Embed Size (px)
Citation preview

JOURNAL OF MORPHOLOGY 222:327-335 (1994)
Detection of Sugar Residues in Lizard Tooth Germs (Liolaemus gravenhorsti) Using Lectin Histochemistry
D. LEMUS, R. CABELLO, R. LEMUS, M. SOTO, AND M. FUENZALIDA Department of Experimental Morphology, Laboratory of Experimental Embryology, Medical School, University of Chile, Santiago, Chile
ABSTRACT The appearance, cellular distribution, and changes of sugar residues during tooth development in adults of the polyphyodont, Liolaemus gruvenhorsti, were investigated by using horseradish-peroxidase-conjugate lectins (lectin-HRP). With Con A (Cunuualiu enszformis), the ameloblasts (late bell stage) show granular supranuclear positivity and also at the Golgi zone and on their tomes process. Reactivity also appears at the apical surface of the odontoblasts and odontoblastic process. With WGA (Tritzcum vulgaris), the tooth germs (late bell stage) show cytoplasmatic granular positivity in the ameloblast cells, Golgi regions, and in a lesser extent of the cytoplasm. Also, the apical surface and the odontoblastic process react. WGA reaction is de- pressed following sialidase treatment.
The significance in tooth germs of a-D-mannose, a-D-glucose as well as p-D-N-acetylglucosamine and sialic acid is difficult to ascertain. These oligosac- charides may have some significance in odontogenesis. In fact, Con A-HRP- and WGA-HRP-binding components in ameloblasts and odontoblasts may be func- tionally related to molecules that are thought to contribute to odontogenesis in lizards. o 1994 Wiley-~iss, ~ n c .
The developing tooth is an excellent tool with which to study the mechanisms govern- ing organogenesis. Evidence suggests that developmental processes related to odonto- genesis are similar in all vertebrates. It is been well established that morphogenesis as well as cell differentiation in the tooth germ are under the control of reciprocal interac- tions between epithelial and mesenchymal tissues (Kollar and Baird, '70b; Slavkin, '74; Ruch, '87; Mina and Kollar, '87; Fuenzalida et d., '90; Thesleff et d., '91). The molecular mechanisms as well as signal substances in- volved in these interactions are mostly un- known at present (Thesleffet al., '90). Some extracellular matrix (ECM) components and growth factors (Thesleff et al., 1991; Hu et al., '93; Slavkin, '93) are good candidates for signal and mediator molecules in tissue inter- actions. Moreover, protein modifications such as phosphorylation and desphosphorylation are known to control several cell functions including regulation of the cell cycle, signal transduction and enzyme activationlinactiva- tion (Zeichner-David et al., '92). By contrast, lectins have been employed as powerful probes for the cytochemical demonstration of
carbohydrate moieties on cell surfaces and in cytoplasmic organelles during cellular differ- entiation in various developing systems (Takata et al., '83; Faze1 et al., '89; Gheri et al., '90, '91; Sasano et al., '92a). Some au- thors suggest that glycoconjugates recog- nized by Aruchis hipogeu (PNA) and Ma- cluru pomifera (MPA) may be functionally related to molecules which contribute to root formation and cementogenesis during mouse molar development (Sasano et al., '92b). The majority of experimental work has employed embryonic teeth of mammals, such as rats, mice, rabbits, guinea pigs, cats, and dogs.
Experimental studies on lizard teeth have not been conducted despite the fact that most species are polyphyodont (i.e., the tooth is replaced continuously) (Fig. 1). In recent years, our interest has been centered on the embryology of two species: Liolaemus t. tenuis and L. gruvenhorsti (Lemus and Duvauchelle, '66; Lemus, '67; Lemus et al., '81). These species are polyphyodont, a condition that is
Address reprint requests to Dr. D. Lemus, Department of Experi- mental Morphology, Laboratory of Experimental Embryology, Casilla 70023-Santiago, Correo 7, Santiago, Chile.
o 1994 WILEY-LISS, INC

328 D. LEMUS ET AL
accentuated in L. gravenhorsti a viviparous species in which tooth germ involution and reabsorption occur during intrauterine devel- opment (Lemus et al., '77). We also have shown that isolated and cultured tooth rudi- ments from adult L. grauenhorsti are able to differentiate teeth (Lemus et al., '80). These results imply that lizard dental tissues main- tain a continuous morphogenetic field throughout life. Moreover, heterologous com- binations of whole tooth buds of adult L. tenuis and L. grauenhorsti with neural crest cells from embryos of the quail Coturnix c. jap6nica showed that some quail cells ex- pressed certain dental characteristics. These results suggest that during in vitro culture of polyphyodont lizard tooth germs, some trans- missible morphogenetic constituents are pro- duced (Lemus et d . , '83). We have cultured heterologous tissue recombinations between quail epithelia and lizard dental papillae in order to examine the hypothetical induction role of the papilla. These experiments show that lizard dental papilla and quail epithe- lium are capable of producing developmental instructions responding to reciprocal interac- tions, resulting in odontogenesis and amelo- genesis (Lemus et al., '86).
In this study, we investigate the binding pattern of dental tissues with four sugar- specific HRP-lectins. This is intended to es- tablish a relationship between morphogen- esis and specific changes in sugar residues during odontogenesis in polyphyodont liz- ards.
MATERIALS AND METHODS
Adult Liolaemus t. tenuis were collected in the foothill of the Andes mountains near Santiago, Chile. A total of 11 males were employed in this study. Lizards were killed by decapitation, and their mandibular arches were dissected. The jaws were fixed either in Carnoy's fluid or in 6% mercuric chloride in 1% sodium acetate containing 0.1% glutaral- dehyde, followed by decalcification in 5% EDTA buffered with 0.01 M phosphate-buff- ered saline (PBS), pH 7.4, at 4°C for 3 weeks. Following ethanol dehydration, tissues were embedded in paraffin. Serial sections (5 pm) were cut parallel to the long axis of the jaw and processed for lectin histochemistry. Sec- tions from tissue fixed in buffered glutaralde- hyde-mercuric chloride were treated with Lugol's solution prior to staining.
After hydration, sections were treated with 0.3% hydrogen peroxide for 10 min (to in- hibit the endogenous peroxidase), rinsed in
distilled water and washed with 1% bovine serum albumin (BSA) in 0.1 M PBS pH 7.2. The sections were incubated for 30 min at room temperature in HRP-lectin-conjugated (Con A Canaualia ensiformis and WGA Triti- cum vulgaris) dissolved in 0.1 M PBS pH 7.2, containing0.1 M NaC1,O.l mM CaC12, MgC12, and MnC12, and then rinsed three times in PBS. The optimal concentration for each lec- tin (Sigma Chemical Co., St. Louis, MO), which allowed maximum staining with mini- mum background was as follows: Con A 50 pg/ml, WGA 20 pg/ml. Staining of the sites containing bound lectin-HRP was obtained by incubating the slides with PBS (pH 7.01, containing 3,3'-diaminobenzidine (DAB) (25 pg/dl) and 0.003% hydrogen peroxide, for 10 min at room temperature. Specimens were rinsed in distilled water, dehydrated using graded ethanol solutions, cleared in xylene, and mounted in Permount.
Control experiments Control for lectin staining included expo-
sure to HRP and substrate medium without lectin; oxidation with 1% periodic acid for 10 min prior to lectin staining and substitution of unconjugated lectin-HRP conjugates. As a control to confirm the sugar specificity for each of the lectins, adjacent sections were incubated with a mixture of lectin-HRP con- jugate (20 pg/ml) and appropriate inhibitory sugars (0.1 M). Methyl-a-D-mannose and methyl-a-D-glucoside were used as the inhibi- tory sugars for Con A, and N-acetyl-D-glu- cosamine was used for WGA. In all controls incubated in lectin-HRP plus inhibitory sug- ars, only melanophores reacted (Fig. 2).
In some experiments, sidic acid was re- moved by pretreating the sections for 18 hr at 37°C in a solution of sodium acetate buffer 0.25 M, pH 5.5, containing 0.1 U/ml sialidase (neuraminidase type X from Clostridiumper- fn'ngens, Sigma Chemical Co., St. Louis, MO), 5.0 mM CaC12, and 154 mM NaCl, prior to staining with lectin-HRP conjugate. Con- trols containing the sialidase buffer without the enzyme were also prepared.
RESULTS
Con A reacts with the a-D-mannOSe and a-D-glucose moieties of glycoconjugates. This lectin binds to the ameloblast and odonto- blast cells of rudiments (early and late bell stage) and well developed teeth (Fig. 3). The ameloblast cells of tooth germs in these stages show granular supranuclear positivity. In-
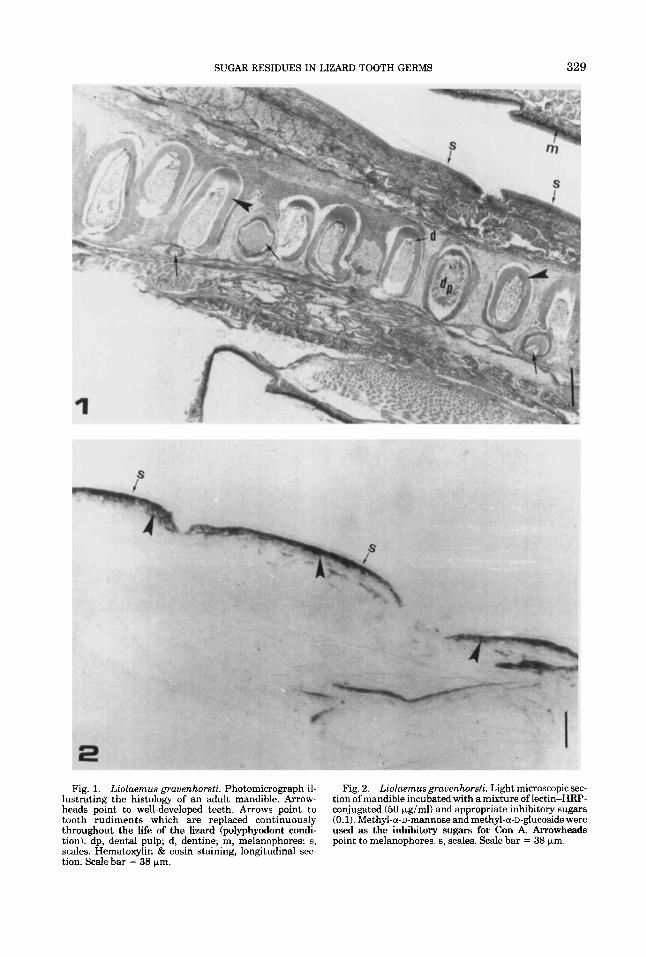
SUGAR RESIDUES IN LIZARD TOOTH GERMS 329
Fig. 1. Liolaemus gruuenhorsti. Photomicrograph il- Fig. 2 . Liolaemusgrauenhorsti. Light microscopic sec- lustrating the histology of an adult mandible. Arrow- tion of mandible incubated with a mixture of lectin-HRP- heads point to well-developed teeth. Arrows point to conjugated (50 pgiml) and appropriate inhibitory sugars tooth rudiments which are replaced continuously (0.1). Methyl-a-D-mannose and methyl-a-D-glucoside were throughout the life of the lizard Cpolyphyodont eondi- used as the inhibitory sugars for Con A. Arrowheads tion). dp, dental pulp; d, dentine; m, melanophores; s, point to melanophores. s, scales. Scale bar = 38 pm. scales. Hematoxylin & eosin staining, longitudinal sec- tion. Scale bar = 38 pm.
330 D. LEMUS ET AL
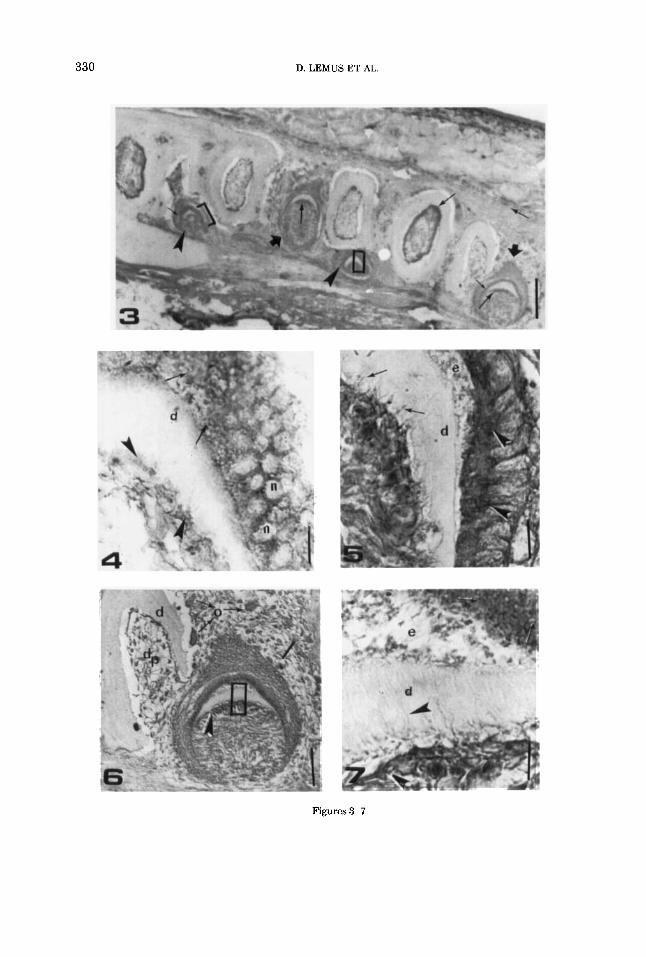
Figures 3-7

SUGAR RESIDUES IN LIZARD TOOTH GERMS 331
tense staining also occurs at the Golgi zone as well as on Tomes processes (Figs. 4, 5, 7). Reactivity is also observable at the apical surface of the odontoblast cells and odonto- blastic processes (Figs. 4,6, 7).
WGA, specific for N-acetyl-D-glucosamine and sialic acid is bound to the ameloblast and odontoblast cells of all the teeth observed. The tooth germs in the late bell stage of development show supranuclear cytoplas- matic granular positivity of the ameloblast cells. The Golgi region and, to a lesser extent the cytoplasm, are reactive (Figs. 8,9). Reac- tivity is also observable at the apical surface and at the odontoblastic process (Figs. 9, 11, 12). Following sialidase treatment, WGA re- activity is slightly depressed in both the am- eloblast and the odontoblast cells (Figs. 9, 10).
Cytochemical controls When sections are stained with the lectin
in one of the haptenic sugars pertinent to each lectin, positive reactions disappear com-
Fig. 3. Liolaemus grauenhorsti. Con A-HRP. Light microscopic section of mandible. The odontoblast cells (long arrows) and ameloblast cells (short arrows) cells of well developed tooth and teeth rudiments are intensely stained. Arrowheads and thick arrows point to tooth germs in early and late stages of development respec- tively. Scale bar = 36 pm.
Liolaemus grauenhorsti. Higher magnifica- tion of the area indicated by the bracket in Figure 3. Positive granular material is observable at the supra- nuclear cytoplasm of the functional ameloblasts (ar- rows). Moderate reaction is also shown by the surface of the functional cdontoblasts and their odontoblastic pro- cess (arrowheads). d, dentin; e, enamel; n, nucleus. Scale bar = 0.6 pm.
Liolaemus grauenhorsti. Higher magnifica- tion of the area encompassed by the rectangle in Figure 3. The section shows ameloblastic cells with a strong reac- tion with Con A-HRP on presumptive Golgi zones as well as on Tomes processes (arrows). Reactivity is also observ- able at the apical surface of the odontoblast cells (arrow- heads). d, dentin; e, enamel. Scale bar = 0.6 pm.
Fig. 6. Liolaemus grauenhorsti. Con A-HRP reactiv- ity in teeth in different stages of development. Odonto- blasts (arrowheads) and ameloblasts (arrow) are in- tensely stained. El area indicated by the rectangle is magnified in Figure 7. d, dentin; dp, dental pulp; 0, odontoclasts. Scale bar = 8 pm.
Fig. 4.
Fig. 5.
Fig. 7. Liolaemus gmuenhorsti. Ameloblasts show positive supranuclear granular material (arrows). Bright reaction is shown by the odontoblasts, and a diffuse positivity is also observable by the odontoblastic process (arrowheads). d, dentin; e, enamel. Scale bar = 0.6 pm.
pletely at the ameloblast and odontoblast (Fig. 2). No staining occurs in sections ex- posed to substrate medium alone or to uncon- jugated lectins. Pretreatment of sections with periodic acid abolishes the affinity of the his- tological sites for the lectins.
DISCUSSION
These results show that the content and distribution of the saccharic moieties changes during the development of a lizard's teeth. All regions that stain with Con A also stain with WGA. Lectin histochemistry shows the presence of a-D-mannose (a-D-man), a-D- glucose (a-~-Glc) as well as P-D-N-aCetylgh- cosamine (P-D-GlcNAch and sialic acid at the cell surface and/or at the cytoplasm of the ameloblast and odontoblast cells.
The significance of these saccharic moi- eties is difficult to ascertain. The presence of these oligosaccharides in tooth germs at early and late bell adult of polyphyodont lizards Liolaemus grauenhorsti may indicate that they have some significance in odontogen- esis, inducing and regulating cellular differen- tiation and cell-cell interaction. In fact, cell- surface glycoconjugates, and in particular their constituent carbohydrate moieties, are known to play important roles during cell- cell recognition, interaction, and pinocytosis during embryonic development, inducing cel- lular differentiation in various developing sys- tem, such as the skin (Takata and Hirano, '83)' the kidney (Holthofer and Virtanen, '87), the heart (Faze1 et al., '891, chick em- bryo lingual epithelium (Gheri et al., '911, early chick embryo development (Griffith and Sanders, '91)' human fetal olfactory epithe- lium (Gheri et al., '92), and mouse molar root development (Sasano et al., '92). Require- ments of this type are particularly well illus- trated in the case of tooth formation, in which interactions between the ectomesenchymal components (preodontoblasts) and epithelial cells (preameloblasts) initiate a series of bio- chemical changes that lead to synthesis of dentine matrix by odontoblasts (Koch, '67; Kollar and Baird, '69; Thesleff et al., '90; Ruch, '87). The mechanism of this interac- tion remains unknown in any epidermal or- gan system (Lemus et al., '80; Thesleff et al., '91).
a-D-man, a-~-Glc and (~-D-GLcNAc), and sialic acid are a widely distributed in odonto- blasts and ameloblasts. High lectin-binding capacity in these cells may be due to the
332 D. LEMUS ET AL.
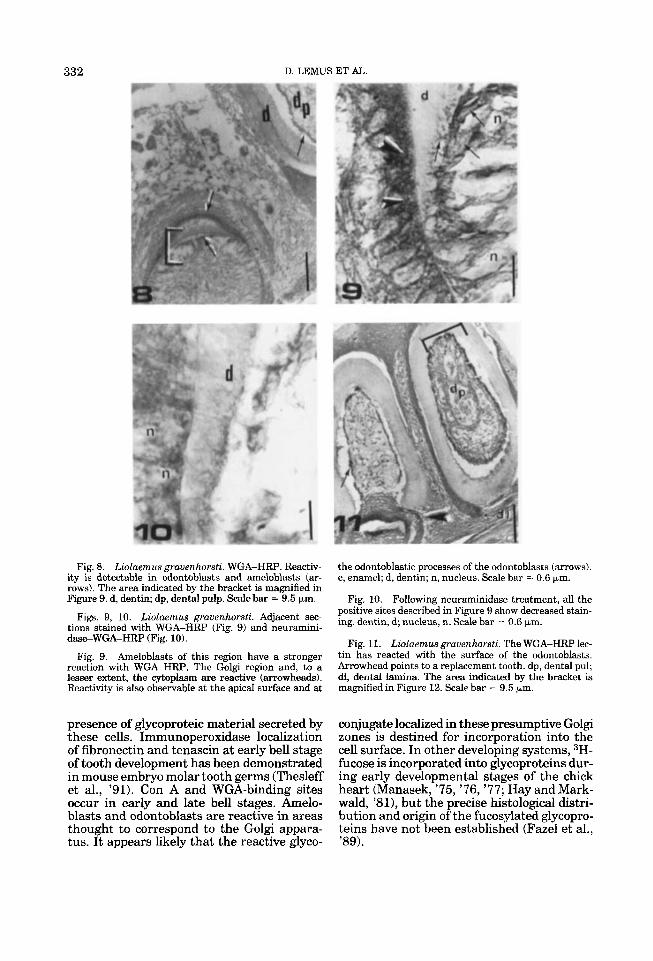
Fig. 8. Liolaemus grauenhorsti. WGA-HRP. Reactiv- ity is detectable in odontoblasts and ameloblasts (ar- rows). The area indicated by the bracket is magnified in Figure 9. d, dentin; dp, dental pulp. Scale bar = 9.5 km.
the odontoblastic processes of the odontoblasts (arrows). e, enamel; d, dentin; n, nucleus. Scale bar = 0.6 pm.
Fig. 10. Following neuraminidase treatment, all the positive sites described in Figure 9 show decreased stain- Figs. 9, 10. Lioluemus grauenhorsti. Adjacent sec- ing, dentin, d; nucleus, n, scale bar = o,6 ~ m , tions stained with WGA-HRP (Fig. 9) and neuramini-
daeWGA-HRP (Fig. 10). Fie 11. Liolaemus grauenhorsti. The WGA-HRP lec- Fig, 9. Ameloblasts of this region have a stronger
reaction with WGA-HRP. The Golgi region and, to a lesser extent, the cytoplasm are reactive (arrowheads). Reactivity is also observable at the apical surface and at
tin 6as reacted with ihe surface of the odontoblasts. Arrowhead points to a replacement tooth. dp, dental pul; dl, dental lamina. The area indicated by the bracket is magnified in Figure 12. Scale bar = 9.5 km.
presence of glycoproteic material secreted by these cells. Immunoperoxidase localization of fibronectin and tenascin at early bell stage of tooth development has been demonstrated in mouse embryo molar tooth germs (Thesleff et al., '91). Con A and WGA-binding sites occur in early and late bell stages. Amelo- blasts and odontoblasts are reactive in areas thought to correspond t o the Golgi appara- tus. It appears likely that the reactive glyco-
conjugate localized in these presumptive Golgi zones is destined for incorporation into the cell surface. In other developing systems, 3H- fucose is incorporated into glycoproteins dur- ing early developmental stages of the chick heart (Manasek, '75, '76, '77; Hay and Mark- wald, '811, but the precise histological distri- bution and origin of the fucosylated glycopro- teins have not been established (Faze1 et al., '89).

SUGAR RESIDUES IN LIZARD TOOTH GERMS 333
Fig. 12. Liolaemus grauenhorsti. Apical portions of the odontoblasts are intensely stained (arrows). Reactivity is also observable at the Tomes processes (arrowheads). d, dentin. Scale bar = 0.6 pn.
Ultrastructural localization of concavalin A binding sites on the surface of odontoblasts demonstrates an apical accumulation of these receptors during polarization in embryonic mouse molar (Meyer et al., '81, cited by Ruch, '87). Sasano et al. ('92), localized glycoconju- gates recognized by Arachis hipogea (PNA) and Maclura pomifera (MPA) lectins during mouse molar root development. PNA pos- sesses high affinity with D-gahCtOSe-( pl-3)-N- acetyl-D-galactosamine residues, whereas MPA does so with residues of D-galactose-(pl- 3)-N-acetyl-D-galactosamine, N-acetyl-D-ga- lactosamine and D-galactose. PNA specifi- cally stains Hertwig's epithelial root sheath (HERS), whereas MPA stained a number of tissues (outermost layer of root dentin, form- ing cellular cementum, alveolar bone and HERS) showed positive reactions with MPA. These glicoconjugates localized by the lectins may be functionally related to molecules with contribute to root formation and cementogen- esis during mouse molar root formation (Sasano et al., '92).
In Liolaemus gravenhorsti, the presence of p-D-GLcNAcIn and sialic acid sugar residues characterized the Golgi region and, to a lesser extent, the cytoplasm of ameloblast cells dur- ing the developmental stages considered. Re- activity was also observed at the apical sur- face and at the odontoblastic processes of the odontoblast cells. By contrast, all the positive sites show decreased staining following siali- dase digestion.
These sugar residues may be involved in- ducing and regulating cellular differentiation and cell-cell interactions in the odontogen- esis of lizards. One residue, sialic acid, may be involved in the early developmental pro- cesses because of its position in the side chain of many glycoconjugates. In fact, glyconju- gates containing sialic acid and fucose have been of particular interest in developmental studies since these sugars invariably occupy a terminal position on oligosaccharide side chain (Bennett et al., '74). Sialic acid is of particular interest in developmental studies. The terminal position of this sugar in the side chain of many structurally defined glyco- conjugates makes them potential candidates as ligands for developmentally regulated rec- ognition molecules such as endogenous lectins, glycosyltransferases, and exoglycosi- dases on the surface of similar or different embryologic cell types (Reutter et al., '82; Faze1 et al., '89).
The present study determined the tissue distribution of glycoconjugates in the tooth germs of lizards during the formation of den- tine and enamel. Sugar residues recognized by Con A and WGA may serve important functions in crown formation. Tooth morpho- genesis and cell differentiation are under the control of reciprocal interactions between the epithelial and mesenchymal tissues. These interactions initiate a series of biochemical changes that lead to the synthesis of dentine by the odontoblasts and of the enamel by the

334 D. LEMUS ET AL.
ameloblasts. Con A-HRP and WGA-HRP- binding components in ameloblasts and odon- toblasts may be functionally related to mol- ecules which are thought to contribute to odontogenesis in polyphyodont lizards. Fur- ther characterization of the other lectin- HRP-binding glycoconjugates may increase understanding of the complex processes re- lated to odontogenesis.
ACKNOWLEDGMENTS
This investigation was supported by grant B-3191-9334 from the DTI, and Vicerrec- toria Academica y Estudiantil, Departamento de Post-Grado y Post Titulo, Universidad de Chile. The authors thank Dr. C. Gans and two anonymous reviewers for their critical review of this manuscript. This paper is a portion of the Master of Biological Sciences thesis (Morphology mention) of Rosa Ca- bello.
LITERATURE CITED
Bennett, G., C.P. Leblond, and A. Haddad (1974) Migra- tion of glycoproteins from the Golgi apparatus to the surface of various cell types as shown by radioautogra- phy after labeled fucose onjection into rats. J. Cell Biol.
Fazel, A.R., H. Sumida, B.A. Schulte, and R.P. Thomp- son (1989) Lectin histochemistry of the embryonic heart: fucose-specific lectin binding in developing rats andchicks. Am. J . Anat. 184:76-84.
Fuenzalida, M., R. Lemus, S. Romero, R. Fernandez- Valencia, and D. Lemus (1990) Behavior of rabbit dental tissues in heterospecific association with embry- onic quail ectoderm. J . Exp. Zool. 256:264-272.
Gheri, G., S. Gheri-Bryk, and V. Petrelli (1990) Histo- chemical detection of sugar residues in the chick em- bryo mesonephros with lectin-horseradish peroxidase conjugates. Histochemistry 95r63-71.
Gheri, G., S. Gheri-Bryk, and E. Sgambati (1991) Use of horseradish peroxitlase conjugated lectins for detection of glycoconjugate changes in developing lingual epithe- lium of the chick embryo. Dev. Growth Diff. 33:371- 377.
Gheri, G., S. Gheri-Bryk, and G.C. Balboni (1992) Identi- fication of sugar residues in human fetal olfactory epithelium using lectin histochemistry. Acta Anat. 145:
Griffith, C.M., and E.J. Sanders (1991) Changes in glyco- conjugate expression during early chick embryo devel- opment: A lectin-binding study. Anat. Rec. 231.238- 250.
Hay, D.A., and R.R. Markwald (1979) Localization of fucose-containing substances in developing atrioven- tricular cushion tissue. In T. Pexider (ed): Perspectives in Cardiovascular Research. Vol. 5: Mechanisms of Cardiac Morphogenesis and Teratogenesis. New York
60~258-284.
16 7-1 74.
Raven Press; pp.-197-211. Holthofer, H., and I. Virtanen (1987) Glycosilation of
developing human glomeruli: lectin binding sites dur- ing cell induction and maduration. J . Histochem. Cyto- chem. 35:33-37.
Hu, C.C., Y. Sakakura, Y. Sasano, L. Shum, P. Bringas, Jr., Z. Werb, and H.C. Slavkin (1992) Endogenous
epidermal growth factor regulates the timing and pat- tern of embryonic mouse molar tooth morphogenesis. Int. J . Biol. 36:505-516.
Koch, W.E. (1967) In vitro differentiation of tooth rudi- ments of embryonic mice. I. Transfilter interactions of embryonic incisor tissues. Anat. Rec. 165:155-170.
Kollar, E., and G. Baird (1970 b) Tissue interactions in embryonic mouse tooth germs. 11. The inductive role of the dental papilla. J. Embryol. Exp. Morphol. 24:173- 186.
Lemus, D. (1967) Tabla de desarrollo del iguanido vi- viparo Liolaenus grauenhorsti. Biologica 40:39-61.
Lemus, D., and R. Duvaucbelle (1966) Desarrollo intra- uterino de Liolaemus tenuis tenuis. Contribucion al estudio del desarrollo embriol6gico de reptiles chilenos. Biologica 39230-98.
Lemus, D., C. Mery, G. Grunert, andM.A. Montenegro (1977) Involution of developing tooth buds in the liz- ard. International Research Communications System Medical Science (IRCS Med. Sci.) 5:442-443.
Lemus, D., Y. Paz de la Vega, M. Fuenzalida, and J. Illanes (1980) In vitro differentiation of tooth buds from embryos and adult lizards Lwlaemus grauen- horsti: an ultrastructural comparison. J. Morphol. 165: 225-236.
Lemus, D., J. Illanes, M. Fuenzalida, Y. Paz de la Vega, and M. Garcia (1981) Comparative analysis of the development of the lizard Liolaenw tenuis tenuis. 11. A series of normal postlaying stages in embryonic devel- opment. J . Morphol. 169:337-349.
Lemus, D., M. Fuenzalida, J . Illanes, and Y. Paz de la Vega (1983) Ultrastructural aspects of dental tissues and their behavior in xenoplastic association (lizard- quail). J . Morphol. 176:341-350.
Lemus, D., L. Coloma, M. Fuenzalida, J . Illanes, Y. Paz de la Vega, A. Ondarza, and M.J. Blanquez (1986) Odontogenesis and amelogenesis in interacting lizarcL quail tissue combinations. J . Morphol. 189t121-129.
Manasek, F.J. (1975) The extracellular matrix of the early embryonic heart. In M. Lieberman and T. Sano (eds): Developmental and Physiological Correlates of Cardiac Muscle. New York, Raven Press, pp. 1-20.
Manasek, F.J. (1976) Glycoprotein synthesis and tissue interactions during establishment of the functional embryonic chick heart. J. Mol. Cell Cardiol. 8t389402.
Manasek, F.J. (1977) Structural glycoproteins of the embryonic cardiac extracellular matrix. J. Mol. Cell Cardiol. 9:425-439.
Mina, M., and E.J. Kollar (1987) The induction of odonto- genesis in non-dental mesenchyme combined with early murine mandibular arch epithelium. Arch. Oral Biol. 32: 123-127.
Reutter, W., E. Kottgen, C. Bauer, and W. Gerok (1982) Biological significance of sialic acids. In R. Schauer (ed): Cell Biology Monographs. Vol. 10: Sialic Acids. Wen: Springer-Verlag, pp. 263-305.
Ruch, J.V. (1987) Determinism of odontogenesis. In E. Barberg-Guillem (ed): Cell Biology Reviews. Vol. 14. Vizcaya, Spain: Leioa pp. 1-112.
Sasano, Y., I. Mizoguchi, M. Kagayama, L. Shum, P. Bringas, Jr., and H.C. Slavkin (1992) Distribution of type I collagen, type I1 collagen and PNA binding glycoconjugates during chondrogenesis of three dis- tinct embryonic cartilages. Anat. Embryol. 186:205- 213.
Sasano, Y., Y. Kaji, M. Nakamura, K. Kindaichi, H.C. Slavkin, and M. Kagayama (1992a) Distribution of glycoconjugates localized by Peanut and Maclurapomlfi era agglutinins during mouse molar root development. Acta Anat. 145:149-155.

SUGAR RESIDUES IN LIZARD TOOTH GERMS 335
Slavkin, H.C. (1974) Embryonic tooth formation. In A.H. Melcher and G.A. Zarb (eds): Oral Sciences Reviews. Vol. 4. Toronto: pp. 1-136.
Slavkin, H.C. (1993) Rieger syndrome revisited: experi- mental approaches using pharmacologic and antisense strategies to abrogate EGF and TGF-alpha functions resulting in dysmorphogenesis during embryonic mouse craniofacial morphogenesis. A. J. Med. Genet. 47r689- 697.
Takata, K., and H. Hirano (1983) Changes in soybean agglutinin (SBA) and peanut agglutinin (PNA) binding pattern in the epidermis of the developing chick em- bryo. Dev. Growth Diff. 25.299-305.
Thesleff, I., A. Vaahtokari, and S. Vainio (1990) Molecu- lar changes during determination and differentiation of the dental mesenchymal cell lineage. J. Biol. Buccale 18r179-188.
Thesleff, I., A.M. Partanen, and S. Vainio (1991) Epithe- lial-mesenchymal interactions in tooth morphogen- esis: the roles of extracellular matrix, growth factors, and cell surface receptors. J. Craniofac. Genet. Dev. Biol. 11:229-237.
Zeichner-David, M., M. MacDougall, S. Yen, F. Hall, and H.C. Slavkin (1992) Protein kinases in dentinogenesis. Proc. Finn. Dent. SOC. 88:295-303.





























