Embed Size (px)
Citation preview

RESEARCH ARTICLE
Chromatin insulators specifically associate with differentlevels of higher-order chromatin organization in Drosophila
Heather A. Wallace & Maria P. Plata &
Hyuck-Joon Kang & Misty Ross & Mariano Labrador
Received: 24 August 2009 /Revised: 20 October 2009 /Accepted: 30 October 2009 /Published online: 23 December 2009# Springer-Verlag 2009
Abstract Chromatin insulators are required for propertemporal and spatial expression of genes in metazoans.Here, we have analyzed the distribution of insulatorproteins on the 56F–58A region of chromosome 2R inDrosophila polytene chromosomes to assess the role ofchromatin insulators in shaping genome architecture. Datashow that the suppressor of Hairy-wing protein [Su(Hw)] isfound in three structures differentially associated withinsulator proteins: bands, interbands, and multi-genedomains of coexpressed genes. Results show that bands aregenerally formed by condensation of chromatin that belongsto genes containing one or more Su(Hw) binding sites,whereas, in interbands, Su(Hw) sites appear associated withopen chromatin. In addition, clusters of coexpressed genes inthis region form bands characterized by the lack of CP190and BEAF-32 insulator proteins. This pattern correlates withthe distribution of specific chromatin marks and is conservedin nurse cells, suggesting that this organization may not belimited to one cell type but represents the basic organizationof interphasic chromosomes.
Introduction
The ability of organisms to precisely regulate geneexpression is central to their development. Proper temporaland spatial expression of genes in higher eukaryotesrequires activation of transcription during the appropriatedevelopmental stages. Gene regulation is establishedthrough the activity of cis-regulatory elements includingproximal promoters, enhancers, repressors, and silencers(Markstein and Levine 2002; Ptashne and Gann 1997;Walhout 2006). However, these processes alone are notsufficient to explain the complexity of gene expressionregulation in a chromatin context given mounting evi-dence supporting that higher-order chromatin structuresand long-range interactions are an important aspect in theprocess of gene transcription regulation during develop-ment and cell differentiation in metazoans (Apostolou andThanos 2008; Osborne et al. 2004; Schuettengruber et al.2007; Spilianakis et al. 2005).
Chromatin insulators are regulatory elements found inDrosophila and in vertebrates that are considered to have amajor role in higher-order chromatin organization basedon their capacity to mediate long-range interactions withinthe chromatin fiber (Bartkuhn and Renkawitz 2008; Caiand Shen 2001; Kuhn-Parnell et al. 2008; Kyrchanova etal. 2008; Ling et al. 2006; Maksimenko et al. 2008;Muravyova et al. 2001; Splinter et al. 2006). Chromatininsulators are DNA sequences that have the ability toblock communication between enhancers and promoterswhen located between them and to prevent heterochroma-tin spreading (Chung et al. 1993; Gaszner and Felsenfeld2006; Gerasimova et al. 1995; Geyer and Corces 1992;Wallace and Felsenfeld 2007; Zhao et al. 1995). It isbelieved that these properties result from cis-interactionsbetween insulator proteins, which loop out the intervening
Communicated by R. Paro
Electronic supplementary material The online version of this article(doi:10.1007/s00412-009-0246-0) contains supplementary material,which is available to authorized users.
H. A. Wallace :M. P. Plata :H.-J. Kang :M. Ross :M. Labrador (*)Department of Biochemistry, Cellular and Molecular Biology,The University of Tennessee,M407 Walters Life Sciences, 1414 Cumberland Avenue,Knoxville, TN 37996, USAe-mail: [email protected]
Present Address:M. RossQuillen College of Medicine, East Tennessee State University,Johnson City, TN 37614, USA
Chromosoma (2010) 119:177–194DOI 10.1007/s00412-009-0246-0

DNA sequences to form functionally independent chro-matin domains (Capelson and Corces 2004; Gaszner andFelsenfeld 2006; Gerasimova and Corces 2001; Wallaceand Felsenfeld 2007).
In Drosophila, suppressor of Hairy-wing [Su(Hw)], Mod(mdg4), and centrosomal protein 190 (CP190) are proteinsassociated with the insulator activity initially identifiedwithin the gypsy retrovirus (Gerasimova and Corces 1998;Gerasimova and Corces 2001; Gerasimova et al. 1995;Geyer and Corces 1992; Ghosh et al. 2001; Pai et al. 2004;Spana et al. 1988). In addition to the gypsy insulator,however, these proteins are present in multiple locationsthroughout the genome. Similarly, the boundary elementassociated factor-32 (BEAF-32) and the Zw5 protein wereinitially identified as components of the scs–scs’ insulatorat the hsp70 locus, but are also found through the entiregenome (Gaszner et al. 1999; Kellum and Schedl 1991;Zhao et al. 1995). Finally, the insulator activity of theCCCTC-binding factor (CTCF) was initially described invertebrates but is also found in Drosophila, where it hasbeen shown to have insulator properties similar to that invertebrates, suggesting that insulator function and proper-ties are conserved across species (Bell and Felsenfeld 2000;Bell et al. 1999; Gerasimova et al. 2007; Moon et al. 2005).
The distribution of insulator protein binding sites in theDrosophila genome has been recently revealed by chromatinimmunoprecipitation on tiling microarrays (Adryan et al.2007; Bartkuhn et al. 2009; Bushey et al. 2009; Jiang et al.2009; Smith et al. 2009) and by the modENCODEconsortium. The analysis of these results suggests thatinsulator sites have the ability to define boundaries ofregulatory units through complex interactions. However,the precise role that insulator proteins play in chromosomeorganization, if any, remains largely speculative (Phillips andCorces 2009). It has been suggested, for example, that one ofthe roles for chromatin insulators is providing boundariesbetween different levels of chromatin organization, such asthe transitions between interbands and condensed chromatinin bands from polytene chromosomes (Gilbert et al. 2006;Labrador and Corces 2002; Pai et al. 2004; Spana et al.1988; Zhao et al. 1995). In addition, it has been hypothesizedthat the functional integrity of multi-gene chromatindomains, such as those described in Drosophila and otherorganisms (Boutanaev et al. 2002; Caron et al. 2001; Doruset al. 2006; Lee and Sonnhammer 2003; Miller et al. 2004;Roy et al. 2002; Versteeg et al. 2003; Yi et al. 2007), isprotected by chromatin insulators to facilitate the coordinateexpression of genes without interference from regulatorysequences adjacent to the domains (Capelson and Corces2004; Kuhn and Geyer 2003; Labrador and Corces 2002;West et al. 2002).
Whereas modern techniques allow for detailed analysisof the local organization of the chromatin fiber, studies of
the role that higher-order chromatin organization plays ingene transcription regulation are hindered by our inabilityto directly visualize chromatin organization within theinterphase nucleus. Polytene chromosomes, however, pro-vide a unique opportunity to examine different levels ofchromatin organization during interphase directly under themicroscope. Polytene chromosomes form in cells that growin size without dividing and therefore remain permanentlyin interphase. In a large number of cell types in Drosophilaand other dipterans, multiple replication rounds without celldivision result in more than 1,000 DNA strands perchromosome. These strands remain aligned and attachedto each other forming the large structures that we know aspolytene chromosomes. This alignment of multiple chro-matin fibers amplifies the basic organization of thechromosome and allows differences in chromatin compac-tion to be seen as a series of bands and interbandsextending across the width of the chromosome arms. Inaddition, because transcription only takes place in the moredecondensed chromatin found in interbands, it is clear thatthese two levels of organization also translate into twodifferent levels of transcriptional activity and chromatinstructure (Kaplan et al. 2000; Labrador and Corces 2003;Lis 2007; Saunders et al. 2003; Yao et al. 2008).
Here, we take advantage of the recently describedgenome-wide distribution of insulator sites in Drosophilaand use this information to devise a series of experimentsassessing the role that insulator proteins have in theorganization of higher-order chromatin structures in inter-phase chromosomes. We have analyzed the distribution ofthe Drosophila insulator protein Su(Hw) along the 56F–58A region of chromosome 2R and have established acorrespondence between Su(Hw), CP190, BEAF-32, andthe CTCF insulator sites, transcriptional units, and theband–interband pattern observed in polytene chromosomes.Our results suggest that most Su(Hw) binding sites arelargely associated with compacted chromatin and that Su(Hw) binding sites associated with open chromatin ininterbands may have properties different from Su(Hw)binding sites found in condensed chromatin. Based on ourfindings, we propose a model in which different levels ofhigher-order chromatin organization mediated by insulatorsand other proteins alternate with each other to generatefunctional differences in chromatin compaction along thechromatin fiber, which are amplified to form the classicalternate sequence of bands and interbands in polytenechromosomes.
Materials and methods
Analysis of the distribution of Su(Hw) binding sites andgene clusters ChIP-on-chip data files for Su(Hw), CP190,
178 Chromosoma (2010) 119:177–194

BEAF-32, and CTCF (accession number GEO GSE16245)from embryos were downloaded from the modENCODEwebsite (http://www.modENCODE.org). Signal intensitieswere converted to normalized log2 ratios using TilingAnalysis Software (Affymetrix). BED files containing Su(Hw), CP190, BEAF-32, and CTCF peak data wereobtained from supplemental material included in Busheyet al. (2009). Peak data files were uploaded into IntegratedGenome Browser (http://igb.bioviz.org/) for visualizationalongside the April 2006 version of the Drosophilamelanogaster genome. Graph thresholding was set tovisualize peaks above the 95th percentile. The distributionof coexpressed genes was analyzed using Excel in whichclusters of coexpressed genes from Spellman and Rubin(2002) and merged with the annotated D. melanogastergenome project (release 5.5).
In situ hybridization and immunostaining of polytenechromosomes Approximately 500 bp DNA fragmentscorresponding to each endogenous insulator were obtainedby PCR (see primers and probe sizes in SupplementalTable 2). Biotin-labeled DNAwas prepared using the BiotinHigh-Prime random priming kit (Roche). The labeled probeDNA was ethanol-precipitated and resuspended in hybrid-ization buffer (4× SSC, 50% formamide, 1× Denhardts, and0.4 mg/ml of salmon sperm DNA). Polytene chromosomesobtained from salivary glands of third instar larvae weredissected in 0.7% NaCl and fixed in a 1:2:3 mixture of aceticacid/water/lactic acid. Polytene chromosomes from nurse cellswere obtained from the ovaries of 3 to 5-day-old otu7/otu11
females maintained in vials with males and in the presence ofdry yeast to stimulate oogenesis. Ovaries were dissected inphosphate-buffered saline (PBS), fixed, and squashed in a1:2:3 mixture of acetic acid/water/lactic acid. Slides wereheated at 65°C in 2× SSC for 30 min, dehydrated in anethanol series, and denatured in 0.07 M NaOH.
For hybridization of DNA, boiled probes were added to theslide, covered immediately with a coverslip, and incubated at37°C overnight in a humidified chamber. Following hybrid-ization, coverslips were removed, and the slides were washedin 2× SSC at 37°C, then at room temperature in 1× PBS, andfinally in antibody dilution buffer. The slides were incubatedovernight in dilution buffer containing a 1:300 dilution of Su(Hw) primary antibody. Slides were then washed in antibodydilution buffer and incubated with a 1:300 dilution ofFluorescein Isothiocyanate (FITC)- or Texas red-conjugatedgoat anti-rabbit IgG (Jackson Laboratories) for 2 h at roomtemperature. Slides were stained for 30 s with 4′,6-diamidino-2-phenylindole (DAPI; 0.5 μg/ml) and mounted in Vecta-shield Mounting Medium (Vector Laboratories). Slides wereanalyzed using a Leica DM6000B fluorescence microscope.Antibodies specific against Su(Hw) protein were raised usingan N-terminal peptide containing the first 218 amino acids of
the protein. Antibodies specific against CP190 protein wereraised using a C-terminal peptide containing amino acids 488to 1084. Antibodies specific against modifier of mdg4 protein(Mod(mdg4)67.2 protein) were raised using a C-terminalpeptide containing amino acids 458 to 610. Antibodies wereraised in rats and rabbits by Pocono Rabbit Farm andLaboratory (Canadensis, PA 18325, USA) and were validatedusing westerns and by co-immunostainings with previouslycharacterized rat or rabbit antibodies.
Immunostaining of polytene chromosomes Antibodies usedfor immunostaining were as follows: rabbit anti-Su(Hw) andanti-CP190 (1:300 dilution) rat anti-Mod(mdg4)67.2 (1:300dilution); mouse anti-H14 (RNA Pol II; 1:50 dilution) waspurchased from Covance (Princeton, New Jersey), mouseanti-heterochromatin protein 1 (HP1; 1:25 dilution). The HP1antibody was developed by L. L. Wallrath and was obtainedfrom the Developmental Studies Hybridoma Bank developedunder the auspices of the National Institute of Child Healthand Human Development and maintained by The Universityof Iowa, Department of Biological Sciences, Iowa City, IA;the following antibodies were purchased from Upstate(Millipore Corporation, Billerica, MA): rabbit anti-H3K4me3 (dilution1:25), rabbit anti-H3K9ac (dilution1:50), rabbit anti-H3K27me1 (dilution 1:50), rabbit anti-H3K4ac (dilution 1:50), and rabbit anti-H3K27me3 (dilution1:50); rabbit anti-Polycomb (dilution 1:50) was purchasedfrom Santa Cruz Biotechnology (Santa Cruz, CA.). Secondaryantibodies, FITC-conjugated goat anti-rabbit IgG, Texas redgoat anti-rabbit IgG, Texas red donkey anti-mouse IgM, andTexas red donkey anti-rat IgG were purchased from TheJackson Laboratory (Bar Harbor, ME).
Polytene chromosomes obtained from salivary glands ofthird instar larvae were dissected in 0.7% NaCl and fixedand squashed in fixative solution containing 3.7% formal-dehyde and 45% acetic acid. Primary antibodies werediluted in PBS containing 0.1% Igepal and 1% milk andincubated on the slides overnight at 4°C. Slides were thenwashed in PBS+0.1% Igepal and incubated with a 1:300dilution of the appropriate FITC- or Texas red-conjugatedsecondary antibody for 2 h at room temperature. Slideswere stained for 30 s with DAPI (0.5 μg/ml) and mountedin Vectashield mounting medium (Vector Laboratories;Burlingame, CA). Slides were analyzed using a LeicaDM6000B fluorescence microscope.
Chromatin immunoprecipitation Chromatin was preparedfrom 17-h embryos collected on grape juice agar media.Embryos were homogenized in buffer A1 (60 mM KCl,15 mM NaCl, 4 mM MgCl2, 15 mM HEPES (pH 7.6),0.5% Triton X-100, 0.5 mM DTT, 10 mM sodium butyrate,and 1× EDTA-free protease inhibitor cocktail (Roche)containing 1.8% formaldehyde). Crosslinking was stopped
Chromosoma (2010) 119:177–194 179

by adding 225 mM glycine solution. Cells were lysed, andchromatin was sheared to an average length of 500–700 bpby sonication. In each chromatin immunoprecipitation(ChIP) experiment, a chromatin solution corresponding to200 mg of live material was incubated with either Su(Hw)antibody or normal rabbit IgG. Immunoprecipitation andwashing were performed as described elsewhere (Cavalli etal. 1999). The same Su(Hw) antibody used in immunos-taining experiments was used in ChIP assays.
Real-time PCR quantification analysis of immunopreci-pitated DNA Real-time PCR quantification of immunopre-cipitated DNAwas carried out with ABGene (Rockford, IL)SYBR green PCR master mix. For input PCRs, a 1/100dilution was used as template. Primers were designed toamplify 100–200 bp fragments. PCR conditions for eachprimer pair were tested to determine the efficiency ofamplification and to ensure amplification was in the linearrange. PCR products for each primer pair were amplifiedfrom at least three separate immunoprecipitation productsfrom at least two different chromatin preparations using theBioRad iQ5 Multicolor Real-Time PCR Detection System(primers listed in Supplemental Table 3). Enrichment ofimmunoprecipitated DNA fragments was calculated usingthe ∆Ct method based on the threshold cycle (Ct) value foreach PCR reaction (BioRad real-time PCR applicationguide). Results are presented as percentage of total input.The statistical significance of the results was calculated byStudent’s t test.
Results
Only a fraction of the Su(Hw) binding sites identified inembryos correspond to major Su(Hw) bands visible inpolytene chromosomes We have used fluorescence in situhybridization (FISH) combined with immunostaining toperform an analysis of the distribution of the Su(Hw) proteinlocalized within a total of two contiguous Mb along the 56F–58A cytological region on polytene chromosome 2R. Thischromosome region has a specific morphology as well as aband–interband pattern that makes it easily identifiable inpolytene chromosome spreads. With the help of ChIP–chipdata on insulator site distribution in the Drosophila genomefrom the modENCODE consortium and Bushey et al. (2009),we have used 29 specific probes containing individual Su(Hw) binding sites from this region to perform FISHcombined with immunostaining (Supplementary Table 1 andSupplementary Figs. 1 and 2). Probes for in situ hybridizationwere designed as 500-bp sequences containing a singlebinding site identified based on ChIP–chip peak data. Weused these data with the intention of, first, directly mappingthe specific Su(Hw) binding sites in the chromosomes, in
relation to the band–interband pattern; second, to determinewhether the specific sites are associated with a Su(Hw)immunostaining signal; and finally, using the in situhybridized sites as landmarks to determine the relativeposition of other insulator sites and genes within the sameregion. One of the arguments against the suitability ofimmunostaining and in situ hybridization on polytenechromosomes is the low resolution of the technique.However, the level of resolution of this technique in ourexperiments is well below 15 kb, even in chromosomeregions in which chromatin appears as DAPI condensedbands (Supplementary Figs. 1 and 2). This high level ofresolution greatly facilitates the analysis of the distribution ofchromatin proteins directly on chromosomes as a comple-ment to analysis performed using ChIP.
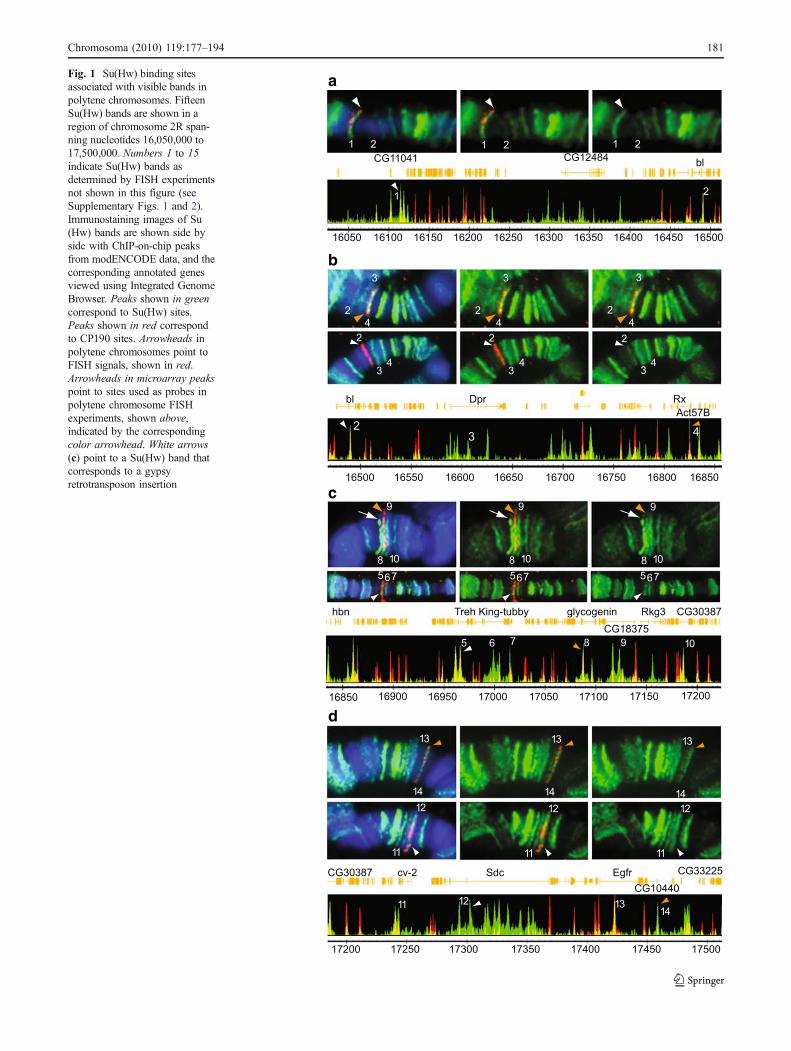
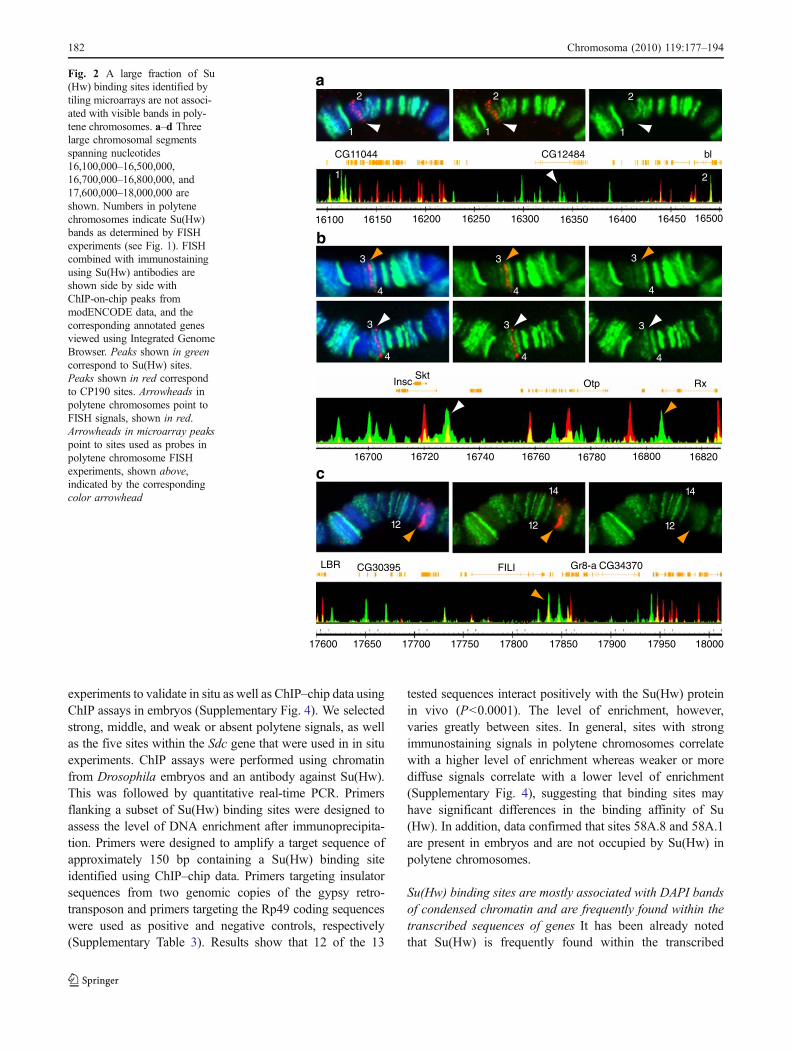
Results show that the 29 in situ hybridization probes used inthese experiments are sufficient to identify all the major Su(Hw) immunostaining signals found within the 56F–58Acytological region (Figs. 1 and 2). In general, there is anoticeable genome-wide variability in the intensity ofimmunostaining signals when antibodies against Su(Hw) areused to immunostain polytene chromosomes (Gerasimovaand Corces 1998; Gerasimova et al. 1995). The same level ofvariation is observed among the Su(Hw) immunostainingsignals that we have tested along the 56F–58A cytologicalregion. Su(Hw) signals can appear as low-, medium-, or high-intensity bands (Fig. 1). In addition, a large number of sitesshow very weak and diffuse or no signal (Fig. 2). Only 15 Su(Hw) bands are present within the 56F–58A cytologicalregion (Fig. 1), to which 18 of the probes localize, while theremaining 11 probes are located in regions with a weak orabsent Su(Hw) signal. Similarly, using ChIP–chip tiling arraydata from Bushey et al. (2009) and modENCODE consor-tium, we found that, out of the 78 binding sites mapped tothis region, only 45 are associated with the 15 immunostain-ing bands visible in polytene chromosomes, whereas 33 donot localize with any visible Su(Hw) band. Sites with animmunostaining signal that can be identified as a band arelisted in Table 1, and sites with no identifiable immunostain-ing signal are listed in Table 2.
In order to further understand the differences in signalintensity between sites observed as immunostaining signalsin salivary glands, we have compared the distribution ofimmunostaining signals in polytene chromosomes with thatof sites obtained from microarray data using the IntegratedGenome Browser (http://igb.bioviz.org/). Fluorescence isvery intense at some binding sites in polytene chromosomes(Fig. 1), whereas it is largely absent or very faint at others(Fig. 2). Strong bands correspond mostly to intragenic sites,whereas sites with no Su(Hw) immunostaining signal arefound mostly in intergenic regions (see Tables 1 and 2).
Since we observed differences between polytenechromosome results and ChIP–chip data, we performed
180 Chromosoma (2010) 119:177–194
a
b
c
d
Fig. 1 Su(Hw) binding sitesassociated with visible bands inpolytene chromosomes. FifteenSu(Hw) bands are shown in aregion of chromosome 2R span-ning nucleotides 16,050,000 to17,500,000. Numbers 1 to 15indicate Su(Hw) bands asdetermined by FISH experimentsnot shown in this figure (seeSupplementary Figs. 1 and 2).Immunostaining images of Su(Hw) bands are shown side byside with ChIP-on-chip peaksfrom modENCODE data, and thecorresponding annotated genesviewed using Integrated GenomeBrowser. Peaks shown in greencorrespond to Su(Hw) sites.Peaks shown in red correspondto CP190 sites. Arrowheads inpolytene chromosomes point toFISH signals, shown in red.Arrowheads in microarray peakspoint to sites used as probes inpolytene chromosome FISHexperiments, shown above,indicated by the correspondingcolor arrowhead. White arrows(c) point to a Su(Hw) band thatcorresponds to a gypsyretrotransposon insertion
Chromosoma (2010) 119:177–194 181
experiments to validate in situ as well as ChIP–chip data usingChIP assays in embryos (Supplementary Fig. 4). We selectedstrong, middle, and weak or absent polytene signals, as wellas the five sites within the Sdc gene that were used in in situexperiments. ChIP assays were performed using chromatinfrom Drosophila embryos and an antibody against Su(Hw).This was followed by quantitative real-time PCR. Primersflanking a subset of Su(Hw) binding sites were designed toassess the level of DNA enrichment after immunoprecipita-tion. Primers were designed to amplify a target sequence ofapproximately 150 bp containing a Su(Hw) binding siteidentified using ChIP–chip data. Primers targeting insulatorsequences from two genomic copies of the gypsy retro-transposon and primers targeting the Rp49 coding sequenceswere used as positive and negative controls, respectively(Supplementary Table 3). Results show that 12 of the 13
tested sequences interact positively with the Su(Hw) proteinin vivo (P<0.0001). The level of enrichment, however,varies greatly between sites. In general, sites with strongimmunostaining signals in polytene chromosomes correlatewith a higher level of enrichment whereas weaker or morediffuse signals correlate with a lower level of enrichment(Supplementary Fig. 4), suggesting that binding sites mayhave significant differences in the binding affinity of Su(Hw). In addition, data confirmed that sites 58A.8 and 58A.1are present in embryos and are not occupied by Su(Hw) inpolytene chromosomes.
Su(Hw) binding sites are mostly associated with DAPI bandsof condensed chromatin and are frequently found within thetranscribed sequences of genes It has been already notedthat Su(Hw) is frequently found within the transcribed
16100 16150 16200 16250 16300 16350 16400 16450 16500
CG11044 blCG12484
1
1 2
2
1
2
1
2a
3
4
3
4
3
4
3
4
3
4
3
4
16700 16720 16740 16760 16780 16800 16820
InscSkt
Otp Rx
b
17600
17650 17700 17750 17800 17850 17900 17950 1800017600
LBR FILI CG34370Gr8-aCG30395
12
14
12
14
12
c
Fig. 2 A large fraction of Su(Hw) binding sites identified bytiling microarrays are not associ-ated with visible bands in poly-tene chromosomes. a–d Threelarge chromosomal segmentsspanning nucleotides16,100,000–16,500,000,16,700,000–16,800,000, and17,600,000–18,000,000 areshown. Numbers in polytenechromosomes indicate Su(Hw)bands as determined by FISHexperiments (see Fig. 1). FISHcombined with immunostainingusing Su(Hw) antibodies areshown side by side withChIP-on-chip peaks frommodENCODE data, and thecorresponding annotated genesviewed using Integrated GenomeBrowser. Peaks shown in greencorrespond to Su(Hw) sites.Peaks shown in red correspondto CP190 sites. Arrowheads inpolytene chromosomes point toFISH signals, shown in red.Arrowheads in microarray peakspoint to sites used as probes inpolytene chromosome FISHexperiments, shown above,indicated by the correspondingcolor arrowhead
182 Chromosoma (2010) 119:177–194
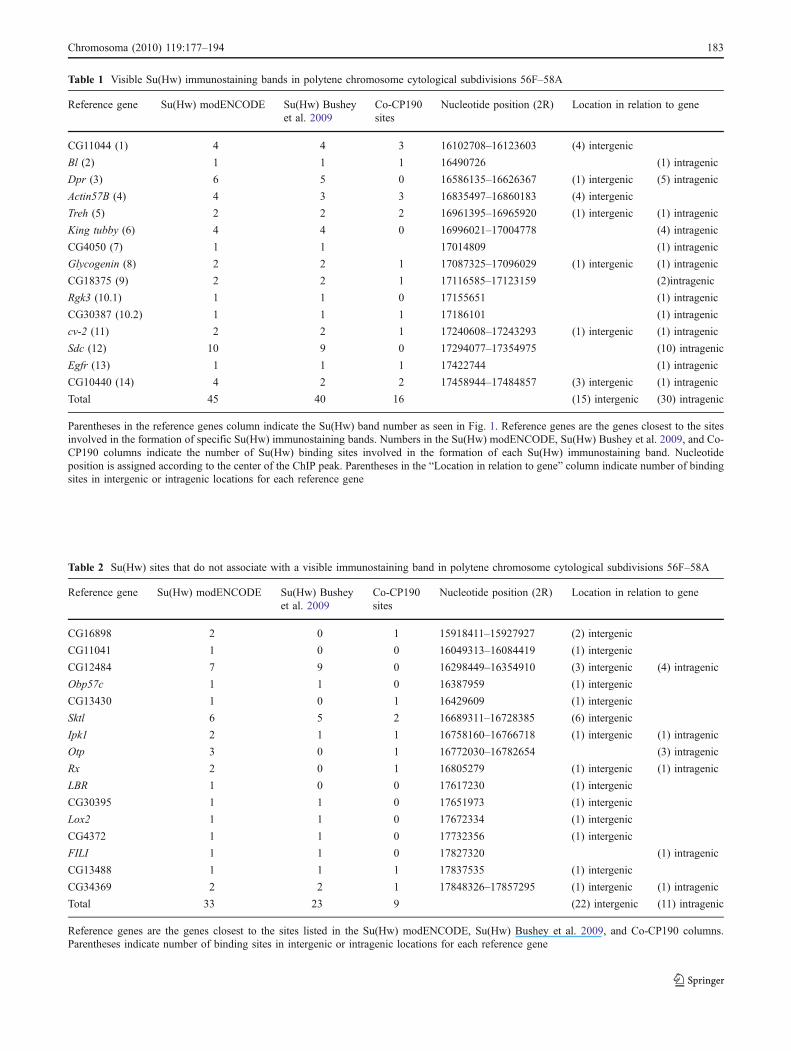
Table 1 Visible Su(Hw) immunostaining bands in polytene chromosome cytological subdivisions 56F–58A
Reference gene Su(Hw) modENCODE Su(Hw) Busheyet al. 2009
Co-CP190sites
Nucleotide position (2R) Location in relation to gene
CG11044 (1) 4 4 3 16102708–16123603 (4) intergenic
Bl (2) 1 1 1 16490726 (1) intragenic
Dpr (3) 6 5 0 16586135–16626367 (1) intergenic (5) intragenic
Actin57B (4) 4 3 3 16835497–16860183 (4) intergenic
Treh (5) 2 2 2 16961395–16965920 (1) intergenic (1) intragenic
King tubby (6) 4 4 0 16996021–17004778 (4) intragenic
CG4050 (7) 1 1 17014809 (1) intragenic
Glycogenin (8) 2 2 1 17087325–17096029 (1) intergenic (1) intragenic
CG18375 (9) 2 2 1 17116585–17123159 (2)intragenic
Rgk3 (10.1) 1 1 0 17155651 (1) intragenic
CG30387 (10.2) 1 1 1 17186101 (1) intragenic
cv-2 (11) 2 2 1 17240608–17243293 (1) intergenic (1) intragenic
Sdc (12) 10 9 0 17294077–17354975 (10) intragenic
Egfr (13) 1 1 1 17422744 (1) intragenic
CG10440 (14) 4 2 2 17458944–17484857 (3) intergenic (1) intragenic
Total 45 40 16 (15) intergenic (30) intragenic
Parentheses in the reference genes column indicate the Su(Hw) band number as seen in Fig. 1. Reference genes are the genes closest to the sitesinvolved in the formation of specific Su(Hw) immunostaining bands. Numbers in the Su(Hw) modENCODE, Su(Hw) Bushey et al. 2009, and Co-CP190 columns indicate the number of Su(Hw) binding sites involved in the formation of each Su(Hw) immunostaining band. Nucleotideposition is assigned according to the center of the ChIP peak. Parentheses in the “Location in relation to gene” column indicate number of bindingsites in intergenic or intragenic locations for each reference gene
Table 2 Su(Hw) sites that do not associate with a visible immunostaining band in polytene chromosome cytological subdivisions 56F–58A
Reference gene Su(Hw) modENCODE Su(Hw) Busheyet al. 2009
Co-CP190sites
Nucleotide position (2R) Location in relation to gene
CG16898 2 0 1 15918411–15927927 (2) intergenic
CG11041 1 0 0 16049313–16084419 (1) intergenic
CG12484 7 9 0 16298449–16354910 (3) intergenic (4) intragenic
Obp57c 1 1 0 16387959 (1) intergenic
CG13430 1 0 1 16429609 (1) intergenic
Sktl 6 5 2 16689311–16728385 (6) intergenic
Ipk1 2 1 1 16758160–16766718 (1) intergenic (1) intragenic
Otp 3 0 1 16772030–16782654 (3) intragenic
Rx 2 0 1 16805279 (1) intergenic (1) intragenic
LBR 1 0 0 17617230 (1) intergenic
CG30395 1 1 0 17651973 (1) intergenic
Lox2 1 1 0 17672334 (1) intergenic
CG4372 1 1 0 17732356 (1) intergenic
FILI 1 1 0 17827320 (1) intragenic
CG13488 1 1 1 17837535 (1) intergenic
CG34369 2 2 1 17848326–17857295 (1) intergenic (1) intragenic
Total 33 23 9 (22) intergenic (11) intragenic
Reference genes are the genes closest to the sites listed in the Su(Hw) modENCODE, Su(Hw) Bushey et al. 2009, and Co-CP190 columns.Parentheses indicate number of binding sites in intergenic or intragenic locations for each reference gene
Chromosoma (2010) 119:177–194 183
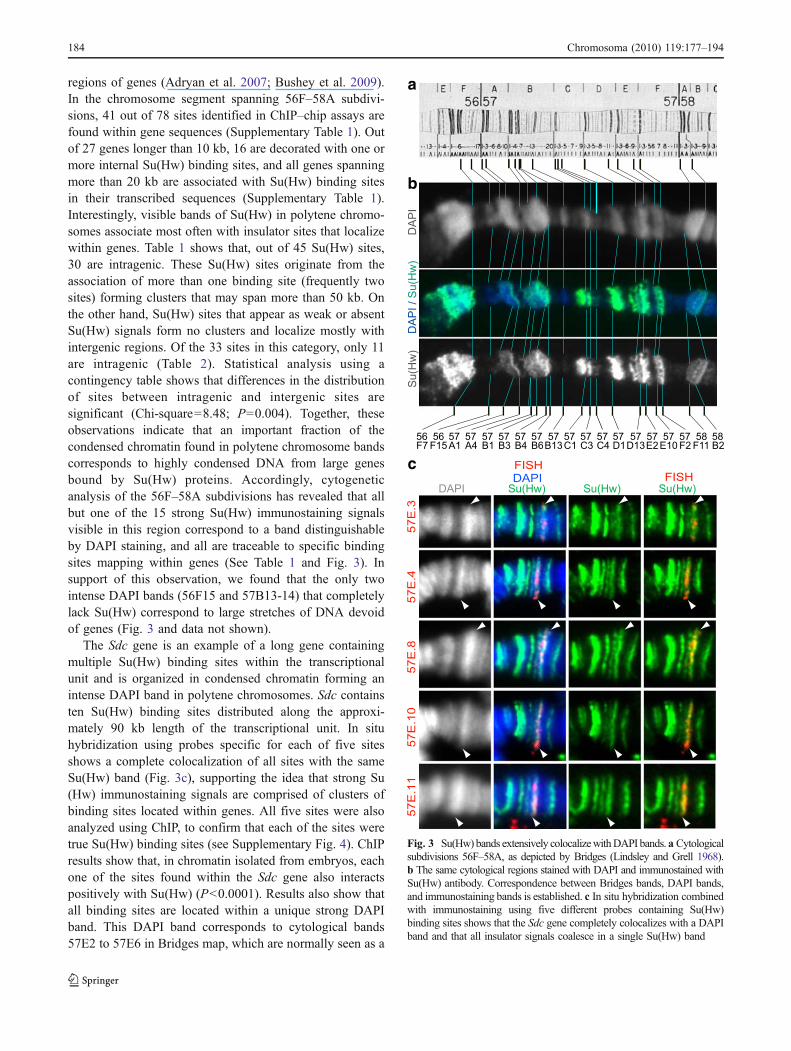
regions of genes (Adryan et al. 2007; Bushey et al. 2009).In the chromosome segment spanning 56F–58A subdivi-sions, 41 out of 78 sites identified in ChIP–chip assays arefound within gene sequences (Supplementary Table 1). Outof 27 genes longer than 10 kb, 16 are decorated with one ormore internal Su(Hw) binding sites, and all genes spanningmore than 20 kb are associated with Su(Hw) binding sitesin their transcribed sequences (Supplementary Table 1).Interestingly, visible bands of Su(Hw) in polytene chromo-somes associate most often with insulator sites that localizewithin genes. Table 1 shows that, out of 45 Su(Hw) sites,30 are intragenic. These Su(Hw) sites originate from theassociation of more than one binding site (frequently twosites) forming clusters that may span more than 50 kb. Onthe other hand, Su(Hw) sites that appear as weak or absentSu(Hw) signals form no clusters and localize mostly withintergenic regions. Of the 33 sites in this category, only 11are intragenic (Table 2). Statistical analysis using acontingency table shows that differences in the distributionof sites between intragenic and intergenic sites aresignificant (Chi-square=8.48; P=0.004). Together, theseobservations indicate that an important fraction of thecondensed chromatin found in polytene chromosome bandscorresponds to highly condensed DNA from large genesbound by Su(Hw) proteins. Accordingly, cytogeneticanalysis of the 56F–58A subdivisions has revealed that allbut one of the 15 strong Su(Hw) immunostaining signalsvisible in this region correspond to a band distinguishableby DAPI staining, and all are traceable to specific bindingsites mapping within genes (See Table 1 and Fig. 3). Insupport of this observation, we found that the only twointense DAPI bands (56F15 and 57B13-14) that completelylack Su(Hw) correspond to large stretches of DNA devoidof genes (Fig. 3 and data not shown).
The Sdc gene is an example of a long gene containingmultiple Su(Hw) binding sites within the transcriptionalunit and is organized in condensed chromatin forming anintense DAPI band in polytene chromosomes. Sdc containsten Su(Hw) binding sites distributed along the approxi-mately 90 kb length of the transcriptional unit. In situhybridization using probes specific for each of five sitesshows a complete colocalization of all sites with the sameSu(Hw) band (Fig. 3c), supporting the idea that strong Su(Hw) immunostaining signals are comprised of clusters ofbinding sites located within genes. All five sites were alsoanalyzed using ChIP, to confirm that each of the sites weretrue Su(Hw) binding sites (see Supplementary Fig. 4). ChIPresults show that, in chromatin isolated from embryos, eachone of the sites found within the Sdc gene also interactspositively with Su(Hw) (P<0.0001). Results also show thatall binding sites are located within a unique strong DAPIband. This DAPI band corresponds to cytological bands57E2 to 57E6 in Bridges map, which are normally seen as a
a
b
c
Fig. 3 Su(Hw) bands extensively colocalizewithDAPI bands.a Cytologicalsubdivisions 56F–58A, as depicted by Bridges (Lindsley and Grell 1968).b The same cytological regions stained with DAPI and immunostained withSu(Hw) antibody. Correspondence between Bridges bands, DAPI bands,and immunostaining bands is established. c In situ hybridization combinedwith immunostaining using five different probes containing Su(Hw)binding sites shows that the Sdc gene completely colocalizes with a DAPIband and that all insulator signals coalesce in a single Su(Hw) band
184 Chromosoma (2010) 119:177–194
single band that appears to be generated by the condensa-tion of the DNA of the Sdc gene (Fig. 3c).
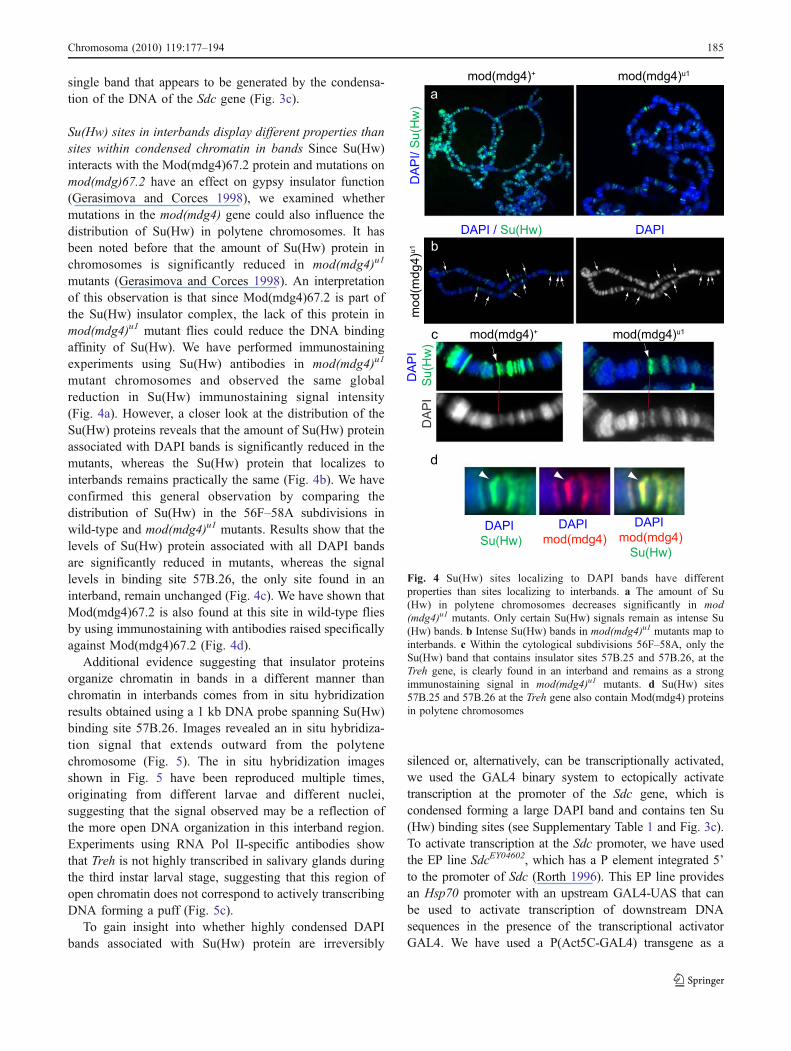
Su(Hw) sites in interbands display different properties thansites within condensed chromatin in bands Since Su(Hw)interacts with the Mod(mdg4)67.2 protein and mutations onmod(mdg)67.2 have an effect on gypsy insulator function(Gerasimova and Corces 1998), we examined whethermutations in the mod(mdg4) gene could also influence thedistribution of Su(Hw) in polytene chromosomes. It hasbeen noted before that the amount of Su(Hw) protein inchromosomes is significantly reduced in mod(mdg4)u1
mutants (Gerasimova and Corces 1998). An interpretationof this observation is that since Mod(mdg4)67.2 is part ofthe Su(Hw) insulator complex, the lack of this protein inmod(mdg4)u1 mutant flies could reduce the DNA bindingaffinity of Su(Hw). We have performed immunostainingexperiments using Su(Hw) antibodies in mod(mdg4)u1
mutant chromosomes and observed the same globalreduction in Su(Hw) immunostaining signal intensity(Fig. 4a). However, a closer look at the distribution of theSu(Hw) proteins reveals that the amount of Su(Hw) proteinassociated with DAPI bands is significantly reduced in themutants, whereas the Su(Hw) protein that localizes tointerbands remains practically the same (Fig. 4b). We haveconfirmed this general observation by comparing thedistribution of Su(Hw) in the 56F–58A subdivisions inwild-type and mod(mdg4)u1 mutants. Results show that thelevels of Su(Hw) protein associated with all DAPI bandsare significantly reduced in mutants, whereas the signallevels in binding site 57B.26, the only site found in aninterband, remain unchanged (Fig. 4c). We have shown thatMod(mdg4)67.2 is also found at this site in wild-type fliesby using immunostaining with antibodies raised specificallyagainst Mod(mdg4)67.2 (Fig. 4d).
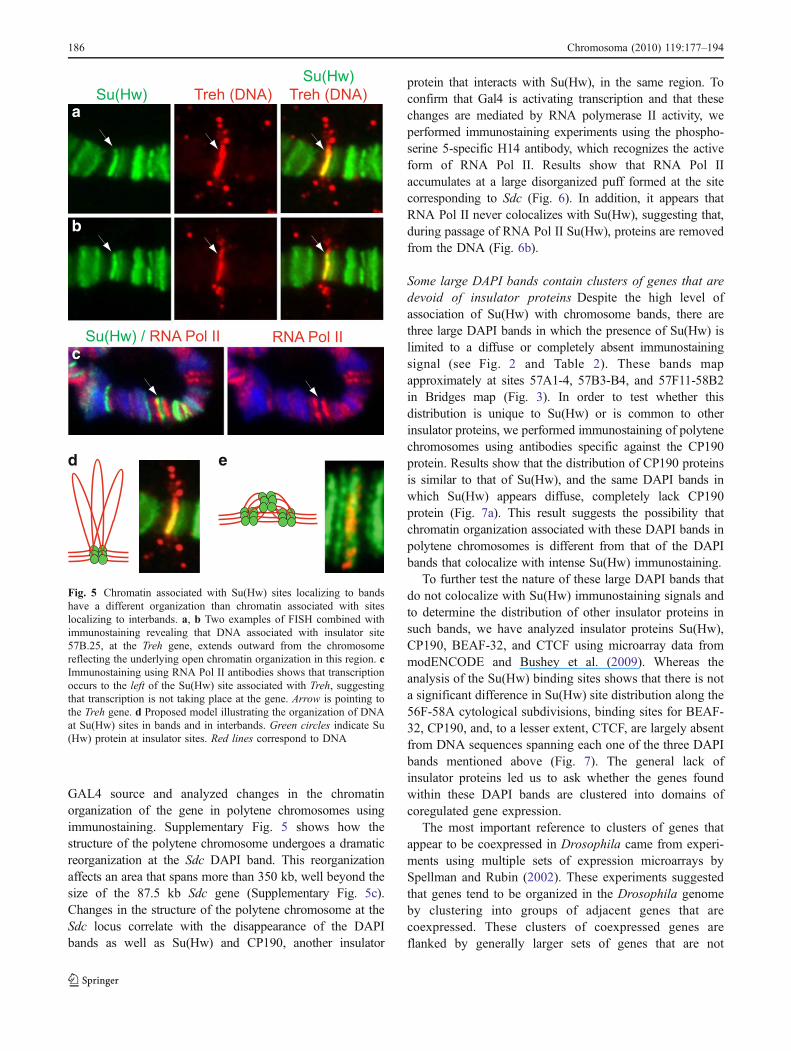
Additional evidence suggesting that insulator proteinsorganize chromatin in bands in a different manner thanchromatin in interbands comes from in situ hybridizationresults obtained using a 1 kb DNA probe spanning Su(Hw)binding site 57B.26. Images revealed an in situ hybridiza-tion signal that extends outward from the polytenechromosome (Fig. 5). The in situ hybridization imagesshown in Fig. 5 have been reproduced multiple times,originating from different larvae and different nuclei,suggesting that the signal observed may be a reflection ofthe more open DNA organization in this interband region.Experiments using RNA Pol II-specific antibodies showthat Treh is not highly transcribed in salivary glands duringthe third instar larval stage, suggesting that this region ofopen chromatin does not correspond to actively transcribingDNA forming a puff (Fig. 5c).
To gain insight into whether highly condensed DAPIbands associated with Su(Hw) protein are irreversibly
silenced or, alternatively, can be transcriptionally activated,we used the GAL4 binary system to ectopically activatetranscription at the promoter of the Sdc gene, which iscondensed forming a large DAPI band and contains ten Su(Hw) binding sites (see Supplementary Table 1 and Fig. 3c).To activate transcription at the Sdc promoter, we have usedthe EP line SdcEY04602, which has a P element integrated 5’to the promoter of Sdc (Rorth 1996). This EP line providesan Hsp70 promoter with an upstream GAL4-UAS that canbe used to activate transcription of downstream DNAsequences in the presence of the transcriptional activatorGAL4. We have used a P(Act5C-GAL4) transgene as a
Fig. 4 Su(Hw) sites localizing to DAPI bands have differentproperties than sites localizing to interbands. a The amount of Su(Hw) in polytene chromosomes decreases significantly in mod(mdg4)u1 mutants. Only certain Su(Hw) signals remain as intense Su(Hw) bands. b Intense Su(Hw) bands in mod(mdg4)u1 mutants map tointerbands. c Within the cytological subdivisions 56F–58A, only theSu(Hw) band that contains insulator sites 57B.25 and 57B.26, at theTreh gene, is clearly found in an interband and remains as a strongimmunostaining signal in mod(mdg4)u1 mutants. d Su(Hw) sites57B.25 and 57B.26 at the Treh gene also contain Mod(mdg4) proteinsin polytene chromosomes
Chromosoma (2010) 119:177–194 185
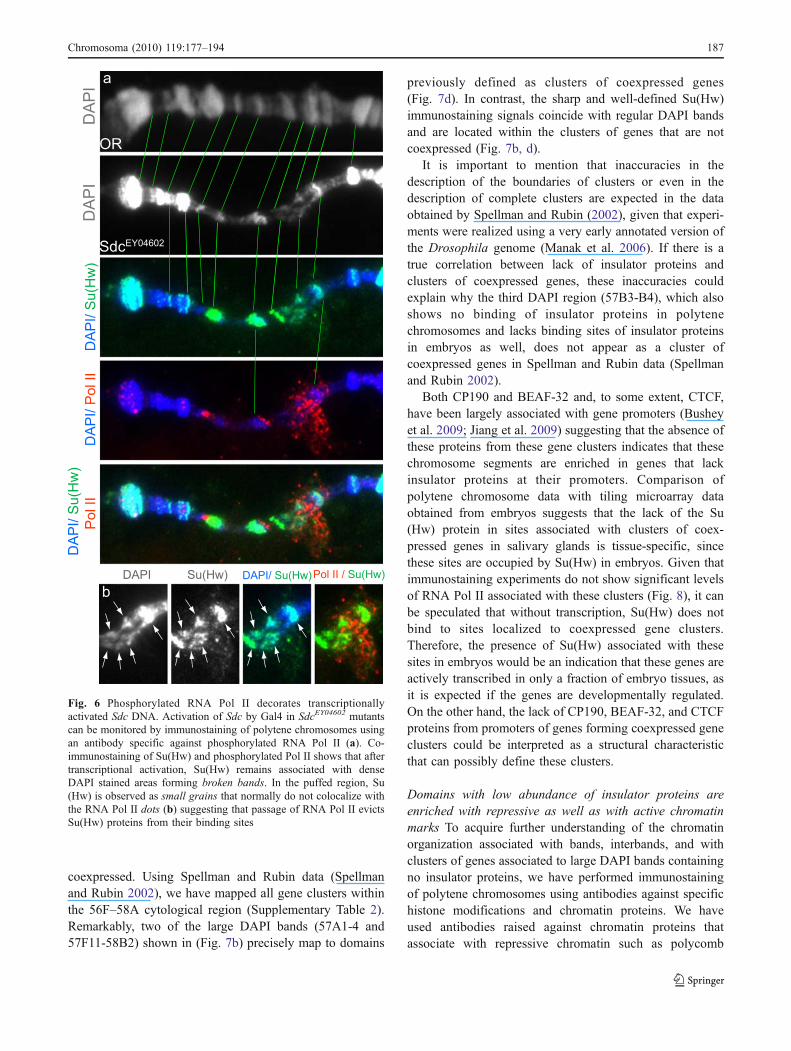
GAL4 source and analyzed changes in the chromatinorganization of the gene in polytene chromosomes usingimmunostaining. Supplementary Fig. 5 shows how thestructure of the polytene chromosome undergoes a dramaticreorganization at the Sdc DAPI band. This reorganizationaffects an area that spans more than 350 kb, well beyond thesize of the 87.5 kb Sdc gene (Supplementary Fig. 5c).Changes in the structure of the polytene chromosome at theSdc locus correlate with the disappearance of the DAPIbands as well as Su(Hw) and CP190, another insulator
protein that interacts with Su(Hw), in the same region. Toconfirm that Gal4 is activating transcription and that thesechanges are mediated by RNA polymerase II activity, weperformed immunostaining experiments using the phospho-serine 5-specific H14 antibody, which recognizes the activeform of RNA Pol II. Results show that RNA Pol IIaccumulates at a large disorganized puff formed at the sitecorresponding to Sdc (Fig. 6). In addition, it appears thatRNA Pol II never colocalizes with Su(Hw), suggesting that,during passage of RNA Pol II Su(Hw), proteins are removedfrom the DNA (Fig. 6b).
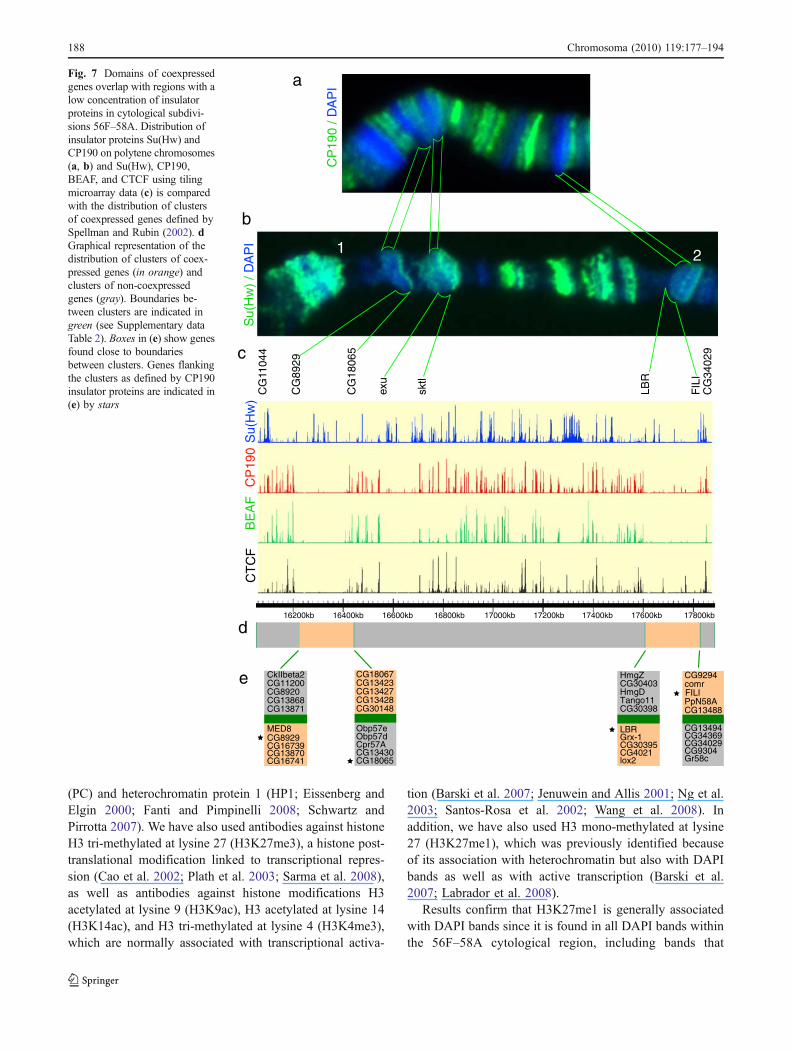
Some large DAPI bands contain clusters of genes that aredevoid of insulator proteins Despite the high level ofassociation of Su(Hw) with chromosome bands, there arethree large DAPI bands in which the presence of Su(Hw) islimited to a diffuse or completely absent immunostainingsignal (see Fig. 2 and Table 2). These bands mapapproximately at sites 57A1-4, 57B3-B4, and 57F11-58B2in Bridges map (Fig. 3). In order to test whether thisdistribution is unique to Su(Hw) or is common to otherinsulator proteins, we performed immunostaining of polytenechromosomes using antibodies specific against the CP190protein. Results show that the distribution of CP190 proteinsis similar to that of Su(Hw), and the same DAPI bands inwhich Su(Hw) appears diffuse, completely lack CP190protein (Fig. 7a). This result suggests the possibility thatchromatin organization associated with these DAPI bands inpolytene chromosomes is different from that of the DAPIbands that colocalize with intense Su(Hw) immunostaining.
To further test the nature of these large DAPI bands thatdo not colocalize with Su(Hw) immunostaining signals andto determine the distribution of other insulator proteins insuch bands, we have analyzed insulator proteins Su(Hw),CP190, BEAF-32, and CTCF using microarray data frommodENCODE and Bushey et al. (2009). Whereas theanalysis of the Su(Hw) binding sites shows that there is nota significant difference in Su(Hw) site distribution along the56F-58A cytological subdivisions, binding sites for BEAF-32, CP190, and, to a lesser extent, CTCF, are largely absentfrom DNA sequences spanning each one of the three DAPIbands mentioned above (Fig. 7). The general lack ofinsulator proteins led us to ask whether the genes foundwithin these DAPI bands are clustered into domains ofcoregulated gene expression.
The most important reference to clusters of genes thatappear to be coexpressed in Drosophila came from experi-ments using multiple sets of expression microarrays bySpellman and Rubin (2002). These experiments suggestedthat genes tend to be organized in the Drosophila genomeby clustering into groups of adjacent genes that arecoexpressed. These clusters of coexpressed genes areflanked by generally larger sets of genes that are not
a
b
c
d e
Fig. 5 Chromatin associated with Su(Hw) sites localizing to bandshave a different organization than chromatin associated with siteslocalizing to interbands. a, b Two examples of FISH combined withimmunostaining revealing that DNA associated with insulator site57B.25, at the Treh gene, extends outward from the chromosomereflecting the underlying open chromatin organization in this region. cImmunostaining using RNA Pol II antibodies shows that transcriptionoccurs to the left of the Su(Hw) site associated with Treh, suggestingthat transcription is not taking place at the gene. Arrow is pointing tothe Treh gene. d Proposed model illustrating the organization of DNAat Su(Hw) sites in bands and in interbands. Green circles indicate Su(Hw) protein at insulator sites. Red lines correspond to DNA
186 Chromosoma (2010) 119:177–194
coexpressed. Using Spellman and Rubin data (Spellmanand Rubin 2002), we have mapped all gene clusters withinthe 56F–58A cytological region (Supplementary Table 2).Remarkably, two of the large DAPI bands (57A1-4 and57F11-58B2) shown in (Fig. 7b) precisely map to domains
previously defined as clusters of coexpressed genes(Fig. 7d). In contrast, the sharp and well-defined Su(Hw)immunostaining signals coincide with regular DAPI bandsand are located within the clusters of genes that are notcoexpressed (Fig. 7b, d).
It is important to mention that inaccuracies in thedescription of the boundaries of clusters or even in thedescription of complete clusters are expected in the dataobtained by Spellman and Rubin (2002), given that experi-ments were realized using a very early annotated version ofthe Drosophila genome (Manak et al. 2006). If there is atrue correlation between lack of insulator proteins andclusters of coexpressed genes, these inaccuracies couldexplain why the third DAPI region (57B3-B4), which alsoshows no binding of insulator proteins in polytenechromosomes and lacks binding sites of insulator proteinsin embryos as well, does not appear as a cluster ofcoexpressed genes in Spellman and Rubin data (Spellmanand Rubin 2002).
Both CP190 and BEAF-32 and, to some extent, CTCF,have been largely associated with gene promoters (Busheyet al. 2009; Jiang et al. 2009) suggesting that the absence ofthese proteins from these gene clusters indicates that thesechromosome segments are enriched in genes that lackinsulator proteins at their promoters. Comparison ofpolytene chromosome data with tiling microarray dataobtained from embryos suggests that the lack of the Su(Hw) protein in sites associated with clusters of coex-pressed genes in salivary glands is tissue-specific, sincethese sites are occupied by Su(Hw) in embryos. Given thatimmunostaining experiments do not show significant levelsof RNA Pol II associated with these clusters (Fig. 8), it canbe speculated that without transcription, Su(Hw) does notbind to sites localized to coexpressed gene clusters.Therefore, the presence of Su(Hw) associated with thesesites in embryos would be an indication that these genes areactively transcribed in only a fraction of embryo tissues, asit is expected if the genes are developmentally regulated.On the other hand, the lack of CP190, BEAF-32, and CTCFproteins from promoters of genes forming coexpressed geneclusters could be interpreted as a structural characteristicthat can possibly define these clusters.
Domains with low abundance of insulator proteins areenriched with repressive as well as with active chromatinmarks To acquire further understanding of the chromatinorganization associated with bands, interbands, and withclusters of genes associated to large DAPI bands containingno insulator proteins, we have performed immunostainingof polytene chromosomes using antibodies against specifichistone modifications and chromatin proteins. We haveused antibodies raised against chromatin proteins thatassociate with repressive chromatin such as polycomb
Fig. 6 Phosphorylated RNA Pol II decorates transcriptionallyactivated Sdc DNA. Activation of Sdc by Gal4 in SdcEY04602 mutantscan be monitored by immunostaining of polytene chromosomes usingan antibody specific against phosphorylated RNA Pol II (a). Co-immunostaining of Su(Hw) and phosphorylated Pol II shows that aftertranscriptional activation, Su(Hw) remains associated with denseDAPI stained areas forming broken bands. In the puffed region, Su(Hw) is observed as small grains that normally do not colocalize withthe RNA Pol II dots (b) suggesting that passage of RNA Pol II evictsSu(Hw) proteins from their binding sites
Chromosoma (2010) 119:177–194 187
(PC) and heterochromatin protein 1 (HP1; Eissenberg andElgin 2000; Fanti and Pimpinelli 2008; Schwartz andPirrotta 2007). We have also used antibodies against histoneH3 tri-methylated at lysine 27 (H3K27me3), a histone post-translational modification linked to transcriptional repres-sion (Cao et al. 2002; Plath et al. 2003; Sarma et al. 2008),as well as antibodies against histone modifications H3acetylated at lysine 9 (H3K9ac), H3 acetylated at lysine 14(H3K14ac), and H3 tri-methylated at lysine 4 (H3K4me3),which are normally associated with transcriptional activa-
tion (Barski et al. 2007; Jenuwein and Allis 2001; Ng et al.2003; Santos-Rosa et al. 2002; Wang et al. 2008). Inaddition, we have also used H3 mono-methylated at lysine27 (H3K27me1), which was previously identified becauseof its association with heterochromatin but also with DAPIbands as well as with active transcription (Barski et al.2007; Labrador et al. 2008).
Results confirm that H3K27me1 is generally associatedwith DAPI bands since it is found in all DAPI bands withinthe 56F–58A cytological region, including bands that
b
1 2
c
CG
1104
4
CG
8929
CG
1806
5
exu
sktl
LBR
FIL
I
Su(
Hw
)C
P19
0B
EA
F
CG
3402
9
Su(
Hw
) / D
AP
I
CP
190
/ DA
PIa
CkIIbeta2CG11200CG8920CG13868CG13871
MED8CG8929CG16739CG13870CG16741
CG18067CG13423CG13427CG13428CG30148
Obp57eObp57dCpr57ACG13430CG18065
HmgZCG30403HmgDTango11CG30398
LBRGrx-1CG30395CG4021lox2
CG9294comrFILIPpN58ACG13488
CG13494CG34369CG34029CG9304Gr58c
e
d16200kb 16400kb 16600kb 16800kb 17000kb 17200kb 17400kb 17600kb 17800kb
CT
CF
Fig. 7 Domains of coexpressedgenes overlap with regions with alow concentration of insulatorproteins in cytological subdivi-sions 56F–58A. Distribution ofinsulator proteins Su(Hw) andCP190 on polytene chromosomes(a, b) and Su(Hw), CP190,BEAF, and CTCF using tilingmicroarray data (c) is comparedwith the distribution of clustersof coexpressed genes defined bySpellman and Rubin (2002). dGraphical representation of thedistribution of clusters of coex-pressed genes (in orange) andclusters of non-coexpressedgenes (gray). Boundaries be-tween clusters are indicated ingreen (see Supplementary dataTable 2). Boxes in (e) show genesfound close to boundariesbetween clusters. Genes flankingthe clusters as defined by CP190insulator proteins are indicated in(e) by stars
188 Chromosoma (2010) 119:177–194
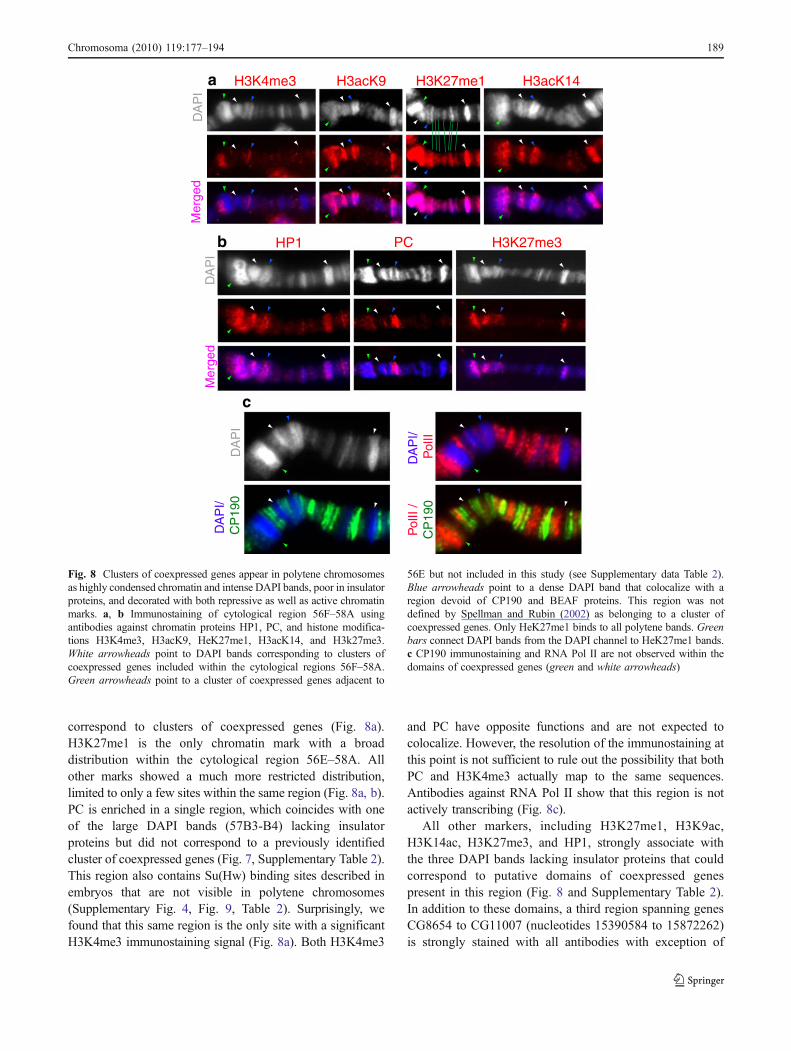
correspond to clusters of coexpressed genes (Fig. 8a).H3K27me1 is the only chromatin mark with a broaddistribution within the cytological region 56E–58A. Allother marks showed a much more restricted distribution,limited to only a few sites within the same region (Fig. 8a, b).PC is enriched in a single region, which coincides with oneof the large DAPI bands (57B3-B4) lacking insulatorproteins but did not correspond to a previously identifiedcluster of coexpressed genes (Fig. 7, Supplementary Table 2).This region also contains Su(Hw) binding sites described inembryos that are not visible in polytene chromosomes(Supplementary Fig. 4, Fig. 9, Table 2). Surprisingly, wefound that this same region is the only site with a significantH3K4me3 immunostaining signal (Fig. 8a). Both H3K4me3
and PC have opposite functions and are not expected tocolocalize. However, the resolution of the immunostaining atthis point is not sufficient to rule out the possibility that bothPC and H3K4me3 actually map to the same sequences.Antibodies against RNA Pol II show that this region is notactively transcribing (Fig. 8c).
All other markers, including H3K27me1, H3K9ac,H3K14ac, H3K27me3, and HP1, strongly associate withthe three DAPI bands lacking insulator proteins that couldcorrespond to putative domains of coexpressed genespresent in this region (Fig. 8 and Supplementary Table 2).In addition to these domains, a third region spanning genesCG8654 to CG11007 (nucleotides 15390584 to 15872262)is strongly stained with all antibodies with exception of
a
DA
PI
H3K27me1 H3acK14H3acK9H3K4me3
Mer
ged
PC H3K27me3HP1
DA
PI
Mer
ged
bD
AP
I
DA
PI/
Pol
II
DA
PI/
C
P19
0
Pol
II /
CP
190
c
Fig. 8 Clusters of coexpressed genes appear in polytene chromosomesas highly condensed chromatin and intense DAPI bands, poor in insulatorproteins, and decorated with both repressive as well as active chromatinmarks. a, b Immunostaining of cytological region 56F–58A usingantibodies against chromatin proteins HP1, PC, and histone modifica-tions H3K4me3, H3acK9, HeK27me1, H3acK14, and H3k27me3.White arrowheads point to DAPI bands corresponding to clusters ofcoexpressed genes included within the cytological regions 56F–58A.Green arrowheads point to a cluster of coexpressed genes adjacent to
56E but not included in this study (see Supplementary data Table 2).Blue arrowheads point to a dense DAPI band that colocalize with aregion devoid of CP190 and BEAF proteins. This region was notdefined by Spellman and Rubin (2002) as belonging to a cluster ofcoexpressed genes. Only HeK27me1 binds to all polytene bands. Greenbars connect DAPI bands from the DAPI channel to HeK27me1 bands.c CP190 immunostaining and RNA Pol II are not observed within thedomains of coexpressed genes (green and white arrowheads)
Chromosoma (2010) 119:177–194 189
H3K4me3 and also corresponds to a cluster of coexpressedgenes that is equally poor in insulator proteins (SeeSupplementary Table 2). These results suggest that thechromosome regions previously identified as containingclusters of coexpressed genes are largely defined by a lowabundance of insulator proteins and are associated with aspecific subset of histone modifications that include bothtranscriptionally active as well as repressive chromatinmarks. The roles that this chromatin structure and absenceof insulator proteins play in these domains is still unclearbut, intriguingly, is reminiscent of the chromatin structureassociated with bivalent domains in vertebrates (Bernsteinet al. 2006; Ku et al. 2008).
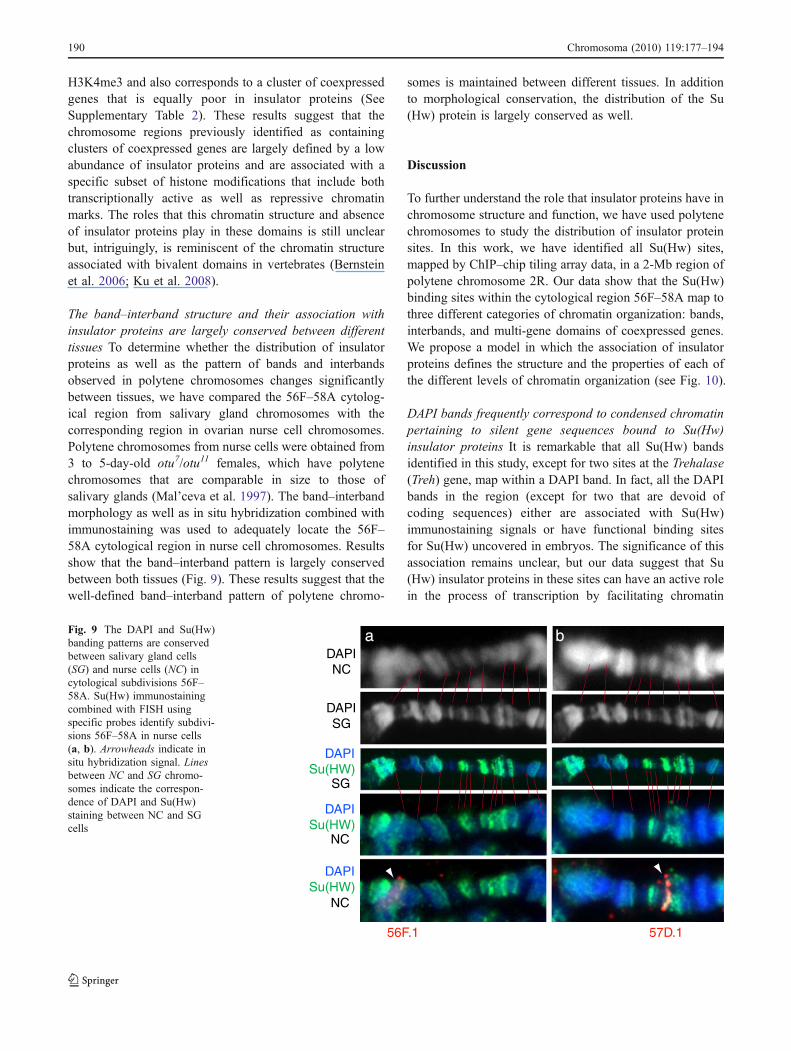
The band–interband structure and their association withinsulator proteins are largely conserved between differenttissues To determine whether the distribution of insulatorproteins as well as the pattern of bands and interbandsobserved in polytene chromosomes changes significantlybetween tissues, we have compared the 56F–58A cytolog-ical region from salivary gland chromosomes with thecorresponding region in ovarian nurse cell chromosomes.Polytene chromosomes from nurse cells were obtained from3 to 5-day-old otu7/otu11 females, which have polytenechromosomes that are comparable in size to those ofsalivary glands (Mal’ceva et al. 1997). The band–interbandmorphology as well as in situ hybridization combined withimmunostaining was used to adequately locate the 56F–58A cytological region in nurse cell chromosomes. Resultsshow that the band–interband pattern is largely conservedbetween both tissues (Fig. 9). These results suggest that thewell-defined band–interband pattern of polytene chromo-
somes is maintained between different tissues. In additionto morphological conservation, the distribution of the Su(Hw) protein is largely conserved as well.
Discussion
To further understand the role that insulator proteins have inchromosome structure and function, we have used polytenechromosomes to study the distribution of insulator proteinsites. In this work, we have identified all Su(Hw) sites,mapped by ChIP–chip tiling array data, in a 2-Mb region ofpolytene chromosome 2R. Our data show that the Su(Hw)binding sites within the cytological region 56F–58A map tothree different categories of chromatin organization: bands,interbands, and multi-gene domains of coexpressed genes.We propose a model in which the association of insulatorproteins defines the structure and the properties of each ofthe different levels of chromatin organization (see Fig. 10).
DAPI bands frequently correspond to condensed chromatinpertaining to silent gene sequences bound to Su(Hw)insulator proteins It is remarkable that all Su(Hw) bandsidentified in this study, except for two sites at the Trehalase(Treh) gene, map within a DAPI band. In fact, all the DAPIbands in the region (except for two that are devoid ofcoding sequences) either are associated with Su(Hw)immunostaining signals or have functional binding sitesfor Su(Hw) uncovered in embryos. The significance of thisassociation remains unclear, but our data suggest that Su(Hw) insulator proteins in these sites can have an active rolein the process of transcription by facilitating chromatin
DAPINC
DAPISG
DAPISu(HW)
DAPISu(HW)
DAPISu(HW)
56F.1
a
57D.1
SG
NC
NC
bFig. 9 The DAPI and Su(Hw)banding patterns are conservedbetween salivary gland cells(SG) and nurse cells (NC) incytological subdivisions 56F–58A. Su(Hw) immunostainingcombined with FISH usingspecific probes identify subdivi-sions 56F–58A in nurse cells(a, b). Arrowheads indicate insitu hybridization signal. Linesbetween NC and SG chromo-somes indicate the correspon-dence of DAPI and Su(Hw)staining between NC and SGcells
190 Chromosoma (2010) 119:177–194

decondensation during the process of transcriptional acti-vation or, alternatively, a repressive role by activelyparticipating in the process of chromatin condensation thatleads to the formation of bands. We have shown thatcondensed chromatin associated with Su(Hw) remainspotentially active, since it can be efficiently decondensedby passage of RNA Pol II. When the Sdc gene isectopically activated by Gal4, the RNA Pol II is capableof inducing the formation of large loops of DNA,generating a prominent puff at the site that was originallyoccupied by a large DAPI band. DNA in the induced puff ishighly enriched in RNA Pol II, which rarely colocalizeswith Su(Hw) proteins. This observation suggests that, inlarge genes containing Su(Hw) sites, the interaction of Su(Hw) with the DNA can be disrupted after transcriptionalactivation. The eviction of Su(Hw) would also disrupthigher-order interactions between Su(Hw) sites, which, inturn, may help in the process of chromatin decondensation.
Su(Hw) in interbands associates to open chromatin Experi-ments in Fig. 4 show that, in the absence of Mod(mdg4)67.2, Su(Hw) proteins remain stably bound to DNA ininterbands, whereas the binding to DNA in bands becomesostensibly weaker, suggesting that Su(Hw) may havealternative functions depending on whether it is targetedto chromosome bands or to interbands. We have shown thatthe Su(Hw) site 57B.26 associated with the gene Trehunequivocally maps to an interband and that in situhybridization images show a signal which may be indica-
tive of the open chromatin structure of interbands.Interestingly, microarray data show that there is anadditional Su(Hw) binding site, 57B.25, approximately5 kb upstream of the 57B.26 site, which maps within thetranscribed region of the Treh gene (SupplementaryTable 2). These two sites cannot be resolved as twoindependent immunostaining signals (see also Supplemen-tary Fig. 3 and Fig. 1). It is possible that the distancebetween these two sites is too small to allow resolution ofthe two independent Su(Hw) bands using our technique.However, an interesting possibility is that the signalobserved by in situ hybridization actually corresponds toDNA loops that are anchored to the two interacting Su(Hw)binding sites (see Fig. 5a, b, and d). Previous reports haveshown that insulator proteins differentially localize to bandsor to the band–interband boundaries in polytene chromo-somes (Capelson and Corces 2004; Pai et al. 2004; Zhao etal. 1995), but, to our knowledge, this is the first time thatthe distribution of insulator proteins has suggested that theymay have differentiated roles in bands and interbands.
Insulator proteins may help organize the genome intodifferentiated transcriptional domains In situ hybridizationcombined with immunostaining revealed that three chro-matin domains in the cytological region 56F–58A appear aslarge DAPI bands that lack Su(Hw) and CP190 proteins.Analysis of tiling microarray data from embryos revealsthat insulator proteins CP190, BEAF-32, and CTCF arealso largely absent from the same domains (Bushey et al.
BEAF-32 / CP190 Su(Hw) / CP190
Interband
DAPI bandDAPI band
Su(Hw) Coexpressiondomain
H3K9ac H3K27me1H3K14ac H3K27me3
Fig. 10 A model integrating the band–interband pattern in polytenechromosomes with the distribution of insulator proteins and geneorganization. Chromatin fiber is indicated as a gray cord. Orangefragments within the chromatin fiber correspond to transcriptionalunits. Broken arrows correspond to gene promoters. Differentinsulator proteins are indicated by spheres and are color-encoded.
Short red segments correspond to Su(Hw) binding sites unoccupied bythe Su(Hw) protein. Histone modifications are indicated by color-encoded circles. Condensed chromatin is indicated by spirals, whichmap to bands, whereas non-condensed chromatin localizes tointerbands, Su(Hw) localizes to all three types of chromatinorganization
Chromosoma (2010) 119:177–194 191

2009 and modENCODE consortium). In contrast, the samedata show that Su(Hw) binding sites are found both withinthe domains and outside the domains. Therefore, true Su(Hw) binding sites within the domains, as determined bymicroarray experiments in embryos, are not bound by Su(Hw) in polytene chromosomes where genes are transcrip-tionally silent (Fig. 7).
We also show that two of these DAPI bands map exactly tothe only two clusters of coexpressed genes described bySpellman and Rubin in this region (Spellman and Rubin2002). Clusters of coexpressed genes are common toeukaryotic organisms including Drosophila, Caenorhabditiselegans, and humans. These clusters are ordered non-randomly in the genome and consists of similarly expressedmulti-gene domains (Boutanaev et al. 2002; Caron et al.2001; Dorus et al. 2006; Lee and Sonnhammer 2003; Milleret al. 2004; Roy et al. 2002; Versteeg et al. 2003; Yi et al.2007). This result opens the possibility that the domains ofcoexpressed genes could actually be defined by a lowabundance or a complete absence of insulator proteins suchas CP190, BEAF-32, and CTCF. The lack of all insulatorproteins would explain the coexpression of all the genes inthe domain, and the activity of insulator proteins within theclusters of non-coexpressed genes would explain the lack ofcoexpression, but this possibility raises the question of therole that Su(Hw) plays in the domains.
A rapid inspection of the classes of genes found in eachtype of multi-gene domain defined by their coexpression levelsuggests that genes found in non-coexpressed multi-genedomains frequently have CP190 and BEAF-32 at theirpromoters, are highly expressed, show very short intergenicdistances between them, and encode proteins with essentialcellular functions. For example, it has been shown that mostgenes regulated by BEAF-32 are involved in basic functions,such as cell cycle regulation or chromosome segregation(Emberly et al. 2008). Genes found in the domains ofcoexpressed genes, however, lack CP190 and BEAF-32, aregenerally tissue-specific, are developmentally regulated, andshow large intergenic distances.
Our data show that the organization described above islargely conserved between tissues given the minimaldifferences found when salivary gland cells and nurse cellsare compared (Fig. 9). In both tissues, the band–interbandpattern as well as the distribution of Su(Hw) protein ispractically identical. It is tempting to speculate that thisorganization is not specific to salivary glands or nurse cellsbut instead may largely represent how Drosophila chromo-somes are generally organized regardless of cell lineage. Ifthis hypothesis is correct, it would be expected that majordifferences in the band–interband pattern as well as in thedistribution of Su(Hw) protein between cell lineages wouldonly occur upon induction or tissue-specific activation ofgenes, such as Sdc, residing within the bands, whereas there
would be genes that will always localize to interbandsregardless of the cell type. Such genes will correspond togenes encoding proteins required for basic cell functionssuch as those having BEAF-32 at their promoters orinducible genes such as Treh. Inducible genes have toremain in a chromatin state that will allow rapid activationbut would stay off in the absence of induction in a varietyof tissues and cell types (see proposed model in Fig. 10).The specific role of Su(Hw) in the formation of thesehigher-order structures or domains remains obscure, sincemutations at the su(Hw) gene alone do not significantlychange the band–interband pattern or the morphology ofpolytene chromosomes, nor do they have severe effects onviability (Smith and Corces 1995). However, since mostbands and interbands are conserved between much unrelatedtissues, it is expected that the condensed chromatin associatedto bands or to domains of coexpressed genes would formbefore cell differentiation, when there is still a supply ofmaternally deposited Su(Hw) and would be transmittedepigenetically during cell division.
The fact that histone modifications normally associatedwith transcriptional activation are found within the samedomains as histone modifications normally associated withtranscriptional repression is reminiscent of the bivalentdomains initially found in stem cells and deserves additionalcharacterization. In-depth analysis assessing the chromatinproperties of bands, interbands, and clusters of coexpressedgenes should provide further insights into the role of insulatorsin the organization of these chromatin domains and whetherthey play an active role in gene expression regulation duringdevelopment and cell differentiation.
Acknowledgements We thank Kevin White and Nicolas Negre forgraciously allowing us to use their unpublished data released by theModENCODE consortium. We also thank Tom Dockendorff and threeanonymous reviewers for valuable comments on earlier versions ofthis manuscript. This work was supported by US National ScienceFoundation Award 0616081 to ML.
References
Adryan B, Woerfel G, Birch-Machin I, Gao S, Quick M, MeadowsL, Russell S, White R (2007) Genomic mapping of suppressorof Hairy-wing binding sites in Drosophila. Genome Biol 8:R167
Apostolou E, Thanos D (2008) Virus infection induces NF-kappaB-dependent interchromosomal associations mediating monoallelicIFN-beta gene expression. Cell 134:85–96
Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, Wei G,Chepelev I, Zhao K (2007) High-resolution profiling of histonemethylations in the human genome. Cell 129:823–837
Bartkuhn M, Renkawitz R (2008) Long range chromatin interactionsinvolved in gene regulation. Biochim Biophys Acta 1783:2161–2166
192 Chromosoma (2010) 119:177–194

Bartkuhn M, Straub T, Herold M, Herrmann M, Rathke C,Saumweber H, Gilfillan GD, Becker PB, Renkawitz R (2009)Active promoters and insulators are marked by the centrosomalprotein 190. EMBO J 28:877–888
Bell AC, Felsenfeld G (2000) Methylation of a CTCF-dependentboundary controls imprinted expression of the Igf2 gene. Nature405:482–485
Bell AC, West AG, Felsenfeld G (1999) The protein CTCF is requiredfor the enhancer blocking activity of vertebrate insulators. Cell98:387–396
Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J,Fry B, Meissner A, Wernig M, Plath K, Jaenisch R, Wagschal A,Feil R, Schreiber SL, Lander ES (2006) A bivalent chromatinstructure marks key developmental genes in embryonic stemcells. Cell 125:315–326
Boutanaev AM, Kalmykova AI, Shevelyov YY, Nurminsky DI (2002)Large clusters of co-expressed genes in the Drosophila genome.Nature 420:666–669
Bushey AM, Ramos E, Corces VG (2009) Three subclasses of aDrosophila insulator show distinct and cell type-specific genomicdistributions. Genes Dev 23:1338–1350
Cai HN, Shen P (2001) Effects of cis arrangement of chromatininsulators on enhancer-blocking activity. Science 291:493–495
Cao R, Wang L, Wang H, Xia L, Erdjument-Bromage H, Tempst P,Jones RS, Zhang Y (2002) Role of histone H3 lysine 27methylation in Polycomb-group silencing. Science 298:1039–1043
Capelson M, Corces VG (2004) Boundary elements and nuclearorganization. Biol Cell 96:617–629
Caron H, van Schaik B, van der Mee M, Baas F, Riggins G, van SluisP, Hermus MC, van Asperen R, Boon K, Voute PA, HeisterkampS, van Kampen A, Versteeg R (2001) The human transcriptomemap: clustering of highly expressed genes in chromosomaldomains. Science 291:1289–1292
Cavalli G, Orlando V, Paro R (1999) Mapping DNA target sites ofchromatin-associated proteins by formaldehyde cross-linking inDrosophila embryos. In: Bickmore WA (ed) Chromosomestructural analysis: a practical approach. Oxford University Press,UK, pp 20–37
Chung JH, Whiteley M, Felsenfeld G (1993) A 5’ element of thechicken beta-globin domain serves as an insulator in humanerythroid cells and protects against position effect in Drosophila.Cell 74:505–514
Dorus S, Busby SA, Gerike U, Shabanowitz J, Hunt DF, Karr TL(2006) Genomic and functional evolution of the Drosophilamelanogaster sperm proteome. Nat Genet 38:1440–1445
Eissenberg JC, Elgin SC (2000) The HP1 protein family: getting agrip on chromatin. Curr Opin Genet Dev 10:204–210
Emberly E, Blattes R, Schuettengruber B, Hennion M, Jiang N, HartCM, Kas E, Cuvier O (2008) BEAF regulates cell-cycle genesthrough the controlled deposition of H3K9 methylation marksinto its conserved dual-core binding sites. PLoS Biol 6:2896–2910
Fanti L, Pimpinelli S (2008) HP1: a functionally multifaceted protein.Curr Opin Genet Dev 18:169–174
Gaszner M, Felsenfeld G (2006) Insulators: exploiting transcriptionaland epigenetic mechanisms. Nat Rev Genet 7:703–713
Gaszner M, Vazquez J, Schedl P (1999) The Zw5 protein, acomponent of the scs chromatin domain boundary, is able toblock enhancer–promoter interaction. Genes Dev 13:2098–2107
Gerasimova TI, Corces VG (1998) Polycomb and trithorax groupproteins mediate the function of a chromatin insulator. Cell92:511–521
Gerasimova TI, Corces VG (2001) Chromatin insulators and bound-aries: effects on transcription and nuclear organization. Annu RevGenet 35:193–208
Gerasimova TI, Gdula DA, Gerasimov DV, Simonova O, Corces VG(1995) A Drosophila protein that imparts directionality on achromatin insulator is an enhancer of position–effect variegation.Cell 82:587–597
Gerasimova TI, Lei EP, Bushey AM, Corces VG (2007) Coordinatedcontrol of dCTCF and gypsy chromatin insulators in Drosophila.Mol cell 28:761–772
Geyer PK, Corces VG (1992) DNA position-specific repression oftranscription by a Drosophila zinc finger protein. Genes Dev6:1865–1873
Ghosh D, Gerasimova TI, Corces VG (2001) Interactions between theSu(Hw) and Mod(mdg4) proteins required for gypsy insulatorfunction. Embo J 20:2518–2527
Gilbert MK, Tan YY, Hart CM (2006) The Drosophila boundaryelement-associated factors BEAF-32A and BEAF-32B affectchromatin structure. Genetics 173:1365–1375
Jenuwein T, Allis CD (2001) Translating the histone code. Science293:1074–1080
Jiang N, Emberly E, Cuvier O, Hart CM (2009) Genome-widemapping of boundary element-associated factor (BEAF) bindingsites in Drosophila melanogaster links BEAF to transcription.Mol Cell Biol 29:3556–3568
Kaplan CD, Morris JR, Wu C, Winston F (2000) Spt5 and spt6 areassociated with active transcription and have characteristics ofgeneral elongation factors in D. melanogaster. Genes Dev14:2623–2634
Kellum R, Schedl P (1991) A position-effect assay for boundaries ofhigher order chromosomal domains. Cell 64:941–950
Ku M, Koche RP, Rheinbay E, Mendenhall EM, Endoh M, MikkelsenTS, Presser A, Nusbaum C, Xie X, Chi AS, Adli M, Kasif S,Ptaszek LM, Cowan CA, Lander ES, Koseki H, Bernstein BE(2008) Genomewide analysis of PRC1 and PRC2 occupancyidentifies two classes of bivalent domains. PLoS Genet 4:e1000242
Kuhn EJ, Geyer PK (2003) Genomic insulators: connecting propertiesto mechanism. Curr Opin Cell Biol 15:259–265
Kuhn-Parnell EJ, Helou C, Marion DJ, Gilmore BL, Parnell TJ, WoldMS, Geyer PK (2008) Investigation of the properties of non-gypsy suppressor of Hairy-wing-binding sites. Genetics179:1263–1273
Kyrchanova O, Chetverina D, Maksimenko O, Kullyev A, Georgiev P(2008) Orientation-dependent interaction between Drosophilainsulators is a property of this class of regulatory elements.Nucleic Acids Res 36:7019–7028
Labrador M, Corces VG (2002) Setting the boundaries of chromatindomains and nuclear organization. Cell 111:151–154
Labrador M, Corces VG (2003) Phosphorylation of histone H3 duringtranscriptional activation depends on promoter structure. GenesDev 17:43–48
Labrador M, Sha K, Li A, Corces VG (2008) Insulator and ovoproteins determine the frequency and specificity of insertion ofthe gypsy retrotransposon in Drosophila melanogaster. Genetics180:1367–1378
Lee JM, Sonnhammer EL (2003) Genomic gene clustering analysis ofpathways in eukaryotes. Genome Res 13:875–882
Lindsley DL, Grell EH (1968) Genetic variations of Drosophilamelanogaster. Washington, DC: Carnegie Institution. 472 p.Publication no 627. Biology Division, Oak Ridge NationalLaboratory, Oak Ridge, TN
Ling JQ, Li T, Hu JF, Vu TH, Chen HL, Qiu XW, Cherry AM,Hoffman AR (2006) CTCF mediates interchromosomal colocal-ization between Igf2/H19 and Wsb1/Nf1. Science 312:269–272
Lis JT (2007) Imaging Drosophila gene activation and polymerasepausing in vivo. Nature 450:198–202
Maksimenko O, Golovnin A, Georgiev P (2008) Enhancer–promotercommunication is regulated by insulator pairing in a Drosophilamodel bigenic locus. Mol Cell Biol 28:5469–5477
Chromosoma (2010) 119:177–194 193

Mal’ceva NI, Belyaeva ES, King RC, Zhimulev IF (1997) Nurse cellpolytene chromosomes of Drosophila melanogaster otu mutants:morphological changes accompanying interallelic complementa-tion and position effect variegation. Dev Genet 20:163–174
Manak JR, Dike S, Sementchenko V, Kapranov P, Biemar F, Long J,Cheng J, Bell I, Ghosh S, Piccolboni A, Gingeras TR (2006)Biological function of unannotated transcription during the earlydevelopment of Drosophila melanogaster. Nat Genet 38:1151–1158
Markstein M, Levine M (2002) Decoding cis-regulatory DNAs in theDrosophila genome. Curr Opin Genet Dev 12:601–606
Miller MA, Cutter AD, Yamamoto I, Ward S, Greenstein D (2004)Clustered organization of reproductive genes in the C. elegansgenome. Curr Biol 14:1284–1290
Moon H, Filippova G, Loukinov D, Pugacheva E, Chen Q, SmithST, Munhall A, Grewe B, Bartkuhn M, Arnold R, Burke LJ,Renkawitz-Pohl R, Ohlsson R, Zhou J, Renkawitz R,Lobanenkov V (2005) CTCF is conserved from Drosophilato humans and confers enhancer blocking of the Fab-8 insulator.EMBO rep 6:165–170
Muravyova E, Golovnin A, Gracheva E, Parshikov A, Belenkaya T,Pirrotta V, Georgiev P (2001) Loss of insulator activity by pairedSu(Hw) chromatin insulators. Science 291:495–498
Ng HH, Robert F, Young RA, Struhl K (2003) Targeted recruitment ofSet1 histone methylase by elongating Pol II provides a localizedmark and memory of recent transcriptional activity. Mol cell11:709–719
Osborne CS, Chakalova L, Brown KE, Carter D, Horton A, Debrand E,Goyenechea B, Mitchell JA, Lopes S, Reik W, Fraser P (2004)Active genes dynamically colocalize to shared sites of ongoingtranscription. Nat Genet 36:1065–1071
Pai CY, Lei EP, Ghosh D, Corces VG (2004) The centrosomal proteinCP190 is a component of the gypsy chromatin insulator. Mol cell16:737–748
Phillips JE, Corces VG (2009) CTCF: master weaver of the genome.Cell 137:1194–1211
Plath K, Fang J, Mlynarczyk-Evans SK, Cao R, Worringer KA, WangH, de la Cruz CC, Otte AP, Panning B, Zhang Y (2003) Role ofhistone H3 lysine 27 methylation in X inactivation. Science300:131–135
Ptashne M, Gann A (1997) Transcriptional activation by recruitment.Nature 386:569–577
Rorth P (1996) A modular misexpression screen in Drosophiladetecting tissue-specific phenotypes. Proc Natl Acad Sci U S A93:12418–12422
Roy PJ, Stuart JM, Lund J, Kim SK (2002) Chromosomal clusteringof muscle-expressed genes in Caenorhabditis elegans. Nature418:975–979
Santos-Rosa H, Schneider R, Bannister AJ, Sherriff J, Bernstein BE,Emre NC, Schreiber SL, Mellor J, Kouzarides T (2002) Activegenes are tri-methylated at K4 of histone H3. Nature 419:407–411
Sarma K, Margueron R, Ivanov A, Pirrotta V, Reinberg D (2008) Ezh2requires PHF1 to efficiently catalyze H3 lysine 27 trimethylationin vivo. Mol Cell Biol 28:2718–2731
Saunders A, Werner J, Andrulis ED, Nakayama T, Hirose S, ReinbergD, Lis JT (2003) Tracking FACT and the RNA polymerase IIelongation complex through chromatin in vivo. Science301:1094–1096
Schuettengruber B, Chourrout D, Vervoort M, Leblanc B, Cavalli G(2007) Genome regulation by polycomb and trithorax proteins.Cell 128:735–745
Schwartz YB, Pirrotta V (2007) Polycomb silencing mechanismsand the management of genomic programmes. Nat Rev Genet8:9–22
Smith PA, Corces VG (1995) The suppressor of Hairy-wing proteinregulates the tissue-specific expression of the Drosophila gypsyretrotransposon. Genetics 139:215–228
Smith ST, Wickramasinghe P, Olson A, Loukinov D, Lin L, Deng J,Xiong Y, Rux J, Sachidanandam R, Sun H, Lobanenkov V, Zhou J(2009) Genome wide ChIP-chip analyses reveal important rolesfor CTCF in Drosophila genome organization. Dev Biol328:518–528
Spana C, Harrison DA, Corces VG (1988) The Drosophila mela-nogaster suppressor of Hairy-wing protein binds to specificsequences of the gypsy retrotransposon. Genes Dev 2:1414–1423
Spellman PT, Rubin GM (2002) Evidence for large domains ofsimilarly expressed genes in the Drosophila genome. J Biol 1:5
Spilianakis CG, Lalioti MD, Town T, Lee GR, Flavell RA (2005)Interchromosomal associations between alternatively expressedloci. Nature 435:637–645
Splinter E, Heath H, Kooren J, Palstra RJ, Klous P, Grosveld F, GaljartN, de Laat W (2006) CTCF mediates long-range chromatinlooping and local histone modification in the beta-globin locus.Genes Dev 20:2349–2354
Versteeg R, van Schaik BD, van Batenburg MF, Roos M, MonajemiR, Caron H, Bussemaker HJ, van Kampen AH (2003) Thehuman transcriptome map reveals extremes in gene density,intron length, GC content, and repeat pattern for domains ofhighly and weakly expressed genes. Genome Res 13:1998–2004
Walhout AJ (2006) Unraveling transcription regulatory networks byprotein–DNA and protein–protein interaction mapping. GenomeRes 16:1445–1454
Wallace JA, Felsenfeld G (2007) We gather together: insulators andgenome organization. Curr Opin Genet Dev 17:400–407
Wang Z, Zang C, Rosenfeld JA, Schones DE, Barski A, Cuddapah S,Cui K, Roh TY, Peng W, Zhang MQ, Zhao K (2008)Combinatorial patterns of histone acetylations and methylationsin the human genome. Nat Genet 40:897–903
West AG, Gaszner M, Felsenfeld G (2002) Insulators: many functions,many mechanisms. Genes Dev 16:271–288
Yao J, Zobeck KL, Lis JT, Webb WW (2008) Imaging transcriptiondynamics at endogenous genes in living Drosophila tissues.Methods (San Diego, Calif) 45:233–241
Yi G, Sze SH, Thon MR (2007) Identifying clusters of functionallyrelated genes in genomes. Bioinformatics 23:1053–1060
Zhao K, Hart CM, Laemmli UK (1995) Visualization of chromosomaldomains with boundary element-associated factor BEAF-32. Cell81:879–889
194 Chromosoma (2010) 119:177–194





























