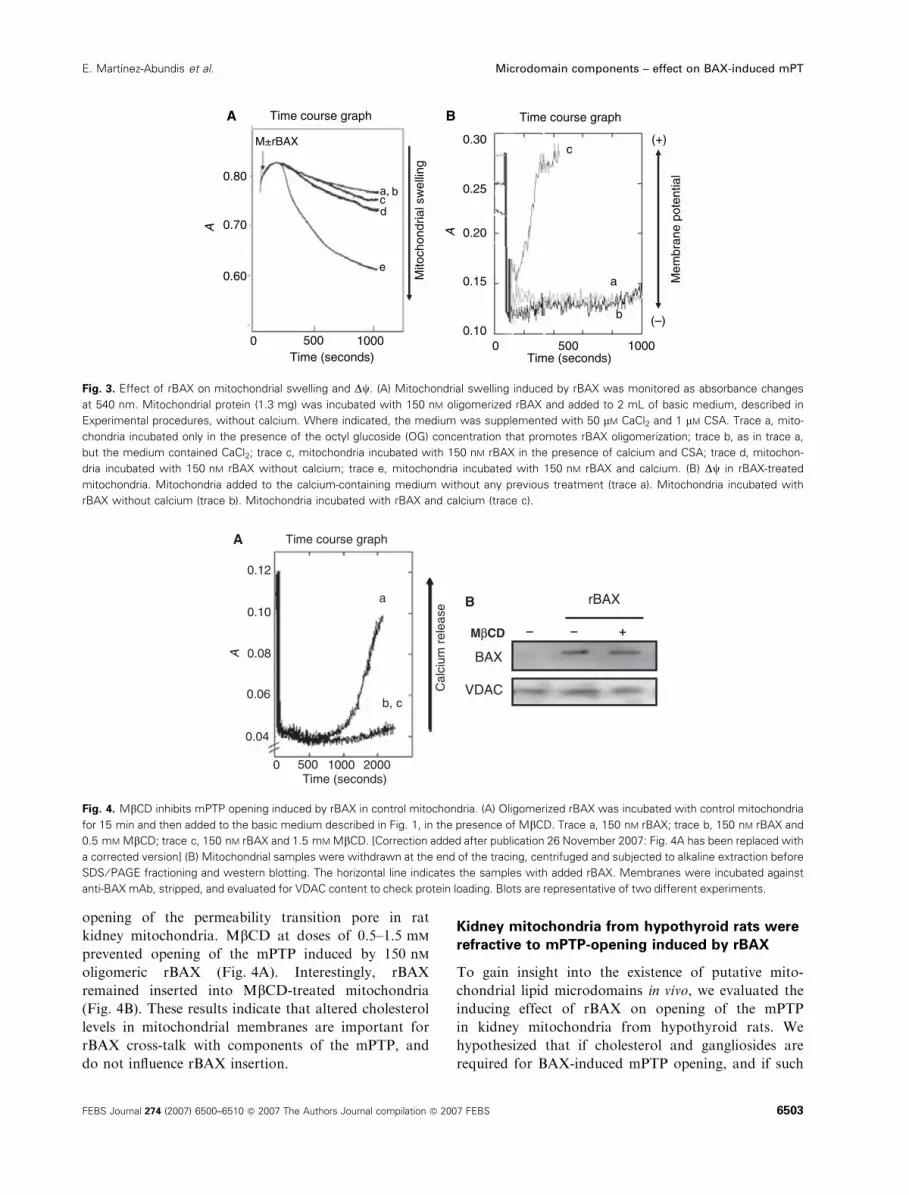
Fig. 3. Effect of rBAX on mitochondrial swelling and Dw. (A) Mitochondrial swelling induced by rBAX was monitored as absorbance changes
at 540 nm. Mitochondrial protein (1.3 mg) was incubated with 150 nM oligomerized rBAX and added to 2 mL of basic medium, described in
Experimental procedures, without calcium. Where indicated, the medium was supplemented with 50 lM CaCl2 and 1 lM CSA. Trace a, mito-
chondria incubated only in the presence of the octyl glucoside (OG) concentration that promotes rBAX oligomerization; trace b, as in trace a,
but the medium contained CaCl2; trace c, mitochondria incubated with 150 nM rBAX in the presence of calcium and CSA; trace d, mitochon-
dria incubated with 150 nM rBAX without calcium; trace e, mitochondria incubated with 150 nM rBAX and calcium. (B) Dw in rBAX-treated
mitochondria. Mitochondria added to the calcium-containing medium without any previous treatment (trace a). Mitochondria incubated with
rBAX without calcium (trace b). Mitochondria incubated with rBAX and calcium (trace c).
A
a
b, c
rBAX
BAX
MβCD – – +
A
B
VDAC
0
0.04
0.06
0.08
0.10
0.12
500 1000 2000Time (seconds)
Time course graph
Cal
cium
rel
ease
Fig. 4. MbCD inhibits mPTP opening induced by rBAX in control mitochondria. (A) Oligomerized rBAX was incubated with control mitochondria
for 15 min and then added to the basic medium described in Fig. 1, in the presence of MbCD. Trace a, 150 nM rBAX; trace b, 150 nM rBAX and
0.5 mM MbCD; trace c, 150 nM rBAX and 1.5 mM MbCD. [Correction added after publication 26 November 2007: Fig. 4A has been replaced with
a corrected version] (B) Mitochondrial samples were withdrawn at the end of the tracing, centrifuged and subjected to alkaline extraction before
SDS ⁄ PAGE fractioning and western blotting. The horizontal line indicates the samples with added rBAX. Membranes were incubated against
anti-BAX mAb, stripped, and evaluated for VDAC content to check protein loading. Blots are representative of two different experiments.
E. Martınez-Abundis et al. Microdomain components – effect on BAX-induced mPT
FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS 6503

domains could be key sites for BAX docking and
anchoring to the membrane, we should be able to find
a different response from either hypothyroid or control
mitochondria, on account of their modified mitochon-
drial lipid composition [23]. Figure 5A shows that
150 nm rBAX did not have any effect on mPTP open-
ing in hypothyroid mitochondria; furthermore, rBAX
insertion was dramatically diminished in mitochondrial
membranes (Fig. 5B).
Comparison between cholesterol and ganglioside
content in chemically modified rat kidney mito-
chondria and hypothyroid kidney mitochondria
The cholesterol content of control and hypothyroid
mitochondria was measured using gas chromatogra-
phy. In hypothyroid kidney mitochondria, cholesterol
content was 39% lower than in control (euthyroid)
mitochondria, i.e. 43.1 ± 1.6 versus 70.5 ± 14.7
lgÆmg)1 protein). Diminished cholesterol levels in
mitochondria correlated with inhibition of the ‘open
state’ of the mPTP induced by rBAX. Interestingly,
chemical cholesterol depletion of control mitochondria
membranes by MbCD (34%) produced resistance to
rBAX-induced permeability transition (Table 1). These
results suggest that specific lipid domains are required
for BAX interaction with mPTP components; alterna-
tively, cholesterol may be important for correct assem-
bly of the pore components after rBAX induction of
the ‘open state’.
Along with cholesterol, glycosphingolipids are
concentrated in specific lipid domains in the plasma
membrane. Recently, evidence has indicated that gly-
cosphingolipids and their precursor, ceramide, are also
associated with intracellular organelles, particularly
mitochondria [24]. Because GD3 is the main glyco-
sphingolipid associated with mitochondria and it has
been shown to have a role in mPTP regulation, we
analyzed the polar glycosphingolipid fraction from
total mitochondrial lipid extracts. The obtained glycos-
phingolipids were dried and separated by TLC. Only
one positive band was found after sulfuric acid detec-
tion that comigrated with the GD3 standard in control
mitochondria (not shown). Indeed, GD3 in hypothy-
roid mitochondria was almost undetectable after TLC.
To verify that GD3 signal variation was not due to
variable loading, we performed densitometric scanning
analysis of plates in which equal volumes of extracted
Control Hypothyroid
M BAX
VDAC
rBAX - - - + + + +
A B
A
0
0.10
0.15
0.20
0.05
500 1000 1750Time (seconds)
Cal
cium
rel
ease
a
b
cd
M±rBAX
Time course graph
Fig. 5. Hypothyroid mitochondria are resistant to mPT induced by rBAX. (A) Calcium release was measured in control and hypothyroid rat
kidney mitochondria as described in Fig. 1. Trace a, control mitochondria incubated with 150 nM rBAX; trace b, control mitochondria incu-
bated with medium without rBAX; trace c, hypothyroid mitochondria incubated in the presence of 150 nM rBAX; trace d, hypothyroid mito-
chondria incubated without rBAX. Results are representative of at least three independent experiments in which different mitochondria and
rBAX preparations were used. (B) Mitochondrial samples were withdrawn at the end of the tracing, centrifuged and subjected to alkaline
extraction before SDS ⁄ PAGE fractioning and western blotting. Membranes were incubated against anti-BAX mAb, stripped, and evaluated
for VDAC content to check loading. Blots are representative of three different experiments. M, mitochondria.
Table 1. Cholesterol content in control and hypothyroid mitochon-
dria after MbCD treatment. Mitochondria from control and hypothy-
roid rats were incubated with 1.5 mM MbCD, as indicated
previously. Total lipids were recovered and evaluated for choles-
terol content as indicated in Experimental procedures. Data repre-
sent mean ± SD of the indicated number of independent
experiments.
Cholesterol (lgÆmg protein)1)
Control
Hypothyroid
(n ¼ 5)
Dimethylsulfoxide-treated
mitochondria
70.55 ± 14.7 (n ¼ 5) 43.1 ± 1.6
MbCD-treated mitochondria 46.54 ± 17.6* (n ¼ 5) 37.6 ± 1.9*
*P < 0.05 versus mitochondria control.
Microdomain components – effect on BAX-induced mPT E. Martınez-Abundis et al.
6504 FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS

glycosphingolipid fractions, obtained from the same
amount of mitochondrial protein, were loaded and
developed with sulfuric acid (unspecific) and resorcinol
(ganglioside-specific stain). By increasing the volumes
applied to the plates we detected resorcinol-positive
signaling (Fig. 6A). The ratio between the signals is
plotted in Fig. 6B.
The lower GD3 content observed in hypothyroid
mitochondria is consistent with reports that hypothy-
roid conditions affect the biosynthesis and expression
of gangliosides in specific tissues and cell types [23].
This, along with lower cholesterol levels in the mem-
branes, may translate into a diminished ability to form
lipid microdomains in which specific proteins could be
recruited and possibly, to interact.
Because cardiolipin is relevant for mPTP regulation
and this inner membrane phospholipid is diminished in
thyroid insufficiency, a condition that favors the closed
mPTP conformation, we sought to determine the effect
of MbCD treatment on cardiolipin content in control
and hypothyroid mitochondria. Figure 7A, shows the
chromatogram of the phospholipids extracted from
both mitochondrial types after MbCD incubation. As
expected, cardiolipin content in hypothyroid mitochon-
dria was significantly diminished compared with con-
trol mitochondria, but no changes were observed in
MbCD-treated mitochondria. We conclude that
changes in cardiolipin content could not account for
MbCD protection from mPTP opening, at least in
control mitochondria, but may be important for rBAX
insertion into the mitochondrial membranes.
Discussion
The purpose of this study was to demonstrate the
participation of lipids, specifically those related to
microdomain components, in the activation of the per-
meability transition pore by the pro-apoptogenic pro-
tein BAX, using lipid-modified mitochondria in vitro
and in vivo.
It is widely known that membrane lipids have major
structural and functional roles modulating signaling
pathways. In the plasma membrane, apoptotic death
mediated by lipids is propagated by activation of the
surface receptor CD95 (FAS ⁄APO-1), and includes the
formation of ceramide and GD3 [24]. Ceramide is pro-
duced by the hydrolysis of membrane sphingomyelin
[25] and by means of a GD3 synthase (a2,8-sialyltrans-ferase), rapidly converted to gangliosides [26]. GD3
and cholesterol are relatively enriched in plasma mem-
branes and are concentrated in specialized domains
called rafts or lipid microdomains [15]. In this sense,
new evidence suggests that raft domains are not exclu-
sive to the plasma membrane [16,27]. In particular, the
existence of GD3-enriched microdomains in mito-
chondria [16], and furthermore, data suggesting the
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
A
B
*
**
Hypothyroid+ MβCD
Hypothyroid Control+ MβCD
Control
Sig
nal r
atio
(re
sorc
inol
/H2S
O4)
Control Hypo GD3
Resorcinol
H2SO4
MβCD - - + + - - + +
n= 9 n= 6 n= 5 n= 6
Fig. 6. GD3 content in control and hypothyroid mitochondria after
MbCD treatment. Mitochondrial ganglioside content in which 5 lg
standard GD3 was run in parallel with mitochondrial extracted lipids.
(A) The plate is representative of three independent experiments.
(B) The densitometric ratio between sulfuric acid detection for total
lipids and ganglioside-specific detection with resorcinol is pre-
sented. A value of P < 0.05 was considered statistically significant.
1.5
1
0.5
19.1 ± 5.2*
17.8 ± 6.0*
19.4 ± 2.0*
15.3 ± 4.3*
31.1 ± 6.4
30.1 ± 1.9
25.9 ± 6.4
29.2 ± 5.20
Hypo
(µg CL/mgprotein)
Control
(µg CL/mg protein)
MβCD
(mM)
MβCD (mM)
CL
A
B
0 0.5 1.0 1.5 0 0.5 1.0 1.5 CL
Control Hypo
Fig. 7. Mitochondrial cardiolipin analysis after MbCD-treatment. (A)
Cardiolipin was separated by TLC as described in the Experimental
procedures from control and hypothyroid mitochondria treated with
MbCD. (B) Quantification was performed using acridine orange. Val-
ues are expressed as the mean ± SD of three different experi-
ments. CL, cardiolipin. *P < 0.005 versus respective control.
E. Martınez-Abundis et al. Microdomain components – effect on BAX-induced mPT
FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS 6505

existence of a metabolic pathway of sphingolipids,
including several enzyme activities of sphingolipid
metabolism, have been reported in these organelles
[28]. Thus, we measured the oligomeric rBAX-inducing
effect on the mitochondrial permeability transition in
chemically modified renal cortex mitochondria and
renal cortex mitochondria from hypothyroid rats,
which show an in vivo modified mitochondrial lipid
composition. According to other groups [20,29,30], our
results indicate that rBAX promoted opening of the
mPTP, and not only formed supramolecular openings
in the outer mitochondrial membranes, as proposed
previously [31]. rBAX insertion into mitochondrial
membranes induced calcium release from the matrix
space, while cytochrome c content diminished. The
rBAX effect was similar to that exerted by carboxya-
tractyloside when used to induce mPTP opening
(Fig. 1). We also observed that rBAX-induced perme-
ability was totally abolished by the classic mPTP
inhibitor CSA (Fig. 2) and depended on calcium over-
load (Fig. 3).
When the lipid mitochondrial composition was dis-
rupted, a different response was observed. Control
mitochondria showed resistance to the rBAX-inducing
action on the mPTP, following removal of cholesterol
with MbCD. Diminution in cholesterol and GD3 levels
in treated membranes, correlated with low cholesterol
and GD3 content in mitochondrial membranes isolated
from hypothyroid rats that were naturally protected
against mPTP opening induced by rBAX (Fig. 5). It is
conceivable that the cholesterol ⁄ gangliosides ratio may
be relevant to maintaining the structure of the lipid
microdomains, in such a way that disruption of one of
the elements could be translated into a loss of the local
formation of specific and functional raft-associated
complexes. Although the MbCD properties to chelate
cholesterol are well known, there is, to our knowledge,
no report indicating that it also decreases GD3 con-
tent. Indeed, our results show that MbCD induced a
marked decrease in GD3, which represent a novel find-
ing. In this respect, it has been described that MbCD–
cholesterol extraction is accompanied by release of
GM1 molecules and proteins from plasma membranes
[32].
With respect to the relevance of cholesterol in a
mitochondrial lipid raft, it is known that addition of
cholesterol to membrane models containing only phos-
pholipids and sphingolipids permits the formation of a
liquid-order phase in which saturated acyl chains are
highly organized, as in a highly ordered gel phase, but
exhibit lateral mobility more similar to that in the
liquid-ordered crystalline phase [21]. If cholesterol is
the determinant for the generation of a liquid-ordered
phase, it follows that alterations in the cholesterol con-
tent of membranes should lead to changes in their
properties. Our findings suggest that cholesterol
decrease correlated with mPTP opening inhibition in
the presence of rBAX, which is consistent with studies
of cholesterol depletion that alters the function of a
raft-associated potassium channel [33]. Modifications
in the cholesterol content in vivo may lead to altera-
tions in the physical environment of the membrane,
consequently changing the likelihood of certain pro-
teins partitioning into these domains. Furthermore, it
has been reported that hypothyroid conditions affect
the biosynthesis and expression of gangliosides in spe-
cific tissues and cell types [34].
Our findings could be organized into three different
scenarios. First, cholesterol and GD3 in the mitochon-
drial membranes favored rBAX insertion and interac-
tion with the mPTP (control mitochondria); second, a
diminished cholesterol and GD3 content, resulted in a
lower likelihood of raft formation, and hence in cross-
talk disruption between rBAX and PTPm (control
mitochondria + MbCD). This is supported by recent
data indicating that the macromolecular complex
conformed by GD3, the voltage dependent anion chan-
nel-1 (VDAC-1) and the fission protein hFis1 could be
targets in which Bcl-2 family proteins are recruited
[16]. Relevant to this issue is that GD3 and ceramide
have been shown to boost the ability of BAX to
induce the mPTP [20]. Finally, in membranes with
endogenously low cholesterol and ganglioside content
(hypothyroid mitochondria) the closed state of the
mPTP was also favored. It has been reported that
hypothyroidism is associated not only with low choles-
terol and ganglioside levels, but also with decreased
mitochondrial functional activity, oxygen consumption,
ATP synthesis and, relevant for mPTP regulation, with
low cardiolipin content and a diminished amount of
some of the mPTP components (e.g. ANT) [35,36].
The striking difference between rBAX association in
control and hypothyroid mitochondria might be
explained by the differences in cholesterol and ganglio-
side content, along with modifications characteristic of
hypothyroidism, in particular cardiolipin content. A
potential function of GD3 could be to enhance translo-
cation of the pro-apoptotic protein BAX to mitochon-
dria. An attractive hypothesis to explain the results
obtained in hypothyroid mitochondria could be that
cardiolipin is relevant for BAX docking into the mem-
branes. Regarding the relevance of cardiolipin and
mPTP on BAX insertion, there are reports suggesting
that BAX permeabilizes synthetic liposomes only if
cardiolipin is present [31,37] and that oligomeric BAX
does not permeabilize outer membrane vesicles if the
Microdomain components – effect on BAX-induced mPT E. Martınez-Abundis et al.
6506 FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS

contact sites have been removed [38]. In this sense,
cardiolipin clusters associated with apoptoptic and
energy-flux process proteins have been found at con-
tact sites in the mitochondrial membranes [39].
In conclusion, our results indicate that cholesterol
and GD3 are relevant for the interaction of BAX with
the mPTP. The data provide evidence that specific lip-
ids play a key role in cross-talk between rBAX and the
mPTP. Indeed, more experimental data supporting the
idea that BAX and mPTP converge into mitochondrial
like-raft domains are imperative.
Experimental procedures
Antibodies and reagents
Chemicals were of reagent or higher grade from Sigma-
Aldrich (St. Louis, MO), unless otherwise specified. Glu-
tathione S-transferase (GST)–Sepharose, thrombin prote-
ase and enhanced chemioluminescence system detection
were obtained from Amersham Biosciences (Chalfont St
Giles, UK); protease inhibitors set was from Roche
(Mannheim, Germany) silica gel 60F254 was purchased
from Merck (Darmstadt, Germany); anti-ANT polyclonal
IgG (N-19) was from Santa Cruz Biotechnology Inc.
(Santa Cruz, CA); anti-BAX mAb (Clone 6A7) was from
Alexis Biochemicals (San Diego, CA), anti-(cytochrome c)
mAb (Clone 7H8.2C12) and biotin-conjugated secondary
antibodies were from Zymed Laboratories (San Francisco,
CA).
Induction of hypothyroidism
All animal procedures were performed in accordance with
the Guide for the Care and Use of Laboratory Animals
published by the US National Institutes of Health [publica-
tion no. 85 (23) revised 1996]. Male Wistar rats weighing
380 ± 17 g underwent surgical thyroidectomy with para-
thyroid re-implant, as described previously [40]. Briefly, the
trachea was exposed under anesthesia. Parathyroid glands
were visualized by means of a stereoscopic microscope
(Wild M5, Wild Heerbrugg, Switzerland), dissected from
the thyroid gland and re-implanted into the surrounding
neck muscles. The thyroid gland was then carefully dis-
sected to avoid injury to the laryngeal nerves and com-
pletely excised. The effectiveness of this procedure was
assessed by determining the concentration of calcium, phos-
phorous and thyroxin in 10 sham-operated control and 10
hypothyroid rats, using standard techniques. The results
obtained 15 days after surgery were: Ca2+, 10.2 ± 3 mm in
control versus 10.3 ± 0.2 mm in hypothyroid; phospho-
rous, 6.5 ± 0.3 mm in control versus 6.3 ± 0.5 mm in
hypothyroid and thyroxine 6.4 ± 7 lgÆL)1 in control versus
11.8 ± 1.9 lgÆL)1 in hypothyroid, P < 0.05. The sham
group (375 ± 14 g) underwent a surgical procedure in
which the animals were anesthetized, the trachea was
exposed and the incision was closed, simulating thyroidec-
tomy.
Purification of BAX-DC (rBAX) protein
Recombinant GST–BAX-DC (protein lacking the C-termi-
nal 20 amino acids) was prepared according to Xie et al.
[41] with slight modifications. Briefly, Escherichia coli
BL21(DE3)pLysS cells carrying the plasmid pGEX-4T1–
BAX-DC were grown overnight at 37 �C in Luria Bertani
medium supplemented with 100 lgÆmL)1 ampicillin,
25 lgÆmL)1 chloramphenicol and 1% glucose. Cells were
cultured overnight at 37 �C after induction with 0.4 mm
isopropyl-b-d-thiogalactoside. Harvested cells were dis-
rupted with lysozyme in the presence of 1% TX 100 and
sonicated in NaCl ⁄Pi, pH 7.4. Unbroken cells were elimi-
nated after centrifugation at 5000 g, for 10 min. High speed
centrifugation (100 000 g for 30 min at 4 �C) was per-
formed to clear the supernatant, before glutathione–Sepha-
rose affinity chromatography. Unbound protein was
washed extensively using NaCl ⁄Pi, pH 7.4, supplemented
with 0.1% TX 100 and the protein was recovered by over-
night proteolytic cleavage with thrombin at room tempera-
ture. Eluted rBAX was dialyzed against 250 vol 10 mm
Tris ⁄HCl (pH 8.0), 1 mm EDTA, and 0.1% (v ⁄ v) 2-b-mer-
captoethanol. rBAX was aliquoted and stored at )70 �C.
rBAX oligomerization
rBAX oligomerization was induced by incubating the pro-
tein with 1% octylglucoside for 60 min at 4 �C. The protein
was then diluted 10· in the assay buffer and incubated with
isolated mitochondria, before measurement of mitochon-
drial permeability transition.
Mitochondrial permeability transition pore
opening induced by rBAX
Mitochondria were isolated from rat kidney cortex by dif-
ferential centrifugation in a sucrose-based medium, as
described previously [42]. Mitochondrial protein (1.3 mg)
was incubated for 15 min in the presence of oligomerized
rBAX at the indicated concentration. Opening of the mPTP
was evaluated by measuring mitochondrial calcium move-
ments, mitochondrial swelling and Dw in a basic medium
containing, 125 mm KCl, 10 mm Hepes, 10 mm succinate,
3 mm Pi, pH 7.4, plus 200 lm ADP, 3 lgÆmL)1 rotenone
and, where indicated, 50 lm CaCl2.
Mitochondrial calcium uptake was evaluated spectropho-
tometrically in a double beam spectrophotometer at 675–
685 nm, by using the metallochromic dye Arsenazo III [43].
Mitochondrial swelling was followed by changes in
E. Martınez-Abundis et al. Microdomain components – effect on BAX-induced mPT
FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS 6507

absorbance at 540 nm and Dw was determined by using the
dye safranine, at 524–554 nm [44].
Measurement of NAD+ content in mitochondria
Mitochondrial NAD+ was measured after perchloric acid
extraction as described previously [45], with minor modifi-
cations. We added 0.5 mL of 21% (v ⁄ v) HClO4 to 10 mg
mitochondrial protein per mL of suspension and incubated
during 30 min in an ice-cold bath. The suspension was cen-
trifuged at 8000 g and the supernatans neutralized. NAD+
was determined fluorometrically at kex ¼ 340 nm and
kem ¼ 460 nm, by measuring NAD+-dependent lactate
dehydrogenase activity, in a medium containing 3 lg of lac-
tate dehydrogenase from rabbit muscle, 400 mm Hydrazine,
500 mm glycine and 10 mm l-lactate pH 9.0, at 25 �C.
rBAX insertion into mitochondria and
cytochrome c release
rBAX insertion and cytochrome c content in mitochondria
were evaluated by western blot analysis, using a primary
anti-BAX mAb (1 lgÆmL)1) or a mAb against cyto-
chrome c (1 : 1000 dilution). Biotin-conjugated secondary
antibodies and streptavidin–peroxidase conjugate were used
followed by an enhanced chemiluminescence system detec-
tion. After maximal calcium release, 1.8 mL of the suspen-
sion was withdrawn and centrifuged at 18 000 g for 10 min.
To discard the protein loosely bound to the membranes,
the mitochondrial pellets were incubated with 0.1 m
Na2CO3, pH 11.5 (alkaline extraction), for 10 min at room
temperature, a procedure that assures that only full inserted
BAX would be detected. Mitochondria were recovered by
centrifugation, washed once with 250 mm sucrose, 10 mm
Tris, pH 7.4, and suspended in sample buffer for electro-
phoresis. Total mitochondrial protein (50 lg) was loaded
into each lane of the SDS ⁄PAGE gels and transferred to
poly(vinylidene difluoride) membranes for immunodetec-
tion. To assess protein loading, the membranes were
‘stripped’ in a buffer containing 62.5 mm Tris ⁄HCl,
100 mm b-mercaptoethanol, 2% SDS, pH 6.7, for 20 min
at 50 �C. The membranes were incubated against anti-ANT
or anti-VDAC polyclonal IgG.
Cholesterol and ganglioside measurements
Control and hypothyroid mitochondria were incubated with
the cholesterol-chelator MbCD for 25 min at 25 �C and
mild shaking. After depletion, mitochondria were centri-
fuged at 8000 g and suspended in NaCl ⁄Pi, pH 7.4, for
further analysis. Total cholesterol was determined from
mitochondrial lipid extracts obtained with chloro-
form ⁄methanol (2 : 1) in the presence of 0.02% butyl-
hydroxy-toluene and using 50 lg stigmasterol as the
internal standard, as described previously [46]. Lipid
extracts were incubated overnight at room temperature with
hexamethyldisilazane and trimethylchlorsilane in dry pyri-
dine. Cholesterol was purified in a fused silica noncapillary
column SE 54 (30 cm, 0.35 mm id) and quantified by gas
chromatography using a gas chromatograph (Carlo Erba
2003), equipped with a flame ionization detector and a
split-less inlet system.
Gangliosides were extracted from control and hypo-
thyroid mitochondria previously treated with MbCD and
analyzed by preparative TLC on silica gel plates with chlo-
roform ⁄methanol ⁄ 0.2% CaCl2 ⁄ acetic acid (7 : 3 : 0.5 : 0.2
v ⁄ v ⁄ v ⁄ v), as described previously [47]. The plates were
sprayed with 10% H2SO4 (v ⁄ v) and heated for spot detec-
tion. GD3 standard gave two equivalent spots (RF ¼ 0.85
and 0.81). The double band is known to result from the
heterogeneity of fatty acid composition in gangliosides.
Positive signal in the samples correlated with the band pre-
senting a retention factor of 0.85 from the GD3 standard.
No signal was detected comigrating with the GM1 stan-
dard. Alternatively, gangliosides were specifically detected
with resorcinol reagent [48] and compared with the signal
developed by H2SO4 using a simple ‘dot’ assay. Cardiolipin
content was analyzed after inner mitochondrial phospho-
lipid extraction with chloroform ⁄methanol (2 : 1 v ⁄ v), thenindividual phospholipids were separated by TLC on silica
gel plates with chloroform ⁄methanol ⁄H2O (65 : 25 : 4
v ⁄ v ⁄ v) and visualized with iodine. Spots comigrating with
the cardiolipin standard were scraped out from the chro-
matogram and quantified spectrophotometrically at
472 ⁄ 492 nm using acridine orange, as described [49].
Statistical analysis
Values are given as means ± SD and were evaluated by
one-way ANOVA. P < 0.05 was considered the threshold
for statistical significance between the groups indicated.
Acknowledgments
We greatly acknowledge the generous gift of pGEX-
4T-I BAX plasmid from Dr John Reed (Burnham
Institute for Medical Research, La Jolla, CA) and par-
tial support from the Doctorate Program in Biomedi-
cal Sciences of the National Autonomous University
of Mexico (UNAM) to Eduardo Martınez-Abundis.
We also thank Mr Jose Santamarıa (Nephrology
Department, National Institute of Cardiology, Ignacio
Chavez) for performing the surgical procedures. This
study was partially supported by grant 46456-M to CZ
from the National Council of Science and Technology
(CONACYT), Mexico. The authors state no conflict
of interest.
Microdomain components – effect on BAX-induced mPT E. Martınez-Abundis et al.
6508 FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS

References
1 Wolter G, Hsu Y, Smith C, Nechushtan A, Xi X &
Youle K (1997) Movement of Bax from the cytosol to
mitochondria during apoptosis. J Cell Biol 139, 1281–
1292.
2 Zhang H, Heim J & Meyhack B (1998) Redistribution
of BAX from cytosol to membranes is induced by apop-
totic stimuli and is an early step in the apoptotic path-
way. Biochem Biophys Res Commun 251, 254–259.
3 Nechushtan A, Smith C, Hsu Y & Youle K (1999)
Conformation of the BAX C-terminus region regulates
subcellular location and cell death. EMBO J 18, 2330–
2341.
4 Schinzel A, Kaufmann T, Schuler M, Martinalbo J,
Grubb D & Borner C (2005) Conformational control of
Bax localization and apoptotic activity by Pro168.
J Cell Biol 164, 1021–1032.
5 Zou H, Li Y, Liu X & Wang X (1999) An Apaf-1–cyto-
chrome c multimeric complex is a functional apopto-
some that activates pro-caspase-9. J Biol Chem 274,
11549–11556.
6 Yi X, Yin XM & Dong Z (2003) Inhibition of Bid-
induced apoptosis by Bcl-2. tBid insertion, Bax translo-
cation and Bax ⁄Bak oligomerization supressed. J Biol
Chem 278, 16992–16999.
7 Jiang M, Pabla N, Murphy RF, Yang T, Yin XM,
Degenhardt K, White E & Dong Z (2007) Nutlin-3 pro-
tects kidney cells during cisplatin therapy by suppressing
Bax ⁄Bak activation. J Biol Chem 282, 2636–2645.
8 Li H, Zhu H, Xu CJ & Yuan J (1998) Cleavege of BID
by caspase 8 mediates the mitochondrial damage in the
Fas pathway of apoptosis. Cell 94, 491–501.
9 Wei MC, Zong WX, Cheng EH, Lindsten T, Papaut-
sakopoulou V, Ross AJ, Roth KA, MacGregor GR,
Thompson CB & Korsmeyers J (2001) Proapoptotic
BAX and BAK: a requisite gateway to mitochondrial
disfunction and death. Science 292, 727–730.
10 Zong WX, Lindsten T, Ross AJ, MacGregor GR &
Thompson CB (2001) BH-only proteins that bind pro-
survival Bcl-2 family members fail to induce apoptosis
in the absence of Bax and Bak. Genes Dev 15, 1481–
1486.
11 Basanez G, Sharpe J, Galanis J, Brandt T, Hardwick J
& Zimmerberg J (2002) Bax-type apoptotic proteins
porate pure lipid bilayers through a mechanism sensitive
to intrinsic monolayer curvature. J Biol Chem 277,
49360–49365.
12 Gogvadze V, Robertson J, Zhivotovsky B & Orrenius S
(2001) Cytochrome c release occurs via Ca2+-dependent
and Ca2+-independent mechanisms that are regulated
by Bax. J Biol Chem 276, 19066–19071.
13 Vyssokikh M, Zorova L, Zorov D, Heimlich G,
Jurgensmeier J & Brdiczka D (2002) Bax releases
cytochrome c preferentially from a complex between
porin and adenine nucleotide translocator. Hexokinase
activity suppresses this effect. Mol Biol Rep 29, 93–96.
14 Shimizu S, Ide T, Yanagida T & Tsujimoto Y (2000)
Electrophysiological study of a novel large pore formed
by BAX and the voltage-dependent anion channel that
is permeable to cytochrome c. J Biol Chem 275, 1231–
1235.
15 Pike L (2004) Lipid rafts: heterogeneity on the high
seas. Biochem J 378, 281–292.
16 Garofalo T, Giammarioli A, Misasi R, Tinari A, Man-
ganelli V, Gambardella L, Pavan A, Malorni W &
Sorice M (2005) Lipid microdomains contribute to
apoptosis-associated modifications of mitochondria in
T cells. Cell Death Differ 12, 1378–1389.
17 Scorrano L, Petronilli V, Di Lisa F & Bernardi P (1999)
Commitment to apoptosis by GD3 ganglioside depends
on the opening of the mitochondrial transition pore.
J Biol Chem 274, 22581–22585.
18 Garcıa-Ruiz C, Colell A, Parıs R & Fernandez-Checa J
(2000) Direct interaction of GD3 ganglioside with mito-
chondria generates reactive oxygen species followed by
mitochondrial permeability transition, cytochrome c
release and caspase activation. FASEB J 14, 847–858.
19 Rippo M, Malisan F, Ravagnan L, Tomassini B, Condo
I, Costantini P, Susin S, Rufino A, Todazo M, Kroemer
G et al. (2000) GD3 ganglioside directly targets mito-
chondria in bcl-2-controlled fashion. FASEB J 14,
2047–2054.
20 Pastorino J, Tafani M, Rothman R, Marcinkeviciute A,
Hoek J & Farber J (1999) Functional consequences of
the sustained or transient activation by Bax of the mito-
chondrial transition pore. J Biol Chem 274, 31734–
31739.
21 Brown D & London E (2000) Structure and function of
sphingolipid- and cholesterol-rich membrane rafts.
J Biol Chem 275, 17221–17224.
22 Keller R, Arnold T & Fliester S (2004) Formation of
7-dehydrocholesterol containing membrane rafts in vitro
and in vivo, with relevance to the Smith–Lemli–Opitz
syndrome. J Lipid Res 45, 347–355.
23 Paradies G, Ruggiero F & Dinoi P (1991) The influence
of hypothyroidism on the transport of phosphate and
on the lipid composition in rat-liver mitochondria.
Biochim Biophys Acta 1070, 180–186.
24 Ardail D, Popa I, Bodennee J, Louisot P, Schmitt D &
Portoukalian J (2003) The mitochondria-associated
endoplasmic reticulum subcompartment (MAM frac-
tion) of rat liver contains highly active sphingolipid-spe-
cific glycosyltransferases. Biochem J 371, 1013–1019.
25 Cifone M, Roncaioli P, De Maria R, Camarda G, San-
toni A, Ruberti G & Testi R (1995) Multiple pathways
originate at the Fas ⁄APO1 (CD95) receptor: sequential
involvement of phosphatidylcholine-specific phospholi-
pase C and acidic sphingomyelinase in the programation
of the apoptotic signal. EMBO J 14, 5859–5868.
E. Martınez-Abundis et al. Microdomain components – effect on BAX-induced mPT
FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS 6509

26 Testi R (1996) Sphingomyelin breakdown and cell fate.
Trends Biochem Sci 21, 468–471.
27 Nakayama J, Fukuda M, Hirabayashi Y, Kanamori A,
Sasaki K, Nishi T & Fukuda M (1996) Expression clon-
ing of a human GT3 synthase. GD3 and GT3 are synthe-
sized by a single enzyme. J Biol Chem 271, 3684–3691.
28 Birbes H, El Bawab S, Hannun Y & Obeid L (2001) A
mitochondrial pool of sphingomyelin is involved in
TNF-a-induced BAX translocation to mitochondria.
FASEB J 14, 2669–2679.
29 Brenner C, Cadiou H, Vieira H, Zamzani N, Marzo I,
Xie Z, Leber B, Andrews D, Duclohier H, Reed J et al.
(2000) Bcl-2 and Bax regulate the channel activity of the
mitochondrial adenine nucleotide translocator. Onco-
gene 19, 329–336.
30 Guha M, Kumar S, Coubev V, Maity P & Bandyopad-
hyay U (2006) Apoptosis in liver during malaria: role of
oxidative stress and implication of mitochondrial path-
way. FASEB J 20, 1224–1226.
31 Kuwana T, Mackey MR, Perkins G, Ellisman MH,
Latterich M, Schneiter R, Green DR & Newmeyer DD
(2002) Bid, Bax and lipids cooperate to form supramo-
lecular openings in the outer mitochondrial membrane.
Cell 111, 331–342.
32 Ilangumaran S & Hoessli DC (1998) Effects of
cholesterol depletion by cyclodextrin on sphingolipid
microdomains of the plasma membrane. Biochem J 335,
433–440.
33 Martens J, Navarro-Polanco R, Coppock E, Nishiyama
A, Parshley L & Grobaski T (2000) Differential target-
ing of shaker-like potassium channels to lipid rafts.
J Biol Chem 275, 7443–7446.
34 Saito M & Sugiyama K (2000) Specific ganglioside
changes in extraneural tissues of adult rats with hypo-
thyroidism. Biochim Biophys Acta 1523, 230–235.
35 Brand MD, Steverding D, Kadenbach S, Stevenson P &
Hafner RP (1992) The mechanism of the increased pro-
ton permeability induced by thyroid hormone. Eur J
Biochem 264, 7826–7830.
36 Chavez E, Franco M, Reyes-Vivas H, Zazueta C, Ram-
ırez J & Carrillo R (1998) Hypothyroidism renders liver
mitochondria resistant to the opening of membrane per-
meability transition pore. Biochim Biophys Acta 1407,
243–248.
37 Terrones O, Antonsson B, Yamaguchi H, Wang H, Liu
Y, Lee R, Herrmann A & Basanez G (2004) Lipid pore
formation by the concerted action of pro-apoptotic
BAX and tBID. J Biol Chem 279, 30081–30091.
38 Wieckowski M, Vyssokikh M, Dymkowska D, Antons-
son B, Brdiczka D & Wojtczak L (2001) Oligomeric
C-terminal truncated Bax preferently releases cyto-
chrome c but no adenylate kinase from mitochondria,
outer membrane vesicles and proteoliposomes. FEBS
Lett 505, 453–459.
39 Epand RF, Schlattner U, Wallimann T & Epand RM
(2007) Cardiolipin clusters and membrane domain for-
mation induced by mitochondrial proteins. J Mol Biol
365, 968–980.
40 Franco M, Bobadilla N, Suarez J, Tapia E, Sanchez L
& Herrera-Acosta J (1996) Participation of adenosine in
the renal hemodynamic abnormalities of hypothyroid-
ism. Am J Physiol 270, F254–F262.
41 Xie Z, Schendel S, Matsuyama S & Reed J (1998)
Acidic pH promotes dimerization of Bcl-2 family pro-
teins. Biochemistry 37, 6410–6418.
42 Chavez E, Briones R, Michel B, Bravo C & Jay D
(1985) Evidence for the involvement of dithiol groups in
mitochondrial calcium transport: studies with cadmium.
Arch Biochem Biophys 242, 293–297.
43 Kendrick N (1976) Purification of arsenazo III, a cal-
cium sensitive dye. Ann Biochem 76, 487–501.
44 Zanotti A & Azzone GF (1980) Safranine as a mem-
brane potential probe in rat liver mitochondria. Arch
Biochem Biophys 201, 255–265.
45 Di Lisa F, Menabo R, Canton M, Barile M & Bernardi
P (2001) Opening of the mitochondrial permeability
transition pore causes depletion of mitochondrial and
cytosolic NAD+ and is a causative event in the death
of myocytes in postischemic reperfusion of the heart.
J Biol Chem 276, 2571–2575.
46 Carvajal K, El Hafidi M, Marın-Hernandez A &
Moreno-Sanchez R (2005) Structural and functional
changes in heart mitochondria from sucrose-fed hyper-
triglyceridemic rats. Biochim Biophys Acta 1709, 231–
239.
47 Miyazaki K, Okamura N, Kishimoto Y & Lee Y (1986)
Determination of gangliosides as 2,4-dinitrophenylhyd-
razides by high-performance liquid chromatography.
Biochem J 235, 755–761.
48 Svennerholm L (1957) Quantitative stimation of sialic
acids. II. A colorimetric resorcinol–hydrochloric acid
method. Biochim Biophys Acta 617, 97–109.
49 Petit JM, Huet O, Gallet P, Maftah A, Ratinaud M &
Julien R (1994) Direct analysis and significance of car-
diolipin transverse distribution in mitochondrial inner
membrane. Eur J Biochem 220, 871–879.
Microdomain components – effect on BAX-induced mPT E. Martınez-Abundis et al.
6510 FEBS Journal 274 (2007) 6500–6510 ª 2007 The Authors Journal compilation ª 2007 FEBS





























