Embed Size (px)
Citation preview

ORIGINAL PAPER
Comparative study on the antibiotic susceptibility and plasmidprofiles of Vibrio alginolyticus strains isolated from four Tunisianmarine biotopes
Rim Lajnef • Mejdi Snoussi • Jesus Lopez Romalde •
Cohen Nozha • Abdennaceur Hassen
Received: 10 April 2012 / Accepted: 6 August 2012
� Springer Science+Business Media B.V. 2012
Abstract The antibiotic resistance patterns and the plas-
mids profiles of the predominant etiological agent respon-
sible for vibriosis in Tunisia, V. alginolyticus were studied
to contribute to control their spread in some Mediterranean
aquaculture farms and seawater. The sixty-nine V. algino-
lyticus strains isolated from different marine Tunisian
biotopes (bathing waters, aquaculture and conchylicole
farms and a river connected to the seawater during the cold
seasons) were multi-drug resistant with high resistance rate
to ampicillin, kanamycin, doxycyclin, erythromycin, imi-
pinem, and nalidixic acid. The multiple resistance index
ranged from 0.3 to 0.7 for the isolates of Khenis, from 0.5
to 0.8 for those of Menzel Jmil, from 0.5 to 0.75 (Hergla)
and from 0.3 to 0.7 for the isolates of Oued Soltane. The
high value of antibiotic resistance index was recorded for
the V. alginolyticus population isolated from the fish farm
in Hergla (ARI = 0.672) followed by the population iso-
lated from the conchylicole station of Menzel Jmil
(ARI = 0.645). The results obtained by the MIC tests
confirmed the resistance of the V. alginolyticus to ampi-
cillin, erythromycin, kanamycin, cefotaxime, streptomycin
and trimethoprim. Plasmids were found in 79.48 % of the
strains analyzed and 30 different plasmid profiles were
observed. The strains had a high difference in the size of
plasmids varying between 0.5 and 45 kb. Our study reveals
that the antibiotic-resistant bacteria are widespread in the
aquaculture and conchylicole farm relatively to others
strains isolated from seawater.
Keywords Vibrio � Disk diffusion test � Antibiotic
resistance � Plasmids � MICs � MBCs � MAR index � ARI
Introduction
Disease outbreaks in marine organisms appear to be esca-
lating worldwide (Harvell et al. 2002) and a growing
number of human bacterial infections have been associated
with recreational and commercial uses of marine resources
(Baffone et al. 2005; Ben Kahla-Nakbi et al. 2007). A
surprising number of Vibrio species have been reported
from marine environments (Gomez-Leon et al. 2005;
Hidalgo et al. 2008; Balcazar et al. 2010), and the proba-
bility of their transmission to humans is correlated with
abiotic factors that affect their distribution, especially the
temperature of seawater during the summer (Croci et al.
2001; Thompson et al. 2004).
In the other hand, Vibrio species have been described as
important fish and shellfish pathogens (Woo and Kelly
1995; Nakayama et al. 2006), as in the case of V. harveyi
in shrimp (Austin and Zhang 2006) and V. alginolyticus in
prawns (Lee et al. 1996) and clams (Gomez-Leon et al.
2005), accounting for the widespread use of antibiotics in
such aquaculture setting (Ferrini et al. 2008).
Vibrio alginolyticus is considered as marine fish and
shellfish pathogen (Gomez-Leon et al. 2005). This bacterium
is a common inhabitant of the marine environment in both
R. Lajnef (&) � M. Snoussi � A. Hassen
Laboratoire de Traitement des Eaux Usees, Centre de
Recherches et des Technologies des Eaux, Technopole de
Borj-Cedria, BP 901, 2050 Hammam-Lif, Tunisia
e-mail: [email protected]
R. Lajnef � J. L. Romalde
Departamento de Microbiologia y Parasitologia,
CIBUS-Facultad de Biologia, Universidad de Santiago,
Santiago de Compostela, 15782 Santiago de Compostela, Spain
C. Nozha
Laboratoire de Microbiologie et d’Hygiene des Aliments et de
l’Environnement, Institut Pasteur, Casablanca, Morocco
123
World J Microbiol Biotechnol
DOI 10.1007/s11274-012-1147-6

temperate and tropical waters (Zanetti et al. 2000) and is
associated with high mortality in aquaculture systems
through the Tunisian seacoasts causing a several economic
losses and high mortality in larvae of many species espe-
cially: Sparus aurata and Dicentrarchus labrax (Bakhrouf
et al. 1995; Snoussi et al. 2006; Ben Kahla-Nakbi et al. 2007).
Vibrio alginolyticus is associated with human infections
related to consumption of raw or undercooked sea products
(fishes and shellfishes) causing severe gastroenteritis and
extra-intestinal diseases (Wounds, intracranial infection in
immunocompromised and cirrhotic patients). These ill-
nesses occur frequently during the summer related to an
increase in the seawater temperature (Croci et al. 2001;
Thompson et al. 2004). This microorganism produces
many extracellular proteases responsible for interaction
between the bacterium and cell hosts (human and animals)
and plays an important role in human infection and fish
pathology (Ottaviani et al. 2001; Thompson et al. 2004).
The mechanism of pathogenicity induced by Vibrio infec-
tions is still complex and related to several factors
including cytotoxins, enterotoxins and lytic enzymes
(Ottaviani et al. 2001).
Antibiotics and other chemotherapeutic agents com-
monly used in fish farms either as feed additives or
immersion baths to achieve either prophylaxis or therapy
may result in an increase of drug-resistant bacteria as well
as R-plasmids (Son et al. 1997; Saitanu et al. 1994). Marine
vibrios have long been recognized as important reservoirs
and vehicles of antibiotic resistance because of their
importance as potential human/or marine animal pathogens
(Thompson et al. 2004), their abundance and diversity in
coastal waters, their ability to readily develop and acquire
antibiotic resistance in response to selective pressure and
their ability to spread resistance by horizontal genetic
material exchanges (Aoki 2000). Traditionally, Vibrio is
considered highly susceptible to all antimicrobials (Oliver
2006). Tetracycline has been recommended as the antimi-
crobial of choice to treat severe Vibrio human infections
(Morris and Tenney 1985), and alternative treatments are a
combination of third-generation cephalosporins (e.g.,
ceftazidime) and doxycycline, or a Fluoroquinolones alone
(Tang et al. 2002).
The increase in multi-antibiotics resistance bacteria in
recent years is worrisome and the presence of resistance gene
in bacteria has further enhanced the transmission and spread
of drugs resistance among microbial pathogens. Resistance
to antibacterial can, in fact, be reached either with a step wise
progression from low to high resistance levels through
sequential mutations in chromosomal genes (Wang et al.
2001), or through the acquisition of mobile genetic elements
such as bacteriophages, plasmid, naked DNA or transposons
(Levy and Marshall 2004), whose transmission between
bacteria, even belonging to different taxonomic and
ecological groups, contributes to the diffusion of antibiotic
resistance gene in the environment (Wang et al. 2006).
Furthermore, inappropriate use of antibiotics is likely to
cause an unnecessary impact on the environment. Therefore,
standardisation and safety of drugs used in aquaculture for
protection of the environment and human has recently been
emphasised (Scholtfeldt 1992).
The aim of the present study was to investigate the
antibiotic susceptibilities of 69 V. alginolyticus strains
isolated from different marine Tunisian biotopes (seawater,
aquaculture and conchylicole farms, sediment, river con-
nected to the Mediterranean seawater) using both the disc
diffusion assay and the microdilution method. Moreover,
the presence of multiple antibiotic resistance of V. algi-
nolyticus in Tunisian biotopes was assessed. In addition,
the correlation between antibiotic resistance and presence
of plasmids was undertaken.
Materials and methods
Sampling sites and strains identification
The strains were isolated from four marine biotopes
including two fish farms where S. aurata and D. labrax are
reared (Khenis and Hergla), from the conchylicole station
of Menzel Jmil (M. edulis and C. gigas) and from Oued
Soltane which is in connection with Mediterranean sea-
water during the cold seasons.
Seawater samples were filtered through a 0.45 lm
membranes, cultured in alkaline peptone water (1 % NaCl,
pH 8.6) and incubated at 37 �C for 18–24 h. A loopful of
the enrichment culture was streaked onto Thiosulphate-
citrate-bile salt-sucrose agar (TCBS, Difco, Spain). Yellow
colonies were randomly selected then subcultured on
Tryptic soy agar (TSA, Difco, Spain) supplemented with
1 % NaCl. Confirmation of the purity of cultures was
obtained for each strain by re-streaking on tryptic soy agar
added with 1 % NaCl. The isolated bacteria were frozen at
-80 �C with 20 % (v/v) glycerol for further analysis.
Seventy-eight Vibrio strains were analyzed in this study
including sixty-nine strains of V. alginolyticus, nine refer-
ence strains including seven strains of V. alginolyticus (CCM
2575, CCM 2576, CCM 2578T, ATCC 33787, ATCC
17749T, I12, I14), one V. parahaemolyticus type strain
(ATCC 43969) and one V. harveyi (CAIM 86). The strains
were identified by the following phenotypic tests: cell mor-
phology and motility, Gram staining (KOH method: Fluharty
and Packard 1967), oxidase, growth on TCBS, susceptibility
to the vibriostatic agent 0/129 (150 lg/disc), production of
arginine dihydrolase, lysine and ornithine decarboxylase,
glucose fermentation, indole, hydrolysis of gelatin, starch,
esculin and Tween 80, reduction of nitrate to nitrite,
World J Microbiol Biotechnol
123

production of gas from glucose, methyl red, growth at dif-
ferent temperatures (4, 37, 44 �C) and at different salinities
(0, 6, 8 and 10 %). These tests were the main assays
employed to identify the organisms belonging to Vibrio
genus (Thompson et al. 2004). The DNA extraction and
molecular identification of V. alginolyticus strains was done
according to the protocol described by Di-Pinto et al. (2005)
targeting the collagenase gene.
Determination of antibiotic susceptibility
The antibiotic susceptibility was determined by using the
Kirby-Bauer method and Mueller–Hinton agar plates sup-
plemented with 1 % NaCl as described by Ottaviani et al.
(2001). Antibiotics tested are as follow: Ampicilline
(AMP) 10 lg, Cefotaxime (CTX) 30 lg, Chloramphenicol
(C) 30 lg, Fosfomycin (FOS) 200 lg, Gentamycin (CN)
10 lg, Imipenem (IMI) 10 lg, Kanamycin (K) 30 lg,
Nalidixic Acid (NA) 30 lg, Norfloxacine (NOR) 10 lg,
Streptomycin (S) 10 lg, Sulfamethoxazole (SMX)
50 lg, Trimethoprime (TM) 5 lg, Doxycycline (DXT)
30 lg, Nitrofurantoine (F) 300 lg, Cephalothin (KF) 30 lg,
Erythromycin (E) 15 lg, Ticarcilline (TC) 75 lg, Cipro-
floxacin (CIP) 5 lg, Co-Trimoxazole Trimethoprime ?
Sulfamethoxazole (SXT) 25 lg, Amikacin (AK) 30 lg
(Liofilchem s.r.l., Roseto, Italy).
After incubation at 37 �C for 18–24 h, the diameter of the
inhibition zone was measured with 1 mm flat rule and the
diameters were interpreted according to CLSI: Performance
Standards for Antimicrobial Disk and Dilution Susceptibility
Tests for Bacteria Isolates From animals (2008). For the two
antibiotics (Fosfomycin and Doxycycline), results of the
diameters of inhibition were interpreted according the
diameters indicated by the Liofilchem company.
The antibiotic resistance index (ARI) of each bacterial
population was determined using the following formula:
ARI = y/nx, where y was the actual number of resistance
determinants recorded in a population of a size n, and x
was the total number of antibacterial tested for in the
sensitivity test. Based on the occurrence of resistance to
more than three antibiotics the isolates were grouped as
multiple antibiotic resistant isolates. The multiple antibi-
otic resistance (MAR) an d ARI indexes were done as
reported by Snoussi et al. (2011) for Vibrio strains. The
MAR index was defined as a/b where a represents the
number of multiple antibiotics to which the particular
isolate is resistant and b as the number of multiple antibi-
otics to which the particular isolates were exposed. A MAR
index value of B0.2 was an indication that the antibiotics
were seldom or never been used for animals treatment
whereas the MAR index value of [0.2 was considered as
an indication that the animals received high exposure to the
antibiotics (Sarter et al. 2007).
Minimum inhibitory concentration determination
The broth Microdilution method was used to determine the
Minimum inhibitory concentration (MIC) and Minimum
bactericidal concentration (MBC) of eleven antibiotics. An
overnight culture at 37 �C of each strain was diluted ten-
fold in fresh Mueller–Hinton broth (Biorad, France) sup-
plemented with 1 % NaCl and incubated at 37 �C until
they reached exponential phase. Serial twofold dilutions of
the tested antibiotics were prepared on 96-wells plate
(190 ll per well). Ten microlitres of the inocula
(OD600 = 1) were added to each well and the tested anti-
biotic. In each plate, two wells were reserved to control the
sterility of the medium used (no inoculum added) and the
viability of the inoculum (no antibiotic added). After 24 h
of incubation at 37 �C, bacterial growth was visually
evaluated by the presence of turbidity and a pellet on the
U-bottom of the 96-wells plate. The MIC value was defined
as the lowest concentration of antibiotic that inhibited
visible cell growth after 24 h of incubation at 37 �C
comparatively to the control well without antibiotic.
Minimum bactericidal concentration determination
The minimum bactericidal concentration was defined as the
lowest concentration of antibiotic able to kill 99 % of
bacteria in the well. For this, 10 microtiter of each well
medium with no visible growth was plated on MH-1 %
NaCl plates and the survived bacteria were estimated after
24 h of incubation at 37 �C.
Plasmid profiling
Cells were grown on overnight in 3 ml of Luria Broth. The
plasmid-DNA extraction was performed as described by
the protocol of alkaline exraction method described by
Birnboim and Doly (1979) and modified by Sambrook
et al. (1989). DNA was electrophoresed on 0.7 % agarose
gel. DNA bands were visualized under ultraviolet transil-
lumination and were photographed. V. alginolyticus
plasmids sizes were estimated by comparaison with
Lambda-DNA-HindIII Marker (Promega, Madison, WI,
USA). The analyses were repeated three times.
Result and discussion
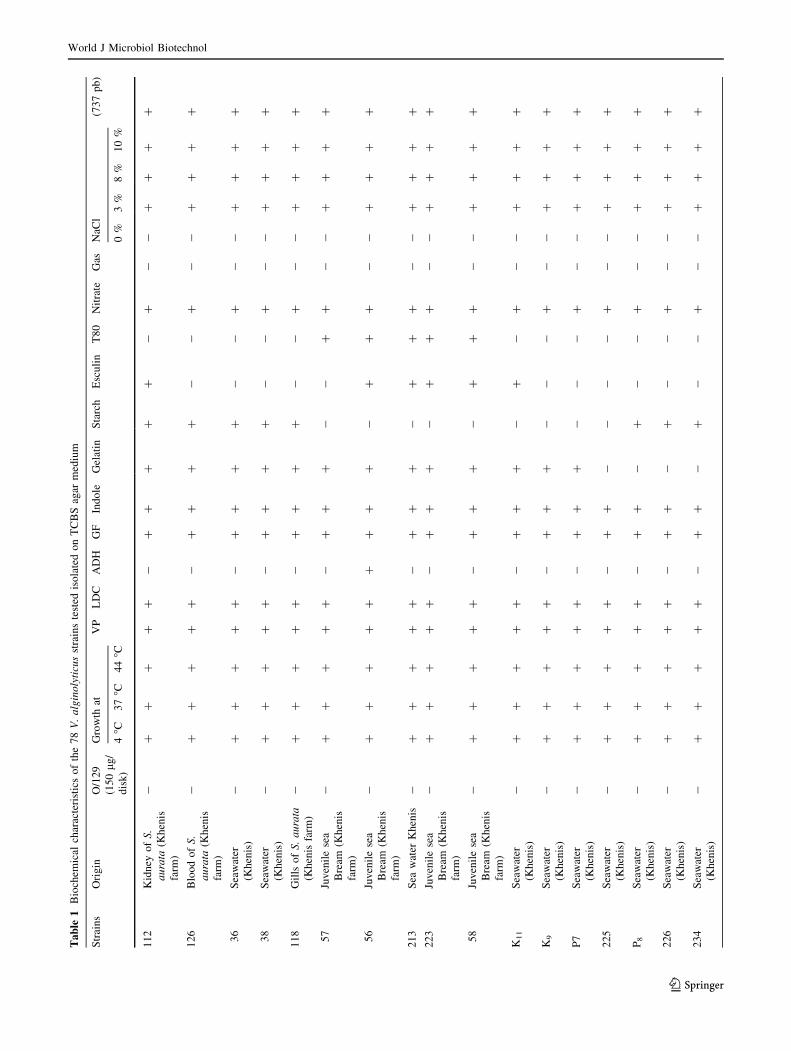
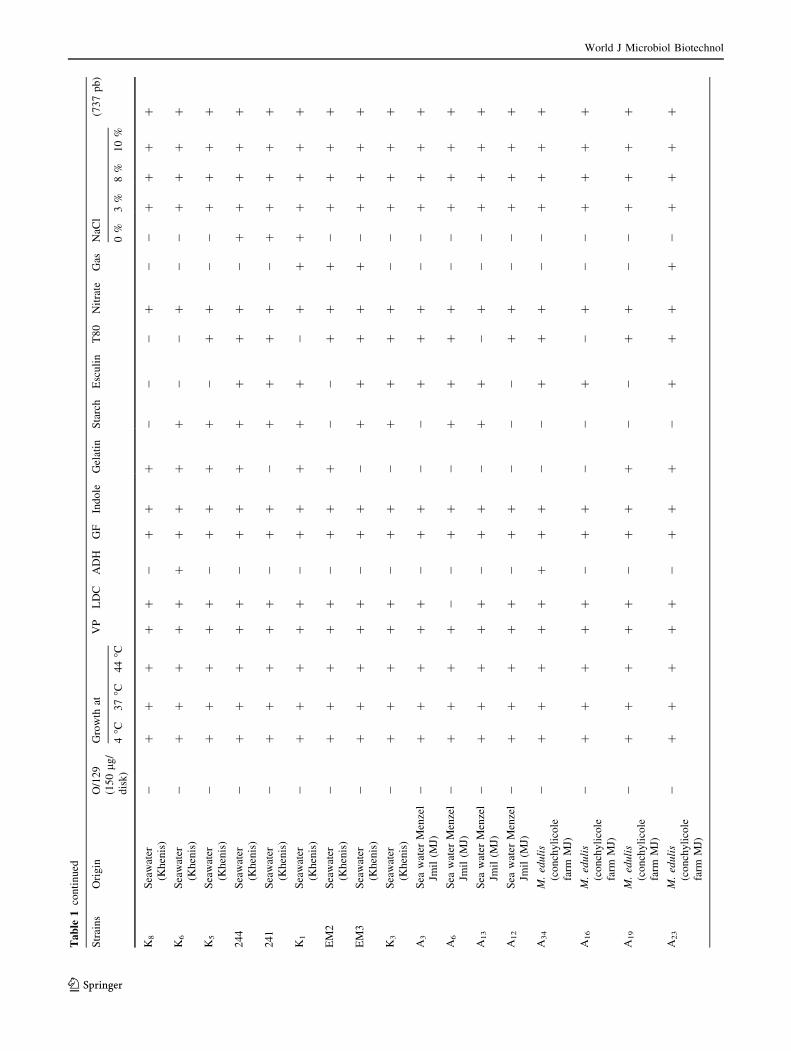
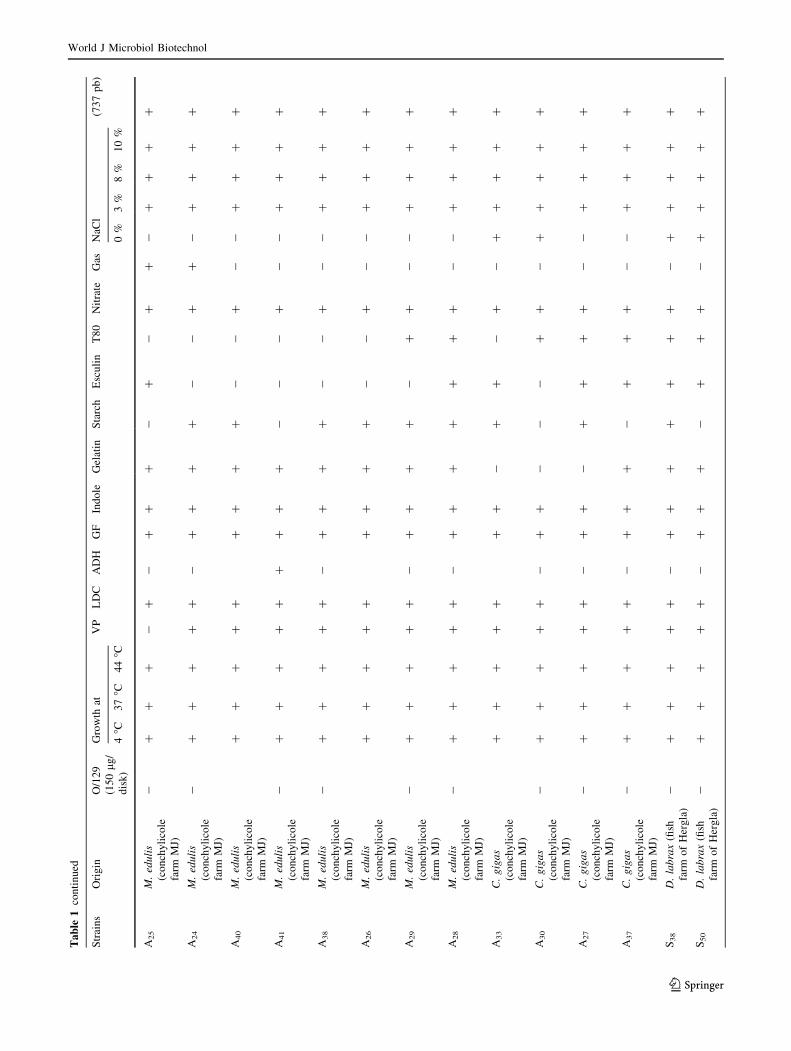
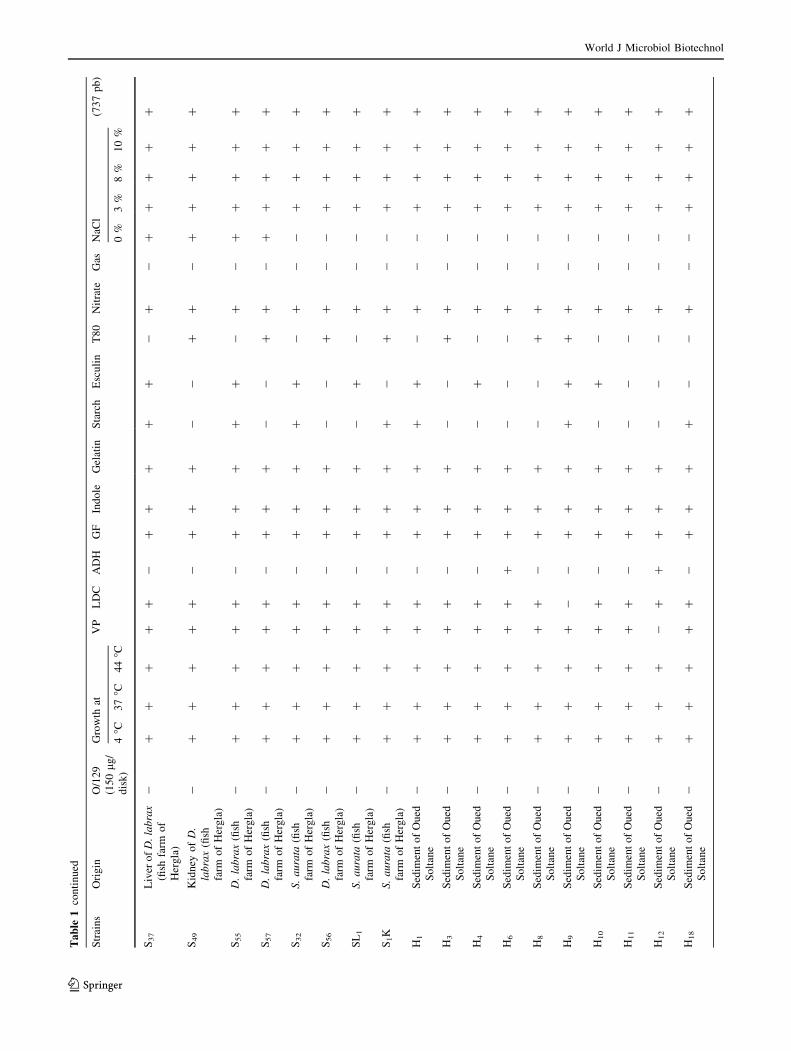
Biochemical and molecular identification
of the isolated strains
Sixty-nine V. alginolyticus strains isolated from four mar-
ine biotopes including two fish farms where S. aurata and
D. labrax are reared (Khenis and Hergla), conchylicole
World J Microbiol Biotechnol
123

station (M. edulis and C. gigas) and from a river ‘‘Soltane’’
which is in connection with Mediterranean seawaters dur-
ing the cold seasons (Fig. 1). Biochemical and phenotypic
identification were based on many specific traits (Table 1).
Yellow colonies isolated from TCBS agar were identified
as Gram-negative motile fermentative rods, producing
enzymes like catalase and oxidase, susceptible to vibrio-
static compounds O/129 (150 lg/disk) and swarming col-
onies on TSA 1 % NaCl at different temperature: 4, 37,
44 �C. Most strains (67/69) were Voges-Proskauer and
lysine decarboxylase positive. Only six strains were orni-
thine decarboxylase positive. However, all strains were
negative for arginine dihydrolase. All, the strains grew in
peptone water prepared respectively with 3, 8 and 10 % of
NaCl (Table 1). All strains tested amplify a 737-pb size
fragment showing the characteristic profile of V. algino-
lyticus (Di-Pinto et al. 2005).
Antibiotic susceptibility
The antibiotic susceptibility of V. alginolyticus strains
showed wide resistance to previously tested antibiotics
(Fig. 2). In fact, the results showed that most strains (59/
69; 85.5 %) were resistant to at least six antimicrobials
agents (Table 2). Fifteen strains isolated from the seawater
in the region of Khenis (strains: 57, 244, k11, k5, 213), from
the mussels in Menzel Jmil (strains: A16, A19, A30, A29),
from fish in Hergla (strains: S50, S38) and from the Oued
sultan in Borj-Cedria (strains: H1, H18, H20) were resistant
to all antibiotic tested in this study. The strains tested were
resistant to ampicillin (94.2 %), erythromycin (85.5 %),
kanamycin (84 %), gentamycin (76.8 %) and cefotaxim
(75.3 %). The lowest percentages of resistance were noted
for trimethoprim-sulfamethoxazole (20.2 %), ciprofloxacin
(26 %), and nalidixic acid (31.8 %).
Fig. 1 Map of Tunisia showing the different sites of study and the number of V. alginolyticus strains isolated from each sample (seawater, fish
and shellfish samples)
World J Microbiol Biotechnol
123
Ta
ble
1B
ioch
emic
alch
arac
teri
stic
so
fth
e7
8V
.a
lgin
oly
ticu
sst
rain
ste
sted
iso
late
do
nT
CB
Sag
arm
ediu
m
Str
ain
sO
rig
inO
/12
9
(15
0lg
/
dis
k)
Gro
wth
atV
PL
DC
AD
HG
FIn
do
leG
elat
inS
tarc
hE
scu
lin
T8
0N
itra
teG
asN
aCl
(73
7p
b)
4�C
37
�C4
4�C
0%
3%
8%
10
%
11
2K
idn
eyo
fS
.a
ura
ta(K
hen
is
farm
)
-?
??
??
-?
??
??
-?
--
??
??
12
6B
loo
do
fS
.a
ura
ta(K
hen
is
farm
)
-?
??
??
-?
??
?-
-?
--
??
??
36
Sea
wat
er
(Kh
enis
)
-?
??
??
-?
??
?-
-?
--
??
??
38
Sea
wat
er
(Kh
enis
)
-?
??
??
-?
??
?-
-?
--
??
??
11
8G
ills
of
S.
au
rata
(Kh
enis
farm
)
-?
??
??
-?
??
?-
-?
--
??
??
57
Juv
enil
ese
a
Bre
am(K
hen
is
farm
)
-?
??
??
-?
??
--
??
--
??
??
56
Juv
enil
ese
a
Bre
am(K
hen
is
farm
)
-?
??
??
??
??
-?
??
--
??
??
21
3S
eaw
ater
Kh
enis
-?
??
??
-?
??
-?
??
--
??
??
22
3Ju
ven
ile
sea
Bre
am(K
hen
is
farm
)
-?
??
??
-?
??
-?
??
--
??
??
58
Juv
enil
ese
a
Bre
am(K
hen
is
farm
)
-?
??
??
-?
??
-?
??
--
??
??
K11
Sea
wat
er
(Kh
enis
)
-?
??
??
-?
??
-?
-?
--
??
??
K9
Sea
wat
er
(Kh
enis
)
-?
??
??
-?
??
--
-?
--
??
??
P7
Sea
wat
er
(Kh
enis
)
-?
??
??
-?
??
--
-?
--
??
??
22
5S
eaw
ater
(Kh
enis
)
-?
??
??
-?
?-
--
-?
--
??
??
P8
Sea
wat
er
(Kh
enis
)
-?
??
??
-?
?-
?-
-?
--
??
??
22
6S
eaw
ater
(Kh
enis
)
-?
??
??
-?
?-
?-
-?
--
??
??
23
4S
eaw
ater
(Kh
enis
)
-?
??
??
-?
?-
?-
-?
--
??
??
World J Microbiol Biotechnol
123
Ta
ble
1co
nti
nu
ed
Str
ain
sO
rig
inO
/12
9
(15
0lg
/
dis
k)
Gro
wth
atV
PL
DC
AD
HG
FIn
do
leG
elat
inS
tarc
hE
scu
lin
T8
0N
itra
teG
asN
aCl
(73
7p
b)
4�C
37
�C4
4�C
0%
3%
8%
10
%
K8
Sea
wat
er
(Kh
enis
)
-?
??
??
-?
??
--
-?
--
??
??
K6
Sea
wat
er
(Kh
enis
)
-?
??
??
??
??
?-
-?
--
??
??
K5
Sea
wat
er
(Kh
enis
)
-?
??
??
-?
??
?-
??
--
??
??
24
4S
eaw
ater
(Kh
enis
)
-?
??
??
-?
??
??
??
-?
??
??
24
1S
eaw
ater
(Kh
enis
)
-?
??
??
-?
?-
??
??
-?
??
??
K1
Sea
wat
er
(Kh
enis
)
-?
??
??
-?
??
??
-?
??
??
??
EM
2S
eaw
ater
(Kh
enis
)
-?
??
??
-?
??
--
??
?-
??
??
EM
3S
eaw
ater
(Kh
enis
)
-?
??
??
-?
?-
??
??
?-
??
??
K3
Sea
wat
er
(Kh
enis
)
-?
??
??
-?
?-
??
??
--
??
??
A3
Sea
wat
erM
enze
l
Jmil
(MJ)
-?
??
??
-?
?-
-?
??
--
??
??
A6
Sea
wat
erM
enze
l
Jmil
(MJ)
-?
??
?-
-?
?-
??
??
--
??
??
A13
Sea
wat
erM
enze
l
Jmil
(MJ)
-?
??
??
-?
?-
??
-?
--
??
??
A12
Sea
wat
erM
enze
l
Jmil
(MJ)
-?
??
??
-?
?-
--
??
--
??
??
A34
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
-?
??
??
??
?-
-?
??
--
??
??
A16
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
-?
??
??
-?
?-
-?
-?
--
??
??
A19
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
-?
??
??
-?
??
--
??
--
??
??
A23
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
-?
??
??
-?
??
-?
??
?-
??
??
World J Microbiol Biotechnol
123
Ta
ble
1co
nti
nu
ed
Str
ain
sO
rig
inO
/12
9
(15
0lg
/
dis
k)
Gro
wth
atV
PL
DC
AD
HG
FIn
do
leG
elat
inS
tarc
hE
scu
lin
T8
0N
itra
teG
asN
aCl
(73
7p
b)
4�C
37
�C4
4�C
0%
3%
8%
10
%
A25
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
-?
??
-?
-?
??
-?
-?
?-
??
??
A24
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
-?
??
??
-?
??
?-
-?
?-
??
??
A40
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
??
??
??
??
?-
-?
--
??
??
A41
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
-?
??
??
??
??
--
-?
--
??
??
A38
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
-?
??
??
-?
??
?-
-?
--
??
??
A26
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
??
??
??
??
?-
-?
--
??
??
A29
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
-?
??
??
-?
??
?-
??
--
??
??
A28
M.
edu
lis
(co
nch
yli
cole
farm
MJ)
-?
??
??
-?
??
??
??
--
??
??
A33
C.
gig
as
(co
nch
yli
cole
farm
MJ)
??
??
??
?-
??
-?
-?
??
??
A30
C.
gig
as
(co
nch
yli
cole
farm
MJ)
-?
??
??
-?
?-
--
??
-?
??
??
A27
C.
gig
as
(co
nch
yli
cole
farm
MJ)
-?
??
??
-?
?-
??
??
--
??
??
A37
C.
gig
as
(co
nch
yli
cole
farm
MJ)
-?
??
??
-?
??
-?
??
--
??
??
S38
D.
lab
rax
(fish
farm
of
Her
gla
)
-?
??
??
-?
??
??
??
-?
??
??
S50
D.
lab
rax
(fish
farm
of
Her
gla
)
-?
??
??
-?
??
-?
??
-?
??
??
World J Microbiol Biotechnol
123
Ta
ble
1co
nti
nu
ed
Str
ain
sO
rig
inO
/12
9
(15
0lg
/
dis
k)
Gro
wth
atV
PL
DC
AD
HG
FIn
do
leG
elat
inS
tarc
hE
scu
lin
T8
0N
itra
teG
asN
aCl
(73
7p
b)
4�C
37
�C4
4�C
0%
3%
8%
10
%
S37
Liv
ero
fD
.la
bra
x(fi
shfa
rmo
f
Her
gla
)
-?
??
??
-?
??
??
-?
-?
??
??
S49
Kid
ney
of
D.
lab
rax
(fish
farm
of
Her
gla
)
-?
??
??
-?
??
--
??
-?
??
??
S55
D.
lab
rax
(fish
farm
of
Her
gla
)
-?
??
??
-?
??
??
-?
-?
??
??
S57
D.
lab
rax
(fish
farm
of
Her
gla
)
-?
??
??
-?
??
--
??
-?
??
??
S32
S.
au
rata
(fish
farm
of
Her
gla
)
-?
??
??
-?
??
??
-?
--
??
??
S56
D.
lab
rax
(fish
farm
of
Her
gla
)
-?
??
??
-?
??
--
??
--
??
??
SL
1S
.a
ura
ta(fi
sh
farm
of
Her
gla
)
-?
??
??
-?
??
-?
-?
--
??
??
S1K
S.
au
rata
(fish
farm
of
Her
gla
)
-?
??
??
-?
??
?-
??
--
??
??
H1
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
-?
??
??
-?
--
??
??
H3
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
-?
??
--
??
--
??
??
H4
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
-?
??
-?
-?
--
??
??
H6
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
??
??
--
-?
--
??
??
H8
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
-?
??
--
??
--
??
??
H9
Sed
imen
to
fO
ued
So
ltan
e
-?
??
?-
-?
??
??
??
--
??
??
H10
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
-?
??
-?
-?
--
??
??
H11
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
-?
??
--
-?
--
??
??
H12
Sed
imen
to
fO
ued
So
ltan
e
-?
??
-?
??
??
--
-?
--
??
??
H18
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
-?
??
?-
-?
--
??
??
World J Microbiol Biotechnol
123

Ta
ble
1co
nti
nu
ed
Str
ain
sO
rig
inO
/12
9
(15
0lg
/
dis
k)
Gro
wth
atV
PL
DC
AD
HG
FIn
do
leG
elat
inS
tarc
hE
scu
lin
T8
0N
itra
teG
asN
aCl
(73
7p
b)
4�C
37
�C4
4�C
0%
3%
8%
10
%
H20
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
-?
??
?-
??
--
??
??
H21
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
-?
??
??
-?
--
??
??
H22
Sed
imen
to
fO
ued
So
ltan
e
-?
??
??
-?
??
--
-?
--
??
??
AT
CC
17
74
9
Ty
pe
stra
in-
??
??
?-
??
-?
--
?-
-?
??
?
AT
CC
33
78
7
Ty
pe
stra
in-
??
??
?-
??
?-
-?
??
-?
??
?
AT
CC
43
96
9
Ty
pe
stra
in-
??
??
?-
??
??
??
?-
-?
??
?
CA
IM8
6T
yp
est
rain
-?
??
??
-?
??
-?
??
--
??
??
CC
M2
57
5T
yp
est
rain
-?
??
??
-?
??
-?
-?
-?
??
??
CC
M2
57
6T
yp
est
rain
-?
??
??
-?
??
?-
??
--
??
??
CC
M2
57
8T
yp
est
rain
-?
??
??
-?
?-
??
-?
--
??
??
I 12
Ital
y-
??
??
?-
??
??
-?
?-
-?
??
?
I 14
Ital
y-
??
??
?-
??
?-
??
?-
-?
??
?
VP
Vo
ges
-Pro
skau
er,
LD
CL
ysi
ne
Dec
arb
ox
yla
se,
AD
HA
rgin
ine
Dih
yd
rola
se,
GF
glu
cose
ferm
enta
tio
n,
T8
0T
wee
n8
0,
Ga
zG
asp
rod
uct
ion
fro
mK
lig
ler-
Haj
na,
Nit
rate
Nit
rate
red
uct
ion
World J Microbiol Biotechnol
123
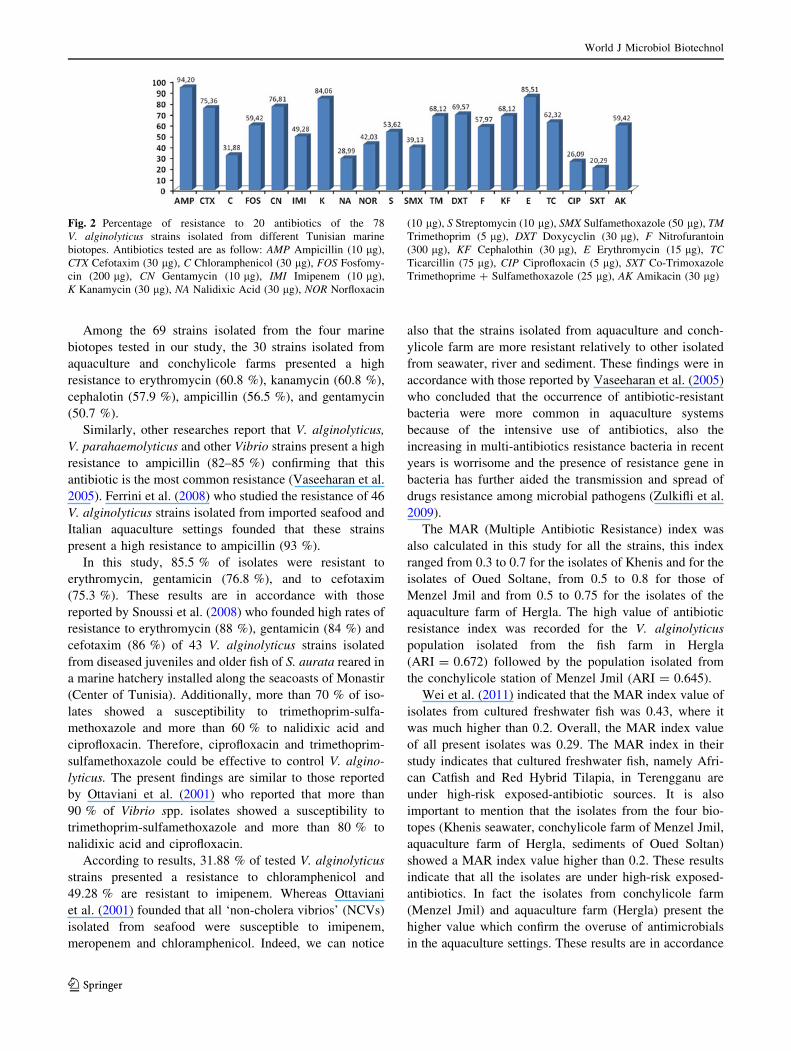
Among the 69 strains isolated from the four marine
biotopes tested in our study, the 30 strains isolated from
aquaculture and conchylicole farms presented a high
resistance to erythromycin (60.8 %), kanamycin (60.8 %),
cephalotin (57.9 %), ampicillin (56.5 %), and gentamycin
(50.7 %).
Similarly, other researches report that V. alginolyticus,
V. parahaemolyticus and other Vibrio strains present a high
resistance to ampicillin (82–85 %) confirming that this
antibiotic is the most common resistance (Vaseeharan et al.
2005). Ferrini et al. (2008) who studied the resistance of 46
V. alginolyticus strains isolated from imported seafood and
Italian aquaculture settings founded that these strains
present a high resistance to ampicillin (93 %).
In this study, 85.5 % of isolates were resistant to
erythromycin, gentamicin (76.8 %), and to cefotaxim
(75.3 %). These results are in accordance with those
reported by Snoussi et al. (2008) who founded high rates of
resistance to erythromycin (88 %), gentamicin (84 %) and
cefotaxim (86 %) of 43 V. alginolyticus strains isolated
from diseased juveniles and older fish of S. aurata reared in
a marine hatchery installed along the seacoasts of Monastir
(Center of Tunisia). Additionally, more than 70 % of iso-
lates showed a susceptibility to trimethoprim-sulfa-
methoxazole and more than 60 % to nalidixic acid and
ciprofloxacin. Therefore, ciprofloxacin and trimethoprim-
sulfamethoxazole could be effective to control V. algino-
lyticus. The present findings are similar to those reported
by Ottaviani et al. (2001) who reported that more than
90 % of Vibrio spp. isolates showed a susceptibility to
trimethoprim-sulfamethoxazole and more than 80 % to
nalidixic acid and ciprofloxacin.
According to results, 31.88 % of tested V. alginolyticus
strains presented a resistance to chloramphenicol and
49.28 % are resistant to imipenem. Whereas Ottaviani
et al. (2001) founded that all ‘non-cholera vibrios’ (NCVs)
isolated from seafood were susceptible to imipenem,
meropenem and chloramphenicol. Indeed, we can notice
also that the strains isolated from aquaculture and conch-
ylicole farm are more resistant relatively to other isolated
from seawater, river and sediment. These findings were in
accordance with those reported by Vaseeharan et al. (2005)
who concluded that the occurrence of antibiotic-resistant
bacteria were more common in aquaculture systems
because of the intensive use of antibiotics, also the
increasing in multi-antibiotics resistance bacteria in recent
years is worrisome and the presence of resistance gene in
bacteria has further aided the transmission and spread of
drugs resistance among microbial pathogens (Zulkifli et al.
2009).
The MAR (Multiple Antibiotic Resistance) index was
also calculated in this study for all the strains, this index
ranged from 0.3 to 0.7 for the isolates of Khenis and for the
isolates of Oued Soltane, from 0.5 to 0.8 for those of
Menzel Jmil and from 0.5 to 0.75 for the isolates of the
aquaculture farm of Hergla. The high value of antibiotic
resistance index was recorded for the V. alginolyticus
population isolated from the fish farm in Hergla
(ARI = 0.672) followed by the population isolated from
the conchylicole station of Menzel Jmil (ARI = 0.645).
Wei et al. (2011) indicated that the MAR index value of
isolates from cultured freshwater fish was 0.43, where it
was much higher than 0.2. Overall, the MAR index value
of all present isolates was 0.29. The MAR index in their
study indicates that cultured freshwater fish, namely Afri-
can Catfish and Red Hybrid Tilapia, in Terengganu are
under high-risk exposed-antibiotic sources. It is also
important to mention that the isolates from the four bio-
topes (Khenis seawater, conchylicole farm of Menzel Jmil,
aquaculture farm of Hergla, sediments of Oued Soltan)
showed a MAR index value higher than 0.2. These results
indicate that all the isolates are under high-risk exposed-
antibiotics. In fact the isolates from conchylicole farm
(Menzel Jmil) and aquaculture farm (Hergla) present the
higher value which confirm the overuse of antimicrobials
in the aquaculture settings. These results are in accordance
Fig. 2 Percentage of resistance to 20 antibiotics of the 78
V. alginolyticus strains isolated from different Tunisian marine
biotopes. Antibiotics tested are as follow: AMP Ampicillin (10 lg),
CTX Cefotaxim (30 lg), C Chloramphenicol (30 lg), FOS Fosfomy-
cin (200 lg), CN Gentamycin (10 lg), IMI Imipenem (10 lg),
K Kanamycin (30 lg), NA Nalidixic Acid (30 lg), NOR Norfloxacin
(10 lg), S Streptomycin (10 lg), SMX Sulfamethoxazole (50 lg), TMTrimethoprim (5 lg), DXT Doxycyclin (30 lg), F Nitrofurantoin
(300 lg), KF Cephalothin (30 lg), E Erythromycin (15 lg), TCTicarcillin (75 lg), CIP Ciprofloxacin (5 lg), SXT Co-Trimoxazole
Trimethoprime ? Sulfamethoxazole (25 lg), AK Amikacin (30 lg)
World J Microbiol Biotechnol
123

Table 2 Correlation between resistance patterns and plasmid profiles
Strains Resistance patterns Plasmid profiles
112 R9: AMP-FOS-CN-S-TM-DXT-E-CIP Plasmid free
126 R10: AMP-CTX-IMI-K-NOR-DXT-F-E-TC-CIP P11: 2.3
36 R11: AMP-FOS-CN-K-NA-SMX-F-KF-TC-CIP P1: 1.5
38 R11: AMP-FOS-CN-K-NA-SMX-F-KF-TC-CIP P1: 1.5
118 R12: AMP-FOS-CN-K-S-TM-DXT-E-CIP P11: 2.3
57 R13: AMP-CTX-C-NA-S-TM-DXT-F-TC-CIP-SXT P18: 9.4; 3; 2.3
56 R14: AMP-IMI-K-NOR-DXT-KF-SXT P10: 1; 0.8; 0.6; 0.5
213 R15: AMP-CTX-FOS-K-NOR-S-DXT-F-E-SXT-AK P3: 45; 1.5
223 R16: AMP-FOS-K-S-SMX-TC-CIP-AK P18: 9.4; 3; 2.3
58 R17: AMP-C-K-NA-NOR-DXT-F-E-CIP-AK P13: 9.4
K11 R18:AMP-CTX-C-CN-IMI-S-DXT-F-E-TC-CIP-AK P20: 23.2; 3
K9 R18:AMP-CTX-C-CN-IMI-S-DXT-F-E-TC-CIP-AK P4: 4.3; 1.5
P7 R19: AMP-CTX-C-IMI-K-S-SMX-DXT-KF-E-TC-CIP P8: 0.8; 0.6; 0.5
225 R20: AMP-K-NA-NOR-S-DXT-TC-CIP-AK P12: 6.5
P8 R21: AMP-CTX-K-NA-DXT-F-TC-AK P19: 9.4; 3
226 R15: AMP-CTX-FOS-K-NOR-S-DXT-F-E-SXT-AK P3: 45; 1.5
234 R11: AMP-FOS-CN-K-NA-SMX-F-KF-TC-CIP P1: 1.5
K8 R22: AMP-CTX-C-FOS-CN-K-TM-DXT-KF-CIP-AK P19: 9.4; 3
K6 R23: AMP-CTX-C-FOS-CN-IMI-K-S-SMX-DXT Plasmid free
K5 R23: AMP-CTX-C-FOS-CN-IMI-K-S-SMX-DXT Plasmid free
244 R24: AMP-CTX-K-NA-K-S-TM-DXT-F-KF-E-TC-CIP P12: 6.5
241 R22: AMP-CTX-C-FOS-CN-K-TM-DXT-KF-CIP-AK Plasmid free
K1 R24: AMP-CTX-K-NA-K-S-TM-DXT-F-KF-E-TC-CIP P6: 23.2; 2.3
EM2 R25: AMP-CTX-CN-K-S-SMX-TC P21: 9.4; 2.3
EM3 R25: AMP-CTX-CN-K-S-SMX-TC P21: 9.4; 2.3
K3 R26: AMP-CTX-C-FOS-K-NOR-TM-DXT-E-TC P7: 0.8; 0.6
A3 R27: AMP-CN-IMI-K-TM-F-E-TC-AK Plasmid free
A6 R28: AMP-CTX-FOS-IMI-K-KF-E-TC P2: 45
A13 R28: AMP-CTX-FOS-IMI-K-KF-E-TC P2: 45
A12 R29: AMP-CTX-FOS-IMI-K-NA-SMX-TM-DXT-TC-CIP P5: 23.2
A34 R29: AMP-CTX-FOS-IMI-K-NA-SMX-TM-DXT-TC-CIP P5: 23.2
A16 R30: CTX-CN-IMI-K-NA-DXT-F-KF-E-TC-CIP Plasmid free
A19 R30: CTX-CN-IMI-K-NA-DXT-F-KF-E-TC-CIP P29: 45; 15
A23 R31: AMP-FOS-IMI-K-TM-DXT-F-KF-E-TC-CIP-SXT P23: 23.2; 9.4; 2.3
A25 R31: AMP-FOS-IMI-K-TM-DXT-F-KF-E-TC-CIP-SXT P5: 23.2
A24 R32: AMP-CTX-C-FOS-CN-IMI-K-NA-SMX-TM-F-KF-E-TC-AK P2: 45
A40 R33: AMP-FOS-IMI-K-NA-S-DXT-KF-E-TC Plasmid free
A41 R34: AMP-CN-IMI-K-S-SMX-TM-DXT-F-E-TC-CIP-SXT P5: 23.2
A38 R35: AMP-CN-IMI-K-NA-S-TM-DXT-F-E-TC-AK Plasmid free
A26 R36: AMP-FOS-CN-IMI-K-TM-DXT-F-KF-E-TC-AK P29: 45; 15
A29 R36: AMP-FOS-CN-IMI-K-TM-DXT-F-KF-E-TC-AK P28: 15
A28 R37: C-FOS-IMI-K-S-SMX-TM-KF-E-SXT Plasmid free
A33 R38: AMP-FOS-IMI-K-NA-TM-DXT-F-KF-E-TC-CIP-AK P28: 15
A30 R39:CN-IMI-K-NA-S-SMX-F-TC- CIP-AK Plasmid free
A27 R40: AMP-C-IMI-K-NA-S-SMX-TM-F-E-TC-CIP-SXT Plasmid free
A37 R41: AMP-CTX-CN-IMI-K-DXT-F-KF-E-TC-CIP P22: 45; 30; 23; 6.5; 1.5
S38 R42: AMP-C-FOS-IMI-K-NA-TM-DXT-F-KF-E-TC-CIP-SXT P6: 23.2; 2.3
S50 R42: AMP-C-FOS-IMI-K-NA-TM-DXT-F-KF-E-TC-CIP-SXT P6: 23.2; 2.3
World J Microbiol Biotechnol
123

with those of Manjusha et al. (2005) who reported that
strains isolated from various tissue samples collected from
site of highest antibiotic resistance emphasizes the fact that
antibiotic resistance in fish and tissue samples augment at
alarming levels as compared to water samples.
According to Mukherji et al. (2000), V. alginolyticus has
been etiologically associated with cellulitis and acute otitis
media or externa. As a whole, these infections have
responded well to appropriate antibiotics. Seven Korean
cases of V. alginolyticus infection have previously been
reported. Four cases were related with otitis media (Doh
et al. 1997) or myringitis (Lee and Choi 1995) two with
soft tissue infection (Cho et al. 1995) and the last one was
related with gastroenteritis (Kim et al. 2000).
Lee et al. (2008) present a case of septic shock due to
V. alginolyticus in a patient in whom the clinical presentation
did not suggest the presence of this organism. He was treated
with the appropriate antibiotics, but his late visit to the hospital
and failure to achieve surgical debridement may have caused
his death. V. alginolyticus was susceptible to a variety of
antibiotics including ampicillin, amoxillin-clavulanate, ceph-
alothin, cefuroxime, gentamicin, ciprofloxacin, and trimeth-
oprim sulfamethoxazole. Immunocompromised hosts should
be careful about eating raw fish, especially during the warm
seasons. At present, thorough cooking of seafood is the only
effective means of prevention. Early administration of antibi-
otics and surgical intervention, if needed, is critical for con-
trolling these invasive vibrios infections.
Table 2 continued
Strains Resistance patterns Plasmid profiles
S37 R42: AMP-C-FOS-IMI-K-NA-TM-DXT-F-KF-E-TC-CIP-SXT P13: 9.4
S49 R42: AMP-C-FOS-IMI-K-NA-TM-DXT-F-KF-E-TC-CIP-SXT P13: 9.4
S55 R43: AMP-FOS-IMI-K- S-DXT-KF-E-TC Plasmid free
S57 R44: AMP-CTX-FOS-CN-IMI-K-NA-SMX-TM-DXT-K-KF-E-TC-CIP P26: 3
S32 R41: AMP-CTX-CN-IMI-K-DXT-F-KF-E-TC-CIP P22: 45; 30; 23; 6.5; 1.5
S56 R43: AMP-FOS-IMI-K-S-DXT-KF-E-TC Plasmid free
SL1 R9: AMP-FOS-CN-S-TM-DXT-E-CIP Plasmid free
S1K R9: AMP-FOS-CN-S-TM-DXT-E-CIP Plasmid free
H1 R45: AMP-CTX-K-DXT-F-TC-AK Plasmid free
H3 R46: AMP-CTX-FOS-CN-NA-TM-F P6: 23.2; 2.3
H4 R47: AMP-FOS-CN-K-NOR-S-DXT-E P9: 1; 0.8; 0.6
H6 R47: AMP-FOS-CN-K-NOR-S-DXT-E P9: 1; 0.8; 0.6
H8 R48: AMP-FOS-NA-F-E-TC- CIP-AK P11: 2.3
H9 R49: AMP-CTX-C-IMI-K-S-SMX-F-E-TC P12: 6.5
H10 R50: AMP-CTX-FOS-NOR-S-F-E-AK P11: 2.3
H11 R51: AMP-FOS-K-S-SMX-TC-CIP-AK P25: 4.3; 2.3
H12 R51: AMP-FOS-K-S-SMX-TC-CIP-AK P25: 4.3; 2.3
H18 R52:AMP-K-NA-NOR-S-DXT-TC-CIP-AK P24: 4.3
H20 R22: AMP-CTX-C-FOS-CN-K-TM-DXT-KF-CIP-AK P24: 4.3
H21 R22: AMP-CTX-C-FOS-CN-K-TM-DXT-KF-CIP-AK P24: 4.3
H22 R22: AMP-CTX-C-FOS-CN-K-TM-DXT-KF-CIP-AK P24: 4.3
ATCC 17749 R4: AMP-FOS-K-S-SMX-TC-CIP P13: 9.4
ATCC 33787 R5: AMP-K-NA-NOR-S-DXT-TC-AK P14: 6.5; 5.7; 4.3
ATCC 43969 R7: AMP-CTX-C-FOS-CN-IMI-TM-DXT-E-TC- AK P27: 3; 2.3; 2; 1.5
CAIM 86 R3: AMP-CTX-FOS-K-NA-NOR-S-TM-DXT-F-E-TC-CIP P30: 30; 23.2; 15; 3; 2.3; 1.5
CCM 2575 R1: AMP-FOS-CN-K-S-TM-DXT-E-CIP P16: 9.4; 2.3
CCM 2576 R1: AMP-FOS-CN-K-S-TM-DXT-E-CIP P16: 9.4; 2.3
CCM 2578 R2: AMP-FOS-NA-S-SMX-F-E-TC P16: 9.4; 2.3
I12 R6: AMP-CTX-C-FOS-CN-K-NOR-TM-DXT-CIP-AK P17: 23.2; 9.4
I14 R6: AMP-CTX-C-FOS-CN-K-NOR-TM-DXT-CIP-AK P15: 6.5; 2.3
AMP Ampicillin (10 lg), CTX Cefotaxim (30 lg), C Chloramphenicol (30 lg), FOS Fosfomycin (200 lg), CN Gentamycin (10 lg), IMIImipenem (10 lg), K Kanamycin (30 lg), NA Nalidixic Acid (30 lg), NOR Norfloxacin (10 lg), S Streptomycin (10 lg), SMX Sulfameth-
oxazole (50 lg), TM Trimethoprime (5 lg), DXT Doxycycline (30 lg), F Nitrofurantoine (300 lg), KF Cephalothin (30 lg), E Erythromycin
(15 lg), TC Ticarcillin (75 lg), CIP Ciprofloxacin (5 lg), SXT Co-Trimoxazole Trimethoprime ? Sulfamethoxazole (25 lg), AK Amikacin
(30 lg)
World J Microbiol Biotechnol
123

MICs of antibiotics
The results of the MICs and MBCs values tested using test
11 antimicrobials agents are listed in (Table 3). The results
obtained in this study are in accordance with those reported
by Ferrini et al. (2008) who found that MBC of ampicillin,
streptomycin, kanamycin, and ciprofloxacin are respec-
tively (C256, C128, 128 and 1 mg/l). In contrast, we noted
some differences between MIC90 reported by these
Authors for some antimicrobials such as tetracycline,
chloramphenicol, oxytetracycline and doxycycline. We can
notice that the results of the MIC values confirmed the
resistance of the V. alginolyticus tested to ampicillin, to
erythromycin, to kanamycin, to cefotaxime, to streptomy-
cin, to trimethoprim.
According to the results founded in the present work,
there is no difference in the MIC and MBC values of
ampicillin: 256 and [256 mg/l, respectively, which are in
accordance with those reported by Zanetti et al. (2001), but
a significant difference was observed between our MBC for
cefotaxime and doxycycline which were higher (64 and
8 mg/l respectively) than those reported by Zanetti et al.
(2001) in V. alginolyticus strains (0.12 and 0.25 mg/l,
respectively). Furthermore, the MIC range of oxytetracy-
cline to control V. alginolyticus was 0.12–1 mg/l, this
range is lower than that found in the study of Vaseeharan
et al. (2005) (22.8–33.5 mg/l). Roque et al. (2001), founded
that the MIC value of oxytetracycline was 301.0 mg/l
which is an extremely high value compared with other
reported results. This worldwide resistance to oxytetracy-
cline of Vibrio isolates from shrimp is probably due to its
frequent use; resistance is plasmid mediated and inducible,
allowing horizontal transfer of resistance (Towner. 1995).
Whereas, in the present study the majority of strains were
sensitive to the oxytetracycline and the MIC range of this
antibiotic was 0.12–1 mg/l.
In 2005, Vaseeharan and colleagues founded the MIC
range of ciprofloxacin of 0.32–0.43 mg/l was able to con-
trol effectively the Vibrio and Aeromonas species. This
result is in accordance with that reported by Zanetti et al.
(2001) who reported that the MIC of ciprofloxacin to
control Vibrio spp. isolated from the environment was
0.38 mg/l. Our result of the MIC of ciprofloxacin
(\0.06–1) was near to those founded in previous studies
(Zanetti et al. 2001; Vaseeharan et al. 2005) which confirm
that the ciprofloxacin is the most active of quinolones and
could be effective in the case of environmental V. algino-
lyticus. Additionally, we founded that the majority of
strains were sensitive to the oxytetracycline and the MIC
range of this antibiotic: 0.12–1 mg/l.
In a recent study on Vibrio spp. and Photobacterium
damsela ssp. picicida isolated from Italian aquaculture
farms (fish, shellfish and crustaceans), Lagana et al. (2011)
founded that the tested strains were resistant to ampicillin,
carbenicillin, cephalotin, kanamycin, while they were
sensitive to chloramphenicol, nitrofurantoin and to
tobramycin.
As a conclusion, the results obtained by the MIC tests
confirmed the resistance of the V. alginolyticus tested to
ampicillin (256 and [256 mg/l), erythromycin (64 and
[128 mg/l), kanamycin ([32 and 128), cefotaxime (1 and
2 mg/l), streptomycin (64 and [128 mg/l) and trimetho-
prim (16 and 32 mg/l).
Plasmid profiling
The plasmids profiles in vibrios have been studied in some
species such as Vibrio ordalii (Tianen et al. 1995), Vibrio
vulnificus (Radu et al. 1998) and Vibrio salmonicida
(Sorum et al. 1990), and most extensively in V. anguilla-
rum (Pedersen 1999) where a high diversity of profiles was
observed (Pedersen et al. 1996).
Plasmid profiling has proven to be useful proven to
differentiate between V. salmonicida, but their discrimi-
natory power has also been questioned (Pedersen 1999).
Our result (Table 2) showed 30 plasmid profiles among
them 12 profiles with the same resistance patterns. Addi-
tionally, sixty-two V. alginolyticus strains (79.5 %) har-
bored one to six plasmids with molecular sizes ranging
from 0.5 to 45 kb. According to plasmid content, strains
were classified into 30 clusters. We showed the presence of
fifteen plasmid with different sizes and only 16 strains
among 78 tested were plasmideless.
Zanetti et al. (2001) showed that only 24 of 48 strains
resistant to ampicillin harbored plasmids. The molecular
weight of these plasmids ranged from 1.5 to 26 kb. There are
also strains which had the same resistance patterns but dif-
ferent plasmids profiles. Zorilla et al. (2003) revealed the
presence of twelve different sized plasmids which were
detected in V. alginolyticus strains, originating eight differ-
ent plasmid patterns. A high percentage of the isolates tested
(53 %) did not carry plasmids and among the strains har-
boured plasmids, eight different plasmid profiles were
recorded. The plasmid content of V. alginolyticus was het-
erogeneous in number and size, varying from five to one, and
lower than 2.1–53.78 kb. The same authors showed that
some V. alginolyticus harbored the plasmid responsible for
virulence, and others strains possessed plasmid which had no
relationship with pathogenicity and this plasmid was sup-
posed to be responsible to antibiotic-resistance.
The results obtained showed that among 78 strains, 62
had one or more plasmids and there is a high difference in
the size of this plasmids variance between 0.5 and 45 kb.
These results are in accordance with those of Molina-Aja
et al. (2002) who found 24 strains (80 %) among 30 strains
with one or more plasmid and showed that there is a high
World J Microbiol Biotechnol
123
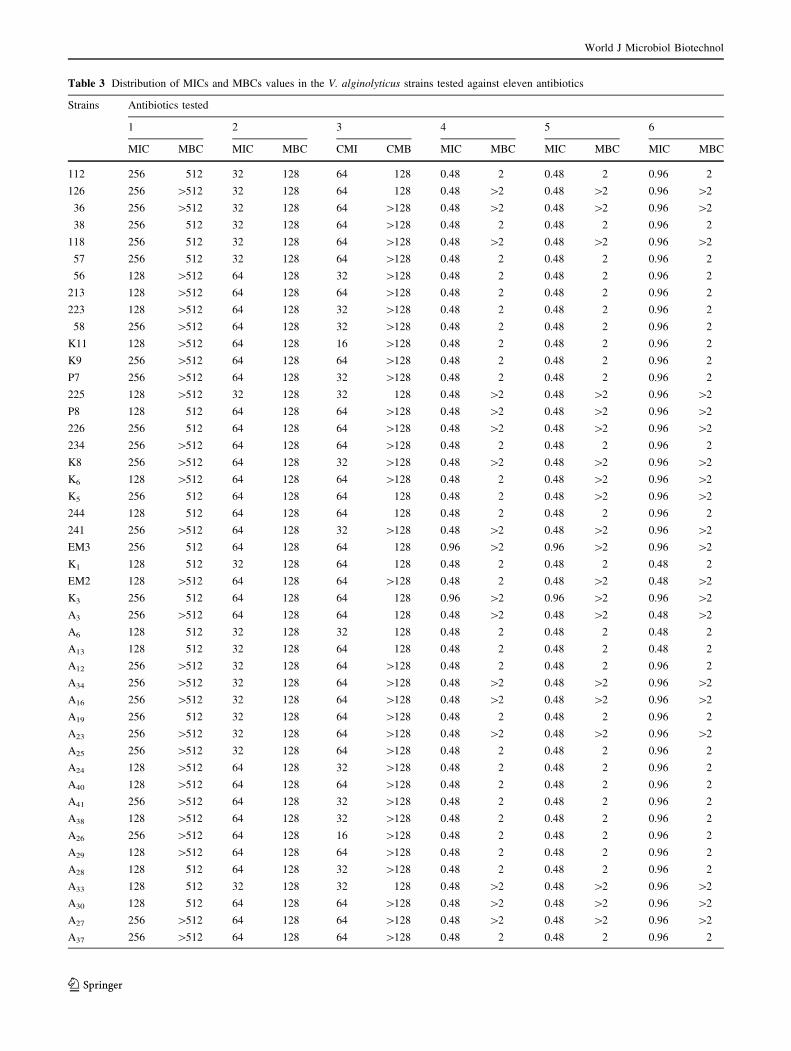
Table 3 Distribution of MICs and MBCs values in the V. alginolyticus strains tested against eleven antibiotics
Strains Antibiotics tested
1 2 3 4 5 6
MIC MBC MIC MBC CMI CMB MIC MBC MIC MBC MIC MBC
112 256 512 32 128 64 128 0.48 2 0.48 2 0.96 2
126 256 [512 32 128 64 128 0.48 [2 0.48 [2 0.96 [2
36 256 [512 32 128 64 [128 0.48 [2 0.48 [2 0.96 [2
38 256 512 32 128 64 [128 0.48 2 0.48 2 0.96 2
118 256 512 32 128 64 [128 0.48 [2 0.48 [2 0.96 [2
57 256 512 32 128 64 [128 0.48 2 0.48 2 0.96 2
56 128 [512 64 128 32 [128 0.48 2 0.48 2 0.96 2
213 128 [512 64 128 64 [128 0.48 2 0.48 2 0.96 2
223 128 [512 64 128 32 [128 0.48 2 0.48 2 0.96 2
58 256 [512 64 128 32 [128 0.48 2 0.48 2 0.96 2
K11 128 [512 64 128 16 [128 0.48 2 0.48 2 0.96 2
K9 256 [512 64 128 64 [128 0.48 2 0.48 2 0.96 2
P7 256 [512 64 128 32 [128 0.48 2 0.48 2 0.96 2
225 128 [512 32 128 32 128 0.48 [2 0.48 [2 0.96 [2
P8 128 512 64 128 64 [128 0.48 [2 0.48 [2 0.96 [2
226 256 512 64 128 64 [128 0.48 [2 0.48 [2 0.96 [2
234 256 [512 64 128 64 [128 0.48 2 0.48 2 0.96 2
K8 256 [512 64 128 32 [128 0.48 [2 0.48 [2 0.96 [2
K6 128 [512 64 128 64 [128 0.48 2 0.48 [2 0.96 [2
K5 256 512 64 128 64 128 0.48 2 0.48 [2 0.96 [2
244 128 512 64 128 64 128 0.48 2 0.48 2 0.96 2
241 256 [512 64 128 32 [128 0.48 [2 0.48 [2 0.96 [2
EM3 256 512 64 128 64 128 0.96 [2 0.96 [2 0.96 [2
K1 128 512 32 128 64 128 0.48 2 0.48 2 0.48 2
EM2 128 [512 64 128 64 [128 0.48 2 0.48 [2 0.48 [2
K3 256 512 64 128 64 128 0.96 [2 0.96 [2 0.96 [2
A3 256 [512 64 128 64 128 0.48 [2 0.48 [2 0.48 [2
A6 128 512 32 128 32 128 0.48 2 0.48 2 0.48 2
A13 128 512 32 128 64 128 0.48 2 0.48 2 0.48 2
A12 256 [512 32 128 64 [128 0.48 2 0.48 2 0.96 2
A34 256 [512 32 128 64 [128 0.48 [2 0.48 [2 0.96 [2
A16 256 [512 32 128 64 [128 0.48 [2 0.48 [2 0.96 [2
A19 256 512 32 128 64 [128 0.48 2 0.48 2 0.96 2
A23 256 [512 32 128 64 [128 0.48 [2 0.48 [2 0.96 [2
A25 256 [512 32 128 64 [128 0.48 2 0.48 2 0.96 2
A24 128 [512 64 128 32 [128 0.48 2 0.48 2 0.96 2
A40 128 [512 64 128 64 [128 0.48 2 0.48 2 0.96 2
A41 256 [512 64 128 32 [128 0.48 2 0.48 2 0.96 2
A38 128 [512 64 128 32 [128 0.48 2 0.48 2 0.96 2
A26 256 [512 64 128 16 [128 0.48 2 0.48 2 0.96 2
A29 128 [512 64 128 64 [128 0.48 2 0.48 2 0.96 2
A28 128 512 64 128 32 [128 0.48 2 0.48 2 0.96 2
A33 128 512 32 128 32 128 0.48 [2 0.48 [2 0.96 [2
A30 128 512 64 128 64 [128 0.48 [2 0.48 [2 0.96 [2
A27 256 [512 64 128 64 [128 0.48 [2 0.48 [2 0.96 [2
A37 256 [512 64 128 64 [128 0.48 2 0.48 2 0.96 2
World J Microbiol Biotechnol
123
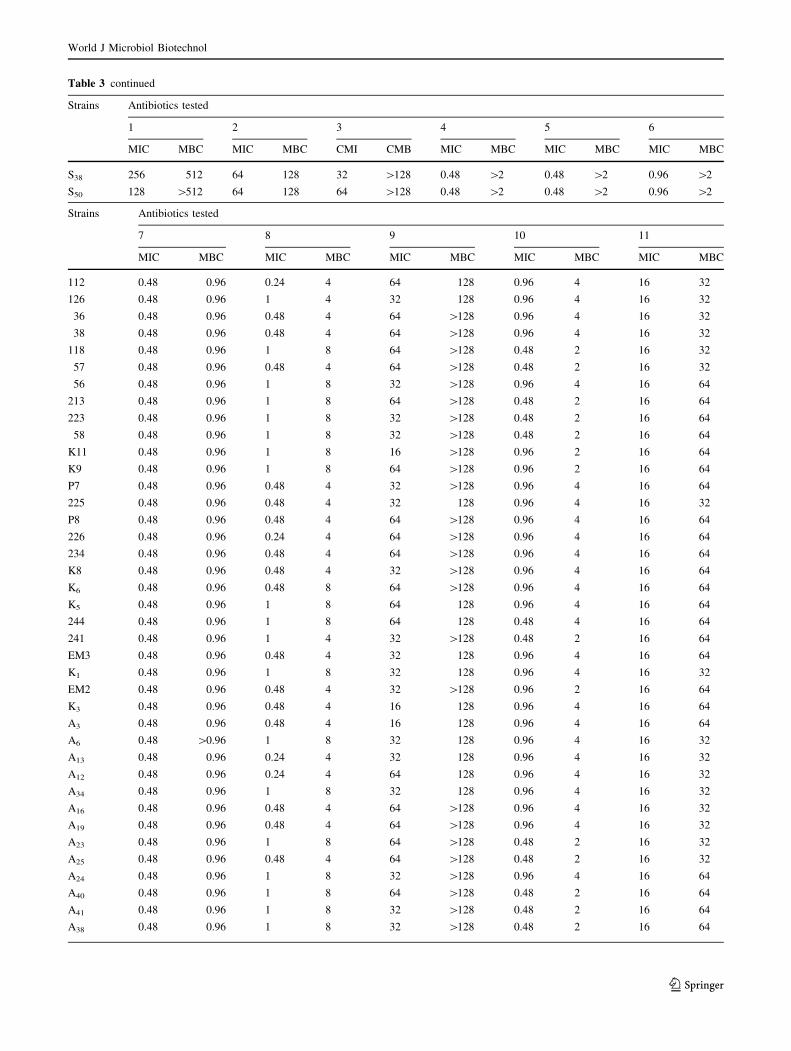
Table 3 continued
Strains Antibiotics tested
1 2 3 4 5 6
MIC MBC MIC MBC CMI CMB MIC MBC MIC MBC MIC MBC
S38 256 512 64 128 32 [128 0.48 [2 0.48 [2 0.96 [2
S50 128 [512 64 128 64 [128 0.48 [2 0.48 [2 0.96 [2
Strains Antibiotics tested
7 8 9 10 11
MIC MBC MIC MBC MIC MBC MIC MBC MIC MBC
112 0.48 0.96 0.24 4 64 128 0.96 4 16 32
126 0.48 0.96 1 4 32 128 0.96 4 16 32
36 0.48 0.96 0.48 4 64 [128 0.96 4 16 32
38 0.48 0.96 0.48 4 64 [128 0.96 4 16 32
118 0.48 0.96 1 8 64 [128 0.48 2 16 32
57 0.48 0.96 0.48 4 64 [128 0.48 2 16 32
56 0.48 0.96 1 8 32 [128 0.96 4 16 64
213 0.48 0.96 1 8 64 [128 0.48 2 16 64
223 0.48 0.96 1 8 32 [128 0.48 2 16 64
58 0.48 0.96 1 8 32 [128 0.48 2 16 64
K11 0.48 0.96 1 8 16 [128 0.96 2 16 64
K9 0.48 0.96 1 8 64 [128 0.96 2 16 64
P7 0.48 0.96 0.48 4 32 [128 0.96 4 16 64
225 0.48 0.96 0.48 4 32 128 0.96 4 16 32
P8 0.48 0.96 0.48 4 64 [128 0.96 4 16 64
226 0.48 0.96 0.24 4 64 [128 0.96 4 16 64
234 0.48 0.96 0.48 4 64 [128 0.96 4 16 64
K8 0.48 0.96 0.48 4 32 [128 0.96 4 16 64
K6 0.48 0.96 0.48 8 64 [128 0.96 4 16 64
K5 0.48 0.96 1 8 64 128 0.96 4 16 64
244 0.48 0.96 1 8 64 128 0.48 4 16 64
241 0.48 0.96 1 4 32 [128 0.48 2 16 64
EM3 0.48 0.96 0.48 4 32 128 0.96 4 16 64
K1 0.48 0.96 1 8 32 128 0.96 4 16 32
EM2 0.48 0.96 0.48 4 32 [128 0.96 2 16 64
K3 0.48 0.96 0.48 4 16 128 0.96 4 16 64
A3 0.48 0.96 0.48 4 16 128 0.96 4 16 64
A6 0.48 [0.96 1 8 32 128 0.96 4 16 32
A13 0.48 0.96 0.24 4 32 128 0.96 4 16 32
A12 0.48 0.96 0.24 4 64 128 0.96 4 16 32
A34 0.48 0.96 1 8 32 128 0.96 4 16 32
A16 0.48 0.96 0.48 4 64 [128 0.96 4 16 32
A19 0.48 0.96 0.48 4 64 [128 0.96 4 16 32
A23 0.48 0.96 1 8 64 [128 0.48 2 16 32
A25 0.48 0.96 0.48 4 64 [128 0.48 2 16 32
A24 0.48 0.96 1 8 32 [128 0.96 4 16 64
A40 0.48 0.96 1 8 64 [128 0.48 2 16 64
A41 0.48 0.96 1 8 32 [128 0.48 2 16 64
A38 0.48 0.96 1 8 32 [128 0.48 2 16 64
World J Microbiol Biotechnol
123
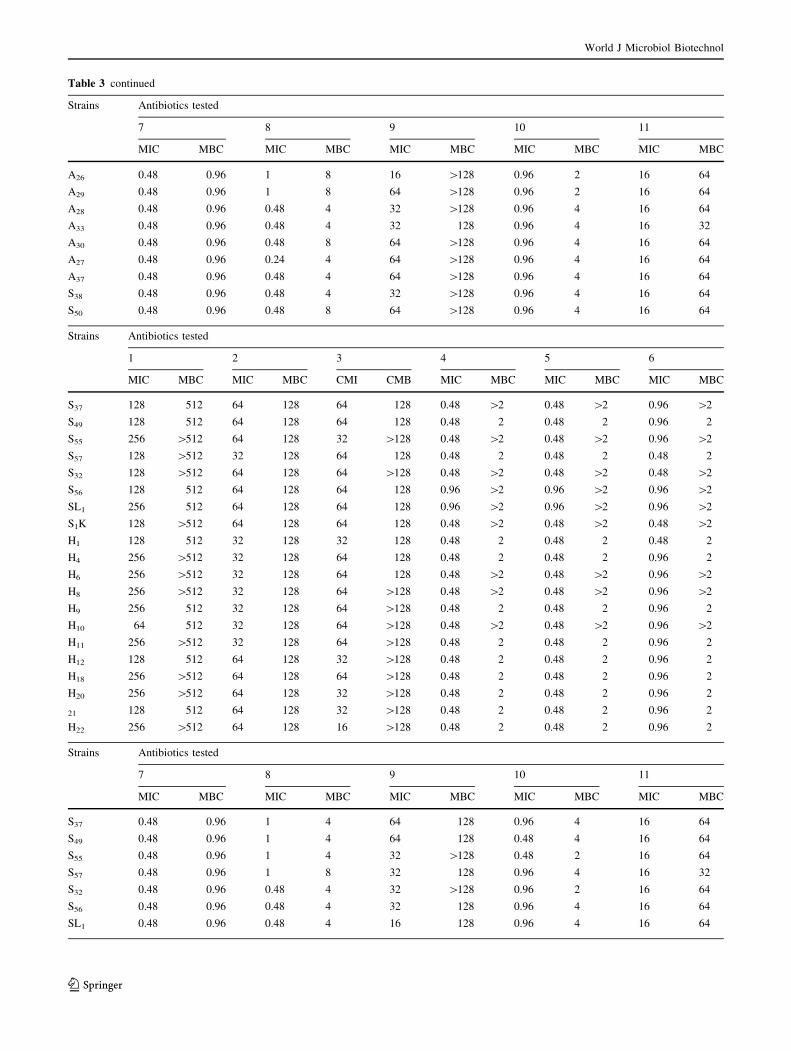
Table 3 continued
Strains Antibiotics tested
7 8 9 10 11
MIC MBC MIC MBC MIC MBC MIC MBC MIC MBC
A26 0.48 0.96 1 8 16 [128 0.96 2 16 64
A29 0.48 0.96 1 8 64 [128 0.96 2 16 64
A28 0.48 0.96 0.48 4 32 [128 0.96 4 16 64
A33 0.48 0.96 0.48 4 32 128 0.96 4 16 32
A30 0.48 0.96 0.48 8 64 [128 0.96 4 16 64
A27 0.48 0.96 0.24 4 64 [128 0.96 4 16 64
A37 0.48 0.96 0.48 4 64 [128 0.96 4 16 64
S38 0.48 0.96 0.48 4 32 [128 0.96 4 16 64
S50 0.48 0.96 0.48 8 64 [128 0.96 4 16 64
Strains Antibiotics tested
1 2 3 4 5 6
MIC MBC MIC MBC CMI CMB MIC MBC MIC MBC MIC MBC
S37 128 512 64 128 64 128 0.48 [2 0.48 [2 0.96 [2
S49 128 512 64 128 64 128 0.48 2 0.48 2 0.96 2
S55 256 [512 64 128 32 [128 0.48 [2 0.48 [2 0.96 [2
S57 128 [512 32 128 64 128 0.48 2 0.48 2 0.48 2
S32 128 [512 64 128 64 [128 0.48 [2 0.48 [2 0.48 [2
S56 128 512 64 128 64 128 0.96 [2 0.96 [2 0.96 [2
SL1 256 512 64 128 64 128 0.96 [2 0.96 [2 0.96 [2
S1K 128 [512 64 128 64 128 0.48 [2 0.48 [2 0.48 [2
H1 128 512 32 128 32 128 0.48 2 0.48 2 0.48 2
H4 256 [512 32 128 64 128 0.48 2 0.48 2 0.96 2
H6 256 [512 32 128 64 128 0.48 [2 0.48 [2 0.96 [2
H8 256 [512 32 128 64 [128 0.48 [2 0.48 [2 0.96 [2
H9 256 512 32 128 64 [128 0.48 2 0.48 2 0.96 2
H10 64 512 32 128 64 [128 0.48 [2 0.48 [2 0.96 [2
H11 256 [512 32 128 64 [128 0.48 2 0.48 2 0.96 2
H12 128 512 64 128 32 [128 0.48 2 0.48 2 0.96 2
H18 256 [512 64 128 64 [128 0.48 2 0.48 2 0.96 2
H20 256 [512 64 128 32 [128 0.48 2 0.48 2 0.96 2
21 128 512 64 128 32 [128 0.48 2 0.48 2 0.96 2
H22 256 [512 64 128 16 [128 0.48 2 0.48 2 0.96 2
Strains Antibiotics tested
7 8 9 10 11
MIC MBC MIC MBC MIC MBC MIC MBC MIC MBC
S37 0.48 0.96 1 4 64 128 0.96 4 16 64
S49 0.48 0.96 1 4 64 128 0.48 4 16 64
S55 0.48 0.96 1 4 32 [128 0.48 2 16 64
S57 0.48 0.96 1 8 32 128 0.96 4 16 32
S32 0.48 0.96 0.48 4 32 [128 0.96 2 16 64
S56 0.48 0.96 0.48 4 32 128 0.96 4 16 64
SL1 0.48 0.96 0.48 4 16 128 0.96 4 16 64
World J Microbiol Biotechnol
123
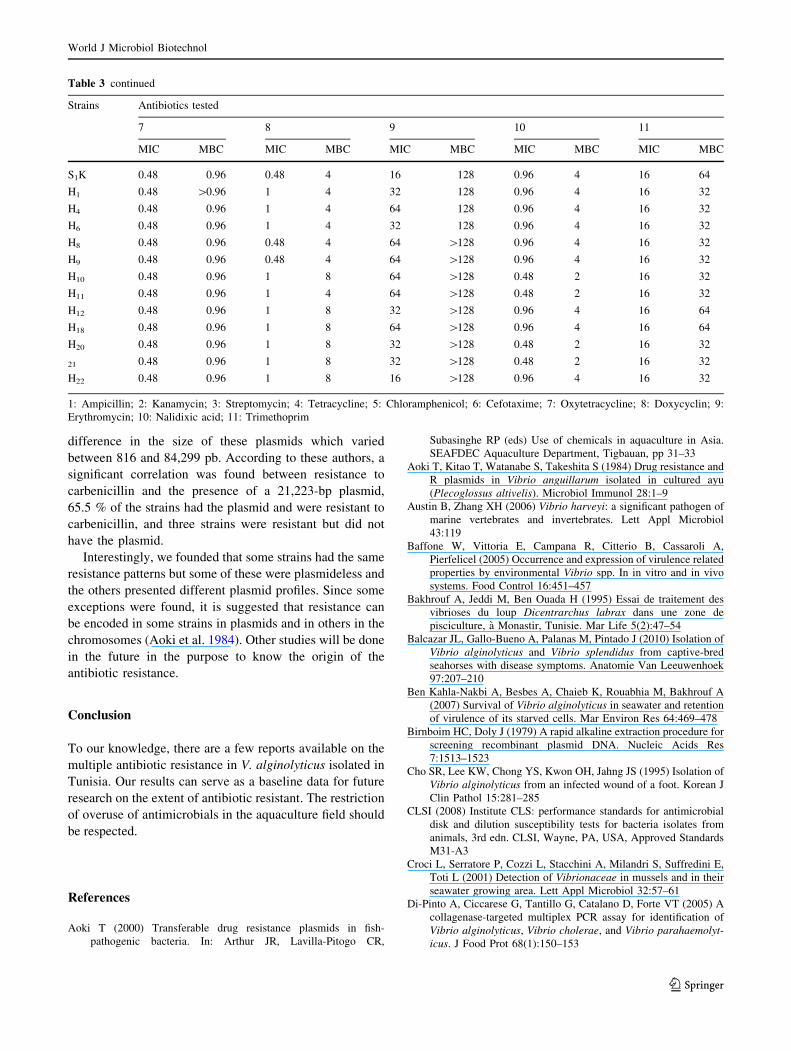
difference in the size of these plasmids which varied
between 816 and 84,299 pb. According to these authors, a
significant correlation was found between resistance to
carbenicillin and the presence of a 21,223-bp plasmid,
65.5 % of the strains had the plasmid and were resistant to
carbenicillin, and three strains were resistant but did not
have the plasmid.
Interestingly, we founded that some strains had the same
resistance patterns but some of these were plasmideless and
the others presented different plasmid profiles. Since some
exceptions were found, it is suggested that resistance can
be encoded in some strains in plasmids and in others in the
chromosomes (Aoki et al. 1984). Other studies will be done
in the future in the purpose to know the origin of the
antibiotic resistance.
Conclusion
To our knowledge, there are a few reports available on the
multiple antibiotic resistance in V. alginolyticus isolated in
Tunisia. Our results can serve as a baseline data for future
research on the extent of antibiotic resistant. The restriction
of overuse of antimicrobials in the aquaculture field should
be respected.
References
Aoki T (2000) Transferable drug resistance plasmids in fish-
pathogenic bacteria. In: Arthur JR, Lavilla-Pitogo CR,
Subasinghe RP (eds) Use of chemicals in aquaculture in Asia.
SEAFDEC Aquaculture Department, Tigbauan, pp 31–33
Aoki T, Kitao T, Watanabe S, Takeshita S (1984) Drug resistance and
R plasmids in Vibrio anguillarum isolated in cultured ayu
(Plecoglossus altivelis). Microbiol Immunol 28:1–9
Austin B, Zhang XH (2006) Vibrio harveyi: a significant pathogen of
marine vertebrates and invertebrates. Lett Appl Microbiol
43:119
Baffone W, Vittoria E, Campana R, Citterio B, Cassaroli A,
Pierfelicel (2005) Occurrence and expression of virulence related
properties by environmental Vibrio spp. In in vitro and in vivo
systems. Food Control 16:451–457
Bakhrouf A, Jeddi M, Ben Ouada H (1995) Essai de traitement des
vibrioses du loup Dicentrarchus labrax dans une zone de
pisciculture, a Monastir, Tunisie. Mar Life 5(2):47–54
Balcazar JL, Gallo-Bueno A, Palanas M, Pintado J (2010) Isolation of
Vibrio alginolyticus and Vibrio splendidus from captive-bred
seahorses with disease symptoms. Anatomie Van Leeuwenhoek
97:207–210
Ben Kahla-Nakbi A, Besbes A, Chaieb K, Rouabhia M, Bakhrouf A
(2007) Survival of Vibrio alginolyticus in seawater and retention
of virulence of its starved cells. Mar Environ Res 64:469–478
Birnboim HC, Doly J (1979) A rapid alkaline extraction procedure for
screening recombinant plasmid DNA. Nucleic Acids Res
7:1513–1523
Cho SR, Lee KW, Chong YS, Kwon OH, Jahng JS (1995) Isolation of
Vibrio alginolyticus from an infected wound of a foot. Korean J
Clin Pathol 15:281–285
CLSI (2008) Institute CLS: performance standards for antimicrobial
disk and dilution susceptibility tests for bacteria isolates from
animals, 3rd edn. CLSI, Wayne, PA, USA, Approved Standards
M31-A3
Croci L, Serratore P, Cozzi L, Stacchini A, Milandri S, Suffredini E,
Toti L (2001) Detection of Vibrionaceae in mussels and in their
seawater growing area. Lett Appl Microbiol 32:57–61
Di-Pinto A, Ciccarese G, Tantillo G, Catalano D, Forte VT (2005) A
collagenase-targeted multiplex PCR assay for identification of
Vibrio alginolyticus, Vibrio cholerae, and Vibrio parahaemolyt-icus. J Food Prot 68(1):150–153
Table 3 continued
Strains Antibiotics tested
7 8 9 10 11
MIC MBC MIC MBC MIC MBC MIC MBC MIC MBC
S1K 0.48 0.96 0.48 4 16 128 0.96 4 16 64
H1 0.48 [0.96 1 4 32 128 0.96 4 16 32
H4 0.48 0.96 1 4 64 128 0.96 4 16 32
H6 0.48 0.96 1 4 32 128 0.96 4 16 32
H8 0.48 0.96 0.48 4 64 [128 0.96 4 16 32
H9 0.48 0.96 0.48 4 64 [128 0.96 4 16 32
H10 0.48 0.96 1 8 64 [128 0.48 2 16 32
H11 0.48 0.96 1 4 64 [128 0.48 2 16 32
H12 0.48 0.96 1 8 32 [128 0.96 4 16 64
H18 0.48 0.96 1 8 64 [128 0.96 4 16 64
H20 0.48 0.96 1 8 32 [128 0.48 2 16 32
21 0.48 0.96 1 8 32 [128 0.48 2 16 32
H22 0.48 0.96 1 8 16 [128 0.96 4 16 32
1: Ampicillin; 2: Kanamycin; 3: Streptomycin; 4: Tetracycline; 5: Chloramphenicol; 6: Cefotaxime; 7: Oxytetracycline; 8: Doxycyclin; 9:
Erythromycin; 10: Nalidixic acid; 11: Trimethoprim
World J Microbiol Biotechnol
123

Doh YJ, Kim MH, Kim ES (1997) A case of Vibrio alginolyticusisolated from otorrhea of chronic otitis media. Korean J Infect
Dis 29:153–157
Ferrini AM, Mannoni V, Suffredini E, Cozzi L, Croci L (2008)
Evaluation of antibacterial resistance in Vibrio strains isolated
from imported seafood and Italian aquaculture settings. Food
Anal Methods 1:164–170
Fluharty DM, Packard WL (1967) Differentiation of gram-positive
and gram-negative bacteria without staining. Am J Vet Clin
Pathol 1:31–35
Gomez-Leon J, Villamil L, Lemos ML, Novoa B, Figueras A (2005)
Isolation of Vibrio alginolyticus and Vibrio splendidus from
aquacultured carpet shell clam (Ruditapes decussatus) larvae
associated with mass mortalities. Appl Environ Microbiol 71:98–104
Harvell CD, Mitchell CE, Ward JR, Altizer S, Dobson AP, Ostfeld
RS, Samuel MD (2002) Ecology-climate warming and disease
risks for terrestrial and marine biota. Science 296:2158–2162
Hidalgo RB, Cleenwerck I, Balboa S, De wachter M, Thompson FL,
Swings J, De Vos P, Romalde JL (2008) Diversity of Vibrios
associated with reared clams in Galicia (NW Spain). Syst Appl
Microbiol 31:215–222
Kim JS, Park SY, Kil YC, Lee HJ, Suh JT (2000) A case of
simultaneous isolation of Vibrio parahemolyticus and Vibrioalginolyticus. Korean J Clin Microbiol 3:147–152
Lagana P, Caruso G, Minutoli E, Zaccone R, Delia S (2011)
Susceptibility to antibiotics of Vibrio spp. and Photobacteriumdamsela spp. piscicida strains isolated from Italian aquaculture
farms. New Microbiol 34:53–63
Lee HJ, Choi JW (1995) A case of Vibrio alginolyticus myringitis.
Korean J Clin Pathol 15:101–105
Lee KK, Yu SR, Yang TL, Liu PC, Chen FR (1996) Isolation and
characterization of Vibrio alginolyticus isolated from diseased
kuruma prawn, Penaeus japonicus. Lett Appl Microbiol
22:111–114
Lee DY, Moon SY, Lee SO, Yang HY, Lee HJ, Lee MS (2008) Septic
shock due to Vibrio alginolyticus in a cirrhotic patient: the first
case in Korea. Yonsei Med J 49(2):329–332
Levy SB, Marshall B (2004) Antimicrobial resistance worldwide:
causes, challenges and responses. Nat Med 10:S122
Manjusha S, Sarita GB, Elyass KK, Chandrasekaran M (2005)
Multiple antibiotic resistance of Vibrio isolates from Coastal and
Brackish Water Areas. Am J Biochem Biotechnol 1(4):201–206
Molina-Aja A, Garcia-Gasca A, Grobois-Abreu A, Bolan-Mejia C,
Roque A, Gomez-Gil B (2002) Plasmid profiling and antibiotic
resistance of Vibrio strains isolated from cultured penaeid
shrimp. FEMS Microbiol Lett 213:7–12
Morris JG, Tenney J (1985) Antibiotic therapy for Vibrio vulnificusinfection. JAMA 253:1121–1122
Mukherji A, Schroeder S, Deyling C, Procop GW (2000) An unusual
source of Vibrio alginolyticus-associated otitis: prolonged col-
onization or freshwater exposure. Arch Otolaryngol Head Neck
Surg 126:790–791
Nakayama T, Ito E, Nomura N (2006) Comparison of Vibrio harveyistrains isolated from shrimp farms and from culture collection in
terms of toxicity and antibiotic resistance. FEMS Microbiol Lett
258:194
Oliver JD (2006) Vibrio vulnificus. In: Thompson FL, Austin B,
Swings J (eds) The biology of vibrios. ASM Press, Washington,
DC, pp 349–366
Ottaviani D, Bacchiocchi I, Masini L, Leoni F, Carraturo A,
Giammarioli M, Giovanni S (2001) Antimicrobial susceptibility
of potentially pathogenic halophilic vibrios isolated from
seafood. Int J Antimicrob Agents 18:135–140
Pedersen K (1999) The fish pathogen Vibrio anguillarum. DoctoralThesis. The Royal Veterinary and Agricultural University,
Denmark
Pedersen K, Tiainen T, Larsen JL (1996) Plasmid profiles restriction
fragment length polymorphism and O-serotypes among Vibrioanguillarum isolates from cultured Sparus sarba. Mar Poll Bull
39:245–249
Radu S, Elhadi N, Hassen Z, Rusul G, Lihan S, Fifadara N, Yuherman
PurwatiE (1998) Characterization of Vibrio vulnificus isolated
from cockles (Anadara gramosa): antimicrobial resistance
plasmid profiles and random amplification of polymorphic
DNA analysis. FEMS Microbiol Lett 165:139–143
Roque A, Molina-Aja A, Bolan-Mejia C, Gomez-Gil B (2001)
In vitro susceptibility to 15 antibiotics of vibrios isolated from
penaeid shrimps in Northwestern Mexico. Int J Antimicrob
Agents 17:383–387
Saitanu K, Chongthaleong A, Endo M, Umeda T, Takami K, Aoki T,
Kitao T (1994) Antimicrobial susceptibilities and detection of
transferable R-plasmids from Aeromonas hydrophila in Thai-
land. Asian Fish Sci 7:41–46
Sambrook PL, Peterson BC, Gerding DN, Cleary PP (1989)
Molecular cloning: a laboratory manual, 2nd edn. Cold Spring
Harbor Laboratory, NY
Sarter S, Nguyen HNK, Hung LT, Lazard J, Montet D (2007)
Antibiotic resistance in gram-negative bacteria isolated from
farmed catfish. Food Control 18:1391–1396
Scholtfeldt HJ (1992) Current practices of chemotherapy in fish
culture. In: Michel C, Alderman DJ (eds) Chemotherapy in
aquaculture: from theory to reality. Office International des
Epizooties, Paris, pp 25–38
Snoussi M, Chaieb K, Rouabhia M, Bakhrouf A (2006) Quantitative
study, identification and antibiotics sensitivity of some Vibrion-aceae associated to a marine fish hatchery. Ann Microbiol
56(4):289–293
Snoussi M, Hajlaoui H, Noumi E, Zanetti S, Bakhrouf A (2008)
Phenotypic and molecular characterization of Vibrio alginolyt-icus strains recovered from juveniles and older Sparus auratareared in a Tunisian marine farm. Ann Microbiol 58:141–146
Snoussi M, Noumi E, Lajnef R, Bellila A, Yazidi N, Bakhrouf A
(2011) Phenotypic characterization and enterobacterial repetitive
intergenic consensus PCR of Aeromonas spp. and Vibrio spp.
strains isolated from Tunisian sea bream (Sparus aurata) fish
farm. Afr J Microbiol Res 5(19):2920–2928
Son R, Rusul G, Sahilah AM, Zainuri A, Raha AR, Salmah I (1997)
Antibiotic resistance and plasmid profile of Aeromonas hydro-phila isolates from cultured fish, Tilapia (Tilapia mossambica).
Lett Appl Microbiol 24:479–482
Sorum H, Hvaal AB, Heum M, Daak FL, Wiik R (1990) Plasmid
profiling of Vibrio salmonicida for epidemiological studies of
cold water vibriosis in Atlantic salmon (Salmo salar) and cod
(Gaihis morua). Appl Environ Microbiol 56:1033–1037
Tang HJ, Chang MC, Ko WC, Huang KY, Lee CL, Chuang YC
(2002) In vitro and in vivo activities of newer fluoroquinolones
against Vibrio vulnificus. Antimicrob Agents Chemother
46:3580–3584
Tianen T, Pedersen K, Larsen JL (1995) Ribotyping and plasmid
profiling of Vibrio anguillarum serovar O2 and Vibrio ordalii. J
Appl Bacteriol 79:384–392
Thompson FL, Iida T, Swings J (2004) Biodiversity of Vibrios.
Microbiol Mol Biol Rev 68:3403–3431
Towner KJ (1995) Mechanisms of acquired resistance. In: Greenwood
D (ed) Antimicrobial chemotherapy. Oxford University Press,
Oxford
Vaseeharan B, Ramasamy P, Murugan T, Chen JC (2005) In vitro
susceptibility of antibiotics against Vibrio spp. and Aeromonasspp. isolated from Penaeus monodon hatcheries and ponds. Int J
Antimicrob Agents 26:285–291
Wang H, Dzink-Fox JL, Chen M (2001) Genetic characterization of
highly fluoroquine-resistant clinical Esherichia coli strains from
World J Microbiol Biotechnol
123

China: role of acr R mutations. Antimicrob Agents Chemother
45:1515
Wang Y, Leung PC, Qian PY (2006) Antibiotic resistance and
plasmid profile of environmental isolates of Vibrio species from
Mai po Nature Reserve, HongKong. Ecotoxicology 15:371
Wei R, Ge F, Huang S, Chen M, Wanga R (2011) Occurrence of
veterinary antibiotics in animal waste-water and surface water
around farms in Jiangsu Province, China. Chemosphere
82:1408–1414
Woo NYS, Kelly SP (1995) Effects of salinity and nutritional status
on growth and methabolism of Sparus sarba in a closed seawater
system. Aquaculture 135:229
Zanetti S, Deriu A, Volterra L, Falchi MP, Molicotti P, Fadda G,
Sechi L (2000) Virulence factors in Vibrio alginolyticus strains
isolated from aquatic environments. Ann Ig 12(6):487–491
Zanetti S, Spanu T, Deriu A, Romano L, Sechi LA, Fadda G (2001)
In vitro susceptibility of Vibrio spp. isolated from the environ-
ment. Int J Antimicrob Agents 17:407–409
Zorilla I, Morinigo MA, Castro D, Balebona MC, Borrego JJ (2003)
Intraspecific characterization of Vibrio alginolyticus isolates
recovered from cultured fish in Spain. J Appl Microbiol
95:1106–1116
Zulkifli Y, Alitheen NB, Son R, Raha AR, Samuel L, Yeap SK,
Nishibuchi M (2009) Random amplified polymorphic DNA-PCR
and ERIC PCR analysis on Vibrio parahaemolyticus isolated
from cockles in Padang, Indonesia. Int Food Res J 16:141–150
World J Microbiol Biotechnol
123


























![[Risk of hyperthyroidism in a Tunisian population of smokers]](https://img.dokumen.tips/doc/110x75/635a37e13c61073a160c93b6/risk-of-hyperthyroidism-in-a-tunisian-population-of-smokers.jpg)


