Embed Size (px)
Citation preview

Drug Development Research 8:287-298 (1986)
Cytochrome P-450: Target for ltraconazole Hugo Vanden Bossche, Dany Bellens, Willy Cools, Jos Gorrens, Patrick Marichal, Hugo Verhoeven, Gustaaf Willemsens, Roland De Coster, Dominiek Beerens, Christian Haelterman, Marie-Claire Coene, William Lauwers, and Ludo Le Jeune
Laboratories of Comparative Biochemistry, (H. V. B., D. B., WC., J.G., P. M., H. V , G. W), Endocrinology (R. D. C., D. B., C. H.), Immunology (M.-C. C.), and Analytical Department (W. L., L. L. J.), Janssen Pharmaceutica Research Laboratories, Beerse, Belgium
ABSTRACT
Vanden Bossche, H., D. Bellens, W. Cools, J. Gorrens, P. Marichal, H. Verhoeven, G. Willemsens, R. De Coster, D. Beerens, C. Haelterman, M.4 . Coene, W. Lauwers, and L. Le Juene: Cytochrome P-450: Target for itraconazole. Drug Dev. Res. 8:287- 298, 1986.
The N-substituted triazole, itraconazole, has high affinity for the cytochrome P-450 (cyt. P- 450) isozyme involved in the 14a-demethylation of lanosterol in Candida albicans micro- somes. Fifty per cent inhibition was already observed at itraconazole concentations < 5 x lo- ’ M. Higher concentrations (> lo-’ M) of this antifungal are needed to interfere with the 14 a-demethylation in mammalian cells. Unlike ketoconazole, itraconazole does not significantly affect in vitro androgen, gluco- and mineralocorticoid steroidogenesis. ltraconazole also does not affect the cyt. P-450-dependent 19-hydroxylation of testoster- one, a step in the conversion of androgens to estrogens. The l-hydroxylation of testoster- one by pig testes microscomes is only slightly inhibited. It is hypothesized that itraconazole’s selective activity on ergosterol biosynthesis is due to its high affinity for the apoprotein of the C. albicans cyt. P-450 involved in the 1401- demethylation of lanosterol.
Key words: androgens, estrogens, glucocorticoids, ketoconazole, rnineralocorticoids, sterols
INTRODUCTION
Cytochrome P-450 (cyt. P-450) isozymes are hemoproteins that possess catalytic activity towards lipid-soluble substrates including a large number of endobiotics such as fatty acids,
Received final version March 3,1986; accepted March 3, 1986.
Address reprint requests to Hugo Vanden Bossche, Laboratory of Comparative Biochemistry, Janssen Pharmaceutica Research Laboratories, Turnhoutseweg 30, B-2340 Beerse, Belgium.
0 1986 Alan R. Liss, Inc.

288 Vanden Bossche et al.
sterols and steroids. The term P-450 derives from the observation that this cytochrome has an unusual absorption spectrum with a peak at about 450 nrn when the iron atom is reduced and sixth coordination position is occupied by carbon monoxide [White and Coon, 19801.
In yeast and fungi a cyt. P-450 isozyme is involved in the l4a-demethylation of lanosterol, a key step in the biosynthesis of ergosterol. It has been shown previously that N- substituted imidazole and triazole antifungal agents, including itraconazole, interfere at low concentrations (< 5 X M) with this microsomal cyt. P-450-dependent lanosterol 14a- demethylase system. Inhibition of this enzyme results in the depletion of ergosterol and a concomitant accumulation of l4a-methylsterols. Consequently this produces alterations in membrane functions and in the activity of membrane bound enzymes [Vanden Bossche, 19851. It was also found that azole derivatives, such as ketoconazole and itraconazole, when added to cyt, P-450 containing suspensions of Saccharomyces cerevisiae and Candida albicans micro- somes, occupy the sixth coordination position of the iron atom in the protoporphyrin moiety of cyt. P-450 [Vanden Bossche and Willemsens, 1982; Vanden Bossche, et al., 1985al.
In the present study the competition between carbon monoxide and the N-substituted azole derivatives N-phenylimidazole, ketoconazole, and itraconazole is studied in a microsomal fraction of C. albicans. The interaction of itraconazole with cyt. P-450 isozymes in microsomal and mitochondrial membranes of adrenals, liver, and testes was also studied.
The affinity of itraconazole for the cyt. P-450 involved in the 14a-demethylation of lanosterol was studied by measuring its effects on sterol synthesis in C. albicans and human lymphocytes. The low affinity of this triazole derivative for cyt. P-450 isozymes involved in testicular and adrenal steroidogenesis was proven by studying its effects on the hydroxylation reactions in sub-cellular fractions of bovine adrenal cortex and pig testes and in dispersed rat testicular cells.
MATERIALS AND METHODS Cyt. P-450 lsozyrnes
The isolation of microsomes from C. albicans (strain ATCC 28516) and of mitochondria and microsomes from mammalian tissues was according to Vanden Bossche et al. [1984a,b]. Piglet testes were obtained at castration (< 21 days). The testes were decapsulated, minced in 0.15 M KCI washed and homogenized in 2 vol of 0.25 M sucrose containing 20 mM KCI, 1 mM EDTA, and 20 mM Tris-buffer (SPET, pH 7.4). The homogenate was centrifuged at 1,500 g for 10 min and the cell-free supernatant at 10,OOO g for 20 min. The pellet was washed with SPET and centrifuged as before (mitochondrial fraction). The combined supernatants were centrifuged at 105,000 g for 60 min, the pellet was resuspended in 0.15 M KCl and was centrifuged as before. The pellet containing the microsomal membranes or the mitochondrial fraction was suspended in 0.1 M potassium phosphate buffer and stored at -80°C.
Spectrophotometric analysis: the cyt. P-450 content of the subcellular fractions was determined according to Omura and Sat0 [ 19641 by measuring the reduced carbon monoxide difference spectrum. The extinction coefficient used was 91 cm-' .mM- I . The interactions(s) of itraconazole or other N-substituted azoles with the cyt. P-450 isozymes in the isolated membrane fractions was examined by analyzing spectral changes induced by the azoles. The membranal fractions were diluted in 0.1 M potassium phosphate buffer (PH 7.4) to obtain a cyt. P-450 content of 0.1 nmole/ml. The suspension was divided between the reference and sample cuvettes, a base line of equal light absorbance was established, and increasing concen- trations of the azole derivatives dissolved in dimethylsulfoxide (DMSO containing 15 p1 of 6 M HCI per ml = DMSO + HCI) were added to the sample cuvette. Equal amounts of DMSO were added to the reference cuvette. The resulting difference spectra were recorded with an Aminco DW-2C UV-VIS spectrophotometer coupled with an IBM-XT and Hewlett-Packard plotter. The same amounts of azole derivative and DMSO were added to the reference and sample cuvettes respectively and the cyt. P-450 isozymes were reduced with a few grains of

Cyt. P-450 and Itraconazole 289
sodium dithionite. The sample cuvette was bubbled for 30 sec with CO and then tightly closed. The difference spectrum was recorded 45 sec later and at different time intervals up to 60 min after the addition of the reductant. Control spectra were obtained in the presence of DMSO. ,I
Sterol Metabolism
C. albicuns (ATCC 44859). Growth conditions in Eagle's minimum essential medium (EMEM), supplemented with glutamine and fetal calf serum (F.C.S.), incubation circumstan- ces, disruption of cells, extraction and saponification of the lipid-fraction, thin layer chroma- tography (TLC), and determination of radioactivity derived from [ 14C]-acetate were as described previously [Vanden Bossche et al., 1984al.
Human lymphocytes. Human peripheral blood lymphocytes were isolated using the method described by Van Wauwe et al. [1980]. Lymphocytes (5.106 cells) were cultured at 37°C in a humidified 5% C02 atmosphere in 2 ml RPMI-Hepes buffer medium (Gibco) supplemented with 5 % FCS, 2 pg/ml phytohemagglutinin (PHA) and itraconazole and/or solvent (DMSO + HC1). After 24 hr of culture, 4 pCi of ['4C]-acetate (S.A. 58.6 pcilmmol) were added, the cells were incubated for an additional 6-hr period. The cells were collected by centrifugation, washed with RPMI-medium, and saponified. The non-saponified lipids were extracted and separated on TLC and the radioactivity of the different lipid fractions was determined [Vanden Bossche et al., 1984al.
Steroid Metabolism
Mitochondrial steroid metabolism. The effects of itraconazole, dissolved in DMSO + HC1 on the cholesterol side-chain cleavage (SCC) in a reconstituted system, containing adrenodoxin and adrenodoxin reductase from bovine adrenal cortex and cyt. P-450 SCC from mitochondria of bovine adrenal cortex or pig testes, were studied as described by Willemsens and Vanden Bossche [1985]. The effects of itraconazole on the 11 0-hydroxylase in intact mitochondria isolated from bovine adrenal cortex were measured as described before [Vanden Bossche et al., 1984bl.
Steroid metabolism in S-10 and microsomal fractions. Androgen synthesis. The interaction of itraconazole with the androgen biosynthesis from pregnenolone in a S-10 fraction (i.e., the supernatant of a 10,OOO g centrifugation) of pig testes or in microsomes of the adrenal cortex was studied with the methods described for the S-10 fraction of rat testes [Vanden Bossche et al., 1985bl. However, a 30-min instead of a 3-hr incubation period was used.
Hydroxylation of testosterone. The microsomal fraction of pig testes was obtained by the method described for piglet testes microsomes. However, potassum phosphate buffer (0.1 M, pH 7.4) was used instead of SPET. The 19-hydroxylase activity (and other hydroxylases) was measured in a mixture containing, in a total volume of 10 ml:8 ml of the microsomal preparation (1 mg protein/ml, the protein content being determined with the Bio-Rad assay- cat. no. 500.0005), 1.6 mM NADPH, 5 mM ATP, 3 mh4 MgC12, 0.2 pCi [4-'4C]-tesosterone (56.9 mCi/mmol), itraconazole, and/or DMSO + HCl. After 30 min of incubation at 37"C, the steroids were extracted (3 times) with 50 ml of ethylacetate, the solvent evaporated, and the residue redissolved in a mixture of 25 ml hexane and 25 ml of CH30H:HzO (9:l). The CH30H:H20 layer was evaporated, and the steroids were re-dissolved in a mixture of 50% ethylacetate and 50% CHC13:CH30H (2: 1) and separated by 2-dimensional-TLC. First the silicagel plates were run 2 times with CH2Clz:ether (85:15) [Verhoeven and Cailleau, 19831. For the second dimension, CHC13:CH30H:H20 (94:6: .5) was used. To identify the steroids, the Rf-values were compared with those of reference compounds. One steroid, first called 1- hydroxtestosterone, was identified by HPLC-gas chromatographic-mass spectrometric analy- sis. For a first separation the HPLC method described by Schoneshofer et al. [1981] was used. The fractions with a retention time between 20 and 22 min were collected and applied to a reversed phase Nucleosil 5 CIS-column. A linear gradient elution technique was used starting

290 Vanden Bossche et al.
from CH30H:H20 (40:60) and reaching 100% CH30H in 50 min. The retention time of the radioactive fraction was 22.8 min. A gas chromatographic-mass spectrometric analysis of this isolated fraction was performed on a fused silica column coupled to a Finnigan 4,500 mass spectrometer.
Testosterone biosynthesis in testicular cells. The effects of itraconazole on testosterone biosynthesis were investigated in dispersed rat testicular cells as described by Azhar and Menon [1972]. Itraconazole was dissolved in DMSO + HCl (final DMSO concentra- tion:O. 1 %). Steroids and cyclic AMP (CAMP) were measured with radioimunoassays [NEN- CAMP set, DeCoster et al., 19841. The viability of the cells was controlled by the Trypan blue exclusion test. At the end of the experiment 82.5 f 7.1 % was found viable.
RESULTS Spectrophotometric Study of the Interactions of Itraconazole, Ketoconazole, and N- Phenylimidazole With Cytochrome P-450 lsozymes
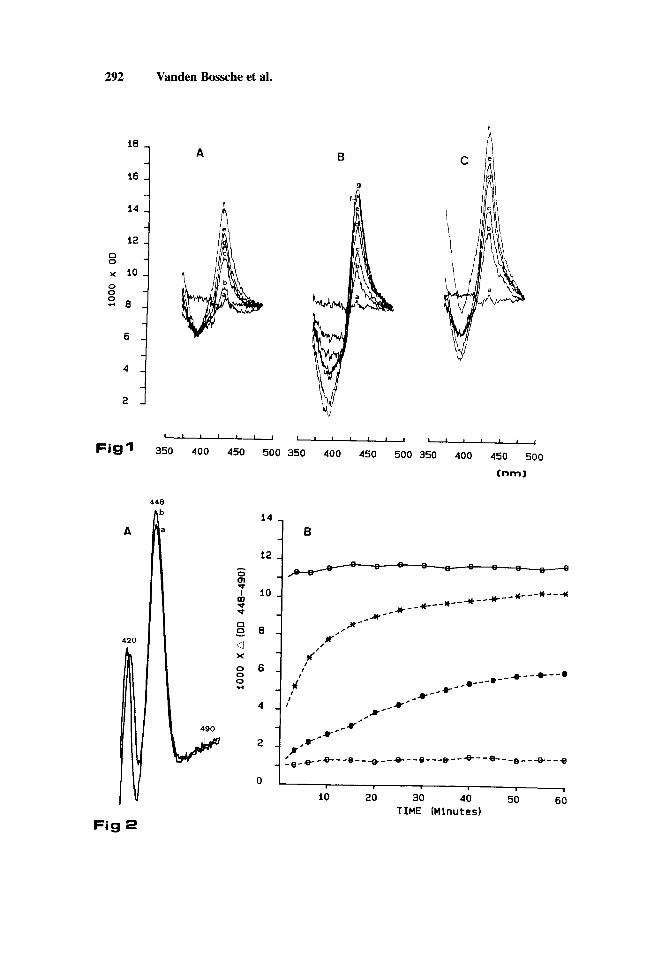
Addition of increasing concentrations of itraconazole to the microsomal suspension of C. ulbicuns containing cyt. P-450 isozymes (0.1 nmol/ml) in their oxidized state caused characteristic type I1 spectra with maxima (OD max.) and minima (OD min.) at respectively +_ 428 nm and k 394 nm (Fig. 1, Table 1). Similar type 11 spectra were obtained with ketoconazole and N-phenylimidazole. However, both imidazole derivatives gave a greater A (OD max - OD min) at their saturation concentrations (1O’M and 5 X 10 -3M respectively) than itraconazole (Fig. 1).
Upon addition of the reductant, dithionite, and saturation with carbon monoxide, the reduced cyt. P-450-CO complex showed a typical spectrum with an absorption peak at 448 nm (Fig. 2A). As shown in Figure 2A and B, this reduced CO-complex was stable over the 60- min measurement period. When the cyt. P-450 isozymes were saturated with ketoconazole (lo-’ M) or itraconazole (5 X 10-6M, i.e., the limit of solubility) and then reduced, a small absorption peak at 448 nm was observed 45 sec after bubbling with CO. In the presence of N- phenylimidazole (5 X 10-3M), this absorption peak was almost 50% of that in the presence of solvent. With the latter imidazole derivative and with ketoconazole a gradual spectral change was observed during the measurement period. This change represents the displacement of both azole derivatives from the complex by CO. After 60 min CO was able to displace N- phenylimidazole completely whereas ketoconazole was only replaced partly. Even after 60 min of contact, CO was unable to remove itraconazole from the binding place (Fig. 2B).
Itraconazole also caused type II spectra when added to suspensions of microsomal and mitochondria1 fractions of bovine adrenal cortex or pig testes. However, the A (OD rnax - OD min) values were lower than those found with C. ulbicuns microsomes (Table 1). Under the present conditions (cyt. P-450 c0ntent:O.l nmoles/ml), almost no type I1 spectrum was
TABLE 1. Maxima and Minima and A(0D max-OD min) of Type II Spectra Induced by Itraconazole
Species Absorbance (nm)a or glands Organelle Maximum Minimum AOD X loo0
C. olbicans Microsomes 428 394 7.3 Piglet testes Mitochondria 426 394 6.4
Microsomes 425 394 6.3 Bovine adrenals Mitochondria 423 397 3.1 (Cortex) Microsomes 42 1 405 1.8
aIncreasing concentrations of itraconazole (lo-* - 5 X M) were added to the sample cuvette, DMSO was added to the reference cuvette and the resulting difference spectra were traced. The values given here are those recorded in the presence of the highest itraconazole concentration.

Cyt. P-450 and Itraconazole 291
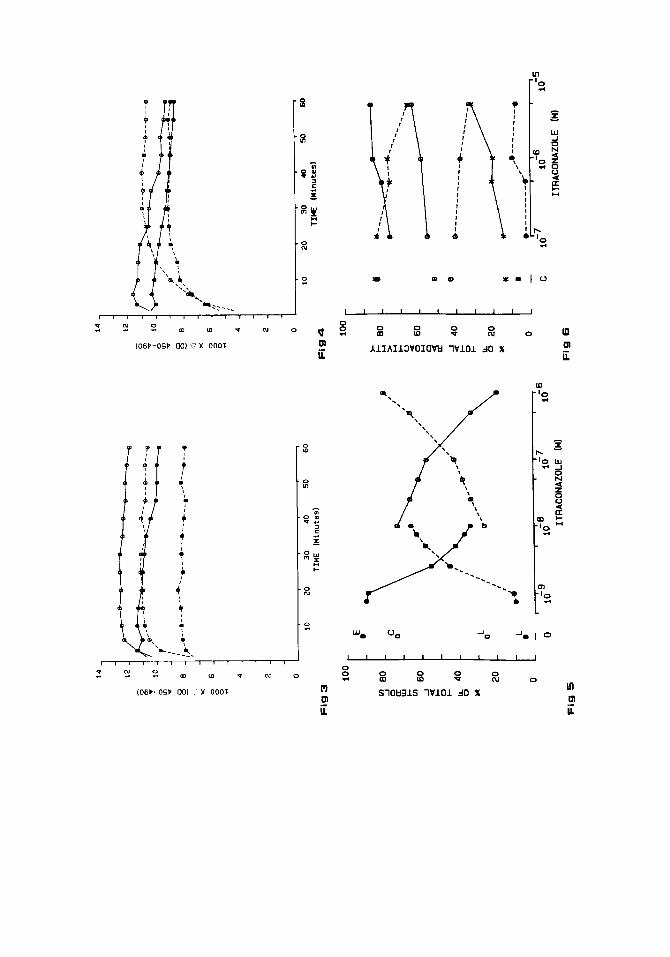
induced in microsomal fractions of livers from phenobarbital-pretreated and -untreated rabbits. This corresponds extremely well with a small change in the spectrum measured after addition of dithionite and CO to the rabbit liver microsomal suspensions, to which increasing itracona- zole concentrations were added (up to 5 X 1OP6M). It should be noted that CO seemed to be unable to replace completely the itraconazole bound (Fig. 3). CO could easily displace itraconazole from the cyt. P-450 isozymes in adrenal cortex mitochondria and microsomes (data not shown). It was also able to replace completely itraconazole from its binding place(s) on the cyt. P-450 in subcellular fractions of piglet testes (Fig. 4). However, the effect of itraconazole, at the onset of the measurement period, i.e., 45 sec after bubbling with CO, was much higher and it took a longer time (> 20 min) for CO to displace the triazole derivative from the testicular cyt. P-450 isozymes than from those present in the other mammalian cell membranes tested.
Effects of ltraconazole on Sterol Biosynthesis in Candida albicans and Human Peripheral Blood Lymphocytes
The effects of itraconazole on ergosterol biosynthesis in C. albicans grown for 24 hr in EMEM + serum and on cholesterol synthesis in PHA-stimuated human peripheral blood lymphocytes grown for 30 hr are summarized in Figure 5. A 50% inhibition of the ergosterol synthesis in C. albicans was observed at 3.2 X 10 -9 M whereas 4 X M itraconazole was needed to affect cholesterol biosynthesis in human lymphocytes. It should be noted that itraconazole at 5 X M inhibited in C. albicans ergosterol biosynthesis for 100%. Indeed, a study using TLC and gas chromatographic-mass spectrometric analysis to identify the sterols revealed that ergosterol in the extracts from these treated cells was completely replaced by 14a-methylsterols [Vanden Bossche et al., 1984al.
Effects of ltraconazole on Steroid Biosynthesis in Sub-Cellular Fractions of Adrenals and Testes
When adrenal cortex mitochondria were incubated in the presence of C3H]-1 l-deoxycor- tisol, almost 83% of the radioactivity was found in cortisol (Fig. 6). No inhibition of the 11- hydroxylase-dependent cortisol synthesis was observed after 30 min of incubation with 5 X
M of itraconazole. The cholesterol-SCC was almost not inhibited (< 24%) when itraconazole was added to a mixture containing NADPH, adrenodoxin and adrenodoxin reductase from bovine adrenal cortex mitochondria and cyt. P-450 SCC from either bovine adrenal cortex or pig testes mitochondria.
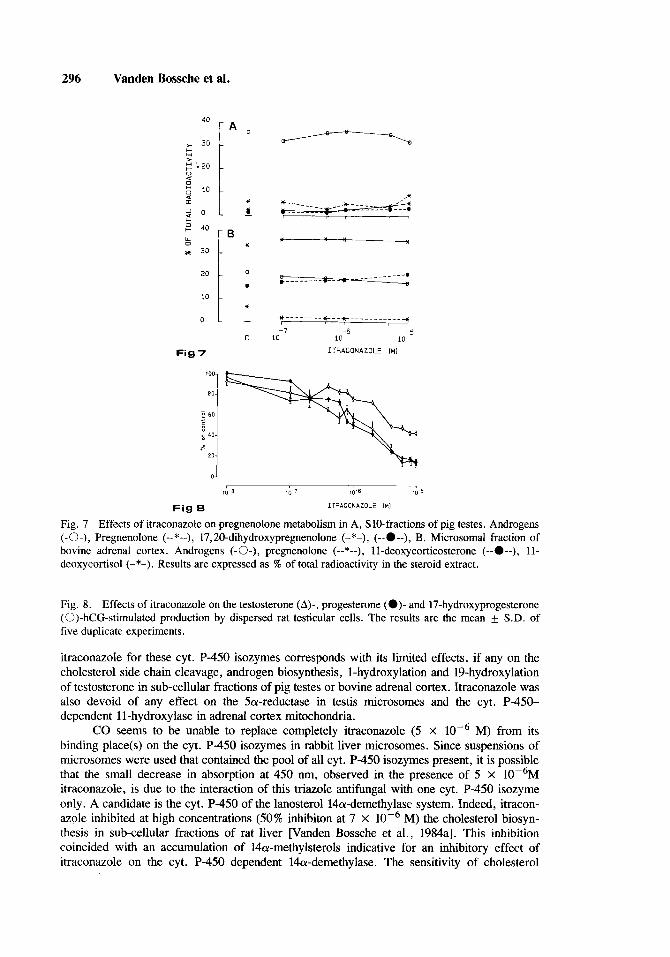
Up to 5 x lop6 M itraconazole did not affect androgen synthesis from pregnenolone in an incubation mixture containing a 10,OOO g supernatant (Sl0 fraction) of pig testes homoge- nates (Fig. 7A) or microsomes of bovine adrenal cortex (Fig. 7B)
Itraconazole did not interfere with the 19-hydroxylation and 5a-reduction of testosterone by pig testes microsomes (Table 2). The results in Table 2 show a small effect of itraconazole (26% at 5 X M) on the synthesis of a testosterone derivative of which a preliminary study revealed a steroid with one keto and two free hydroxyl functions. Comparisons of the mass spectrum of the oxime-silyl derivative with that published by Lisboa and Gustafsson [ 19681 revealed that the structure of this steroid is most probably consistent with l-hydroxytes- tosterone (More details on the identification of this C19-steroid will be published elsewhere). No significant effect of itraconazole was found on the synthesis of dihydrotestosterone (Table 2), indicating that itraconazole did not affect the 5a-reductase activity in pig testes microsomes.
Effects of ltraconazole on Testosterone Synthesis in Dispersed Testicular Cells
Incubation of dispersed rat testicular cells resulted in a testosterone production of 5.6 f 1.5 nmoles/106 cells/3 h (mean f S.D., number of experiments =7). After hCG (human chorionic gonadotropin) stimulation, 42.3 f 13.8, 1.24 f 0.54, and 1.03 f 0.41 nmoles/106 cells/3 h of testosterone, 17a-hydroxyprogesterone, and progesterone were found. Itraconazole
292 Vanden Bossche et al.
A
420
1
I
J
A
b
l a
B
9
f
v I I I I , , , , , , , , , 1-
Fig1 350 400 450 500 350 400 450 500 350 400 450 500
Cnml
448
- 0
I W v cl
0 0 - Q X 1 d'
20 30 40 50 60 10 TIME [Minutes)
Fig 2

Cyt. P-450 and Itraconazole 293
showed a dose-dependent inhibition of testosterone synthesis. Without stimulation, 50% inibi- tion was found at 4.5 x M itraconazole. In the presence of hCG, 50% inhibition was obtained at 1.6 X lop6 M (Fig. 8). The production of the precursors 17 a-hydroxyprogester- one and progesterone was affected in the same way as that of testosterone (Fig. 8). It should be noted that itraconazole did not affect the amount of CAMP excreted into the medium.
DISCUSSION
The results presented further show that itraconazole interacts with cyt. P-450 in C. albicans microsomes, yielding type II binding spectra. A type I1 spectral change suggests that the observed interaction of itraconazole and other N-substituted nitrogen heterocycles [for a review see Vanden Bossche, 19851 with cyt. P-450 is due to binding of the unhindered nitrogen to the sixth coordination positon of the heme iron atom. The intensity of these difference spectra, expressed by A (OD max - OD min) was proportional to the inhibitor concentration. However, the value that appeared at the saturation concentrations was different for itracona- zole, ketoconazole, and N-phenylimidazole. Itraconazole, being the biggest molecule, gave a much lower intensity. From the results presented here it can be deduced that the type II difference spectra obtained with these hydrophobic substances are not related to their capacity to compete with carbon monoxide for the sixth coordination position of the iron in the protoporphyrin moiety of the C. albicans cyt. P-450. Indeed, itraconazole inducing the lowest type 11 spectrum could not be displaced by CO; ketoconazole was replaced partly only and N- phenylimidazole was intirely removed from the sixth position. This corresponds with the low effect, if any, of N-phenylimidazole (up to 2 X M) on the growth of C. albicans (ATTC 28516) in casein-hydrolysate-yeast-extract-glucose-medium (data not shown). Under the same
TABLE 2. Effect of Itraconazole on the Incorporation of Radioactivity Derived From [‘4C]- Testosterone Into Steroids by pig Testes Microsomes*
Itraconazole (M) 19-OH-testo 1-OH-testo DHT testo
0 24.1 i- 2.8 24.3 i- 5.8 6.4 f 1.5 2.8 * 0.6 10-7 24.5 i- 3.6 25.0 * 1.3 5.5 2.7 2.5 f 0.5 5. lop7 27.1 f 4.2 19.9 3.5 5.1 f 1.7 2.5 * 0.8 10-6 25.1 + 5.0 18.7 i- 3.1 5.3 & 1.9 2.5 f 1.1 5.1OP6 21.2 * 4.7 18.0 * 0.8 4.6 & 2.1 3.5 f 0.9
*Abbreviations: 19-OH-testo, 19-hydroxy-testosterone; 1-OH-testo, 1-hydroxytestosterone; DHT, 5a- dihydrotestosterone; testo, testosterone. aResults are expressed as percent of total radioactivity recovered after separation of all the methanol- extractable steroids. Results are the mean * SD of at least three experiments.
Per cent total radioactivitya
Fig. 1. Type I1 difference spectra obtained with DMSO (a); A, itraconazole-(b) : 5 X
10W7 M, (d) : 5 x (8) : (f) : 5 x
M (c) : M (f) : 5 X lop6 M; B, ketoconazole-(b-f, see itraconazole,
M, (e) : M, M, (e) :
M. C, N-phenylimidazole-(b) :5 X lo-’ M, (c) : 10K4 M, (d) : 5 X M . C. albicuns microsomes with a cyt. P-450 content of 0.1 nmoles/ml were used.
Fig. 2. A, CO-difference spectra of a microsomal preparation of C. albicans. a is the spectrum recorded 45 sec after the sample cuvette was bubbled with CO; b is the spectrum recorded 60 min later. B, Data taken from difference spectra obtained with microsomal preparations of C. albicans. The cyt. P-450 isozymes were titrated (see Fig. 1) with DMSO (-0-), itraconazole (--0--), ketoconazole (--O--), or N-phenylimidazole (--X--), reduced with dithionite and bubbled with CO. The differences in absorption at 448 nm and 490 nm are plotted against time in minutes.
y\ I I I
I I
i
1 1 1 1 1 1 1 1 1 1 1
O U I m ii
0 N 0 9 0 (D 0 0 s: rn
AlIhI13VOIWtl lVlO1 dO X

Cyt. P-450 and Itraconazole 295
conditions, ketoconazole and itraconazole induced 50% inhibition of growth at about 3.5 x lo-' M [Vanden Bossche et al., 1984al.
Recently Yoshida and Aoyama [ 19861, using cyt. P-450 purified from Saccharomyces cerevisiae microsomes, found that triadimefon, being also a triazole antifungal, caused a type I1 spectral change similar to that induced by itraconazole. However, striking differences were observed by studying the reactivity of the reduced cyt. P-450-itraconazole and triadimefon- complexes with CO. Indeed, the authors found that the triadimefon-complex was readily converted into the CO-complex, whereas the itraconazole-complex was not changed by the addition of CO. This property of itraconazole was not simply due to binding of its triazole nitrogen to the iron, because triadimefon was readily displaced. The results obtained by Yoshida and Aoyama [I9861 suggest that the differences, reported in the present paper, between the two imidazole derivatives, N-phenylimadazole and ketoconazole, and the triazole deriva- tive, itraconazole, are not due to differences in binding capacity of the nitrogen heterocycles to the heme iron atom. Differences in their interactions with the apoprotein seem to be involved. So, binding of the azole derivatives to the heme iron atom is responsible for the spectral changes and inhibition of oxygen binding and activation whereas binding to the apoprotein determines the affinity of the chemicals to the cyt. P-450 isozyme(s).
The results discussed here also suggest that the ability of an azole derivative to induce a type I1 difference spectrum is not necessarily indicative for a possible inhibitory effect on a cyt. P-450-dependent reaction. The degree and velocity of the convertion of the reduced cyt. P-450 -drug complex into the cyt. P-450-CO complex might be a better indication.
At higher concentrations (> 10-7M) than those needed to inhibit the cyt. P-450- dependent ergosterol biosynthesis in yeasts and fungi, ketoconazole inhibits in vitro cyt. P- 450-dependent steroidogenic enzymes in mitochondria and microsomes of testicular and adrenal cortex cells [Vanden Bossche et al., 1985b,c 1986; Willemsens and Vanden Bossche, 19851. As previously shown, itraconazole does not affect testosterone synthesis in the sub- cellular fractions of rat testes [Vanden Bossche et al., 1985bl. The results presented in this paper indicate that although itraconazole induced type II difference spectra, indicative for binding to the heme iron atom in cyt. P-450 of testes and adrenal cortex microsomes and mitochondria, CO could displace itraconazole from its binding place(s). This low affinity of
Fig. 3. Differences in absorption at 450 nm and 490 nm are plotted against time in minutes. The cyt. P-450 isozymes in liver microsomes from untreated (0) and with phenobarbital pretreated (0) rabbits were titrated with DMSO (-) or with itraconazole (---) to a final concentration of 5 X M, reduced with dithionite and bubbled with CO.
Fig. 4. Differences in absorption obtained with cyt. P-450 isozymes in pig testes mitochondria (0) or microsomes (0) are plotted against time. DMSO (-), itraconazole (--). More details are given in the legend to Figure 3.
Fig. 5. Effects of itraconazole on sterol synthesis in C. albicuns (0) and PHA stimulated human peripheral blood lymphocytes (0). E, ergosterol; C, cholesterol; L, lanosterol. Solid lines represent the effects of itraconazole on ergosterol (0) or cholesterol (0) synthesis. Broken lines represent the accumulation of 4, 4', 14-trimethylsterols (e.g., lanosterol) and 4,14-dimethylsterols found in C. albicans (0 ) and of lanosterol in lymphocytes (0). Results are expressed as % of the radioactivity found in all the sterols present.
Fig. 6. Effects of itraconazole on the conversion of cholesterol (-*-, -o-), into pregnenolone (--*--, --0--) in a reconstituted system containing cyt. P-450 isozymes from pig testes (*) or bovine adrenal cortex (0) mitochondria. Effects of itraconazole on the 11-hydroxylase in bovine adrenal cortex mitochondria (0). 11-Deoxycortisol (--0--), cortisol (-0-). C = control. Results are expressed as % of the total radioactivity found in the steroid extract.
296 Vanden Bossche et al.
40 [ A 0 30
6 ," 40
:30[ *
,*
c I
,*
c I
70-8 10.7 10.6 10-5
Fig 8 ITRACONAZOLE [MI
Fig. 7 Effects of itraconazole on pregnenolone metabolism in A, SlO-fractions of pig testes. Androgens (-0-), Pregnenolone (--*--), 17,20-dihydroxypregnenolone (-*-), (--O--), B. Microsomal fraction of bovine adrenal cortex. Androgens (-0-), pregnenolone (--*--), 11-deoxycorticosterone (--O--), 11- deoxycortisol (-*-). Results are expressed as % of total radioactivity in the steroid extract.
Fig. 8. Effects of itraconazole on the testosterone (A)-, progesterone (0)- and 17-hydroxyprogesterone (0)-hCG-stimulated production by dispersed rat testicular cells. The results are the mean f S.D. of five duplicate experiments.
itraconazole for these cyt. P-450 isozymes corresponds with its limited effects, if any on the cholesterol side chain cleavage, androgen biosynthesis, 1-hydroxylation and 1Phydroxylation of testosterone in sub-cellular fractions of pig testes or bovine adrenal cortex. Itraconazole was also devoid of any effect on the 5a-reductase in testis microsomes and the cyt. P-450- dependent 11-hydroxylase in adrenal cortex mitochondria.
M) from its binding place(s) on the cyt. P-450 isozymes in rabbit liver microsomes. Since suspensions of microsomes were used that contained the pool of all cyt. P-450 isozymes present, it is possible that the small decrease in absorption at 450 nm, observed in the presence of 5 X 10P6M itraconazole, is due to the interaction of this triazole antifungal with one cyt. P-450 isozyme only. A candidate is the cyt. P-450 of the lanosterol l4a-demethylase system. Indeed, itracon- azole inhibited at high concentrations (50% inhibiton at 7 X loP6 M) the cholesterol biosyn- thesis in sub-cellular fractions of rat liver [Vanden Bossche et al., 1984al. This inhibition coincided with an accumulation of 14a-methylsterols indicative for an inhibitory effect of itraconazole on the cyt. P-450 dependent 14a-demethylase. The sensitivity of cholesterol
CO seems to be unable to replace completely itraconazole (5 X

Cyt. P-450 and Itraconazole 297
biosynthesis for itraconazole was further proven in lymphocytes (this paper) and almost similar results were obtained with human primary vulva carcinoma cells and human fibroblasts (data not shown).
The inhibitory effect of itraconazole on cholesterol synthesis might produce the de- creased synthesis of progesterone, 17a-hydroxy-progesterone, and testosterone observed when dispersed testicular cells were incubated in the presence of itraconazole. However, in vivo studies did not reveal any effect of itraconazole either on androgen or on corticosteroid plasma levels in rats and volunteers [Van Cauteren et al., 19861. The glands involved have the ability to derive their cholesterol from the blood, compensating in this way for the inhibition of sterol synthesis. Even for some tissues, such as adrenals and kidney, synthesis of sterols contributes only a relatively small proportion of the total tissue cholesterol [Turley et al. 19811. The use of cholesterol from the diet might also compensate for its possible decreased synthesis. Indeed, after 3 months of treatment of patients with itraconazole doses up to 400 mg daily, the serum cholesterol level did not decrease [Cauwenbergh, personal communication].
ACKNOWLEDGMENTS
The authors are grateful to J. Van Cutsem and J. Van Wauwe for providing the Cundida albicuns strains and human lymphocytes respectively and to G . Cauwenbergh for giving permission to use unpublished results. Grateful appreciation is expressed to L. DonnC for typing the text and to L. Leijssen for preparing the photographic prints. This work was supported in part by a grant from the “Instituut voor aanmoediging van het wetenschappelijk onderzoek in nijverheid en landbouw . ”
REFERENCES
Azhar, S. , Menon, K.M.J. : Receptor mediated gonadotropin action in gonadal tissues: Relationship between blood cholestrol levels and gonadotropin stimulated steroidogenesis in isolated rat Leydig and luteal cells. J. Steroid. Biochem. 16:175-184, 1972.
De Coster, R., Beerens, D., Dom, J., Willemsens, G.: Endocrinological effects of single daily ketocon- azole administration ia male beagle dogs. Acta Endocrinol. 107:275-281 1984.
Lisboa, B.P., Gustafsson, J. A .: Biosynthesis of two new steroids in the human foetal liver, lp- and 2p- hydroxytestosterone. Eur. J. Biochem. 6:419-424, 1968.
Omura, T., Sato, R.: The carbon monoxide-binding pigment of liver microsomes. I. Evidence for its hemoprotein nature. J. Biol. Chem. 239:2370-2378, 1964.
Schoneshofer, M., Fenner, A., Duke, H.J.: Assessment of eleven adrenal steroids from a single serum sample by combination automatic high-performance liquid chromatography and radioimmunoas- say (HPLC-RIA). 5. Steroid. Biochem. 14:377-386, 1981.
Turley, S.D., Andersen, J.M., Dietschy, J.M: Rates of strol synthesis and uptake in the major organs of the rat in vivo. J. Lip. Res. 22:551-569, 1981.
Van Cauteren, H., Heykants, J., De Coster, R., Cauwenbergh, G.: Itraconazole: Animal and human pharmacological studies. Rev. Infect. Dis., 1986.
Vanden Bossche, H.: Biochemical targets for antifungal azole derivatives: Hypothesis on the mode of action. In McGinnis, M.R., (ed.): “Current Topics in Medical Mycology,” Vol. 1. New York; Springer-Verlag, 1985, pp. 313-351.
Vanden Bossche, H., Willemsens, G.: Effects of the antimycotics, miconazole and ketoconazole, on cytochrome P-450 in yeast microsomes and rat liver microsomes. Arch. Int. Physiol. Biochim.
Vanden Bossche, H . , Willemsens, G., Marichal, P., Cools, W., Gorrens, J: The molecular basis for the antifungal activities of N-substituted azole derivatives. Focus on R 51 211. In Trinci, A.P.J., Ryley, J.F. (eds.): “Mode of Action of Antifungal Agents. ” Cambridge: Cambridge University Press, 1984a, pp. 321-341.
Vanden Bossche, H . , Willemsens, G., Cools, W., Bellens, D.: Effects of etomidate on steroid biosyn- thesis in sub-cellular fractions of bovine adrenals. Biochem. Pharmaco1.33:3861-3868, 1984b.
90: B218-B219, 1982.

298 Vanden Bossche et al.
Vanden Bossche, H., Bellens, D., Gorrens, J., Marichal, P., Verhoeven H., Willemsens, G . : Yeast and plant cytochrome P-450 enzymes: Targets for azole derivatives. In Vereczkey, L., Magyar, K. , (eds.): “Cytochrome P-450, Biochemistry, Biophysics and Induction. Budapest: AkadCmia Kiad6, 1985a, pp. 423-429.
Vanden Bossche, H., Lauwers W., Willemsens, G . , Cools, W.: The cytochrome P-450 dependent C 17,20-lyase in subcellular fractions of the rat testis: Differences in sensitivity to ketoconazole and itraconazole. In Boobis, A.R., Caldwell, J., De Matteis, F., Elcombe, C.R. (eds.): “Micro- somes and Drug Oxidations.” London: Taylor and Francis, 1985b, pp. 63-73.
Vanden Bossche, H., Lauwers, W., Willemsens, G . , Cools, W.: Ketoconazole, an inhibitor of the cytochrome P-450 dependent testosterone biosynthesis. In Schroeder, F.H., Richards, B. (eds.): “EORTC Genitourinary Group Monograph 2, Part. A. The Therapeutic Principles in Metastatic Prostatic Cancer.” New York: Alan R. Liss, Inc., 1985c, pp. 187-196.
Vanden Bossche, H., De Coster, R., Amery, W.K.: Pharmacology and clinical uses of ketoconazole. In Furr, B.J.A., Wakeling, A. (eds.): “Pharmacology and Clinical Uses of Inhibitors of Hormone Secretion and Action. Eastbourne: Praeger, 1986.
Van Wauwe, J., De Mey, J., Goossens, J . : OKT3: Monoclonal anti-human T-lymphocyte antibody with potent mitogenic properties. J. Immunol. 124: 2708-2713, 1980.
Verhoeven, G . . Cailleau, J. : Inhibition by a 4-methyl-4-aza-steroid of NADPH: A4-3-oxosteroid-cw- oxido-reductase activity in cultured interstitial cells derived from immature rat testis. J. Steroid. Biochem. 118:365-367, 1983.
Willemsens, G. , Vanden Bossche, H.: Effects of ketoconazole on the testicular and adrenal cholesterol side-chain cleavage. In Vereczkey, L., Magyar, K. (eds.): “Cytochrome P450, Biochemistry, Biophysics and Induction. ” Budapest: Akadtmia Kiad6, 1985, pp. 203-206.
White, R.E., Coon, M.J.: Oxygen activation by cytochrome P450. Ann. Rev. Biochem. 49:315-356, 1980.
Yoshida, Y., Aoyama, Y.: Interaction of azole fungicides with yeast cytochrome P-450 which catalyzes lanosterol l4a-demethylation. In Iwata, K., Vanden Bossche, H. (eds.): “In Vitro and In Vivo Evaluation of Antifungal Agents. Amsterdam: Elsevier Science Publishers, 1986, pp. 125-134.





























