Embed Size (px)
Citation preview

Changes of soil microbial biomass C and basal soil
respiration in different land use and land cover
R. Kızılkaya *
, O. Dengiz , T. Alparslan, M. Durmuş, V. Işıldak, S. Aksu
Ondokuz Mayıs University, Faculty of Agriculture, Department of Soil Science, Samsun,Turkey
Abstract
Soil microbial biomass (Cmic) and basal soil respiration (BSR) are being increasingly recognized as a sensitive
indicator of soil quality. Theirs knowledge are fundamental for sustainable environmental management. This
study aimed to determine the impact of different land uses (forest, pasture, and agricultural lands) on Cmic and
BSR. In this study, the changes in the properties of four different pedons classified as Lithic Xerorthent, Typic
Dystroxerept and Typic Haploxerept located on three adjacent land use types which are native forest, pasture
and cultivated fields include some physico-chemical properties (texture, pH, EC, lime content and organic
matter), Cmic and BSR. It was determined that Cmic and BSR of the soils varied from 29 – 786 mg C kg-1
and
67-2198 µg CO2-C g-1
, respectively. In addition, it was found that changes of land use and land cover associated
with organic matter content can alter the Cmic and BSR within the soil profile. It was concluded that Cmic and
BSR decreased through profile depth, and Cmic and BSR were shown to be significantly correlated to the
physico-chemical properties of the soil. Present study clearly shows that land use has a significant effect Cmic
and BSR in soil by altering natural soil characteristics under the same ecological conditions.
Key words: land use, microbial biomass, basal soil respiration, profile, organic matter
INTRODUCTION The complex integration of the primary natural resources — soil, water and vegetation, is
vital for maintaining terrestrial ecosystem functions and productivity. Human poverty and a
continuous decline in the amount of agricultural land per person have led to indiscriminate
exploitation of natural resources in developing countries of the world (Mahtab and Karim,
1992). Land use changes, especially cultivation of deforested land may rapidly diminish soil
quality, as ecologically sensitive components of ecosystem are not able to buffer the effects
of agricultural practices. As a result, severe deterioration in soil quality may lead to a
* Corresponding author: Rıdvan Kızılkaya E-mail: [email protected]
Kızılkaya, R., Dengiz, O., Alparslan, T., Durmuş, M., Işıldak, V., Aksu, S.,2010. Changes of soil microbial biomass
C and basal soil respiration in different land use and land cover. In: Proceedings of the International Soil
Science Congress on Management of Natural Resources to Sustain Soil Health and Quality. R.Kizilkaya,
C.Gulser, O.Dengiz (eds.), May 26-28, 2010. Ondokuz Mayis University, Samsun, Turkey. pp. 1039-1046

R. Kızılkaya et al.
Changes of soil microbial biomass C and basal soil respiration in different land...
1040
permanent degradation of land productivity (Kang and Juo, 1986; Nardi et al., 1996; Islam et
al., 1999).
The important role that soil microorganisms play in the nutrient and energy-flow
relationships of natural as well as man-manipulated environments has given rise to the need
for easily measured biological indicators of ecosystem development and disturbance (Yan et
al., 2003). Soil microorganisms are also agents that promote aggregate stability and good soil
structure. Several studies have shown that the soil microbial biomass changes more quickly
than soil organic matter as a whole due to changes in soil management. Soil microbial
biomass (Cmic) and basal soil respiration (BSR) are being increasingly recognized as a
sensitive indicator of soil quality. Theirs knowledge are fundamental for sustainable
environmental management. For example, Powlson et al. (1987) showed that 18 years of
straw incorporation increased the biomass by about 50%, while changes in total organic
matter were undetectable. Chander and Brookes (1991) demonstrated that the ratio of
biomass C to soil organic C was a sensitive indicator of the effects of heavy metals on the
microbial biomass using two different field experiment soils. Powlson et al. (1987) and
Wardle (1992) pointed out that the soil microbial biomass and biomass C/organic C ratio can
provide an effective early warning of the deterioration of soil quality. Killham (1985) and
Killham and Firestone (1984) showed that soil microorganisms divert more energy from
growth into maintenance as stress increases and thus the ratio of respired C to biomass can
be a much more sensitive indicator of stress.
This study aimed to determine the impact of different land uses (forest, pasture, and
agricultural lands) on Cmic and BSR. In this study, the changes in the properties of four
different pedons classified as Lithic Xerorthent, Typic Dystroxerept and Typic Haploxerept
located on three adjacent land use types which are native forest, pasture and cultivated
fields include some physico-chemical properties (texture, pH, EC, lime content and organic
matter), Cmic and BSR.
MATERIALS AND METHODS Description of the study field
The study area is located in Çankırı-Uludere watershed, which extend from 40o
45ı to 40
o 52
ı
N latitude and has an elevation between 1200-1846m above sea level within longitude 33o
37ı and 33
o 52
ı E in Central Anatolia of Turkey (Figure 1). According to Thorntwaite (1948),
half humid micro thermal having abundant water supply in winter land climate and that was
coded with C2B1ıs2d
ı. The long-term mean annual temperature and precipitation were 9.1
oC and 530.8 mm, respectively. Topography and slope show great variations and hilly and
rolling physiographic units are particularly common in the study area. Geology of the study
area is dominantly composed of ophiolitic series and basalts. In addition, there are marls,
conglomerates and stone in the study area. The research area is located in the Iran-Turan
flora zone that is one of the three major flora zones of Turkey and lies in the A4 square
according to the Davis’s grid sistem (Davis, 1965). Dominant tree species of natural forest
are Pinus sylevestris L.and Abies nordmanniana in Uludere watershed. Some part of natural
forest has been fragmented and degraded by such human disturbances as clearance for
agriculture activities and pasture.
Soil physico-chemical analysis
Four soil profiles were selected for this study from four sites in each of four adjacent land
use types which are native forest, pasture and cultivated fields. These adjacent four profiles
locate on the similar aspect, elevation and slope. Disturbed soil samples were then air-dried

R. Kızılkaya et al.
Changes of soil microbial biomass C and basal soil respiration in different land...
1041
and passed through a 2 mm sieve to prepare for laboratory analysis. Soil samples were then
air-dried and passed through a 2 mm sieve, particle size distribution was determined by the
hydrometer method (Bouyoucos, 1951). Soil organic carbon was measured by wet digestion
(Nelson and Sommers, 1982). Values of soil organic carbon were multiplied by a factor 1.72
to obtain soil organic matter. pH, EC-electrical conductivity (of the saturation) by method of
the (Soil Survey Labrotory, 1992).
The Study Area
Figure 1. Location map of the study area
Microbial biomass and basal respiration analysis
Microbial biomass carbon (Cmic) was determined by the substrate-induced respiration
method of by Anderson and Domsch (1978). A moist sample equivalent to 10 g oven-dry
soil or cats was amended with a powder mixture containing 40 mg glucose. The CO2
production rate was measured hourly using the method described by Anderson (1982).
The pattern of respiratory response was recorded for 4 h. Microbial biomass carbon (Cmic)
was calculated from the maximum initial respiratory response in terms of mg C g-1 soil as
40.04 mg CO2 g-1 + 3,75. Data are expressed as mg C g-1 dry sample.
Basal soil respiration (BSR) at field capacity (CO2 production at 22 0C without addition of
glucose) was measured, as reported by Anderson (1982); by alkali (Ba(OH)2.8H2O + BaCI2)
absorption of the CO2 produced during the 24h incubation period, followed by titration of
the residual OH- with standardized hydrochloric acid, after adding three drops of
phenolphthalein as an indicator. Data are expressed as µg CO2-C g-1
dry sample.
All determinations of Cmic and BSR were performed for the each soil sample in triplicate,
and all values reported are averages of the three determinations expressed on an oven-dried
sample basis at 105 0C for 24 h.

R. Kızılkaya et al.
Changes of soil microbial biomass C and basal soil respiration in different land...
1042
NATURAL FOREST LAND (Pinus sylvestris L.)
Soil class: Lithic Xerorthent
3,80
1,07
0,75
0,00 0,50 1,00 1,50 2,00 2,50 3,00 3,50 4,00 4,50
20-38 cm
13-20 cm
0-13 cm
C2
C1
A
Total organic C, g. 100 g-1
NATURAL FOREST LAND (Abies nordmanniana )
Soil class: Typic Dystroxerept
0,30
0,56
1,14
3,02
3,62
0,00 0,50 1,00 1,50 2,00 2,50 3,00 3,50 4,00
70 + cm
58-70 cm
38-58 cm
28-38 cm
0-28 cm
C2
C1
Bw
2B
w1
A1
Total organic C, g. 100 g-1
PASTURE LAND
Soil class: Typic Dystroxerept
0,32
0,35
0,4
0,9
1,05
0 0,2 0,4 0,6 0,8 1 1,2
100+ cm
80-100 cm
46-80 cm
20-46 cm
0-20 cm
CB
w2
Bw
1A
2A
1
Total organic C, g. 100 g-1
CULTIVATED LAND
Soil class: Typic Haploxerept
0,32
0,35
0,4
0,9
1,63
0 0,5 1 1,5 2
70+ cm
50-70 cm
43-50 cm
10-43 cm
0-10 cm
CB
CB
wB
CA
p
Total organic C, g. 100 g-1
RESULTS AND DISCUSSION The natural forest soils were considerably lower in silt and slightly lower in clay than the
adjacent soils under cultivated and pasture soils, most likely as a result of preferential
removal of silt and clay by accelerated water erosion. Because, depths of the natural forest
soils have less thick than other soils that leads to low water retention in soil and occurs
runoff. Similarly, Jaiyeoba (2003) indicated that clay contents of deeper depths increase
with the increases of cultivation year due to either increases of clay translocation from the
surface horizon or removal of clay from the surface by runoff. In this study, soil pH tends to
increase in the cultivated lands. The pH values of the natural forest, pasture and cultivated
lands varied significantly from 6.03 to 7.71 (Table 1). Natural forest and pasture soils were
more acidic than those of the cultivated sites. However, soil pH slightly increases with soil
depth due to accumulation of basic cations in cultivated lands. Lime content in forest and
pasture soil were the similar while lime content significantly increased under cultivated land.
The highest lime content was obtained at 43 – 50 cm depth of the cultivated land.
Conversion of the natural forest into continuous cultivation had resulted in significant
reductions of both the concentration and stock of organic matter and organic carbon (Table
1 and Figure 2). Lobe et al (2001) reported that the organic matter and organic carbon
content in soils decreased rapidly in the first few years they were cultivated. Cultivation
lands and pasture, the topsoil contained less organic matter than the continuously natural
forestland due to forest clearance and high decompositions in the study area.
Figure 2. Total organic carbon of the three different adjacent land use types

R. Kızılkaya et al.
Changes of soil microbial biomass C and basal soil respiration in different land...
1043
NATURAL FOREST LAND (Pinus sylvestris L.)
Soil class: Lithic Xerorthent
786
230
114
0 100 200 300 400 500 600 700 800 900
20-38 cm
13-20 cm
0-13 cm
C2
C1
A
Microbial biomass C, mg C kg-1
NATURAL FOREST LAND (Abies nordmanniana )
Soil class: Typic Dystroxerept
45
56
98
323
698
0 100 200 300 400 500 600 700 800
70 + cm
58-70 cm
38-58 cm
28-38 cm
0-28 cm
C2
C1
Bw
2B
w1
A1
Microbial biomass C, mg C kg-1
PASTURE LAND
Soil class: Typic Dystroxerept
37
45
89
263
478
0 100 200 300 400 500 600
100+ cm
80-100 cm
46-80 cm
20-46 cm
0-20 cm
CB
w2
Bw
1A
2A
1
Microbial biomass C, mg C kg-1
CULTIVATED LAND
Soil class: Typic Haploxerept
29
45
55
96
239
0 50 100 150 200 250 300
70+ cm
50-70 cm
43-50 cm
10-43 cm
0-10 cm
CB
CB
wB
CA
p
Microbial biomass C, mg C kg-1
Table 1. Some soil physico-chemical properties for the four typical soil profiles under different land uses and
land covers
Particle size (%) Horizon Depth
(cm) Sand Silt Clay Class
pH EC
(dS m-1
)
O.M
(%)
CaCO3
(%)
Land use type: Natural forest (Pinus sylvestris L.) / Soil class: Lithic Xerorthent
A 0-13 58 25 17 SL 6,68 1,63 6,55 0.72
C1 13-20 58 27 15 SL 6,83 1,45 1,84 0.61
C2 20-38 59 30 11 SL 6,24 1,19 1,30 0.15
R 38+ - - - - - - - -
Land use type: Natural forest (Abies nordmanniana) / Soil class: Typic Dystroxerept
A1 0-28 47 28 25 SCL 6,03 1,11 6,24 0.45
Bw1 28-38 48 38 13 L 6,24 1,19 5,21 0.40
Bw2 38-58 51 30 19 L 6,67 1,25 1,96 0.45
C1 58-70 72 7 22 SCL 6,66 1,12 0,96 0.41
C2 70 + 73 4 23 SL 6,72 1,14 0,52 0.53
Land use type: Pasture area / Soil class: Typic Dystroxerept
A1 0-20 41 34 25 L 6,13 1,45 1,81 0.91
A2 20-46 47 36 17 L 6,28 1,36 1,55 0.96
Bw1 46-80 45 28 27 L 6,60 1,23 0,69 0.61
Bw2 80-100 34 26 40 CL 6,78 1,21 0,61 0.00
C 100+ 55 27 18 SL 7,46 1,23 0,56 0.46
Land use type: Cultivated area / Soil class: Typic Haploxerept
Ap 0-10 45 26 30 L 6,94 1,88 2,81 0.40
Ad 10-43 31 28 41 L 7,46 1,76 1,55 1.45
Bw 43-50 37 24 39 CL 7,71 1,75 0,69 3.62
BC 50-70 43 25 32 CL 7,09 1,68 0,61 2.53
C 70+ 39 24 37 L 7,48 1,47 0,56 1.57
EC: Electric conductivity, OM: Organic Matter.
Figure 3. Soil microbial biomass C of the three different adjacent land use types
R. Kızılkaya et al.
Changes of soil microbial biomass C and basal soil respiration in different land...
1044
NATURAL FOREST LAND (Pinus sylvestris L.)
Soil class: Lithic Xerorthent
2198
1945
785
0 500 1000 1500 2000 2500
20-38 cm
13-20 cm
0-13 cm
C2
C1
A
Basal soil respiration, µµµµg CO2-C g-1
NATURAL FOREST LAND (Abies nordmanniana )
Soil class: Typic Dystroxerept
233
298
732
1785
1974
0 500 1000 1500 2000 2500
70 + cm
58-70 cm
38-58 cm
28-38 cm
0-28 cm
C2
C1
Bw
2B
w1
A1
Basal soil respiration, µµµµg CO2-C g-1
PASTURE LAND
Soil class: Typic Dystroxerept
96
148
337
567
1534
0 500 1000 1500 2000
100+ cm
80-100 cm
46-80 cm
20-46 cm
0-20 cm
CB
w2
Bw
1A
2A
1
Basal soil respiration, µµµµg CO2-C g-1
CULTIVATED LAND
Soil class: Typic Haploxerept
67
188
545
673
785
0 200 400 600 800 1000
70+ cm
50-70 cm
43-50 cm
10-43 cm
0-10 cm
CB
CB
wB
CA
p
Basal soil respiration, µµµµg CO2-C g-1
Soil microbial biomass C was affected significantly by three different adjacent land use
types.The microbial biomass C content of intercropped three different adjacent land use
soils (forest, pasture and cultivated land) ranged from 239 to 786 mg C kg-1
soil and
increased significantly from cultivated area to the natural forest areas (Figure 3). The highest
microbial biomass C content was found under the natural forest lands whereas cultivated
soil has the lowest value. There were no significant differences between Pinus sylvestris L.
and Abies nordmanniana.
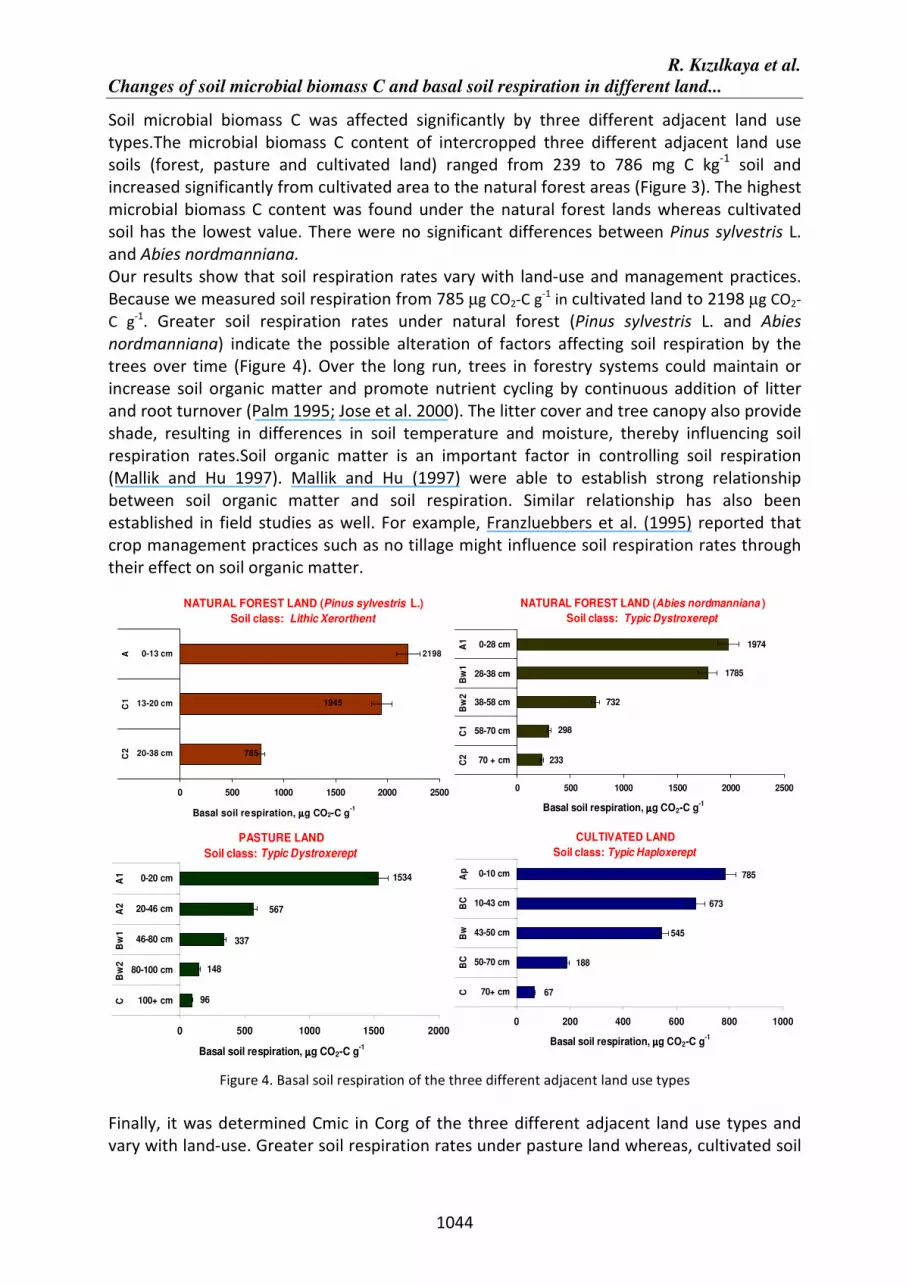
Our results show that soil respiration rates vary with land-use and management practices.
Because we measured soil respiration from 785 µg CO2-C g-1
in cultivated land to 2198 µg CO2-
C g-1. Greater soil respiration rates under natural forest (Pinus sylvestris L. and Abies
nordmanniana) indicate the possible alteration of factors affecting soil respiration by the
trees over time (Figure 4). Over the long run, trees in forestry systems could maintain or
increase soil organic matter and promote nutrient cycling by continuous addition of litter
and root turnover (Palm 1995; Jose et al. 2000). The litter cover and tree canopy also provide
shade, resulting in differences in soil temperature and moisture, thereby influencing soil
respiration rates.Soil organic matter is an important factor in controlling soil respiration
(Mallik and Hu 1997). Mallik and Hu (1997) were able to establish strong relationship
between soil organic matter and soil respiration. Similar relationship has also been
established in field studies as well. For example, Franzluebbers et al. (1995) reported that
crop management practices such as no tillage might influence soil respiration rates through
their effect on soil organic matter.
Figure 4. Basal soil respiration of the three different adjacent land use types
Finally, it was determined Cmic in Corg of the three different adjacent land use types and
vary with land-use. Greater soil respiration rates under pasture land whereas, cultivated soil

R. Kızılkaya et al.
Changes of soil microbial biomass C and basal soil respiration in different land...
1045
NATURAL FOREST LAND (Pinus sylvestris L.)
Soil class: Lithic Xerorthent
2,07
2,16
1,51
0,00 0,50 1,00 1,50 2,00 2,50
20-38 cm
13-20 cm
0-13 cm
C2
C1
A
Cmic in Corg, g. 100 g-1
NATURAL FOREST LAND (Abies nordmanniana )
Soil class: Typic Dystroxerept
1,49
1,01
0,86
1,07
1,93
0,00 0,50 1,00 1,50 2,00 2,50
70 + cm
58-70 cm
38-58 cm
28-38 cm
0-28 cm
C2
C1
Bw
2B
w1
A1
Cmic in Corg, g. 100 g-1
PASTURE LAND
Soil class: Typic Dystroxerept
1,14
1,27
2,22
2,93
4,55
0 1 2 3 4 5 6
100+ cm
80-100 cm
46-80 cm
20-46 cm
0-20 cm
CB
w2
Bw
1A
2A
1
Cmic in Corg, g. 100 g-1
CULTIVATED LAND
Soil class: Typic Haploxerept
0,89
1,27
1,37
1,07
1,47
0 0,5 1 1,5 2
70+ cm
50-70 cm
43-50 cm
10-43 cm
0-10 cm
CB
CB
wB
CA
p
Cmic in Corg, g. 100 g-1
has the lowest value. There were no significant differences between Pinus sylvestris L. and
Abies nordmanniana (Figure 5).
Figure 5. Cmic in Corg of the three different adjacent land use types
CONCLUSION In summary, some soil physico-chemical properties were affected modification of land use
and land covers. In addition soil respiration, soil organic matter, organic carbon and
microbial biomass have also been changed under different land uses and land covers. The
results of this study indicate, changing of land cover resulted in deterioration of soil
properties compared to soils under natural forest, pasture and cultivated lands. Organic
matter, organic C and microbial biomass were all reduced in cultivated land. At the same
time, specific maintenance respiration was greater on natural forest sites than cultivated
soils. Lee and Jose (2003) reported in their study that differences observed in soil properties,
including organic matter and microbial biomass, may have developed as a result of land-use
changes and management practices over time. It is clear that over time trees will
significantly increase the soil organic matter content and improve the soil biological activity
of agricultural fields. Thus, it is clear that deforestation caused adverse changes in soil
physico-chemical properties and microbiological quality which included microbial biomass
and activity. Since, microbiological parameters are indices of biological stability that
ultimately controls the sustainable fertility status of soil (Hart et al., 1989; Harris and Birch,
1990), decline in microbial parameters reveals poor unstable soil which may further lead to
development of degraded land that can neither support soil life forms nor any overlying
vegetation.

R. Kızılkaya et al.
Changes of soil microbial biomass C and basal soil respiration in different land...
1046
REFERENCES Anderson, J.P.E. 1982. Soil respiration. In. Methods of soil analysis, Part 2, Chemical and microbiological
properties, ed. A.L. Page, 831-871. Madison, Wisconsin: ASA-SSSA.
Anderson, J.P.E. and K.H. Domsch. 1978. A physiological method for the quantative measurement of microbial
biomass in soils. Soil Biology and Biochemistry, 10: 215 – 221.
Bouyoucos, G, J. 1962. Hydrometer method improved for making particle size analyses of soils, Argon. J. 54,
464–465.
Chander, K., Brookes, P.C. 1991. Effects of heavy metals from past applications of sewage sludge on microbial
biomass and organic matter accumulation in a sandy loam and silty loam U.K. soil. Soil Biology &
Biochemistry 23, 927–932.
Franzluebbers A.J., Hons F.M. and Zuberer D.A. 1995. Tillage and crop effect on seasonal dynamics of soil CO2
evolution, water content, temperature, and bulk density. Appl Soil Ecol 2: 95– 109.
Jaiyeoba, I.A. 2003. Changes in soil properties due to continuous cultivation in Nigerian semiarid Savannah. Soil
Till Res 70: 91-98.
Jose S., Gillespie R, Seifert J.R., Mengel D.B. and Pope P.E. 2000. Defining competition vectors in a temperate
alley cropping system in the Midwestern USA. 3. Competition for nitrogen and litter decomposition
dynamics. Agrofor Syst 48: 61–77.
Harris, J.A., Brich, P. 1990. The effects of civil engineering and stock-piling on the soil microbial community. In
Microbiology. In Civil Engineering, Howsan P, Spon FN (eds). Chapman and Hall: London; 247-287.
Hart, P.B.S., August, J.A., West, A.W. 1989. Long term consequence of top soil mining on biological and physical
characteristics of two New Zealand loessial soils undergrazed pasture. Land Degradation and Reliability 1:
77-88.
Islam, K.R., Kamaluddin, M., Bhuiyan, M.K., Badruddin, Abu., 1999. Comparative performance of exotic and
indigenous forest species for tropical semi-evergreen degraded forest land reforestation in Bangladesh.
Land Degrad. Dev. 10, 241–249.
Kang, B.T., Juo, A.S.R., 1986. Effect of forest clearing on soil chemical properties and crop performance. In: Lal,
R., Sanchez, P.A., Cummings, Jr., R.W. (Eds.), Land Clearing and Development in the Tropics. Belkema,
Rotterdam, pp. 383–394.
Killham, K., 1985. A physiological determination of the impact of environmental stress on the activity of
microbial biomass. Environmental Pollution. Series A 38, 283– 294.
Killham, K., Firestone, M.K., 1984. Salt stress control of intracellular solutes in streptomycetes indigenous to
saline soils. Applied and Environmental Microbiology 47, 301–306.
Lee, K.H and S. Jose. 2003. Soil respiration and microbial biomass in a pecan – cotton alley cropping system in
Southern USA. Agroforestry Systems 58: 45–54.
Lobe, I., Amelung, W. and C.C. Du Preez. 2001. Losses of carbon and nitrogen with prolonged arable cropping
from sandy soils of the South African Highveld. Europ. J. Soil Sci. 52, 93.
Mallik A.U. and Hu D. 1997. Soil respiration following site preparation treatments in boreal mixedwood forest.
For Ecol Manage 97: 265–275.
Mahtab, F.U., Karim, Z., 1992. Population and agricultural land use: towards a sustainable food production
system in Bangladesh. Ambio 21, 50–55.
Nardi, S., Cocheri, G., Dell’Agnola, G., 1996. Biological activity of humus. In: Piccolo, A. (Ed.), Humic Substances
in Terrestrial Ecosystems. Elsevier, Amsterdam, pp. 361–406.
Nelson, D.W., Sommers, L. E. 1982. Total carbon, organic carbon, and organic matter, In: Page, A.L, (Ed,),
Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties, second ed, Agronomy
Monographs, 9, ASA-SSA, Madison, WI, pp, 539–579.
Palm C.A. 1995. Contribution of agroforestry trees to nutrient requirements of intercropped plants. Agrofor
Syst 30: 105–124.
Powlson, D.S., Brookes, P.C., Jenkinson, D.S., 1987. Measurement of soil microbial biomass provides an early
indication of changes in total soil organic matter due to straw incorporation. Soil Biology & Biochemistry
19, 159–164.
Soil Survey Staff. 1992. Procedures for collecting soil samples and methods of analysis for soil survey, Soil Surv,
Invest, Rep, I, U,S, Gov, Print, Office, Washington D.C.
Wardle, D.A., 1992. A comparative assessment of factors which influence microbial biomass carbon and
nitrogen levels in soils. Biological Reviews 67, 321– 358.
Yan, T., Yang, L., C.D. Campbell. 2003. Microbial biomass and metabolic quotient of soils under different land
use in the Three Gorges Reservoir area. Geoderma 115; 129– 138.





























