Embed Size (px)
Citation preview

C
PCa
Cb
c
d
e
a
ARRA
KCHMT
1
afTLlmOCT
i
K
1d
Journal of Clinical Virology 45 (2009) 16–22
Contents lists available at ScienceDirect
Journal of Clinical Virology
journa l homepage: www.e lsev ier .com/ locate / j cv
oxsackievirus B4 in Southern Taiwan: Molecular Epidemiology
ei-Yu Chua,∗, Yu-Ling Tsaib, Hsiu-Lin Chenc, Guan-Ming Ked, Chien-Ying Hsua, Yi-Ting Chena,hu-Feng Wangd, Hui-Ju Sud, Li-Chiu Choud, Li-Ching Hsue, Kuei-Hsiang Lind,∗∗
Department of Medical Laboratory Science and Biotechnology, College of Health Science, Kaohsiung Medical University,S604, 100 Shih-Chuan 1st Rd, Kaohsiung 80705, TaiwanDepartment of Food Science and Nutrition, Hungkuang University, Taichung, TaiwanDepartment of Pediatrics, Neonatology, Kaohsiung Medical University Hospital, Kaohsiung, TaiwanDepartment of Clinical Laboratory, Kaohsiung Medical University, Kaohsiung, TaiwanDivision of Viral Diseases, Center for Disease Control, Department of Health, Taiwan
r t i c l e i n f o
rticle history:eceived 10 November 2008eceived in revised form 18 February 2009ccepted 25 February 2009
eywords:oxsackievirus B4uman enterovirusolecular epidemiology
aiwan
a b s t r a c t
Background: Enterovirus outbreaks caused by Coxsackievirus B4 (CB4) in Taiwan in 2004 and 2008.Objective: To retrospectively analyze the molecular epidemiology and pathogenicity of CB4 in Taiwan.Study design: This study analyzed twenty-three CB4 strains isolated in Taiwan during 1993–2004.Sequence variations data were obtained using 420 bp of VP4/VP2 region and 331 bp of 3′ VP1 region.Phylogenetic dendrograms were constructed with other CB4 sequences in Genebank. The clinical man-ifestations of CB4 infection were examined by retrospectively reviewing medical records of infectedpatients.Results: Three CB4 genotypes were identified: genotypes II, IVb and VIII. Genotype VIII, a new and geo-graphically distinct cluster, has been isolated in South Korea, China and Taiwan. This genotype was isolatedin twelve of twenty-three CB4 patients treated in Taiwan during 1997–2004. Eight of twenty-three strainsbelonging to genotype II, now the major genotype worldwide, were first identified in Taiwan in 2000.
Three isolates (identified 1993–1994) analyzed in this study belonged to genotype IVb. In this retrospec-tive follow-up study of sixteen patients with CB4 infection, the median patient age at the time of infectiondiagnosis was 4-year-old (range, 18 days to 10-year-old), and male–female ratio was 1:1. None of the six-teen patients suffered IDDM or myocarditis after their B4 infection episodes; four had Attention DeficitHyperactivity Disorder (ADHD) and/or tic disorders (TDs) at follow-up.and Vible a
Conclusions: Genotypes IIneeded to evaluate a poss
. Background
Human enteroviruses (HEVs) are common clinical pathogensssociated with a wide spectrum of health consequences rangingrom respiratory disease in mild cases to death in extreme cases.he HEVs also reportedly vary according to their geographic origin.arge-scale outbreaks may be associated with changes in circu-
ating HEV serotypes. Epidemiological surveillance is crucial foronitoring rapidly changing patterns of enterovirus prevalence.1
utbreaks of CB4 infections are rare, and most diseases caused byB4 are either asymptomatic or mild and self-limiting. However,he CB4 viruses are known to cause neonatal infection, myocardi-
Abbreviations: ADHD, Attention Deficit Hyperactivity Disorder; CB4, Coxsack-evirus B4; HEV, human enteroviruses; TD, tic disorder.∗ Corresponding author. Tel.: +886 7 3121101x2351; fax: +886 7 3113449.
∗∗ Corresponding author. Tel.: +886 7 3121101x7250; fax: +886 7 3114449.E-mail addresses: [email protected] (P.-Y. Chu),
[email protected] (K.-H. Lin).
386-6532/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.jcv.2009.02.012
III of CB4 have co-circulated in Taiwan since 2000. Controlled studies aressociation between ADHD and TDs with CB4 infection.
© 2009 Elsevier B.V. All rights reserved.
tis, encephalitis and IDDM.2–7 Further, CB4 reportedly has a higherfatality rate than other HEV infections.1,8 The affected populationis geographically diverse; cases have been reported frequently inFinland,9 South Korea,10 Cyprus,11 western Germany,12 Spain,13 andFrance.14 However, CB4 was not the top five prevalent HEV in thesecountries except France.14 According to the CDC-TW SurveillanceSystem installed since 1999, CB4 was one of the five most prevalententeroviruses in 2000, 2001, 2004 and 2008. CB4 is found almostevery year in Taiwan, sporadic cases can be found throughout theyear. In both 2004 and 2008 outbreaks, CB4 cases steadily increasedduring spring-to-summer and peaked at June. Although CB4 activ-ity was moderate in 2000, the reported CB4 cases were associatedwith myopericarditis.4 A higher proportion of patients were asso-ciated with myopericarditis (33.6%) in Taiwan during 2000–2005.
However, no clear clinical manifestation was found in the 2008outbreak.Molecular and epidemiologic studies of CB4 isolates haveidentified at least seven genotypes (genotypes I–VII) circulatingworldwide since 1951.15 However, the molecular epidemiology of

P.-Y. Chu et al. / Journal of Clinical Virology 45 (2009) 16–22 17
Table 1Data for twenty-three clinical isolates.
Designation Genotype VP4a VP1a Specimen Gender Ageb Clinical diagnosis Note
1684/93/TW IVb AB456020 AB456001 TS M Lost to follow-up1716/93/TW IVb AB456021 AB455999 RS M 2 months Acute gastroenteritis Admission0016/94/TW IVb AB456022 AB456000 TS Lost to follow-up0260/97/TW VIII AB456033 AB456003 CSF F 18 days Meningoencephalitis Admission; abnormal EEG
indicating epileptogenicity for 2years
0295/97/TW VIII AB456034 AB456002 TS F Lost to follow-up4200/00/TW VIII AB456025 AB456009 TS F 6 years Herpangina OPD4338/00/TW VIII AB456031 AB456008 TS F 7 years URI Admission4463/00/TW II AB456015 AB455993 TS F 1 year 7 months Acute tonsillitis Admission1034/01/TW VIII AB456030 AB456006 TS F 3 years 8 months Acute pharyngitis OPD1217/01/TW II AB456018 AB455998 TS M Lost to follow-up2277/01/TW II AB456019 AB455997 TS F 1 year 2 months Herpangina OPD2570/01/TW VIII AB456032 AB456011 TS M 4 years 4 months Herpangina OPD; lost to follow-up2712/01/TW VIII AB456026 AB456007 TS F 4 years HFMD; pneumonia Admission; lost to follow-up2830/01/TW II AB456028 ND CSF F 5 years Acute bronchitis Admission3055/01/TW II AB456017 AB455995 TS M 1 year 3 months Herpangina Admission3315/01/TW II AB456016 AB455996 TS F 3 years 4 months Acute pharyngitis Admission4512/02/TW II AB456012 AB455992 TS F 7 years Acute pharyngitis Lost to follow-up5141/02/TW II AB456013 AB455991 TS M 10 months Herpangina Admission1717/04/TW VIII AB456023 AB456010 TS M Lost to follow-up4107/00/TW II AB456014 AB455994 TS M 5 years 4 months Acute tonsillitis OPD; ADHD diagnosed 5 years later2235/01/TW VIII AB456027 AB456005 TS M 6 years Herpangina OPD; TD diagnosed 1 year later2823/01/TW VIII AB456029 AB456004 TS M 5 years Herpangina OPD; ADHD and TD diagnosed 1
year later2522/04/TW VIII AB456024 ND TS M 8 years Severe urticaria and URI Admission; ADHD diagnosed 1 year
later; then TD also noted 2 years later
T ment;T
Caihc
2
rbCef
3
3
cscesbws
3r
to
S = throat swab; RS = rectal swab; CSF = cerebrospinal fluid; OPD = outpatient departD = tic disorder; HFMD = hand–foot–mouth disease; ND = not determined.a Accession number in genebank.b Age at isolation of virus.
B4 in Taiwan has not been reported. Amplification and subsequentnalysis of the 3′ VP1 gene fragment have proven reliable for analyz-ng HEV type and molecular epidemiology.13,16 The VP4/VP2 regionsave been amplified by a single primer set and are also useful forlassifying enteroviruses.15,17
. Objective
To investigate the molecular epidemiology of CB4 in Taiwan, thisetrospective study characterized genotype distribution patternsased on sequence variations in the VP4/VP2 and 3′VP1 regions ofB4 isolates. The clinical manifestations of CB4 infection were alsoxamined by retrospectively reviewing medical charts, particularlyor subsequent occurrences of cardiomyopathy and IDDM.
. Study design
.1. Specimens and viruses
Twenty-three CB4 strains were randomly selected from clini-al specimens isolated in Kaohsiung University Hospital (KMUH) inouthern Taiwan during 1993–2004. Changes in KMUH policy pre-luded further serotyping for non-polio enteroviruses other thannterovirus 71 and coxsackievirus A16 since 2004. Therefore, thetudy did not analyze isolates thereafter. All strains were confirmedy immunofluorescence assay. Twenty of the twenty-three strainsere isolated from throat swabs, one was isolated from a rectal
wab, and two were isolated from cerebrospinal fluid (Table 1).
.2. RNA isolation, reverse transcription-polymerase chain
eaction (RT-PCR) and sequencingViral RNA was extracted according to manufacturer instruc-ions (QIAamp viral RNA purification kit, Qiagen Co.). The 330 bpf 3′VP1 region was amplified using the primer sets 08840 and
URI = upper respiratory infection; ADHD = Attention Deficit Hyperactivity Disorder;
71851. A second primer set, 040 and 011, was used if product yieldwas insufficient.15,18 For simplicity, the uniform RT-PCR conditionwas performed for 3′VP1 as reported previously.15 The 420 bp ofVP4/VP2 gene was amplified by a primer set of EVP2 and OL68-1,19
and the RT-PCR was performed as described previously.20
The RT-PCR products were purified with commercial reagents(Qiagen Gmbh, HIlden, Germany). Cycle sequencing reactions wereperformed using the (Big) Dye terminator kit (PE Applied Biosys-tems). The sequences for forward and reverse strands were obtainedsimultaneously and then edited using Sequence Navigator (ver-sion 1; PE Applied Biosystems). The sequences were compared toGenebank sequences by BLAST. The deduced amino acid sequenceswere also compared.
3.3. Phylogenetic analysis
The nucleotide sequence data for the prototype and Genebankstrains were used for phylogenetic analysis. Phylogenetic anal-ysis was performed with the 330 bp fragments of the 3′ VP1region for the strains identified in this study. The analysis includedthe CB4 prototype X5690/JVB/51/USA (JVB), a diabetogenic strainS76772/E2/58/USA (E2)21 and sixty-four others strains includ-ing one from China,22 forty-four from Europe10,14,22–25 six fromUSA,15,23 five from South Korea10 and nine previously unreportedstrains (eight from France and one from South Korea). The phy-logenetic analysis was similarly performed using the VP4/VP2regions for the Taiwan strains identified in this study, JVB, E2and forty previously reported strains.15 The ClustalW program wasused to perform multiple sequence alignments. The MEGA Ver-sion 4 program was used for pairwise comparison, phylogenetic
tree construction and bootstrap.25 Genetic distance was calculatedusing maximum likelihood model of nucleotide substitution. Thephylogenetic tree was constructed by UPGMA method, and the sig-nificance of phylogenetic analysis was estimated by bootstrap with1000 pseudoreplicates.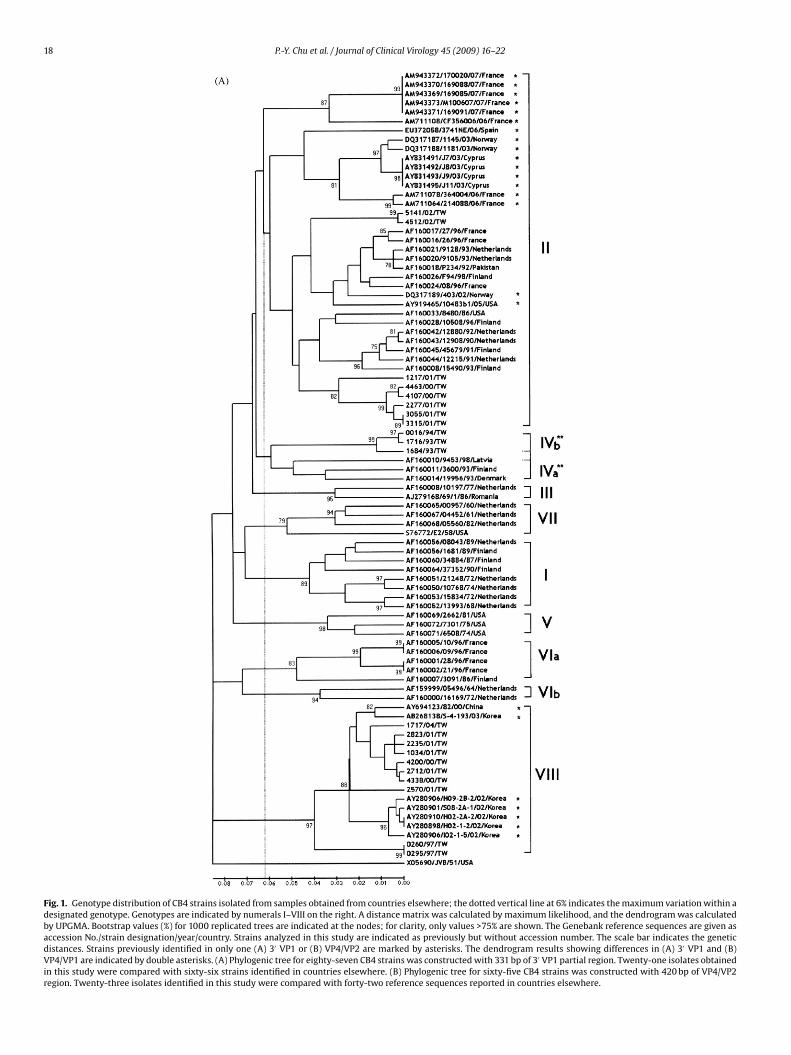
18 P.-Y. Chu et al. / Journal of Clinical Virology 45 (2009) 16–22
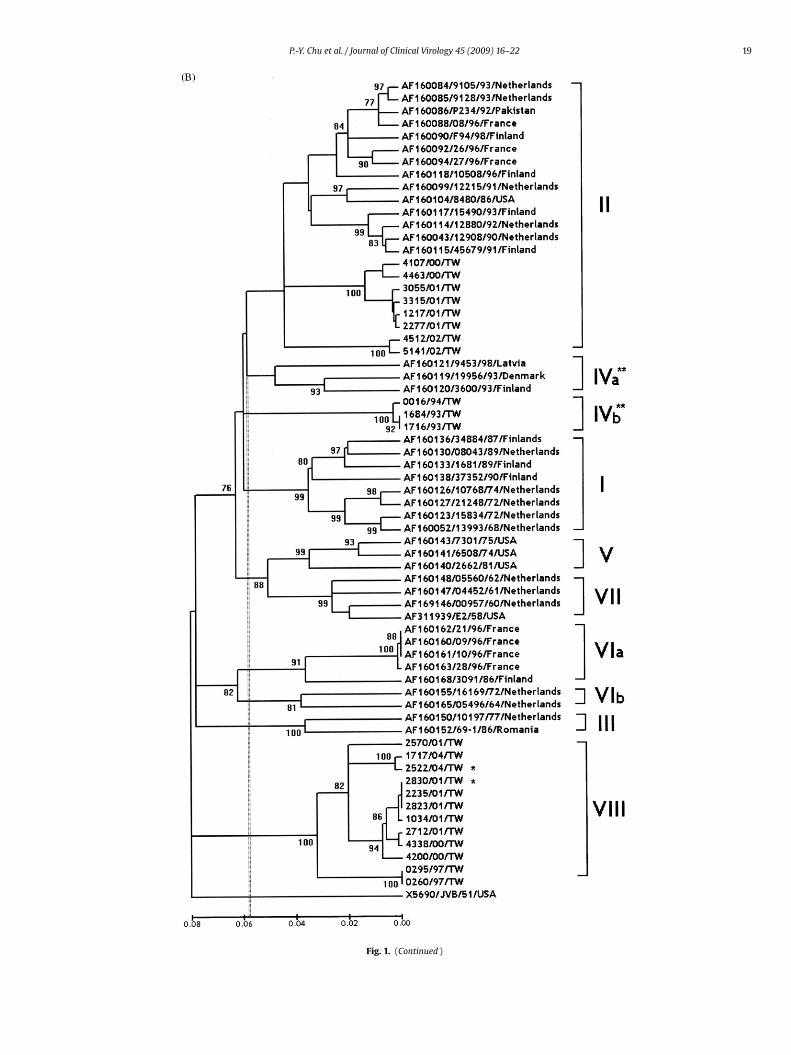
Fig. 1. Genotype distribution of CB4 strains isolated from samples obtained from countries elsewhere; the dotted vertical line at 6% indicates the maximum variation within adesignated genotype. Genotypes are indicated by numerals I–VIII on the right. A distance matrix was calculated by maximum likelihood, and the dendrogram was calculatedby UPGMA. Bootstrap values (%) for 1000 replicated trees are indicated at the nodes; for clarity, only values >75% are shown. The Genebank reference sequences are given asaccession No./strain designation/year/country. Strains analyzed in this study are indicated as previously but without accession number. The scale bar indicates the geneticdistances. Strains previously identified in only one (A) 3′ VP1 or (B) VP4/VP2 are marked by asterisks. The dendrogram results showing differences in (A) 3′ VP1 and (B)VP4/VP1 are indicated by double asterisks. (A) Phylogenic tree for eighty-seven CB4 strains was constructed with 331 bp of 3′ VP1 partial region. Twenty-one isolates obtainedin this study were compared with sixty-six strains identified in countries elsewhere. (B) Phylogenic tree for sixty-five CB4 strains was constructed with 420 bp of VP4/VP2region. Twenty-three isolates identified in this study were compared with forty-two reference sequences reported in countries elsewhere.
P.-Y. Chu et al. / Journal of Clinical Virology 45 (2009) 16–22 19
Fig. 1. (Continued )

2 linica
3
GAat
3
ppviiBpuawfcfi
4
4
aPTpb(
sgfcttibVtSsgtwTra
4
cCbt
i
0 P.-Y. Chu et al. / Journal of C
.4. Accession numbers of nucleotide sequences
The sequences described in this study were deposited in theenebank nucleotide sequence database with accession numbersB455912–AB456034 for VP4/VP2 region (positions 744–1163) andccession numbers AB455991–AB456011 for 3′ VP1 region (posi-ions 2880–3210) (Table 1).
.5. Clinical records review
Specimen collection for viral isolation included pediatricatients treated at KMUH during 1993–2004 for clinically sus-ected viral respiratory infection. In patients with positive CB4irus isolates, sixteen of twenty-three clinical indications, includ-ng time of infection and time of outpatient medical follow-up afternfection, were retrospectively reviewed by a single pediatrician.ecause the other seven throat swab isolates were obtained fromatients treated by a local physician rather than by this medicalniversity hospital, medical records for these patients were unavail-ble for review. All KMUH patients were followed up at least onceithin the week after discharge. Thereafter, follow-up was per-
ormed irregularly. All symptoms and signs at the time of specimenollection were recorded. Medical records of all patients were care-ully reviewed for events occurring after the episode of enteroviralnfection.
. Results
.1. Sequence clustering in the 3′ VP1 region
For 3′ VP1 region, primer sets 08840 and 71851 were used tomplify a 330 bp product in twenty of the twenty-three isolates.rimer sets 011 and 040 were used to amplify one additional isolate.wo of the twenty-three isolates were not amplified by any of therimer sets. The differences in nucleotide and amino acid sequencesetween the Taiwan strains and the prototype, X05690/JVB/51/USAJVB),24 were 11.6–20.2% and 0.9–2.9%, respectively.
The phylogenic tree for eighty-seven CB4 strains was con-tructed with 331 bp fragment of 3′ VP1 region. The extrapolatedenotype demarcation was 6.2%. The results showed that, exceptor JVB, which constituted a unique branch, all CB4 isolates werelustered into eight distinct genotypes (Fig. 1A) designated geno-ypes I–VIII. Of these, genotype VIII is reported here for the firstime. This study identified CB4 strains of genotypes II, IVb and VIIIn Taiwan. The new lineage, genotype VIII, was supported by a highootstrap value of 97%. The nucleotide distances within genotypeIII were less than 9.0% (data not shown). All strains in this geno-
ype were isolated in Asia: ten, six and one were isolated in Taiwan,outh Korea and China, respectively. Ten of the twenty-one Taiwantrains were isolated during 1997–2004. Genotype II, the largestenotype, included strains reported elsewhere in the world such ashose recently identified in France in 2007. Eight of twenty-one Tai-an strains classified as genotype II were isolated in 2000–2002.
he three Taiwan strains, which were isolated in 1993 and 1994, areelated to genotype IV, which includes strains isolated in Denmarknd Finland in 1993.
.2. Sequence clustering in the VP4/VP2 region
In the VP4/VP2 region, the EVP2 and OL68-1 primer sets suc-essfully amplified 420nt product from all twenty-three strains of
B4 isolates. The variations in nucleotide and amino acid sequencesetween Taiwan and JVB were 12.2–22.5% and 0.7–4.6%, respec-ively.The phylogenetic analysis of VP4/VP2 in sixty-five CB4 strainsncluded twenty-three Taiwan strains, JVB, E2 and forty previ-
l Virology 45 (2009) 16–22
ously reported strains.15 The extrapolated genotype demarcationof VP4/VP2 was 5.8%. Clustering patterns were similar to thoseobtained by VP1 sequence alignment (Fig. 1B). As in the 3′ VP1 tree,the newly identified genotype VIII was supported by a 100% boot-strap value. In contrast with the 3′ VP1 tree, genotype IVb formedan entirely separate cluster in the VP4/VP2 tree and had a bootstrapvalue of 100%. In this tree, the average distance between geno-types IVa and II was smaller than that between genotypes IVa andIVb (12.1% vs. 12.4%, respectively). In both analyzed regions, thenucleotide differences were almost tenfold that of the amino aciddifferences, which indicated that most nucleotide variations weresynonymous mutations in the 3′ VP1 and VP4/VP2 regions. Com-parison of the 140 deduced amino acid sequence in the VP4/VP2region among sixty-four strains identified worldwide with proto-type JVB strain revealed a distinctive neutral mutation at residue138 (V → G) in all three of genotype IVb strains. A common neutralmutation was found at residue 20 (1 S → P, 2 S → A and 9 S → T)in all twelve strains genotype VIII strains in Taiwan. Interestingly, amissense mutation was revealed at residue 16 (2 S → N and 6 S → G)in all eight strains of genotype II strains. The variance in genotypeII strains is 100% in Taiwan but is lower (four/fourteen, 33.3%) incountries elsewhere.
4.3. Clinical manifestations and patient follow-up
Of the remaining sixteen subjects treated and followed up atKMUH, the median age at time of CB4 infection was 4 years old(range, 18 days–10 years), and the male–female ratio was 1:1.Interestingly, a review of follow-up records revealed no IDDM ormyocarditis, although CB4 is reported to associate with these twodiseases. However, neuropsychological disorders were noted in fiveof the sixteen CB4 infection cases. One 18-day-old newborn withCNS infection had abnormal electroencephalogram (EEG) consis-tent with epileptogenicity for 2 years in subsequent follow-ups. Noapparent predisposing factor or other clinical symptoms or signs,other than this CB4 infection, was remarkable in this patient. Theother four boys who were older than 5 years at the time of CB4infection had been diagnosed with Attention Deficit HyperactivityDisorder (ADHD) and/or tic disorder (TD) within 1–5 years after CB4infection (Table 1). Clinical diagnosis of ADHD and/or TD had beenmade by clinical observation and physical examinations in thesefour patients when they were taken back to our clinic for suspect-ing symptoms including inattention, hyperactivity, and impulsivity.The risk factors of prenatal toxin exposure such as maternal alcoholor tobacco use as well as postnatal lead exposure and family historyof ADHD/TD in these four patients were ruled out by the review ofpatient medical history from parents and medical chart. Three ofthem also had received behavior rating scales evaluation. One 6-year-old boy had EEG and nerve conduction velocity examinationto rule out other neurological disorders.
5. Conclusions
To date, at least seven distinct genotypes are known to have cir-culated in Europe during 1959–1998. Current data indicate that CB4outbreaks are intermittent and are followed by interruption of viruscirculation until the population of susceptible patients increasesor until a new virus variant is introduced. Genotype IV was onlyrecently identified in Taiwan in 1993–1994. The 2000 and 2004outbreaks involved both genotypes II and VIII. Genotype VIII has
been the dominant Asian strain since 1997 while genotype II strainshave spread worldwide since 1986. Moreover, ten new genotypeVIII strains were identified in Taiwan in 2004, one new genotypeII strains was identified in 2006, thereafter two new genotype IIstrains and eight new genotype VIII strains were identified in 2008
linica
(i1rficairvcrmcosttbmctE
onG5∼ecmsaCfv2brtEqoIdiata
C
A
dKp
aa
P.-Y. Chu et al. / Journal of C
data from CDC-TW). Three novel findings were deduced here: first,n previous report,15 genotypes I, II and VI were predominant before998, whereas genotype II has been predominant since 2000 in cur-ent study. Second, genotype VIII, which is reported here for therst time, has been present in Asia since 1997. Third, in a high per-entage (six/eight, 75%) of genotype II strains in Taiwan, the 16thmino residue in VP4 changed from S to G. The 16th residue of VP4s reportedly associated with CB4 diabetogenic virulence: the Seresidue at the 16th position mutates into Arg in the non-virulentariant.26 CB4 infection can cause death of � cells by virus-inducedytolysis and thus lead to diabetes mellitus.7 VP4 plays a criticalole in viral maturation and cell entry,27,28 therefore a missenseutation in the 16th amino acid residue of VP4 may affect its repli-
ation cycles in the � cell. On the other hand, the reported incidencef IDDM is low in Asian populations.29 It is well known that thetrength of affinity between TCR and MHC/epitope complex dictateshe outcome of the immune response.30,31 A missense mutation inhe VP4 may affect peptide loading onto the MHC grove and/or theinding affinity/avidity of TCR and MHC/peptide complex. Further-ore, the recent works has demonstrated that the sequence of VP4
orresponding to amino acids 11–30, or a part of it plays a role inhe plasma-dependent enhancement of CB4 diabetogenic strains2 induced IFN-� synthesis by PBMC.32,33
In this clinical retrospective study, we found four boys who werelder than 5 years at the time of CB4 infection had been diag-osed with ADHD and/or TD within 1–5 years after CB4 infection.lobally, ADHD is a highly prevalent psychiatric condition affecting–7% of school-age children.34 Although genetic factors account for80% of ADHD etiology, the growing understanding of ADHD nownables diagnosis by symptom clusters used in various immunehallenges. Biological adversity, such as trauma, metabolic derange-ent, toxin exposure, and CNS infection, may also cause ADHD in
ome children.34,35 There were few studies on reporting the associ-tion of enteroviral infection and the development of ADHD. EV71NS infection had been suggested as a trigger and perpetuating
actor in ADHD in a follow-up study of 142 children who had sur-ived EV71 CNS infections during enteroviral outbreak in Taiwan in007.36 The TDs are neurodevelopmental disorders characterizedy childhood onset and the presence of tics such as sudden, rapid,epetitive and non-rhythmic movements or vocalizations. Symp-oms of ADHD are usually observed 2–3 years before onset of TD.37
pidemiological data shows that ADHD and TD co-occur more fre-uently than expected by chance. The co-morbidity of TD and ADHDutcome remains a subject of great scientific and clinical interest.38
n this retrospective study, four boys older than 5 years old wereiagnosed with ADHD and/or TD after episodes of CB4 enteroviral
nfection at 5 or more years of age. All four boys were only mildlyffected by their acute infection and most did not require hospi-alization. A possible association between CB4 infection and ADHDnd TD requires further controlled study.
onflict of interest
None declared.
cknowledgements
We gratefully acknowledge the Taiwan CDC for providingata regarding enterovirus prevalence. The Virology Laboratory at
aohsiung Medical University Hospital, Taiwan is appreciated forroviding the enterovirus isolates and valuable technical assistance.Funding: Kaohsiung Medical University grants KMU-Q095014,nd partly supported by the grants from Center for Disease Controlnd Prevention of Taiwan (CDC94-RM-010).
Ethical approval: Not required.
l Virology 45 (2009) 16–22 21
References
1. Khetsuriani N, Ashley L-F, Oberst MS, Pallansch MA. Enterovirus surveillance—United States, 1970–2005. MMWR Surveill Summ 2006;15:1–20.
2. Verboon-Maciolek M, Krediet T, van Loon A, Kaan J, Galama J, Gerards L, etal. Epidemiological survey of neonatal non-polio enterovirus infection in theNetherlands. J Med Virol 2002;66:241–5.
3. Cree B, Bernardini G, Hays A, Lowe G. A fatal case of coxsackievirus B4 menin-goencephalitis. Arch Neurol 2003;60:107–12.
4. Tseng F, Huang H, Chi C, Lin T, Liu C, Jian J, et al. Epidemiological survey ofenterovirus infections occurring in Taiwan between 2000 and 2005. J Med Virol2007;79:1850–60.
5. Baboonian C, McKenna W. Eradication of viral myocarditis: is there hope? J AmColl Cardiol 2003;42:473–6.
6. Kaski J, Burch M. Viral myocarditis in childhood. J Paediatr Child Health2007;17:11–8.
7. Dotta F, Censini S, van Halteren AG, Marselli L, Masini M, Dionisi S, et al.Coxsackie B4 virus infection of beta cells and natural killer cell insulitis inrecent-onset type 1 diabetic patients. Proc Natl Acad Sci USA 2007;104:5115–20.
8. Khetsuriani N, Lamonte A, Oberste M, Pallansch M. Neonatal enterovirus infec-tions reported to the national enterovirus surveillance system in the UnitedStates, 1983–2003. Pediatr Infect Dis J 2006;25:889–93.
9. Hovi T, Stenvik M, Rosenlew M. Relative abundance of enterovirus serotypes insewage differs from that in patients: clinical and epidemiological implications.Epidemiol Infect 1996;116:91–7.
10. Lee C, Lee SH, Han E, Kim SJ. Use of cell culture-PCR assay based on combinationof A549 and BGMK cell lines and molecular identification as a tool to monitorinfectious adenoviruses and enteroviruses in river water. Appl Environ Microbiol2004;70:6695–705.
11. Bolanaki E, Kottaridi C, Markoulatos P, Margaritis L, Katsorchis T. Evolution of2B and 2C genomic parts of species B Coxsackie viruses, phylogenetic study andcomparison with other regions. Virus Genes 2006;32:249–59.
12. Roth B, Enders M, Arents A, Pfitzner A, Terletskaia-Ladwig E. Epidemiologicaspects and laboratory features of enterovirus infections in Western Germany,2000–2005. J Med Virol 2007;79:956–62.
13. Cabrerizo M, Echevarria J, González I, de Miguel T, Trallero G. Molecularepidemiological study of HEV-B enteroviruses involved in the increase inmeningitis cases occurred in Spain during 2006. J Med Virol 2008;80:1018–24.
14. Antona D, Lévêque N, Chomel J, Dubrou SDL-B, Lina B. Surveillanceof enteroviruses in France, 2000–2004. Eur J Clin Microbiol Infect Dis2007;26:403–12.
15. Mulders MN, Salminen M, Kalkkinen N, Hovi T. Molecular epidemiology of cox-sackievirus B4 and disclosure of the correct VP1/2A(pro) cleavage site: evidencefor high genomic diversity and long-term endemicity of distinct genotypes. JGen Virol 2000;81:803–12.
16. Cabrerizo M, Echevarria J, Otero A, Lucas P, Trallero G. Molecular charac-terization of a coxsackievirus A24 variant that caused an outbreak of acutehaemorrhagic conjunctivitis in Spain, 2004. J Clin Virol 2008;43:323–7.
17. Kubo H, Iritani N, Seto Y. Molecular classification of enteroviruses not identifiedby neutralization tests. Emerg Infect Dis 2002;8:298–304.
18. Oberste MS, Maher K, Flemister MR, Marchetti G, Kilpatrick DR, Pallansch MA.Comparison of classic and molecular approaches for the identification of unty-pable enteroviruses. J Clin Microbiol 2000;38:1170–4.
19. Kitamura A, Narisawa T, Hayashi A, Ashihara Y, Ishiko H. Serotype determi-nation of enteroviruses that cause hand–foot–mouth disease: identificationof enterovirus 71 and Coxsackie virus A 16 from clinical specimens by usingspecific probe. J Jpn Assoc Infect Dis 1997;71:715–23.
20. Chu PY, Lin KH, Hwang KP, Chou LC, Wang CF, Shih SR, et al. Molecular epidemi-ology of enterovirus 71 in Taiwan. Arch Virol 2001;146:589–600.
21. Hartig PC, Madge GE, Webb SR. Diversity within a human isolate of coxsackieB4: relationship to viral-induced diabetes. J Med Virol 1983;11:23–30.
22. Tian B, Hiromu Y, Wu Y, Lu L, Takao T, Hiroyuki S, et al. Molecular typing andepidemiology of non-polio enteroviruses isolated from Yunnan Province, thePeople’s Republic of China. J Med Virol 1998;80:670–9.
23. Oberste M, Maher K, Williams A, Dybdahl-Sissoko N, Brown BA, Gookin M,et al. Species-specific RT-PCR amplification of human enteroviruses: a toolfor rapid species identification of uncharacterize enteroviruses. J Gen Virol2006;87:119–28.
24. Jenkins O, Booth JD, Minor PD, Almond JW. The complete nucleotide sequence ofcoxsackievirus B4 and its comparison to other members of the Picornaviridae.J Gen Virol 1987;68:1835–48.
25. Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary GeneticsAnalysis (MEGA) software version 4.0. Mol Biol Evol 2007;24:1596–9.
26. Ramsingh A, Slack J, Silkworth J, Hixson A. Severity of disease induced by apancreatropic Coxsackie B4 virus correlates with the H-2Kq locus of the majorhistocompatibility complex. Virus Res 1989;14:347–58.
27. Davis MP, Bottley G, Beales LP, Killington RA, Rowlands DJ, Tuthill TJ. Recom-binant VP4 of human rhinovirus induces permeability in model membranes. J
Virol 2008;82:4169–74.28. Hogle JM. Poliovirus cell entry: common structural themes in viral cell entrypathways. Annu Rev Microbiol 2002;56:677–702.
29. Bessaoud K, Boudraa G, de Ropolo M, de Sereday M, Marti M, Moser M, et al. Inci-dence and trends of childhood Type 1 diabetes worldwide 1990–1999. DiabetMed 2006;23:857–66.

2 linica
37. Gillberg C, Gillberg I, Rasmussen P, Kadesjo B, Soderstrom H, Rastam M, et al.
2 P.-Y. Chu et al. / Journal of C
30. Buslepp J, Zhao R, Donnini D, Loftus D, Saad M, Appella E, et al. T cell activitycorrelates with oligomeric peptide-major histocompatibility complex bindingon T cell surface. J Biol Chem 2001;276:47320–8.
31. Bullock T, Mullins D, Colella T, Engelhard V. Manipulation of avidityto improve effectiveness of adoptively transferred CD8(+) T cells formelanoma immunotherapy in human MHC class I-transgenic mice. J Immunol2001;167:5824–31.
32. Sauter P, Chehadeh W, Lobert PE, Lazrek M, Goffard A, Soumillon M, et al. A
part of the VP4 capsid protein exhibited by coxsackievirus B4 E2 is the target ofantibodies contained in plasma from patients with type 1 diabetes. J Med Virol2008;80:866–78.33. Chehadeh W, Lobert PE, Sauter P, Goffard A, Lucas B, Weill J, et al. Viral pro-tein VP4 is a target of human antibodies enhancing coxsackievirus B4- andB3-induced synthesis of alpha interferon. J Virol 2005;79:13882–91.
l Virology 45 (2009) 16–22
34. Biederman J, Faraone S. Attention-deficit-hyperactivity disorder. Lancet2005;366:237–48.
35. Biederman J, Faraone S. Current concepts on the neurobiology of attention-deficit/hyperactivity disorder. J Atten Disord 2002;6(Suppl. 1):S7–16.
36. Gau S, Chang L, Huang L, Fan T, Wu Y, Lin T. Attention-deficit/hyperactivity-related symptoms among children with enterovirus 71 infection of the centralnervous system. Pediatrics 2008;122:e452–8.
Coexisting disorders in ADHD—implications for diagnosis and intervention. EurChild Adolesc Psychiatry 2004;13(Suppl. 1):180–92.
38. Banaschewski T, Neale B, Rothenberger A, Roessner V. Comorbidity of tic dis-orders and ADHD. Conceptual and methodological considerations. Eur ChildAdolesc Psychiatry 2007;16(Suppl. 1):I/5–14.





























