Embed Size (px)
Citation preview

ELSEVIER 0024-3205(95)02305-4
Life Sciences, Vol. 58, No. 5, pp. 405414, 1996
Copyright Q 1995 Elsevia Science Inc. Printed in the USA. All rights reserved
0024-3205/96 $15.00 + .oO
DESENSITIZATION EFFECT OF IN VIVO TREATMENT WITH METAPROTERENOL
ON p,, p2 AND Pa-ADRENERGIC RESPONSIVENESS IN RAT ADIPOCYTES
Maria Puy Portillol, Antonio S. Del Barrio 1, Maria Angeles Garcia-Calongel and
J. Alfred0 Martinez2
Dep. Nutrition and Food Science, University of Pais Vasco, 01006 Vitoria,Spainl
Dep. Physiology and Nutrition, University of Navarra, 31008 Pamplona, Spain2
(Received in final form November 10, 1995)
Summary
The desensitization process of j%adrenergic system was assessed by in vivo administration to 7-week old rats of a mixed P-agonist, metaproterenol (3,5-dyhydroxyphenyl-N-isopropyl-amine-P-ethanol sulphate; T1/2=6 hours), (2 mg/kg/d) in treatments of 12 hours, 2 days and 10 days. The in vitro lipolytic effect of selective j3-adrenergic agonists, dobutamine, salbutamol and BRL 37344, as well as plasma free fatty acid concentrations were measured in treated and control animals given vehicle. Different times of exposure to a P-agonist induced a loss of responsiveness on lipolytic response mediated by 61 and j32-adrenoceptors, as demonstrated by decreased affinity and intrinsic activity (maximal effect) of dobutamine and salbutamol. In contrast, no changes were found in Ps-mediated lipolysis. These observations suggest that 61, p2 and P3-adrenoceptors follow different regulatory patterns. Lack of j33-adrenoceptor desensitization may have important physiological and therapeutic consequences in the treatment of diseases such as obesity and heart failure.
Key Words: p-adrcnoceptors, adipose tissue, dcscnsiriyation, lipolysis, mctaproterenol
The prolonged exposure of cells to 6-adrenergic agonists produces a decrease of responsiveness to stimulation with time. This phenomenon, which is referred as desensitization, tachyphylaxis or refractoriness has been observed in either in vivo and/or in vitro studies (1, 2).
Different processes are believed to contribute to this phenomenon: a reversible alteration in the state of the receptor, so that it is not able to activate adenylate-cyclase, which is referred as an uncoupling state (3, 4) and an apparent loss in the number of j3-adrenoceptors (ARs), which is referred to as down-regulation process could be the most important (5).
All correspondence to: Dra. Maria del Puy Portillo. Nutrition. Facultad de Farmacia. Universidad del Pais Vasco. Marques de Urquijo s/n. 01006 VITORIA (SPAIN).

406 Lipolytic Response after Metaprotercnol Vol. 58, No. 5, 1996
Studies of the time course of multiple regulation steps and the major enzymes involved in desensitization have been reported in detail mainly for j12-ARs (6,7). In contrast to the well documented desensitization of the f3*-AR, less information concerning j3l-ARs is available (8, 9). A different regulation of j31 and j32-AR subtypes has been proposed in several studies (10, 11, 12). In the earlier studies on mammal species, the existence of an additional j3s-AR subtype involved in the control of lipolysis was poorly considered in the extrapolation of the regulatory process.
The desensitization process has an essential physiological relevance, it avoids hyperstimulation of target cells (7). On the other hand, desensitization is particularly significant because it can be the basis for tolerance to effects of drugs used in the treatment of a variety of diseases, such as obesity, where long-term treatments could be needed (13). So, the clinical use of those drugs in long-term treatment may be of limited value because tolerance to their action will develop (14, 15). In this context, several strategies have been proposed to deal with this process. An example is intermittent administration of P-AR antagonists, which allows up-regulation of P-ARs and therefore, more effectiveness of subsequent agonist therapy (16). Also it is of great importance to find drugs that are capable to prevent desensitization process accompanying long-term treatment with P-AR agonists (15).
The aim of the present study was to assess the in vitro lipolytic response from isolated rat adipocytes to j3,-selective (dobutamine), b2-selective (salbutamol) and j3s-selective (BRL 37344) adrenergic agonists after different in viva treatment periods with the mixed P-agonist metaproterenol (3,5-dyhydroxyphenyl-N-isopropyl-amine-P-ethanol sulphate; T112 = 6 hours), as well as to compare the desensitization process for PI, p2 and j3s-ARs, through lipolysis measurements. Evaluation of desensitization in intact animals is of interest because data obtained from isolated adipocytes cannot be extrapolated to long-term treatment under in vivo conditions.
Methods
Animals Specific pathogen free (SPF) female outbred Wistar rats, obtained from Charles River (Spain), weighing about 225 g (7 weeks old) were used after 3 days of acclimatation. NIH guidelines for care and use of laboratory animals were followed. Rats were randomly allocated into 4 groups of 7 animals each: control (vehicle), a 12-hour metaproterenol treatment (2 mg/kg body weight), a 2-day metaproterenol treatment (2 mg/kg body weight) and a lo-day metaproterenol-treatment (2 mg/kg body weight). Compounds or vehicle (NaCI 9 g/L) were administered subcutaneously in a single dose for the 12-hour treated group (9 a.m.) and two doses in the 2-day and lo-day treated groups (9 a.m. and 16 p.m.). At the end of the treatment period, rats were fasted overnight with free access to water and then killed by cervical dislocation. Blood was immediately collected and epididymal adipose tissue was quickly removed.
Adipocvtes isolation and lipolytic exoeriments Isolated fat cells were obtained according to the method of Rodbell (17) with minor modifications (18) by collagenase digestion (1 mg/mL; 37” C) from rat perirenal adipose tissue in Krebs Ringer Bicarbonate buffer, containing 3.5 g/100 mL of bovine serum albumin (BSA V) and 0.6 mmol/lOO mL of glucose at pH 7.4 (KRBA).
Under our experimental conditions, isolated rat fat cells were obtained after 60 minutes of incubation. Fat cells were filtered through nylon mesh and washed twice with the same incubation buffer (KRBA). Measurements of lipolytic activity were performed by incubating isolated adipocytes (20-30 mg total lipid) in 1 mL of KRBA

Vol. 58, No. 5, 1996 Lipolytic Response after Metaproterenol 407
buffer. After 90 minutes of incubation with dobutamine, salbutamol and BRL 37344 (10-s M to 10-4 M) at 37” C, the reaction was stopped with ice and an aliquot (200 uL) was taken to determine glycerol release in the incubation buffer by the method of Wieland (19) with minor modifications (18). The metabolic activity was expressed as micromoles of glycerol released per 100 mg of total lipid, which were determined gravimetrically after the extraction, according to the method of Dole and Meinertz (20). Plasma concentrations of non-esterified free fatty acids (NEFA) were evaluated using an enzymatic method.
Dobutamine was purchased from Lilly (Alcobendas, Spain). Salbutamol and BRL 37344 were generous gifts from Glaxo (Aranda de Duero, Spain) and Smithkline Beecham (Madrid, Spain). Bovine serum albumin (fraction V) was obtained from Sigma Chemical Co (St. Louis, MO). Crude collagenase (0.52 U/mg) and free fatty acid kit were supplied by Boehringer-Mannheim (Barcelona, Spain). All other chemicals were reagent grade.
Calculations and statistical analysis Concentration-response curves (n= 7) for each P-agonist were used to determine an EC50 value (M) as the concentration of agonist causing 50% of its maximal response. These EC50 values were used to calculate pD2 values (- log E&o). Statistical analysis was performed using analysis of variance (ANOVA test); when ANOVA yielded significant values (confidence intervals 95%) variables were compared by Duncan’s t test. Differences between groups with a p value less than 0.05 were considered as statistically significant.
In order to study the possibility of 6-adrenegic system desensitization after different periods of in viva treatment with the mixed P-agonist metaproterenol, we evaluated the in vitro lipolytic response to different P-agonists from isolated adipocytes. We also measured plasma free fatty acid concentrations in control and treated animals.
TABLE
Affinity of the Assayed 8-Adrenergic Agonists, Expressed as pD2 values
in vivo metaproterenol treated groups
@Agonist -___ -p--
control 12 hours 2 days 10 days _~__~~_~_____~__
Dobutamine 6.3 & 0.04 a 6.2 f 0.06 a 6.4 + 0.05 a 5.7 + 0.04 b Salbutamol 6.1 f0.04a 5.4 f 0.07 b 5.3 4 0.05 b 5.2 f 0.04 b BRL 37344 8.8+O.lOa 8.8f0.12a 9.1 + 0.09 a 8.7 f 0.08 a
pD2 values of the S-agonists were estimated from each concentration-response curve. The results are meansfSE of 7 separate experiments. Statistical analysis was performed using ANOVA test and Duncan’s t test. For each P-agonist values not sharing a common letter are statistically different (~~0.05).
408 Lipolytic Response after Mctaproterenol Vol. 58, No. 5, 1996
3- 0 B ,a .
i-z 2- 8 z . e 8 g l-
a _
E,
0 ! I I I I I
-9 -a -7 -6 -5 -4
-LOG [SALBUTAMOL] (M)
-12 -10 -0 -6 -4
-LOG [BRL 373441 (M)
FIG.1
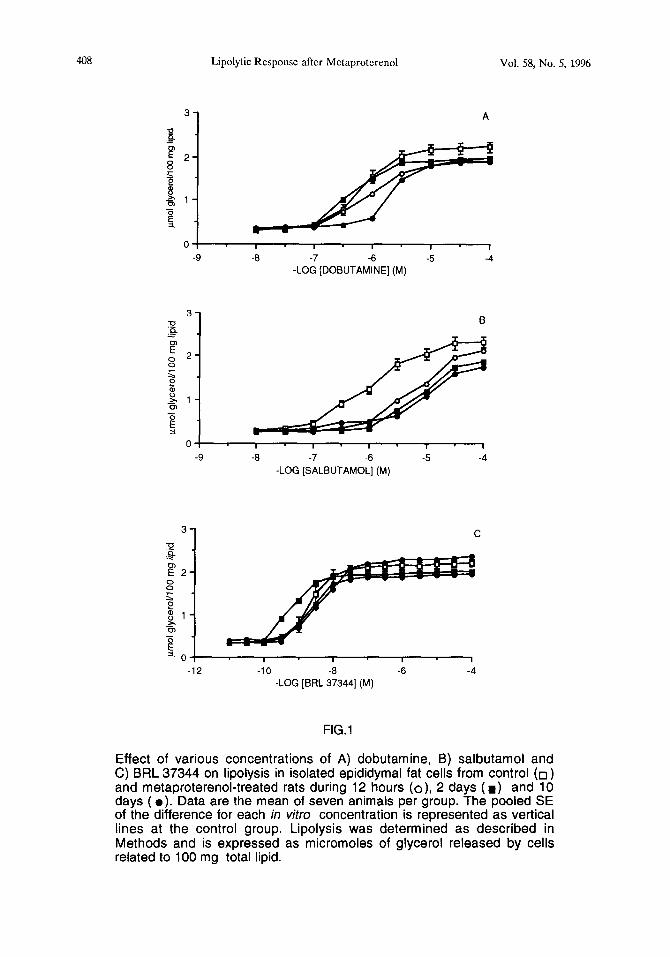
Effect of various concentrations of A) dobutamine, B) salbutamol and C) BRL 37344 on lipolysis in isolated epididymal fat cells from control (0) and metaproterenol-treated rats during 12 hours (o), 2 days ( W) and 10 days ( l ). Data are the mean of seven animals per group. The pooled SE of the difference for each in vitro concentration is represented as vertical lines at the control group. Lipolysis was determined as described in Methods and is expressed as micromoles of glycerol released by cells related to 100 mg total lipid.
Vol. 58, No. 5, 1996 Lipolytic Response after Mctaprotcrenol 409
As shown in FIG.lA, different in vivo treatment periods produced changes in the concentration-response curves of in vitro dobutamine-induced lipolysis. There is a shift towards the right in treated rat adipocytes compared with the controls.
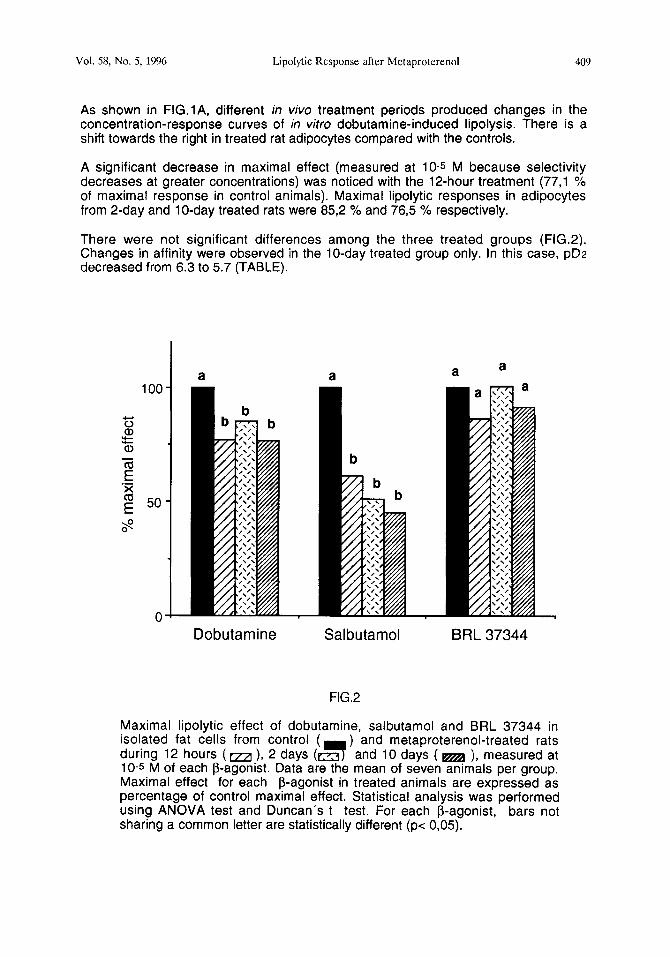
A significant decrease in maximal effect (measured at 10-s M because selectivity decreases at greater concentrations) was noticed with the 12-hour treatment (77,l % of maximal response in control animals). Maximal lipolytic responses in adipocytes from 2-day and lo-day treated rats were 852 % and 765 % respectively.
There were not significant differences among the three treated groups (FIG.2). Changes in affinity were observed in the lo-day treated group only. In this case, pD2 decreased from 6.3 to 5.7 (TABLE).
0
a a
-
Dobutamine Salbutamol BRL 37344
FIG.2
Maximal lipolytic effect of dobutamine, salbutamol and BRL 37344 in isolated fat cells from control (
-7 ) and metaproterenol-treated rats
during 12 hours (m ), 2 days (e;3 and 10 days ( m ), measured at 10-s M of each P-agonist. Data are the mean of seven animals per group. Maximal effect for each P-agonist in treated animals are expressed as percentage of control maximal effect. Statistical analysis was performed using ANOVA test and Duncan’s t test. For each P-agonist, bars not sharing a common letter are statistically different (p< 0,05).
410 Lipolytic Response after Mctaproterenol Vol. 58, No. 5, 1996
Following the same in viva treatments, there was a greater loss of Ps-adrenergic responsiveness, because concentration-response curves of salbutamol-induced lipolysis were more shifted towards the right (FIG.lB). In fact, decrease of maximal effects (61,3 %, 51,3 % and 45,l % for each treatment vs control) was larger than those observed in response to dobutamine (FIG. 2). Also there were changes in affinity, which follow a different modification pattern from the dobutamine study. In this case, the decrease of pD2 was already observed after a 12-hour treatment. More prolonged in vivo exposure to metaproterenol did not lead to an additional decrease of in vitro lipolytic salbutamol affinity in rat adipocytes. Thus, pD2 decreases from 6.1 to 5.2 (TABLE).
We have reported the effects of some classical b-agonists. In contrast to the above-mentioned changes, there was not significant modifications in BRL 37344 concentration-response curves (FIG.lC), nor pD2 (TABLE) nor maximal effect (FIG.2), suggesting no alteration of “b-atypical”-adrenergic responsiveness in rat fat cells.
b b
FIG.3
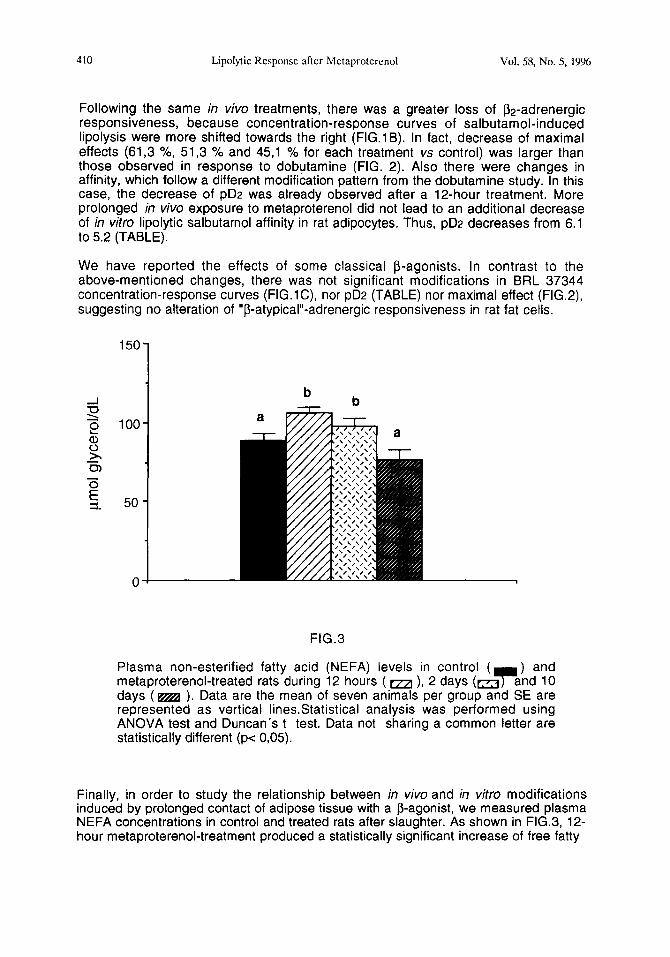
Plasma non-esterified fatty acid (NEFA) levels in control ( 7
) and metaproterenol-treated rats during 12 hours ( aa ), 2 days (m and 10 days (m ). Data are the mean of seven animals per group and SE are represented as vertical linesStatistical analysis was performed using ANOVA test and Duncan’s t test. Data not sharing a common letter are statistically different (pe 0,05).
Finally, in order to study the relationship between in viva and in vitro modifications induced by prolonged contact of adipose tissue with a P-agonist, we measured plasma NEFA concentrations in control and treated rats after slaughter. As shown in FIG.3, 12- hour metaproterenol-treatment produced a statistically significant increase of free fatty

Vol. 58, No. 5, 1996 Lipolytic Response after Metaprotcrenol 411
acids in plasma (106 umol/dL vs 88,3 umol/dL) which slightly diminished prolonging the treatment up to 2 days (98,0 umol/dL). In contrast, following a lo-day treatment plasma free fatty acids fell down near the control levels (76,5 umol/dL).
Discussion
The three subtypes of j3-ARs are involved in the regulation of lipolytic process in rats (7, 18, 21), so that rat adipose tissue and in vitro lipolytic measurements are good tools to carry out desensitization studies of P-adrenergic system. Many desensitization studies with P-agonists have been performed in vitro with isolated cells (9, 22, 23, 24). In contrast, the consequences of chronic in vivo exposure to j3-agonists are not as well characterized. Examination of desensitization in intact animals is of interest because it is difficult to extrapolate from information obtained in isolated fat cells, after a long-term P-agonist exposure, to the more complex intact organism (8). So, for this study we used intermittent treatment with a high dose of the mixed P-agonist metaproterenol (2 mg/kg BW). This process could be partially masked due to a possible recovery from desensitization between the last injection (16 p.m.) and use of fat cells of animals.
Metaproterenol is a mixed P-agonist which binds the three subtypes of P-adrenoceptors, although a major affinity to P2-ARs than to 8,-A& has been found (25). This p-agonist has been selected due to previous protein and lipid metabolism impacts found in previous studies (26,27,28,29). In the literature is shown that in vitro lipolytic effects of high doses of a mixed P-agonist are not antagonized by high doses of selective pi and p2 antagonists (31,32).
However, in in vivo studies using intermittent bolus doses of P-agonists, it is difficult to assess whether suitable levels of the infused drug reached a in situ concentration in white adipose tissue for activation of fat cell j3-ARs, because of wide fluctuations in blood can occur (33).
Our results indicate that j31 and @ARs coupled to adenylate-cyclase system are desensitized in rat adipocytes following exposure to metaproterenol. The rapid agonist mediated desensitization observed in our conditions is reflected by decreased maximal stimulation of in vitro lipolysis (intrinsic activity). Also, a decreased affinity of j31 and fin-agonists was observed as shown by the shift towards the right of the concentration-response curves and pD2 values. As shown, P2-adrenergic loss of responsiveness is greater than PI because decreases of maximal j32-adrenergic lipolytic effect are larger. Furthermore, salbutamol affinity disminished after a 12-hour treatment, whereas dobutamine affinity only decreased with the longest treatment.
These results are in good agreement with other studies performed in different conditions and species. In this way, Valet et el. (10) reported a strong decrease in procaterol induced lipolysis from dog adipocytes exposed to the sustained stimulation of the sympathetic nervous system that occurs after sino-aortic denervation. A similar phenomenon was observed in human adipose tissue after in situ administration of catecholamines (11). Also, prolonged in vivo infusion of isoproterenol in rats has been shown to induce a down-regulation of > 70 % of the heart j32-ARs with a concomitant reduction of c 40 % of pi-ARs (30). Similarly, Suzuki et al. (9) found a much lower down-regulation of PI-ARs and of minor amplitude than that of the 82-ARs from transfected Chinese hamster fibroblasts (CHW) incubated with isoproterenol.

412 Lipolytic Response after Metaprotercnol Vol. 58, No. 5, 1996
As indicated previously, the possible desensitization pattern of P3-ARs has been poorly studied. In contrast to 81 and j32-adrenergic responses, in the present report, agonist stimulation of P3-ARs by BRL 37344 was not affected by different times of in viva exposure to metaproterenol, because no differences in its maximal response and affinity were found. Some studies failed to find a loss of p3 responsiveness in different conditions. Granneman (34) observed that treatment of isolated adipocytes with isoproterenol greatly reduced activation of adenylate-cyclase by j31-ARs but not by j3s-ARs. Nantel et al. (24) showed that the human @-AR expressed in CHW is much less prone to rapid agonist-mediated desensitization than j%AR. Carp&e et al. (35) obtained similar results in studies performed in intact animals after norepinephrine infusions.
Although the molecular basis for the desensitization of j31-ARs and j32-ARs was not addressed in this study, in the published literature different specific mechanisms underliyng this phenomenon have been postulated, one or more of which may be involved in given dependent conditions: a) loss of the number of receptors -down regulation- (5) b) uncoupling of the receptor from GTP binding protein (Gs) due to a P-AR phosphoylation in consensus regions located adjacent to regions of the receptor proposed to be involved in coupling to the Gs (4,36), c) modification of adenylate-cyclase enzyme (37), d) increase in the activity of phosphodiesterase (3,38). Differences in the primary sequence of the three subtypes of 8-ARs could explain, at least in part, their differential desensitization.
The in vitro results of this work are in good accordance with data of in viva effects of metaproterenol on plasma NEFA concentrations, which were increased on 12-hour and 2-day treated groups. This effect is easily explained because of the lipolysis stimulation by the administered P-agonist. The decrease of those plasma levels in 2- day treated rats vs those of 12-hour treated group could be related to loss of 81 and p2 responsiveness, demonstrated by our in vitro lipolytic experiments. The longest treatment (10 days) induced no changes in plasma NEFA vs controls. Probably, the strong desensitization of classical j3-adrenergic response avoids to detect, by blood biochemical determinations, the remaining 82-adrenergic-mediated lipolysis.
Desensitization of 8-ARs mediated responses poses a serious therapeutic problem in the treatment of obesity and heart failure, so its prevention would be of great therapeutic value (39). As different P-AR subtypes are regulated differently, it may be possible to adopt strategies to overcome desensitization (40,41,42).
@-AR may represent the physiological receptor for noradrenaline secreted from sympathetic nerve endings when noradrenaline concentrations in the synaptic cleft are very high and/or when the high affinity j3-ARs are desensitized by prolonging adrenergic stimulation (43,44). However, elucidation of the molecular mechanism involved in the possible down-regulation and biological consequences requires further investigation.
Acknowledaements
The authors are grateful to Glaxo and Smithkline Beecham laboratories for the generous gift of salbutamol and BRL 37344 respectively. We also acknowledge the finantial support from CICYT (GAN-91/C02), UPV/EHU (101.123-EA 152/92) and Government of Pais Vasco (Euskadi-Aquitania Cooperation/Nutrition and Health/94- 96).

Vol. 58, No. 5, 1996 Lipolytic Response after Metaproterenol 413
References
1. W. P. HAUSDORFF, M. G. CARON and R. J. LEFKOWITZ, Faseb J. 4 2881-2889 (1990).
2. R. J. LEFKOWITZ, W. P. HAUSDORFF and M. G. CARON, Trends Pharmacol. Sci. 11 190-194 (1990).
3. V. HOMBURGER, M. LUCAS, B. CANTAU, J. BARABE, J. PENIT and J. BOCKAERI, J. Biol. Chern.255 10436-10444 (1980).
4. R. L. H UGANIR and P. GREENGARD, Neuron 5, 555-567 (1990). 5. Y. F. HU, K. R. PURUSHOTHAN, P .L. WANG, R. DAWSON and
M. G. HUMPHEYSBEHER, Am. J. Physiol. a G433-G443 (1994). 6. E. BENOVIC, MAYOR, C. STANISZEWSKI, R.J. LEFKOWITZ and M.C. CARON, J.
Biol. Chem. m 9026-9032 (1987). 7. M. LAFONTAN and M. BERLAN, J. Lipid Res. 34 1057-1092 (1993). 8. M.L. HEREPATH and K.J. BROADLEY, Eur. J. Pharmacol. 215 209-219 (1992). 9. T. SUZUKI, C. T. NGUYEN, F. NANTEL, H. BONIN, M. VALIQUET-TE, T. FRIELLE
and M. BOUVIER, Mol. Pharmacol. 41542-548 (1992). 10. P. VALET, J. L. MONTASTRUC, M. BERLAN, M. A TRAN, M. LAFONTAN
and P. MONTASTRUC, J. Pharmacol. Exp. Ther. 249 271-277 (1989). 11. P. ARNER, E. KRIEGHOLM and P. EGFELDT, J. Pharmacol. Exp. Ther. 252
317-322 (1991). 12. J.G. GRANNEMAN, and K.N. LAHNERS, Endocrinology m 109-l 14 (1992). 13. J. P. REVELLI, P. MUZZIN and J.P. GIACOBINO, Biochem. J. m 743-746 (1992). 14. H. S. NELSON, S. J. SZEFLER and R. J. MARTIN, Am. Rev. Resp. Dis. 144
249-250 (1991). 15.0. E. BRODDE, S. PETROSH, H. J. BAUCH, A. PAUL, M. GRADT, D. OEFLER
and M.C. MICHEL, J. Cardiol. Pharmacol. a 434-439 (1992). 16. C. R. JONES, P. MOLENAAR and R. J. SUMMERS, J. Mol. Cell. Cardiol. 21
519-535 (1989). 17. M. RODBELL, J. Biol. Chem. a 375-380 (1964). 18. D. LANGIN, M. P. PORTILLO, J. S. SAULNIER-BLACHE and M. LAFONTAN,
Eur. J. Pharmacol. 199291-301 (1991). 19. 0. WIELAND, Biochem. Zeitung 239 313-319 (1957). 20. V.P. DOLE and H. MEINERTZ, J. Biol. Chem. 235 2595-2599 (1960). 21. M. SANO, T. YOSHIMASA, T. YAGURA and I. YAMAMOTO, Life Sci. 52
1063- 1070 ( 1993). 22. P. ARNER, Am. J. Clin. Nutr. s 228S-2368 (1992). 23. T. W. BURNS, P. E. LANGLEY, B. E. TERRY and D.B. BYLUND Metabolism 31
288-293 (1982). 24. F. NANTEL, H. BONIN, L. J. EMORINE, V. ZILBERFARB, A. STROSBERG,
M. BOUVIER and S. MARULLO, Mol. Pharmacol. g 548-555 (1993). 25. H. C. GILLIES, H. J. ROGERS, R. G. SPECTOR and J. R. TROUNCE, A
Textbook ofTClinical Pharmacolgl! H. C. Gillies, H. J. Rogers, R. G. Spector and J.R.Trounce (Eds), 467-481, Habder and Stoughton, London (1986).
26. M. P. PORTILLO, J. A. MARTINEZ and J. LARRALDE, Reprod. Nutr. Dev. 31 509-519 (1991).
27. J. A. MARTINEZ, M. P. PORTILLO and J. LARRALDE, Horm. Metab, Res. a 594-599 (1991).
28. A. S. DEL BARRIO, M. RODRIGUEZ-MARISCAL, J. A. MARTINEZ and J. LARRALDE, Growth Dev. Aging 56 141-l 48 (1992).
29. M. RODRIGUEZ-MARISCAL, A. S. DEL BARRIO, J. LARRALDE and J. A. MARTINEZ, Experientia 49 308-312 (1993).

414 Lipolytic Response after Mctaprotcrcnol Vol. 58, No. 5, lW6
30. M. P. PORTILLO, A. S. DEL BARRIO and J. A. MARTINEZ, J. Physiol. Biochem. fl (in press) (1995).
31. C. HOLLENGA and J ZAAGSMA, Br. J. Pharmacol. 981420-1424 (1989). 32. I. VAN LIEFDE, A.V. WITZENBURG and G. VAUQUELIN, J. Pharmacol. Exp. Ther.
m 552-558 (1992). 33. G. TSUJIMOTO, W. M. MANGER and B. B. HOFFMAN, Endocrinol.
114 1272-1278 (1984). 34. J.G. GRANNEMAN, J. Pharmacol. Exp. Ther. a 638-642 (1992). 35. C. CARPENE, M.C. REBOURCET, C. GUICHARD, M. LAFONTAN an M. LAVAU,
J. J. Pharmacol. Exp. Ther. a 237-247 (1993). 36. M. J. LOHSE, J.L. BENOVIC, M.C. CARON and R.J. LEFKOWITZ, J. Biol. Chem.
m 3202-3209 (1990). 37. M.G. MURPHY and M. HORACKOVA, Eur. J. Pharmacol. m 121-128 (1994). 38. R . J. LEFKOWITZ and R. J. WILLIAMS, Advances in Cyclic Nucleotide
Research, W. J. George and L. J. lgnarro (Eds), 1-17, Raven Press, New York (1978).
39. G. PELA, C. MISSALE, R. RADINO, E. CONDORELLI, P. F. SPAN0 and 0. VISIOLI, J. Cardiovasc. Pharmacol. E 839-846 (1990).
40.0. E. BRODDE, S. SCHULER, R. KRETSCH, M. BRINKMANN, H. G. BORST, R. HETEER, J. C. REIDEMEISTER, H. WARNECKE and H. R. ZERKOWSKI, J. Cardiovasc. Pharmacol. 8 1235-1242 (1986).
41. M. R. BRISTOW, R. GINSBURG, V. UMANS, M. FOWLER, M. MINOBE, W. RASSMUSSEN, R. ZERA, P. HENLOVE, R. SHAN, P. JAMIESON and E.B. STINSON, Cir. Res. % 297-309 (1986).
39. T. YOSHIDA, N. SAKANE, Y. WAKABAYASHI, T. UMEKAWA and M. KONDO, Life Sci. 5fl. 491-498 (1994).
42. P. MUZZIN, J. P. REVELLI, F. KUHNE, J. D. GOCAYNE, W. R. McCOMBIC, J. C. VENTER, J.P. GIACOBINO and CM. FRASSER, J. Biol. Chem.
m 24053-24058 (1991). 43. L. J. EMORINE, S. MARULLO, M. N. BRIEND-SUTREN, G. PATCY, K. M
TATE, C. DELAVIER-KLUTCHKO and A.D. STROSBERG, Science 245 1118-l 12 (1989).





























