Embed Size (px)
Citation preview

doi:10.1152/ajpheart.00221.2012 303:H256-H270, 2012. First published 25 May 2012;Am J Physiol Heart Circ Physiol
Hatzistergos and Joshua M. HareVasileios Karantalis, Wayne Balkan, Ivonne H. Schulman, Konstantinos E.myocardial remodelingCell-based therapy for prevention and reversal of
You might find this additional info useful...
for this article can be found at:Supplemental material/content/suppl/2012/08/01/ajpheart.00221.2012.DC1.html
188 articles, 61 of which can be accessed free at:This article cites /content/303/3/H256.full.html#ref-list-1
5 other HighWire hosted articlesThis article has been cited by
[PDF] [Full Text] [Abstract]
, January 24, 2013; .HeartElie R Chemaly, Roger J Hajjar and Larissa LipskaiaMolecular targets of current and prospective heart failure therapies
[PDF] [Full Text] [Abstract], March 21, 2013; .Perfusion
N Sikora, A Lacis, V Kasyanov, V Groma, E Ligere, V Ozolins and L Smitsarteriesinvestigation of biomechanical and structural properties of neonatal and adult coronary
an−The proper delivery pressure for cardioplegic solution in neonatal cardiac surgery
[PDF] [Full Text], April 15, 2013; 304 (8): H1045-H1049.Am J Physiol Heart Circ Physiol
William C. Stanley and Kara Hansell KeehanCirculatory Physiology
American Journal of Physiology-Heart andUpdate on innovative initiatives for the
[PDF] [Full Text] [Abstract], July , 2013; 28 (4): 357-367.Perfusion
N Sikora, A Lacis, V Kasyanov, V Groma, E Ligere, V Ozolins and L Smitsarteriesinvestigation of biomechanical and structural properties of neonatal and adult coronary
an−The proper delivery pressure for cardioplegic solution in neonatal cardiac surgery
[PDF] [Full Text] [Abstract], July 15, 2013; 99 (14): 992-1003.Heart
Elie R Chemaly, Roger J Hajjar and Larissa LipskaiaMolecular targets of current and prospective heart failure therapies
including high resolution figures, can be found at:Updated information and services /content/303/3/H256.full.html
can be found at:AJP - Heart and Circulatory Physiologyabout Additional material and information http://www.the-aps.org/publications/ajpheart
This information is current as of August 22, 2013.
ISSN: 0363-6135, ESSN: 1522-1539. Visit our website at http://www.the-aps.org/.Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2012 by the American Physiological Society. intact animal to the cellular, subcellular, and molecular levels. It is published 12 times a year (monthly) by the Americanlymphatics, including experimental and theoretical studies of cardiovascular function at all levels of organization ranging from the
publishes original investigations on the physiology of the heart, blood vessels, andAJP - Heart and Circulatory Physiology
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

Cell-based therapy for prevention and reversal of myocardial remodelingVasileios Karantalis,1 Wayne Balkan,1 Ivonne H. Schulman,1,2 Konstantinos E. Hatzistergos,1
and Joshua M. Hare1
1Interdisciplinary Stem Cell Institute, University of Miami Miller School of Medicine, Miami, Florida;and 2Nephrology-Hypertension Section, Miami Veterans Affairs Healthcare System, Miami, Florida
Submitted 15 March 2012; accepted in final form 21 May 2012
Karantalis V, Balkan W, Schulman IH, Hatzistergos KE, Hare JM. Cell-based therapy for prevention and reversal of myocardial remodeling. Am J PhysiolHeart Circ Physiol 303: H256–H270, 2012. First published May 25, 2012;doi:10.1152/ajpheart.00221.2012.—Although pharmacological and interventionaladvances have reduced the morbidity and mortality of ischemic heart disease, thereis an ongoing need for novel therapeutic strategies that prevent or reverse progres-sive ventricular remodeling following myocardial infarction, the process that formsthe substrate for ventricular failure. The development of cell-based therapy as astrategy to repair or regenerate injured tissue offers extraordinary promise for apowerful anti-remodeling therapy. In this regard, the field of cell therapy has mademajor advancements in the past decade. Accumulating data from preclinical studieshave provided novel insights into stem cell engraftment, differentiation, andinteractions with host cellular elements, as well as the effectiveness of variousmethods of cell delivery and accuracy of diverse imaging modalities to assesstherapeutic efficacy. These findings have in turn guided rationally designed trans-lational clinical investigations. Collectively, there is a growing understanding of theparameters that underlie successful cell-based approaches for improving heartstructure and function in ischemic and other cardiomyopathies.
myocardial infarction; stem cells; mesenchymal stem cells; bone marrow cells;heart failure
THIS ARTICLE is part of a collection on Physiological Basis ofCardiovascular Cell and Gene Therapies. Other articles ap-pearing in this collection, as well as a full archive of all collec-tions, can be found online at http://ajpheart.physiology.org/.
Introduction
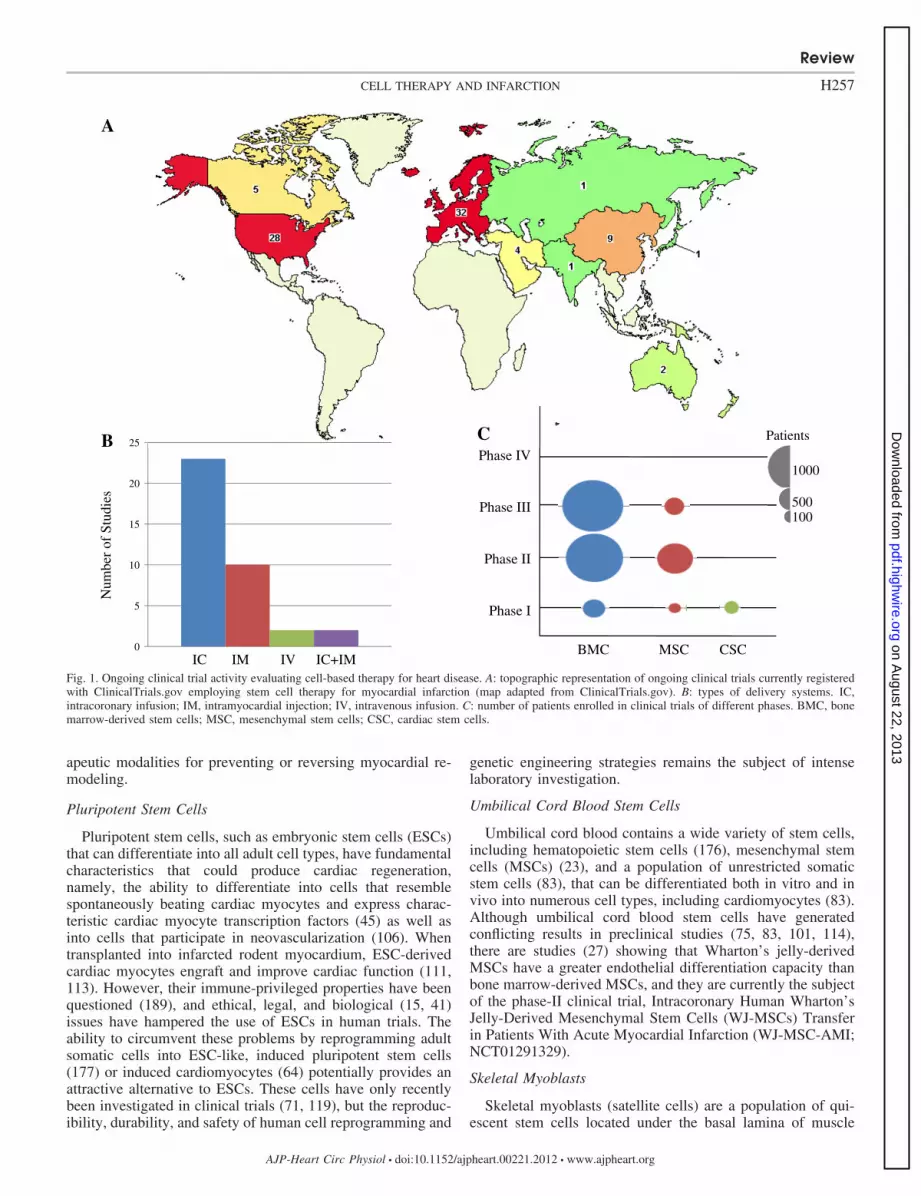
A recent update from the American Heart Association (140)reported that in 2008, 2.7% of Americans suffered a myocar-dial infarction (MI), �8 million survived an MI, and anestimated 82.6 million were living with one or more types ofcardiovascular disease, costing a staggering $300 billion thatyear. These statistics underscore the challenges facing ourhealth care system today. While advances in both pharmaco-logical and interventional approaches for revascularizationhave significantly reduced the morbidity and mortality ofischemic heart disease over the past half-century, they do notdirectly reverse the major reason for ventricular failure andelectrical system disturbance, the progressive ventricular re-modeling that follows an MI. Therefore, the need for noveltherapeutic modalities is paramount. In this regard, encourag-ing results from preclinical studies and phase I–III clinicaltrials (Fig. 1) suggest that stem cell therapy provides anti-remodeling effects leading to the restoration of contractilefunction in patients with ischemic cardiomyopathy.
With the discovery of cardiac stem cells (CSCs), the notionthat adult myocardial tissue has endogenous regenerative ca-pability (12) signaled the beginning of a new era in cardiovas-cular medicine. A stem cell is defined by its ability to self-renew and to differentiate into one or more lineages of spe-cialized cell types (151). In addition, stem cells are generallyorganized in niches where their interaction with other cell types(including other stem cells) and the stroma maintains their“stemness” (61, 110, 173). Characteristics of an ideal stem cellfor use in cardiac cell-based therapy are availability, tissueresidence or homing specifically to the injured tissue, ability tomobilize endogenous repair mechanisms that are otherwiseoverwhelmed by the magnitude of damage, and providing asustainable benefit that translates as prevention or reversal ofpathological remodeling and as improvement of left ventricular(LV) function. Previous and ongoing preclinical studies andclinical trials suggest that there are many stem cells that couldplay this role. Determining the most effective stem cell(s) andalso understanding the mechanisms by which they exert theirtherapeutic effects are the focus of current studies (96, 104).Each stem cell type, in theory, exhibits some of the character-istics that contribute to a capacity to exert a therapeuticresponse. As is typical in the evolution of new therapeuticstrategies, the ultimate proof derives from clinical testing, andin this regard, some cell therapeutic strategies have not pro-duced the hypothesized improvements in clinical endpoints(161). Nevertheless, in this early stage of stem cell therapy,both the successes and the failures are informative. This articlewill review the different types of stem cells and recent ad-vances and molecular mechanisms underlying cell-based ther-
Address for reprint requests and other correspondence: J. M. Hare, Biomed-ical Research Bldg., 1501 N.W. 10th Ave., Rm. 824, PO Box 016960 (R125),Miami, FL, 33101 (e-mail: [email protected]).
Am J Physiol Heart Circ Physiol 303: H256–H270, 2012.First published May 25, 2012; doi:10.1152/ajpheart.00221.2012.Review
http://www.ajpheart.orgH256
on August 22, 2013
pdf.highwire.org
Dow
nloaded from
apeutic modalities for preventing or reversing myocardial re-modeling.
Pluripotent Stem Cells
Pluripotent stem cells, such as embryonic stem cells (ESCs)that can differentiate into all adult cell types, have fundamentalcharacteristics that could produce cardiac regeneration,namely, the ability to differentiate into cells that resemblespontaneously beating cardiac myocytes and express charac-teristic cardiac myocyte transcription factors (45) as well asinto cells that participate in neovascularization (106). Whentransplanted into infarcted rodent myocardium, ESC-derivedcardiac myocytes engraft and improve cardiac function (111,113). However, their immune-privileged properties have beenquestioned (189), and ethical, legal, and biological (15, 41)issues have hampered the use of ESCs in human trials. Theability to circumvent these problems by reprogramming adultsomatic cells into ESC-like, induced pluripotent stem cells(177) or induced cardiomyocytes (64) potentially provides anattractive alternative to ESCs. These cells have only recentlybeen investigated in clinical trials (71, 119), but the reproduc-ibility, durability, and safety of human cell reprogramming and
genetic engineering strategies remains the subject of intenselaboratory investigation.
Umbilical Cord Blood Stem Cells
Umbilical cord blood contains a wide variety of stem cells,including hematopoietic stem cells (176), mesenchymal stemcells (MSCs) (23), and a population of unrestricted somaticstem cells (83), that can be differentiated both in vitro and invivo into numerous cell types, including cardiomyocytes (83).Although umbilical cord blood stem cells have generatedconflicting results in preclinical studies (75, 83, 101, 114),there are studies (27) showing that Wharton’s jelly-derivedMSCs have a greater endothelial differentiation capacity thanbone marrow-derived MSCs, and they are currently the subjectof the phase-II clinical trial, Intracoronary Human Wharton’sJelly-Derived Mesenchymal Stem Cells (WJ-MSCs) Transferin Patients With Acute Myocardial Infarction (WJ-MSC-AMI;NCT01291329).
Skeletal Myoblasts
Skeletal myoblasts (satellite cells) are a population of qui-escent stem cells located under the basal lamina of muscle
Fig. 1. Ongoing clinical trial activity evaluating cell-based therapy for heart disease. A: topographic representation of ongoing clinical trials currently registeredwith ClinicalTrials.gov employing stem cell therapy for myocardial infarction (map adapted from ClinicalTrials.gov). B: types of delivery systems. IC,intracoronary infusion; IM, intramyocardial injection; IV, intravenous infusion. C: number of patients enrolled in clinical trials of different phases. BMC, bonemarrow-derived stem cells; MSC, mesenchymal stem cells; CSC, cardiac stem cells.
Review
H257CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

fibers that proliferate in response to injury to help regeneratemuscular tissue (19). Although they have limited differentia-tion ability, skeletal myoblasts, based on the concept that thesecells would have sufficient plasticity for cardiac myocytedifferentiation, have been investigated as a potential therapeu-tic approach for myocardial regeneration (1, 53, 162). Studiesindicate that skeletal myoblasts are unable to couple electro-mechanically with the endogenous cardiomyocytes, and thisdeficit is likely the basis of the detrimental ventricular tachy-arrhythmias observed in early studies (154, 156). Thus, despitemore than 40 preclinical studies, questions regarding the safetyand efficacy of these cells remain (154, 156), and there isrelatively less enthusiasm for this cell type compared withother alternatives.
Bone Marrow-Derived Stem Cells
There is intense interest in the bone marrow as a source ofcells with capacity for cardiac repair, and bone marrow hasbeen widely tested in mechanistic studies and clinical trialsfor cardiac disease. Bone marrow harbors a wide variety ofstem/precursor cells with potential for cardiac differentia-tion. Some, but not all, studies support the idea that bonemarrow-derived stem/progenitor cells (BMSCs) (20) havethe capacity to differentiate into cardiac myocytes andendothelial cells (123, 124). These characteristics and therelative accessibility of bone marrow have supported thehypothesis that autologous BMSCs and/or its stem/progenitor cell populations could produce phenotypic andclinical recovery in patients with remodeled ventricles.Preclinical studies have demonstrated that whole bone mar-row (8, 65, 70, 118, 123) improves hemodynamic parame-ters and cardiac morphology. Kamihata et al. (70) showedthat whole bone marrow improved myocardial perfusion andreduced infarct size in a swine model. In addition, the newlyformed capillaries, but not the fibroblasts, in the infarct areawere derived from donor BMSCs, supporting the notion thatBMSC therapy produces a proangiogenic rather than afibrosis-inducing microenvironment.
Endothelial progenitor cells and mononuclear cells. In ad-dition to whole bone marrow, numerous studies have beenconducted examining specific constituents of bone marrow.Endothelial progenitor cells (EPCs) share certain phenotypicand functional characteristics with fetal angioblasts (80) andexpress the surface markers CD34, CD133 (5), Flk-1 [vascularendothelial growth factor (VEGF) receptor-2] (128), andAC133 (hematopoietic stem cell marker not expressed onmature endothelial cells) (51). In preclinical studies, EPCspromote neovascularization, leading to the prevention of LVremodeling and to some degree of cardiomyocyte regeneration(147). Although several phase-1 clinical trials have reportedpreliminary feasibility and safety data with EPC therapy afterMI, most of these studies actually investigated bone marrow-derived mononuclear cells (MNCs), not isolated EPCs (150).In the Transplantation of Progenitor Cells and RegenerationEnhancement in Acute Myocardial Infarction (TOPCARE-AMI) trial (91), patients with acute MI received an intracoro-nary infusion of ex vivo expanded bone marrow MNCs orculture-enriched EPCs derived from peripheral blood MNCs.With this strategy, there was sustained improvement in LVfunction, reduction in end-systolic volume, and prevention of
remodeling one year later in both the bone marrow MNCs andculture-enriched circulating EPC treatment groups. Given thepromising findings of the TOPCARE-AMI trial, additionalclinical investigations are underway. Notably, the EnhancedAngiogenic Cell Therapy-Acute Myocardial Infarction Trial(ENACT-AMI) is a phase-II clinical trial that will investigatethe efficacy and safety of autologous EPCs and autologousEPCs transfected with human endothelial nitric oxide synthase(NCT00936819).
While there is substantial evidence for the safety of bonemarrow MNCs from clinical trials, there are inconsistent find-ings as to their efficacy. The TOPCARE-AMI (91) and theBOne marrOw transfer to enhance ST-elevation infarct regen-eration (BOOST) trials (144) recently completed their 5-yearfollow-up. These trials, which together totaled 119 patients,illustrated the long-term safety of this cell-based therapy ap-proach. In TOPCARE-AMI, the improvement in LV ejectionfraction (LVEF) in the treated group was sustained, but resultsfrom BOOST showed that the early (6 mo) improvement inLVEF in the treated group was not sustained at 5 years. Theseinconsistent findings emphasize the importance of examiningin future trials the long-term sustainability of cell-based ther-apy.
Furthermore, the First Mononuclear Cells injected in theUnited States conducted by the Cardiovascular Cell TherapyResearch Network (FOCUS-CCTRN) phase-2 trial evaluatedwhether transendocardial delivery of bone marrow MNCsimproves LV performance and perfusion at 6 mo in patientswith chronic ischemic cardiomyopathy (130). Although therewas no improvement compared with placebo in LV end-systolic volume, maximal oxygen consumption, or myocardialperfusion, exploratory analyses showed significant improve-ment in LVEF and stroke volume. Interestingly, the improve-ment in ejection fraction was associated with higher bonemarrow CD34� and CD133� progenitor cell counts. Theseresults suggest that the cellular composition of the bone mar-row may determine clinical end points, with certain cell pop-ulations providing a greater regenerative benefit.
MSCs for ischemic heart disease. MSCs comprise �0.01%of bone marrow and are operationally defined as plastic-adherent, hematopoietic lineage negative cells of the MNCfraction of the bone marrow (133). MSCs have also beenisolated from virtually every tissue type (33) and are thought toparticipate in niches, regulating the function of hematopoieticcells (26) and other specific lineages (40). Interestingly, MSCsalso differentiate into bone, tendon, cartilage, fat, and muscle,raising the possibility of their use in disorders of organ systemscomprising those tissues (26). While these cells express nu-merous cell surface markers, including CD105, CD73, andCD90 (190), no single cell surface marker indisputably char-acterizes MSCs. Furthermore, MSCs may lose the expressionof certain surface markers or even acquire the expression ofnew ones as they are isolated and expanded in vitro (67).Therefore, MSCs are not a homogenous population of cells(120) and whether MSCs from different tissues are biologicallyequivalent remains to be determined (191).
Multilineage differentiation potential has become one of thedefining characteristics of MSCs (7). Under specific cultureconditions in vitro, these cells can differentiate into osteocytes,chondrocytes, adipocytes, smooth muscle cells, fibroblasts,endothelial cells, (132) and cardiomyocyte-like cells (102).
Review
H258 CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

Caplan (21) showed that differentiation depends on the cellularmilieu. For example, bone marrow-derived MSCs (BM-MSCs)cultured in the presence of 5-azacytidine differentiate intomultinucleated myotubes that express �-myosin heavy chain,desmin, and �-cardiac actin and also display spontaneousrhythmic calcium fluxes (180). Murine BM-MSCs culturedunder the same conditions started beating spontaneously, con-nected to adjoining cells, and became synchronized after 2 to 3wk (102). Those cells adopted two types of action potentials,sinus node-like and ventricular myocyte-like, and exhibitedcardiomyocyte-specific gene expression (atrial natriuretic pep-tide, �- and �-myosin heavy chain, Nkx2–5, and GATA4)(102). Rat BM-MSCs cocultured with neonatal rat ventric-ular myocytes differentiated into cardiomyocytes that con-tracted, expressed sarco(endo)plasmic reticulum calcium-ATPase 2 and ryanodine receptor 2 and were positive fortroponin T, �-actinin, and desmin (98). Another factor thatmay promote cardiomyocyte differentiation is mechanicalloading (131). Several studies have revealed two mainregulatory pathways of MSCs differentiation, the Wnt (17,37, 44) and the transforming growth factor (TGF)-� path-ways (4, 7), but there is a plethora of other molecules thathave been shown to play a regulatory role, such as epider-mal growth factor, platelet-derived growth factor, and fibro-blast growth factor (108, 165).
Animal studies have established that MSCs are capable ofdifferentiating into cardiomyocytes and/or vascular structuresin both allogeneic (61, 145) and xenotransplantation (166)models, resulting in significant functional improvement andreduction of ischemic scar area. Interestingly, animals sub-jected to allogeneic MSC transplantation did not manifestevidence of rejection (3, 141), consistent with observations byseveral groups that MSCs exhibit immune-privileged proper-ties in vitro and in vivo (56), likely because of the absence ofMHC-II, B-7 costimulatory molecules, and CD40 ligand (170).The lack of costimulatory molecules prevents T-cell responsesand also induces an immunosuppressive local microenviron-ment through the production of prostaglandins and other sol-uble mediators such as nitric oxide, indoleamine 2,3-dioxyge-nase, and heme oxygenase-1 (24, 79, 84, 138, 143). MSCsreduce the respiratory burst that follows neutrophilic responsesby releasing interleukin-6 (136). They also inhibit the differ-entiation of immature monocytes into dendritic cells, hence theantigen presentation to naïve T cells is greatly impaired (66). Inaddition, MSCs release soluble factors, such as hepatocytegrowth factor and TGF-�1 (39) that suppress the proliferationof cytotoxic and helper T cells.
The potential clinical application of MSCs was exploredearly in the developing field of cell-based therapy. Chen et al.randomized 69 patients post-MI to receive intracoronary ad-ministration of either autologous MSCs or placebo. They usedsingle positron emission computer tomography and detected asignificant improvement in LV function and perfusion defect,suggesting that MSC therapy induces regeneration of infarctedmyocardium and protects against LV remodeling (28). Wecompared the therapeutic benefit of intravenous administrationof allogeneic MSCs to placebo in 60 patients after acute MI(60). In addition to establishing the safety of allogeneic MSCdelivery to humans, echocardiography revealed a 6% increasein LVEF at 3 mo for MSC-treated patients. In addition, patientsreceiving MSCs had improved quality of life, better respiratory
function, and a near eradication of ventricular arrhythmias.Recently, Williams et al. (179) reported the results from thefirst eight patients of the Transendocardial Autologous Cells inIschemic Heart Failure Trial (TAC-HFT), in which autologousBMSCs (either culture expanded MSCs or MNCs) were in-jected intramyocardially into patients with chronic ischemiccardiomyopathy. These patients demonstrated improved re-gional myocardial contractility and decreased infarct size.Notably, early improvements in regional function predicted thedegree of reverse remodeling 12 mo following cell therapy.These early investigations have engendered a plethora of on-going trials testing the capacity of MSCs or MSC precursorsfor myocardial regeneration. Importantly, the findings fromthese studies have also indicated that the selection of endpoints, such as direct measures of infarct size and ventricularremodeling, that accurately reflect clinical outcomes couldrepresent more suitable measures of efficacy for cell therapy(161, 179) than LVEF, which most cell therapy trials have usedas the primary end-point marker of efficacy.
As previously mentioned, MSCs have the capacity to beused as an allogeneic cell therapeutic. To test whether alloge-neic MSCs are as safe and effective as autologous MSCs, ourgroup is conducting The Percutaneous Stem Cell Injection DeliveryEffects on Neomyogenesis Pilot Study (The POSEIDON-PilotStudy; NCT01087996), the first direct randomized head-to-head comparison of autologous versus allogeneic MSCs deliv-ered by transendocardial, intramyocardial injection. This studywill help determine whether or not allogeneic MSCs, a cost-effective “off the shelf” cell product, are equally effective as atherapeutic regimen as autologous MSCs and will also definethe relative advantages of the autologous versus allogeneicapproach in various patient populations.
MSCs for nonischemic heart disease. Although the majorityof cardiac cell-based therapy studies have targeted acute MIand ischemic cardiomyopathy, it is important to point out thata significant number of heart failure patients have nonischemiccardiomyopathy. Idiopathic dilated cardiomyopathy (DCM) isa leading cause of heart failure morbidity and mortality andaccounts for �50% of heart transplants (163). The primaryphenotype of DCM is an enlarged, remodeled ventricle withincreased end-diastolic and end-systolic volumes, reducedejection fraction, and impaired contractile and diastolic func-tion. Given that cell-based therapy produces durable and sus-tainable improvements in cardiac function and causes reverseremodeling in ischemic cardiomyopathy, it is attractive tohypothesize the same for patients with nonischemic cardiomy-opathies. If these effects can be clinically established andoptimized, there is enormous potential for improving clini-cal outcomes for the many patients suffering from DCM. Acase report and small clinical study on the safety andfeasibility of cell therapy for DCM (86, 109) were encour-aging and indicate that larger scale randomized trials arewarranted. Accordingly, we have initiated a clinical trial toestablish the safety and efficacy of bone marrow-derivedMSC therapy for patients with DCM. The Percutaneous Stem CellInjection Delivery Effects on Neomyogenesis-Dilated Cardiomy-opathy (POSEIDON-DCM; NCT01392625) study is a random-ized comparison of autologous versus allogeneic MSC therapyemploying percutaneous delivery with a transendocardial cath-eter delivery system.
Review
H259CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

Cardiac Stem Cells
The realization that adult myocardial tissue(s) have endog-enous regenerative capability (12) dramatically shifted thelong-held view that the heart is a terminally differentiatedorgan without regenerative capability. It has now becomeevident that there are myocytes in the heart expressing the cellcycle proteins CDC6, Ki67, MCM5, serine 10-phosphorylatedhistone H3 (PH3; Fig. 2B), and aurora B kinase, indicating thatthese cells can replicate (11). The rate of replication of thesemyocytes increases substantially in pathological states (93).Two possibilities could account for the presence of these cells:replicating CSCs (Fig. 2C) evolving into transient amplifyingcells or mature myocytes that have dedifferentiated and reen-tered the cell cycle (14, 43, 85). A landmark study by Beltramiet al. (12) was the first to challenge the dogma that the adultmyocardium lacks regenerative capacity. The authors reporteda population of cardiac Lin� c-kit� cells possessing the prop-erties of CSCs. These cells gave rise to cardiomyocytes,smooth muscle cells, and endothelial cells; indicating theirability to undergo trilineage differentiation, reconstitute myo-cardium, and form new blood vessels and cardiomyocyteswhen injected into an ischemic heart. Accordingly, this studyestablished that the adult mammalian heart does indeed containa compartment of stem cells.
CSCs are a diverse cell population. They are approximatelyone-tenth the size of mature cardiac myocytes and express thestem cell factor receptor c-kit (CD117) (12), stem cell antigen1 (Sca-1; an antigen present in rodents but not mammals)(122), and in some cases the transcription factor Islet-1 (89),which in neonatal hearts marks the second heart field. Islet-1 isalso reported in cardiosphere-derived cells (discussed below)derived from postnatal hearts (112). Hierlihy et al. (62) iden-tified a side population of cells that form colonies in vitro(semisolid media) and differentiate into cardiomyocytes. Inter-estingly, these cells were found to be activated when growth ofthe postnatal murine heart was attenuated, through overexpres-sion of a dominant negative cardiac transcription factor
(MEF2C), and subsequently depleted. These findings weresuggestive of a responsive CSC pool in the adult myocardium.Additionally, Martin et al. (107) detected another side popula-tion of cells that are capable of proliferating and differentiatinginto cardiac and hematopoietic lineages in vitro. These cellswere identified based on their expression of Abcg2, an ATP-binding cassette transporter.
Another source of CSCs may emerge from studying theintegral role of the epicardium in heart development (158),which in lower vertebrates (68, 74) participates in cardiacregeneration. Although several groups have studied the pres-ence of c-kit� cells in the epicardium with variable results(100, 184), there is now mounting evidence for the presence ofepicardial progenitor cells that resemble MSCs (29). FollowingMI, these cells undergo endothelial-to-mesenchymal cell trans-formation (22). The acquisition of a mesenchymal phenotypeand subsequent migration from the subepicardial space into themyocardial wall is followed by proliferation and formation ofmyocardial precursors and vascular cells (22, 38, 100). Fur-thermore, MI triggers the activation of fetal epicardial genes(Tbx18, WT1, and retinaldehyde dehydrogenase type 2) (99).Limana et al. (99) demonstrated in infarcted murine hearts thatepicardial progenitor cells coexpress c-kit and WT1 and/orTbx18, suggesting that these cell populations are epicardialcardiogenic precursors.
The identification of progenitor cells in the epicardium withESC characteristics suggests that signaling factors that modu-late embryonic epicardial cell functions may also contribute tothe activation of adult epicardial progenitors. Several growthfactors that contribute to coronary vessel formation [erythro-poietin, VEGF, fibroblast growth factor (FGF), thymosin �4]have been administered in animal models of MI and heartfailure resulting in enhanced vascularization (2, 90, 139, 171).Smart et al. (155) studied cardiac-specific thymosin �4 knock-down mice and demonstrated that the loss of thymosin �4 mayresult in impaired paracrine signaling to the epicardium, lead-ing to impaired coronary vascular formation. Furthermore, it
Fig. 2. Mechanisms of cardiac regeneration. Mammalian myocyte renewal may result from stem cell differentiation and/or myocyte mitosis. These processes arelikely linked in that CSCs become mature myocytes by becoming transient amplifying cells, a process enhanced by MSCs. A: Masson’s trichrome (Masson’sTri) histological stain obtained from the border zone between infarcted and viable myocardium in the porcine heart. In this image viable new myocytes are shownin red and scarred myocardium in blue. Immunohistochemical analysis for the mitotic marker serine 10 phosphorylated histone H3 (PH3; white nucleus) in Band c-kit (green) in C suggests regeneration of new myocardium from replicating myocytes and adult c-kit� CSCs, respectively. The relative contribution of CSCsor adult dedifferentiation to myocyte entry into the cell cycle is not definitively defined. The spatial and temporal colocalization of c-kit cells and the PH3myocytes in an animal treated with MSCs is suggestive that the mitotic cells represent transient amplifying cells (61, 94).
Review
H260 CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

has been shown that systemic administration of thymosin �4 inmice after MI results in decreased cardiomyocyte death (159)and reactivation of the embryonic program, as demonstrated bythe epicardial increased expression of VEGF, Flk-1, FGF17,FGF receptor-2, FGF receptor-4, TGF-� and �-catenin (16).
CSCs are likely supported and maintained within cardiactissue in specialized microenvironments termed “stem cellniches” (48, 115). Niches are located in the myocardial inter-stitium where CSCs can maintain their “stemness” by formingconnections with other stem and supporting cells, i.e., maturecardiomyocytes and fibroblasts, through connexins and cad-herins. Niches have been detected throughout the myocardiumand are concentrated at the atria and apex (172), and it is fromwithin these niches that endogenous stem cells proliferate andthen migrate to the injury site after an ischemic incident (122).CSCs and side population cells most likely mediate endoge-nous mechanisms for minor repair and for replacement ofongoing cell turnover within the adult heart. The endogenousrepair appears to be limited to noninfarcted tissue (13) and mayattenuate the progression to heart failure. However, significantcellular injury, as inflicted by an MI, could overwhelm theregenerative ability and contribute to stem cell and nichedepletion or destruction (94).
It has been proposed that cardiospheres mimic cardiacniches (95). Cardiospheres contain a central cluster of c-kit�
cells, layers of differentiating cells expressing myocyte pro-teins and the gap junction protein, connexin 43, and an externallayer of mesenchymal cells. When transplanted into murine-injured hearts, cardiospheres exhibit enhanced engraftment andimproved myocardial function compared with cells grown in amonolayer. This difference may in part be due to the presenceof connexin-43 gap junctions. Connexin 43 plays a differenti-ation-dependent role. In more differentiated cardiomyocytes,connexin 43 promotes cell-cell electrical coupling (72), whereas inless mature cells it promotes proliferation and differentiation(55, 125). Dissociation of cardiospheres into single cells de-creased the expression of adhesion molecules and underminedresistance to oxidative stress, thereby reducing their ability toengraft and their functional benefit (95). On the other hand, Yeat al. (182) isolated and cloned Sca-1�CD45� cardiosphere-derived cells and injected them into post-MI mice hearts. Theyobserved that cloned Sca1�CD45� cells induced angiogenesis,differentiated into endothelial and smooth muscle cells, andimproved cardiac function. Interestingly, the expression of thetranscription factor Islet-1 was threefold higher in theSca1�CD45� cells than in whole cardiospheres. Islet-1 isexpressed in a cell population that gives rise to both myocardialand vascular tissues (89). These studies suggest that cardio-spheres provide a niche-like environment that helps resistoxidative stress and maintain “stemness,” whereas isolation ofcells from cardiospheres promotes their differentiation andlineage commitment. Further studies are needed to elucidatethe mechanisms involved in the balance between resistance tooxidative stress and relevant lineage commitment.
Clinical trials employing CSCs. CSCs can be harvested frompatient cardiac biopsies and expanded ex vivo to generate largenumbers of autologous cells (10), which can then be deliveredback to the patient. Recently published results from the phase-Iclinical trial Cardiac Stem Cell Infusion in Patients WithIschemic CardiOmyopathy (SCIPIO) (18) demonstrated thatintracoronary infusion of autologous c-kit� CSCs is effective
at improving LV systolic function and reducing infarct size inpatients with heart failure after MI. These results are veryencouraging and provide rationale for larger randomized trialsthat will extend these observations to test whether clinicalbenefits derive in patients receiving infusions of these cells.
A phase-I randomized clinical trial of cardiospheres as acell-based therapeutic was also recently completed. The CAr-diosphere-Derived aUtologous stem CElls to reverse ventricU-lar dySfunction (CADUCEUS) trial (103) included 25 patientswho had suffered an MI and who were randomized to receivecardiosphere-derived cells (by intracoronary infusion) or stan-dard care. Magnetic resonance imaging (MRI) analyses 6 moafter treatment with cardiospheres showed a reduction in scarmass and an increase in viable heart mass, regional contractil-ity, and regional systolic wall thickening. In contrast to thestudy of culture-expanded c-kit� CSCs, cardiospheres did notaugment parameters of integrated cardiac performance such asLVEF.
Mechanisms of Action of Cell-Based Therapy
The initial motivation for stem cell research and cell-basedtherapy was the idea that the injection of stem cells into ascarred myocardium would promote regeneration throughtransdifferentiation of the exogenously delivered stem cells, ahypothesis supported by Orlic et al. (123). However, thetotality of ongoing mechanistic and translational studies hashighlighted the fact that the therapeutic process is a multifac-eted, orchestrated process involving multiple mechanisms ofaction.
The in vivo mechanism of action of MSCs is under intenseinvestigation in an effort to shed light on the multifacetednature of cell-based stimulation of cardiac repair (3, 135, 166,181). MSCs engraft and differentiate when transplanted in theheart (61, 135). In this regard, Toma et al. (166) used immu-nofluorescence staining with anti-�-galactosidase to identifyengrafted MSCs (labeled with lacZ) intramyocardially deliv-ered to murine hearts. Engrafted MSCs expressed markers ofmyocardial differentiation as early as 14 days postinjection.Building on these early results, many studies followed inrodent (149) as well as swine models (3, 61, 145, 185). Thestudy by Quevedo et al. (135), using a swine model of ischemiccardiomyopathy, showed that 14% of engrafted MSCs demon-strated evidence of cardiomyocyte differentiation. In addition,there was evidence of cell-to-cell coupling between thesedifferentiated MSCs and host myocytes via connexin-43 gapjunctions. In a swine model of acute MI, a combination of astatin and intramyocardial injections of BM-MSCs improvedcardiomyocyte-differentiation capability by fourfold (181).However, not all studies show in vivo differentiation of MSCs.For example, MSC differentiation was not evident in a caninemodel by Silva and colleagues (152), revealing potential dif-ferences between species and/or study designs.
Despite the evidence for differentiation of engrafted MSCs(69, 105, 135), the collective results from studies of MSCshave called into question the correlation between the frequencyof engraftment and differentiation of transplanted MSCs andthe reported functional recovery early posttransplantation. Forinstance, reports of LV functional improvement within 72 hpostinjection cannot be explained by MSC transdifferentiation(117). Based on these observations, some investigators ques-
Review
H261CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

tioned whether differentiation occurs at all, raising the theorythat cell fusion with resident myocytes could explain, at least inpart, cardiac recovery (121). On the other hand, the fact that ofhundreds of millions of cells fewer than 10% remain after 2 wkhas lent credence to the paracrine signaling hypothesis (54),whereby exogenous MSCs secrete factors that induce neovas-cularization and prevent cardiomyocyte apoptosis, thereby re-ducing remodeling and improving cardiac function (80, 147).However, the observation of long-term MSC survival, engraft-ment, and trilineage differentiation and that MSC engraftmentcorrelated with functional recovery in contractility and myo-cardial blood flow in a swine model of chronic ischemiccardiomyopathy (105) indicate that stem cells do provide morethan simply paracrine support for repair. Moreover, otherinvestigators have suggested that it may simply be the sheer
mechanical strengthening of the MI scar that prevents furtherdeterioration in cardiac function (121). It is likely that thecombination of short- and long-term effects of MSCs maintainsthe process of reverse remodeling.
With regard to the paracrine signaling hypothesis, variousstudies indicate that MSCs secrete proangiogenic, antifibrotic,and antiapoptotic factors, such as VEGF, stromal-derived fac-tor-1 (77), angiopoietin 1 (73), insulin-like growth factor 1(57), hepatocyte growth factor, periostin (76), and thymosin b4(63). Amado et al. (3) reported that MSCs express VEGF,which is linked to both neoangiogenesis and stem cell homingand migration, but also provided strong evidence of cell dif-ferentiation.
Perhaps the most important insight into the mechanism ofaction derives from studies of cell-cell interactions. In a study
Fig. 3. MSCs stimulate endogenous cardiomyocyte cell (CM) cycling. A and B: quantification of newly formed myocytes of both host (red bar graph,phospho-H3�) and donor (green bar graph, Y chromosome�) origin 2- and 8-wk postinjection, respectively. MSCs stimulated host cardiomyocytes to amplifyduring the first 2 wk following transendocardial injection. The new CMs were mainly distributed at the ischemic zone (IZ) and border zone (BZ) of the treatedhearts, indicating active regeneration of injured myocardium. By 8 wk, endogenous cycling CMs levels had returned to normal values. C and D: mitotic featuresin endogenous CMs from the BZ of an MSC-treated and cell-conditioned media (CCM)-treated heart, respectively. MLC, myosin light chain; DAPI,4,6-diamidino-2-phenylindole. Values are means � SE (n � 3 hearts, each). *P � 0.05, †P � 0.005 between groups; ‡P � 0.05 between groups. Reproducedwith permission from Hatzistergos et al., Circulation Research, 2010 (61).
Review
H262 CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

comparing MSC conditioned medium versus MSCs them-selves, Hatzistergos et al. (61) demonstrated in a swine modelof MI that intramyocardial injection of MSCs reduced infarctsize by recruiting endogenous c-kit� CSCs, whereas an injec-tion of concentrated MSC-conditioned media had no effect. Inthis study, histological examination revealed chimeric clusterscomprised of adult cardiomyocytes, transplanted MSCs, andhost c-kit� CSCs that expressed connexin-43-mediated gapjunctions and N-cadherin mechanical connections betweencells. Importantly, MSC treatment enhanced substantially theabundance of host c-kit� CSCs and their cardiomyocyte lin-eage commitment capacity. In addition, MSCs and CSCsappeared to create cell clusters closely resembling stem cellniches (Fig. 3). The latter underscores the emerging conceptthat MSCs may act both as progenitors for certain cell lineagesand, through their participation in niches, as supporting/regu-latory cells for other lineages (94). All of the aforementionedmechanistic insights (Fig. 4) allow us to generate a novelhypothesis that cell therapy reconstitutes endogenous stem cellniches—by way of transdifferentiation and paracrine signal-ing—and in that manner facilitate cardiac self-healing.
Timing of Delivery
It is recognized that practical issues such as timing, deliverymethod, and dosing could contribute substantially to the out-come of clinical investigation. In the setting of acute MI, celltherapy seems to have a specific time frame of therapeuticopportunity, a period after MI when the myocardium is more“ripe” for cell therapy (6, 42). In this regard, two clinical trialsare investigating the optimal timing of cell delivery of bonemarrow MNCs with the intent of resolving the questionsregarding their therapeutic potential. The Transplantation inMyocardial Infarction Evaluation (TIME) study is a phase-IItrial developed by the CCTRN to test the efficacy, safety, andmost appropriate timing of bone marrow MNCs in patientsafter acute MI (169). This trial is comparing the effects ofintracoronary delivery of MNCs at 3 and 7 days post-MI inpatients with ST-segment elevation, whereas the recently pub-
lished LateTIME trial tested whether delaying MNC deliveryfor 2 to 3 wk following MI, and primary percutaneous coronaryintervention improves global and regional LV function (59,168). In LateTIME, there were no significant changes betweenbaseline and 6-mo measures in the MNC group compared withplacebo in LVEF, wall motion in the infarct zone, and wallmotion in the border zone, as measured by cardiac MRI. Theseresults suggest that the timing of cell therapy at 2 to 3 wkpost-MI may exceed the therapeutic window.
Insights on the most appropriate timing can be gleaned frompreclinical studies. For instance, Zang et al. (188) attempted todetermine the timing and frequency of cell injections necessaryfor optimal therapeutic efficacy. They examined the relation-ship between frequency of injection and therapeutic potentialby injecting bone marrow cells into murine ischemic hearts atdays 3, 7, or 14 post-MI (monotherapy), at days 3 and 7 (dualtherapy), and at days 3, 7, and 14 (triple therapy). Injectionsolely at day 3 reduced infarct size and improved LV function.However, multiple injections of bone marrow cells had noadditive effect, and delaying cell therapy post-MI resulted inno functional benefit. Moreover, a study by Urbanek et al.(172) showed that endogenous CSCs proliferate immediatelyafter MI, but in the chronic phase, their numbers fall and theremaining CSCs have less regenerative potential. This notion ischallenged by the preliminary results of the TAC-HFT clinicaltrial (179) where functional improvement was achieved even ata very late stage post-MI, when heart failure was well estab-lished.
Delivery Methods
While there is general agreement that stem cells are apromising therapeutic regimen post-MI, determining the mostappropriate stem cell delivery method is still a matter ofvigorous debate. To date many techniques have been de-scribed, including intravenous (9), transarterial (175), intra-coronary (160), intramyocardial—either transepicardial (129)or catheter-based transendocardial (153)—and transvenous in-jection into coronary veins (164). The least invasive, trans-
Fig. 4. Mechanisms underlying cardiac re-generation due to cell-based therapy. Theimprovement in myocardial function andventricular geometry after transplantation ofstem results from multiple coordinated ac-tions of cells used as a therapeutic. Success-ful cell-base therapy, as observed as a resultof MSC and CSC therapy, likely arises fromthe actions of both administered cells andhost cellular elements. Notably, MSCs bothhave the capacity for trilineage differentia-tion, as well as stimulating the recruitment,survival, and differentiation of host CSCs.This action of MSCs results from both se-cretion of cytokines and growth factors [vas-cular endothelial growth factor (VEGF),stromal-derived factor 1 (SDF1), insulin-growth factor 1 (IGF1), angiopoietin 1] andantifibrotic mediators that together promoteneovascularization and reduce fibrosis. To-gether these actions and the recruitment ofendogenous CSCs could represent the recon-stitution of stem cell niches in the myocar-dium.
Review
H263CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

venous, has the disadvantage that cells may be trapped in thepulmonary circulation (9) before they even reach the systemiccirculation. Thus far, the most frequently used technique inclinical trials is the percutaneous coronary delivery of cells.The cells are injected via an over-the-wire balloon catheter intothe vessel supplying the ischemic territory. This techniquerequires a transient ischemic period through the inflation of theballoon to give the cells the chance to be distributed and notwashed out, although Tossios et al. (167) proposed that theocclusion is unnecessary.
Intramyocardial delivery of cells is a technique that hasgained momentum in the clinical setting (Fig. 1) (3, 179). Theadvantage of this method lies in the fact that unlike theintracoronary approach, which requires transmigration of the endo-thelial barrier, cells that undergo intramyocardial injection arelargely found in the interstitial space. Conversely, the maindisadvantage is that larger scarred areas require multiple injec-tions. The addition of electromechanical mapping will renderthis approach the most accurate on tracking the ischemic areas(156). Determining the most suitable method and the mostappropriate route of cell delivery will likely depend on theclinical setting and the cell type and no consensus has beenreached.
The common issues surrounding every cardiac deliverymethod employed to date are the reduction of inflammation,stimulation of neovascularization, and improvement in cellengraftment, proliferation, and survival rates (174). To addressthese issues, new experimental strategies are emerging usingcardiac tissue engineering. The general strategy involves acombination of cells and biomaterials (hydrogel or three-dimensional scaffolds) (148). This approach promotes greatercell retention (higher viscosity than saline-based suspension)and survival (antiapoptotic biomaterials) (31, 137). A varietyof biomaterials have been investigated, including fibrin (142,186), collagen (34), Matrigel (82), self-assembling peptides(35), chitosan (183), and alginate (92) in combination withskeletal myoblasts (30), endothelial cells (36), bone marrowMNCs (186), MSCs (187), ESCs (81), and neonatal cardiomy-ocytes (36). Although there are obvious advantages to thisstrategy, there are also some disadvantages. The biomaterialslack flexibility and thus cannot successfully mimic the myo-cardial mechanical microenvironment (50). In addition, theydegrade relatively quickly, and, more importantly, the en-grafted cells create isolated “islands” with no direct cell-cellinteraction with the surrounding tissue (49).
Optimal Dosage of Cell Therapy
Another issue defining an effective therapy is the appropriatequantity of cells transplanted. Instinctively, in stem cell therapyit may be assumed that the more cells transplanted the largertheir beneficial impact would be. Two studies that addressedcell dosage reported interesting results that introduce the notionof the threshold effect. Hamamoto and colleagues (58) injectedsheep with one of four different dosages (25, 75, 225, and 450million allogeneic STRO-3-positive MSCs) or cell media, 1 hpost-induced MI. When compared with control, low-dose (25and 75 million) cell treatment significantly attenuated infarctexpansion and increases in LV end-diastolic volume and LVend-systolic volume. LVEF was improved at all cell doses. Onthe other hand, Schuleri et al. (146) reported a significant
reduction in infarct size in high-dose (200 million) autologousMSC-treated swine compared with low-dose (20 million) au-tologous MSC-treated animals. The regional contractility, asassessed by tagged MRI-derived peak Eulerian circumferentialshortening, was improved in both groups, although the con-tractility of the infarct zone was improved only in the higherdose-treated animals (Fig. 5).
Emerging Trends
Although cell therapy has progressed substantially since thelandmark trials at the beginning of the 21st century, manyquestions remain unanswered regarding the best cell type, thesource of cells, the route of delivery, the timing of the inter-vention, and the number of the cells needed. Studies havesuggested that cells harvested from patients do not show thesame benefit as those from healthy individuals (52), highlight-ing the need for a better understanding of the mechanismsleading to stem cell function and dysfunction. There is aplethora of preclinical studies that suggest strategies to im-
Fig. 5. Dose responsiveness of cardiac functional recovery to intramyocardialMSCs. Plots depict peak circumferential shortening (peak Ecc) in the infarct,border, and remote zones. Peak negative Ecc values represent myocardialshortening and increased contractility, whereas increasingly positive valuesindicate myocardial dysfunction. Ecc improves after cell therapy in both low-and high-dose cell groups in border zones. In contrast, Ecc improves in infarctzones only in the high- and not the low-dose MSC group, consistent with adose-response effect to MSC therapy in that zone. *P � 0.05 vs. placebo;†P � 0.05 week 12 vs. week 24. Reproduced with permission from Schuleri etal, European Heart Journal, 2009 (146).
Review
H264 CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

prove homing and engraftment capacity of stem cells, whichwill potentially translate into better clinical results (71, 134,157). Two potentially exciting possibilities are the combiningof different stem cells or the combination of cell and genetherapy. Recently, microRNAs have assumed a role as poten-tial regulators of cardiovascular biology, vascular growth, andstem cell differentiation and may be attractive targets to opti-mize cell-based therapies (25).
A major issue with cell-based therapy is cell survival, andthe recipient environment seems to play a major role. HumanMSCs were injected into rat infarcted or healthy myocardiumand after 28 days; 90% of human MSCs were still found in thehealthy myocardium, whereas only 18% of those injected inthe infarcted zone were detectable (88). This observationsupports the idea that ischemia creates an unfavorable envi-ronment, likely because of locally expressed proinflammatorycytokines inducing cell apoptosis (47). Preconditioning, byincubation of stem cells with prosurvival factors (e.g., stromalderived factor 1a) (126), increased cell survival by 20% andreduced the number of dying cells in the peri-infarct region by33%. A more complicated approach is transfection of stemcells with prosurvival or antiapoptotic (i.e., Bcl-2) genes (97).Another focus of genetic modification of stem cells is theserine-threonine kinases, Akt, and its downstream targetPim-1, both of which appear to play a crucial prosurvivalsignaling role in the myocardium (32, 116). Normally, theexpression of Pim-1 is low in mature human and murinemyocardium. MI and its sequelae are accompanied by a reac-tivation of Akt and Pim-1 signaling, but this overexpressionafter injury is apparently insufficient to (completely) reversethe damage (116). Fischer et al. (46) modified CSCs to stablyoverexpress Pim-1 and injected them into murine infarctedmyocardium. They reported greater levels of cellular engraft-ment, survival, and functional improvement for Pim-1-overex-pressing cells relative to control 32 wk postdelivery.
Preconditioning the recipient environment represents an-other therapeutic approach. Activation of the Akt system re-duced post-MI myocardial apoptosis by �41% and infarct sizeby 7% in a rodent model (78). Alternatively, inhibition of localinflammation may increase cell survival. Patel et al. (127) usedan adenosine agonist at the time of coronary reperfusion in acanine MI model and reported a 16% reduction of infarct size.Moreover, simultaneous delivery of MSCs and VEGF to ische-mic myocardium increased MSC survival by almost three-fold (134).
Laflamme et al. (87) coinjected a “cocktail” of six differentagents together with cardiomyocytes derived from ESCs in amurine heart post-MI. Each agent was chosen based on adifferent beneficial function. Z-Val-Ala-Asp-fluoromethylk-etone was used to inhibit caspase-induced apoptosis, cyclo-sporine and Bcl-XL peptide to block different mitochondrialdeath pathways, pinacildil to open ATP dependent potassiumchannels, Matrigel to prevent anoikis, and insulin growthfactor-1 to promote cell proliferation. This combinatorial ap-proach resulted in a 2.5-fold increase in LV wall thickeningand suggests that a combination of cell types and/or factors willprove the most efficacious in eliciting a favorable remodelingresponse. Evidence to this effect is found in a preliminaryreport in a porcine MI model showing that the combination ofMSCs and CSCs is more effective at reverse remodeling thaneither cell type alone (178).
Conclusion
In summary, the use of cell-based therapy in acute MI andchronic ischemic cardiomyopathy is rapidly emerging as atherapeutic approach with enormous clinical potential. Thiscell-based approach also holds promise for the treatment ofnonischemic DCM. The next phase of work will likely focusnot only on determining the best cell type(s) to use but, moreimportantly, the mechanisms by which these cells interact withhost cells and elicit their therapeutic effects. As mechanisticunderstanding evolves, there is the growing opportunity toenhance the production and utilization of novel formulations ofcell-based therapeutics.
GRANTS
J. M. Hare is supported by National Heart, Lung, and Blood Institute GrantsRO1-HL-094849, P20-HL-101443, RO1-HL-084275, RO1-HL-107110, andRO1-HL-110737.
DISCLOSURES
J. M. Hare and K. E. Hatzistergos are listed on a patent for cardiaccell-based therapy. J. M. Hare is a consultant to Kardia and receives researchsupport from Biocardia.
AUTHOR CONTRIBUTIONS
V.K. prepared figures; V.K. drafted manuscript; V.K., W.B., I.H.S., K.H.,and J.M.H. edited and revised manuscript; V.K., W.B., I.H.S., K.H., andJ.M.H. approved final version of manuscript.
REFERENCES
1. Aharinejad S, Abraham D, Paulus P, Zins K, Hofmann M, MichlitsW, Gyongyosi M, Macfelda K, Lucas T, Trescher K, Grimm M,Stanley ER. Colony-stimulating factor-1 transfection of myoblasts im-proves the repair of failing myocardium following autologous myoblasttransplantation. Cardiovasc Res 79: 395–404, 2008.
2. Ahn WS, Jeon JJ, Jeong YR, Lee SJ, Yoon SK. Effect of culturetemperature on erythropoietin production and glycosylation in a perfu-sion culture of recombinant CHO cells. Biotechnol Bioeng 101: 1234–1244, 2008.
3. Amado LC, Saliaris AP, Schuleri KH, St John M, Xie JS, CattaneoS, Durand DJ, Fitton T, Kuang JQ, Stewart G, Lehrke S, Baum-gartner WW, Martin BJ, Heldman AW, Hare JM. Cardiac repair withintramyocardial injection of allogeneic mesenchymal stem cells aftermyocardial infarction. Proc Natl Acad Sci USA 102: 11474–11479,2005.
4. Arita NA, Pelaez D, Cheung HS. Activation of the extracellular signal-regulated kinases 1 and 2 (ERK1/2) is needed for the TGFbeta-inducedchondrogenic and osteogenic differentiation of mesenchymal stem cells.Biochem Biophys Res Commun 405: 564–569, 2011.
5. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T,Witzenbichler B, Schatteman G, Isner JM. Isolation of putativeprogenitor endothelial cells for angiogenesis. Science 275: 964–967,1997.
6. Assmus B, Iwasaki M, Schachinger V, Roexe T, Koyanagi M, IekushiK, Xu Q, Tonn T, Seifried E, Liebner S, Kranert WT, Grunwald F,Dimmeler S, Zeiher AM. Acute myocardial infarction activates progen-itor cells and increases Wnt signalling in the bone marrow. Eur Heart J.2011 December 15. [Epub ahead of print.]
7. Augello A, De Bari C. The regulation of differentiation in mesenchymalstem cells. Hum Gene Ther 21: 1226–1238, 2010.
8. Balsam LB, Wagers AJ, Christensen JL, Kofidis T, Weissman IL,Robbins RC. Haematopoietic stem cells adopt mature haematopoieticfates in ischaemic myocardium. Nature 428: 668–673, 2004.
9. Barbash IM, Chouraqui P, Baron J, Feinberg MS, Etzion S, TessoneA, Miller L, Guetta E, Zipori D, Kedes LH, Kloner RA, Leor J.Systemic delivery of bone marrow-derived mesenchymal stem cells tothe infarcted myocardium: feasibility, cell migration, and body distribu-tion. Circulation 108: 863–868, 2003.
Review
H265CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

10. Barile L, Chimenti I, Gaetani R, Forte E, Miraldi F, Frati G, MessinaE, Giacomello A. Cardiac stem cells: isolation, expansion and experi-mental use for myocardial regeneration. Nat Clin Pract Cardiovasc Med4, Suppl 1: S9–S14, 2007.
11. Bearzi C, Rota M, Hosoda T, Tillmanns J, Nascimbene A, De AngelisA, Yasuzawa-Amano S, Trofimova I, Siggins RW, Lecapitaine N,Cascapera S, Beltrami AP, D’Alessandro DA, Zias E, Quaini F,Urbanek K, Michler RE, Bolli R, Kajstura J, Leri A, Anversa P.Human cardiac stem cells. Proc Natl Acad Sci USA 104: 14068–14073,2007.
12. Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, ChimentiS, Kasahara H, Rota M, Musso E, Urbanek K, Leri A, Kajstura J,Nadal-Ginard B, Anversa P. Adult cardiac stem cells are multipotentand support myocardial regeneration. Cell 114: 763–776, 2003.
13. Beltrami AP, Urbanek K, Kajstura J, Yan SM, Finato N, Bussani R,Nadal-Ginard B, Silvestri F, Leri A, Beltrami CA, Anversa P.Evidence that human cardiac myocytes divide after myocardial infarc-tion. N Engl J Med 344: 1750–1757, 2001.
14. Bersell K, Arab S, Haring B, Kuhn B. Neuregulin1/ErbB4 signalinginduces cardiomyocyte proliferation and repair of heart injury. Cell 138:257–270, 2009.
15. Blum B, Benvenisty N. The tumorigenicity of human embryonic stemcells. Adv Cancer Res 100: 133–158, 2008.
16. Bock-Marquette I, Shrivastava S, Pipes GC, Thatcher JE, BlystoneA, Shelton JM, Galindo CL, Melegh B, Srivastava D, Olson EN,DiMaio JM. Thymosin beta4 mediated PKC activation is essential toinitiate the embryonic coronary developmental program and epicardialprogenitor cell activation in adult mice in vivo. J Mol Cell Cardiol 46:728–738, 2009.
17. Boland GM, Perkins G, Hall DJ, Tuan RS. Wnt 3a promotes prolif-eration and suppresses osteogenic differentiation of adult human mesen-chymal stem cells. J Cell Biochem 93: 1210–1230, 2004.
18. Bolli R, Chugh AR, D’Amario D, Loughran JH, Stoddard MF,Ikram S, Beache GM, Wagner SG, Leri A, Hosoda T, Sanada F,Elmore JB, Goichberg P, Cappetta D, Solankhi NK, Fahsah I,Rokosh DG, Slaughter MS, Kajstura J, Anversa P. Cardiac stem cellsin patients with ischaemic cardiomyopathy (SCIPIO): initial results of arandomised phase 1 trial. Lancet 378: 1847–1857, 2011.
19. Buckingham M. Skeletal muscle formation in vertebrates. Curr OpinGenet Dev 11: 440–448, 2001.
20. Bui QT, Gertz ZM, Wilensky RL. Intracoronary delivery of bone-marrow-derived stem cells. Stem Cell Res Ther 1: 29, 2010.
21. Caplan AI. Mesenchymal stem cells. J Orthop Res 9: 641–650, 1991.22. Castaldo C, Di Meglio F, Nurzynska D, Romano G, Maiello C,
Bancone C, Muller P, Bohm M, Cotrufo M, Montagnani S. CD117-positive cells in adult human heart are localized in the subepicardium,and their activation is associated with laminin-1 and alpha6 integrinexpression. Stem Cells 26: 1723–1731, 2008.
23. Cetrulo CL Jr. Cord-blood mesenchymal stem cells and tissue engi-neering. Stem Cell Rev 2: 163–168, 2006.
24. Chabannes D, Hill M, Merieau E, Rossignol J, Brion R, Soulillou JP,Anegon I, Cuturi MC. A role for heme oxygenase-1 in the immuno-suppressive effect of adult rat and human mesenchymal stem cells. Blood110: 3691–3694, 2007.
25. Chamorro-Jorganes A, Araldi E, Penalva LO, Sandhu D, Fernandez-Hernando C, Suarez Y. MicroRNA-16 and microRNA-424 regulate cell-autonomous angiogenic functions in endothelial cells via targeting vascularendothelial growth factor receptor-2 and fibroblast growth factor receptor-1.Arterioscler Thromb Vasc Biol 31: 2595–2606, 2011.
26. Charbord P, Livne E, Gross G, Haupl T, Neves NM, Marie P, BiancoP, Jorgensen C. Human bone marrow mesenchymal stem cells: asystematic reappraisal via the genostem experience. Stem Cell Rev 7:32–42, 2011.
27. Chen MY, Lie PC, Li ZL, Wei X. Endothelial differentiation ofWharton’s jelly-derived mesenchymal stem cells in comparison withbone marrow-derived mesenchymal stem cells. Exp Hematol 37: 629–640, 2009.
28. Chen SL, Fang WW, Qian J, Ye F, Liu YH, Shan SJ, Zhang JJ, LinS, Liao LM, Zhao RC. Improvement of cardiac function after trans-plantation of autologous bone marrow mesenchymal stem cells in pa-tients with acute myocardial infarction. Chin Med J (Engl) 117: 1443–1448, 2004.
29. Chong JJ, Chandrakanthan V, Xaymardan M, Asli NS, Li J, AhmedI, Heffernan C, Menon MK, Scarlett CJ, Rashidianfar A, Biben C,
Zoellner H, Colvin EK, Pimanda JE, Biankin AV, Zhou B, Pu WT,Prall OW, Harvey RP. Adult cardiac-resident MSC-like stem cells witha proepicardial origin. Cell Stem Cell 9: 527–540, 2011.
30. Christman KL, Fok HH, Sievers RE, Fang Q, Lee RJ. Fibrin gluealone and skeletal myoblasts in a fibrin scaffold preserve cardiac functionafter myocardial infarction. Tissue Eng 10: 403–409, 2004.
31. Christman KL, Vardanian AJ, Fang Q, Sievers RE, Fok HH, Lee RJ.Injectable fibrin scaffold improves cell transplant survival, reduces in-farct expansion, and induces neovasculature formation in ischemic myo-cardium. J Am Coll Cardiol 44: 654–660, 2004.
32. Cook SA, Matsui T, Li L, Rosenzweig A. Transcriptional effects ofchronic Akt activation in the heart. J Biol Chem 277: 22528–22533,2002.
33. da Silva Meirelles L, Chagastelles PC, Nardi NB. Mesenchymal stemcells reside in virtually all post-natal organs and tissues. J Cell Sci 119:2204–2213, 2006.
34. Dai W, Wold LE, Dow JS, Kloner RA. Thickening of the infarcted wallby collagen injection improves left ventricular function in rats: a novelapproach to preserve cardiac function after myocardial infarction. J AmColl Cardiol 46: 714–719, 2005.
35. Davis ME, Hsieh PC, Takahashi T, Song Q, Zhang S, Kamm RD,Grodzinsky AJ, Anversa P, Lee RT. Local myocardial insulin-likegrowth factor 1 (IGF-1) delivery with biotinylated peptide nanofibersimproves cell therapy for myocardial infarction. Proc Natl Acad Sci USA103: 8155–8160, 2006.
36. Davis ME, Motion JP, Narmoneva DA, Takahashi T, Hakuno D,Kamm RD, Zhang S, Lee RT. Injectable self-assembling peptidenanofibers create intramyocardial microenvironments for endothelialcells. Circulation 111: 442–450, 2005.
37. Day TF, Guo X, Garrett-Beal L, Yang Y. Wnt/beta-catenin signalingin mesenchymal progenitors controls osteoblast and chondrocyte differ-entiation during vertebrate skeletogenesis. Dev Cell 8: 739–750, 2005.
38. Di Meglio F, Castaldo C, Nurzynska D, Romano V, Miraglia R,Montagnani S. Epicardial cells are missing from the surface of heartswith ischemic cardiomyopathy: a useful clue about the self-renewalpotential of the adult human heart? Int J Cardiol 145: e44–e46, 2010.
39. Di Nicola M, Carlo-Stella C, Magni M, Milanesi M, Longoni PD,Matteucci P, Grisanti S, Gianni AM. Human bone marrow stromalcells suppress T-lymphocyte proliferation induced by cellular or nonspe-cific mitogenic stimuli. Blood 99: 3838–3843, 2002.
40. Diaz-Solano D, Wittig O, Ayala-Grosso C, Pieruzzini R, Cardier JE.Human olfactory mucosa multipotent mesenchymal stromal cells pro-mote survival, proliferation and differentiation of human hematopoieticcells. Stem Cells Dev 2012.
41. Draper JS, Pigott C, Thomson JA, Andrews PW. Surface antigens ofhuman embryonic stem cells: changes upon differentiation in culture. JAnat 200: 249–258, 2002.
42. Dulce RA, Balkan W, Hare JM, Schulman IH. Wnt signalling: amediator of the heart-bone marrow axis after myocardial injury? EurHeart J. 2011 December 23. [Epub ahead of print.]
43. Engel FB. Cardiomyocyte proliferation: a platform for mammaliancardiac repair. Cell Cycle 4: 1360–1363, 2005.
44. Etheridge SL, Spencer GJ, Heath DJ, Genever PG. Expression pro-filing and functional analysis of wnt signaling mechanisms in mesenchy-mal stem cells. Stem Cells 22: 849–860, 2004.
45. Fijnvandraat AC, van Ginneken AC, Schumacher CA, Boheler KR,Lekanne Deprez RH, Christoffels VM, Moorman AF. Cardiomyo-cytes purified from differentiated embryonic stem cells exhibit charac-teristics of early chamber myocardium. J Mol Cell Cardiol 35: 1461–1472, 2003.
46. Fischer KM, Cottage CT, Wu W, Din S, Gude NA, Avitabile D,Quijada P, Collins BL, Fransioli J, Sussman MA. Enhancement ofmyocardial regeneration through genetic engineering of cardiac progen-itor cells expressing Pim-1 kinase. Circulation 120: 2077–2087, 2009.
47. Forrester JS, Libby P. The inflammation hypothesis and its potentialrelevance to statin therapy. Am J Cardiol 99: 732–738, 2007.
48. Fuchs E, Tumbar T, Guasch G. Socializing with the neighbors: stemcells and their niche. Cell 116: 769–778, 2004.
49. Fujimoto KL, Ma Z, Nelson DM, Hashizume R, Guan J, Tobita K,Wagner WR. Synthesis, characterization and therapeutic efficacy of abiodegradable, thermoresponsive hydrogel designed for application inchronic infarcted myocardium. Biomaterials 30: 4357–4368, 2009.
Review
H266 CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

50. Gaudette GR, Cohen IS. Cardiac regeneration: materials can improvethe passive properties of myocardium, but cell therapy must do more.Circulation 114: 2575–2577, 2006.
51. Gehling UM, Ergun S, Schumacher U, Wagener C, Pantel K, Otte M,Schuch G, Schafhausen P, Mende T, Kilic N, Kluge K, Schafer B,Hossfeld DK, Fiedler W. In vitro differentiation of endothelial cellsfrom AC133-positive progenitor cells. Blood 95: 3106–3112, 2000.
52. Giannotti G, Doerries C, Mocharla PS, Mueller MF, Bahlmann FH,Horvath T, Jiang H, Sorrentino SA, Steenken N, Manes C, MarzilliM, Rudolph KL, Luscher TF, Drexler H, Landmesser U. Impairedendothelial repair capacity of early endothelial progenitor cells in pre-hypertension: relation to endothelial dysfunction. Hypertension 55:1389–1397, 2010.
53. Gmeiner M, Zimpfer D, Holfeld J, Seebacher G, Abraham D,Grimm M, Aharinejad S. Improvement of cardiac function in thefailing rat heart after transfer of skeletal myoblasts engineered to over-express placental growth factor. J Thorac Cardiovasc Surg 141: 1238–1245, 2011.
54. Gnecchi M, Zhang Z, Ni A, Dzau VJ. Paracrine mechanisms in adultstem cell signaling and therapy. Circ Res 103: 1204–1219, 2008.
55. Gramsch B, Gabriel HD, Wiemann M, Grummer R, Winterhager E,Bingmann D, Schirrmacher K. Enhancement of connexin 43 expres-sion increases proliferation and differentiation of an osteoblast-like cellline. Exp Cell Res 264: 397–407, 2001.
56. Griffin MD, Ritter T, Mahon BP. Immunological aspects of allogeneicmesenchymal stem cell therapies. Hum Gene Ther 21: 1641–1655, 2010.
57. Guo J, Lin G, Bao C, Hu Z, Chu H, Hu M. Insulin-like growth factor1 improves the efficacy of mesenchymal stem cells transplantation in arat model of myocardial infarction. J Biomed Sci 15: 89–97, 2008.
58. Hamamoto H, Gorman JH 3rd, Ryan LP, Hinmon R, Martens TP,Schuster MD, Plappert T, Kiupel M, St. John-Sutton MG, Itescu S,Gorman RC. Allogeneic mesenchymal precursor cell therapy to limitremodeling after myocardial infarction: the effect of cell dosage. AnnThorac Surg 87: 794–801, 2009.
59. Hare JM. Bone marrow therapy for myocardial infarction. JAMA 306:2156–2157, 2011.
60. Hare JM, Traverse JH, Henry TD, Dib N, Strumpf RK, SchulmanSP, Gerstenblith G, DeMaria AN, Denktas AE, Gammon RS, Her-miller JB Jr, Reisman MA, Schaer GL, Sherman W. A randomized,double-blind, placebo-controlled, dose-escalation study of intravenousadult human mesenchymal stem cells (prochymal) after acute myocardialinfarction. J Am Coll Cardiol 54: 2277–2286, 2009.
61. Hatzistergos KE, Quevedo H, Oskouei BN, Hu Q, Feigenbaum GS,Margitich IS, Mazhari R, Boyle AJ, Zambrano JP, Rodriguez JE,Dulce R, Pattany PM, Valdes D, Revilla C, Heldman AW, McNieceI, Hare JM. Bone marrow mesenchymal stem cells stimulate cardiacstem cell proliferation and differentiation. Circ Res 107: 913–922, 2010.
62. Hierlihy AM, Seale P, Lobe CG, Rudnicki MA, Megeney LA. Thepost-natal heart contains a myocardial stem cell population. FEBS Lett530: 239–243, 2002.
63. Ho JH, Ma WH, Su Y, Tseng KC, Kuo TK, Lee OK. Thymosin beta-4directs cell fate determination of human mesenchymal stem cells throughbiophysical effects. J Orthop Res 28: 131–138, 2010.
64. Ieda M, Fu JD, Delgado-Olguin P, Vedantham V, Hayashi Y, Bru-neau BG, Srivastava D. Direct reprogramming of fibroblasts intofunctional cardiomyocytes by defined factors. Cell 142: 375–386, 2010.
65. Jackson KA, Majka SM, Wang H, Pocius J, Hartley CJ, MajeskyMW, Entman ML, Michael LH, Hirschi KK, Goodell MA. Regener-ation of ischemic cardiac muscle and vascular endothelium by adult stemcells. J Clin Invest 107: 1395–1402, 2001.
66. Jiang XX, Zhang Y, Liu B, Zhang SX, Wu Y, Yu XD, Mao N. Humanmesenchymal stem cells inhibit differentiation and function of monocyte-derived dendritic cells. Blood 105: 4120–4126, 2005.
67. Jones EA, Kinsey SE, English A, Jones RA, Straszynski L, MeredithDM, Markham AF, Jack A, Emery P, McGonagle D. Isolation andcharacterization of bone marrow multipotential mesenchymal progenitorcells. Arthritis Rheum 46: 3349–3360, 2002.
68. Jopling C, Sleep E, Raya M, Marti M, Raya A, Izpisua Belmonte JC.Zebrafish heart regeneration occurs by cardiomyocyte dedifferentiationand proliferation. Nature 464: 606–609, 2010.
69. Kajstura J, Rota M, Whang B, Cascapera S, Hosoda T, Bearzi C,Nurzynska D, Kasahara H, Zias E, Bonafe M, Nadal-Ginard B,Torella D, Nascimbene A, Quaini F, Urbanek K, Leri A, Anversa P.
Bone marrow cells differentiate in cardiac cell lineages after infarctionindependently of cell fusion. Circ Res 96: 127–137, 2005.
70. Kamihata H, Matsubara H, Nishiue T, Fujiyama S, Tsutsumi Y,Ozono R, Masaki H, Mori Y, Iba O, Tateishi E, Kosaki A, ShintaniS, Murohara T, Imaizumi T, Iwasaka T. Implantation of bone marrowmononuclear cells into ischemic myocardium enhances collateral perfu-sion and regional function via side supply of angioblasts, angiogenicligands, and cytokines. Circulation 104: 1046–1052, 2001.
71. Kanashiro-Takeuchi RM, Schulman IH, Hare JM. Pharmacologicand genetic strategies to enhance cell therapy for cardiac regeneration. JMol Cell Cardiol 51: 619–625, 2011.
72. Kapoor N, Galang G, Marban E, Cho HC. Transcriptional suppressionof connexin43 by TBX18 undermines cell-cell electrical coupling inpostnatal cardiomyocytes. J Biol Chem 286: 14073–14079, 2011.
73. Khoury M, Drake A, Chen Q, Dong D, Leskov I, Fragoso MF, Li Y,Iliopoulou BP, Hwang W, Lodish HF, Chen J. Mesenchymal stemcells secreting angiopoietin-like-5 support efficient expansion of humanhematopoietic stem cells without compromising their repopulating po-tential. Stem Cells Dev 20: 1371–1381, 2011.
74. Kikuchi K, Holdway JE, Werdich AA, Anderson RM, Fang Y,Egnaczyk GF, Evans T, Macrae CA, Stainier DY, Poss KD. Primarycontribution to zebrafish heart regeneration by gata4� cardiomyocytes.Nature 464: 601–605, 2010.
75. Kim BO, Tian H, Prasongsukarn K, Wu J, Angoulvant D, Wnendt S,Muhs A, Spitkovsky D, Li RK. Cell transplantation improves ventric-ular function after a myocardial infarction: a preclinical study of humanunrestricted somatic stem cells in a porcine model. Circulation 112:I96–I104, 2005.
76. Kinnaird T, Stabile E, Burnett MS, Lee CW, Barr S, Fuchs S,Epstein SE. Marrow-derived stromal cells express genes encoding abroad spectrum of arteriogenic cytokines and promote in vitro and in vivoarteriogenesis through paracrine mechanisms. Circ Res 94: 678–685,2004.
77. Kitaori T, Ito H, Schwarz EM, Tsutsumi R, Yoshitomi H, Oishi S,Nakano M, Fujii N, Nagasawa T, Nakamura T. Stromal cell-derivedfactor 1/CXCR4 signaling is critical for the recruitment of mesenchymalstem cells to the fracture site during skeletal repair in a mouse model.Arthritis Rheum 60: 813–823, 2009.
78. Klopsch C, Furlani D, Gabel R, Li W, Pittermann E, Ugurlucan M,Kundt G, Zingler C, Titze U, Wang W, Ong LL, Wagner K, Li RK,Ma N, Steinhoff G. Intracardiac injection of erythropoietin induces stemcell recruitment and improves cardiac functions in a rat myocardialinfarction model. J Cell Mol Med 13: 664–679, 2009.
79. Klyushnenkova E, Mosca JD, Zernetkina V, Majumdar MK, BeggsKJ, Simonetti DW, Deans RJ, McIntosh KR. T cell responses toallogeneic human mesenchymal stem cells: immunogenicity, tolerance,and suppression. J Biomed Sci 12: 47–57, 2005.
80. Kocher AA, Schuster MD, Szabolcs MJ, Takuma S, Burkhoff D,Wang J, Homma S, Edwards NM, Itescu S. Neovascularization ofischemic myocardium by human bone-marrow-derived angioblasts pre-vents cardiomyocyte apoptosis, reduces remodeling and improves car-diac function. Nat Med 7: 430–436, 2001.
81. Kofidis T, de Bruin JL, Hoyt G, Lebl DR, Tanaka M, Yamane T,Chang CP, Robbins RC. Injectable bioartificial myocardial tissue forlarge-scale intramural cell transfer and functional recovery of injuredheart muscle. J Thorac Cardiovasc Surg 128: 571–578, 2004.
82. Kofidis T, Lebl DR, Martinez EC, Hoyt G, Tanaka M, Robbins RC.Novel injectable bioartificial tissue facilitates targeted, less invasive,large-scale tissue restoration on the beating heart after myocardial injury.Circulation 112: I173–I177, 2005.
83. Kogler G, Sensken S, Airey JA, Trapp T, Muschen M, Feldhahn N,Liedtke S, Sorg RV, Fischer J, Rosenbaum C, Greschat S, KnipperA, Bender J, Degistirici O, Gao J, Caplan AI, Colletti EJ, Almeida-Porada G, Muller HW, Zanjani E, Wernet P. A new human somaticstem cell from placental cord blood with intrinsic pluripotent differenti-ation potential. J Exp Med 200: 123–135, 2004.
84. Krampera M, Cosmi L, Angeli R, Pasini A, Liotta F, Andreini A,Santarlasci V, Mazzinghi B, Pizzolo G, Vinante F, Romagnani P,Maggi E, Romagnani S, Annunziato F. Role for interferon-gamma inthe immunomodulatory activity of human bone marrow mesenchymalstem cells. Stem Cells 24: 386–398, 2006.
85. Kuhn B, del Monte F, Hajjar RJ, Chang YS, Lebeche D, Arab S,Keating MT. Periostin induces proliferation of differentiated cardio-myocytes and promotes cardiac repair. Nat Med 13: 962–969, 2007.
Review
H267CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

86. Lacis A, Erglis A. Intramyocardial administration of autologous bonemarrow mononuclear cells in a critically ill child with dilated cardio-myopathy. Cardiol Young 21: 110–112, 2011.
87. Laflamme MA, Chen KY, Naumova AV, Muskheli V, Fugate JA,Dupras SK, Reinecke H, Xu C, Hassanipour M, Police S, O’SullivanC, Collins L, Chen Y, Minami E, Gill EA, Ueno S, Yuan C, Gold J,Murry CE. Cardiomyocytes derived from human embryonic stem cellsin pro-survival factors enhance function of infarcted rat hearts. NatBiotechnol 25: 1015–1024, 2007.
88. Laflamme MA, Gold J, Xu C, Hassanipour M, Rosler E, Police S,Muskheli V, Murry CE. Formation of human myocardium in the ratheart from human embryonic stem cells. Am J Pathol 167: 663–671,2005.
89. Laugwitz KL, Moretti A, Lam J, Gruber P, Chen Y, Woodard S, LinLZ, Cai CL, Lu MM, Reth M, Platoshyn O, Yuan JX, Evans S, ChienKR. Postnatal isl1� cardioblasts enter fully differentiated cardiomyocytelineages. Nature 433: 647–653, 2005.
90. Lavine KJ, White AC, Park C, Smith CS, Choi K, Long F, Hui CC,Ornitz DM. Fibroblast growth factor signals regulate a wave of Hedge-hog activation that is essential for coronary vascular development. GenesDev 20: 1651–1666, 2006.
91. Leistner DM, Fischer-Rasokat U, Honold J, Seeger FH, SchachingerV, Lehmann R, Martin H, Burck I, Urbich C, Dimmeler S, ZeiherAM, Assmus B. Transplantation of progenitor cells and regenerationenhancement in acute myocardial infarction (TOPCARE-AMI): final5-year results suggest long-term safety and efficacy. Clin Res Cardiol100: 925–934, 2011.
92. Leor J, Tuvia S, Guetta V, Manczur F, Castel D, Willenz U, Petne-hazy O, Landa N, Feinberg MS, Konen E, Goitein O, Tsur-Gang O,Shaul M, Klapper L, Cohen S. Intracoronary injection of in situforming alginate hydrogel reverses left ventricular remodeling aftermyocardial infarction in Swine. J Am Coll Cardiol 54: 1014–1023, 2009.
93. Leri A, Kajstura J, Anversa P. Cardiac stem cells and mechanisms ofmyocardial regeneration. Physiol Rev 85: 1373–1416, 2005.
94. Leri A, Kajstura J, Anversa P. Role of cardiac stem cells in cardiacpathophysiology: a paradigm shift in human myocardial biology. CircRes 109: 941–961, 2011.
95. Li TS, Cheng K, Lee ST, Matsushita S, Davis D, Malliaras K, ZhangY, Matsushita N, Smith RR, Marban E. Cardiospheres recapitulate aniche-like microenvironment rich in stemness and cell-matrix interac-tions, rationalizing their enhanced functional potency for myocardialrepair. Stem Cells 28: 2088–2098, 2010.
96. Li TS, Cheng K, Malliaras K, Smith RR, Zhang Y, Sun B, Matsu-shita N, Blusztajn A, Terrovitis J, Kusuoka H, Marban L, Marban E.Direct comparison of different stem cell types and subpopulations revealssuperior paracrine potency and myocardial repair efficacy with cardio-sphere-derived cells. J Am Coll Cardiol 59: 942–953, 2012.
97. Li W, Ma N, Ong LL, Nesselmann C, Klopsch C, Ladilov Y, FurlaniD, Piechaczek C, Moebius JM, Lutzow K, Lendlein A, Stamm C, LiRK, Steinhoff G. Bcl-2 engineered MSCs inhibited apoptosis andimproved heart function. Stem Cells 25: 2118–2127, 2007.
98. Li X, Yu X, Lin Q, Deng C, Shan Z, Yang M, Lin S. Bone marrowmesenchymal stem cells differentiate into functional cardiac phenotypesby cardiac microenvironment. J Mol Cell Cardiol 42: 295–303, 2007.
99. Limana F, Bertolami C, Mangoni A, Di Carlo A, Avitabile D, MociniD, Iannelli P, De Mori R, Marchetti C, Pozzoli O, Gentili C, ZacheoA, Germani A, Capogrossi MC. Myocardial infarction induces embry-onic reprogramming of epicardial c-kit� cells: role of the pericardialfluid. J Mol Cell Cardiol 48: 609–618, 2010.
100. Limana F, Zacheo A, Mocini D, Mangoni A, Borsellino G, Diaman-tini A, De Mori R, Battistini L, Vigna E, Santini M, Loiaconi V,Pompilio G, Germani A, Capogrossi MC. Identification of myocardialand vascular precursor cells in human and mouse epicardium. Circ Res101: 1255–1265, 2007.
101. Ma N, Ladilov Y, Kaminski A, Piechaczek C, Choi YH, Li W,Steinhoff G, Stamm C. Umbilical cord blood cell transplantation formyocardial regeneration. Transplant Proc 38: 771–773, 2006.
102. Makino S, Fukuda K, Miyoshi S, Konishi F, Kodama H, Pan J, SanoM, Takahashi T, Hori S, Abe H, Hata J, Umezawa A, Ogawa S.Cardiomyocytes can be generated from marrow stromal cells in vitro. JClin Invest 103: 697–705, 1999.
103. Makkar RR, Smith RR, Cheng K, Malliaras K, Thomson LE,Berman D, Czer LS, Marban L, Mendizabal A, Johnston PV, RussellSD, Schuleri KH, Lardo AC, Gerstenblith G, Marban E. Intracoro-
nary cardiosphere-derived cells for heart regeneration after myocardialinfarction (CADUCEUS): a prospective, randomised phase 1 trial. Lan-cet 379: 895–904, 2012.
104. Malliaras K, Kreke M, Marban E. The stuttering progress of celltherapy for heart disease. Clin Pharmacol Ther 90: 532–541, 2011.
105. Mangi AA, Noiseux N, Kong D, He H, Rezvani M, Ingwall JS, DzauVJ. Mesenchymal stem cells modified with Akt prevent remodeling andrestore performance of infarcted hearts. Nat Med 9: 1195–1201, 2003.
106. Marchetti S, Gimond C, Iljin K, Bourcier C, Alitalo K, PouyssegurJ, Pages G. Endothelial cells genetically selected from differentiatingmouse embryonic stem cells incorporate at sites of neovascularization invivo. J Cell Sci 115: 2075–2085, 2002.
107. Martin CM, Meeson AP, Robertson SM, Hawke TJ, Richardson JA,Bates S, Goetsch SC, Gallardo TD, Garry DJ. Persistent expression ofthe ATP-binding cassette transporter, Abcg2, identifies cardiac SP cellsin the developing and adult heart. Dev Biol 265: 262–275, 2004.
108. Martin I, Muraglia A, Campanile G, Cancedda R, Quarto R. Fibro-blast growth factor-2 supports ex vivo expansion and maintenance ofosteogenic precursors from human bone marrow. Endocrinology 138:4456–4462, 1997.
109. Martino HF, Oliveira PS, Souza FC, Costa PC, Assuncao ESE,Villela R, Gaze M, Weitzel LH, Oliveira A Jr, Muccillo FB, ArveloSN, Sa R, Guimaraes TC, Tura BR, Campos de Carvalho AC. Asafety and feasibility study of cell therapy in dilated cardiomyopathy.Braz J Med Biol Res 43: 989–995, 2010.
110. Mazhari R, Hare JM. Mechanisms of action of mesenchymal stem cellsin cardiac repair: potential influences on the cardiac stem cell niche. NatClin Pract Cardiovasc Med 4, Suppl 1: S21–S26, 2007.
111. Menard C, Hagege AA, Agbulut O, Barro M, Morichetti MC,Brasselet C, Bel A, Messas E, Bissery A, Bruneval P, Desnos M,Puceat M, Menasche P. Transplantation of cardiac-committed mouseembryonic stem cells to infarcted sheep myocardium: a preclinical study.Lancet 366: 1005–1012, 2005.
112. Messina E, De Angelis L, Frati G, Morrone S, Chimenti S, FiordalisoF, Salio M, Battaglia M, Latronico MV, Coletta M, Vivarelli E, FratiL, Cossu G, Giacomello A. Isolation and expansion of adult cardiacstem cells from human and murine heart. Circ Res 95: 911–921, 2004.
113. Min JY, Yang Y, Converso KL, Liu L, Huang Q, Morgan JP, XiaoYF. Transplantation of embryonic stem cells improves cardiac functionin postinfarcted rats. J Appl Physiol 92: 288–296, 2002.
114. Moelker AD, Baks T, Wever KM, Spitskovsky D, Wielopolski PA,van Beusekom HM, van Geuns RJ, Wnendt S, Duncker DJ, van derGiessen WJ. Intracoronary delivery of umbilical cord blood derivedunrestricted somatic stem cells is not suitable to improve LV functionafter myocardial infarction in swine. J Mol Cell Cardiol 42: 735–745,2007.
115. Moore KA, Lemischka IR. Stem cells and their niches. Science 311:1880–1885, 2006.
116. Muraski JA, Rota M, Misao Y, Fransioli J, Cottage C, Gude N,Esposito G, Delucchi F, Arcarese M, Alvarez R, Siddiqi S, Emman-uel GN, Wu W, Fischer K, Martindale JJ, Glembotski CC, Leri A,Kajstura J, Magnuson N, Berns A, Beretta RM, Houser SR, SchaeferEM, Anversa P, Sussman MA. Pim-1 regulates cardiomyocyte survivaldownstream of Akt. Nat Med 13: 1467–1475, 2007.
117. Murry CE, Reinecke H, Pabon LM. Regeneration gaps: observationson stem cells and cardiac repair. J Am Coll Cardiol 47: 1777–1785, 2006.
118. Murry CE, Soonpaa MH, Reinecke H, Nakajima H, Nakajima HO,Rubart M, Pasumarthi KB, Virag JI, Bartelmez SH, Poppa V,Bradford G, Dowell JD, Williams DA, Field LJ. Haematopoietic stemcells do not transdifferentiate into cardiac myocytes in myocardialinfarcts. Nature 428: 664–668, 2004.
119. Mushtaq M, Oskouei BN, Hare JM. Cell therapy for heart disease: togenetically modify or not, that is the question. Circ Res 108: 398–401,2011.
120. Nombela-Arrieta C, Ritz J, Silberstein LE. The elusive nature andfunction of mesenchymal stem cells. Nat Rev Mol Cell Biol 12: 126–131,2011.
121. Nygren JM, Jovinge S, Breitbach M, Sawen P, Roll W, Hescheler J,Taneera J, Fleischmann BK, Jacobsen SE. Bone marrow-derivedhematopoietic cells generate cardiomyocytes at a low frequency throughcell fusion, but not transdifferentiation. Nat Med 10: 494–501, 2004.
122. Oh H, Bradfute SB, Gallardo TD, Nakamura T, Gaussin V, MishinaY, Pocius J, Michael LH, Behringer RR, Garry DJ, Entman ML,Schneider MD. Cardiac progenitor cells from adult myocardium: hom-
Review
H268 CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

ing, differentiation, and fusion after infarction. Proc Natl Acad Sci USA100: 12313–12318, 2003.
123. Orlic D, Kajstura J, Chimenti S, Jakoniuk I, Anderson SM, Li B,Pickel J, McKay R, Nadal-Ginard B, Bodine DM, Leri A, Anversa P.Bone marrow cells regenerate infarcted myocardium. Nature 410: 701–705, 2001.
124. Orlic D, Kajstura J, Chimenti S, Limana F, Jakoniuk I, Quaini F,Nadal-Ginard B, Bodine DM, Leri A, Anversa P. Mobilized bonemarrow cells repair the infarcted heart, improving function and survival.Proc Natl Acad Sci USA 98: 10344–10349, 2001.
125. Park JH, Lee MY, Heo JS, Han HJ. A potential role of connexin 43 inepidermal growth factor-induced proliferation of mouse embryonic stemcells: involvement of Ca2�/PKC, p44/42 and p38 MAPKs pathways. CellProlif 41: 786–802, 2008.
126. Pasha Z, Wang Y, Sheikh R, Zhang D, Zhao T, Ashraf M. Precon-ditioning enhances cell survival and differentiation of stem cells duringtransplantation in infarcted myocardium. Cardiovasc Res 77: 134–142,2008.
127. Patel RA, Glover DK, Broisat A, Kabul HK, Ruiz M, Goodman NC,Kramer CM, Meerdink DJ, Linden J, Beller GA. Reduction inmyocardial infarct size at 48 hours after brief intravenous infusion ofATL-146e, a highly selective adenosine A2A receptor agonist. Am JPhysiol Heart Circ Physiol 297: H637–H642, 2009.
128. Peichev M, Naiyer AJ, Pereira D, Zhu Z, Lane WJ, Williams M, OzMC, Hicklin DJ, Witte L, Moore MA, Rafii S. Expression ofVEGFR-2 and AC133 by circulating human CD34� cells identifies apopulation of functional endothelial precursors. Blood 95: 952–958,2000.
129. Perin EC, Lopez J. Methods of stem cell delivery in cardiac diseases.Nat Clin Pract Cardiovasc Med 3, Suppl 1: S110–S113, 2006.
130. Perin EC, Willerson JT, Pepine CJ, Henry TD, Ellis SG, Zhao DX,Silva GV, Lai D, Thomas JD, Kronenberg MW, Martin AD, Ander-son RD, Traverse JH, Penn MS, Anwaruddin S, Hatzopoulos AK,Gee AP, Taylor DA, Cogle CR, Smith D, Westbrook L, Chen J,Handberg E, Olson RE, Geither C, Bowman S, Francescon J, Bara-niuk S, Piller LB, Simpson LM, Loghin C, Aguilar D, Richman S,Zierold C, Bettencourt J, Sayre SL, Vojvodic RW, Skarlatos SI,Gordon DJ, Ebert RF, Kwak M, Moye LA, Simari RD. Effect oftransendocardial delivery of autologous bone marrow mononuclear cellson functional capacity, left ventricular function, and perfusion in chronicheart failure: The FOCUS-CCTRN Trial. JAMA 307: 1717–1726, 2012.
131. Pijnappels DA, Schalij MJ, Ramkisoensing AA, van Tuyn J, de VriesAA, van der Laarse A, Ypey DL, Atsma DE. Forced alignment ofmesenchymal stem cells undergoing cardiomyogenic differentiation af-fects functional integration with cardiomyocyte cultures. Circ Res 103:167–176, 2008.
132. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, MoscaJD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilin-eage potential of adult human mesenchymal stem cells. Science 284:143–147, 1999.
133. Pittenger MF, Mosca JD, McIntosh KR. Human mesenchymal stemcells: progenitor cells for cartilage, bone, fat and stroma. Curr TopMicrobiol Immunol 251: 3–11, 2000.
134. Pons J, Huang Y, Arakawa-Hoyt J, Washko D, Takagawa J, Ye J,Grossman W, Su H. VEGF improves survival of mesenchymal stemcells in infarcted hearts. Biochem Biophys Res Commun 376: 419–422,2008.
135. Quevedo HC, Hatzistergos KE, Oskouei BN, Feigenbaum GS, Ro-driguez JE, Valdes D, Pattany PM, Zambrano JP, Hu Q, McNiece I,Heldman AW, Hare JM. Allogeneic mesenchymal stem cells restorecardiac function in chronic ischemic cardiomyopathy via trilineage dif-ferentiating capacity. Proc Natl Acad Sci USA 106: 14022–14027, 2009.
136. Raffaghello L, Bianchi G, Bertolotto M, Montecucco F, Busca A,Dallegri F, Ottonello L, Pistoia V. Human mesenchymal stem cellsinhibit neutrophil apoptosis: a model for neutrophil preservation in thebone marrow niche. Stem Cells 26: 151–162, 2008.
137. Reffelmann T, Kloner RA. Cellular cardiomyoplasty—cardiomyo-cytes, skeletal myoblasts, or stem cells for regenerating myocardium andtreatment of heart failure? Cardiovasc Res 58: 358–368, 2003.
138. Ren G, Zhang L, Zhao X, Xu G, Zhang Y, Roberts AI, Zhao RC, ShiY. Mesenchymal stem cell-mediated immunosuppression occurs viaconcerted action of chemokines and nitric oxide. Cell Stem Cell 2:141–150, 2008.
139. Riley PR, Smart N. Thymosin beta4 induces epicardium-derived neo-vascularization in the adult heart. Biochem Soc Trans 37: 1218–1220,2009.
140. Roger VL, Go AS, Lloyd-Jones DM, Benjamin EJ, Berry JD, BordenWB, Bravata DM, Dai S, Ford ES, Fox CS, Fullerton HJ, Gillespie C,Hailpern SM, Heit JA, Howard VJ, Kissela BM, Kittner SJ, Lack-land DT, Lichtman JH, Lisabeth LD, Makuc DM, Marcus GM,Marelli A, Matchar DB, Moy CS, Mozaffarian D, Mussolino ME,Nichol G, Paynter NP, Soliman EZ, Sorlie PD, Sotoodehnia N, TuranTN, Virani SS, Wong ND, Woo D, Turner MB. Heart disease andstroke statistics—2012 update: a report from the American Heart Asso-ciation. Circulation 125: e2–e220, 2012.
141. Ryan JM, Barry FP, Murphy JM, Mahon BP. Mesenchymal stemcells avoid allogeneic rejection. J Inflamm (Lond) 2: 8, 2005.
142. Ryu JH, Kim IK, Cho SW, Cho MC, Hwang KK, Piao H, Piao S, LimSH, Hong YS, Choi CY, Yoo KJ, Kim BS. Implantation of bonemarrow mononuclear cells using injectable fibrin matrix enhances neo-vascularization in infarcted myocardium. Biomaterials 26: 319–326,2005.
143. Sato K, Ozaki K, Oh I, Meguro A, Hatanaka K, Nagai T, Muroi K,Ozawa K. Nitric oxide plays a critical role in suppression of T-cellproliferation by mesenchymal stem cells. Blood 109: 228–234, 2007.
144. Schaefer A, Zwadlo C, Fuchs M, Meyer GP, Lippolt P, Wollert KC,Drexler H. Long-term effects of intracoronary bone marrow cell transferon diastolic function in patients after acute myocardial infarction: 5-yearresults from the randomized-controlled BOOST trial–an echocardio-graphic study. Eur J Echocardiogr 11: 165–171, 2010.
145. Schuleri KH, Amado LC, Boyle AJ, Centola M, Saliaris AP, GutmanMR, Hatzistergos KE, Oskouei BN, Zimmet JM, Young RG, Held-man AW, Lardo AC, Hare JM. Early improvement in cardiac tissueperfusion due to mesenchymal stem cells. Am J Physiol Heart CircPhysiol 294: H2002–H2011, 2008.
146. Schuleri KH, Feigenbaum GS, Centola M, Weiss ES, Zimmet JM,Turney J, Kellner J, Zviman MM, Hatzistergos KE, Detrick B,Conte JV, McNiece I, Steenbergen C, Lardo AC, Hare JM. Autolo-gous mesenchymal stem cells produce reverse remodelling in chronicischaemic cardiomyopathy. Eur Heart J 30: 2722–2732, 2009.
147. Schuster MD, Kocher AA, Seki T, Martens TP, Xiang G, Homma S,Itescu S. Myocardial neovascularization by bone marrow angioblastsresults in cardiomyocyte regeneration. Am J Physiol Heart Circ Physiol287: H525–H532, 2004.
148. Segers VF, Lee RT. Biomaterials to enhance stem cell function in theheart. Circ Res 109: 910–922, 2011.
149. Shake JG, Gruber PJ, Baumgartner WA, Senechal G, Meyers J,Redmond JM, Pittenger MF, Martin BJ. Mesenchymal stem cellimplantation in a swine myocardial infarct model: engraftment andfunctional effects. Ann Thorac Surg 73: 1919–1925, 2002.
150. Shantsila E, Watson T, Lip GY. Endothelial progenitor cells in cardio-vascular disorders. J Am Coll Cardiol 49: 741–752, 2007.
151. Sherman LS, Munoz J, Patel SA, Dave MA, Paige I, Rameshwar P.Moving from the laboratory bench to patients’ bedside: considerationsfor effective therapy with stem cells. Clin Transl Sci 4: 380–386, 2011.
152. Silva GV, Litovsky S, Assad JA, Sousa AL, Martin BJ, Vela D,Coulter SC, Lin J, Ober J, Vaughn WK, Branco RV, Oliveira EM,He R, Geng YJ, Willerson JT, Perin EC. Mesenchymal stem cellsdifferentiate into an endothelial phenotype, enhance vascular density, andimprove heart function in a canine chronic ischemia model. Circulation111: 150–156, 2005.
153. Silva GV, Perin EC, Dohmann HF, Borojevic R, Silva SA, Sousa AL,Assad JA, Vaughn WK, Mesquita CT, Belem L, Carvalho AC,Dohmann HJ, Barroso do Amaral E, Coutinho J, Branco R, OliveiraE, Willerson JT. Catheter-based transendocardial delivery of autologousbone-marrow-derived mononuclear cells in patients listed for heart trans-plantation. Tex Heart Inst J 31: 214–219, 2004.
154. Siminiak T, Kalawski R, Fiszer D, Jerzykowska O, Rzezniczak J,Rozwadowska N, Kurpisz M. Autologous skeletal myoblast transplan-tation for the treatment of postinfarction myocardial injury: phase Iclinical study with 12 months of follow-up. Am Heart J 148: 531–537,2004.
155. Smart N, Rossdeutsch A, Riley PR. Thymosin beta4 and angiogenesis:modes of action and therapeutic potential. Angiogenesis 10: 229–241,2007.
156. Smits PC, van Geuns RJ, Poldermans D, Bountioukos M, Onderwa-ter EE, Lee CH, Maat AP, Serruys PW. Catheter-based intramyocar-
Review
H269CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from

dial injection of autologous skeletal myoblasts as a primary treatment ofischemic heart failure: clinical experience with six-month follow-up. JAm Coll Cardiol 42: 2063–2069, 2003.
157. Spyridopoulos I, Haendeler J, Urbich C, Brummendorf TH, Oh H,Schneider MD, Zeiher AM, Dimmeler S. Statins enhance migratorycapacity by upregulation of the telomere repeat-binding factor TRF2 inendothelial progenitor cells. Circulation 110: 3136–3142, 2004.
158. Srivastava D. Making or breaking the heart: from lineage determinationto morphogenesis. Cell 126: 1037–1048, 2006.
159. Srivastava D, Saxena A, Michael Dimaio J, Bock-Marquette I.Thymosin beta4 is cardioprotective after myocardial infarction. Ann NYAcad Sci 1112: 161–170, 2007.
160. Strauer BE, Brehm M, Zeus T, Kostering M, Hernandez A, Sorg RV,Kogler G, Wernet P. Repair of infarcted myocardium by autologousintracoronary mononuclear bone marrow cell transplantation in humans.Circulation 106: 1913–1918, 2002.
161. Suncion VY, Schulman IH, Hare J. Concise Review: The role ofclinical trials in deciphering mechanisms of action of cardiac cell-basedtherapy. Stem Cells Transl Med 2: 29–35, 2012.
162. Tambara K, Sakakibara Y, Sakaguchi G, Lu F, Premaratne GU, LinX, Nishimura K, Komeda M. Transplanted skeletal myoblasts can fullyreplace the infarcted myocardium when they survive in the host in largenumbers. Circulation 108, Suppl 1: II259–II263, 2003.
163. Taylor DO, Edwards LB, Aurora P, Christie JD, Dobbels F, Kirk R,Rahmel AO, Kucheryavaya AY, Hertz MI. Registry of the InternationalSociety for Heart and Lung Transplantation: twenty-fifth official adult hearttransplant report—2008. J Heart Lung Transplant 27: 943–956, 2008.
164. Thompson CA, Nasseri BA, Makower J, Houser S, McGarry M,Lamson T, Pomerantseva I, Chang JY, Gold HK, Vacanti JP,Oesterle SN. Percutaneous transvenous cellular cardiomyoplasty. Anovel nonsurgical approach for myocardial cell transplantation. J AmColl Cardiol 41: 1964–1971, 2003.
165. Tokunaga A, Oya T, Ishii Y, Motomura H, Nakamura C, Ishizawa S,Fujimori T, Nabeshima Y, Umezawa A, Kanamori M, Kimura T,Sasahara M. PDGF receptor beta is a potent regulator of mesenchymalstromal cell function. J Bone Miner Res 23: 1519–1528, 2008.
166. Toma C, Pittenger MF, Cahill KS, Byrne BJ, Kessler PD. Humanmesenchymal stem cells differentiate to a cardiomyocyte phenotype inthe adult murine heart. Circulation 105: 93–98, 2002.
167. Tossios P, Krausgrill B, Schmidt M, Fischer T, Halbach M, FriesJW, Fahnenstich S, Frommolt P, Heppelmann I, Schmidt A, Scho-macker K, Fischer JH, Bloch W, Mehlhorn U, Schwinger RH,Muller-Ehmsen J. Role of balloon occlusion for mononuclear bonemarrow cell deposition after intracoronary injection in pigs with reper-fused myocardial infarction. Eur Heart J 29: 1911–1921, 2008.
168. Traverse JH, Henry TD, Ellis SG, Pepine CJ, Willerson JT, ZhaoDX, Forder JR, Byrne BJ, Hatzopoulos AK, Penn MS, Perin EC,Baran KW, Chambers J, Lambert C, Raveendran G, Simon DI,Vaughan DE, Simpson LM, Gee AP, Taylor DA, Cogle CR, ThomasJD, Silva GV, Jorgenson BC, Olson RE, Bowman S, Francescon J,Geither C, Handberg E, Smith DX, Baraniuk S, Piller LB, Loghin C,Aguilar D, Richman S, Zierold C, Bettencourt J, Sayre SL, VojvodicRW, Skarlatos SI, Gordon DJ, Ebert RF, Kwak M, Moye LA, SimariRD. Effect of intracoronary delivery of autologous bone marrow mono-nuclear cells 2 to 3 weeks following acute myocardial infarction on leftventricular function: the LateTIME randomized trial. JAMA 306: 2110–2119, 2011.
169. Traverse JH, Henry TD, Vaughan DE, Ellis SG, Pepine CJ, Willer-son JT, Zhao DX, Piller LB, Penn MS, Byrne BJ, Perin EC, Gee AP,Hatzopoulos AK, McKenna DH, Forder JR, Taylor DA, Cogle CR,Olson RE, Jorgenson BC, Sayre SL, Vojvodic RW, Gordon DJ,Skarlatos SI, Moye LA, Simari RD. Rationale and design for TIME: Aphase II, randomized, double-blind, placebo-controlled pilot trial evalu-ating the safety and effect of timing of administration of bone marrowmononuclear cells after acute myocardial infarction. Am Heart J 158:356–363, 2009.
170. Tse WT, Pendleton JD, Beyer WM, Egalka MC, Guinan EC. Suppressionof allogeneic T-cell proliferation by human marrow stromal cells: implica-tions in transplantation. Transplantation 75: 389–397, 2003.
171. Ueda K, Takano H, Niitsuma Y, Hasegawa H, Uchiyama R, Oka T,Miyazaki M, Nakaya H, Komuro I. Sonic hedgehog is a criticalmediator of erythropoietin-induced cardiac protection in mice. J ClinInvest 120: 2016–2029, 2010.
172. Urbanek K, Torella D, Sheikh F, De Angelis A, Nurzynska D,Silvestri F, Beltrami CA, Bussani R, Beltrami AP, Quaini F, Bolli R,Leri A, Kajstura J, Anversa P. Myocardial regeneration by activationof multipotent cardiac stem cells in ischemic heart failure. Proc NatlAcad Sci USA 102: 8692–8697, 2005.
173. Wagers AJ. The stem cell niche in regenerative medicine. Cell Stem Cell10: 362–369, 2012.
174. Wang F, Guan J. Cellular cardiomyoplasty and cardiac tissue engineer-ing for myocardial therapy. Adv Drug Deliv Rev 62: 784–797, 2010.
175. Wang X, Jameel MN, Li Q, Mansoor A, Qiang X, Swingen C,Panetta C, Zhang J. Stem cells for myocardial repair with use of atransarterial catheter. Circulation 120: S238–S246, 2009.
176. Watt SM, Contreras M. Stem cell medicine: umbilical cord blood andits stem cell potential. Semin Fetal Neonatal Med 10: 209–220, 2005.
177. Wernig M, Meissner A, Foreman R, Brambrink T, Ku M,Hochedlinger K, Bernstein BE, Jaenisch R. In vitro reprogramming offibroblasts into a pluripotent ES-cell-like state. Nature 448: 318–324,2007.
178. Williams A, Hatzistergos K, Carvalho D, Suncion VY, McCall F,Mather J, Morales AR, Sussman MA, Da Silva J, Heldman AW,Hare J. Synergistic effect of human cardiac stem cells and bone marrowmesenchymal stem cells to reduce infarct size and restore cardiacfunction. Circulation 124: A13079, 2011.
179. Williams AR, Trachtenberg B, Velazquez DL, McNiece I, Altman P,Rouy D, Mendizabal AM, Pattany PM, Lopera GA, Fishman J,Zambrano JP, Heldman AW, Hare JM. Intramyocardial stem cellinjection in patients with ischemic cardiomyopathy: functional recoveryand reverse remodeling. Circ Res 108: 792–796, 2011.
180. Xu W, Zhang X, Qian H, Zhu W, Sun X, Hu J, Zhou H, Chen Y.Mesenchymal stem cells from adult human bone marrow differentiateinto a cardiomyocyte phenotype in vitro. Exp Biol Med (Maywood) 229:623–631, 2004.
181. Yang YJ, Qian HY, Huang J, Li JJ, Gao RL, Dou KF, Yang GS,Willerson JT, Geng YJ. Combined therapy with simvastatin and bonemarrow-derived mesenchymal stem cells increases benefits in infarctedswine hearts. Arterioscler Thromb Vasc Biol 29: 2076–2082, 2009.
182. Ye J, Boyle A, Shih H, Sievers RE, Zhang Y, Prasad M, Su H, ZhouY, Grossman W, Bernstein HS, Yeghiazarians Y. Sca-1 cardiosphere-derived cells are enriched for isl1-expressing cardiac precursors andimprove cardiac function after myocardial injury. PLoS One 7: e30329,2012.
183. Yeo Y, Geng W, Ito T, Kohane DS, Burdick JA, Radisic M. Photo-crosslinkable hydrogel for myocyte cell culture and injection. J BiomedMater Res B Appl Biomater 81: 312–322, 2007.
184. Zaruba MM, Soonpaa M, Reuter S, Field LJ. Cardiomyogenic poten-tial of C-kit�-expressing cells derived from neonatal and adult mousehearts. Circulation 121: 1992–2000, 2010.
185. Zeng L, Hu Q, Wang X, Mansoor A, Lee J, Feygin J, Zhang G,Suntharalingam P, Boozer S, Mhashilkar A, Panetta CJ, Swingen C,Deans R, From AH, Bache RJ, Verfaillie CM, Zhang J. Bioenergeticand functional consequences of bone marrow-derived multipotent pro-genitor cell transplantation in hearts with postinfarction left ventricularremodeling. Circulation 115: 1866–1875, 2007.
186. Zhang G, Hu Q, Braunlin EA, Suggs LJ, Zhang J. Enhancing efficacyof stem cell transplantation to the heart with a PEGylated fibrin biom-atrix. Tissue Eng Part A 14: 1025–1036, 2008.
187. Zhang G, Wang X, Wang Z, Zhang J, Suggs L. A PEGylated fibrinpatch for mesenchymal stem cell delivery. Tissue Eng 12: 9–19, 2006.
188. Zhang Y, Sievers RE, Prasad M, Mirsky R, Shih H, Wong ML,Angeli FS, Ye J, Takagawa J, Koskenvuo JW, Springer ML, Gross-man W, Boyle AJ, Yeghiazarians Y. Timing of bone marrow celltherapy is more important than repeated injections after myocardialinfarction. Cardiovasc Pathol 20: 204–212, 2011.
189. Zhu WZ, Hauch KD, Xu C, Laflamme MA. Human embryonic stemcells and cardiac repair. Transplant Rev (Orlando) 23: 53–68, 2009.
190. Zimmet JM, Hare JM. Emerging role for bone marrow derived mes-enchymal stem cells in myocardial regenerative therapy. Basic ResCardiol 100: 471–481, 2005.
191. Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ, Benhaim P,Lorenz HP, Hedrick MH. Multilineage cells from human adipose tissue:implications for cell-based therapies. Tissue Eng 7: 211–228, 2001.
Review
H270 CELL THERAPY AND INFARCTION
AJP-Heart Circ Physiol • doi:10.1152/ajpheart.00221.2012 • www.ajpheart.org
on August 22, 2013
pdf.highwire.org
Dow
nloaded from





























