Embed Size (px)
Citation preview

Cyclic ADP-ribose, a putative Ca2�-mobilizing second messenger, operates insubmucosal gland acinar cells
Kan Sasamori,1 Tsukasa Sasaki,1 Shin Takasawa,2 Tsutomu Tamada,1 Masayuki Nara,1
Toshiya Irokawa,1 Sanae Shimura,1 Kunio Shirato,3 and Toshio Hattori1
Divisions of 1Respiratory and Infectious Diseases and 3Cardiovascular Medicine, Tohoku UniversityGraduate School of Medicine, Sendai 980-8574; and 2Department of Biochemistry,Tohoku University Graduate School of Medicine, Sendai 980-8575, Japan
Submitted 24 December 2003; accepted in final form 18 February 2004
Sasamori, Kan, Tsukasa Sasaki, Shin Takasawa, TsutomuTamada, Masayuki Nara, Toshiya Irokawa, Sanae Shimura,Kunio Shirato, and Toshio Hattori. Cyclic ADP-ribose, a putativeCa2�-mobilizing second messenger, operates in submucosal glandacinar cells. Am J Physiol Lung Cell Mol Physiol 287: L69–L78,2004. First published February 27, 2004; 10.1152/ajplung.00454.2003.—Cyclic ADP-ribose (cADPR), a putative Ca2�-mobilizingsecond messenger, has been reported to operate in several mammaliancells. To investigate whether cADPR is involved in electrolyte secre-tion from airway glands, we used a patch-clamp technique, themeasurement of microsomal Ca2� release, quantification of cellularcADPR, and RT-PCR for CD38 mRNA in human and feline trachealglands. cADPR (�6 �M), infused into the cell via the patch pipette,caused ionic currents dependent on cellular Ca2�. Infusions of lowerconcentrations (2–4 �M) of cADPR or inositol 1,4,5-trisphosphate(IP3) alone were without effect on the baseline current, but a com-bined application of cADPR and IP3 mimicked the cellular responseto low concentrations of acetylcholine (ACh). Microsomes derivedfrom the isolated glands released Ca2� in response to both IP3 andcADPR. cADPR released Ca2� from microsomes desensitized to IP3
or those treated with heparin. The mRNA for CD38, an enzymeprotein involved in cADPR metabolism, was detected in humantissues, including tracheal glands, and the cellular content of cADPRwas increased with physiologically relevant concentrations of ACh.We conclude that cADPR, in concert with IP3, operates in airwaygland acinar cells to mobilize Ca2�, resulting in Cl� secretion.
patch clamp; CD38; calcium store; inositol 1,4,5-trisphosphate; cal-cium; acetylcholine
TRANSEPITHELIAL Cl� secretion is critical for the production ofproperly hydrated secretion in airways, thus contributing to theairway defense mechanisms. Recurrent intractable airway in-fection in cystic fibrosis (CF), a hereditary disease with com-promised airway Cl� secretion, attests to the importance of thisfunction since lung disease is the primary cause of mortality inCF patients (8, 64). Because human airway epithelium is likelyto be primarily absorptive (5, 22, 26, 62, 63), a major fractionof the airway fluid seems to be derived from the submucosalgland (2, 48) and follows an active Cl� secretion from theglandular acini (11, 14, 48). The airway secretion appears to beunder tonic regulation by cholinergic nerves because oraladministration of atropine reduces the secretion (35) and vagalcooling reduces it by �40% (59). An in vivo observationindicated that cholinergic agents were much more potent stim-
ulators of gland secretion than were adrenergic agonists whenestimated using hillock formations of a powdered tantalumlayer coating the airway surface (43). Also, primary cultures ofhuman submucosal gland secreted Cl� in response to bothcholinergic and adrenergic stimulation, with cholinergic stim-ulation being more potent (65). The acinar cells of the airwaysubmucosal gland are the predominant site for the expressionof cystic fibrosis transmembrane conductance regulator(CFTR), a protein that is flawed in CF disease (2, 12, 64).Recently, CFTR has been shown to be involved in cholinergi-cally stimulated, Ca2�-activated Cl� secretion in culturedhuman airway cells (42) and in porcine bronchial submucosalgland (2). These findings demonstrate the major role of Ca2�-mediated Cl� secretion stimulated by cholinergic agents inairway submucosal glands. However, the cellular mechanismof Ca2� mobilization that ultimately leads to activation of Cl�
channels is only partly understood in airway submucosalglands.
The muscarinic receptors have been identified on swine (67),ferret (49), and feline (21) submucosal glands. Muscarinicagonists activate, via a guanine nucleotide-binding protein (Gprotein) coupled with the receptor, the plasma membranephospholipase C, generating inositol 1,4,5-trisphosphate (IP3)and diacylglycerol. IP3 binds to its specific receptor on thecytosolic Ca2�-storage compartment to release Ca2� into thecytosol, thus initiating the Ca2� signal (4). Inositol phosphateformation in submucosal glands has been shown in response toagents, including the muscarinic agonist carbachol, in bovineairway (18). Moreover, IP3-mediated Ca2� release has beenfound to play a critical role in starting acetylcholine (ACh)-evoked Cl� secretion in human and feline tracheal glands inpatch-clamp experiments and intracellular Ca2� measurements(51). Interestingly, however, prevention of the IP3-mediatedsignaling pathway by a MAb against IP3 receptors (IP3R) didnot totally abolish the ACh-induced response in tracheal glands(51). This observation raised the possibility that there may bean additional Ca2�-mobilizing mechanism independent of theIP3-mediated one in airway gland acinar cells.
A likely candidate for such a second messenger is cyclicADP-ribose (cADPR), a metabolite of the ubiquitous pyridinenucleotide �-NAD� (7, 46, 53). Since the identification ofcADPR as a potent mediator of Ca2� release in sea urchin eggmicrosomes (7), its Ca2�-mobilizing activity has been shownin rodent pituitary cell (27), dorsal root ganglion cell (10),
Address for reprint requests and other correspondence: K. Sasamori, Divi-sion of Respiratory and Infectious Diseases, Tohoku Univ. Graduate School ofMedicine, 1-1 Seiryo-machi, Aoba-ku, Sendai 980-8574, Japan (E-mail:[email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Lung Cell Mol Physiol 287: L69–L78, 2004.First published February 27, 2004; 10.1152/ajplung.00454.2003.
1040-0605/04 $5.00 Copyright © 2004 the American Physiological Societyhttp://www.ajplung.org L69

pancreatic �-cell (46, 53), and successively in rodent exocrineglands, including pancreas (56), salivary (32), and lacrimal(17) glands. Moreover, the CD38 protein, originally identifiedon leukocytes as one of the surface antigens, was revealed toretain enzymatic activities in both the synthesis and degrada-tion of cADPR (19, 54). CD38 appears to work in rat parotidacinar cells in raising the cellular level of cADPR (34). Thesefindings suggest that cADPR may be involved in the Ca2�
mobilization in exocrine acinar cells. However, none of thesereports (17, 32, 56) addressed whether cADPR can occurnaturally in response to physiological stimuli in exocrineglands. It is also unclear whether cADPR can operate not onlyin rodent glands but also in human acinar cells. To addressthese issues, electrophysiological, biochemical, and molecularbiological experiments were carried out using freshly isolatedhuman and feline tracheal submucosal glands. This is the firstreport that describes cADPR involvement in Ca2� homeostasisin human acinar cells, which may give some insights into themechanism underlying deteriorated Ca2�-activated Cl� secre-tion in CF submucosal glands.
METHODS
Cell preparations. Submucosal glands were isolated either from thetracheae of human surgical specimens from patients with laryngealcancer or those of cats (2–5 kg body wt) anesthetized with intramus-cular ketamine hydrochloride (30 mg/kg) and intravenous thiopentalsodium (30 mg/kg). The external surface of the cat trachea wascleaned of fat and connective tissues, cut into rings 3–4 cm long, andpinned in the extracellular solution with the posterior (membranous)wall side up. Light through a flexible fiber bronchoscope (FBS-1;Machida, Tokyo, Japan) placed inside the tracheal ring was used totransilluminate the membranous portion. In the human tracheal piece,the mucosal membrane together with the underlying connective tis-sues were peeled off from the cartilage layer and pinned on asilicon-rubber-coated chamber with the submucosal aspect directedupward. Light was shed from the lateral side of the tissue. Thesubmucosal gland could then be easily distinguished from the sur-rounding connective tissue under a stereoscopic microscope (�60–80magnification). Fresh, unstained submucosal glands were isolatedusing two pairs of tweezers and microscissors (20, 21, 51, 55). Theisolated glands were further dispersed enzymatically into single orclustered acinar cells by incubating them with enzyme solutioncontaining collagenase (200 U/ml), DL-dithiothreitol (0.31 mg/ml),and trypsin inhibitor (1 mg/ml) for 30 min at 37°C. After dispersionand being washed three times with centrifugation at 180 g, the cells wereresuspended in a standard extracellular solution (see below) until use.
Electrical recordings. Ionic currents were measured with a patch-clamp amplifier (EPC9; HEKA Electronic), low-pass filtered at 2.9kHz, and monitored on both a built-in software oscilloscope and athermal pen recorder (RECTI-HORIZ-8K; Nippondenki San-ei, To-kyo, Japan). Patch pipettes were made of glass capillary with an outerdiameter of 1.5 mm using a vertical puller (PP-83; Narishige ScientificInstruments, Tokyo, Japan) and had a tip resistance of 2–6 M�. Thejunction potential between the patch pipette and bath solution wasnulled by the amplifier circuitry. After establishing a high resistance(�1 G�) tight seal, the whole cell configuration was obtained byrupturing the patch membrane with negative pressure applied to thepipette tip. Membrane currents were monitored at two differentholding potentials (Hp), i.e., 0 and/or �80 mV, each of which roughlycorresponded to the Cl� and K� equilibrium potential, respectively,under the present electrolyte conditions. The double current monitor-ing, i.e., alternate recording of the ionic currents corresponding to Hpof 0 and �80 mV, was accomplished by applying 200-ms voltagepulses of �80 mV at a frequency of 2 Hz to the pipette voltage of 0
mV (20, 50, 51, 55). The upward or downward deflection of thecurrent tracing represents outward (Io) or inward current (Ii), respec-tively. The solutions employed were of the following compositions (inmM): extracellular (bath) solution, 120 NaCl, 4.7 KCl, 1.13 MgCl2,1.2 CaCl2, 10 glucose, and 10 HEPES; and intracellular (pipette)solution, 120 KCl, 1.13 MgCl2, 0.5 EGTA, 1 Na2ATP, 10 glucose,and 10 HEPES. For the extracellular solution used in the experimentswith external free Ca2�, Ca2� was removed from the extracellularsolution and replaced with 1 mM EGTA. The fluids were superfusedover the cell(s) by hydrostatic pressure-driven application (20–30cmH2O) through polyethylene tubes. All solutions were at pH 7.2, andall experiments were carried out at room temperature (22–25°C).
Preparation of cADPR. cADPR was prepared enzymatically fromNAD� with ADP-ribosyl cyclase purified from ovotestis of Aplysiakurodai, a species common around the Japan coast (53). cADPR wasthen purified by HPLC and characterized by 1H nuclear magneticresonance and fast atom bombardment mass spectrometry.
Ca2� release assay. Microsomes were prepared as previouslydescribed (52, 53). In brief, cells were homogenized with a pelletmixer (Treff, Degersheim, Switzerland) in 0.2 ml of acetate intracel-lular medium composed of 250 mM potassium acetate, 250 mMN-methylglucamine, 1 mM MgCl2, and 20 mM HEPES (pH 7.2)supplemented with 0.5 mM ATP, 4 mM phosphocreatine, creatinephosphokinase (2 U/ml), 2.5 mM benzamidine, and 0.5 mM phenyl-methylsulfonyl fluoride. After the homogenates had been centrifugedfor 45 s at 13,000 g, the microsomes were prepared by Percoll densitygradient centrifugation at 20,000 g for 40 min at 10°C. Ca2� releasewas monitored in 3 ml of intracellular medium composed of 250 mMpotassium gluconate, 250 mM N-methylglucamine, 1 mM MgCl2, and20 mM HEPES (pH 7.2) supplemented with 1 mM ATP, 4 mMphosphocreatine, creatine phosphokinase (2 U/ml), 2.5 mM benzami-dine, 0.5 mM phenylmethylsulfonyl fluoride, and 3 �M fluo 3, afluorescent dye Ca2� indicator. Fluo 3 fluorescence was measured at490 nm excitation and 535 nm emission with a Jasco CAF-110intracellular ion analyzer (Tokyo, Japan) at 37°C with a circulatingwater bath. The amounts of Ca2� released were measured as the peakresponses to either agent. We added a 3-�l aliquot of 1 �M standardCa2� solution into the cuvette, giving rise to a final amount of 3 nmolof Ca2� in the bath. The Ca2� released in response to IP3 and/orcADPR was estimated by comparing with the standard 3 nmol ofCa2� added.
Detection of CD38 mRNA. Total RNAs were isolated as describedpreviously (54). RT-PCR was carried out as described (54). Thesequences of sense and antisense primers used for CD38 were 5-GCTCTAGAGCCCTATGGCCAACTGCGAGTT-3 and 5-GCTCTAGAGCTCAGATCTCAGATGTGCA-3, respectively (54),and those for glyceraldehyde-3-phosphate dehydrogenase (GAPDH)were 5-CATTGACCTCAACTACATGGT-3 and 5-TTGTCATAC-CAGG-AAATGAGC-5, corresponding to nucleotides 112–132 and928–948 of human GAPDH cDNA (1), respectively. The PCR con-ditions were as follows: denaturation at 94°C for 30 s, annealing at68°C (CD38) or at 60°C (GAPDH) for 1 min, and extension at 72°Cfor 2 min. The cycle was repeated 40 times for CD38 and 30 times forGAPDH as an internal control. The reaction mixture was analyzed ona 1.5% agarose gel in HCl/acetate/EDTA buffer.
Quantification of cellular IP3. The cellular content of IP3 wasestimated with a D-myo-inositol 1,4,5-trisphosphate [3H] assay system(TRK 1000; Amersham Pharmacia Biotech UK, Buckinghamshire,UK) either in the presence or absence of ACh (10�9, 10�8, or 10�7
M). The suspension of enzymatically dispersed acinar cells in 1 ml ofexternal solution (with or without ACh) in 2-ml test tubes wasincubated for 1 min, and the supernatant was discarded after centrif-ugation at 2,000 g for 5 min at 4°C and stocked at �80°C. The cellswere homogenized in 200 �l of ice-cold 10% (vol/vol) perchloric acidwith an ultrasonic processor (Astrason; Heat Systems, Farmingdale,NY) for 10 s. The samples were then neutralized by the addition of0.15 M KOH (1 ml), and a 2-�l aliquot was kept for protein
L70 IP3 AND CADPR MIMICS THE ACH-INDUCED RESPONSES IN SUBMUCOSAL GLANDS
AJP-Lung Cell Mol Physiol • VOL 287 • JULY 2004 • www.ajplung.org

quantification. After sedimentation of KClO4 by centrifugation at2,000 g for 15 min at 4°C, the supernatant was used for IP3 measure-ment. The assay was performed according to the manufacturer’sinstructions.
Quantification of cellular cADPR. The cells were prepared with thesame protocol as described above. The frozen pellet samples werehomogenized in 370 �l of perchloric acid solution (2.5%, vol/vol) andcentrifuged for 10 min at 13,000 g. cADPR was extracted andconcentrated as follows (52). The supernatant (300–350 �l) of thehomogenates was then mixed with 150 �l of a suspension of Norit A(27 mg/ml in H2O; Nacalai tesque, Kyoto, Japan). After a 30-minincubation at 37°C, the samples were again centrifuged, and thesupernatant was discarded. The pellet was washed three times with 1.0ml of H2O, resuspended in a pyridine/ethanol/H2O mixture (10:50:40,vol/vol/vol), and incubated for 120 min at 37°C. After a furthercentrifugation, the supernatant was collected and evaporated (Speed-vac; Savant Instrument, Farmingdale, NY). The recovery of cADPR,monitored by the recovery of [3H]cADPR added in each homogenate,was 55.8 2.61% (n � 20). Correction was introduced for therecovery of cADPR. The cADPR content of the cell extracts wasmeasured by a radioimmunoassay. Briefly, the evaporated materialseluted from Norit A charcoal were resuspended with 50 �l of H2O andthen incubated at 25°C for 2 h with bovine alkaline phosphatase andvenom phosphoesterase (Worthington) at final concentrations of 50and 2 U/ml, respectively, in 100 mM imidazole HCl (pH 7.5), 2 mMMgCl2, 100 mM NaCl, and 400 mM KCl. The reaction was termi-nated by adding a solution of trichloroacetic acid (at a final concen-tration of 4%, wt/vol) and was kept on ice for 20 min. A clearsupernatant was obtained after centrifugation at 13,000 g for 10 min.An aliquot (10–20 �l) of the supernatant was immediately neutralizedwith a solution of 2 M Tris base and subjected to the procedure forcADPR measurement (52).
Animal care and human surgical specimen. This study was ap-proved by the Animal Care and Use Committee and the EthicsCommittee on Human Investigations of the Tohoku University Schoolof Medicine. The care and handling of the animals were performed inaccordance with National Institutes of Health guidelines for the careand handling of animals (60).
Reagents. Fluo 3 was purchased from Molecular Probes (Eugene,OR). Tetraethylammonium (TEA)-Cl and collagenase were fromWako Pure Chemicals, Osaka, Japan. HEPES was from DojindoLaboratories, Kumamoto, Japan. Charybdotoxin was from PeptideInstitute, Osaka, Japan. FK-506 was a generous gift from FujisawaPharmaceuticals, Osaka, Japan. All other reagents were from SigmaChemical, St. Louis, MO.
RESULTS
ACh-induced ionic currents in submucosal glands. The freshsubmucosal gland acinar cells generated ionic currents in thepresence of a wide range of ACh concentrations (4 � 10�9-10�6 M). Lower concentrations of ACh (up to several tens ofnM) induced repetitive transient activations of the ionic cur-rents recorded at Hp of 0 and �80 mV. The ionic currentactivated at 0 mV Hp (the upward deflection of the current-trace shown in Fig. 1A; Io) was abolished by charybdotoxin, aspecific inhibitor of the Ca2�-activated large conductance K�
channel (BK channel) (39) (100–200 nM, n � 5; Fig. 1A). Thedownward deflection of the current recorded at �80 mV (Ii)was inhibited in the presence of diphenylamine-2-carboxylate(DPC, 1 mM, n � 6), a Cl� channel inhibitor (2) (Fig. 1B).TEA (2–5 mM), a known inhibitor of BK channels (28),abolished both Io and Ii (data not shown, n � 8). As reportedpreviously (20, 51), extremely high concentrations of AChinduced a sustained (not oscillatory) activation (see Fig. 2, A
and B), and the sustained Io and Ii were carried by K� and Cl�,respectively, as evidenced by experiments using channelblockers and electrolyte-replacement studies (51). Thus theregenerative current spikes (or the oscillation) activated by lowconcentrations of ACh shared the same characteristics with thesustained currents stimulated by high doses of ACh. Theseionic currents were abolished totally either with the extracel-lular membrane-permeable Ca2� chelator BAPTA-AM (50�M, n � 3) or with intracellular high EGTA (10 mM, n � 3),indicating Ca2� dependence of the ACh-stimulated ionic cur-rents (data not shown). As shown in Fig. 1, C–E, the oscillatoryIi increased in both frequency and amplitude as the concentra-tion of ACh was raised, although the sensitivity to ACh waslargely variable from cell to cell. A sustained current compo-nent became visible and was accompanied by oscillatory acti-vation with stronger stimuli (Fig. 1, B and E).
Effects of FK-506 and ryanodine on the oscillatory Cl�
current. FK-506 (tacrolimus, 10 �g/ml), an immunosuppres-sant drug, abolished the ACh-induced oscillatory Cl� current(n � 8, Fig. 1F). Ryanodine (10 �M), an agent that acts via theendoplasmic reticulum Ca2� release channel ryanodine recep-tor (RyR), also inhibited Ii in a way similar to FK-506 (n � 12,Fig. 1G), although the duration required for the recovery wasfar longer with FK-506.
Actions of IP3 and/or cADPR introduced into cytoplasm.FK-506 has been shown to bind to one of the immunophilinFK-506-binding proteins (FKBP) associated with the RyR (29,45) or IP3R (6), thus regulating the intracellular release ofCa2�. We, therefore, performed experiments with intracellularinfusion of IP3 and/or cADPR, a putative RyR agonist, intohuman tracheal gland acinar cells. As shown in Fig. 2, wechose relatively high (�20 �M, Fig. 2, C–E) and low (2–4�M, Fig. 2, F–H) doses of IP3 (or cADPR) for infusing into thecytoplasm via the patch pipette. This was intended to recon-stitute the bioelectric responses of the gland to ACh in high andlow concentrations. The infusion of highly concentrated IP3
alone activated abrupt but transient responses in both directions(n � 7, Fig. 2C), which closely mimicked the initiation of AChresponses in both human and feline tracheal acinar cells (Fig.2, A and B) but lacked the successive activation. A high doseof cADPR (23 �M) included in the patch pipette activated bothIo and Ii (n � 7) with a gradual onset without the initial surgein Ii (Fig. 2D). In contrast, the combined infusion of IP3 andcADPR (both 20 �M) induced a rapid and long-lasting currentin the presence of extracellular Ca2� (n � 4, Fig. 2E), whereasthe current decayed rapidly in the absence of external Ca2�
(n � 4, data not shown).As shown in Fig. 2, F and G, infusions of 2–4 �M of IP3
alone or 2 �M of cADPR alone did not induce any responsesup to 12 min after the establishment of the whole cell config-uration (n � 11 for IP3 and n � 16 for cADPR). However, thecombined infusion of IP3 and cADPR (2 �M each) evokedoscillatory current responses with quiet intervals of 1–2 minafter the establishment of the whole cell configuration (4positive responses out of 9 experiments, Fig. 2H). The pipettecADPR concentration required for the activation of the ioniccurrent was �5 �M. As shown in Fig. 2I, 6 �M cADPRincluded in the pipette, however, did not induce an oscillationbut stimulated gradually the Io alone without Ii (n � 3).
In a separate series of experiments, we applied cADPR onthe cytosolic aspect of excised inside-out patches (n � 18).
L71IP3 AND CADPR MIMICS THE ACH-INDUCED RESPONSES IN SUBMUCOSAL GLANDS
AJP-Lung Cell Mol Physiol • VOL 287 • JULY 2004 • www.ajplung.org

However, we did not find any channels activated in the pres-ence of cADPR (4–20 �M). cADPR did not affect the activityof the Ca2�-dependent large conductance K� channels con-tained in the isolated patches (n � 7).
Measurement of microsomal Ca2� release. We then inves-tigated the presence or absence of the intracellular Ca2� poolthat was sensitive to these agents. The microsome fractionobtained from the feline tracheal glands responded to both IP3
L72 IP3 AND CADPR MIMICS THE ACH-INDUCED RESPONSES IN SUBMUCOSAL GLANDS
AJP-Lung Cell Mol Physiol • VOL 287 • JULY 2004 • www.ajplung.org

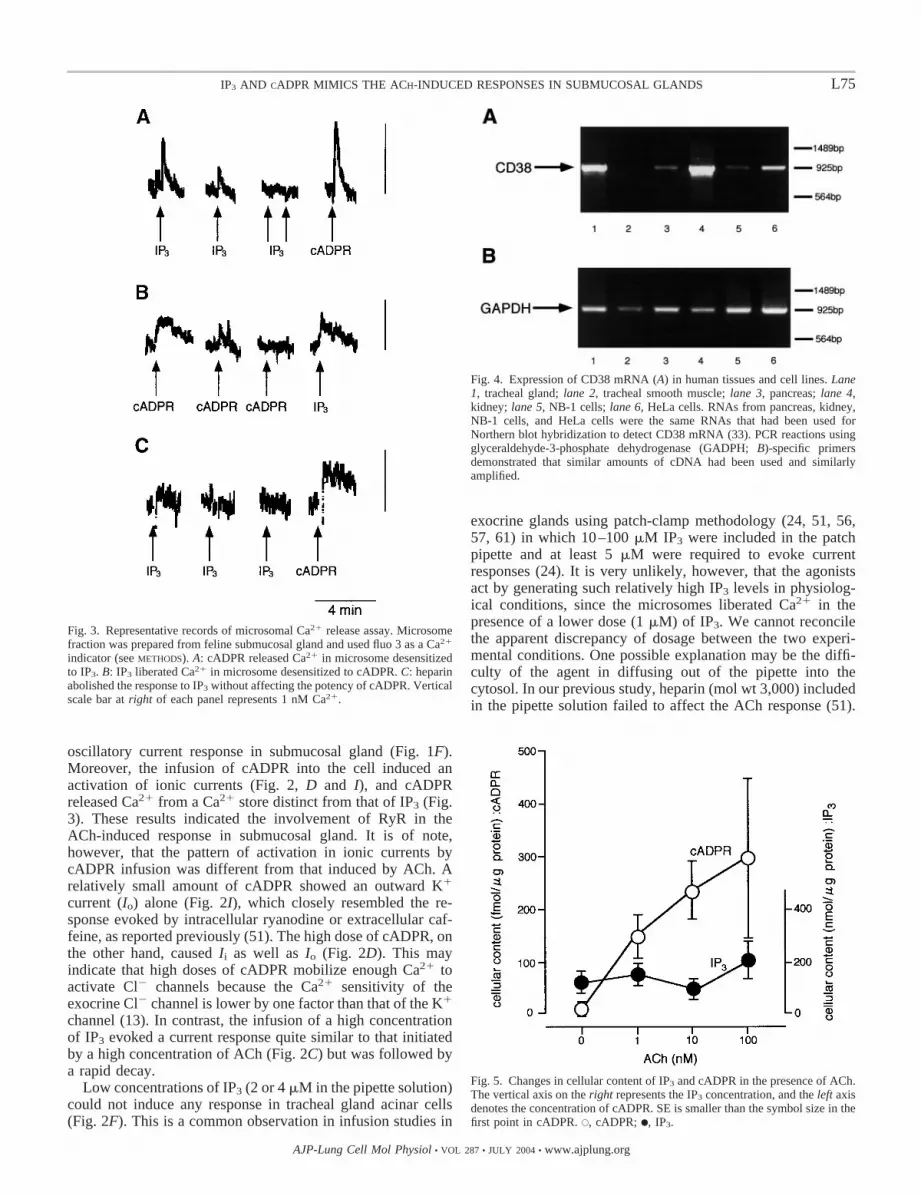
and cADPR to release Ca2�. IP3 and cADPR (1 �M each)released 0.65 0.13 (n � 4, means SE) and 0.86 0.10nmol (n � 4) of Ca2�, respectively. The protein content in thecuvette was 105 �g/3 ml. The Ca2� stores sensitive to eitheragent were independent of each other because the microsomesdesensitized with IP3 by repetitive stimulations still respondedto cADPR, or vice versa (n � 5, Fig. 3, A and B). TheIP3-induced Ca2� release was abolished by adding heparin(250 �g/ml) to the bath, but cADPR still released Ca2� fromthe heparin-treated microsomes (n � 2, Fig. 3C).
Detection of CD38 mRNA. CD38 has been shown to catalyzethe formation and degradation of cADPR (19, 25, 34, 54). Toexamine whether the airway glands have the ability to synthe-size cADPR, we carried out experiments to detect the mRNAof CD38 using human materials, including submucosal glands(Fig. 4). CD38 mRNA was expressed strongly in humantracheal submucosal glands by RT-PCR (Fig. 4) as well as inexocrine pancreas but not in tracheal smooth muscle.
Changes in cellular content of IP3 and cADPR by ACh. Wenext examined whether cADPR can occur naturally in thepresence of physiological concentrations of ACh. We mea-sured the cellular content of IP3 and cADPR stimulated for 1min with 10�9 to 10�7 M ACh. The IP3 contents were notchanged within the concentration range of ACh used, i.e.,116.6 24.9, 126.8 41.1, 85.7 32.4, and 222.7 81.6nmol/�g protein for control, 10�9, 10�8, and 10�7 M of ACh,respectively (n � 4 each; P � 0.05 by ANOVA). The meanprotein content in each sample was 124.0 21.1 �g/ml (n �16). In contrast, the cADPR content was markedly raised byACh at a concentration as low as 10�9 M, i.e., the nontreatmentcontrol gland yielded 4.4 1.2 fmol/�g of protein (n � 10)and was increased to 152.0 40.5, 233.7 54.9, and 295.0 154.7 in the presence of 10�9, 10�8, and 10�7 M ACh,respectively (n � 3–4). These results are summarized in Fig. 5.
DISCUSSION
The airway submucosal gland is a mixed serous and mucousgland, and the serous phenotype consists of �60% of the cellpopulation (3). We could not discriminate the cell types underthe present experimental settings. However, both cell typesappeared to be activated by ACh to generate ionic currentsbecause most of the cells (�95%) responded to high concen-trations of ACh (10�7 to 10�6 M) with a strikingly similarpattern, although �20% of the cells failed to respond to a lowconcentration of ACh (10�8 M) (51, 55). Both the mucous and
serous cells have been shown to respond equally to cholinergicstimuli to cause secretions in a rheological (33) and a morpho-logical (58) experiment. In addition, an earlier bioelectric studyperformed on submaxillary salivary gland, which is a mixedgland similar to the submucosal gland, revealed that all theacinar cells responded to chorda tympani stimulation throughrandomized intracellular microelectrode recordings (36). Theseobservations support our notion that the ionic currents could beinduced by ACh regardless of the cell type.
The rise in intracellular Ca2� concentration ([Ca2�]i) canactivate Ca2�-sensitive ion channels on the plasma membrane,and, in exocrine acinar cells, the opening of Ca2�-gated Cl�
channels leads to a release of Cl� out of the cell, which hasbeen correlated with water secretion physiologically (47, 66).In the present study, ACh stimulated Io and Ii in trachealsubmucosal glands, corresponding to the efflux of K� and Cl�
from the cell, respectively, since Io was inhibited by charyb-dotoxin and Ii was blocked by DPC. In addition, the totalreplacement of K� with Cs� abolished Io, and the substitutionof Cl� with gluconate (or SO
4
2�) inhibited the major fraction ofIi (51). Both the currents were dependent on [Ca2�]i becausethe membrane-permeable Ca2� chelator BAPTA-AM abol-ished the currents and because the cells perfused with a highEGTA could no longer respond to ACh. In the presence ofphysiologically relevant concentrations of ACh, Io and Ii
showed oscillatory activations (Fig. 1), the magnitudes ofwhich increased with the augmentation of the stimulus, andfinally gave rise to sustained activations with extreme concen-trations of ACh (Fig. 2, A and B). These steps may reflect theregulation of fluid secretion in physiological situations. Inter-estingly, both the oscillatory Io and Ii in the presence oflow-ACh concentrations were inhibited concurrently by TEA,a known BK channel blocker (28). A similar observation hasbeen reported in sheep parotid gland (9) and was attributable tothe atropine-like action of TEA.
Mobilization of intracellular Ca2� in mammalian exocrineglands is mediated by two major mechanisms: IP3-inducedCa2� release and Ca2�-induced Ca2� release (CICR) (61),although the latter mechanism seemed absent in avian nasalgland (37). Accumulating evidence suggests that cADPR maybe an endogenous regulator of CICR through RyR (15, 29, 30,31, 41, 53, 56). In addition, FK-506 has been shown to disturbthe Ca2� release due to a competition with cADPR in bindingto FKBP that is allied with the RyR (29, 45) or IP3R (6). In thepresent study, FK-506 and ryanodine reversibly abolished the
Fig. 1. Representative original recordings showing acetylcholine (ACh)-induced ionic currents in tracheal submucosal gland acinarcells. A conventional whole cell patch-clamp technique was used. A: membrane currents were monitored at 2 different holdingpotentials (Hp), i.e., 0 and �80 mV, each of which roughly corresponded to the Cl� and K� equilibrium potential, respectively.This dual current monitoring was accomplished by applying 200-ms voltage pulses of �80 mV at a frequency of 2 Hz to the pipetteHp of 0 mV. The upward or downward deflection of the current tracing represents outward (Io) or inward current (Ii), respectively.The periodic spikes imposed on the current traces (looking like a band of shadow) are parts of the capacitance currents of the plasmamembrane. These artificial currents arise at the respective moments, just after the switching of the Hp from 0 to �80 mV, or theopposite. In B–G, ionic currents were monitored with Hp fixed at �80 mV so that only Ii was recorded. In the presence of lowconcentrations of ACh, the submucosal gland generated repetitive spiky ionic currents. A: Io was abolished by charybdotoxin(ChTx), a K� channel blocker. B: Ii was abolished by diphenylamine-2-carboxylate (DPC), one of the Cl� channel blockers. C–E:original current recordings showing the effect of increasing doses of ACh. These traces were obtained from the same cell withincreasing concentrations of ACh. The oscillatory response was increased in frequency and amplitude with incremental ACh, givingrise to a sustained component mixed with the spiky currents (E). The sustained component is also shown in B, in which the activatedIi returned to baseline with the Cl� channel blocker DPC. As discernible from these recordings, the sensitivity to ACh was variablefrom cell to cell, which may reflect differences in the cell population, i.e., serous and mucous cells. F and G: representativerecordings showing the effects of extracellular FK-506 and ryanodine on the ACh-induced oscillatory Ii. Both the agents inhibitedIi in a reversible manner. FK-506 required a longer interval for the cell to regain activity.
L73IP3 AND CADPR MIMICS THE ACH-INDUCED RESPONSES IN SUBMUCOSAL GLANDS
AJP-Lung Cell Mol Physiol • VOL 287 • JULY 2004 • www.ajplung.org
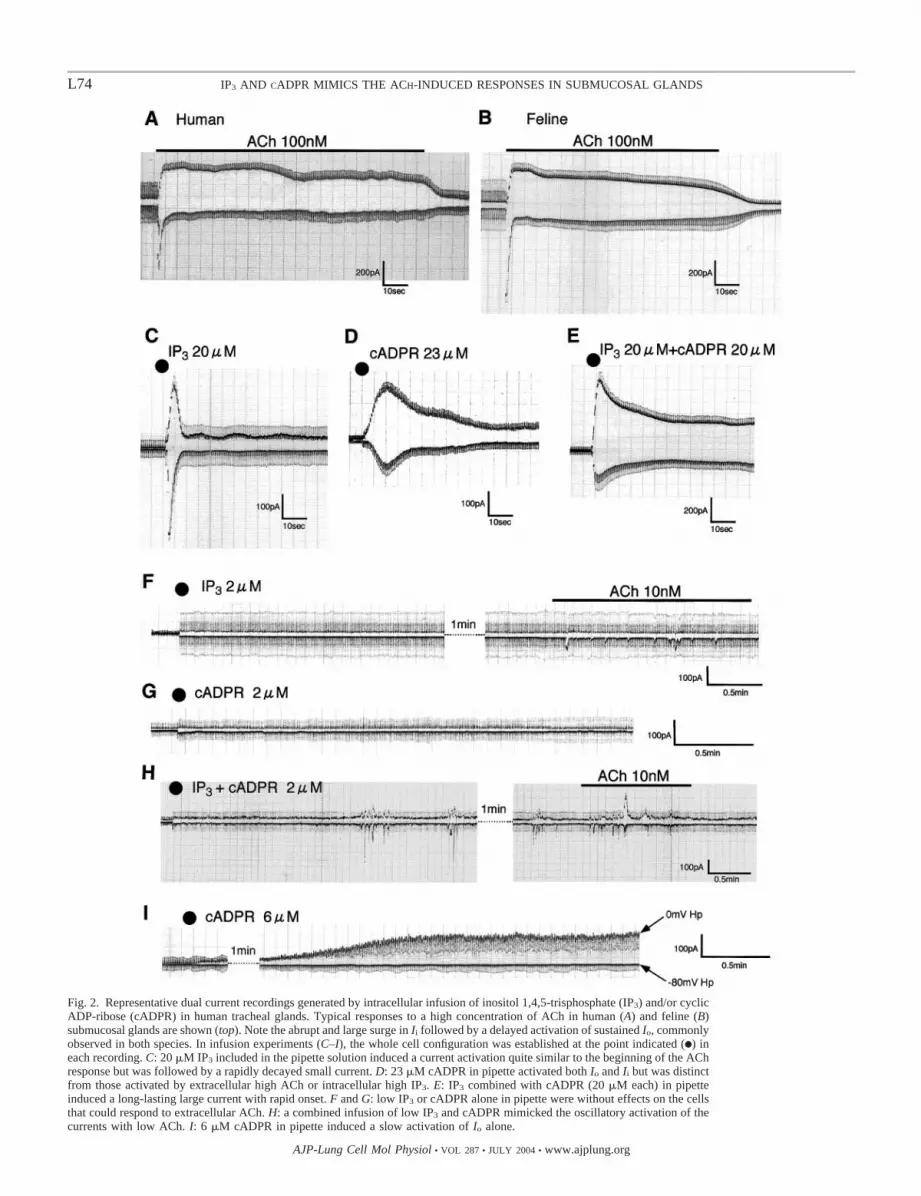
Fig. 2. Representative dual current recordings generated by intracellular infusion of inositol 1,4,5-trisphosphate (IP3) and/or cyclicADP-ribose (cADPR) in human tracheal glands. Typical responses to a high concentration of ACh in human (A) and feline (B)submucosal glands are shown (top). Note the abrupt and large surge in Ii followed by a delayed activation of sustained Io, commonlyobserved in both species. In infusion experiments (C–I), the whole cell configuration was established at the point indicated (F) ineach recording. C: 20 �M IP3 included in the pipette solution induced a current activation quite similar to the beginning of the AChresponse but was followed by a rapidly decayed small current. D: 23 �M cADPR in pipette activated both Io and Ii but was distinctfrom those activated by extracellular high ACh or intracellular high IP3. E: IP3 combined with cADPR (20 �M each) in pipetteinduced a long-lasting large current with rapid onset. F and G: low IP3 or cADPR alone in pipette were without effects on the cellsthat could respond to extracellular ACh. H: a combined infusion of low IP3 and cADPR mimicked the oscillatory activation of thecurrents with low ACh. I: 6 �M cADPR in pipette induced a slow activation of Io alone.
L74 IP3 AND CADPR MIMICS THE ACH-INDUCED RESPONSES IN SUBMUCOSAL GLANDS
AJP-Lung Cell Mol Physiol • VOL 287 • JULY 2004 • www.ajplung.org
oscillatory current response in submucosal gland (Fig. 1F).Moreover, the infusion of cADPR into the cell induced anactivation of ionic currents (Fig. 2, D and I), and cADPRreleased Ca2� from a Ca2� store distinct from that of IP3 (Fig.3). These results indicated the involvement of RyR in theACh-induced response in submucosal gland. It is of note,however, that the pattern of activation in ionic currents bycADPR infusion was different from that induced by ACh. Arelatively small amount of cADPR showed an outward K�
current (Io) alone (Fig. 2I), which closely resembled the re-sponse evoked by intracellular ryanodine or extracellular caf-feine, as reported previously (51). The high dose of cADPR, onthe other hand, caused Ii as well as Io (Fig. 2D). This mayindicate that high doses of cADPR mobilize enough Ca2� toactivate Cl� channels because the Ca2� sensitivity of theexocrine Cl� channel is lower by one factor than that of the K�
channel (13). In contrast, the infusion of a high concentrationof IP3 evoked a current response quite similar to that initiatedby a high concentration of ACh (Fig. 2C) but was followed bya rapid decay.
Low concentrations of IP3 (2 or 4 �M in the pipette solution)could not induce any response in tracheal gland acinar cells(Fig. 2F). This is a common observation in infusion studies in
exocrine glands using patch-clamp methodology (24, 51, 56,57, 61) in which 10–100 �M IP3 were included in the patchpipette and at least 5 �M were required to evoke currentresponses (24). It is very unlikely, however, that the agonistsact by generating such relatively high IP3 levels in physiolog-ical conditions, since the microsomes liberated Ca2� in thepresence of a lower dose (1 �M) of IP3. We cannot reconcilethe apparent discrepancy of dosage between the two experi-mental conditions. One possible explanation may be the diffi-culty of the agent in diffusing out of the pipette into thecytosol. In our previous study, heparin (mol wt 3,000) includedin the pipette solution failed to affect the ACh response (51).
Fig. 4. Expression of CD38 mRNA (A) in human tissues and cell lines. Lane1, tracheal gland; lane 2, tracheal smooth muscle; lane 3, pancreas; lane 4,kidney; lane 5, NB-1 cells; lane 6, HeLa cells. RNAs from pancreas, kidney,NB-1 cells, and HeLa cells were the same RNAs that had been used forNorthern blot hybridization to detect CD38 mRNA (33). PCR reactions usingglyceraldehyde-3-phosphate dehydrogenase (GADPH; B)-specific primersdemonstrated that similar amounts of cDNA had been used and similarlyamplified.
Fig. 5. Changes in cellular content of IP3 and cADPR in the presence of ACh.The vertical axis on the right represents the IP3 concentration, and the left axisdenotes the concentration of cADPR. SE is smaller than the symbol size in thefirst point in cADPR. E, cADPR; F, IP3.
Fig. 3. Representative records of microsomal Ca2� release assay. Microsomefraction was prepared from feline submucosal gland and used fluo 3 as a Ca2�
indicator (see METHODS). A: cADPR released Ca2� in microsome desensitizedto IP3. B: IP3 liberated Ca2� in microsome desensitized to cADPR. C: heparinabolished the response to IP3 without affecting the potency of cADPR. Verticalscale bar at right of each panel represents 1 nM Ca2�.
L75IP3 AND CADPR MIMICS THE ACH-INDUCED RESPONSES IN SUBMUCOSAL GLANDS
AJP-Lung Cell Mol Physiol • VOL 287 • JULY 2004 • www.ajplung.org

Nevertheless, the microsome fraction treated with heparinshowed an attenuation of Ca2� release as also shown in thepresent experiments (Fig. 3). Presumably, heparin could notreach its effector sites due, perhaps, to the large-molecular-weight and/or to the electric charge. Another likely explanationfor the necessity of an extremely high amount of IP3 in evokingthe response is that an additional factor other than IP3 isinvolved in the signaling pathway employed by ACh.
Importantly, the combined application of IP3 and cADPRmimicked the responses induced by ACh both at higher con-centrations (Fig. 2E) and lower concentrations (oscillatoryresponse; Fig. 2H). These currents decayed rapidly in theabsence of external Ca2�, indicating the requirement of areplenishment of the internal Ca2� stores to maintain thecontinuous activation of the currents. Thus the ionic currentsstimulated by ACh could be reconstituted by IP3 with cADPRbut not by either agent alone.
In the present Ca2�-release assay, we demonstrated thepresence of a cADPR-sensitive Ca2� pool in submucosal glandmicrosomes (Fig. 3). The microsome fraction obtained fromthe whole gland is derived from morphologically distinct cellpopulations, i.e., serous and mucous cells. There is a possibilitythat IP3 and cADPR released Ca2� from different cell types.However, if IP3 and cADPR were synthesized separately indistinct cell types by ACh, we would have found at least twopatterns of current activations, because intracellular applica-tions of IP3 or cADPR alone elicited different and distinctpatterns of responses by the respective agents. Yet, as de-scribed above, almost all the cells responded to ACh with apattern strikingly similar to each other that was mimicked bythe combined infusion of IP3 and cADPR. This strongly sug-gested the coexistence of IP3- and cADPR-sensitive Ca2�
stores in a single cellular cytoplasmic compartment.CD38 has been found to be involved in cADPR metabolism
(19, 25, 54), acting as ADP-ribosyl cyclase for the synthesis ofcADPR and as cADPR hydrolase for its degradation. Weidentified CD38 mRNA in submucosal gland, which suggeststhe possibility that the cells synthesize cADPR. In pancreatic�-cells, the relative balance of the two catalytic activities isregulated by cellular ATP to control the cellular level ofcADPR (54). We do not have any evidence, so far, concerninga regulatory mechanism of the cADPR concentration throughmuscarinic receptor activation. It has long been known that thecGMP level is raised after muscarinic receptor activation inexocrine gland (23), but the physiological relevance of theincreased cGMP in these cells has not yet been established.Recently, cGMP was reported to increase [Ca2�]i by stimulat-ing cADPR synthesis in sea urchin eggs (16). We also foundthat nitric oxide, a cGMP enhancer, may act as a potentsecretagogue in human airway submucosal glands (44). Itwould be of interest to study the possibility that cGMP may actdirectly or via a cGMP-dependent phosphorylation on CD38protein, resulting in the increase in cADPR. Whatever thepossible mechanism, it was evident that the cellular content ofcADPR was markedly increased in response to physiologicalACh stimuli (Fig. 5). In contrast, the IP3 content was un-changed within the concentration range of ACh used in thepresent experiment (10�9 to 10�7 M). This was compatiblewith the report that measured 3H-labeled inositol phosphateformation in bovine submucosal gland (18). A significantincrease in the cellular inositol phosphate was observed at 10�6
M or higher concentrations of carbachol. Similarly, in a studyusing physiologically relevant concentrations of cholecystoki-nin on pancreatic acini, the investigators could not detect asignificant rise in IP3 with a radioreceptor assay, although anincrease in [Ca2�]i was evident (38).
We have shown that cADPR operates in submucosal glandacinar cells by acting on a Ca2� store distinct from that of IP3.The resultant increase in [Ca2�]i may activate K� channels inthe immediate vicinity of the cADPR-sensitive Ca2� store. Theactivation of the K� channels induces membrane hyperpolar-ization (51), forming an electrochemical environment favor-able to the efflux of Cl� ions (47) and to the influx of Ca2� intothe cytosol (40), thus assuring continuous Cl� secretion. Wepropose that IP3 and cADPR are both involved in a cooperativemanner in developing the ACh-induced response.
ACKNOWLEDGMENTS
We thank Dr. Hiroshi Okamoto for the cADPR measurements and for manyhelpful discussions. We also thank Brent K. Bell for reading the manuscript.
GRANTS
This work was supported by Grant-in-Aid 09670593 for Scientific Researchfrom the Ministry of Education, Science, Sports, and Culture, Japan.
REFERENCES
1. Allen RW, Trach KA, and Hoch JA. Identification of the 37-kDa proteindisplaying a variable interaction with the erythroid cell membrane asglyceraldehyde-3-phosphate dehydrogenase. J Biol Chem 262: 649–653,1987.
2. Ballard ST, Trout L, Bebok Z, Sorscher EJ, and Crews A. CFTRinvolvement in chloride, bicarbonate, and liquid secretion by airwaysubmucosal glands. Am J Physiol Lung Cell Mol Physiol 277: L694–L699, 1999.
3. Basbaum CB, Jany B, and Finkbeiner WE. The serous cell. Annu RevPhysiol 52: 97–113, 1990.
4. Berridge MJ and Irvine RF. Inositol phosphates and cell signaling.Nature 341: 197–205, 1989.
5. Boucher RC. Human airway ion transport. I. Am J Respir Crit Care Med150: 271–281, 1994.
6. Cameron AM, Steiner JP, Roskams AJ, Ali SM, Ronnet GV, andSnyder SH. Calcineurin associated with the inositol 1,4,5-trisphosphatereceptor-FKBP12 complex modulates calcium flux. Cell 83: 463–472,1995.
7. Clapper DL, Walseth TF, Dargie PJ, and Lee HC. Pyridine nucleotidemetabolites stimulates calcium release from sea urchin egg microsomesdesensitized to inositol trisphosphate. J Biol Chem 262: 9561–9568, 1987.
8. Collins FS. Cystic fibrosis: molecular biology and therapeutic implica-tions. Science 265: 774–779, 1992.
9. Cook DI, Wegman EA, Ishikawa T, Poronnik P, Allen DG, and YoungJA. Tetraethylammonium blocks muscarinically evoked secretion in thesheep parotid gland by a mechanism additional to its blockade of BKchannels. Pflugers Arch 420: 167–171, 1992.
10. Currie K, Swann K, Galione A, and Scott RH. Activation of Ca2�-dependent currents in cultured rat dorsal root ganglion neurones by asperm factor and cyclic ADP-ribose. Mol Biol Cell 3: 1415–1425, 1992.
11. Davis B. Mucous secretion and ion transport in airways. In: Textbook ofRespiratory Medicine, edited by Murray JF and Nadel JA. Philadelphia,PA: Saunders, 1988, p. 374–388.
12. Engelhard JF, Yankaskas JR, Ernst SA, Yang Y, Marino CR,Boucher RC, Cohn JA, and Wilson JM. Submucosal glands are thepredominant site of CFTR expression in human bronchus. Nat Genet 2:240–248, 1992.
13. Evans MG and Marty A. Calcium-dependent chloride currents in iso-lated cells from rat lacrimal glands. J Physiol 378: 437–460, 1986.
14. Finkbeiner WE, Shen BQ, and Widdicombe JH. Chloride secretion andfunction of serous and mucous cells of human airway glands. Am J PhysiolLung Cell Mol Physiol 267: L206–L210, 1994.
15. Galione A, Lee HC, and Busa WB. Ca2�-induced Ca2� release in seaurchin egg homogenate: modulation by cyclic ADP-ribose. Science 253:1143–1146, 1991.
L76 IP3 AND CADPR MIMICS THE ACH-INDUCED RESPONSES IN SUBMUCOSAL GLANDS
AJP-Lung Cell Mol Physiol • VOL 287 • JULY 2004 • www.ajplung.org

16. Galione A, White A, Willmott N, Turner M, Potter BVC, and WatsonSP. cGMP mobilizes intracellular Ca2� in sea urchin eggs by stimulatingcyclic ADP-ribose synthesis. Nature 365: 456–459, 1993.
17. Gromada J, Jørgensen TJ, and Dissing S. The release of intracellularCa2� in lacrimal acinar cells by �-, �-adrenergic and muscarinic cholin-ergic stimulation: the roles of inositol trisphosphate and cyclic ADP-ribose. Pflugers Arch 429: 751–761, 1995.
18. Hall IP. Agonist-induced inositol phosphate responses in bovine airwaysubmucosal glands. Am J Physiol Lung Cell Mol Physiol 262: L257–L262,1992.
19. Howard H, Grimaldi JC, Bazan JF, Lund FE, Santos-Argumedo L,Parkhouse RME, Walseth TF, and Lee HC. Formation and hydrolysisof cyclic ADP-ribose catalyzed by lymphocyte antigen CD38. Science262: 1056–1059, 1993.
20. Irokawa T, Sasaki T, Shimura S, Sasamori K, Oshiro T, Nara M,Tamada T, and Shirato K. Cholinomimetic action of macrolide antibi-otics on airway gland electrolyte secretion. Am J Physiol Lung Cell MolPhysiol 276: L951–L957, 1999.
21. Ishihara H, Shimura S, Satoh M, Masuda T, Nosaka H, Kase H,Sasaki T, Sasaki H, Takishima T, and Tamura K. Muscarinic receptorsubtypes in feline tracheal submucosal gland secretion. Am J Physiol LungCell Mol Physiol 262: L223–L228, 1992.
22. Iwase N, Sasaki T, Shimura S, Fushimi T, Okayama H, Hoshi H,Irokawa T, Sasamori K, Takahashi K, and Shirato K. Signature currentof SO2-induced bronchitis in rabbit. J Clin Invest 99: 1651–1661, 1997.
23. Jones CJ and Kealey T. Electrophysiological and dye-coupling studieson secretory, myoepithelial and duct cells in human eccrine sweat glands.J Physiol 389: 461–481, 1987.
24. Kasai H, Li YX, and Miyashita Y. Subcellular distribution of Ca2�
release channels underlying Ca2� waves and oscillations in exocrinepancreas. Cell 74: 669–667, 1993.
25. Kato I, Yamamoto Y, Fujimura M, Noguchi N, Takasawa S, andOkamoto H. CD38 disruption impairs glucose-induced increases in cyclicADP-ribose, [Ca2�]i and insulin secretion. J Biol Chem 274: 1869–1872,1999.
26. Kerem E, Bistritzer T, Hanukoglu A, Hofmann T, Zhou Z, Bennet W,MacLaughlin E, Barker P, Nash M, Quittel L, Boucher R, andKnowles MR. Pulmonary epithelial sodium-channel dysfunction andexcess airway liquid in pseudohypoaldosteronism. N Engl J Med 341:156–162, 1999.
27. Koshiyama H, Lee HC, and Tashjian Jr AH. Novel mechanism ofintracellular calcium release in pituitary cells. J Biol Chem 266: 16985–16988, 1991.
28. Kunzelmann K, Pavenstadt H, and Gregor R. Characterization ofpotassium channels in respiratory cells. II. Inhibitors and regulation.Pflugers Arch 414: 297–303, 1989.
29. Lam E, Martin MM, Timerman AP, Sabers C, Fleischer S, Lukas T,Abraham RT, O’Keefe SJ, O’Neill EA, and Widerrecht GJ. A novelFK506 binding protein can mediate the immunosuppressive effects ofFK506 and is associated with the cardiac ryanodine receptor. J Biol Chem270: 26511–26522, 1995.
30. Lee HC. Potentiation of calcium- and caffeine-induced calcium release bycyclic ADP-ribose. J Biol Chem 268: 293–299, 1993.
31. Lee HC, Aarhus R, and Walseth TF. Calcium mobilization by dualreceptors during fertilization of sea urchin eggs. Science 261: 352–355,1993.
32. Lee MG, Xu X, Zeng W, Diaz J, Wojeikievicz RJH, Kuo TH, WuytackF, Racymaekers L, and Muallem S. Polarized expression of Ca2�
channels in pancreatic and salivary gland cells. J Biol Chem 272: 15765–15770, 1997.
33. Leikauf GD, Ueki IF, and Nadel JA. Autonomic regulation of viscoelas-ticity of cat tracheal gland secretion. J Appl Physiol 56: 426–430, 1984.
34. Looms D, Nauntofte B, and Dissing S. ADP ribosyl cyclase activity inrat parotid acinar cells. Eur J Morphol 36, Suppl: 181–185, 1998.
35. Lopez-Vidriero MT, Costello J, Clark TJH, Das I, Keal EE, and ReidL. Effect of atropine on sputum production. Thorax 30: 543–547, 1975.
36. Lundberg A. The electrophysiology of the submaxillary gland of the cat.Acta Physiol Scand 35: 1–25, 1955.
37. Martin SC and Shuttleworth TJ. Ca2� influx drives agonist-activated[Ca2�]i oscillations in an exocrine cell. FEBS Lett 352: 32–36, 1994.
38. Matozaki T, Goke B, Tsunoda Y, Rodriguez M, Martinez J, andWilliams JA. Two functionally distinct cholecystokinin receptors showdifferent modes of action on Ca2� mobilization and phospholipid hydro-
lysis in isolated rat pancreatic acini. Studies using a new cholecystokininanalog, JMV-180. J Biol Chem 265: 6247–6254, 1990.
39. McCann JD, Matsuda J, Garcia M, Kaczorowski G, and Welsh MJ.Basolateral K� channels in airway epithelia. I. Regulation by Ca2� andblock by charybdotoxin. Am J Physiol Lung Cell Mol Physiol 258:L334–L342, 1990.
40. Mertz LM, Baum BJ, and Ambudkar IS. Membrane potential modu-lates divalent cation entry in rat parotid acini. J Membr Biol 126: 183–193,1992.
41. Meszaros LG, Bak J, and Chen A. Cyclic ADP-ribose as an endogenousregulator of the non-skeletal type ryanodine receptor Ca2� channel.Nature 364: 76–79, 1993.
42. Moon S, Singh M, Krouse ME, and Wine JJ. Calcium-stimulated Cl�
secretion in Calu-3 human airway cells requires CFTR. Am J Physiol LungCell Mol Physiol 273: L1208–L1219, 1997.
43. Nadel JA and Davis B. Parasympathetic and sympathetic regulation ofsecretion from submucosal glands in airways. Fed Proc 39: 3075–3079,1980.
44. Nagaki M, Shimura S, Irokawa T, Sasaki T, and Shirato K. Nitricoxide regulation of glycoconjugate secretion from feline and humanairways in vitro. Respir Physiol 102: 89–95, 1995.
45. Noguchi N, Takasawa S, Nata K, Tohgo A, Kato I, Ikehata F,Yonekura H, and Okamoto H. Cyclic ADP-ribose binds to FK506-binding protein 12.6 to release Ca2� from islet microsomes. J Biol Chem272: 3133–3136, 1997.
46. Okamoto H, Takasawa S, Nata K, and Yonekura H. Cyclic ADP-ribose, a novel second messenger for intracellular Ca2� mobilization inpancreatic islets. In: 1st IUBMB Conference: Biochemistry of Diseases.Nagoya, Japan. 1992, Abstract.
47. Petersen OH. Stimulus-secretion coupling: cytoplasmic calcium signalsand the control of ion channels in exocrine acinar cells. J Physiol 448:1–51, 1992.
48. Quinton PM. Composition and control of secretions from tracheal bron-chial submucosal glands. Nature 279: 551–552, 1979.
49. Ramnarine SI, Haddad EB, Khawaja AM, Mak JCW, and Rogers DF.On muscarinic control of neurogenic mucus secretion in ferret trachea.J Physiol 494: 577–586, 1996.
50. Sasaki T and Gallacher DV. The ATP-induced inward current in mouselacrimal acinar cells is potentiated by isoproterenol and GTP. J Physiol447: 103–118, 1992.
51. Sasaki T, Shimura S, Wakui M, Ohkawara Y, Takishima T, andMikoshiba K. Apically localized IP3 receptors control chloride current inairway gland acinar cells. Am J Physiol Lung Cell Mol Physiol 267:L152–L158, 1994.
52. Takasawa S, Akiyama T, Nata K, Kuroki M, Tohgo A, Noguchi N,Kobayashi S, Kato I, Katada T, and Okamoto H. Cyclic ADP-riboseand inositol 1,4,5-trisphosphate as alternate second messenger for intra-cellular Ca2� mobilization in normal and diabetic �-cells. J Biol Chem273: 2497–2500, 1998.
53. Takasawa S, Nata K, Yonekura H, and Okamoto H. Cyclic ADP-ribosein insulin secretion from pancreatic � cells. Science 259: 370–373, 1993.
54. Takasawa S, Tohgo A, Noguchi N, Koguma T, Nata K, Sugimoto T,Yonekura H, and Okamoto H. Synthesis and hydrolysis of cyclicADP-ribose by human leukocyte antigen CD38 and inhibition of thehydrolysis by ATP. J Biol Chem 268: 26052–26054, 1993.
55. Tamada T, Sasaki T, Saitoh H, Ohkawara Y, Irokawa T, Sasamori K,Oshiro T, Tamura G, Shimura S, and Shirato K. A novel function ofthyrotropin as a potentiator of electrolyte secretion from tracheal gland.Am J Respir Cell Mol Biol 22: 566–573, 2000.
56. Thorn P, Gerasimenko O, and Petersen OH. Cyclic ADP-ribose regu-lation of ryanodine receptors involved in agonist evoked cytosolic Ca2�
oscillation in pancreatic acinar cells. EMBO J 13: 2038–2043, 1994.57. Thorn P, Lawrie AM, Smith PM, Gallacher DV, and Petersen OH.
Local and global cytosolic Ca2� oscillations in exocrine cells evoked byagonists and inositol trisphosphate. Cell 74: 661–668, 1993.
58. Tom-Moy M, Basbaum CB, and Nadel JA. Localization and release oflysozyme from ferret trachea: effects of adrenergic and cholinergic drugs.Cell Tissue Res 228: 549–562, 1983.
59. Ueki I, German VF, and Nadel JA. Micropipette measurement of airwaysubmucosal gland secretion. Autonomic effects. Am Rev Respir Dis 121:351–357, 1980.
60. United States Department of Health and Human Services. Guide forthe care and use of laboratory animals. In: National Institutes of Health
L77IP3 AND CADPR MIMICS THE ACH-INDUCED RESPONSES IN SUBMUCOSAL GLANDS
AJP-Lung Cell Mol Physiol • VOL 287 • JULY 2004 • www.ajplung.org

Publication No. 86–23. Washington, DC: U.S. Government PrintingOffice, 1985.
61. Wakui M, Osipchuck YV, and Petersen OH. Receptor-activated cyto-plasmic Ca2� spiking mediated by inositol trisphosphate is due to Ca2�-induced Ca2� release. Cell 63: 1025–1032, 1990.
62. Welsh MJ. Electrolyte transport by airway epithelia. Physiol Rev 67:1143–1184, 1987.
63. Widdicombe JH, Bastacky SJ, Wu DX, and Lee CY. Regulation ofdepth and composition of airway surface liquid. Eur Respir J 10: 2892–2897, 1997.
64. Wine JJ. The genesis of cystic fibrosis lung disease. J Clin Invest 103:309–312, 1999.
65. Yamaya M, Finkbeiner WE, and Widdicombe JH. Ion transport bycultures of human tracheobronchial submucosal glands. Am J PhysiolLung Cell Mol Physiol 261: L485–L490, 1991.
66. Yang CM, Farley JM, and Dwyer TM. Acetylcholine-stimulated chlo-ride flux in tracheal submucosal gland cells. J Appl Physiol 65: 1890–1894, 1988.
67. Yang CM, Farley JM, and Dwyer TM. Muscarinic stimulation ofsubmucosal glands in swine trachea. J Appl Physiol 64: 200–209, 1988.
L78 IP3 AND CADPR MIMICS THE ACH-INDUCED RESPONSES IN SUBMUCOSAL GLANDS
AJP-Lung Cell Mol Physiol • VOL 287 • JULY 2004 • www.ajplung.org





























