Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Oct. 2011, p. 5683–5691 Vol. 193, No. 200021-9193/11/$12.00 doi:10.1128/JB.00428-11Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Brucella melitensis Cyclic di-GMP Phosphodiesterase BpdA ControlsExpression of Flagellar Genes�†
Erik Petersen,1 Pallab Chaudhuri,2 Chris Gourley,1 Jerome Harms,1 and Gary Splitter1*Department of Pathobiological Sciences, University of Wisconsin—Madison, 1656 Linden Dr., Madison,
Wisconsin 53706,1 and Genetic Engineering of Bacteria Lab, Division of Bacteriology,Indian Veterinary Research Institute, Izatnagar, India2
Received 29 March 2011/Accepted 7 August 2011
Brucella melitensis encounters a variety of conditions and stimuli during its life cycle—including environ-mental growth, intracellular infection, and extracellular dissemination—which necessitates flexibility of bac-terial signaling to promote virulence. Cyclic-di-GMP is a bacterial secondary signaling molecule that plays animportant role in adaptation to changing environments and altering virulence in a number of bacteria. Toinvestigate the role of cyclic-di-GMP in B. melitensis, all 11 predicted cyclic-di-GMP-metabolizing proteins wereseparately deleted and the effect on virulence was determined. Three of these cyclic-di-GMP-metabolizingproteins were found to alter virulence. Deletion of the bpdA and bpdB genes resulted in attenuation of virulenceof the bacterium, while deletion of the cgsB gene produced a hypervirulent strain. In a Vibrio reporter systemto monitor apparent alteration in levels of cyclic-di-GMP, both BpdA and BpdB displayed a phenotypeconsistent with cyclic-di-GMP-specific phosphodiesterases, while CgsB displayed a cyclic-di-GMP synthasephenotype. Further analysis found that deletion of bpdA resulted in a dramatic decrease in flagellar promoteractivities, and a flagellar mutant showed similar phenotypes to the bpdA and bpdB mutant strains in mousemodels of infection. These data indicate a potential role for regulation of flagella in Brucella melitensis viacyclic-di-GMP.
Brucella spp. are Gram-negative, facultative intracellularpathogens and the causative agents responsible for the zoo-notic disease brucellosis. Brucella melitensis is endemic in manyareas of the world, causing sterility, abortions, and chronicdisease in infected animals. Human infection with B. melitensisprogresses from an initial period of undulating fever to a pro-longed period of chronic infection potentially resulting in en-docarditis, osteomyelitis, and meningitis (45). Brucella spp. areroutinely passed from animals to humans through the con-sumption of contaminated milk or the inhalation of aerosol-ized bacteria generated from aborted placenta or during butch-ering practices (64). While the economic, environmental, andhealth costs of Brucella spp. are high, more remains to beknown about Brucella virulence factors and their regulation.
Brucella spp. must encounter several different environmentsduring the course of an infection. The bacteria first adjustduring transmission from the reservoir (either an infected an-imal or an environmental contamination) to a new host. Later,during infection, Brucella spp. must adapt to intracellular lifein a wide variety of cellular types and tissues, including bothprofessional and nonprofessional phagocytes and organs rang-ing from spleen and heart to placenta (6, 24, 26, 48, 56, 57). Tosurvive this wide range of hostile encounters, Brucella spp.have evolved to swiftly adapt in response to a rapidly changingenvironment.
Recently, work in several systems has identified the bacterialsecondary messenger cyclic-di-GMP (c-di-GMP) as a powerfulmediator used by bacteria to respond to varied environments(75). c-di-GMP is generated by c-di-GMP synthases (diguany-late cyclases) containing GGDEF domains and degraded byc-di-GMP-specific phosphodiesterases containing EAL or HD-GYP domains (60, 62, 63). The number of c-di-GMP-meta-bolizing proteins in bacterial species varies widely, from a sin-gle protein in some species up to nearly 100 in others (59). Thiscomplexity in signaling is also demonstrated in the variety ofphenotypes that are reportedly regulated by c-di-GMP, includ-ing motility, biofilm formation, cell division, virulence, andintracellular survival (reviewed in references 58 and 67).
To date, the role of c-di-GMP in B. melitensis has not beenexamined. The presence of 11 c-di-GMP-metabolizing proteinsencoded by B. melitensis suggests a system of previously un-known regulation in Brucella. Further analysis of the identifiedproteins responsible for c-di-GMP synthesis and degradationfound that deletion of two genes, BMEI1453 (Brucella phos-phodiesterase A [bpdA]) and BMEI1448 (Brucella phosphodi-esterase B [bpdB]), resulted in the attenuation of virulenceduring mouse infection, while the deletion of a third gene,BMEI1520 (c-di-GMP synthase of Brucella [cgsB]), resulted inincreased virulence. Expression of the BpdA and BpdB pro-teins in a Vibrio reporter system for apparent alterations ofc-di-GMP levels resulted in phenotypes typical of c-di-GMP-specific phosphodiesterases. CgsB expression in the sameVibrio system resulted in a phenotype consistent with that of ac-di-GMP synthase. Furthermore, deletion of the bpdA generesulted in a dramatic downregulation of flagellar promoteractivities and a deletion mutant with mutation of several fla-
* Corresponding author. Mailing address: Department of Pathobio-logical Sciences, University of Wisconsin—Madison, 203 AHABS,1656 Linden Dr., Madison, WI 53706. Phone: (608) 262-1837. Fax:(608) 262-7420. E-mail: [email protected].
† Supplemental material for this article may be found at http://jb.asm.org/.
� Published ahead of print on 19 August 2011.
5683

gellar genes also resulted in attenuation of virulence in ourmouse models of infection.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions. The strains and plasmidsused in this study are listed in Table S1 in the supplemental material. Escherichiacoli strain DH5� was used for propagation of cloning plasmids. ccdB Survival E.coli (Invitrogen) was used to propagate plasmids with the Gateway recombina-tion region or ccdB gene. E. coli was grown in Luria-Bertani broth at 37°C.Brucella melitensis strain 16M (ATCC 23456) was used as the wild-type strain. AllB. melitensis strains were maintained on brucella agar and grown in brucellabroth (Becton Dickinson) at 37°C. Vibrio parahaemolyticus strains were grown at30°C in heart infusion medium (Becton Dickinson) supplemented with 15g/literNaCl. Kanamycin (50 �g/ml), zeocin (50 �g/ml for E. coli, 250 �g/ml for Bru-cella), spectinomycin (100 �g/ml), ampicillin (100 �g/ml), chloramphenicol (20�g/ml), and gentamicin (25 �g/ml) were added to the medium as needed.
Identification of B. melitensis c-di-GMP-metabolizing domains. Protein-pro-tein BLAST analysis of the B. melitensis 16M genome was conducted to identifyc-di-GMP-metabolizing domains (3). The c-di-GMP synthase GGDEF domainfrom Caulobacter crescentus protein PleD and the c-di-GMP-specific phosphodi-esterase EAL and HD-GYP domains from Pseudomonas aeruginosa proteinPA2567 and Xanthomonas campestris protein RpfG were used to identify therespective domains in B. melitensis proteins (22, 32, 60). Further analysis ofidentified B. melitensis proteins was conducted using the SMART domain tool(39). Three genes were found to show a phenotype in this study, and to easefurther discussion, these three genes were renamed BMEI1520 (cyclic-di-GMPsynthase of Brucella [cgsB]), BMEI1453 Brucella phosphodiesterase A [bpdA]),and BMEI1448 Brucella phosphodiesterase B [bpdB]).
Generation of deletion mutants of B. melitensis c-di-GMP-metabolizing pro-teins. Deletion mutants of each of the 11 B. melitensis genes found to encode ac-di-GMP-metabolizing domain were generated by double homologous recom-bination, as described previously (50). The oligonucleotides used to generate thedeletion vectors are listed in Table S2 in the supplemental material. In short, thegene of interest and approximately 1 kb of DNA flanking either end of the genewere amplified by PCR. This PCR product was cloned into the pZErO-1 vectorusing KpnI and XhoI restriction sites found in the vector and oligonucleotidescontaining these restriction sites and sequence complementary to the targetedgene. An inverse PCR (IPCR) of the resulting vector was used to remove �95%of the gene of interest, leaving only the pair of 1 kb of flanking DNAs. Akanamycin resistance cassette was cloned into the place of the gene of interestusing PstI sites found flanking the kanamycin resistance cassette in pUC4K(Amersham) and in the IPCR oligonucleotides. The resulting deletion vector waselectroporated into B. melitensis strain 16M. Kanamycin-resistant clones wereselected for the loss of the pZErO-1 vector backbone by testing for zeocinsensitivity, indicating that replacement of the targeted gene with the kanamycincassette had occurred. The resulting clones were tested by PCR for confirmationof deletion of the gene of interest.
For in vivo imaging experiments, bpdA, bpdB, cgsB, and flgBCG fliE genemutants were made in the GR023 background. GR023 is a strain of B. melitensisthat carries the transposon-based Photorhabdus luminescens luxCDABE operonproviding luminescence to the bacterium while preserving virulence (51). Mutantstrains were generated as described above, using either a zeocin or ampicillinmarker for deletion as GR023 is kanamycin resistant. All mutant strains werefound to produce luminescence at the same levels as the wild-type GR023 strain(data not shown).
Mouse infection with B. melitensis c-di-GMP-metabolizing mutants. The vir-ulence of the B. melitensis c-di-GMP-metabolizing mutants was evaluated usingan IRF-1�/� murine model of infection as described previously (52). One millionCFU of overnight brucella broth-grown culture in 0.2 ml of phosphate-bufferedsaline (PBS) of each of the 11 B. melitensis c-di-GMP-metabolizing mutants wasinjected intraperitoneally (i.p.) into groups of 6- to 8-week-old IRF-1�/� mice(n � 7 or 8). Mice were monitored for mortality up to 60 days. Mutants withmutation of three genes (BMEI1448 [bpdB], BMEI1453 [bpdA], and BMEI1520[cgsB]) were found to have altered virulence in the IRF-1�/� murine model.These three mutants and wild-type strain 16M were analyzed for splenic colo-nization of IRF-1�/� mice. Mice were infected as described above. At specifiedtime points, mice (n � 3) were euthanized and their spleens were removed. Thespleens were ground in 3 ml of PBS, and dilutions were plated to determine theamount of CFU in each spleen. Survival of IRF-1�/� mice was analyzed usingthe Mantel-Cox test, and splenic CFU values were analyzed by Student’s t test.
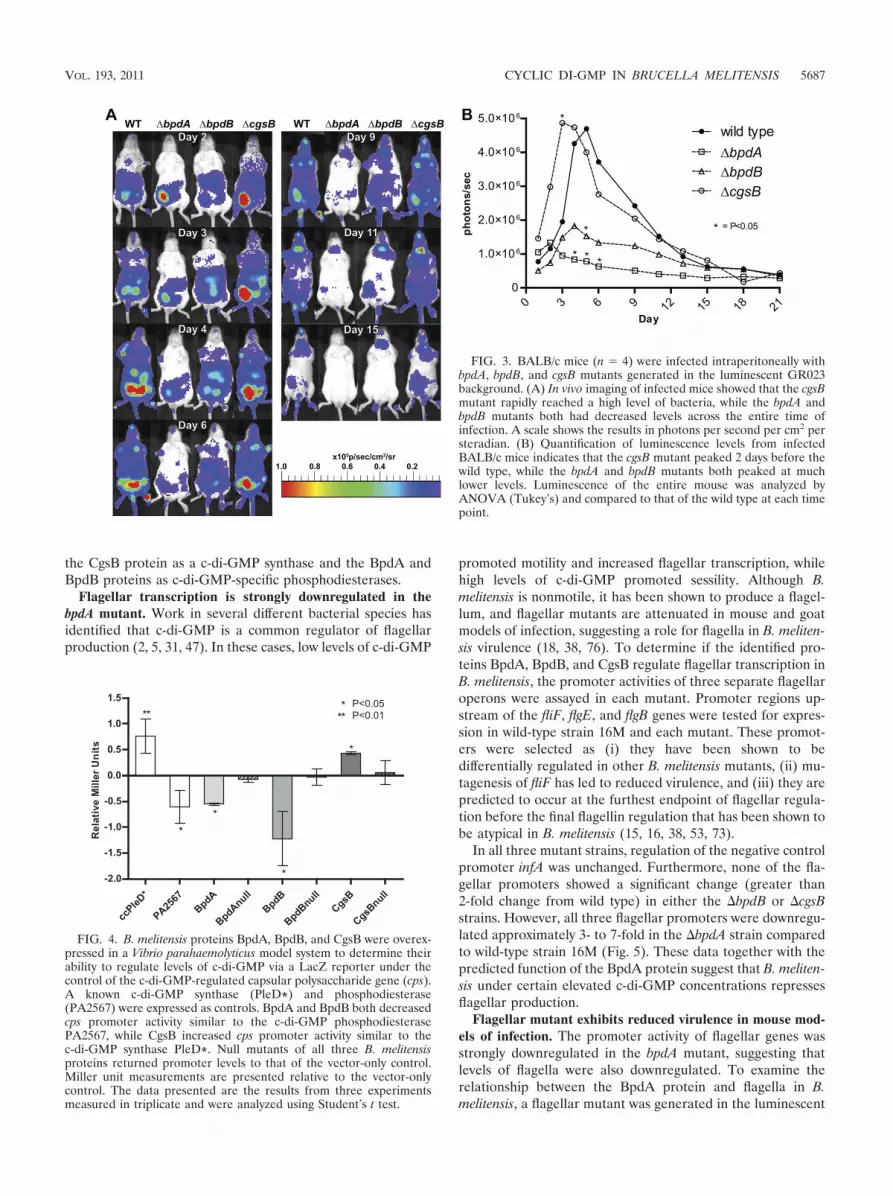
BALB/c mice (n � 4) were infected with luminescent bpdA, bpdB, and cgsBmutants and compared to wild-type B. melitensis GR023 infections. Mice were
infected as described above. Imaging of the mice was done using the Caliper LifeScience in vivo imaging system. Quantification of the level of luminescence ofeach mouse was conducted at each imaging point using Living Image software(Caliper Life Science). Analysis of the data was conducted using analysis ofvariance (ANOVA) with Tukey’s comparison and compared to the wild-typelevel at each time point.
Generation of expression plasmids for a Vibrio parahaemolyticus c-di-GMPreporter system. To determine the activity of the B. melitensis c-di-GMP-metab-olizing proteins identified to play a role in virulence, each protein was expressedin a Vibrio parahaemolyticus reporter system. V. parahaemolyticus strain LM5984contains a genomic transcriptional lacZ fusion to the capsular polysaccharidesynthase (cps) promoter (8, 17, 20, 27). The V. parahaemolyticus cps promoter inthis cps::lacZ strain has been shown to alter levels of LacZ in response tochanging c-di-GMP concentrations induced by expression of c-di-GMP-metab-olizing proteins (17, 27).
Plasmid pLM1877 has been used in V. parahaemolyticus to express proteinsfrom the IPTG(isopropyl-�-D-thiogalactopyranoside)-inducible Ptac promoter(7). To ease cloning of genes into pLM1877, the Gateway recombination regionfrom pRH003 was cloned into pLM1877 downstream of the Ptac promoter, givingplasmid pEP95 (21). Proper orientation was confirmed by restriction enzymedigestion. Full-length B. melitensis proteins were transferred by Gateway reaction(Invitrogen) into pEP95 from pDONR201-based plasmids generated as part ofthe B. melitensis ORFeome (14). As positive controls for the V. parahaemolyticuscps::lacZ system, the c-di-GMP synthase PleD* and the c-di-GMP-specific phos-phodiesterase PA2567 were also expressed. PleD* is a constitutive variant of thec-di-GMP synthase from Caulobacter crescentus and has been shown in several invitro experiments to synthesize c-di-GMP (44, 46, 65). PA2567 is a c-di-GMP-specific phosphodiesterase from Pseudomonas aeruginosa that has been shownexperimentally to degrade c-di-GMP (54, 60).
Mutation of bpdA, bpdB, and cgsB to generate inactive (null) proteins foranalysis in the Vibrio reporter system. The genes encoding the BpdA, BpdB, andCgsB proteins were mutated in an effort to determine whether inactivation of theprotein would result in loss of a phenotype in the Vibrio reporter system. As theBpdB and CgsB proteins only carry transmembrane domains and the c-di-GMP-metabolizing domain, conserved signature motifs in each domain were mutatedby alanine substitution of the active site residues. Site-directed mutagenesis(SDM) oligonucleotides were generated that spanned the active site residues,and nucleotides were replaced in the oligonucleotide that would result in aconversion of the active site residues to alanine residues. The EAL residues ofBpdB were mutated to AAA, while the RMGGEEF residues of CgsB weremutated to AMGAAEF, both of which have been shown to abrogate activity inother domains (9, 28, 55, 68).
pDONR-based plasmids containing either the bpdB or cgsB gene were ex-tracted from a dam/dcm methylation-competent background to ensure parentDNA was methylated. PCR of the entire plasmid using the SDM oligonucleo-tides followed by methylation-specific restriction digest of the parent plasmidresulted in bpdB and cgsB active site mutations that were confirmed by DNAsequencing. These plasmids were then used in Gateway-based recombination forfuture use in the Vibrio reporter system.
As BpdA contains several regulatory domains in addition to the EAL phos-phodiesterase domain, the EAL domain was simply removed from the protein,leaving the transmembrane domain cluster, PAS fold, and inactive GGDEFdomain. Oligonucleotides corresponding to the start codon and to the codonsimmediately upstream of the EAL domain were used to clone the C-terminaltruncated bpdA gene into the plasmid pCR8 (Invitrogen). DNA sequencingconfirmed deletion of the EAL domain. This plasmid was also used in Gateway-based recombination reactions for use in the Vibrio reporter system.
Analysis of B. melitensis c-di-GMP-metabolizing protein activity using theVibrio reporter system. The expression plasmids generated above were trans-formed into the V. parahaemolyticus cps::lacZ strain (LM5984). Each strain wasspread on a heart infusion agar plate, harvested after 18 h of growth at 30°C, andassayed for LacZ expression as described by Miller (43). Briefly, the A600 of theharvested bacteria was determined, and a 1:10 dilution was lysed using Koch’slysis solution (49). The resulting preparation was incubated with ortho-nitrophe-nyl-�-galactoside (ONPG) at a final concentration of 0.67 mg/ml. After thereaction, the breakdown of ONPG was determined by measuring the A420. Millerunits were quantified as (1,000 � A420)/(A600 � 0.1 � t), where t is the time ofreaction, and relative Miller units were calculated by determining the fold dif-ference from the vector-only control.
As positive controls for the Vibrio assay, the genes encoding the constitutivelyactive Caulobacter crescentus protein PleD* and the Pseudomonas aeruginosaPA01 protein PA2567 were cloned into the Gateway-ready donor vector pCR8according to the manufacturer’s instructions (Invitrogen). The PleD* and
5684 PETERSEN ET AL. J. BACTERIOL.

PA2567 proteins, respectively, generate and degrade c-di-GMP in both in vitroand in vivo assays (2, 44, 46, 54, 60). All samples were measured in triplicate, andthe results of three replicate experiments are graphed. Data were analyzed usingStudent’s t test.
Generation of luminescent reporter plasmids to monitor activity of threeflagellar promoters within Brucella. Expression from three flagellar promoters inthe bpdA, bpdB, and cgsB mutants and wild-type strain 16M were measured usingthe luminescent reporter vector pEP3 (53). Plasmid pEP3 contains a promoter-less luxCDABE operon and a multiple-cloning site (MCS) upstream of theoperon for insertion of promoters of interest. Three separate flagellar promoters(fliF, flgE, and flgB) were selected for analysis, and the infA (initiation factor A)promoter was used as a negative control (53). Approximately 500 bp upstream ofthe respective start codon was cloned into the MCS of pEP3 upstream of theluxCDABE operon in order to encompass a majority of the potential transcrip-tion factor binding sites (11). The four reporter plasmids were electroporatedinto the bpdA, bpdB, and cgsB mutants and the 16M wild-type strain for analysis.
Flagellar promoter analysis in the bpdA, bpdB, and cgsB mutants using aluminescent reporter system. Strains were grown to stationary phase and thendiluted to an optical density (OD) of 0.1 in brucella broth. Samples were platedin triplicate into a 96-well plate, and then the plate was transferred to a DTX 880plate reader (Beckman Coulter). Samples were grown to an OD595 of 0.2, atwhich point the luminescence and OD595 values were recorded. The experimentwas repeated in triplicate from three separate cultures grown from a streakedplate. To quantify luminescence of the promoter strains, relative light units(RLU) were calculated by determining the luminescence/OD595. The ratio ofpromoter activity in each mutant to wild-type activity was then calculated to givethe fold difference in RLU for each promoter. Data were analyzed by Student’st test.
RESULTS
Brucella melitensis encodes 11 c-di-GMP-metabolizing pro-teins. c-di-GMP has been identified within several bacterialspecies as a secondary signaling molecule that regulates a shiftbetween various lifestyles. To determine what role c-di-GMPplays in B. melitensis, BLAST analysis was conducted on thepublished 16M genome to identify c-di-GMP-metabolizingproteins (3). c-di-GMP is generated by c-di-GMP synthases(diguanylate cyclases) containing a GGDEF domain, whiledegradation of c-di-GMP is governed by c-di-GMP-specificphosphodiesterases containing either an EAL domain or anHD-GYP domain. Analysis of the B. melitensis 16M genomeidentified 11 genes that encode a GGDEF and/or EAL domainand none with an HD-GYP domain (Fig. 1). Six proteins con-tain a GGDEF domain, two proteins contain an EAL domain,and three proteins contain both a GGDEF domain and anEAL domain. Analysis of the latter group identified one gene,bpdA (BMEI1453), in which the usually conserved residues inthe c-di-GMP synthase active site (GG[D/E]EF) instead en-code the amino acids SSDQF. Mutation of GGDEF domainactive site residues abrogates c-di-GMP synthase activity inother proteins and was therefore predicted to be inactive inBpdA (9, 28, 40).
Two c-di-GMP-metabolizing B. melitensis mutants are atten-uated and one mutant is hypervirulent in a mouse model ofBrucella infection. Mutants with deletion of each of the 11c-di-GMP-metabolizing proteins were generated in the B.melitensis 16M parent strain as described in Materials andMethods. Growth of each of the mutants in brucella broth anda minimal medium was identical to that of the parent strain. Togauge any attenuation in the mutant strains to infect and rep-licate within phagocytic cells, each was tested for intracellularlevels after infection of RAW 264.7 macrophages. Each strainexhibited similar levels to the parent strain during macrophageinfection, indicating that intracellular infection and growth
were not dramatically affected (data not shown). To furthergauge the level of virulence of each of the mutants, an IRF-1�/� mouse model assay was conducted. IRF-1�/� mice havebeen used previously to successfully determine whether Bru-cella mutants are still virulent, as these immunocompromisedmice succumb to wild-type B. melitensis infection but surviveinfection with a variety of attenuated mutants (29, 30). For thisassay, IRF-1�/� mice (n � 8) were infected with each of the 11mutants or the parent strain, and the time point to death afterinfection was determined. Three c-di-GMP-metabolizing mu-tants possessed a significant change in virulence (Fig. 2A).Mice infected with the wild-type strain 16M have a mean timeto death of 10.7 � 0.5 days. The bpdA mutant was avirulent,with all mice living until the end of the experiment at 60 days.Mice infected with the bpdB mutant possessed intermediatevirulence, with mice dying on average a week later than theparent strain-infected mice, resulting in a mean time to deathof 18.0 � 1.3 days. cgsB mutant-infected mice displayed ahypervirulent phenotype, dying several days earlier than theparent strain-infected mice, with a mean time to death of 8.3 �0.5 days. Complementation with a plasmid-based copy of thedeleted gene fully restored the phenotype of the bpdA and cgsBmutants to wild-type levels and restored the phenotype of thebpdB mutant to a statistically nonsignificant level (see Fig. S1in the supplemental material).
Further characterization of the phenotype of the bpdA,bpdB, and cgsB mutants was accomplished by determining lev-els of bacteria in infected spleens at time points during IRF-1�/� mouse infection (Fig. 2B). Consistent with the shortertime to death and increase in virulence, mice infected with thecgsB hypervirulent mutant contained higher initial levels ofsplenic bacteria than the parent strain. Mice infected with thebpdB mutant had very low initial levels of splenic bacteria, butthe splenic bacterial levels eventually increased and killed the
FIG. 1. BLAST analysis of the B. melitensis 16M genome identified11 genes that encode proteins predicted to regulate levels of c-di-GMP. Six genes encode a c-di-GMP synthase (GGDEF) domain, twogenes encode a c-di-GMP phosphodiesterase (EAL) domain, andthree genes encode both domains. Several other regulatory domainsare also present, including REC domains (response regulator phos-phorylation), PAS/PAC domains (cofactor regulatory domain, i.e.,heme, flavin, etc.), and GAF domains (cyclic nucleotide or small mol-ecule binding). Horizontal bars indicate predicted transmembrane he-lices.
VOL. 193, 2011 CYCLIC DI-GMP IN BRUCELLA MELITENSIS 5685

mice. bpdA mutant-infected mice also had lower levels ofsplenic bacteria, but those levels stayed fairly consistent for 2weeks before declining over the course of the 60-day infection.However, these remaining levels of splenic bacteria are fargreater than those exhibited by strains exhibiting rough lipo-polysaccharide mutations or mutations in the type IV secretionsystem gene virB, two commonly studied avirulent Brucellamutants (data not shown).
bpdA and bpdB mutants exhibit decreased disseminationwithin immunocompetent mice. bpdA, bpdB, and cgsB deletionmutants were generated in the GR023 luminescent back-ground (51). These mutants were used to infect BALB/c miceto examine the virulence and dissemination of the mutants inan immunocompetent mouse (Fig. 3). Unlike the immunocom-promised IRF-1�/� mice, BALB/c mice typically exhibit a peaklevel of bacteria between 5 and 7 days postinfection, followedby the subsequent clearance below detectable levels. The wild-type GR023 strain possessed classical dissemination and viru-lence levels, initially localizing near the site of injection (lower
left abdomen), spreading to the rest of the mouse, peaking atday 5, and declining by the second week. The hypervirulentcgsB mutant infected at a higher level than the wild type,disseminating and peaking before the wild type at day 3. ThebpdB mutant spread throughout the mouse similarly to the wildtype, but at a lower level of infection, similar to observations inthe IRF-1�/� mice. The highly attenuated bpdA mutant neverdisseminated far from the site of injection before rapidly de-creasing.
CgsB exhibits the c-di-GMP synthase phenotype, whileBpdA and BpdB exhibit the c-di-GMP phosphodiesterase phe-notype in a Vibrio reporter system. After determining thatmutants with deletion of the three B. melitensis c-di-GMP-metabolizing proteins BpdA, BpdB, and CgsB possessed asignificant phenotype in mouse models of Brucella infection,the functional activity of the proteins toward c-di-GMP wasdetermined. Vibrio spp. have been well studied with regard toc-di-GMP due to the very recognizable phenotypes displayedunder high-c-di-GMP (biofilm formation) and low-c-di-GMP(motility) conditions. A V. parahaemolyticus strain with a lacZtranscriptional fusion to the capsular polysaccharide synthasepromoter (cps) was used for analysis of cps promoter activity(20). Several previously published reports have indicated thatthe level of LacZ production from the cpsA3::lacZ-Genr fusionis directly tied to the level of cyclic-di-GMP that has beenaltered by the expression of c-di-GMP-metabolizing proteins(17, 27).
Two control proteins (the constitutive c-di-GMP synthasePleD* and the phosphodiesterase PA2567) and the three B.melitensis proteins (BpdA, BpdB, and CgsB) were expressedwithin the V. parahaemolyticus cpsA3::lacZ-Genr strain (Fig. 4).PleD* is a constitutively active c-di-GMP synthase from Cau-lobacter crescentus that has been shown in several assays togenerate c-di-GMP (44, 46, 65). The increased level of LacZactivity seen with expression of PleD* is consistent with thepredicted result of expression of a c-di-GMP synthase whichgenerates c-di-GMP, increasing activity from the cps promoterand generating a higher level of LacZ activity. Similarly, ex-pression of CgsB supported the fact that it was a functionalc-di-GMP synthase as it exhibited higher levels of LacZ activ-ity, similar to that of the control PleD*. Expression of theprotein PA2567 from Pseudomonas aeruginosa resulted in adecrease in LacZ activity, consistent with the expected resultsof expression of a c-di-GMP-phosphodiesterase in which alowering of c-di-GMP levels by PA2567 reduces the activity ofthe cps promoter (54, 60). Similarly, expression of both BpdAand BpdB decreased levels of LacZ activity similar to that ofPA2567, supporting the case that both of these proteins arec-di-GMP phosphodiesterases.
To further characterize the BpdA, BpdB, and CgsB proteins,null mutants of each were generated and expressed in theVibrio reporter system. The BpdB and CgsB null mutants eachcontained alanine substitutions in the active site residues oftheir respective c-di-GMP-metabolizing domains. The BpdAnull mutant had the entire EAL domain truncated from theC-terminal portion of the protein, leaving the transmembranedomains, PAS fold, and the inactive GGDEF domain. Allthree null mutants had their activity reduced to the level of thevector-only control, further supporting our characterization of
FIG. 2. bpdA and bpdB mutants are attenuated in IRF-1�/� mice,while the cgsB mutant is hypervirulent. (A) IRF-1�/� mice (n � 7 or 8)were infected with mutants containing mutations of each of the 11identified c-di-GMP-metabolizing genes. Three mutants possessed asignificant change in the amount of time that it took for the mice tosuccumb to infection. (B) Spleens from IRF-1�/� mice (n � 3 or 4/timepoint) infected with each of the three mutants were removed at thespecified time points and plated for CFU determinations. The cgsBstrain had a higher initial level of CFU, the bpdB strain had lowerinitial level until eventually reaching the wild-type level, and the bpdAstrain had lower but persistent level throughout the course of infection.Survival data were analyzed using the Mantel-Cox test, and CFU datawere analyzed using Student’s t test.
5686 PETERSEN ET AL. J. BACTERIOL.
the CgsB protein as a c-di-GMP synthase and the BpdA andBpdB proteins as c-di-GMP-specific phosphodiesterases.
Flagellar transcription is strongly downregulated in thebpdA mutant. Work in several different bacterial species hasidentified that c-di-GMP is a common regulator of flagellarproduction (2, 5, 31, 47). In these cases, low levels of c-di-GMP
promoted motility and increased flagellar transcription, whilehigh levels of c-di-GMP promoted sessility. Although B.melitensis is nonmotile, it has been shown to produce a flagel-lum, and flagellar mutants are attenuated in mouse and goatmodels of infection, suggesting a role for flagella in B. meliten-sis virulence (18, 38, 76). To determine if the identified pro-teins BpdA, BpdB, and CgsB regulate flagellar transcription inB. melitensis, the promoter activities of three separate flagellaroperons were assayed in each mutant. Promoter regions up-stream of the fliF, flgE, and flgB genes were tested for expres-sion in wild-type strain 16M and each mutant. These promot-ers were selected as (i) they have been shown to bedifferentially regulated in other B. melitensis mutants, (ii) mu-tagenesis of fliF has led to reduced virulence, and (iii) they arepredicted to occur at the furthest endpoint of flagellar regula-tion before the final flagellin regulation that has been shown tobe atypical in B. melitensis (15, 16, 38, 53, 73).
In all three mutant strains, regulation of the negative controlpromoter infA was unchanged. Furthermore, none of the fla-gellar promoters showed a significant change (greater than2-fold change from wild type) in either the bpdB or cgsBstrains. However, all three flagellar promoters were downregu-lated approximately 3- to 7-fold in the bpdA strain comparedto wild-type strain 16M (Fig. 5). These data together with thepredicted function of the BpdA protein suggest that B. meliten-sis under certain elevated c-di-GMP concentrations repressesflagellar production.
Flagellar mutant exhibits reduced virulence in mouse mod-els of infection. The promoter activity of flagellar genes wasstrongly downregulated in the bpdA mutant, suggesting thatlevels of flagella were also downregulated. To examine therelationship between the BpdA protein and flagella in B.melitensis, a flagellar mutant was generated in the luminescent
FIG. 4. B. melitensis proteins BpdA, BpdB, and CgsB were overex-pressed in a Vibrio parahaemolyticus model system to determine theirability to regulate levels of c-di-GMP via a LacZ reporter under thecontrol of the c-di-GMP-regulated capsular polysaccharide gene (cps).A known c-di-GMP synthase (PleD*) and phosphodiesterase(PA2567) were expressed as controls. BpdA and BpdB both decreasedcps promoter activity similar to the c-di-GMP phosphodiesterasePA2567, while CgsB increased cps promoter activity similar to thec-di-GMP synthase PleD*. Null mutants of all three B. melitensisproteins returned promoter levels to that of the vector-only control.Miller unit measurements are presented relative to the vector-onlycontrol. The data presented are the results from three experimentsmeasured in triplicate and were analyzed using Student’s t test.
FIG. 3. BALB/c mice (n � 4) were infected intraperitoneally withbpdA, bpdB, and cgsB mutants generated in the luminescent GR023background. (A) In vivo imaging of infected mice showed that the cgsBmutant rapidly reached a high level of bacteria, while the bpdA andbpdB mutants both had decreased levels across the entire time ofinfection. A scale shows the results in photons per second per cm2 persteradian. (B) Quantification of luminescence levels from infectedBALB/c mice indicates that the cgsB mutant peaked 2 days before thewild type, while the bpdA and bpdB mutants both peaked at muchlower levels. Luminescence of the entire mouse was analyzed byANOVA (Tukey’s) and compared to that of the wild type at each timepoint.
VOL. 193, 2011 CYCLIC DI-GMP IN BRUCELLA MELITENSIS 5687

GR023 background. A four-gene operon encoding flagellarproteins that constitute the transmembrane flagellar rod (flg-BCGfliE, corresponding to BMEI1086 to -9, respectively) wasdeleted, and the mutant was examined alongside the bpdAmutant in both the BALB/c and IRF-1�/� mouse models ofBrucella infection (Fig. 6).
In vivo imaging of BALB/c mice (n � 4) with the lumines-cent mutant shows that initial infection with the flagellar mu-tant is also restricted to the site of injection (lower left, i.p.).While the flagellar mutant eventually lightly disseminated out-side the site of injection, it never reached the level of wild-typeinfection (Fig. 6A). Infection of immunocompromised IRF-1�/� mice with the flagellar mutant resulted in attenuatedvirulence, with the mice eventually succumbing to infection,with a mean time to death of 19.0 � 4.8 days (Fig. 6B). These
data are in contrast to bpdA mutant-infected mice that neversuccumbed to infection, possibly indicating that additional fac-tors are regulated by the BpdA protein in addition to flagellathat result in additional attenuation of virulence.
DISCUSSION
The identification of c-di-GMP-metabolizing proteins thatplay a role in virulence of B. melitensis suggests a previouslyunidentified mechanism(s) of regulation of virulence. Signa-ture-tagged mutagenesis of B. melitensis during acute infectionfirst identified the “EAL-domain hypothetical protein” BpdB(BMEI1448) as a potential regulator of virulence (38). Thiswork has identified a second c-di-GMP-specific phosphodies-terase, BpdA (BMEI1453), that shows an even further level ofattenuation of virulence, and a c-di-GMP synthase, CgsB(BMEI1520), that shows increased virulence in mouse modelsof infection.
Together with the phenotypes seen during expression of theB. melitensis proteins in the Vibrio reporter system for c-di-GMP regulation, we can begin to hypothesize what role c-di-GMP may play in Brucella virulence. Deletion of the cgsBc-di-GMP synthase would be expected to lower potential levelsof c-di-GMP, leading to an increase in virulence. On the otherhand, deletion of the bpdA or bpdB c-di-GMP phosphodies-terases would potentially increase the level of c-di-GMP andresult in a corresponding decrease in virulence. This hypothesiswould be consistent with that proposed for other models inwhich production of virulence factors (i.e., cholera toxin andflagella) requires a reduction in c-di-GMP and that loss ofthese virulence factors is accompanied by a decrease in viru-lence (5, 10, 35, 71).
Further analysis of the bpdA mutant indicated that activa-tion levels of several flagellar promoters were strongly down-regulated compared to wild-type levels, suggesting that flagel-lar production is decreased in the mutant. Previously publishedresults indicate that flagellar mutants of B. melitensis do not
FIG. 5. Loss of BpdA results in a 3- to 7-fold decrease in activity ofthree flagellar promoters. Three separate flagellar operons were testedfor promoter activity in the wild-type strain 16M and mutant strainsusing a luminescent reporter assay. All three operons were highlydownregulated in the bpdA strain, indicating that the BpdA proteinregulates flagellar gene expression in B. melitensis. Promoter levelswere graphed as fold difference relative to wild-type levels of promoteractivity. The data presented are the results from three experiments,each measured in triplicate and analyzed using Student’s t test.
FIG. 6. Deletion of B. melitensis flagellar genes results in attenuation in virulence in mouse models of infection. A four-gene operon encodingflagellar components of the central rod (flgB, flgC, flgG, and fliE) was deleted in the GR023 luminescent, virulent B. melitensis strain. (A) Infectionof BALB/c mice (n � 4) with the flagellar mutant strain results in an attenuated phenotype in which the bacteria fail to fully disseminate andinstead localize at the site of infection. A scale shows the results in photons per second per cm2 per steradian. (B) Infection of IRF-1�/� mice withthe flagellar mutant results in prolonged survival but eventual death of the mice.
5688 PETERSEN ET AL. J. BACTERIOL.

show a phenotype different from the wild type in cellular mod-els of infection, similar to the bpdA mutant, but do show aphenotype in mice (18, 38). To establish a correlation betweenflagellar production and virulence, a flagellar deletion mutantwas generated and compared to the bpdA mutants. While theflagellar mutant disseminates similar to the bpdA mutant, IRF-1�/� mice infected with the flagellar mutant still succumb toinfection while bpdA mutant-infected mice do not, potentiallyindicating a role of the BpdA protein in virulence in additionto flagellar regulation (18, 38). These data suggest a possiblerole for c-di-GMP in the regulation of virulence through fla-gellar expression in B. melitensis.
The role of flagella in B. melitensis is still an evolving field.Lack of a chemotactic system, truncation of several flagellargenes, and historical classification as a nonmotile bacteriummeant that identification of flagellar genes within the genomeof B. melitensis was initially hypothesized to either be the resultof cryptic remnants or as part of a type III secretory system (1,13). More recent work found that not only does B. melitensisproduce a sheathed flagellum under the proper conditions, butdeletion of flagellar genes results in attenuation of virulence inboth mouse and goat models of infection (18, 38, 76). Ratherthan motility, it is possible that B. melitensis uses flagella forattachment, secretion, or as a source of immune system acti-vation through either Toll-like receptor 5 (TLR5) or IpaF (42).
The exact role of c-di-GMP in the regulation of flagella in B.melitensis is still unknown. Flagella are often regulated in tiers,with the proteins of the hook-basal body occupying tier II,downstream of regulatory factors in tier I but upstream of thechemotaxis and filament proteins of tier III (12). The masterregulator FtcR was found to regulate expression of severalflagellar genes, possibly under the control of the quorum-sens-ing regulator VjbR (37). A second quorum-sensing regulator,BlxR, also regulates expression levels of flagellar genes (53).Recent work has furthered our knowledge of the regulation offlagella in B. melitensis, identifying both a sigma factor(RpoE1) that represses expression of flagellar genes and aflagellin activator responsible for regulation of the fliC gene(FlbT) (15, 16). As the flagellin filament protein FliC wasshown to be produced even in B. melitensis mutants lackingassembly of the flagellar hook, promoter activity of third tierproteins was not examined in this study (16). By selectingstructural proteins downstream of several regulatory check-points for our assay, the cumulative effect of c-di-GMP onflagellar expression was determined. However, detailed workat each of the checkpoints in flagellar synthesis will be requiredto pinpoint the location(s) of c-di-GMP involvement.
The mechanism of signaling achieved by c-di-GMP in B.melitensis is also unknown. Three separate mechanisms havebeen shown in other bacterial species, although it is likely thatmore remain. The first is at the protein level, where c-di-GMPbinds directly to a protein and activates an adjacent enzymaticor regulatory domain. These proteins include PilZ domain-encoding proteins in components of cellulose synthase andtype IV pilin apparatus and the PelD polysaccharide regulator(4, 36, 61, 74). Unfortunately, none of these c-di-GMP-bindingdomains show homology to any B. melitensis protein. The sec-ond method of regulation is the recently discovered c-di-GMPriboswitch. This upstream mRNA element regulates termina-tor formation in the mRNA based upon binding to c-di-GMP
(66). Again though, previous sequence analysis of the B.melitensis genome did not identify any c-di-GMP-responsiveriboswitches (66). The third mechanism of c-di-GMP regula-tion is through the binding of c-di-GMP to transcription fac-tors like the FleQ flagellar synthesis regulator and cyclic AMPreceptor protein (CRP)-like proteins (Clp) (23). While B.melitensis doesn’t encode a protein with any similarity to theFleQ protein, it encodes several proteins with homology to theClp family of proteins. A subset of these B. melitensis Clpproteins conserve a glutamate residue shown to be required forc-di-GMP binding, but further analysis of these proteins wouldbe required to suggest that they possess the ability to bindc-di-GMP (69).
In addition to regulation of flagella, it is likely that othercellular processes are controlled by c-di-GMP in B. melitensis.CgsB and BpdB do not appear to affect flagellar production inour assay under the conditions tested, but both contribute tothe virulence of the bacterium. BMEII0660 encodes a putativec-di-GMP synthase with homology to the family of PleD re-sponse regulators whose activity has been shown to play a rolein motility and replication in Caulobacter crescentus and hostcell infection and survival in Anaplasma phagocytophilum andEhrlichia chaffeensis (22, 33, 34). Additional c-di-GMP-meta-bolizing proteins encoded by B. melitensis also contain domainslike PAS and GAF domains that have been found in otherinstances to respond to oxygen levels, redox, and cyclic nucle-otides (41, 70). Recent reports have also identified the abilityof B. melitensis to form biofilms after certain genetic modifi-cations, another network commonly regulated by c-di-GMP(19, 25, 72). While there have been no identified instances inwhich B. melitensis is naturally found in biofilms, it is possiblethat B. melitensis uses c-di-GMP to regulate production of abiofilm in specific, yet unidentified, environments.
In conclusion, we have identified three B. melitensis proteinsthat actively regulate levels of the secondary messenger c-di-GMP, opening up a wealth of new possibilities for B. melitensisregulation. A c-di-GMP-specific phosphodiesterase, BpdA,was found to regulate the expression of several flagellar com-ponents, suggesting a role for c-di-GMP in the regulation ofthe B. melitensis flagellum. Further experimentation will seekto identify the mechanisms and stimuli of this regulation. Thepresence of an additional 8 predicted c-di-GMP-metabolizingproteins indicates that there may be several additional path-ways regulated by c-di-GMP that are yet to be uncovered.
ACKNOWLEDGMENTS
This work was supported by NIH 1R01AI073558, NIH R21AI088038, and BARD grant US-4378-11. Erik Petersen was supportedby a Molecular Biosciences Training grant (NIH T32 GM07215-34).
We also thank Urs Jenal at the University of Basel, Switzerland, forthe gift of the PleD* gene, Linda McCarter at the University of Iowafor the Vibrio parahaemolyticus strains, and Susan West at the Univer-sity of Wisconsin—Madison for the Pseudomonas aeruginosa PA01strain. Plasmid construction was aided by Diogo Magnani. Vibrio para-haemolyticus Miller assays were accomplished with help from MichaelCho.
REFERENCES
1. Abdallah, A. I., et al. 2003. Type III secretion homologs are present inBrucella melitensis, B. ovis, and B. suis biovars 1, 2, and 3. Curr. Microbiol.46:241–245.
2. Aldridge, P., R. Paul, P. Goymer, P. Rainey, and U. Jenal. 2003. Role of the
VOL. 193, 2011 CYCLIC DI-GMP IN BRUCELLA MELITENSIS 5689

GGDEF regulator PleD in polar development of Caulobacter crescentus.Mol. Microbiol. 47:1695–1708.
3. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990.Basic local alignment search tool. J. Mol. Biol. 215:403–410.
4. Amikam, D., and M. Y. Galperin. 2006. PilZ domain is part of the bacterialc-di-GMP binding protein. Bioinformatics 22:3–6.
5. Beyhan, S., A. D. Tischler, A. Camilli, and F. H. Yildiz. 2006. Transcriptomeand phenotypic responses of Vibrio cholerae to increased cyclic di-GMP level.J. Bacteriol. 188:3600–3613.
6. Billard, E., C. Cazevieille, J. Dornand, and A. Gross. 2005. High suscepti-bility of human dendritic cells to invasion by the intracellular pathogensBrucella suis, B. abortus, and B. melitensis. Infect. Immun. 73:8418–8424.
7. Boles, B. R., and L. L. McCarter. 2000. Insertional inactivation of genesencoding components of the sodium-type flagellar motor and switch of Vibrioparahaemolyticus. J. Bacteriol. 182:1035–1045.
8. Boles, B. R., and L. L. McCarter. 2002. Vibrio parahaemolyticus scrABC, anovel operon affecting swarming and capsular polysaccharide regulation. J.Bacteriol. 184:5946–5954.
9. Christen, M., B. Christen, M. Folcher, A. Schauerte, and U. Jenal. 2005.Identification and characterization of a cyclic di-GMP-specific phosphodies-terase and its allosteric control by GTP. J. Biol. Chem. 280:30829–30837.
10. Claret, L., et al. 2007. The flagellar sigma factor Flia regulates adhesion andinvasion of Crohn’s disease-associated Escherichia coli via a c-di-GMP-de-pendent pathway. J. Biol. Chem. 282:33275–33283.
11. Collado-Vides, J., B. Magasanik, and J. D. Gralla. 1991. Control site loca-tion and transcriptional regulation in Escherichia coli. Microbiol. Rev. 55:371–394.
12. Dasgupta, N., et al. 2003. A four-tiered transcriptional regulatory circuitcontrols flagellar biogenesis in Pseudomonas aeruginosa. Mol. Microbiol.50:809–824.
13. DelVecchio, V. G., et al. 2002. Brucella proteomes—a review. Vet. Microbiol.90:593–603.
14. Dricot, A., et al. 2004. Generation of the Brucella melitensis ORFeomeversion 1.1. Genome Res. 14:2201–2206.
15. Ferooz, J., J. Lemaire, M. Delory, X. De Bolle, and J. J. Letesson. 2011.RpoE1, an extracytoplasmic sigma factor, is a repressor of the flagellarsystem in Brucella melitensis. Microbiology 157:1263–1268.
16. Ferooz, J., J. Lemaire, and J. J. Letesson. 2011. Role of FlbT in flagellinproduction in Brucella melitensis. Microbiology 157:1253–1262.
17. Ferreira, R. B., L. C. Antunes, E. P. Greenberg, and L. L. McCarter. 2008.Vibrio parahaemolyticus ScrC modulates cyclic dimeric GMP regulation ofgene expression relevant to growth on surfaces. J. Bacteriol. 190:851–860.
18. Fretin, D., et al. 2005. The sheathed flagellum of Brucella melitensis isinvolved in persistence in a murine model of infection. Cell. Microbiol.7:687–698.
19. Godefroid, M., et al. 2010. Brucella melitensis 16M produces a mannan andother extracellular matrix components typical of a biofilm. FEMS Immunol.Med. Microbiol. 59:364–377.
20. Guvener, Z. T., and L. L. McCarter. 2003. Multiple regulators control cap-sular polysaccharide production in Vibrio parahaemolyticus. J. Bacteriol. 185:5431–5441.
21. Hallez, R., J. J. Letesson, J. Vandenhaute, and X. De Bolle. 2007. Gateway-based destination vectors for functional analyses of bacterial ORFeomes:application to the Min system in Brucella abortus. Appl. Environ. Microbiol.73:1375–1379.
22. Hecht, G. B., and A. Newton. 1995. Identification of a novel response regu-lator required for the swarmer-to-stalked-cell transition in Caulobacter cres-centus. J. Bacteriol. 177:6223–6229.
23. Hickman, J. W., and C. S. Harwood. 2008. Identification of FleQ fromPseudomonas aeruginosa as a c-di-GMP-responsive transcription factor. Mol.Microbiol. 69:376–389.
24. Hong, P. C., R. M. Tsolis, and T. A. Ficht. 2000. Identification of genesrequired for chronic persistence of Brucella abortus in mice. Infect. Immun.68:4102–4107.
25. Jonas, K., O. Melefors, and U. Romling. 2009. Regulation of c-di-GMPmetabolism in biofilms. Future Microbiol. 4:341–358.
26. Kim, S., et al. 2005. Interferon-gamma promotes abortion due to Brucellainfection in pregnant mice. BMC Microbiol. 5:22.
27. Kim, Y. K., and L. L. McCarter. 2007. ScrG, a GGDEF-EAL protein,participates in regulating swarming and sticking in Vibrio parahaemolyticus. J.Bacteriol. 189:4094–4107.
28. Kirillina, O., J. D. Fetherston, A. G. Bobrov, J. Abney, and R. D. Perry. 2004.HmsP, a putative phosphodiesterase, and HmsT, a putative diguanylatecyclase, control Hms-dependent biofilm formation in Yersinia pestis. Mol.Microbiol. 54:75–88.
29. Ko, J., A. Gendron-Fitzpatrick, T. A. Ficht, and G. A. Splitter. 2002. Viru-lence criteria for Brucella abortus strains as determined by interferon regu-latory factor 1-deficient mice. Infect. Immun. 70:7004–7012.
30. Ko, J., A. Gendron-Fitzpatrick, and G. A. Splitter. 2002. Susceptibility ofIFN regulatory factor-1 and IFN consensus sequence binding protein-defi-cient mice to brucellosis. J. Immunol. 168:2433–2440.
31. Kuchma, S. L., et al. 2007. BifA, a c-di-GMP phosphodiesterase, inversely
regulates biofilm formation and swarming motility by Pseudomonas aerugi-nosa PA14. J. Bacteriol. 189:8165–8178.
32. Kulesekara, H., et al. 2006. Analysis of Pseudomonas aeruginosa diguanylatecyclases and phosphodiesterases reveals a role for bis-(3-5)-cyclic-GMP invirulence. Proc. Natl. Acad. Sci. U. S. A. 103:2839–2844.
33. Kumagai, Y., J. Matsuo, Y. Hayakawa, and Y. Rikihisa. 2010. Cyclic di-GMPsignaling regulates invasion by Ehrlichia chaffeensis of human monocytes. J.Bacteriol. 192:4122–4133.
34. Lai, T. H., Y. Kumagai, M. Hyodo, Y. Hayakawa, and Y. Rikihisa. 2008. TheAnaplasma phagocytophilum PleC histidine kinase and PleD diguanylatecyclase two-component system and role of cyclic di-GMP in host-cell infec-tion. J. Bacteriol. 191:693–700.
35. Lee, H. S., F. Gu, S. M. Ching, Y. Lam, and K. L. Chua. 2010. CdpA is aBurkholderia pseudomallei cyclic di-GMP phosphodiesterase involved inautoaggregation, flagellum synthesis, motility, biofilm formation, cell inva-sion, and cytotoxicity. Infect. Immun. 78:1832–1840.
36. Lee, V. T., et al. 2007. A cyclic-di-GMP receptor required for bacterialexopolysaccharide production. Mol. Microbiol. 65:1474–1484.
37. Leonard, S., et al. 2007. FtcR is a new master regulator of the flagellarsystem of Brucella melitensis 16M with homologs in Rhizobiaceae. J. Bacte-riol. 189:131–141.
38. Lestrate, P., et al. 2003. Attenuated signature-tagged mutagenesis mutantsof Brucella melitensis identified during the acute phase of infection in mice.Infect. Immun. 71:7053–7060.
39. Letunic, I., T. Doerks, and P. Bork. 2009. SMART 6: recent updates and newdevelopments. Nucleic Acids Res. 37:D229–D232.
40. Liu, N., T. Pak, and E. M. Boon. 2010. Characterization of a diguanylatecyclase from Shewanella woodyi with cyclase and phosphodiesterase activi-ties. Mol. Biosyst. 6:1561–1564.
41. Martinez, S. E., J. A. Beavo, and W. G. Hol. 2002. GAF domains: two-billion-year-old molecular switches that bind cyclic nucleotides. Mol. Interv. 2:317–323.
42. Miao, E. A., E. Andersen-Nissen, S. E. Warren, and A. Aderem. 2007. TLR5and Ipaf: dual sensors of bacterial flagellin in the innate immune system.Semin. Immunopathol. 29:275–288.
43. Miller, J. F. 1972. Experiments in molecular genetics. Cold Spring HarborLaboratory Press, Cold Spring Harbor, NY.
44. Neunuebel, M. R., and J. W. Golden. 2008. The Anabaena sp. strain PCC7120 gene all2874 encodes a diguanylate cyclase and is required for normalheterocyst development under high-light growth conditions. J. Bacteriol.190:6829–6836.
45. Pappas, G., N. Akritidis, M. Bosilkovski, and E. Tsianos. 2005. Brucellosis.N. Engl. J. Med. 352:2325–2336.
46. Paul, R., et al. 2004. Cell cycle-dependent dynamic localization of a bacterialresponse regulator with a novel di-guanylate cyclase output domain. GenesDev. 18:715–727.
47. Pesavento, C., et al. 2008. Inverse regulatory coordination of motility andcurli-mediated adhesion in Escherichia coli. Genes Dev. 22:2434–2446.
48. Pizarro-Cerda, J., et al. 1998. Brucella abortus transits through the au-tophagic pathway and replicates in the endoplasmic reticulum of nonprofes-sional phagocytes. Infect. Immun. 66:5711–5724.
49. Putnam, S. L., and A. L. Koch. 1975. Complications in the simplest cellularenzyme assay: lysis of Escherichia coli for the assay of beta-galactosidase.Anal. Biochem. 63:350–360.
50. Rajashekara, G., D. A. Glover, M. Banai, D. O’Callaghan, and G. A. Splitter.2006. Attenuated bioluminescent Brucella melitensis mutants GR019 (virB4),GR024 (galE), and GR026 (BMEI1090-BMEI1091) confer protection inmice. Infect. Immun. 74:2925–2936.
51. Rajashekara, G., D. A. Glover, M. Krepps, and G. A. Splitter. 2005. Tem-poral analysis of pathogenic events in virulent and avirulent Brucella meliten-sis infections. Cell. Microbiol. 7:1459–1473.
52. Rajashekara, G., et al. 2005. Unraveling Brucella genomics and pathogenesisin immunocompromised IRF-1�/� mice. Am. J. Reprod. Immunol. 54:358–368.
53. Rambow-Larsen, A. A., G. Rajashekara, E. Petersen, and G. Splitter. 2008.Putative quorum-sensing regulator BlxR of Brucella melitensis regulates vir-ulence factors including the type IV secretion system and flagella. J. Bacte-riol. 190:3274–3282.
54. Rao, F., et al. 2009. The functional role of a conserved loop in EAL domain-based cyclic di-GMP-specific phosphodiesterase. J. Bacteriol. 191:4722–4731.
55. Rao, F., Y. Yang, Y. Qi, and Z. X. Liang. 2008. Catalytic mechanism of cyclicdi-GMP-specific phosphodiesterase: a study of the EAL domain-containingRocR from Pseudomonas aeruginosa. J. Bacteriol. 190:3622–3631.
56. Reguera, J. M., et al. 2003. Brucella endocarditis: clinical, diagnostic, andtherapeutic approach. Eur. J. Clin. Microbiol. Infect. Dis. 22:647–650.
57. Rittig, M. G., M. T. Alvarez-Martinez, F. Porte, J. P. Liautard, and B. Rouot.2001. Intracellular survival of Brucella spp. in human monocytes involvesconventional uptake but special phagosomes. Infect. Immun. 69:3995–4006.
58. Romling, U. 2009. Cyclic di-GMP (c-di-GMP) goes into host cells: c-di-GMPsignaling in the obligate intracellular pathogen Anaplasma phagocytophilum.J. Bacteriol. 191:683–686.
5690 PETERSEN ET AL. J. BACTERIOL.

59. Romling, U., M. Gomelsky, and M. Y. Galperin. 2005. C-di-GMP: the dawn-ing of a novel bacterial signalling system. Mol. Microbiol. 57:629–639.
60. Ryan, R. P., et al. 2006. Cell-cell signaling in Xanthomonas campestris in-volves an HD-GYP domain protein that functions in cyclic di-GMP turnover.Proc. Natl. Acad. Sci. U. S. A. 103:6712–6717.
61. Ryjenkov, D. A., R. Simm, U. Romling, and M. Gomelsky. 2006. The PilZdomain is a receptor for the second messenger c-di-GMP: the PilZ domainprotein YcgR controls motility in enterobacteria. J. Biol. Chem. 281:30310–30314.
62. Ryjenkov, D. A., M. Tarutina, O. V. Moskvin, and M. Gomelsky. 2005. Cyclicdiguanylate is a ubiquitous signaling molecule in bacteria: insights into bio-chemistry of the GGDEF protein domain. J. Bacteriol. 187:1792–1798.
63. Schmidt, A. J., D. A. Ryjenkov, and M. Gomelsky. 2005. The ubiquitousprotein domain EAL is a cyclic diguanylate-specific phosphodiesterase: en-zymatically active and inactive EAL domains. J. Bacteriol. 187:4774–4781.
64. Seleem, M. N., S. M. Boyle, and N. Sriranganathan. 2009. Brucellosis: are-emerging zoonosis. Vet. Microbiol. 140:392–398.
65. Spangler, C., A. Bohm, U. Jenal, R. Seifert, and V. Kaever. 2010. A liquidchromatography-coupled tandem mass spectrometry method for quantita-tion of cyclic di-guanosine monophosphate. J. Microbiol. Methods 81:226–231.
66. Sudarsan, N., et al. 2008. Riboswitches in eubacteria sense the secondmessenger cyclic di-GMP. Science 321:411–413.
67. Tamayo, R., J. T. Pratt, and A. Camilli. 2007. Roles of cyclic diguanylate inthe regulation of bacterial pathogenesis. Annu. Rev. Microbiol. 61:131–148.
68. Tamayo, R., A. D. Tischler, and A. Camilli. 2005. The EAL domain proteinVieA is a cyclic diguanylate phosphodiesterase. J. Biol. Chem. 280:33324–33330.
69. Tao, F., Y. W. He, D. H. Wu, S. Swarup, and L. H. Zhang. 2010. The cyclicnucleotide monophosphate domain of Xanthomonas campestris global re-gulator Clp defines a new class of cyclic di-GMP effectors. J. Bacteriol.192:1020–1029.
70. Taylor, B. L., and I. B. Zhulin. 1999. PAS domains: internal sensors ofoxygen, redox potential, and light. Microbiol. Mol. Biol. Rev. 63:479–506.
71. Tischler, A. D., and A. Camilli. 2005. Cyclic diguanylate regulates Vibriocholerae virulence gene expression. Infect. Immun. 73:5873–5882.
72. Uzureau, S., et al. 2007. Mutations of the quorum sensing-dependent regu-lator VjbR lead to drastic surface modifications in Brucella melitensis. J.Bacteriol. 189:6035–6047.
73. Weeks, J. N., et al. 2010. Brucella melitensis VjbR and C12-HSL regulons:contributions of the N-dodecanoyl homoserine lactone signaling moleculeand LuxR homologue VjbR to gene expression. BMC Microbiol. 10:167.
74. Weinhouse, H., et al. 1997. c-di-GMP-binding protein, a new factor regulat-ing cellulose synthesis in Acetobacter xylinum. FEBS Lett. 416:207–211.
75. Yildiz, F. H. 2008. Cyclic dimeric GMP signaling and regulation of surface-associated developmental programs. J. Bacteriol. 190:781–783.
76. Zygmunt, M. S., S. D. Hagius, J. V. Walker, and P. H. Elzer. 2006. Identi-fication of Brucella melitensis 16M genes required for bacterial survival in thecaprine host. Microbes Infect. 8:2849–2854.
VOL. 193, 2011 CYCLIC DI-GMP IN BRUCELLA MELITENSIS 5691





























