Embed Size (px)
Citation preview

Toxicology Letters 95 (1998) 31–39
Cypermethrin increases apo A-1 and apo B mRNA but nothyperlipidemia in rats
Lourdes Aldana a, Elvira Gonzalez de Mejıa a,*, Arthur Craigmill b, Vıctor Tsutsumi c,Juan Armendariz-Borunda d, Arturo Panduro d, Ana Rosa Rincon d
a Uni6ersidad Autonoma de Queretaro, Queretaro, Mexicob Uni6ersity of California Da6is, Da6is, CA, USA
c CINVESTAV-IPN, Mexicod Institute of Molecular Biology in Medicine, Centro Uni6ersitario de Ciencias de la Salud,
Uni6ersidad de Guadalajara and Hospital Ci6il de Belen, Guadalajara, Jalisco, Mexico
Received 17 September 1997; received in revised form 8 January 1998; accepted 13 January 1998
Abstract
The hepatotoxic effect of cypermethrin and the expression of hepatic genes at the mRNA level, as molecularmarkers of liver damage, were evaluated in rats following exposure to cypermethrin. The expression of hepatic geneswas compared with conventional liver functional tests, and correlations were made by studying the liver at theultrastructural level. Cypermethrin treated rats presented a significant decrease, of 79% and 22%, on the expressionof albumin and apo E genes at 5 days, respectively. The levels of apo A-1 and apo B mRNA were increased up tofour- and fivefold, respectively. This increase did not correlate with the serum values of HDL and VLDL lipoproteinparticles. Intracytoplasmic lipid droplets were observed after the first 2 days following cypermethrin administration,suggesting that apo A-1 and B mRNA were translated but not secreted. There were significant correlations betweenthe low values of the albumin gene expression, the decrease in the HDL concentrations, and the ultrastructuralalterations, respectively. These alterations were mainly a large amount and increased size of mitochondria in theanimals exposed to cypermethrin. It is concluded that under the experimental conditions used, cypermethrin may alterthe metabolism of lipids and proteins in rat liver. © 1998 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Cypermethrin; Pyrethroids; Acute phase proteins; Hepatic ultrastructure
* Corresponding author. Departamento de Investigacion yPosgrado en Alimentos, Universidad Autonoma de Queretaro.Centro Universitario, Cerro de las Campanas, 76040 Quere-taro, Queretaro, Mexico. Tel.: +52 62 156867; fax: +52 62163730; e-mail: [email protected]
1. Introduction
Synthetic pyrethroids have been consideredamong the safest pesticides available, and are used
0378-4274/98/$19.00 © 1998 Elsevier Science Ireland Ltd. All rights reserved.
PII S0378-4274(98)00013-7

L. Aldana et al. / Toxicology Letters 95 (1998) 31–3932
worldwide (Dorman and Beasley, 1991). Cyper-methrin contains a cyano group substituent onthe alpha-carbon of the phenoxy-benzyl moietyand it is classified as a Type II pyrethroid(Gray, 1985; World Health Organization, 1989).Administration of Type II pyrethroids inducesataxia, convulsions, hyperactivity, and profusesalivation in rats and mice (Casida et al., 1983;Vershoyle and Aldridge, 1980), and has beenwidely studied (Eells et al., 1992; Narahashi,1985; Shing and Agarwal, 1993).
The biochemical and histopathological effectsof pyrethroids include slight increases in serumtransaminase activities (El-Toukhy and Girgis,1993; Miyanoto, 1976; Reddy et al., 1991;Shakoori et al., 1988), as well as in other en-zymes (Flodstrom et al., 1988; Krechniak andWrzesniowska, 1991), and cytoplasmic hypertro-phy with intracytoplasmic vacuolation, occa-sional patchy necrosis, and polymorphonuclearcellular infiltration (El-Toukhy and Girgis,1993).
The liver plays an important role in the syn-thesis of apolipoproteins and in lipidmetabolism (Shepherd, 1994). Like albumin, theapolipoprotein E (apo E) gene is expressedmainly in liver cells (Elshourbagy et al., 1985a),whereas apo A-1 and apo B genes are expressedin liver as well as intestine (Elshourbagy et al.,1985b).
In partially hepatectomized rats, and in ratstreated with a single dose of carbon tetrachlo-ride (CCl4) to induce acute liver damage, thesteady state levels (SSL) of apo A-1 mRNA in-crease several fold, whereas the SSL of apo EmRNA decrease (Panduro et al., 1993). Such aresponse in the expression of apolipoproteingenes at the mRNA level during acute liverdamage and liver regeneration, is similar to thatdetected during liver development (Panduro etal., 1987), and to the expression of a-fetoproteinand albumin during liver development and re-generation (Panduro et al., 1987, 1986). Basedon such observations, determinations of apo A-1and apo E mRNAs have been suggested to beused as molecular markers of liver damage andregeneration (Panduro et al., 1992).
Besides the evaluation of liver function, using
molecular markers, the analysis of the expres-sion of apo A-1 at the mRNA level, and theserum lipid profile, help us understand the regu-lation of apolipoprotein gene expression underdifferent pathophysiological conditions. The ob-jective of the present study was to analyze thehepatotoxic effect of cypermethrin on the ex-pression of albumin, apolipoprotein A-1, E andB genes, and hepatic changes at the ultrastruc-tural level in the rat.
2. Materials and methods
2.1. Test material
Cypermethrin [(R,S) a-cyano-3-phenoxy-benzyl (1R,S)-cis, trans-3-(2,2-dichlorovinyl)-2,2-dimethyl-cyclopropane carboxylate) was suppliedby ICI Agricultural Products Group (Bucks,AL) to Zeneca Company (ICI of Mexico). Thepurity analyses were performed by the ICIgroup. The test material was technical grade andhad 91% purity (49.9:50.1, cis:trans). The com-pound was produced synthetically and did notcontain any toxicologically significant impurities.Cypermethrin was stored at 20°C in the dark.
2.2. Animals and maintenance
Male Wistar rats were obtained from the Uni-versity of Puebla farm, Mexico, weighing 280–300 g. Animals were acclimatized for aminimum of 7 days before treatment. The ani-mals were randomly selected, based on theirbody weight, and housed in stainless-steel wirecages during the 7 days of treatment. The ani-mals were maintained on day-night cycles of 12h, at a relative humidity of 30–60% and a tem-perature of 19–22°C; they also had free accessto a rodent lab diet (Ralston Rations, Purina,St. Louis, MO) and water. The experimentaland control rats were kept separately. Twice aweek the cages were cleaned and the sawdustreplaced. At least seven animals were used foreach time point, and three independent experi-ments were carried out.

L. Aldana et al. / Toxicology Letters 95 (1998) 31–39 33
2.3. Treatment
The substance was dissolved in Pluronic F-68(20%) (Sigma, St. Louis, MO), which was used asa vehicle (v/v, 1:1). A daily dose was administeredby intraperitoneal injection (i.p.) (300 mg/kg/day)for 7 days. This exposure concentration was de-termined by preliminary range-finding studiesconducted at the same laboratory. The controlanimals received a corresponding amount of thevehicle (i.p.). Rats were killed daily by decapita-tion and exsanguinated. Serum was obtained bycentrifugation at 2000 rpm for 2 min.
2.4. Biochemical assays of li6er function
Total protein, albumin, aspartate aminotrans-ferase (AST), and alanine aminotransferase(ALT) were measured spectrophotometrically inserum using commercial kits (Lakeside Laborato-ries, Boehringer Mannheim, Germany).
2.5. Serum lipid profile
Cholesterol, triacylglycerides, high density lipo-protein (HDL) and low density lipoprotein(LDL)-cholesterol determinations were performedusing fresh serum samples. Serum total choles-terol was measured as described by Panduro et al.(1993), using the monotest cholesterol kit pro-vided by Lakeside Laboratories (BoehringerMannheim, Germany). Triacylglycerides werecompletely hydrolyzed and the liberated glycerolwas determined by colorimetry (Lehnus andSmith, 1978). HDL-cholesterol concentration wasseparated from very low density lipoproteins(VLDL-cholesterol) and LDL-cholesterol by pre-cipitation with phosphotungstic acid (0.55 mmol/l) (Burnstein et al., 1970). On the other hand, theLDL-cholesterol was precipitated by addingpolyvinyl sulphate to another sample; the concen-tration was calculated from the difference betweenthe serum total cholesterol and the cholesterol inthe supernatant after centrifugation. VLDL-cholesterol concentration was calculated as thedifference between total serum cholesterol andLDL+HDL-cholesterol (Bronzert and Brewer,1977).
2.6. Isolation of total RNA and Northern blotanalysis
Small fragments of liver tissue were frozen us-ing liquid nitrogen, and stored at −70°C untilRNA extraction. RNA was extracted by themethod described by Chomczynski and Sacchi(1987). Total RNA (10 mg) was denatured for 15min at 65°C, as previously described (Panduro etal., 1986), and the sample run in separated lanesin a 1% agarose gel prepared in 1× MOPS buffercontaining 6% formaldehyde, and electrophoresedfor 3–4 h at 40 V according to the methodreported by Thomas (1980). After electrophoresis,the RNA was transferred to a Gene Screen mem-brane (Du Pont Company, Wilmington, DE) fol-lowing the instructions of the manufacturer. Themembranes were pre-hybridized and hybridizedunder stringent conditions with the correspondingradiolabeled probe. The albumin, apo A-1, apo Band apo E cDNAs probes were radioactively la-beled by random primer extension (Amersham,Buckinghamshire, UK), as described by the man-ufacturer, using [32P]dCTP (specific activity 3000Ci/mmol) (Amersham, Buckinghamshire, UK) toobtain a specific activity of 3×108 cpm/mg ofDNA. After hybridization, the membranes werewashed, and a photographic plate was exposed at−20°C using XAR-5 Kodak film (Du Pont inten-sifier screens). In each set of experiments, thesame membrane was washed at 95°C and rehy-bridized with different radiolabeled cDNAs. Atthe end of each experiment the autoradiographicbands were evaluated by densitometry (Spec-trophotometer Beckman DU 650, Palo Alto, CA).
2.7. Histology and electron microscopy processing
Tissues, for light microscopy examination, werefixed in 10% phosphate-buffer formalin (pH 7.0).Blocks were embedded in paraffin, then 4–6-mmthick sections were stained with hematoxylin andeosin. For electron microscopy, selected liver frag-ments measuring less than 1 mm thick were fixedby immersion in 2.5% glutaraldehyde using a 0.1M sodium cacodylate buffer, pH 7.2, at roomtemperature, postfixed with 1% osmium tetroxidein the same buffer and embedded in Epon 812.
L. Aldana et al. / Toxicology Letters 95 (1998) 31–3934
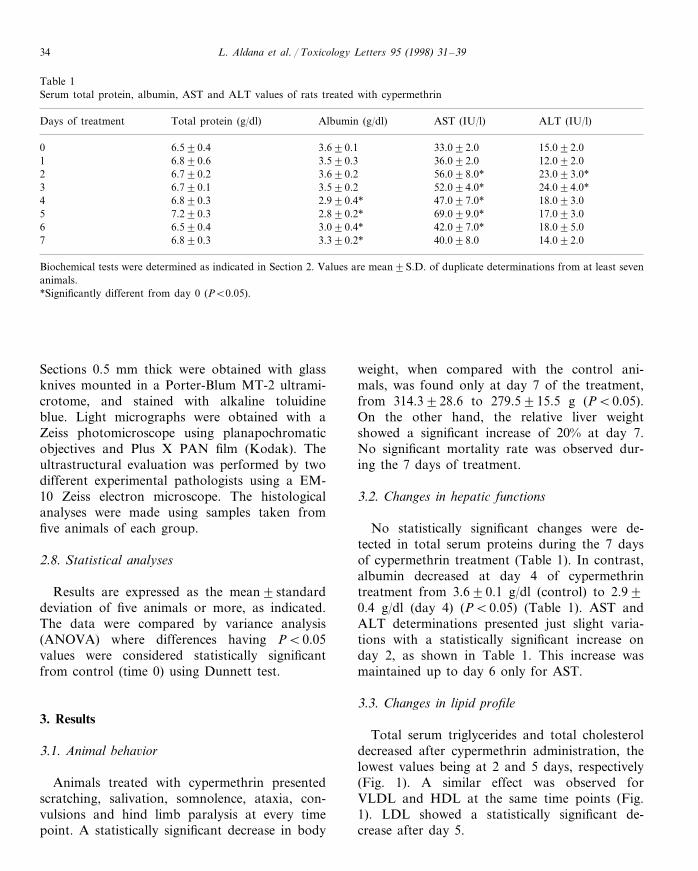
Table 1Serum total protein, albumin, AST and ALT values of rats treated with cypermethrin
Albumin (g/dl) AST (IU/l)Days of treatment ALT (IU/l)Total protein (g/dl)
3.690.10 33.092.06.590.4 15.092.01 6.890.6 3.590.3 36.092.0 12.092.02 6.790.2 3.690.2 56.098.0* 23.093.0*
3.590.2 52.094.0*6.790.1 24.094.0*36.890.34 2.990.4* 47.097.0* 18.093.07.290.35 2.890.2* 69.099.0* 17.093.0
3.090.4* 42.097.0*6.590.4 18.095.063.390.2*7 40.098.06.890.3 14.092.0
Biochemical tests were determined as indicated in Section 2. Values are mean9S.D. of duplicate determinations from at least sevenanimals.*Significantly different from day 0 (PB0.05).
Sections 0.5 mm thick were obtained with glassknives mounted in a Porter-Blum MT-2 ultrami-crotome, and stained with alkaline toluidineblue. Light micrographs were obtained with aZeiss photomicroscope using planapochromaticobjectives and Plus X PAN film (Kodak). Theultrastructural evaluation was performed by twodifferent experimental pathologists using a EM-10 Zeiss electron microscope. The histologicalanalyses were made using samples taken fromfive animals of each group.
2.8. Statistical analyses
Results are expressed as the mean9standarddeviation of five animals or more, as indicated.The data were compared by variance analysis(ANOVA) where differences having PB0.05values were considered statistically significantfrom control (time 0) using Dunnett test.
3. Results
3.1. Animal beha6ior
Animals treated with cypermethrin presentedscratching, salivation, somnolence, ataxia, con-vulsions and hind limb paralysis at every timepoint. A statistically significant decrease in body
weight, when compared with the control ani-mals, was found only at day 7 of the treatment,from 314.3928.6 to 279.5915.5 g (PB0.05).On the other hand, the relative liver weightshowed a significant increase of 20% at day 7.No significant mortality rate was observed dur-ing the 7 days of treatment.
3.2. Changes in hepatic functions
No statistically significant changes were de-tected in total serum proteins during the 7 daysof cypermethrin treatment (Table 1). In contrast,albumin decreased at day 4 of cypermethrintreatment from 3.690.1 g/dl (control) to 2.990.4 g/dl (day 4) (PB0.05) (Table 1). AST andALT determinations presented just slight varia-tions with a statistically significant increase onday 2, as shown in Table 1. This increase wasmaintained up to day 6 only for AST.
3.3. Changes in lipid profile
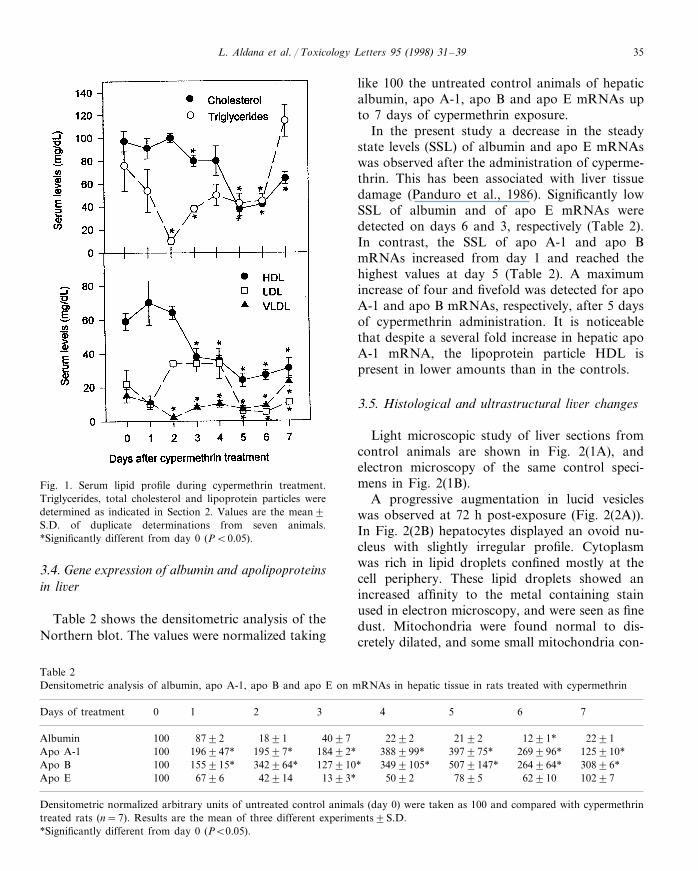
Total serum triglycerides and total cholesteroldecreased after cypermethrin administration, thelowest values being at 2 and 5 days, respectively(Fig. 1). A similar effect was observed forVLDL and HDL at the same time points (Fig.1). LDL showed a statistically significant de-crease after day 5.
L. Aldana et al. / Toxicology Letters 95 (1998) 31–39 35
Fig. 1. Serum lipid profile during cypermethrin treatment.Triglycerides, total cholesterol and lipoprotein particles weredetermined as indicated in Section 2. Values are the mean9S.D. of duplicate determinations from seven animals.*Significantly different from day 0 (PB0.05).
like 100 the untreated control animals of hepaticalbumin, apo A-1, apo B and apo E mRNAs upto 7 days of cypermethrin exposure.
In the present study a decrease in the steadystate levels (SSL) of albumin and apo E mRNAswas observed after the administration of cyperme-thrin. This has been associated with liver tissuedamage (Panduro et al., 1986). Significantly lowSSL of albumin and of apo E mRNAs weredetected on days 6 and 3, respectively (Table 2).In contrast, the SSL of apo A-1 and apo BmRNAs increased from day 1 and reached thehighest values at day 5 (Table 2). A maximumincrease of four and fivefold was detected for apoA-1 and apo B mRNAs, respectively, after 5 daysof cypermethrin administration. It is noticeablethat despite a several fold increase in hepatic apoA-1 mRNA, the lipoprotein particle HDL ispresent in lower amounts than in the controls.
3.5. Histological and ultrastructural li6er changes
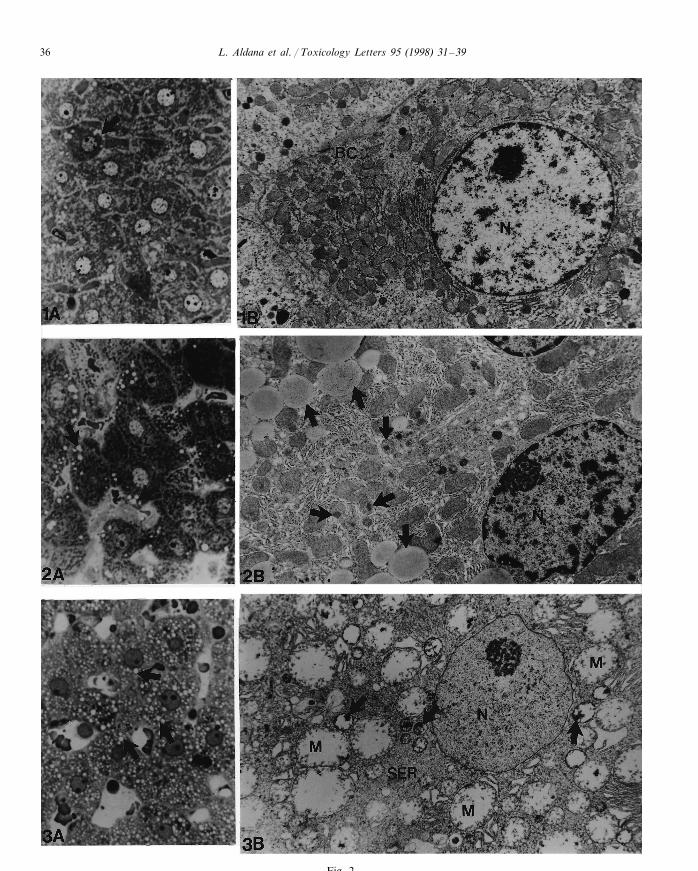
Light microscopic study of liver sections fromcontrol animals are shown in Fig. 2(1A), andelectron microscopy of the same control speci-mens in Fig. 2(1B).
A progressive augmentation in lucid vesicleswas observed at 72 h post-exposure (Fig. 2(2A)).In Fig. 2(2B) hepatocytes displayed an ovoid nu-cleus with slightly irregular profile. Cytoplasmwas rich in lipid droplets confined mostly at thecell periphery. These lipid droplets showed anincreased affinity to the metal containing stainused in electron microscopy, and were seen as finedust. Mitochondria were found normal to dis-cretely dilated, and some small mitochondria con-
3.4. Gene expression of albumin and apolipoproteinsin li6er
Table 2 shows the densitometric analysis of theNorthern blot. The values were normalized taking
Table 2Densitometric analysis of albumin, apo A-1, apo B and apo E on mRNAs in hepatic tissue in rats treated with cypermethrin
5 743210Days of treatment 6
100 8792 1891 4097 2292 2192 1291* 2291Albumin196947* 19597*Apo A-1 18492*100 388999* 397975* 269996* 125910*
100 155915* 342964* 127910* 3499105* 5079147* 264964* 30896*Apo BApo E 1393*429146796 10297629107895100 5092
Densitometric normalized arbitrary units of untreated control animals (day 0) were taken as 100 and compared with cypermethrintreated rats (n=7). Results are the mean of three different experiments9S.D.*Significantly different from day 0 (PB0.05).
L. Aldana et al. / Toxicology Letters 95 (1998) 31–3936
Fig. 2.

L. Aldana et al. / Toxicology Letters 95 (1998) 31–39 37
tained electron dense inclusions. Hepatocytes werepractically free from lipid inclusions and insteadthey showed a large amount of dilated round orovoid mitochondria with short cristae and clearmatrix. Small damaged mitochondria were alsonoticed to contain electron dense inclusions. Anincrease in mitochondria swelling was observedafter 72 h of cypermethrin exposure (Fig. 2(3B)).Endoplasmic reticulum cisternae was associatedto mitochondria (Fig. 2(2B)). Liver parenchymaobserved at 120 h after treatment displayed hepa-tocytes with abundant clear vesicles of differentsizes (Fig. 2(3A)). Smooth endoplasmic reticulumalso increased in number (Fig. 2(3B)). As the rattreatment with cypermethrin continued, the pro-liferation and swelling of smooth endoplasmicreticulum were more evident on day 5 andsubsequently.
4. Discussion
The main objective of this study was to use amolecular approach to understand if exposure toa pyrethroid (cypermethrin) could modulate theexpression of several apolipoprotein genes in vivo.
The evaluation of liver function using conven-tional biochemical studies indicates that subacutedoses of cypermethrin induce only a slight in-crease in transaminases, as reported by other au-thors (Shakoori et al., 1988; Cantalamessa, 1993).Such results, taken together with the minor alter-ation in albumin and total serum proteins, corre-sponded to the hepatotoxic effect ofcypermethrin. Normally, the plasma albumin con-
centration remains constant at 3.5–4.0 g/dl, sug-gesting an active regulation of albuminmetabolism. The present study has shown thatstress, due to exposure to a xenobiotic, such as thepesticide cypermethrin, may influence albuminsynthesis.
In previous studies we have also shown thatwhen liver tissue damage occurs, albumin tran-scription and the steady state levels of albuminmRNA decrease to 85% (Panduro et al., 1992).
However, the decrease of hepatic albumin andapo E mRNAs, associated with the increased SSLof apo A-1 and apo B mRNAs, indicates thatcypermethrin induces an important hepatotoxiceffect, which is reflected in the altered expressionof these genes. The increase in the ratio apoA-1/apo E mRNA has been shown to occur underthose physiopathological conditions that firstcause liver damage and then regeneration, such asin partially hepatectomized and acutely CCl4treated rats (Panduro et al., 1993, 1990), andwhen the liver is preserved at different tempera-tures and with different preservation solutions(Panduro et al., 1992).
In the rat, the main apolipoprotein of the newlysecreted HDL is apo E, whereas apo A-1 is themajor apolipoprotein of mature plasma HDL(Panduro et al., 1993). Thus, the reduced value ofplasma HDL during cypermethrin exposure ap-pears to correlate with the reduced steady statelevels of hepatic apo E mRNA. This finding hasbeen reported to occur also during acute liverdamage in CCl4 treated rats (Panduro et al.,1993). However, once the liver regenerates, thereis a close correlation between apo A-1 mRNA
Fig. 2. (1) Liver from control animal. (1A) Light microscopy of epoxy-embedded semithin section showed normal hepatocytes withround clear nucleus, containing scarce chromatin and a conspicuous nucleolus. Cytoplasm was granular and few cells presentedscanty lucid vesicles at the periphery. Few cells presented small vesicles (arrows) (×1000). (1B) Electron micrograph showed wellcircumscribed hepatocyte with abundant small mitochondria and flattened cisternae of rough endoplasmic reticulum (RER)randomly scattered in the cytoplasm with an increase around the cell nucleus. BC, bile canaliculus; N, nucleus (×9400). (2) Rat liverat 72 h after treatment with cypermethrin. (2A) Light micrograph of liver parenchyma shows hepatocytes with small clear vesicles,mostly localized at cell periphery (arrows) (×1000). (2B) Ultrastructure of hepatocyte with many lipid vacuoles (arrows). Nucleus(N) profile is slightly irregular. Several small mitochondria contain dense inclusions (arrows-heads). Rough endoplasmic reticulum(RER) is closely associated to mitochondrial surfaces. G, Golgi complex (×9400). (3) Rat liver at 120 h after treatment withcypermethrin. (3A) Light microscopy of liver parenchyma shows hepatocytes with abundant vesicles of different sizes (arrows)(×1000). (3B) At electron microscopy level, the cytoplasm of hepatocyte is occupied by dilated swollen mitochondria (M) withvestiges of cristae. Small mitochondria present dense inclusion (arrows). Smooth endoplasmic reticulum (SER). N, Nucleus(×9400).

L. Aldana et al. / Toxicology Letters 95 (1998) 31–3938
and HDL (Panduro et al., 1993). In the presentstudy, such a close correlation was not detectedduring the 7 days of cypermethrin treatment, per-haps due to the continued administration ofcypermethrin, and the subsequent accumulativedegree of liver damage.
Previous studies have shown that apo A-1 geneexpression is regulated at the post-transcriptionallevel (Panduro et al., 1990), and that low physio-logical viscosity values induce the expression ofapo A-1 at the mRNA level in vivo and in vitro(Nuno et al., 1997). Since lipoprotein particles,and therefore hyperlipidemia, are important fac-tors for plasma viscosity, the observed low serumvalues of total cholesterol, triglycerides and theprincipal HDL and VLDL lipoprotein particles incypermethrin treated rats, suggest that viscosity isdecreased and could be responsible for the induc-tion of apo A-1 gene expression at the mRNAlevel.
Our results are in agreement with those ofShakoori et al. (1988) with regard to the histolog-ical structure of the liver of cypermethrin-fed ani-mals. These authors also found hypertrophy ofhepatic cells and nuclei after the insecticidetreatment.
At the structural level, the lipid droplets weremostly observed at the cell periphery during thefirst 3 days of cypermethrin treatment. Such find-ings, together with the lipid profile and SSL of theapolipoproteins mRNA analyzed, suggest that atleast during those days, such mRNAs are trans-lated and the lipoprotein particles are producedbut not secreted.
The large amount and size of mitochondria, asobserved previously by other authors (El-Toukhyand Girgis, 1993), clearly indicate the hepatotoxiceffect of cypermethrin on liver cells. These mor-phological alterations as well as the proliferationand swelling of smooth endoplasmic reticulumwere associated with the growth of liver tissueobserved precisely at day 5 of cypermethrin ad-ministration. Such findings also correlated withlow values in the SSL of albumin mRNA, indicat-ing that albumin gene expression is inhibited dur-ing acute liver damage, as well as the proliferationof hepatic cells as reported previously (Panduro etal., 1986, 1988).
The present study indicates that cypermethrin isan important hepatotoxic pesticide that inducesthe expression of apo A-1 and B genes at themRNA level, and inhibits apo E synthesis butdoes not originate hyperlipidemia. Such a hepato-toxic effect was confirmed by the ultrastructuralalterations associated with cypermethrinadministration.
Acknowledgements
Lourdes Aldana was recipient of a scholarshipfrom the National Council for Science and Tech-nology (CONACYT) and the University ofSonora. The research was supported by the Stateof Queretaro’s Council for Science and Technol-ogy (CONCyTEQ) and in part by the Universityof Guadalajara (research program). The authorswish to express their recognition to Dr. ArnulfoAlbores from CINVESTAV-IPN, Mexico, for hiskind review and comments to the manuscript.
References
Bronzert, T., Brewer, H.B., 1977. New micromethod for mea-suring cholesterol in plasma lipoprotein fractions. Clin.Chem. 23, 2089–2098.
Burnstein, M., Scholnick, H.R., Morfin, R., 1970. Rapidmethod for the isolation of lipoproteins from human serumby precipitation with polyanions. J. Lipid Res. 11, 583–595.
Cantalamessa, F., 1993. Acute toxicity of two pyrethroids,permethrin, and cypermethrin in neonatal and adult rats.Arch. Toxicol. 67, 510–513.
Casida, J.E., Gammon, D.W., Glickman, A.H., Lawrence,L.J., 1983. Mechanisms of selective action of pyrethroidinsecticides. Ann. Rev. Pharmacol. Toxicol. 23, 413–438.
Chomczynski, P., Sacchi, N., 1987. Single-step method ofRNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162, 156–159.
Dorman, D.C., Beasley, V.R., 1991. Neurotoxicology ofpyrethrin and the pyrethroid insecticides. Vet. Hum. Toxi-col. 33, 238–243.
Eells, J.T., Bandettini, P.A., Holman, P.A., Propp, J.M., 1992.Pyrethroid insecticide-induced alterations in mammaliansynaptic membrane potential. J. Pharmacol. Exp. Ther.262, 1173–1181.
Elshourbagy, N.A., Liao, W.S., Mahley, R.W., Taylor, J.M.,1985a. Apolipoprotein E mRNA is abundant in the brainand adrenals, as well as in the liver, and is present in other

L. Aldana et al. / Toxicology Letters 95 (1998) 31–39 39
peripheral tissues of rats and marmosets. Proc. Natl. Acad.Sci. USA 82, 203–207.
Elshourbagy, N., Boguski, M.S., Liao, W.S.L., Jefferson, L.S.,Gordon, J.I., Taylor, J.M., 1985b. Expression of ratapolipoprotein A-IV and A-1 genes mRNA induction dur-ing development and in response to glucocorticoids andinsulin. Proc. Natl. Acad. Sci. USA 82, 8242–8246.
El-Toukhy, M.A., Girgis, R.S., 1993. In vivo and in vitrostudies on the effect of larvin and cypermethrin onadenosine triphosphatase activity of male rats. J. Environ.Sci. Health B28, 599–619.
Flodstrom, S., Warngard, L., Ljungquist, S., Ahlborg, U.G.,1988. Inhibition of metabolic cooperation in vitro andenhancement of enzyme altered foci incidence in rat liverby the pyrethroid insecticide fenvalerate. Arch. Toxicol. 61,218–223.
Gray, A.J., 1985. Pyrethroid structure-toxicity relationships inmammals. Neurotoxicology 6, 127–138.
Krechniak, J., Wrzesniowska, K., 1991. Effects of pyrethroidinsecticides on hepatic microsomal enzymes in rats. Envi-ron. Res. 55, 129–134.
Lehnus, G., Smith, L., 1978. Automated procedure for kineticmeasurement of total triglycerides (as glycerol) in serumwith the Gilford system 3500. Clin. Chem. 24, 27–31.
Miyamoto, J., 1976. Degradation, metabolism and toxicity ofsynthetic pyrethroids. Environ. Health Perspect. 14, 15–28.
Narahashi, T., 1985. Nerve membrane ionic channels as theprimary target of pyrethroids. Neurotoxicology 6, 3–22.
Nuno, P., Hernandez, A., Mendoza-Figueroa, T., Panduro,A., 1997. Viscosity regulates apolipoprotein A-1 gene ex-pression in experimental models of secondary hyperlipi-demia and in cultured hepatocytes. Biochim. Biophys. Acta1344, 262–269.
Panduro, A., Shalaby, F., Weiner, F., Biempica, L., Zern, M.,Shafritz, D., 1986. Transcriptional switch from albumin toa-fetoprotein and changes in transcription of other genesduring carbon tetrachloride induced liver regeneration.Biochemistry 25, 1414–1420.
Panduro, A., Shalaby, F., Shafritz, D.A., 1987. Changingpatterns of transcriptional and post-transcriptional controlof liver-specific gene expression during rat development.Genes Dev. 1, 1172–1182.
Panduro, A., Shalaby, F., Biempica, L., Shafritz, D.A., 1988.Changes in albumin, a-fetoprotein and collagen gene tran-scription in CCl4-induced fibrosis. Hepatology 8, 259–265.
Panduro, A., Lin-Lee, Y., Chan, L., Shafritz, D.A., 1990.Transcriptional and post-transcriptional regulation ofapolipoprotein E, A-1 and A-II gene expression in normalrat liver and during several pathophysiologic states. Bio-chemistry 29, 8430–8435.
Panduro, A., Contreras, J.L., Sigler, A., Gonzalez, L., Poo,J.L., Diliz, H.S., 1992. Molecular markers of liver trans-plantation: effect of rapid cooling on the expression ofhepatic genes. Transplant. Proc. 24, 1976–1978.
Panduro, A., Castrillon, L., Gonzalez, L., Shafritz, D.A.,1993. Regulation of hepatic and non-hepatic apolipo-protein A-1 and E gene expression during liver regenera-tion. Biochim. Biophys. Acta 1167, 37–42.
Reddy, A.T.V., Ayyanna, K., Yellamma, K., 1991. Cyperme-thrin induced modulations in lipid metabolism of freshwa-ter teleost, Tilapia mossambica. Biochem. Int. 23, 963–967.
Shakoori, A.R., Ali, S.S., Saleem, M.A., 1988. Effects of sixmonths feeding of cypermethrin on the blood and liver ofalbino rats. J. Biochem. Toxicol. 3, 59–71.
Sheperd, J., 1994. Lipoprotein metabolism, an overview.Drugs 47 (Suppl. 2), 1–10.
Shing, A., Agarwal, R.A., 1993. Effect of cypermethrin onlactate and succinic dehydrogenase and cytochrome oxi-dase of snail and fish. Bull. Environ. Contam. Toxicol. 51,445–452.
Thomas, P.S., 1980. Hybridization of denatured RNA andsmall DNA fragments transferred to nitrocellulose. Proc.Natl. Acad. Sci. USA 77, 5201–5205.
Vershoyle, R.D., Aldridge, W.N., 1980. Structure-activity rela-tionships of some pyrethroids in rats. Arch. Toxicol. 45,325–329.
World Health Organization, 1989. Cypermethrin. Environ.Health Criteria 82, 85–92.
.





























